
Genetica di popolazioni 3
32
Genetica di popolazioni 3: Inbreeding
-
Upload
genetica-ferrara-university-italy -
Category
Education
-
view
7.212 -
download
2
Transcript of Genetica di popolazioni 3

Genetica di popolazioni 3: Inbreeding

Programma del corso
1. Diversità genetica
2. Equilibrio di Hardy-Weinberg
3. Inbreeding
4. Linkage disequilibrium
5. Mutazione
6. Deriva genetica
7. Flusso genico e varianze genetiche
8. Selezione
9. Mantenimento dei polimorfismi e teoria neutrale
10. Introduzione alla teoria coalescente
11. Struttura e storia della popolazione umana
+ Lettura critica di articoli

• Quando la scelta del partner riproduttivo non è casuale rispetto al suo genotipo si parla di unione assortativa
Unione non casuale

L’unione assortativa è positiva quando si scelgono
preferenzialmente partner geneticamente affini,
negativa quando avviene il contrario
Aumentato rischio di omozigosi per alleli patologici recessiviSuddivisione del pool genico che favorisce la speciazione

• L’unione assortativa positiva provoca un deficit di eterozigoti rispetto alle attese di Hardy-Weinberg
• Il deficit di eterozigoti viene misurato dal coefficiente F di inbreeding
• Coefficienti di inbreeding possono essere stimati dalle frequenze genotipiche o dagli alberi genealogici
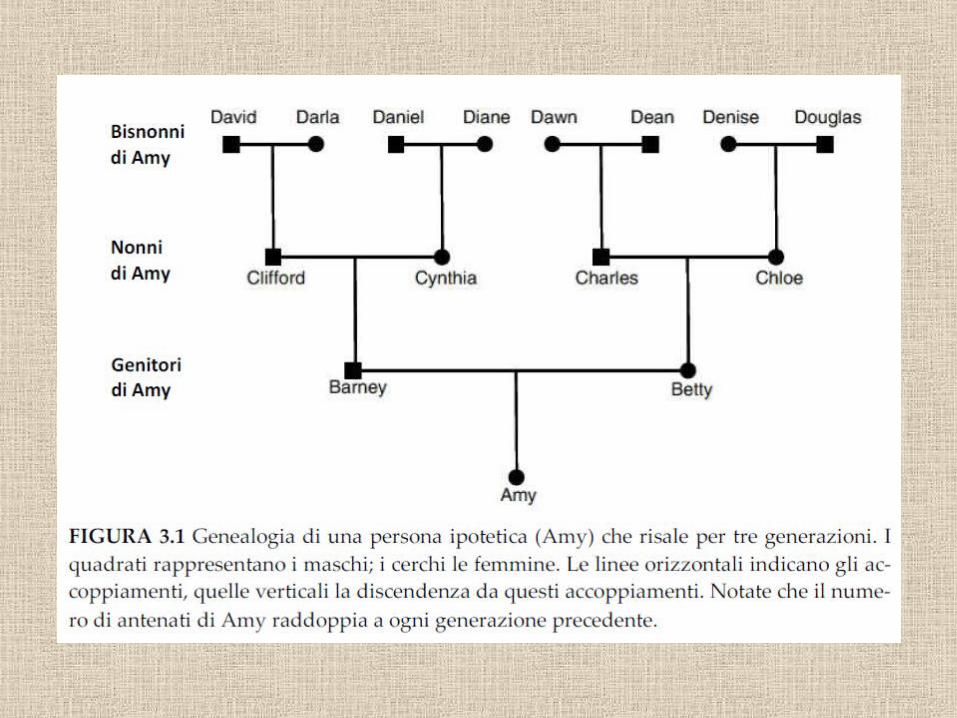
• L’inbreeding è conseguenza anche del fatto che il numero di antenati di ognuno raddoppia ad ogni generazione, mentre le popolazioni hanno dimensioni finite
Unione non casuale

Unione assortativa positiva per le dimensioni corporee:
Asellus aquaticus Gammarus pulex Plecia nearctica (isopodi) (gambero d’acqua dolce, amfipodi) (lovebug, ditteri)
Unione assortativa positiva per la morfologia
Salvelinus alpinus(salmerino, salmonidi)

In molti uccelli le femmine preferiscono accoppiarsi con maschi che cantano lo stesso dialetto cantato dai loro padri.
Zonotrichia leucophrys (white-crowned sparrow)
Agelaius phoeniceus(merlo ala rossa)
Unione assortativa positiva per caratteristiche comportamentali

An incompatible pollination in Brassica showing the inhibition of self-pollen (Po) and the inability of the emerging pollen tube (Pt) to invade the wall of a stigma epidermal cell (SE).
Effect of the S (sterility) locus.
The number of S-locus alleles is usually large, being estimated at 22 in Iberis, 34 in Raphanus, 50 in B. oleracea and 30 in B. campestris.
Da Nasrallah (1997) PNAS
Unione assortativa negativa: alleli di autoincompatibilità nelle piante

Il numero di alleli al locus S è generalmente elevato usually large, e si stima che sia 50 in Brassica oleracea.La fecondazione fallisce se il granulo pollinico ha lo stesso genotipo delle cellule ovariche.Qual è la probabilità P(x) che ci sia incrocio fra due individui che, per caso, hanno genotipo identico?
Vediamo se ci siamo capiti
Bisogna fare qualche assunzione: p. es., ogni allele ha la stessa frequenza.Con 50 alleli, ognuno ha frequenza q=0.02
P(x) = 0.022 x 50 = 0.0004 x 50 = 0.02
E, più in generale, se n è il numero degli alleli: P(x) = 1/n2 x n = 1/n
Dunque, 1 diviso il numero degli alleli è la frazione di incroci possibili che falliscono per autoincompatibilità, cioè il prezzo (riduzione di fertilità) che la pianta paga al meccanismo che evita l’autofecondazione

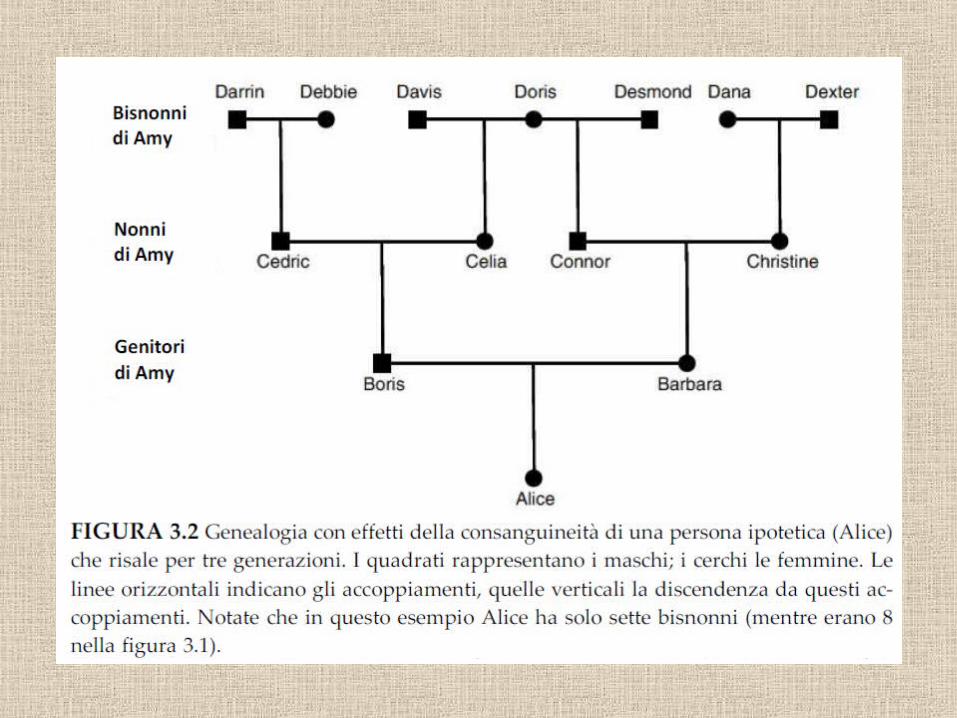
16 trisavoli
Figlio
PadreMadre
4 nonni 8 bisnonni
32 antenati 4 generazioni fa
Abbiamo tanti antenati
1750: 1024 antenati1500: 1 milione1240: 1 miliardo1000: 1000 miliardi250 aC: 1030
e ciascuno ci ha trasmesso un pezzetto del suo genoma
6 miliardi di nucleotidi nel genoma umano



Probabilità di identità per discesa di un allele
P = 1/26 = 1/64
Vale sia per A che per a
La probabilità complessiva è F = 1/32 = 0.03125
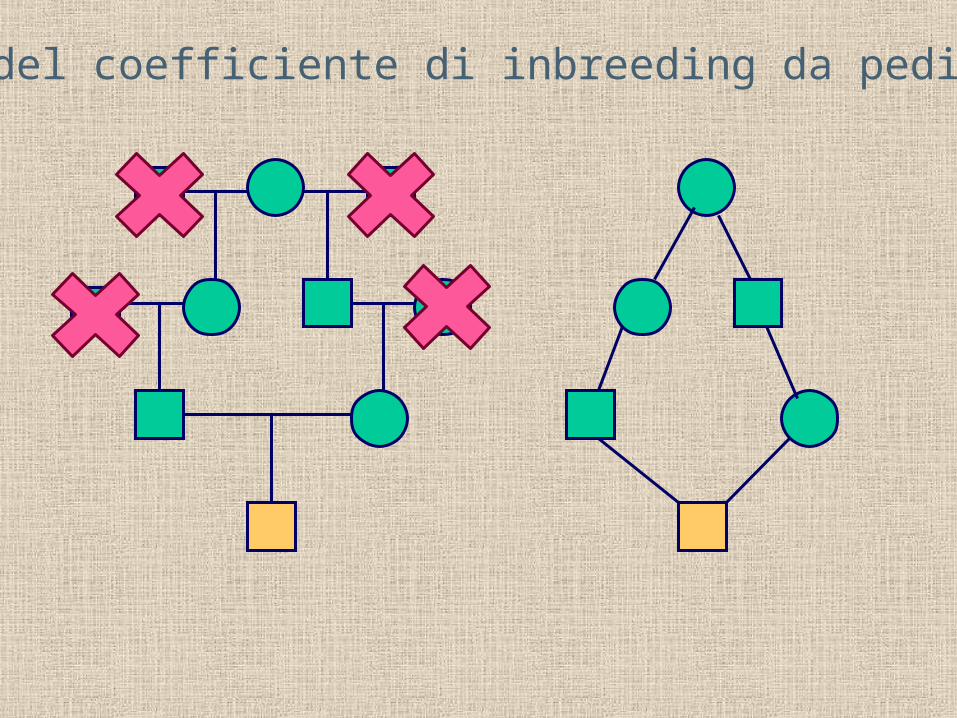
Stima del coefficiente di inbreeding da pedigree

Stima del coefficiente di inbreeding da pedigree
½
½ ½
½ ½
Aa
F = (½)5 = 1/32



Stima del coefficiente di inbreeding da pedigree
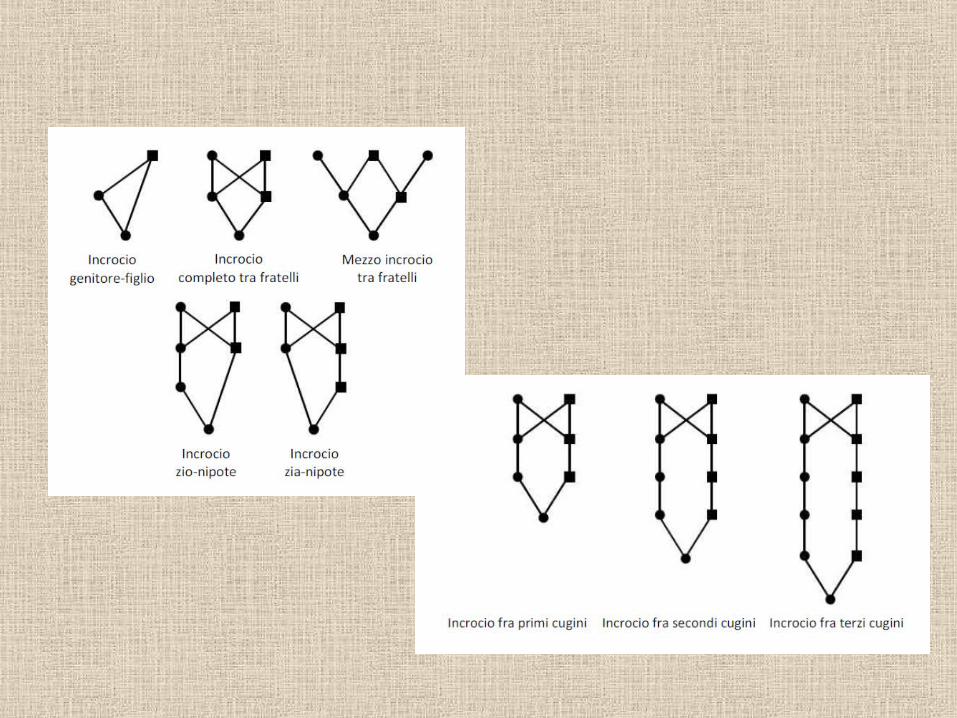
Il valore di F è pari a ½ elevato a una potenza pari al numero di passaggi nel pedigree.
Valore di F nella progenie di varie unioni consanguinee:
Autofecondazione: ½Fra fratello e sorella: ¼Fra zio e nipote: 1/8Fra cugini primi: 1/16Fra cugini 1 e ½: 1/32Fra cugini secondi: 1/64…

Inbreeding medio di una popolazione
Totale 1246 14,28125

f(AA) = ¼ f(Aa) = ½ f(aa) = ¼
Unione assortativa positiva: autofecondazione
¼ AA x AA 100% AA½ Aa x Aa ¼ AA, ½ Aa, ¼ aa¼ aa x aa 100% aa
f(AA) = 3/8 f(Aa) = ¼ f(aa) = 3/8
f(AA) = ¼ + (½ x ¼) f(Aa) = 1/4 f(aa) = ¼ + (½ x ¼)

Unione assortativa positiva: autofecondazione
3/8 AA x AA 100% AA¼ Aa x Aa ¼ AA, ½ Aa, ¼ aa3/8 aa x aa 100% aa
f(AA) = 3/8 f(Aa) = ¼ f(aa) = 3/8
f(AA) = 3/8 + (¼ x ¼) f(Aa) = 1/8 f(aa) = 3/8 + (¼ x ¼)
f(AA) = 7/16 f(Aa) = 1/8 f(aa) = 7/16

Unione assortativa positiva: autofecondazione
Generazione AA Aa aa1 ¼ ½ ¼ 2 3/8 1/4 3/83 7/16 1/8 7/164 15/32 1/16 15/32
N 1/2N

Effetti dell’inbreeding
La tendenza ad accoppiarsi fra consanguinei determina la comparsa nella progenie di un eccesso di omozigoti:

Crescita del coefficiente di inbreeding in funzione del tempo e del sistema di accoppiamento

Unione assortativa positiva: inbreeding
Se Foss(Aa) = H
Fatt(Aa) = H0 = 2pq
(H0 – H) / H0 = F coefficiente di inbreeding
FH0 = H0 – H
H = H0 – FH0 , ma H0 = 2pq
H = 2pq - 2pqF = 2pq(1-F)Un coefficiente di inbreeding pari a F porta a un deficit di eterozigoti pari a (1-F): metà AA e metà aa
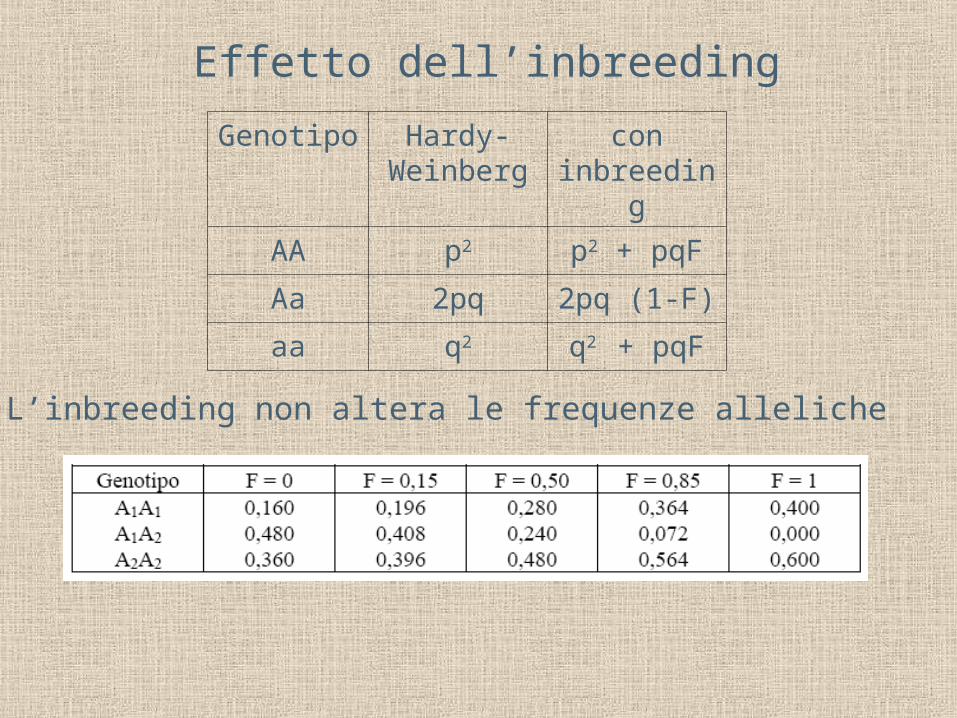
Effetto dell’inbreeding
Genotipo Hardy-Weinberg
con inbreeding
AA p2 p2 + pqF
Aa 2pq 2pq (1-F)
aa q2 q2 + pqF
L’inbreeding non altera le frequenze alleliche

Depressione da inbreeding
Pony delle Shetland

Nessuno è immune dall’inbreeding40 generazioni fa (1000 dC): 1 000 000 000 000 antenatiPopolazione stimata della terra: 100 000 000
80 generazioni fa: 1030 antenatiPopolazione stimata della terra: 10 000 000
1000 generazioni fa: 10300 antenatiPopolazione stimata della terra: 1 000 000 Quindi:
Del milione di individui presenti 25 000 anni fa, molti non hanno lasciato discendenti, molti non sono nostri antenati, altri lo sono miliardi di volte
Le nostre genealogie sono tutte fortemente intrecciate

• L’unione assortativa positiva provoca un deficit di eterozigoti rispetto alle attese di Hardy-Weinberg
• Il deficit di eterozigoti viene misurato dal coefficiente F di inbreeding
• Coefficienti di inbreeding possono essere stimati dalle frequenze genotipiche o dagli alberi genealogici
• L’inbreeding è conseguenza anche del fatto che il numero di antenati di ognuno raddoppia ad ogni generazione, mentre le popolazioni hanno dimensioni finite
Riassunto


















