
Glioma therapy and real-time imaging of neural precursor cell migration and tumor regression
8
Glioma Therapy and Real-Time Imaging of Neural Precursor Cell Migration and Tumor Regression Khalid Shah, PhD, 1,2 Emilie Bureau, MSc, 1 Dong-Eog Kim, MD, 2 Katherine Yang, BSc, 1 Yi Tang, MD, 2 Ralph Weissleder, MD, PhD, 2 and Xandra O. Breakefield, PhD 1,2 Despite many refinements in current therapeutic strategies, the overall prognosis for a patient with glioblastoma is dismal. Neural precursor cells (NPCs) are capable of tracking glioma tumors and thus could be used to deliver thera- peutic molecules. We have engineered mouse NPCs to deliver a secreted form of tumor necrosis factor–related apoptosis– inducing ligand (S-TRAIL); S-TRAIL is optimized to selectively kill neoplastic cells. Furthermore, we have developed means to simultaneously monitor both the migration of NSCs toward gliomas and the changes in glioma burden in real time. Using a highly malignant human glioma model expressing Renilla luciferase (Rluc), intracranially implanted NPC- FL-sTRAIL expressing both firefly luciferase (Fluc) and S-TRAIL was shown to migrate into the tumors and have profound antitumor effects. These studies demonstrate the potential of NPCs as therapeutically effective delivery vehicles for the treatment of gliomas and also provide important tools to evaluate the migration of NPCs and changes in glioma burden in vivo. Ann Neurol 2005;57:34 – 41 Glioblastomas are the most common primary malig- nant brain tumors. Over the past 20 years, the inci- dence of gliomas has increased considerably and the median survival for patients with glioblastomas is less than 2 years. 1 Gliomas are diffuse and infiltrating with no clear border between normal brain and tumor. Sur- gical resection is almost always followed by regrowth of tumor cells residing in adjacent regions of normal brain. New therapies are needed that specifically target tumor cells, especially those cells that have escaped the main tumor mass. Although oncolytic viral therapy ap- pears promising, 2 it is limited by relatively short sur- vival of viral vectors caused by immune rejection and by their difficulty in reaching glioblastoma cells infil- trating the brain parenchyma. Neural precursor cells (NPCs) have the ability to mi- grate widely throughout diseased and aged brain 3,4 and to differentiate into neurons, glia, and oligodendro- glia. 5 NPCs also can be used to deliver therapeutic pro- teins, particularly those with selective antineoplastic ef- fects. One such protein that has been used in this context is tumor necrosis factor–related apoptosis- inducing ligand (TRAIL) 6 which selectively induces apoptosis in transformed cells 7–9 and compared with other proapoptotic proteins, such as FasL, has little toxic side effects on normal cells. 10 The presence of death domain containing TRAIL receptors in trans- formed cells such as gliomas makes them susceptible to TRAIL-mediated apoptosis. 11,12 In contrast, the ab- sence of such receptors in normal cells such as NPCs makes them resistant to TRAIL-mediated apoptosis, thus allowing them to be used as therapeutic vehicles to deliver TRAIL. We have recently engineered a secretable form of TRAIL (S-TRAIL) and shown that it has an enhanced apoptosis-inducing and bystander effect for tumor cells. 13 To extend the release time of S-TRAIL and de- liver it directly to invasive tumor cells, we evaluated NPCs expressing S-TRAIL as a unique delivery modal- ity for brain tumor therapy. Quantitative evaluation was facilitated by engineering cell lines with lumines- cent and fluorescent transgenes and by the use of dual bioluminescence imaging to track growth of highly ma- lignant gliomas, Gli36EGFR, and migration of NPCs. Results demonstrate that intracranially im- From the 1 Department of Neurology, and 2 Center for Molecular Imaging Research (CMIR), Massachusetts General Hospital, Har- vard Medical School, Boston, MA. Received Jul 2, 2004, and in revised form Sep 1. Accepted for pub- lication Sep 1, 2004. Published online Dec 27, 2004 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/ana.20306 Address correspondence to Dr Shah, 5421CMIR, Massachusetts General Hospital-East, 13th Street, Building 149, Charlestown, MA 02129. E-mail: [email protected] 34 © 2004 American Neurological Association Published by Wiley-Liss, Inc., through Wiley Subscription Services
-
Upload
khalid-shah -
Category
Documents
-
view
213 -
download
0
Transcript of Glioma therapy and real-time imaging of neural precursor cell migration and tumor regression

Glioma Therapy and Real-Time Imaging ofNeural Precursor Cell Migration and
Tumor RegressionKhalid Shah, PhD,1,2 Emilie Bureau, MSc,1 Dong-Eog Kim, MD,2 Katherine Yang, BSc,1 Yi Tang, MD,2
Ralph Weissleder, MD, PhD,2 and Xandra O. Breakefield, PhD1,2
Despite many refinements in current therapeutic strategies, the overall prognosis for a patient with glioblastoma isdismal. Neural precursor cells (NPCs) are capable of tracking glioma tumors and thus could be used to deliver thera-peutic molecules. We have engineered mouse NPCs to deliver a secreted form of tumor necrosis factor–related apoptosis–inducing ligand (S-TRAIL); S-TRAIL is optimized to selectively kill neoplastic cells. Furthermore, we have developedmeans to simultaneously monitor both the migration of NSCs toward gliomas and the changes in glioma burden in realtime. Using a highly malignant human glioma model expressing Renilla luciferase (Rluc), intracranially implanted NPC-FL-sTRAIL expressing both firefly luciferase (Fluc) and S-TRAIL was shown to migrate into the tumors and haveprofound antitumor effects. These studies demonstrate the potential of NPCs as therapeutically effective delivery vehiclesfor the treatment of gliomas and also provide important tools to evaluate the migration of NPCs and changes in gliomaburden in vivo.
Ann Neurol 2005;57:34–41
Glioblastomas are the most common primary malig-nant brain tumors. Over the past 20 years, the inci-dence of gliomas has increased considerably and themedian survival for patients with glioblastomas is lessthan 2 years.1 Gliomas are diffuse and infiltrating withno clear border between normal brain and tumor. Sur-gical resection is almost always followed by regrowth oftumor cells residing in adjacent regions of normalbrain. New therapies are needed that specifically targettumor cells, especially those cells that have escaped themain tumor mass. Although oncolytic viral therapy ap-pears promising,2 it is limited by relatively short sur-vival of viral vectors caused by immune rejection andby their difficulty in reaching glioblastoma cells infil-trating the brain parenchyma.
Neural precursor cells (NPCs) have the ability to mi-grate widely throughout diseased and aged brain3,4 andto differentiate into neurons, glia, and oligodendro-glia.5 NPCs also can be used to deliver therapeutic pro-teins, particularly those with selective antineoplastic ef-fects. One such protein that has been used in thiscontext is tumor necrosis factor–related apoptosis-inducing ligand (TRAIL)6 which selectively induces
apoptosis in transformed cells7–9 and compared withother proapoptotic proteins, such as FasL, has littletoxic side effects on normal cells.10 The presence ofdeath domain containing TRAIL receptors in trans-formed cells such as gliomas makes them susceptible toTRAIL-mediated apoptosis.11,12 In contrast, the ab-sence of such receptors in normal cells such as NPCsmakes them resistant to TRAIL-mediated apoptosis,thus allowing them to be used as therapeutic vehiclesto deliver TRAIL.
We have recently engineered a secretable form ofTRAIL (S-TRAIL) and shown that it has an enhancedapoptosis-inducing and bystander effect for tumorcells.13 To extend the release time of S-TRAIL and de-liver it directly to invasive tumor cells, we evaluatedNPCs expressing S-TRAIL as a unique delivery modal-ity for brain tumor therapy. Quantitative evaluationwas facilitated by engineering cell lines with lumines-cent and fluorescent transgenes and by the use of dualbioluminescence imaging to track growth of highly ma-lignant gliomas, Gli36�EGFR, and migration ofNPCs. Results demonstrate that intracranially im-
From the 1Department of Neurology, and 2Center for MolecularImaging Research (CMIR), Massachusetts General Hospital, Har-vard Medical School, Boston, MA.
Received Jul 2, 2004, and in revised form Sep 1. Accepted for pub-lication Sep 1, 2004.
Published online Dec 27, 2004 in Wiley InterScience(www.interscience.wiley.com). DOI: 10.1002/ana.20306
Address correspondence to Dr Shah, 5421CMIR, MassachusettsGeneral Hospital-East, 13th Street, Building 149, Charlestown, MA02129. E-mail: [email protected]
34 © 2004 American Neurological AssociationPublished by Wiley-Liss, Inc., through Wiley Subscription Services

planted S-TRAIL–secreting NPCs have significant an-titumor effects.
Materials and MethodsGeneration of Tumor Necrosis Factor–RelatedApoptosis–Inducing Ligand AmpliconsThe cDNA sequence encoding amino acids 114 to 281TRAIL fused to a cDNA fragment encoding a leucine zipperand the extracellular domain of the hFlt3 ligand were clonedinto the multiple cloning site (MCS) of the pKSR2–Herpessimplex virus type 1 amplicon resulting in the S-TRAIL con-struct.13 Similarly the cDNA fragment encoding Renilla lu-ciferase (Rluc) was cloned into the MCS of the pHGCX am-plicon resulting in the Rluc construct.6
Cell Lines and Cell CultureNPC-FL is a vmyc immortalized mouse neuroprogenitor cellline (NPC line) derived from the C17.2 cell line14 whichstably expresses �-galactosidase (�-gal 3) and firefly luciferase(Fluc).15 NPC-FLs were cotransfected with the S-TRAILamplicon plasmid and pcDNA3.1 Hyg (�) (Invitrogen,Carlsbad, CA). Resulting stable S-TRAIL–positive cloneswere selected using 200�g/ml hygromycin in the growth me-dium, isolated, and checked for S-TRAIL secretion in themedium by ELISA (described below). To create a glioma cellline expressing Rluc, we stably cotransfected highly malig-nant human primary glioma cells, Gli36�EGFR,16,17 withthe Rluc amplicon6 and pcDNA3.1 Hyg (�) (Invitrogen) asdescribed above. The clone showing the highest Rluc activity(see luciferase activity section below), Gli36-RL cell line (alsogreen fluorescent protein (GFP)-positive), was used for fur-ther studies.
Cell Viability Assay and ELISAGli36�EGFR cells and NPC-FLs were plated in 96-well Pri-maria plates (Falcon, Bedford, MA) at a concentration of1.5 � 104 cells/well and 3 � 103 cells/well, respectively.Twenty-four hours later, cells were incubated in a mediumcontaining 0 to 240ng/ml of recombinant TRAIL (rTRAIL;PeproTech, Rocky Hill, NJ). Forty-eight hours after TRAILaddition, cell viability was assessed using 10�l WST (Tetra-zolium salt; Roche, Indianapolis, IN) in a final volume of100�l growth medium. After 4 hours of incubation, plateswere read at 450nm using a Vmax kinetic microplate reader(Molecular Devices, Sunnyvale, CA). TRAIL concentrationin the conditioned culture medium from NPC-FLs or NPC-FL-sTRAILs was measured by ELISA with the TRAIL Im-munoassay Kit (Biosource International, Camarillo, CA) ac-cording to manufacturer’s protocol using recombinanthuman (h) TRAIL expressed in Escherichia coli as a standard.
ImmunocytochemistryTo assess transgene expression and cell type, we fixed NPC-FL-sTRAILs and permeabilized and incubated them with:(1) a rabbit anti–firefly luciferase antibody (1:100, MolecularProbes, Eugene, OR); (2) a rabbit anti–�-gal antibody (1;100; Research Products International, Mount Prospect, IL);or (3) a antinestin monoclonal antibody (MAb) (1:200,Chemicon International, Temecula, CA) for 1 hour at 37°C.
Cells then were washed and incubated with either goat anti–rabbit Alexa dye 488nm (1, 2) or goat anti–mouse Alexa dye496nm (3) conjugated secondary antibodies (MolecularProbes, Eugene, OR) for 1 hour, then washed, mounted, andexamined microscopically.
To assess apoptotic effects of S-TRAIL secreted fromNPCs on glioma cells, we grew either 1 � 106 NPC-FLs orNPC-FL-sTRAILs in a total volume of 1ml, and 72 hourslater 400�l of either of the growth medium was added to thecultures of 2 � 105 Gli36�EGFR cells. After 48 hours, cellswere fixed and immunostained using a rabbit anti–caspase-3primary antibody (Cell Signaling, Beverly, MA) and a goatanti–rabbit Alexa dye 488nm-conjugated secondary anti-body, as described above.
Sodium Dodecyl Sulfate Polyacrylamide GelElectrophoresis (SDS-PAGE) and Western blottingGli36�EGFR, NPC-FL, or NPC-FL-sTRAIL cells werelysed, and proteins were denatured, resolved by SDS-PAGE,transferred to membranes, and blocked, as described previ-ously13 using rabbit anti–TRAIL-R1 (1:500; AXXORA,LLC, San Diego, CA) or goat anti–TRAIL-R2 (1:500; AlexisBiochemicals, San Diego, CA) antibodies.
Cell ImplantationAthymic nude mice (nu/nu; 6–7 weeks old; Charles RiverLaboratories, Wilmington, MA) were stereotactically im-planted with Gli36-RL glioblastoma cells and NPC-FL orNPC-FL-sTRAILs. Two to 3 days after glioma cell implan-tations, mice were imaged for Rluc activity before their dis-tribution into various experimental groups as described be-low. All animal protocols were approved by an institutionalreview board. Three sets of experiments were performed. (1)For studying migration of NPC-FL-sTRAILs towardGli36-RL gliomas, Gli36-RL cells (0.7 � 106 in 4�lphosphate-buffered saline [PBS]) were implanted stereotacti-cally into the right frontal lobe of nude mice (n �20) and 2days later, NPC-FL-sTRAIL (n �8) or NPC-FLs (n �8) (1�106 in 4�l PBS) or saline (n �4) were injected into theleft frontal lobe of tumor-bearing mice. NPC-FL-sTRAILswere also implanted into the left frontal lobe of non–tumor-bearing mice (n �4). (2) For studying the effect of S-TRAILsecreted by NPCs within the tumors 0.25 � 106 Gli36-RLcells (in 4�l PBS; n �5) or a mix of 0.25 � 106 Gli36-RLcells with 0.75 �106 NPC-FL-sTRAILs (in 5�l PBS; n �5)or NPC-FLs (in 5�l PBS; n �5) were implanted stereotac-tically into the right frontal lobe of nude mice. (3) To de-termine the influence of S-TRAIL secreted by NPC-FL-sTRAILs implanted in the close vicinity of the gliomas, weimplanted Gli36-RL cells (0.25 �106 in 4�l PBS) into theright frontal lobe of nude mice (n �20), and 3 days laterNPC-FL-sTRAILs or NPC-FLs (0.75 � 106 in 4�l PBS)were stereotactically implanted into the right frontal lobe.
In Vivo Photonflux ImagingMice were imaged for Rluc activity by injecting coelentera-zine (100�g/animal in 150�l saline) intravenously via thetail vein, and 5 minutes later photon counts were recordedfor 5 minutes using a cryogenically cooled high efficiencycharge-coupled device camera system (Roper Scientific,
Shah et al: TRAIL-Secreting NPCs 35
Trenton, NJ). For Fluc imaging, mice were given intraperi-toneal injection of D-luciferin (4.5mg/animal in 150�l sa-line), and photon counts were recorded 10 minutes afterD-luciferin administration. Postprocessing and visualizationwere performed as described previously.6 Mice were imagedevery 5 to 7 days for Fluc and or Rluc activity.
Tissue ProcessingImmediately after the last imaging session, mice were sacri-ficed, and brains were immersed in Tissue Freezing Medium(Triangle Biomedical Sciences, Durham, NC) on dry ice and30�m coronal brain sections were cut. For X-gal staining,brain sections on slides were incubated overnight in X-galsolution. After washing slides were counterstained with eosin,dried, and mounted for microscopy. Parallel sections alsowere visualized for GFP fluorescence on a fluorescence mi-croscope (Axiovert, Carl Zeiss, Ag, Germany). Brain sectionsfrom NPC-Fl-sTRAIL cell implanted mice were also stainedwith caspase-3 antibody (Cell Signaling Technology, Beverly,MA) according to manufacturer’s protocol and counter-stained with eosin.
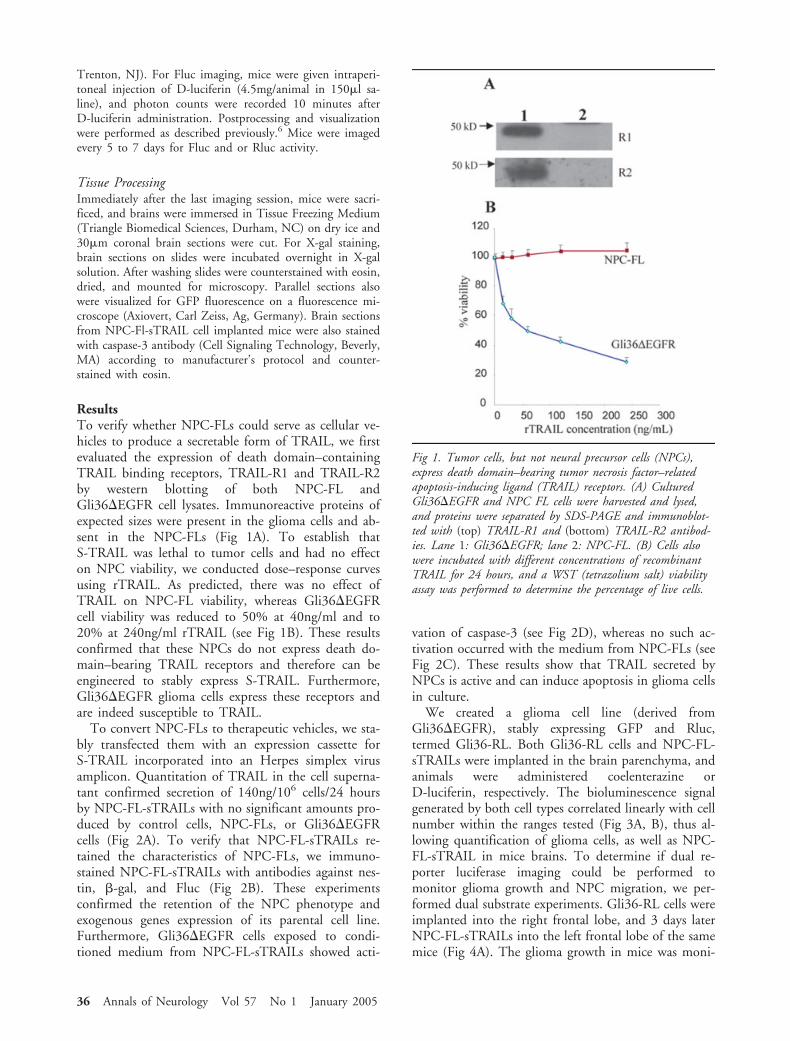
ResultsTo verify whether NPC-FLs could serve as cellular ve-hicles to produce a secretable form of TRAIL, we firstevaluated the expression of death domain–containingTRAIL binding receptors, TRAIL-R1 and TRAIL-R2by western blotting of both NPC-FL andGli36�EGFR cell lysates. Immunoreactive proteins ofexpected sizes were present in the glioma cells and ab-sent in the NPC-FLs (Fig 1A). To establish thatS-TRAIL was lethal to tumor cells and had no effecton NPC viability, we conducted dose–response curvesusing rTRAIL. As predicted, there was no effect ofTRAIL on NPC-FL viability, whereas Gli36�EGFRcell viability was reduced to 50% at 40ng/ml and to20% at 240ng/ml rTRAIL (see Fig 1B). These resultsconfirmed that these NPCs do not express death do-main–bearing TRAIL receptors and therefore can beengineered to stably express S-TRAIL. Furthermore,Gli36�EGFR glioma cells express these receptors andare indeed susceptible to TRAIL.
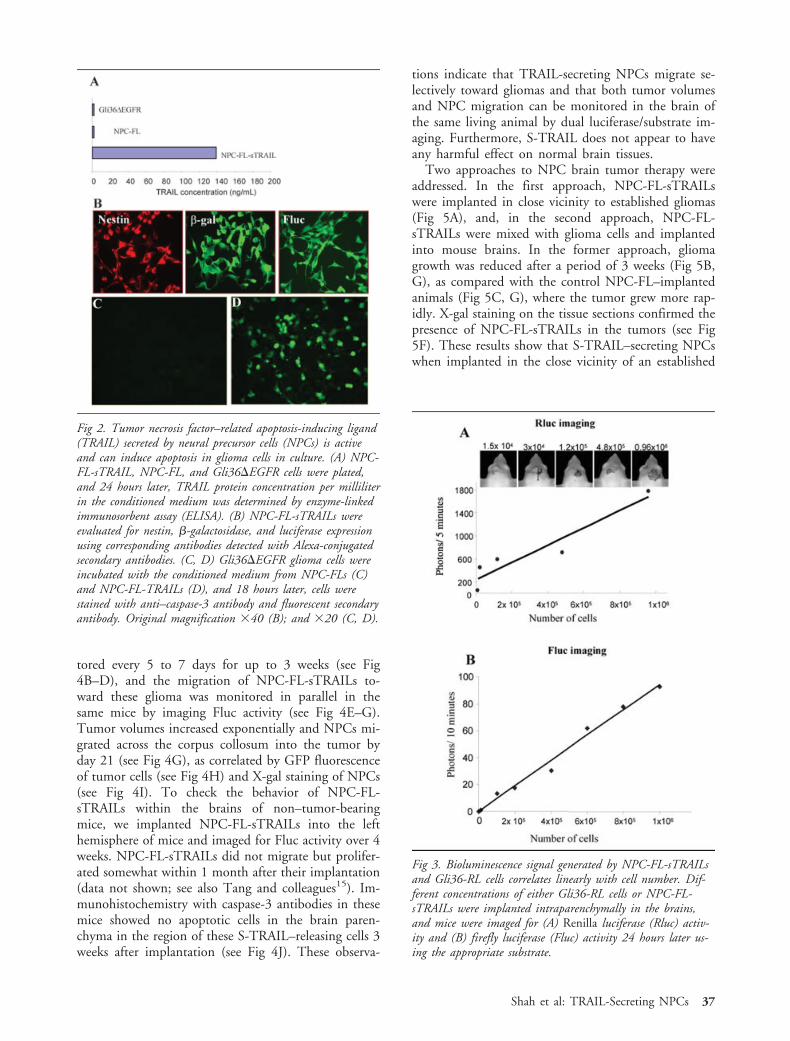
To convert NPC-FLs to therapeutic vehicles, we sta-bly transfected them with an expression cassette forS-TRAIL incorporated into an Herpes simplex virusamplicon. Quantitation of TRAIL in the cell superna-tant confirmed secretion of 140ng/106 cells/24 hoursby NPC-FL-sTRAILs with no significant amounts pro-duced by control cells, NPC-FLs, or Gli36�EGFRcells (Fig 2A). To verify that NPC-FL-sTRAILs re-tained the characteristics of NPC-FLs, we immuno-stained NPC-FL-sTRAILs with antibodies against nes-tin, �-gal, and Fluc (Fig 2B). These experimentsconfirmed the retention of the NPC phenotype andexogenous genes expression of its parental cell line.Furthermore, Gli36�EGFR cells exposed to condi-tioned medium from NPC-FL-sTRAILs showed acti-
vation of caspase-3 (see Fig 2D), whereas no such ac-tivation occurred with the medium from NPC-FLs (seeFig 2C). These results show that TRAIL secreted byNPCs is active and can induce apoptosis in glioma cellsin culture.
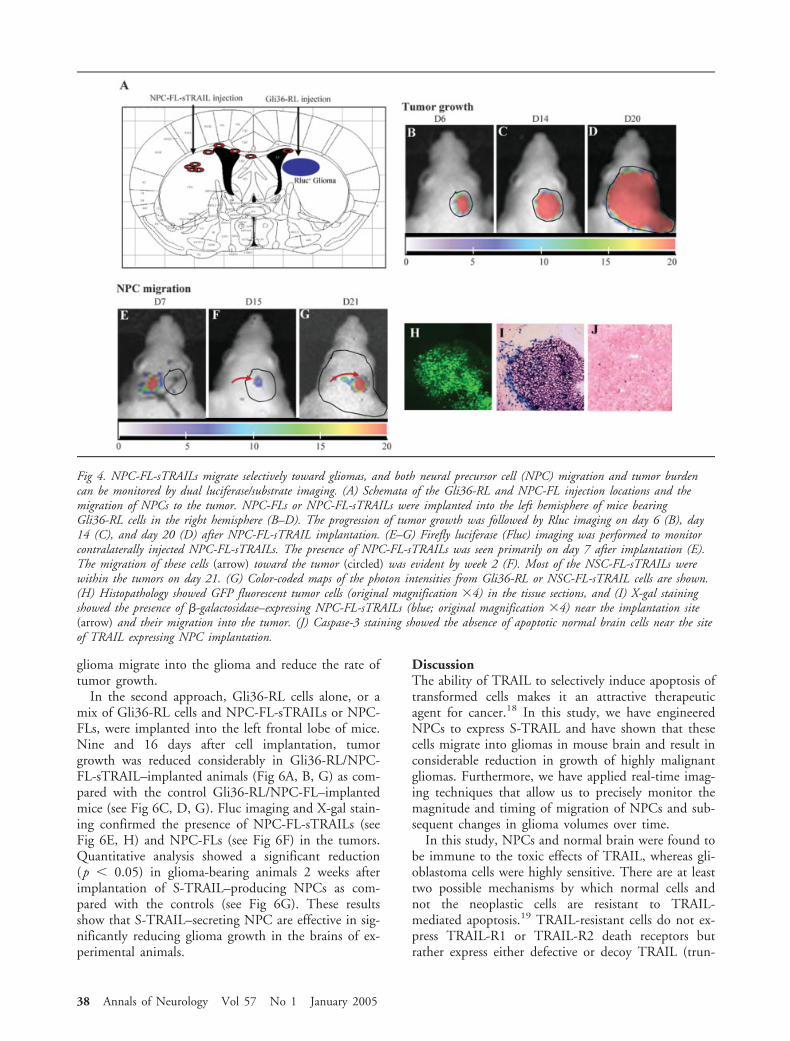
We created a glioma cell line (derived fromGli36�EGFR), stably expressing GFP and Rluc,termed Gli36-RL. Both Gli36-RL cells and NPC-FL-sTRAILs were implanted in the brain parenchyma, andanimals were administered coelenterazine orD-luciferin, respectively. The bioluminescence signalgenerated by both cell types correlated linearly with cellnumber within the ranges tested (Fig 3A, B), thus al-lowing quantification of glioma cells, as well as NPC-FL-sTRAIL in mice brains. To determine if dual re-porter luciferase imaging could be performed tomonitor glioma growth and NPC migration, we per-formed dual substrate experiments. Gli36-RL cells wereimplanted into the right frontal lobe, and 3 days laterNPC-FL-sTRAILs into the left frontal lobe of the samemice (Fig 4A). The glioma growth in mice was moni-
Fig 1. Tumor cells, but not neural precursor cells (NPCs),express death domain–bearing tumor necrosis factor–relatedapoptosis-inducing ligand (TRAIL) receptors. (A) CulturedGli36�EGFR and NPC FL cells were harvested and lysed,and proteins were separated by SDS-PAGE and immunoblot-ted with (top) TRAIL-R1 and (bottom) TRAIL-R2 antibod-ies. Lane 1: Gli36�EGFR; lane 2: NPC-FL. (B) Cells alsowere incubated with different concentrations of recombinantTRAIL for 24 hours, and a WST (tetrazolium salt) viabilityassay was performed to determine the percentage of live cells.
36 Annals of Neurology Vol 57 No 1 January 2005
tored every 5 to 7 days for up to 3 weeks (see Fig4B–D), and the migration of NPC-FL-sTRAILs to-ward these glioma was monitored in parallel in thesame mice by imaging Fluc activity (see Fig 4E–G).Tumor volumes increased exponentially and NPCs mi-grated across the corpus collosum into the tumor byday 21 (see Fig 4G), as correlated by GFP fluorescenceof tumor cells (see Fig 4H) and X-gal staining of NPCs(see Fig 4I). To check the behavior of NPC-FL-sTRAILs within the brains of non–tumor-bearingmice, we implanted NPC-FL-sTRAILs into the lefthemisphere of mice and imaged for Fluc activity over 4weeks. NPC-FL-sTRAILs did not migrate but prolifer-ated somewhat within 1 month after their implantation(data not shown; see also Tang and colleagues15). Im-munohistochemistry with caspase-3 antibodies in thesemice showed no apoptotic cells in the brain paren-chyma in the region of these S-TRAIL–releasing cells 3weeks after implantation (see Fig 4J). These observa-
tions indicate that TRAIL-secreting NPCs migrate se-lectively toward gliomas and that both tumor volumesand NPC migration can be monitored in the brain ofthe same living animal by dual luciferase/substrate im-aging. Furthermore, S-TRAIL does not appear to haveany harmful effect on normal brain tissues.
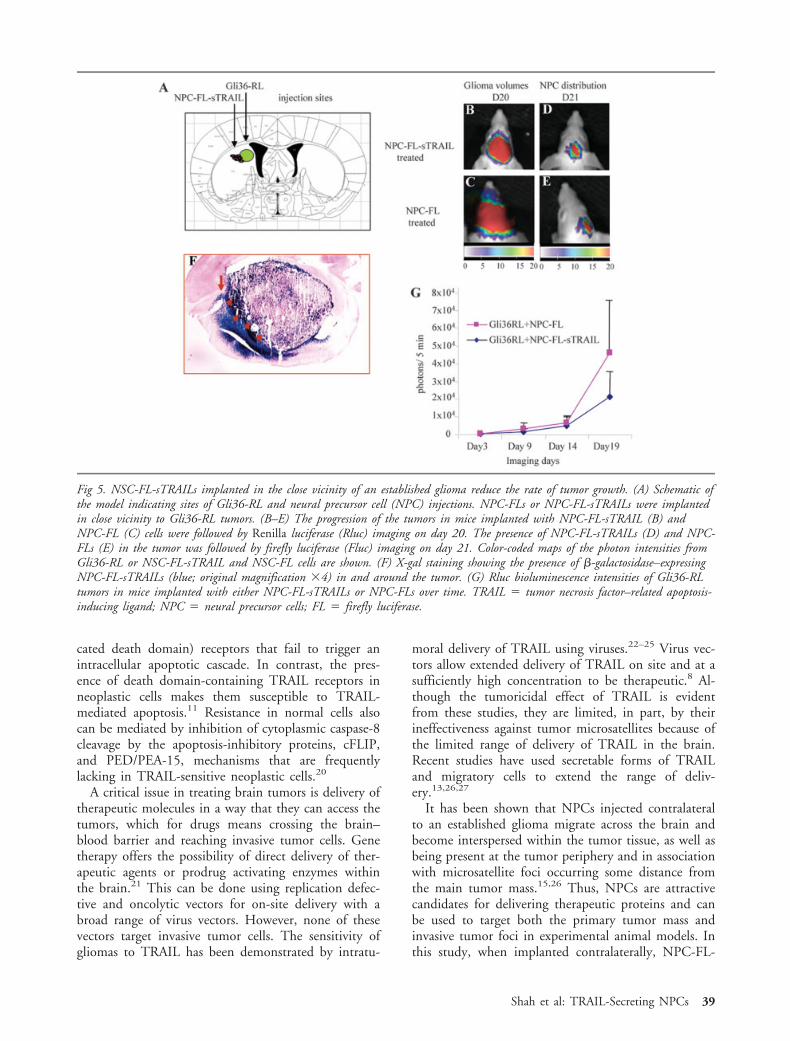
Two approaches to NPC brain tumor therapy wereaddressed. In the first approach, NPC-FL-sTRAILswere implanted in close vicinity to established gliomas(Fig 5A), and, in the second approach, NPC-FL-sTRAILs were mixed with glioma cells and implantedinto mouse brains. In the former approach, gliomagrowth was reduced after a period of 3 weeks (Fig 5B,G), as compared with the control NPC-FL–implantedanimals (Fig 5C, G), where the tumor grew more rap-idly. X-gal staining on the tissue sections confirmed thepresence of NPC-FL-sTRAILs in the tumors (see Fig5F). These results show that S-TRAIL–secreting NPCswhen implanted in the close vicinity of an established
Fig 2. Tumor necrosis factor–related apoptosis-inducing ligand(TRAIL) secreted by neural precursor cells (NPCs) is activeand can induce apoptosis in glioma cells in culture. (A) NPC-FL-sTRAIL, NPC-FL, and Gli36�EGFR cells were plated,and 24 hours later, TRAIL protein concentration per milliliterin the conditioned medium was determined by enzyme-linkedimmunosorbent assay (ELISA). (B) NPC-FL-sTRAILs wereevaluated for nestin, �-galactosidase, and luciferase expressionusing corresponding antibodies detected with Alexa-conjugatedsecondary antibodies. (C, D) Gli36�EGFR glioma cells wereincubated with the conditioned medium from NPC-FLs (C)and NPC-FL-TRAILs (D), and 18 hours later, cells werestained with anti–caspase-3 antibody and fluorescent secondaryantibody. Original magnification �40 (B); and �20 (C, D).
Fig 3. Bioluminescence signal generated by NPC-FL-sTRAILsand Gli36-RL cells correlates linearly with cell number. Dif-ferent concentrations of either Gli36-RL cells or NPC-FL-sTRAILs were implanted intraparenchymally in the brains,and mice were imaged for (A) Renilla luciferase (Rluc) activ-ity and (B) firefly luciferase (Fluc) activity 24 hours later us-ing the appropriate substrate.
Shah et al: TRAIL-Secreting NPCs 37
glioma migrate into the glioma and reduce the rate oftumor growth.
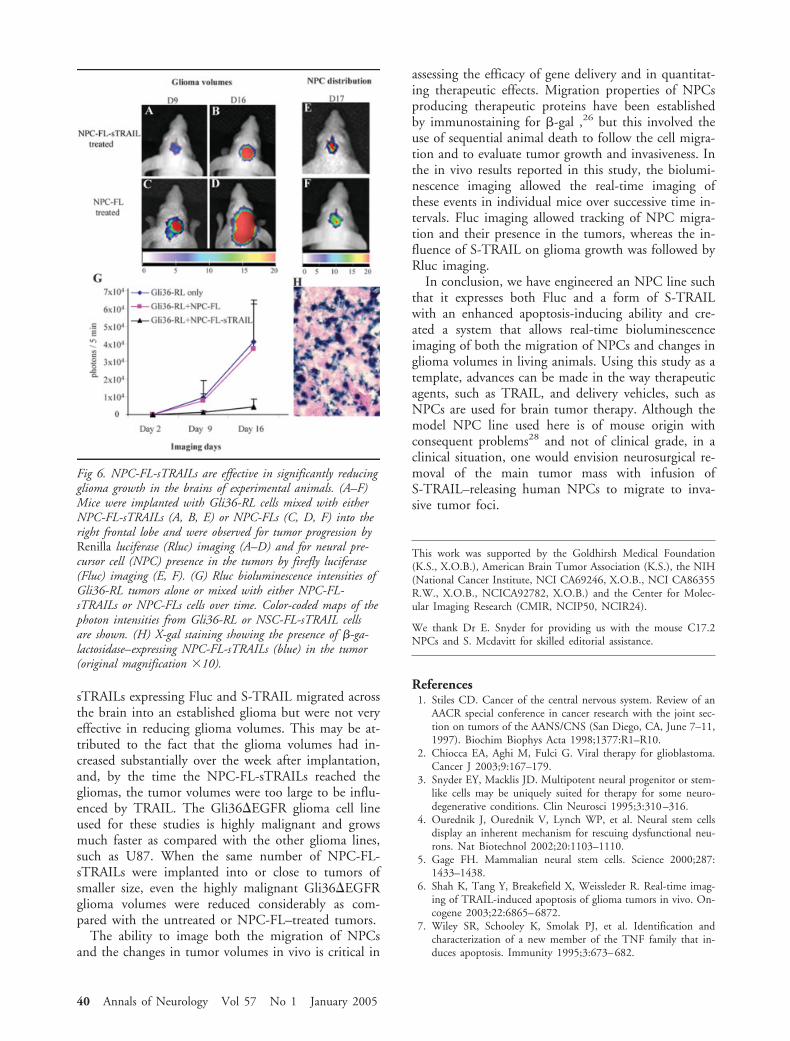
In the second approach, Gli36-RL cells alone, or amix of Gli36-RL cells and NPC-FL-sTRAILs or NPC-FLs, were implanted into the left frontal lobe of mice.Nine and 16 days after cell implantation, tumorgrowth was reduced considerably in Gli36-RL/NPC-FL-sTRAIL–implanted animals (Fig 6A, B, G) as com-pared with the control Gli36-RL/NPC-FL–implantedmice (see Fig 6C, D, G). Fluc imaging and X-gal stain-ing confirmed the presence of NPC-FL-sTRAILs (seeFig 6E, H) and NPC-FLs (see Fig 6F) in the tumors.Quantitative analysis showed a significant reduction(p � 0.05) in glioma-bearing animals 2 weeks afterimplantation of S-TRAIL–producing NPCs as com-pared with the controls (see Fig 6G). These resultsshow that S-TRAIL–secreting NPC are effective in sig-nificantly reducing glioma growth in the brains of ex-perimental animals.
DiscussionThe ability of TRAIL to selectively induce apoptosis oftransformed cells makes it an attractive therapeuticagent for cancer.18 In this study, we have engineeredNPCs to express S-TRAIL and have shown that thesecells migrate into gliomas in mouse brain and result inconsiderable reduction in growth of highly malignantgliomas. Furthermore, we have applied real-time imag-ing techniques that allow us to precisely monitor themagnitude and timing of migration of NPCs and sub-sequent changes in glioma volumes over time.
In this study, NPCs and normal brain were found tobe immune to the toxic effects of TRAIL, whereas gli-oblastoma cells were highly sensitive. There are at leasttwo possible mechanisms by which normal cells andnot the neoplastic cells are resistant to TRAIL-mediated apoptosis.19 TRAIL-resistant cells do not ex-press TRAIL-R1 or TRAIL-R2 death receptors butrather express either defective or decoy TRAIL (trun-
Fig 4. NPC-FL-sTRAILs migrate selectively toward gliomas, and both neural precursor cell (NPC) migration and tumor burdencan be monitored by dual luciferase/substrate imaging. (A) Schemata of the Gli36-RL and NPC-FL injection locations and themigration of NPCs to the tumor. NPC-FLs or NPC-FL-sTRAILs were implanted into the left hemisphere of mice bearingGli36-RL cells in the right hemisphere (B–D). The progression of tumor growth was followed by Rluc imaging on day 6 (B), day14 (C), and day 20 (D) after NPC-FL-sTRAIL implantation. (E–G) Firefly luciferase (Fluc) imaging was performed to monitorcontralaterally injected NPC-FL-sTRAILs. The presence of NPC-FL-sTRAILs was seen primarily on day 7 after implantation (E).The migration of these cells (arrow) toward the tumor (circled) was evident by week 2 (F). Most of the NSC-FL-sTRAILs werewithin the tumors on day 21. (G) Color-coded maps of the photon intensities from Gli36-RL or NSC-FL-sTRAIL cells are shown.(H) Histopathology showed GFP fluorescent tumor cells (original magnification �4) in the tissue sections, and (I) X-gal stainingshowed the presence of �-galactosidase–expressing NPC-FL-sTRAILs (blue; original magnification �4) near the implantation site(arrow) and their migration into the tumor. (J) Caspase-3 staining showed the absence of apoptotic normal brain cells near the siteof TRAIL expressing NPC implantation.
38 Annals of Neurology Vol 57 No 1 January 2005
cated death domain) receptors that fail to trigger anintracellular apoptotic cascade. In contrast, the pres-ence of death domain-containing TRAIL receptors inneoplastic cells makes them susceptible to TRAIL-mediated apoptosis.11 Resistance in normal cells alsocan be mediated by inhibition of cytoplasmic caspase-8cleavage by the apoptosis-inhibitory proteins, cFLIP,and PED/PEA-15, mechanisms that are frequentlylacking in TRAIL-sensitive neoplastic cells.20
A critical issue in treating brain tumors is delivery oftherapeutic molecules in a way that they can access thetumors, which for drugs means crossing the brain–blood barrier and reaching invasive tumor cells. Genetherapy offers the possibility of direct delivery of ther-apeutic agents or prodrug activating enzymes withinthe brain.21 This can be done using replication defec-tive and oncolytic vectors for on-site delivery with abroad range of virus vectors. However, none of thesevectors target invasive tumor cells. The sensitivity ofgliomas to TRAIL has been demonstrated by intratu-
moral delivery of TRAIL using viruses.22–25 Virus vec-tors allow extended delivery of TRAIL on site and at asufficiently high concentration to be therapeutic.8 Al-though the tumoricidal effect of TRAIL is evidentfrom these studies, they are limited, in part, by theirineffectiveness against tumor microsatellites because ofthe limited range of delivery of TRAIL in the brain.Recent studies have used secretable forms of TRAILand migratory cells to extend the range of deliv-ery.13,26,27
It has been shown that NPCs injected contralateralto an established glioma migrate across the brain andbecome interspersed within the tumor tissue, as well asbeing present at the tumor periphery and in associationwith microsatellite foci occurring some distance fromthe main tumor mass.15,26 Thus, NPCs are attractivecandidates for delivering therapeutic proteins and canbe used to target both the primary tumor mass andinvasive tumor foci in experimental animal models. Inthis study, when implanted contralaterally, NPC-FL-
Fig 5. NSC-FL-sTRAILs implanted in the close vicinity of an established glioma reduce the rate of tumor growth. (A) Schematic ofthe model indicating sites of Gli36-RL and neural precursor cell (NPC) injections. NPC-FLs or NPC-FL-sTRAILs were implantedin close vicinity to Gli36-RL tumors. (B–E) The progression of the tumors in mice implanted with NPC-FL-sTRAIL (B) andNPC-FL (C) cells were followed by Renilla luciferase (Rluc) imaging on day 20. The presence of NPC-FL-sTRAILs (D) and NPC-FLs (E) in the tumor was followed by firefly luciferase (Fluc) imaging on day 21. Color-coded maps of the photon intensities fromGli36-RL or NSC-FL-sTRAIL and NSC-FL cells are shown. (F) X-gal staining showing the presence of �-galactosidase–expressingNPC-FL-sTRAILs (blue; original magnification �4) in and around the tumor. (G) Rluc bioluminescence intensities of Gli36-RLtumors in mice implanted with either NPC-FL-sTRAILs or NPC-FLs over time. TRAIL � tumor necrosis factor–related apoptosis-inducing ligand; NPC � neural precursor cells; FL � firefly luciferase.
Shah et al: TRAIL-Secreting NPCs 39
sTRAILs expressing Fluc and S-TRAIL migrated acrossthe brain into an established glioma but were not veryeffective in reducing glioma volumes. This may be at-tributed to the fact that the glioma volumes had in-creased substantially over the week after implantation,and, by the time the NPC-FL-sTRAILs reached thegliomas, the tumor volumes were too large to be influ-enced by TRAIL. The Gli36�EGFR glioma cell lineused for these studies is highly malignant and growsmuch faster as compared with the other glioma lines,such as U87. When the same number of NPC-FL-sTRAILs were implanted into or close to tumors ofsmaller size, even the highly malignant Gli36�EGFRglioma volumes were reduced considerably as com-pared with the untreated or NPC-FL–treated tumors.
The ability to image both the migration of NPCsand the changes in tumor volumes in vivo is critical in
assessing the efficacy of gene delivery and in quantitat-ing therapeutic effects. Migration properties of NPCsproducing therapeutic proteins have been establishedby immunostaining for �-gal ,26 but this involved theuse of sequential animal death to follow the cell migra-tion and to evaluate tumor growth and invasiveness. Inthe in vivo results reported in this study, the biolumi-nescence imaging allowed the real-time imaging ofthese events in individual mice over successive time in-tervals. Fluc imaging allowed tracking of NPC migra-tion and their presence in the tumors, whereas the in-fluence of S-TRAIL on glioma growth was followed byRluc imaging.
In conclusion, we have engineered an NPC line suchthat it expresses both Fluc and a form of S-TRAILwith an enhanced apoptosis-inducing ability and cre-ated a system that allows real-time bioluminescenceimaging of both the migration of NPCs and changes inglioma volumes in living animals. Using this study as atemplate, advances can be made in the way therapeuticagents, such as TRAIL, and delivery vehicles, such asNPCs are used for brain tumor therapy. Although themodel NPC line used here is of mouse origin withconsequent problems28 and not of clinical grade, in aclinical situation, one would envision neurosurgical re-moval of the main tumor mass with infusion ofS-TRAIL–releasing human NPCs to migrate to inva-sive tumor foci.
This work was supported by the Goldhirsh Medical Foundation(K.S., X.O.B.), American Brain Tumor Association (K.S.), the NIH(National Cancer Institute, NCI CA69246, X.O.B., NCI CA86355R.W., X.O.B., NCICA92782, X.O.B.) and the Center for Molec-ular Imaging Research (CMIR, NCIP50, NCIR24).
We thank Dr E. Snyder for providing us with the mouse C17.2NPCs and S. Mcdavitt for skilled editorial assistance.
References1. Stiles CD. Cancer of the central nervous system. Review of an
AACR special conference in cancer research with the joint sec-tion on tumors of the AANS/CNS (San Diego, CA, June 7–11,1997). Biochim Biophys Acta 1998;1377:R1–R10.
2. Chiocca EA, Aghi M, Fulci G. Viral therapy for glioblastoma.Cancer J 2003;9:167–179.
3. Snyder EY, Macklis JD. Multipotent neural progenitor or stem-like cells may be uniquely suited for therapy for some neuro-degenerative conditions. Clin Neurosci 1995;3:310–316.
4. Ourednik J, Ourednik V, Lynch WP, et al. Neural stem cellsdisplay an inherent mechanism for rescuing dysfunctional neu-rons. Nat Biotechnol 2002;20:1103–1110.
5. Gage FH. Mammalian neural stem cells. Science 2000;287:1433–1438.
6. Shah K, Tang Y, Breakefield X, Weissleder R. Real-time imag-ing of TRAIL-induced apoptosis of glioma tumors in vivo. On-cogene 2003;22:6865–6872.
7. Wiley SR, Schooley K, Smolak PJ, et al. Identification andcharacterization of a new member of the TNF family that in-duces apoptosis. Immunity 1995;3:673–682.
Fig 6. NPC-FL-sTRAILs are effective in significantly reducingglioma growth in the brains of experimental animals. (A–F)Mice were implanted with Gli36-RL cells mixed with eitherNPC-FL-sTRAILs (A, B, E) or NPC-FLs (C, D, F) into theright frontal lobe and were observed for tumor progression byRenilla luciferase (Rluc) imaging (A–D) and for neural pre-cursor cell (NPC) presence in the tumors by firefly luciferase(Fluc) imaging (E, F). (G) Rluc bioluminescence intensities ofGli36-RL tumors alone or mixed with either NPC-FL-sTRAILs or NPC-FLs cells over time. Color-coded maps of thephoton intensities from Gli36-RL or NSC-FL-sTRAIL cellsare shown. (H) X-gal staining showing the presence of �-ga-lactosidase–expressing NPC-FL-sTRAILs (blue) in the tumor(original magnification �10).
40 Annals of Neurology Vol 57 No 1 January 2005

8. Walczak H, Miller RE, Ariail K, et al. Tumoricidal activity oftumor necrosis factor-related apoptosis-inducing ligand in vivo.Nat Med 1999;5:157–163.
9. Pollack IF, Erff M, Ashkenazi A. Direct stimulation of apopto-tic signaling by soluble Apo2l/tumor necrosis factor-relatedapoptosis-inducing ligand leads to selective killing of gliomacells. Clin Cancer Res 2001;7:1362–1369.
10. Norris JS, Hyer ML, Voelkel-Johnson C, et al. The use of Fasligand, TRAIL and Bax in gene therapy of prostate cancer. CurrGene Ther 2001;1:123–136.
11. Ashkenazi A, Dixit VM. Apoptosis control by death and decoyreceptors. Curr Opin Cell Biol 1999;11:255–260.
12. LeBlanc HN, Ashkenazi A. Apo2L/TRAIL and its death anddecoy receptors. Cell Death Differ 2003;10:66–75.
13. Shah K, Tung CH, Yang K, et al. Inducible release of TRAILfusion proteins from a proapoptotic form for tumor therapy.Cancer Res 2004;64:3236–3242.
14. Ryder EF, Snyder EY, Cepko CL. Establishment and character-ization of multipotent neural cell lines using retrovirus vector-mediated oncogene transfer. J Neurobiol 1990;21:356–375.
15. Tang Y, Shah K, Messerli SM, et al. In vivo tracking of neuralprogenitor cell migration to glioblastomas. Hum Gene Ther2003;14:1247–1254.
16. Nagane M, Coufal F, Lin H, et al. A common mutant epider-mal growth factor receptor confers enhanced tumorigenicity onhuman glioblastoma cells by increasing proliferation and reduc-ing apoptosis. Cancer Res 1996;56:5079–-5086.
17. Ichikawa T, Hogemann D, Saeki Y, et al. MRI of transgeneexpression: correlation to therapeutic gene expression. Neoplasia2002;4:523–530.
18. Gliniak B, Le T. Tumor necrosis factor-related apoptosis-inducing ligand’s antitumor activity in vivo is enhanced by thechemotherapeutic agent CPT-11. Cancer Res 1999;59:6153–6158.
19. Wang S, El-Deiry WS. TRAIL and apoptosis induction byTNF-family death receptors. Oncogene 2003;22:8628–8633.
20. Xiao C, Yang BF, Asadi N, et al. Tumor necrosis factor-relatedapoptosis-inducing ligand-induced death-inducing signalingcomplex and its modulation by c-FLIP and PED/PEA-15 inglioma cells. J Biol Chem 2002;277:25020–25025.
21. Aghi M, Hochberg F, Breakefield XO. Prodrug activation en-zymes in cancer gene therapy. J Gene Med 2000;2:148–164.
22. Hao C, Beguinot F, Condorelli G, et al. Induction and intra-cellular regulation of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) mediated apotosis in human malig-nant glioma cells. Cancer Res 2001;61:1162–1170.
23. Roth W, Isenmann S, Naumann U, et al. Locoregional Apo2L/TRAIL eradicates intracranial human malignant glioma xeno-grafts in athymic mice in the absence of neurotoxicity. BiochemBiophys Res Commun 1999;265:479–483.
24. Lee J, Hampl M, Albert P, Fine HA. Antitumor activity andprolonged expression from a TRAIL-expressing adenoviral vec-tor. Neoplasia 2002;4:312–323.
25. Naumann U, Waltereit R, Schulz JB, Weller M. Adenoviral(full-length) Apo2L/TRAIL gene transfer is an ineffective treat-ment strategy for malignant glioma. J Neurooncol 2003;61:7–15.
26. Aboody KS, Brown A, Rainov NG, et al. Neural stem cellsdisplay extensive tropism for pathology in adult brain: evidencefrom intracranial gliomas. Proc Natl Acad Sci USA 2000;97:12846–12851.
27. Wu X, He Y, Falo LD, Hui KM, Huang L. Regression of hu-man mammary adenocarcinoma by systemic administration of arecombinant gene encoding the hFlex-TRAIL fusion protein.Mol Ther 2001;3:368–374.
28. Isacson O, Breakefield XO. Benefits and risks of hosting animalcells in the human brain. Nat Med 1997;3:964–969.
Shah et al: TRAIL-Secreting NPCs 41


















