
Ginger Phenylpropanoids Inhibit IL-1b and Prostanoid Secretion and Disrupt Arachidonate
12
of January 11, 2019. This information is current as 2 Targeting Phospholipases A Arachidonate-Phospholipid Remodeling by Prostanoid Secretion and Disrupt and β Ginger Phenylpropanoids Inhibit IL-1 Karl-Heinz Altmann and Jürg Gertsch Andreas Nievergelt, Janine Marazzi, Roland Schoop, ol.1100880 http://www.jimmunol.org/content/early/2011/09/09/jimmun published online 9 September 2011 J Immunol Material Supplementary 0.DC1 http://www.jimmunol.org/content/suppl/2011/09/09/jimmunol.110088 average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2011 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on January 11, 2019 http://www.jimmunol.org/ Downloaded from by guest on January 11, 2019 http://www.jimmunol.org/ Downloaded from
Transcript of Ginger Phenylpropanoids Inhibit IL-1b and Prostanoid Secretion and Disrupt Arachidonate

of January 11, 2019.This information is current as 2Targeting Phospholipases A
Arachidonate-Phospholipid Remodeling byProstanoid Secretion and Disrupt
andβGinger Phenylpropanoids Inhibit IL-1
Karl-Heinz Altmann and Jürg GertschAndreas Nievergelt, Janine Marazzi, Roland Schoop,
ol.1100880http://www.jimmunol.org/content/early/2011/09/09/jimmun
published online 9 September 2011J Immunol
MaterialSupplementary
0.DC1http://www.jimmunol.org/content/suppl/2011/09/09/jimmunol.110088
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 11, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Ginger Phenylpropanoids Inhibit IL-1b and ProstanoidSecretion and Disrupt Arachidonate-PhospholipidRemodeling by Targeting Phospholipases A2
Andreas Nievergelt,* Janine Marazzi,* Roland Schoop,† Karl-Heinz Altmann,‡ and
Jurg Gertsch*
The rhizome of ginger (Zingiber officinale) is employed in Asian traditional medicine to treat mild forms of rheumatoid arthri-
tis and fever. We have profiled ginger constituents for robust effects on proinflammatory signaling and cytokine expression in
a validated assay using human whole blood. Independent of the stimulus used (LPS, PMA, anti-CD28 Ab, anti-CD3 Ab, and
thapsigargin), ginger constituents potently and specifically inhibited IL-1b expression in monocytes/macrophages. Both the
calcium-independent phospholipase A2 (iPLA2)-triggered maturation and the cytosolic phospholipase A2 (cPLA2)-dependent
secretion of IL-1b from isolated human monocytes were inhibited. In a fluorescence-coupled PLA2 assay, most major ginger
phenylpropanoids directly inhibited i/cPLA2 from U937 macrophages, but not hog pancreas secretory phospholipase A2. The
effects of the ginger constituents were additive and the potency comparable to the mechanism-based inhibitor bromoenol lactone
for iPLA2 and methyl arachidonyl fluorophosphonate for cPLA2, with 10-gingerol/-shogaol being most effective. Furthermore,
a ginger extract (2 mg/ml) and 10-shogaol (2 mM) potently inhibited the release of PGE2 and thromboxane B2 (>50%) and
partially also leukotriene B4 in LPS-stimulated macrophages. Intriguingly, the total cellular arachidonic acid was increased 2- to
3-fold in U937 cells under all experimental conditions. Our data show that the concurrent inhibition of iPLA2 and prostanoid
production causes an accumulation of free intracellular arachidonic acid by disrupting the phospholipid deacylation-reacylation
cycle. The inhibition of i/cPLA2, the resulting attenuation of IL-1b secretion, and the simultaneous inhibition of prostanoid
production by common ginger phenylpropanoids uncover a new anti-inflammatory molecular mechanism of dietary ginger that
may be exploited therapeutically. The Journal of Immunology, 2011, 187: 000–000.
The rhizome of ginger (Zingiber officinale) is widely usedin diet, but also in Asian traditional medicine, where it isemployed therapeutically to treat mild forms of rheuma-
toid arthritis, fever, and upper respiratory tract infections (1). Inconventional Western medicine, ginger preparations are adminis-tered clinically to treat emesis, nausea, and migraine headache (2,3). Based on the widespread therapeutic use of ginger in Asiafor the treatment of inflammatory diseases, several studies havebeen dedicated to the elucidation of potential anti-inflammatorymechanisms (4–7). At relatively high mM concentrations, gingerconstituents such as gingerols, shogaols, and diarylheptanoids
have been reported to weakly inhibit components of proinflam-matory signal transduction pathways in vitro, such as NF-kB,protein kinase C, and MAPKs (8, 9). Moreover, ginger con-stituents have been shown to inhibit inducible NO synthase,cyclooxygenase (COX)-1/-2, and lipoxygenase in vitro (9–12). Asignificant inhibition of PGE2 formation by ginger constituentshas already been shown in vivo (13). Furthermore, different gin-gerols and shogaols at low mM concentrations have been shown todirectly bind to and partially activate the cation channel transientreceptor potential vanilloid 1 (14–16), the molecular clue for thespiciness of ginger, as well as the serotonin 5-HT1A G protein-coupled receptor (17). Although the latter studies have elucidatedfeasible molecular mechanisms of action of the major biologicallyactive ginger constituents, they cannot explain the range of anti-inflammatory effects observed in vivo. In particular, the reportedantipyretic effects of ginger remain poorly understood.In this study, we have profiled a standardized food-grade
commercial ginger extract in a high content assay using freshlyisolated human peripheral whole blood. Human blood was in-cubated with discrete stimuli targeting specific or nonspecific sig-nal transduction pathways, and the resulting cytokine patterns werequantified. The assay was validated using blood from differentdonors and by profiling the effects of anti-inflammatory drugs withknown targets. Using this profiling setup, we were able to identifycalcium-independent phospholipase A2 (iPLA2) and cytosolicphospholipase A2 (cPLA2) enzymes as major biological targetsof ginger phenylpropanoids.PLA2 enzymes play a central role in inflammatory processes
(18, 19) and are classified into three main groups, as follows:secretory phospholipase A2 (sPLA2), cPLA2, and iPLA2. Thesedistinctly different enzymes hydrolyze the sn-2 ester bond of
*Institute of Biochemistry and Molecular Medicine, University of Bern, 3012 Bern,Switzerland; †Bioforce AG, 9325 Roggwil, Switzerland; and ‡Department of Chem-istry and Applied Biosciences, Swiss Federal Institute of Technology, 8093 Zurich,Switzerland
Received for publication March 25, 2011. Accepted for publication August 7, 2011.
This work was supported by a research grant from Bioforce AG (Roggwil, Switzer-land). The establishment of lipid analyses by gas chromatography/mass spectrometrywas made possible through a grant from the Novartis Research Foundation.
Address correspondence and reprint requests to Prof. Jurg Gertsch, Institute of Bio-chemistry and Molecular Medicine, University of Bern, 3012 Bern, Switzerland.E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: AA, arachidonic acid; BEL, bromoenol lactone;CBA, cytometric bead array; COX, cyclooxygenase; cPLA2, cytosolic phospholipaseA2; DBA, 2,49-dibromoacetophenone; iPLA2, calcium-independent phospholipase A2;MAFP, methyl arachidonyl fluorophosphonate; NMR, nuclear magnetic resonance;PAK, palmitoyl-6-O-ascorbate potassium salt; PEA, palmitoylethanolamide; PLA2,phospholipase A2; RT, room temperature; sPLA2, secretory PLA2; thio-PC, 1-hexa-decyl-2-arachidonolythio-2-deoxy-sn-glycero-3-phosphorylcholine; TXB2, thrombox-ane B2.
Copyright� 2011 by The American Association of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1100880
Published September 9, 2011, doi:10.4049/jimmunol.1100880 by guest on January 11, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

phospholipids, thus liberating lysophospholipids and free fattyacids, which are precursors for a variety of different mediatorsof physiological and pathological processes (20, 21). PLA2 iso-enzymes are not only involved in the generation of free arach-idonic acid (AA), which is metabolized by cyclo- and lipoxy-genases to eicosanoids (22), but also in the incorporation of freeAA into membranes and redistribution into specific compart-ments (remodeling) (23, 24). Besides regulatory functions in lipidcatabolism/metabolism, PLA2 enzymes are critically involved insignal transduction, phospholipid and cell membrane bilayerremodeling (25), store-operated cation channels and Ca2+ release-activated Ca2+ channels (26), Fas-induced apoptosis (27), cellularproliferation and differentiation (28, 29), glucose-induced insulinsecretion (30), and, most important for this study, they play acentral role in acute (18) and chronic inflammation (31).Both iPLA2 and cPLA2 are crucial for IL-1b maturation and
secretion (membrane translocation and fusion of storage vesicles)(32, 33). IL-1b is mainly secreted by monocytes/macrophages anddendritic cells, but also fibroblasts; is involved in a variety ofinflammatory processes, such as costimulus for T cell activation;and is one of the key players in rheumatoid arthritis and in-flammatory bowel disease (34, 35). IL-1b further induces COX-2expression in endothelial cells and is involved in edema forma-tion, where it synergizes with PGE2 (36). IL-1b is also a majormediator of fever induction (34, 37). In this study, we provideevidence that the inhibition of i/cPLA2 by phenylpropanoids is di-rectly linked to the global inhibition of IL-1b secretion by gingerextracts in vitro. Moreover, ginger phenylpropanoids induce dra-matic changes in arachidonate-phospholipid remodeling, due totheir simultaneous inhibition of iPLA2 and prostanoid synthesis.This apparently robust mechanism of action could provide a ra-tional basis for the reported antipyretic and anti-inflammatory(i.e., immunomodulatory) effects of ginger preparations.
Materials and MethodsChemicals and reagents
Ethanol 99%, KOH, NaCl, D-(+)-glucose, EDTA, LPS from Escherichiacoli, ATP, PMA from Euphorbiaceae, BSA fraction V (BSA), and 2,49-dibromoacetophenone (DBA) were obtained from Fluka Chemie AG(Buchs, Switzerland). DMSO, HEPES, MOPS, chloroform-d, mono-bromobimane, cylcosporin A, aspirin, diclofenac sodium salt, cetirizine,curcumin, parthenolide, fMLP, and dexamethasone 21-phosphate diso-dium salt were purchased from Sigma-Aldrich Chemie. All kinaseinhibitors and methyl arachidonyl fluorophosphonate were purchased fromTocris Biosciences. Thapsigargin was purchased from Alexis Biochem.Water LiChrosolv, TLC silica aluminum sheets 60F254, silica gel 60 (15–40 mm), CaCl2, and acetonitrile were from Merck. Chloroform was pur-chased from Scharlau SA (Barcelona, Spain). The DTT high purity wasobtained from GERBU Biochemicals (Gaiberg, Germany). Authenticstandards of 6-shogaol and 6-, 8-, and 10-gingerol were purchased fromChromaDex (Santa Ana, CA). Vacutainer LH 170 I.U. and Precision-Glide were from BD Biosciences (Plymouth, U.K.). KCl was fromHanseler AG (Herisau, Switzerland), and FCS was from Omnilab. The 1-hexadecyl-2-arachidonoylthio-2-deoxy-sn-glycero-3-phosphorylcholine(thio-PC) was purchased from Cayman Chemicals. The 8- and 10-shogaolwere isolated by liquid chromatography and HPLC, as described pre-viously (17).
Plant material
The extract of Z. officinale (ginger Hot Flavor CO2 extract, type 014.088,batch number 120507) was provided by FLAVEX (Rehlingen, Germany).It was produced by supercritical fluid extraction with carbon dioxide,according to Manninen et al. (38). According to the analysis certificate,it contains 38.7% 6-, 8-, 10-gingerol, 4.3% 6-, 8-, 10-shogaol, 0.7% zin-gerone, and 60 ml/g essential oil. Ginger essential oil was a commercialgrade water vapor distillate from Farfalla. Extracts of organic Harpa-gophytum procumbens were obtained from Bioforce AG (Roggwil,Switzerland). Extract of Salix cortex was obtained from Bionorica SE(Neumarkt, Germany).
Whole blood, assay conditions, and purification of immunecells
Human venous whole blood was drawn from healthy volunteers with theBD vacutainer system. Isolation of PBMCs was conducted in accordancewith the guidelines of the World Medical Association’s Declaration ofHelsinki and approved by ETH Zurich. PBMCs were isolated with Opti-Prep (Axis-Shield, Oslo, Norway) density gradient flotation, and CD14+
monocytes were enriched by adhesion to culture flasks (FACS analysis wasused to check for purity). Whole blood was immediately pipetted ina 96-well plate at 200 ml/well. Ethanolic extracts, pure compounds, orethanol as vehicle control were added at different concentrations with amaximal concentration of 0.5% ethanol and incubated for 30 min at 37˚C.Where indicated, stimuli (LPS [312 ng/ml]) or a combination of PMA[15 ng/ml] and thapsigargin [1 mM], anti-CD3 Ab, or anti-CD28 Ab [each1 mg/ml]) were added, and blood was incubated for another 18 h at 37˚C.For cytokine measurements, the plates were centrifuged, and an ap-propriate amount (usually 50 ml) of the supernatant serum was removedfor immediate analysis. Isolation of PBMCs and T lymphocytes fromperipheral whole blood was performed, as previously reported (39).
Cell cultures. U937 cells (CRL-1593.2) (40) obtained from American TypeCulture Collection and isolated human monocytes/macrophages werecultured in RPMI 1640 medium containing penicillin (100 U/ml), strep-tomycin (100 mg/ml), 2 mM glutamate (all from Life Technologies Invi-trogen, Basel, Switzerland), amphotericin B (2 mg/ml; Sigma-Aldrich,Steinheim, Germany), and 10% FCS (heat inactivated for 30 min at 56˚C)at 37˚C, 5% CO2, and humidified atmosphere. In the experiments mea-suring AA and eicosanoids, the cells were washed and incubated in serum-free medium (see below).
Cytokine measurements. Cytokine quantification in whole blood was donewith BD cytometric bead array (CBA) human inflammatory cytokine kits(BD kit 551811) for IL-8, IL-1b, IL-6, IL-10, TNF-a, IL-12p70, and theBD CBA human allergy mediators kit (BD kit 558022) for IL-3, IL-4,IL-5, IL-7, IL-10, and GM-CSF, or with isolated monocytes using theBD human IL-1b Flex Set (558279), according to the manufacturer’smanuals, measured with a FACScan flow cytometer equipped with anargon laser, and evaluated with the CBA software V1.4 (all by BDBiosciences).ELISA measurements of thromboxane B2 and leukotriene B4. The throm-boxane B2 (TXB2) ELISA colorimetric kit was from Cayman Chemicals(10004023), limit of detection 35 pg/ml, and leukotriene B4 (LTB4) ELISAcolorometric kit (EHLTB4) was from Thermo Scientific (Pierce ProteinResearch Products), limit of detection 20 pg/ml. ELISA measurementswere carried out as specified by the manufacturer’s instructions. A total of23 106 U937 macrophages was stimulated with LPS (1 mg/ml) to measureTXB2 and 10 3 106 primary CD14+ monocytes/macrophages to measureLTB4. Cells were preincubated with DMSO (vehicle) or test compoundsfor 30 min prior to stimulation with LPS (1 mg/ml) and for full LTB4
release costimulation with fMLP (1 mM). LTB4 could not be measuredfrom stimulated U937 macrophages.
Western blots
Gels for Western blots were run on denaturing NuPage Novex 4–12%Bis-Tris precast gels and corresponding MOPS buffer from Invitrogen,according to the manufacturer’s recommendations, using 10 ml samplesdissolved in LDS buffer. Blotting was done on nitrocellulose mem-branes in a NuPage blotting chamber, according to the instructionmanual.
Staining was done according to the ECL manual from PerkinElmer usingTBST (pH 7.6 at used temperatures) and nitrocellulose membranes blockedwith TBST plus 5% defatted milk powder. Ab labeling was done withTBST; 1% defatted milk powder and primary Ab (anti–IL-1b; AbnovaMaxPab and Sigma-Aldrich) were used at 2 ml/ml; and secondary Ab(anti-mouse IgG [goat] HRP labeled by GE Healthcare) at 1/8 ml/ml.Chemiluminescence detection was done with ECL Plus Western blottingdetection reagents (Amersham Biosciences by GE Healthcare). For IL-1bdetection with Western blot analysis, isolated monocytes, in 96-well platesat a density of 2 3 106 cells/ml and 100 ml/well, were stimulated in freshRPMI 1640 with 100 ng/ml LPS for 4 h. Medium was changed after LPSpriming to a gluconate basal salt solution described earlier (33), omittingglycine. Further processing was in analogy to the CBA (BD Biosciences)protocol, but lyophilisates were directly dissolved in 10 ml LDS samplebuffer for gel electrophoresis.
Ethidium bromide uptake to measure P2X7-mediated ion fluxes
Ethidium+ uptake was measured in isolated CD14+ human monocytes(according to 41), but without measuring additional cell markers. Primary
2 GINGER PHENYLPROPANOIDS INHIBIT PHOSPHOLIPASES A2
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

human cells were used for this assay because they gave a better signal-to-noise ratio than U937 cells.
Measurement of IL-1b maturation and secretion
Cytometric bead arrays. For IL-1b detection with CBA, the isolatedmonocytes, in 96-well plates at a density of 2 3 106 cells/ml and 100 ml/well, were stimulated in fresh RPMI 1640 for 4 h under different primingconditions (vehicle, 100 ng/ml LPS and/or 10 mg/ml ginger Hot Flavorextract). Cells were spun down for 5 min at 200 3 g, and medium wasreplaced (to be processed as described below) with 100 ml fresh onecontaining different stimuli (vehicle, 1 mM ATP and/or 10 mg/ml gingerHot Flavor extract), then incubated for 30 min and centrifuged for 5 min at200 3 g, and supernatant was removed. Medium samples and cells wereshock frozen and lyophilized, to be stored at 280˚C until measurement.Lyophilisates were reconstituted in 10 ml hypotone buffer (10 mM HEPES,0.3 mM EGTA, 0.1% Triton X-100, adjusted with KOH to pH 7.4 at RT),and IL-1b was quantified with IL-1b BD FlexSet.
Establishment of PLA2 assay
Phospholipase preparation. U-937 cells (106 cells/ml) were kept in culturemedium, according to the recommendations by American Type CultureCollection. Twenty-four hours before membrane processing, cells werestimulated with 35 ng/ml PMA, according to published protocols (42, 43).Two times 2.83 107 U937 cells were lysed in a hypotone lysis buffer (340mM sucrose, 10 mM HEPES, 1 mM EDTA, 1 mM DTT, pH adjusted withKOH to 7.4 at 4˚C) and centrifuged at 1000 3 g for 30 min at 4˚C. Thepellets were kept as whole membrane preparations because cPLA2 doescolocalize with cell membranes. Supernatants were ultrafiltrated with a 50-kDa molecular mass cut-off, resulting in lysate concentrates of 0.625–1.5mg protein/ml for the U937 (quantified with the reducing agent and de-tergent-compatible protein assay by Bio-Rad).
Substrate synthesis and isolation procedure. Palmitoyl-6-O-ascorbic acidwas synthesized (according to 44). Reaction conditions were as follows:35˚C was the reaction temperature; 8.8 ml 95% sulphuric acid, 2.29 gpalmitic acid, and 1.76 g ascorbic acid; 100% ultrasonic power output withstirring for 15 min. Identity was confirmed by electrospray ionization massspectrometry and two-dimensional nuclear magnetic resonance (NMR).Palmitoyl-6-O-ascorbic acid potassium salt (PAK) was generated beforeuse by mixing appropriate solutions of palmitoyl-6-O-ascorbic acid inchloroform with KOH in methanol to result in a 100 mM mixture of 95:5chloroform to methanol. Crude phosphatidylcholine was isolated fromcommercial grade lecithin (Hanseler AG) by quantitative de-oiling withacetone, enrichment of phosphatidylcholine by quantitative extractionwith boiling ethanol, and finally, precipitation of impurities at 0˚C. Crudephosphatidylcholine was separated over a silica gel column with a solventsystem of dichloromethane/methanol/acetic acid (5:5:1). The extract wasneutralized and dried by filtration over sodium carbonate and anhydrousmagnesium sulfate. The phosphatidylcholine fraction had a TLC purity of.95% and was stabilized with 10 ppm a-tocopherol, dried, and stored at280˚C.
Buffers. The final phospholipase buffer consisted of 10 ml lysate and 10 mlmixed micelle buffer (for iPLA2: 190 mM MOPS, 152 mM KOH, 3 mMDTT, 2 mM ATP, 1 mg/ml BSA, and for cPLA2: 190 mM MOPS, 152 mMKOH, 3 mM DTT, 1.2 mM CaCl2, 1 mg/ml BSA, 20 mM bromoenollactone [BEL]) containing mixed micelles (16 mM PAK, 2 mM Tween 80,and 2 mM thio-PC, self-forming at ∼T . 35˚C). (Final concentrations aretherefore 100 mM Good’s Buffer adjusted with KOH to pH 7.4 at 40˚C,170 mM sucrose, 2 mM DTT, 0.5 mg/ml BSA, 8 mM PAK, 1 mM Tween80, and 1 mM thio-PC, and either 1 mM ATP and 0.5 mM EDTA or 0.1mM free calcium and 10 mM BEL.)
Mixed micelles. Stock solutions of PAK in chloroform, Tween 80, andphospholipid in ethanol were mixed and dried under a nitrogen flow. Themixed lipid film was hydrated in mixed micelle buffer at 40˚C, brieflysonicated to detach film from glass surface if necessary, and gently (toavoid foam formation) shaken until the solution became totally clear.
Size and z potential of the micelles were analyzed on a zetasizer, givingone single sharp peak in both cases with an average size distribution of32.3 6 9.6 nm by volume (area of 99.3%) and 27.9 6 6.25 nm by number(area of 100%) and an average z potential of 239.4 6 3.25. Estimated bytheir size, they consist of ∼500 detergent molecules (quod est demon-strandum), and their z potential most likely traps c/sPLA2 in the scootingmode (45, 46). Notably, these micelles are self-forming, sonication is notmandatory, and they are readily hydrolyzed by all three phospholipasesubtypes tested.
Fluorescence detection and HPLC. Solutions were mixed at room tem-perature (RT), heated to 40˚C (giving a clear solution), and again incubated
at RT under gentle shaking. The enzymatic reaction was stopped after 3 h(.50% hydrolysis) with 16.6 ml ice-cold MeOH. A total of 0.2 ml DTT(100 mM aq. sol., for reduction of dithio-lyso-phosphatidylcholine), 0.2 mlCs4EDTA (100 mM aqueous solution, to chelate free calcium, whichotherwise would interfere with the reaction), and 2 ml Cs2CO3 (200 mMaq. sol., to achieve a pH .8) was added and incubated for 1 h at RT. Then20 ml monobromobimane (10 mM stock in MeOH) was added and in-cubated for 1 h at RT. The reaction was stopped and stabilized by acidi-fication with 1 ml trichloroacetic acid (2.2 M aqueous solution, to achievea pH ,3) and centrifuged at 500 3 g for 5 min. The supernatant could bestored with no detectable decomposition for 3 d at 218˚C until HPLCmeasurement. Analytical HPLC was performed on an Elite LaChromedevice (VWR International, Hitachi) equipped with a fluorescence detectorusing a Nucleodur Sphinx column (4 mm particle size, 4.6 3 100-mmdimension; Macherey-Nagel). A 15-min gradient of 0.25% NH3 in water/acetonitril (LiChrosolv; Merck) from 30:70 to 10:90 was run for analysis,and UV detection at lex 385 nm and lem 485 nm was carried out.
The enzyme activity was calculated from the area under the curve inabsolute values (nmol product total) and (nmol product/min/mg protein).
TLC. TLC was done with a solvent mixture (according to 47). For detectionof phosphate-containing lipids, a molybdenum blue spray solution andcharring were used.
Phospho-MAPKs
MAPK phosphorylation in leukocytes was quantified using BD FlexSets,according to the manufacturer’s recommendations, but with 5 times smallervolumes.
Isolated human lymphocytes were adjusted to 4 3 106 cells/ml andaliquoted at 50 ml into 96-well plates. The 50 ml diluted stimuli wereadded as 23 stock and incubated at 37˚C for the indicated time. Con-ditions for p38 phosphorylation were 10 ml/ml anti-CD3 Ab and 2 nMPMA for 20 min; for ERK1/2 phosphorylation, 10 nM PMA for 30 min;and for JNK1/2 phosphorylation, 10 ml/ml anti-CD3 Ab and 10 nM PMAfor 60 min. Subsequently, samples were cooled on ice and centrifuged at300 3 g for 3 min at 4˚C. The pellets were lysed with 5 ml diluted BDdenaturation buffer containing phosphatase inhibitors (4 mM sodium po-tassium tartrate, 2 mM imidazole, 1.15 mM sodium molybdate, 1 mMsodium fluoride, and 1 mM sodium orthovanadate) and heated to 80˚C for5 min. The samples were cooled on ice, diluted with 5 ml assay diluent,and shock frozen in liquid nitrogen. After rethawing, the lysates werevigorously pipetted to give a clear solution. Finally, phosphorylation wasdetermined with BD FlexSets.
T cell proliferation
CD4+ human lymphocytes were cultured in 96-well plates at 13 106 cells/ml, and 100 ml/well was substituted with additional 50 ml RPMI 1640medium containing 0.625 ml BrdU stock solution and 2 ml anti-CD3 Abper ml or 50 ml RPMI 1640 medium containing vehicle control or testcompounds prior to incubation for 5 d.
Proliferation was determinedwith the BD FITCBrdU FlowKit by FACS,according to the recommendations by the manufacturer (48).
A clear correlation between the proliferation rate and residual CD14+
monocytes could be observed and is in agreement with earlier reports (49, 50).
Characterization of compounds
Nuclear magnetic resonance. Palmitatoyl-6-O-ascorbate was verified bytwo-dimensional NMR done on a Bruker 400 UltraShield and BrukerTOPSPIN 1.3 software.
Mass spectrometry. For in process controls for palmitatoyl-6-O-ascor-bate synthesis and phospholipase assay establishment, mass spectrometry(Waters Alliance HT with separation module 2795 coupled to a dual labsorption detector 2487 and MassLynx V4.0 software) was used. Solventwas 30% water with 0.1% formic acid and 70% acetonitrile; 20 ml samplesolution was injected. The mass range measured was m/z 100–500 and/orm/z 200–600, electrospray ionization (cone voltage): 620 eV, +40 eV, and+80 eV; capillary voltage 3.0 kV; extractor 1 V; and RF lens 0.2 V.Temperature was set at 120˚C and desolvatation temperature at 250˚C. N2
was used as carrier gas at a flow rate of 600 l/h for desolvatation and forcone flow at 40 l/h.
Quantification of lipids (AA, PGE2, palmitoylethanolamide) bygas chromatography/mass spectrometry
Quantification was performed by a gas chromatography/mass spectrometrymethod. A total of 5 3 106 U937 cells was cultured in complete RPMI1640 medium and differentiated with 2 nM PMA for 48 h and washed withPBS, and the medium was replaced with FBS-free medium. Cells were
The Journal of Immunology 3
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
incubated with test compounds (DMSO, 2 mM 10-shogaol, 2 mg/ml gingerHot Flavor extract, 50 and 100 mM aspirin, 0.5 mM methyl arachidonylfluorophosphonate [MAFP]) and incubated for 30 min. Then LPS (1 ug/ml) was added, and the cells were incubated for another 4 h. Cells werescraped off and separated from the supernatant by centrifugation for 5 minat 1000 rpm in a Heraeus Biofuge fresco (rotor 3325).
Lipids were extracted from the supernatants and the cell fractions asfollows: 1 ml ethanol was added to internal standards (vide infra) and mixedwith 9 ml supernatant. The pH was brought to 3 with hydrochloric acid andthe samples added to C 18 Sep-Pak cartridge (Waters) (preactivated with3 ml methanol and equilibrated with 3 ml 10% ethanol). Cartridges werewashed with 10% ethanol and eluted with 3 ml acetonitrile/ethyl acetate(1:1). Supernatant samples were evaporated to dryness.
Cells were extracted similar to the lipid extraction described by Folchet al. (51). In short, cell pellets were sonicated for 5 min at 4˚C ice-coldchloroform (1 ml containing the internal standards), methanol (0.5 ml), andPBS (0.25 ml), and then centrifuged for 5 min at 800 3 g. The organicphase was dried in a glass vial and dried under N2 and reconstituted in 1 mlethanol by vortexing at RT (,5 min), diluted with 9 ml water, and ex-tracted by solid-phase extraction, as described for the supernatant.
Internal standards were as follows: PGE2-d4, 4 ng/ml (100 ng/sample);AA-d8, 4 ng/ml (100 ng/sample); and palmitoylethanolamide-d5, 1 ng/ml(25 ng/sample).
Derivatization of the hydroxyl group of palmitoylethanolamide (PEA)was performed with the silylating agent dimethylisopropylsilyl imidazole.As described by Obata et al. (52), the molecular stability is increased andlarge fragments could be detected. Derivatization of AA was achieved byesterification with pentafluorobenzylbromide and N,N-diisopropylethyl-amine (53). The hydroxyl groups of PGE2 were readily silylated withdimethylisopropylsilyl imidazole and the carboxylic acid as for AA withpentafluorobenzylbromide. The remaining ketone was transformed into anO-methyl oxime by methoxiyamine hydrochloride in pyridine (52).
The samples were analyzed byGC/electron ionizationmass spectrometryusing an Agilent 6890N GC equipped with a 30 m HP-5MS column anda 5975C MS with triple-axis detector. As carrier, gas helium was used ata constant flow rate of 1.5 ml/min with splitless injection at an inlettemperature of 250˚C. Optimal separation of the three analytes was ach-ieved with the following oven program: initial temperature, 150˚C for1 min, followed by an increase to 280˚C at 8˚C/min, with a final time of20 min. Specific ions were used for selected ion monitoring (54).
Quantification of TXB2 and LTB4
TXB2 and LTB4 were quantified from cell culture supernatants usingcommercially available ELISA kits.
ResultsSetup of whole blood assay and immunopharmacologicalprofiling of ginger extract
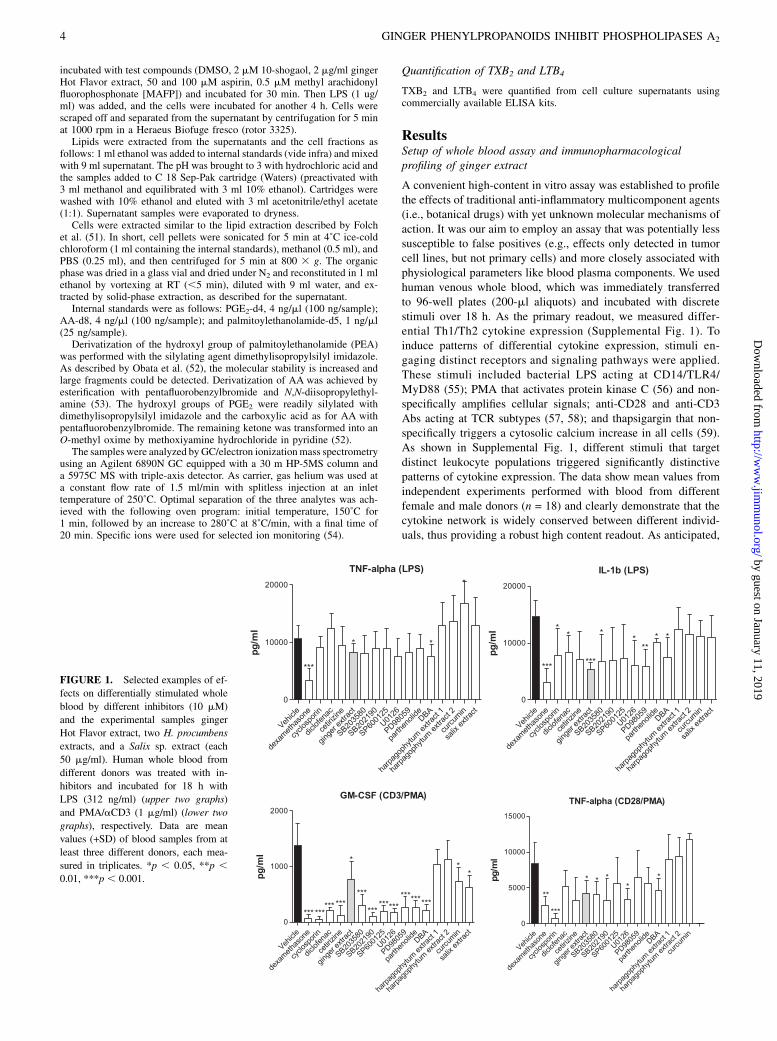
A convenient high-content in vitro assay was established to profilethe effects of traditional anti-inflammatory multicomponent agents(i.e., botanical drugs) with yet unknown molecular mechanisms ofaction. It was our aim to employ an assay that was potentially lesssusceptible to false positives (e.g., effects only detected in tumorcell lines, but not primary cells) and more closely associated withphysiological parameters like blood plasma components. We usedhuman venous whole blood, which was immediately transferredto 96-well plates (200-ml aliquots) and incubated with discretestimuli over 18 h. As the primary readout, we measured differ-ential Th1/Th2 cytokine expression (Supplemental Fig. 1). Toinduce patterns of differential cytokine expression, stimuli en-gaging distinct receptors and signaling pathways were applied.These stimuli included bacterial LPS acting at CD14/TLR4/MyD88 (55); PMA that activates protein kinase C (56) and non-specifically amplifies cellular signals; anti-CD28 and anti-CD3Abs acting at TCR subtypes (57, 58); and thapsigargin that non-specifically triggers a cytosolic calcium increase in all cells (59).As shown in Supplemental Fig. 1, different stimuli that targetdistinct leukocyte populations triggered significantly distinctivepatterns of cytokine expression. The data show mean values fromindependent experiments performed with blood from differentfemale and male donors (n = 18) and clearly demonstrate that thecytokine network is widely conserved between different individ-uals, thus providing a robust high content readout. As anticipated,
FIGURE 1. Selected examples of ef-
fects on differentially stimulated whole
blood by different inhibitors (10 mM)
and the experimental samples ginger
Hot Flavor extract, two H. procumbens
extracts, and a Salix sp. extract (each
50 mg/ml). Human whole blood from
different donors was treated with in-
hibitors and incubated for 18 h with
LPS (312 ng/ml) (upper two graphs)
and PMA/aCD3 (1 mg/ml) (lower two
graphs), respectively. Data are mean
values (+SD) of blood samples from at
least three different donors, each mea-
sured in triplicates. *p , 0.05, **p ,0.01, ***p , 0.001.
4 GINGER PHENYLPROPANOIDS INHIBIT PHOSPHOLIPASES A2
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

LPS stimulation induced mainly Th1 monokines with strong TNF-a, IL-1b, IL-6, IL-8, and IL-10 expressions (.10,000 pg/ml), butweak IL-12 expression (Supplemental Fig. 1). Under these con-ditions, the T cell cytokines IL-3, IL-4, IL-5, IL-7, and GM-CSFwere not induced. In contrast, PMA/anti-CD3 Ab stimulation ledto a robust Th2 cell response with similar IL-8, IL-12, and TNF-aexpressions; less pronounced IL-1b, IL-6, and IL-10 expressions;but significantly stronger IL-3, IL-4, IL-5, IL-7, and GM-CSFexpressions.To validate our assay, we applied specific inhibitors of key in-
flammatory processes or signal transduction events. Clinically andexperimentally used inhibitors were dexamethasone, cyclosporin,diclofenac, cetirizine, parthenolide, the MAPK kinase inhibitorsSB203580 (p38 MAPK inhibitor), SB202190 (p38 MAPK in-hibitor), SP600125 (JNK inhibitor), U0126 (MEK inhibitor), andPD98059 (selective MEK1 inhibitor) (Fig. 1). As expected, dexa-methasone potently inhibited the expression of proinflammatorycytokines independent of the stimulus used. Intriguingly, in thisassay, only dexamethasone strongly inhibited LPS-stimulatedTNF-a expression. The MEK1/2 inhibitor U0126 and the p38inhibitor SB203580 weakly inhibited TNF-a. Distinct MEK andp38 inhibitors like U0126 and PD98059, as well as the broad-spectrum enzyme inhibitor 2,49-dibromoacetophenone (4-bromo-phenacyl bromide or DBA), significantly inhibited LPS-stimulatedIL-1b expression, but with a high interassay variability. Based onour assumption that certain ginger constituents may be able tomodulate proinflammatory cytokine expression (vide supra), thecommercial food-grade ginger Hot Flavor extract (50 mg/ml) wasanalyzed in this assay. Additional anti-inflammatory medicinalplant extracts, like Harpagopyhtum procumbens and Salix spp.,were tested in the same setup. The ginger extract potently in-hibited IL-1b expression with little interassay variability, whereasthe H. procumbens and Salix extracts (both 50 mg/ml) and cur-cumin (25 mg/ml) were largely ineffective (Fig. 1). Moreover, theginger extract differentially modulated the expression of severalcytokines, depending on the stimulus used (vide infra). All in-hibitors of kinases (SB203580, SB202190, SP600125, U0126) andNF-kB (parthenolid and curcumin), as well as DBA and cetirizine,potently inhibited the PMA/anti-CD3 Ab-stimulated GM-CSF(Fig. 1), which may be explained by the fact that this factor isleast strongly induced (,1500 pg/ml). As anticipated, cyclo-
sporine potently inhibited the expression of CD3/CD28-inducedfactors, as exemplified by GM-CSF and TNF-a (Fig. 1), but alsoother calcium-dependent cytokines (IL-3, IL-4, IL-5, IL-7) (datanot shown). The COX-2 inhibitor diclofenac and the H1-antag-onist cetirizine were largely ineffective, whereas the broad-spectrum enzyme inhibitor DBA significantly inhibited most cy-tokines (Fig. 1). As blood from different donors was employed, weconcluded that the inhibitory effects observed were robust andmeaningful.
Ginger extract globally inhibits IL-1b expression in humanwhole blood and in primary monocytes
Intriguingly, the ginger extract (50 mg/ml) potently inhibited IL-1bexpression ($35%) in all experiments in which whole blood wasstimulated, irrespective of the stimulus applied (Fig. 2). In con-trast, the other cytokines were not or only partially inhibited (Figs.1, 2), also reflecting the lack of general cytotoxicity of gingerextracts under these assay conditions (data not shown). Based onthis finding, we concluded that ginger constituents could selec-tively and globally interfere with the IL-1b expression machineryby concrete, yet unknown mechanisms (Supplemental Fig. 2).Because the stimuli employed induced distinctly different signal
FIGURE 2. Global inhibition of IL-1b expression by ginger Hot Flavor
extract in differentially stimulated whole blood. A total of 50 mg/ml extract
was incubated together with different stimuli (z-axis) for 18 h, and cyto-
kine expression was determined by CBA. Arrow indicates that IL-1b
is robustly inhibited by 30–50%, independent of the stimulus applied,
whereas TNF-a, IL-6, and IL-10 are only partially inhibited. IL-8 ex-
pression was not modulated by ginger. Data are the mean values from
blood of at least three different donors, each measured in triplicates. The
SD was ,20%.
FIGURE 3. A, Relative expression of IL-1b in culture medium of hu-
man monocytes. Cells were incubated for 4 h with vehicle control, LPS
(312 ng/ml), ginger Hot Flavor extract (10 mg/ml), or both, and then
stimulated for 30 min (e.g., with vehicle control, ATP [2 mM], ginger Hot
Flavor extract [10 mg/ml], or both). Data show mean values + SD (n = 4);
*p , 0.05. B, Representative graphs of the time-resolved (x-axis) ethidium
bromide fluorescence (y-axis) of vehicle control (1) and ginger Hot Flavor
extract (10 mg/ml) in LPS-primed and ATP-stimulated monocytes (ATP
added at 0 s). The time versus fluorescence mean value plots were iden-
tical, and an inhibition of either ATP at its receptor or fast ion fluxes
(potassium out, calcium in) can be excluded. C, Western blot for IL-1b
after 60-min stimulation in gluconate basal salt solution in the cytosol (a)
and in supernatant medium (b) of isolated monocytes. Upper row, 31-kDa
pro–IL-1b; lower row, 17-kDa mature IL-1b. Lane 1, Vehicle control;
lanes 2–4, priming with ginger extract, LPS, and ginger extract prior to
LPS; lane 5, LPS priming plus ginger extract stimulation; lane 6, only ATP
stimulation; lanes 7–9, LPS priming with ATP stimulation; alone, with
ginger extract prior to LPS, and ginger extract prior to ATP; and lane 10,
pro- and mature IL-1b standards.
The Journal of Immunology 5
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

transduction events, we excluded the possibility that upstreamevents, such as, for example, inhibition of MAPKs or transcriptionfactors, could be responsible for this effect.
The inhibition of IL-1b expression by ginger extract ismediated by phenylpropanoids that inhibit i/cPLA2, but notsPLA2
Both the purinoreceptor P2X7 and PLA2 enzymes have beenshown to be crucial for efficient IL-1b expression (maturation andsecretion) from monocytes/macrophages (60). To explore theeffects of ginger on IL-1b, experiments in which LPS stimulationwas coactivated by ATP were performed in U937 cells, a humanmonocyte/macrophage cell line, to differentiate between IL-1bmaturation and secretion. U937 cells that are stimulated by LPSempty the IL-1b stores (by yet unknown mechanisms), leadingonly to partial IL-1b section (Fig. 3A) (61). When the cells arecostimulated by ATP (which is elevated under inflammatoryconditions and present in whole blood), the purinoceptor P2X7,a ligand-gated ion channel, is activated, leading to activation of
caspase-1 that cleaves pro–IL-1b into the mature IL-1b. In par-allel, this causes an increase in [Ca2+]i, which triggers the releaseof the IL-1b storage vesicles. As shown in Fig. 3A, the gingerextract significantly and specifically inhibited the ATP/LPS-stimulated IL-1b secretion from U937 cells, thus indicating po-tential effects on P2X7, PLA2 enzymes, and/or caspase-1. Thelevel of ethidium bromide uptake after ATP stimulation can beregarded as equivalent to general cation influx through large P2X7
receptor adjacent pores (41). The assay was performed in isolatedhuman monocytes, as they showed a better signal-to-noise ratiothan U937 cells. In this assay, P2X7 activity was not inhibited bythe ginger extract (Fig. 3B).Western blot and cytometric bead analyses showed that the
ginger extract inhibits maturation and release of IL-1b from ac-tivated monocytes by ∼60% when the ginger extract was addedbefore ATP (Fig. 3C). Under nonstimulated conditions, the iso-lated human monocytes (2 3 105) secrete ∼4 pg mature IL-1bwithin 4 h in culture and ∼60 pg/ml when incubated (primed) withLPS. The constitutive secretion was statistically unchanged by 10mg/ml ginger extract (Fig. 3A). A total of 1 mM ATP did notmodulate cytokine secretion in nonprimed cells. However, stim-ulating LPS-primed cells with ATP caused an increase in cytokinesecretion (180 pg/ml) (Fig. 3A). This stimulated secretion wassignificantly reduced by .50% when ginger extract (10 mg/ml)was added prior to the LPS priming and by ∼40% when addedafter LPS and prior to ATP. These levels were significantly dif-ferent between LPS plus ATP, but insignificant when comparedwith LPS alone, ruling out an effect on caspase-1. A differentialeffect on intra- and extracellular pro- and mature IL-1b specieswas clearly visible in Western blots (Fig. 3C) even though the
FIGURE 5. A, Fluorescence-coupled PLA2 assay.
The substrate thio-PC is incorporated into mixed
micelles and incubated with a PLA2 containing cell
lysate. The reaction is quenched with methanol,
and the product, a free thiol, is coupled to mono-
bromobimane and quantified by HPLC with fluo-
rescence detection. B, Size and homogeneity of
mixed micelles were measured by zetasizer. C,
Absolute activities of iPLA2 (left) and cPLA2
(right) in absence and presence of either 10 mg/ml
ginger Hot Flavor extract or 10 mM pure com-
pounds or controls are shown as mean values + SD
(n = 3). If not indicated otherwise, mean values are
significantly different from controls (p , 0.05). D,
Concentration-dependent inhibition of 10-shogaol
(mM) and ginger extract (mg/ml) on iPLA2 and
cPLA2, showing positive controls BEL and MAFP,
respectively. Data show mean values 6 SD (n = 3).
FIGURE 4. Chemical structures of ginger phenylpropanoids. The alkyl
side chain length varies by two carbon atoms between 6-(n = 1), 8-(n = 2),
and 10-(n = 3) gingerol and shogaol, respectively.
6 GINGER PHENYLPROPANOIDS INHIBIT PHOSPHOLIPASES A2
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

inhibition of maturation and secretion was less pronounced than inthe assay conditions used for quantitative CBA (Fig. 3A). In iso-lated human monocytes, the iPLA2-dependent IL-1b maturationand the cPLA2-dependent IL-1b secretion were reduced to ∼60%(at 10 mg/ml ginger extract), whereas transcription/translationand constitutive maturation/secretion were unaffected (Fig. 3C).Therefore, we concluded that the major ginger constituents (Fig.4) could be potential inhibitors of PLA2 enzymes (SupplementalFig. 2).To measure i/cPLA2 enzyme activities, a suitable assay was
established using mixed micelles and a fluorescence-coupled as-say (Fig. 5A, 5B) (for experimental details, see Materials andMethods). The major lipophilic ginger constituents (Fig. 4) werepurchased or isolated, as previously reported (17), and tested at 10mM. As shown in Fig. 5C, the ginger extract and its main gingerconstituents inhibited both iPLA2 and cPLA2 enzyme activities.Overall, the phenylpropanoids more strongly inhibited iPLA2. Theinhibition of ∼50% was in the same range as the inhibition by theirreversible PLA2 inhibitors MAFP and BEL. The relatively lowefficacy of the latter two may be due to the preincubation schemeused (62, 63) or more likely residual calcium- and phospholipase-independent hydrolytic activities. Nevertheless, the assay employedwas robust enough to detect i/cPLA2 inhibitors and to comparepotencies of individual compounds. The inhibitor concentrationwas as low as 0.1 molar percent of the total detergent moleculesa possible inhibition by surface dilution kinetics could be ex-cluded (64). The ginger extract and 10-shogaol concentration-dependently inhibited both iPLA2 and cPLA2 with similar poten-cies as BEL and MAFP (Fig. 5D). However, the inhibitionof iPLA2 was clearly stronger at lower concentrations than theinhibition of cPLA2, in particular with the ginger extract (EC50
values 0.7 mg/ml versus 3 mg/ml). For iPLA2, the eight homologswere less active than the controls at equimolar concentrations(10 mM), but 6-shogaol and 10-gingerol were somewhat morepotent, with an overall inhibition of ∼65% (Fig. 5C). Although6-,8-gingerols did not inhibit cPLA2 and 10-shogaol was the mostpotent inhibitor, structure-activity relationships were not apparent.
To assess the specificity of this effect, we also measured sPLA2.Porcine pancreatic sPLA2 activity was tested using isolated phos-phatidylcholine, and an established TLC detection method (65,66) was not inhibited by ginger constituents up to 20 mM, butstrongly inhibited by the nonspecific enzyme inhibitor DBA (datanot shown).
Weak effects of ginger extract on T cell proliferation andMAPKs
iPLA2 inhibition has been shown to contribute to the prolifera-tion of lymphocytes, Jurkat T cells (28), and monocytes (29). Asshown in Fig. 6, the proliferation of anti-CD3 Ab-stimulatedhuman lymphocytes was weakly, but significantly inhibited by theginger CO2 extract (10 mg/ml) and 10-shogaol (.5 mM), but notby a totum extract containing a higher essential oil content. Toaddress the effect of ginger constituents on MAPKs previouslyreported with cancer cells (54), we also measured the effects ofgingerols and shogaols on ERK, JNK, and p38 in primary humanT cells. PMA/anti-CD3 Ab-stimulated lymphocytes were used todetermine the modulation of MAPKs by measuring kinase phos-phorylation states using a commercial CBA assay (from BDBiosciences). As shown in Fig. 6, experiments using primaryhuman T cells did not show significant modulations. Phosphory-lation of p38 was even increased by ginger constituents. Only10-shogaol significantly inhibited JNK phosphorylation by ∼35%,but the whole extract showed a trend toward activation of thiskinase, and thus, no conclusive picture.
Inhibition of prostanoid secretion and modulation ofarachidonate-phospholipid remodeling by gingerphenylpropanoids
Based on the finding that ginger phenylpropanoids (Fig. 4) inhibitthe PLA2 enzymes in the mixed micelles assay (Fig. 5), we nextassessed the effects of ginger extract and 10-shogaol on free AAlevels and PGE2 release directly in differentiated U937 macro-phages (differentiated with 5 nM PMA for 48 h). A quantitativegas chromatography/mass spectrometry analysis was employed to
FIGURE 6. A, Proliferation of anti-CD3 Ab-
stimulated human CD4+ T lymphocytes, as de-
termined by BrdU incorporation. The ginger Hot
Flavor extract (10 mg/ml) and 10-shogaol (5–10
mM) show weak, but significantly reduced pro-
liferation, whereas essential ginger oil, a ginger
totum extract, and the other phenylpropanoids had
no effect. Shown are the mean values + SEM of
triplicates performed with lymphocytes of at least
three different donors. B–D, MAPK phosphoryla-
tion in isolated human CD4+ lymphocytes stimu-
lated with PMA/anti-CD3 Ab together with the
ginger Hot Flavor extract (10 mg/ml) (2) and four
of its main constituents (10 mM). Phosphorylation of
kinases was measured using selective Abs and CBA
(BD Biosciences). Shown are the mean values 6SEM of triplicates performed with lymphocytes
from at least three different donors. *p , 0.05.
The Journal of Immunology 7
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

determine both AA and PGE2 levels in cell supernatants (cellmedium) and in the cellular fractions (see Materials and Meth-ods). Whereas free AA could be detected in both supernatant (5.4nmol/13 107 cells) and in the thoroughly washed cellular fraction(0.4 nmol/1 3 107 cells) of undifferentiated U937 cells, PGE2 wasonly found in the supernatant of U937 macrophages. U937 mac-rophages constitutively released PGE2 (0.1 nmol/1 3 107 cells)even without LPS stimulation (Fig. 7A). Upon stimulation by LPS,the PGE2 levels stably increased ∼3-fold. As shown in Fig. 7B, thepositive control acetyl salicylic acid (aspirin) at concentrations atwhich COX-1/2 is fully inhibited in vitro (.10 mM) inhibitedPGE2 release from LPS-stimulated U937 cells. The PLA2 andnonspecific hydrolase inhibitor MAFP (0.5 mM) and 10-shogaol
(2 mM) inhibited the release of PGE2 and the prostanoid metab-olite TXB2 by .50% (Figs. 7A, 7B, 8). In nonstimulated U937macrophages, 10-shogaol inhibited constitutive PGE2 expressionby .50% (Fig. 7A). Noteworthily, even concentrations of gingerextract as low as 2 mg/ml showed a significant inhibition of PGE2
and TXB2 release, whereas LTB4 was only weakly inhibited. Asexpected, BEL, which more specifically inhibits iPLA2, had noeffect on PGE2, TXB2, or LTB4 production, whereas MAFP alsoinhibited LTB4 (Figs. 7, 8). Moreover, PEA, which is also con-stitutively released by U937 macrophages (0.1 nmol/1 3 107
cells), was not modulated by ginger extract or 10-shogaol (Fig. 7F,7G). LPS stimulation did not increase PEA secretion. Somewhatunexpectedly, ginger extract and 10-shogaol significantly increased
FIGURE 7. Effects of ginger extract and 10-sho-
gaol on PEG2, AA, and PEA levels in LPS-stimulated
U937 macrophages (PMA differentiated). A and B,
PGE2 levels in supernatant. C and D, Free AA levels
in supernatant. E, Free AA levels in cells. F, PEA
levels in supernatant. G, PEA levels in cells. Super-
natant levels of AA, PGE2, and PEA and the in-
tracellular level of AA and PEA were measured by
GC/MC. Ginger Hot Flavor extract (ginger, 2 mg/ml),
10-shogaol (2 mM), MAFP (0.5 mM), and acetyl sal-
icylic acid (aspirin, 50 and 100 mM) were used as
controls. Indicated are the relative mean values + SD
of three independent experiments compared with un-
treated controls (one sample t test). *p , 0.05, **p ,0.01, ***p , 0.001.
8 GINGER PHENYLPROPANOIDS INHIBIT PHOSPHOLIPASES A2
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

the free AA levels in U937 macrophages in the cellular fractionand in the supernatant under both untreated and LPS-stimulatedconditions (Fig. 7C–E). A PLA2 inhibitor would be expectedto rather decrease free AA levels. MAFP reduced free AA inthe supernatant, but aspirin, as expected, did not influence free AAlevels. Because the ginger extract and 10-shogaol inhibit bothiPLA2 and COX, we speculated that this double effect may leadto an increase of free AA from the pool that constitutively feedsCOX catalyzing prostanoid synthesis. We therefore combined theiPLA2 inhibitor BEL and aspirin. As shown in Fig. 7C and 7D,this combination led to an increase of free AA similar to what wasobserved with ginger phenylpropanoids. We concluded that thedisruption of arachidonate-phospholipid remodeling, that is, theblockage of phospholipid reacylation by iPLA2 inhibition, and theconcurrent inhibition of COX is responsible for the unexpectedeffect on free AA by ginger phenylpropanoids (SupplementalFig. 3).
DiscussionA validated high-content whole blood assay was established toprofile robust cytokine modulating effects of reported immuno-modulatory agents, including botanical drugs. This has led to theelucidation of a new anti-inflammatory molecular mechanism ofaction of gingerols and shogaols, which are the major phenyl-propanoids in dietary ginger (67, 68). Our study indicates that thepotent inhibition of i/cPLA2 by phenylpropanoids is related to thespecific inhibition of IL-1b secretion from monocytes/macrophagesby ginger (Supplemental Fig. 2). Moreover, our data confirm theinhibition of COX (10, 11, 67–69) by ginger phenylpropanoids.This is shown by the inhibition of LPS-stimulated PGE2 and TXB2
secretion (Figs. 7, 8) from U937 macrophages, indicating non-selective inhibition of COX enzymes. LPS-stimulated LTB4 fromprimary human monocytes/macrophages (U937 macrophages didnot secrete LTB4) was only weakly inhibited by the ginger extract.The double inhibition of prostanoid secretion and IL-1b by gingerphenylpropanoids is interesting because IL-1b is known to re-inforce PGE2 synthesis (70), which plays multiple roles in in-flammatory processes (71, 72).Our study further shows that the significant cellular inhibition of
prostanoid production by ginger phenylpropanoids together with
the inhibition of iPLA2 leads to a dramatic and unexpected in-crease of free AA. This was corroborated in an experiment inwhich BEL and aspirin were coincubated, and in this combinationalso led to significant increase of free AA levels (Fig. 7). Free fattyacids are first bound to CoA and then incorporated into lipids byeither de novo synthesis of triglycerides (Kennedy pathway) orremodeling of phospholipids (Lands cycle) (23, 24) (SupplementalFig. 3). In the case of AA, the latter involves incorporation ofarachidonate into phosphatidylcholine by a deacetylation/reacety-lation reaction, followed by a phospholipid class switch via CoA-independent transacylation (73, 74). This is the main pathway forAA incorporation in most cell types (75). In both of these pro-cesses, phospholipases, and especially iPLA2, play major regulatoryroles by providing free fatty acid acceptor molecules (e.g., lyso-phosphatidylcholine) (25). In contrast, immune cells generate freeAA mainly from phospholipids by cPLA2 IVA, the only PLA2
with a preference for AA (46, 76–79), and only under certain con-ditions by sPLA2 IIA, V, and X and iPLA2 VIA (74). Because weobserved a significant increase of free cellular AA by ginger phenyl-propanoids in U937 macrophages (Fig. 7E), the iPLA2 inhibitionis likely to be predominant in cells, and sPLA2 enzymes arenot affected. The sPLA2 enzymes may be the cause of the freeAA observed in our assays. The reason that MAFP at concentra-tions around the IC50 of reported i/cPLA2 inhibition (80, 81) onlyweakly inhibits free AA in the supernatant is not clear, but maybe due to the combined i/cPLA2 isoform inhibition and a result-ing indirect reduction of extracellular sPLA2 activity. Moreover,a fraction of the AA that is released by PLA2 activation will berapidly reincorporated into phospholipid, whereas the remainderwill be lost by conversion to eicosanoids or other products, orby b-oxidation (82, 83). Unesterified AA in plasma rapidlyreplaces the amount lost, and this replacement is proportionalto PLA2 activation (84, 85). In our experiments, BEL alone in-hibited free AA, but increased free AA when COX was inhibitedat the same time. This dramatic change (Fig. 7C, 7D) showsthe role of iPLA2 for AA phospholipid reacylation. Therefore,in the presence of COX inhibitors, blockage of iPLA2 deprivesthe cell of phospholipid acceptor molecules and subsequentlyaugments the intracellular AA concentration (75).Given that free AA in cells induces apoptosis (29), the pro-
nounced increase of free cellular AA may also explain some ofthe differential antiproliferative effects of gingerols and shogaolson cancer cells (86, 87). In contrast, PEA levels were not affectedby ginger phenylpropanoids. Its biosynthesis mainly relies onN-acyltransferases, N-acylphosphatidylethanolamine phospholi-pase D, and fatty acid amide hydrolases, and only to a minorextent on lyso-phospholipase D and sPLA2, but not on other PLA2
classes (88, 89). Consequently, our data indicate selectivity towardi/cPLA2 of the ginger extract and its main constituents and ex-clude unspecific perturbation of lipid homeostasis. The action ofginger phenylpropanoids is in line with the already known anti-oxidant and radical scavenging effects of ginger (90, 91) and mayfurther increase its effect on iPLA2 in cells. Reactive oxygenspecies produced by cyclo-/lipoxygenases are known to eitheractivate iPLA2 in a positive feedback loop (42) or act as mediatorbetween cPLA2 and sPLA2 (92–95).The ginger rhizome and its extracts have been shown to be
safe, as exemplified by its widespread dietary use in Asia (1). Incontrast, many tested synthetic i/cPLA2 inhibitors exert unwantedside effects due to unspecific toxicity/reactivity (e.g., BEL or MAFP),poor selectivity (e.g., MAFP and arachidonyl trifluoromethyl ke-tone), or lack of oral availability [e.g., EXPLIS (96)]. Gingerextracts or isolated compounds show similar in vitro potenciesagainst iPLA2 enzymes as standard PLA2 inhibitors, but are
FIGURE 8. Effects of ginger extract and 10-shogaol and PLA2 inhib-
itors MAFP and BEL on production of TXB2 and LTB4 in LPS-stimulated
macrophages. A total of 2 3 106 U937 macrophages (TXB2) and 10 3 106
primary human CD14+ monocytes/macrophages (LTB4) was incubated
with test compounds for 30 min prior to stimulation by LPS (1 mg/ml). To
measure LTB4, the cells were costimulated with fMLP (1 mM). After 3 h,
TXB2 and LTB4 were quantified from cell-free culture supernatants by
ELISA. Data show mean values of three independent experiments + SD,
*p , 0.05, ***p , 0.001.
The Journal of Immunology 9
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

nontoxic. Ginger as a botanical drug has a great acceptance inthe population, and might therefore be used as a physically andmentally well-tolerated augmentation to conventional anti-inflam-matory medication in cases where first-line therapy is not suffi-cient. In particular, treatment of inflammatory bowel syndromeand celiac disease in which IL-1b and PGE2 play a major role(97) or autoimmune inner ear disease (98) could be novel thera-peutic applications of ginger. Overall, the inhibition of PLA2 en-zymes provides a rational basis for several reported propertiesof ginger [anti-inflammatory, antipyretic, analgesic, or cardio-vascular effects (86, 87)].
AcknowledgmentsWe acknowledge the numerous blood donors and medical assistants, in
particular Maria Feher. The ginger extracts were provided by FLAVEX.
DisclosuresThe authors have no financial conflicts of interest.
References1. Duke, J. A., and E. S. Ayensu. 1985. Medicinal Plants of China. Reference
Publications, Algonac, MI.2. Boone, S. A., and K. M. Shields. 2005. Treating pregnancy-related nausea and
vomiting with ginger. Ann. Pharmacother. 39: 1710–1713.3. Bryer, E. 2005. A literature review of the effectiveness of ginger in alleviating
mild-to-moderate nausea and vomiting of pregnancy. J. Midwifery WomensHealth 50: e1–e3.
4. Altman, R. D., and K. C. Marcussen. 2001. Effects of a ginger extract on kneepain in patients with osteoarthritis. Arthritis Rheum. 44: 2531–2538.
5. Bliddal, H., A. Rosetzsky, P. Schlichting, M. S. Weidner, L. A. Andersen,H. H. Ibfelt, K. Christensen, O. N. Jensen, and J. Barslev. 2000. A randomized,placebo-controlled, cross-over study of ginger extracts and ibuprofen in osteo-arthritis. Osteoarthritis Cartilage 8: 9–12.
6. Penna, S. C., M. V. Medeiros, F. S. Aimbire, H. C. Faria-Neto, J. A. Sertie, andR. A. Lopes-Martins. 2003. Anti-inflammatory effect of the hydralcoholic ex-tract of Zingiber officinale rhizomes on rat paw and skin edema. Phytomedicine10: 381–385.
7. Wigler, I., I. Grotto, D. Caspi, and M. Yaron. 2003. The effects of Zintona EC(a ginger extract) on symptomatic gonarthritis. Osteoarthritis Cartilage 11: 783–789.
8. Kim, J. K., Y. Kim, K. M. Na, Y. J. Surh, and T. Y. Kim. 2007. [6]-Gingerolprevents UVB-induced ROS production and COX-2 expression in vitro andin vivo. Free Radic. Res. 41: 603–614.
9. Lee, T. Y., K. C. Lee, S. Y. Chen, and H. H. Chang. 2009. 6-Gingerol inhibitsROS and iNOS through the suppression of PKC-alpha and NF-kappaB pathwaysin lipopolysaccharide-stimulated mouse macrophages. Biochem. Biophys. Res.Commun. 382: 134–139.
10. Nurtjahja-Tjendraputra, E., A. J. Ammit, B. D. Roufogalis, V. H. Tran, andC. C. Duke. 2003. Effective anti-platelet and COX-1 enzyme inhibitors frompungent constituents of ginger. Thromb. Res. 111: 259–265.
11. van Breemen, R. B., Y. Tao, and W. Li. 2011. Cyclooxygenase-2 inhibitors inginger (Zingiber officinale). Fitoterapia 82: 38–43.
12. Heubl, G., G. Abel, W. Blaschek, and H. Hager. 1997. Hagers Handbuch derPharmazeutischen Praxis. Springer, Berlin.
13. Aimbire, F., S. C. Penna, M. Rodrigues, K. C. Rodrigues, R. A. Lopes-Martins,and J. A. Sertie. 2007. Effect of hydroalcoholic extract of Zingiber officinalisrhizomes on LPS-induced rat airway hyperreactivity and lung inflammation.Prostaglandins Leukot. Essent. Fatty Acids 77: 129–138.
14. Denniff, P., I. Macleod, and D. A. Whiting. 1981. Syntheses of the (6)-[n]-gingerols (pungent principles of ginger) and related compounds through regio-selective aldol condensations: relative pungency assays. J.C.S. Perkin I: 82–87.
15. Iwasaki, Y., A. Morita, T. Iwasawa, K. Kobata, Y. Sekiwa, Y. Morimitsu,K. Kubota, and T.Watanabe. 2006. A nonpungent component of steamed ginger—[10]-shogaol—increases adrenaline secretion via the activation of TRPV1.Nutr. Neurosci. 9: 169–178.
16. Morita, A., Y. Iwasaki, K. Kobata, H. Yokogoshi, and T. Watanabe. 2007. Newlysynthesized oleylgingerol and oleylshogaol activate TRPV1 ion channels. Biosci.Biotechnol. Biochem. 71: 2304–2307.
17. Nievergelt, A., P. Huonker, R. Schoop, K. H. Altmann, and J. Gertsch. 2010.Identification of serotonin 5-HT1A receptor partial agonists in ginger. Bioorg.Med. Chem. 18: 3345–3351.
18. Gilroy, D. W., J. Newson, P. Sawmynaden, D. A. Willoughby, and J. D. Croxtall.2004. A novel role for phospholipase A2 isoforms in the checkpoint control ofacute inflammation. FASEB J. 18: 489–498.
19. Amandi-Burgermeister, E., U. Tibes, B. M. Kaiser, W. G. Friebe, andW. V. Scheuer. 1997. Suppression of cytokine synthesis, integrin expression andchronic inflammation by inhibitors of cytosolic phospholipase A2. Eur. J.Pharmacol. 326: 237–250.
20. Honda, Z., M. Nakamura, I. Miki, M. Minami, T. Watanabe, Y. Seyama,H. Okado, H. Toh, K. Ito, T. Miyamoto, et al. 1991. Cloning by functional ex-pression of platelet-activating factor receptor from guinea-pig lung. Nature 349:342–346.
21. Caughey, G. E., M. J. James, and L. G. Cleland. 2005. Prostaglandins and leu-kotrienes. In Encyclopedia of Human Nutrition, 2nd Ed. B. Caballero, L. Allen,and A. Prentice, eds. Elsevier, Amsterdam, p. 42–45.
22. Smith, W. L., and R. C. Murphy. 2004. Prostaglandins and Leukotrienes. InEncyclopedia of Biological Chemistry. W. J. Lennarz and M. D. Lane, eds.Elsevier, Amsterdam, p. 452–456.
23. Lands, W. E. 1958. Metabolism of glycerolipides; a comparison of lecithin andtriglyceride synthesis. J. Biol. Chem. 231: 883–888.
24. Lands, W. E. 1960. Metabolism of glycerolipids. 2. The enzymatic acylation oflysolecithin. J. Biol. Chem. 235: 2233–2237.
25. Balsinde, J., I. D. Bianco, E. J. Ackermann, K. Conde-Frieboes, andE. A. Dennis. 1995. Inhibition of calcium-independent phospholipase A2 pre-vents arachidonic acid incorporation and phospholipid remodeling in P388D1macrophages. Proc. Natl. Acad. Sci. USA 92: 8527–8531.
26. Smani, T., S. I. Zakharov, E. Leno, P. Csutora, E. S. Trepakova, andV. M. Bolotina. 2003. Ca2+-independent phospholipase A2 is a novel de-terminant of store-operated Ca2+ entry. J. Biol. Chem. 278: 11909–11915.
27. Atsumi, G., M. Murakami, K. Kojima, A. Hadano, M. Tajima, and I. Kudo. 2000.Distinct roles of two intracellular phospholipase A2s in fatty acid release in thecell death pathway: proteolytic fragment of type IVA cytosolic phospholipaseA2alpha inhibits stimulus-induced arachidonate release, whereas that of type VICa2+-independent phospholipase A2 augments spontaneous fatty acid release. J.Biol. Chem. 275: 18248–18258.
28. Roshak, A. K., E. A. Capper, C. Stevenson, C. Eichman, and L. A. Marshall.2000. Human calcium-independent phospholipase A2 mediates lymphocyteproliferation. J. Biol. Chem. 275: 35692–35698.
29. Balboa, M. A., R. Perez, and J. Balsinde. 2008. Calcium-independent phos-pholipase A2 mediates proliferation of human promonocytic U937 cells. FEBS J.275: 1915–1924.
30. Poitout, V. 2008. Phospholipid hydrolysis and insulin secretion: a step towardsolving the Rubik’s cube. Am. J. Physiol. Endocrinol. Metab. 294: E214–E216.
31. Malaviya, R., J. Ansell, L. Hall, M. Fahmy, R. L. Argentieri, G. C. Olini, Jr.,D. W. Pereira, R. Sur, and D. Cavender. 2006. Targeting cytosolic phospholipaseA2 by arachidonyl trifluoromethyl ketone prevents chronic inflammation inmice. Eur. J. Pharmacol. 539: 195–204.
32. Kahlenberg, J. M., K. C. Lundberg, S. B. Kertesy, Y. Qu, and G. R. Dubyak.2005. Potentiation of caspase-1 activation by the P2X7 receptor is dependent onTLR signals and requires NF-kappaB-driven protein synthesis. J. Immunol. 175:7611–7622.
33. Qu, Y., L. Franchi, G. Nunez, and G. R. Dubyak. 2007. Nonclassical IL-1 betasecretion stimulated by P2X7 receptors is dependent on inflammasome activa-tion and correlated with exosome release in murine macrophages. J. Immunol.179: 1913–1925.
34. Dinarello, C. A. 1996. Biologic basis for interleukin-1 in disease. Blood 87:2095–2147.
35. Fernandes, J. C., J. Martel-Pelletier, and J. P. Pelletier. 2002. The role of cyto-kines in osteoarthritis pathophysiology. Biorheology 39: 237–246.
36. Molina-Holgado, E., S. Ortiz, F. Molina-Holgado, and C. Guaza. 2000. Inductionof COX-2 and PGE(2) biosynthesis by IL-1beta is mediated by PKC and mitogen-activated protein kinases in murine astrocytes. Br. J. Pharmacol. 131: 152–159.
37. Dinarello, C. A. 1998. Interleukin-1, interleukin-1 receptors and interleukin-1receptor antagonist. Int. Rev. Immunol. 16: 457–499.
38. Manninen, P., E. Haivala, S. Sarimo, and H. Kallio. 1997. Distribution ofmicrobes in supercritical CO2 extraction of sea buckthorn (Hippophae rham-noides) oils. Zeitschrift fur Lebensmitteluntersuchung und -Forschung A 204:202–205. DOI: 10.1007/s002170050063.
39. Gertsch, J., R. Schoop, U. Kuenzle, and A. Suter. 2004. Echinacea alkylamidesmodulate TNF-alpha gene expression via cannabinoid receptor CB2 and multiplesignal transduction pathways. FEBS Lett. 577: 563–569.
40. Sundstrom, C., and K. Nilsson. 1976. Establishment and characterization ofa human histiocytic lymphoma cell line (U-937). Int. J. Cancer 17: 565–577.
41. Jursik, C., R. Sluyter, J. G. Georgiou, S. J. Fuller, J. S. Wiley, and B. J. Gu. 2007.A quantitative method for routine measurement of cell surface P2X7 receptorfunction in leucocyte subsets by two-colour time-resolved flow cytometry. J.Immunol. Methods 325: 67–77.
42. Balboa, M. A., and J. Balsinde. 2002. Involvement of calcium-independentphospholipase A2 in hydrogen peroxide-induced accumulation of free fattyacids in human U937 cells. J. Biol. Chem. 277: 40384–40389.
43. Perez, R., R. Melero, M. A. Balboa, and J. Balsinde. 2004. Role of group VIAcalcium-independent phospholipase A2 in arachidonic acid release, phospholipidfatty acid incorporation, and apoptosis in U937 cells responding to hydrogenperoxide. J. Biol. Chem. 279: 40385–40391.
44. Wen, B., W. Eli, Q. Xue, X. Dong, and W. Liu. 2007. Ultrasound acceleratedesterification of palmitic acid with vitamin C. Ultrason. Sonochem. 14: 213–218.
45. Jain, M. K., B. Z. Yu, M. H. Gelb, and O. G. Berg. 1992. Assay of phospholi-pases A(2) and their inhibitors by kinetic analysis in the scooting mode. Medi-ators Inflamm. 1: 85–100.
46. Bayburt, T., and M. H. Gelb. 1997. Interfacial catalysis by human 85 kDa cy-tosolic phospholipase A2 on anionic vesicles in the scooting mode. Biochemistry36: 3216–3231.
47. Weerheim, A. M., A. M. Kolb, A. Sturk, and R. Nieuwland. 2002. Phospholipidcomposition of cell-derived microparticles determined by one-dimensional high-performance thin-layer chromatography. Anal. Biochem. 302: 191–198.
10 GINGER PHENYLPROPANOIDS INHIBIT PHOSPHOLIPASES A2
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

48. Eidukevicius, R., D. Characiejus, R. Janavicius, N. Kazlauskaite, V. Pasukoniene,M. Mauricas, and W. Den Otter. 2005. A method to estimate cell cycle time andgrowth fraction using bromodeoxyuridine-flow cytometry data from a singlesample. BMC Cancer 5: 122.
49. Thiele, D. L., M. Kurosaka, and P. E. Lipsky. 1983. Phenotype of the accessorycell necessary for mitogen-stimulated T and B cell responses in human periph-eral blood: delineation by its sensitivity to the lysosomotropic agent, L-leucinemethyl ester. J. Immunol. 131: 2282–2290.
50. Dixon, J. F., and K. Uyemura. 1987. Highly purified human T-lymphocytes donot respond to mitogens—including calcium ionophore and phorbol ester.Immunol. Invest. 16: 189–200.
51. Folch, J., M. Lees, and G. H. Sloane Stanley. 1957. A simple method for theisolation and purification of total lipides from animal tissues. J. Biol. Chem. 226:497–509.
52. Obata, T., Y. Sakurai, Y. Kase, Y. Tanifuji, and T. Horiguchi. 2003. Simulta-neous determination of endocannabinoids (arachidonylethanolamide and 2-arach-idonylglycerol) and isoprostane (8-epiprostaglandin F2alpha) by gas chromato-graphy-mass spectrometry-selected ion monitoring for medical samples. J.Chromatogr. B Analyt. Technol. Biomed. Life Sci. 792: 131–140.
53. Balazy, M. 1991. Metabolism of 5,6-epoxyeicosatrienoic acid by the humanplatelet: formation of novel thromboxane analogs. J. Biol. Chem. 266: 23561–23567.
54. Wubert, J., E. Reder, A. Kaser, P. C. Weber, and R. L. Lorenz. 1997. Simulta-neous solid phase extraction, derivatization, and gas chromatographic massspectrometric quantification of thromboxane and prostacyclin metabolites,prostaglandins, and isoprostanes in urine. Anal. Chem. 69: 2143–2146.
55. Aderem, A., and R. J. Ulevitch. 2000. Toll-like receptors in the induction of theinnate immune response. Nature 406: 782–787.
56. Castagna, M., Y. Takai, K. Kaibuchi, K. Sano, U. Kikkawa, and Y. Nishizuka.1982. Direct activation of calcium-activated, phospholipid-dependent proteinkinase by tumor-promoting phorbol esters. J. Biol. Chem. 257: 7847–7851.
57. Los, M., W. Droge, and K. Schulze-Osthoff. 1994. Inhibition of activation oftranscription factor AP-1 by CD28 signalling in human T-cells. Biochem. J. 302:119–123.
58. Nusslein, H. G., K. H. Frosch, W. Woith, P. Lane, J. R. Kalden, and B. Manger.1996. Increase of intracellular calcium is the essential signal for the expressionof CD40 ligand. Eur. J. Immunol. 26: 846–850.
59. Inesi, G., R. Wade, and T. Rogers. 1998. The sarcoplasmic reticulum Ca2+pump: inhibition by thapsigargin and enhancement by adenovirus-mediated genetransfer. Ann. N. Y. Acad. Sci. 853: 195–206.
60. Andrei, C., P. Margiocco, A. Poggi, L. V. Lotti, M. R. Torrisi, and A. Rubartelli.2004. Phospholipases C and A2 control lysosome-mediated IL-1 beta secretion:implications for inflammatory processes. Proc. Natl. Acad. Sci. USA 101: 9745–9750.
61. Perregaux, D. G., R. E. Laliberte, and C. A. Gabel. 1996. Human monocyteinterleukin-1beta posttranslational processing: evidence of a volume-regulatedresponse. J. Biol. Chem. 271: 29830–29838.
62. Yang, H. C., M. Mosior, C. A. Johnson, Y. Chen, and E. A. Dennis. 1999. Group-specific assays that distinguish between the four major types of mammalianphospholipase A2. Anal. Biochem. 269: 278–288.
63. Lucas, K. K., and E. A. Dennis. 2005. Distinguishing phospholipase A2 types inbiological samples by employing group-specific assays in the presence ofinhibitors. Prostaglandins Other Lipid Mediat. 77: 235–248.
64. Carman, G. M., R. A. Deems, and E. A. Dennis. 1995. Lipid signaling enzymesand surface dilution kinetics. J. Biol. Chem. 270: 18711–18714.
65. Skipski, V. P., R. F. Peterson, and M. Barclay. 1964. Quantitative analysis ofphospholipids by thin-layer chromatography. Biochem. J. 90: 374–378.
66. Nakagawa, Y., and K. Waku. 1989. Phospholipids. In Lipids and RelatedCompounds. Springer Protocols, Berlin, p. 149–178.
67. Jolad, S. D., R. C. Lantz, G. J. Chen, R. B. Bates, and B. N. Timmermann. 2005.Commercially processed dry ginger (Zingiber officinale): composition andeffects on LPS-stimulated PGE2 production. Phytochemistry 66: 1614–1635.
68. Jolad, S. D., R. C. Lantz, A. M. Solyom, G. J. Chen, R. B. Bates, andB. N. Timmermann. 2004. Fresh organically grown ginger (Zingiber officinale):composition and effects on LPS-induced PGE2 production. Phytochemistry 65:1937–1954.
69. Lantz, R. C., G. J. Chen, M. Sarihan, A. M. Solyom, S. D. Jolad, andB. N. Timmermann. 2007. The effect of extracts from ginger rhizome on in-flammatory mediator production. Phytomedicine 14: 123–128.
70. Dinarello, C. A. 1985. An update on human interleukin-1: from molecular bi-ology to clinical relevance. J. Clin. Immunol. 5: 287–297.
71. Ha, H., J. H. Lee, H. N. Kim, H. M. Kim, H. B. Kwak, S. Lee, H. H. Kim, andZ. H. Lee. 2006. alpha-Lipoic acid inhibits inflammatory bone resorption bysuppressing prostaglandin E2 synthesis. J. Immunol. 176: 111–117.
72. White, K. E., Q. Ding, B. B. Moore, M. Peters-Golden, L. B. Ware,M. A. Matthay, and M. A. Olman. 2008. Prostaglandin E2 mediates IL-1beta-related fibroblast mitogenic effects in acute lung injury through differentialutilization of prostanoid receptors. J. Immunol. 180: 637–646.
73. Balsinde, J. 2002. Roles of various phospholipases A2 in providing lysophos-pholipid acceptors for fatty acid phospholipid incorporation and remodelling.Biochem. J. 364: 695–702.
74. Perez-Chacon, G., A. M. Astudillo, D. Balgoma, M. A. Balboa, and J. Balsinde.2009. Control of free arachidonic acid levels by phospholipases A2 and lyso-phospholipid acyltransferases. Biochim. Biophys. Acta 1791: 1103–1113.
75. Chilton, F. H., A. N. Fonteh, M. E. Surette, M. Triggiani, and J. D. Winkler.1996. Control of arachidonate levels within inflammatory cells. Biochim. Bio-phys. Acta 1299: 1–15.
76. Diez, E., P. Louis-Flamberg, R. H. Hall, and R. J. Mayer. 1992. Substrate spe-cificities and properties of human phospholipases A2 in a mixed vesicle model.J. Biol. Chem. 267: 18342–18348.
77. Burdge, G. C., A. Creaney, A. D. Postle, and D. C. Wilton. 1995. Mammaliansecreted and cytosolic phospholipase A2 show different specificities for phos-pholipid molecular species. Int. J. Biochem. Cell Biol. 27: 1027–1032.
78. Burke, J. R., M. R. Witmer, and J. A. Tredup. 1999. The size and curvature ofanionic covesicle substrate affects the catalytic action of cytosolic phospholipaseA2. Arch. Biochem. Biophys. 365: 239–247.
79. Ghosh, M., D. E. Tucker, S. A. Burchett, and C. C. Leslie. 2006. Properties of thegroup IV phospholipase A2 family. Prog. Lipid Res. 45: 487–510.
80. Lio, Y. C., L. J. Reynolds, J. Balsinde, and E. A. Dennis. 1996. Irreversibleinhibition of Ca(2+)-independent phospholipase A2 by methyl arachidonyl flu-orophosphonate. Biochim. Biophys. Acta 1302: 55–60.
81. Surette, M. E., N. Dallaire, N. Jean, S. Picard, and P. Borgeat. 1998. Mechanisms ofthe priming effect of lipopolysaccharides on the biosynthesis of leukotriene B4 inchemotactic peptide-stimulated human neutrophils. FASEB J. 12: 1521–1531.
82. Rapoport, S. I. 2001. In vivo fatty acid incorporation into brain phosholipids inrelation to plasma availability, signal transduction and membrane remodeling. J.Mol. Neurosci. 16: 243–261, discussion 279–284.
83. Rapoport, S. I. 2003. In vivo approaches to quantifying and imaging brainarachidonic and docosahexaenoic acid metabolism. J. Pediatr. 143: S26–S34.
84. Holman, R. T. 1986. Control of polyunsaturated acids in tissue lipids. J. Am.Coll. Nutr. 5: 183–211.
85. Basselin, M., L. Chang, R. Seemann, J. M. Bell, and S. I. Rapoport. 2005.Chronic lithium administration to rats selectively modifies 5-HT2A/2C receptor-mediated brain signaling via arachidonic acid. Neuropsychopharmacology 30:461–472.
86. Aggarwal, B. B., A. B. Kunnumakkara, K. B. Harikumar, S. T. Tharakan,B. Sung, and P. Anand. 2008. Potential of spice-derived phytochemicals forcancer prevention. Planta Med. 74: 1560–1569.
87. Aggarwal, B. B., and S. Shishodia. 2006. Molecular targets of dietary agents forprevention and therapy of cancer. Biochem. Pharmacol. 71: 1397–1421.
88. Schmid, H. H. 2000. Pathways and mechanisms of N-acylethanolamine bio-synthesis: can anandamide be generated selectively? Chem. Phys. Lipids 108: 71–87.
89. LoVerme, J., M. Guzman, S. Gaetani, and D. Piomelli. 2006. Cold exposurestimulates synthesis of the bioactive lipid oleoylethanolamide in rat adiposetissue. J. Biol. Chem. 281: 22815–22818.
90. Dugasani, S., M. R. Pichika, V. D. Nadarajah, M. K. Balijepalli, S. Tandra, andJ. N. Korlakunta. 2010. Comparative antioxidant and anti-inflammatory effectsof [6]-gingerol, [8]-gingerol, [10]-gingerol and [6]-shogaol. J. Ethnopharmacol.127: 515–520.
91. Patro, B. S., S. Rele, G. J. Chintalwar, S. Chattopadhyay, S. Adhikari, andT. Mukherjee. 2002. Protective activities of some phenolic 1,3-diketones againstlipid peroxidation: possible involvement of the 1,3-diketone moiety. Chem-BioChem 3: 364–370.
92. Balboa, M. A., R. Perez, and J. Balsinde. 2003. Amplification mechanisms ofinflammation: paracrine stimulation of arachidonic acid mobilization by secretedphospholipase A2 is regulated by cytosolic phospholipase A2-derived hydro-peroxyeicosatetraenoic acid. J. Immunol. 171: 989–994.
93. Kuwata, H., S. Yamamoto, Y. Miyazaki, S. Shimbara, Y. Nakatani, H. Suzuki,N. Ueda, S. Yamamoto, M. Murakami, and I. Kudo. 2000. Studies on a mecha-nism by which cytosolic phospholipase A2 regulates the expression and functionof type IIA secretory phospholipase A2. J. Immunol. 165: 4024–4031.
94. Murakami, M., Y. Nakatani, H. Kuwata, and I. Kudo. 2000. Cellular componentsthat functionally interact with signaling phospholipase A(2)s. Biochim. Biophys.Acta 1488: 159–166.
95. Burke, J. E., and E. A. Dennis. 2009. Phospholipase A2 structure/function,mechanism, and signaling. J. Lipid Res. 50: S237–S242.
96. Chakraborti, S. 2003. Phospholipase A(2) isoforms: a perspective. Cell. Signal.15: 637–665.
97. Harris, K. M., A. Fasano, and D. L. Mann. 2008. Cutting edge: IL-1 controls theIL-23 response induced by gliadin, the etiologic agent in celiac disease. J.Immunol. 181: 4457–4460.
98. Pathak, S., E. Goldofsky, E. X. Vivas, V. R. Bonagura, and A. Vambutas. 2011.IL-1b is overexpressed and aberrantly regulated in corticosteroid nonresponderswith autoimmune inner ear disease. J. Immunol. 186: 1870–1879.
The Journal of Immunology 11
by guest on January 11, 2019http://w
ww
.jimm
unol.org/D
ownloaded from


















