
Growth Media and Economical Impact Plasmids
8
490 JOURNAL OF BIOSCIENCE AND BIOENGINEERING © 2007, The Society for Biotechnology, Japan Vol. 104, No. 6, 490–497. 2007 DOI: 10.1263/jbb.104.490 Growth Medium Selection and Its Economic Impact on Plasmid DNA Production Michael K. Danquah 1 * and Gareth M. Forde 1 Bioengineering Laboratory, Department of Chemical Engineering, Monash University, Clayton campus, Wellington Rd., Victoria 3800, Australia 1 Received 22 January 2007/Accepted 27 September 2007 Current developments in gene medicine and vaccination studies are utilizing plasmid DNA (pDNA) as the vector. For this reason, there has been an increasing trend towards larger and larger doses of pDNA utilized in human trials: from 100–1000 µg in 2002 to 500–5000 µg in 2005. The increasing demand of pDNA has created the need to revolutionalize current production levels under optimum economy. In this work, different standard media (LB, TB and SOC) for culturing recombinant Escherichia coli DH5α harbouring pUC19 were compared to a medium optimised for pDNA production. Lab scale fermentations using the standard media showed that the highest pDNA volumetric and specific yields were for TB (11.4 µg/ml and 6.3 µg/mg dry cell mass respec- tively) and the lowest was for LB (2.8 µg/ml and 3.3 µg/mg dry cell mass respectively). A fourth medium, PDMR, designed by modifying a stoichiometrically-formulated medium with an optimised carbon source concentration and carbon to nitrogen ratio displayed pDNA volumetric and spe- cific yields of 23.8 µg/ml and 11.2 µg/mg dry cell mass respectively. However, it is the economic ad- vantages of the optimised medium that makes it so attractive. Keeping all variables constant except medium and using LB as a base scenario (100 medium cost [MC] units/mg pDNA), the optimised PDMR medium yielded pDNA at a cost of only 27 MC units/mg pDNA. These results show that greater amounts of pDNA can be obtained more economically with minimal extra effort simply by using a medium optimised for pDNA production. [Key words: plasmid DNA, cultivation medium, fermentation] Recombinant DNA technology and the sequencing of the human genome have led to revolutionary discoveries espe- cially in the fields of gene therapy and nucleic acid vaccines (1, 2). Plasmid DNA has acquired appreciable interest due to its attractive potential application in gene therapy and DNA vaccines applications (3). Gene therapy processes in- volve the introduction of one or more functional and spe- cific genes into a human recipient to repair certain genetic defects and aberrations. A pDNA vaccine can be developed from a pathogen’s genes to provide immunity against dis- eases (4, 5). Plasmid DNA vaccines allow the foreign genes to be expressed transiently in transfected cells, mimicking intracellular pathogenic infection and inducing humoral and cellular immune responses (6, 7). Plasmid DNA vaccines offer a new opportunity to immunize with materials that are mainly gene-based and are expressed by the cells of the re- cipient. This gives greater control over the entire immuniza- tion process (2). Considerable attention has been given to the potential of pDNA vaccines to mitigate and prevent a number of infec- tions but substantially less examination has been given to the practical challenges of producing large quantities of pDNA for therapeutic use in humans, for both clinical studies and ultimately full-scale production under optimum economy. Plasmid DNA is produced from a fermentation process and the success of this fermentation process hinges on the inter- actions between the host organism harbouring the recombi- nant plasmid vector and the growth environment (8). A major advantage of fermentation is that conditions that influence cell growth, plasmid yield, quality and stability can be ex- amined and controlled. These include media composition, temperature, pH, dissolved oxygen and build-up of waste metabolites. Plasmid DNA vaccine doses are increasing more than the amounts currently produced by fermentations (2). The cultivation medium formulation dramatically affects the performance and nature of microbial processes. This im- plies that, the type and source of nutrients available in the growth medium can have a profound effect on the fermenta- tion yields (1, 9). Media containing yeast extract and hy- drolysed protein are often used because they are relatively simple to prepare and generally lead to high cell densities. Meat extracts are also rich sources of nutrients for fermenta- tion, but there is the risk of contamination with animal viruses (1). In addition to complex nutrients glucose, glycerol or other sugars are included as a source of carbon and energy. Nitrogen is usually provided in inorganic form such as am- monium salts (9). Trace metals and vitamins may also con- tribute to cell growth and plasmid yield. Defined media pre- * Corresponding author. e-mail: [email protected] phone: +61-399053440 fax: +61-399055686
-
Upload
alex-issamu-kanno -
Category
Documents
-
view
30 -
download
1
Transcript of Growth Media and Economical Impact Plasmids

490
JOURNAL OF BIOSCIENCE AND BIOENGINEERING © 2007, The Society for Biotechnology, Japan
Vol. 104, No. 6, 490–497. 2007
DOI: 10.1263/jbb.104.490
Growth Medium Selection and Its Economic Impacton Plasmid DNA Production
Michael K. Danquah1* and Gareth M. Forde1
Bioengineering Laboratory, Department of Chemical Engineering, Monash University,Clayton campus, Wellington Rd., Victoria 3800, Australia1
Received 22 January 2007/Accepted 27 September 2007
Current developments in gene medicine and vaccination studies are utilizing plasmid DNA(pDNA) as the vector. For this reason, there has been an increasing trend towards larger andlarger doses of pDNA utilized in human trials: from 100–1000 µg in 2002 to 500–5000 µg in 2005.The increasing demand of pDNA has created the need to revolutionalize current production levelsunder optimum economy. In this work, different standard media (LB, TB and SOC) for culturingrecombinant Escherichia coli DH5α harbouring pUC19 were compared to a medium optimisedfor pDNA production. Lab scale fermentations using the standard media showed that the highestpDNA volumetric and specific yields were for TB (11.4 µg/ml and 6.3 µg/mg dry cell mass respec-tively) and the lowest was for LB (2.8 µg/ml and 3.3 µg/mg dry cell mass respectively). A fourthmedium, PDMR, designed by modifying a stoichiometrically-formulated medium with an optimisedcarbon source concentration and carbon to nitrogen ratio displayed pDNA volumetric and spe-cific yields of 23.8 µg/ml and 11.2 µg/mg dry cell mass respectively. However, it is the economic ad-vantages of the optimised medium that makes it so attractive. Keeping all variables constant exceptmedium and using LB as a base scenario (100 medium cost [MC] units/mg pDNA), the optimisedPDMR medium yielded pDNA at a cost of only 27 MC units/mg pDNA. These results show thatgreater amounts of pDNA can be obtained more economically with minimal extra effort simply byusing a medium optimised for pDNA production.
[Key words: plasmid DNA, cultivation medium, fermentation]
Recombinant DNA technology and the sequencing of thehuman genome have led to revolutionary discoveries espe-cially in the fields of gene therapy and nucleic acid vaccines(1, 2). Plasmid DNA has acquired appreciable interest dueto its attractive potential application in gene therapy andDNA vaccines applications (3). Gene therapy processes in-volve the introduction of one or more functional and spe-cific genes into a human recipient to repair certain geneticdefects and aberrations. A pDNA vaccine can be developedfrom a pathogen’s genes to provide immunity against dis-eases (4, 5). Plasmid DNA vaccines allow the foreign genesto be expressed transiently in transfected cells, mimickingintracellular pathogenic infection and inducing humoral andcellular immune responses (6, 7). Plasmid DNA vaccinesoffer a new opportunity to immunize with materials that aremainly gene-based and are expressed by the cells of the re-cipient. This gives greater control over the entire immuniza-tion process (2).
Considerable attention has been given to the potential ofpDNA vaccines to mitigate and prevent a number of infec-tions but substantially less examination has been given to thepractical challenges of producing large quantities of pDNAfor therapeutic use in humans, for both clinical studies and
ultimately full-scale production under optimum economy.Plasmid DNA is produced from a fermentation process andthe success of this fermentation process hinges on the inter-actions between the host organism harbouring the recombi-nant plasmid vector and the growth environment (8). A majoradvantage of fermentation is that conditions that influencecell growth, plasmid yield, quality and stability can be ex-amined and controlled. These include media composition,temperature, pH, dissolved oxygen and build-up of wastemetabolites. Plasmid DNA vaccine doses are increasingmore than the amounts currently produced by fermentations(2).
The cultivation medium formulation dramatically affectsthe performance and nature of microbial processes. This im-plies that, the type and source of nutrients available in thegrowth medium can have a profound effect on the fermenta-tion yields (1, 9). Media containing yeast extract and hy-drolysed protein are often used because they are relativelysimple to prepare and generally lead to high cell densities.Meat extracts are also rich sources of nutrients for fermenta-tion, but there is the risk of contamination with animal viruses(1). In addition to complex nutrients glucose, glycerol orother sugars are included as a source of carbon and energy.Nitrogen is usually provided in inorganic form such as am-monium salts (9). Trace metals and vitamins may also con-tribute to cell growth and plasmid yield. Defined media pre-
* Corresponding author. e-mail: [email protected]: +61-399053440 fax: +61-399055686

MEDIUM SELECTION AND ECONOMICS FOR pDNA PRODUCTIONVOL. 104, 2007 491
pared from purified components instead of complex bio-logical mixtures may be more reproducible since differentpreparations of complex substrates may vary somewhat incomposition of vital elements. However, defined media usu-ally require more components, are more difficult to prepare,and often lead to lower cell densities than can be achievedwith complex media (1). Hence there is a challenging multi-variable trade-off or decision to be made regarding growthof biomass, plasmid yield, quality, stability and cost in theselection of medium for pDNA production.
Escherichia coli is a micro-organism that grows in bothrich complex organic media and salt-based chemically de-fined media as long as carbon source is present (2). Besidesan energy source, it requires nutrients for the biosynthesis ofcellular matter, formation of products and maintenance sothe content of the medium must supply the nutrients neededto accomplish this. The vital chemical elements needed forthe cultivation of E. coli are hydrogen, carbon, nitrogen, oxy-gen, sodium, magnesium, phosphorus, potassium and cal-cium. These have specific functions during the bioactivityof cell growth and plasmid yield. Physiologically, hydrogenand oxygen form the basis of cellular water and with carbonas the main constituent of organic cell materials. Sodium,potassium, magnesium and calcium are cellular cations andcofactors for some enzymes. Phosphorous constitutes phos-pholipids, coenzymes and nucleotides in nucleic acids (9,10). The type and concentration of these elemental ingredi-ents used in cultivation medium determines the amount ofbiomass produced, as well as plasmid volumetric yield andspecific yield. It is also likely that medium composition willdirectly bear on the physiology of the microorganisms byinfluencing their intricate regulatory systems, and thereforewill control plasmid copy number (2). Plasmids DNA forgene therapy or vaccination harbouring genes are very largemolecules in comparison to proteins. Knowledge about cul-ture media and conditions for the fermentation of recombi-nant E. coli has been obtained on the basis of studies, whichoptimised the expression of proteins (11–13). However,nutrient conditions for optimisation of pDNA production inE. coli could be significantly different from those of proteinproduction. Replication is the only process required whenpDNA is the final product. Transcription and translation asin the case of proteins are generally undesirable duringpDNA production in bacterial cells (4). Media suitable forE. coli can be purchased or their formulations can be ob-tained from the literature. Compositions of complex and de-fined media are extensively discussed in many publications(14–17). Due to the simplicity of use associated with off-the-shelf cultivation media, a number of laboratory-scalepDNA vaccine production schemes rely on the use of un-optimised small-scale processes employing commerciallyavailable complex formulations (18, 19).
Although many works on pDNA production in E. coli byfermentation are reported using different media, reports onthe experimental examination of comparative effect of dif-ferent media on cell growth and pDNA yield kinetics aswell as economic consideration of medium formulation arelimited. This limitation makes cultivation medium selectionmore of a trial and error process without any medium costper pDNA yield analysis based on previous fermentation
works in the literature. This constitutes generally to the lowpDNA yield that is encountered even for expensive mediumformulations and high copy number plasmids. Studies onpDNA yield kinetics of a particular medium dictates whenfermentation can be halted for maximum yield if it is em-ployed rather than the normal random stoppage which con-stantly produces small amounts of pDNA. This work isgeared toward investigating the effects of different culturemedia on biomass growth as well as yield and purity ofpDNA. An economic comparison between different media(cost of medium per mg of pDNA produced), generally notconsidered in the literature to date, is presented here. Eco-nomic modification of stoichiometric medium and improve-ment of standard and stoichiometric media performances oncell growth and plasmid yield based on glucose addition andoptimisation of carbon to nitrogen ratio and cost analysisare also reported. Economic evaluation of growth mediumfor large scale pDNA production is discussed.
MATERIALS AND METHODS
Materials Maximum efficiency E. coli DH5α (EndA–), pUC19plasmid (0.01 µg/l) were purchased from Invitrogen (Victoria,Australia), tryptone (Difco, NJ, USA), yeast extract (Difco), NH
4Cl
(Sigma-Aldrich, NSW, Australia), K2HPO
4 (Merck, Whitehouse
Station, NJ, USA), ethidium bromide (Sigma-Aldrich), NaOH(Sigma-Aldrich), KH
2PO4 (Merck), HCl (Sigma-Aldrich), glycerol
(Amresco, OH, USA), NaCl (Amresco), KCl (Sigma-Aldrich), MgCl2
(Univar, WA, USA), MgSO4 (Sigma-Aldrich), glucose (Merck),
propylene glycol (Sigma-Aldrich) and Bradford reagent (Sigma-Aldrich).
Media Preparation Compositions of LB, TB and SOC mediaused for shake flask fermentation are as shown in Table 1. Wang etal. (4) reported the rational design of a defined medium optimisedfor pDNA production by E. coli strain JM109 harbouring thepcDNA3S plasmid using a stoichiometric approach. They consid-ered the effect of glucose, amino acid and nucleosides concentra-tions on cell growth and pDNA yields. Maximum growth yield co-efficient of glucose in the synthesis of cell biomass θglc was obtainedas 0.40 g DCW/g glucose. After identifying six key amino acids(Asp, Glu, Gly, His, Leu, Try) to add to a glucose-basal saltmedium, the designed formulation supported higher pDNA volu-metric and specific yields when compared with those routinelyachieved in the complex LB medium. When the defined mediumwas supplemented with nucleosides (adenosine, guanosine, cytidine,and thymidine), the pDNA volumetric and specific yields werefurther enhanced. The final concentration of nutrients in the me-dium (MW1) obtained from the stoichiometric model were 1.240 g/lasparate, 1.152 g/l glutamine, 0.378 g/l glycine, 0.111 g/l histidine,0.693 g/l leucine, 0.03 g/l tryptophan, 0.096 g/l adenosine, 0.099 g/lguanosine, 0.146 g/l cytidine, 0.156 g/l thymidine, 10 g/l glucose,12.8 g/l Na
2HPO
4⋅7H
2O, 3 g/l KH
2PO
4, 0.5 g/l NH
4Cl, 0.24 g/l
MgSO4, and 0.004 g/l thiamine. This medium gave a high plasmid
yield of 60 mg/l in batch fermentation of E. coli JM109 pcDDA3S.
TABLE 1. Composition of LB, TB and SOC media preparedat room temperature
Medium Composition (l–1)
LB 10 g tryptone, 5 g yeast extract, 5 g NaClTB 12 g tryptone, 24 g yeast extract, 9.4 g K
2HPO
4,
2.2 g KH2PO
4, 4 ml glycerol
SOC 20 g tryptone, 5 g yeast extract, 0.6 g NaCl, 0.2 g KCl, 0.95 g MgCl
2, 1.2 g MgSO
4, 3.6 g glucose

DANQUAH AND FORDE J. BIOSCI. BIOENG.,492
Following a critical economic consideration of medium MW1 inboth analytical and large scale production of pDNA on the groundsof high cost of amino acids, nucleosides and thiamine, this paperreports the formulation of an optimised and economically viablemedium PDM (plasmid DNA medium) for pDNA production viafermentation. Yeast extract and tryptone were identified to containthe required amino acids (Asp, Glu, Gly, His, Leu, Try), nucleo-sides (adenosine, guanosine, cytidine and thymidine) and thiaminefor cell growth and pDNA yield as in medium MW1 whilst beingeconomically preferable. Tryptone contains approximately 35%amino acids (Asp, Glu, Gly, His, Leu, Try), and 0.33 µg/g thiamine.Yeast extract contains approximately 19% amino acids (Asp, Glu,Gly, His, Leu, Try) and 3.2 µg/gm thiamine. Tryptone and yeastextract contain 0.75% and 0.29% respectively of phosphorus whichsupport the building of nucleotides (Difco & BBL manual, 2003).Calculated amounts of yeast extract and tryptone were used formedium PDM (Table 2) based on the aforementioned compositionsto make up the content of amino acid, nucleoside and thiamine asfor medium MW1.
Cell transformation Maximum efficiency E. coli, DH5α(EndA–), cells were transformed with pUC19 plasmid as per themanufacturer’s instructions (Invitrogen). Cell lines were created(LB-glycerol), frozen in a dry ice/ethanol bath for 5 min thenstored at −75°C (Nuaire, Tokyo) until further use.
Shake flask fermentation process A single bacterial colonycarrying the pUC19 plasmid (E. coli DH5α-pUC19) was pickedfrom the LB-agar-ampicillin plate and subcultured with 100 ml ofLB culture containing an appropriate amount of ampicillin (100µg/ml) at 37°C overnight under 200 rpm shaking. Subsequently,2 ml of the culture was inoculated into each of 200 ml LB, TB,SOC and PDM containing 100 µg/ml of ampicillin. Fermentationwas allowed overnight at 37°C and 200 rpm shaking. Fermentationprocess was halted after 15 h.
Fermentation monitoring Two milliliter sample of each cul-ture was pipetted aseptically after every 30 min and part was usedfor optical density determination (OD
600) using a spectrophotome-
ter (Shimadzu, Kyoto). The remaining was spun in a centrifuge(Heraeus, Victoria, Australia) at 10,000×g for 5 min at 4°C. Thesupernant liquid was gently decanted and the cell resuspended inPBS buffer and stored in −75°C freezer. Cells were later lysed, pu-rified for pDNA and analysed for quantity and purity.
Batch fermentation process One single bacterial colony car-rying the plasmid was picked and subcultured with 1 l of LB cul-ture containing an appropriate amount of ampicillin at 37°C over-night under 200 rpm shaking. Subsequently, 500 ml of the culturewas inoculated into a 20 l fermentor (New Brunswick Scientific,Edison, NJ, USA) vessel containing 15 l of PDMR medium (PDMwith optimum C/N ratio) and 100 µg/ml of ampicillin. The temper-ature was set at 37°C and the dissolved oxygen (DO) value wasmaintained at 30% by the proportional integral derivative (PID)controller, which changed the speed of agitation to maintain the setDO value. The pH was maintained at 7.0 by the addition of 4 MNaOH and 1 M HCl. The inflow sparge air at a flow rate of 7.2slpm was set at 20 psia and foaming was checked by using poly-
propylene glycol as antifoam. Culture sampling was performed afterevery 30 min to monitor biomass growth as well as pDNA yieldkinetics. The cultivation was terminated 15 h after inoculation offermentor. The culture broth was harvested and then concentratedby ultrafiltration. The obtained cell paste was packaged and storedat −75°C prior to lysis.
Dry cell concentration determination Ten milliliter aliquotsof cell culture with known OD
600 were spun at 3700×g in a centri-
fuge for 10 min at 4°C. Supernant was decanted and the cell pelletswere washed with equal volume of sterile water. Cell were trans-ferred to a pre-weighed glass plate and dried over night to a con-stant weight at 105°C.
Cell lysis and plasmid DNA purification Plasmid DNApurification from bacterial cell was performed with Wizard PlusSV Minipreps (Promega) according to the manufacturer’s instruc-tions. Briefly, cells were alkaline lysed, clarified, plasmid was col-umn purified and eluted prior to quantification.
Plasmid DNA analysis Quantity and purity of pDNA sam-ples from LB, TB, SOC and PDM media were determined fromoptical density analysis, OD
260 and OD
280. Nature and size of plas-
mid DNA were determined by ethidium bromide agarose gel elec-trophoresis using a 1 kbp DNA ladder. Gel was made up in ×50dilution of TAE buffer (242 g of Tris base, 57.1 ml acetic acid,9.305 g of EDTA), stained with 3 µg/ml EtBr and run at 66 V for2 h. Gel was scanned with gel analyser (Bio-Rad, Segrate, Italy).Protein quantification analysis was performed using Bradford assayaccording to the manufacturer’s instructions.
RESULTS AND DISCUSSION
Transformation efficiency A transformation efficiencyof 5×108 CFU/µg pUC19 obtained shows that appreciableamount of bacterial cells (E. coli DH5α) were transformedwith pUC19 and this reveals favourable cell viability.
Biomass growth kinetics: shake flask fermentationOptical density values at 600 nm measured for samplesfrom each of the bacterial culture, LB, TB, SOC and PDMat 30 min interval were converted to dry cell concentration(mg/ml). Different biomass growth profiles of E. coli DH5αpUC19 were portrayed by the different media (Fig. 1A).The growth kinetics of the cells in the different media wasabout the same in the first 3 h and this similarity continuedfor TB, SOC and PDM up to the 5th hour. Cell growth inLB medium started stabilizing after 10 h reaching a finalcell concentration of 0.85 mg/ml. Significant variation ex-isted in the cell densities for the different media after 10 hand persisted until fermentation was halted after 15 h. Thisvariation is attributed to the difference in the amount of car-bon source present in the different media since biomassgrowth is boosted mainly by the presence of carbon source.The slow biomass growth in SOC after 9 h is due to theexhaustion of its glucose (carbon source) content. The finaldry cell concentrations for PDM and TB were 1.94 mg/mland 1.80 mg/ml respectively with the expectation of furtherincrement. There is some level of similarity in PDM and TBmedia support for biomass growth considering their growthprofiles and final cell optical densities. Both TB and PDMattained a maximum growth rate of 0.22 h–1.
Plasmid DNA yield kinetics: shake flask fermentationThe concentration of pDNA was analysed spectrophoto-metrically via absorbance at 260 nm. The optical density of1.0 measured at 260 nm with light path of 1 cm represents
TABLE 2. Composition of medium PDM preparedat room temperature
Composition Amount (g/l)
Yeast extract 4.41Tryptone 7.93Glucose 10.00Na
2HPO
4⋅7H
2O 12.80
KH2PO
43.00
NH4Cl 0.50
MgSO4
0.24

MEDIUM SELECTION AND ECONOMICS FOR pDNA PRODUCTIONVOL. 104, 2007 493
50 mg/l of double stranded DNA. Analysis of samples ofpDNA purified from LB, TB, SOC and PDM bacterial cellcultures showed different plasmid yield kinetics for the dif-ferent media (Fig. 1B). After 3 h of near negligible pDNAproduction, the volumetric yield of pDNA increased overthe final 12 h reaching 2.8 µg/ml, 11.4 µg/ml, 6.4 µg/ml and14.6 µg/ml for LB, TB, SOC and PDM media respectively.
There is a clear distinction between the pDNA volumetricyield characteristics of PDM and TB media; this may beattributed to the stoichiometrically designed PDM contain-ing optimised amino acid, nucleoside and thiamine concen-trations for pDNA production. Figure 1C shows a similartrend for the specific yield of pDNA for the different media.There exists some level of similarity in the progression ofpDNA specific yield for the LB, TB and SOC, most notablya kink in the specific yield profile between 4–6 h. This phe-nomenon is ascribed to the fact that in the exponential phaseof a batch profile, growth rate of biomass is maximal and itovertakes the replication rate of pDNA thereby causing adecrease in specific yield. This scenario was not observed inPDM profile as the rate of pDNA volumetric yield matchesthe increase in biomass during the exponential growth phase.
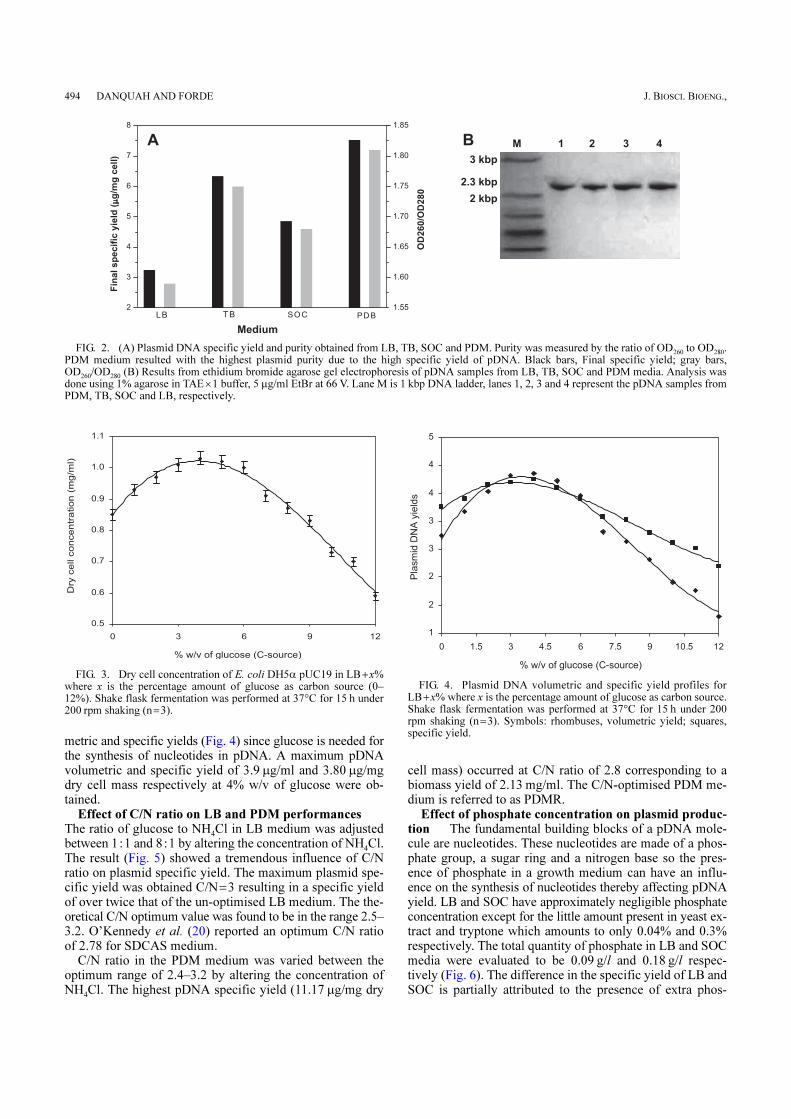
Absorption measurements taken at wavelengths of 260 nmand 280 nm were used to determine the purity of doublestranded DNA based on the ratio OD260/OD280 which is ex-pected to be within 1.7–1.9 to indicate pure double strandedDNA. A low OD260/OD280 ratio indicates protein contam-ination whilst a high OD260/OD280 shows possible RNAor residual organics contamination. The final purity of DNAfrom the PDM medium was the highest and that from theLB was the lowest (Fig. 2A). Ethidium bromide agarose gelelectrophoresis (Fig. 2B) shows no RNA or gDNA contami-nation and the purification protocol ensures that organicsare removed from the system before elution; hence the highOD260/OD280 ratio for PDM indicates the presence ofsupercoiled pDNA. Protein quantification analysis usingBradford assay shows protein concentration of ~0.4%, whichis keeping with the regulatory standard of <1%. The highestspecific yield of plasmid obtained from the PDM mediumhas an advantageous effect on plasmid purity since the quan-tity of impurities existing with the plasmid per unit mass ofcell is minimal.
Effect of glucose on LB performance LB performancewas modified by the gradual addition of glucose (LB+x%where x is the % w/v of glucose as carbon source) and sub-jected to the same condition of shake flask fermentation ofE. coli DH5α-pUC19. The growth of biomass was greatlyinfluenced by the right proportion of carbon source. Muchof the carbon source taken up by E. coli enters the pathwaysof energy yielding (ATP) metabolism and is eventually se-creted from the cell as CO
2 (the major product of energy
yielding metabolism). According to Fig. 3, the maximumdry cell concentration (1.03 mg/ml) was obtained at a glu-cose concentration of 4% w/v. The decrease in dry cell den-sity at glucose concentrations above 4% w/v is as a result ofthe excess glucose concentration in the growth medium whichposses an extra metabolic burden to the cells, thereby re-tarding their proliferation rate and hence the dry cell con-centration.
The addition of glucose also boosted both pDNA volu-
FIG. 1. (A) Dry cell concentration profiles obtained from OD600
measurements for the growth of E. coli DH5α pUC19 in LB, TB, SOCand PDM media in shake flask fermentation at 37°C for 15 h under200 rpm shaking. Samples were collected at 30 min intervals (n=3).(B) Volumetric yield characteristic of pDNA purified from LB, TB,SOC and PDM media. Samples were collected at 30 min intervals pu-rified and analysed spectrophometrically at OD
260 under room temper-
ature. 1 OD260
unit = 50 µg/ml double stranded DNA (n=3). (C) Spe-cific yield characteristic of pDNA purified from LB, TB, SOC andPDM media. Samples were collected at 30 min intervals. Specific yieldis the ratio of the volumetric yield to the cell concentration. Symbols:rhombuses, LB; squares, TB; triangles, SOC; crosses, PDM.
DANQUAH AND FORDE J. BIOSCI. BIOENG.,494
metric and specific yields (Fig. 4) since glucose is needed forthe synthesis of nucleotides in pDNA. A maximum pDNAvolumetric and specific yield of 3.9 µg/ml and 3.80 µg/mgdry cell mass respectively at 4% w/v of glucose were ob-tained.
Effect of C/N ratio on LB and PDM performancesThe ratio of glucose to NH
4Cl in LB medium was adjusted
between 1:1 and 8:1 by altering the concentration of NH4Cl.
The result (Fig. 5) showed a tremendous influence of C/Nratio on plasmid specific yield. The maximum plasmid spe-cific yield was obtained C/N=3 resulting in a specific yieldof over twice that of the un-optimised LB medium. The the-oretical C/N optimum value was found to be in the range 2.5–3.2. O’Kennedy et al. (20) reported an optimum C/N ratioof 2.78 for SDCAS medium.
C/N ratio in the PDM medium was varied between theoptimum range of 2.4–3.2 by altering the concentration ofNH
4Cl. The highest pDNA specific yield (11.17 µg/mg dry
cell mass) occurred at C/N ratio of 2.8 corresponding to abiomass yield of 2.13 mg/ml. The C/N-optimised PDM me-dium is referred to as PDMR.
Effect of phosphate concentration on plasmid produc-tion The fundamental building blocks of a pDNA mole-cule are nucleotides. These nucleotides are made of a phos-phate group, a sugar ring and a nitrogen base so the pres-ence of phosphate in a growth medium can have an influ-ence on the synthesis of nucleotides thereby affecting pDNAyield. LB and SOC have approximately negligible phosphateconcentration except for the little amount present in yeast ex-tract and tryptone which amounts to only 0.04% and 0.3%respectively. The total quantity of phosphate in LB and SOCmedia were evaluated to be 0.09 g/l and 0.18 g/l respec-tively (Fig. 6). The difference in the specific yield of LB andSOC is partially attributed to the presence of extra phos-
FIG. 2. (A) Plasmid DNA specific yield and purity obtained from LB, TB, SOC and PDM. Purity was measured by the ratio of OD260
to OD280
.PDM medium resulted with the highest plasmid purity due to the high specific yield of pDNA. Black bars, Final specific yield; gray bars,OD
260/OD
280 (B) Results from ethidium bromide agarose gel electrophoresis of pDNA samples from LB, TB, SOC and PDM media. Analysis was
done using 1% agarose in TAE×1 buffer, 5 µg/ml EtBr at 66 V. Lane M is 1 kbp DNA ladder, lanes 1, 2, 3 and 4 represent the pDNA samples fromPDM, TB, SOC and LB, respectively.
FIG. 3. Dry cell concentration of E. coli DH5α pUC19 in LB+x%where x is the percentage amount of glucose as carbon source (0–12%). Shake flask fermentation was performed at 37°C for 15 h under200 rpm shaking (n=3).
FIG. 4. Plasmid DNA volumetric and specific yield profiles forLB+x% where x is the percentage amount of glucose as carbon source.Shake flask fermentation was performed at 37°C for 15 h under 200rpm shaking (n=3). Symbols: rhombuses, volumetric yield; squares,specific yield.

MEDIUM SELECTION AND ECONOMICS FOR pDNA PRODUCTIONVOL. 104, 2007 495
phate concentration in SOC and the presence of other essen-tial nutrients such as carbon source and trace metals. TB andPDMR showed higher pDNA specific yields than LB andSOC; possibly due to the availability of extra phosphatesgroups. However, the pDNA specific yield from TB is foundto be lower than that of PDMR though it contains a higherphosphate concentration. TB as a commercially availablemedium contains nutrients which are not perhaps optimizedsolely for pDNA replication but for cell proliferation andother biomolecules synthesis such as proteins. On the otherhand, PDMR contains the right nutrients (P:C:N) whosequantities are stoichiometrically evaluated and tuned in theright proportion for plasmid replication, hence the highestpDNA specific yield.
Performance of PDMR employed in batch fermentationBatch cultivation of E. coli DH5α-pUC 19 using PDMRmedium showed a massive increase in biomass yield as wellas plasmid volumetric and specific yields. After an initial lagof 3 h the biomass yield increased to 0.26 mg/ml and to 4.55mg/ml after the next 12 h of cultivation (Fig. 7). Biomassyield increased continuously throughout the entire cultiva-
tion period with the expectation of further increment. Thiscontinuous increase in biomass is obviously due to the avail-able amount of carbon source present in PDMR medium forcell growth. Glucose uptake rate and metabolism by cell wereenhanced due to oxygen availability resulting from sparged airforced into the system. The maximum growth rate attainedduring cultivation was 0.45 h–1.
As shown in Fig. 8, there was a general correspondence be-tween plasmid volumetric yield and biomass growth withinthe first 12 h of cultivation. After the initial 3 h of nearlyzero plasmid production, the volumetric yield of plasmid in-creased to 1.1 µg/ml and to a maximum of 62.6 µg/ml after9 h. However, plasmid volumetric yield started declining fromthe 12th hour to a final value of 35.9 µg/ml. This is possiblydue to the use of glucose as a carbon source which usuallyincreases the maximum specific growth rate during the ex-ponential phase of batch fermentation (where rate of metab-olism is improved) and maximizes acetate production whichis detrimental to pDNA replication. An alternative carbonsource will be glycerol (21). Similar trend was observed for
FIG. 5. Effect of C/N ratio on pDNA production from LB andPDM medium. C/N ratios were achieved by adjusting concentration ofNH
4Cl. Shake flask fermentation was performed at 37 °C for 15 h
under 200 rpm shaking. Symbols: rhombuses, LB; squares, PDM.
FIG. 6. Dependency of pDNA specific yield on phosphate concen-tration of different growth media LB, TB, SOC and PDMR. Black bars,Specific yield; gray bars, total phosphate concentration.
FIG. 7. Dry cell concentration profiles obtained from OD600
mea-surements for the growth of Escherichia coli DH5α pUC19 in PDMRmedium. Batch fermentation was carried out in a 20 l fermentor at37°C for 15 h. Samples were collected at 30 min intervals (n =3).
FIG. 8. Plasmid DNA volumetric and specific yield profiles forPDMR medium. Batch fermentation was carried out in a 20 l fermen-tor at 37°C for 15 h. Samples were collected at 30 min intervals (n=3).Symbols: squares, volumetric yield; triangles, specific yield.

DANQUAH AND FORDE J. BIOSCI. BIOENG.,496
the pDNA specific yield. The maximum pDNA specific yieldof 18.07 µg/mg obtained from the batch cultivation of PDMRcompared to those routinely achieved reveals the potentialof PDMR for use in large scale pDNA production with min-imum cultivation period.
Economic evaluation The cost inventory for a typicalbatch shake flask fermentation process for pDNA produc-tion encompasses of all materials and equipment for the ac-tual fermentation work as well as downstream purification.The medium is a key critical input as it has direct impactboth on the pDNA yield (as shown earlier) and the cost. Me-dium selection is often based on following previously de-veloped protocols or, at best, trial and error without any costper yield of pDNA analysis prior to fermentation. This is themain reason for the low pDNA yields attained even for ex-pensive medium formulations and high copy number plas-mids. A medium cost per pDNA yield analysis was per-formed on LB, TB, SOC, PDM and PDMR using standardLB medium as a base scenario of 100 medium cost units,where 100 MC units ~ US$ 0.58. The cost of medium wascalculated based on the following relation.
Ck= αi,kmi,k+ βj,kνj,k (1)
Where Ck is the total cost of medium k, αi,k is the cost perunit weight of components i existing in medium k, mi,k is theweight of component i present in medium k, βj,k is the costper unit volume of components j existing in medium k andνj,k is the volume of component j present in medium k. Fromthe results obtained from the economic study as shown inFig. 9, the cost of medium per mg pDNA from PDM me-dium was 29 MC units and that of TB was 65 MC units. C/Nratio optimised PDM medium (PDMR) gave the highest plas-mid yield at 27 MC units/mg pDNA (Table 3). PDMR me-dium increased pDNA yield to an extent that it reduced thecost of medium per mg pDNA. Application of PDMR inbatch fermentations displayed an average volumetric yieldof 62.6 mg/l which is slightly higher than that of the originalMW1 in Wang et al. (~60 mg/l) (4). Aside this raw volumet-ric yield comparison, the cost per yield analysis which is themain thrust of this paper, favours outstandingly the PDMRmedium (7 medium cost (MC)/mg pDNA) more than theoriginal MW1 (35.2 MC/mg pDNA). However, costs of com-pressed air/oxygen and utilities can not be omitted in a com-plete cost evaluation of the batch run. Considering this eco-
nomic evaluation, there is a great cost per yield advantageof using PDMR medium for small scale batch production,which shows promise for larger scale production of pDNA.Sedegah et al. (22) reported that pDNA vaccine doses usedfor human trials are commonly 500–5000 µg. For mediumalone, this corresponds to 8.5–85 MC units (~US$ 0.05–0.50)when PDMR is used compared to 50–500 MC units (~US$0.29–2.90) for LB medium.
Growth medium selection and economics for large-scaleproduction of pDNA From a manufacturing perspective,large scale production of pDNA starts with high throughputfermentation. Medium formulation is a fundamental and chal-lenging consideration in large scale fermentation as it re-quires a decision to be made between the cost of nutrients,plasmid yield and quality. Considerations that must be putforward in developing a medium for large scale pDNA pro-duction are: (i) The optimum nutrient requirements of thehost organism; (ii) Comparison of nutrient requirements tothe composition of available industrial and off-the-shelf nu-trients. If not compatible, plan a formulation; (iii) Nutrientproperties with respect to storage, handling, sterilization,processing and pDNA purification; and (iv) cost inventoryof the ingredients.
Computation of the cost of the medium for large scaleproduction of pDNA involves the recognition of all cost re-
i 1=
∑j 1=
∑
FIG. 9. Medium cost per mg plasmid DNA from LB, TB, SOC,PDM and PDMR. The result shows medium cost per plasmid DNAyield advantage of PDMR. 100 units = US$ 0.58 for standard LB me-dium. Shake flask fermentation was performed at 37°C for 15 h under200 rpm shaking (n=3).
TABLE 3. Summary of pDNA yield and economic study results obtained for the various media investigatedin shake flask and batch fermentation
MediumDry cell concn
(g/l)pDNA vol. yield
(mg/l)pDNA specific yield
(mg/g)Medium cost/mg pDNA
(units)
LB 0.85 2.8 3.29 100LB (C optimised) 1.03 3.9 3.80 135LB (C/N optimised) 1.18 6.1 5.17 117TB 1.08 11.4 6.33 65SOC 1.31 6.4 4.86 81PDM 1.94 14.6 7.53 29PDMR 2.13 23.8 11.17 27PDMR (batch) (maximum yield) 3.66 62.6 17.10 7PDMR (batch) (final yield) 4.55 35.9 7.89 12
PDMR medium is obtained from C/N optimisation of PDM. Data shown are the average values of three replicates (n=3).

MEDIUM SELECTION AND ECONOMICS FOR pDNA PRODUCTIONVOL. 104, 2007 497
lating to the medium. Thus, in addition to the main purchaseprice or cost of formulation, cost of material handling andstorage, labour and analytical requirements must be esti-mated. Medium storage and handling depends on the typeand nature of the nutrients. Liquid nutrients require largerstorage capacities than slurry or concentrated nutrients. How-ever, most of the nutrients required for bacteria fermentationfor pDNA production exist in powder forms; hence lessstorage capacity is required. The rheological properties ofthe medium may affect the cost of operations such as mix-ing, aeration and temperature control. The concentration ofthe pDNA influences its recovery and purification costs.Productivity per unit process capacity helps determine theamount of capital, labour, and indirect costs assignable topDNA. Reducing manufacturing costs by replacing nutrientswith cheaper ones may not be the solution. A better approachwould be to explore how the impact of a change in nutrientwould affect the product yield and purity in addition to thecost analysis.
ACKNOWLEDGMENTS
Funding for this research was kindly provided by the VictorianEndowment for Science, Knowledge and Innovation (VESKI) andvia the Monash University Early Career Researcher Grant Scheme.
REFERENCES
1. Durland, R. H. and Eastman, E. M.: Manufacturing andquality control of plasmid-based gene expression systems. Adv.Drug Deliv. Rev., 30, 33–48 (1998).
2. Prather, K. J., Sagar, S., Murphy, J., and Chartrain, M.:Industrial scale production of plasmid DNA for vaccine andgene therapy: plasmid design, production and purification.Enzyme Microb. Technol., 33, 865–883 (2003).
3. Prazeres, D., Ferreira, G. N. M., Monteiro, G. A., Cooney,C. L., and Cabral, J. M. S.: Large-scale production of phar-maceutical-grade plasmid DNA for gene therapy: problems andbottlenecks. Trends Biotechnol., 17, 169–174 (2003).
4. Wang, Z., Le, G., Shi, W., and Wegrzyn, G.: Medium de-sign for plasmid DNA production based on stoichiometricmodel. Process Biochem., 36, 1085–1093 (2001).
5. Mor, G.: Plasmid DNA, a new era in vaccinology. Biochem.Pharm., 55, 1151–1153 (1998).
6. Butts, C., Zubkoff, I., and Robbins, D. S.: DNA immuniza-tion of infants: potential and limitations. Vaccine, 16, 1444–1449 (1998).
7. Schodel, F., Aguado, M. T., and Lambert, P. H.: Introduc-tion: nucleic acid vaccine, WHO, Geneva. Vaccine, 12, 1491–1492 (1994).
8. O’Kennedy, R., Ward, J., and Keshavarz-Moore, E.:Effect of fermentation strategy on the characteristics of plas-mid DNA production. Biotechnol. Appl. Biochem., 37, 83–90(2003).
9. Stanier, R. Y., Doudoroff, M., and Adelberg, E. A.: Themicrobial world, 3rd ed. Prentice Hall, Englewood Cliffs, NJ(1976).
10. West-Robert, C.: Handbook of chemistry and physics, 60thed. CRC Press, Boca Raton, Florida (1979).
11. Makrides, S.: Strategies for achieving high-level expressionof genes in Escherichia coli. Microbiol. Rev., 60, 512–538(1996).
12. Balbas, P. and Bolivar, F.: Design and construction of ex-pression plasmid vectors in Escherichia coli. Methods Enzymol.,185, 14–37 (1990).
13. Venetianer, P.: Possibilities of increasing the expression ofcloned foreign genes in Escherichia coli. Acta Biotechnol.,11, 129–133 (1991).
14. Riesenberg, D., Menzel, K., Schultz, V., Schumann, K.,Veith, G., and Zuber, G.: High cell density fermentation ofrecombinant Escherichia coli expressing human interferonalpha 1. Appl. Microbiol. Biotechnol., 34, 77–82 (1990)
15. Zabriskie, D., Wareheim, D., and Polanski, M.: Effects offermentation feeding strategies prior to induction of expres-sion of a recombinant malaria antigen in Escherichia coli.Ind. Microbiol., 2, 87–95 (1987).
16. Nakagawa, S., Oda, H., and Anazawa, H.: High cell den-sity cultivation and high recombinant protein production ofEscherichia coli strain expressing uricase. Biosci. Biotechnol.Biochem., 59, 2263–2267(1995).
17. Korz, D., Hellmurh, R., Sanders, E., and Deckwer, W.:Simple fed-batch technique for high cell density cultivation ofEscherichia coli. Biotechnol., 39, 59–65 (1995).
18. Diogo, M., Queiroz, J., Monteiro, G., Martins, S., Ferreira,G., and Prazeres, D.: Purification of a cystic fibrosis plasmidvector for gene therapy using hydrophobic interaction chro-matography. Biotechnol. Bioeng., 68, 576–583 (2000).
19. Drew, D., Lighttowlers, M., and Strugnell, R.: Humoral im-mune responses to DNA vaccines expressing secreted, mem-brane bound and non-secreted forms of the Taeniaovis 45Wantigen. Vaccine, 18, 2522–2532 (2000).
20. O’Kennedy, R., Baldwin, C., and Keshavarz-Moore, E.:Effect of growth medium selection on plasmid DNA pro-duction and initial processing step. Biotechnology, 76, 175–183 (2000).
21. Carnes, A. E., Hodgson, C. P., and Williams, J. A.: Induc-ible Escherichia coli fermentation for increased plasmid DNAproduction. Biotechnol. Appl. Biochem., 45, 155–166 (2006).
22. Sedegah, M., Rogers, W. O., Belmonte, A., Belmonte, M.,Banania, G., Patterson, N., Ferrari, M., Kaslow, D. C.,Carucci, D. J., Richie, T. L., and Doolan, D. L.: Vaxfectin™
enhances immunogenicity and protective efficacy of P. yoeliicircumsporozoite DNA vaccines. Vaccine, 24, 1921–1927(2006).


















