
Hatıra sağlamlaşması - Hatırla ve değiştir · Hatıra sağlamlaşması - Hatırla ve...
57
Hatıra sağlamlaşması - Hatırla ve değiştir Sinaptik ve sistem (yeniden) sağlamlaşması [Synaptic and systems memory (re)consolidation] Monday, November 7, 2011
Transcript of Hatıra sağlamlaşması - Hatırla ve değiştir · Hatıra sağlamlaşması - Hatırla ve...

Hatıra sağlamlaşması - Hatırla ve değiştir
Sinaptik ve sistem (yeniden) sağlamlaşması[Synaptic and systems memory (re)consolidation]
Monday, November 7, 2011

Hayvanların deneyimlerini kaydedebilme ve bu deneyimlerin doğasına göre davranışlarını değiştirebilme
yetenekleri net bir şekilde biyolojinin en önemli ve heyecan verici fenomenlerinden biri durumundadır
(James L. McGaugh, 1966, Science)
Monday, November 7, 2011

Konular
Monday, November 7, 2011

Konular
•Nöronlar ve sinapslar
Monday, November 7, 2011

Konular
•Nöronlar ve sinapslar•Öğrenmeyi mümkün kılan nöral süreçler
Monday, November 7, 2011

Konular
•Nöronlar ve sinapslar•Öğrenmeyi mümkün kılan nöral süreçler•Hatıraların sağlamlaştırılmaları (sabitlenmeleri)
Monday, November 7, 2011

Konular
•Nöronlar ve sinapslar•Öğrenmeyi mümkün kılan nöral süreçler•Hatıraların sağlamlaştırılmaları (sabitlenmeleri)•Hafıza her hatırlandığında yeniden mi yazılıyor?
Monday, November 7, 2011

Konular
•Nöronlar ve sinapslar•Öğrenmeyi mümkün kılan nöral süreçler•Hatıraların sağlamlaştırılmaları (sabitlenmeleri)•Hafıza her hatırlandığında yeniden mi yazılıyor? •Hatıraların yeniden sağlamlaştırılmaları
Monday, November 7, 2011

Bir nöron
24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
Hücre gövdesi
Mesaj yönü
Akson uçları (ileticiler)
Monday, November 7, 2011

Bir nöron
•Duysal hücreler, hareket hücreleri, ve aralarında onları “birleştiren” sinir hücreleri (nöronlar)24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
Hücre gövdesi
Mesaj yönü
Akson uçları (ileticiler)
Monday, November 7, 2011

Bir nöron
•Duysal hücreler, hareket hücreleri, ve aralarında onları “birleştiren” sinir hücreleri (nöronlar)
•Nöron doktrini: Bireysel nöronlar, sinir sisteminin iletişim birimleridir. (Cajal)
24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
Hücre gövdesi
Mesaj yönü
Akson uçları (ileticiler)
Monday, November 7, 2011

Bir nöron
•Duysal hücreler, hareket hücreleri, ve aralarında onları “birleştiren” sinir hücreleri (nöronlar)
•Nöron doktrini: Bireysel nöronlar, sinir sisteminin iletişim birimleridir. (Cajal)
24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
24 Part I / The Neurobiology of Behavior
Figure 2-4 Neurons can be classified as unipolar, bipolar, or multipolar accord- ing to the number of processes that originate from the cell body. A. Unipolar cells have a single process, with different segments serving as recep- tive surfaces or releasing terminals. Unipolar cells are characteristic of the in- vertebrate nervous system. B. Bipolar cells have two processes that are functionally specialized: the dendrite carries information to the cell, and the axon transmits information to other cells. C. Certain neurons that carry sensory in- formation, such as information about touch or stretch, to the spinal cord belong to a subclass of bipolar cells designated as pseudo-unipolar. As such cells develop, the two processes of the embryonic bipo- lar cell become fused and emerge from the cell body as a single process. This out- growth then splits into two processes, both of which function as axons, one go- ing to peripheral skin or muscle, the other going to the central spinal cord. D. Multipolar cells have an axon and many dendrites. They are the most common type of neuron in the mammalian nervous system. Three examples illustrate the large diversity of these cells. Spinal motor neurons (left) innervate skeletal muscle fibers. Pyramidal cells (middle) have a roughly triangular cell body; dendrites emerge from both the apex (the apical dendrite) and the base (the basal den- drites). Pyramidal cells are found in the hippocampus and throughout the cerebral cortex. Purkinje cells of the cerebellum (right) are characterized by the rich and ex- tensive dendritic tree in one plane. Such a structure permits enormous synaptic in- put. (Adapted from Ramon y Cajal 1933.)
A Unipolar cell B Bipolar cell
-Cell body
Axon
Invertebrate neuron Bipolar cell of retina
D Three types of multipolar cells
Motor neuron of s p n a cord
Unipolar neurons are the simplest nerve cells because they have a single primary process, which usually gives rise to many branches. One branch serves as the axon; other branches function as dendritic receiving struc- tures (Figure 2-4A). These cells predominate in the ner- vous systems of invertebrates; in vertebrates they occur in the autonomic nervous system.
Bipolar neurons have an oval-shaped soma that gives rise to two processes: a dendrite that conveys informa- tion from the periphery of the body, and an axon that carries information toward the central nervous system (Figure 2-4B). Many sensory cells are bipolar cells, in- cluding those in the retina of the eye and in the olfactory
Pyramidal cell of h~ppocampus
Peripheral axon to s k n and muscle
S n g e bifurcated 11- process
Central
Ganglion cell of dorsal root
Axon
Purknje cell of cerebellum
epithelium of the nose. The mechanoreceptors that con- vey touch, pressure, and pain to the spinal cord are vari- ants of bipolar cells called pseudo-unipolar cells. These cells develop initially as bipolar cells; later the two cell processes fuse to form one axon that emerges from the cell body. The axon then splits into two; one branch runs to the periphery (to sensory receptors in the skin, joints, and muscle), the other to the spinal cord (Figure 2-4C).
Multipolar neuvons predominate in the nervous sys- tem of vertebrates. They have a single axon and, typi- cally, many dendrites emerging from various points around the cell body (Figure 2-4D). Multipolar cells vary greatly in shape, especially in the length of their
Hücre gövdesi
Mesaj yönü
Akson uçları (ileticiler)
Monday, November 7, 2011

Hücreyi saran yarı geçirgen zar
Hücre dışı
Hücre içi
Monday, November 7, 2011

Hücreye iyon girişi sağlayan kanallar
Hücre dışı
Hücre içi
Kapalı
Açık
Monday, November 7, 2011

Nöronlar konuşurken...
•İki nöronun “konuştuğu” noktaya sinaps denir•“Sinaptik benlik” hipotezi (Changeux)
Monday, November 7, 2011

İletişimin iki yüzü
Part 111
Monday, November 7, 2011

İletişimin iki yüzü
•Elektriksel
Part 111
Monday, November 7, 2011

İletişimin iki yüzü
•Elektriksel
Part 111
Monday, November 7, 2011

İletişimin iki yüzü
•Elektriksel
• Kimyasal
Part 111
Monday, November 7, 2011

İletişimin iki yüzü
•Elektriksel
• Kimyasal
Part 111
Chapter 10 / Overview of Synaptic Transmission 183
Presynaptic acton potential
Excitatory postsynaptic potential
Act~on potenta n nerve termnal opens Ca2+channels
ca2+ entry causes vesicle fuson and transmtter release
Receptor-channels open, Nat enters the postsynaptic cell and vesces recycle
@ e' e @
N a i Na+ NaT cell
Figure 10-7 Synapt ic t ransmission at chemical synapses in- volves several steps. An action potential arriving at the termi- nal of a presynaptic axon causes voltage-gated Ca2+ channels at the active zone to open. The influx of Ca2- produces a high concentration of Ca2+ near the active zone, which in turn causes vesicles containing neurotransmitter t o fuse with the presynaptic cell membrane and release their contents into the synaptic cleft (a process termed exocytosis). The released neurotransmitter molecules then diffuse across the synaptic
Chemical Transmitters Bind to Postsynaptic Receptors
Chemical synaptic transmission can be divided into two steps: a tvnnsmitting step, in which the presynaptic cell releases a chemical messenger, and a veceptiue step, in which the transmitter binds to the receptor molecules in the postsynaptic cell.
The transmitting process resembles the release process of an endocrine gland, and chemical synaptic transmission can be seen as a modified form of hormone secretion. Both endocrine glands and presynaptic termi- nals release a chemical agent with a signaling function, and both are examples of regulated secretion (Chapter 4). Similarly, both endocrine glands and neurons are usually some distance from their target cells. There is one important difference, however. The hormone re- leased by the gland travels through the blood stream until it interacts with all cells that contain an appropri- ate receptor. A neuron, on the other hand, usually com- municates only with specific cells, the cells with which it forms synapses. Communication consists of a presynap- tic neuron sending an action potential down its axon to the axon terminal, where the electrical signal triggers
cleft and bind to specific receptors on the post-synaptic mem- brane. These receptors cause ion channels to open (or close), thereby changing the membrane conductance and membrane potential of the postsynaptic cell. The complex process of chemical synaptic transmission is responsible for the delay be- tween action potentials in the pre- and post-synaptic cells com- pared wi th the virtually instantaneous transmission of signals at electrical synapses (see Figure 10-2B). The gray filaments rep- resent the docking and release sites of the active zone.
the focused release of the chemical transmitter onto a target cell. Thus the chemical signal travels only a small distance to its target. Neuronal signaling, therefore, has two special features: It is fast and precisely directed.
To accomplish this highly directed or focused re- lease, most neurons have specialized secretory machin- ery, the active zones. In neurons without active zones the distinction between neuronal and hormonal trans- mission becomes blurred. For example, the neurons in the autonomic nervous system that innervate smooth muscle reside at some distance from their postsynaptic cells and do not have specialized release sites in their terminals. Synaptic transmission between these cells is slower and more diffuse. Furthermore, at one set of ter- minals a transmitter can be released at an active zone, as a conventional transmitter acting directly on neighbor- ing cells; at another locus it can be released in a less fo- cused way as a modulator, producing a more diffuse ac- tion; and at a third locus it can be released into the blood stream as a neurohormone.
Although a variety of chemicals serve as neuro- transmitters, including both small molecules and pep- tides (see Chapter 15), the action of a transmitter in the
Monday, November 7, 2011

Öğrenmenin “Hebb Kuralı”•Öğrenmenin (hafızanın) hücresel temeli, sinaps sonrası nöron ateşlenirken her defasında aktif olan sinapsın güçlendirilmesidir
Sinaps öncesi
Sinaps sonrası
Monday, November 7, 2011

“Hebb Kuralı”
•Birlikte ateş eden nöronlar birbirlerine bağlanırlar•Neurons that fire together, wire together
Monday, November 7, 2011

Nereden başlayalım? Hippokampus (deniz atı)
Monday, November 7, 2011

Sinaptik plastisite
•LTP = Uzun süreli potansiyalizasyon: Bir nöronun belli bir sinaptik girdi sayesinde harekete geçirilebilirliğinde bu girdinin yüksek frekansta tekrarlanması sonucu oluşan uzun süreli artış
Monday, November 7, 2011

Ne gerekiyor?
1) Bir sinapsin uyarılması
2) Sonraki hücrede aynı anda depolarizasyon
Monday, November 7, 2011

Nasıl?
AP zaten güçlü sinapsın
ileticisine geliyor (depolarizasyona
yol açıyor)
Güçlü sinaps
Depolarizasyon sonraki hücrenin ateş etmesi için
yeterli
Geri ateşleme dendrit boyunca geri dönüyor ve buradaki NMDA
reseptörlerini “hazırlıyor”
Zayıf sinapstan gelen AP geri ateşleme ile
karşılaşınca LTP meydana geliyor
LTP gerçekleşiyor ve sinaps güçleniyor
Monday, November 7, 2011

Sinaps nasıl güçlenir?
•Nitrik-oxit sayesinde salgılanan NT (haberleşme kimyasalı) miktarı artıyor (burada soru işaretleri yok değil)
•Uzun süreli değişiklikler
•Reseptör artışı (AMPA)
•Yapısal değişiklikler
Monday, November 7, 2011

“O” sinaps nasıl tanınıyor?•LTP’nin protein sentezi ile daha da uzun süreli kılınması (erken LTP’den geç LTP’ye) •Frey & Morris’in ‘tagleme’ hipotezi
LTP bu sinapsta
gerçekleştiBu olduktan
sonra kimyasal ‘tag’ler
üretiliyor
Çekirdeğe protein sentezlenmesi için
mesaj gidiyor
Proteinler tagler bu etiketler sayesinde
yakalanıyor
Monday, November 7, 2011
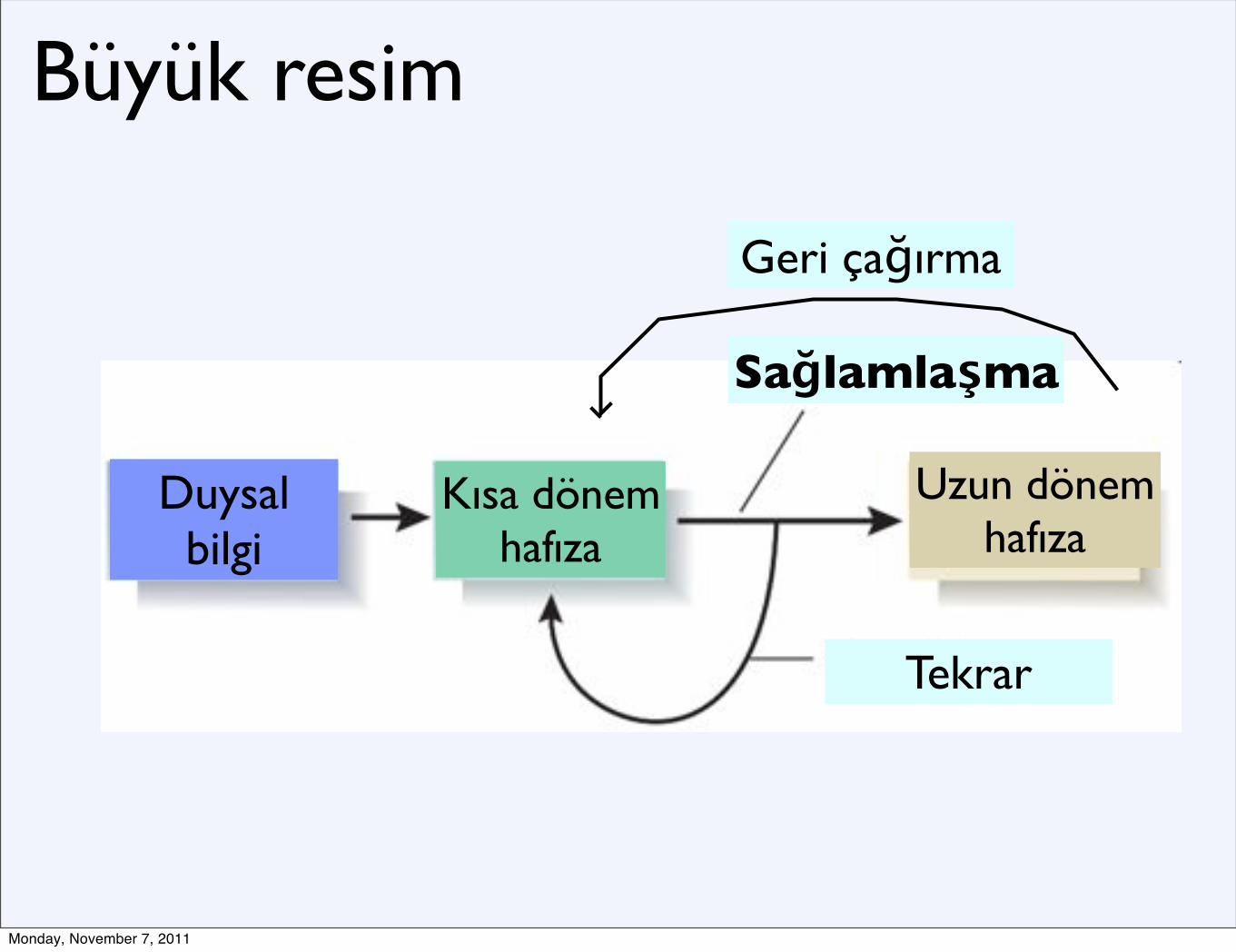
Büyük resim
Duysal bilgi
Kısa dönem hafıza
Sağlamlaşma
Uzun dönem hafıza
Tekrar
Geri çağırma
Monday, November 7, 2011

Hatıra Sağlamlaşması
•Uzun süreli hafızanın edinildikten sonra giderek daha sabit bir hale gelmesi, ve bu sabitlenmenin içerisinde gerçekleştiği bellek süreçleri (Dudai, 2002, 2004)•Sinaptik: Kısa sürede (dakikalardan saatlere)•Sistemsel: Uzun sürede (saatlerden yıllara)
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
54 DUDAI
ON THE TERMINOLOGY OF CONSOLIDATION
The term “consolidation” is currently used in the neuroscience literature to re-fer to two types of processes, or a family of processes (Dudai 1996, Dudai &Morris 2000; Figure 1). One type is accomplished within the first minutes tohours after the encoding has occurred or practice ended. Ample evidence indicatesthat this relatively fast type of process takes place in local nodes in the neuronal
Figure 1 Types of consolidation. (A) The time course of synaptic (cellular, local) con-solidation, determined by measuring the sensitivity of memory to the inhibition of proteinsynthesis. Consolidated memory is defined as treatment-resistant long-term memory. Thedata are from experiments on shuttle-box learning in the goldfish (Agranoff et al. 1966).The protein synthesis inhibitor was administered to separate groups of fish at the indicatedtime points after training. The sensitivity of memory to protein synthesis inhibition was overby about one hour. A consolidation process that depends on protein synthesis during andimmediately after training is a universal property of the nervous system. (B) The time courseof system consolidation, determined by measuring the sensitivity of long-term memory tohippocampal damage. The data are from experiments on contextual fear conditioning in therat (Kim & Fanselow 1992). The lesion was inflicted to separate groups at the indicated timepoints after training. The dependence of long-term memory on the hippocampus in this casewas over by about one month. System consolidation, lasting weeks or longer, during whichthe memory becomes independent of the hippocampus, is observed in declarative memory.(Figure adapted from Dudai 2002a.)
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Protein sentezi engellemesi
(balık)
Hipokampus hasarı (sıçan)
Monday, November 7, 2011

Sürecin bozulması
•Ögrenme sonrasi amnezik (unutkanlik yaratan) bir müdahelenin yarattığı etkinin gücü zamanın bir fonksiyonu olmalı
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
54 DUDAI
ON THE TERMINOLOGY OF CONSOLIDATION
The term “consolidation” is currently used in the neuroscience literature to re-fer to two types of processes, or a family of processes (Dudai 1996, Dudai &Morris 2000; Figure 1). One type is accomplished within the first minutes tohours after the encoding has occurred or practice ended. Ample evidence indicatesthat this relatively fast type of process takes place in local nodes in the neuronal
Figure 1 Types of consolidation. (A) The time course of synaptic (cellular, local) con-solidation, determined by measuring the sensitivity of memory to the inhibition of proteinsynthesis. Consolidated memory is defined as treatment-resistant long-term memory. Thedata are from experiments on shuttle-box learning in the goldfish (Agranoff et al. 1966).The protein synthesis inhibitor was administered to separate groups of fish at the indicatedtime points after training. The sensitivity of memory to protein synthesis inhibition was overby about one hour. A consolidation process that depends on protein synthesis during andimmediately after training is a universal property of the nervous system. (B) The time courseof system consolidation, determined by measuring the sensitivity of long-term memory tohippocampal damage. The data are from experiments on contextual fear conditioning in therat (Kim & Fanselow 1992). The lesion was inflicted to separate groups at the indicated timepoints after training. The dependence of long-term memory on the hippocampus in this casewas over by about one month. System consolidation, lasting weeks or longer, during whichthe memory becomes independent of the hippocampus, is observed in declarative memory.(Figure adapted from Dudai 2002a.)
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Protein sentezi engellemesi
(balık)
Hipokampus hasarı (sıçan)
Monday, November 7, 2011

Sinaptiğe dair
Monday, November 7, 2011

Sinaptiğe dair
•Öğrenmenin ardından belirli bir süre içinde çesitli müdaheleler hatıralarda amnezik etki yaratıyor, süre sonunda ise etkisizler (ECS, davranışsal dikkat bozucular, çeşitli ilaçlar)
Monday, November 7, 2011

Sinaptiğe dair
•Öğrenmenin ardından belirli bir süre içinde çesitli müdaheleler hatıralarda amnezik etki yaratıyor, süre sonunda ise etkisizler (ECS, davranışsal dikkat bozucular, çeşitli ilaçlar)
•Müdaheleler ya hep ya hiç değil, dereceli bir etki gösteriyor.
Monday, November 7, 2011

Sinaptiğe dair
•Öğrenmenin ardından belirli bir süre içinde çesitli müdaheleler hatıralarda amnezik etki yaratıyor, süre sonunda ise etkisizler (ECS, davranışsal dikkat bozucular, çeşitli ilaçlar)
•Müdaheleler ya hep ya hiç değil, dereceli bir etki gösteriyor. •RNA veya protein sentezini engelleyen maddeler, gen ekspresyonunun öğrenme ardından durdurulması USB’in oluşmasını engellerken, algı, KSB, hatıranın saklanması, ve geri çağırılmasında etkili değil.
Monday, November 7, 2011

Ne sorun ne söyleyeyim
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
58 DUDAI
involve coordination between the activated synapse and the nucleus, whichpossibly optimize exploitation of the metabolic resources of the neuron andthe specificity of the long-term change (Dudai & Morris 2000).
f. The long-term changes in the synapse involve trafficking of new receptormolecules and possibly other proteins into the synaptic membrane, and al-teration in the association of receptors with cellular cytoskeleton and signaltransduction cascades (e.g., El-Husseini et al. 2002, Shi et al. 1999). In ad-dition, there is evidence that long-term synaptic plasticity and long-termmemory are correlated with morphological changes in synapses (e.g., Bailey& Kandel 1993, Weiler et al. 1995).
THE STANDARD MODEL OF SYNAPTIC CONSOLIDATION
The above and additional findings served as the basis for the formulation of thestandard model of synaptic consolidation (Figure 2). This model posits that mem-ory traces can exist in at least two forms: short-term and labile, and long-term and
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Monday, November 7, 2011

Sisteme dair
•Sadece MTL hasarı genelde geriye doğru (retrograde) dereceli unutkanlık gösterirken, neokorteks hasarı da varsa unutkanlık düz olabiliyor
Beyin hasar öncesi olaylar
hatırlanmıyor
Beyin hasarının gerçekleştiği
an
Beyin hasarı sonrası olaylar KSB
sonrası hatırlanamıyor
Monday, November 7, 2011

Geriye dönük hafıza kaybı
Sağlıklı kontrollerGeriye dönük hafıza kaybı hastaları
Toplumsal olayın yaşandığı onyıl
Hat
ırla
ma
oran
ı
Monday, November 7, 2011

Ne oluyor?
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
62 DUDAI
Figure3 The standardmodel of systemconsolidation is depicted in aflowchart. Initialstorage, i.e., encoding and registration of the perceived information (Dudai 2002a),occurs in both the hippocampal system and the neocortical system. Sh(0) and Sc(0)represent the strength of the initial hippocampal and neocortical traces, respectively.These traces are expected to differ, with the hippocampal one probably representinga compressed version of the internal representation. The hippocampal representationlater becomes active either in explicit recall, or in implicit processes such as sleep.This gives rise to reinstatement of the corresponding neocortical memory, resulting inincremental adjustment of neocortical connections, probably involving local, synapticconsolidation. In parallel, memory also decays, faster in the hippocampus (Dh) than inthe cortex (Dc). The net result is that memories initially dependent on the hippocampusgradually become independent of it. In reality this happens over weeks or longer. Thehippocampal system can hence be viewed not only as a memory store but also as ateacher of the neocortical system. This process (C, rate of consolidation) is proposed toallow the hippocampal system to rapidly learn new information without disrupting oldmemory stored in the neocortex, while at the same time allowing gradual integration ofthe new information into the older, structured information. Adapted from McClellandet al. (1995).
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Monday, November 7, 2011

Ne oluyor?
•Algılanan olay hem hippokampusta hem neokortekste iz bırakıyor
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
62 DUDAI
Figure3 The standardmodel of systemconsolidation is depicted in aflowchart. Initialstorage, i.e., encoding and registration of the perceived information (Dudai 2002a),occurs in both the hippocampal system and the neocortical system. Sh(0) and Sc(0)represent the strength of the initial hippocampal and neocortical traces, respectively.These traces are expected to differ, with the hippocampal one probably representinga compressed version of the internal representation. The hippocampal representationlater becomes active either in explicit recall, or in implicit processes such as sleep.This gives rise to reinstatement of the corresponding neocortical memory, resulting inincremental adjustment of neocortical connections, probably involving local, synapticconsolidation. In parallel, memory also decays, faster in the hippocampus (Dh) than inthe cortex (Dc). The net result is that memories initially dependent on the hippocampusgradually become independent of it. In reality this happens over weeks or longer. Thehippocampal system can hence be viewed not only as a memory store but also as ateacher of the neocortical system. This process (C, rate of consolidation) is proposed toallow the hippocampal system to rapidly learn new information without disrupting oldmemory stored in the neocortex, while at the same time allowing gradual integration ofthe new information into the older, structured information. Adapted from McClellandet al. (1995).
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Monday, November 7, 2011

Ne oluyor?
•Algılanan olay hem hippokampusta hem neokortekste iz bırakıyor•Hippokampustaki iz diğerinden çok daha güçlü
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
62 DUDAI
Figure3 The standardmodel of systemconsolidation is depicted in aflowchart. Initialstorage, i.e., encoding and registration of the perceived information (Dudai 2002a),occurs in both the hippocampal system and the neocortical system. Sh(0) and Sc(0)represent the strength of the initial hippocampal and neocortical traces, respectively.These traces are expected to differ, with the hippocampal one probably representinga compressed version of the internal representation. The hippocampal representationlater becomes active either in explicit recall, or in implicit processes such as sleep.This gives rise to reinstatement of the corresponding neocortical memory, resulting inincremental adjustment of neocortical connections, probably involving local, synapticconsolidation. In parallel, memory also decays, faster in the hippocampus (Dh) than inthe cortex (Dc). The net result is that memories initially dependent on the hippocampusgradually become independent of it. In reality this happens over weeks or longer. Thehippocampal system can hence be viewed not only as a memory store but also as ateacher of the neocortical system. This process (C, rate of consolidation) is proposed toallow the hippocampal system to rapidly learn new information without disrupting oldmemory stored in the neocortex, while at the same time allowing gradual integration ofthe new information into the older, structured information. Adapted from McClellandet al. (1995).
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Monday, November 7, 2011

Ne oluyor?
•Algılanan olay hem hippokampusta hem neokortekste iz bırakıyor•Hippokampustaki iz diğerinden çok daha güçlü •Hippokampus izi, gerek hatırlama, gerekse uykuda yeniden aktifleşince neokorteks bağlantıları da ayarlanıyor
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
62 DUDAI
Figure3 The standardmodel of systemconsolidation is depicted in aflowchart. Initialstorage, i.e., encoding and registration of the perceived information (Dudai 2002a),occurs in both the hippocampal system and the neocortical system. Sh(0) and Sc(0)represent the strength of the initial hippocampal and neocortical traces, respectively.These traces are expected to differ, with the hippocampal one probably representinga compressed version of the internal representation. The hippocampal representationlater becomes active either in explicit recall, or in implicit processes such as sleep.This gives rise to reinstatement of the corresponding neocortical memory, resulting inincremental adjustment of neocortical connections, probably involving local, synapticconsolidation. In parallel, memory also decays, faster in the hippocampus (Dh) than inthe cortex (Dc). The net result is that memories initially dependent on the hippocampusgradually become independent of it. In reality this happens over weeks or longer. Thehippocampal system can hence be viewed not only as a memory store but also as ateacher of the neocortical system. This process (C, rate of consolidation) is proposed toallow the hippocampal system to rapidly learn new information without disrupting oldmemory stored in the neocortex, while at the same time allowing gradual integration ofthe new information into the older, structured information. Adapted from McClellandet al. (1995).
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Monday, November 7, 2011

Ne oluyor?
•Algılanan olay hem hippokampusta hem neokortekste iz bırakıyor•Hippokampustaki iz diğerinden çok daha güçlü •Hippokampus izi, gerek hatırlama, gerekse uykuda yeniden aktifleşince neokorteks bağlantıları da ayarlanıyor
•İz hippokampusta daha hızlı kayboluyor
2 Dec 2003 18:37 AR AR207-PS55-03.tex AR207-PS55-03.sgm LaTeX2e(2002/01/18) P1: GCE
62 DUDAI
Figure3 The standardmodel of systemconsolidation is depicted in aflowchart. Initialstorage, i.e., encoding and registration of the perceived information (Dudai 2002a),occurs in both the hippocampal system and the neocortical system. Sh(0) and Sc(0)represent the strength of the initial hippocampal and neocortical traces, respectively.These traces are expected to differ, with the hippocampal one probably representinga compressed version of the internal representation. The hippocampal representationlater becomes active either in explicit recall, or in implicit processes such as sleep.This gives rise to reinstatement of the corresponding neocortical memory, resulting inincremental adjustment of neocortical connections, probably involving local, synapticconsolidation. In parallel, memory also decays, faster in the hippocampus (Dh) than inthe cortex (Dc). The net result is that memories initially dependent on the hippocampusgradually become independent of it. In reality this happens over weeks or longer. Thehippocampal system can hence be viewed not only as a memory store but also as ateacher of the neocortical system. This process (C, rate of consolidation) is proposed toallow the hippocampal system to rapidly learn new information without disrupting oldmemory stored in the neocortex, while at the same time allowing gradual integration ofthe new information into the older, structured information. Adapted from McClellandet al. (1995).
Ann
u. R
ev. P
sych
ol. 2
004.
55:5
1-86
. Dow
nloa
ded
from
ww
w.a
nnua
lrevi
ews.o
rgby
WIB
6113
- U
nive
rsita
tsbib
lioth
ek O
snab
ruck
on
09/2
3/11
. For
per
sona
l use
onl
y.
Monday, November 7, 2011

Sisteme dair
•Amnezi hastaları•Laboratuvar hayvanlarında hippokampus hasarı sonrası aynı dereceli unutkanlığın gözlenmesi •Hippokampal hemodinamik aktivite 3 yıl öncesi gibi yakın zamanlı olaylarda bir kaç on yıl önceki olaylarin hatırlanmasına oranla çok daha aktif (Smith & Squire, 2009)
•Derhal gen ekspresyonu (IEG) öğrenme sonrasındaki haftalarda hippokampal formasyondan neokorteks kayıyor
Monday, November 7, 2011

Sağlamlaşma akla yatkın mı?
•Hafıza değişebilir demedik mi?•Sahte hafızalar vs.
Monday, November 7, 2011

Peki bu ne anlama geliyor?
•Misanin, Miller & Lewis; 1968
Hafıza yok
Öğrenme
Öğrenme
Öğrenme Sadece koşullu uyaran uyarılması
Hafıza var
Hafıza yok
Monday, November 7, 2011

Yeniden sağlamlaşma!
•Sağlamlaşma esnasında hatıranın değişebilir, zarar görebilir olduğunu gördük
•Bu aşamadan sonra ise hatıranın bir daha değismemek üzere sabitlendiği fikri 100 yıla yakın doğru kabul edildi•Ama bu hafızanin dinamik şekilde değişebildiği bulguları ile uyumlu değil•Son deneyde ise bir gün sonra hatırlatılan ögrenmenin yeniden aynı kırılgan, değişebilir duruma geldiğini gördük•30 sene kadar uyuyan bu erken bulgular 2000’de mihen taşı bir çalışma ile şu anda çok yoğun bir şekilde yeniden ele alınır oldu
Nader, 2003, TiNSMonday, November 7, 2011

Korkuyu silmek
•Kullanılan bilgiler
!"##"$% #& '(#)$"
*++ !"#$%& ' ()* +,- ' ./ "$0$1# 2,,, ' 333456789:4;<=
>?7:> ?5 :6;@ ;<857AB C:D:5C?5E <5 7@: D<D8F69?7A <G 6 E?H:5 =?I789: 3?7@ G69=:9>4 JF<7>965E:C G9<= .,, 7< +K, =2 :6;@B C:D:5C?5E <5 L:FC >?M:4
189H:A DF<7> 3:9: 6>>:>>:C ?5 F67: "8E8>7 G<9 7@: >:H:9?7A <G NF6>7 >A=D7<=>B :ID9:>>:C 6>7@: D:9;:576E: <G D65?;F: N965;@:> 7@67 3:9: 5:;9<7?; C8: 7< 7@: :GG:;7> <G!" #$%&'(4 O?>:6>:36> 6>>:>>:C 67 LH: >6=DF?5E D<?57> ?5 :6;@ DF<7B C?>79?N87:C ?5 6 85?G<9= D677:954 #3:57A@?FF> 9:>8F7?5E G9<= 7@: 7965>DF657?5E D9<;:>> 3:9: :H6F867:C 67 :6;@ >6=DF?5E D<?57B 3?7@:6;@ @?FF ;<576?5?5E 6N<87 ., D65?;F:> D:9 @?FFB 7< E?H: 6 7<76F <G 6DD9<I?=67:FA .B,,,D65?;F:> :H6F867:C D:9 DF<74 &6;@ >6=DF:C D65?;F: 36> H?>86FFA :I6=?5:C NA :ID:9?:5;:CD:9><55:F 7< :>7?=67: 7@: D:9;:576E: <G N965;@:> 7@67 3:9: 5:;9<7?; C8: 7< ?5G:;7?<5 NA!"#$%&'(4 &6;@ D65?;F: 36> E?H:5 6 967?5E2P G9<= , 7< KB 3@:9: , ?> 5< C?>:6>:Q . ?> F:>> 7@65 KR<G D65?;F: N965;@:> 5:;9<7?;Q 2 ?> KST,R 5:;9<7?;Q T ?> T,SK,R 5:;9<7?;Q + ?> E9:67:9 7@65K,R 5:;9<7?;Q 65C K ?> .,,R 5:;9<7?;4 O?>:6>: >:H:9?7A 36> >8==69?M:C 3?7@?5 :6;@ DF<7 6>U!"). ! .# $ ")2 ! 2# $ ")T ! T# $ ")+ ! +# $ ")K ! K#%!!),!)KV ! .,,B 3@:9: ),444)K ?>7@: 58=N:9 <G ;8F=> ?5 :6;@ <G 7@: 9:>D:;7?H: C?>:6>: ;67:E<9?:>4 #@8>B 6 C?>:6>: >:H:9?7A <G,R 3<8FC ?5C?;67: 5< C?>:6>: 65C .,,R 3<8FC ?5C?;67: 7@67 .,,R <G D65?;F: N965;@:>3:9: 5:;9<7?;4
!"#$% #&'$(')"*+JF<7> 3:9: @65CW@69H:>7:CB 7@9:>@:C 65C 3:?E@:C 7< C:7:9=?5: E96?5 A?:FC4 X5C?H?C86FH69?:7?:> 3:9: :H6F867:C >:D6967:FA ?5 =?I789:>4 *65C :Y8?H6F:57 967?<>22 3:9: ;6F;8F67:C 6>ZA?:FC @6". <G H69?:7A " ?5 =?I789:[A?:FC @6".<G H69?:7A " ?5 =<5<;8F789:\ ] ZA?:FC @6". <GH69?:7A ^ ?5 =?I789:[A?:FC @6". <G H69?:7A ^ ?5 =<5<;8F789:\4
,)')"-)".'$ '+'$/-#-&6;@ >89H:A DF<7 36> ;<5>?C:9:C 7< N: 65 :ID:9?=:576F 85?7B 65C 656FA>:> 3:9: N6>:C <5=:65 C?>:6>: >:H:9?7?:> 65C E96?5 A?:FC G<9 :6;@ DF<74 1767?>7?;6F 656FA>:> 3:9: ;<5C8;7:C>:D6967:FA NA A:69 65C ;<857A <3?5E 7< C?GG:9:5;:> ?5 C?>:6>: F:H:F4 )5:W76?F:C *W7:>7> 3:9:8>:C 7< C:7:9=?5: ?G NF6>7 >:H:9?7A G<9 :6;@ <G 7@: 73< H69?:7?:> ?5 :6;@ <G 7@: G<89 =?I789:>C?GG:9:C >?E5?L;657FA G9<= ?7> ;<99:>D<5C?5E =<5<;8F789: ;<579<F4
%:;:?H:C ._ "D9?FQ 6;;:D7:C T, `85: 2,,,4
.4 ^9<35?5EB `4 "4 a b9:AB c4 `4 d8F7?F?5: ;8F7?H69> 6> 6 =:65> <G C?>:6>: ;<579<F4 +))," -'." /01*23(*024"
!" TKKST_2 Z.P-P\4
24 e<FG:B d4 14 #@: ;899:57 >7678> 65C D9<>D:;7> <G =8F7?F?5: ;8F7?H69> 65C H69?:7A =?I789:> G<9 C?>:6>:
9:>?>765;:4 +))," -'." /01*23(*024" #$" 2K.S2/T Z.P_K\4
T4 d85C7B f4 f4 ?5 -%5' 64(&* 7%&'(&' Z:C> g:?EF:9B %4 14B *:<5EB 14 "4 a #:5EB J4 14\ 2PTST,_ Zf"^
X57:9567?<56FB e6FF?5EG<9CB .PP+\4
+4 0699:77B c4 "4 a d85C7B f4 f4 &D?C:=?<F<EA ?5 =?I:C @<>7 D<D8F67?<5>4 /01*23(*0242#1 %&" P_+SPP,
Z.PPP\4
K4 )8B 14 h4 -%5' 7%&'(&'& 25C :C5 Zf<==<53:6F7@ dA;<F<E?;6F X5>7?787:B c:3B .P_K\4
-4 J<F?>B 04 "4B "5C:9><5B e4 ^4 a h<F7B %4 O4 #<369C 65 ?57:E967?<5 <G F65C>;6D: 65C G<<C 3:N :;<F<EAi
#@: CA56=?;> <G >D67?6FFA >8N>?C?M:C G<<C 3:N>4 +))," -'." 8524" 91&*" #%" 2_PST.- Z.PP/\4
/4 O3A:9B 04B &Fj?57<5B `4 14 a h6k:jB "4 &4 1D67?6F >;6F: 65C 7@: >D9:6C <G 6 G85E6F D67@<E:5 <G EAD>A
=<7@4 +:" ;(*" '(#" +_KS+P+ Z.PP_\4
_4 1;@3697MB d4 e4 f@<<>?5E 7@: 6DD9<D9?67: >;6F: <G 9:>:9H:> G<9 ;<5>:9H67?<54 +))," -'." 8524" 91&*" $)"
_TS.,_ Z.PPP\4
P4 e6?C:B %4 ^4 '* (4" #@: 9:F67?<5>@?D N:73::5 D9<C8;7?H?7A 65C >D:;?:> 9?;@5:>>4 +))," -'." 8524" 91&*"
$)" 2K/ST,, Z.PPP\4
.,4 d67><5B J4 "4B J697<5B e4 4B J<3:9B "4 04 a 13?G7B d4 4 "E9?;8F7896F ?57:5>?L;67?<5 65C :;<>A>7:=
D9<D:97?:>4 95%')5' #!!" K,+SK,_ Z.PP/\4
..4 #?F=65B O4 #@: E9::5?5E <G 7@: E9::5 9:H<F87?<54 ;(*,$' $&*" 2..S2.2 Z.PP_\4
.24 X57:9567?<56F %?;: %:>:69;@ X5>7?787:4 <--< -%5' =(5*& ZX57:9567?<56F %?;: %:>:69;@ X5>7?787:B d65?F6B
.PP/\4
.T4 ^6j:9B ^4B g6=N9A>j?B J4B 176>j63?;M a O?5:>@Wc8=69B 14 J4 1?E56F?5E ?5 DF657W=?;9<N: ?57:96;7?<5>4
95%')5' #!*" /2-S/TT Z.PP/\4
.+4 176>j63?;MB ^4 `4B "8>8N:FB b4 d4B ^6j:9B ^4 `4B &FF?>B 4 04 a `<5:>B 4 O4 04 d<F:;8F69 E:5:7?;> <G DF657
C?>:6>: 9:>?>765;:4 95%')5' #*%" --.S--/ Z.PPK\4
.K4 )8B 14 h4 J67@<E:5?;?7A 65C @<>7 DF657 9:>?>765;: ?5 9?;: NF6>7 C?>:6>:4+))," -'." /01*23(*024" '%" .-/S
._/ Z.P_,\4
.-4 ^<5=65B `4 d4B c@8>@B 04 14 a !:F><5B %4 `4 ^9::C?5E 9?;: G<9 9:>?>765;: 7< D:>7>4 +))," -'."
/01*23(*024" $)" K,/SK2_ Z.PP2\4
./4 c?A<>636B 14 0:5:7?;> 65C :D?C:=?<F<E?;6F =<C:F?5E <G N9:6jC<35 <G DF657 C?>:6>: 9:>?>765;:4 +)),"
-'." /01*23(*024" #)" PTS../ Z.P_2\4
._4 ^<8C9:68B d4 "4 a d85C7B f4 f4 ?5 8).%$2):')*(441 9(>' +33$2(50'& *2 7%&'(&' ?2)*$24 Z:C> %:;@;?EFB
4 a %:;@;?EFB !4\ TTS-2 Zf%fB ^<;6 %67<5B .PP/\4
.P4 f@?5B c4 d4 a e<FG:B d4 14 1:F:;7?<5 <5 8$1&%30' #$(:%)%& ?5 D89: 65C =?I:C >765C> <G N69F:A4 /4()*
/(*024" $$" _PS.,, Z.P_+\4
2,4 f6F<55:;B "4B 0<A:68B h4 a C: (6FF6H?:?FF:WJ<D:B f4 &GG:;7> <G ?5C8;:C 9:>?>765;: <5 ?5G:;7?<5
:GL;?:5;A 65C >D<98F67?<5 <G /,55%)%( &*$%%>2$:%& <5 >::CF?5E> ?5 H69?:76F =?I789:> 65C <5 L:FC
:D?C:=?;> ?5 D89: >765C>4 8,$" @" /4()* /(*024" ')#" /TTS/+. Z.PP-\4
2.4 0:<9E:B d4 *4B !:F><5B %4 4B g:?EF:9B %4 14 a *:85EB h4 %6D?C D<D8F67?<5 656FA>?> <G!(#)(32$*0' #$%&'(
NA 8>?5E 9:DWJf% 65C :5C<E:5<8> 9:D:7?7?H: O!" >:Y8:5;:>4 /01*23(*0242#1 %%" 22TS22P Z.PP_\4
224 b965;?>B f4 "4 ?5 !,4*%34' ?$233%)# 91&*':& Z:C4 b965;?>B f4 "4\ .S.P Zd6;d?FF65B !:3 l<9jB .P_-\4
2T4 d85C7B f4 f4 ?5 /4()* 7%&'(&' 83%A':%242#1 Z:C> *:<569CB c4 `4 a b9AB e4 &4\ .K,S._. Zd;0963Wh?FFB
.P_P\4
2+4 d85C7B f4 f a ^9<35?5EB `4 "4 ?5 B0' ?'$'(4 -,&*&(<F4 2 Z:C> %<:FG>B "4 J4 a ^8>@5:FFB e4 %4\ K2/SK-,
Z";6C:=?;B )9F65C<B .P_K\4
2K4 ^9<35?5EB `4 "4 a b9:AB c4 4 ?5 9*$(*'#%'& >2$ *0' ?2)*$24 2> ?'$'(4 7%&'(&' Z:C> `:5jA5B 4 b4 a JF8=NB %4
#4\ T/S+- Z^F6;j3:FFB )IG<9CB .P_.\4
2-4 e<FG:B d4 14 ?5 6($4'1 C')'*%5& D< Z:C4 d85;jB *4\ .,KKS.,-/ Zd85j>E669CB f<D:5@6E:5B .PP2\4
2/4 d655B f4 %:>::C?5E 7@: 09::5 %:H<F87?<54 95%')5' #!!" .,T_S.,+2 Z.PP/\4
2_4 1;@=?C7B %4 "4 ?5 /4()* 7%&'(&'E +) +A.()5'A B$'(*%&' Z:C> h<9>G6FFB `4 0 a f<3F?5EB &4 ^4\ 2_/ST.K
Z";6C:=?;B !:3 l<9jB .P/_\4
2P4 B0' 9*(*' 9*()A($A 2> *0' /'234'F& -'3,G4%5 2> ?0%)(H ;2" C6IB JKLMNOJMMK" .S.T Zf@?56 1765C69C
J9:>>B ^:?k?5EB .PP-\4
0.1+*2$#%3#4#+)-#@?> 3<9j 36> >8DD<97:C NA 7@: ">?65 O:H:F<D=:57 ^65jB 7@: l85565 J9<H?5;:0<H:95=:57B #@: d?5?>79A <G 1;?:5;: 65C #:;@5<F<EA <G f@?56B 7@: X57:9567?<56F %?;:%:>:69;@ X5>7?787: ZX%%X\B 65C 6 >;?:57?L; 6E9::=:57 N:73::5 X%%X 65C )9:E<5 1767:$5?H:9>?7A4 e: 7@65j 7@: D:9><55:F <G 7@: D9<H?5;?6F 65C ;<857A JF657 J9<7:;7?<5 1767?<5>65C D697?;?D67?5E G69=:9> G<9 7@:?9 ;<579?N87?<5> 7< 7@?> D9<k:;7B 65C d4 h<GG:9 G<9;<=D87:9 6>>?>765;: 65C E96D@?;>4
f<99:>D<5C:5;: 65C 9:Y8:>7> G<9 =67:9?6F> >@<8FC N: 6CC9:>>:C 7< f4f4d4Z:W=6?Fi =85C7;mN;;4<9>74:C8\4
44444444444444444444444444444444444444444444444444444444444444444!"#$%"%&$'"( $")*'$" +$&,"'-(.-,/"('( '- ,/" #%.01#2# 3&$$"4&-(&2'1#,'&- #3,"$ $",$'"5#26#$'% 7#1"$8 92"-- :; <4/#3" = >&("+/ :; ?" @&*A
P" !" Q'5R =2,)A(*%2) S(G2$(*2$1 2> ;',$2G%242#1H ?')*'$ >2$ ;',$(4 95%')5'H;'T U2$R V)%.'$&%*1H ;'T U2$RH ;'T U2$R JNNNWH V9+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
,-./0 1.1234.5 63. 47484699: 96;49. 67< 5.75484=. 82 <453>?8427;.@23. ;.47A B275294<68.< 4782 586;9. 927AC8.31 1.1234.5'D(+E>BF .=4<.7B. 47<4B68.5 8F68 8F45 B275294<68427 47=29=.5 8F.5:78F.545 2@ 7./ ?328.475 47 7.>3275*D&+ GF. 968.369 67< ;65697>B9.4 2@ 8F. 61:A<696 HIJKL 63. ;.94.=.< 82 ;. 6 548. 2@ 1.123:58236A. 47 @.63 9.63747A')+ M7@>5427 2@ 8F. ?328.47 5:78F.54547F4;4823 674521:B47 4782 8F. IJK 5F2389: [email protected] 8364747A ?3.=.785
C
L
B
L C
L
B
L
L
B
L
L
B
L
–2.8
–3.8
CL
B
L C
L
B
L
–3.3
C
L
B
L
BC
–2.3
5"3(6# 7 !"#$%&'(" )$*)$+$,'&'(-, -. '#$ &%/01&2& &' .-3) 1(..$)$,' )-+')&24"&31&2
*2&,$+5 6#$ ,3%7$)+ )$*)$+$,' '#$ *-+'$)(-) "--)1(,&'$ .)-% 7)$0%&5 8,9$"'-)
*2&"$%$,'+ (, '#$ :;< &)$ )$*)$+$,'$1 7/ '#$ =22$1 +/%7-2+> 72&"? =22$1 +@3&)$+
)$*)$+$,' <!AB 0)-3* *2&"$%$,'+C 0)$/ =22$1 ')(&,02$+ )$*)$+$,' '#$ 2-DE1-+$
&,(+-%/"(,C &,1 72&"? =22$1 "()"2$+ )$*)$+$,' #(0#E1-+$ 0)-3*5 :C 2&'$)&2 ,3"2$3+> ;C 7&+&2
,3"2$3+> AC "$,')&2 ,3"2$3+5 6#$ *2&"$%$,'+ .-) +37+$@3$,' $F*$)(%$,'+ &22 1$%-,+')&'$
+(%(2&) 1(+')(73'(-,+ &+ (, '#(+ $F*$)(%$,' &,1 '#$)$.-)$ &)$ ,-' +#-D,5
© 2000 Macmillan Magazines Ltd
Nature, 2000
Protein sentezi sadece LBA’da engellenecek!
Monday, November 7, 2011

Sonuçlar
•Nader et al., 2000, Nature
!"##"$% #& '(#)$"
!"#$%& ' ()* +,- ' ./ "$0$1# 2,,, ' 333456789:4;<= *+,
!"#$"%&'()&"# "* *+(, -+-",&+$../ 0+,+ 1+ $2"1 )2() !"#$"%&'()+'*+(, -+-",&+$3 12+# ,+(!)&4()+' '5, ,+),&+4(%3 ,+)5,# )" (%(7&%+ $)()+ &# 12&!2 &#*5$&"# "* (#&$"-8!&# $2",)%8 (*)+, -+-",8,+(!)&4()&"# 9,"'5!+$ (-#+$&( "# %()+, )+$)$3 ,+6(,'%+$$ "*12+)2+, ,+(!)&4()&"# 1($ 9+,*",-+' . ", .: '(8$ (*)+, !"#'&)&"#;/ <2+ $(-+ ),+()-+#) 1&)2 (#&$"-8! &# )2+ (7$+#!+ "*-+-",8 ,+(!)&4()&"#3 %+*) -+-",8 &#)(!)/ ="#$&$)+#) 1&)2 ()&-+;%&-&)+' ,"%+ *", 9,")+&# $8#)2+$&$ 9,"'5!)&"# &# !"#$"%&'(;)&"#3 '+%(8 "* )2+ &#*5$&"# 5#)&% $&> 2"5,$ (*)+, -+-",8 ,+(!)&4(;)&"# 9,"'5!+' #" (-#+$&(/ ?5, '()( $2"1 )2() !"#$"%&'()+' *+(,-+-",&+$3 12+# ,+(!)&4()+'3 ,+)5,# )" ( %(7&%+ $)()+ )2() ,+@5&,+$'+ #"4" 9,")+&# $8#)2+$&$ *", ,+!"#$"%&'()&"#/ <2+$+ A#'$ (,+#") 9,+'&!)+' 78 ),('&)&"#(% )2+",&+$ "* -+-",8 !"#$"%&'()&"#/#>: ?@:6 7>67 5:3 =:=<9?:A B< 7>9<8B> 65 ?5?7?6C C6D?C: E:9?<@
D:F<9: D:?5B ;<5A<C?@67:@ ?57< A76DC: C<5BG7:9= =:=<9?:A ?A 65:579:5;>:@ E697 <F EAH;><C<B?;6C 65@ 5:89<D?<C<B?;6C =<@:CA <F=:=<9H.24 I<9 :J6=EC:K 7>:9: ?A ;<5A?@:96DC: :L?@:5;: 7>67 7>:F<9=67?<5 <F 6 C<5BG7:9= =:=<9H ;65 D: @?A98E7:@ DH ;:976?579:67=:57AK A8;> 6A AHA7:=?; @98B ?5M:;7?<5A <9 :C:;79<;<5L8CA?L:A><;NK B?L:5 A><97CH 6F7:9 796?5?5BK D87 7>67 7>: A6=: 79:67=:57AB?L:5 A:L:96C ><89A <9 @6HA C67:9 >6L: 5< :FF:;74 )5: <F 7>: =<A7;<==<5CH 8A:@ @98B =65?E8C67?<5A ?5L<CL:A 7>: 6@=?5?A7967?<5 <F@98BA 7>67 DC<;N 7>: 7965AC67?<5 <F %!" ?57< E9<7:?54 178@?:A <F7>?A 7HE: ?5@?;67: 7>67 =:=<9H ;<5A<C?@67?<5 ?5L<CL:A E9<7:?5AH57>:A?AOPQ4R7 >6A 6CA< D::5 9:E<97:@ 7>67 :C:;79<;<5L8CA?L: A><;N <9 AHA7:=?;
@98B 6@=?5?A7967?<5 B?L:5 6F7:9 =:=<9H 9:6;7?L67?<5 S9:79?:L6CT ;65;68A: 65 6=5:A?6 F<9 7>: <9?B?56C C:695?5B.UP.-K 3>?;> ?5@?;67:A 7>67;<5A<C?@67:@ =:=<9?:A =?B>7 D:;<=: C6D?C: 3>:5 9:79?:L:@K 65@=?B>7 :L:5 9:V8?9: 9:;<5A<C?@67?<54 W:9: 3: :J6=?5: 3>:7>:99:;<5A<C?@67?<5 ?5L<CL?5B E9<7:?5 AH57>:A?A ?A 9:V8?9:@ F<9 9:79?:L:@=:=<9?:A 7< E:9A?A74 X: 8A: 6 D:>6L?<896C E696@?B=K 68@?7<9H F:69;<5@?7?<5?5BK F<9 3>?;> 7>: 5:896C ;?9;8?7 85@:9CH?5B =:=<9HF<9=67?<5 ?A 3:CC ;>696;7:9?Y:@./P.Q4 #>?A 6CC<3A 8A 7< =65?E8C67:
=:=<9H 67 ?7A E9:A8=:@ C<;8A <F A7<96B:K ?5 ;<5796A7 7< E6A7 A78@?:A?5 3>?;> @98BA 3:9: 6@=?5?A7:9:@ AHA7:=?;6CCH4 1E:;?Z;6CCHK 3:769B:7 ?5F8A?<5A <F 65?A<=H;?5K 65 ?5>?D?7<9 <F E9<7:?5 AH57>:A?AK7< 7>: *["K 6 9:B?<5 ?=EC?;67:@ ?5 F:69 C:695?5B DH C:A?<5K E>69=6G;<C<B?;6C 65@ E>HA?<C<B?;6C Z5@?5BA./P.Q4 \9:L?<8ACHK 3: A><3:@ 7>67?5F8A?<5A <F 65?A<=H;?5 B?L:5 6F7:9 796?5?5B DC<;N C<5BG7:9= D875<7 A><97G7:9= =:=<9H <F 68@?7<9H F:69 ;<5@?7?<5?5B..4 W:9: 3::J6=?5: 7>: :FF:;7A <F A?=?C69 =65?E8C67?<5A 6@=?5?A7:9:@ 6F7:99:79?:L6C4%67A 3:9: B?L:5 6 A?5BC: E6?9?5B <F 6 7<5: S;<5@?7?<5:@ A7?=8C8AK
]1T 65@ F<<7GA><;N S85;<5@?7?<5:@ A7?=8C8AK $1T4 )5 7:A7 @6HAK?==<D?C?7H SF9::Y?5BT 36A 8A:@ 6A 65 ?5@:J <F F:69 C:695?5B2,4#3:57HGF<89 ><89A C67:9K 7>: 967A 9:;:?L:@ 6 A?5BC: ]1 E9:A:5767?<5S7:A7 .T ?==:@?67:CH F<CC<3:@ DH D?C67:96C ?5F8A?<5A <F 65?A<=H;?5<9 L:>?;C: S697?Z;?6C ;:9:D9<AE?56C ^8?@_ "]1IT ?57< 7>: *["4I9::Y?5B ?5 7:A7 . 36A AE:;?Z; 7< 7>: ]1 65@ ;<=E696DC: 6;9<AAB9<8EA4 "5 656CHA?A <F L69?65;: S"!)("T 7>67 ;<=E69:@ F9::Y?5B@89?5B 7>: E9:G]1 <9 ]1 E:9?<@A 6;9<AA B9<8EA ?5@?;67:@ 7>67 7>:9:36A 5< ?57:96;7?<5 D:73::5 7>:A: 73< L69?6DC:A S!!2! .`" # ."-TK 5<965 :FF:;7 <F B9<8E S!!2! .`" # ."+T4 W<3:L:9K 7>:9: 36A 6 A?B5?Z;657:FF:;7 <F E:9?<@ S!!.! .`" # .-,K "! ,",.T4 #3:57HGF<89 ><89A 6F7:97:A7 .K 7>: 967A 3:9: E9:A:57:@ 3?7> 7>9:: ]1A S7:A7 2T4 "5?A<=H;?5E9<@8;:@ 6 @<A:G@:E:5@:57 @:;9:6A: ?5 F9::Y?5B ?5 9:AE<5A: 7< 7>:]1 ?5 7:A7 2 SI?BA .K 26P;T4 "5 "!)(" 9:L:6C:@ 6 =6?5 :FF:;7 <FB9<8E S!!2! .`" # .2K "! ,",.T4 " !:3=65Pa:8CA E<A7 ><; 7:A79:L:6C:@ 7>67 7>: C<3G@<A: 65?A<=H;?5 65@ "]1I B9<8EA 3:9:A?=?C69 7< :6;> <7>:9 S"" ,",OTK D87 D<7> 3:9: A?B5?Z;657CH@?FF:9:57 F9<= 7>: >?B>G@<A: B9<8E S" L6C8:A! ,",.T4 &J7?5;7?<536A <DA:9L:@ <L:9 7>: 7>9:: ]1 E9:A:5767?<5A S=6?5 :FF:;7 <F 79?6CS!!2! U-" # /K "! ,",.TK D87 7>:9: 36A 5< ?57:96;7?<5 D:73::5 79?6CA65@ B9<8E S! ! .T4 #>?A :FF:;7 <F 65?A<=H;?5 9:V8?9:A 7>67 7>:=:=<9H D: 6;7?L:CH 9:79?:L:@K 6A <=?AA?<5 <F 7>: ]1 D:F<9: 65?A<G=H;?5 ?5F8A?<5 ?5 7:A7 . C:@ 7< 5<9=6C ;<5@?7?<5:@ F:69 9:AE<5A:A ?57:A7 2 SI?B4 2@K :_ 5< =6?5 :FF:;7 <F B9<8E S! ! .T4 #>: C677:9 Z5@?5B
!"#$%& ' ! "#$" %& '(#"(#) *%+$%,-./"#. &#/) 0#0%)-#$ */+ 1#*%0# ,/1-,# '(#+
)#/*"-2/"#.3 (4 5(# 1#(/2-%6)/, 7)%*#.6)# 6$#. &%) #87#)-0#+" 9!3 )4 :)##;-+< "% "(# =>
%+ "#$" 9 '/$ *%07/)/1,# /*)%$$ <)%67$ /+. '/$ $7#*-?* "% "(# =>3 *4 @+")/ABC!/+-$%0D*-+ -+&6$-%+$ /&"#) )#/*"-2/"-%+ %& / *%+$%,-./"#. &#/) 0#0%)D 7)%.6*# /0+#$-/
&%) "(# %)-<-+/, ,#/)+-+<4 /$ 0#/$6)#. %+ "#$" E3 +4 &4 F/"$ .#0%+$")/"#. +%)0/, 0#0%)D -&
"(# => '/$ %0-""#. 1#&%)# /+-$%0D*-+3 +4 5(# 1#(/2-%6)/, 7)%*#.6)# 6$#. &%) #87#)-0#+"
9C3 F/"$ '#)# 7,/*#. -+ "(# "#$" *(/01#) /+. )#*#-2#. -+&6$-%+$ %& /+-$%0D*-+3 &4 G#)*#+"&)##;-+< %+ "#$" E3 :-<6)# ,#<#+. -$ /77,-*/1,# "% 1%"( * /+. &3 H#)"-*/, %7#+A(#/.#./))%'$ )#7)#$#+" -+&6$-%+$3 !,, ./"/ 7%-+"$ )#7)#$#+" <)%67 0#/+$ # $3#303
© 2000 Macmillan Magazines Ltd
Monday, November 7, 2011

Sonuçlar
•Nader et al., 2000, Nature
!"##"$% #& '(#)$"
!"#$%& ' ()* +,- ' ./ "$0$1# 2,,, ' 333456789:4;<= *+,
!"#$"%&'()&"# "* *+(, -+-",&+$../ 0+,+ 1+ $2"1 )2() !"#$"%&'()+'*+(, -+-",&+$3 12+# ,+(!)&4()+' '5, ,+),&+4(%3 ,+)5,# )" (%(7&%+ $)()+ &# 12&!2 &#*5$&"# "* (#&$"-8!&# $2",)%8 (*)+, -+-",8,+(!)&4()&"# 9,"'5!+$ (-#+$&( "# %()+, )+$)$3 ,+6(,'%+$$ "*12+)2+, ,+(!)&4()&"# 1($ 9+,*",-+' . ", .: '(8$ (*)+, !"#'&)&"#;/ <2+ $(-+ ),+()-+#) 1&)2 (#&$"-8! &# )2+ (7$+#!+ "*-+-",8 ,+(!)&4()&"#3 %+*) -+-",8 &#)(!)/ ="#$&$)+#) 1&)2 ()&-+;%&-&)+' ,"%+ *", 9,")+&# $8#)2+$&$ 9,"'5!)&"# &# !"#$"%&'(;)&"#3 '+%(8 "* )2+ &#*5$&"# 5#)&% $&> 2"5,$ (*)+, -+-",8 ,+(!)&4(;)&"# 9,"'5!+' #" (-#+$&(/ ?5, '()( $2"1 )2() !"#$"%&'()+' *+(,-+-",&+$3 12+# ,+(!)&4()+'3 ,+)5,# )" ( %(7&%+ $)()+ )2() ,+@5&,+$'+ #"4" 9,")+&# $8#)2+$&$ *", ,+!"#$"%&'()&"#/ <2+$+ A#'$ (,+#") 9,+'&!)+' 78 ),('&)&"#(% )2+",&+$ "* -+-",8 !"#$"%&'()&"#/#>: ?@:6 7>67 5:3 =:=<9?:A B< 7>9<8B> 65 ?5?7?6C C6D?C: E:9?<@
D:F<9: D:?5B ;<5A<C?@67:@ ?57< A76DC: C<5BG7:9= =:=<9?:A ?A 65:579:5;>:@ E697 <F EAH;><C<B?;6C 65@ 5:89<D?<C<B?;6C =<@:CA <F=:=<9H.24 I<9 :J6=EC:K 7>:9: ?A ;<5A?@:96DC: :L?@:5;: 7>67 7>:F<9=67?<5 <F 6 C<5BG7:9= =:=<9H ;65 D: @?A98E7:@ DH ;:976?579:67=:57AK A8;> 6A AHA7:=?; @98B ?5M:;7?<5A <9 :C:;79<;<5L8CA?L:A><;NK B?L:5 A><97CH 6F7:9 796?5?5BK D87 7>67 7>: A6=: 79:67=:57AB?L:5 A:L:96C ><89A <9 @6HA C67:9 >6L: 5< :FF:;74 )5: <F 7>: =<A7;<==<5CH 8A:@ @98B =65?E8C67?<5A ?5L<CL:A 7>: 6@=?5?A7967?<5 <F@98BA 7>67 DC<;N 7>: 7965AC67?<5 <F %!" ?57< E9<7:?54 178@?:A <F7>?A 7HE: ?5@?;67: 7>67 =:=<9H ;<5A<C?@67?<5 ?5L<CL:A E9<7:?5AH57>:A?AOPQ4R7 >6A 6CA< D::5 9:E<97:@ 7>67 :C:;79<;<5L8CA?L: A><;N <9 AHA7:=?;
@98B 6@=?5?A7967?<5 B?L:5 6F7:9 =:=<9H 9:6;7?L67?<5 S9:79?:L6CT ;65;68A: 65 6=5:A?6 F<9 7>: <9?B?56C C:695?5B.UP.-K 3>?;> ?5@?;67:A 7>67;<5A<C?@67:@ =:=<9?:A =?B>7 D:;<=: C6D?C: 3>:5 9:79?:L:@K 65@=?B>7 :L:5 9:V8?9: 9:;<5A<C?@67?<54 W:9: 3: :J6=?5: 3>:7>:99:;<5A<C?@67?<5 ?5L<CL?5B E9<7:?5 AH57>:A?A ?A 9:V8?9:@ F<9 9:79?:L:@=:=<9?:A 7< E:9A?A74 X: 8A: 6 D:>6L?<896C E696@?B=K 68@?7<9H F:69;<5@?7?<5?5BK F<9 3>?;> 7>: 5:896C ;?9;8?7 85@:9CH?5B =:=<9HF<9=67?<5 ?A 3:CC ;>696;7:9?Y:@./P.Q4 #>?A 6CC<3A 8A 7< =65?E8C67:
=:=<9H 67 ?7A E9:A8=:@ C<;8A <F A7<96B:K ?5 ;<5796A7 7< E6A7 A78@?:A?5 3>?;> @98BA 3:9: 6@=?5?A7:9:@ AHA7:=?;6CCH4 1E:;?Z;6CCHK 3:769B:7 ?5F8A?<5A <F 65?A<=H;?5K 65 ?5>?D?7<9 <F E9<7:?5 AH57>:A?AK7< 7>: *["K 6 9:B?<5 ?=EC?;67:@ ?5 F:69 C:695?5B DH C:A?<5K E>69=6G;<C<B?;6C 65@ E>HA?<C<B?;6C Z5@?5BA./P.Q4 \9:L?<8ACHK 3: A><3:@ 7>67?5F8A?<5A <F 65?A<=H;?5 B?L:5 6F7:9 796?5?5B DC<;N C<5BG7:9= D875<7 A><97G7:9= =:=<9H <F 68@?7<9H F:69 ;<5@?7?<5?5B..4 W:9: 3::J6=?5: 7>: :FF:;7A <F A?=?C69 =65?E8C67?<5A 6@=?5?A7:9:@ 6F7:99:79?:L6C4%67A 3:9: B?L:5 6 A?5BC: E6?9?5B <F 6 7<5: S;<5@?7?<5:@ A7?=8C8AK
]1T 65@ F<<7GA><;N S85;<5@?7?<5:@ A7?=8C8AK $1T4 )5 7:A7 @6HAK?==<D?C?7H SF9::Y?5BT 36A 8A:@ 6A 65 ?5@:J <F F:69 C:695?5B2,4#3:57HGF<89 ><89A C67:9K 7>: 967A 9:;:?L:@ 6 A?5BC: ]1 E9:A:5767?<5S7:A7 .T ?==:@?67:CH F<CC<3:@ DH D?C67:96C ?5F8A?<5A <F 65?A<=H;?5<9 L:>?;C: S697?Z;?6C ;:9:D9<AE?56C ^8?@_ "]1IT ?57< 7>: *["4I9::Y?5B ?5 7:A7 . 36A AE:;?Z; 7< 7>: ]1 65@ ;<=E696DC: 6;9<AAB9<8EA4 "5 656CHA?A <F L69?65;: S"!)("T 7>67 ;<=E69:@ F9::Y?5B@89?5B 7>: E9:G]1 <9 ]1 E:9?<@A 6;9<AA B9<8EA ?5@?;67:@ 7>67 7>:9:36A 5< ?57:96;7?<5 D:73::5 7>:A: 73< L69?6DC:A S!!2! .`" # ."-TK 5<965 :FF:;7 <F B9<8E S!!2! .`" # ."+T4 W<3:L:9K 7>:9: 36A 6 A?B5?Z;657:FF:;7 <F E:9?<@ S!!.! .`" # .-,K "! ,",.T4 #3:57HGF<89 ><89A 6F7:97:A7 .K 7>: 967A 3:9: E9:A:57:@ 3?7> 7>9:: ]1A S7:A7 2T4 "5?A<=H;?5E9<@8;:@ 6 @<A:G@:E:5@:57 @:;9:6A: ?5 F9::Y?5B ?5 9:AE<5A: 7< 7>:]1 ?5 7:A7 2 SI?BA .K 26P;T4 "5 "!)(" 9:L:6C:@ 6 =6?5 :FF:;7 <FB9<8E S!!2! .`" # .2K "! ,",.T4 " !:3=65Pa:8CA E<A7 ><; 7:A79:L:6C:@ 7>67 7>: C<3G@<A: 65?A<=H;?5 65@ "]1I B9<8EA 3:9:A?=?C69 7< :6;> <7>:9 S"" ,",OTK D87 D<7> 3:9: A?B5?Z;657CH@?FF:9:57 F9<= 7>: >?B>G@<A: B9<8E S" L6C8:A! ,",.T4 &J7?5;7?<536A <DA:9L:@ <L:9 7>: 7>9:: ]1 E9:A:5767?<5A S=6?5 :FF:;7 <F 79?6CS!!2! U-" # /K "! ,",.TK D87 7>:9: 36A 5< ?57:96;7?<5 D:73::5 79?6CA65@ B9<8E S! ! .T4 #>?A :FF:;7 <F 65?A<=H;?5 9:V8?9:A 7>67 7>:=:=<9H D: 6;7?L:CH 9:79?:L:@K 6A <=?AA?<5 <F 7>: ]1 D:F<9: 65?A<G=H;?5 ?5F8A?<5 ?5 7:A7 . C:@ 7< 5<9=6C ;<5@?7?<5:@ F:69 9:AE<5A:A ?57:A7 2 SI?B4 2@K :_ 5< =6?5 :FF:;7 <F B9<8E S! ! .T4 #>: C677:9 Z5@?5B
!"#$%& ' ! "#$" %& '(#"(#) *%+$%,-./"#. &#/) 0#0%)-#$ */+ 1#*%0# ,/1-,# '(#+
)#/*"-2/"#.3 (4 5(# 1#(/2-%6)/, 7)%*#.6)# 6$#. &%) #87#)-0#+" 9!3 )4 :)##;-+< "% "(# =>
%+ "#$" 9 '/$ *%07/)/1,# /*)%$$ <)%67$ /+. '/$ $7#*-?* "% "(# =>3 *4 @+")/ABC!/+-$%0D*-+ -+&6$-%+$ /&"#) )#/*"-2/"-%+ %& / *%+$%,-./"#. &#/) 0#0%)D 7)%.6*# /0+#$-/
&%) "(# %)-<-+/, ,#/)+-+<4 /$ 0#/$6)#. %+ "#$" E3 +4 &4 F/"$ .#0%+$")/"#. +%)0/, 0#0%)D -&
"(# => '/$ %0-""#. 1#&%)# /+-$%0D*-+3 +4 5(# 1#(/2-%6)/, 7)%*#.6)# 6$#. &%) #87#)-0#+"
9C3 F/"$ '#)# 7,/*#. -+ "(# "#$" *(/01#) /+. )#*#-2#. -+&6$-%+$ %& /+-$%0D*-+3 &4 G#)*#+"&)##;-+< %+ "#$" E3 :-<6)# ,#<#+. -$ /77,-*/1,# "% 1%"( * /+. &3 H#)"-*/, %7#+A(#/.#./))%'$ )#7)#$#+" -+&6$-%+$3 !,, ./"/ 7%-+"$ )#7)#$#+" <)%67 0#/+$ # $3#303
© 2000 Macmillan Magazines Ltd
!"##"$% #& '(#)$"
*+, !"#$%& ' ()* +,- ' ./ "$0$1# 2,,, ' 333456789:4;<=
98>:? <87 7@: A<??BCB>B7D 7@67 7@: E:F;B7 3: <C?:9G:E B? E8: 7< 6EB?98A7B<5 <H >67: 36G:? <H A9<7:B5 ?D57@:?B?2. 7@67 =6D C: 5:;:??69DH<9 7@: ;<5?<>BE67B<5 <H 7@: <9BIB56> >:695B5IJ <9 7< 5<5?A:;BF;:HH:;7? ?8;@ 6? E6=6I: 7< 7@: 6=DIE6>6 CD 7@: E98I4 K7 B? 85>BL:>D7@67 7@: @BI@ E<?: <H 65B?<=D;B5 E6=6I:E 7@: 6=DIE6>6 6H7:9 M17965?=B??B<5J 6? 7@:9: 36? 5< @B?7<><IB;6> :GBE:5;: <H 6=DIE6>6E6=6I:4 N897@:9=<9:J 967? 9:>:695 H:69 ;<5EB7B<5B5I 5<9=6>>D 3@:57@: ?6=: E<?: <H 65B?<=D;B5 B? B5H8?:E B57< 7@: 6=DIE6>6 6H7:9B5B7B6> >:695B5I4 #@: H6;7 7@67 65B?<=D;B5 B5H8?B<5? 6H7:9 9:6;7BG67B<5<H 7@: H:69 =:=<9D A9<E8;:E 6=5:?B6 H<9 7@: <9BIB56> >:695B5IB5EB;67:? 7@67 9:6;7BG67B<5 <H 6 ;<5?<>BE67B<5 H:69 =:=<9D =6DA>6;: B7 B5 6 >6CB>: ?767:J <5: 7@67 @6? 7< C: 9:;<5?<>BE67:E CD A9<7:B5?D57@:?B? 7< 9:=6B5 8?6C>: 7< 7@: <9I65B?= B5 H8789: ?B7867B<5?4O9<7:B5 ?D57@:?B? B5@BCB7<9? 7DAB;6>>D B=A6B9 7@: ;<5?<>BE67B<5 <H
5:3 =:=<9B:? 3@:5 7@:D 69: 6E=B5B?7:9:E E89B5I 6 ?A:;BF; 7B=:3B5E<3 P3@B;@ G69B:? H9<= =B587:? 7< @<89?Q 6H7:9 >:695B5IR4"E=B5B?7967B<5 <H ?8;@ E98I? 6H7:9 7@B? 7B=: 3B5E<3 E<:? 5<76HH:;7 =:=<9D4 S: 5:T7 6?L:E 3@:7@:9 6 7B=: 3B5E<3 6>?< :TB?7? H<99:;<5?<>BE67B<5J CD E:>6DB5I 65B?<=D;B5 B5H8?B<5? H<9 - @ 6H7:99:79B:G6>4 N9::UB5I B5 7:?7 . 36? ?A:;BF; 7< 7@: M1 65E ;<=A696C>:6;9<?? I9<8A?4 "!)("? 9:G:6>:E 5< A:9B<E ! I9<8A B57:96;7B<5P! ! .Q 65E 5< :HH:;7 <H I9<8A P! ! .Q C87 6 ?BI5BF;657 :HH:;7 <HA:9B<E P!".! .V# ! .2+J "! ,",.Q4 K5 ;<5796?7 7< 65B?<=D;B5 B5H8W?B<5 B==:EB67:>D 6H7:9 9:79B:G6>J B5H8?B<5 - @ 6H7:9 9:79B:G6> @6E 5<:HH:;7 PNBI4 VQ4 "!)("? 9:G:6>:E 7@67 B5 7:?7 2 7@:9: 36? 5< =6B5:HH:;7 <H I9<8A P!".! .V# ! +QJ 5< 79B6> ! I9<8A B57:96;7B<5P!"2! 2-# ! 2"/Q 65E 5< =6B5 :HH:;7 <H 79B6> P!"2! 2-# ! VQ4 !<7:7@67 7@: 5<5?BI5BF;657 B=A6B9=:57 A9<E8;:E CD E:>6DB5I 65B?<=DW;B5 B? =8;@ ?=6>>:9 7@65 7@67 ?::5 3@:5 65B?<=D;B5 B? IBG:5B==:EB67:>D 6H7:9 M1 9:6;7BG67B<5 P;<=A69: 3B7@ NBI4 2;Q4 #@8?JC<7@ ;<5?<>BE67B<5 65E 9:;<5?<>BE67B<5 @6G: 7B=: 3B5E<3? 3B7@B53@B;@ A9<7:B5 ?D57@:?B? B? 9:X8B9:E BH 6 =:=<9D B? 7< A:9?B?74K5 6>> <H 7@: A9:GB<8? :TA:9B=:57?J 7@: 7B=: C:73::5 796B5B5I 65E
M1 A9:?:5767B<5 H<9 7:?7 . 36? 6C<87 2+ @4 S: 5:T7 B5G:?7BI67:E3@:7@:9 9:;<5?<>BE67B<5 @6? 6 7:=A<96> I96EB:574 N<9 :T6=A>:J<>E:9 =:=<9B:? =6D C: =<9: 7@<9<8I@>D ;<5?<>BE67:E 65E 7@8?=6D C: 9:?B?7657 7< C:;<=B5I 85?76C>: 3@:5 9:79B:G:E4 #< 7:?7 7@B?J3: 36B7:E H<9 .+ E6D? C:73::5 ;<5EB7B<5B5I 65E 7:?7 . PNBI4 +Q4
O<?7A<5B5I 7:?7 . CD .+ E6D? A9<E8;:E 65 B5;8C67B<5 :HH:;722 ?8;@7@67 H9::UB5I B5 7:?7 . 36? @BI@:9 7@65 B5 7@: A9:GB<8? :TA:9B=:574N9::UB5I E89B5I 7:?7 . 36? ?A:;BF; 7< 7@: M1 65E 36? ;<=A696C>:6;9<?? I9<8A?4 #@:9: 36? 5< A:9B<E ! I9<8A B57:96;7B<5P!".! Y# ! .".Q 65E 5< :HH:;7 <H I9<8A P! ! .QJ C87 6 ?BI5BF;657:HH:;7 <H A:9B<E P!".! Y# ! ++,J "! ,",.Q4 Z<3:G:9J 7@: A:9H<9=65;:<H 7@: I9<8A? B5 7:?7 2 EBG:9I:E[7@: I9<8A 9:;:BGB5I B5796W*\"B5H8?B<5? <H 65B?<=D;B5 ?@<3:E ?BI5BF;657>D >:?? H9::UB5I 7@65 7@:;<579<>?4 "!)("? 9:G:6>:E 6 =6B5 :HH:;7 <H I9<8A P!".! Y# ! 2,"-J"! ,",.Q 65E 79B6>? P!"2! .R# ! /J "! ,",.QJ C87 5< B57:96;7B<5C:73::5 7@:?: G69B6C>:? P! ! .Q4 #@8?J C><;L6E: <H A9<7:B5 ?D57@:?B?B5 7@: *\" 6H7:9 =:=<9D 9:6;7BG67B<5 ;68?:E 6=5:?B6 <H 7@: <9BIB56>>:695B5IJ :G:5 7@<8I@ 7@: >:695B5I 7<<L A>6;: .+ E6D? C:H<9: 7@:9:6;7BG67B<5 65E E98I 79:67=:574 &G:5 3:>> ;<5?<>BE67:E =:=<9B:?69: >6CB>: 65E ?8C]:;7 7< EB?98A7B<5 3@:5 9:6;7BG67:E4">7@<8I@ 65B?<=D;B5 C><;L? 9:;<5?<>BE67B<5J B7 B? A<??BC>: 7@67
7@: B=A6B9=:57 B? E8: 7< 5<5?A:;BF; :HH:;7? 7@67 9:5E:9 7@:6=DIE6>6 7:=A<969B>D ED?H85;7B<56> H<9 9:6?<5? <7@:9 7@65 A9<7:B5?D57@:?B? B5@BCB7B<54 #< A9<GBE: ;<=A:>>B5I :GBE:5;: 7@67 65D=65BA8>67B<5 6;7? ?A:;BF;6>>D <5 7@: =<>:;8>69 =:;@65B?=? =:EBW67B5I ;<5?<>BE67B<5J 6? <AA<?:E 7< A9<E8;B5I 5<5?A:;BF; :HH:;7?J 676 =B5B=8= B7 B? 5:;:??69D 7< ?@<3 7@67 =:=<9D B? B576;7 ?@<97>D6H7:9 796B5B5I C87 B=A6B9:E >67:92V^2-4 N<9 :T6=A>:J 967? H9::U:5<9=6>>D + @ C87 69: B=A6B9:E 2+ @ 6H7:9 9:;:BGB5I B5796W*\"65B?<=D;B5 B5H8?B<5? B==:EB67:>D 6H7:9 796B5B5I..4 $?B5I 7@: ?6=:><IB; 65E 6AA>DB5I B7 7< 9:;<5?<>BE67B<5J BH 7@: :HH:;7? <H 65B?<=D;B5B5H8?:E B57< 7@: *\" ?@<97>D 6H7:9 =:=<9D 9:6;7BG67B<5 69: ?A:;BF;7< 9:;<5?<>BE67B<5J 7@:5 H9::UB5I ?@<8>E C: 5<9=6> 67 + @J C87B=A6B9:E 67 2+ @J 6H7:9 M1 A9:?:5767B<54 S: 9:H:9 7< 7@:?: 73<7B=: A<B57? 69: A<?7W9:6;7BG67B<5 ?@<97W7:9= =:=<9D PO%W1#_Q65E A<?7W9:6;7BG67B<5 ><5IW7:9= =:=<9D PO%W*#_QJ 9:?A:;7BG:>D4#@: 73< I9<8A? :T@BCB7:E ;<=A696C>: H9::UB5I E89B5I 7:?7 .
!"#$%& ' !"#$%# &'&()* +, $"+-(&*%+" +",.-+("- $)' /'0$*'/ 1* 2 34 (5 63' 1'3$7+(.)$0
8)(%'/.)' .-'/ ,() '98')+&'"# :4 ;')#+%$0 (8'"<3'$/'/ $))(=- )'8)'-'"# +",.-+("-4
)5 >)''?+"@ (" #'-# A =$- -8'%+B% #( #3' CD $"/ %(&8$)$10' $%)(-- @)(.8-4 *5 E')%'"#
,)''?+"@ /.)+"@ #'-# :4 63' @)(.8- $)' "(# -+@"+B%$"#0* /+,,')'"#4 F00 /$#$ 8(+"#- )'8)'-'"#
@)(.8 &'$"- " -4'4&4
!"#$%& + >(.)#''" /$*- $,#') #)$+"+"@5 $"+-(&*%+" +",.-+("- $,#') )'$%#+7$#+(" (, #3'
&'&()* -#+00 8)(/.%' $&"'-+$4 (5 63' 1'3$7+(.)$0 8)(%'/.)' .-'/ ,() '98')+&'"# G4
;')#+%$0 (8'"<3'$/'/ $))(=- )'8)'-'"# +",.-+("-4 )5 >)''?+"@ /.)+"@ #'-# A =$- -8'%+B% #(
#3' CD $"/ =$- %(&8$)$10' $%)(-- @)(.8-4 *5 E')%'"# ,)''?+"@ (" #'-# :4 F00 /$#$ 8(+"#-
)'8)'-'"# @)(.8 &'$"- " -4'4&4
© 2000 Macmillan Magazines Ltd
Monday, November 7, 2011

KSB’de sorun yok!
!"##"$% #& '(#)$"
!"#$%& ' ()* +,- ' ./ "$0$1# 2,,, ' 333456789:4;<= *+,
>?@A4 BC4 "5 "!)(" 9:D:6E:F 6 G@A5@H;657 :II:;7 <I J:9@<F>!!.! .," # .+/K "! ,",.CK 5< :II:;7 <I A9<8J >! ! .C 65F 5<@57:96;7@<5 L:73::5 7M:G: 73< D69@6LE:G >! ! .C4 %67G 79:67:F 3@7M65@G<=N;@5 @==:F@67:EN 6I7:9 7:G7 . :OM@L@7:F @576;7 P%Q1#R L87F:H;@:57 P%Q*#R4 "!)("G <5 7M: G;<9:G <I 7M: P%Q1#R 7:G7GM<3:F 7M67 7M:9: 36G 5< =6@5 :II:;7 <I A9<8J >! ! .CK 5< @57:9Q6;7@<5 L:73::5 79@6E ! A9<8J >! ! .C 65F 5< :II:;7 <I 79@6E >! ! .C4S<5D:9G:ENK 6 G@=@E69 656ENG@G <5 7M: P%Q*#R G;<9:G F@F 9:D:6E 6G@A5@H;657 :II:;7 <I A9<8J >!!.! .+" # .+K "! ,",.C4 ?897M:9=<9:K7M:9: 36G 5< @57:96;7@<5 L:73::5 A9<8J 65F 79@6E >! ! .C L87 7M:9:36G 6 =6@5 :II:;7 <I 79@6E >!!2! 2T" # BK "! ,",BC4 #M: I6;7 7M6765@=6EG ;65 6;;8967:EN J:9;:@D:K :D6E867: 65F 9:GJ<5F 7< 7M: S1 + M6I7:9 65@G<=N;@5 @5I8G@<5 GM<3G 7M67 7M: 6=NAF6E6 @G I85;7@<56EEN@576;7 67 7M: 7@=: <I 7M: P%Q1#R 7:G7K 65F 7M8G 7M67 65@G<=N;@5 F@F5<7 6II:;7 9:;<5G<E@F67@<5 LN J9<F8;@5A 5<5GJ:;@H; :II:;7G4 #M@GJ677:95 <I H5F@5AGK @576;7 P%Q1#R 65F @=J6@9:F P%Q*#RK E<;6E@U:G7M: :II:;7G <I 65@G<=N;@5 <5 I:69 L:M6D@<89 7< 7M: =<E:;8E69J9<;:GG:G =:F@67@5A 9:;<5G<E@F67@<54#M: ;<5D:57@<56E D@:3 <I =:=<9N ;<5G<E@F67@<5 J9:F@;7G 7M67
LE<;V6F: <I J9<7:@5 GN57M:G@G GM<8EF LE<;V 5:3 E:695@5A @5 7:G7 . <I7M:G: :OJ:9@=:57GK 3M@;M @G 65 :O7@5;7@<5 7:G74 #M: <5EN 5:3E:695@5A 7M67 <;;89G @G 6L<87 7M: I6@E89: <I 7M: S1 7< J9:F@;7 7M:$14 WE<;V6F: <I J9<7:@5 GN57M:G@G GM<8EF 7M:9:I<9: LE<;V :O7@5;7@<5K65F 7M8G :5M65;: =:=<9NK 6;;<9F@5A 7< 7M: ;<5D:57@<56E D@:34 X5;<5796G7K M<3:D:9K LE<;V6F: <I J9<7:@5 GN57M:G@G M6F 7M: <JJ<G@7::II:;7Y@7 :E@=@567:F 7M: =:=<9N 967M:9 7M65 =6V@5A @7 G79<5A:94#M:G: 9:G8E7G J9<D@F: :D@F:5;: 7M67 I:69 =:=<9@:GK <5;:
9:79@:D:FK =8G7 85F:9A< J9<7:@5QGN57M:G@GQF:J:5F:57 9:;<5G<E@F6Q7@<5 @5 7M: *W" <9 5:69LN 69:6G 7< 9:=6@5 6;;:GG@LE: 67 E67:9 7@=:G4#M: I8EE @=JE@;67@<5G <I 7M@G H5F@5A I<9 I:69 65F <7M:9 =:=<9@:G 69:5<7 85F:9G7<<F 67 J9:G:574 X7 @G J<GG@LE: 7M67 5<7 6EE =:=<9@:G9:Z8@9: 9:;<5G<E@F67@<54 #M:9: =6N L: 6 965A: <I J696=:7:9G 3@7M@53M@;M 9:6;7@D67@<5 <I 6 =:=<9N ;<5D:97G @7 @57< 6 E6L@E: G767:KJ<GG@LEN @5D<ED@5A 7M: :O7:57 <I :OJ:9@:5;: 3@7M 7M: J697@;8E69
E:695@5A G@7867@<5K 7M: V@5F <I E:695@5A GNG7:= :5A6A:F 65F 7M:=<7@D67@<56E G767: <I 7M: G8L[:;7 67 7M: 7@=: <I E:695@5A 65F 9:79@:D6E4%:;<5G<E@F67@<5 =6N 9:\:;7 7M: FN56=@; 56789: <I 7M: J9<;:GG LN
3M@;M 5:3 @5I<9=67@<5 @G 6FF:F 7< :O@G7@5A G7<9:G4 X7 M6G E<5A L::5L:E@:D:F 7M67 =:=<9N 9:79@:D6E @G 65 6;7@D: <9 ;<5G798;7@D: J9<;:GGLN 3M@;M <EF @5I<9=67@<5 @G @57:A967:F 3@7M 7M: ;899:57 V5<3E:FA:L6G: <I 7M: <9A65@G=2/4 %:;<5G<E@F67@<5 =6N L: J697 <I 7M: 5:896E=:;M65@G= 7M9<8AM 3M@;M ;<5G798;7:F =:=<9@:G 69: G7<9:F I<9E67:9 ;<5G798;7@<5G4S899:57 =<F:EG <I E:695@5A J9<J<G: 7M67 7M: J9<F8;7@<5 <I 5:3
J9<7:@5G @G 5:;:GG69N I<9 G798;7896E :5;<F@5A <I 9:;:57 :OJ:9@:5;:G @5E<5AQ7:9==:=<9N-]^4 X5 6FF@7@<5K @7 5<3 6JJ:69G 7M67 5:3 J9<7:@5G69: 6EG< 9:Z8@9:F 7< =6@576@5 =:=<9@:G 7M67 M6D: L::5 9:6;7@D67:F4X7 G::=G 85E@V:EN 7M67 9:79@:D6E 9:D:9G:G 7M: G798;7896E ;M65A:G@5F8;:F LN <9@A@56E E:695@5A4 %67M:9K G<=: J9<J:97N <I 9:79@:D6E=6N F:G76L@E@U: 7M: G798;7896E ;M65A:G G8;M 7M67 7M:N 5<3 M6D: 7< L:9:;<5G<E@F67:F 3@7M 7M: 6@F <I 5:3 J9<7:@5G4 #M: I6;7 7M67 65@=6EGF:=<5G7967: @576;7 I9::U@5A + M 6I7:9 65@G<=N;@5 @5F@;67:G 7M67 7M:G798;7896E ;M65A:G =6N 9:=6@5 I85;7@<56E I<9 67 E:6G7 + M4 P697@;8E69EN@=J<97657 I<9 I8789: 3<9V3@EE L: 7M: ;E69@H;67@<5 <I 7M: JMNG@<E<A@;6EL6G@G <I =:=<9N E6L@E@7N F89@5A 9:79@:D6E 65F 7M: 9:Z8@9:=:57 I<99:;<5G<E@F67@<54 X7 @G J<GG@LE: 7M67 G<=: =<F@H;67@<5 <I 7M: GN56JQ7@; 76AA@5A MNJ<7M:G@G2TK2^K 3M@;M J9<J<G:G 7M67 6;7@D: GN56JG:G 69:A@D:5 =<E:;8E69 =69V:9G 7M67 M:EJ G76L@E@U: GN56JG:G LN ;6J789@5AJ9<7:@5G =6F: @5 7M: ;:EE 58;E:8GK =@AM7 6;;<857 I<9 E6L@E@7N 65F9:;<5G<E@F67@<5K 6E7M<8AM 7M@G 9:=6@5G 7< L: G::54_: M6D: GM<35 7M67 9:6;7@D67@<5 ;M65A:G 7M: G7678G <I 6 ;<5Q
G<E@F67:F I:69 =:=<9N 7< 6 E6L@E: <5: 7M67 =8G7 L: 9:;<5G<E@F67:F8G@5A F: 5<D< J9<7:@5 GN57M:G@G 7< J:9G@G74 *@V: ;<5G<E@F67@<5 @7G:EIK9:;<5G<E@F67@<5 M6G 6 7:=J<96E 3@5F<3 F89@5A 3M@;M LE<;V6F: <IJ9<7:@5 GN57M:G@G @G :II:;7@D:K 65F L:N<5F 3M@;M @7 @G 5<74 ?897M:9Q=<9: 7M: 9:;<5G<E@F67@<5 J9<;:GGK E@V: 7M: ;<5G<E@F67@<5 J9<;:GGKM6G 6 GM<97Q7:9= JM6G: 7M67 @G 5<7 F:J:5F:57 <5 J9<7:@5 GN57M:G@G4" F:H5@7@<5 <I ;<5G<E@F67@<5 L6G:F <5 `5:3a =:=<9@:G @G @5G8IHQ;@:57EN L9<6F 7< F:G;9@L: 7M:G: F6764 _: J9<J<G:K @5 V::J@5A 3@7M7M: <9@A@56E G8AA:G7@<5 LN R@G65@5 #$ %&'.bK 7M67 6G 6 H9G7 6JJ9<O@=6Q7@<5 `6;7@D:a 967M:9 7M65 `5:3a =:=<9@:G L: D@:3:F 6G E6L@E:KG8L[:;7 7< F@G98J7@<5K 65F 9:Z8@9@5A J9<7:@5QGN57M:G@GQF:J:5F:57;<5G<E@F67@<5 J9<;:GG:G4 !
!"#$%&'()*+",#'18L[:;7G 3:9: 6F8E7 =6E: 1J96A8:]c63E:N 967G I9<= d@EE7<J *6LG4 %67G 3:9: M<8G:F@5F@D@F86EEN @5 JE6G7@; !6EA:5: ;6A:G 65F =6@576@5:F <5 6 .2e.2 M E@AM7eF69V ;N;E:4 ?<<F65F 367:9 3:9: J9<D@F:F %( &)*)$+,4
()-."-/ 01& $2'#%3%./$5F:9 !:=L876E 656:G7M:G@6 >+B=A VA".CK 967G 3:9: @=JE657:F L@E67:96EEN 3@7M 22QA68A:G76@5E:GG G7::E ;6558E6G @57< 7M: E67:96E 6=NAF6764 S<<9F@567:G 3:9: b4,== J<G7:9@<9 7<L9:A=6K B4b== E67:96E 7< 7M: =@FE@5: 65F T4,== D:5796E 7< 7M: GV8EE G89I6;:4 %67G 3:9:A@D:5 67 E:6G7 B F6NG 7< 9:;<D:9 L:I<9: :OJ:9@=:576E J9<;:F89:G4 "EE J9<;:F89:G 3:9: @56;;<9F65;: 3@7M 7M: !Xd 08@F:K 65F 3:9: 6JJ9<D:F LN 7M: !f$ "5@=6E S69: 65F $G:S<==@77::4 "7 7M: :5F <I 7M: :OJ:9@=:57K 8G@5A G765F69F M@G7<E<A@;6E =:7M<FGK 65@=6EG3:9: J:9I8G:F 65F 7M:@9 L96@5G G:;7@<5:F 67 B,"=7M@;V5:GG4 #M: G:;7@<5G3:9: G76@5:F 8G@5A;9:GNE D@<E:7 65F :O6=@5:F LN E@AM7 =@;9<G;<JN I<9 ;6558E6 J:5:7967@<5 @57< 7M: *W"4
41#-05678 219)'2%1'c98AG 3:9: @5I8G:F GE<3EN 8G@5A 65 @5I8G@<5 J8=J @57< 7M: *W" 67 ,42B"E =@5".4?<EE<3@5A F98A @5I8G@<5K ;6558E6G 3:9: E:I7 @5 JE6;: I<9 65 6FF@7@<56E =@587: 7< 6EE<3F@II8G@<5 <I 7M: F98A 636N I9<= 7M: ;6558E6 7@J4 "5@G<=N;@5 >1@A=6C 36G F@GG<ED:F @5:Z8@=<E69 dSEK F@E87:F 3@7M "S1? 65F 6F[8G7:F 7< Jd /4+ 3@7M !6)d4 "E7M<8AM 7M:E67:96E 58;E:8G 36G 7M:=6@5 769A:7K 7M: ,4BQ"E @5I8G@<5G 6EG< J9<L6LEN 6II:;7:F 7M: 6F[6;:57L6G6E 58;E:8G4 _: 7M:9:I<9: 9:I:9 7< 7M: 6II:;7:F 69:6 6G 7M: E67:96E 65F L6G6E 6=NAF6E6>*W"C4
8::0-0#)'S<5F@7@<5@5A 65F 7<5: 7:G7@5A 3:9: ;<5F8;7:F @5 F@II:9:57 ;M6=L:9G4 ?<9 ;<5F@7@<5@5AK967G 3:9: JE6;:F @5 6 PE:O@AE6G 9<F:57 ;<5F@7@<5@5A ;M6=L:9 >;M6=L:9 "C 3@7M 6=:76E A9@F\<<9 >R<F:E &.,Q.,K S<8EL<895 X5G798=:57GC 7M67 36G :5;E<G:F 3@7M@5 6 G<85F677:5867@5A ;M6=L:9 >R<F:E &.,Q2,C4 #M: ;M6=L:9 36G F@=EN @EE8=@567:F LN 6 G@5AE:M<8G: E@AM74 ?<9 7<5: 7:G7@5AK 967G 3:9: JE6;:F @5 6 F@II:9:57 PE:O@AE6G ;M6=L:9 >&!(Q,,.K
;2.)-" < !"#$%&' ()**)+&#, '#&%)"-.&# &% #)/ 01$ /) #)#%2$.&3. $(($./%4 05 67$
8$7'9&)1:'* 2:).$01:$ 1%$0 (): $;2$:&"$#/ <4 =$:/&.'* )2$#>7$'0$0 '::)+% :$2:$%$#/
&#(1%&)#%4 !#&%)"-.&# &#(1%&)#% &"2'&:$0 2)%/>:$'./&9'/&)# *)#,>/$:" "$"):- ?@A>B6CD5
81/ 7'0 #) $(($./ )# 2)%/>:$'./&9'/&)# %7):/>/$:" "$"):- ?@A>E6CD4 *5 !** :'/%
0$")#%/:'/$0 .)"2':'8*$ (:$$F&#, %.):$% )# /$%/ G4 ,5 E.):$% )# /7$ @A>E6C /$%/ < 7
'(/$: :$'./&9'/&)# '#0 '#&%)"-. &5 E.):$% )# /7$ @A>B6C /$%/ H< 7 '(/$: :$'./&9'/&)#
'#0 '#&%)"-. 67$ I$- &% '22*&.'8*$ /) 8)/7 , '#0 &4 !** 0'/' 2)&#/% :$2:$%$#/ ,:)12
"$'#% # %4$4"4
© 2000 Macmillan Magazines Ltd
!"##"$% #& '(#)$"
!"#$%& ' ()* +,- ' ./ "$0$1# 2,,, ' 333456789:4;<= *+,
>?@A4 BC4 "5 "!)(" 9:D:6E:F 6 G@A5@H;657 :II:;7 <I J:9@<F>!!.! .," # .+/K "! ,",.CK 5< :II:;7 <I A9<8J >! ! .C 65F 5<@57:96;7@<5 L:73::5 7M:G: 73< D69@6LE:G >! ! .C4 %67G 79:67:F 3@7M65@G<=N;@5 @==:F@67:EN 6I7:9 7:G7 . :OM@L@7:F @576;7 P%Q1#R L87F:H;@:57 P%Q*#R4 "!)("G <5 7M: G;<9:G <I 7M: P%Q1#R 7:G7GM<3:F 7M67 7M:9: 36G 5< =6@5 :II:;7 <I A9<8J >! ! .CK 5< @57:9Q6;7@<5 L:73::5 79@6E ! A9<8J >! ! .C 65F 5< :II:;7 <I 79@6E >! ! .C4S<5D:9G:ENK 6 G@=@E69 656ENG@G <5 7M: P%Q*#R G;<9:G F@F 9:D:6E 6G@A5@H;657 :II:;7 <I A9<8J >!!.! .+" # .+K "! ,",.C4 ?897M:9=<9:K7M:9: 36G 5< @57:96;7@<5 L:73::5 A9<8J 65F 79@6E >! ! .C L87 7M:9:36G 6 =6@5 :II:;7 <I 79@6E >!!2! 2T" # BK "! ,",BC4 #M: I6;7 7M6765@=6EG ;65 6;;8967:EN J:9;:@D:K :D6E867: 65F 9:GJ<5F 7< 7M: S1 + M6I7:9 65@G<=N;@5 @5I8G@<5 GM<3G 7M67 7M: 6=NAF6E6 @G I85;7@<56EEN@576;7 67 7M: 7@=: <I 7M: P%Q1#R 7:G7K 65F 7M8G 7M67 65@G<=N;@5 F@F5<7 6II:;7 9:;<5G<E@F67@<5 LN J9<F8;@5A 5<5GJ:;@H; :II:;7G4 #M@GJ677:95 <I H5F@5AGK @576;7 P%Q1#R 65F @=J6@9:F P%Q*#RK E<;6E@U:G7M: :II:;7G <I 65@G<=N;@5 <5 I:69 L:M6D@<89 7< 7M: =<E:;8E69J9<;:GG:G =:F@67@5A 9:;<5G<E@F67@<54#M: ;<5D:57@<56E D@:3 <I =:=<9N ;<5G<E@F67@<5 J9:F@;7G 7M67
LE<;V6F: <I J9<7:@5 GN57M:G@G GM<8EF LE<;V 5:3 E:695@5A @5 7:G7 . <I7M:G: :OJ:9@=:57GK 3M@;M @G 65 :O7@5;7@<5 7:G74 #M: <5EN 5:3E:695@5A 7M67 <;;89G @G 6L<87 7M: I6@E89: <I 7M: S1 7< J9:F@;7 7M:$14 WE<;V6F: <I J9<7:@5 GN57M:G@G GM<8EF 7M:9:I<9: LE<;V :O7@5;7@<5K65F 7M8G :5M65;: =:=<9NK 6;;<9F@5A 7< 7M: ;<5D:57@<56E D@:34 X5;<5796G7K M<3:D:9K LE<;V6F: <I J9<7:@5 GN57M:G@G M6F 7M: <JJ<G@7::II:;7Y@7 :E@=@567:F 7M: =:=<9N 967M:9 7M65 =6V@5A @7 G79<5A:94#M:G: 9:G8E7G J9<D@F: :D@F:5;: 7M67 I:69 =:=<9@:GK <5;:
9:79@:D:FK =8G7 85F:9A< J9<7:@5QGN57M:G@GQF:J:5F:57 9:;<5G<E@F6Q7@<5 @5 7M: *W" <9 5:69LN 69:6G 7< 9:=6@5 6;;:GG@LE: 67 E67:9 7@=:G4#M: I8EE @=JE@;67@<5G <I 7M@G H5F@5A I<9 I:69 65F <7M:9 =:=<9@:G 69:5<7 85F:9G7<<F 67 J9:G:574 X7 @G J<GG@LE: 7M67 5<7 6EE =:=<9@:G9:Z8@9: 9:;<5G<E@F67@<54 #M:9: =6N L: 6 965A: <I J696=:7:9G 3@7M@53M@;M 9:6;7@D67@<5 <I 6 =:=<9N ;<5D:97G @7 @57< 6 E6L@E: G767:KJ<GG@LEN @5D<ED@5A 7M: :O7:57 <I :OJ:9@:5;: 3@7M 7M: J697@;8E69
E:695@5A G@7867@<5K 7M: V@5F <I E:695@5A GNG7:= :5A6A:F 65F 7M:=<7@D67@<56E G767: <I 7M: G8L[:;7 67 7M: 7@=: <I E:695@5A 65F 9:79@:D6E4%:;<5G<E@F67@<5 =6N 9:\:;7 7M: FN56=@; 56789: <I 7M: J9<;:GG LN
3M@;M 5:3 @5I<9=67@<5 @G 6FF:F 7< :O@G7@5A G7<9:G4 X7 M6G E<5A L::5L:E@:D:F 7M67 =:=<9N 9:79@:D6E @G 65 6;7@D: <9 ;<5G798;7@D: J9<;:GGLN 3M@;M <EF @5I<9=67@<5 @G @57:A967:F 3@7M 7M: ;899:57 V5<3E:FA:L6G: <I 7M: <9A65@G=2/4 %:;<5G<E@F67@<5 =6N L: J697 <I 7M: 5:896E=:;M65@G= 7M9<8AM 3M@;M ;<5G798;7:F =:=<9@:G 69: G7<9:F I<9E67:9 ;<5G798;7@<5G4S899:57 =<F:EG <I E:695@5A J9<J<G: 7M67 7M: J9<F8;7@<5 <I 5:3
J9<7:@5G @G 5:;:GG69N I<9 G798;7896E :5;<F@5A <I 9:;:57 :OJ:9@:5;:G @5E<5AQ7:9==:=<9N-]^4 X5 6FF@7@<5K @7 5<3 6JJ:69G 7M67 5:3 J9<7:@5G69: 6EG< 9:Z8@9:F 7< =6@576@5 =:=<9@:G 7M67 M6D: L::5 9:6;7@D67:F4X7 G::=G 85E@V:EN 7M67 9:79@:D6E 9:D:9G:G 7M: G798;7896E ;M65A:G@5F8;:F LN <9@A@56E E:695@5A4 %67M:9K G<=: J9<J:97N <I 9:79@:D6E=6N F:G76L@E@U: 7M: G798;7896E ;M65A:G G8;M 7M67 7M:N 5<3 M6D: 7< L:9:;<5G<E@F67:F 3@7M 7M: 6@F <I 5:3 J9<7:@5G4 #M: I6;7 7M67 65@=6EGF:=<5G7967: @576;7 I9::U@5A + M 6I7:9 65@G<=N;@5 @5F@;67:G 7M67 7M:G798;7896E ;M65A:G =6N 9:=6@5 I85;7@<56E I<9 67 E:6G7 + M4 P697@;8E69EN@=J<97657 I<9 I8789: 3<9V3@EE L: 7M: ;E69@H;67@<5 <I 7M: JMNG@<E<A@;6EL6G@G <I =:=<9N E6L@E@7N F89@5A 9:79@:D6E 65F 7M: 9:Z8@9:=:57 I<99:;<5G<E@F67@<54 X7 @G J<GG@LE: 7M67 G<=: =<F@H;67@<5 <I 7M: GN56JQ7@; 76AA@5A MNJ<7M:G@G2TK2^K 3M@;M J9<J<G:G 7M67 6;7@D: GN56JG:G 69:A@D:5 =<E:;8E69 =69V:9G 7M67 M:EJ G76L@E@U: GN56JG:G LN ;6J789@5AJ9<7:@5G =6F: @5 7M: ;:EE 58;E:8GK =@AM7 6;;<857 I<9 E6L@E@7N 65F9:;<5G<E@F67@<5K 6E7M<8AM 7M@G 9:=6@5G 7< L: G::54_: M6D: GM<35 7M67 9:6;7@D67@<5 ;M65A:G 7M: G7678G <I 6 ;<5Q
G<E@F67:F I:69 =:=<9N 7< 6 E6L@E: <5: 7M67 =8G7 L: 9:;<5G<E@F67:F8G@5A F: 5<D< J9<7:@5 GN57M:G@G 7< J:9G@G74 *@V: ;<5G<E@F67@<5 @7G:EIK9:;<5G<E@F67@<5 M6G 6 7:=J<96E 3@5F<3 F89@5A 3M@;M LE<;V6F: <IJ9<7:@5 GN57M:G@G @G :II:;7@D:K 65F L:N<5F 3M@;M @7 @G 5<74 ?897M:9Q=<9: 7M: 9:;<5G<E@F67@<5 J9<;:GGK E@V: 7M: ;<5G<E@F67@<5 J9<;:GGKM6G 6 GM<97Q7:9= JM6G: 7M67 @G 5<7 F:J:5F:57 <5 J9<7:@5 GN57M:G@G4" F:H5@7@<5 <I ;<5G<E@F67@<5 L6G:F <5 `5:3a =:=<9@:G @G @5G8IHQ;@:57EN L9<6F 7< F:G;9@L: 7M:G: F6764 _: J9<J<G:K @5 V::J@5A 3@7M7M: <9@A@56E G8AA:G7@<5 LN R@G65@5 #$ %&'.bK 7M67 6G 6 H9G7 6JJ9<O@=6Q7@<5 `6;7@D:a 967M:9 7M65 `5:3a =:=<9@:G L: D@:3:F 6G E6L@E:KG8L[:;7 7< F@G98J7@<5K 65F 9:Z8@9@5A J9<7:@5QGN57M:G@GQF:J:5F:57;<5G<E@F67@<5 J9<;:GG:G4 !
!"#$%&'()*+",#'18L[:;7G 3:9: 6F8E7 =6E: 1J96A8:]c63E:N 967G I9<= d@EE7<J *6LG4 %67G 3:9: M<8G:F@5F@D@F86EEN @5 JE6G7@; !6EA:5: ;6A:G 65F =6@576@5:F <5 6 .2e.2 M E@AM7eF69V ;N;E:4 ?<<F65F 367:9 3:9: J9<D@F:F %( &)*)$+,4
()-."-/ 01& $2'#%3%./$5F:9 !:=L876E 656:G7M:G@6 >+B=A VA".CK 967G 3:9: @=JE657:F L@E67:96EEN 3@7M 22QA68A:G76@5E:GG G7::E ;6558E6G @57< 7M: E67:96E 6=NAF6764 S<<9F@567:G 3:9: b4,== J<G7:9@<9 7<L9:A=6K B4b== E67:96E 7< 7M: =@FE@5: 65F T4,== D:5796E 7< 7M: GV8EE G89I6;:4 %67G 3:9:A@D:5 67 E:6G7 B F6NG 7< 9:;<D:9 L:I<9: :OJ:9@=:576E J9<;:F89:G4 "EE J9<;:F89:G 3:9: @56;;<9F65;: 3@7M 7M: !Xd 08@F:K 65F 3:9: 6JJ9<D:F LN 7M: !f$ "5@=6E S69: 65F $G:S<==@77::4 "7 7M: :5F <I 7M: :OJ:9@=:57K 8G@5A G765F69F M@G7<E<A@;6E =:7M<FGK 65@=6EG3:9: J:9I8G:F 65F 7M:@9 L96@5G G:;7@<5:F 67 B,"=7M@;V5:GG4 #M: G:;7@<5G3:9: G76@5:F 8G@5A;9:GNE D@<E:7 65F :O6=@5:F LN E@AM7 =@;9<G;<JN I<9 ;6558E6 J:5:7967@<5 @57< 7M: *W"4
41#-05678 219)'2%1'c98AG 3:9: @5I8G:F GE<3EN 8G@5A 65 @5I8G@<5 J8=J @57< 7M: *W" 67 ,42B"E =@5".4?<EE<3@5A F98A @5I8G@<5K ;6558E6G 3:9: E:I7 @5 JE6;: I<9 65 6FF@7@<56E =@587: 7< 6EE<3F@II8G@<5 <I 7M: F98A 636N I9<= 7M: ;6558E6 7@J4 "5@G<=N;@5 >1@A=6C 36G F@GG<ED:F @5:Z8@=<E69 dSEK F@E87:F 3@7M "S1? 65F 6F[8G7:F 7< Jd /4+ 3@7M !6)d4 "E7M<8AM 7M:E67:96E 58;E:8G 36G 7M:=6@5 769A:7K 7M: ,4BQ"E @5I8G@<5G 6EG< J9<L6LEN 6II:;7:F 7M: 6F[6;:57L6G6E 58;E:8G4 _: 7M:9:I<9: 9:I:9 7< 7M: 6II:;7:F 69:6 6G 7M: E67:96E 65F L6G6E 6=NAF6E6>*W"C4
8::0-0#)'S<5F@7@<5@5A 65F 7<5: 7:G7@5A 3:9: ;<5F8;7:F @5 F@II:9:57 ;M6=L:9G4 ?<9 ;<5F@7@<5@5AK967G 3:9: JE6;:F @5 6 PE:O@AE6G 9<F:57 ;<5F@7@<5@5A ;M6=L:9 >;M6=L:9 "C 3@7M 6=:76E A9@F\<<9 >R<F:E &.,Q.,K S<8EL<895 X5G798=:57GC 7M67 36G :5;E<G:F 3@7M@5 6 G<85F677:5867@5A ;M6=L:9 >R<F:E &.,Q2,C4 #M: ;M6=L:9 36G F@=EN @EE8=@567:F LN 6 G@5AE:M<8G: E@AM74 ?<9 7<5: 7:G7@5AK 967G 3:9: JE6;:F @5 6 F@II:9:57 PE:O@AE6G ;M6=L:9 >&!(Q,,.K
;2.)-" < !"#$%&' ()**)+&#, '#&%)"-.&# &% #)/ 01$ /) #)#%2$.&3. $(($./%4 05 67$
8$7'9&)1:'* 2:).$01:$ 1%$0 (): $;2$:&"$#/ <4 =$:/&.'* )2$#>7$'0$0 '::)+% :$2:$%$#/
&#(1%&)#%4 !#&%)"-.&# &#(1%&)#% &"2'&:$0 2)%/>:$'./&9'/&)# *)#,>/$:" "$"):- ?@A>B6CD5
81/ 7'0 #) $(($./ )# 2)%/>:$'./&9'/&)# %7):/>/$:" "$"):- ?@A>E6CD4 *5 !** :'/%
0$")#%/:'/$0 .)"2':'8*$ (:$$F&#, %.):$% )# /$%/ G4 ,5 E.):$% )# /7$ @A>E6C /$%/ < 7
'(/$: :$'./&9'/&)# '#0 '#&%)"-. &5 E.):$% )# /7$ @A>B6C /$%/ H< 7 '(/$: :$'./&9'/&)#
'#0 '#&%)"-. 67$ I$- &% '22*&.'8*$ /) 8)/7 , '#0 &4 !** 0'/' 2)&#/% :$2:$%$#/ ,:)12
"$'#% # %4$4"4
© 2000 Macmillan Magazines Ltd
Monday, November 7, 2011

Gereklilikler
•Hatıranın sağlamlaşmış olduğundan emin ol•Hatırlama olmazsa amnezik etkinin görülmediğinden emin ol
•Kısa süreli bellek (ve tabi diğer hafıza süreçleri)
Memory traces unboundKarim Nader
Department of Psychology, McGill University, Montreal, Quebec, Canada H3A 1B1
The idea that new memories are initially ‘labile’ andsensitive to disruption before becoming permanentlystored in the wiring of the brain has been dogma for>100 years. Recently, we have revisited the hypothesisthat reactivation of a consolidated memory can return itto a labile, sensitive state – in which it can be modified,strengthened, changed or even erased! The data gener-ated from some of the best-described paradigms inmemory research, in conjunction with powerful neuro-biological technologies, have provided striking supportfor a very dynamic neurobiological basis of memory,which is beginning to overturn the old dogma.
For .100 years, generations of behavioural paradigmsand technologies have been used to address questionsabout the mechanisms that mediate learning and memory[1–3]. Repeatedly, evidence has been found to suggest thatthe properties of the memory trace change in a time-dependent manner, such that new memories are initiallyin a dynamic ‘labile’ form for a short time [short-termmemory (STM)], after which the memory trace is ‘fixed’ or‘consolidated’ into the physical structure of the brain [long-term memory (LTM)] [4–6]. For example, electroconvul-sive shock (ECS) is effective in inducing amnesia ifpresented shortly after training (during STM) but not ifgiven a few hours later (during LTM) [7]. Time-dependenteffects such as these are the cornerstone of memoryconsolidation theory (now called cellular consolidationtheory [8]). During the past 40 years, incredible effortshave been made to describe across all levels of analysisthe processes that contribute to the transformation of atrace from being labile to being fixed [9,10]. Of note isthe finding that the transcription factor Ca2!-response-element-binding protein (CREB), transcription and trans-lation all seem to be universal neuronal requirements fortraces to enter LTM [11–15] (Fig. 1a).
Early studies on reconsolidationIn 1968, the view that memories are consolidated overtime into a permanent state was challenged by Lewisand colleagues [16]. In agreement with previous studies,when ECS was given 24 h after fear conditioning it wasineffective in generating amnesia. However, if the memorywas reactivated before ECS administration, amnesia wasobserved the following day. Given that amnesia was notproduced in the absence of memory reactivation, thememory is defined as being consolidated by that time.Therefore, reactivation of a consolidated memory presum-ably returned it to a labile state, which initiated another
time-dependent memory process similar to that seen afternew learning. This phenomenon is now referred to asreconsolidation [17–19]. Lewis’ study defined a paradigmfor experimentally differentiating consolidation and recon-solidation: a necessary criterion if an effect is to be attri-buted to reconsolidation is that the amnesic agent must be
Fig. 1. Two models of memory processing. (a) The traditional consolidation theory,which posits a labile, short-term memory (STM) state and a later, consolidatedlong-term memory (LTM) state. Once fixed in LTM, the memory is posited to bepermanent. Below each memory state is a list that is typically used to describesome of the properties of the two states. (b) The memory model proposed byLewis [33]. The active state (AS) and inactive state (IS) are analogous to STM andLTM, respectively. The molecular descriptors in brackets were not part of theoriginal model but have been inserted for comparison with (a). New memoriesenter a labile AS and then with time enter the IS [top red arrow, again similar to(a)]. Reactivation of memories that are in an IS returns them to the AS (bottom redarrow). Both new and reactivated memories require protein-synthesis-dependentmechanisms in order to enter the IS. Contrary to consolidation theory, whichcannot explain the reconsolidation data, this model incorporates both the datafrom consolidation and reconsolidation experiments.
TRENDS in Neurosciences
Short-term memory (STM)
• Lasts for seconds to hours
• ‘Labile’ (sensitive to disruption)
• Does not require new RNA or protein synthesis
Active state (AS)
• Lasts for seconds to hours
• ‘Labile’ (sensitive to disruption)
(Does not require new RNA or protein synthesis)
Long-term memory (LTM)
• Lasts for days to weeks
• Consolidated (insensitive to disruption)
• Does require new RNA or protein synthesis
Inactive state (IS)
• Lasts for days to weeks
• Inactive (insensitive to disruption)
(Does require new RNA or protein synthesis)
(a)
(b)
Corresponding author: Karim Nader ([email protected]).
Opinion TRENDS in Neurosciences Vol.26 No.2 February 2003 65
http://tins.trends.com 0166-2236/03/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved. PII: S0166-2236(02)00042-5
Nader, 2003, TiNS
KSBSaniyelerden saatlere
Hasara açık Yeni RNA veya protein sentezine ihtiyaç yok
USBGünlerden haftalaraHasara kapalı (sabit)
Yeni RNA veya protein sentezine ihtiyaç var
Aktif DurumSaniyelerden saatlere
Hasara açık Yeni RNA veya protein sentezine ihtiyaç yok
İnaktif DurumSaniyelerden saatlere
Hasara açık Yeni RNA veya protein sentezine ihtiyaç yok
Monday, November 7, 2011

Hem sinaptik hem sistem
Neuron, Vol. 36, 527–538, October 24, 2002, Copyright !2002 by Cell Press
Cellular and Systems Reconsolidationin the Hippocampus
studies demonstrating that amnesia could also occur ifa fully consolidated and stable LTM was reactivatedprior to amnesic treatments (Misanin et al., 1968). This
Jacek Debiec,1,2 Joseph E. LeDoux,2
and Karim Nader3,4
1Department of Psychiatryphenomenon has been described in a large number ofJagiellonian University Collegium Medicumspecies, using a wide array of behavioral paradigms andCracow 31-501amnesic agents (Sara, 2000). These findings suggestedPolandthat old, reactivated memories undergo another round of2 W.M. Keck Foundation Laboratory of Neurobiologyconsolidation, a process referred to as reconsolidationCenter for Neural Science(Nader et al., 2000b; Przybyslawski and Sara, 1997).New York UniversityConsistent with the reconsolidation hypothesis is ourNew York, New York 10003recent demonstration that consolidated memories for3 Department of Psychologyauditory fear conditioning, which are stored in the amyg-McGill Universitydala (Fanselow and LeDoux, 1999; Maren, 2001; SchafeMontreal, Quebec H3A 1B1et al., 2000), undergo protein synthesis-dependent re-Canadaconsolidation in the amygdala and that this process iscontingent on memory reactivation (Nader et al., 2000a).Indeed, reconsolidation and consolidation have beenSummaryfound to share a number of common properties, includ-ing: (1) requirement of protein synthesis in order forCellular theories of memory consolidation posit thatthe memory to persist, (2) time windows during whichnew memories require new protein synthesis in orderprotein synthesis blockade is effective, and (3) that pro-to be stored. Systems consolidation theories posit thattein synthesis blockage in the same brain region, thethe hippocampus has a time-limited role in memoryamygdala, disrupts both. Given these similarities, itstorage, after which the memory is independent of theseemed parsimonious to conclude that a new memoryhippocampus. Here, we show that intra-hippocampaland a reactivated, consolidated memory share a com-infusions of the protein synthesis inhibitor anisomycinmon memory state, as originally proposed by Lewiscaused amnesia for a consolidated hippocampal-(1979). Thus, instead of just occurring once, memorydependent contextual fear memory, but only if thestorage may instead be a process that is reiterated withmemory was reactivated prior to infusion. The effecteach use of the memory.occurred even if reactivation was delayed for 45 days
A key issue is whether reconsolidation also occursafter training, a time when contextual memory is inde-in other brain systems. The most extensively studiedpendent of the hippocampus. Indeed, reactivation ofmemory system of the brain involves the hippocampus.a hippocampus-independent memory caused the traceResults from previous studies have suggested thatto again become hippocampus dependent, but onlymemories for hippocampus-dependent tasks can un-for 2 days rather than for weeks. Thus, hippocampaldergo reconsolidation (Mactutus et al., 1979; Przybys-memories can undergo reconsolidation at both thelawski et al., 1999; Schneider and Sherman, 1968). Forcellular and systems levels.example, using a radial arm maze, systemic postreacti-vation injections of propranol were effective at produc-Introductioning amnesia if the memory was first reactivated (Przy-byslawski et al., 1999). Because the treatment wasThe formation of long-term memory (LTM) is generallysystemic, however, it is not known whether the effectsbelieved to involve a process by which a labile short- of the drug on reconsolidation occurred in the hippo-
term memory (STM) is converted into a lasting stable campus or in some other structure that contributes totrace (Ebbinghaus, 1885; Hebb, 1949; Muller and Pil- this task. Similarly, recent findings that disruption ofzecker, 1900). Evidence for this time-dependent process CREB-mediated transcription in the forebrain interfereshas come from numerous studies, showing that treat- with the reconsolidation of contextual fear memoriesments such as electroconvulsive shock (ECS) produce (Kida et al., 2002) suffer from the same drawback. Inamnesia shortly after learning, but the same treatment support of the possibility that memories stored withinseveral hours later has no effect (Duncan, 1949; McGaugh, the hippocampus itself might undergo reconsolidation1966). The dominant view of how the conversion from are the recent findings showing that reactivation of con-STM to LTM occurs is that new RNA and proteins are textual memories induces the expression of zif268, asynthesized and transform temporary alterations in syn- gene implicated in consolidation of new hippocampal-aptic transmission into persistent modifications of syn- dependent memories (Hall et al., 2001).aptic architecture (Davis and Squire, 1984; Flexner et In the present study, we first asked if hippocampal-al., 1965; Goelet et al., 1986). This is called consolidation mediated memories undergo protein synthesis-depen-theory, or more precisely, cellular consolidation theory dent reconsolidation in the hippocampus. The task we(Dudai and Morris, 2000). used was contextual fear conditioning, a variant of audi-
Cellular consolidation theory was challenged by early tory fear conditioning in which the footshock comes tobe associated with the chamber (context) in which theshock occurred. The hippocampus is thought to estab-4 Correspondence: [email protected]
Monday, November 7, 2011

İnsanlarda da oluyor
•1. listeyi ögren (masa, kulak, çiçek, düdük)•Bir gün sonra hatırlat veya hatırlatma, ve ardından yeni bir liste ögren•Ertesi gün•Hatırlatma oldu ise, birinci listeye ikinci listeden öğe ekleme
•Olmadı ise böyle birşey yok
Reconsolidation of episodic memories: A subtlereminder triggers integration of new informationAlmut Hupbach,1,3 Rebecca Gomez,1 Oliver Hardt,2 and Lynn Nadel11Department of Psychology, The University of Arizona, Tucson, Arizona 85721, USA; 2Department of Psychology, McGillUniversity, Montreal, Quebec H3A 2T5, Canada
Recent demonstrations of “reconsolidation” suggest that memories can be modified when they are reactivated.Reconsolidation has been observed in human procedural memory and in implicit memory in infants. This study askswhether episodic memory undergoes reconsolidation. College students learned a list of objects on Day 1. On Day 2,they received a reminder or not, and then learned a second list. Memory for List 1 was tested immediately on Day 2(Experiment 2) or on Day 3 (Experiment 1). Although the reminder did not moderate the number of items recalledfrom List 1 on either day, subjects who received a reminder incorrectly intermixed items from the second list whenrecalling List 1 on Day 3. Experiment 2 showed that this effect does not occur immediately and thus istime-dependent. The reminder did not affect memory for List 2 on Day 3 (Experiment 3), demonstrating thatmodification occurred only for the original memory (List 1). The study demonstrates the crucial role of remindersfor the modification of episodic memory, that reconsolidation of episodic memory is time-dependent, and, incontrast to previous reconsolidation findings, that reconsolidation is also a constructive process, one that supportsthe incorporation of new information in memory.
Consolidation theory assumes that memories are labile during alimited window after encoding, but as time passes, memories areconsolidated and become resistant to change (e.g., McGaugh2000). The rediscovery of reactivation-induced reconsolidationhas challenged this view (cf. Przybyslawski and Sara 1997; Sara2000; Nader 2003; cf. Lewis et al. 1968; Misanin et al. 1968, forearly demonstrations of the reconsolidation phenomenon). Incontrast to the consolidation account, reactivation is thought toreturn memories to a labile state, which allows them to change.In the normal course of events, reactivated memories are retainedthrough a process similar to initial consolidation, i.e., reconsoli-dation. This view of memory formation as a dynamic processrather than a static one has implications for theories of memoryand for our understanding of memory change.
Nader et al. (2000) showed that memories are labile by dem-onstrating that fear memory in the rat can be severely impairedat the time the memory is recalled. Rats received several tone-footshock pairings on Day 1. The following day, half of the ani-mals were again exposed to the tone. The tone elicited freezing,indicating that the rats recalled the conditioned fear memory. Allrats were then injected with a protein-synthesis inhibitor or itsvehicle into the amygdala. When tested for fear memory in re-sponse to the tone on Day 3, rats who had received the remind-ing tone before injection of the inhibitor showed less freezingthan the vehicle-injected rats and, most importantly, than ratswho were not reminded. This demonstrated that reactivation ofthe fear memory returned the memory to a labile state, whichlike a newly acquired, nonconsolidated memory requires de novoprotein-synthesis in order to be maintained. The protein-synthesis inhibitor blocked reconsolidation, and thus led tomemory impairment. This finding has now been replicated in avariety of species and paradigms (see Dudai and Eisenberg 2004;Alberini 2005).
Walker et al. (2003) recently demonstrated reconsolidation
effects in humans. Participants were trained on a procedural mo-tor-skill task that involved finger-tapping a simple sequence (e.g.,4-1-3-2). Twenty-four hours later they briefly rehearsed the se-quence (reactivating it) and learned a second sequence (e.g., 2-3-1-4). When tested on Day 3, accuracy performance for Sequence1 was significantly impaired in comparison to a group of partici-pants who did not rehearse Sequence 1 before learning Sequence2. This shows that the reactivation of the memory for Sequence1 on Day 2 destabilized it such that a competing motor patterncould interfere.
Galluccio (2005) and Galluccio and Rovee-Collier (2005) in-vestigated the fate of reactivated memories in infants trained tokick their foot to activate a mobile. After a delay, infants werereminded of the event: The moving mobile was presented for abrief period during which it was no longer attached to the baby’sfoot. After reactivation, one group of infants learned to move anovel mobile. One day later, infants who were exposed to thenovel mobile showed a modification of the reactivated memoryin that they no longer recognized the original mobile and solelyreacted to the novel one.
Thus, reconsolidation effects have been shown in humans,but only for procedural memory (Walker et al. 2003) and condi-tioning (Galluccio 2005). These are both forms of implicitmemory that do not require conscious recollection, making itimportant to ask whether reconsolidation also applies to explicitmemory, a form of memory that allows for the conscious recol-lection of events (episodic memory) and facts (semanticmemory).
Although the idea of fixed consolidated memory traces hasbeen the dominant view in neurobiology, it has long beenknown in cognitive psychology that episodic memories can bereconstructed. For instance, Bartlett’s (1932) classic study inwhich participants were repeatedly asked to recall a NativeAmerican folktale showed that, over time, participants recon-structed their recall to better fit their own cultural schema. It isalso well known, from the study of retroactive interference, thata subsequent learning experience can impair performance forpreviously acquired material (e.g., Barnes and Underwood 1959;Delprato 2005). And we know that episodic memories can be
3Corresponding author.E-mail [email protected]; fax (520) 621-9306.Article published online in January 2007. Article and publication date are athttp://www.learnmem.org/cgi/doi/10.1101/lm.365707.
Research
14:47–53 ©2007 by Cold Spring Harbor Laboratory Press ISSN 1072-0502/07; www.learnmem.org Learning & Memory 47www.learnmem.org
on August 21, 2007 www.learnmem.orgDownloaded from
Learning & Memory, 2007
Monday, November 7, 2011

Hafıza değişimi•Sağlamlaşma => BDNF (beyin-türetilmiş nörotrofik faktör
•Yeniden sğlamlaşma => zif268•Lee, 2008, Nature Neuroscience
fear-conditioning procedure was used, in which both trials werecondensed into a single session, memory consolidation remaineddependent on hippocampal BDNF and not on zif268 (Fig. 1b,c andSupplementary Fig. 5 online). ANOVA revealed an overall significantsession! gene! ASO interaction (F2,44 " 11.11, Po 0.001). Analysisof the zif268 groups alone revealed nomain effect of ASO (F1,12" 1.05,P " 0.32) or session ! ASO interaction (F2,24 " 1.23, P " 0.31),whereas analysis of the BDNF groups revealed a significant session !ASO interaction (F2,20 " 19.95, P o 0.001) that was driven by asignificant effect of ASO during the long-term memory (LTM) tests(simple effects one-way ANOVA, Po 0.05), with no effect on the STMtest. Therefore, these data reveal a double dissociation between thecellular mechanisms of initial memory consolidation (BDNF) andmemory strengthening through additional learning (zif268). Moreover,the selective dependence of the two processes on their relative cellularmechanisms was not a result of nonspecific or quantitative factors.A further contention might be that zif268 is not required for
memory strengthening per se, but is instead required for synapticplasticity in neural circuits that have been modified recently bybehavioral experience. Thus, the selective dependence of additionallearning on zif268may result not from the factthat an existing memory is being updated andstrengthened, but arises because some learn-ing experience, which need not have beenrelated, engaged the dorsal hippocampus onthe previous day. To test this hypothesis, wefirst conditioned rats in a separate context(different operant chambers12), before return-ing them to the standard fear-conditioningapparatus (context 2) for the second day oftraining. Infusion of zif268 ASO into thedorsal hippocampus before this second con-ditioning trial had no effect on subsequentconditioned freezing in context 2, whereasknockdown of hippocampal BDNF resultedin severe amnesia (Fig. 2a and Supplemen-tary Fig. 6 online). Although ANOVArevealed no gene ! ASO interaction duringthe first conditioning session (F o 1), therewas an overall significant session ! gene !ASO interaction in context 2 (F1,22 " 11.31,P " 0.003). Analysis of the zif268 groupsalone revealed no main effect of ASO or
session ! ASO interaction (F values o 1), whereas the BDNF groupshad a significant session ! ASO interaction (F1,12 " 22.17, Po 0.001)that was driven by a significant effect of ASO during the LTM test(simple effects one-way ANOVA, Po 0.05). Thus, even when there hasbeen recent synaptic plasticity in the hippocampus, contextual-fearconditioning functionally recruits consolidation, but not reconsolida-tion, mechanisms. Moreover, these results provide strong evidence thatmemory strengthening has underlying mechanisms that are separatefrom those of memory acquisition and consolidation.If the mechanisms of reconsolidation are the same as those that
strengthen the memory trace, impairing memory destabilizationshould prevent the modification of memory strength. It has beenshown recently that hippocampal synaptic protein degradation is acritical process in the destabilization of contextual-fear memories8.Infusion of the proteasome inhibitor clasto-Lactacystin-b-lactone intothe dorsal hippocampus prevented the amnesic action of anisomycin ina standard memory reconsolidation setting. Thus, although anisomy-cin infusion alone, immediately after contextual re-exposure, resultedin subsequent amnesia, the co-infusion of blac rendered the memoryinvulnerable to protein synthesis inhibition8.
80
a b c
60
Per
cent
age
time
free
zing
Per
cent
age
time
free
zing
Per
cent
age
time
free
zing
40
20
0
80
100
60
40
20
STM LTM LTM2
Test
STM LTM LTM2
Test
0
80
100
60
40
20
0
Cond1 Cond2
Session
Test
BDNF MSO
BDNF ASOBDNF MSO
BDNF ASOzif268 MSO
zif268 ASOZif268 MSO
Zif268 ASO
Figure 1 The cellular mechanisms of memory consolidation and strengthening differ. (a) Knockdown of zif268, but not BDNF, during a second conditioningsession resulted in subsequent amnesia. Rats were fear conditioned on 2 consecutive days (Cond1 and Cond2) and were tested 24 h later (Test). Behavioralmeasurements are shown for the periods before foot shock presentation during conditioning and for the whole test (n " 5–6 per group). (b,c) Knockdown ofBDNF (b), but not zif268 (c), during a single two-trial conditioning session resulted in subsequent amnesia. Behavioral measurements are shown for the STMtest (3 h after conditioning) and for two LTM tests (24 h, LTM; 7 d, LTM2; n " 6–7 per group). Data presented as mean ± s.e.m.
70
a b
60
50
40
30
20
10
Cond1 Cond2 Test Cond1 Cond2 Test
CX1 CX2 Session
0
60
80
40
20
0
Per
cent
age
time
free
zing
Per
cent
age
time
free
zing
BDNF MSO Veh/Veh
!lac/Veh
!lac/ANI
Veh/ANIBDNF ASO
Zif268 MSO
Zif268 ASO
Figure 2 Memory strengthening requires destabilization and reconsolidation. (a) Knockdown of BDNF,but not zif268, during fear conditioning to a changed context resulted in subsequent amnesia. Rats werefear conditioned first to context 1 (CX1, Cond1), then to CX2 on the next day (Cond2), and were tested24 h later (Test) in CX2. Behavioral measurements are shown for the periods before foot shockpresentation during conditioning and for the whole test (n " 6–7 per group). (b) Proteasome inhibitionprotected a contextual-fear memory against both additional learning and amnesia. Rats were fearconditioned on 2 consecutive days (Cond1 and Cond2), and were tested 24 h later (Test). Behavioralmeasurements are shown for the periods before foot shock presentation during conditioning and for thewhole test (n " 7 per group). Data presented as mean ± s.e.m. MSO, control missenseoligodeoxynucleotide sequences; Veh, vehicle.
NATURE NEUROSCIENCE VOLUME 11 [ NUMBER 11 [ NOVEMBER 2008 1265
BR I E F COMMUNICAT IONS
Monday, November 7, 2011

Kısaca
Brief Communications
Sensory-Specific Associations Stored in the LateralAmygdala Allow for Selective Alteration of Fear Memories
Lorenzo Díaz-Mataix,1 Jacek Debiec,1,2,3 Joseph E. LeDoux,1,4 and Valerie Doyere1,5,6
1Center for Neural Science, New York University, New York, New York 10003, 2New York University Child Study Center, New York, New York 10016,3Copernicus Center for Interdisciplinary Studies, Krakow 30387, Poland, 4Emotional Brain Institute, Nathan S. Kline Institute for Psychiatry Research,Orangeburg, New York 10962, 5Universite Paris-Sud, Centre de Neurosciences Paris-Sud, UMR 8195, Orsay F-91405, France, and 6CNRS, Orsay F-91405,France
Consolidated long-term fear memories become labile and can be disrupted after being reactivated by the presentation of the uncondi-tioned stimulus (US). Whether this is due to an alteration of the conditioned stimulus (CS) representation in the lateral amygdala (LA) isnot known. Here, we show in rats that fear memory reactivation through presentation of the aversive US, like CS presentation, triggers aprocess which, when disrupted, results in a selective depotentiation of CS-evoked neural responses in the LA in correlation with a selectivesuppression of CS-elicited fear memory. Thus, an aversive US triggers the reconsolidation of its associated predictor representation in LA.This new finding suggests that sensory-specific associations are stored in the lateral amygdala, allowing for their selective alteration byeither element of the association.
IntroductionIn Pavlovian auditory fear conditioning, an initially neutral con-ditioned acoustic stimulus (CS), after being associated with anaversive unconditioned stimulus (US), typically electric shock,acquires the ability to induce conditioned fear responses (Pavlov,1927). This form of associative learning critically involves thelateral nucleus of the amygdala (LA). Although the LA is believedto be a storage site for such fear memories (Davis, 1997; Fanselowand LeDoux, 1999; LeDoux, 2000; Maren, 2001; Fanselow andPoulos, 2005), the nature of the information stored in LA is un-clear. A key issue is whether CS–US associations encoding spe-cific sensory features of both stimuli are stored in the LA.
After consolidation of the fear memory, presentation of theUS triggers a reconsolidation process which, when disrupted,leads to a decrease in CS-evoked behavioral fear memory in anamygdala-dependent manner (Debiec et al., 2010). It has beenproposed that the neural representation of the CS activatessensory-specific representations of the US, which, in turn, elicitsthe corresponding behavioral conditioned responses (Konorski,1967). Thus, the loss of fear behavior when US-triggered recon-solidation is disrupted may conceivably result from the disruption of
either CS or US neural representation. Using a second-order con-ditioning paradigm, we have previously shown that not allthe fear associations are rendered labile in an amygdala-dependent manner after reactivation of one of them (Debiec etal., 2006). It is therefore possible, as one could argue, thatalthough amygdala-dependent, the behavioral disruption mayreflect alteration of processes in brain networks other than theLA. Furthermore, even if some altered processes reside in theLA, they may not necessarily affect the CS representation, as adisrupted US representation may solely be responsible for the loss ofbehavioral-conditioned responses after disruption of US-triggeredreconsolidation processes. Alternatively, stored first-order CS–USassociation in the LA could permit the CS representation to be ren-dered labile after US reactivation and produce a sensory-specific fearmemory loss. A selective updating of CS representation would pro-vide evidence that specific CS–US associations linked through neuralplasticity are stored in the LA.
In the present study, we thus asked whether US reactivation trig-gers an update of the CS fear representation in the LA and producesa reconsolidation process during which the CS representation is de-stabilized. To do this, we recorded CS-evoked field potentials in LAand tracked the effects of US-induced reconsolidation blockade onthe CS representation in the LA. We used an MAPK inhibitor toblock the reconsolidation process, allowing the comparison of ourresult with previous data showing that the MAPK/ERK pathway isinvolved in reconsolidation triggered by the CS (Duvarci et al., 2005;Doyere et al., 2007). To control for the selectivity of the effects, weused a dual-fear conditioning protocol in which rats were trained totwo different conditioned stimuli paired with two different aversiveunconditioned stimuli (Debiec et al., 2010). We found that theblockade of US-triggered reconsolidation results in a selective era-sure of the CS neural representation previously linked to that US butnot of the CS linked to the other US.
Received Nov. 4, 2010; revised April 16, 2011; accepted May 9, 2011.Author contributions: L.D.-M., J.D., and V.D. designed research; L.D.-M. and V.D. performed research; L.D.-M. and
V.D. analyzed data; L.D.-M., J.E.L., and V.D. wrote the paper.This work was supported by the National Institute for Mental Health Grants R01 MH46516 and R01 MH38774, and
McDonnell Foundation Grant 220020155. L.D.-M. was recipient of a Fulbright-Spanish Ministry of Science postdoc-toral fellowship. We thank Dr. Joshua Johansen for his helpful suggestions revising the manuscript and Claudia Farbfor her excellent assistance with histology.
The authors declare no competing financial interests.This article is freely available online through the J Neurosci Open Choice option.Correspondence should be addressed to Lorenzo Díaz-Mataix, Center for Neural Science, New York University, 4
Washington Place, Room 809, New York, NY 10003. E-mail: [email protected]:10.1523/JNEUROSCI.5808-10.2011
Copyright © 2011 the authors 0270-6474/11/319538-06$15.00/0
9538 • The Journal of Neuroscience, June 29, 2011 • 31(26):9538 –9543
2011, Haziran
©20
11 N
atur
e A
mer
ica,
Inc.
All
righ
ts r
eser
ved.
NATURE NEUROSCIENCE ADVANCE ONLINE PUBLICATION 1
A R T I C L E S
Aversive associative memories formed by the association between a neutral conditioned stimulus and an aversive unconditioned stimulus are progressively made permanent by a process of consolidation1. However, upon retrieval, intervention by amnestic agents2–7, either before or immediately after retrieval, results in disruption of the pre-viously consolidated fear memory. This suggests that a consolidated memory returns to a transient destabilized state shortly after reac-tivation, necessitating a dynamic time-dependent process of recon-solidation to persist further. During this reconstruction, a memory is vulnerable to experimental intervention8–10 leading to amnesia, but can also be enhanced11–13 or modified in the long-term14–16, thereby updating the previous memory with new information14–17. In clini-cal terms, the bidirectional and adaptive nature of reconsolidation is ideally placed to mediate the modification of both memory strength12 and memory content16,18, rendering this process a promising thera-peutic target for counteracting the hyper-responsive fear system. To fully exploit reconsolidation-based therapies that adapt the content of fear memories, leading to a loss of fear response in the long term, it is crucial to elucidate the molecular underpinnings of reconsolidation, which remain obscure.
Long-lasting changes in synaptic efficacy brought about by gene transcription, protein synthesis and changes in strength of hippo-campal glutamatergic synapses through AMPA receptor trafficking are believed to be the cellular substrates of learning and memory19–21. Although reconsolidation is not merely a recapitulation of the initial consolidation process22, it has been shown that transcription, de novo protein synthesis and synaptic protein degradation in the hippocampus
are necessary for memory remodeling after retrieval4,7,17,23–25. Here, we investigated whether the temporal profile of reconsolidation, which is hypothesized to be limited to a 6 h time window5,8, reflects a sequential profile of defined dorsohippocampal AMPA receptor synaptic plastic-ity that is crucial to the synaptic remodeling that underlies subsequent fear expression (changes in memory strength) and reinterpretation of fear memory after retrieval (changes in memory content).
RESULTSMemory recall induces acute hippocampal AMPAR endocytosisTo analyze whether glutamate receptors are regulated during reconsolida-tion in mice receiving the unconditioned stimulus and retrieval (US-R), we dissected the dorsal hippocampus at 1 and 4 h after retrieval and analyzed the synaptic membrane fraction, including membrane-bound proteins and associated proteins26,27, by immunoblotting for subunits of AMPA receptors. A no-shock group experiencing retrieval (NS-R) was used to control for the specificity of an aversive associative memory (Supplementary Fig. 1). These two time points were chosen because they fall within the 6-h time window after retrieval during which the memory undergoes reconsolidation5. After retrieval, subsequent recon-solidation requires protein synthesis for the memory to persist further7 (Supplementary Fig. 1). First, the 1 h time point was analyzed. All AMPAR subunits (GluA1–GluA3) were downregulated (31.4%, 16.4% and 50.20%, respectively; P < 0.05), indicating a weakened state of the synapse28,29 (Fig. 1a,b and Supplementary Fig. 2). The observed downregulation was specific to retrieval of an associative contextual conditioned stimulus–unconditioned stimulus representation, with
1Department of Molecular & Cellular Neurobiology, Center for Neurogenomics & Cognitive Research, Neuroscience Campus Amsterdam, VU University (Vrije Universiteit), Amsterdam, The Netherlands. 2Department of Integrative Neurophysiology, Center for Neurogenomics & Cognitive Research, Neuroscience Campus Amsterdam, VU University, Amsterdam, The Netherlands. 3Department of Functional Genomics, Center for Neurogenomics & Cognitive Research, Neuroscience Campus Amsterdam, VU University, Amsterdam, The Netherlands. 4These authors contributed equally to this work. Correspondence should be addressed to S.S. ([email protected]).
Received 2 May; accepted 18 July; published online 11 Septmber 2011; doi:10.1038/nn.2907
Retrieval-specific endocytosis of GluA2-AMPARs underlies adaptive reconsolidation of contextual fearPriyanka Rao-Ruiz1,4, Diana C Rotaru1,2,4, Rolinka J van der Loo1, Huibert D Mansvelder2, Oliver Stiedl1,3, August B Smit1 & Sabine Spijker1
Upon retrieval, fear memories are rendered labile and prone to modification, necessitating a restabilization process of reconsolidation to persist further. This process is also crucial for modulating both strength and content of an existing memory and forms a promising therapeutic target for fear-related disorders. However, the molecular and cellular mechanism of adaptive reconsolidation still remains obscure. Here we show that retrieval of fear memory induces a biphasic temporal change in GluA2-containing AMPA-type glutamate receptor (AMPAR) membrane expression and synaptic strength in the mouse dorsal hippocampus. Blockade of retrieval-induced, regulated, GluA2-dependent endocytosis enhanced subsequent expression of fear. In addition, this blockade prevented the loss of fear response after reconsolidation-update of fear memory content in the long-term. Thus, endocytosis of GluA2-containing AMPARs allows plastic changes at the synaptic level that exerts an inhibitory constraint on memory strengthening and underlies the loss of fear response by reinterpretation of memory content during adaptive reconsolidation.
2011, Eylül
Monday, November 7, 2011

Teşekkürler!!!
Monday, November 7, 2011














![SİNEMATOGRAFİK MASAL - Turuz€¦ · Sinemanın Masalları, Bir Yüzyılın Tarihleri [Hikayeleri] Belgesel Kurmaca: Marker ve Hatıra Kurmacası Kıssadan Hissesi Olmayan Bir Masal:](https://static.fdocuments.net/doc/165x107/61494182080bfa6260147e41/snematografk-masal-turuz-sinemann-masallar-bir-yzyln-tarihleri.jpg)


