
Next generation sequencing & microarray-- Genotypic Technology

Vol. 175, No. 15JOURNAL OF BACTERIOLOGY, Aug. 1993, p. 4895-49040021-9193/93/154895-10$02.00/0Copyright ) 1993, American Society for Microbiology
Genotypic Identification and Phylogenetic Analysis of theSpotted Fever Group Rickettsiae by Pulsed-Field
Gel ElectrophoresisV. ROUX AND D. RAOULT*
Unites des Rickettsies, Faculte de M&decine, 27 Boulevard Jean Moulin, 13385 Marseille, France
Received 12 October 1992/Accepted 29 April 1993
Using pulsed-field gel electrophoresis, we studied the chromosomes of spotted fever group rickettsiae. Wedigested the DNA of 16 species currently known to belong to this group with SmaI, EagI, and BssHII. Thegenome size of 13 rickettsiae was between 1,200 and 1,300 kb. "Ricketisia massiliae" and "R. helvetica"genome sizes were 1,370 and 1,397 kb, respectively, and that ofR. bellii was 1,660 kb. It was possible to obtaindistinctive patterns for each species, but in R. conorii, 10 isolates exhibited the same profiles, showing thatpulsed-field gel electrophoresis is a good interspecies identification tool. We achieved a phylogenetic analysis ofthese bacteria by using the Dice coefficient and UPGMA and Package Philip programming. We established adendrogram of the genetic relationships between the different species showing the existence of a cluster in thespotted fever group rickettsiae including R. conorii, R. rickettsii, R. parkeri, R. sibirica, "R. africae," "R.slovaca," Thai tick typhus rickettsia, and Israeli tick typhus rickettsia. We located three genes previouslycloned and sequenced (genes encoding the R. rickettsii surface proteins of 120 and 190 kDa and the R.prowazekii citrate synthase gene), using Southern hybridization. The genes encoding citrate synthase and thesurface protein of 190 kDa were usually located on the same band, and it is hypothesized that they are relativelyclose on the chromosome.
The family Rickettsiaceae comprises several genera in-cluding the genus Rickettsia, which is subdivided into threegroups: typhus, scrub typhus, and spotted fever group (SFG)rickettsiae. SFG rickettsiae have been grouped taxonomi-cally on the basis of morphological, ecological, and antigeniccriteria. These bacilli are gram-negative short rods, 0.3 to 0.5pum in diameter and 0.8 to 2.0 pum in length (sometimeslonger when cell division is impaired), which retain basicfuchsin when stained by the method of Gimenez (28) andwhich grow both in the nucleus and in the cytoplasm of thehost cells (60). They are transmitted to humans by infected-arthropod bites or feces.The usual identification methods used in bacteriology are
not applicable for rickettsiae because of their strictly intra-cellular position. The actual classification of SFG rickettsiaeis exclusively based on mouse serotyping (41). The antigenicdeterminant of this serotyping is constituted by two majorenvelope proteins of high molecular weight called rOmp Aand rOmp B (26). In fact, this method allows the identifica-tion of new isolates, but no information on the relationshipsbetween the different strains is given. It is necessary todefine strict criteria of classification, because with the de-velopment of a new cell culture isolation technique (the shellvial technique) (36) over the past few years, there has beendetection of new isolates everywhere in the world, both fromticks ("Rickettsia massiliae" in the south of France [6, 7],MC6 in Morocco [37], GS in Greece [5], and "R. helvetica"in Switzerland [15]) and from humans ("R africae" inZimbabwe [34] and R japonica in Japan [54]). Consideringthe few phenotypic characteristics expressed by these bac-teria, a coherent classification should proceed from genomiccriteria. Several studies of the chromosomes intended toestablish a phylogenetic analysis of the rickettsiae have been
* Corresponding author.
performed. 16S rRNA sequence investigation has been usedto reexamine the actual classification of the rickettsiae (58).The first results established by a study of six species showthat the family Rickettsiaceae belongs to the phylum of theProteobacteria phylum and that the three representatives ofthe genus Rickettsia (R. typhi, R prowazekii, and R rick-ettsii) are within the a subdivision. A second study, per-formed by Regnery et al., is based on the comparison ofpatterns obtained after polymerase chain reaction (PCR)followed by restriction fragment length polymorphism(RFLP) analysis of two genes (44). Another approach hasbeen proposed by Ralph et al. (43). They studied rickettsiaegenomes using randomly chosen DNA probes and RFLPanalysis to determine genetic similarity on the basis ofpercent base pair mismatch. The principal disadvantage ofthese last two approaches is that only a small part of thegenome is studied.
Pulsed-field gel electrophoresis (PFGE) appears to be thebest way to study the entire genome of SFG rickettsiae,since this has already been performed for other microorgan-isms (1, 11, 14, 17, 33, 47, 52, 57). Several species of thefamily Rickettsiaceae have been studied by PFGE. Thiswork has allowed the estimation of the molecular weight ofthe genome of bacteria belonging to the genera Rickettsiella(24), Coxiella (31), and Rickettsia (48), and it also allowedthe determination of six different genomic groups within thegenus Coxiella. The purpose of our work was to use PFGEto establish a phylogenetic analysis of the bacteria of thisgroup, including several species which have never beentested ("R helvetica," R japonica, and "R massiliae"),and to localize three previously identified genes (35).
MATERIALS AND METHODS
Rickettsial strains. The rickettsial strains used for thisstudy are listed in Table 1.
4895
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

4896 ROUX AND RAOULT
TABLE 1. Rickettsial strains studied
Rickettsia Strain Source Geographical origin Reference(s)
R. rickettsii Sheila Smith Human Montana 9R. cononi Moroccan Unknown Morocco 9Itt rickettsia ISTT CDC1 Human Israel 29Ttt rickettsia TT-118 Ixodes or Rhipicephalus sp. Thailand 46"R. africae" ESF-5 A. variegatum Shulu Province, Ethiopia 34, 39R. parkeri Maculatum 20 A. maculatum Mississippi 9R. sibirica 232 Human USSR 8"R. slovaca" 13-B Dermacentor marginalis Slovakia 45R. japonica YM Human Japan 54R. Montana ATCC VR-611 Dermacentor varabilis or Ohio 9
Dermacentor andersoniR. rhipicephali 3-7-6 Rhipicephalus sanguineus Mississippi 16"R. massiliae" Mtu 1 Rhipecephalus turanicus France 6, 7R. australis Phillips Human Queensland, Australia 4R. akari MK (Kaplan) Human New York, N.Y. 32"R. helvetica" C9P9 Ixodes ricinus Switzerland 15R. belli 369L42-1 D. vanabilis Ohio 40
(i) Recognized species. Four strains were obtained from theAmerican Type Culture Collection: R. conorii (Moroccanstrain VR-141), R. rickettsii (Sheila Smith strain VR-149), R.akani (VR-148), and R. sibirica (strain 232). R. australis andR. rhipicephali were supplied by G. A. Dasch (Naval Med-ical Research Institute, Bethesda, Md.). Four strains weresupplied by D. H. Walker (University of Texas, Galveston):R. parkeri, R. Montana, R belli, and R. japonica.
(ii) Unrecognized species. One strain, "R. helvetica" (pro-posed name for the "Swiss agent"), was given by W.Burgdorfer (Rocky Mountain Laboratory, Hamilton, Mon-tana) (15). Four strains were given by G. A. Dasch: "R.slovaca" (45), Thai tick typhus (Ttt) rickettsia (46) andIsraeli tick typhus (Itt) rickettsia (29) (considered R. conorii,but antigenically different), and "R. africae" (34, 39). Onestrain, "R. massiliae" (proposed name for Mtu 1) (6, 7), wasisolated in the laboratory from a tick found in France.
(iii) R. conorii isolates. Several isolates of R. conorii wereobtained in our laboratory from the blood of French patients(130114, 151820, 180522, 060518, and 130126) and a Spanishpatient (isolate 16B). One isolate originated from a tick inKenya (Kenya tick typhus rickettsia [10]). Another strainwas obtained from the American Type Culture Collection(R. cononi 7 [VR-613]), and yet another, Ml, from theGamaleya Research Institute Collection, was isolated from atick in Suchumi (Black Sea region, former USSR) (30).
Rickettsial cultivation and purification. The rickettsialstrains were propagated on Vero cell monolayers at 320C in150-cm2 tissue culture flasks with minimal essential mediumsupplemented with 4% fetal calf serum and 2 mM glutamine.Infection of Vero cells was monitored by Gimenez staining,and the rickettsiae were harvested when multiplication wasoptimal (3 to 5 days), centrifuged (12,000 x g for 10 min),suspended in K36 buffer (16.5 mM KH2PO4, 33.5 mMK2HPO4, 100 mM KCl, 15.5 mM NaCl), and immediatelypurified or stored at -70'C.
Cultures harvested from 10 150-cm2 flasks were pooled,sonicated, and centrifuged twice for 10 min at 1,200 x g.Centrifugation through a 25% sucrose cushion (30 min at7,000 x g) was performed in order to eliminate cell debris.The final purification was performed by Renografin densitygradient centrifugation (59) (the gradient was obtained fromtwo solutions, of 28 and 45% Radioselectan in phosphate-buffered saline with a Gilson Minipuls 2 apparatus). After the
bacterial suspension was deposited on the gradient, the tubewas centrifuged for 1 h at 120,000 x g at 40C (Beckman L3-50 centrifuge). After this, the band corresponding to therickettsiae was recovered and the bacteria were washedtwice and suspended again in K36 buffer.PFGE. The bacterial suspension was added to 1 volume of
Incert agarose (FMC Bioproducts, Rockland, Maine) at 420Cto form plugs in a mold apparatus. These were transferred toESP lysis buffer (0.5 M EDTA [pH 8], 1% N-lauroylsar-cosine, proteinase K [1 mg/ml]) and incubated twice for 24 hat 50'C and then washed in TE buffer (10 mM Tris-HCl, 1mM EDTA [pH 7.6]), incubated twice for 1 h in TE buffersupplemented with 0.04 mg of phenylmethylsulfonyl fluorideper ml to inactivate the proteinase K, washed again in TEbuffer, and stored at 40C in EDTA or digested with thechosen endonuclease. The plugs were individually treated inan Eppendorf tube (270 pAl of 1x enzymatic buffer and 20 Uof enzyme) three times for 2 h each at the appropriatetemperature and then stored at 40C in ESP buffer or equili-brated in 0.5x Tris-borate-EDTA (TBE), pH 8 (44.5 mMTris-HCl, 44.5 mM boric acid, 1 mM EDTA), twice for 30min each before electrophoresis. The migration was per-formed in a contour-clamped homogeneous electric fieldsystem (CHEF DRII; Bio-Rad Laboratories, Richmond,Calif.) at 14WC in 0.5x TBE, pH 8. Three endonucleaseswere used: SmaI (CCCGGG) (Boehringer Mannheim Bio-chemicals, Indianapolis, Ind.), EagI (CGGCCG), and BssHII(GCGCGC) (New England BioLabs, Inc., Beverly, Mass.).Three molecular size markers were used: low-range PFGmarker (0.13 to 194 kb), lambda ladder PFG marker (48.5 to1,018 kb), and yeast chromosome PFG marker (225 to 1,900kb) (New England BioLabs, Inc.). Five sets of migrationconditions were tested, depending on the profiles obtained:ramped pulse times of from 3 to 10 s at 5.7 V/cm for 24 h, 5.4V/cmfor 5to20sfor33 h, 5.8V/cmfor3 to5 sfor25 h, 4.5V/cm for 5 to 120 s for 48 h, 6 V/cm for 70 s for 15 h, and then6 V/cm for 120 s for 15 h. At least five independent digestionswere performed with each restriction endonuclease for eachstrain.
Estimation of the band sizes and of the number of frag-ments of identical size inside a given band was performedwith The Imager apparatus (APPLIGENE, Illkirch, France)and the QGEL-1D program (Quantigel Corporation, Madi-son, Wis.).
J. BACTERIOL.
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

PHYLOGENETIC ANALYSIS OF SFG RICKETTSIAE 4897
DNA extraction. The DNA was extracted from rickettsiaepurified by Renografin density gradient centrifugation. Thebacterial wall was lysed during a 2-h incubation in a mixtureof 0.5% sodium dodecyl sulfate (SDS) and 0.5 mg of protein-ase K per ml in an Eppendorf tube at 50'C. A phenolextraction and then a phenol-chloroform extraction werecarried out after aqueous-phase recovery. The DNA wasprecipitated by the addition of 2 volumes of prechilledethanol. The tube was stored overnight at -20'C. The pelletofDNA obtained after centrifugation (12,000 x g for 10 min)was washed with cold 70% ethanol (12,000 x g for 5 min).After elimination of the upper phase, the pellet was dried andsuspended in TE buffer, pH 8 (49). The DNA concentrationwas determined by spectrophotometry at 260 nm.
Southern hybridization. The probes were PCR-amplifiedsequences of the citrate synthase gene and the 120- and190-kDa surface protein genes. The primers were RpCs.877pand RpCs.1258n obtained from the R prowazekii citratesynthase gene (62), Rr 190.70p and Rr 190.602n from the190-kDa surface protein gene ofR. rickettsii (2), and BG 1-21and GB 2-20 from the 120-kDa surface protein gene of Rrickettsii (27) (Bioprobe System, Montreuil-sous-Bois,France). The amplification conditions were those previouslydescribed by Regnery et al. (44): 35 cycles of denaturation(20 s at 950C), annealing (30 s at 480C), and extension (2 minat 60°C). Nonradioactive digoxigenin DNA labeling wasused (DNA labeling and detection kit, nonradioactive; Boe-hringer Mannheim) with a random priming system (22, 23).DNA depurination was performed in 0.25 N HCl for 10 minto enhance the large DNA fragment transfer. The gel wasdenatured in 1.5 M NaCl-0.5 N NaOH and then neutralizedin 0.5 M Tris-1.5 M NaCl (53). DNA was transferred to aHybond-N membrane (Amersham International plc, Amer-sham, United Kingdom). The transfer buffer was 20x SSC(lx SSC is 0.15 M NaCl plus 0.015 M sodium citrate, pH 7).Hybridization at high stringency was carried out at 68°C in abuffer containing 5x SSC, 0.1% N-lauroylsarcosine, 0.02%SDS, and 1% blocking reagent. Hybridization at low strin-gency was carried out at 55°C in the same buffer but with 8 xSSC. The filters were washed twice for 5 min in 2x SSC-0.1% SDS at room temperature and then twice for 15 min in0.1x SSC-0.1% SDS at 68°C after hybridization at highstringency and in lx SSC-0.1% SDS at 55°C after hybrid-ization at low stringency. The hybrids were detected byenzyme-linked immunoassay using antibody conjugate (anti-digoxigenin conjugate with alkaline phosphatase) and subse-quent enzyme-catalyzed color reaction with 5-bromo-4-chloro-3-indolylphosphate and nitroblue tetrazolium salt.Destaining was performed with dimethylformamide and re-moval of the probe by incubation of the filter in 0.2 NNaOH-0.1% SDS for 30 min. It was possible to use TheImager apparatus to locate the stained band on the restric-tion profiles by superimposition of the gel and the Southernblot pictures taken at the same cinecamera distance from thegel or the nylon membrane.
Numerical analysis. A phenogram of the genetic relation-ships between the different species of the SFG rickettsiaewas established from the restriction profiles obtained afterthe three enzymatic DNA digestions and PFGE. The gelswere divided into sections of 5 kb, and we determinedwhether a section was occupied by a band. In this way, itwas possible to define the number of comigrating bands foreach pair of species. DNA divergence between rickettsialgenomes was estimated by using the Dice coefficient (13),and the phenogram was built by using UPGMA and PackagePhilip commercial computer programming.
KB
145.5-
48.5-
9.4-ABC DE FGH I J K L MN O P Q RS T U
FIG. 1. PFGE ofSmaI restriction fragments of rickettsial DNAs.Lanes A, K, and R, lambda ladder PFG marker; lanes B to I, R.ricketsii, R conorii, Ttt rickettsia, "R. africae," Itt rickettsia, R.parkeri, K sibirica, and "R slovaca," respectively; lanes J and Q,low-range PFG marker; lanes L to P, R. bellii, R. japonica, "R.helvetica," "R massiliae," and R. montana, respectively; lanes Sto U, R akari, R. australis, and R. rhipicephali, respectively;Migration was performed in 0.5x TBE at 14WC at 5.7 V/cm for 24 hwith ramped pulse times from 3 to 10 s. The positions and sizes ofmarker bands are indicated at the left.
RESULTS
Digestion of rickettsial DNA by low-frequency cleavagerestriction endonucleases. We chose SmaI, EagI, and BssHII,as for previous work in this laboratory (48), to obtain asuitable number of fragments allowing the determination ofthe molecular weight and the comparison of the profiles ofthe different strains to establish a phylogenetic analysis. Inorder to obtain a good separation of the different fragments,it was necessary to perform several migrations for eachstrain digested with a given enzyme. We changed severalparameters: pulse time, voltage, and time of migration (12).The conditions of migration, pulses of 5.7 V/cm for 3 to 10 sfor 24 h, were tested for the three enzymes, allowingvisualization of almost all of the bands. Shorter pulses (3 to5 s) gave better separation of small fragments. Longer pulses(5 to 120 s) with longer migration times allowed separation oflarge fragments. In fact, a 48-h migration at 4.5 V/cm withramped pulse times of 5 to 120 s allowed visualization of thebands from 48 to 1,000 kb, and migration conditions of 6V/cm for 70 s for 15 h followed by 6 V/cm for 120 s for 15 hallowed separation of DNA fragments up to 2,000 kb. It wassometimes impossible to separate bands migrating at thesame level, despite running several gels under differentconditions. We obtained restriction profiles with the chosenenzymes for the 16 strains tested. To estimate the averagegenome size, the best enzyme was BssHII, because therewas no doublet and the digestion generated a small numberof fragments. Therefore, it was necessary only to determinethe sizes and total them. For comparing strains, the profilesobtained with EagI and SmaI were superior, because it waspossible to have bands with the same size for several strains.For the 10 newly studied strains, using SmaI, we observedbetween 20 and 30 DNA bands whose sizes ranged from 8 to182 kb (Fig. 1 and 2). When using EagI, we observedbetween 15 and 20 bands with sizes from 8 to 330 kb;however, for R befiii two large fragments of 1,300 and 382kb (Fig. 3 and 4) were noted. After digestion of the DNAwith BssHII, we observed between three ("R slovaca") andseven ("R massiliae") bands for eight of the strains. For R.bellii and "R. helvetica," only one band was noted, and ithas been difficult to determine by comparing the patterns
VOL. 175, 1993
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

4898 ROUX AND RAOULT
R cononi Ttt rickettsia R. parked 'R. slovaca R. montana "R. massiliae " R. akai R. belliikb R. rickettsii Itt rickettsia "R. africae" R. sibirica R. japonica R. rhipicephali R. australis "R. helvetica "
500
400
300
250
200
150
120100
80706050
40
30 .._-*. . ..-*
20
15
10 *_.. 10
1255 1230 1217 1220 1237 1246 1238 1242 1272 1252 1257 1363 1256 1221 1361 1639
FIG. 2. Schematic representation of PFGE profiles obtained after SMaI digestion of 16 rickettsial strain genomes. The number of asterisksindicates the number of fragments of identical size inside a given band, as revealed by densitometry using The Imager apparatus and theQGEL-1D program. Numbers at the bottom of the figure indicate the estimated sizes (in kilobases) of the bands obtained after enzymaticdigestion and PFGE.
obtained with digested and nondigested plugs (Fig. 5 and 6)whether the single cut of the chromosome was due to theenzyme or a mechanical alteration during the preparation ofthe plugs.
Isolates of R conorii from different areas (southernFrance, Spain, Kenya, Morocco, the United States, and the
KB
242.5-
145.5-
48.5
ABC D EF G H I J K L MN O PQ RS
FIG. 3. PFGE of EagI restriction fragments of rickettsial DNAs.Lanes A, L, N, and 0, lambda ladder PFG marker; lanes B to K, R.befiii, R. japonica, "R helvetica," "R. massiliae," R. montana,"R slovaca," R parkeri, Itt rickettsia, "R africae," and Tttrickettsia, respectively; lane M, R rickettsii; lanes P to S: Rconorii, R australis, R sibirica, and R. akan, respectively. Migra-tion was performed in 0.5x TBE at 14WC at 5.4 V/cm for 33 h withramped pulse times from 5 to 20 s. The positions and sizes of markerbands are indicated at the left.
former USSR) obtained from both humans (130114, 130126,151820, 180522, 060518, and 16B) and ticks (Kenya ticktyphus, Ml, and strain 7) were compared. PFGE patternsafter enzymatic digestion with SmaI (Fig. 7), EagI, andBssHII were extremely well conserved.We obtained identical patterns for all the strains when we
repeated enzymatic digestion and plug preparation.Estimation of chromosome size. It was possible to obtain
consistent values with the three endonucleases tested. Forseven of the new strains tested, the average genome size wasbetween 1,222 and 1,276 kb ("R slovaca," R. japonica, R.montana, Ttt rickettsia, "R. africae," R parkeri, and Ittrickettsia). For two strains, it was close to 1,400 kb (1,370and 1,397 kb for "R. massiliae" and "R. helvetica," respec-tively), and the calculated size forR beiii was 1,660 kb. ForR rickettsii it was 1,272 kb (Table 2).
Numerical analysis. The graphic representation of thenumerical analysis is presented in Fig. 8. There is a clusterincluding 8 of the 16 SFG rickettsiae tested (R. conorii, Rrickettsii, Itt rickettsia, R parkepi, "R africae," "R. slo-vaca," R. sibirica, and Ttt rickettsia). Among these strains,R conorii and Itt rickettsia are relatively close (28% diver-gence). The other species, except R montana and "R.massifiae" (50% divergence), which are grouped together,are isolated. R beiii and "R helvetica" are the most distantfrom the cluster described above.Mapping of amplified genes on the rickettsial chromosome.
We determined the locations of three genes, the genes
kb500
400
300
250
200
150
120
100
80*** 70
- ** 60
50
40
30
20
15
J. BACTERIOL.
-.*.. -*...
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

PHYLOGENETIC ANALYSIS OF SFG RICKETTSIAE 4899
R. conori Ttt rickettsia R. parkefi "R. slovaca R. montana "R. massiliae R. akar R. be/lli
kb R. rickettsfi itt rickettsia "R. africae R. sibirica R. japonica R. rhipicephali R. australis "R. helvetica '1000
800
600
500
400
300
250
200
150
120
100
807060 - -
50
40 - -* - -
30 **
20
15
10
1273 1234 1234 1238 1259 1219 1233 1235 1268 1238 1252 1349 1276 1235 1387 1682
FIG. 4. Schematic representation of PFGE profiles obtained after EagI digestion of 16 rickettsial strain genomes. The number of asterisksindicates the number of fragments of identical size inside a given band, as revealed by densitometry using The Imager apparatus and theQGEL-1D program. Numbers at the bottom of the figure indicate the estimated sizes (in kilobases) of the bands obtained after enzymaticdigestion and PFGE.
encoding surface proteins of 120 and 190 kDa and the citratesynthase gene (Table 3). The hybridizations were performedwith the PFGE gels after migration at 5.7 V/cm for 3 to 10 s
for 24 h after digestion with EagI and SmaI, except for "Rafricae" and R montana (for the citrate synthase and190-kDa surface protein genes) and for EagI digestion of "R.helvetica" (for the three genes considered), for which it wasnecessary to use 5.4 V/cm for 5 to 20 s for 33 h. Afterdigestion with BssHII, the migration conditions were 4.5V/cm for 5 to 120 s for 40 h. It was possible to locate thecitrate synthase gene under high-stringency conditions forthe 16 rickettsial strains. After SmaI DNA digestion, it waslocated on a band of between 19 and 88 kb (Fig. 9), and afterEagI digestion, it was on a band of between 43 and 305 kb.After BssHII digestion, it was found on a larger fragment ofmore than 500 kb for R. rickettsii, "R. africae," Ttt rickett-sia,R sibinica, "R. slovaca," and R. montana and less than500 kb for the other strains. Under high-stringency condi-tions, we obtained stained bands for 12 strains after hybrid-ization of the PCR products derived from the 190- and120-kDa surface protein genes. WithR australis, the signalexisted but was very weak. The gene encoding the 190-kDasurface protein was on the same fragment as the citratesynthase gene. No signal was detected in "R helvetica,"Rakari, or R bellii, but under low-stringency conditions welocated the 190-kDa surface protein gene on a band different
from the one where the citrate synthase gene for "R.helvetica" and R. akari was located after SmaI digestion.The size of the fragment was identical for R. beiii after SmiaIdigestion, but it was impossible to determine whether it wasthe same one, because we obtained comigrating bands at thislevel. The same observation was valid for R australis.Hybridization with the 120-kDa surface protein gene on gelsobtained after SmaI digestion showed its location on a
100-kb fragment for six rickettsiae (R. conorii, Itt rickettsia,"R africae," R parkeri, R. sibirica, and R. japonica), on a
140-kb fragment for five rickettsiae (R. rickettsii, Ttt rickett-sia, "R slovaca," R. montana, and "R. massiliae"), on a
band of 128 kb forR rhipicephali, and on a band of 260 kbfor R australis. Under low-stringency conditions, we lo-cated the genes of "R helvetica," R. akari, andR belii on
bands of 221, 225, and 60 kb, respectively. As with EagIdigestion, the gene was located on a band of 104 to 111 kb formost strains. It was noted that the three genes were locatedon the same fragment after EagI digestion of "R. helvetica."For Ttt rickettsia, we obtained comigrating bands at thislevel, so it was impossible to conclude whether the threegenes were on the same fragment. For the profiles obtainedafter digestion with BssHII, the gene was located on thesame band as the other two genes for R. nickettsii, R.cononii, "R africae," R. sibirica, "R slovaca," R. mon-
tana, andR akari and on another band for the other strains.
kb1300
1000
800
600
500
400
300
250
200
150
120
100
8070
60
50
40
30
20
15
10
VOL. 175, 1993
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

4900 ROUX AND RAOULT
A
KB
485
97-
ABC D E F G H I J K L M NO P Q R S T UV
B
KB
145.5-
48.5-
A B C D EF G H I J K LM N 0 PQRST UV
FIG. 5. PFGE of BssHII restriction fragments of rickettsialDNAs. (A) Lanes A, L, N, P, and V, lambda ladder PFG marker;lanes B to F, R. bellii, R. japonica, R montana, "R massiliae,"and "R. helvetica," respectively; lane G, yeast chromosome PFGmarker; lanes H to K, "R. slovaca," R. parkeri, Itt rickettsia, and"R africae," respectively; lanes M and 0, R nckettsii and Rrhipicephali, respectively; lanes Q to U, Ttt rickettsia, R. cononi, Rak/an, R. australis, and R sibirica, respectively. Migration was
performed in 0.5 x TBE at 14'C at 4.5 V/cm for 48 h with rampedpulse times from 5 to 120 s. (B) Lanes A, G, M, 0, R, and V, lambdaladder PFG marker; lanes B to F, R. bellii, R. japonica,R montana,"R. massiliae," and "R. helvetica," respectively; lanes H to L, "R.slovaca," R. parken, Itt rickettsia, "R. africae," and R. nckettsii,respectively; lanes N, R. rhipicephali; lanes P and Q, Ttt rickettsiaandR cononii, respectively; lanes S to U,R ak/an, R australis, andR sibinica, respectively. Migration was performed in 0.5 x TBE at14'C at 5.7 V/cm for 24 h with ramped pulse times from 3 to 10 s. Thepositions and sizes of marker bands are indicated at the left.
DISCUSSION
PFGE is a good technique for studying total bacterialDNA, and the DNA preparation method as modified bySchwartz and Cantor (50) makes it possible to preserve
genome integrity with a view to subsequent investigations.We used the contour-clamped homogeneous electric fieldsystem described by Chu et al. in 1986 (18), which gives goodresolution of the fragments generated by enzymatic diges-tion. This allows good evaluation of their size and compar-
ison of the profiles of different species. In this work, we
estimated the genome sizes for 13 of the strains to bebetween 1,200 and 1,300 kb. This is low compared with sizesfor other bacteria of the family Rickettsiaceae, such as
Rickettsiella (1,700 to 2,100 kb) (24) and Coxiella (1,600 kb)(31). The "R massiliae" (1,370-kb) and "R. helvetica"(1,397-kb) chromosomes are a little larger. The size of theRbelli genome (1,660 kb) is significantly different from thoseof other rickettsiae. This is not surprising, since most
rickettsiologists think that this bacterium does not belong in
the SFG (40, 44). In our previous work (48), we estimatedthe genome size of R. nckettsdi as 2,163 kb. At that time, wespeculated that the differences between this species andmost of the others could be due to mycoplasma contamina-tion. This hypothesis was abandoned, because after hybrid-ization with R. conorii total DNA on the PFGE patterns ofR rickettsii we stained all the bands. Moreover, the esti-mated size corresponded with that established by the rena-turation rate method of Gillis et al. and Myers and Wisseman(25, 38). Unfortunately, the R conorii strain was alsomycoplasma contaminated, but this was not apparent, be-cause mycoplasmas did not grow in the agar. This problemdid not occur when PFGE was used, because only a fewexperiments were done with intracellular bacteria. The studyof the R. rickettsii genome with a mycoplasma-free strainshows a mean genome size of 1,272 kb.
It was possible to obtain different patterns with all thestrains studied, showing that PFGE is actually the best wayto identify rickettsial strains. In fact, PCR-RFLP analysis isa good method for rapid identification of new isolates, butsometimes it is not appropriate. For example, "R. africae"andR parken PCR amplification products obtained by usingthe three primer pairs described in Materials and Methodsproduced identical patterns after enzymatic digestion (20).Using PFGE, we also found small differences between theGS strain and "R. massiliae" when PCR-RFLP analysisshowed none (5). However, PFGE showed identical profilesfor isolates of the same species (10 isolates of R. conoriiexhibited exactly the same profiles). Moreover, the sameobservation was made with different isolates of R sibinca(21) and R. ak/an (19). This favors the use of this method foran interspecies classification tool.Using the profiles obtained by PFGE, we established a
phylogenetic dendrogram of the SFG rickettsiae. The SFGrickettsiae were first classified according to geographicalcriteria. Serotyping by complement fixation (42) and thentoxin neutralization tests in mice (10) allowed differentiationof serotypes, serogroups, and strains. In 1978, Philip et al.reported a method for serological typing using microimmu-nofluorescence with mouse polyclonal antisera (41). This iscurrently the reference method for identifying SFG rickett-siae. Ten species are currently recognized by this technique(51): R nickettsii, R. cononii, R sibinrca, R. parkeni, R.australis, R. akari, R montana, R rhipicephali, R. beiii,andR japonica (54). An additional six species are proposed:"R. helvetica" (15), "R slovaca" (45), Itt rickettsia (29), Tttrickettsia (46), "R africae" (34, 39), and "R massiliae" (6,7). Among these rickettsiae, some (such as R. conorni, R.rickettsii, R australis, R. akari, R. sibirica, R. japonica, Ittrickettsia, and "R africae") are pathogenic for humans andare responsible for tick-borne spotted fevers in which theprincipal symptoms consist of fever, headache, and erup-tion. The others have been isolated only from ticks (R.montana, "R massiliae," R. parkeri, "R. helvetica," R.rhipicephali, "R slovaca," R beiii, and Ttt rickettsia), andtheir pathogenic role in humans is unknown. In their work,Philip et al. (41) described 12 serotypes clustered in severalsubgroups; one includedR fickettsii, R conorii, R sibirica,and R. parkeri, with R. montana, R. australis, R. rhipiceph-ali, andR ak/an each in independent subgroups. Sequencingof the 16S rRNA has allowed the comparison of R. fickettsiiwith other members of the family Rickettsiaceae but notwith other SFG rickettsiae (58). By genetic criteria, twostrains belong to the same species if they are characterizedby >70% relatedness by DNA-DNA hybridization (56).Preliminary reports of these values have been made for R.
J. BACTERIOL.
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

PHYLOGENETIC ANALYSIS OF SFG RICKETTSIAE 4901
R. conori Ttt nckettsia R. parked 'R.kb R. rlckettsii Itt nckettsia "R. africae " R. sibirica
1500
1200
1000
800
600
500
400
300
250
200
150
120100
80706050
40
slovaca" R. montana "R. massiliae " R. akari R. be/llR. japonica R. rhipicephali R. australis "R. helvetica "
30
20 - 20
1515
1288 1246 1216 1257 1258 1240 1258 1268 1289 1259 1258 1398 1276 1232 1444 1660
FIG. 6. Schematic representation ofPFGE profiles obtained after BssHII digestion of 16 rickettsial strain genomes. Numbers at the bottomof the figure indicate the estimated sizes (in kilobases) of the bands obtained after enzymatic digestion and PFGE.
rickettsii and other species. Thus,R rickettsii, R. conorii (91to 94%), R. sibirica (70 to 74%), and R montana (73%)would belong to the same species, butR australis (53%) andR akari (46%) would be considered different species (55).The study of Ralph et al. comparing Southern hybridizationprofiles allowed the identification of a subgroup of rickettsiae(2.1% divergence) including R rickettsii, R parkeri, "R.
africae," R sibirica, "R. slovaca," and R. conorhi; R.
montana and R. rhipicephali were distant but related to this
KB
97
48.5
23.1-
A B C D E F G H I J KFIG. 7. PFGE of SmaI restriction fragments of different isolates
of R conorii. Lane A, lambda ladder PFG marker; lanes B to J,Moroccan strain, MAZ, ORT, FER, REV, 16B, Kenya tick typhusrickettsia, Ml, and strain 7, respectively; lane K, low-range PFGmarker. Migration was performed in 0.5 x TBE at 14WC at 5.7 V/cmfor 24 h with ramped pulse times from 3 to 10 s. The positions andsizes of marker bands are indicated at the left.
subgroup. R australis, R akari, andR belii were indepen-dent and distant from the aforesaid strains (43).The dendrogram established by Regnery et al. (44) after
study of the patterns obtained after PCR-RFLP analysis ofthe citrate synthase gene showed that R befii was distantfrom the SFG and the typhus group. R. australis and R
TABLE 2. Estimated rickettsia genome sizes after DNAdigestion with EagI, SmaI, and BssHII and separation
by PFGE of the fragments obtained
Genome size (kb) for enzyme Mean sizeRickettsia
EagI SmaI BssHII (kb)
R rickettsii 1,273 1,255 1,288 1,272R. conorii 1,234 1,230 1,246 1,237Itt rickettsia 1,234 1,217 1,216 1,222Ttt rickettsia 1,238 1,220 1,257 1,238"R africae" 1,259 1,237 1,258 1,251R parkeri 1,219 1,246 1,240 1,235R sibirica 1,233 1,238 1,258 1,243"R slovaca" 1,235 1,242 1,268 1,248R japonica 1,268 1,272 1,289 1,276R montana 1,238 1,252 1,259 1,250R rhipicephali 1,252 1,257 1,258 1,256"R. massiliae" 1,349 1,363 1,398 1,370R australis 1,276 1,256 1,276 1,269R akari 1,235 1,221 1,232 1,229"R helvetica" 1,387 1,361 1,444 1,397R. beMNi 1,682 1,639 1,660 1,660
kb1500
12001000
800
600500
400
300250
200
150
120100
80706050
40
30
VOL. 175, 1993
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
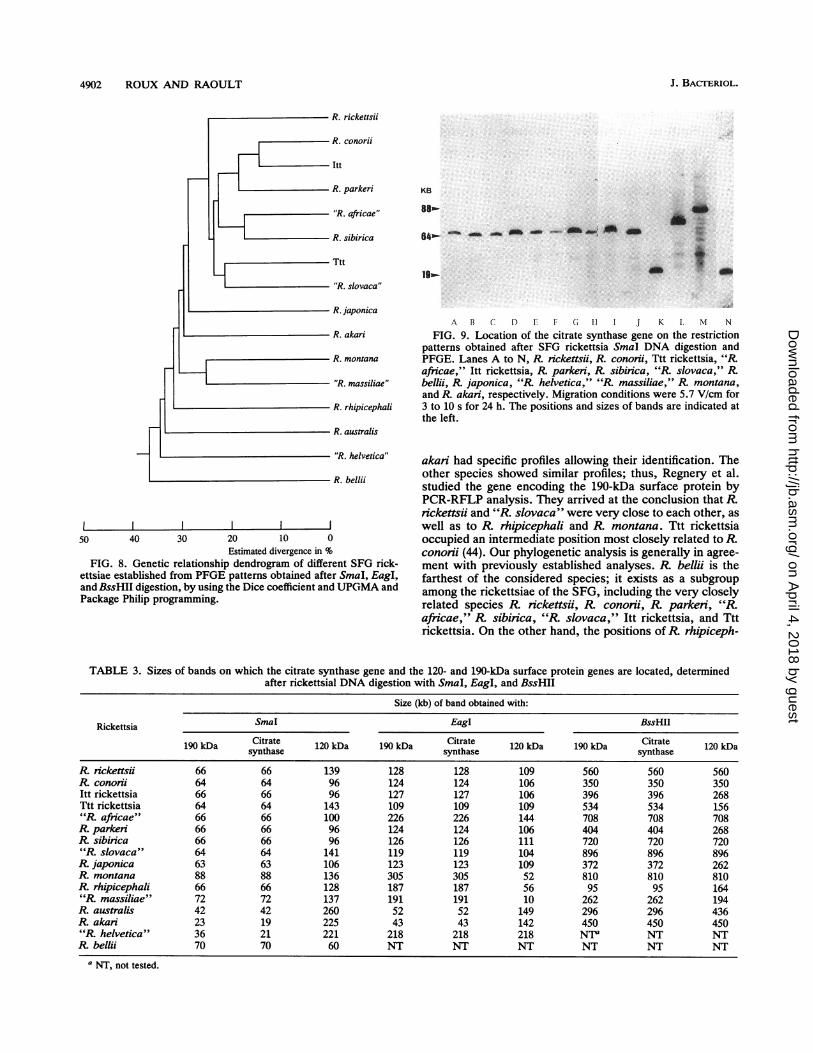
4902 ROUX AND RAOULT
A.
KB i$
I
50 40 30 20 10 0
Estimated divergence in %FIG. 8. Genetic relationship dendrogram of different SFG rick-
ettsiae established from PFGE patterns obtained after SmaI, EagI,and BssHII digestion, by using the Dice coefficient and UPGMA andPackage Philip programming.
A B C D E F G H I J K L M N
FIG. 9. Location of the citrate synthase gene on the restrictionpatterns obtained after SFG rickettsia SmaI DNA digestion andPFGE. Lanes A to N, R rickettsii, R. conorii, Ttt rickettsia, "Rafricae," Itt rickettsia, R parkeri, R sibirica, "R. slovaca," Rbeli, R japonica, "R helvetica," "R. massiliae," R. montana,and R. akari, respectively. Migration conditions were 5.7 V/cm for3 to 10 s for 24 h. The positions and sizes of bands are indicated atthe left.
akari had specific profiles allowing their identification. Theother species showed similar profiles; thus, Regnery et al.studied the gene encoding the 190-kDa surface protein byPCR-RFLP analysis. They arrived at the conclusion that R.rickettsii and "R. slovaca" were very close to each other, aswell as to R rhipicephali and R. montana. Ttt rickettsiaoccupied an intermediate position most closely related to R.conorii (44). Our phylogenetic analysis is generally in agree-ment with previously established analyses. R. beiii is thefarthest of the considered species; it exists as a subgroupamong the rickettsiae of the SFG, including the very closelyrelated species R. rickettsii, R conorii, R. parkeri, "R.africae," R sibinica, "R slovaca," Itt rickettsia, and Tttrickettsia. On the other hand, the positions ofR rhipiceph-
TABLE 3. Sizes of bands on which the citrate synthase gene and the 120- and 190-kDa surface protein genes are located, determinedafter rickettsial DNA digestion with SmaI, EagI, and BssHII
Size (kb) of band obtained with:
Rickettsia SmaI EagI BssHII
190 kDa Citrate 120 kDa 190 kDa Citrate 120 kDa 190 kDa Citrate 120 kDasynthase synthase synthase
R rickettsii 66 66 139 128 128 109 560 560 560R. conorii 64 64 96 124 124 106 350 350 350Itt rickettsia 66 66 96 127 127 106 396 396 268Ttt rickettsia 64 64 143 109 109 109 534 534 156"K africae" 66 66 100 226 226 144 708 708 708R. parkeri 66 66 96 124 124 106 404 404 268R sibirica 66 66 96 126 126 111 720 720 720"R slovaca" 64 64 141 119 119 104 896 896 896R japonica 63 63 106 123 123 109 372 372 262R montana 88 88 136 305 305 52 810 810 810R rhipicephali 66 66 128 187 187 56 95 95 164"R massiliae" 72 72 137 191 191 10 262 262 194R australis 42 42 260 52 52 149 296 296 436R akan 23 19 225 43 43 142 450 450 450"R helvetica" 36 21 221 218 218 218 NF NT NTR belldi 70 70 60 NT NT NT NT NT NT
a NT, not tested.
J. BACTERIOL.
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

PHYLOGENETIC ANALYSIS OF SFG RICKETTSIAE 4903
ali, R. montana, R. australis, and R. akari vary dependingon the methods of testing but are distinctly different from theothers. As for R. japonica, "R. helvetica," and "R. massil-iae," our work is the first in which these species have beentaken into consideration, so no comparison is possible.However, "R helvetica" is as distant as R. beMii from theother species. R japonica is close to the subgroup definedabove. "R massiliae" is grouped with R montana. Finally,our work allowed us to classify SFG rickettsiae into eightsubgroups, which probably represent different species. Thefirst consists of eight serotypes, including five pathogenicrickettsiae, isolated in Europe, Africa, Asia, and/or theUnited States. The second includes R montana, and "R.massiliae." R rhipicephali, R. australis, R. ak/an,R japon-ica, "R. helvetica," and R. beNii are sole members of theirsubgroups.
Five genes have currently been cloned and sequenced: thegenes encoding citrate synthase (62), ADP/ATP translocase(61), and three surface proteins of 17 (3), 120 (27), and 190 (2)kDa in R nickettsii. We have located three of them on ourprofiles. Knowing the high percentage of DNA-DNA hybrid-ization between strains of the SFG rickettsiae, we hoped thatit would be possible to obtain stained bands with all thestrains by using R. conorii PCR amplification products. Wefound that the genes encoding citrate synthase and the190-kDa surface protein were usually located on the samefragment, showing that these two genes are close on therickettsial chromosome. After EagI digestion of the "R.helvetica" genome, we stained the same band with the threegenes. This observation shows that the gene encoding the120-kDa surface protein would be close to the other two. Itwas not possible to locate the 120- and 190-kDa surfaceprotein genes after hybridization under high-stringency con-ditions for "R. helvetica," R. ak/an, and R belii, and weobtained a weak signal for R. australis. This is not surpris-ing, since the primers described here do not amplify thesespecies, and it shows that the percentages of hybridizationbetween R australis, R ak/an, "R. helvetica," or R. belliandR conori are low. We conclude that these two genes arepresent on the rickettsial genomes but that the primers beingused currently are not suitable. However, it was possible tolocate the citrate synthase gene for all the species underhigh-stringency conditions. Thus, it can be deduced that thisgene is more highly conserved than the other two.We obtained a size evaluation of the SFG rickettsia
genomes, characteristic profiles for each strain, the locationof three genes, and the basis for a genomic classification.PFGE is a powerful technique which gives interesting resultsbecause, in contrast to other methods, it allows the study ofthe total intact rickettsial DNA. However, it is not the bestmethod to define criteria for species identification among theSFG rickettsiae. These criteria can probably be establishedby sequencing the 16S and 23S rRNAs.
ACKNOWLEDGMENTS
We are grateful to Herv6 Tissot-Dupont for his technical assis-tance and to David Walker for reviewing the manuscript.
REFERENCES1. Allardet-Servent, A., G. Bourg, M. Ramuz, M. Pages, M. Bellis,
and G. Roizes. 1988. DNA polymorphism in strains of the genusBrucella. J. Bacteriol. 170:4603-4607.
2. Anderson, B. E., G. A. McDonald, D. C. Jones, and R. L.Regnery. 1990. A protective protein antigen of Rickettsia rick-ettsii has tandemly repeated, near-identical sequences. Infect.Immun. 58:2760-2769.
3. Anderson, B. E., R. L. Regnery, G. M. Carlone, T. Tzianabos,J. E. McDade, Z. Y. Fu, and W. J. Bellini. 1987. Sequenceanalysis of the 17-kilodalton-antigen gene from Rickettsia rick-ettsii. J. Bacteriol. 169:2385-2390.
4. Andrew, R., J. M. Bonnin, and S. Williams. 1946. Tick typhus inNorth Queensland. Med. J. Aust. 2:253.
5. Babalis, T., Y. Tselensis, V. Roux, A. Psaroulaki, and D. Raoult.Submitted for publication.
6. Beati, L., J.-P. Finidori, B. Gilot, and D. Raoult. 1992. Compar-ison of serologic typing, sodium dodecyl sulfate-polyacrylamidegel electrophoresis protein analysis, and genetic restrictionfragment length polymorphism analysis for identification ofrickettsiae: characterization of two new rickettsial strains. J.Clin. Microbiol. 30:1922-1930.
7. Beati, L., V. Roux, M. Eremeeva, X. J. Yu, H. Tissot-Dupont,and D. Raoult. 1992. Rickettsia massiliae (sp. nov.), poster 19.Tenth sesquiannual meeting of the American Society for Rick-ettsiology and Rickettsial Diseases, Hamilton, Montana.
8. Bell, E. J., G. M. Kohls, H. G. Stoenner, and D. B. Lackman.1963. Non pathogenic rickettsias related to the spotted fevergroup isolated from ticks, Dernnacentor vaniabilis and Derma-centor andersoni from Eastern Montana. J. Immunol. 90:770-781.
9. Bell, E. J., and E. G. Pickens. 1953. A toxic substance associ-ated with the rickettsias of the spotted fever group. J. Immunol.70:461.
10. Bell, E. J., and H. G. Stoenner. 1960. Immunologic relationshipsamong the spotted fever group of rickettsias determined bytoxin neutralisation tests in mice with convalescent animalserums. J. Immunol. 84:171-182.
11. Birkelund, S., and R. S. Stephens. 1992. Construction of phys-ical and genetic maps of Chlamydia trachomatis serovar L2 bypulsed-field gel electrophoresis. J. Bacteriol. 174:2742-2747.
12. Birren, B. W., E. Lai, S. M. Clark, L. Hood, and M. I. Simon.1988. Optimized conditions for pulsed field gel electrophoreticseparations of DNA. Nucleic Acids Res. 15:7563-7581.
13. Bollet, C., and P. de Micco. 1992. Taxonomie bacterienne, p.23-57. In J. Freney, F. Reynaud, W. Hansen, and C. Bollet(ed.), Manuel de bacteriologie clinique. Elsevier, Paris.
14. Brosch, R., C. Buchrieser, and J. Rocourt. 1991. Subtyping ofListeria monocytogenes serovar 4b by use of low-frequency-cleavage restriction endonucleases and pulsed-field gel electro-phoresis. Res. Microbiol. 142:667-675.
15. Burgdorfer, W., A. Aeschlimann, 0. Peter, S. F. Hayes, andR. N. Philip. 1979. Ixodes ricinus: vector of a hitherto unde-scribed spotted fever group agent in Switzerland. Acta Trop.39:357-367.
16. Burgdorfer, W., L. P. Brinton, W. L. Krinsky, and R. N. Philip.1978. Rickettsia rhipicephali: a new spotted fever group rickett-sia from the brown dog tick Rhipicephalus sanguineus, p.307-316. In J. Kazar, R. A. Ormsbee, and I. N. Tarasevich(ed.), Rickettsiae and rickettsial diseases. Publishing House ofthe Slovak Academy of Sciences, Bratislava, Slovakia.
17. Bygraves, J. A., and M. C. J. Maiden. 1992. Analysis of theclonal relationships between strains of Neissenia meningitidisby pulsed-field gel electrophoresis. J. Gen. Microbiol. 138:523-531.
18. Chu, G., D. Vollrath, and R. W. Davis. 1986. Separation of largeDNA molecules by contour-clamped homogeneous electricfield. Science 234:1582-1585.
19. Eremeeva, M., and D. Raoult. Unpublished data.20. Eremeeva, M., X. Yu, and D. Raoult. Submitted for publication.21. Eremeeva, M. E., N. M. Balayeva, V. F. Ignatovich, and D.
Raoult. Submitted for publication.22. Feinberg, A. P., and B. Vogelstein. 1983. A technique for
radiolabeling DNA restriction endonuclease fragments to highspecific activity. Anal. Biochem. 132:6-13.
23. Feinberg, A. P., and B. Vogelstein. 1984. A technique forradiolabeling DNA restriction endonuclease fragments to highspecific activity. Anal. Biochem. 137:266-267. (Addendum.)
24. Frutos, R., M. Pages, M. Bellis, G. Roizes, and M. Bergoin. 1989.Pulsed-field gel electrophoresis determination of the genomesize of obligate intracellular bacteria belonging to the genera
VOL. 175, 1993
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

4904 ROUX AND RAOULT
Chlamydia, Rickettsiella, and Porochlamydia. J. Bacteriol. 171:4511-4513.
25. Gillis, M., J. De Lay, and M. De Cleene. 1970. The determinationof molecular weight of bacterial genome DNA from renaturationrates. Eur. J. Biochem. 12:143-153.
26. Gilmore, R. D., Jr., and T. Hackstadt. 1991. DNA polymor-phism in the conserved 190 kDa antigen gene repeat regionamong spotted fever group rickettsiae. Biochim. Biophys. Acta1097:77-80.
27. Gilmore, R. D., Jr., N. Joste, and G. A. McDonald. 1989.Cloning, expression and sequence analysis of the gene encodingthe 120 kDa surface-exposed protein of Rickettsia nckettsii.Mol. Microbiol. 3:1579-1586.
28. Gimenez, D. F. 1964. Staining Rickettsiae in yolk-sac cultures.Stain Technol. 39:135-140.
29. Goldwasser, R. A., Y. Steiman, W. Klingberg, T. A. Swartz, andM. A. Klingberg. 1974. The isolation of strains of rickettsiae ofthe spotted fever group in Israel and their differentiation fromother members of the group by immunofluorescence methods.Scand. J. Infect. Dis. 6:53-62.
30. Golinevitch, H. 1960. A propos de la differenciation de quelquesrickettsies du groupe de la fievre pourpree a tiques. Arch. Inst.Pasteur Tunis 37:13-22.
31. Heinzen, R., G. L. Stiegler, L. L. Whiting, S. A. Schmitt, L. P.Mallavia, and M. E. Frazier. 1990. Use of pulsed field gelelectrophoresis to differentiate Coxiella bumetii strains. Ann.N.Y. Acad. Sci. 590:504-513.
32. Huebner, R. J., and C. Armstrong. 1946. Rickettsialpox-anewly recognized rickettsial disease. I. Isolation of the etiolog-ical agent. Public Health Rep. 61:1605.
33. Kauc, L., M. Mitchell, and S. H. Goodgal. 1989. Size andphysical map of the chromosome of Haemophilus influenzae. J.Bacteriol. 171:2474-2479.
34. Kelly, P., L. Matthewman, L. Beati, D. Raoult, P. Mason, M.Deary, and R. Makombe. 1992. African tick-bite fever: a newspotted fever group rickettsiosis under an old name. Lancet340:382-383.
35. Mallavia, L. P. 1991. Genetics of rickettsiae. Eur. J. Epidemiol.7:213-221.
36. Marrero, M., and D. Raoult. 1989. Centrifugation shell vialtechnique for rapid detection of mediterranean spotted fever inblood culture. Am. J. Trop. Med. Hyg. 40:197-199.
37. Meskini, M., and D. Raoult. Unpublished data.38. Myers, W. F., and C. L. Wisseman, Jr. 1981. The taxonomic
relationship of Rickettsia canada to the typhus and spottedfever groups of the genus Rickettsia, p. 313-325. In W. Burg-dorfer and L. R. Anacker (ed.), Rickettsiae and rickettsialdiseases. Academic Press, New York.
39. Philip, C. B., H. Hoogstraal, R. Reiss-Gutfreund, and C. M.Cliford. 1966. Evidence of rickettsial disease agents in ticksfrom Ethiopian cattle. Bull. W.H.O. 35:127-131.
40. Philip, R. N., E. A. Casper, R. L. Anacker, J. Cory, S. F. Hayes,W. Burgdorfer, and C. E. Yunker. 1983. Rickettsia bellii sp.nov.: tick-borne rickettsia, widely distributed in the UnitedStates, that is distinct from the spotted fever and typhusbiogroups. Int. J. Syst. Bacteriol. 33:94-106.
41. Philip, R. N., E. A. Casper, W. Burgdorfer, R. K. Gerloff, L. E.Hughes, and E. J. Bell. 1978. Serologic typing of Rickettsiae ofspotted fever group by microimmunofluorescence. J. Immunol.121:1961-1968.
42. Plotz, H., R. L. Reagan, and K. Wertman. 1944. Differentiationbetween fievre boutonneuse and rocky mountain spotted feverby means of complement fixation. Proc. Soc. Exp. Biol. Med.55:173-176.
43. Ralph, D., C. Pretzman, N. Daugherty, and K. Poetter. 1990.Genetic relationships among the members of the family Rickett-siaceae as shown by DNA restriction fragment polymorphismanalysis. Ann. N. Y. Acad. Sci. 590:541-553.
44. Regnery, R. L., C. L. Spruill, and B. D. Plikaytis. 1991.Genotypic identification of rickettsiae and estimation of in-
traspecies sequence divergence for portions of two rickettsialgenes. J. Bacteriol. 173:1576-1589.
45. Rehacek, J. 1984. Rickettsia slovaca, the organism and itsecology. Acta Sci. Nat. Acad. Sci. Bohemoslov. Brno 18(2):1-50.
46. Robertson, R. G., and C. L. Wisseman, Jr. 1972. Tick-bornerickettsiae of the spotted fever group in West Pakistan. II.Serological classification of isolates from West Pakistan andThailand: evidence for two new species. Am. J. Epidemiol.97:55-64.
47. Romling, U., and B. Tfimmler. 1991. The impact of two-dimensional pulsed-field gel electrophoresis techniques for theconsistent and complete mapping of bacterial genomes: refinedphysical map of Pseudomonas aeruginosa PAO. Nucleic AcidsRes. 19:3199-3206.
48. Roux, V., M. Drancourt, and D. Raoult. 1992. Determination ofgenome sizes of Rickettsia spp. within the spotted fever group,using pulsed-field gel electrophoresis. J. Bacteriol. 174:7455-7457.
49. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning. A laboratory manual, 2nd ed. Cold Spring HarborLaboratory Press, Cold Spring Harbor, N.Y.
50. Schwartz, D. C., and C. R. Cantor. 1984. Separation of yeastchromosome-sized DNAs by pulsed field gradient gel electro-phoresis. Cell 37:67-75.
51. Skerman, V. B. D., V. McGowan, and P. H. A. Sneath (ed.).1989. Approved lists of bacterial names. American Society forMicrobiology, Washington, D.C.
52. Smith, C. L., J. G. Econome, A. Schutt, S. Klco, and C. R.Cantor. 1987. A physical map of the Escherichia coli K12genome. Science 236:1448-1453.
53. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
54. Uchida, T., T. Uchiyama, K. Kumano, and D. H. Walker. 1992.Rickettsia japonica sp. nov., the etiological agent of spottedfever group rickettsiosis in Japan. Int. J. Syst. Bacteriol.42:303-305.
55. Walker, D. H. 1989. Rocky mountain spotted fever: a disease inneed of microbiological concern. Clin. Microbiol. Rev. 2:227-240.
56. Wayne, L. G., D. J. Brenner, R. R. Colwell, P. A. D. Grimont,0. Kandler, M. I. Krichevsky, L. H. Moore, W. E. C. Moore,R. G. E. Murray, E. Stackebrandt, M. P. Starr, and H. G.Triiper. 1987. Report of the ad hoc committee on reconciliationof approaches to bacterial systematics. Int. J. Syst. Bacteriol.37:463-464.
57. Weil, M. D., and M. McClelland. 1989. Enzymatic cleavage of abacterial genome at a 10-base-pair recognition site. Proc. Natl.Acad. Sci. USA 86:5-55.
58. Weisburg, W. G., M. E. Dobson, J. E. Samuel, G. A. Dasch,L. P. Mallavia, 0. Baca, L. Mandelco, J. E. Sechrest, E. Weiss,and C. R. Woese. 1989. Phylogenetic diversity of the rickettsiae.J. Bacteriol. 171:4202-4206.
59. Weiss, E., J. C. Coolbaugh, and J. C. Williams. 1975. Separationof viable Rickettsia typhi from yolk sac and L cell host compo-nents by Renografin density gradient centrifugation. Appl. Mi-crobiol. 30:456-463.
60. Weiss, E., and J. W. Moulder. 1984. Order I. RickettsialesGieszczkiewicz 1939, 25k', p. 687-701. In N. R. Kreig and J. G.Holt (ed.), Bergey's manual of systematic bacteriology, vol. 1.The Williams and Wilkins Co., Baltimore.
61. Williamson, L. R., G. V. Plano, H. H. Winkler, D. C. Krause,and D. 0. Wood. 1989. Nucleotide sequence of the Rickettsiaprowazekii ATP/ADP translocase-encoding gene. Gene 80:269-278.
62. Wood, D. O., L. R. Williamson, H. H. Winkler, and D. C.Krause. 1987. Nucleotide sequence of the Rickettsia prowazekiicitrate synthase gene. J. Bacteriol. 169:3564-3572.
J. BACTERIOL.
on April 4, 2018 by guest
http://jb.asm.org/
Dow
nloaded from


















