
FUNCTIONAL ANALYSIS OF STEROLS IN AUXIN SIGNALING · Grebe, 2009, Schaller, 2003). Sterol...
81
Ghent University – Department of Plant Biotechnology and Bioinformatics VIB – Center for Plant Systems Biology Research Group: Root Development FUNCTIONAL ANALYSIS OF STEROLS IN AUXIN SIGNALING Martha Elena Ibarra-Vaele Student number: 01314148 Promoters: Prof. Dr. Tom Beeckman and Dr. Steffen Vanneste Scientific supervisor: Dr.Steffen Vanneste Master’s dissertation submitted to Ghent University to obtain the degree of Master of Science in Biochemistry and Biotechnology. Major Plant Biotechnology Academic year: 2016-2017
Transcript of FUNCTIONAL ANALYSIS OF STEROLS IN AUXIN SIGNALING · Grebe, 2009, Schaller, 2003). Sterol...

Ghent University – Department of Plant Biotechnology and Bioinformatics VIB – Center for Plant Systems Biology
Research Group: Root Development
FUNCTIONAL ANALYSIS OF STEROLS IN AUXIN
SIGNALING
Martha Elena Ibarra-Vaele Student number: 01314148
Promoters: Prof. Dr. Tom Beeckman and Dr. Steffen Vanneste
Scientific supervisor: Dr.Steffen Vanneste
Master’s dissertation submitted to Ghent University to obtain the degree of Master of Science in
Biochemistry and Biotechnology. Major Plant Biotechnology
Academic year: 2016-2017

i
Acknowledgements
I want to express my gratitude to Dr. Steffen Vanneste for his expertise, scope and kind guidance
that made possible the completion of this work. Additionally, I am thankful to Kjell De Vriese for his patience and good disposition concerning laboratory work.

ii
Table of contents
Section Page
Resume 1 1. Introduction 2
1.1 Plant sterols 2 1.1.1 Phytosterol biosynthesis 2 1.1.2 Sterol biosynthetic pathway disruptions 4 1.1.3 Phytosterols and other hormone pathways 5 1.2 Auxin 6
1.2.1 Auxin signaling 6 1.2.2 Auxin distribution 8
1.3 Endocytosis, endosomal recycling, phytosterols and auxin 10 1.3.1 An overview of endocytosis in plants 10
1.3.2 Plant endosomal trafficking 12 1.3.3 Connections between auxin transport, sterol trafficking and endocytic pathways
12
2. Aim of the study 16 3. Results 19 3.1 Screening of potential sterol biosynthesis inhibitors in Arabidopsis 19 3.1.1 Response to voriconazole 21 3.1.2 Effects of other compounds on FvCYP51A overexpressing lines 23 3.1.3 Effects of other compounds on Arabidopsis lines overexpressing sterol biosynthesis pathway genes
24
3.1.4 Effects of other compounds on Arabidopsis mutants in the sterol biosynthesis pathway
26
3.2 Root length response to potential sterol biosynthesis inhibitor compounds 28 3.2.1 Root length response to inhibitors in sterol biosynthesis mutants 29 3.3 Effects of auxin treatment 31 3.3.1 Effect of auxin on the root length of wild type plants and sterol biosynthesis mutants
31
3.3.2 Effect of auxin on the root length and hypocotyl of wild type plants and cvp1 under candidate inhibitor treatment
31
3.4 Calcium supplementation experiments 34 3.4.1 Root length on diluted medium 35
3.4.2 Root length on medium without calcium 36 3.4.2 Root length on diluted medium, medium with no calcium and under sterol
biosynthesis inhibitor treatment
37
3.4.3 Calcium supplementation on root length 39
3.5 cvp1 response to tamoxifen 40 3.6 Effect of candidate inhibitor compounds on auxin transporter expression in
roots.
41

iii
3.6.1 Long term and short term exposure of PIN2-GFP plants to candidate
inhibitors
42
3.6.2 Long term exposure of ABCB19-GFP plants to candidate inhibitors 44 3.6.3 Stigmasterol and β-sitosterol supplementation assay 45 3.6.4 Effects of the candidate inhibitors in PIN endocytic recycling 49 4. Discussion 53 4.1 Identification of novel plant sterol biosynthesis inhibitors 53 4.2 Sterol biosynthesis as a possible target for control over PIN2 homeostasis 55 4.3 Sterol defect-induced PIN degradation possibly reflects activation of ethylene-dependent trafficking
57
4.4 Calcium supplementation experiments 58 5. Materials and methods 60
5.1 Plant material 60 5.2 Media and growing conditions 60
5.3 Solutions of utilized compounds 60 5.4 Root/hypocotyl imaging and measurements 61
5.5 Confocal laser microscopy 61 5.6 Gene amplification and cloning 61
5.7 Statistical analysis 61
6. References 62 7. Addendum 73
7.1 Differential expression of sterol biosynthesis genes after auxin treatment 73 7.2 Fusion of sterol biosynthesis genes to fluorescent tags to generate
overexpressing/reporter lines
74

iv
Table of Abbreviations Term Abbreviation 2,4-Dichlorophenoxyacetic acid 2,4-D 24-Dehydrocholesterol reductase DHCR24 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase HMGR 4-chloroindole-3-acetic acid 4-CI-IAA 4-chloroindole-3-acetic acid PAA
Adaptor protein AP Analysis of variance ANOVA
ATP-binding cassette type B ABCB AUXIN RESPONSE FACTORS ARF
Auxin/INDOLE ACETIC ACID Aux/IAA Auxin-binding protein 1 ABP1
Brassinosteroids BR Brefeldin-A BFA
Carboxy terminal propeptide CPP
Clathrin heavy chain CHC Clathrin light chain CLC
Clathrin-mediated endocytosis CME Columbia Col-0
COTYLEDON VASCULAR PATTERN 1 CVP1 CYCLOARTENOL SYNTHASE 1 CAS1
Cyclopropyl sterol isomerase CPI Deoxyribonucleic acid DNA
Dimethyl sulfoxide DMSO DIMINUTO DIM
DNA binding domain DBD Domain II dII DWARF DWF Dynamin related protein DRP
Endoplasmic reticulum ER
ETHYLENE RESPONSE FACTOR-associated EAR FACKEL Fk
Fragaria vesca CYP51A FvCYP51A gas chromatography-mass spectrometry GC-MS
Green fluorescent protein GFP Guanine nucleotide exchange factor GEF
guanosine triphosphatases GTPases HYDRA1 HYD1 indole-3-acetic acid IAA

v
indole-3-butyric acid IBA
Methyl jasmonate MeJA Multivesicular endosomes MVE Murashige and Skoog MS Naphtalene acetic acid NAA Overexpressing lines OE P-glycoprotein PGP phosphoinositides PIPs PIN-FORMED PIN PINOID PID
Plasma membrane PM Red fluorescent protein RFP
Rho-like GTPases from plants ROPs Ribosomal large subunit 4 Rpl4d
Selective estrogen receptor modulators SERMs Sorting nexin SNX
Standard deviation SD STEROL 1 STE1
STEROL METHYLTRANSFERASE SMT
Sterol-4 α-methyl oxidases SMO TOPLESS TPL
Trans-Golgi network TGN Transmembrane kinase members TMK
TRANSPORT INHIBITOR RESISTANT 1/ AUXIN SIGNALING F-BOX TIR1/AFB Tukey multiple comparisons correction TMC
Vacuolar protein sorting VPS Vacuolar trafficking VT
Yellow fluorescent protein YFP

1
Resume
Sterols are isoprenoid derived molecules that exist in all eukaryotes and that have important
structural and signaling roles. In plants, multiple end-sterol products exist and they are involved
in the synthesis of secondary metabolites and hormones such as brassinosteroids (BR). Sterol
biosynthesis disruption has developmental consequences that are dependent on the step that
has being interrupted. When early steps of the pathway are interrupted, fertility is decreased,
and growth defects cannot be rescued by external BR application. Defects in later steps on the
pathway can be partially rescued by BR and in general show less aberrant phenotypes. All mutants
along the pathway have their own characteristics and show different sterol profiles reflecting
which step was disrupted. Another way to disrupt the synthesis of sterols in an immediate,
controlled and switchable manner is through the usage of inhibitor compounds. Inhibitor
compounds circumvent the pleiotropic effects shown in mutants, are the only way to see
inhibition effects at short term and their use can be easily combined with multiple treatments
and in a variety of plant lines and conditions. In animals and fungi, sterol inhibitors have been
known and used as fungicides. In plants, only two inhibitors with known targets had been
characterized: fenpropimorph and voriconazole. We show here a preliminary characterization of
four compounds: artemether, tamoxifen, oxiconazole, and clotrimazole, as candidate inhibitors
of sterol biosynthesis, based on their effects in hypocotyl elongation (BR response) and root
elongation (structural sterols response dependent on cell division), impairing of NAA-dependent
PIN2 recycling after brefeldin-A application in roots, and overall cellular defects after treatment
in roots. Additionally, we can suggest a possible target for tamoxifen: STEROL
METHYLTRANSFERASE 2 (SMT2) as shown by the hypersensitivity to the smt2 mutant roots to
tamoxifen. This hypersensitivity is not present in hypocotyls, which suggests a BR independent,
structural sterol-related manner of action, which coincides with the placement of SMT2 in the
synthetic pathway. We found also that the response to the candidate inhibitors in the root,
depicted by the PIN2-GFP signal, is first seen as a collection of cellular division defects and cellular
mis-organizations, and after a while, PIN turnover, agreeing with previous reports that show PIN
degradation in sterol mutants. The cellular defects were not rescued by the application of end-
sterols β-sitosterol and stigmasterol. Instead, PIN2 degradation was triggered by exogenous β-
sitosterol supplementation. Together, all these facts add to the known link between auxin
signaling, auxin efflux carrier trafficking and sterols which act harmoniously to promote correct
development. These inhibitor candidates should now be further validated by biochemical studies
such as gas chromatography/ mass spectrometry and genetic expression evaluation to mark a
positive target so that effects of sterol biosynthesis interruption can be discretely assessed based
on the inactivation of these enzymes in plants with consequences in a variety of physiological
processes, such as vesicle trafficking and auxin carrier recycling .

2
Part 1. Introduction
1.1 Plant Sterols
Sterols are essential molecules for all eukaryotes. They have a well acknowledged structural role
as major components of cell membranes that are critical for membrane reinforcement, fluidity
and permeability regulation, and most recently, their capacity to finely attune a variety of metabolic and ontogenetic processes. In the case of animals, cholesterol is a very good example.
Cholesterol is their predominant sterol, and functions as a precursor and a signal in cell division, cell growth, cell death and various developmental processes (Hartmann, 2004).
Sterols are part of the large isoprenoid family, and as they participate as intermediates in the
biosynthesis of many vital compounds. Plants sterol diversity is set apart because animals and
fungi possess each only one major multifunctional end-product of the sterol biosynthesis
pathway: cholesterol and ergosterol, respectively, while plants produce numerous end-product sterols (phytosterols) such as campesterol, sitosterol, stigmasterol and isofucosterol in
proportions and with functions that are dependent on the species (Schaller, 2004). In plants, sterols are also vital for cellular and developmental processes as precursors of brassinosteroids
(BR) and substrates for metabolites such as saponins, cardenolides and glycoalkaloids (Hartmann,
2004).
1.1.1 Phytosterol biosynthesis
Phytosterols are found most abundantly as free sterols but they are also present as conjugated forms. In Arabidopsis, phytosterols are synthesized in the smooth endoplasmic reticulum (ER) and
are then carried to the plasma membrane through the trans-Golgi network (TGN) (Boutte & Grebe, 2009, Schaller, 2003). Sterol intracellular endocytic trafficking is actin dependent (Grebe
et al., 2003).
A diagram of the currently known Arabidopsis sterol biosynthesis pathway is shown in Figure 1. Sterol biosynthesis is complex, comprising of at least 25 steps from isopentenyl diphosphate,
precursor of all isoprenoids, to end product phytosterols. All eukaryotes share the same steps in the mevalonate pathway from isopentenyl diphosphate to (S)-Squalene 2,3 epoxide (Benveniste,
2002). Thereafter, two possible cyclization pathways exist, one that is dependent on CYCLOARTENOL SYNTHASE 1 (CAS1) to produce stigmasterol and campesterol as end products,
and another that goes through lanosterol to produce cholesterol and sitosterol. The latter pathway (through lanosterol) contributes to a small fraction of total sitosterol production (1.5%)
and is inessential to the biosynthesis of membrane sterols, indicating a possible involvement of
its end products, possibly steroids, as secondary metabolites with a role in plant defense mechanisms (Ohyama et al., 2009).

3
Cycloartenol is the substrate of C-24 STEROL METHYLTRANSFERASE 1 (SMT1) , which catalyzes
the addition of a single methyl at C-24. This step in the pathway is especially crucial to maintain a balanced phytosterol production (Diener et al., 2000). The product, 24-methylene cycloartenol,
undergoes the removal of two methyl groups at position 4 and one methyl at position 14. The enzymes involved in this step are part of one of two existent families of sterol-4 α-methyl
oxidases (SMOs), SMO1, with three members identified in Arabidopsis (Darnet & Rahier, 2004). The reaction yields cycloeucalenol, which through the action of a cyclopropyl sterol isomerase
(CPI) is transformed into obtusifoliol when its cyclopropane ring is opened (Benveniste, 2002).
The obtusifoliol 14 α-demethylase CYP51A2 is a cytochrome P-450 enzyme that catalyzes the
removal of the 14 α and 4 methyl groups from obtusifoliol and produces δ-E14-sterol. This enzyme
is an inhibition target for diverse azole compounds with fungicide and herbicide use, which prevent the interaction between oxygen and the heme group iron of the enzyme by nitrogen
binding (Kim et al., 2005, Kushiro et al., 2001, Rozhon et al., 2013). The next step is the NADPH-mediated hydrogenation of the Δ14 double bond of δ-E14-sterol to give fecosterol, and this is
catalyzed by FACKEL, a sterol C-4 reductase that has been found to be inhibited by fenpropimorph
Figure 1. The Sterol Biosynthetic Pathway. Model of the sterol biosynthetic pathway in Arabidopsis and
known rice orthologs. All major components of the phytosterol pathway have been identified and
functionally characterized in Arabidopsis (dark-blue boxes). Activations at the transcriptional and
posttranslational levels are represented by dashed green arrows and green arrows, respectively. Repression
at the posttranslational level is represented by magenta blunt-end lines. Alternative gene/enzyme names in
Arabidopsis: SQE1/XF1, STE/DWF7, DWF1/DIM/CBB1. Taken from (Vriet et al., 2013).

4
in yeast and that is a compound that is also used as a fungicide (Baloch & Mercer, 1987, Schrick
et al., 2000). Fecosterol is then transformed into a Δ5 sterol, 24-methylene lophenol, by the action of the Δ8-Δ7 sterol isomerase HYDRA1, which catalyzes the reduction of the Δ7 double bond
(Souter et al., 2002). The next step is where the balance between stigmasterol (C-24 ethylsterols) and campesterol (C-24 methysterols and downstream brassinosteroids) amounts lies: the
pathway presents a bifurcation.
Following the first branch implies to undergo the extra C-24 transmethylation via redundant factors, COTYLEDON VASCULAR PATTERN 1/STEROL METHYLTRANSFERASE 2 (CVP1/SMT2) and
SMT3 to produce 24-ethylidenelophenol (Carland et al., 2010, Carland et al., 2002). Thereafter, a member of the second family of sterol-4 α-methyl oxidases or methylsterol monooxygenases
(SMO2) catalyzes the sequential double demethylation of 24-ethylidenelophenol yielding δ-7-
avenasterol (Darnet et al., 2001, Darnet & Rahier, 2004, Sonawane et al., 2016). A subsequent desaturation by the Δ7 C-5 sterol desaturase STEROL 1 (STE1) gives rise to 5-dehydroavenasterol
(Choe et al., 1999b, Gachotte et al., 1996). As a next step, 5-dehydrovenasterol is converted into
isofucosterol by DWARF 5 (DWF5), a sterol Δ7 reductase (Choe et al., 2000). Then, the Δ5-sterol
Δ24-reductase DIMINUTO/DWARF 1 (DIM/DWF1) catalyzes the reduction of the C-24 double bond of isofucosterol, transforming it into sitosterol (Choe et al., 1999a). A further C-22 desaturation
by CYP710A1 gives rise to stigmasterol, the end product of the branch and, jointly with sitosterol, the most abundant components of the phytosterol profile in Arabidopsis (Clouse, 2002, Morikawa
et al., 2006).
The second branch takes 24-methylene lophenol directly under the demethylating action of SMO2, generating episterol (Darnet et al., 2001, Darnet & Rahier, 2004, Sonawane et al., 2016).
STE1 desaturates its C-5 to produce 5-dehydrosterol and then DWF5 reduces C-7, yielding 14-methylene-cholesterol (Choe et al., 1999b, Choe et al., 2000). The C-24 double bond of methylene
cholesterol is reduced by the action of DIM1/DWF1 to finally produce campesterol, which
subsequently enters the BR biosynthesis pathway as its progenitor (Choe et al., 1999a, Clouse, 2011).
1.1.2 Sterol biosynthetic pathway disruptions
Sterol biosynthetic mutants have been characterized. Depending on the position of the mutation
in the pathway and the existence of redundant alleles, a variety of developmental alterations can be observed, which point to involvement of sterols in different developmental processes.
When the mutation is located in the late steps of the biosynthetic pathway, downstream the
synthesis of 24-methylene lophenol, the mutants show a BR-deficient like phenotype that can be partially rescued by BRs (fertility and organ size was nothing comparable to the wild type plants
after rescue in contrast to pedicels and hypocotyls) (Schaller, 2003). The mutants, dwf7/ste1, dim/dwf1 and dwf5 share a number of traits like short stems, reduced fertility, prolonged l ife
cycle, curled, dark leaves and a distorted sterol profile, that in the case of dwf7 was characterized

5
by almost a total absence of detectable campesterol (Choe et al., 1999a, Choe et al., 1999b, Choe
et al., 2000). The fact that their sterol compositions are primarily characterized by campesterol depletion, that their phenotypes are partially rescued by BR application and that fertility was not
restored indicates an important role for sterols in plant reproduction that is BR independent (Schaller, 2004).
Mutations in the early steps of the biosynthetic pathway cannot be rescued by BR
supplementation. These include smt1, fackel (fk), hydra1 (hyd1), cpi, cvp1/smt2 (Figure 1 for position). These mutants show phenotypes that indicate sterol involvement in embryo, vein,
shoot and root patterning, embryogenesis, cell expansion, cell division, polarity and proliferation, fertility, gravitropism and hormone signaling (Carland et al., 2010, Carland et al., 2002, Diener et
al., 2000, He et al., 2003, Jang et al., 2000, Men et al., 2008, Nakamoto et al., 2015, Schrick et al.,
2000, Souter et al., 2002, Willemsen et al., 2003, Zhang & Li, 2016). This BR-independent behavior could indicate a role for early synthesized phytosterols as signal molecules, mirroring cholesterol
in mammalians (Vriet et al., 2013).
The other obvious way to disrupt sterol biosynthesis is through chemical inhibitors. The best
known inhibitors are azole compounds. Azole fungicides such as oxiconazole, ketoconazole,
fluconazole, clotrimazole and voriconazole are the most widely used class of antifungal agents for the control of diseases of higher eukaryotes, being the preferred treatment owing to their
relatively low cost and effectiveness against a broad range of fungi (Price et al., 2015) . They have a single-site mode of action: the nitrogen of the azole heterocyclic ring directly bound to the heme
ferric ion of CYP51 as a sixth ligand, with the varying azole compound side chains interacting with the CYP51 structure, preventing it to access its substrate. The affinity between the agent and the
enzyme is highly variable with the species and the drug (Warrilow et al., 2013).
Other kind of inhibitors are the morpholine compounds. Morpholine fungicides such as
dodemorph, tridemorph, aldimorph and fenpropimorph inhibit either FACKEL or HYDRA1 by
binding to the enzymes with an affinity many times higher than their true substrates, blocking their access to the active sites. As a consequence and in common with the mutant, their sterol
composition is aberrant and growth is inhibited. Plants tolerate morpholine compounds better
than fungi do (Mercer, 1991).
1.1.3 Phytosterols and other hormone pathways
There have been revealed interactions between phytosterol biosynthetic pathway components
and other hormone pathways. FACKEL has been shown to be upregulated by hormones such as
BR, auxin, gibberellins, cytokinin and ethylene (He et al., 2003). Sterol mutants hyd1 and hyd2/fk show an enhanced auxin response dependent on ethylene, and their BR defective phenotypes
were partially rescued by ethylene and auxin supplementation (Souter et al., 2002). Mutants smo2-1 and smo2-2 have phenotypes resembling those of auxin defective mutants and display an
aberrant expression and localization of the PIN-FORMED 1 (PIN1) auxin efflux carrier. The double

6
mutant smo2-1smo2-2 is embryonically lethal with a dwarf phenotype, and this can be completely
rescued by endogenous or exogenous auxin supplementation (Zhang & Li, 2016). Additionally, fk/hyd2, cpi and cvp1/smt2 present alterations as well in the polar localization of PIN proteins,
possibly related to membrane structural disruptions that also seem to affect vasculature formation (Carland et al., 2010, Men et al., 2008, Pullen et al., 2010, Willemsen et al., 2003). In
the case of the double mutant smt2 smt3, the incorrect localization of PIN2 is due to disturbances in cell division and a failure to distribute PIN2 asymmetrically after cytokinesis, which leads to
reduction of PIN2 in the plasma membrane, and the associated lateral root proliferation defects
can be rescued by sitosterol supplementation (Nakamoto et al., 2015).
1.2 Auxin
Auxin (from the Greek αὐξάνω, to grow) is a plant hormone involved in regulation and triggering of many of the developmental and growth programs that dynamically change as plants cope with
environmental challenges. They participate in all aspects of plant life, such as cell division and differentiation, organ development, embryogenesis, gravitropism and lateral root emergence
(Salehin et al., 2015, Vanneste & Friml, 2009). Its mechanism of action allows the plant cell to swiftly transition from transcriptional repression of auxin-responsive genes to their activation
(Lavy & Estelle, 2016).
1.2.1 Auxin signaling
The most known auxin response is the result of the genome-wide triggering of transcriptional activation via changes of activity of the AUXIN RESPONSE FACTORS (ARFs) that are dependent on
auxin concentrations (Figure 2). When auxin levels are low, the Auxin/INDOLE ACETIC ACID
(Aux/IAA) transcriptional repressors interact with ARFs, inhibiting their activity and quenching the expression of responsive genes. On the other hand, when auxin is perceived in the nucleus, the
TRANSPORT INHIBITOR RESISTANT 1/ AUXIN SIGNALING F-BOX (TIR1/AFB) auxin receptors stably bind to Aux/IAAs facilitated by auxin, which results in the polyubiquitination and subsequent
proteolysis of Aux/IAA and finally releasing ARF repression (Lavy & Estelle, 2016, Salehin et al., 2015). ARF repressing implies the recruiting of TOPLESS (TPL), a co-repressor, by Aux/IAA-ARFs
complexes to the chromatin (Szemenyei et al., 2008).
Activation of ARFs occurs when the Aux/IAA repressors are degraded, and for this is critical the
activity of an E3 ubiquitin protein ligase called SCFTIR1/AFB. The aforementioned TIR1 F-box protein
is the substrate recognition subunit of the E3. The degradation signal of the Aux/IAAs is located in a conserved domain called domain II (dII). Auxin has a special manner to promote degradation:
instead of causing changes in the substrate as many other E3 ligases require to happen in order to recognize it, auxin boosts the interaction between SCFTIR1/AFB and the dII through direct
attachment to TIR1, which means that TIR1 is an auxin receptor, confirmed by affinity studies.

7
Supporting this, mutants from the TIR1 family display developmental and growth disturbances and increased auxin resistance (Dharmasiri et al., 2005, Kepinski & Leyser, 2005, Ruegger et al.,
1998). It was also discovered that to ensure an efficient binding to auxin, TIR1 must be previously bound to an Aux/IAA as a co-receptor complex. Different combinations of TIR1 and Aux/IAA
proteins show different affinities for auxin, which would signify a fine orchestration and a wide
range of auxin responses determined by available receptors in each cell, potentially explaining how auxin is behind the control of so many developmental events (Calderon Villalobos et al.,
2012, Lee et al., 2014).
There exist, however, a TIR1 independent signaling pathway that responds to extracellular auxin and regulates in a non-transcriptional manner plasma membrane and cytoplasmic responses. The
proposed receptor for auxin in this pathway is auxin-binding protein 1 (ABP1). ABP1, and importantly, its auxin binding pocket region, is essential for many developmental processes such
as root development and cell morphogenesis and expansion, and it seems to coordinate with the TIR1 pathway to regulate gene transcription(Chen et al., 2001, Chen et al., 2014, Chen et al., 2012,
Grones et al., 2015, Tromas et al., 2013). ABP1 interacts with transmembrane kinase members (TMK) of the receptor-like kinase family at the PM, being TMK the docking protein for ABP1, which
is essential for the activation of Rho-like guanosine triphosphatases (GTPases) from plants (ROPs)
upon auxin perception at the PM (Xu et al., 2014). GTPase action leads to regulation of cytoskeletal organization (through their RIC effectors) and clathrin-mediated endocytosis of PINs,
which in turn has an influence on the subcellular distribution of PIN transporters (Chen et al.,
2012, Fu et al., 2005, Fu et al., 2009, Robert et al., 2010).
Figure 3. General mechanism of auxin perception and response. (A) Domain structure of the Aux/IAA and ARF proteins. EAR is the ETHYLENE RESPONSE
FACTOR-associated repression motif that interacts with the TPL co-repressor. The dII domain facil itates interaction with the TIR1/AFB protein in response to auxin. The PB1 domain has both positive and
negative electrostatic interfaces for directional protein interaction. DBD is the B3 DNA binding domain, and MR is the middle region that determines the activity of the ARF.
(B) At low auxin levels, the Aux/IAA proteins form multimers with ARFs and recruit TPL to the chromatin.
(C) High levels of auxin promote ubiquitination and degradation of Aux/IAAs through SCFTIR1/AFB and the proteasome. ARFs are free to activate transcription of target genes. The site of Aux/IAA ubiquitination is
arbitrary. The actual sites are unknown. Auxin is represented by the red oval. Modified from (Salehin et al., 2015).

8
1.2.2 Auxin distribution
At the moment, four native auxins have been identified in plants: indole-3-acetic acid(IAA), indole-3-butyric acid (IBA), 4-chloroindole-3-acetic acid (4-CI-IAA) and phenylacetic acid (PAA) ,
being indole-3-acetic acid (IAA) the most active form in Arabidopsis, where IBA functions mostly as a precursor and a regulator of IAA (Simon & Petrasek, 2011). Asymmetric distribution of auxin
can be observed from punctual accumulation in single cells to auxin gradients in certain tissues,
such as the locations presenting cell division, expansion and differentiation (van Berkel et al., 2013). Auxin biosynthesis and the accompanying release of its precursors are highly localized and
play a key role in shaping auxin gradients after responding to environmental and developmental signals (Zhao, 2010) but the current information suggests that the asymmetric auxin distribution
is a product mainly from intercellular transport (Tanaka et al., 2006). There are two different ways by which auxin can be transported. One is the nonpolar, non-regulated, rapid transport in which
Figure 3. Cellular auxin transport. A general panorama of auxin transport proteins. PIN efflux carriers
are shown in red or pink. ER depicts endoplasmic reticulum, in gray. ER derived endos omes also in gray.
ER marks endoplasmic reticulum, pale gray structures represent ER and endosomes, curved bold full
arrows show constitutive protein cycling, and dashed arrows symbolize the process of transcytosis.
Possible collaboration between ABCBs and PINs is suggested by placing the symbols close to each other.
Taken from (Zazimalova et al., 2010).

9
most IAA from the source tissues as young leaves and flowers reaches the sink tissues by bulk
flow through the phloem, and there exists another system that moves actin in a slower, more controlled manner through long and short distances: active transport (Figure 3) (Petrasek & Friml,
2009).
Active transport relies on the fact that auxins, being weak acids, possess proton-dissociated and associated forms, and as such, their ability to penetrate through a hydrophobic membrane
depends on pH. Plant apoplast pH is 5.5, and at this pH the calculated proportion of IAA molecules in equilibrium is 83% dissociated (anionic) and 17% proton-associated (IAA pKa=4.85). The anionic
IAA forms cannot cross the membrane because the negative charge of the carboxyl group prevents it from interacting with the membrane, whereas the proton-associated IAA molecules
can enter the cell by lipophilic diffusion without the help of a carrier protein.
Once in the cytoplasm, the pH is 7 and the proportion of IAA in equilibrium shifts to an almost entirety of anionic forms, preventing them from escaping across the PM and making the cell a
weak acid anion trap that makes necessary the existence of active efflux transporters to avoid unilateral auxin accumulation. This way, and if the carriers would be located preferentially on one
side of the cells, auxin flow could be unidirectional, or polar. This is what is called the
chemiosmotic hypothesis (Raven, 1975, Vanneste & Friml, 2009, Zazimalova et al., 2010).
The aforementioned auxin efflux carriers have been characterized. Two main protein families
have been found to possess auxin exporting capacities: PIN and ATP-binding cassette (ABC), primarily by the B type (ABCB). PIN proteins are membrane proteins with eight members with
are divided in two categories characterized by the length of a hydrophilic loop in the middle of
two hydrophobic regions consisting each of five transmembrane parts (Krecek et al., 2009). Long PINs (PIN1-4 and 7) show a polar PM localization, and there is strong evidence for direct auxin
transport by PIN1, 2, 4 and 7 (Zazimalova et al., 2007, Zazimalova et al., 2010). PIN proteins
determine the flow direction through their polar localization and are involved in a wide variety
of plant developmental processes (Friml et al., 2002a, Friml et al., 2002b, Sauer et al., 2006, Wisniewska et al., 2006).
The family of plant orthologs of the ATP-binding cassette subfamily B (ABCB or PGP from p-
glycoprotein) includes 21 members in three clusters, but only three of them have been well characterized as auxin transporters: ABCB1, ABCB4 and ABCB19 (Geisler & Murphy, 2006,
Titapiwatanakun & Murphy, 2009). They have a non-polar localization in the membrane. A direct role for ABCB1 and ABC19 in cellular efflux was indicated by the abnormal auxin accumulation in
protoplasts from Arabidopsis mutant abcb1 and abcb19 (Geisler et al., 2005). ABCBs are
important for several developmental processes, such as embryogenesis and gravitropism, and capable to direct auxin traffic in heterologous systems (Cho & Cho, 2013, Petrasek & Friml, 2009,
Petrasek et al., 2006).
There is evidence of interaction and coordinated auxin transported between ABCB19 and PIN1.
They have independent functions but their subcellular co-localization and co-
immunoprecipitation suggest physical interaction. This is also strongly suggested by synergistically abnormal phenotypes of double mutants pin1 abcb19 and an increase auxin efflux

10
and inhibitor sensitivity in HeLa cells that co-express ABCB19 and PIN1. There is no evidence of
direct interaction between PIN1 and ABCB1, but when co-expressed in the heterologous system, there was also an increase of auxin efflux (Bandyopadhyay et al., 2007, Blakeslee et al., 2007).
While auxin can passively enter the PM, it is also brought inside by members of the AUX1/LAX
family , permeases with H+ symport activity. This is particularly useful when pointed cells need a copious and fast auxin influx, e.g. at the lateral root cap, where AUX1 controls and directs polar
auxin fluxes (Zazimalova et al., 2010).
A critical aspect the capacity of plants to adapt their growth to their environment, is the ability
auxin to control its transport. Canonical auxin signaling mediates auxin-responsive transcriptional
changes, that cause changes PIN abundance at the level of biosynthesis (Vieten et al., 2005), degradation (Baster et al., 2013), and polarity (Sauer et al., 2006). On the short term, this signaling
pathway results in increased PIN abundance at the PM (Vieten et al., 2005), and has been associated with PIN repolarization (Sauer et al., 2006).
On the other hand, prolonged auxin signaling stimulates PIN degradation (Abas et al., 2006, Baster
et al., 2013). In addition to transcriptional auxin signaling, PIN abundance is further fine-tuned via an ABP1 mediated, non-transcriptional auxin signaling pathway that controls CME (Chen et al.,
2012, Paciorek et al., 2005, Robert et al., 2010) The molecular mechanisms by which distinct auxin signaling mechanisms are integrated to bring about such contrasting effects on PIN abundance
are currently unknown.
1.3 Endocytosis, endosomal recycling, phytosterols and auxin
1.3.1 An overview of endocytosis in plants
Endocytosis occurs when cells internalize portions of their plasma membrane and extracellul ar substances by forming endocytic vesicles. In plants, it happens at the plasma membrane that
surrounds the cell and at the cell plate membrane as well, where it helps removing the excess membrane derived from the fusion of Golgi vesicles . Endocytic vesicles leave the plasma
membrane, move into the cell and fuse to membrane-bound organelles called endosomes. Early endosomes receive the endocytic cargo from the PM, while late endosomes fuse with vacuoles
and lysosomes to deliver proteins destined for degradation. Endocytosis intersects with exocytosis, the process by which material, recycled or newly synthesized, is delivered to the PM.
Jointly, they modulate the composition and surface area of the PM. Choosing which molecules
are to be internalized by endocytosis is determined by adaptor proteins, intrinsic domains, coating and other associated proteins that decide the sequestering, trafficking and fate of the
internalized cargo. It is also required, though, that the cargo molecules are concentrated in a
patch of the donor membrane. (Kelly & Owen, 2011, McMahon & Boucrot, 2011, Paez Valencia
et al., 2016, Traub & Bonifacino, 2013).

11
In plants and animals, the most prominent endocytic pathways depends on clathrin, a coat
protein. Clathrin-mediated endocytosis (Figure 4) can be divided in five steps: nucleation, cargo selection, clathrin coat assembly, membrane scission and uncoating. Clathrin does not interact
directly with the cargo. The membrane starts shaping pits around it through adaptor (AP) and accessory protein recruitment which interact with membrane lipids and the different sorting
motifs or signals belonging to cargo proteins, and then fetch clathrin from the cytoplasm (Kelly & Owen, 2011, McMahon & Boucrot, 2011).
Cargo selection is dependent on the signals that are present in the cytos olic domains of PM
proteins. There are three signal classes: linear amino acid motifs, conformational motifs and posttranslational modification such as phosphorylation and ubiquitination. A signal can be
present alone or accompanied by other signals in the same cargo protein, and it also can be
recognized by other organelles in the transcytosis pathway, as endosomes and TGN. The presence of internalization motifs will lead to constitutive endocytosis, in the absence of factors that
modulate accessibility of the motif to clathrin adaptors. In contrast, dynamic tagging of cargo by
ubiquitination provides a route to internalization that is regulated at the level of the signal itself
(Kelly & Owen, 2011, Traub & Bonifacino, 2013).
Once the cargo is selected, the clathrin coat starts to assemble. Clathrin triskelia consist of three light chains (CLC) and three heavy chains (CHC) subunits , which are localized in the TGN,
endosomes and PM foci. They are brought from the cytosol by APs and other accessory proteins,
like TPLATE complexes. Clathrin polymerization stabilizes membrane curvature, displacing
undesired proteins to the periphery of the nascent vesicle (Gadeyne et al., 2014, Holstein, 2002,
Ito et al., 2012). Thereafter, dynamin, a large GTPase, assembles into multimeric helices that wrap around the closing gap of the nascent clathrin-coated vesicle. The dynamin GTPase
Figure 4. Endocytic and endosomal trafficking in plants. Cargo (green) with internalizing
signal (ubiquitination, Ub) is is surrounded by a clathrin-coated pit, encased in a clathrin coated vesicle after membrane scission
and transported to the TGN. From it, if It possesses a recycling signal, it is re-trafficked to the PM, mediated by GNOM. If it possesses
a degradation signal, it is recognized by ESCRTs and transported to multivesicular
endosomes (MVE) and then to the vacuole to be destroyed. Retromer-dependent recycling redirects cargo from the MVE to
the TGN to be re-located to the PM. Modified from (Paez Valencia et al., 2016).

12
hydrolysis drives membrane fission. There exist six types of Dynamin Related Proteins (DRPs) in
plants, with DRP1 and DRP2 involvement in clathrin-mediated trafficking, even with a yet poorly understood recruiting mechanism to clathrin-coated pits (Gadeyne et al., 2014, McMahon &
Boucrot, 2011, Paez Valencia et al., 2016). The vesicle then detaches from the PM, is stripped from its clathrin coat and can freely merge with endosomes. The uncoating in Arabidopsis was
shown to be facilitated by an auxilin-like protein and a protein called SH3, the latter colocalizing with clathrin (Lam et al., 2001).
1.3.2 Plant endosomal trafficking
Recycling of endosomal cargo proteins in plants is mediated by the TGN and MVEs, contrasting with animals, which possess dedicated recycling tabulated endosomes (Figure 4). ARF-guanine
nucleotide exchange factor (GEF) GNOM is the best known regulator of recycling and a proposed marker for recycling endosomes. ARF-GEFs recruit coat proteins and cargos at the membrane and
promote vesicle formation through activation of ARF GTPase by exchanging bound GDP for GTP. It localizes to the Golgi apparatus and is important for the maintenance of the functional integrity
of the TGN, supported by the TGN abnormalities displayed by gnom mutants. It regulates
trafficking from the ER to the TGN but a fraction of GNOM also controls endocytosis at the PM (Geldner et al., 2003, Naramoto et al., 2010, Naramoto et al., 2014).
The retromer complex orchestrates an alternative recycling pathway. In mammals, it is composed of two subcomplexes: a core constituted by Vacuolar Protein Sorting proteins (VPSs) and a dimer
of Sorting nexin (SNX). In Arabidopsis, all members of the retromer are conserved, but its
retromer complex has the particularity of having a dispensable SNX dimer but indispensable VPSs to maintain endosome homeostasis and organogenesis. The SNX dimer is the part of the complex
that directly interacts with the phosphoinositides (PIPs) of the membrane. The core (specifically VPS35) binds to the cytosolic tail of cargo proteins. VPS26 and VPS29 are important to membrane
recruitment (Zelazny et al., 2013a, Zelazny et al., 2013b). The putative mechanism states that the retromer complex coat endosomal membranes enriched in cargo proteins and transport them
from the MVEs and away from vacuolar degradation back to the TGN. There exist another end to
cargo: degradative sorting. The primary signal for endosomal-mediated degradation is ubiquitin. Cargo that is not recycled and remains ubiquitinated is recognized and directed to MVEs by the
ESCRT proteins for its degradation. ESCRT complexes mediate membrane invagination instead of budding, sequestering cargo into intraluminal vesicles to subsequently be degraded when the
MVE containing it matures and fuses to the vacuole. This is a pH dependent process, where the late endosome must be alkalinized in order to be competent for fusion(Paez Valencia et al., 2016).
1.3.3 Connections between auxin transport, sterol trafficking and endocytic pathways

13
PIN proteins are not permanently docked to the PM. They are constantly being trafficked and
recycled between the PM and ER derived endosomes, and this allows them to be swiftly relocated to other parts of the cell through a transcytosis-like mechanism (Figure 5) (Dhonukshe et al., 2007,
Geldner et al., 2001, Kleine-Vehn & Friml, 2008). PIN endocytosis is clathrin-dependent, as evidenced by the endocytic recycling defects and abnormal polar distribution of PIN transporters
that arise from CME interference in chc mutants and expression of a dominant negative CHC fragment (Kitakura et al., 2011). Auxin inhibits PIN CME, regulating its own flow by controlling the
abundance of PIN transporters at the plasma membrane through the auxin receptor ABP1 and
downstream effectors ROP6 and RIC1 (Chen et al., 2012, Paciorek et al., 2005, Robert et al., 2010). However, recently the functionality of ABP1 in auxin signaling processes has been questioned
casting serious doubt on our current understanding of the pathway underlying auxin-regulated endocytosis (Gao et al., 2015).
There is evidence that sterol levels are critical components in the establishment of PIN
polarization. While phytosterol biosynthesis occurs at the ER as indicated by biochemical
fractionation studies and by subcellular localization studies of biosynthetic enzymes (Boutte &
Grebe, 2009, Men et al., 2008), they do not accumulate at the ER. They are transported via the TGN to the PM where they accumulate (Grebe et al., 2003, Moreau et al., 1998). smo2-1 and
smo2-2 have phenotypes resembling those of auxin defective mutants and display an aberrant expression and localization of PIN1. The double mutant smo2-1smo2-2 is embryonically lethal
with a dwarf phenotype, and this can be completely rescued by endogenous or exogenous auxin
supplementation (Zhang & Li, 2016). fk/hyd2, cpi and cvp1/smt2 show an incorrect polar localization of PIN proteins. The vascular pattern defects in cvp1 and smt3 mutants could be
associated to alterations in the membrane when veins are formed (Carland et al., 2010, Men et al., 2008, Pullen et al., 2010, Willemsen et al., 2003).
Figure 5. CME, sterols and auxin transport. The PIN2
auxin efflux carrier (in green) localizes to the apical
membrane. PIN2 is internalized by CME and recycled
at the PM or sent to degradation. Altered sterol
composition in sterol biosynthesis mutants (cpi1 for
reference) or under the action of biosynthesis
inhibitors (fenpropimorph for reference) interferes
with PIN2 endocytosis and auxin inhibition of PIN2
endocytosis requires sterol function. Modified from
(Boutte & Grebe, 2009)

14
Sterol endocytic trafficking involves a Brefeldin-A (BFA) sensitive, actin dependent pathway
(Grebe et al., 2003). BFA targets GNOM, that mediates the endosomal recycling to the PM (Kleine-Vehn & Friml, 2008). Interestingly, PIN exocytic trafficking is also BFA-sensitive but auxin-
stimulated (Paciorek et al., 2005), and PIN1 and PIN3 polarization requires correct sterol biosynthesis, as shown by defects in cell polarity and auxin distribution in smt1 mutants root cells
(Willemsen et al., 2003). In plants that harbor a fully functional, BFA-resistant version of GNOM, PIN1 localization and auxin transport are no longer affected by BFA (Geldner et al., 2003). Partial
loss of function gnom mutants show developmental defects related to auxin and strongly
diminished PIN1 internalization (Geldner et al., 2004, Naramoto et al., 2010).
Similarly, a biosynthesis mutant, cpi1, fails to re-establish PIN2 polarity after cytokinesis. This was
associated with a defect in PIN2 internalization in the cpi1 mutant, which has a sterol content of
almost 99% cyclopropylsterols (Boutte & Grebe, 2009, Men et al., 2008). Another sterol biosynthesis double mutant, smt2 smt3, shows PIN2 mis-localization due to disrupted cell division
and failure to selectively remove PIN2 after cytokinesis. Although endocytosis of PIN2 from the
plasma membrane (PM) is apparently unaffected in smt2 smt3, strong inhibition of the endocytic
recycling is associated with a remarkable reduction in the level of PIN2 on the PM (Nakamoto et al., 2015).
The fate of auxin carriers can also be determined by the retromer complex, which mediates the
recycling of biosynthetic cargo between the TGN, PM and the vacuole and is BFA-insensitive. In Arabidopsis, one of the components of the retromer complex, VPS29, is required for PIN1
repolarization after containment in pre-vacuolar compartments (PVC) and endosomal homeostasis (Jaillais et al., 2007), and another one, SORTING NEXIN 1 (SNX1), colocalizes with
VPS29, defines the identity of PVCs and targets PIN2 for vacuolar degradation, but not PIN1 (Kleine-Vehn & Friml, 2008, Kleine-Vehn et al., 2008, Paez Valencia et al., 2016). C-24 ethylsterol
defective smt2 smt3 presents mis-localizations of the retromer associated protein CLASP, which
interacts with SNX1, possibly explaining the enhanced PIN degradation in this mutant as CLASP is involved in PIN2 recycling (Ambrose et al., 2013, Nakamoto et al., 2015).
Notably, the translational Arabidopsis mutant ribosomal large subunit 4 (rpl4d) which is defective
in vacuolar trafficking (VT) and auxin signaling, has reduced levels of lipid metabolism intermediates, and its phenotype can be mimicked in wild type plants when grown in the
presence of sterol biosynthesis inhibitors. Its vacuolar trafficking impairment could also be
rescued partially by complementation with lipid biosynthesis enzymes, such as SMT2 (Li et al.,
2015). An auxin signaling mutant, axr1-12 also displays significantly reduced sterol levels, while having increased PIN degradation (Pan et al., 2009). This suggests that sterol levels are not only
involved in PIN recycling but also in PIN turnover via VT in an auxin-respondent manner.
The location and behavior of ABCB auxin transporters are also related to sterol content. ABCB19
has been shown to stabilize PIN1 at the PM, has been isolated in sterol- and sphingolipid-enriched
detergent resistant membranes, and is required for PIN1 retention in those membranes. Further,
ABCB19 auxin transport activity in heterologous systems is enhanced by structural sterol

15
enrichment similar to what is observed in animal cells (Blakeslee et al., 2007, Titapiwatanakun &
Murphy, 2009).
ABCB19 trafficking between the TGN and PM is also regulated by membrane sterol content. abcb19 plants are partially resistant to fenpropimorph, and previous studies show that
endocytosis of ABCB19 is not affected by loss of sterols caused by fenpropimorph, suggested by the co-localization of ABCB19-GFP with the endocytic marker FM4-64 at the TGN after FEN
treatment and no changes of PM ABCB19 signals upon FEN treatment (Yang et al., 2013).
Calcium (Ca+2) is a second messenger that is transient, local and triggered by a multitude of stimuli, making its responses difficult to delimit. It is known that Ca+2 stimulates exocytosis in
animal cells e.g. during neurotransmission. In plants, there are indications for a possible role of Ca+2 as a coordinator of trafficking during tip growth in pollen and root hairs, simultaneously
stimulating exocytosis while inhibiting endocytosis in the growing tip where Ca2+ levels are highest. In the subapical zone of these cells, CME is coinciding with a low Ca+2 concentration
area. As a result, cell polarity is established by the specific disposition and removal of membrane
proteins (Himschoot et al., 2015). Several components of the CME machinery are calcium sensitive and are localized subapically in pollen tubes. Caffeine, an intracellular Ca+2 leak inductor,
enables the dislodgement of the TPC subunit, T-PLATE and CLC3 from the cell plate during cytokinesis, suggesting calcium-mediated inactivation of CME (Van Damme et al., 2011).
Indirect interaction of Ca2+ and CME can be attributed to phosphatidylinositides (PIPs), PIPs are
lipid molecules that mediate protein to vesicle recruitment and are regulated by phosphorylation. pip5k2, a mutant in a PIP kinase, has decelerated vesicular trafficking and altered response to BFA
and the pip5k1pip5k2 double mutant shows decelerated endocytic recycling of both PIN1 and PIN2. PIP5K1 and PIP5K2 are essential for cargo localization, such as PINs. This causes a
disturbance of auxin gradients and as a consequence, incorrect embryonic and postembryonic patterning. Auxin regulates itself PIP5K transcription. PI(4,5) biphosphate is required for CME and
PIN polarity in pollen tubes as well as root cells, and, at least in pollen tubes, Ca2+ signals control
PI hydrolyzing enzymes localization (Ischebeck et al., 2013, Tejos et al., 2014).
Intracellular PIN trafficking is also influenced by PINOID (PID) phosphorylation. PID-mediated
phosphorylation in the root yields PINs that are insensitive to GNOM driven cargo traffic, which leads to PIN3 polarity towards the shoot. After photostimulation and subsequent PID inactivation,
GNOM can recruit PIN3 and shuttle it towards the inner and lateral faces of the cell. PID activity
can be regulated by interaction with different Ca2+ binding proteins, suggesting that Ca2+ signals could control PIN polarization via effects on PID activity (Vanneste & Friml, 2013). This would not
be the first reported intertwining of Ca+2 and auxin signaling pathways. It is already known that in Arabidopsis roots, gravistimulation results in auxin flux redistribution in the cap and subsequent
auxin accumulation on the lower face of the root. Interestingly, areas that display sharp Ca 2+ level peaks after gravistimulation coincided with auxin redistribution areas and with pH increases,
possibly regulating the root growth tropic response by cell wall alkalinization (Monshausen et al.,
2011).

16
Sterol biosynthesis has also links to calcium signaling. Arabidopsis smt1 mutant has been shown
to be hypersensitive to calcium ions, and it has been proposed that this behavior is derived from an altered membrane permeability attributed to sterol balance deficiencies and accompanying
aberrant calcium transmembrane traffic (Diener et al., 2000). DWF1 is a Ca2+/calmodulin-binding protein and this binding is critical for its function, campesterol synthesis (on the 24-methylsterol
branch of the pathway). Analysis of site-directed and deletion mutants revealed that loss of calmodulin binding completely abolished the function of DWF1 in planta, and loss of calmodulin
binding resulted in dwarf phenotypes, indicating calcium/calmodulin signaling need for DWF1 to
function correctly (Du & Poovaiah, 2005).

17
Part 2 . Aim of the study
Besides their known structural function as component of the plasma membranes, sterols are also
known as signaling molecules in eukaryotes. Plants possess a variety of end-sterols like campesterol, β-sitosterol and stigmasterol, with known important functions such as
brassinosteroid precursors and metabolite substrates (Schaller, 2004). Their biosynthesis pathway is a complex sequence or reactions that is shared by all eukaryotes until the production
of squalene 2-3 epoxide, where by means of CAS1 oxide squalene cyclase plants take an
alternative way diverging from lanosterol and ultimately cholesterol or ergosterol, and instead going further into cycloartenol, which depending on the chosen downstream pathway, is
ultimately converted into 24-methylsterols (BR precursors such as campesterol) or 24-ethylsterols such as β-sitosterol or stigmasterol (Vriet et al., 2013).
Sterol biosynthesis deficiency has been characterized in mutants affected in different steps of the
pathway. When the mutation is situated before the bifurcation of 24-ethyl and 24-methylsterols, it can be rescued by BRs, but not if it is situated earlier in the pathway. As expected, all sterol
biosynthesis mutants have abnormal sterol profiles. They show developmental involvement of sterols indicated by low fertility, vein, shoot and root patterning defects, cell division and
proliferation abnormalities and deficiencies in gravitropism and hormone signaling (Carland et al., 2010, Carland et al., 2002, Diener et al., 2000, He et al., 2003, Jang et al., 2000, Men et al., 2008,
Nakamoto et al., 2015, Schrick et al., 2000, Souter et al., 2002, Willemsen et al., 2003, Zhang & Li,
2016). When wild-type plants have their sterol biosynthesis pathway disturbed through sterol biosynthesis inhibitors like voriconazole and fenpropimorph, they show similar development
impairments such as dwarf stature with stunted shoots and roots product of cell elongation inhibition (He et al., 2003, Rozhon et al., 2013).
These phenotypes have been related to interactions between the phytosterol biosynthetic
pathway and hormone signaling pathways such as auxin signaling. Auxin supplementation was
able to partially rescue the BR deficient phenotype of hyd1 and fk mutants, which cannot be
rescued by BR supplementation itself (Souter et al., 2002). The embryonic lethality and dwarfism of mutant smo2-1smo2-2 can be completely rescued by endogenous or exogenous auxin
supplementation (Zhang & Li, 2016). In sterol mutants, there is often an non-polar PIN
localization in the membrane. PIN transporters and sterols are both intracellularly trafficked by
an endocytic pathway that is BFA-sensitive. Auxin exocytic pathway is stimulated by auxin even
in the presence of BFA, and lack of sterols either by mutation or by the usage of sterol inhibitors
promote PIN internalization even in the presence of exogenous auxin (Pan et al., 2009).
In summary, sterols are intimately connected to the precise manner in which auxin flux is directed
by its carriers, and that has ignited our interest in the dissection of the role of sterol biosynthesis
intermediates in auxin signaling, PIN polarity and trafficking. Sterol biosynthesis intermediates
are plenty, and while mutants affected in a variety of steps in the pathway are well characterized,
only sterol biosynthesis inhibitors that cover two steps along the pathway had been described.

18
Sterol biosynthesis inhibitors possess the flexibility of exogenous agents that can be applied in
various concentrations to any plant whichever its genotype in an immediate manner and can be
withdrawn and combined at any moment with other agents. A wider catalog of inhibitors would
be useful to be able to toggle the activity of determined enzymes if the targets are known, observing the consequences of the target activities under countless treatments and conditions.
The first aim of this study is the gathering of a number of compounds that could present sterol
biosynthesis activity based on current reports and chemical screenings. Only two sterol inhibitors
under the lanosterol synthesis step, fenpropimorph and voriconazole, that had been reported in
plants. Based on their characterization, we will test our candidate inhibitors looking for similar
activity in Arabidopsis, such as BR deficiency phenotypes and cell division and morphology
defects. Looking for possible redundancy on targets or interaction of targets, the activity of
candidate inhibitors will be tested on sterol biosynthesis mutants, expecting a change in the
behavior of the inhibitor potentiated by an abolishment of the activity of certain enzyme if their
interact in some way to provide certain combination of sterol intermediates necessary for the
preservation of normal phenotypes or if the targets are the same. The activity of candidate
inhibitors will also be tested on Arabidopsis lines that overexpress certain enzymes in the sterol
biosynthesis pathway, looking for changes in the activity of the inhibitor, possibly due to a
restoring of the balance of sterols that the overexpressed enzyme provides to buffer the
disrupting of the pathway caused by the inhibitor. This could be also an indicator of the target of
the enzyme. Additionally, we will clone genes which lines are not available in order to generate overexpressing lines along the whole pathway.
We will compare the response of the mutants to the inhibitors with and without auxin
supplementation in search for potential auxin signaling activating responses that could be
abolished in either the mutant and/or the plant suffering from sterol inhibition by one of our
compounds and that could be rescued by the presence of exogenous auxin. We will test also the
localization of auxin carriers in the membrane of inhibitor-treated plants and the consequences
of the treatment in their exocytic trafficking upon auxin supplementation, to see if all our
candidates respond in the same manner, and if it corresponds to the canonical fk/fenpropimorph exocytosis inhibiting, auxin resistant manner.
Once having evaluated the effects that our candidate inhibitor have in plants, we will try to
counteract their activity by the application of exogenous phytosterols expecting to see rescue of the potential growth deficiencies caused by the compounds.
All this information will be the beginning of a thorough characterization of sterol biosynthesis
inhibitors focused in their effects on growth and cell structure and organization, the influence
that they would have in auxin-derived exocytosis stimulation, their response to exogenous sterol
and auxin supplementation and their behavior of altered sterol-related gene expressing plants
when confronted to the compounds.

19
Part 3. Results
3.1 Screening of potential sterol biosynthesis inhibitors in Arabidopsis
Sterols are critical to assure the formation and maintenance of structurally correct cells with adequate intracellular vesicle trafficking in eukaryotes . In plants their biosynthesis notably
overlaps with that plant growth signals (brassinosteroids (BR)), adding to the importance of the pathway. Sterol biosynthesis disruption has direct and severe consequences in mammals, as
cholesterol biosynthesis deficiencies such as Smith-Lemli-Opitz syndrome (A mutation in the
HMG-CoA reductase (3-hydroxy-3-methyl-glutaryl-coenzyme A reductase, HMGR) are associated with major developmental malformations that are unusual for metabolic disorders (Herman,
2003). It is of course known that sterol biosynthesis inhibitors like azoles and morpholine compounds affect fungi greatly enough to be used as fungicides. In plants, only fenpropimorph
and more recently, voriconazole, have been characterized concerning their inhibitory effects over plant sterol biosynthesis (He et al., 2003, Rozhon et al., 2013).
We aimed to expand the catalog of compounds that disrupt the sterol biosynthetic pathway in
plants for further characterization the functional relevance of sterols, ideally identifying compounds targeting different steps along the way. We had two sources to help us with the
selection of the candidate compounds. First, a recent report described the screening of a small molecule library by assessing the effect of each molecule on the sterol profile of murine neuronal
ells. Different sterol profiles (each with more or less abundance of one or other cholesterol
synthesis intermediary) would signify a disruption of a certain step on the synthetic pathway. The
compounds that showed disrupted activity were validated by observing their effect on cholesterol
abundance in murine fibroblasts (Korade et al., 2016). Second, in our laboratory, a previous massive chemical screening was performed by Kjell De Vriese. In the screening, auxin-induced
calcium signaling responses were recorded in tobacco BY-2 cell cultures after the supplementation with the compounds. Some of the compounds shown by this latter screening
overlapped with the recent report on mouse cells. We decided on selecting clotrimazole, oxiconazole and artemether from our local screening and tamoxifen and fluphenazine from the
screening in murine cells. We used voriconazole and fenpropimorph as positive controls in our
experiments as they had been already described as sterol biosynthesis inhibitors in plants (Figure 6).
This candidate list, as mentioned, includes fungicides that have known targets in fungi, such as
fenpropimorph that targets the fungal delta 14 reductase (Kerkenaar, 1990) and clotrimazole and oxiconazole, azole compounds that inhibit the fungal lanosterol 14 α-demethylase, CYP51A1
(Becher & Wirsel, 2012, Lepesheva & Waterman, 2007). They appeared as hits in the BY-2 cell
calcium screening and in the mouse cell-inhibitor screening. On the other hand, tamoxifen is a
synthetic antagonistic ligand for estrogen receptor α that belongs to the selective estrogen receptor modulators (SERMs) compounds and that is used to treat certain types of breast cancer

20
(Shiau et al., 1998). Estrogen is also one of the multiple products of sterol biosynthesis in
mammals, which is notably also dependent on cytochrome P450 family enzymes (Grogan et al., 1993). Tamoxifen was shown to potentially disrupt either the Δ8-7 isomerase or the 24-
Dehydrocholesterol Reductase (DHCR24)- mediated steps in sterol biosynthesis (Korade et al.,
2016), putatively corresponding to the HYDRA or DWF1 enzymatic activities in Arabidopsis.
Fluphenazine is an antipsychotic that was shown to inhibit calcium responses in the previous
chemical screening (De Vriese, K. and Vanneste, S.; unpublished results) and appears also as a potential inhibitor of sterol biosynthesis in mouse cells (Korade et al., 2016). It is metabolized in
humans by members of the cytochrome P450 family (Attia et al., 2012). It was shown to
potentially disrupt the the Δ8-7 isomerase or the 24-Dehydrocholesterol Reductase (DHCR24)- mediated steps in sterol biosynthesis (Korade et al., 2016). Finally, artemether is an artemisinin,
a lactone compound that is used as an antimalaria agent with an incompletely known mechanism of action that involves reactive oxygen species (ROS) generation in order to provoke membrane
damage in the parasite (Haynes et al., 2012). The last two compounds have no known direct link to any sterol biosynthetic pathways, but artemether was discovered to potentially disrupt the
step mediated by HMGR in mouse cells (Korade et al., 2016).
To test our candidates, we relied on the fact that a potent effect on Arabidopsis sterol biosynthesis that could be evidenced by a strong inhibition of hypocotyl elongation while grown
in the dark, directly associated to BR deprivation (Asami et al., 2000, Rozhon et al., 2013). It is unfortunately not specific enough to point to a determined step to be disrupted but it is a good
indicator of sterol /BR content disruption, as they share many enzymes.
Wild type Columbia ecotype (Col-0) Arabidopsis seedlings were sown on 0.5X Murashige and
Skoog (0.5 MS) agar medium (Murashige & Skoog, 1962), were vernalized for three days followed
by four hours of light to trigger germination, and subsequently grown under darkness for eight days in the presence of different concentrations of each of the compound, ranging between 0.1
Figure 6. Chemical structures of the chosen potential inhibitors. A) artemether, B) oxiconazole, C) tamoxifen, D) voriconazole, E) clotrimazole, F) fenpropimorph, G) fluphenazine. Taken from (Kim et al., 2016).
A B C D
E G F

21
and 10µM, and as a control, 0.1% dimethyl sulfoxide (DMSO). The elongated hypocotyls were
measured after 8 days of darkness. The effect of the compounds can be seen in Figure 7. We
observed evident reductions in hypocotyl elongation in all treatments but not for fluphenazine. The strongest effect was observed on the seedlings grown with artemether, oxiconazole and
clotrimazole, with concentrations as low as 0.5µM. We decided to keep working with oxiconazole, clotrimazole, artemether and tamoxifen. This experiment also had the aim to indicate us which
concentrations of each compound were sufficient to carry out subsequent assays with evident effects.
3.1.1 Response to voriconazole
Azole fungicides such as voriconazole inhibit the activity of CYP51A, a cytochrome 450 enzyme
that catalyzes the synthesis of ergosterol. We verified the previously reported hypocotyl length
reduction related to sterol biosynthesis inhibition caused by voriconazole presence in Arabidopsis (Rozhon et al., 2013) with the object of having a compound with known effects and to assure that
Figure 7. Hypocotyl growth in response to potential inhibitors. Sterol biosynthesis inhibiting
compounds in different concentrations were supplemented to 0.5MS medium in which Col -0 seedlings
were sown. Their hypocotyl length was measured after eight days of growth under darkness. Values
are plotted as Mean ± SD.
0.000
0.500
1.000
1.500
2.000
2.500
0.00 0.10 0.50 1.00 5.00 10.00
Hyp
oco
tyl
len
gth
(cm
)
Compound concentration in medium (µM)
Hypocotyl length in response to sterol inhibitors
Fenpropimorph
Clotrimazole
Artemether
Fluphenazine
Oxiconazole
Tamoxifen
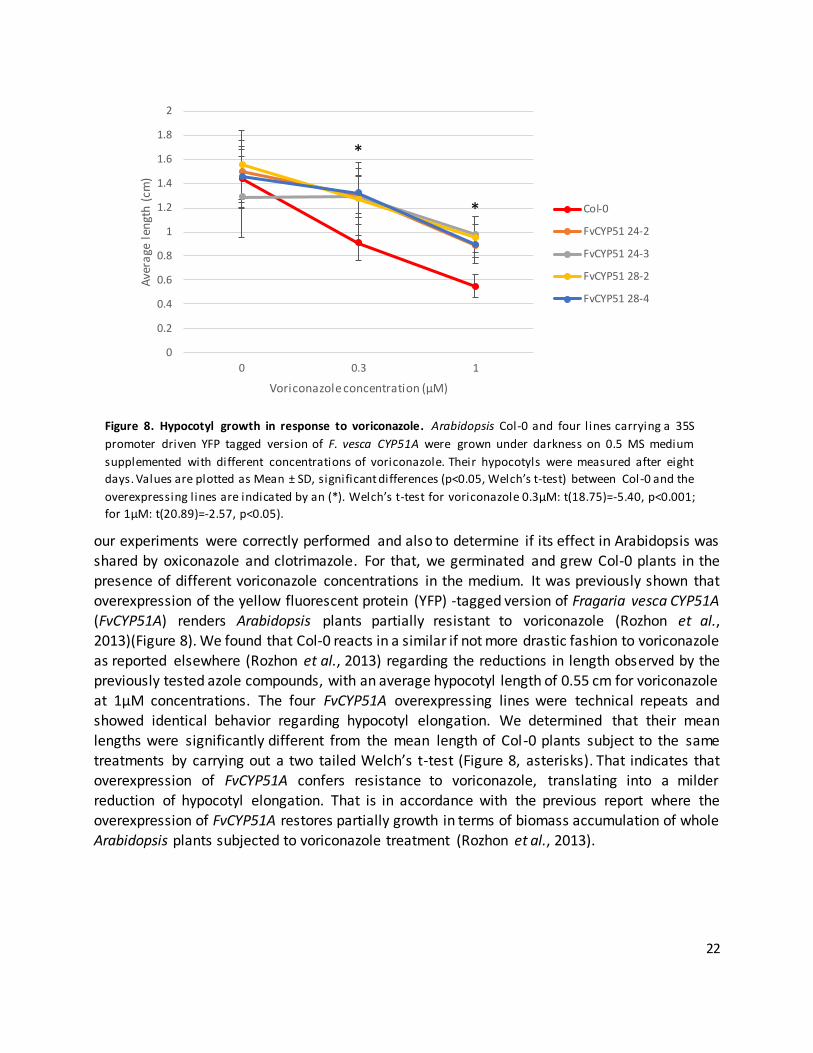
22
our experiments were correctly performed and also to determine if its effect in Arabidopsis was
shared by oxiconazole and clotrimazole. For that, we germinated and grew Col-0 plants in the
presence of different voriconazole concentrations in the medium. It was previously shown that overexpression of the yellow fluorescent protein (YFP) -tagged version of Fragaria vesca CYP51A
(FvCYP51A) renders Arabidopsis plants partially resistant to voriconazole (Rozhon et al., 2013)(Figure 8). We found that Col-0 reacts in a similar if not more drastic fashion to voriconazole
as reported elsewhere (Rozhon et al., 2013) regarding the reductions in length observed by the previously tested azole compounds, with an average hypocotyl length of 0.55 cm for voriconazole
at 1µM concentrations. The four FvCYP51A overexpressing lines were technical repeats and showed identical behavior regarding hypocotyl elongation. We determined that their mean
lengths were significantly different from the mean length of Col-0 plants subject to the same
treatments by carrying out a two tailed Welch’s t-test (Figure 8, asterisks). That indicates that overexpression of FvCYP51A confers resistance to voriconazole, translating into a milder
reduction of hypocotyl elongation. That is in accordance with the previous report where the overexpression of FvCYP51A restores partially growth in terms of biomass accumulation of whole
Arabidopsis plants subjected to voriconazole treatment (Rozhon et al., 2013).
Figure 8. Hypocotyl growth in response to voriconazole. Arabidopsis Col-0 and four l ines carrying a 35S
promoter driven YFP tagged version of F. vesca CYP51A were grown under darkness on 0.5 MS medium
supplemented with different concentrations of voriconazole. Their hypocotyls were measured after eight
days. Values are plotted as Mean ± SD, significant differences (p<0.05, Welch’s t-test) between Col-0 and the
overexpressing l ines are indicated by an (*). Welch’s t-test for voriconazole 0.3µM: t(18.75)=-5.40, p<0.001;
for 1µM: t(20.89)=-2.57, p<0.05).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 0.3 1
Ave
rage
len
gth
(cm
)
Voriconazole concentration (µM)
Col-0
FvCYP51 24-2
FvCYP51 24-3
FvCYP51 28-2
FvCYP51 28-4
*
*

23
3.1.2 Effects of other compounds on FvCYP51A overexpressing lines
In order to determine if the overexpression of either FvCYP51A gave any resistance to hypocotyl
stunted elongation caused by our other compounds, we grew Arabidopsis seedlings, either Col-0
or overexpressing line FvCYP51A 28-2 in medium supplemented with 10µM tamoxifen, 5µM clotrimazole, 1µM artemether or 0.1% DMSO as control. After eight days, their hypocotyls were
measured (Figure 9). We found that the overexpression of FvCYP51A yielded significantly longer hypocotyls than those of the Col-0 plants for any of the treatments, this being verified after
carrying out a two tailed Welch’s t-test ,suggesting partial resistance to these compounds due to the overexpressed CYP51A. It is interesting to mention that from these three compounds ,
clotrimazole is the only one that shares a known common target with voriconazole in the sterol
biosynthetic pathway.
Figure 9. Hypocotyl growth in CYP51A overexpressing lines in response to potential inhibitors.
Arabidopsis Col-0 l ines carrying a 35S promoter driven YFP tagged version of FvCYP51A (FvCYP51A28-
2 OE) were grown under darkness on 0.5 MS medium supplemented wi th different compounds (x-
axis). Their hypocotyls were measured after eight days. Values are plotted as Mean ± SD, significant
differences (p<0.05 and p <0.01) between Col -0 and the overexpressing l ines are indicated by (*) or
(**), respectively. Welch’s t-test: for tamoxifen 10µM: t(26.96)=21.79, p<0.0001; for artemether 1µM:
t(10.42)=-2.90, p<0.05; for clotrimazole 5µM: t(13.51)=-3.62, p<0.01.
0
0.5
1
1.5
2
2.5
DMSO 0.1% Artemether 1µM Clotrimazole 5µM Tamoxifen 10µM
Hyp
oco
tyl
len
gth
(cm
)
Compound and concentration in medium
Col-0
FvCYP51A OE
**
**
*

24
3.1.3 Effects of other compounds on Arabidopsis lines overexpressing sterol biosynthesis pathway
genes
We obtained three Arabidopsis lines that overexpress other YFP-tagged sterol biosynthetic
enzymes : STE1, DWF1/DIM1 and DWF5. Their gene products act downstream of CYP51A in the
pathway and are involved in the synthesis of both campesterol and sitosterol (Figure 1) (Vriet et
al., 2013). We decided to test if the overexpressed gene products conferred resistance to some
of the compounds we selected in a similar manner to the one given by the overexpression of
FvCYP51A. First, we grew Arabidopsis seedlings, either Col-0 or overexpressing lines for
DIM1/DWF1 and DWF5 in medium supplemented with 10µM tamoxifen, 5µM clotrimazole, 1µM
artemether or 0.1%DMSO as control. After eight days, their hypocotyls were measured (Figure
10). Both lines showed significantly longer hypocotyls than Col-0 when grown under artemether
1µM. There was no significant difference between the mean lengths of the overexpressing lines and Col-0 in response to tamoxifen 10µM treatment.
0
0.5
1
1.5
2
2.5
3
DMSO 0.1% Artemether1µM
Clotrimazole5µM
Tamoxifen10µM
Hyp
oco
tyl
len
gth
(cm
)
Compound and concentration in medium
Col-0
DIM1/DWF1 OE
DWF5 OE
*** ***
* *
Figure 10. Hypocotyl growth in DIM1/DWF1 and DWF5 overexpressing lines in response to potential
inhibitors. Arabidopsis Col-0 l ines carrying a 35S promoter driven YFP tagged version of either DIM1/DWF1
or DWF5 were grown under darkness on 0.5 MS medium supplemented with different compounds (x -axis).
Their hypocotyls were measured after eight days. Values are plotted as Mean ± SD, significant differences
(p<0.05 and p <0.001, Welch’s t-test) between Col-0 and the overexpressing l ines are indicated by (*) or
(***), respectively. Welch’s t-test for DIM1: t(37.034)=-16.316, p<0.0001; DWF1: t(11.23964)=-19.3,
p<0.0001) and clotrimazole 5µM(DIM1: t(30.72)=-7.34, p<0.05; DWF1: t(6.46)=-1.69, p<0.05).

25
The line overexpressing YFP-STE1 (STE1) showed another behavior when challenged with the compounds. Instead of promoting resistance to the inhibitors through overexpressing a sterol
biosynthesis enzyme, the hypocotyl length was significantly reduced in the overexpressing line at
high and low concentrations of each compound (Figure 11 A-D). After a two-way analysis of
variance (ANOVA), the short length can be attributed to both the treatments and the STE1
Figure 11. Hypocotyl growth in a STE1 overexpressing line in response to potential inhibitors. Both Col-0 and
STE1-YFP were grown under darkness and on 0.5 MS supplemented with different concentrations of diverse
inhibitors. Their hypocotyls were measured after eight days. Values are plotted as Mean ± SD. Asterisks (***)
show P-value significance (p<0.001) after a two-way ANOVA analysis. ANOVA was conducted on the influence of
treatment (each concentration of compound) and line (Col -0 and STE1-YFP) on hypocotyl length after 8 days after
germination. Main effect for voriconazole treatment: ratio of F(2,86)= 103.89, p<0.0001, main effect for plant
l ine: F ratio of F(1, 86)=143.28, p<0.0001. The interaction was not significant. Main effect for artemether
treatment: F ratio of F(2,86)= 83.73, p<0.0001, Main effet of plant l ine: F ratio of F(1,86)=172.55, p<0.0001. The
interaction was not significant. Main effect for clotrimazole treatment: F ratio of F(1,86)=159.48, p<0.0001, main
effect for plant l ine: F ratio of F(2,86)=163.02, p<0.0001. The interaction was not significant. Main effect for
oxiconazole treatment: F ratio of F(2,86)=51.92, p<0.0001, main effect of plant l ine: F ratio of F(1,86)=78.01,
p<0.0001),
0
0.5
1
1.5
2
Col-0 STE1
Ro
ot
len
gth
(cm
)
Plant l ines
Voriconazole treatment
0
0.3µM
1µM
0
0.5
1
1.5
2
Col-0 STE1
Ro
ot
len
gth
(cm
)
Plant l ines
Oxiconazole treatment
0
0.3µM
1µM
0
0.5
1
1.5
2
Col-0 STE1
Ro
ot
len
gth
(cm
)
Plant l ines
Artemether treatment
0
0.3µM
1µM
0
0.5
1
1.5
2
Col-0 STE1
Ro
ot
len
gth
(cm
)
Plant l ines
Clotrimazole treatment
0
0.3µM
1µM
A B
C D
*** *** *** ***
*** ***
*** ***

26
overexpressing line, showing a hypersensitivity linked to STE1 overexpression against all the
tested compounds. STE1 is naturally expressed in structures that surround the nucleus and that coincide with the ER (Silvestro et al., 2013). These observations were made in leaf tissue. Other
expression databases indicate that STE1 is also expressed, though mildly, in the hypocotyl (Winter et al., 2007). In general, overexpression assays provide an artificial setting wherein the challenging
conditions (in this case the inhibitor presence) can be tested and results can be a starting point to further investigate the role of the overexpressed gene product on the behavior of the plant
regarding the challenge, in this case hyper or hyposensitivity to the inhibitor presence. Further
assays can assess if these responses are linked specifically to the overexpressed gene products and the manner that this occurs.
3.1.4. Effects of other compounds on Arabidopsis mutants in the sterol biosynthesis pathway
Overexpressing genes in the sterol biosynthesis pathway gave us divergent results in terms of hyper- or hyposensitivity to our compounds in terms of hypocotyl length. If the over-abundance
of one of the enzymes of the pathway can modify the response of the plants, then mutations on the various catalyzers that shape the pathway could also have repercussions on the plant
sensitivity to the compound. The pathway then would be disrupted twice and the effects of the compound could be even more severe. We chose to work with mutants that belong to the post-
bifurcation part of the sterol biosynthesis pathway (cvp1/smt2, dwf1, cyp710a1-1 and cyp710a2-2), as they belong to the proper C-24 ethylsterol pathway that is different from the
brassinosteroid-related branch and they catalyze the production of intermediates that are
exclusive of plants. These mutants were grown along Col-0 under different concentrations of tamoxifen, artemether, clotrimazole, oxiconazole and voriconazole and under darkness for eight
days as described previously, and their hypocotyls were measured (Figure 12). After a two way
ANOVA, we found a strong genotype effect that means that dwf1 behaves differently from the
other mutants. All the other mutants and the wild type were undistinguishable from each other (Tukey multiple comparisons correction (TMC) between main effects after the analysis of
variance, significance level=0.05). For the treatments, we found that the effect of the compound s
was significant for all concentrations and the mutants (except for dwf1) and Col-0 respond equally to the treatments except for voriconazole, where a difference between Col-0 and cyp710a1-1 and
the other two mutants was found. There was no difference between cvp1 and Col-0 regarding
tamoxifen, neither between treatments or genotypes. The behavior of both lines was statistically
indistinguishable. Voriconazole and clotrimazole affected the length of the root more drastically in every genotype, with more pronounced effects than oxiconazole and artemether at 0.3µM. At
1µM, the reduction of the hypocotyl reached a level where the compounds were indistinguishable. Tamoxifen was an exception to this: the reduction at the hypocotyl was not
significant at any concentration used, neither in Col-0 nor in cvp1.

27
Figure 12. Hypocotyl length in response to potential inhibitors in mutants, per compound. Sterol biosynthesis
inhibiting compounds in different concentrations were supplemented to 0.5 MS medium in which Col-0 seedlings
were sown and grown in the dark. Their hypocotyl was measured after eight days. Values are plotted as Mean ±
SD. Asterisks (***) show P-value significance (p<0.001) after a two-way ANOVA on the influence of treatment
(each concentration of compound) and genotype (Col -0 and the cvp1/smt2, dwf1, cyp710a1-1 and cyp710a2-2
mutants) on hypocotyl length. Artemether main effect of treatment: ratio of F(2, 143)= 99.23 , p<0.0001, main
effect for plant l ine: F ratio of F(3, 143)=49.86, p<0.0001. The interaction between effects was not significant.
Main effect for clotrimazole treatment: F ratio of F(2, 143)= 228.67 , p<0.0001. Main effect for plant l ine: F ratio
of F(3, 143)=38.76, p<0.0001. Main effect for voriconazole treatment: F ratio of F(2, 143)= 185.09 , p<0.0001.
0
0.5
1
1.5
2
2.5
0µM 0.3µM 1µM
Hyp
oco
tyl
len
gth
(cm
)
concentration in medium (µM)
Artemether treatment
Col-0
cvp1
dwf1
710a1-1
710a2-2 0
0.5
1
1.5
2
2.5
0µM 0.3µM 1µM
Hyp
oco
tyl
len
gth
(cm
)
concentration in medium (µM)
Clotrimazole treatment
Col-0
cvp1
dwf1
710a1-1
710a2-2
0
0.5
1
1.5
2
2.5
0µM 0.5µM 1µM
Hyp
oo
oty
l le
ngt
h (
cm)
concentration in medium (µM)
Tamoxifen treatment
Col-0
cvp1
0
0.5
1
1.5
2
2.5
0µM 0.3µM 1µM
Hyp
oco
tyl
len
gth
(cm
)
concentration in medium (µM)
Oxiconazole treatment
Col-0
cvp1
dwf1
710a1-1
710a2-20
0.5
1
1.5
2
2.5
0µM 0.3µM 1µM
Hyp
oco
tyl
len
gth
(cm
)
concentration in medium (µM)
Voriconazole treatment
Col-0
cvp1
dwf1
710a1-1
710a2-2
***
*** ***
***
***
*** ***
***
*** ***
***
*** ***

28
3.2 Root length response to potential sterol biosynthesis inhibitor compounds
Longitudinally, the root is characterized by a series of developmental zones. The quiescent center (QC) enables its adjacent cells to continuously produce initial cells that are gradually displaced
upwards, giving rise to a meristematic zone where they undergo additional rounds of division and eventually become displaced from the root meristematic zone. Subsequently, these displaced
cells become part of the elongation zone and then the differentiation zone. As such, the longitudinal axis of a root represents a constantly renewing gradient of cell differentiation that
gives length as a result. The rate of cell division and cell elongation is determined by a fine
regulation of signaling and developmental cues, such as auxin, and is also dependent on sterol availability, as sterols are critical to ensure correct cell division which in turn influences auxin
trafficking (Jaillais & Gaude, 2008, Nakamoto et al., 2015, Overvoorde et al., 2010).
Thus, root length is also a good indicator of sterol abundancy with a potential link to auxin signaling. We decided to test the effect of our inhibitor candidates and to find the concentrations
of each compound that we should use for further assays given that an effect on root length was found. We grew Col-0 seedlings in the presence of different concentrations of the candidate
compounds in continuous light conditions and after eight days we measured their roots (Figure 13). Every compound caused a reduction in root length, having the most drastic mean length
decrease the azole fungicides voriconazole, oxiconazole and clotrimazole, with reductions from
70-80% at concentrations as low as 0.5µM. Tamoxifen and artemether also showed reduction in
length, reaching the same level of reduction at 5µM. At 10µM, the root mean length was not significantly different regardless of the treatment used. To further dissect the effect of the
0
5
10
15
20
25
30
35
0mM .5µM 1µM 5µM 10µM
Ro
ot
len
gth
(m
M)
Concentration in medium
Tamoxifen
Oxiconazole
Voriconazole
Clotrimazole
Artemether
Figure 13. Root growth in response to potential inhibitors. Sterol biosynthesis inhibiting compounds in
different concentrations were supplemented to 0.5 MS medium in which Col -0 seedlings were sown.
Their root length was measured after eight days of growth under continuous light. Values are plotted as
Mean ± SD.

29
compound on the root, lower concentrations of each compound (0.1 and 0.3 µM) were
supplemented to the medium on which the seedlings were grown for eight days under continuous light. The roots were measured (Figure 14) and we could see reduction in the mean length under
every 0.3µM treatment, more pronounced in the case of clotrimazole and less pronounced in the case of tamoxifen, voriconazole and oxiconazole. (Welch’s t-test, figure 14) This helped us to
know correct concentrations to use for other growth experiments.
3.2.1. Root length response to inhibitors in sterol biosynthesis mutants
Further disruption of sterol biosynthesis pathways could give us insight of effects of the
compounds on plants that already possess a defect in a known step on the pathway in terms of sensitivity to the shortening effect of the compound. For this, we grew Col-0, cvp1 and dwf1
seedlings for 8 days under continuous light on 0.5 MS solid medium in the presence of different concentrations of our inhibitor candidates (Figure 15). A Two-way ANOVA was performed to
assess the effects of genotype and treatment per compound. Again, dwf1 was significantly different from Col-0 and cvp1 in terms of root length. The response to oxiconazole 0.3µM was
significantly different in both mutants compared to the wild type, being more sensitive. Both mutants were also more sensitive to clotrimazole at 0.3µM. Both mutants were more sensitive to
both voriconazole and tamoxifen at 0.3µM and 0.1µM. Notably, there was an clear
Figure 14. Root growth in response to low concentrations of potential inhibitors. Sterol biosynthesis
inhibiting compounds in different concentrations were supplemented to 0.5 MS medium in which Col -0
seedlings were sown. Their root length was measured after eight days of growth under continuous light.
Values are plotted as Mean ± SD, significant di fferences (p <0.001, Welch’s t-test) between clotrimazole
0.3µM and the rest of the compounds are indicated by (***). Welch’s t-test for differences with
oxiconazole 0.3µM: t(51)=14.3, p<0.0001; voriconazole 0.3µM: t(51)=-5.38, p<0.0001;and tamoxifen
0.3µM: t(30.)=9.346, p<0.0001.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Oxiconazole Clotrimazole Voriconazole Tamoxifen
Ro
ot
len
ght
(cm
)
Compound0µM 0.1µM 0.3µM
*** ***
***

30
hypersensitivity effect of tamoxifen on root elongation of cvp1 that was not present in hypocotyl
elongation, where the hypocotyls were not at all reduced in response to these concentrations of tamoxifen (Figure 12). There also appeared to be a significant effect of clotrimazole and
oxiconazole that was not noticeable when testing hypocotyl elongation, as the hypocotyls were indeed shorter but not significantly different from the response found in Col-0. Voriconazole
0
0.5
1
1.5
2
2.5
Col-0 cvp1 dwf1
Ro
t le
ngt
h (
cm)
Plant l ine
Oxiconazole treatment
0µM 0.1µM 0.3µM
0
0.5
1
1.5
2
2.5
Col-0 cvp1 dwf1
Ro
ot
len
gth
(cm
)
Plant l ine
Tamoxifen treatment
0µM 0.5µM 1µM
0
0.5
1
1.5
2
2.5
Col-0 cvp1 dwf1
Ro
ot
len
gth
(cm
)
Plant l ine
Voriconazole treatment
0µM 0.1µM 0.3µM
0
0.5
1
1.5
2
2.5
Col-0 cvp1 dwf1
Ro
ot
len
gth
(cm
)
Plant l ine
Clotrimazole treatment
0µM 0.1µM 0.3µM
Figure 15. Root length of sterol biosynthesis mutants . Col-0, cvp1 and dwf1 seedlings were sown and grown on 0.5 MS medium supplemented with different sterol biosynthesis inhibitors in different concentrations for 8 days and their roots were measured. Values are mean ±SD. Asterisks (***, **, *) show P-value significance
(p<0.001, p<0.01 p<0.05) after a two-way ANOVA analysis for genotype and treatment effects. Voriconazole main treatment effect: F ratio of F(2, 103)= 18.31 , p<0.0001. F ratio of F(2, 103)=19.37, p<0.001. The interaction between effects was not significant. Tamoxifen main effect for treatment: F ratio of F(2, 103)= 18.57
, p<0.0001. Plant l ine effect: F ratio of F(2, 103)=21.19, p<0.001. The interaction between effects was not significant. Main effect for oxiconazole treatment: F ratio of F(2, 103)= 35.44 , p<0.0001. Main effect for plant l ine: F ratio of F(2, 103)=3.35, p=0.0389. Main effect for clotrimazole treatment: F ratio of F(2, 103)= 57.70 , p<0.0001. Main effect for plant l ine: F ratio of F(2, 103)=6.17, p=0.0039. The interaction between effects was not significant. The data regarding Col -0 length in response to compounds is the same reported in figure 14.
*** *
**
***
*** *** *** ***
*** *** *** ***

31
effect was both significant regarding hypocotyls and root lengths. In all cases, the effect of the
compounds was significant when treating dwf1, as it was when the response of the hypocotyls was tested.
3.3 Effects of auxin treatment
3.3.1 Effect of auxin on the root length of wild type plants and sterol biosynthesis mutants
Auxin is a signal that acts primarily as a transcriptional activator. It also orchestrates root
architectural changes by precisely tuning gradients where activation or repressing of genes are needed, by biosynthesis, transport or degradation, resulting in changes such as length, as
mentioned before. Auxin supplementation, depending on the site and concentration, could promote or repress the expression of genes that could help rescue or further incapacitate the
sterol biosynthetic pathway in the context of auxin treatment of mutants.
Moreover, if sterol biosynthesis and availability has a direct effect on auxin transport and auxin is exogenously supplemented, it could indicate that certain intermediate of the pathway is
presumably involved in the ensuring of a correct auxin signaling. For this, we decided to test the response of our Arabidopsis plants to auxin by assessing its effect on root length. The auxin 2,4-
Dichlorophenoxyacetic acid (2,4-D) displays an inhibitory effect on plant growth via the TIR1/AFB
auxin-mediated signaling pathway and promotes cell division but no root elongation, leading to root shortening and plant death after certain concentrations (Campanoni & Nick, 2005).
Seedlings of the following sterol biosynthesis gene mutants were grown in the presence of different concentrations of 2,4-D in order to evaluate its effect on cell elongation: dwf1, cvp1,
cyp710A1-1, A2-1 and A2-2 along with Col-0. Two experiments were done in order to test a variety
of auxin concentrations. The effects of root growth compared to mock-treated roots are shown in Figure 16. We observed an evident reduction on root elongation in all lines, being dwf1 the
most sensitive one and cvp1 the least auxin sensitive. The mean lengths of cvp1 and dwf1 were determined to be significantly different from the mean length of Col-0 roots subject to 50nM of
2,4-D by carrying out a two tailed Welch’s t-test. That indicates that dwf1 is more sensitive to auxin than Col-0 and that cvp1 is more resistant to auxin supplementation than Col-0.
3.3.2 Effect of auxin on the root length and hypocotyl of wild type plants and cvp1 under
candidate inhibitor treatment
Knowing that both cvp1 and Col-0 responded distinctly to auxin and the inhibitors both at the
concentrations previously shown, we decided to test if auxin supplementation caused any anomaly in the behavior of cvp1 and Col-0 when confronted with the compounds. We sowed
both lines on compound-supplemented and/or 50nM 2-4 D-supplemented solid 0.5MS and grew
them either under the dark (for hypocotyl elongation) or under continuous light (for root growth) for 8 days (Figure 17). A three-way ANOVA was conducted on the influence of treatment (each

32
concentration of compound), auxin supplementation and line (Col-0 and cvp1) on hypocotyl
length 8 days after germination (Results in table 1). We found that artemether-treated roots under auxin treatment are significantly longer than the ones without auxin, this attributable to
auxin treatment. Hypocotyls under tamoxifen treatment are similarly long in both auxin-treated
and non-treated Col-0 and cvp1. Tamoxifen application has no effect on auxin resistance in roots, as he roots of the auxin-treated and non-treated plants of each lines behave the same.
Additionally, cvp1 shows the same hypersensitivity to tamoxifen with and without auxin supplementation, as shown by their significantly shorter roots compared to those of Col -0 due to
an effect of Tamoxifen and the genotype. Apparently, auxin supplementation gives artemether resistance to the plant at least to maintain long hypocotyls under darkness, but confers no
resistance to the plant to avoid root length reduction under artemether treatment. Tamoxifen treatment causes no effect on the plant with or without auxin supplementation, effectively
yielding plants with long roots compared to non-auxin treated plants. On the other hand, cvp1
plants were tamoxifen resistant concerning their hypocotyl length but hypersensitive to tamoxifen in their root length, both with or without auxin supplementation, reaching in both
cases comparable levels of root length stunting. Roots from cvp1 and Col-0 responded in an identical manner regarding auxin under oxiconazole, voriconazole and clotrimazole treatments.
0
0.5
1
1.5
2
2.5
0 10 50 100
Ro
ot
len
gth
(cm
)
2-4 D in medium (nM)
Col-0
710a1-1
710a2-1
710a2-2
cvp1
dwf1
Figure 16. Root length after auxin treatment. Sterol biosynthesis mutants and Col -0 were sown
in 0.5MS medium in the presence of different concentrations of 2,4-D. Their roots were
measured eight days after germination and the relative length calculated respect to untreated
seedlings of the same line. A) First experiment. B) Second experiment. Values are plotted as
mean percentages ± SEM. Asterisks denote significant reduction or increase in length (Welch’s t
test P value < 0.001; for cvp1, 50µM 2,4-D: t(59.36)=-6.93, p<0.001; for dwf1, 50µM 2,4-D:
t(36.17)=8.72, p<0.01).
***
***

33
0
1
2
3
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Ro
ot
len
gth
(cm
)
Plant l ine and treatment
Voriconazole
DMSO
0.3µM
0.5µM
0
0.5
1
1.5
2
2.5
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Hyp
oco
tyl
len
gth
(cm
)
Plant l ine and treatment
Voriconazole
DMSO
0.3µM
0.5µM
0
1
2
3
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Ro
ot
len
gth
(cm
)
Plant l ine and treatment
Clotrimazole
DMSO
0.3µM
0.5µM
0
1
2
3
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Ro
ot
len
gth
(cm
)
Plant l ine and treatment
Oxiconazole
DMSO
0.3µM
0.5µM
00.5
11.5
22.5
3
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Ro
ot
len
gth
(cm
)
Plant l ine and treatment
Artemether
DMSO
0.3µM
0.5µM
0
1
2
3
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Ro
ot
len
gth
(cm
)
Plant l ine and treatment
Tamoxifen
DMSO
0.5µM
1µM
0
0.5
1
1.5
2
2.5
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Hyp
oco
tyl
len
gth
(cm
)
Plant l ine and treatment
Clotrimazole
DMSO
0.3µM
0.5µM
0
0.5
1
1.5
2
2.5
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Hyp
oco
tyl
len
gth
(cm
)
Plant l ine and treatment
Oxiconazole
DMSO
0.3µM
0.5µM
0
0.5
1
1.5
2
2.5
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-DH
ypo
coty
l le
ngt
h (
cm)
Plant l ine and treatment
Artemether
DMSO
0.3µM
0.5µM
0
0.5
1
1.5
2
2.5
Col-0 Col-0 +2,4-D
cvp1 cvp1 +2,4-D
Hyp
oco
tyl
len
gth
(cm
)
Plant l ine and treatment
Tamoxifen
DMSO
0.5µM
1µM
***
***
***
***
***
***

34
Table 1. ANOVA results for root and hypocotyl length of cvp1 and Col-0 in the presence of supplemented auxin and
sterol biosynthesis inhibitors.
Compound Source of variation
Main effect on hypocotyl length Main effect on root length
Artemether Genotype F(1, 235)=31.36 p<0.0001 F(1, 235)=5.67 p=0.0181
Concentration F(2, 235)= 78.27 p<0.0001 F(2, 235)= 165.26 p<0.0001
Auxin F(1, 235)=1.03 p=0.3108 F(1, 235)=72.47 p<0.0001
Clotrimazole Genotype F(1, 235)=25.76 p<0.001 F(1, 235)=3.18 p=0.0759
Concentration F(2, 235)= 322.45 p<0.0001 F(2, 235)=897.20 p<0.0001
Auxin F(1, 235)=11, p=0.0011 F(1, 235)=132.32 p<0.0001
Oxiconazole Genotype F(1, 235)=9.91 p=0.0019 F(1, 235)=72.28 p<0.0001
Concentration F(2, 235)= 135.26 p<0.0001 F(2, 235)= 193.02 p<0.0001
Auxin F(1, 235)=6.56 p=0.0111 F(1, 235)=72.28 p<0.0001
Tamoxifen Genotype F(1, 235)=56.98 p<0.001 F(1, 235)=208.81 p<0.0001
Concentration F(2, 235)= 4.12 p=0.0175 F(2, 235)= 126.7 p<0.0001
Auxin F(1, 235)=56.98 p<0.001 F(1, 235)=28.19 p<0.0001
Voriconazole Genotype F(1, 235)=44.06 p<0.0001 F(1, 235)=0.15 p=0.7012
Concentration F(2, 235)= 491.32 p<0.0001 F(2, 235)=725.02 p<0.0001
Auxin F(1, 235)=9.16 p=0.0027 F(1, 235)=111.04 p<0.0001
3.4 Calcium supplementation experiments
A previous report described the importance of SMT1 as a regulator of cholesterol levels in
Arabidopsis by characterizing the smt1 mutant (Diener et al., 2000). Arabidopsis contains three
genes that are homologous to yeast ERG6: sterol methyltransferase 1 (SMT1), SMT2, and SMT3
which are known to encode an S-adenosylmethionine–dependent C-24 SMT that catalyzes a
single methyl addition. SMT1 is a C-24 methyltransferase responsible for the conversion of
cycloartenol to 24-methylenecycloartenol in the first step of sterol biosynthesis, whereas SMT2
and SMT3 are the enzymes responsible of the branching that gives place to C24 ethyl sterols
(Carland et al., 2010, Clouse, 2002). It has been described that smt1 mutants have root defects
that impair length, being at least in part due to calcium ion sensitivity. The basis of the claim is
that roots of the smt1 mutant that had been grown in 0.05MS medium are longer and with overall
better aspect than the ones grown in 0.5MS. The explanation relies on calcium sensitivity claiming
that other elements in the nutrient mix had been also tested and shown not to be responsible for
the change of root length (Diener et al., 2000). The smt2/cvp1 mutants show no root grow
impairments (the double mutant cvp1/smt3 does) and is comparable to Col-0 roots in length
Figure 17. Root and hypocotyl length of cvp1 and Col-0 in the presence of supplemented auxin and sterol
biosynthesis inhibitors. Col-0 and cvp1 seedlings were sown and grown on 0.5 MS medium supplemented with
different sterol biosynthesis inhibitors and /or 2,4-D in different concentrations for 8 days and their hypocotyls
(left) and roots (right) were measured. Values are Mean ±SD. Arrows indicate hypersensitivity (red) or resistance
(green) to compound or auxin. Asterisks (***, **, *) show P-value significance (p<0.001, p<0.01 p<0.05) after a
three-way ANOVA analysis for genotype, auxin and treatment (compound) effects.

35
when grown in 0.5MS. The mutation affects a downstream step on the pathway from smt1
(Carland et al., 2010).
3.4.1 Root length on diluted medium
We decided to test if calcium depletion would affect growth of the root in the cvp1 mutant given
that SMT1 and SMT2 are both homologs of ERG6 and therefore possibly affected by the same
environmental stimuli. Additionally, the response of downstream Δ24 reductase dwf1 mutants
would be tested too. For this, we followed the already described procedure and sown Col-0, dwf1
and cvp1 seeds either on solid 0.5MS medium or solid MS with a ten-fold diluted nutrient salt
mix and same sucrose content (0.05MS; see methods). After a week of continuous light growth,
their roots were measured (Figure 18). Surprisingly, in every line, diluted medium caused
increased root elongation.
A two-way ANOVA was conducted on the influence of medium (0.05 MS or 0.5MS) and line (Col-
0 and cvp1) on root length 8 days after germination. We found that again, dwf1 was set apart on
behavior from Col-0 and cvp1, which responded identically to medium dilution. However,
medium dilution had an effect on all genotypes, causing root lengthening.
Figure 18. Root length of dwf1, cvp1 and Col-0 in 0.5MS and 0.05MS (diluted medium). Col-0, dwf1 and cvp1 seedlings were sown and grown on either 0.5 MS or 0.05MS medium for 8 days and their roots were
measured. Values are Mean ±SD. Asterisks (***, **, *) show P-value significance (p<0.001, p<0.01 p<0.05) after a two-way ANOVA analysis. The main effect for medium yielded an F ratio of F(1, 86)= 51.11, p<0.0001, main effect of genotype: F ratio of F(2, 86)=5.65, p=0.0050.The interaction between effects was not significant.
0
0.5
1
1.5
2
2.5
3
Col-0 cvp1 dwf1
Ro
ot
len
gth
(cm
)
0.5MS 0.05MS
***
*** ***
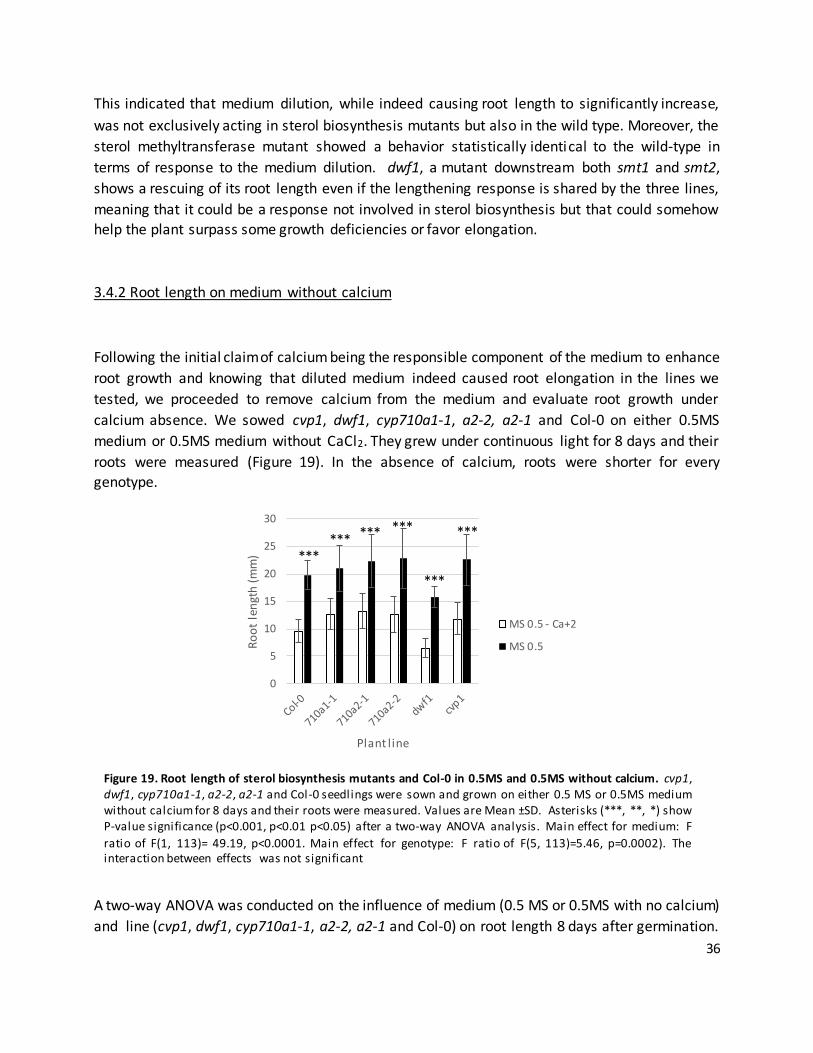
36
This indicated that medium dilution, while indeed causing root length to significantly increase,
was not exclusively acting in sterol biosynthesis mutants but also in the wild type. Moreover, the
sterol methyltransferase mutant showed a behavior statistically identical to the wild-type in
terms of response to the medium dilution. dwf1, a mutant downstream both smt1 and smt2,
shows a rescuing of its root length even if the lengthening response is shared by the three lines,
meaning that it could be a response not involved in sterol biosynthesis but that could somehow
help the plant surpass some growth deficiencies or favor elongation.
3.4.2 Root length on medium without calcium
Following the initial claim of calcium being the responsible component of the medium to enhance
root growth and knowing that diluted medium indeed caused root elongation in the lines we
tested, we proceeded to remove calcium from the medium and evaluate root growth under
calcium absence. We sowed cvp1, dwf1, cyp710a1-1, a2-2, a2-1 and Col-0 on either 0.5MS
medium or 0.5MS medium without CaCl2. They grew under continuous light for 8 days and their
roots were measured (Figure 19). In the absence of calcium, roots were shorter for every
genotype.
A two-way ANOVA was conducted on the influence of medium (0.5 MS or 0.5MS with no calcium)
and line (cvp1, dwf1, cyp710a1-1, a2-2, a2-1 and Col-0) on root length 8 days after germination.
0
5
10
15
20
25
30
Ro
ot
len
gth
(m
m)
Plant l ine
MS 0.5 - Ca+2
MS 0.5
****** *** ***
***
***
Figure 19. Root length of sterol biosynthesis mutants and Col-0 in 0.5MS and 0.5MS without calcium. cvp1, dwf1, cyp710a1-1, a2-2, a2-1 and Col-0 seedlings were sown and grown on either 0.5 MS or 0.5MS medium without calcium for 8 days and their roots were measured. Values are Mean ±SD. Asterisks (***, **, *) show P-value significance (p<0.001, p<0.01 p<0.05) after a two-way ANOVA analysis. Main effect for medium: F
ratio of F(1, 113)= 49.19, p<0.0001. Main effect for genotype: F ratio of F(5, 113)=5.46, p=0.0002). The interaction between effects was not significant

37
The behavior of dwf1 was different from the rest of the mutants due to its genotype, but the
effect of the medium affected every plant line with an effect of length shortening. Calcium
absence, thus, at least for all the sterol biosynthesis mutant lines we tested, caused a reaction
that differed from the response that was reported for smt1 on 0.005MS, a reaction that was
shared by the wild type as well. It also affected greatly the length of dwf1 mutants, with the
possibility of being related to the previous results with diluted medium enhancing root elongation
especially in that line.
3.4.3 Root length on diluted medium, medium with no calcium and under sterol biosynthesis
inhibitor treatment
Knowing that calcium absence caused root shortening and medium dilution caused root
lengthening, we wished to test in the wild-type if there was some difference in root length under
sterol inhibitor treatment with different nutrient availability. For this, we sowed Col -0 seeds on
either 0.5MS, 0.5MS without calcium or 0.05MS solid medium supplemented with different
concentrations of sterol biosynthesis inhibitors for 8 days under continuous light and we
measured their roots (Figure 20).
When sown on diluted MS medium, the response to the compounds was more sensitive in terms
of root shortening and compound concentration (except for clotrimazole) but at 0.3µM all
compounds caused identical root growth stunting (Figure 20, left). When sown on calcium-free
medium, the root was extremely short even under low concentrations of the compounds, being
significantly shorter than the ones in 0.5MS even under higher concentrations of the compound
(0.5µM for oxiconazole, clotrimazole and voriconazole and tamoxifen 1µM) (Figure 20, right).
A two-way ANOVA was conducted on the influence of medium (0.5 MS, 0.05MS or 0.5MS with no
calcium) and treatment (different concentrations of the compound) on root length 8 days after
germination. It showed that in all cases, the differences in root length were due the medium, either diluted or calcium free, and the treatment being the different concentrations in medium.
When grown in diluted (0.05 MS) medium, the roots were longer because of the medium but
rapidly affected by the compounds, reaching both the same stunting under higher concentrations.
In contrast, the calcium-free medium caused roots to stunt even with very low concentrations of
each compound, and even under higher concentrations of the inhibitor the stunting of the root
never reached that under calcium-free conditions, perhaps due an effect of calcium on root
elongation that could potentiate the effect of the inhibitor, being the calcium depletion not
enough to potentiate the effect of the compound when the medium was just diluted. The effect
of the dilution do not protect the root against the compound even if causes root lengthening.

38
Figure 20. Root length of Col-0 calcium free and diluted medium. Col-0 seedlings were sown and grown on either
0.5 MS, calcium free 0.5MS or 0.05MS medium supplemented with different concentrations of candidate inhibitors for 8 days and their roots were measured. Values are Mean ±SD. Medium and treatment had significant effects on root length in both diluted and calcium-free medium. ANOVA for treatment and medium in terms of root length, per medium. Effects from diluted medium (left) and per compound as follows: clotrimazole
treatment, medium main effect: F(1, 146)= 44.99, p<0.0001. treatment main effect: F(2, 146)=5.46, p<0.0001. Oxiconazole, medium main effect:F(1, 146)= 37.12, p<0.0001. Main effect for treatment: F(2, 146)=46.42, p<0.0001. Tamoxifen treatment, medium main effect: F(1, 146)= 123.46, p<0.0001. treatment main effect: F ratio of F(2, 146)=16.86, p<0.0001. Voriconazole treatment, medium main effect:F(1, 146)= 146.57, p<0.0001. Main
effect for treatment: F(2, 146)=69.23, p<0.0001. The interaction between effects was not significant. Effects from calcium-free medium (right) and per compound as follows: Clotrimazole treatment, medium main effect: F(1, 171)= 326.11, p<0.0001. Treatment main effect: F(3, 146)=189.35, p<0.0001. Oxiconazole treatment, medium
main effect: F(1, 171)= 326.11, p<0.0001. Main effect for treatment: F(3, 146)=189.35, p<0.0001. Tamoxifen treatment, medium main effect: F(1, 171)= 500.45, p<0.0001. Main effect for treatment:F(3, 146)=18.72 p<0.0001. Voriconazole treatment, main effect of medium: F(1, 171)= 600.98, p<0.0001. Main effect of treatment: F(3, 146)=179.98, p<0.0001. The interaction between effects was not significant.

39
3.4.4 Calcium supplementation on root length
Medium dilution and absence of calcium were shown to affect root length in a contrasting way
but they did not affect the response to inhibitors in a particular manner in the wild-type. It was
interesting to see if there existed a concentration of calcium in the medium that turned the tables
on root length from shrinkage to normality and further to lengthening.
For this, we set up a calcium supplementation gradient experiment, where Col-0 Seeds were sown
in either 0.05MS or calcium-free 0.5MS medium with gradually increasing concentrations of
calcium added, from 0mM to the normal concentration of CaCl2 in 0.5MS (1.5mM), grown under
continuous light for 8 days and their roots measured (Figure 21). With no supplemented calcium,
root growth was as expected with stunted roots grown on calcium-free medium and longer roots
grown on diluted medium. As the CaCl2 increased, roots grown in calcium-free medium gradually
approached the same length of roots grown in 0.5MS, until reaching the same length around
supplemented 0.5mM CaCl2 (statistically not different, Welch’s two tailed t-test, p>0.1). However,
no supplemented CaCl2 concentration to 0.05MS medium grown roots was able to reduce root
length elongation comparable to non-supplemented 0.05MS medium-grown roots, which
Figure 21. Root length of Col-0 in calcium free and diluted medium after calcium supplementation. Col-0 seedlings were sown and grown on either 0.5 MS, calcium free 0.5MS or 0.05MS medium s upplemented with different concentrations of CaCl 2 for 8 days and their roots were measured. Values are Mean ±SEM.
Asterisks denote significant difference with respect to 0.5MS (Welch’s t test P value < 0.001) For calcium-free medium, 0.0 mM: t(34)=15.0499, p<0.001; 0.01mM: t(37)=8.3370, p<0.001; 0.05mM: t(34)=4.72, p<0.001; 0.1mM: t(38)=3.207, p=0.0027; 0.5mM: t(33)=1.7545, p=0.08; 1mM: t(37)=0.4889, p=0.627; 1.5mM: t(24)=0.0840, p=0.933; for 0.05MS medium, 0.0mM: t(19)=8.6137, p<0.001; 0.01mM: t(20)=7.014,
p<0.001; 0.05mM: t(27)=7.398, p<0.001; 0.1mM: t(28)=5.27, p<0.001; 0.5mM: t(24)=7.7002, p<0.001; 1mM: t(30)=9.1652, p<0.001; 1.5mM: t(26)=5.3412, p<0.001;
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.5MS 0 0.01 0.05 0.1 0.5 1 1.5Ro
ot
gro
wth
re
lati
ve to
co
ntr
ol (
0.5
MS)
Supplemented CaCl2 (mM)
0.05MS
No Ca+2
***
***
*** *
***
*** ***
***
*** ***
***

40
indicates that calcium is not the element causal to the enhanced root elongation phenotype in
the 0.05MS medium. This shows again that root elongation due to medium dilution cannot be responsibility of calcium depletion.
3.5 cvp1 response to tamoxifen
The Arabidopsis mutant in the SMT2 gene, cvp1, showed no response to tamoxifen regarding
hypocotyl length reduction, (Figure 12). However, when root length under tamoxifen treatment
was measured, a drastic decrease was shown even in low (0.5µM) concentrations, which in Col-0
did not affect root growth (Figure 15). Auxin supplementation did not affect hypocotyl or root
length response to tamoxifen in cvp1 or Col-0, being their hypocotyls both relatively unaffected
by tamoxifen with and without auxin supplementation and cvp1 roots being very sensitive to
tamoxifen regardless of auxin supplementation (Figure 17). We wished to further address the
question about the possibility of calcium depletion to affect in any way the apparent tamoxifen
hypersensitivity of cvp1.
0
5
10
15
20
25
30
0 0.5 1 5
Ro
ot
len
gth
(m
m)
concentration (µM)
Col-0, 0.5MS No Ca+2
Col-0, 0.5MS
cvp1, 0.5MS No Ca+2
cvp1, 0.5MS
Figure 22. Root length of Col-0 and cvp1 in calcium free and 0.5MS medium supplemented with tamoxifen.
Col-0 and cvp1 seedlings were sown and grown on either 0.5 MS or calcium free 0.5MS medium supplemented with different concentrations of tamoxifen for 8 days and their roots were measured. Values are Mean ±SD. Asterisks (***, **, *) show P-value significance (p<0.001, p<0.01 p<0.05) after a three-way ANOVA analysis on
the influence of medium (0.5 MS or calcium-free 0.5MS), genotype (cvp1 and Col -0) and treatment (different concentrations of tamoxifen) on root length 8 days after germination. Main effect for medium: F ratio of F(1, 346)= 266.50, p<0.0001. Main effect for treatment: F ratio of F(3, 346)=55.65, p<0.0001. Main effect for genotype: F ratio of F(1, 346)=104.77, p<0.0001. The interaction between effects was not significant.
.
***
***
***

41
We sowed and grew both Col-0 and cvp1 plants on solid 0.5 MS or calcium-free 0.5MS medium
supplemented with four different concentrations of tamoxifen for 8 days under continuous light and then we measured the roots (Figure 22).
Consistent with our earlier experiments, low concentrations (0.5µM) of tamoxifen, did not affect
the root length of Col-0, while dramatically reducing the length of cvp1. Under calcium starvation
conditions, no effect of tamoxifen could be observed on Col-0 roots, in addition to the reduced
root length due to calcium starvation. Under calcium starvation, the level of root length reduction
of cvp1 could also be observed, with the length of the root being even shorter in higher
concentrations (1µM, 5µM) of tamoxifen. However, the effect was less pronounced that on
normal 0.5MS
In summary, cvp1 appears to be extremely hypersensitive to tamoxifen, perhaps due being its
target, even when earlier reports in mammals point to the mammalian orthologs of HYD1 ( Δ8-7
isomerase) or DWF1 (Δ24-sterol reductase, DHCR24) as possible targets, or due the possible
imbalance of sterols caused by SMT2 absence that has a consequence on target abundance
(DWF1 is situated downstream in the biosynthetic pathway). Additional data of ours indicate no
particular sensitivity to tamoxifen in dwf1 mutants, either in diluted or complete medium (data
not shown). This could be also due to the extremely disturbed phenotype natural to dwf1
mutants, which could mask any extra sensitivity under an already stunted root growth.
3.6 Effect of candidate inhibitor compounds on auxin transporter expression in roots.
The plasma membrane possess an important amount of phytosterols, which are important to
provide structural and chemical viability to the membrane to support protein-protein
interactions, meaning that they determine the localization and abundance of auxin carriers in the
membrane and therefore how effective the auxin flux across the membrane will be. The sterol
composition of membranes has been shown to be crucial for the positioning of certain auxin
transporting PIN proteins in the plasma membrane (Men et al., 2008, Petrasek & Friml, 2009,
Willemsen et al., 2003). Plasma membrane sterols are also important for the localization of
another type of auxin transporters, ABCB19, which has been found to be present in plasma
membrane microsomal fractions that are sterol- and sphingolipid rich,-and presumably constitute
microdomains at which ABCB19 and PIN1 interact physically (Bandyopadhyay et al., 2007,
Blakeslee et al., 2007). Moreover, ABCB19 stabilizes PIN1 in these fractions, and presumably
influences the rate of PIN1 endocytosis and thus its incidence at the plasma membrane
(Titapiwatanakun & Murphy, 2009). Sterol biosynthesis mutants are generally defective in cell
division and cell-to-cell auxin transport; The smt1 mutant exhibits incomplete cell wall structures
and aberrant membrane localization of the PIN1 and PIN3 auxin efflux carriers (Willemsen et al.,
2003). The cpi1-1 mutant presents severe dwarfism, loss of polar localization of PIN2, incorrect

42
endocytosis, and abnormal cytokinesis (Men et al., 2008). Other Arabidopsis mutants, such as
fackel/hydra2 and hydra1, also suffer severe developmental defects (Jang et al., 2000, Schrick et
al., 2000, Souter et al., 2002). This shows that interrupting sterol biosynthesis in any step with the
help of an synthesis inhibitor could cause developmental aberrations and mislocalizations of auxin transporters, or even a halt or perturbation of the constant PIN transport recycling.
3.6.1 Long term and short term exposure of PIN2-GFP plants to candidate inhibitors
To explore the origin of the root growth defects seen after inhibitor treatments, we analyzed the
effects of these inhibitors on the expression levels and localization pattern on the PIN2 auxin
transporter. Seeds of an Arabidopsis plant line carrying a PIN2::PIN2-GFP (PIN2-GFP) insert were
sown and grown on solid 0.5MS medium supplemented with different concentrations of the
candidate inhibitors for 8 days under constant light and then their fluorescent signal was
visualized via confocal laser microscopy (Figure 23).
Clotrimazole 0.5µM (Fig. 23B) and 1µM (23C), Oxiconazole 0.5µM (Fig. 23D) and 1µM (Fig. 23E),
and Voriconazole 0.5µM (Fig. 23H) and 1µM (Fig. 23I) treatments rendered plants that were so
severely damaged and deformed that imaging them became difficult due to the impossibility to
adequately focus and seriously disturbed cell shapes and organization, and low PIN2-GFP levels.
PIN2 localization was irregularly distributed, sometimes being the signal a blurry spot without
definition. Only tamoxifen treatment (Figure 23F-G, inset) yielded roots that although presenting
Figure 23. Long term effect of candidate inhibitors on PIN-GFP localization. Arabidopsis PIN2-GFP seedlings were sown and grown under continuous light in 0.5 MS in the presence of different concentrations and inhibitors. Their roots were photographed using confocal laser microscopy 8 days after germination. A)DMSO 0.1%, B) Clotrimazole 0.5 µM, C) Clotrimazole 1µM D) Oxiconazole 0.5µM, E)Oxiconazole 1µM, F) Tamoxifen 0.5µM, G) Tamoxifen 1µM, H) Voriconazole 0.5µM, F) Voriconazole 1µM. Scale bar=100µm.

43
cell division defects (as evidenced by misplaced cell membrane orientations and irregularities in
cell files) conserved certain organization and PIN polarity. We decided then to use lower concentrations for long-term experiments.
A short term time-course of exposure to inhibitors was then set up (Figure 24). PIN2-GFP
seedlings were sown and grown for 8 days in solid 0.5MS medium. They were then transferred
to solid 0.5MS medium supplemented with either 0.1 or 0.5 µM clotrimazole, voriconazole, and
oxiconazole or either 0.5 or 1µM tamoxifen. After 3, 6 or 24 hours, their fluorescence was
Figure 24. Short term timeline of inhibitor effects on PIN-GFP localization. Arabidopsis PIN2-GFP seedlings were sown and grown under continuous light in 0.5 MS for 8 days and transferred to medium supplemented with different concentrations of inhibi tors. Their roots were photographed using confocal laser microscopy 3,6 or 24
hr after transferring. A)DMSO 0.1% 3, 6 and 24 hrs (from left to right), B) Clotrimazole 0.1 µM 3, 6 and 24 hrs (from left to right), C) Clotrimazole 0.5µM 3, 6 and 24 hrs (from left to right), D) Oxiconazole 0.1µM 3, 6 and 24 hrs (from left to right), E)Oxiconazole 0.5 µM 3, 6 and 24 hrs (from left to right), F) Tamoxifen 0.5µM 3, 6 and 24 hrs (from left to right), G) Tamoxifen 1µM 3, 6 and 24 hrs (from left to right), H) Voriconazole 0.1µM 3, 6 and 24 hrs (from left to right), F) Voriconazole 1µM 3, 6 and 24 hrs (from left to right).Scale bar = 100µm.

44
recorded. Only after 24 hours of exposure to the higher concentrations of the compounds (0.5µM
clotrimazole, oxiconazole and voriconazole and 1µM tamoxifen) cell division and growth defects
started to be seen, with roundish cells, disorganized cell files, curved membranes and misoriented
cell division planes (Fig.24 C,D,E, G,H,I, insets), much like those shown by the smt2smt3 double
mutant (Nakamoto et al., 2015) all this while still conserving PIN2 polarity. This could signify that
inhibitors have effects on the structural arrangement and division pattern of root cells before
interfering with auxin transport, or that the concentrations used are on the verge of causing auxin
transporter defects by not completely shutting down sterol biosynthesis, a threshold of still
permitting the existence of the sphingolipid and sterol-rich microdomains needed to maintain PIN polarity.
3.6.2 Long term exposure of ABCB19-GFP plants to candidate inhibitors
Interestingly, ABCB-type auxin transporter ABCB19, has been shown to stabilize PIN1 at the PM,
and is also regulated by sterols (Yang et al. 2013). This prompted us to evaluate the effects of our
putative sterol biosynthesis inhibitors on ABCB19-GFP. For this, we sowed and grew seeds from
Figure 25. Long term effect of inhibitors on
ABCB19-GFP localization. Arabidopsis ABCB-GFP
seedlings were sown and grown under continuous light in 0.5 MS in the presence of
different concentrations and inhibitors. Their roots
were photographed using confocal laser microscopy 8 days after germination. A)
DMSO 0.1%, B) Clotrimazole 0.3 µM, C) Clotrimazole 0.5µM D) Oxiconazole
0.3µM, E) Oxiconazole 0.5µM, F) Tamoxifen 0.5µM, G) Tamoxifen
1µM, H) Voriconazole 0.3µM, F) Voriconazole 0.5µM. Scale bar=100µm.

45
an Arabidopsis line carrying ABCB19-GFP on solid 0.5MS medium containing different
concentrations of tamoxifen, clotrimazole, oxiconazole and voriconazole for 8 days under
continuous light, and then we visualized their roots via confocal laser scanning microscopy (Figure
259). Similarly to the PIN2-GFP experiments, we observed a clear defects in the organization and
the shape of the root cells between the mock-treated (Figure 25A) and the treated roots,
especially at higher compound concentrations (Figure 29 C, E, G, I, insets), with roundish, swollen,
invaginated cells and disorganized cell files due to aberrant cell divisions. However, the ABCB19-
GFP signal intensity and membrane localization was not obviously affected by the treatments.
This is in accordance with the previous report that showed no change in PM signal of ABCB19
after fenpropimorph treatment, a known sterol inhibitor, suggesting that the effects of lowering
sterol levels on PIN2-GFP intensity are not caused by reduced ABCB19 levels in the PM. ABCB19-
GFP trafficking between the TGN and PM is regulated by membrane sterol content. abcb19 plants
are partially resistant to fenpropimorph (Yang et al., 2013) and endocytosis of sterol-packed
ABCB19 is not affected by loss of sterols caused by fenpropimorph, suggested by the co-
localization of ABCB19-GFP with the endocytic marker FM4-64 at the TGN after FEN treatment
and no changes of PM ABCB19 signals upon FEN treatment (Yang et al., 2013).
3.6.3 Stigmasterol and β-sitosterol supplementation assay
Phytosterols, as already mentioned, are fundamental for the plasma membrane functionally
correct constitution, as they are involved in regulating membrane fluidity and permeability and
also determining, like in the case of PIN and ABCB auxin transporters, the localization of
transmembrane proteins. Not all eukaryotes possess the same sterol composition: cholesterol is
the principal sterol in vertebrates, whereas ergosterol is the main sterol in yeast. The variety of
sterols found in plants is much more complex. Plant sterols generally occur as a mixture of several
major species differing in the presence or position of double bonds, methylations or other
structural modifications, like the side-chain length. Sitosterol, stigmasterol, isofucosterol and 24-
methylcholesterol are end sterols common to most plants (Benveniste, 2002, Schaller, 2003,
Schaller, 2004). Sterol biosynthesis mutants show developmental and cellular alterations which
can partially be attributed to the defects in the synthesis of BR (Clouse, 2002). However,
mutations early in the sterol synthesis pathway and smt mutations lead to defects which cannot
be rescued by BR addition, suggesting that sterols also influence responses unrelated to BR
(Carland et al., 2010, Clouse, 2002).
A recent report describes the cell division and auxin transport defects of smt2smt3 mutants,
which under culture conditions with synthetic auxins yielded only disorganized tissue bumps
instead of the expected lateral root development. Exogenous β-sitosterol administration partially
rescued the failed lateral root development without restoring normal C24-ethylsterol levels, as

46
did in a milder manner stigmasterol (another significant C24-ethylsterol) but not by campesterol
(C24-methylsterol and BR precursor) (Nakamoto et al., 2015).
Having compared the overall defects that our candidate inhibitor-treated root cells presented to
those of the double smt3smt2 mutants, we decided to supplement the medium with both
stigmasterol and β-sitosterol and the compounds to see if there was any rescue of the cell
organization and morphology defects . PIN2-GFP seedlings were grown on solid 0.5MS medium
for 4 days then transferred to solid 0.5MS medium supplemented with different concentrations
of the candidate inhibitors accompanied or not by 3µg/ml stigmasterol, a concentration
previously used to rescue the smt2smt3 lateral root phenotype (Nakamoto et al, 2015). One
hundred hours after transfer, PIN2-GFP was visualized under confocal laser microscopy (Figure
26). The DMSO control treated with stigmasterol did not display any obvious differences than the
untreated DMSO control (Figure 26A). All concentrations of the compounds provoked cellular
division, organizational and structural defects such as misoriented cell walls, invaginations in the
membrane, cellular roundness and malformed cell files, more evident for voriconazole and
clotrimazole as described above (Figure 26 B-I; left). However, none of these defects could be rescued by the application of exogenous stigmasterol (Figure 26B-I; right, insets).
β-sitosterol supplementation was carried out in the exact same manner as the stigmasterol
experiment. 4 day old PIN2-GFP Arabidopsis seedlings were transferred to solid 0.5MS medium
containing certain concentrations of the candidate inhibitors with or without 3µg/ml of β-
sitosterol for 100 hours and their fluorescence was analyzed by confocal laser microscopy (Figure
27). We included to the experiment the known sterol inhibitor fenpropimorph, which led in a slighter manner to the same defects as our candidates (Fig. 27D).
Remarkably, the β-sitosterol treatment led to a dramatic reduction in PIN2-GFP signal in the
DMSO control experiment (Figure 27A). Artemether, shown to reduce the length of the root in a
drastic manner in previous experiments with the same concentration, showed the less defective
roots among the compounds (Figure 27B). Again, the root appeared to suffer the most under treatment by the azole compounds (Figure 27C,E,G).
While the exogenous supplementation of β-sitosterol did not obviously rescue any of the cellular
defects seen in the treatments, it had an interesting effect on the presence of PIN2, with a
remarkable decrease of PIN2-GFP under β-sitosterol treatment. This strong signal decrease
related to β-sitosterol addition was also seen when the root is treated with clotrimazole (Figure
27C, right) and in a slighter amount when it was treated with voriconazole (Figure 27G, right).
Interestingly, oxiconazole (another azole) did not provoke the same response when β-sitosterol
was added (Figure 27E, right), and neither did fenpropimorph (Figure 27D, right), tamoxifen (Figure 27F, right) or artemether (Figure 27B, right).
Notably, over-abundance of auxin itself promotes PIN2 degradation in an endosomal cycling,
ubiquitination manner (Abas et al., 2006), which could insinuate an exacerbated auxin perception

47
Figure 26. Stigmasterol supplementation experiment. Arabidopsis PIN2-GFP seedlings were sown and grown
under continuous light in 0.5 MS for 4 days and transferred to medium supplemented with different concentrations of inhibitors and/or 3 µg/ml stigmasterol. Their roots were photographed using confocal laser microscopy 100 hours after transferring. A) DMSO 0.1% without (left) and with (right) stigmasterol, B) Clotrimazole 0.1 without (left) and with (right) stigmasterol, C) Clotrimazole 0.5 µM without (left) and with (right)
stigmasterol, D) Oxiconazole without (left) and with (right) stigmasterol, E) Oxiconazole 0.5 µM without (left) and with (right) stigmasterol, F) Tamoxifen 0.5µM without (left) and with (right) stigmasterol, G) Tamoxifen 1µM without (left) and with (right) stigmasterol, H) Voriconazole 0.1µM without (left) and with (right) stigmasterol, F) Voriconazole 1µM without (left) and with (right) stigmasterol. Scale bar = 100µm.

48
upon β-sitosterol application leading to PIN2 degradation in the mock-treated plants, but this is
of course to be tested further.
Figure 27. β-Sitosterol supplementation experiment . Arabidopsis PIN2-GFP seedlings were sown and grown under continuous light in 0.5 MS for 4 days and transferred to medium supplemented with different concentrations of inhibitors and/or 30mg/ml β-sitosterol. Their roots were photographed using confocal laser microscopy 100 hours after transferring. A) DMSO 0.1% without (left) and with (right) β-sitosterol, B) Artemether
0.5µM without (left) and with (right) β-sitosterol, C) Clotrimazole 0.5µM without (left) and with (right) β-sitosterol, D) Fenpropimorph 0.5 µM without (left) and with (right) β-sitosterol, E) Oxiconazole 0.5 µM without (left) and with (right) β-sitosterol, F) Tamoxifen 1µM without (left) and with (right) β-sitosterol, G) Voriconazole 0.5µM without (left) and with (right) β-sitosterol. Scale bar = 100µm.

49
Stigmasterol and sitosterol were not capable of rescuing the defects caused by our inhibitors,
even if they are two of the most abundant phytosterols and important end-products of the
pathway, leading to think that the defects on the roots are caused by the lacking of other kind of
intermediates of because of the overall sterol imbalance caused by the compound.
3.6.4 Effects of the candidate inhibitors in PIN endocytic recycling
We found that the candidate inhibitors produced effects on cell morphology and division patterns
that resembled the ones presented by smt2smt3 mutants and the ones caused by the known
sterol inhibitor fenpropimorph (Nakamoto et al., 2015). However, the inhibitors did not cause
obvious a loss of PIN2-GFP polarity. This in in contrast to reported PIN2 polarity defects and
reduced endocytic rates in the sterol biosynthesis mutant cpi1-1 (Men et al., 2008). Importantly,
other sterol biosynthesis mutants smt2smt3, fk and fenpropimorph treatments do not display PIN
endocytosis defects from the plasma membrane, but impair auxin inhibition of PIN2 endocytosis,
which was suggested to reflect an enhanced trafficking to the vacuole for degradation (Abas et
al., 2006, Men et al., 2008, Nakamoto et al., 2015, Paciorek et al., 2005, Pan et al., 2009).
Therefore, we aimed to explore the short-term effects of the putative inhibitors on PIN2 endocytic dynamics.
The experiment was set up as follows: 4 day old PIN2-GFP seedlings were transferred to liquid
0.5MS medium containing the candidate inhibitor (10 µM for artemether, voriconazole,
clotrimazole and oxiconazole, 20 µM for fenpropimorph and tamoxifen) for an hour pre-
treatment, followed by another hour treatment by soaking in liquid 0.5MS medium that included
that same compound in the same concentration (Figure 28), 25µg/ml BFA (Figure 29) and/or 10µM NAA (Figure 30).
Pre-treatment with the compounds caused the roots to showed increased intracellular PIN2-GFP
positive signals (Figure 28, insets) especially after treatment with clotrimazole, tamoxifen,
voriconazole and fenpropimorph (Fig. 28 B, F, G, D insets). These are seen as small fluorescent
dots overall the cell, suggesting increased endocytic PIN2-GFP dynamics after inhibitor treatment.
As expected, BFA treatment resulted in big, circular intracellular PIN2-GFP positive signals, in so-
called BFA bodies (Figure 29). Treatment with any of the inhibitors did not inhibit PIN2-GFP
accumulation in BFA bodies, suggesting that they do not inhibit PIN2 internalization (Figure 29B-
I, arrowheads point to BFA bodies). Under tamoxifen treatment, the PIN2-GFP positive signals
appeared to be irregularly shaped and diffuse and ultimately not so well aggregated into circular
BFA bodies, being also positive signals outside of the BFA bodies themselves (Fig. 29F) compared
to the roundness and definition of for example, BFA bodies under treatment with voriconazole
and clotrimazole (Figure 29 G, I). When roots were cotreated with BFA and NAA and DMSO
(Figure 30 A), PIN2-GFP no longer accumulated in BFA bodies due to impaired endocytosis, as
previously reported (Paciorek et al., 2005, Robert et al., 2010). In combination with

50
fenpropimorph treatment, some PIN2-GFP signal in BFA bodies could be seen in presence of NAA
(Figure 30B, arrowheads). Similarly, co-treatments with artemether, clotrimazole, oxiconazole,
voriconazole and tamoxifen led to different degrees of PIN2-GFP accumulation in BFA bodies in
the presence of auxin (Figure 30C-I).
Together, these results show that all tested compounds display effects on PIN2-GFP trafficking,
albeit to different efficiencies, that are also seen in sterol biosynthesis mutants fk, smt2 and
smt2smt3 (Carland et al., 2010, Nakamoto et al., 2015, Pan et al., 2009). They promote PIN2-GFP
internalization even in the presence of
auxin. The additional PIN2-GFP
signals outside the regular BFA
bodies, seen upon tamoxifen
treatment, reflect the presence of
PIN2-GFP in BFA insensitive
endocytic compartments, such as
late endosomes (Grebe et al., 2003)
suggesting that tamoxifen is highly
efficient in stimulating PIN2 degradation.
Figure 28. PIN recycling under sterol inhibitor compounds and auxin
supplementation. Part 1: compound pretreatment. Arabidopsis PIN2-GFP seedlings were sown and grown under continuous light in 0.5 MS for 4 days and
transferred to l iquid MS medium supplemented with different concentrations of inhibitors for an hour. After the pretreatment, they were
transferred to l iquid MS medium containing the same concentration of inhibitor. Their roots were imaged using
confocal laser microscopy. A) DMSO 0.1%, B) Fenpropimorph 20µM, C) Artemether 10µM, D) Clotrimazole 10µM, E) Oxiconazole 10µM, F)
Tamoxifen 20µM, G) Voriconazole 10µM. Insets detail vacuolar signals. Scale bar= 100µm.

51
Figure 29. PIN recycling under sterol inhibitor compounds and auxin supplementation. Part 2: compound
pretreatment plus BFA treatment. Arabidopsis PIN2-GFP seedlings were sown and grown under continuous light
in 0.5 MS for 4 days and transferred to l iquid MS medium supplemented with different concentrations of inhibitors for an hour. After the pretreatment, they
were transferred to l iquid MS medium containing the same concentration of inhibitor plus 25µg/ml BFA. Their roots were imaged using confocal laser
microscopy. A) DMSO 0.1%, B) Fenpropimorph 20µM, C) Artemether 10µM, D) Clotrimazole 10µM, E)
Oxiconazole 10µM, F) Tamoxifen 20µM, G) Voriconazole 10µM. Arrowheads point to BFA bodies. Scale bar= 100µm.

52
Figure 30. PIN recycling under sterol
inhibitor compounds and auxin supplementation. Part 3: compound pretreatment plus BFA treatment +NAA treatment. Arabidopsis PIN2-GFP
seedlings were sown and grown under continuous light in 0.5 MS for 4 days and transferred to l iquid MS medium supplemented with different
concentrations of inhibitors for an hour. After the pretreatment, they were transferred to l iquid MS medium
containing the same concentration of inhibitor plus 25µg/ml BFA and 10µM NAA. Their roots were imaged using confocal laser microscopy. A) DMSO
0.1%, B) Fenpropimorph 20µM, C) Artemether 10µM, D) Clotrimazole 10µM, E) Oxiconazole 10µM, F)
Tamoxifen 20µM, G) Voriconazole 10µM. Arrowheads point to BFA bodies. Scale bar= 100µm.

53
Part 4. Discussion
4.1 Identification of novel plant sterol biosynthesis inhibitors
Sterol biosynthesis inhibitors, unlike the mutants, have the benefit to allow dissecting the role of
sterols in plants in a temporary, switchable manner, that opens the possibil ity to combine sterol
deprivation and other treatments and conditions immediately or whenever it is desired. The use
of sterol biosynthesis inhibitors is the only way to evaluate short term effects of sterol
deprivation. A mutant is permanently under sterol deprivation conditions, and that can cause
pleiotropic developmental defects which interfere and be confused with real effects of the sterol deprivation in a short term.
Only two sterol biosynthesis inhibitors with targets downstream lanosterol synthesis and
therefore acting on the proper sterol biosynthesis pathway are well described for effects in plants,
voriconazole and fenpropimorph. They disrupt the sterol biosynthetic pathway with
developmental and cell morphology impairments as consequences (Figure 31) (He et al., 2003,
Rozhon et al., 2013). A recent chemical screen performed on murine cells that interprets their
sterol profiles after the supplementation with a variety of compounds points to inhibition of
catalyzers along the sterol biosynthetic pathway on several potential targets scattered in several
steps of the synthesis (Korade et al., 2016). Further analysis would then position the action of the
compound against a specific enzymatic action, and the compound could be properly called a
synthesis inhibitor with known consequences. A varied catalog of inhibitors disturbing discrete
biochemical steps with defined characteristics would then reflect various behaviors, and this is
one of the objectives of the work: characterize the compounds and compare their responses to reflect inhibition of sterol biosynthesis at different steps.
Animal and plant sterol biosynthesis pathways share a number of steps but there exist several
conversion steps that are a specific plant feature and that allow plants to have more than one
primary sterol in contrast to animals having only cholesterol. The C-24 transmethylation catalyzed
by SMT2/SMT3 and the last C-22 desaturation catalyzed by CYP710A are steps that are present
in plants (and the desaturation in fungi) but not in animals. The consequence of this is a variety
of end-products such as campesterol, stigmasterol, isofucosterol and sitosterol, and their
influence on the synthesis of sterol-derived secondary metabolites and BR. Cholesterol is,
nevertheless, not absent in plants. They produce it in minute amounts, reflecting the converging
point between both kingdom pathways and the existence of parallel, conserved targets for an
inhibitor compound that affect cholesterol and other sterols synthesis. The divergence in the
pathways would further allow for pathway specificity in targets for the compounds, opening a
possibility to find plant-specific sterol biosynthesis inhibitors that would facilitate the analysis of

54
plant-specific physiology. In the aforementioned chemical screen, oxiconazole, clotrimazole,
fluphenazine, artemether and tamoxifen are among the compounds with potential targets in
mammal cells (Figure 31). For artemether, HMGR, an enzyme that catalyzes the reduction of
HCoA, an early step of the cholesterol biosynthesis pathway. Tamoxifen was found to cause sterol
profiles coincident with a disturbance in a delta-8 isomerase and DHCR24, late steps in cholesterol
synthesis. Fluphenazine was found to coincide with delta-8 isomerase inactivation, and
clotrimazole and oxiconazole, as expected, were found to act on CYP51 (Korade et al., 2016). We
chose fluphenazine, artemether, tamoxifen, clotrimazole and oxiconazole as candidate inhibitors
of sterol biosynthesis. Given that BR and structural sterol biosynthesis pathways majorly overlap,
we screened the selected chemicals for BR deprivation phenotypes, such as a strong inhibition of
hypocotyl elongation. We found that artemether, clotrimazole and oxiconazole were highly
effective in preventing hypocotyl elongation, tamoxifen had a mild response, and fluphenazine
was not able to prevent hypocotyl elongation, so it was discarded from further analysis.
We then tested the compound action against root elongation, and we found that the roots were
severely shrunk due to growth in the presence of all the compounds, including tamoxifen. After
visualizing the root cell organization with the help of PIN2-GFP and ABCB19-GFP, which are
located on the PM, on compound-treated roots we can see that cell division defects occur
Figure 31. Sterol biosynthesis inhibitors targets in plants and mammals. Juxtaposed for comparison, simplified schemes of both biosynthetic pathways of mammals and plants. In plants, a bifurcation of the synthetic pathway gives rise to BR precursors (C24-methylsterols, in green) and C24-ethylsterols (in red). Known inhibitors of sterol
biosynthesis (purple) are shown with equivalent targets for both kingdoms. Potential targets in mammal cells for artemether and tamoxifen as reported by (Korade et al., 2016) are shown in pink. In brown is shown tamoxifen as a suggested target by findings from this work.

55
throughout the root, the cell structure is aberrant, and after various days, the cells are effectively
destroyed by the compound. Further work is necessary to delimit the direct causes of these
cellular effects, but it is known that some sterol biosynthesis mutants present cytokinesis defects
(Nakamoto et al., 2015). It is notable that the cell division orientation is apparently more sensitive
to the inhibitors than the PIN2-GFP turnover, being then PIN2 localization more resilient to sterol
deficiency than structural cell organization dependent on sterol content. This behavior was seen
to be similar in known inhibitors fenpropimorph and voriconazole. Together, these findings
suggest that artemether, clotrimazole, oxiconazole and tamoxifen could qualify as sterol
biosynthesis inhibitors.
Tamoxifen is a special case in which while the BR deficiency response (based on the hypocotyl
phenotype) was very mild, its action on the roots was much stronger. This could suggest that
tamoxifen affects specifically the synthesis of structural sterols that makes the root growth and
structure especially sensitive to its supplementation. Two options are specific enzymatic steps for
structural sterols: SMT2/3 and CYP710A. Our mutant analysis shows cvp1 , but not cyp710a1 or
cyp710a2, hypersensitivity to tamoxifen, suggesting SMT2/3 as possible targets of tamoxifen.
In summary, our selection of chemicals are possibly highly potent inhibitors of sterol biosynthesis
in plants, but to validate the activity of our candidate inhibitors, we need a confirmed target. This
can be approached by obtaining the sterol profile of plants treated with the compounds via
biochemical analyses such as GC-MS, which will tell us exactly the proportions of each of the
intermediaries that are present in the plant and therefore where is the missing link. That could
be confirmed by doing the same assays in mutants, treated and untreated, and comparing their
sterol profiles. This would expand the toolbox to explore the functional importance of sterol
biosynthesis in cell biological processes such and endocytic trafficking, and the interplay with auxin signaling.
4.2 Sterol biosynthesis as a possible target for control over PIN2 homeostasis
PIN auxin efflux carriers are localized in the cell depending on their hydrophilic regions. PIN1-type
are localized on the PM and determine the direction of auxin flow (Wisniewska et al., 2006). Their
polarity depends on the cell type and PIN-specific cues such as developmental signals and
hormones translate in dynamic adjustments in directional vesicle trafficking of the PIN proteins.
For example, developmental triggers in embryonic cells control PIN1 polarizing for root definition,
PIN3 and PIN7 are rapidly polarized in response to gravity in columella cells (and subsequently
trigger auxin-dependent differential growth responses)(Friml et al., 2002a, Kleine-Vehn et al.,
2010, Kleine-Vehn & Friml, 2008). PIN1 and PIN7 expression respond differentially to exogenous
cytokinin (Simaskova et al., 2015), high concentrations of methyl jasmonate (MeJA) reduce PIN2
at the plasma membrane whereas low concentrations of MeJA inhibits PIN2 endocytosis (Sun et

56
al., 2011), and it has been demonstrated that gibberellic acid perception decreases PIN2 by
targeting it to the vacuole for lytic degradation, with possible implication in gravitropic auxin response in the root (Willige et al., 2011).
Auxin itself, at long term high concentrations, promotes PIN degradation through vacuolar
trafficking, which surprisingly occurs at auxin suboptimal levels as well (Baster et al., 2013). This
concurs with the auxin-response impaired mutant axr1-12 showing BFA bodies in presence of BFA
and NAA, pointing to internalization and further degradation of PIN without an adequate auxin
signaling (Pan et al., 2009). These altered PIN dynamics were also correlated with lower levels of
β-sitosterol, stigmasterol and campesterol, suggesting an auxin regulation of sterol biosynthesis.
Pan et al. (2009) raised the possibility of sterol biosynthesis as an important target for auxin
signaling as seen by the incapacity of NAA to inhibit PIN PM trafficking in presence of BFA and the
sterol biosynthesis inhibitor fenpropimorph. Moreover, the sterol biosynthesis mutant
smt2(cvp1-3) showed the same response to NAA and BFA, further linking auxin signaling and a
proper sterol content (Pan et al., 2009). A ribosomal unit mutant, rpl4d, that is defective in lipid
synthesis, auxin regulated development and vacuolar trafficking (aberrantly secreting a Carboxy
Terminal Propeptide (CPP)-red fluorescent protein (RFP) fusion that should be transported to the
vacuole), is rescued from its developmental and trafficking defects when complemented with
SMT2 overexpression. Interestingly, auxin signaling mutant tir1 vacuolar trafficking defects were
also partially restored by SMT2 overexpression (Li et al., 2015).
We found that exogenous application of stigmasterol did not alter PIN2 trafficking, while β-
sitosterol dramatically increased PIN2 degradation. This highlights β-sitosterol homeostasis as a
target to modulate PIN2 levels. Consistently, exogenous treatment with bacterial epitopes were
reported to trigger CYP710A activity and transcription in Arabidopsis (as fast as 4 hours for
transcription and 24 hours for activity) (Griebel & Zeier, 2010) demonstrating CYP710A activity
to be an in vivo functionally relevant target for regulation of beta-sitosterol levels.
However, total β-sitosterol levels cannot account for the observed PIN2 degradation as we found
that some combinations with our putative inhibitors could stabilize PIN2 degradation. This
suggests that the β-sitosterol-induced PIN2 degradation likely requires additional sterol
biosynthesis intermediates. To better understand this puzzling observation, it will be imperative
to determine the effect of each of the inhibitors on sterol profiles. Overexpression of CYP710A1
in terms of PIN degradation would be also interesting, as it is the enzyme that mediates the
conversion from β-sitosterol to stigmasterol, along with the crossing of the cyp710a mutants with
PIN2-GFP carrying lines when confronted with both end-sterols to determine if the degradation
of PIN relies solely on β-sitosterol conversion to stigmasterol.
When treated with diverse concentrations of exogenous 2,4-D, the root length of both Col-0 and
the sterol mutants is gradually decreased due to its action as a promoter of cell division instead
of cell elongation. We found that auxin treatment gave Col-0 resistance to hypocotyl shrinkage
when treated with artemether, but not to cvp1, which remained sensitive, possibly indicating

57
gene expresssion of sterol biosynthesis enzymes promoted by auxin that somehow counteracts
the effects of artemether, but that due to the absence of cvp1 or the scarcity of any of the sterols
downstream SMT2 (even with the presence of its homologous SMT3) is ineffective. Tamoxifen
treatment gave both Col-0 and cvp1 resistance to hypocotyl shrinkage caused by auxin
supplementation, but it gave no resistance to root reduction caused by auxin. In fact, the root
hypersensitivity of cvp1 to tamoxifen was present in both auxin treated and non-treated cvp1.
Tamoxifen could possibly target cvp1 in such a drastic way, quenching the action of SMT2 and
possibly SMT3, that auxin gene activation cannot rescue it, and that is translated in hypocotyls as
an impossibility of auxin to activate genes that directly affect SMT2 and 3 in a way that reduces
cell elongation. Gene overexpression of sterol biosynthesis enzymes gave us divergent
information, notably with STE1 overexpressing plants being hypersensitive to the compounds .
This could be due to an artefact caused by the unnatural expression of the gene, but the absence
of obvious phenotypes suggests either that these steps are not rate-limiting for the balancing
effect.
To further address how overexpressing sterol enzymes would affect the behavior of the plant
against our inhibitors and auxins, we had already cloned three genes that delimit the ethyl -sterol
branch of the sterol pathway: CVP1/SMT2, SMO1-1 and CYP710A1. We fused them to fluorescent
tags and diverse promoters in order to generate overexpressing plants (Addendum). This is
especially interesting for the CVP1 overexpressing plant, for being confronted with tamoxifen to
find a possible resistance would strengthen the idea of a target. Further analyses that we have
already begun to prepare, is the determining of expression levels of sterol biosynthesis related
genes upon auxin treatment, while under the action of our inhibitors or without it, to unravel the
ways auxin signaling interacts with the sterol pathway when it is disrupted in order to cope with
the stress of sterol depletion. We have already designer primers for genes along the pathway and
the assay can be done in any moment (Addendum).
Together, we hypothesize that the finely regulated balance of different sterol intermediates
explains how so many stimuli converge on PIN2 turnover. By simple mis -regulation of the
biosynthetic enzyme activity, the sterol balance could be modified causing PIN2 turnover.
4.3 Sterol defect-induced PIN degradation possibly reflects activation of ethylene-dependent
trafficking
In the above paragraphs we argued that the balance of different sterol intermediates controls PIN
turnover. How this works remains to be determined. An alternative hypothesis could be raised
that all inhibitor treatments that induce PIN2 turnover simply reflect enhanced ethylene
biosynthesis, and thus ethylene triggered PIN2 degradation. The function of structural sterols is
mainly at the PM for membrane microdomains/lipid order. Filipin based visualization of β-

58
hydroxysterols (cholesterol, β-sitosterol, campesterol and stigmasterol) shows presence of
sterols in BFA sensitive endosomes (Grebe et al., 2003). The localization of sterol biosynthesis
enzymes would also give an idea of sterol profile modifications being only controlled at the ER or
also in the PM to allow for the membrane microdomains formation in a rapid way. In the case all
enzymes are in the ER, the ratio of different sterols in the PM will then depend on de novo
synthesis in the ER and delivery of the end-product to the secretory pathway, and further passive
turnover, or targeted turnover via modifications such as glycosylation, which occurs at the PM (DeBolt et al., 2009).
Characterization of different sterol biosynthesis mutants has revealed reduced PIN2 levels
(Carland et al., 2010, Men et al., 2008, Nakamoto et al., 2015, Souter et al., 2002, Zhang & Li,
2016) similarly to our inhibitor treatments. Interestingly, the PIN2-GFP degradation in hydra1
was reported to be largely reverted by inhibition of ethylene signaling via AgNO3. There is no
rescue when ethylene biosynthesis is inhibited, though. In addition, pin3, fk and hydra1 show
columella identity loss that suggests that these defects may be in part due to a loss of PIN3
localization and activity and ethylene could be regulating PIN3 response through controlling
cytoskeleton reorganization as PIN trafficking depends on microtubules integrity and localization
(Souter et al., 2004). smt2smt3 mutants also show microtubule defects and microtubule -
associated protein CLASP defects (Nakamoto et al., 2015) , both involved in vesicle trafficking and
more specifically, involved in PIN2 recycling (Ambrose et al., 2013). Therefore, it will be
interesting to analyze ethylene dependence of PIN2-GFP turnover after inhibitor treatment.
Notably, no reports could be found for plant sterol biosynthesis enzymes residing outside of the
ER. Thus interference with their activity likely results in a massive accumulation of sterol
biosynthetic intermediates in the ER, which possibly causes ER stress which in turn triggers
ethylene biosynthesis (Ref for link ER Stress and ethylene biosynthesis). Recently, ethylene was
shown to specifically modify the activity of ARF-GEF BIG1-4 at the TGN and thus endosomal
trafficking at the PM. BIG1-4 mediates AUX1 secretion to the PM from the TGN (Jonsson et al., 2017).
4.4 Calcium supplementation experiments
Trying to link calcium signaling and sterol biosynthesis, and based on a report that showed the
hypersensitivity to calcium ions of a sterol mutant (smt1) (Diener et al., 2000), we tested the
sensitivity to calcium of our sterol mutants. We replicated their experiment, to find that the root
elongation supposedly caused by calcium depletion (in this case achieved by medium dilution)
was shown not only by the mutants but also by the wild type, with all plant lines tested having a
significantly longer root when grown on diluted medium than the ones grown in normal 0.5MS
medium. To make sure that calcium depletion was the reason of the elongation, we grew the

59
mutants and wild-type on calcium-free medium, to discover that the length of the root was in fact
shortened instead of lengthened.
A calcium supplementation experiment was then carried out to know if there was a
concentration of calcium in the medium that could change the behavior of the root from
shortening to lengthening. We discovered that no concentration of calcium in the medium was
able to cause the root lengthening observed with diluted medium, and therefore, the
aforementioned root length rescue of the smt1 mutant achieved by medium dilution, could not
be attributable to calcium shortage.

60
Part 5. Materials and Methods
5.1 Plant material
As wild type plants, Columbia ecotype (Col-0) Arabidopsis thaliana plants were used. The
following mutants and transgenic lines were used: the dwf1 mutants were the homozygous offspring of the SALK_006932 T-DNA mutant heterozygous parents as the mutation is
embryonically lethal, and has been reported elsewhere (Du & Poovaiah, 2005). The cyp710a2-1 mutant is the SALK_001175 T-DNA mutant line reported in (Morikawa et al., 2006). The cyp710a1-
1 mutant is the GABI_325E09 T-DNA mutant line (Griebel & Zeier, 2010). The cyp710a2-2 mutant is the GABI_062A09 T-DNA mutant line. The DIM1-YFP, STE1-YFP and DWF5-YFP lines were the
same used elsewhere (Silvestro et al., 2013). The FvCYP51A OE lines were the same reported by (Rozhon et al., 2013). The PIN2-GFP and ABCB19-GFP reporter lines were also previously reported
(Dhonukshe et al., 2008, Xu & Scheres, 2005).
5.2 Media and growing conditions
For the hypocotyl etiolation assays, seeds were sown and placed in square plates with 0.5X
Murashige and Skoog agar medium supplemented with 1% sucrose (w/v) (Murashige & Skoog, 1962). They were placed under vernalization for three days under darkness at 4 degrees Celsius,
then placed for four hours under light to stimulate germination, and then for 8 days under darkness at 21 degrees to stimulate hypocotyl elongation. For the root growth assays and GFP
visualization experiments, the seeds were sown, vernalized and then placed under under
continuous light in a growth chamber at 21 degrees Celsius. They grew on 0.5X Murashige and Skoog agar supplemented with 1% sucrose (w/v).For diluted medium (0.05MS), the procedure
was to use one tenth of the nutrient salt mix and same quantities of sucrose, MES and myo-inositol in the agar. For calcium-free medium, every macronutrient and micronutrient was added
in the normal amounts, except for the 1.5mM CaCl2, which was omitted.
5.3 Solutions of utilized compounds
Every compound was purchased from Sigma-Aldrich. Fenpropimorph, fluphenazine, dicyclomine,
artemether, voriconazole, oxiconazole, clotrimazole and tamoxifen were dissolved in dimethylsulfoxide (DMSO) and 20mM stock solutions were prepared. Stigmasterol was dissolved
in chloroform and a stock solution of 30mg/ml was prepared. Β-sitosterol was dissolved in ethanol
and a stock solution of 30 mg/ml was prepared. 1-NAA and 2,4-D were dissolved in DMSO, 2,4-D
in Ethanol and 100µM stock solutions were prepared. BFA was dissolved in DMSO and a stock solution of 50 mM was prepared.

61
5.4 Root/hypocotyl imaging and measurements
Agar plates containing vertically grown eight-day old seedlings were digitally scanned with a HP
scanner using a resolution of 300 dots per inch (dpi). Root and hypocotyl lengths were digitally
measured with the help of the ImageJ software and a scanned scale and the measurements were exported to Microsoft Excel to be analyzed.
5.5 Confocal laser microscopy
Images of PIN2-GFP and ABCB19-GFP fluorescence were visualized by confocal laser scanning
microscopy, using a Leica TCS SP2 AOBS with upright microscope stand and a HC PL APO CS 20.0x0.70 IMM/COR immersion objective. Images were processed in GIMP and Microsoft Power
Point. Scale was analyzed with Image J using the experiment data provided by the Leica Software.
5.6 Gene amplification and cloning
Using the primers in Table 2, and genomic Col-0 DNA as a template, the coding regions of SMO1-
1, SMT2 and CYP710A1 were amplified using the iProof High fidelity polymerase (Bio-Rad). The amplified product was then identified and separated by electrophoresis in 1% agarose gels. The
amplified fragments were sliced and purified from the gel with the help of the GeneJet Gel Extraction Kit (Thermo Scientific) and subsequently introduced by a BP reaction into a pDONR221
Gateway vector (Thermo scientific) following the instructions of the manufacturer. The resulting
entry clones were then sequenced to verify the correctness of the cloning (sequencing performed by this institute core facility). After propagating these entry clones by a plasmid DNA
minipreparation using the GeneJet Plasmid Miniprep Kit (Thermo Scientific) , we subcloned the corresponding genes via LR reaction with the entry clones, Gateway vector and a variety of
promoters and tags to be fused to the coding regions of our genes of interest (Table 4). These clones were then checked by restriction digest for correct recombination.
5.7 Statistical analysis
For Welch’s two tailed t-test, the analysis was made using Microsoft Excel and GraphPAD. SAS
University Edition for Windows was used to perform the ANOVA n-way analyses.

62
Part 6. References
Abas L, Benjamins R, Malenica N, Paciorek T, Wisniewska J, Moulinier-Anzola JC, Sieberer T, Friml J, Luschnig C
(2006) Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root gravitropism. Nat Cell Biol 8: 249-56
Ambrose C, Ruan Y, Gardiner J, Tamblyn Laura M, Catching A, Kirik V, Marc J, Overall R, Wasteneys Geoffrey O (2013) CLASP Interacts with Sorting Nexin 1 to Link Microtubules and Auxin Transport via PIN2 Recycling in Arabidopsis thaliana. Developmental Cell 24: 649-659
Arvidsson S, Kwasniewski M, Riano-Pachon DM, Mueller-Roeber B (2008) QuantPrime--a flexible tool for reliable high-throughput primer design for quantitative PCR. BMC Bioinformatics 9: 465
Asami T, Min YK, Nagata N, Yamagishi K, Takatsuto S, Fujioka S, Murofushi N, Yamaguchi I, Yoshida S (2000) Characterization of brassinazole, a triazole-type brassinosteroid biosynthesis inhibitor. Plant Physiol 123: 93-100
Attia TZ, Yamashita T, Miyamoto M, Koizumi A, Yasuhara Y, Node J, Erikawa Y, Komiyama Y, Horii C, Yamada M,
Omar MA, Abdelmageed OH, Derayea SM, Uno T (2012) Comparison of cytochrome p450 mediated metabolism of three central nervous system acting drugs. Chem Pharm Bull (Tokyo) 60: 1544-9
Baloch RI, Mercer EI (1987) Inhibition of sterol Δ8 → Δ7-isomerase and Δ14-reductase by fenpropimorph tridemorph and fenpropidin in cell -free enzyme systems from Saccharomyces cerevisiae. Phytochemistry 26: 663-668
Bandyopadhyay A, Blakeslee JJ, Lee OR, Mravec J, Sauer M, Titapiwatanakun B, Makam SN, Bouchard R, Geisler M, Martinoia E, Friml J, Peer WA, Murphy AS (2007) Interactions of PIN and PGP auxin transport mechanisms. Biochemical Society transactions 35: 137-41
Baster P, Robert S, Kleine-Vehn J, Vanneste S, Kania U, Grunewald W, De Rybel B, Beeckman T, Friml J (2013)
SCF(TIR1/AFB)-auxin signalling regulates PIN vacuolar trafficking and auxin fluxes during root gravitropism. EMBO J 32: 260-74
Becher R, Wirsel SG (2012) Fungal cytochrome P450 sterol 14alpha -demethylase (CYP51) and azole resi stance in plant and human pathogens. Appl Microbiol Biotechnol 95: 825-40
Benveniste P (2002) Sterol Metabolism. The Arabidopsis Book: e0004
Blakeslee JJ, Bandyopadhyay A, Lee OR, Mravec J, Titapiwatanakun B, Sauer M, Makam SN, Cheng Y, Bouchard R, Adamec J, Geisler M, Nagashima A, Sakai T, Martinoia E, Friml J, Peer WA, Murphy AS (2007) Interactions among PIN-FORMED and P-glycoprotein auxin transporters in Arabidopsis. Plant Cell 19: 131-47
Boutte Y, Grebe M (2009) Cellular processes relying on sterol function in plants. Curr Opin Plant Biol 12: 705-13

63
Calderon Villalobos LI, Lee S, De Oliveira C, Ivetac A, Brandt W, Armitage L, Sheard LB, Tan X, Parry G, Mao H, Zheng
N, Napier R, Kepinski S, Estelle M (2012) A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin. Nature chemical biology 8: 477-85
Campanoni P, Nick P (2005) Auxin-dependent cell division and cell elongation. 1-Naphthaleneacetic acid and 2,4-dichlorophenoxyacetic acid activate different pathways. Plant Physiol 137: 939-48
Carland F, Fujioka S, Nelson T (2010) The sterol methyltransferases SMT1, SMT2, and SMT3 influence Arabidopsis development through nonbrassinosteroid products. Plant Physiol 153: 741-56
Carland FM, Fujioka S, Takatsuto S, Yoshida S, Nelson T (2002) The identification of CVP1 reveals a role for sterols in vascular patterning. Plant Cell 14: 2045-58
Chen JG, Ullah H, Young JC, Sussman MR, Jones AM (2001) ABP1 is required for organized cell elongation and division in Arabidopsis embryogenesis . Genes Dev 15: 902-11
Chen X, Grandont L, Li H, Hauschild R, Paque S, Abuzeineh A, Rakusova H, Benkova E, Perrot-Rechenmann C, Friml J (2014) Inhibition of cell expansion by rapid ABP1-mediated auxin effect on microtubules. Nature 516: 90-3
Chen X, Naramoto S, Robert S, Tejos R, Lofke C, Lin D, Yang Z, Friml J (2012) ABP1 and ROP6 GTPase signaling regulate clathrin-mediated endocytosis in Arabidopsis roots. Curr Biol 22: 1326-32
Cho M, Cho HT (2013) The function of ABCB transporters in auxin transport. Plant signaling & behavior 8: e22990
Choe S, Dilkes BP, Gregory BD, Ross AS, Yuan H, Noguchi T, Fujioka S, Takatsuto S, Tanaka A, Yoshida S, Tax FE,
Feldmann KA (1999a) The Arabidopsis dwarf1 mutant is defective in the conversion of 24 -methylenecholesterol to campesterol in brassinosteroid biosynthesis. Plant Physiol 119: 897-907
Choe S, Noguchi T, Fujioka S, Takatsuto S, Tissier CP, Gregory BD, Ross AS, Tanaka A, Yoshida S, Tax FE, Feldmann KA (1999b) The Arabidopsis dwf7/ste1 mutant is defective in the delta7 sterol C-5 desaturation step leading to brassinosteroid biosynthesis. Plant Cell 11: 207-21
Choe S, Tanaka A, Noguchi T, Fujioka S, Takatsuto S, Ross AS, Tax FE, Yoshida S, Feldmann KA (2000) Lesions in the sterol delta reductase gene of Arabidopsi s cause dwarfism due to a block in brassinosteroid biosynthesis. Plant J 21: 431-43
Clouse SD (2002) Arabidopsis mutants reveal multiple roles for sterols in plant development. Plant Cell 14: 1995-2000
Clouse SD (2011) Brassinosteroids. The Arabidopsis Book / American Society of Plant Biologists 9: e0151

64
Darnet S, Bard M, Rahier A (2001) Functional identification of sterol -4alpha-methyl oxidase cDNAs from Arabidopsis thaliana by complementation of a yeast erg25 mutant lacking sterol -4alpha-methyl oxidation. FEBS Lett 508: 39-43
Darnet S, Rahier A (2004) Plant sterol biosynthesis: identification of two distinct families of sterol 4alpha-methyl oxidases. Biochem J 378: 889-98
DeBolt S, Scheible W-R, Schrick K, Auer M, Beisson F, Bischoff V, Bouvier-Navé P, Carroll A, Hematy K, Li Y, Milne J, Nair M, Schaller H, Zemla M, Somerville C (2009) Mutations in UDP-Glucose:Sterol Glucosyltransferase in Arabidopsis Cause Transparent Testa Phenotype and Suberization Defect in Seeds. Plant Physiology 151: 78-87
Dharmasiri N, Dharmasiri S, Estelle M (2005) The F-box protein TIR1 is an auxin receptor. Nature 435: 441-5
Dhonukshe P, Aniento F, Hwang I, Robinson DG, Mravec J, Stierhof YD, Friml J (2007) Clathrin -mediated constitutive endocytosis of PIN auxin efflux carri ers in Arabidopsis. Curr Biol 17: 520-7
Dhonukshe P, Grigoriev I, Fischer R, Tominaga M, Robinson DG, Hasek J, Paciorek T, Petrasek J, Seifertova D, Tejos R, Meisel LA, Zazimalova E, Gadella TW, Jr., Stierhof YD, Ueda T, Oiwa K, Akhmanova A, Brock R, Span g A, Friml J
(2008) Auxin transport inhibitors impair vesicle motil ity and actin cytoskeleton dynamics in diverse eukaryotes. Proc Natl Acad Sci U S A 105: 4489-94
Diener AC, Li H, Zhou W, Whoriskey WJ, Nes WD, Fink GR (2000) Sterol methyltransferase 1 controls the level of cholesterol in plants. Plant Cell 12: 853-70
Du L, Poovaiah BW (2005) Ca2+/calmodulin is critical for brassinosteroid biosynthesis and plant growth. Nature 437: 741-5
Friml J, Benkova E, Bli lou I, Wisniewska J, Hamann T, Ljung K, Woody S, Sandberg G, Scheres B, Jurgens G, Palme K (2002a) AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 108: 661-73
Friml J, Wisniewska J, Benkova E, Mendgen K, Palme K (2002b) Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 415: 806-9
Fu Y, Gu Y, Zheng Z, Wasteneys G, Yang Z (2005) Arabidopsis interdigitating cell growth requires two antagonistic pathways with opposing action on cell morphogenesis. Cell 120: 687-700
Fu Y, Xu T, Zhu L, Wen M, Yang Z (2009) A ROP GTPase Signaling Pathway Controls Cortical Microtubule Ordering and Cell Expansion in Arabidopsis. Current Biology 19: 1827-1832
Gachotte D, Husselstein T, Bard M, Lacroute F, Benveniste P (1996) Isolation and character ization of an Arabidopsis
thaliana cDNA encoding a delta 7-sterol-C-5-desaturase by functional complementation of a defective yeast mutant. Plant J 9: 391-8

65
Gadeyne A, Sanchez-Rodriguez C, Vanneste S, Di Rubbo S, Zauber H, Vanneste K, Van Leene J, De Winne N,
Eeckhout D, Persiau G, Van De Slijke E, Cannoot B, Vercruysse L, Mayers JR, Adamowski M, Kania U, Ehrlich M, Schweighofer A, Ketelaar T, Maere S et al. (2014) The TPLATE adaptor complex drives clathrin-mediated endocytosis in plants. Cell 156: 691-704
Gao Y, Zhang Y, Zhang D, Dai X, Estelle M, Zhao Y (2015) Auxin binding protein 1 (ABP1) is not required for either auxin signaling or Arabidopsis development. Proc Natl Acad Sci U S A 112: 2275-80
Geisler M, Blakeslee JJ, Bouchard R, Lee OR, Vincenzetti V, Bandyopadhyay A, Titapiwatanakun B, Peer WA, Bailly A,
Richards EL, Ejendal KF, Smith AP, Baroux C, Grossniklaus U, Muller A, Hrycyna CA, Dudler R, Murphy AS, Martinoia E (2005) Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1. Plant J 44: 179-94
Geisler M, Murphy AS (2006) The ABC of auxin transport: the role of p-glycoproteins in plant development. FEBS Lett 580: 1094-102
Geldner N, Anders N, Wolters H, Keicher J, Kornberger W, Muller P, Delbarre A, Ueda T, Nakano A, Jurgens G (2003) The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth. Cell 112: 219-30
Geldner N, Friml J, Stierhof YD, Jurgens G, Palme K (2001) Auxin transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 413: 425-8
Geldner N, Richter S, Vieten A, Marquardt S, Torres -Ruiz RA, Mayer U, Jurgens G (2004) Partial loss-of-function
alleles reveal a role for GNOM in auxin transport-related, post-embryonic development of Arabidopsis. Development 131: 389-400
Grebe M, Xu J, Mobius W, Ueda T, Nakano A, Geuze HJ, Rook MB, Scheres B (2003) Arabidopsis sterol endocytosis involves actin-mediated trafficking via ARA6-positive early endosomes. Curr Biol 13: 1378-87
Griebel T, Zeier J (2010) A role for beta-sitosterol to stigmasterol conversion in plant-pathogen interactions. Plant J 63: 254-68
Grogan J, Shou M, Zhou D, Chen S, Korzekwa KR (1993) Use of aromatase (CYP19) metabolite ratios to characterize electron transfer from NADPH-cytochrome P450 reductase. Biochemistry 32: 12007-12
Grones P, Chen X, Simon S, Kaufmann WA, De Rycke R, Nodzynski T, Zazimalova E, Friml J (2015) Auxin -binding pocket of ABP1 is crucial for its gain-of-function cellular and developmental roles. J Exp Bot 66: 5055-65
Hartmann M-A (2004) 5 Sterol metabolism and functions in higher plants. In Lipid Metabolism and Membrane Biogenesis, Daum G (ed) pp 183-211. Berlin, Heidelberg: Springer Berlin Heidelberg

66
Haynes RK, Cheu KW, Chan HW, Wong HN, Li KY, Tang MM, Chen MJ, Guo ZF, Guo ZH, Sinniah K, Witte AB, Coghi P,
Monti D (2012) Interactions between artemisinins and other antimalarial drugs in relation to the cofactor model --a unifying proposal for drug action. ChemMedChem 7: 2204-26
He JX, Fujioka S, Li TC, Kang SG, Seto H, Takatsuto S, Yoshida S, Jang JC (2003) Sterols regulate development and gene expression in Arabidopsis. Plant Physiol 131: 1258-69
Herman GE (2003) Disorders of cholesterol biosynthesis: prototypic metabolic malformation syndromes. Hum Mol Genet 12 Spec No 1: R75-88
Himschoot E, Beeckman T, Friml J, Vanneste S (2015) Calcium is an organizer of cell polarity in plants. Biochimica et biophysica acta 1853: 2168-72
Holstein SE (2002) Clathrin and plant endocytosis. Traffic 3: 614-20
Ischebeck T, Werner S, Krishnamoorthy P, Lerche J, Meijon M, Stenzel I, Lofke C, Wiessner T, Im YJ, Perera IY, Iven T, Feussner I, Busch W, Boss WF, Teichmann T, Hause B, Persson S, Heilmann I (2013) Phosphatidylinositol 4,5 -
bisphosphate influences PIN polarization by controll ing clathrin-mediated membrane trafficking in Arabidopsis. Plant Cell 25: 4894-911
Ito E, Fujimoto M, Ebine K, Uemura T, Ueda T, Nakano A (2012) Dynamic behavior of clathrin in Arabidopsis thaliana unveiled by l ive imaging. Plant J 69: 204-16
Jail lais Y, Gaude T (2008) Plant cell polarity: sterols enter into action after cytokinesis. Dev Cell 14: 318-20
Jail lais Y, Santambrogio M, Rozier F, Fobis -Loisy I, Miege C, Gaude T (2007) The retromer protein VPS29 links cell polarity and organ initiation in plants. Cell 130: 1057-70
Jang JC, Fujioka S, Tasaka M, Seto H, Takatsuto S, Ishii A, Aida M, Yoshida S, Sheen J (2000) A critical role of sterols in embryonic patterning and meristem programming revealed by the fackel mutants of Arabidopsis thaliana. Genes Dev 14: 1485-97
Jonsson K, Boutté Y, Singh RK, Gendre D, Bhalerao R (2017) Ethylene Regulates differential growth via BIG ARF -GEF-dependent post-Golgi secretory trafficking in Arabidopsis. Plant Cell
Kelly BT, Owen DJ (2011) Endocytic sorting of transmembrane protein cargo. Curr Opin Cell Biol 23: 404-12
Kepinski S, Leyser O (2005) The Arabidopsis F-box protein TIR1 is an auxin receptor. Nature 435: 446-51
Kerkenaar A (1990) Inhibition of the sterol delta 14-reductase and delta 8----delta 7-isomerase in fungi. Biochemical Society transactions 18: 59-61

67
Kim HB, Schaller H, Goh CH, Kwon M, Choe S, An CS, Durst F, Feldmann KA, Feyereisen R (2005) Arabidopsis cyp51
mutant shows postembryonic seedling lethality associated with lack of membrane integrity. Plant Physiol 138: 2033-47
Kim S, Thiessen PA, Bolton EE, Chen J, Fu G, Gindulyte A, Han L, He J, He S, Shoemaker BA, Wang J, Yu B, Zhang J, Bryant SH (2016) PubChem Substance and Compound databases. Nucleic Acids Res 44: D1202-13
Kitakura S, Vanneste S, Robert S, Lofke C, Teichmann T, Tanaka H, Friml J (2011) Clathrin mediates endocytosis and polar distribution of PIN auxin transporters in Arabidopsis. Plant Cell 23: 1920-31
Kleine-Vehn J, Ding Z, Jones AR, Tasaka M, Morita MT, Friml J (2010) Gravity-induced PIN transcytosis for polarization of auxin fluxes in gravity-sensing root cells. Proceedings of the National Academy of Sciences 107: 22344-22349
Kleine-Vehn J, Friml J (2008) Polar targeting and endocytic recycling in auxin-dependent plant development. Annu Rev Cell Dev Biol 24: 447-73
Kleine-Vehn J, Leitner J, Zwiewka M, Sauer M, Abas L, Luschnig C, Friml J (2008) Differential degradation of PIN2 auxin efflux carrier by retromer-dependent vacuolar targeting. Proc Natl Acad Sci U S A 105: 17812-7
Korade Z, Kim HY, Tallman KA, Liu W, Koczok K, Balogh I, Xu L, Mirnics K, Porter NA (2016) The Effect of Small Molecules on Sterol Homeostasis: Measuring 7-Dehydrocholesterol in Dhcr7-Deficient Neuro2a Cells and Human Fibroblasts. Journal of medicinal chemistry 59: 1102-15
Krecek P, Skupa P, Libus J, Naramoto S, Tejos R, Friml J, Zazimalova E (2009) The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol 10: 249
Kushiro M, Nakano T, Sato K, Yamagishi K, Asami T, Nakano A, Takatsuto S, Fujioka S, Ebizuka Y, Yoshida S (2001) Obtusifoliol 14alpha-demethylase (CYP51) antisense Arabidopsis shows slow growth and long life. Biochemical and biophysical research communications 285: 98-104
Lam BC, Sage TL, Bianchi F, Blumwald E (2001) Role of SH3 domain-containing proteins in clathrin-mediated vesicle trafficking in Arabidopsis. Plant Cell 13: 2499-512
Lavy M, Estelle M (2016) Mechanisms of auxin signaling. Development 143: 3226-9
Lee S, Sundaram S, Armitage L, Evans JP, Hawkes T, Kepinski S, Ferro N, Napier RM (2014) Defining binding efficiency and specificity of auxins for SCF(TIR1/AFB)-Aux/IAA co-receptor complex formation. ACS Chem Biol 9: 673-82

68
Lepesheva GI, Waterman MR (2007) Sterol 14alpha -demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochimica et biophysica acta 1770: 467-77
Li R, Sun R, Hicks GR, Raikhel NV (2015) Arabidopsis ribosomal proteins control vacuole trafficking and developmental programs through the regulation of l ipid metabolism. Proc Natl Acad Sci U S A 112: E89-98
McMahon HT, Boucrot E (2011) Molecular mechanism and physiological functions of clathrin -mediated endocytosis. Nature reviews Molecular cell biology 12: 517-33
Men S, Boutte Y, Ikeda Y, Li X, Palme K, Stierhof YD, Hartmann MA, Moritz T, Grebe M (2008) Sterol-dependent endocytosis mediates post-cytokinetic acquisition of PIN2 auxin efflux carrier polarity. Nat Cell Biol 10: 237-44
Mercer EI (1991) Morpholine antifungals and their mode of action. Biochemical Society transactions 19: 788-93
Monshausen GB, Miller ND, Murphy AS, Gilroy S (2011) Dynamics of auxin-dependent Ca2+ and pH signaling in root growth revealed by integrating high-resolution imaging with automated computer vision-based analysis. Plant J 65: 309-18
Moreau P, Hartmann MA, Perret AM, Sturbois-Balcerzak B, Cassagne C (1998) Transport of sterols to the plasma membrane of leek seedlings. Plant Physiol 117: 931-7
Morikawa T, Mizutani M, Aoki N, Watanabe B, Saga H, Saito S, Oikawa A, Suzuki H, Sakurai N, Shibata D, Wadano A,
Sakata K, Ohta D (2006) Cytochrome P450 CYP710A encodes the sterol C-22 desaturase in Arabidopsis and tomato. Plant Cell 18: 1008-22
Murashige T, Skoog F (1962) A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiologia plantarum 15: 473-497
Nakamoto M, Schmit AC, Heintz D, Schaller H, Ohta D (2015) Diversification of sterol methyltransferase enzymes in plants and a role for beta-sitosterol in oriented cell plate formation and polarized growth. Plant J 84: 860-74
Naramoto S, Kleine-Vehn J, Robert S, Fujimoto M, Dainobu T, Paciorek T, Ueda T, Nakano A, Van Montagu MC, Fukuda H, Friml J (2010) ADP-ribosylation factor machinery mediates endocytosis in plant cells. Proc Natl Acad Sci U S A 107: 21890-5
Naramoto S, Otegui MS, Kutsuna N, de Rycke R, Dainobu T, Karampelias M, Fujimoto M, Feraru E, Miki D, Fukuda H,
Nakano A, Friml J (2014) Insights into the localization and function of the membrane trafficking regulator GNOM ARF-GEF at the Golgi apparatus in Arabidopsis. Plant Cell 26: 3062-76
Ohyama K, Suzuki M, Kikuchi J, Saito K, Muranaka T (2009) Dual biosynthetic pathways to phytosterol via cycloartenol and lanosterol in Arabidopsis. Proc Natl Acad Sci U S A 106: 725-30

69
Overvoorde P, Fukaki H, Beeckman T (2010) Auxin control of root development. Cold Spring Harb Perspect Biol 2: a001537
Paciorek T, Zazimalova E, Ruthardt N, Petrasek J, Stierhof YD, Kleine-Vehn J, Morris DA, Emans N, Jurgens G, Geldner N, Friml J (2005) Auxin inhibits endocytosis and promotes its own efflux from cells. Nature 435: 1251-6
Paez Valencia J, Goodman K, Otegui MS (2016) Endocytosis and Endosomal Trafficking in Plants. Annu Rev Plant Biol 67: 309-35
Pan J, Fujioka S, Peng J, Chen J, Li G, Chen R (2009) The E3 ubiquitin l igase SCFTIR1/AFB and membrane sterols play key roles in auxin regulation of endocytosis, recycling, and plasma membrane accumulation of the auxin efflux transporter PIN2 in Arabidopsis thaliana. Plant Cell 21: 568-80
Petrasek J, Friml J (2009) Auxin transport routes in plant development. Development 136: 2675-88
Petrasek J, Mravec J, Bouchard R, Blakeslee JJ, Abas M, Seifertova D, Wisniewska J, Tadele Z, Kubes M, Covanova M, Dhonukshe P, Skupa P, Benkova E, Perry L, Krecek P, Lee OR, Fink GR, Geisler M, Murphy AS, Luschnig C et al. (200 6) PIN proteins perform a rate-limiting function in cellular auxin efflux. Science 312: 914-8
Price CL, Parker JE, Warrilow AG, Kelly DE, Kelly SL (2015) Azole fungicides - understanding resistance mechanisms in agricultural fungal pathogens. Pest Manag Sci 71: 1054-8
Pullen M, Clark N, Zarinkamar F, Topping J, Lindsey K (2010) Analysis of vascular development in the hydra sterol biosynthetic mutants of Arabidopsis. PloS one 5: e12227
Raven J (1975) Transport of indoleacetic acid in plant cells in relati on to pH and electrical potential gradients, and its significance for polar IAA transport. New Phytologist 74: 163-172
Robert S, Kleine-Vehn J, Barbez E, Sauer M, Paciorek T, Baster P, Vanneste S, Zhang J, Simon S, Covanova M, Hayashi K, Dhonukshe P, Yang Z, Bednarek SY, Jones AM, Luschnig C, Aniento F, Zazimalova E, Friml J (2010) ABP1 mediates auxin inhibition of clathrin-dependent endocytosis in Arabidopsis. Cell 143: 111-21
Rozhon W, Husar S, Kalaivanan F, Khan M, Idlhammer M, Shumilina D, Lange T, Hoffmann T, Schwab W, Fujioka S,
Poppenberger B (2013) Genetic variation in plant CYP51s confers resistance against voriconazole, a novel inhibitor of brassinosteroid-dependent sterol biosynthesis. PloS one 8: e53650
Ruegger M, Dewey E, Gray WM, Hobbie L, Turner J, Estelle M (1998) The TIR1 protein of Arabidopsis functions in auxin response and is related to human SKP2 and yeast grr1p. Genes Dev 12: 198-207
Salehin M, Bagchi R, Estelle M (2015) SCFTIR1/AFB-based auxin perception: mechanism and role in plant growth and development. Plant Cell 27: 9-19

70
Sauer M, Balla J, Luschnig C, Wisniewska J, Reinohl V, Friml J, Benkova E (2006) Canalization of auxin flow by Aux/IAA-ARF-dependent feedback regulation of PIN polarity. Genes Dev 20: 2902-11
Schaller H (2003) The role of sterols in plant growth and development. Prog Lipid Res 42: 163-75
Schaller H (2004) New aspects of sterol biosynthesis in growth and development of higher plants. Plant physiology and biochemistry : PPB / Societe francaise de physiologie vegetale 42: 465-76
Schrick K, Mayer U, Horrichs A, Kuhnt C, Bellini C, Dangl J, Schmidt J, Jurgens G (2000) FACKEL is a sterol C -14 reductase required for organized cell division and expansion in Arabidopsis embryogenesis. Genes Dev 14: 1471-84
Shiau AK, Barstad D, Loria PM, Cheng L, Kushner PJ, Agard DA, Greene GL (1998) The structural basis of estrogen receptor/coactivator recognition and the antagonism of this interaction by tamoxifen. Cell 95: 927-37
Silvestro D, Andersen TG, Schaller H, Jensen PE (2013) Plant sterol metabolism. Delta(7)-Sterol-C5-desaturase (STE1/DWARF7), Delta(5,7)-sterol-Delta(7)-reductase (DWARF5) and Delta(24)-sterol-Delta(24)-reductase (DIMINUTO/DWARF1) show multiple subcellular localizations in Arabidopsis thaliana (Heynh) L. PloS one 8: e56429
Simaskova M, O'Brien JA, Khan M, Van Noorden G, Otvos K, Vieten A, De Clercq I, Van Haperen JM, Cuesta C, Hoyerova K, Vanneste S, Marhavy P, Wabnik K, Van Breusegem F, Nowack M, Murphy A, Friml J, Weijers D,
Beeckman T, Benkova E (2015) Cytokinin response factors regulate PIN-FORMED auxin transporters. Nature communications 6: 8717
Simon S, Petrasek J (2011) Why plants need more than one type of auxin. Plant science : an international journal of experimental plant biology 180: 454-60
Sonawane PD, Pollier J, Panda S, Szymanski J, Massalha H, Yona M, Unger T, Malitsky S, Arendt P, Pauwels L, Almekias-Siegl E, Rogachev I, Meir S, Cárdenas PD, Masri A, Petrikov M, Schaller H, Schaffer AA, Kamble A, Giri AP et al. (2016) Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. 3: 16205
Souter M, Topping J, Pullen M, Friml J, Palme K, Hackett R, Grierson D, Lindsey K (2002) hydra Mutants of Arabidopsis are defective in sterol profiles and auxin and ethylene signaling. Plant Cell 14: 1017-31
Souter MA, Pullen ML, Topping JF, Zhang X, Lindsey K (2004) Rescue of defective auxin-mediated gene expression and root meristem function by inhibition of ethylene signalling in sterol biosynthesis mutants of Arabidopsis. Planta 219: 773-783
Sun J, Chen Q, Qi L, Jiang H, Li S, Xu Y, Liu F, Zhou W, Pan J, Li X, Palme K, Li C (2011) Jasmonate modulates endocytosis and plasma membrane accumulation of the Arabidopsis PIN2 protein. New Phytologist 191: 360-375
Szemenyei H, Hannon M, Long JA (2008) TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 319: 1384-6

71
Tanaka H, Dhonukshe P, Brewer PB, Friml J (2006) Spatiotemporal asymmetric auxin distribution: a means to coordinate plant development. Cell Mol Life Sci 63: 2738-54
Tejos R, Sauer M, Vanneste S, Palacios-Gomez M, Li H, Heilmann M, van Wijk R, Vermeer JE, Heilmann I, Munnik T,
Friml J (2014) Bipolar Plasma Membrane Distribution of Phosphoinositides and Their Requirement for Auxin -Mediated Cell Polarity and Patterning in Arabidopsis. Plant Cell 26: 2114-2128
Titapiwatanakun B, Murphy AS (2009) Post-transcriptional regulation of auxin transport proteins: cellular trafficking, protein phosphorylation, protein maturation, ubiquitination, and membrane composition. J Exp Bot 60: 1093-107
Traub LM, Bonifacino JS (2013) Cargo recognition in clathrin-mediated endocytosis. Cold Spring Harb Perspect Biol 5: a016790
Tromas A, Paque S, Stierle V, Quettier AL, Muller P, Lechner E, Genschik P, Perrot-Rechenmann C (2013) Auxin-binding protein 1 is a negative regulator of the SCF(TIR1/AFB) pathway. Nature communications 4: 2496
van Berkel K, de Boer RJ, Scheres B, ten Tusscher K (2013) Polar auxin transport: models and mechanisms. Development 140: 2253-68
Van Damme D, Gadeyne A, Vanstraelen M, Inze D, Van Montagu MC, De Jaeger G, Russinova E, Geelen D (2011) Adaptin-like protein TPLATE and clathrin recruitment during plant somatic cytokinesis occurs via two distinct pathways. Proc Natl Acad Sci U S A 108: 615-20
Vanneste S, Friml J (2009) Auxin: a trigger for change in plant development. Cell 136: 1005-16
Vanneste S, Friml J (2013) Calcium: The Missing Link in Auxin Action. Plants (Basel) 2: 650-75
Vieten A, Vanneste S, Wisniewska J, Benkova E, Benjami ns R, Beeckman T, Luschnig C, Friml J (2005) Functional redundancy of PIN proteins is accompanied by auxin-dependent cross-regulation of PIN expression. Development 132: 4521-31
Vriet C, Russinova E, Reuzeau C (2013) From squalene to brassinolide: the steroid metabolic and signaling pathways across the plant kingdom. Mol Plant 6: 1738-57
Warrilow AG, Parker JE, Kelly DE, Kelly SL (2013) Azole affinity of sterol 14alpha -demethylase (CYP51) enzymes from Candida albicans and Homo sapiens. Antimicrob Agents Chemother 57: 1352-60
Willemsen V, Friml J, Grebe M, van den Toorn A, Palme K, Scheres B (2003) Cell polarity and PIN protein positioning in Arabidopsis require STEROL METHYLTRANSFERASE1 function. Plant Cell 15: 612-25

72
Willige BC, Isono E, Richter R, Zourelidou M, Schwechheimer C (2011) Gibberellin Regulates PIN-FORMED
Abundance and Is Required for Auxin Transport–Dependent Growth and Development in <em>Arabidopsis thaliana</em>. Plant Cell 23: 2184-2195
Winter D, Vinegar B, Nahal H, Ammar R, Wilson GV, Provart NJ (2007) An "Electronic Fluorescent Pictograph" browser for exploring and analyzing large-scale biological data sets. PloS one 2: e718
Wisniewska J, Xu J, Seifertova D, Brewer PB, Ruzicka K, Bli lou I, Rouquie D, Benkova E, Scheres B, Friml J (2006 ) Polar PIN localization directs auxin flow in plants. Science 312: 883
Xu J, Scheres B (2005) Dissection of Arabidopsis ADP-RIBOSYLATION FACTOR 1 function in epidermal cell polarity. Plant Cell 17: 525-36
Xu T, Dai N, Chen J, Nagawa S, Cao M, Li H, Zhou Z, Chen X, De Rycke R, Rakusova H, Wang W, Jones AM, Friml J,
Patterson SE, Bleecker AB, Yang Z (2014) Cell surface ABP1-TMK auxin-sensing complex activates ROP GTPase signaling. Science 343: 1025-8
Yang H, Richter GL, Wang X, Mlodzinska E, Carraro N, Ma G, Jenness M, Chao DY, Peer WA, Murphy AS (2013) Sterols and sphingolipids differentially function in trafficking of the Arabidopsis ABCB19 auxin transporter. Plant J 74: 37-47
Zazimalova E, Krecek P, Skupa P, Hoyerova K, Petrasek J (2007) Polar transpor t of the plant hormone auxin - the role of PIN-FORMED (PIN) proteins. Cell Mol Life Sci 64: 1621-37
Zazimalova E, Murphy AS, Yang H, Hoyerova K, Hosek P (2010) Auxin transporters --why so many? Cold Spring Harb Perspect Biol 2: a001552
Zelazny E, Santambrogio M, Gaude T (2013a) Retromer association with membranes: plants have their own rules! Plant signaling & behavior 8
Zelazny E, Santambrogio M, Pourcher M, Chambrier P, Berne-Dedieu A, Fobis-Loisy I, Miege C, Jail lais Y, Gaude T
(2013b) Mechanisms governing the endosomal membrane recruitment of the core retromer in Arabidopsis. J Biol Chem 288: 8815-25
Zhang X, Li H (2016) Role of Arabidopsis sterol 4alpha -methyl oxidase2 family in embryo and postembryonic development. Plant signaling & behavior 11: e1249081
Zhao Y (2010) Auxin biosynthesis and its role in plant development. Annu Rev Plant Biol 61: 49-64

73
Part 7. Addendum
7.1 Differential expression of sterol biosynthesis genes after auxin treatment
We consulted several expression databases such as the Arabidopsis eFP browser (Winter et al.,
2007) and microarray expression databases belonging to our institute searching for genes that
responded in any way to auxin treatment and that were involved in sterol biosynthesis. We chose 9 genes in order to perform qPCR experiments to confirm their behavior upon auxin treatment
and further to test their expression levels after treatment with both auxin and sterol biosynthesis inhibitors (Table 2). We designed primers for each of them to be used in qPCR experiments with
the help of the Quantprime software (Arvidsson et al., 2008).
Table 2. Chosen genes to perform qPCR and their primers
Transcript identifier /
Gene name
Forward primer sequence Fwd Tm
Reverse primer sequence Rev Tm
Amplicon length
AT2G17330 CYP51A1
TCACCGTGACAACTCGAGAAGG 65 TGGCGAATGCAGGAGAAGTTGC 64 78
AT1G76090 SMT3
TACCAAGTGCAACGAGCCAAGC 64 ACCACAAACGACGTTGCAGAGAG 64 72
AT1G20330 CVP1/SMT2
AACTTCCTCCAGATGCCGTTCG 63 ACGTGGCTTCGATGGAATAAGCAC 64 64
AT2G34490
CYP710A2
TGTCCCACGATATCGCCAAAGG 63 ACGTTACGATGCGCTTAGACAAG 61 61
AT4G12110 SMO1-1
GCAACTTCGCTTCAGTGTTCACG 63 TCGACGACTCCTTGATCTGCTC 62 102
AT4G22756 SMO1-2
GTTGGAGGACAAAGCCAGAGTAAC
62 CTTGGACTTCTCCTTCATCTGCTG 61 123
AT2G34500 CYP710A1
AGGCTTCGATCAGACGGTTGTG 63 ACAAGAACACGGTGCACCCATC 64 79
AT5G50375 CPI1
GCTTCCCAATGTTCTTCAGGATGG 62 AGGTCCCAAGAAGAGACGCCATAG
64 134
AT1G11680
CYP51A2
AGCCTCGCACAGTGATTTCAGC 64 TGCAACGATGTGACCCTTTGGG 64 76
74
7.2 Fusion of sterol biosynthesis genes to fluorescent tags to generate overexpressing/
reporter lines
From the aforementioned 9 genes, we were interested in fusing CVP1/SMT2, SMO1-1 and
CYP710A1 (genes that delimit the ethyl-sterol branch biosynthesis pathway) to fluorescent tags
and diverse promoters in order to be able to generate Arabidopsis lines that overexpress or mark
the expression of such genes. These genes were interesting because having tried the response of
STE1, DWF1 and DWF5 overexpressing lines, we had seen that STE1 OE plants were, for example,
hypersensitive to some of the compounds and DWF1 and DWF5 OE plants were resistant to some
of the compounds. Also, to be able to know the location or modulate the expression of the gene
with a reporter would be also useful.
We designed a pair of primers for each gene (Table 3) to amplify their coding region. Each primer
has a border (attb1 or attb2) that allows it to be seamlessly pasted by recombination in a
Gateway™ entry clone vector, which facilitates enormously their fusing to a variety of already
cloned promoters and fluorescent tags.
Table 3. Primers to amplify coding regions of genes of interest
Transcript identifier / Gene name
Forward primer sequence Reverse primer sequence Amplicon length
CYP710A1 GGGGACAAGTTTGTACAAAAAAGCAG
GCTCCAACATGGTTTTCTCTGTTTCTAT
GGGGACCACTTTGTACAAGAAAGCTGGGTC
ACCTCCGCCACCGGAAAAGTTGGGATACTTTG
1.6kbp
SMO 1-1 GGGGACAAGTTTGTACAAAAAAGCAGGCTCCAACATGATTCCTTACGCTACAGT
GGGGACCACTTTGTACAAGAAAGCTGGGTCACCTCCGCCACCATCGGATTTTATTCCTCCGT
851bp
SMT2 GGGGACAAGTTTGTACAAAAAAGCAG
GCTCCAACATGGACTCTTTAACACTCTT
GGGGACCACTTTGTACAAGAAAGCTGGGTC
ACCTCCGCCACCAGAACTCTCCTCCGGTGACT
1051bp
We amplified them (Figure 32), purified the amplified DNA fragments and fused them by
recombination in the Gateway™ entry clone vector (confirmation done by sequencing). We
propagated the entry clones by a DNA minipreparation and performed a second recombination
reaction to fuse the coding regions to several promoters and fluorescent tags, and were
confirmed by sequencing (Table 4). The clones are at the moment ready to be introduced to plants.

75
Table 4. Promoters and tags fused to genes of interest
Promoter Gene Tag
UBQ10 (constitutive) DIM1 (previously cloned) R-GECO (calcium reporter) UBQ10 SMT2 R-GECO
UBQ10 DIM1 GEM-GECO
UBQ10 SMT2 GEM-GECO
UBQ10 CYP710A1 mRuby3 (fluorescent tag)
UBQ10 SMO1-1 mRuby3 (fluorescent tag)
Rx (estradiol inducible) CYP710A1 mRuby3 (fluorescent tag)
Rx SMO1-1 mRuby3 (fluorescent tag)
Rx DIM1 mRuby3 (fluorescent tag)
Rx SMT2 mRuby3 (fluorescent tag)
Figure 32. Amplification of SMT2, SMO1-1 and CYP710A1. Arabidopsis genomic DNA was used as a template to amplify the coding regions of SMT2, SMO1-1 and CYP710A1. Right: Smart Ladder molecular weight marker.
SMT2 SMO1-1 CYP710A1


















