
Germ-Line and Somatic Recombination Induced by in … · 2002-07-08 · Although male recombination...
7
Copyright 0 1990 by the Genetics Society of America Germ-Line and Somatic Recombination Induced by in Vitro Modified P Elements in Drosophila melanogaster John A. Sved,* William B. Egglestont and William R. Engelst *School of Biological Sciences, Sydney University, New South Wales 2006, Australia and +Laboratory of Genetics, University of Wisconsin, Madison, Wisconsin 53706 Manuscript received May 22, 1989 Accepted for publication August 24, 1989 ABSTRACT The P element insertion A2-3(99B) has previously been shown to activate incomplete P elements elsewhere in the genome. Weshowthat this element, in conjunction with a second incomplete P element, P[CaSpeR], also induces recombination in the male germ line. The recombination is induced preferentially in the region of the P[CaSpeR] element. Recombinant chromosomes contain the P[CaSpeR] element in more than 50% of cases, and alternative models of transposon replication and preferential chromosome breakage are put forward to explain this finding. As is the case with male recombination induced by P-M dysgenic crosses, recombination appears to be premeiotic in a high proportion of cases. The A2-3(99B) element is known to act in somatic cells. Correspondingly, we show that the A2-3(99B)-P[CaSpeR] combination elevates the incidence of somatic recombination. H IRAIZUMI (1 97 1) first showed that male recom- bination in Drosophila melanogaster can occur at a rate of around 1% per chromosome in crosses in- volving newly-collected wild type chromosomes. KID- WELL, KIDWELL and SVED (1977) showed that this male recombination was one manifestation of the syn- drome of ‘hybrid dysgenesis,’ and that it occurred specifically in crosses involving wild type males and laboratory females. VOELKER (1974) analyzed the products of male recombination cytologically and showed that most recombinants were not associated with any chromosomal rearrangements. The most recent, and comprehensive, analysis of male recom- bination has been that of SINCLAIR and GRIGLIATTI (1985), who analyzed the chromosomal distribution of male recombination events in crosses involving a multiply marked laboratory strain and a rangeof wild type strains. They showed that the pattern of recom- bination was strain-specific, as would be expected if the factors determiningmale recombination were dif- ferently distributed in different strains. The molecular basis of dysgenesis was elucidated by BINGHAM, KIDWELL and RUBIN (1982), who cloned the P element, and showed that its properties were consistent with many of theattributes of the P-M system of hybrid dysgenesis. The important aspect of P elementbehavior, which allowed its cloning and analysis, was the production of insertion mutations in P-M dysgenesis (SIMMONS and LIM 1980). See ENGELS (1 989) for a review of P elements. Although male recombination is one of the more of page charges. This article must therefore be hereby marked “advertisement” The publication costs of this article were partly defrayed by the payment in accordance with 18 U.S.C. $1734 solely to indicate this fact. Genetics 124: 331-337 (February, 1990) distinctive manifestations of dysgenesis, it is usually measured using very longchromosomeregions, so that the site of the recombination event is unknown. Thus there has been no molecular information which is directly relevant to the study of male recombination. However ENGELS and PRESTON (1 98 1, 1984) showed that the sites of P element insertions were hotspots for chromosome breakage, leading to the production of inversions and other complex rearrangements. Al- though it is not clear that the types of recombination event involved in male recombination and in chro- mosomal rearrangement are similar, the studies of ENGELS and PRESTON provided support to the idea of a direct cause of recombination by P elements. The use of genetically engineered P elements (KA- RESS and RUBIN 1984; LASKI, RIO and RUBIN 1986) has greatly facilitated the study of dysgenesis. In par- ticular, the A2-3(99B) element has enabled the con- ditions of P-M dysgenesis to be manipulated in a very precise way (ROBERTSON et al. 1988). This element has been shown to be capable of mobilizing other incomplete P elements, while remaining essentially unaffected itself. The purpose of the present paper is to study the phe- nomenon of male recombination using the A2-3(99B) element. Weshow that male recombination can be induced at similar levels to that found using strains with many P elements, andfurthermorethat this recombination may occur at the site of incomplete P elements on which transposase is acting. MATERIALS AND METHODS P[ry+ A2-3](99B) insertion: This insertion, hereafter ab- breviated A2-3(99B), has been described by ENCELS et al.
Transcript of Germ-Line and Somatic Recombination Induced by in … · 2002-07-08 · Although male recombination...

Copyright 0 1990 by the Genetics Society of America
Germ-Line and Somatic Recombination Induced by in Vitro Modified P Elements in Drosophila melanogaster
John A. Sved,* William B. Egglestont and William R. Engelst
*School of Biological Sciences, Sydney University, New South Wales 2006, Australia and +Laboratory of Genetics, University of Wisconsin, Madison, Wisconsin 53706
Manuscript received May 22, 1989 Accepted for publication August 24, 1989
ABSTRACT The P element insertion A2-3(99B) has previously been shown to activate incomplete P elements
elsewhere in the genome. We show that this element, in conjunction with a second incomplete P element, P[CaSpeR], also induces recombination in the male germ line. The recombination is induced preferentially in the region of the P[CaSpeR] element. Recombinant chromosomes contain the P[CaSpeR] element in more than 50% of cases, and alternative models of transposon replication and preferential chromosome breakage are put forward to explain this finding. As is the case with male recombination induced by P-M dysgenic crosses, recombination appears to be premeiotic in a high proportion of cases. The A2-3(99B) element is known to act in somatic cells. Correspondingly, we show that the A2-3(99B)-P[CaSpeR] combination elevates the incidence of somatic recombination.
H IRAIZUMI ( 1 97 1 ) first showed that male recom- bination in Drosophila melanogaster can occur at
a rate of around 1 % per chromosome in crosses in- volving newly-collected wild type chromosomes. KID- WELL, KIDWELL and SVED (1977) showed that this male recombination was one manifestation of the syn- drome of ‘hybrid dysgenesis,’ and that it occurred specifically in crosses involving wild type males and laboratory females. VOELKER (1974) analyzed the products of male recombination cytologically and showed that most recombinants were not associated with any chromosomal rearrangements. The most recent, and comprehensive, analysis of male recom- bination has been that of SINCLAIR and GRIGLIATTI (1985), who analyzed the chromosomal distribution of male recombination events in crosses involving a multiply marked laboratory strain and a range of wild type strains. They showed that the pattern of recom- bination was strain-specific, as would be expected if the factors determining male recombination were dif- ferently distributed in different strains.
The molecular basis of dysgenesis was elucidated by BINGHAM, KIDWELL and RUBIN (1982), who cloned the P element, and showed that its properties were consistent with many of the attributes of the P-M system of hybrid dysgenesis. The important aspect of P element behavior, which allowed its cloning and analysis, was the production of insertion mutations in P-M dysgenesis (SIMMONS and LIM 1980). See ENGELS ( 1 989) for a review of P elements.
Although male recombination is one of the more
of page charges. This article must therefore be hereby marked “advertisement” T h e publication costs of this article were partly defrayed by the payment
in accordance with 18 U.S.C. $1734 solely to indicate this fact.
Genetics 124: 331-337 (February, 1990)
distinctive manifestations of dysgenesis, it is usually measured using very long chromosome regions, so that the site of the recombination event is unknown. Thus there has been no molecular information which is directly relevant to the study of male recombination. However ENGELS and PRESTON (1 98 1 , 1984) showed that the sites of P element insertions were hotspots for chromosome breakage, leading to the production of inversions and other complex rearrangements. Al- though it is not clear that the types of recombination event involved in male recombination and in chro- mosomal rearrangement are similar, the studies of ENGELS and PRESTON provided support to the idea of a direct cause of recombination by P elements.
The use of genetically engineered P elements (KA- RESS and RUBIN 1984; LASKI, RIO and RUBIN 1986) has greatly facilitated the study of dysgenesis. In par- ticular, the A2-3(99B) element has enabled the con- ditions of P-M dysgenesis to be manipulated in a very precise way (ROBERTSON et al. 1988). This element has been shown to be capable of mobilizing other incomplete P elements, while remaining essentially unaffected itself.
The purpose of the present paper is to study the phe- nomenon of male recombination using the A2-3(99B) element. We show that male recombination can be induced at similar levels to that found using strains with many P elements, and furthermore that this recombination may occur at the site of incomplete P elements on which transposase is acting.
MATERIALS AND METHODS
P[ry+ A2-3](99B) insertion: This insertion, hereafter ab- breviated A2-3(99B), has been described by ENCELS et al.

332 J. A. Sved, W. B. Eggleston and W. R. Engels
y w sn
29A 35D-E 50c 2 ,NMw" n
a/ b Region 1
cn Reglon 2
b w sp Region 3
FIGURE 1.-Genetic markers and P[CaSpeR] insertions used to study somatic recombination between the centromere and sn on the X chromosome and germ-line recombination in three regions on chromosome 2. The P[CaSpeR] elements are shown as triangles with their cytological locations as determined by in situ hybridiza- tion. The original identifying numbers for these elements are 32.4, 120.10, 32.2, 34.5 and 3 1.4, corresponding to the elements at 12E, 17C, 29A, 35D-E and 50C. The markers bw and sp were used interchangeably to mark the right end of region 3 .
(1987) and ROBERTSON et al. (1988). The P[ry+ A2-31 plas- mid was produced by artificial splicing of the 2-3 intron of the complete P element by LASKI, RIO and RUBIN (1 986). Transformation experiments showed the element to have the ability to activate other P elements somatically and in the germ line. One transformed strain, in which the element was integrated at location 99B, i e . , close to the end of chromosome arm 3R, was found to have stable inheritance of the element, despite having the ability to mobilize other nonautonomous P elements. Thus, the A2-3(99B) element is described as a stable transposase source, although the reason for its stability is not presently understood. The element carries the wild type allele of rosy.
P[CaSpeR] insertions: The P[CaSpeR] plasmid was con- structed by PIRROTTA (1988) by modification of the Car- negie-4 vector (RUBIN and SPRADLINC 1983). It contains P element ends, thew gene, plus a portion of the pUC plasmid.
The transformation of the A2-3(99B) strain using the P[CaSpeR] plasmid has been described by ROBERTSON et al. (1988). Eggs from white females heterozygous for the A2-3(99B) element were injected with the P[CaSpeR] plas- mid. Males and females which showed eye color, indicating the presence of a P[CaSpeR] insertion, and in which the A2-3(99B)-containing chromosome had segregated out, were selected. Each insertion was then assigned to a partic- ular chromosome on the basis of the segregation in the progeny. Stocks were made homozygous by intercrossing. This procedure was facilitated by the fact that the eye color in individuals containing two elements is usually darker than in individuals containing a single element.
Several lines were selected with insertions on chromo- some 2. Chromosome positions were determined by in situ hybridization using a white probe (Xm2.1-see LEVIS, BINCHAM and RUBIN 1982). Conditions for hybridization were as given by ENCELS et al. (1986). Three lines were selected for further study on the basis of chromosome position. The positions of these three elements in relation to the genetic markers used to measure male recombination are shown in Figure 1. The genetic map positions of the P[CaSpeR] insertions are not known, since the insertions were located by in situ hybridization, and the regions in question are not located accurately with respect to visible markers (LINDSLEY and GRELL 1968). However the exact map position of the insertions is unimportant, given that they are located within the three regions defined by the visible markers.
The measurement of germ-line recombination in males: The scheme to study male recombination is shown in Figure 2. The principal genotype of interest is that of the G3a male, which contains the P[CaSpeR] and A2-3(99B) P ele-
a1 b cn bw . [[[a a l b c n b w ' d
J. P[CaSpeR] a1 b cn bw [[[a
P[CaSpeR] Q x -is= d I
a l b c n b w , a1 h cn bw ' Illa
Score recombinants and backcross d d t o w/wfemales
G1
G2
G3a
G3b
determine transposition and excislon rates Score ddfor flCaSpeR] and Cy010
FIGURE 2.-Crossing scheme for production of male recombi- nation using P[CaSpeR] and A2-3(99B) P elements. The second and third chromosomes are shown, except for the female in G3b where the only chromosome shown is the X . The P[CaSpeR] element on chromosome 2 refers in turn to each of the three elements shown in Figure 1. P[CaSpeR] and A2-3(99B) are the only two P elements present in the crosses. The unmarked third chromosomes in the a1 b cn bw and P[CaSpeR] stocks are designated III" and IIP, respec- tively.
ments, together with second chromosome markers a1 b cn bw which allow recombination to be detected. Note that the P[CaSpeR] and A2-3(99B) elements are brought together for the first time in this genotype.
Association of the P[CaSpeR] element with recombi- nant gametes: The procedure of Figure 2 does not allow the presence or absence of the P[CaSpeR] element to be classified directly in recombinant progeny of the G3a males, since the experiment is carried out in the presence of the w+ gene. However for recombinant chromosomes carried by male offspring the classification can be made in an extra generation by backcrossing tow females, and recording the presence or absence of P[CaSpeR] amongst the male prog- eny. About half of the recombinant progeny are expected to carry the A2-3(99B) element, but this does not affect the usefulness of the backcross test.
Measurement of excision and insertion: Cross G3b of Figure 2 is used to estimate the extent of excision and insertion of the P[CaSpeR] elements. Male progeny have the w allele, which allows the P[CaSpeR] element to be scored. Recombination cannot be detected in this cross. The ap- pearance of progeny which contain neither the Cy nor P[CaSpeR] markers indicates excision. The fraction of ex- cision is calculated from the number of progeny lacking both Cy and P[CaSpeR] divided by the total number of non- Cy progeny. Similarly, the appearance of male progeny with both markers indicates an insertion event. Such insertions can be into any chromosome passed on in the gamete, either the Cy chromosome 2 or chromosome 3, and potentially also the Y chromosome or chromosome 4. However insertions into the original chromosome 2, whether or not accom- panied by excision, cannot be detected in this screen. The fraction of insertion is calculated from the number of Cy, P[CaSpeR] progeny divided by the total number of Cy prog- eny.
Tests for somatic recombination: The X-chromosome markers that were used to measure somatic recombination are shown in Figure 1. The markers y and sn' are cell-

P Element-Induced Male Recombination 333
yellow wild type
(four-strand stage shown) _..- Y W + v w TABLE 1
Summary of germ-line recombination
-.. + w s n A w
FIGURE 3.-X chromosome genotype constructed to test for so- matic recombination, showing the possible production of twin- spotting by reciprocal recombination at or near the site of the P[CaSpeR] insertion. Although not shown here, the A2-3(99B) element was also present.
autonomous markers which can both be classified in individ- ual bristles (LINDSLEY and GRELL 1968). Two P[CaSpeR] insertions lying proximal to these markers were used. Both have been shown to excise and insert at high rates in the presence of A2-3(99B) (R. PHILLIS, personal communica- tion).
The genotype of females constructed to test for somatic recombination is shown in Figure 3. To obtain them, we performed the cross
y w + P[CaSpeR] + w sn A2-3 ox”. - y w + P[CaSpeR] Y ’ + d
and selected daughters carrying the A2-3(99B) element, easily identified by eye color mosaicism due to the interac- tion between A2-3(99B) and P[CaSpeR]. Flies lacking the A2-3(99B) element were also scored and treated as a control group.
Figure 3 depicts the production of twin spots, caused by reciprocal recombination between the proximal marker (sn) and the centromere. Note that the production of twin- spotting requires the appropriate segregation of crossover and non’crossover products. Single spots of either yellow or singed can result if recombination is nonreciprocal, or if only one of the two daughter cells survives to give rise to a visible patch of cuticle. Recombination between the y and sn loci can also give rise to yellow patches.
RESULTS
Germ-line recombination: A summary of the re- sults of the experiments to measure germ-line recom- bination in males is given in Table 1. The numbers in columns 2-5 show how many males produced recom- binants in a particular region. For example, the first row shows that out of a total of 155 males tested, 47 gave one or more recombinants in the first region (which contained the P[CaSpeR] element), 8 in the second, and 32 in the third. Note that some males are counted as having recombination in more than one region. The clustering of recombination events, pre- sumably due to pre-meiotic origin of crossingover (HIRAIZUMI 197 l) , makes such counts useful for meas- uring recombination frequency. However it is useful to have also an estimate of the overall number and percentage of recombinants, and these numbers, along with the total numbers of progeny, are shown
No. of crosses with re-
in the indicated region combination
(Figure 1) Progeny counts
Total Recom- Totdl Recom- P[CaSpeR] 1 2 3 crosses binants progeny binants
%
29A 47* 8 32 155 145 17,322 0.84
50C 5 4 79* 179 158 23,651 0.67 12E 0 0 1 43 1 7,421 0.01 None 0 0 0 63 0 7,587 0
35D-E 7 21* 2 70 84 9,446 0.89
Columns 2-4 refer to the number of G3a males (see Figure 2) producing at least one recombinant in the given genetic region of chromosome 2. Intervals containing a P[CaSpeR] element are indi- cated by asterisks. Columns 6-8 refer to the actual progeny num- bers.
in the final three columns. It is worth noting that the overall frequency of recombination is comparable to that found over the entire chromosome in crosses involving P and M strains (KIDWELL, KIDWELL and SVED 1977).
The most important finding from the results of Table 1 is the concentration of recombination events in the region containing the P[CaSpeR] element. We interpret this correlation to mean that most recombi- nation events take place at the sites of the P[CaSpeR] element. However the large size of our genetic inter- vals leaves open other possibilities.
Table 1 also indicates an elevated recombination rate in regions lacking a P[CaSpeR] element, but only in genotypes with a P[CaSpeR] elsewhere on the chro- mosome. These recombinants might represent two- step events in which a jump of the P[CaSpeR] element is followed by recombination.
Recombination clusters: The male recombination observed in these experiments is closely analogous to the recombination found in P-M cross hybrids. A key feature of recombination in such crosses is the pro- duction of clusters of recombinant products (HIRAI- ZUMI 1971), suggesting a premeiotic origin for some, and conceivably all, of the recombinants. Clusters were also found in our experiments, as illustrated by the analysis of the data in Figure 4. Note that the test statistic for each data set was in the far right tail of its null distribution, indicating highly significant cluster- ing. Individually recognizable clusters were found on several occasions, notably a single progeny group with 19 recombinants among 165 total progeny in one of the crosses involving P[CaSpeR](35D-E). The largest cluster in the P[CaSpeR](29A) crosses was 14/241, and in the P[CaSpeR](50C) crosses it was 11/442. Each of these was a reciprocal cluster, having both recom- binant products at about the same frequency.
Excision and insertion: The measurements of ex-
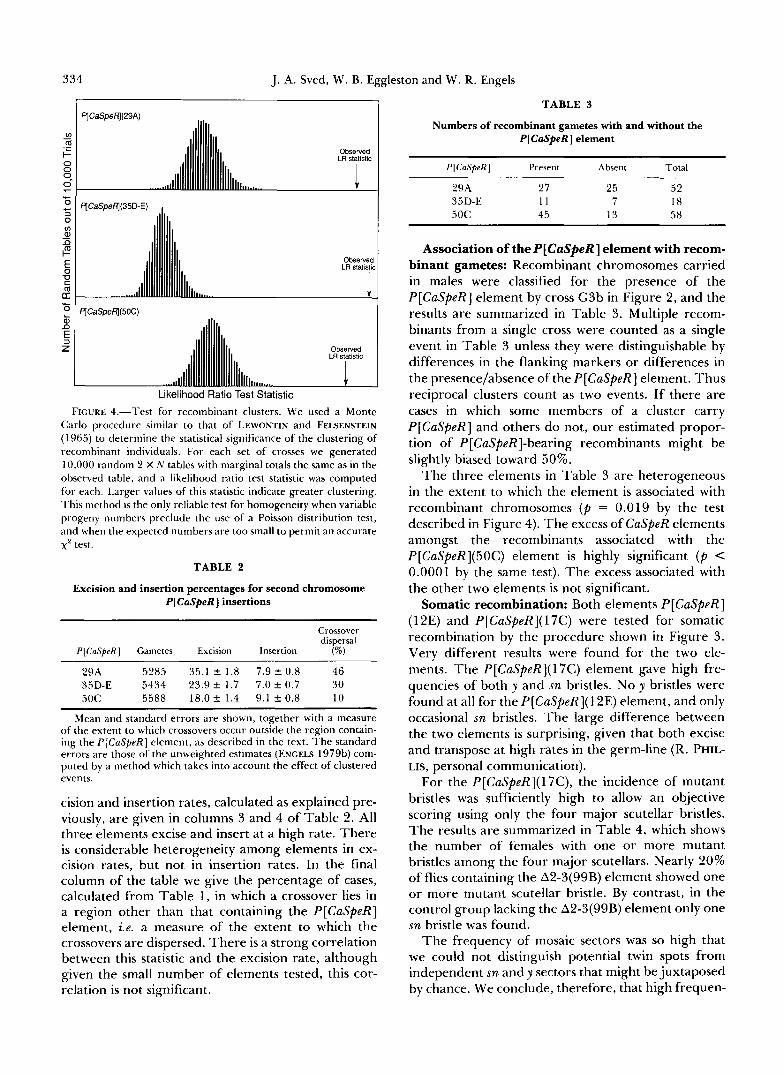
334 J. A. Sved, W. B. Eggleston and W. R. Engels
qCaSpeW(35D-E) , I
qCaSpefl(5OC) I
Likelihood Ratio Test Statistic
FIGURE 4.-Test for recombinant clusters. We used a Monte Carlo procedure similar to that of LEWONTIN and FELSENSTEIN (1965) to determine the statistical significance of the clustering of reconlbinant individuals. For each set of crosses we generated 10,000 random 2 X N tables with marginal totals the same as in the observed table, and a likelihood ratio test statistic was computed for each. Larger values of this statistic indicate greater clustering. This method is the only reliable test for homogeneity when variable progeny numbers preclude the use of a Poisson distribution test, and when the expected numbers are too small to permit an accurate x2 test.
TABLE 2
Excision and insertion percentages for second chromosome P[CaSpeR] insertions
Crossover dispersal
P[CaSpeR] Gametes Excision Insertion (%)
29A 5285 35.1 t 1.8 7.9 t 0.8 46 35D-E 5434 2 3 . 9 t 1.7 7.0 t 0.7 30 50C 5588 18.0 k 1.4 9.1 k 0.8 10
Mean and standard errors are shown, together with a measure of the extent to which crossovers occur outside the region contain- ing the P[CaSpeR] element, as described in the text. The standard errors are those of the unweighted estimates (ENGELS 1979b) com- puted by a method which takes into account the effect of clustered events.
cision and insertion rates, calculated as explained pre- viously, are given in columns 3 and 4 of Table 2. All three elements excise and insert at a high rate. There is considerable heterogeneity among elements in ex- cision rates, but not in insertion rates. In the final column of the table we give the percentage of cases, calculated from Table 1, in which a crossover lies in a region other than that containing the P[CaSpeR] element, i.e. a measure of the extent to which the crossovers are dispersed. There is a strong correlation between this statistic and the excision rate, although given the small number of elements tested, this cor- relation is not significant.
TABLE 3
Numbers of recombinant gametes with and without the P[CaSpeR] element
P[CaSpeR] Present Absent Total
29A 27 25 52 35D-E 11 7 18 50C 45 13 58
Association of theP[CaSpeR] element with recom- binant gametes: Recombinant chromosomes carried in males were classified for the presence of the P[CaSpeR] element by cross G3b in Figure 2, and the results are summarized in Table 3. Multiple recom- binants from a single cross were counted as a single event in Table 3 unless they were distinguishable by differences in the flanking markers or differences in the presence/absence of the P[CaSpeR] element. Thus reciprocal clusters count as two events. If there are cases in which some members of a cluster carry P[CaSpeR] and others do not, our estimated propor- tion of P[CaSpeR]-bearing recombinants might be slightly biased toward 50%.
The three elements in Table 3 are heterogeneous in the extent to which the element is associated with recombinant chromosomes ( p = 0.019 by the test described in Figure 4). The excess of CaSpeR elements amongst the recombinants associated with the P[CaSpeR](50C) element is highly significant ( p < 0.0001 by the same test). The excess associated with the other two elements is not significant.
Somatic recombination: Both elements P[CaSpeR] (12E) and P[CaSpeR]( 17C) were tested for somatic recombination by the procedure shown in Figure 3. Very different results were found for the two ele- ments. The P[CaSpeR](17C) element gave high fre- quencies of both y and sn bristles. No y bristles were found at all for the P[CaSpeR]( 12E) element, and only occasional sn bristles. The large difference between the two elements is surprising, given that both excise and transpose at high rates in the germ-line (R. PHIL- LIS, personal communication).
For the P[CaSpeR](17C), the incidence of mutant bristles was sufficiently high to allow an objective scoring using only the four major scutellar bristles. The results are summarized in Table 4, which shows the number of females with one or more mutant bristles among the four major scutellars. Nearly 20% of flies containing the A2-3(99B) element showed one or more mutant scutellar bristle. By contrast, in the control group lacking the A2-3(99B) element only one sn bristle was found.
The frequency of mosaic sectors was so high that we could not distinguish potential twin spots from independent sn and y sectors that might be juxtaposed by chance. We conclude, therefore, that high frequen-

P Element-Induced Male Recombination
TABLE 4
Results of somatic recombination test
P element - 335 and Mosaic
Y
A2-3(998) Nonmosaic y sn y sn sn frequency
+ 155 6 12 8 10 2 0.1969 - 186 0 1 0 0 0 0.0053
The table shows flies classified for singed and yellow scutellar bristles. The genotype of the test individuals is shown in Figure 3, and the results all refer to the P [CaSpeR]( 17C) insertion.
cies of somatic recombination can be detected in a P[CaSpeR]-containing interval in the presence of A2- 3(99B), but the reciprocal nature of these events has not been established.
DISCUSSION
The mechanism of male recombination: Our re- sults indicate that there are two requirements for elevated frequencies of male recombination: one acts in trans and the other in cis. The trans acting factor is presumably the P element transposase provided by the A2-3(99B) element. The cis requirement is the presence of one or more P elements in the interval of recombination. The P[CaSpeR] elements fulfill this role in our experiments. I t is not known whether immobile elements such as A2-3(99B) can perform the cis function.
The cases in which recombination occurs outside the P[CaSpeR]-containing interval might occur in either of two ways. First, there may be a two step process starting with a transposition of the P[CaSpeR] element to a new site outside the original interval. Subsequently, perhaps several cell generations later, the new P[CaSpeR] site could interact with transposase to produce a recombination event. The second possi- bility is that the sporadic recombination events can occur in the absence of a P element to serve as substrate, but only at relatively low frequencies. There may be occasional genomic sequences which, by chance, have sufficient sequence similarity to the P element so as to interact with P element transposase. MCCARRON et al. (1989) provide evidence for this latter kind of event. In our experiments, however, the rate of recombination in a region lacking CaSpeR elements is increased by the presence of a CaSpeR element elsewhere in the genome. Thus, mobile P elements appear to be able to stimulate at least lower levels of recombination at distant sites.
Our results leave several unanswered questions which can be approached by molecular experiments now in progress. The most important is whether the sites of recombination coincide precisely with the P[CaSpeR] elements, as suggested by the strong cor-
A B
llllllllllllllllllllI 7 IIIIIIIIIIIIIIIIIII llllllllllllllnlll(l~~~~~~~~~.' " 1111111111111111111
FIGURE 5.--Alternative explanations for excess of CaSpeR-con- taining recombinant chromosomes. Only the two strands involved in a recombination event are shown. In A the P element is replicated during the recombination event and remains on both reciprocal products. Hypothesis B involves nonreciprocal recombination and loss of chromosome fragments which fail to rejoin. Only the end products are shown since there are several possibilities for inter- mediates.
relation between the genetic interval of recombination and the genomic location of each P[CaSpeR]. Exami- nation of polymorphic restriction sites flanking the P[CaSpeR] sites in recombinant chromosomes will be needed for confirmation.
The excess of P[CaSpeR]tontaining recombi- nants: Our experiments indicate that, at least for the CaSpeR element at position 5 0 C , well over half of the recombinant chromosomes carried a CaSpeR element capable of generating eye pigmentation. The propor- tion might be even greater if some CaSpeR elements have lost a portion of the w+ gene and were therefore undetected in our testcrosses. The excess of CaSpeR elements might be explained by new transpositions of CaSpeR to other parts of the recombinant chromo- some unrelated to any recombination event. However the frequency of new transpositions of this element is only 9.1% (Table 2), which seems insufficient to ac- count for the excess of CaSpeR-containing recombi- nant chromosomes.
Therefore, we consider two other hypotheses to explain this excess. The first, shown in Figure 5A, is that P element replication occurs in the process of recombination, thus yielding two recombinant chro- mosomes, both of which carry P elements. The model put forward by SHAPIRO (1979) explicitly calls for a recombination intermediate in the process of replica- tive transposition. Under this hypothesis, the non CaSpeR-containing recombinant chromosomes are as- sumed to result from partial or complete deletions of the element. Such deletions can occur either before or after recombination. It is not known whether P elements normally replicate themselves during trans- position, but some prokaryotic transposable elements are known to do so (reviewed in BERG and HOWE 1989). Note from Figure 5 that the end product is a cell in which both homologs carry P elements in the

336 J. A. Sved, W. B. Eggleston and W. R. Engels
same position. Therefore, one testable prediction from this model is that a P element will tend to transpose preferentially to the corresponding site on the homologous chromosome.
Alternatively, the excess of CaSpeR-carrying recom- binants might be explained by non-reciprocal recom- bination as shown in Figure 5B. In this case, an unattached P element terminus produced by a trans- posase-induced break at either end invades the ho- mologous chromosome and is ligated to it. If, in at least some cases, the remaining chromosome frag- ments fail to rejoin there will be loss of the daughter cell that does not carry the P element, resulting in an excess of P[CaSpeR]-containing recombinants. Which of the two recombinant chromosomes is produced depends upon which end of the P element is initially broken.
These two hypotheses make different predictions concerning the reciprocality of recombination events. In hypothesis A, reciprocal exchanges should be com- mon, and both products are expected to carry P elements at the breakpoints. If hypothesis B is correct, reciprocal exchanges should be less frequent, and only one product should carry the P element.
The evidence for at least some reciprocal exchanges is strong for germ-line recombination. SVED (1978) found that reciprocal clusters occurred almost as often as non-reciprocal clusters. The occurrence of recip- rocal clusters argues in favor of a reciprocal recom- bination event. Although non-reciprocal clusters are expected with non-reciprocal recombination, they are also consistent with reciprocal recombination, pro- vided that this occurs at the four-strand stage and that only one of the two recombinant products is incor- porated into a stem cell which gives rise to many sperm.
In the present series of experiments, we could iden- tify at least two putative reciprocal clusters in which we have some information about the association of the CaSpeR element with the recombinant products. The first of these clusters, involving P[CaSpeR](50C), had 7 progeny of genotype +++ bw and 4 progeny of genotype a1 b cn +. The majority of these recombinant chromosomes were, by chance, in males, and we were able to classify 6 and 3, respectively. All were found to have the CaSpeR element. The second cluster, involving the element P[CaSpeR](29A), had 7 a1 +++ and 7 + b cn bw progeny. In this case we could classify only 4 and 2 for the association of CaSpeR, and the numbers were 3:l and 1:l of CaSpeRmon-CaSpeR, respectively. Thus the evidence from the first cluster points to the possibility of replication of the CaSpeR element. The second cluster could also be explained in this way, with loss of the CaSpeR element through excision in some progeny. 111 both cases, however, the
possibility exists that more than one recombination event is involved in the progeny group.
The evidence from the somatic recombination ex- periments is equivocal. True twin-spotting would be an indication of reciprocal recombination. As re- ported above, although the incidence of recombina- tion was high, we could not be certain about the incidence of twin-spots. For somatic recombination, association of the CaSpeR element with the products of recombination was not tested. It is possible that the mechanism of recombination is different in germ-line and somatic tissues, being generally reciprocal in one case and non-reciprocal in the other.
In summary, our results show that the level of germ- line recombination induced by the two-P element system is of a similar order to that given by crosses involving wild type strains (e.g. , HIRAIZUMI 197 1 ; EN- GELS 1979b; SINCLAIR and GRICLIATTI 1985) in which up to 50-60 P elements may be involved and where recombination occurred at approximately 1% per chromosome arm. This model system therefore offers many advantages for the study of the properties and mechanism of P element-induced dysgenic recombi- nation.
We are grateful for advice and assistance from DENA JOHNSON- SCHLITZ, RANDY PHILLIS, CHRISTINE R. PRESTON and DAWN VER- LIN. The work was supported by grants from the Australian Re- search Council and from U.S. Public Health Service grant GM30948. This is paper 3086 from the University of Wisconsin, Laboratory of Genetics.
LITERATURE CITED
BERG, D. E., and M. M. HOWE (Editors), 1989 Mobile DNA. American Society of Microbiology, Washington, D.C.
BINGHAM, P. M., M. G. KIDWELL and G. M . RUBIN, 1982 The molecular basis of P-M hybrid dysgenesis: the role of the P element, a P strain-specific transposon family. Cell 2 9 995- 1004.
ENGELS, W. R., 1979a Germline aberrations associated with a case of hybrid dysgenesis in Drosophila me1anogastt.r males. Genet. Res. 33: 137-146.
ENGELS, W. R. , 1979b The estimation of mutation rates when premeiotic events are involved. Environ. Mutagen. 1: 37-43.
ENGELS, W. R., 1989 P elements in Drosophila, pp 437-484 in Mobile DNA, edited by D. BERG and M. HOWE, American Society of Microbiology, Washington, D.C.
ENGELS, W. R., and C . R. PRESTON, 1981 Identifying P factors in Drosophila by means of chromosome breakage hotspots. Cell 2 6 42 1-428.
ENGELS, W. R. , and C. R. PRESTON, 1984 Formation of chromo- some rearrangements by P factors in Drosophila. Genetics 107:
ENGELS, W. R., C. R. PRESTON, P. THOMPSON and W. B. EGGLES- TON, 1986 In situ hybridization to Drosophila salivary chro- mosomes with biotinylated probes and alkaline phosphatase. Focus 8: 6-8.
ENGELS, W. R., W. K. BENZ, C. R. PRESTON, P. L. GRAHAM, R. w. PHILLIS and H. M. ROBERTSON, 1987 Somatic effects of P element activity in Drosophila melanogaster: pupal lethality. Genetics 117: 745-757.
HIRAIZUMI, Y . , 1971 Spontaneous recombination in Drosophila melanogaster males. Proc. Natl. Acad. Sci. USA 68: 268-270.
657-678.

P Element-Induced Male Recombination 337
KARESS, R. E., and G. M. RUBIN, 1984 Analysis of P transposable element functions in Drosophila. Cell 38: 135-146.
KIDWELL, M. G., J. F. KIDWELL and J. A. SVED, 1977 Hybrid dysgenesis in Drosophila melanogaster: a syndrome of aberrant traits including mutation, sterility, and male recombination. Genetics 8 6 813-833.
LASKI, F. A., D. C. RIO and G. M. RUBIN, 1986 Tissue specificity of Drosophila P element transposition is regulated at the level of mRNA splicing. Cell 44: 7-19.
LEVIS, R., P. M. BINGHAM and G. M. RUBIN, 1982 Physical map of the white locus of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 79: 564-568.
LEWONTIN, R. C., and J. FELSENSTEIN, 1965 T h e robustness of homogeneity tests in 2 X N tables. Biometrics 21: 19-33.
LINDSLEY, D. L., and E. H. GRELL, 1968 Genetic Variations of Drosophila melanogaster. Carnegie Inst. Wash. Publ. 627.
MCCARRON, M. Y . , A. DUTTAROY, G. A. DOUGHTY and A. CHOV- NICK, 1989 P-element transposase induces male recombina- tion in Drosophila melanogaster. Genet. Res. 54: 137-141.
PIRROTTA, V., 1988 Vectors for P-mediated transformation Dro- sophila, pp 437-456, in Vectors, edited by R. L. RODRIGUEZ and D. T. DENHARDT. Butterworth Pub., Stoneham, Mass.
ROBERTSON, H. M., C. R. PRESTON, R. W. PHILLIS, D. JOHNSON-
SCHLITZ, W. K. BENZ and W. R. ENGELS, 1988 A stable genomic source of P element transposase in Drosophila mela- nogaster. Genetics 118: 461-470.
RUBIN, G. M., and A. C. SPRADLING, 1983 Vectors of P element mediated gene transfer in Drosophila. Nucleic Acids Res. 11:
SHAPIRO, J. A,, 1979 Molecular model for the transformation of bacteriophage Mu and other transposable elements. Proc. Natl. Acad. Sci. USA 76: 1933-1937.
SIMMONS, M. J., and J. K. LIM, 1980 Site specificity of mutations arising in dysgenic hybrids of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 77: 6042-6046.
SINCLAIR, D. A. R., and T . A. GRIGLIATTI, 1985 Investigation of the nature of P-induced male recombination in Drosophila melanogaster. Genetics 1 1 0 257-279.
SVED, J. A,, 1978 Male recombination in dysgenic hybrids of Drosophila melanogaster: chromosome breakage or mitotic crossing-over? Aust. J. Biol. Sci. 31: 303-309.
VOELKER, R. A,, 1974 The genetics and cytology of a mutator factor in Drosophila melanogaster. Mutat. Res. 22: 265-276.
6341-6351.
Communicating editor: M. J. SIMMONS











![Nant - Final[1]](https://static.fdocuments.net/doc/165x107/577cdab71a28ab9e78a658a6/nant-final1.jpg)






