
Growth of the fetal calf and its arterial pressure, blood...
5
JOURNAL OF APPLIED PHYSIOLOGY Vol. 32, No. 2, February 1972. Printed in U.S.A. Growth of the fetal calf and its arterial pressure, blood gases, and hematologic data JOHN T. REEVES, FUHEID S. DAOUD, AND MICHAEL GENTRY De$artment of Medicine, University of Kentucky School of Medicine, Lexington, Kentucky 40506 REEVES, JOHN T., FUHEID S. DAOIJD, AND MICHAEL GENTRY. Growth of the fetal calf and its arterial pressure, blood gases, and hemato- logic data. J. Appl. Physiol. 32(2): 240-244. 1972.-Fetal growth, arterial pressure, arterial oxygen tensions, and hematologic data in 35 fetal calves were compared with similar data in fetal lambs. The data of the present report suggested that the fetal calf, its organs, and placenta had rates of growth which were similar to those in the lamb. The increasing arterial pressure during the last third of gestation was similar to that in the lamb and the higher arterial pressure achieved by the mature calf appeared to reflect merely its larger size. The fetal calf had a relatively high arterial oxygen tension which suggested there may be an efficient trans- port of oxygen across the placenta. The hemoglobin concentration was lower than that in the lamb and did not change during the last third of gestation. Thus the differences in oxygen transport in the fetal calf and lamb may reside primarily in the arterial oxygen tensions and the blood oxygen carrying capacities. bovine; fetus; fetal growth; oxygen; hemoglobin concentration MAMMALIAN FETUSES grow rapidly during the last third of gestation despite a relatively slow growth of the placenta (4). Data from the fetal lamb suggest that a progressive in- crease in arterial pressure increases placental blood flow dur- ing the period of rapid fetal growth (4). Increasing placental blood flow is probably a major mechanism “providing an increased oxygen transport across the placenta to the fetus. Acute experiments in fetal lambs suggest there also may be an increase in the oxygen carrying capacity of the fetal blood during the last trimester of gestation (1, 4). One wonders how fetuses larger than the lamb utilize various mechanisms for oxygen transport at a time when growth of the fetus out- strips placental growth. The fetal calf is large and may be conveniently studied (12). However, the growth curves of the fetal calf and its placenta are not available for the last third of gestation. A previous study of fetal calves in the 7 th and 8 th months of gestation suggested that the arterial pressures and oxygen tensions were higher than in fetal lambs (12). More complete data were needed for the fetal calf. The present study reports measurements of bovine fetal growth, arterial pressure, arte- rial blood gas values, and hemoglobin concentrations from 6 through 9 months of gestation. METHODS Methods of studying the liue fetal calf. Thirty-five cesarean set tions were performed under local anesthesia in Jersey cows (age 2-l 1 years) judged to be healthy by the consulting veterinarian. The handling of the mother cows and the sur- gical procedure have been described (12). The cows had been bred by artificial insemination and the ages of the fetuses varied between 163 and 281 days. The duration of gestation is approximately 281 days in the Jersey cow. In the first 12 experiments the fetuses were fully delivered onto a warmed table. In subsequent experiments only the head (12 exp) or the foreleg (11 exp) was delivered from the uterus. After local anesthesia had been administered to the fetus a carotid artery or a brachial artery was exposed and intubated with polyethylene tubing. Fetal arterial pressure was recorded with a Sanborn 150 one-channel or an Elec- tronics for Medicine four-channel recorder. Heart rates were counted at I- to 2-min intervals from the pressure recording. Ten to thirty minutes after intubation of the fetal artery, and at a time when the fetal heart rate and arterial pressure were stable, simultaneous samples were drawn from catheters in the jugular vein (or systemic artery) of the mother and the artery of the fetus. Blood samples for hematologic study were placed in tubes containing sodium citrate. Arterial samples for blood gas analysis were drawn into heparinized glass syringes and placed on ice. The calves were then utilized for other experiments ( 12). Weights of fetal calves and fetal organs. At the end of the ex- periment, the fetus was delivered, the cord tied, and the fetus weighed. After the last gasp, fetuses of 260 days or younger were dissected to obtain fetal organ weights. The skin, in- cluding the tail and the ears and excluding the hooves, was removed by sharp dissection and weighed while still wet. The two lungs were weighed together attached to a short portion of trachea. The heart was weighed unopened after the great vessels and pericardium had been removed. The kidneys were weighed after the fat was removed. The brain weight included the cerebral hemispheres, midbrain, cere- bellum, and the medulla oblongata to the level of the fora- men magnum. The weight of the “guts” included the stomach, intestines, bladder, and their contents, and the pancreas, mesenteric fat, adrenals, and ureters. A portion of the gastric contents was occasionally lost in the dissection. In 10 experiments the placenta was manually removed from each cotyledon. Although 2-4 hr were required to remove the placenta, removal was incomplete since a portion was retained within the depressions in the maternal cotyledons. The placenta weight (expressed as a percent of fetal weight) includes placenta, membranes, and a portion of the umbil- ical cord.
-
Upload
nguyencong -
Category
Documents
-
view
214 -
download
0
Transcript of Growth of the fetal calf and its arterial pressure, blood...

JOURNAL OF APPLIED PHYSIOLOGY Vol. 32, No. 2, February 1972. Printed in U.S.A.
Growth of the fetal calf and its arterial
pressure, blood gases, and hematologic data
JOHN T. REEVES, FUHEID S. DAOUD, AND MICHAEL GENTRY De$artment of Medicine, University of Kentucky School of Medicine, Lexington, Kentucky 40506
REEVES, JOHN T., FUHEID S. DAOIJD, AND MICHAEL GENTRY. Growth of the fetal calf and its arterial pressure, blood gases, and hemato- logic data. J. Appl. Physiol. 32(2): 240-244. 1972.-Fetal growth, arterial pressure, arterial oxygen tensions, and hematologic data in 35 fetal calves were compared with similar data in fetal lambs. The data of the present report suggested that the fetal calf, its organs, and placenta had rates of growth which were similar to those in the lamb. The increasing arterial pressure during the last third of gestation was similar to that in the lamb and the higher arterial pressure achieved by the mature calf appeared to reflect merely its larger size. The fetal calf had a relatively high arterial oxygen tension which suggested there may be an efficient trans- port of oxygen across the placenta. The hemoglobin concentration was lower than that in the lamb and did not change during the last third of gestation. Thus the differences in oxygen transport in the fetal calf and lamb may reside primarily in the arterial oxygen tensions and the blood oxygen carrying capacities.
bovine; fetus; fetal growth; oxygen; hemoglobin concentration
MAMMALIAN FETUSES grow rapidly during the last third of gestation despite a relatively slow growth of the placenta (4). Data from the fetal lamb suggest that a progressive in- crease in arterial pressure increases placental blood flow dur- ing the period of rapid fetal growth (4). Increasing placental blood flow is probably a major mechanism “providing an increased oxygen transport across the placenta to the fetus. Acute experiments in fetal lambs suggest there also may be an increase in the oxygen carrying capacity of the fetal blood during the last trimester of gestation (1, 4). One wonders how fetuses larger than the lamb utilize various mechanisms for oxygen transport at a time when growth of the fetus out- strips placental growth.
The fetal calf is large and may be conveniently studied (12). However, the growth curves of the fetal calf and its placenta are not available for the last third of gestation. A previous study of fetal calves in the 7 th and 8 th months of gestation suggested that the arterial pressures and oxygen tensions were higher than in fetal lambs (12). More complete data were needed for the fetal calf. The present study reports measurements of bovine fetal growth, arterial pressure, arte- rial blood gas values, and hemoglobin concentrations from 6 through 9 months of gestation.
METHODS
Methods of studying the liue fetal calf. Thirty-five cesarean set tions were performed under local anesthesia in Jersey cows
(age 2-l 1 years) judged to be healthy by the consulting veterinarian. The handling of the mother cows and the sur- gical procedure have been described (12). The cows had been bred by artificial insemination and the ages of the fetuses varied between 163 and 281 days. The duration of gestation is approximately 281 days in the Jersey cow. In the first 12 experiments the fetuses were fully delivered onto a warmed table. In subsequent experiments only the head (12 exp) or the foreleg (11 exp) was delivered from the uterus. After local anesthesia had been administered to the fetus a carotid artery or a brachial artery was exposed and intubated with polyethylene tubing. Fetal arterial pressure was recorded with a Sanborn 150 one-channel or an Elec- tronics for Medicine four-channel recorder. Heart rates were counted at I- to 2-min intervals from the pressure recording. Ten to thirty minutes after intubation of the fetal artery, and at a time when the fetal heart rate and arterial pressure were stable, simultaneous samples were drawn from catheters in the jugular vein (or systemic artery) of the mother and the artery of the fetus. Blood samples for hematologic study were placed in tubes containing sodium citrate. Arterial samples for blood gas analysis were drawn into heparinized glass syringes and placed on ice. The calves were then utilized for other experiments ( 12).
Weights of fetal calves and fetal organs. At the end of the ex- periment, the fetus was delivered, the cord tied, and the fetus weighed. After the last gasp, fetuses of 260 days or younger were dissected to obtain fetal organ weights. The skin, in- cluding the tail and the ears and excluding the hooves, was removed by sharp dissection and weighed while still wet. The two lungs were weighed together attached to a short portion of trachea. The heart was weighed unopened after the great vessels and pericardium had been removed. The kidneys were weighed after the fat was removed. The brain weight included the cerebral hemispheres, midbrain, cere- bellum, and the medulla oblongata to the level of the fora- men magnum. The weight of the “guts” included the stomach, intestines, bladder, and their contents, and the pancreas, mesenteric fat, adrenals, and ureters. A portion of the gastric contents was occasionally lost in the dissection. In 10 experiments the placenta was manually removed from each cotyledon. Although 2-4 hr were required to remove the placenta, removal was incomplete since a portion was retained within the depressions in the maternal cotyledons. The placenta weight (expressed as a percent of fetal weight) includes placenta, membranes, and a portion of the umbil- ical cord.

GROWTH OF FETAL CALF 241
Weights of the fetus: weights of fetal organs and placenta as percent of fetal weight TABLE 1.
Mean G~mmal
Range, days Fetal Wt, kg Carcass, % 1 Skin, % 1 Guts, % / Liver, % 1 Lungs, % / Brain, % 1 Heart, % Kidney, % Spleen, % Placenta, y.
178 5.4 zkz 1.6 64.0 (163-187) (4) (2) 207 9.6 =t 1.2 60.7 sf= 3.5 (200-216) (6) (5) 233 16.8 + 2.5 61.1 zt 3.4 (225-238) (7) (7) 252 18.6 =f= 2.1 63.6 zk 2.2 (242-260) (6) (5)
0.8 zt 0.1 1.0 It 0.1 0.4 rfr 0.1 (4) (4) (4)
0.8 + 0.1 0.8 zt 0.1 0.4 * 0.1 (6) (6) (6)
1.0 zt 0.1 0.6 + 0.1 0.2 * 0.1 (7) (7) (6)
0.9 zt 0.1 0.5 zt 0.1 0.2 zk 0.1 (5) (5) (5)
16.4 Az 2.8 (4)
11.7 * 1.9 (3)
9.7 zk 0.8 (3)
10.6 6.2 + 1.43.3 & 0.33.4 zt 0.41.5 zt 0.3 (2) (4) (4) (4) (3)
13.2 zt 0.67.1 zt 2.12.9 zt 0.22.6 =fi 0.31.3 zt 0.1 I (5) (6) (6) (6) (6) ‘16.3 + 2.37.2 A 1.32.4 zt 0.22.9 zt 0.41.0 & 0.1
(7) (7) (7) (7) (6) ~16.8 =t 3.46.5 If= 0.92.5 =t 0.22.6 + 0.5 1.0 =t 0.1 I I (5) (5) (5) (5) (5)
Values are means A 1 SD; number of observations is given in parentheses.
WEIGHT (Kg)
(
1’ r’
r’
I’ /
1’ /
I :
I
:
48 32 FETAL WEIGHT (g:)
46
42
24 -
38
20 - 34
30 16-
26 12 -
8-
18
4-
I . - t I , , , , , , , I I ,
‘0 40 80 120 160 200 240 280
6 FIG. 2. Relationship of fetal weighPI to gestational age of 56 fetal
calves (0) 80 days or older. Linear regression equation (-) indicated a slope of 0.135 and an intercept on ah&s a of 47 days and there was a high correlation coefficient (T = 0.98). Linear regression equation calculated by Huggett and Widdas (8) (---) from less complete data was similar. Linear regression equation for sheep (8) is shown (. l 0).
240(
200(
160(
l20(
80(
FETAL WE (0)
, c
P I
- COW(THIS REPORT W+=.,,,
--- COW(8) W’=.l45(T-50)
l *o=*o* SHEEP( 8) Wf=. ,47( T-37)
(T-47)
GESTATIONAL AGE (DAYS)
GESTATIONAL AGE (DAYS) T POST NATAL AGE (DAYS I I I I I I 1
160 200 240 280 20 40 60
FIG. 1. Relationship of weight of fetal calves (0) to their gesta- tional ages (80-281 days). Thirty-two fetuses are from this report and twenty-four were reported by other authors (7, 16). Curvilinear line drawn through the data was calculated from the equation, W1j3 = 0.135 (T-47), shown in Fig. 2. T is the gestational age in days. Mean weights (&SD) of 18 spontaneously born calves on day of birth and 30 calves at 2, 4, and 9 weeks of age are shown (0).
photometer and the hematocrits were done by the micro- method. The total erythrocyte and leucocyte counts were done manually using an American Optical Bright Line hemocytometer.
Newborn calves. Eighteen spontaneously born male Jersey calves were weighed approximately 12-24 hr after birth. Weights were obtained in 32 spontaneously born male Jersey calves at 2 weeks, 4 weeks, and 9 weeks of age. In 10 new- born calves aged 2 days and in 9 calves 12-24 days, blood samples were drawn for determination of hemoglobin con- centrations, hematocrits, and total and differential leuco- cyte counts.
ha&is of bZood samples. Fetal and maternal arterial blood samples for blood gas analysis were kept on ice for 2-3 hr and analyzed for POT, Pco~, and pH at 37 C using Radio- meter electrodes. The values were corrected for the tempera- ture of mother or fetus. Hemoglobin concentrations were determined colorome trically using a Coleman Jr. spec tro-
RESULTS
Weights of fetus and newborn calves (Table I). There was a rapid increase in the weight of the fetal calf (3.6-30 kg) with increasing gestational age (163-281 days) (Fig. 1). Weights have previously been reported in various breeds of fetal calves from age 1 to 7 months gestation (7, 16) and these have been included in Figs. 1 and 2 for comparison with the present data. The relationship of fetal weight to gestational age (Fig. 1) appeared to be curvilinear as has been reported for this and other species (1, 4, 7, 16). There was a linear relationship (r = 0.98) of fetal weight l/3 and gestational age (Fig. 2) for fetuses older than 2 months (~56 days). The re- gression equation had a slope (0.135) and a calculated inter-
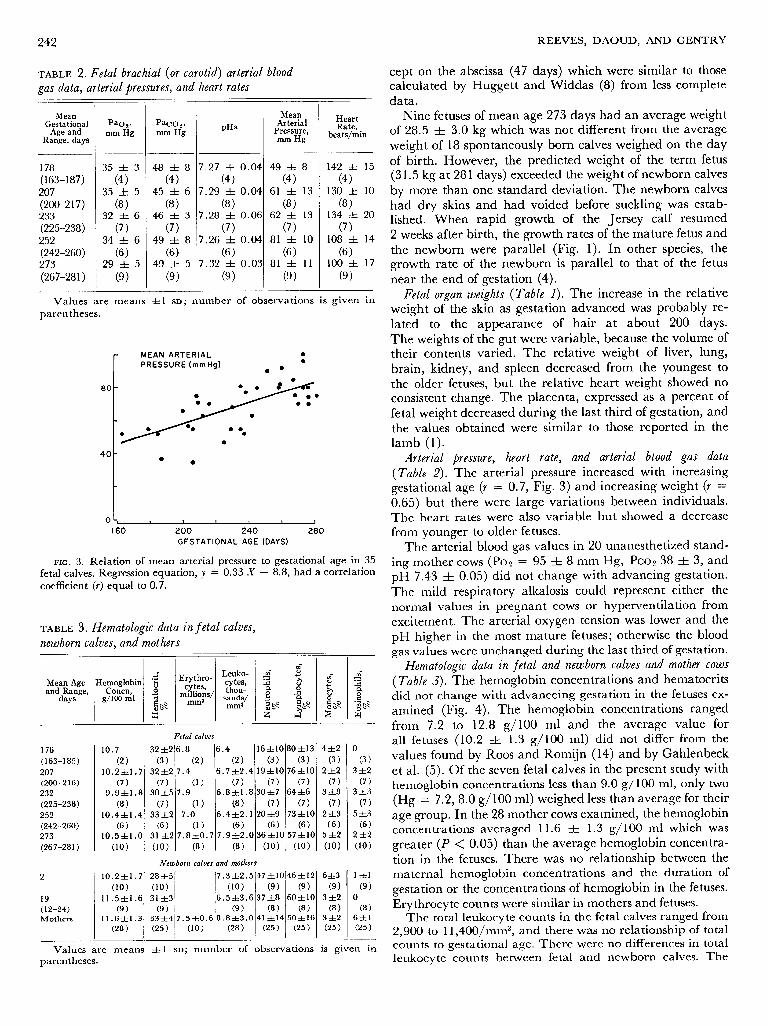
242 REEVES, DAOUD, AND GENTRY
TABLE 2. Fetal brachial (or carotid) arterial blood cept on the abscissa (47 days) which were similar to those gas data, arterial pressures, and heart rates calculated by Huggett and Widdas (8) from less complete .- data.
Mean Gr;es;;;al
Range, days
178 (163-187) 207 (200-2 17) 233 (225-238) 252 (242-260) 273 (267-281)
paop mm Hg
35 * 3 (4)
35 * 5 (8)
32 zk 6 (7)
34 zt 6 (6)
29 =t 5 (9)
pacop mm Hg
48 zk 8 (4)
45 zk 6 (8)
46 zt 3 (7)
49 zt 8 (6)
49 * 5 (9)
PHI
Mean Arterial
Pressure, mm Hg
7.27 + 0.04 49 zk 8 (4) (4)
7.29 + 0.04 61 zt 13 (8) 03)
7.28 zt 0.06 62 h 13 (7) (7)
7.26 A 0.04 81 zt 10 (6) (6)
7.32 zt 0.03 81 31 11 (9) (9)
Heart Rate,
beats/min
142 ztz 15 (4)
130 * 10 (8)
134 * 20 (7)
108 ZJZ 14 . (6)
100 zt 17 (9)
Values are means & 1 SD; number of observations is given in paren theses.
80
40
0
- MEAN ARTERIA PRESSURE (mm
0 0
0 0
160 200 240 280 GESTATIONAL AGE (DAYS)
FIG. 3. Relation of mean arterial pressure to gestational age in 35 fetal calves. Regression equation, y = 0.33 X - 8.8, had a correlation coefficient (r) equal to 0.7.
TABLE 3. Hematologic data newborn calves, and mothers
in f eta1 calves,
Mean Age Hemoglobin and Range, Concn,
days g/100 ml
176 10.7 32zt2 6.8 6.4 16=f=lO 80+13 4&2 0 (163-185) (2) (3) (2 > (2) (3) (3) (3) (3) 207 10.2zt1.7 32zt2 7.4 6.7zt2.4 19ztlO 76ztlO 2zt2 3zt2 (200-2 16) (7) (7) (1) (7) (7) (7) (7) (7) 232 9.9&l .a 30*5 7.9 6.8&l .8 30~1~7 64&6 3*3 3+3 (225-238) (8) (7) (1) (8) (7) (7) (7) (7) 252 10.4&l .4 33zt2 7.0 6.4zt2.1 2Ozt9 73ztlO 2=t3 5*3 (242-260) (6) (6) (1) (6) (6 ) (6) (6) (6) 273 10.5ztl .o 31 &2 7.8ztO.7 7.9zt2.0 36ztlO 57sf:lO 5+2 2*2 (267-281) (10) (10) (8 > (8) (10) (10) 00) (10)
Fetal calves
Newborn calves and mothers
2 10.2ztl.7 28zt5 7.3zt2.5 47ztlO 46&12 6&3 (10) (10) (10) (9) (9) (9)
19 11.5ztl.6 31~3 6.5zk3.6 37&8 6O=tlO 3~2 (12-24) (9) (9) (9) (8 > (8) (8) Mothers 11.6ztl.3 33&4 7.5ztO.6 8.8zt3.0 41 ztl4 50~16 3zt2
(28 > (25 1 (10) (28 > (25) (25) (25)
Values are means A=1 SD; number of observations is gi parentheses.
- V
lzlzl (9)
0 (8 >
63~1 (25 )
en in
Nine fetuses of mean age 273 days had an average weight of 28.5 + 3.0 kg which was not different from the average weight of 18 spontaneously born calves weighed on the day of birth. However, the predicted weight of the term fetus (3 1.5 kg at 28 1 days) exceeded the weight of newborn calves by more than one standard deviation. The newborn calves had dry skins and had voided before suckling was estab- lished. When rapid growth of the Jersey calf resumed 2 weeks after birth, the growth rates of the mature fetus and the newborn were parallel (Fig. 1). In other species, the growth rate of the newborn is parallel to that of the fetus near the end of gestation (4).
Fetal organ weights (Table 1). The increase in the relative weight of the skin as gestation advanced was probably re- lated to the appearance of hair at about 200 days. The weights of the gut were variable, because the volume of their contents varied. The relative weight of liver, lung, brain, kidney, and spleen decreased from the youngest to the older fetuses, but the relative heart weight showed no consistent change. The placenta, expressed as a percent of fetal weight decreased during the last third of gestation, and the values obtained were similar to those reported in the lamb (1).
Arterial pressure, heart rate, and arterial blood gas data (Table 2). The arterial pressure increased with increasing gestational age (r = 0.7, Fig. 3) and increasing weight (r = 0.65) but there were large variations between individuals. The heart rates were also variable but showed a decrease from younger to older fetuses.
The arterial blood gas values in 20 unanesthetized stand- ing mother cows (PO, = 95 & 8 mm Hg, PCO~ 38 + 3, and pH 7.43 & 0.05) did not change with advancing gestation. The mild respiratory alkalosis could represent either the normal values in pregnant cows or hyperventilation from excitement. The arterial oxygen tension was lower and the pH higher in the most mature fetuses; otherwise the blood gas values were unchanged during the last third of gestation.
Hematologic data in fetal and newborn calves and mother cows (Table 3). The hemoglobin concentrations and hematocrits did not change with advanceing gestation in the fetuses cx- amined (Fig. 4). The hemoglobin concentrations ranged from 7.2 to 12.8 g/ 100 ml and the average value for all fetuses (10.2 & 1.3 g/100 ml) did not differ from the values found by Roos and Romijn (14) and by Gahlenbeck et al. (5). Of the seven fetal calves in the present study with hemoglobin concentrations less than 9.0 g/ 100 ml, only two (Hg = 7.2, 8.0 g/100 m 1) weighed less than average for their age group. In the 28 mother cows examined, the hemoglobin concentrations averaged 11.6 & 1.3 g/ 100 ml which was greater (P < 0.05) than the average hemoglobin concentra- tion in the fetuses. There was no relationship between the ma ternal hemoglobin concentrations and the duration of gestation or the concentrations of hemoglobin in the fetuses. Erythrocyte counts were similar in mothers and fetuses.
The total leukocyte counts in the fetal calves ranged from 2,900 to 11,400/ mm3, and there was no relationship of total counts to gestational age. There were no differences in total leukocyte counts between fetal and newborn calves. The

GROWTH OF FETAL CALF
differential leukocyte counts showed the oldest fetuses had more neutrophils (36 & 10 %) than did the 23 younger fe- tuses examined (24 & 10 %). The newborn calves age 2 days had more neutrophils than did fetuses or did newborns at age 2 weeks. No data are available for leukocyte counts in fetal cows, but the counts in the newborn calves are within the normal range (15).
One mother cow had an elevated total leukocyte count of 18,00O/mm 3. However she was afebrile, there was no evi- dence of infection and she recovered well from the experi- ment. The other 27 mother cows had leukocyte counts of 4,500-14,000/ mm3 which were within the normal range (15). The differential leukocyte counts were all within the normal range (15).
DISCUSSION
When the fetal weights from previous reports (7,16) are combined with the present data, the growth of the bovine fetus can be estimated throughout gestation (Figs. 1 and 2). The previously reported fetal weights were from larger breeds of cattle than the Jersey, but the differences in fetal weight between breeds should be small early in gestation. The good agreement of the present data with that previously reported, and the similar growth rates of the fetus near term and of the young calf older than two weeks suggest that the data are reliable.
The regression equation relating W1’3 to gestational age had an earlier intercept on the abscissa for the fetal larnb (37 days (8)) than for the fetal calf (47 days) (Fig. 1). I f the in- tercept represents the approximate demarkation between ernbryonal and fetal growth rates (8) then the lamb enters upon the fetal growth phase a few days earlier than the calf. Thereafter the growth rates, i.e., the slopes of the regression line are approximately similar, but the calf has a longer gestation. The data in the calf agree with the general con- clusion of Huggett and Widdas “that when the birth weight of the young is increased the mammal does not grow its young quicker, along a steeper slope, but must grow its foetus a longer time” (8). The fetal calf was also similar to the lamb in that there was little growth of the placenta dur- ing the last third of gestation while the fetus was rapidly in- creasing in weight.
In both the lamb and the calf the rapid growth of the heart compared to the other organs probably reflected car- diac hypertrophy secondary to increasing arterial pressure. There were large variations in arterial pressure between calves of similar age or size. The data suggest that normal growth of the fetal calf may occur within rather wide limits of mean arterial pressure. It would be of interest to measure the placental flows relative to the observed arterial pressures. The increasing arterial pressure during rapid growth late in gestation is reported for several mammalian species but the average pressures achieved at term vary with the species (4). The fetal calf at term weighs more and has higher arterial pressures than do fetuses (4) of sheep, man, rhesus monkey, dog, cat, rabbit, or rat. For these species there appeared to be a linear relationship between the mean arterial pressure at term and the logarithm of the term fetal weight (Fig. 5). The data for the fetal calf lay along an extension of the line suggesting that the calf’s arterial pressure was at a level ex- netted for its large size.
HCT 40
E*
(%I 0
-
243
I1 I I I I I , , , 1, d , ,
160 200 240 280 20 4 GESTATIONAL AGE (DAYS)
FIG. 4. Relationship of hematocrit (lop) and hemoglobin (bottom) to gestational age in 33 fetal calves. Open circles (0) indicate mean values (&SD) for 10 calves aged 2 days, 9 calves of mean age 19 days, and 28 mothers (M).
loo- MEAN ARTERIAL _ PRESSURE
(mm Hg) 80- 0 cow
/ .SHEEP
60-
/ l MAN
/
.Rh. MONKEY 40-
. DOG - RABBIT. aCAT
20- / l RAT
- 0
0-’ I I I I I IO0 IO’ I02 IO3 IO4 IO5
FETAL WEIGHT AT TERM (gm)
FIG. 5. Relation of mean arterial pressure to logarithm of fetal weights at term in several species. Linear regression, y = 16.9 (log X) had a correlation coefficient (r) equal to 0.97.
In the present study, the values of arterial pH were lower than those reported for fetal calves (5), lambs (2, 3, 4, 1 l), and foals (3). It is possible that the surgical trauma to, and the manipulation of, mother and fetus altered the fetal state. However, the rather high arterial oxygen tensions, the ab- sence of hypercapnia, and of tachycardia suggested the fe- tuses were otherwise in good condition. The fetal oxygen tensions were higher than those reported by Gahlenbeck et al. (5), a finding which probably reflected a higher maternal oxygen tension in the present study (95 vs. 70 mm Hg).
In the fetal calf the arterial oxygen tensions throughout the last third of gestation were higher than those in fetal lambs studied under either acute (4) or chronic experimen- tal conditions (2, 3, 11). Rather the arterial oxygen tensions resembled those reported in the foal (3). Comline and Silver (3) suggested that the fetal arterial Paz was higher in the foal than in the lamb primarily because there was a smaller pressure gradient for oxygen across the placenta from mother to fetus (lamb 17 mm Hg, foal 4 mm Hg). It is pos- sible that the fetal calf, like the foal, has a relatively “em- cient” transfer of oxygen across the placental membranes.
Increases in hemoglobin concentrations that have been reported in mammalian fetuses approaching term (humans (1, 4), goats (9), lambs (1, 4), rabbits (18), rats (18), cats (18) and dogs (18)) are considered to reflect the stimulus of the hvpoxic fetal environment on the hemopoietic system

244 REEVES, DAOUD, AND GENTRY
(1, 4). Indeed hemoglobin concentration increases in the fe-
phatic system ( 15). However, even this interpretation may
tal lamb when the ewe is exposed for a few days to simulated high altitude (10). Further, relief of the fetal hypoxia after
not be correct, because the differential lymphocyte count
birth may account for the fall in hemoglobin concentration in the neonate ( 1, 4). Yet in the last third of gestation the hemoglobin concentration in the calf was lower than that in the lamb (1, 2, 4) and did not rise. Nor did it fall in the new- born period. Reticulocyte counts in newborn calves are low (< 1.4 %) (15). It is difficult to attribute the absence of a rise in the fetal calf’s hemoglobin concentration to immatu- rity of the hemopoietic system. Hemoglobin production was adequate to achieve and maintain a concentration of 10 g/ 100 ml during the period of rapid growth. Erythrocytes of the newborn calf, though small and numerous, do not show signs of immaturity (15). The low lymphocyte count in the newborn calf has been attributed to immaturity of the lym-
or absent hemopoietic response to chronic hypoxia has been observed in young calves made chronically hypoxemic by a surgical right-to-left shunt ( 13) and in healthy steers taken for months to high altitude (6, 17). Possibly, the fetal calf, the young calf, and the mature bovine all have a diminished hematopoietic response to chronic hypoxemia. However, Meschia et al. (11) and Comline and Silver (2) have sug- gested that under normal conditions hemoglobin concen- tration in the fetal lamb may not increase near term. Serial observations utilizing indwelling catheters (2, 11) are needed to evaluate the role of chronic hypoxia as a hematopoietic stimulus to the healthy bovine fetus.
Mrs. Joyce Hutchinson and Mrs. Janet Mobley provided valuable
s”gge~tlons~
technical assistance. Dr. Charles Eastin performed the surgery until December, 1970. The authors thank Dr. G. S. Dawes for his helpful
This study was supported by National Institutes of Health Grants HE-06780 and HE-08932.
was particularly high in the fetus and only temporarily de- creased to a minimum in the early newborn period. A poor Received for publication 13 September 197 1.
REFERENCES
1. BARCROFT, J. Researches on B-e-Natal Life. Oxford : Blackwell, 1946. 2. COMLINE, R. S., AND M. SILVER. Daily changes in foetal and
maternal blood of conscious pregnant ewes with catheters in umbilical and uterine vessels. J. Physiol., London 209: 567-586, 1970.
3. COMLINE, R. S., AND M. SILVER. PO%, PCOZ, and pH levels in the umbilical and uterine blood of the mare and ewe. J. Physiol., London 209 : 587-608, 1970.
4. DAWES, G. S. Foetal and Neonatal Physiology. Chicago, Ill. : Year Book, 1968.
5. GAHLENBECK, H., H. FRERKING, A. M. RATH SCHLAG-SCHAEFER, AND H. BARTELS. Oxygen and carbon dioxide exchange across the cow placenta during the second part of pregnancy. Respiration Physiol. 4: 119-131, 1968.
6. GROVER, R. F., J. T. REEVES, D. H. WILL, AND S. G. BLOUNT, JR. Pulmonary vasoconstriction in steers at high altitude. J. Appk Physiol. 18: 567-574, 1963.
7. HAMMOND, J. The Physiology of Reproduction in the Cow. Cambridge: Cambridge Univ. Press, 1927.
8. HUGGETT, A. ST. G., AND W. F. WIDDAS. The relationship between mammalian foetal weight and conception age. J. Physiol., London 114: 306-317, 1957.
9. KAISER, I. H., AND J. M. CUMMINGS. pH, carbon dioxide, oxygen, hemoglobin, and plasma electrolytes in blood of pregnant goats and their fetuses. Am. J. Physiol. 195 : 481-486, 1958.
10. KAISER, I. H., J. H. CUMMINGS, S. R. M. REYNOLDS, AND J. P.
MARBARGER. Acclimatization response of the pregnant ewe and fetal lamb to diminished ambient pressure. J. A/$. Physiol. 13: 171-178, 1958.
11. MESCHIA, G., J. R. COTTER, C. S. BREATHNACH, AND D. H. BARRON. The hemoglobin, oxygen, carbon dioxide, and hydrogen ion concentrations in the umbilical bloods of sheep and goats as sampled via indwelling plastic catheters. Quart. J. Exptl. Physiol 50: 185-195, 1965.
12. REEVES, J. T., F. DAOUD, AND C. EASTIN. Effects of vagotomy on arterial pressure and blood gases in the fetal calf. Am. J. Physiol. 221; 349-355, 1971.
13. REEVES, J. T., J. E. LEATHERS, B. EISEMAN, AND F. C. SPENCER. Alveolar hypoxia versus hypoxemia in the development of pul- monary hypertension. Med. Thoracalis 19: 56 l-572, 1962.
14. Roes, J., AND C. ROMIJN. Some conditions of foetal respiration in the cow. J. Physiol., London 92: 219-267, 1938.
15. SCHALM, 0. W. Veterinary Hematology. Philadelphia, Pa. : Lea & Febiger, 196 1.
16. SWEET, W. N., C. A. MATHEWS, AND M. H. FOHRMAN. US Dept. Agr. Tech. Bull. 964 : l-34, 1948.
17. WILL, D. H., A. F. ALEXANDER, J. T. REEVES, AND R. F. GROVER. High altitude induced pulmonary hypertension in normal cattle. Circulation Res. 10 : 172-l 77, 1962.
18. WINTROBE, M. M., AND H. B. SHUMACKER, JR. Erythrocyte studies in the mammalian fetus and newborn. Am. J. Anat. 58: 313-328, 1936.


















