
files.transtutors.com · Web viewABSTRACT α-Enolase is an important glycolytic enzyme in the...
26
ABSTRACT α-Enolase is an important glycolytic enzyme in the cytoplasm of prokaryotic and eukaryotic cells and is considered a multifunctional protein. α-Enolase is expressed on several cell surface, where it acts as a plasminogen receptor, hydrolyzing the inactive plasminogen into active proteolytic plasmin. In addition to glycolytic enzyme and plasminogen receptor functions, α-Enolaseperforms other cellular functions and found at varying subcellular localizations that are distinct from its well- established function in glycolysis. Therapeutically α-enolase is an essential enzyme as many streptococcal species such as Streptococcus mutansuses plasminogen binding site of α- enolase to spread infective endocarditis. In this study functional identity of recombinant α-enolase has been confirmed by its ability to catalyze glycolytic reactions and its enzyme kinetics.α-Enolase is localized on the surface of hematopoietic cells such as monocytes, T cells and B cells, neuronal cells, and endothelial cells as a strong plasminogen receptor, modulating pericellularfibrinolytic activity. 1
Transcript of files.transtutors.com · Web viewABSTRACT α-Enolase is an important glycolytic enzyme in the...

ABSTRACT
α-Enolase is an important glycolytic enzyme in the cytoplasm of prokaryotic and eukaryotic cells
and is considered a multifunctional protein. α-Enolase is expressed on several cell surface, where
it acts as a plasminogen receptor, hydrolyzing the inactive plasminogen into active proteolytic
plasmin. In addition to glycolytic enzyme and plasminogen receptor functions, α-
Enolaseperforms other cellular functions and found at varying subcellular localizations that are
distinct from its well-established function in glycolysis. Therapeutically α-enolase is an essential
enzyme as many streptococcal species such as Streptococcus mutansuses plasminogen binding
site of α- enolase to spread infective endocarditis. In this study functional identity of recombinant
α-enolase has been confirmed by its ability to catalyze glycolytic reactions and its enzyme
kinetics.α-Enolase is localized on the surface of hematopoietic cells such as monocytes, T cells
and B cells, neuronal cells, and endothelial cells as a strong plasminogen receptor, modulating
pericellularfibrinolytic activity.
1

Table of content
TOPIC PAGE NO.
ABSTRACT 1
INTRODUCTION 3
METHOD 6
DISCUSSION 14
REFERENCES 15
2

INTRODUCTION
Enolase (2-phospho-D-glycerate hydrolyase, EC4.2.1.11) is a key glycolytic enzyme [1] in the
cytotoplasma of both prokaryotes and eukaryotes and is also considered as multifunctional
protein. It is also one of the most abundantly expressed cytosolic enzyme in many organisms
discovered in 1934 by Lohman and Mayerhof. Itcatalyzes the dehydration of 2-phospho D-
glycerate (2PGA) to phosphoenolpyruvate (PEP) in glycolysis as shown in figure 2, and the
reverse reaction in gluconeogenesis.Before binding with substrate enolase form complex with
divalent metal cations and hence it is characterize as metalloenzyme. It shows highest activity
when bound to metallic cation Mg+2. Enolaseexist in three isoformsin mammals and birds:α
enolase encoded by Eno1 gene found in all human tissues, β enolaseencoded by Eno3 gene found
only in muscle tissues and γ enolase encoded by Eno2 gene found only in neuron and
neuroendocrine tissues [2] .the three isoforms of enolase share high structural and kinetic
similarities [3,4]. Enolase exist in a dimeric form either homodimer or heterodimer composed of
two subunits arranged in antiparallel fashion as shown in figure 1 [4,5].
3
1st α subunit of α enolasehomodimer
2nd α subunit of α enolasehomodimer

Figure 1: PDB 2PSN structural model of α enolase. It’s a homodimer with two structural units
facing each other in antiparallel fashion
4

Figure 2: Metabolic chain reactions of glycolysis, the central pathway for the catabolism of
carbohydrates that takes place in the cytoplasm of almost all prokaryotic and eukaryotic cells.
The insert shows different enolaseisoenzymes in vertebrates.
α-Enolase mRNA translation is under developmental control which is highly unregulated during
cellular growth [6,7]. Recent studies have shown that in addition to its innate glycolytic function
α-enolase play key role in several biological and pathophysiological processes. α-Enolase has an
alternative stop codon by using it α-enolase m-RNA can be translated into an additional 37 kDa
protein which lacks the first 96 amino acids residues and called as c-myc promoter binding
protein 1 (MBP- 1) localized in nucleus and negatively regulates transcription of the
protooncogene [8].
α-Enolaseis a multifunctional protein as is found on the surface of hematopoietic cells such as
monocytes, T cells and B cells, neuronal cells, and endothelial cells as a strong plasminogen
receptor, modulating pericellularfibrinolytic activity. It is also described neurotrophic factor [9],
a heat-shock protein (HSP48) [10], and a hypoxic stress protein [11], it’s a part of the crystallin
lens of vertebrates [12], binds to fragments of F-actin and tubulin [13], and has been detected
associated to centrosomes in HeLa cells [14].α-Enolase also binds with other adjacent glycolytic
enzyme such aspyruvate kinase, phosphoglyceratemutase and andolase (which is associated with
cytoskeletal protein) with high affinity [15] (figure 3).
Sterptococcus mutans is a gram positive bacterium and a causative agent of oral cavity and
infectious endocarditis. S.mutans cells are found to associate with blood component and then
eventually colonizes cardiovascular endothelium causing deadly infectious endocarditis. they
associate with fibronectin and degrade by activating plasminogen, hence spread through blood in
body of host organisum.
5

Figure 3: Shows multifunctional characteristics of α-enolase
Recombinant DNA or Molecular Cloning
In this process Eno1 gene forms covalent interaction with plasmid vector. Many copies of the
hybrid DNA may be produced by the progeny of the recipient cells; hence, the DNA molecule is
cloned. If theinserted fragment contains a functional gene carrying the code for a specific
protein, many copies of that gene and translated protein may be produced in the host cell. This
process has become important for the large-scale production of proteinsthat are of value in
medicine and basic science.
6

Figure 4: Different steps of molecular cloning
7

Materials and Methods
Preparation of α-enolase antiserum
Antisera against S. mutans α-enolase was prepared by immunizing subcutaneously New
Zealand female rabbits with 200 μg of purified his-tagged α-enolase protein emulsified in
Freund's incomplete adjuvant containing 10% muramyl peptide. Rabbits were boosted two
weeks later with the same amount of his-tagged α-enolase in Freund's incomplete adjuvant and
after two additional weeks were bled. Antiserum to the α-enolase of S. pyogenes (anti-SEN)
was a gift from Vijaykumar Pancholi [10].
Isolation of cellular protein fractions
Extracellular, cytoplasmic, and cell wall protein fractions of S. mutans M51 were prepared
from 1-liter cultures grown at 37 ° C for 48 hours in Todd-Hewitt broth as described by
Kuykindoll and Holt [17]. Total cell lysates of S. mutans M 51 were prepared from 20 ml of
culture grown overnight at 37 ° C in Todd-Hewitt broth. Cells were collected by centrifugation
and resuspended in 10 mM phosphate buffered saline, pH 7.0. The cell suspension was
transferred to a 2 ml bead beater tube and 0.4 mg of 0.1 mm zirconia/silica beads per ml of cell
suspension were added. The tube was placed in a Mini-Beadbeater 3110BX and run in one
minute time periods with cooling on ice between time periods for a total of 3 minutes. The
beads were allowed to settle and the supernatant fluid (total cell extract) was removed.
Source of DNA fragment
Eno1 gene is isolated from Streptococcus mutans.
The Polymerase Chain Reaction Amplifies Specific DNA Sequence
PCR amplifies Eno1 gene , given below is α enolase protein sequence.This gene that encodes
alpha enolase is use to design oligonucleotide PCR primer for sequence amplification. There will
be two primers forward primer (5′- ATGTCA ATT ATT ACT GAT GTT TAC GCT CGC-3′)
that starts from base 1 and ends at base 30 and reverse primer (5′- TTATTT TTT CAA GTT
GTA GAA TGA TTT AAG ACC ACG-3′) began at base 1263 and ends at base 1296 of the
sequence including termination codon.The forward primer began at base 1 and ended at base 30,
8

while the backward primer began at base 1263 and ended at base 1296 of the sequence and
includes a termination codon.The primer pair was used in PCR reactions with denaturing,
annealing, and extension temperatures and times of 94 ° C for 1 minute, 50 ° C for 30 seconds,
and 72 ° C for 3 minutes; respectively. The amplified PCR product generated was approximately
1.3 kb in size, which is the correct size based on the nucleotide sequence. The α-enolase PCR
product was cloned into the TA expression vector pCR T7/NT TOPO-TA. The resulting plasmid
DNA was transformed into E. coli TOP 10F' cells and spread on LB plates supplemented with
100 μg/ml of ampicillin. Plasmid DNA was isolated from several transformants and digested
with restriction enzymes to verify the size of the insert DNA (figure 6 and 7). The plasmid DNA
was then sequenced for determination of the sequence of the insert DNA using Big Dye
terminator reactions.
Figure 6: Plasmid vector for Eno1 gene
9

Figure 6: Ligation of amplified PCR product into Invitrogen's TOPO TA Cloning System
For the fast and efficient cloning of polymerase chain reaction product the recommended clonind
system is Invitrogen's TOPO TA Cloning System (TOPO Cloning). This is a linearized plasmid
3’ overhang consisting of deoxythimidine (T) which form covalent interaction with
topoisomerase I. On the other hand PCR product 3’ overhang consist of deoxyadenine (A)
which is complementary to that of the plasmid 3’ polyT overhang, allowing fast ligation in the
presence of topoisomerase 1. Engineered plasmid is then transformed into competent E. coli
TOP 10F' cells [32].
10

Figure 7: Transformation of E.coli
11

RESULTS
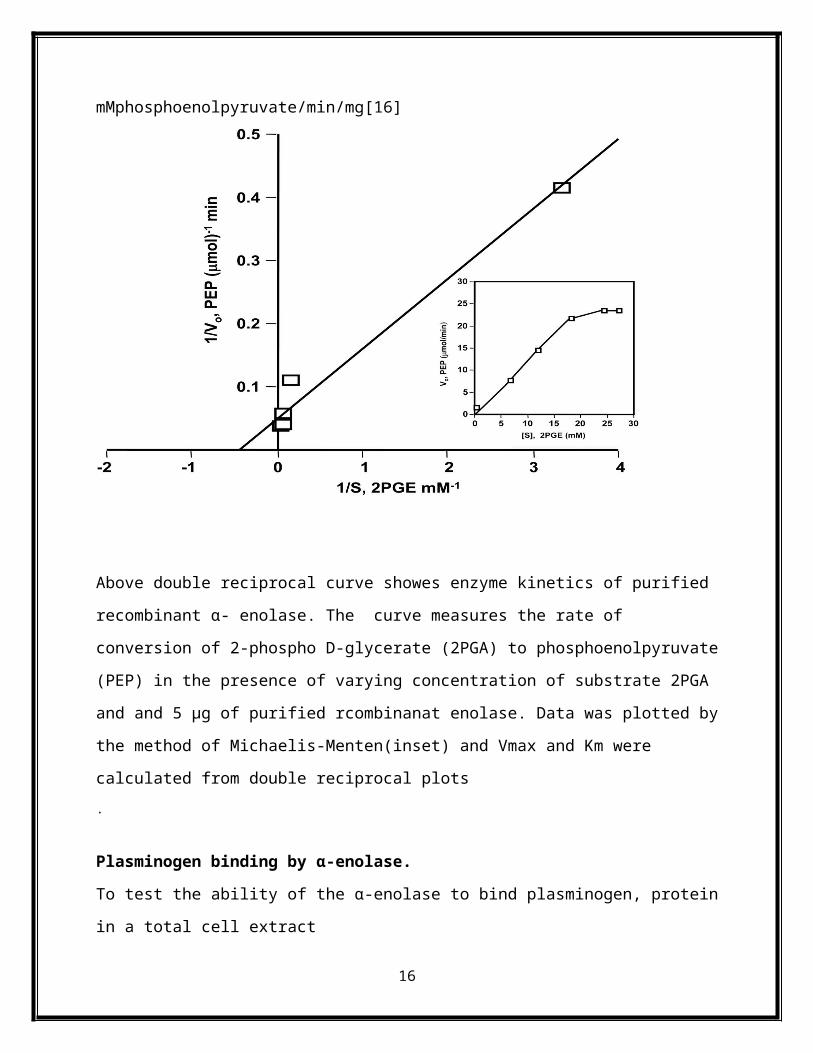
Alpha-enolase activity and enzyme kinetic properties of recombinant Enolase(rEno)
Similar to the enzyme reaction catalyzed by α enolase recombinant proteinrEno also possessed
the same functional capacity to catalyze the conversion of 2-phosphoglycerate to
phosphoenolpyruvate.
Kinetic properties of rEno was determined by enzyme reaction kinetics using purified his-tagged
protein with various concentrations of 2-phosphoglycerate. Values for Km and Vmax were
calculated from reciprocal plots. This analysis revealed a Km value of 9.5 mM for 2-
phosphoglycerate and a Vmax of 31 mMphosphoenolpyruvate/min/mg for rSmEno. On the other
hand α enolaseKmvalue of1.492 mMfor 2-phosphoglycerate andVmaxof31.25
mMphosphoenolpyruvate/min/mg[16]
12

Above double reciprocal curve showes enzyme kinetics of purified recombinant α- enolase. The
curve measures the rate of conversion of 2-phospho D-glycerate (2PGA) to phosphoenolpyruvate
(PEP) in the presence of varying concentration of substrate 2PGA and and 5 μg of purified
rcombinanat enolase. Data was plotted by the method of Michaelis-Menten(inset) and Vmax and
Km were calculated from double reciprocal plots.
Plasminogen binding by α-enolase.
To test the ability of the α-enolase to bind plasminogen, protein in a total cell extract
and an extracellular protein fraction along with purified rSmEno were separated
by SDS-PAGE and transferred to nitrocellulose membranes. The blot was incubated with human
plasminogen and then probed with antibodies against human plasminogen. The
resultsdemonstrated that plasminogen bound to a protein of approximately 50 kDa in the total
cellextract, extracellular protein fraction and with purified rEno that migrated with a mobility
that was very similar to a protein in each of the preparations that reacted with anti-rEno sera.
13

Above blot shows Plasminogen binding to the α-enolase of S. mutans.SDS-polyacrylamide
gelelectrophoresis of a total protein extract and extracellular protein of S. mutans and purified
rSmEno. Lane 1, molecular weight markers.
DISCUSSION
This study has demonstrated that the chemical attributes of α- enolase and recombinant α-
enolase don’t differ considerably. Both α- enolase as well as recombinant α- enolase processes
their innate ability to catalyze dehydration of 2-phospho D-glycerate (2PGA) to
phosphoenolpyruvate (PEP). The enzyme kinetic data reveled both enolase exhibits same Vmax .
The α- enolase of streptococcus specieshave on its C-terminal end two lysine residues that have
been reported to interact with the lysine binding sites of plasminogen [33,34]. Bergmann et al.
[35] have demonstrated that the enolase of S. pneumoniae, designated Eno, has an additional
internal plasminogen binding epitope and that this internal motif is crucial for the interaction of
the Eno protein with human plasminogen. These investigators identified a motif of nine amino
acids, FYDKERKVY (amino acid residues crucial for plasminogen binding are bold and
underlined), that bound human plasminogen. The acidic amino acids, aspartic acid (position 3)
and glutamic acid, and the lysine residues at positions 4 and 7 in the motif were found to be
important for plasminogen binding by the S. pneumoniae enolase. Furthermore, mutant strains of
S.pneumoniae with substitutions in the critical amino acids of the internal plasminogen-binding
motif were substantially less virulent than a wild-type strain of S. pneumoniae. The S.
mutansα-enolase has the sequence, FYDNG**VY, that has five amino acids in common with
the internal plasminogen-binding motif of S. pneumoniae. However of the amino acids required
for plasminogen binding, only the aspartic acid at position 3 is present and two of the amino
acids in the motif are not present at all. A similar sequence was found in the enolase of S.
sobrinus, another caries causing oral streptococcus. This finding indicates that the S. mutans
enolase does not have a functional internal plasminogen-binding site and suggests that enolase
mediated plasminogen binding by S. mutans occurs primarily through interaction of
plasminogen with the C-terminal lysyl residues. Previously, we have shown that plasminogen
activation by S. mutans is almost totally inhibited by lysine or the lysine analog, ε-aminocaproic
14

acid [36].
In conclusion, we have characterized the α-enolase of S. mutans and provided evidence that
this typically cytoplasmic protein can translocate the cell wall of the oral bacterium, S.
mutans, and function as a receptor for plasminogen. This finding suggests that this generally
non-invasive oral organism may have virulence potential similar to that of more pathogenic
streptococci
REFERENCES
1. V. Pancholi, “Multifunctional α-enolase: its role in diseases,” Cellular and Molecular
Life Sciences, vol. 58, no. 7, pp. 902–920, 2001.
2. P. J. Marangos, A. M. Parma, and F. K. Goodwin, “Functional properties of neuronal and
glial isoenzymes of brain enolase,” Journal of Neurochemistry, vol. 31, no. 3, pp. 727–
732, 1978.
3. S. Feo, D. Oliva, G. Barbieri, W. Xu, M. Fried, and A. Giallongo, “The gene for the
muscle-specific enolase is on the short arm of human chromosome 17,” Genomics, vol. 6,
no. 1, pp. 192–194, 1990. View at Publisher · View at Google Scholar ·
4. L. Fletcher, C. C. Rider, and C. B. Taylor, “Enolaseisoenzymes. III. Chromatographic
and immunological characteristics of rat brain enolase,” BiochimicaetBiophysicaActa,
vol. 452, no. 1, pp. 245–252, 1976.
5. K. Kato, Y. Okagawa, F. Suzuki, A. Shimizu, K. Mokuno, and Y. Takahashi,
“Immunoassay of human muscle enolase subunit in serum: a novel marker antigen for
muscle diseases,” ClinicaChimicaActa, vol. 131, no. 1-2, pp. 75–85, 1983.
6. J. P. Holland, L. Labieniec, C. Swimmer, and M. J. Holland, “Homologous nucleotide
sequences at the 5' termini of messenger RNAs synthesized from the yeast enolase and
glyceraldehyde-3-phosphate dehydrogenase gene families. The primary structure of a
third yeast glyceraldehyde-3-phosphate dehydrogenase gene,” Journal of Biological
Chemistry, vol. 258, no. 8, pp. 5291–5299, 1983
7. A. Giallongo, D. Oliva, L. Calì, G. Barba, G. Barbieri, and S. Feo, “Structure of the
human gene for alpha-enolase,” European Journal of Biochemistry, vol. 190, no. 3, pp.
567–573, 1990.
15
8. S. Feo, D. Arcuri, E. Piddini, R. Passantino, and A. Giallongo, “ENO1 gene product
binds to the c-myc promoter and acts as a transcriptional repressor: relationship with Myc
promoter-binding protein 1 (MBP-1),” FEBS Letters, vol. 473, no. 1, pp. 47–52, 2000
9. N. Takei, J. Kondo, K. Nagaike, K. Ohsawa, K. Kato, and S. Kohsaka, “Neuronal
survival factor from bovine brain is identical to neuron-specific enolase,” Journal of
Neurochemistry, vol. 57, no. 4, pp. 1178–1184, 1991
10. H. Iida and I. Yahara, “Yeast heat-shock protein of M(r) 48,000 is an isoprotein of
enolase,” Nature, vol. 315, no. 6021, pp. 688–690, 1985
11. R. M. Aaronson, K. K. Graven, M. Tucci, R. J. McDonald, and H. W. Farber, “Non-
neuronal enolase is an endothelial hypoxic stress protein,” Journal of Biological
Chemistry, vol. 270, no. 46, pp. 27752–27757, 1995.
12. R. L. Mathur, M. C. Reddy, S. Yee, R. Imbesi, B. Groth-Vasselli, and P. N. Farnsworth,
“Investigation of lens glycolytic enzymes: species distribution and interaction with
supramolecular order,” Experimental Eye Research, vol. 54, no. 2, pp. 253–260, 1992.
13. J. L. Walsh, T. J. Keith, and H. R. Knull, “Glycolytic enzyme interactions with tubulin
and microtubules,” BiochimicaetBiophysicaActa, vol. 999, no. 1, pp. 64–70, 1989.
14. S. A. Johnstone, D. M. Waisman, and J. B. Rattner, “Enolase is present at the centrosome
of HeLa cells,” Experimental Cell Research, vol. 202, no. 2, pp. 458–463, 1992.
15. T. Merkulova, M. Lucas, C. Jabet et al., “Biochemical characterization of the mouse
muscle-specific enolase: developmental changes in electrophoretic variants and selective
binding to other proteins,” Biochemical Journal, vol. 323, no. 3, pp. 791–800, 1997.
16. Pancholi V, Fischetti VA. A major surface protein on group A streptococci is a
glyceraldehyde-3-phosphate-dehydrogenase with multiple binding activity. J Exp Med
1992;176:415–426. [PubMed:1500854]
17. P. A. Fontan, V. Pancholi, M. M. Nociari, and V. A. Fischetti, “Antibodies to
streptococcal surface enolase react with human α-enolase: implications in
poststreptococcalsequelae,” Journal of Infectious Diseases, vol. 182, no. 6, pp. 1712–
1721, 2000.
18. A. Kinloch, V. Tatzer, R. Wait et al., “Identification of citrullinated alpha-enolase as a
candidate autoantigen in rheumatoid arthritis,” Arthritis Research & Therapy, vol. 7, no.
6, pp. R1421–1429, 2005
16

19. F. Pratesi, S. Moscato, A. Sabbatini, D. Chimenti, S. Bombardieri, and P. Migliorini,
“Autoantibodies specific for α-enolase in systemic autoimmune disorders,” Journal of
Rheumatology, vol. 27, no. 1, pp. 109–115, 2000
20. K. Wakui, M. Tanemura, K. Suzumori et al., “Clinical applications of two-color
telomeric fluorescence in situ hybridization for prenatal diagnosis: identification of
chromosomal translocation in five families with recurrent miscarriages or a child with
multiple congenital anomalies,” Journal of Human Genetics, vol. 44, no. 2, pp. 85–90,
1999
21. V. Saulot, O. Vittecoq, R. Charlionet et al., “Presence of autoantibodies to the glycolytic
enzyme α-enolase in sera from patients with early rheumatoid arthritis,” Arthritis and
Rheumatism, vol. 46, no. 5, pp. 1196–1201, 2002.
22. N. Wegner, K. Lundberg, A. Kinloch et al., “Autoimmunity to specific citrullinated
proteins gives the first clues to the etiology of rheumatoid arthritis,” Immunological
Reviews, vol. 233, no. 1, pp. 34–54, 2010.
23. X. Chang and C. Wei, “Glycolysis and rheumatoid arthritis,” International Journal of
Rheumatic Diseases, vol. 14, no. 3, pp. 217–222, 2011.
24. C. Roozendaal, M. H. Zhao, G. Horst et al., “Catalase and α-enolase: two novel
granulocyte autoantigens in inflammatory bowel disease (IBD),” Clinical and
Experimental Immunology, vol. 112, no. 1, pp. 10–16, 1998.
25. N. Vermeulen, I. Arijs, S. Joossens et al., “Anti-α-enolase antibodies in patients with
inflammatory bowel disease,” Clinical Chemistry, vol. 54, no. 3, pp. 534–541, 2008.
26. E. Ballot, A. Bruneel, V. Labas, and C. Johanet, “Identification of rat targets of anti-
soluble liver antigen autoantibodies by serologic proteome analysis,” Clinical Chemistry,
vol. 49, no. 4, pp. 634–643, 2003.
27. M. Bruschi, M. L. Carnevali, C. Murtas et al., “Direct characterization of target podocyte
antigens and auto-antibodies in human membranous glomerulonephritis: alfa-enolase and
borderline antigens,” Journal of Proteomics, vol. 74, no. 10, pp. 2008–2017, 2011.
28. S. T. Tsai, I. H. Chien, W. H. Shen et al., “ENO1, a potential prognostic head and neck
cancer marker, promotes transformation partly via chemokine CCL20 induction,”
European Journal of Cancer, vol. 46, no. 9, pp. 1712–1723, 2010.
17

29. S. Jin, R. S. DiPaola, R. Mathew, and E. White, “Metabolic catastrophe as a means to
cancer cell death,” Journal of Cell Science, vol. 120, no. 3, pp. 379–383, 2007.
30. H. Iwabata, M. Yoshida, and Y. Komatsu, “Proteomic analysis of organ-specific post-
translational lysine-acetylation and -methylation in mice by use of anti-acetyllysine and -
methyllysine mouse monoclonal antibodies,” Proteomics, vol. 5, no. 18, pp. 4653–4664,
2005.
31. K. Wakui, M. Tanemura, K. Suzumori et al., “Clinical applications of two-color
telomeric fluorescence in situ hybridization for prenatal diagnosis: identification of
chromosomal translocation in five families with recurrent miscarriages or a child with
multiple congenital anomalies,” Journal of Human Genetics, vol. 44, no. 2, pp. 85–90,
1999.
32. Hoover C, 1998 July 8. Invitrogen web catalog.
<http://www.invitrogen.com/pdf_manuals/topota_man.pdf> Accessed 1999 Feb 14.
33. Bergmann S, Rohde M, Chhatwal GS, Hammerschmidt S. alpha-Enolase of
Streptococcus pneumonia is a plasmin(ogen)-binding protein displayed on the bacterial
cell surface. Mol Microbiol 2001;40:1273–1287. [PubMed: 11442827].
34. Pancholi V, Fischetti VA. alpha-enolase, a novel strong plasmin(ogen) binding protein on
the surfaceof pathogenic streptococci. J Biol Chem 1998;273:14503–14515. [PubMed:
9603964]
35. Bergmann S, Wild D, Diekmann O, Frank R, Bracht D, Chhatwal GS, Hammerschmidt
S.Identification of a novel plasmin(ogen)-binding motif in surface displayed alpha-
enolase ofStreptococcus pneumoniae. Mol Microbiol 2003;49:411–423. [PubMed:
12828639]
36. Jones MN, Holt RG. Activation of plasminogen by Streptococcus mutans. Biochem
Biophys ResCommun 2004;322:37–41. [PubMed: 15313170]
18


















