
Glutathione and Glutathione Transferase Levels - Cancer Research
7
[CANCER RESEARCH 46, 735-739, February 1986] Glutathione and Glutathione Transferase Levels in Mouse Granulocytes following Cyclophosphamide Administration1 James Carmichael,2 David J. Adams,3 John Ansell, and C. Roland Wolf4 Imperial Cancer Research Fund, Laboratory of Molecular Pharmacology and Drug Metabolism, Department ot Biochemistry, George Square [J. C., D. J. A., C. P. W.], and Department of Zoology, The King's Buildings [J. A.], University of Edinburgh, Edinburgh, United Kingdom ABSTRACT Following an initial depletion, glutathione and glutathione trans- ferase levels are transiently increased in mouse bone marrow following the administration of a low dose of Cyclophosphamide. Similar effects are observed on subsequent administration of the drug. The separation of various bone marrow populations on a fluorescence-activated cell sorter has shown that increase in glutathione and glutathione transferase levels are restricted to the granulocytic fraction. This may well provide an explanation for the protective effect of a low 'priming' dose of Cyclophospha mide against a subsequent lethal dose. The changes in granu locytic glutathione and glutathione transferase levels can also be monitored in the peripheral circulation. The enhanced levels of glutathione in cells resulting from cytotoxic insult appear to be a general response of cells to cytotoxins and may be important in both antitumor therapy as well as the initiation of chemical toxicity and carcinogenicity. INTRODUCTION The response of normal cells to cytotoxins and chemical carcinogens is important in our understanding of the changes which result in malignant transformation (1). In addition, the response to cytotoxins is also an important aspect of current cancer chemotherapy in that toxicity to normal tissues is a dose- limiting factor. In this regard we are currently investigating the properties of bone marrow cells which in addition to being sites of malignant transformation are also sensitive targets for che- motherapeutic agents. In a previous communication we reported the response of mouse bone marrow to the cytotoxins Cyclo phosphamide, X-radiation, and 1-0-D-arabinofuranosylcytosine (2). In particular, the effects of these reagents on marrow gluta thione and glutathione transferases were studied as these com ponents play a central role in the protection of cells from a wide variety of alkylating agents as well as radiation (3, 4). The data presented indicated that treatment of mice with a low cytotoxic dose of these reagents resulted in a significant increase in marrow GSH5 and GSH transferase content. These increases were paralleled by an increased resistance to a high dose of the cytotoxic compounds. These experiments raised the following questions: (a) Were the changes observed related to changes in Received 5/29/85; revised 10/23/85; accepted 10/24/85. 1The financial support of the Cancer Research Campaign, Grant No. SP 1610, is gratefully acknowledged. * Present address: National Cancer Institute, Naval Medical Oncology Branch, Bethesda, MD. 3 Present address: Department of Microbiology, University of Leeds, Leeds, LS2 9JT, United Kingdom. 4To whom requests for reprints should be addressed. * The abbreviations used are: GSH, glutathione; FACS, fluorescence activated cell sorter. marrow cell population? (b) Were the changes due to alterations in intracellular GSH and GSH transferase homeostasis and there fore did they represent a general response to cytotoxic com pounds? If so, which cells were affected? In this report we have investigated these possibilities following the administration of Cyclophosphamide. In addition we have determined whether the changes in the marrow cells are reflected in the cells of the peripheral blood and therefore provide a potential method for monitoring the status of the bone marrow during chemotherapy. MATERIALS AND METHODS Male CBA mice (25 g) were used. Animals treated with Cyclophospha mide received i.p. either 75 mg/kg body weight as a nonlethal "priming" dose (2) or 350 mg/kg in 0.9% saline. At various time points after drug administration, animals were killed by cervical dislocation. Blood samples were drawn from the posterior vena cava into a heparinized syringe. Bone marrow cells were obtained by passing cold isotonic saline buffered with 10 HIM phosphate through the femurs as previously described (2). Peritoneal granulocytes were prepared by giving mice i.p. injections of 2 ml of a solution of 2% calcium caseinate (5). After 3 h the animals were killed and 5 ml of 0.168 M NaCI was injected i.p. The peritoneal exúdate was withdrawn after gentle peritoneal massage, and the cells were spun down at 650 x g for 5 min, washed three times with ice cold 0.168 M NH4CI, and left to stand for 10 min to lyse erythrocytes. Cells were then suspended in 0.168 M phosphate buffered saline for GSH assay. For cell sorter (FACS IV; Becton Dickinson) analysis and sorting, approximately 1 ml of blood was washed in 9 ml of cold phosphate buffered isotonic saline supplemented with 0.1% bovine serum albumen and 0.02% EDTA. After centrifugation at 450 x g for 7 min, the blood cell pellet was mixed with 18 ml of distilled water and left for 12 s to lyse erythrocytes. Two ml of 10x concentrated Hanks' balanced salt solution (Grand Island Biological Co.) were then added and mixed. After a second centrifugation at 250 x g for 7 min, the cell pellet was resuspended in 1 ml of the supplemented saline. (If erythrocytes were to be separated on the FACS, peripheral blood was used directly after appropriate dilution.) Bone marrow cells were obtained by flushing femurs with 1 ml of similarly supplemented saline. Various cell types from bone marrow and peripheral blood suspensions were separated on the FACS on the basis of their differential forward and right angle light scattering properties (6, 7). The 488-nm line of an argon ¡on laser was used for the excitation of both the forward and right angle scatter signals and samples were run at approximately 1000 cells/ s. Appropriate gates were set around granulocytes, lymphocytes, eryth rocytes, and a fourth mixed cell population (mainly monocytes) from bone marrow preparations (see Fig. 4), and lymphocytes, granulocytes, and erythrocytes from peripheral blood (see Fig. 6), and these popula tions were sorted into cooled Eppendorf tubes. The purity of the cell fractions was tested cytologically by Leishman staining. The granulocytes were 89.9 ±7.6% (SD) pure (n = 6) and the lymphocyte population was 70.1 ±10.9% pure, the remainder of the cells being monocytes. All procedures were carried out at 4°C.Cells were collected in aliquots CANCER RESEARCH VOL. 46 FEBRUARY 1986 735 on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Transcript of Glutathione and Glutathione Transferase Levels - Cancer Research

[CANCER RESEARCH 46, 735-739, February 1986]
Glutathione and Glutathione Transferase Levels in Mouse Granulocytesfollowing Cyclophosphamide Administration1
James Carmichael,2 David J. Adams,3 John Ansell, and C. Roland Wolf4
Imperial Cancer Research Fund, Laboratory of Molecular Pharmacology and Drug Metabolism, Department ot Biochemistry, George Square [J. C., D. J. A., C. P. W.],and Department of Zoology, The King's Buildings [J. A.], University of Edinburgh, Edinburgh, United Kingdom
ABSTRACT
Following an initial depletion, glutathione and glutathione trans-
ferase levels are transiently increased in mouse bone marrowfollowing the administration of a low dose of Cyclophosphamide.Similar effects are observed on subsequent administration of thedrug. The separation of various bone marrow populations on afluorescence-activated cell sorter has shown that increase in
glutathione and glutathione transferase levels are restricted tothe granulocytic fraction. This may well provide an explanationfor the protective effect of a low 'priming' dose of Cyclophospha
mide against a subsequent lethal dose. The changes in granulocytic glutathione and glutathione transferase levels can also bemonitored in the peripheral circulation. The enhanced levels ofglutathione in cells resulting from cytotoxic insult appear to be ageneral response of cells to cytotoxins and may be important inboth antitumor therapy as well as the initiation of chemical toxicityand carcinogenicity.
INTRODUCTION
The response of normal cells to cytotoxins and chemicalcarcinogens is important in our understanding of the changeswhich result in malignant transformation (1). In addition, theresponse to cytotoxins is also an important aspect of currentcancer chemotherapy in that toxicity to normal tissues is a dose-
limiting factor. In this regard we are currently investigating theproperties of bone marrow cells which in addition to being sitesof malignant transformation are also sensitive targets for che-
motherapeutic agents. In a previous communication we reportedthe response of mouse bone marrow to the cytotoxins Cyclophosphamide, X-radiation, and 1-0-D-arabinofuranosylcytosine
(2). In particular, the effects of these reagents on marrow glutathione and glutathione transferases were studied as these components play a central role in the protection of cells from a widevariety of alkylating agents as well as radiation (3, 4). The datapresented indicated that treatment of mice with a low cytotoxicdose of these reagents resulted in a significant increase inmarrow GSH5 and GSH transferase content. These increases
were paralleled by an increased resistance to a high dose of thecytotoxic compounds. These experiments raised the followingquestions: (a) Were the changes observed related to changes in
Received 5/29/85; revised 10/23/85; accepted 10/24/85.1The financial support of the Cancer Research Campaign, Grant No. SP 1610,
is gratefully acknowledged.* Present address: National Cancer Institute, Naval Medical Oncology Branch,
Bethesda, MD.3 Present address: Department of Microbiology, University of Leeds, Leeds, LS2
9JT, United Kingdom.4To whom requests for reprints should be addressed.* The abbreviations used are: GSH, glutathione; FACS, fluorescence activated
cell sorter.
marrow cell population? (b) Were the changes due to alterationsin intracellular GSH and GSH transferase homeostasis and therefore did they represent a general response to cytotoxic compounds? If so, which cells were affected?
In this report we have investigated these possibilities followingthe administration of Cyclophosphamide. In addition we havedetermined whether the changes in the marrow cells are reflectedin the cells of the peripheral blood and therefore provide apotential method for monitoring the status of the bone marrowduring chemotherapy.
MATERIALS AND METHODS
Male CBA mice (25 g) were used. Animals treated with Cyclophosphamide received i.p. either 75 mg/kg body weight as a nonlethal "priming"
dose (2) or 350 mg/kg in 0.9% saline. At various time points after drugadministration, animals were killed by cervical dislocation. Blood sampleswere drawn from the posterior vena cava into a heparinized syringe.Bone marrow cells were obtained by passing cold isotonic saline bufferedwith 10 HIM phosphate through the femurs as previously described (2).
Peritoneal granulocytes were prepared by giving mice i.p. injectionsof 2 ml of a solution of 2% calcium caseinate (5). After 3 h the animalswere killed and 5 ml of 0.168 M NaCI was injected i.p. The peritonealexúdate was withdrawn after gentle peritoneal massage, and the cellswere spun down at 650 x g for 5 min, washed three times with ice cold0.168 M NH4CI, and left to stand for 10 min to lyse erythrocytes. Cellswere then suspended in 0.168 M phosphate buffered saline for GSHassay.
For cell sorter (FACS IV; Becton Dickinson) analysis and sorting,approximately 1 ml of blood was washed in 9 ml of cold phosphatebuffered isotonic saline supplemented with 0.1% bovine serum albumenand 0.02% EDTA. After centrifugation at 450 x g for 7 min, the bloodcell pellet was mixed with 18 ml of distilled water and left for 12 s to lyseerythrocytes. Two ml of 10x concentrated Hanks' balanced salt solution
(Grand Island Biological Co.) were then added and mixed. After a secondcentrifugation at 250 x g for 7 min, the cell pellet was resuspended in 1ml of the supplemented saline. (If erythrocytes were to be separated onthe FACS, peripheral blood was used directly after appropriate dilution.)Bone marrow cells were obtained by flushing femurs with 1 ml of similarlysupplemented saline.
Various cell types from bone marrow and peripheral blood suspensionswere separated on the FACS on the basis of their differential forwardand right angle light scattering properties (6, 7). The 488-nm line of an
argon ¡onlaser was used for the excitation of both the forward and rightangle scatter signals and samples were run at approximately 1000 cells/s. Appropriate gates were set around granulocytes, lymphocytes, erythrocytes, and a fourth mixed cell population (mainly monocytes) frombone marrow preparations (see Fig. 4), and lymphocytes, granulocytes,and erythrocytes from peripheral blood (see Fig. 6), and these populations were sorted into cooled Eppendorf tubes. The purity of the cellfractions was tested cytologically by Leishman staining. The granulocyteswere 89.9 ±7.6% (SD) pure (n = 6) and the lymphocyte population was
70.1 ±10.9% pure, the remainder of the cells being monocytes.All procedures were carried out at 4°C.Cells were collected in aliquots
CANCER RESEARCH VOL. 46 FEBRUARY 1986
735
on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CYCLOPHOSPHAMIDE EFFECTS ON GSH AND GLUTATHIONE TRANSFERASE
of 3 x 10s for further assays, frozen immediately after collection in solid
C02 and assayed within 24 h. Control experiments showed that therewas no difference in glutathione measurements between cells assayedimmediately and cells stored in this manner.
Cellular glutathione content was determined from the deproteinatedcell homogenates by the fluorimetrie method of Hissin and Hilf (8) usingo-phthalaldehyde. This assay was used for its simplicity, and the values
obtained compared favorably with other published methods describedpreviously (2). Glutathione transferase activity was measured in the cellsamples lysed by addition of nonisotonic buffer, 0.1 M phosphate, pH6.5, using 1-chloro-2,4-dinitrobenzene as substrate as described by
Habig et al. (9).Cell numbers were determined either with the FACS or by hemocy-
tometer.All the reagents used in this study were obtained from commercial
sources and were of the highest purity available.
RESULTS
The effect of nonlethal priming and a subsequent lethal doseof cyclophosphamide on bone marrow GSH and GSH transferaseis shown in Fig. 1. In agreement with previous findings (2) aftera dose of 75 mg/kg an initial depletion in glutathione and GSHtransferase levels was followed by a significant increase overcontrol levels on day 5, the concentrations being increased 1.8-and 2.5-fold for GSH and GSH transferase, respectively. On day
6 a normally lethal dose of cyclophosphamide (350 mg/kg) wasadministered which resulted in a second depletion in both GSH
300
200
•s3
100
35O mg/kg
Time Idaysl
2 4 6 8 10 12 UTime (days)
Fig. 1. The effect of cyclophosphamide on bone marrow glutathione (a) andglutathione transferase (£>)levels. Male CBA mice were initially given ¡.p.injectionsof cyclophosphamide (75 mg/kg). Six days later a second dose (350 mg/kg) wasadministered. Bone marrow samples were obtained on the days shown as described in "Materials and Methods." Three animals were taken per experimental
point. Data are expressed relative to control values obtained at that particular timepoint. Control values were 7.15 ±1.46 x 10"" mol/cell and 6.01 ±1.69 nmol 1-chloro-2,4-dinitrobenzene conjugated/min/106 cells (n = 14) for glutathione and
glutathione transferase, respectively.
and GSH transferase levels to 32 and 40% of control values,respectively. This decrease was much less pronounced whencompared with previously untreated animals where GSH contentwas reduced to 5% of controls (data not shown) (2). The GSHlevels remained suppressed for 3 days but recovered and increased to much higher levels on day 11. In the case of theglutathione transferase activity, the activity was increased approximately 8-fold and GSH approximately 2.5-fold. The primed
cells of the marrow which were more resistant to a lethal doseof cyclophosphamide therefore respond to the high dose in amanner very similar to their response to a priming dose. Thereare several possible explanations for measurement of increasedGSH and GSH transferase levels in the marrow following cyclophosphamide administration, as whole marrow samples weretaken for assay. Experiments were carried out to establishwhether the GSH and GSH transferase activities were intracel-lular (Fig. 2). In these experiments marrow cells from control andprimed animals were spun down. In both cases the glutathionecontent of the pelleted cells closely matched the theoreticalcontent based on the measured cell viability of the original bonemarrow sample, measured using trypan blue, and the GSHcontent of the original sample, demonstrating that the GSHmeasured in these assays was intracellular.
Administration of cyclophosphamide at either 75 or 350 mg/
Control CyclophosphamideI-treated
IFig. 2. Glutathione content of marrow cells from untreated mice or mice treated
with cyclophosphamide 5 days before use. Aliquots of the bone marrow samplewere either taken for direct measurement of glutathione (•)or were pelletedresuspended in ice cold isotonic saline and the glutathione content was determined(D); D, theoretical glutathione level in the cell pellet based on the cell viabilitymeasured with trypan blue and the glutathione content of the original sample. Thisexperiment was repeated and gave the same results, cone., concentration.
190
I
< 2 3 4 5 6 7 8 9 10 11 12 13 14 15Time (days)
Fig. 3. Effect of cyclophosphamide (75 mg/kg) on bone marrow cell number.Experimental details are as those given in Fig. 1.
CANCER RESEARCH VOL. 46 FEBRUARY 1986
736
on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
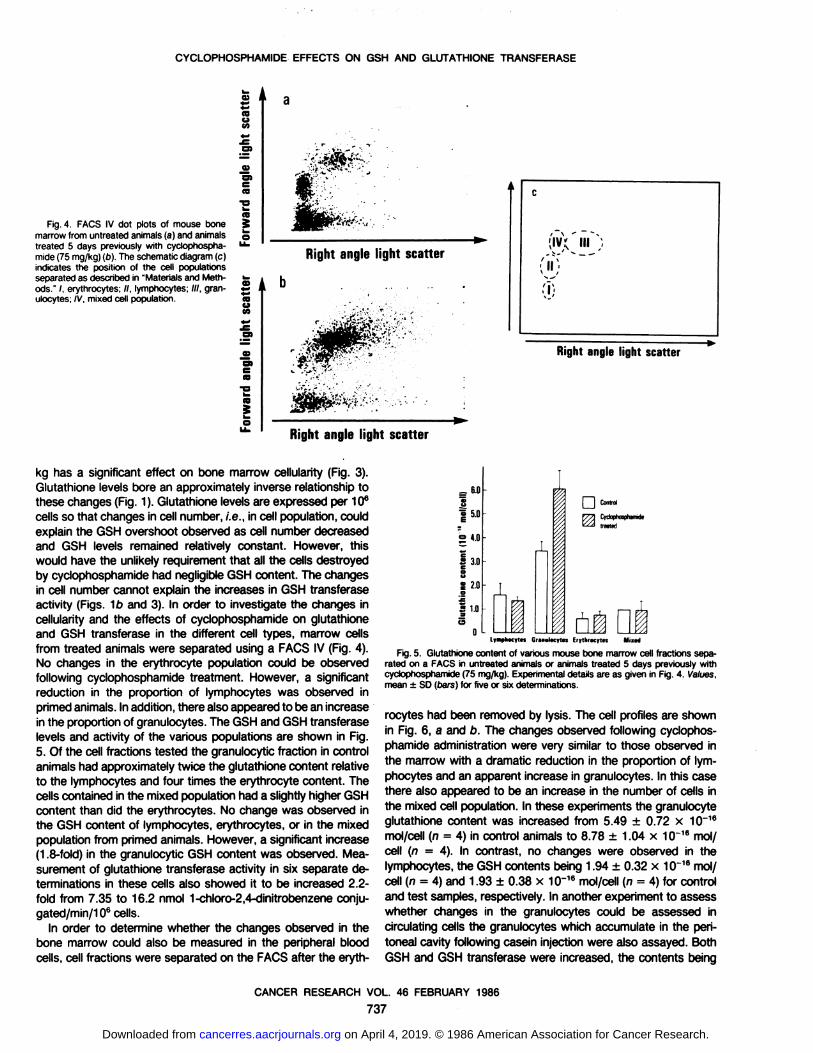
CYCLOPHOSPHAMIDE EFFECTS ON GSH AND GLUTATHIONE TRANSFERASE
Fig. 4. FACS IV dot plots of mouse bonemarrow from untreated animals (a) and animalstreated 5 days previously with cyclophospha-
mide (75 mg/kg) (o). The schematic diagram (c)indicates the position of the cell populationsseparated as described in "Materials and Methods." /. erythrocytes; //, lymphocytes; ///, gran-
ulocytes; IV. mixed cell population.
09 i L
«BUta
en
O)
IB
IBOM
—en
IB-a
L.
IB
-TÕ*1-'-•*•; . ' " '. «*
Rightanglelight scatter
/ \ " N;iv' HI ;
olÃ
-1)
Right angle light scatter
Rightanglelight scatter
kg has a significant effect on bone marrow cellularity (Fig. 3).Glutathione levels bore an approximately inverse relationship tothese changes (Fig. 1). Glutathione levels are expressed per 106
cells so that changes in cell number, i.e.. in cell population, couldexplain the GSH overshoot observed as cell number decreasedand GSH levels remained relatively constant. However, thiswould have the unlikely requirement that all the cells destroyedby cyclophosphamide had negligible GSH content. The changesin cell number cannot explain the increases in GSH transferaseactivity (Figs. 1b and 3). In order to investigate the changes incellularity and the effects of cyclophosphamide on glutathioneand GSH transferase in the different cell types, marrow cellsfrom treated animals were separated using a FACS IV (Fig. 4).No changes in the erythrocyte population could be observedfollowing cyclophosphamide treatment. However, a significantreduction in the proportion of lymphocytes was observed inprimed animals. In addition, there also appeared to be an increasein the proportion of granulocytes. The GSH and GSH transferaselevels and activity of the various populations are shown in Fig.5. Of the cell fractions tested the granulocytic fraction in controlanimals had approximately twice the glutathione content relativeto the lymphocytes and four times the erythrocyte content. Thecells contained in the mixed population had a slightly higher GSHcontent than did the erythrocytes. No change was observed inthe GSH content of lymphocytes, erythrocytes, or in the mixedpopulation from primed animals. However, a significant increase(1.8-fold) in the granulocytic GSH content was observed. Mea
surement of glutathione transferase activity in six separate determinations in these cells also showed it to be increased 2.2-fold from 7.35 to 16.2 nmol 1-chloro-2,4-dinitrobenzene conju-gated/min/106 cells.
In order to determine whether the changes observed in thebone marrow could also be measured in the peripheral bloodcells, cell fractions were separated on the FACS after the eryth-
Glutlthionecontent(10"moliceli)=sssssg•
JLJLI
lymphacyip\ GrMHilKyln Erytlirocyt« Mixed
Fig. 5. Glutathione content of various mouse bone marrow cell fractions separated on a FACS in untreated animals or animals treated 5 days previously withcyclophosphamide (75 mg/kg). Experimental details are as given in Fig. 4. Values,mean ±SD (bars) for five or six determinations.
rocytes had been removed by lysis. The cell profiles are shownin Fig. 6, a and b. The changes observed following cyclophosphamide administration were very similar to those observed inthe marrow with a dramatic reduction in the proportion of lymphocytes and an apparent increase in granulocytes. In this casethere also appeared to be an increase in the number of cells inthe mixed cell population. In these experiments the granulocyteglutathione content was increased from 5.49 ±0.72 x 10~16mol/cell (n = 4) in control animals to 8.78 ±1.04 x 10~16mol/
cell (n = 4). In contrast, no changes were observed in thelymphocytes, the GSH contents being 1.94 ±0.32 x 10~16mol/cell (n = 4) and 1.93 ±0.38 x 10~16mol/cell (n = 4) for control
and test samples, respectively. In another experiment to assesswhether changes in the granulocytes could be assessed incirculating cells the granulocytes which accumulate in the peritoneal cavity following casein injection were also assayed. BothGSH and GSH transferase were increased, the contents being
CANCER RESEARCH VOL. 46 FEBRUARY 1986
737
on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CYCLOPHOSPHAMIDE EFFECTS ON GSH AND GLUTATHIONE TRANSFERASE
couM
Fig. 6. FACS IV dot plots of untreated (a)and cyclophosphamide-treated (b) mouse peripheral blood, prepared as described in "Materials and Methods." The schematic diagram
(c) indicates the position of the populationsseparated. I, erythrocytes; //, lymphocytes; ///,granulocytes.
~m*.
Right angle light scatter
a»en
C'""")
Õ
Right angle light scatter
Right angle light scatter
approximately 2.0- and 1.7-fold higher than controls, respectively. The glutathione values were 5.30 ±1.09 x 10~16(n = 3)and 10.39 ±1.87 x 10~16(n = 3) mol glutathione/cell, and the
glutathione transferase values were 10.42 ±1.87 (n = 3) and16.87 ±4.53 (n = 3) nmol 1-chloro-2,4-dinitrobenzene conju-gated/min/106 cells for control and primed animals, respectively.
DISCUSSION
In this study we have demonstrated that significant changesin intracellular marrow glutathione and glutathione transferaseactivity occur following the administration of a myelotoxic primingdose of cyclophosphamide (2, 10). In agreement with literaturereports (11, 12), cyclophosphamide had significant effects onbone marrow cellularity as well as the relative proportions of thevarious cell types. The marked loss of lymphocytes at the doseused was of particular interest. On the basis of these changesour earlier observation of increased glutathione or GSH transferase activity in marrow fractions (2) could be partially explainedby a changed marrow cell population. However, increases inintracellular granulocytic GSH and GSH transferase levels wouldalso appear to be an important factor. Glutathione is known toprotect against cyclophosphamide-induced bladder damage (13)
and we have recently postulated that it is also protective againstthe myelotoxicity of this compound (2). The lethality of highdoses of cyclophosphamide is associated with granulocytopenia.The protective effect of a priming dose could therefore beexplained by increased resistance of the granulocytes to toxicityconferred by their higher glutathione content. The difference ingranulocytic and lymphocytic GSH content could also explainthe higher susceptibility of the latter cells to cyclophosphamide.The changes in granulocyte GSH and GSH transferase contentcould also be measured in cells isolated from the peripheralcirculation. This fact tends to rule out the possibility that the cells
separated from the marrow by the FACS within the granulocyticfraction following priming represent an enrichment of precursorcells rather than granulocytes with higher GSH content. Moreimportantly, the finding that changes associated with primingcan be monitored in the peripheral blood could be used tooptimize the timing of high dose chemotherapy regimens ¡nwhichdrug priming is used.
The changes in GSH and GSH transferase activities in thegranulocytes were even more marked on a second administrationof the cytotoxin (Fig. 1). This could also be of potential clinicalvalue. The fact that the stimulatory response can be repeatedand each time follows a suppression of enzyme or cofactor levelmay be indicative of a feedback control mechanism leading tooverproduction. Such a mechanism has been identified in thecase of GSH homeostasis (14). It has also been demonstratedthat cyclophosphamide administration stimulates a burst of mi-totic activity (11, 12). This may also be related to the changesobserved. This finding together with other literature reports (15-17) provides an increasing body of evidence that changes inGSH and GSH transferase levels in cells represent a generalresponse to cytotoxic damage caused by radiation or alkylatingagents. Increases in GSH and GSH transferase levels have beenrelated to the initiation of chemically induced neoplastic growth(1,18). Such increases in the tumor would undoubtedly protectit from further therapy. Indeed there are certain examples ofchemically induced drug resistance which have been explainedby increased glutathione levels (19, 20).
ACKNOWLEDGMENTS
The authors wish to thank D. V. van Heyningen for suggesting the use of caseinfor the induction of peritoneal granulocytes, Dr. H. S. Micklem and the CancerResearch Campaign for the use of the FACS IV, A. Sanderson for assistance withsorting, L. Forrester for carrying out the cytological staining and R. A. Ramage forexpert assistance in the preparation of this manuscript.
CANCER RESEARCH VOL. 46 FEBRUARY 1986
738
on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

.-•y?
CYCLOPHOSPHAMIDE EFFECTS ON GSH AND GLUTATHIONE TRANSFERASE
REFERENCES
1. Faber,E. The biochemistryof preneoplastic liver: a common metabolicpatternin hepatocyte nodules. Can. J. Biochem. Cell Biol.. 62: 486-494, 1984.
2. Adams, D. J., Carmichael, J., and Wolf. C. R. Altered mouse bone marrowglutathioneand glutathione transferase levels in response to cytotoxins. Cancer Res., 45:1669-1673,1985.
3. Chasseaud.L. F. The rote of glutathione S-transferases in the metabolism ofchemical carcinogens and other electrophilic agents. Adv. Cancer Res., 29:175-275,1979.
4. Meister, A., and Anderson, M. E. Glutathione. Ann. Rev. Biochem., 52: 711-760,1983.
5. Watt, S. M., Burgess, A. W. and Metcalf, D. Isolation and surface labellingofmurine pcdymorphonudearneutrophils. J. Cell. Phystol., 100:1-21:1979.
6. Ritclue. A. W. S., Gray, R. A., and Micklem. H. S. Rightangle light scatter: anecessary parameter in flow cytofluorimetric analysis of human peripheralblood mononuclearcells. J. Immunol. Methods, 64:109-117,1983.
7. Wan, S. M.. Burgess, A. W., Metcalf, D., and Battye, F. L. Isolationof mousebone marrow neutrophils by light scatter and autofluorescence.J. Histochem.Cytochem.,28: 934-946,1980.
8. Hissin, P. J., and Hilf, R. A fluorimetrie method for determination of oxidizedand reduced glutathione in tissues. Anal. Biochem. 74: 214-226,1976.
9. Habig, W. H., Pabst, M. J., and Jakoby, W. B. Glutathione transferases, thefirst enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139,1974.
10. Millar, J. L., Hudspith, B. N., and Blacknett, N. M. Reduced lethality in micereceiving a combination dose of cyclophosphamide and busulphun. Br. J.Cancer., 32. 193-198,1975.
11. Adam, M., Ernst, H., and Bicker, U. Influenceof cyclophosphamide(Endoxan)and azimexone (BM12.531), 2-|2-cyanaziridinyl-(1)|-2-|2-carbomoylazindinyl-(1)]-propane,on the bone marrow cells of mice. Exp. Pathol.. 79:1-14,1981.
12. Rosenoff, S. H., Bosttek, F., and Young, R. C. Recovery of normal hemato-poietic tissue and tumor following chemotherapeuticinjury from cyclophosphamide(CTX):comparativeanalysisof biochemicalandclinicaltechniques.Blood,45: 465-475,1975.
13. Berrigan, M. J., Mannello,A. J., Pavelic,Z., Williams,C. J., Struck, R. f., andGurtoo, H. L. Protective roteof thtols in cydophosphamide-inducedurotoxiciiyand depression of hepatic drug metabolism. Cancer Res., 42: 3688-3695,1982.
14. Richman,P., and Meister, A. Regulationof •y-glutamyl-cysteinesynthetase bynonallosteric feedback inhibition by glutathione. J. Biol. Chem., 250: 1422-1426,1975.
15. Di Simplicio, P. Glutathione and glutathione S-transferases in rat liver and inplasma after carbon tetrachlondeand thioacetamide intoxication. Pharmacol.Res. Commun., 74: 909-920,1982.
16. Kimball, R. E., Reddy, K., and Pierce,T. H. Oxygen toxicity: augmentationofantioxidant defencemechanismsin rat lung. Am. J. Physiol , 230:1425-1431,1976.
17. Lock, E. A., and ishmaei,J. Hepatic and renal non-protein sulphydryl concentration following toxic doses of hexachloro-1,3-butadtenein the rat: the effectof Arochtor 1254, phenobarbitone or SKF 525A treatment. Toxicol. Appi.Pharmacol.,57: 79-87,1981.
18. Buchmann,A., Kuhlmann,W. D., Schwarz, M., Kunz, H. W.. Wolf, C. R., Moll,E., Friedberg,T., and Oesch. F. Regulationand expressionof four cytochromeP-450 isoenzymes, NADPH cytochrome P-450 reducÃase,the glutathionetransferasesB and C and microsomalepoxide hydrolase in preneoplasticandneoplastic lesions in rat liver. Carcmogenesis(Lond.),6: 513-521,1985.
19. Calcutt, G-, and Connors, T. A. Tumour sulphydryl and sensitivity to nitrogenmustard. Biochem. Pharmacol., 72:839-845, 1963.
20. Hirono, I. Non-proteinsulphydryl group in the original strain and sublineof theaseriestumour resistant to alkylatingagents. Nature (Lond.), 786:1059-1069,1960.
CANCER RESEARCH VOL. 46 FEBRUARY 1986
739
on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANNOUNCEMENTS
pheric genotoxicants. For further information, contact: Dr. Shahbeg S.Sandhu. Scientific Coordinator, U. S. Environmental ProtectionAgency, Genetic Toxicology Division, Mail Drop 68. Research TrianglePark, NC 27711. Telephone: (919)541-3850.
CHEMOTHERAPY FOUNDATION SYMPOSIUM VII
The Chemotherapy Foundation Symposium VII, entitled "Innovative Cancer Chemotherapy for Tomorrow," will be held in New York
City on November 12 to 14. 1986. This meeting is presented by theDivision of Medical Oncology of the Department of Medicine, theDepartment of Neoplastic Diseases, and the Page and William BlackPostgraduate School of Medicine of the Mount Sinai School of Medicine. Registration fees are $300 for physicians and $75 for staff, fellows,and other health care professionals. The symposium meets the criteriafor 20 hours in Category 1 of the Physician's Recognition Award of
the American Medical Association and for 20 cognates in FormalLearning by the American College of Obstetricians and Gynecologists.For more information, contact: Director, the Page and William BlackPostgraduate School of Medicine, One Gustave L. Levy Place, NewYork, NY 10029. Telephone: (212)650-6737 or 650-6772.
THIRTY-FIFTH ANNUAL MEETING OF THE RADIATION RESEARCH SOCIETY
The Thirty-fifth Annual Meeting of the Radiation Research Societywill take place in Atlanta, Georgia, on February 22 to 26, 1987. Formore information, contact: Radiation Research Society, 925 ChestnutStreet, Philadelphia, PA 19107. Telephone: (215)574-3153.
FIFTH INTERNATIONAL CONFERENCE ON THE ADJUVANT THERAPY OF CANCER
The Fifth International Conference on the Adjuvant Therapy ofCancer will be held from March 11 to 14, 1987, in Tucson, Arizona.The deadline for submission of abstracts is December 1, 1986. Forabstract forms and further information, contact: Mary Humphrey,Conference Coordinator, Arizona Cancer Center, University of ArizonaCollege of Medicine, Tucson, AZ 85724. Telephone: (602)626-6044.
FUTURE MEETINGS OF THE AMERICAN ASSOCIATION FOR CANCER RESEARCH
The dates and sites for future annual meetings of the AmericanAssociation for Cancer Research are: May 20 to 23, 1987, Atlanta,Georgia; May 25 to 28, 1988, New Orleans, Louisiana; May 24 to 27,1989, San Francisco, California; May 16 to 19, 1990, Washington,
DC; May 15 to 18, 1991, San Antonio, Texas; and May 13 to 16, 1992,San Diego, California.
Errata
In the article by James Carmichael et a/., entitled "Glutathione and
Glutathione Transferase Levels in Mouse Granulocytes following Cy-clophosphamide Administration," which appeared in the February issue
(Cancer Res., 46: 735-739), it should be noted that part of the workwas carried out at the Imperial Cancer Research Fund Medical Oncology Unit, Western General Hospital, Edinburgh, United Kingdom.
Due to an error in the computer program which the authors used tocalculate intracellular polyamine levels from high performance liquidchromatography chromatograms, the data regarding the spermine content of HT-29 cells are incorrect in the article by Jerome Seidenfeld etal., entitled "Altered Cell Cycle Phase Distributions in Cultured Human
Carcinoma Cells Partially Depleted of Polyamines by Treatment withDifluoromethylornithine," which appeared in the January issue (Cancer
Res., 46: 47-53). For the same reason, all of the data regardingspermine content are incorrect in another article by the same authors,entitled "Reduced Cytocidal Efficacy for Adriamycin in Cultured Hu
man Carcinoma Cells Depleted of Polyamines by Difluoromethylornithine Treatment," which appeared in the March issue (Cancer Res., 46:
1155-1159).As a result of this error in the computer program, the HT-29
spermine content presented in Table 2 (final column, lines 5-8) onpage 51 of the January article, and all of the data regarding sperminecontent in Table 2 (final column) on page 1157 of the March article,are only 50% of the true values.
These corrections do not change the relative decline in sperminecontent caused by difluoromethylornithine treatment in either of thesestudies, nor do they alter the authors' conclusions regarding the effect
of difluoromethylornithine treatment and polyamine depletion on cellcycle phase distributions in the January article and on the efficacy ofAdriamycin in the March article.
Recent Deaths
We regret to announce the deaths of the following members of theAmerican Association for Cancer Research: Dr. Henry G. Kaplan,Mercer Island, Washington; Dr. Nathan O. Kaplan, Chemistry Department, University of California, La Jolla, California; Dr. Clara J.Lynch, Chevy Chase, Maryland; Dr. Pablo Mori-Chavez (emeritus),Laboratorio de Oncologia, Universidad Peruana Cayetano Heredia,Lima, Perú;and Dr. Edward A. Smuckler, Department of Pathology,University of California, School of Medicine, San Francisco, California.
3748

1986;46:735-739. Cancer Res James Carmichael, David J. Adams, John Ansell, et al. Granulocytes following Cyclophosphamide AdministrationGlutathione and Glutathione Transferase Levels in Mouse
Updated version
http://cancerres.aacrjournals.org/content/46/2/735
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/46/2/735To request permission to re-use all or part of this article, use this link
on April 4, 2019. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from


















