
Fall Semester 2011 Lecture 11 Lipids III - Washington...
28
MPS 587 - Advanced Plant Biochemistry Course Fall Semester 2011 Lecture 11 Lipids III 9. Triacylglycerol synthesis 10. Engineering triacylglycerol fatty acid composition
Transcript of Fall Semester 2011 Lecture 11 Lipids III - Washington...

MPS 587 - Advanced Plant Biochemistry Course
Fall Semester 2011
Lecture 11
Lipids III
9. Triacylglycerol synthesis 10. Engineering triacylglycerol fatty acid composition

Today’s topics on the Arabidopsis node map
2

Tissue and function of triacylglycerol accumulation in plants
1. Seed, storage oil – energy for germinating seedlings
1. Soybeans
2. Brassica napus (Canola)
2. Fruit, animal attractant?
1. Avocado mesocarp
2. Palm fruit
3. Leaf, intermediate in lipid breakdown during senescence
4. Anthers and pollen, may be used for energy in pollen tube growth
3

Lipids in oil bodies (oleosomes)
• Contain TAGs.
• Surrounded by protein-rich membrane; size 1-2 um.
• ER contains biosynthetic enzymes; oil bodies are spun off from ER microdomains.
• Oleosins: 15-25 kDa proteins; conserved hydrophobic core; amphipathic flanking sequences; present only in seed and pollen.
4

Plant triacylglycerol biosynthesis – overview
Malonyl-CoA
Acyl-ACP (16:0;18:0)
Acyl-ACP (18:1)
Acetyl-CoA
ACCase
FAS Desaturase
PC
Pyruvate
LACS Acyl-CoA
Thioesterase
FA
G3P
LPA
PA
DAG
PLASTID
PAP
ACYL EDITING LPC
ENDOPLASMIC RETICULUM
FA SYNTHESIS
LIP
ID A
SSEM
BLY
FA MODIFICATION • Lipid linked desaturases
• Acyl-CoA elongases
DAG
TAG
TAG
Oil body
TAG
DGAT
PDAT
GPAT
LPAT
5
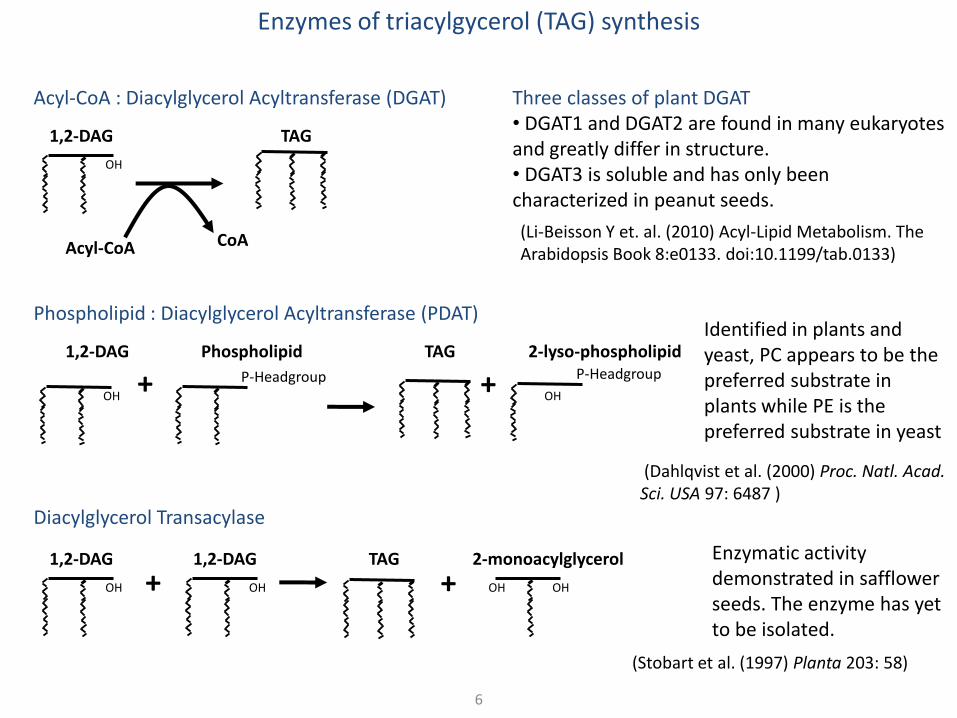
Enzymes of triacylgycerol (TAG) synthesis
Acyl-CoA CoA
1,2-DAG
Acyl-CoA : Diacylglycerol Acyltransferase (DGAT)
Phospholipid : Diacylglycerol Acyltransferase (PDAT)
TAG
OH
OH
1,2-DAG
P-Headgroup P-Headgroup
OH
TAG Phospholipid 2-lyso-phospholipid
+ +
Diacylglycerol Transacylase
OH
1,2-DAG
OH
1,2-DAG TAG
OH
2-monoacylglycerol
+ + OH
Three classes of plant DGAT • DGAT1 and DGAT2 are found in many eukaryotes and greatly differ in structure. • DGAT3 is soluble and has only been characterized in peanut seeds.
Enzymatic activity demonstrated in safflower seeds. The enzyme has yet to be isolated.
Identified in plants and yeast, PC appears to be the preferred substrate in plants while PE is the preferred substrate in yeast
(Li-Beisson Y et. al. (2010) Acyl-Lipid Metabolism. The Arabidopsis Book 8:e0133. doi:10.1199/tab.0133)
(Dahlqvist et al. (2000) Proc. Natl. Acad. Sci. USA 97: 6487 )
(Stobart et al. (1997) Planta 203: 58)
6

Schematic representations of murine DGAT1 and
DGAT2 proteins.
(Yen et al. (2008) Journal of Lipid Research 49: 2283)
DGAT1 and DGAT2 have different structures and may not have overlapping functions
Major DGAT classes involved in TAG synthesis in plants: • Arabidopsis: DGAT1 • Brassica napus: DGAT1 • Castor bean: DGAT2 • Tung Tree: DGAT2
• DGAT1 and DGAT2 localize to separate subdomains of the ER
7

Malonyl-CoA
Acyl-ACP (16:0;18:0)
Acyl-ACP (18:1)
Acetyl-CoA
ACCase
FAS Desaturase
PC
Pyruvate
LACS Acyl-CoA
Thioesterase
FA
G3P
LPA
PA
DAG
PLASTID
PAP
ACYL EDITING LPC
ENDOPLASMIC RETICULUM
FA SYNTHESIS
LIP
ID A
SSEM
BLY
FA MODIFICATION • Lipid linked desaturases
• Acyl-CoA elongases
DAG
TAG
TAG
Oil body
TAG
DGAT
PDAT
GPAT
LPAT
DGAT1 and PDAT1 are the only confirmed acyltransferases known in Arabidopsis TAG synthesis
8

Determination of the enzymes responsible for triacylglycerol synthesis in Arabidopsis
• DGAT1 mutant has ~20% reduction in seed TAG
(Katavic et al. (1995) Plant Physiology 108: 399)
•DGAT2 cannot complement yeast DGAT knockouts as DGAT1 does • http://arabidopsisacyllipids.plantbiology.msu.edu/enzymes/105
•PDAT1 mutant has no effect on amounts of seed TAG
(Mhaske et al. (2005) Plant Physiology and Biochemistry 43: 413)
•DGAT1/PDAT1 double mutant is pollen lethal
•dgat1/dgat1 + PDAT1 RNAi or pdat1/pdat1 + DGAT1 RNAi has 70-80% reduction in seed oil content
(Zhang et al. (2009) The Plant Cell 21: 3885)
9
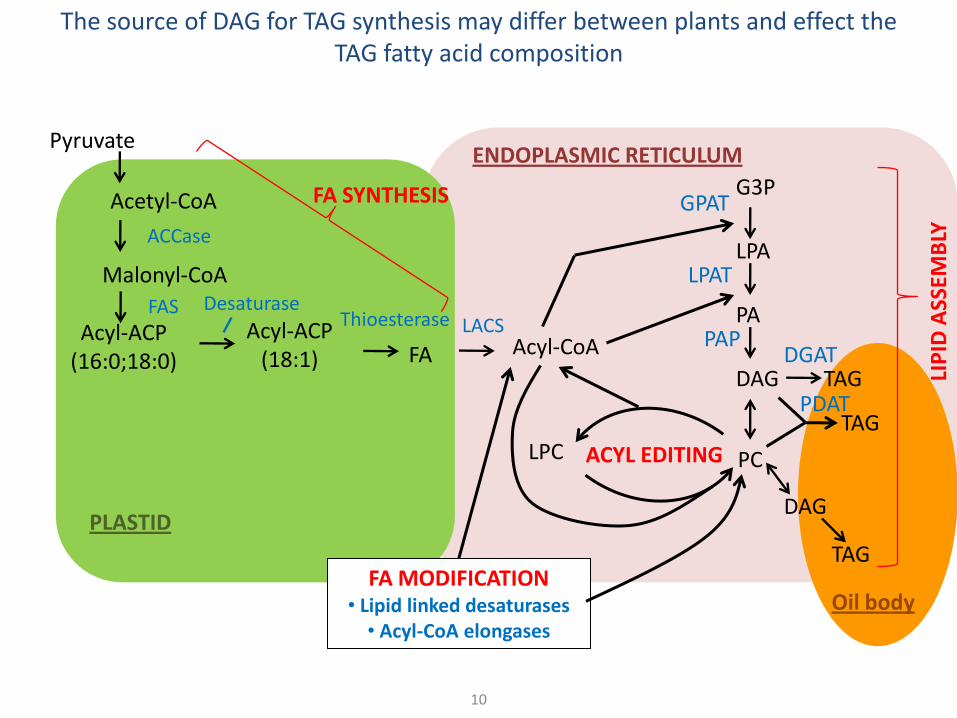
The source of DAG for TAG synthesis may differ between plants and effect the TAG fatty acid composition
Malonyl-CoA
Acyl-ACP (16:0;18:0)
Acyl-ACP (18:1)
Acetyl-CoA
ACCase
FAS Desaturase
PC
Pyruvate
LACS Acyl-CoA
Thioesterase
FA
G3P
LPA
PA
DAG
PLASTID
PAP
ACYL EDITING LPC
ENDOPLASMIC RETICULUM
FA SYNTHESIS
LIP
ID A
SSEM
BLY
FA MODIFICATION • Lipid linked desaturases
• Acyl-CoA elongases
DAG
TAG
TAG
Oil body
TAG
DGAT
PDAT
GPAT
LPAT
10

DAG source can affect TAG composition
P P P GPAT LPAAT PAP
G3P LPA PA de novo DAG
PC derived DAG
P
Cho PC
desaturation, hydroxylation,
acyl editing, etc.
Acyl-CoA pool
DGAT PDAT
TAG
DGAT PDAT
TAG
11

Reduced oleate desaturation mutnat (rod1) encodes an enzyme that affects the flux of DAG through PC into TAG
Phosphatidylcholine : Diacylglycerol CholinephosphoTransferase (PDCT)
P
Cho 1,2-DAG
P
Cho 1,2-DAG
+ +
(Lu et al. (2009) Proceedings of the National Academy of Sciences in the USA 106: 18837)
PC PC
12

Determine relative flux through competing TAG biosynthesis pathways with [14C]glycerol labeling
P P P GPAT LPAAT PAP
G3P LPA PA de novo DAG
DGAT PDAT
TAG
DGAT PDAT
PC derived DAG
TAG
P
Cho PC
CPT PDCT
PDCT
rCPT
PLC PLD/PAP
[14C]glycerol
13

P P P GPAT LPAAT PAP
G3P LPA PA de novo DAG
DGAT PDAT
TAG
DGAT PDAT
PC derived DAG
TAG
P
Cho PC
CPT PDCT
PDCT
rCPT
PLC PLD/PAP
[14C]glycerol
0
5
10
15
20
25
30
35
0 5 10 15 20 25 30 35
DP
M/
tota
l lip
id F
AM
E
PC DAG
TAG
Flux of glycerol through TAG synthesis in WT Arabidopsis (Bates and Browse (2011) The Plant Journal, in press)
14
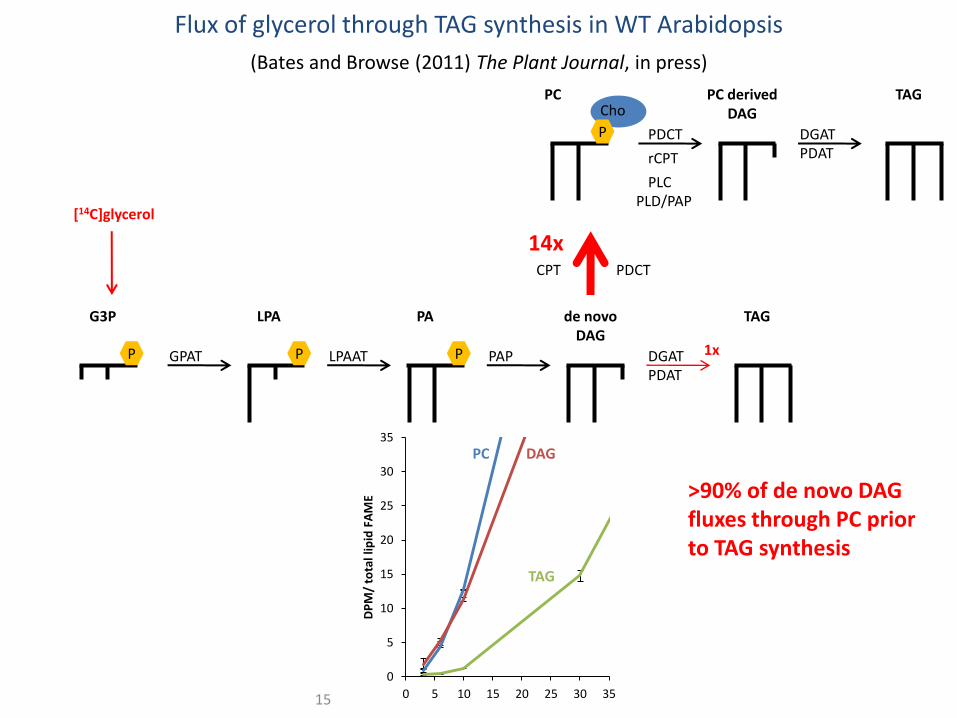
Flux of glycerol through TAG synthesis in WT Arabidopsis
P P P GPAT LPAAT PAP
G3P LPA PA de novo DAG
DGAT PDAT
TAG
DGAT PDAT
PC derived DAG
TAG
P
Cho PC
CPT PDCT
PDCT
rCPT
PLC PLD/PAP
[14C]glycerol
14x
1x
0
5
10
15
20
25
30
35
0 5 10 15 20 25 30 35
DP
M/
tota
l lip
id F
AM
E
PC DAG
TAG
>90% of de novo DAG fluxes through PC prior to TAG synthesis
(Bates and Browse (2011) The Plant Journal, in press)
15

Flux of DAG through PC for TAG synthesis may depend on plant species
Seed Oil content (weight %)
Arabidopsis yes ~20% ~37%
Castor bean No ~90% ~50%
Use of PC-derived DAG
Non-membrane lipid FA in TAG
Soybean yes no ~20%
Plant
(Bafor et al. (1991) Biochemical Journal 280: 507)
(Bates & Browse (2011) The Plant Journal, in press)
(Bates et al. (2009) Plant Physiology 150: 55)
16

World vegetable oil production: Four major crops
0
10
20
30
40
50
60
70
16:0 18:0 18:1 18:2(n-6) 18:3(n-3)
Pe
rce
nt
fatt
y ac
id Palm
0
10
20
30
40
50
60
70
16:0 18:0 18:1 18:2(n-6) 18:3(n-3)
Pce
cen
t fa
tty
acid
Soybean
0
10
20
30
40
50
60
70
16:0 18:0 18:1 18:2(n-6) 18:3(n-3)
Pe
rce
nt
fatt
y ac
id
Canola
0
10
20
30
40
50
60
70
16:0 18:0 18:1 18:2(n-6) 18:3(n-3)
Pe
rce
nt
fatt
y ac
id Sunflower
(http://lipidlibrary.aocs.org/market/fourmain.htm) (http://www.scientificpsychic.com/fitness/fattyacids1.html)
17

(http://www.soystats.com/)
World vegetable oil consumption 2010 146.9 Million Metric Tons
U.S. Fats & Oils Edible Consumption 2010 9.64 Million Metric Tons
U.S. Soybean Oil Consumption 2010 7.54 Million Metric Tons
~ US petroleum based Fuel consumption 2009 770 Million Metric Tons
Vegetable oil consumption
18

http://oilreset.com/blog/?p=159
Vegetable oil can replace petroleum for the chemical industry
19

Uses of vegetable oil and reasons for engineering fatty acid content
Food oils, “Healthy” oils
•Low in saturated fatty acids •High in ω-3 polyunsaturated fatty acids •Fish type oils
•EPA and DHA
Good cooking properties •Low polyunsaturated fatty acids •High in 18:1
Industrial applications •Medium-chain fatty acids (12:0): soaps, detergents, surfactants •Long-chain (22:1): Lubricants, slip agents •Epoxy fatty acids: Plasticizers, coatings, paints, •Hydroxy fatty acids: Lubricants, polymers
Engineering Strategies
• Adjust proportions of endogenous fatty acids in crop plants •Introduce new fatty acids into crop plants
20

Breeding changed rapeseed (Brassica napus) into Canola
CANOLA = CANadian Oil Low Acid
Natural isolates of a β–ketoacyl-CoA synthase used to remove Erucic acid (22:1) content
LEAR Low Erucic Acid Rapeseed HEAR High Erucic Acid Rapeseed
21

https://www.vistivegold.com/Pages/About.aspx
New GM soybeans with improved cooking oil properties from Monsanto about to hit the market
Vistive® Gold soybeans have advanced to Phase IV (pre-launch), the final step within the R&D pipeline. Monsanto has recently completed submissions to the USDA and FDA in support of the trait. The U.S. Food and Drug Administration has issued a positive response letter to Monsanto’s Generally Recognized as Safe notification. The FDA’s letter supports the use of Vistive® Gold under its intended uses.
• Higher oxidative stability • Reduced polymerization during frying
• No need for hydrogenation • Therefore no trans-fats
• Less saturated fat
22

Production of low saturate high oleic soybeans by Monsanto
Malonyl-CoA
Acyl-ACP (16:0;18:0)
Acyl-ACP (18:1)
Acetyl-CoA
ACCase
FAS Desaturase
PC
Pyruvate
LACS Acyl-CoA
Thioesterase
FA
G3P
LPA
PA
DAG
PAP
ACYL EDITING LPC
ENDOPLASMIC RETICULUM
FA SYNTHESIS
LIP
ID A
SSEM
BLY
DAG
TAG
TAG
Oil body
TAG
DGAT
PDAT
GPAT
LPAT
18:1-PC
18:2-PC
18:3-PC
FAD2
FAD3
FA MODIFICATION • Lipid linked desaturases
FATb
•Conventional breeding FAD3 mutant • FAD3: desaturase 18:2 to 18:3
•RNAi FAD2 • FAD2: 18:1 to 18:2 •Only wanted a semi KD therefore specifically targeted 2 of 4 FAD2 isoforms
•RNAi FATb •FATb: 16:0-ACP thioesterase
• Reduces 16:0 and 18:0 fatty acid release from fatty acid synthesis, and promotes production of more 18:1
23

Vernolic acid
Ricinoleic acid
Eleostearic acid
Uses
• Laxatives • Chemotherapy carrier • Lubricants • Polymers • Fuel additives
• Plastics • Adhesives • Paints
• Drying oils • Varnishes • Paints
Ricinus communis (Castor bean)
Vernonia galamensis (Iornweed)
Vernicia fordii (Tung tree)
Industrially useful fatty acids are found throughout the plant kingdom but not always in suitable crop plants
Agronomic features
•~50% oil by weight in seeds •Highly toxic co-product Ricin •Grows in the sub-tropics
Fatty acid
% unusual FA in oil
~90%
~80% •~40% oil by weight in seeds • only grows near the equator
~82%
•Slow growing tree •Only grows in the sub-tropics •Susceptible to hurricanes and condominium plantations
24

Ricinoleic acid (12-hydroxy-9-cis-octadecenoic acid)
Engineering of industrial oils in Arabidopsis: Castor oil a case study
P
Cho
P
Cho
RcFAH12 ~ 90% ricinoleic acid
Phosphatidylcholine (PC)
TAG
RcFAH12 expressed in develoing Arabidopsis seeds, ~17% hydroxy fatty acids (HFA) mostly at sn-2 (Broun and Somerville (1997) Plant Physiology 113: 922) (Smith et al. (2003) Planta 217: 507) Lu et al. (2006) The Plant Journal 45: 847)
25

Malonyl-CoA
Acyl-ACP (16:0;18:0)
Acyl-ACP (18:1)
Acetyl-CoA
ACCase
FAS Desaturase
Pyruvate
LACS Acyl-CoA
Thioesterase
FA
G3P
LPA
PA
DAG
PAP
ACYL EDITING LPC
FA SYNTHESIS
LIP
ID A
SSEM
BLY
RcFAH12 •Hydryoxylation of sn-2 PC
DAG
TAG
TAG
Oil body
TAG
RcDGAT
RcPDAT
GPAT
LPAT
HFA specific TAG biosynthetic increase accumulation of HFA in Arabidopsis TAG and gave clues on HFA metabolism in Arabidopsis (Burgal et al. (2008) Plant Biotechnology Journal 6: 819)
(van Erp et al. (2011) Plant Physiology 155: 683)
PC-HFA •Co-expression of RcFAH12 with Castor DGAT or PDAT increased seed HFA content from 17% to 25-28% • Both RcDGAT and RcPDAT lower the amount of HFA that accumulate in PC
• HFA still mostly at sn-2 + sn-3 26

P P P GPAT LPAAT PAP
G3P LPA PA de novo HFA-DAG
DGAT PDAT
TAG PC derived HFA-DAG
P
Cho PC
CPT PDCT
PDCT
rCPT
PLC PLD/PAP
RcFAH12 sn-2
hydroxylation
HFA-CoA F.A.S.
1. RcFAH12 produces sn-2 HFA PC which can be converted to sn-2 HFA TAG 2. HFA can also be utilized for de novo DAG synthesis
1. HFA-de novo DAG is turned over 3. In efficient utilization of HFA leads to inhibition of FA synthesis and reduced oil 4. HFA specific sn-3 acyltransferases allow HFA-TAG accumulation
Flux analysis to determine bottlenecks in HFA TAG synthesis
[14C]glycerol
(Bates and Browse (2011) The Plant Journal, in press)
27

1. Inhibit enzymes that turnover HFA-containing de novo DAG 2. Allow de novo DAG to flux directly to TAG
P P P GPAT LPAAT PAP
G3P LPA PA de novo HFA-DAG
DGAT PDAT
TAG PC derived HFA-DAG
P
Cho PC
CPT PDCT
PDCT
rCPT
PLC PLD/PAP
RcFAH12 sn-2
hydroxylation
HFA-CoA
Future HFA engineering strategies
28


















