
Factors affecting transduction efficiency of pseudotyped ...
104
Purdue University Purdue e-Pubs Open Access Dissertations eses and Dissertations 3-2016 Factors affecting transduction efficiency of pseudotyped viral vectors incorporating alphaviral glycoproteins Aditi Kesari Purdue University Follow this and additional works at: hps://docs.lib.purdue.edu/open_access_dissertations Part of the Genetics and Genomics Commons , and the Virology Commons is document has been made available through Purdue e-Pubs, a service of the Purdue University Libraries. Please contact [email protected] for additional information. Recommended Citation Kesari, Aditi, "Factors affecting transduction efficiency of pseudotyped viral vectors incorporating alphaviral glycoproteins" (2016). Open Access Dissertations. 666. hps://docs.lib.purdue.edu/open_access_dissertations/666
Transcript of Factors affecting transduction efficiency of pseudotyped ...

Purdue UniversityPurdue e-Pubs
Open Access Dissertations Theses and Dissertations
3-2016
Factors affecting transduction efficiency ofpseudotyped viral vectors incorporating alphaviralglycoproteinsAditi KesariPurdue University
Follow this and additional works at: https://docs.lib.purdue.edu/open_access_dissertations
Part of the Genetics and Genomics Commons, and the Virology Commons
This document has been made available through Purdue e-Pubs, a service of the Purdue University Libraries. Please contact [email protected] foradditional information.
Recommended CitationKesari, Aditi, "Factors affecting transduction efficiency of pseudotyped viral vectors incorporating alphaviral glycoproteins" (2016).Open Access Dissertations. 666.https://docs.lib.purdue.edu/open_access_dissertations/666

Graduate School Form30 Updated
PURDUE UNIVERSITYGRADUATE SCHOOL
Thesis/Dissertation Acceptance
This is to certify that the thesis/dissertation prepared
By
Entitled
For the degree of
Is approved by the final examining committee:
To the best of my knowledge and as understood by the student in the Thesis/Dissertation Agreement, Publication Delay, and Certification Disclaimer (Graduate School Form 32), this thesis/dissertation adheres to the provisions of Purdue University’s “Policy of Integrity in Research” and the use of copyright material.
Approved by Major Professor(s):
Approved by:Head of the Departmental Graduate Program Date
Aditi S. Kesari
Factors affecting Transduction Efficiency of Pseudotyped Viral Vectors Incorporating Alphaviral Glycoproteins.
Doctor of Philosophy
Peter J. HollenbeckChair
David A. Sanders
James F. Leary
Richard J. Kuhn
David A. Sanders
Jeffery Lucas 3/2/2016


FACTORS AFFECTING TRANSDUCTION EFFICIENCY OF PSEUDOTYPED
VIRAL VECTORS INCORPORATING ALPHAVIRAL GLYCOPROTEINS.
A Dissertation
Submitted to the faculty
of
Purdue University
by
Aditi Kesari
In Partial Fulfillment of the
Requirements for the Degree
of
Doctor of Philosophy
May 2016
Purdue University
West Lafayette, Indiana

ii
This Thesis is Dedicated to
P.P. Dr. Suchitdada Dattopadhye
And
My dear Son, Aariv Josyula

iii
ACKNOWLEDGEMENTS
Firstly, I would like to thank the Almighty for continuously showering blessings
on me. It is because of Your grace that I am able to write this today.
I would like express my sincere gratitude to my advisor, Dr. David Sanders for
giving me an opportunity to work in his lab and believing in me. I wouldn’t have been
able to fulfill my dream of pursuing a PhD, if it wasn’t for his support. His immense
knowledge, the thought provoking discussions we had, the freedom he gave me to
explore the field of science and the scientific insights he provided have always been a
source of inspiration. Above all, the friendly atmosphere that he maintained in the lab
made this journey thoroughly enjoyable.
The continuous support and guidance I received from my committee members,
Dr. Peter Hollenbeck, Dr. Richard Kuhn and Dr. James Leary, through all these years
was extremely valuable. Their suggestions and comments broadened the scope of my
research. I really appreciate the help they provided me at different stages of this journey.
I would like to thank all the current and past members of Dr. Sanders’ lab,
especially, Dr. Anita Ghosh, Jason Segura, Dr. C. Matthew Sharkey and Dr. Swapna

iv
Apte for their support and help. Also, it was great working with Dr. Oya Bulut, Sarah
Talamentez, Melanie Bumbalough and Tycho Jaquish. I would also like to thank all
graduate students, post-doctoral researchers and technicians in the Department of
Biological Sciences and the Structural Biology building, who gave me access to different
research facilities, helped me with different techniques, advised me on presentation skills,
etc. I am especially grateful to Dr. Rushika Perera, Dr. Joyce Jose, Dr. Thomas Edwards
and other members from Dr. Kuhn’s laboratory for their guidance and help. I would also
like to thank Dr. Ronald Hullinger, Dr. Laurie Jaeger and other faculty members from
department of Basic Medical Sciences for their support.
My sincere thanks to all my friends and well-wishers, who made these years of
my life so memorable. I sincerely thank Pravinsinh Wagh, Jalaja Padma, Prasanth
Tanikella, Sanmit Ambedkar, Sudhir Bangaru, Chaitra Chavali, Harish Suryanarayana
and Deenal Mannava for helping me during a challenging phase of my life.
Finally I would like to thank my entire family. My father, Shashikant Kesari who
taught me no dream is too big to achieve, my mother, Shubhada Kesari who imbibed in
me the strength and the confidence to achieve those dreams, my brother, Amit Kesari,
who showed me the right path to achieve those dreams, I am forever indebted to them.
Last but not the least, my husband, Aditya Teja Josyula, you have been God’s best gift to
me. Thank you for being my best friend, teacher, mentor, confidant, my audience, my
editor, my ‘tech-guy’, but beyond everything the best partner. Thank you for everything.
Looking forward to spending many more sweet and memorable moments with you.

v
TABLE OF CONTENTS
Page
LIST OF TABLES ........................................................................................................... viii
LIST OF FIGURES ........................................................................................................... ix
LIST OF ABBREVIATION .............................................................................................. xi
ABSTRACT ..................................................................................................................... xiv
CHAPTER 1. GENE THERAPY/ TRANSFER VECTORS AND RETROVIRUSES ..... 1
1.1 Introduction to Gene Therapy ................................................................................... 1
1.2 Gene delivery vectors ................................................................................................ 4
1.3 Retroviruses ............................................................................................................... 5
1.4 Moloney Murine Leukemia Virus (MoMuLV) ......................................................... 6
1.5 Replication cycle of MoMuLV ................................................................................. 8
1.6 Limitations of retroviruses as gene therapy vectors ................................................ 10
CHAPTER 2. ALPHAVIRUSES AND PSEUDOTYPING ............................................ 12
2.1 Introduction ............................................................................................................. 12
2.2 Structure of alphaviruses ......................................................................................... 12

vi
Page
2.3 Replication cycle of Alphaviruses........................................................................... 15
2.4 Pseudotyping ........................................................................................................... 18
2.5 Generation of pseudotyped viruses ......................................................................... 19
CHAPTER 3. MATERIALS AND METHODS .............................................................. 22
3.1 Cell lines and cell culture ........................................................................................ 22
3.2 Plasmids .................................................................................................................. 23
3.3 Production of virus by transient expression of envelope glycoproteins .................. 24
3.4 Transduction assay .................................................................................................. 25
3.5 Immunoblot assay ................................................................................................... 25
3.6 Immunofluorescence assay ..................................................................................... 26
3.7 Addition of Heparinase I to Producer cells ............................................................. 27
3.8 Addition of Heparinase I to target cells .................................................................. 27
3.9 Normalization following quantitative analysis of the immunoblot assay ............... 27
3.10 Transduction assays in cells treated with lipid-lowering drugs ............................ 28
3.11 Production of baculovirus virus pseudotyped with alphavirus envelope
glycoproteins ................................................................................................................. 29
3.12 Transduction assay in C6/36 cells grown in sterol-free conditions ...................... 29
3.13 Transduction of ASMase-/- cells........................................................................... 29

vii
Page
3.14 Verification of cholesterol concentrations in ASMase-/- cells and ASMase+/+
cells................................................................................................................................ 30
CHAPTER 4. ROLE OF HEPARAN SULFATE IN ENTRY AND EXIT OF ROSS
RIVER VIRUS GLYCOPROTEIN-PSEUDOTYPED RETROVIRAL VECTORS ...... 31
4.1 Summary ................................................................................................................. 31
4.2 Introduction ............................................................................................................. 32
4.3 Results ..................................................................................................................... 38
4.4 Discussion ............................................................................................................... 53
CHAPTER 5. ROLE OF CHOLESTEROL IN ENTRY OF PSEUDOTYPES
INCORPORATING ALPHAVIRAL GLYCOPROTEINS AND NIEMANN PICK’S
DISEASE .......................................................................................................................... 57
5.1 Summary ................................................................................................................. 57
5.2 Introduction ............................................................................................................. 58
5.3 Results ..................................................................................................................... 62
5.4 Discussion ............................................................................................................... 70
REFERENCES ................................................................................................................. 72
VITA ................................................................................................................................. 85

viii
LIST OF TABLES
Table Page
1.1 Viral Vectors ................................................................................................................ 5
4.1 Paired T-test ............................................................................................................... 46
4.2 Pearson Coefficient BHK .......................................................................................... 49
4.3 Pearson Coefficient CHO22 ...................................................................................... 49
4.4 Pearson Coefficient CHO18. ..................................................................................... 49

ix
LIST OF FIGURES
Figure Page
Figure 1.1 Types of Gene Therapies ................................................................................... 2
Figure 1.2 MoMuLV particle .............................................................................................. 7
Figure 1.3 MoMuLV genome ............................................................................................. 7
Figure 1.4 Gag polyprotein ................................................................................................. 8
Figure 1.5 Replication cycle of MoMuLV ....................................................................... 10
Figure 2.1 Structure of Alphaviruses ................................................................................ 13
Figure 2.2 Alphaviral genome .......................................................................................... 14
Figure 2.3 Alphaviral Structural Proteins ......................................................................... 15
Figure 2.4 Replication cycle of alphaviruses .................................................................... 17
Figure 2.5 Pseudotyping ................................................................................................... 21
Figure 4.1 Structure of Heparan Sulfate ........................................................................... 35
Figure 4.2 RRV-E2 Envelope glycoprotein amino-acid sequence ................................... 36
Figure 4.3 RRV-E2 Envelope glycoprotein and HS ......................................................... 37
Figure 4.4 T216R-RRV and N218R-RRV pseudotypes have lower transduction titers .. 39
Figure 4.5 T216R-RRV and N218R-RRV envelope glycoproteins are inefficiently
incorporated in pseudotyped virus .................................................................................... 41

x
Figure Page
Figure 4.6 Intracellular retention of T216R-RRV and N218R-RRV in HS-expressing
cells ................................................................................................................................... 43
Figure 4.7 Addition of Heparinase I to Producer ØNXnlslacZ cells releases T216R-RRV
and N218R-RRV pseudotyped viruses ............................................................................. 46
Figure 4.8 Heparinase treatment of target cells reduces transduction by T216R-RRV and
N218R-RRV pseudotyped viruses .................................................................................... 48
Figure 4.9 T216R-RRV and N218R-RRV pseudotyped viruses possess higher
transduction efficiencies when transductions by viruses are normalized by glycoprotein
content ............................................................................................................................... 51
Figure 5.1 Cholesterol and Sphingolipid rich lipid rafts .................................................. 61
Figure 5.2 Alphavirus-MoMuLV pseudotypes require cholesterol for entry in mammalian
cells ................................................................................................................................... 64
Figure 5.3 Alphavirus-baculovirus pseudotypes require cholesterol for entry in insect
cells ................................................................................................................................... 66
Figure 5.4 Transduction efficiency of alphaviral pseudotypes is higher in ASMase-/- cells
........................................................................................................................................... 69

xi
LIST OF ABBREVIATION
SCID Severe Combined Immuno-deficiency syndrome
ADA
Adenosine deaminase
OTC Ornithine transcarbamylase
MoMuLV Moloney Murine Leukemia Virus
RRV Ross River Virus
HIV Human immunodeficiency virus
DNA Deoxyribonucleic acid
RNA Ribonucleic acid
RA Reverse transcriptase
IN Integrase
PR Aspartic protease
SU Surface unit
TM Transmembrane domain
PBS Primer binding site
PPT Polypurine tract
MA Matrix domain
CA Capsid

xii
NC Nucleocapsid
ER Endoplasmic reticulum
ESCRT Endosomal sorting complexes required for transport
SFV Semliki forest virus
SINV Sindbis virus
VEEV Venezuelan equine encephalitis virus
EEEV Eastern equine encephalitis virus
ORF Open reading frames
CMV Cytomegalovirus
LTR Long terminal repeats
BHK Baby hamster kidney cells
CHO Chinese hamster ovary cells
DMEM Dulbecco’s Modified Eagle’s Medium
FBS Fetal bovine serum
ASMase Acid sphingomyelinase
ASMase-/- cells Acid sphingomyelinase deficient cells
PBS Phoshate buffered saline
VSV Vesicular stomatitis virus
TU/mL Transduction Units/ml
SDS-PAGE Sodium dodecyl sulfate polyacrylamide gel electrophoresis
ß-mCD Methyl-beta-cyclodextrin
HS Heparan sulfate

xiii
FIV Feline immunodeficiency virus
SD Standard deviation
HSV Herpes Simplex Virus
NPD Niemann Pick’s disease

xiv
ABSTRACT
Kesari, Aditi PhD. Purdue University, May 2016. Factors Affecting Transduction Efficiency of Pseudotyped Viral Vectors Incorporating Alphaviral Glycoproteins. Major PI: David A. Sanders
The genome of an organism has the complete set of biochemical instructions
required for sustenance of life. Mutations or abnormalities in this genome lead to genetic
disorders. Currently available therapeutic options mostly focus on treating the symptoms,
but not curing them. Gene therapy promises to be a curative form of medicine. In gene
therapy cells carrying a defective gene are targeted and replaced with a healthy copy of
that gene. The vehicles used for delivering this gene are known as vectors. Retroviruses
are popularly used gene therapy/transfer vectors. However, retroviruses are limited in the
range of cells they can enter and infect. The range of cells targeted by the viral vector can
be either expanded or narrowed by replacing the envelope of the virus with the envelope
of another virus that has the desired range of tissue tropism. Such hybrid viruses are
called pseudotyped viruses. Previously, we have pseudotyped Moloney Murine Leukemia
Virus (MoMuLV), a retrovirus with the envelope of Ross River Virus (RRV), an
alphavirus. Here, we substituted amino-acid residues of RR-envelope glycoprotein with

xv
basic amino-acid residues. We show that this makes the pseudotyped virus utilize
heparan sulfate, a ubiquitous molecule present on the surfaces of most cells, as an
attachment factor. Attachment to cellular heparan sulfate helps in concentration of virus
particles on the cell surface and thus enhancing their chances of cell entry, thereby
increasing their transduction efficiency of this viral vector. The same affinity towards
heparan sulfate, however, poses a challenge in terms of release of this pseudotyped virus
from the producer cells. General principles concerning viral adaptation to the use of
attachment factors and improving pseudotyped virus titers through modifying cell
membrane components can be derived from these results. We also show that alphavirus-
glycoprotein retroviral pseudotypes require cholesterol for entry into the cells.
Furthermore, excess cholesterol in cells facilitates the entry of alphavirus-glycoprotein
retroviral pseudotypes. We show that alphavirus-glycoprotein pseudotypes transduce the
acid sphingomyelinase deficient (ASMase-/-) cells derived from Niemann Pick’s disease-
model mice more efficiently than the cells from healthy mice (ASMase+/+). Thus
alphavirus-glycoprotein pseudotypes hold a potential as suitable vectors for gene therapy/
transfer in Niemann Pick’s Disease-Type A, a genetic lipid-storage disorder.

1
CHAPTER 1. GENE THERAPY/ TRANSFER VECTORS AND RETROVIRUSES
1.1 Introduction to Gene Therapy
The genome of an organism has the complete set of biochemical instructions
required for sustenance of life. If this genome carries any abnormality or mutation, it
leads to genetic disorders. Currently available therapeutic options mostly focus on
treating the symptoms but not curing them. Gene therapy promises to be a curative form
of medicine.
In gene therapy, cells carrying a defective gene are targeted and replaced with a
healthy copy of that gene. The gene delivery can be achieved by administering genetic
material into the body. This type of gene therapy is referred to as in vivo gene therapy
(Figure 1.1). Gene delivery can also take place outside the body, where affected cells are
removed from the body, delivered with genetic material followed by re-administering the
cells back in the body. This type of gene therapy is termed as ex vivo gene therapy
(Figure1.1).

2
Figure 1.1 Types of Gene Therapies
In ex vivo gene therapy, defective cells are removed from the body and cultured. The genetic material is introduced into the cells outside the body, before re-injecting the cells back into the body. In in vivo gene therapy the corrective gene is directly administered into the body.
Genetic material can be delivered either to the somatic (non-reproductive) cells or
to the germline cells. The effect of delivery of genetic material to the somatic cells
doesn’t pass on to the next generation, whereas the effect of gene delivery in germline
cells is carried on to the next generation. Thus, the therapeutic effect of germline gene
therapy, unlike somatic gene therapy, is permanent. However one potential hazard of the
former method is that the negative effects can also be permanently passed down to
subsequent generations (www.genetherapynet.com/viral-vectors.html n.d.).
Gene therapy to cure human diseases has been proposed since a long time. In
spite of advancement in this field, currently there is no approved treatment available. The
first attempt at gene therapy to treat diseases in humans was done in 1990. A 4 year old

3
girl suffering from Severe Combined Immuno-deficiency syndrome (SCID), a
monogenetic disease, which is caused due to the deficiency of adenosine deaminase
(ADA), was treated at NIH Clinical Center through an ex vivo gene therapy approach.
Her white blood cells were removed from her body and a healthy gene expressing ADA
was introduced into them. These cells were re-administered into her body (Blaese R
1995).
Multiple gene therapy/transfer studies were performed until late nineties. In 1999,
this field suffered a major set-back when a patient named Jesse Gelsinger, who had
participated in a clinical trial, died within a week of receiving the treatment. He had
deficiency of ornithine transcarbamylase (OTC), one of the enzymes required for
converting excess nitrogen in blood into urea. The gene expressing OTC was
administered using adenovirus vector. His body invoked a major immune reaction in
response to the high dosage of the virus (P. N.-H. Wirth T 2013). The failure of this trial
resulted in withdrawal of many studies.
Despite this, there have been many studies that have shown encouraging results
such as SERCA gene therapy for cardiac failure (Periasamy M 2008), gene therapy for
X-linked Severe Combined immunodeficiency syndrome (Hacein-Bey-Abina S 2010),
etc. China was the first country to approve a gene therapy product for therapeutic use (P.
N.-H. Wirth T 2013). However, this approval wasn’t backed up by clinical trial data.
Cerepro, an adenoviral vector carrying thymidine kinase gene, is the first gene therapy
product used for treating malignant brain tumors to have passed phase III clinical trial (S.
H.-H. Wirth T 2009). Glybera, an adeno-associated viral vector delivering lipoprotein

4
lipase gene, is the first gene therapy product to be recommend by European Medicines
Agency for approval in Europe (P. N.-H. Wirth T 2013). Given the recent advancements
in this field, gene therapy definitely seems to be an attainable therapeutic option in the
near future.
1.2 Gene delivery vectors
The vehicles used for delivering the genes are known as vectors. The gene
therapy/transfer vectors are broadly classified into viral and non-viral vectors. Liposomes
and naked DNA delivery have been used for non-viral gene delivery. Even though these
physical methods tend to be safer and less immunogenic than viral vectors, viral vectors
have proven to be efficient gene delivery vectors as compared to the non-viral ones (Li
SD 2006).
There are many viruses that can be used as gene delivery vectors. Table 1.1
discusses the properties of a few such viruses (www.genetherapynet.com/viral-
vectors.html n.d.).

5
Table 1.1 Viral Vectors
Virus Retrovirus/ lentivirus
Adenoviruses Adeno-associated
Herpesvirus
Family Retroviridae Adenoviredae Parvoviridae Herpesviridae
Types of cells entered
Dividing and non-dividing (only lentivirus)
Dividing and non-dividing
Dividing and non-dividing
Dividing and non-dividing
Integration with the host genome
Yes No Yes (lower frequency)
No
Expression of transgene
Long lasting Transient Long lasting Transient
Packaging capacity
8kb 7.5kb 4.5kb 30kb
Immune response
Low (moderate for lentivirus)
High Low Low
1.3 Retroviruses
Retroviruses are a large group of viruses responsible for causing
immunodeficiency syndromes and leukemias in vertebrates. These viruses contain a
single-stranded positive-sense RNA genome, which through the process of reverse
transcription forms a DNA intermediate. This DNA intermediate can integrate with the
host genome leading to formation of a provirus. Retroviruses are transmitted by coming
in close contact with infected animal, exposure to body fluids of infected animals,

6
maternofetal transmission, accidental inoculation and experimental infection (Coffin JM
1997). Retroviruses are increasingly being used as research tools in cell and molecular
biology.
Retroviruses like Human immunodeficiency virus (HIV), Moloney murine
leukemia virus (MoMuLV) etc. are used as vectors of gene therapy because they allow
fast and stable transfer of genetic material to cells, and have strong promoter system that
results in efficient and long lasting expression of genetic material. However, the host
range and tissue tropism of this viral vector are limited.
1.4 Moloney Murine Leukemia Virus (MoMuLV)
MoMuLV (gammaretrovirus genus) is one of the simplest retroviruses and hence is used
as a model for studying retroviruses. The MoMuLV is a polymorphic virion around 100-
120 A˚ in size (Figure 1.2) (Yeager M 1998). Two strands of single-stranded RNA
genome are incorporated into the virus particle. The viral genome has a coding region
and a non-coding region (Figure 1.3). Coding region has genes gag-pol-env that code for
the three polyproteins, Gag (Figure 1.4), Pol and Env respectively. These are involved in
assembly of the virion. The capsid of the virion is formed from Gag while the envelope is
developed from the Env polyprotein and the lipid bilayer of the host. The three enzymes-
reverse transcriptase (RA), integrase (IN) and aspartic protease (PR) are derived from the
Pol polyprotein (Rein 2011).

7
Figure 1.2 MoMuLV particle
The outermost layer is formed from Env glycoproteins and host-derived lipid bilayer. The Env glycoprotein is made up of the surface unit (SU), which contais the receptor binding domain and transmembrane (TM) unit, which has the fusion peptide. The lipid bilayer and the embedded glycoproteins constitute the envelope of the virus particle. Inside, lies the core made up of the Gag polyprotein. Within the core there are two strands of single-stranded RNA genome. Along with that, there is Pol, the polyprotein which comprises of three enzymes: (i) reverse transcriptase, which converts the viral RNA genome into a DNA copy, (ii) viral aspartic protease, which leads to maturation of the virion by cleaving the polyproteins into their respective domains and (iii) integrase, which processes the DNA copy of viral genome and inserts it into the host genome to form the provirus. (Figures from Rein A, Advances in Virology, 2011 and Silverman RH et al., Nature Reviews urology, 2010) (Rein 2011) (Silverman RH 2010)
Figure 1.3 MoMuLV genome
The coding region of the viral genome includes the gag, pol and env regions that encode Gag, Pol and Env polyproteins respectively. The non-coding region has a LTR sequence, which has all the promoters and enhancers, the psi (Ψ) packaging sequence, a primer binding site (PBS) and the polypurine tract (PPT), which acts a primer for synthesis of second strand of DNA (Shinnick TM 1981)

8
Figure 1.4 Gag polyprotein
Gag polyprotein is made up of a matrix (MA) domain, which lies on the outer side of the core and associates with the lipid bilayer through myristylation, a small proline rich p12 region, which is thought to promote the release of virus particles, a capsid (CA) domain and a nucleocapsid (NC) domain which lies on the inner side and is in association with the RNA genome (Rein 2011, Shinnick TM 1981)
1.5 Replication cycle of MoMuLV
MoMuLV infects the cell by receptor-mediated membrane fusion (Figure 1.5). The SU
subunit of the envelope glycoprotein of the virus binds to the cell surface receptor. This
causes structural change in the SU subunit which leads to disruption of the disulfide bond
between the SU and TM subunits (Freed EO 1987, Pinter A 1997), exposing the fusion
peptide of the TM subunit. Fusion between the viral and cell membranes releases the
capsid into the cytoplasm which later disintegrates and the viral RNA escapes into the
cytoplasm. The RNA genome is copied into DNA by reverse transcriptase. This DNA
enters the nucleus and gets inserted into the host genome with the help of viral integrase
to form a provirus. The integrated DNA is transcribed into mRNA which then travels out
of the nucleus. In the cytoplasm it is translated into the assembly proteins. Envelope
glycoproteins are synthesized from spliced mRNA and translocated to ER. Glycosylation
takes place in the Golgi along with the furin cleavage of Env polyprotein into the SU and
TM subunits. The SU-TM heterodimer is linked with a disulfide bond. Three
heterodimers come together to form trimers and these are then transported to the
membrane. Gag-Pol polyproteins, which are synthesized together as fusion protein co-

9
assemble at the membrane. Myristylation of the N terminal of Gag targets it to the plasma
membrane (Rein A 1986). An alternative glyco-Gag is also synthesized and is thought to
target the virus assembly towards lipid rafts (Nitta T 2011). The psi packaging sequence
helps the viral RNA to compete against the cellular RNA to be incorporated in the
budding virus. The late motif (p12) of Gag interacts with the ESCRT machinery, which is
involved in the budding process of the assembled virus (Rein 2011, Pornillos O 2002).
One of the cleavage products of the Pol polyprotein, the viral aspartic protease (PR),
plays a role in maturation of the virus particle as it cleaves the Gag molecule into its
respective subunits. The maturation step is crucial for the infectivity of the virus (Coffin
JM 1997).

10
Figure 1.5 Replication cycle of MoMuLV
Retrovirus entry is initiated when the virus particle binds to the cell surface receptor. Fusion between the viral and cell membrane releases the capsid into the cytoplasm. The viral RNA escapes into the cytoplasm upon disintegration of the capsid. RNA is copied by into DNA by reverse transcriptase. This DNA enters the nucleus and gets inserted in the host genome with help of viral integrase to form a provirus. The integrated DNA is transcribed into mRNA. mRNA is gets transported to the cytoplasm, where it is translated into the assembly proteins. Envelope proteins are synthesized from spliced mRNA and translocated to ER. Gag-Pol polyprotein which are synthesized together as fusion proteins co-assemble at the membrane. The psi sequence helps in packaging of the viral RNA into compete the budding virus. Viral aspartic protease cleaves the Gag molecule into its respective subunits. This step of maturation of the virus is crucial for its infectivity.
1.6 Limitations of retroviruses as gene therapy vectors
The retroviruses have three major limitations as gene therapy/transfer vectors.
Firstly, the host range and tissue tropism are not very extensive. This limits the types of
cells they can enter into. Secondly, there is a chance of recombination and formation of

11
replicating and competitive virus particles capable of causing active infection in
individuals receiving the treatment (Anson 2004). And finally, clinical trials in the past
have shown instances of insertional mutagenesis leading to oncogenesis (Anson 2004).
Pseudotyping of the virus can be used to overcome the first two limitations. It is a
process in which the envelope protein of the virus is replaced with an envelope protein of
other virus. Previously, we have pseudotyped retroviruses by replacing its envelope with
the envelope of alphaviruses in our lab. The details about alphaviruses and process of
pseudotyping are discussed in the subsequent chapter.

12
CHAPTER 2. ALPHAVIRUSES AND PSEUDOTYPING
2.1 Introduction
Alphaviruses are arthropod-borne viruses that cause severe infections in a wide
range of vertebrates. The clinical manifestations in infected humans include fever, rash,
arthralgia, myalgia, headaches and polyarthritis (Strauss J H 1994). Alphaviruses belong
to the Togaviridae Family and are classified into two categories: (i) Old World viruses
which include Semliki forest virus (SFV), Sindbis virus (SINV) and Ross River virus
(RRV) (ii) New World viruses which include Venezuelan equine encephalitis virus
(VEEV), Eastern equine encephalitis virus (EEEV).
2.2 Structure of alphaviruses
An alphavirus is an enveloped, single-stranded, positive-sense, RNA virus
(11.5kb) (Figure 2.1) (Strauss J H 1994). The RNA genome (Figure 2.2) is surrounded by
the capsid, which is made up of 240 units of capsid protein. This is in turn is surrounded
by an envelope, which is a lipid bilayer membrane with 80 spikes of envelope
glycoprotein embedded into it (Figure 2.3). E1, E2 and E3 are the three envelope
glycoproteins. Each spike is a trimer of an E1/E2 heterodimer (Cheng RH 1995).
Whereas E2 is required for binding to cell-surface receptors, E1 is required for fusion of
the viral and cellular membrane. The envelope and the inner nucleocapsid core have

13
icosahedral symmetry with triangulation number T=4 (Strauss J H 1994, Cheng RH
1995). The E2/E1 heterodimer has an one-one association with each nucleocapsid
monomer via the C-terminal of E2 (Cheng RH 1995).
Figure 2.1 Structure of Alphaviruses
Alphaviruses have 80 spikes of envelope glycoproteins (blue) embedded in the lipid bilayer (green), which forms its envelope. Inside this, lies the nucleocapsid core (yellow and red) made up of 240 units of Capsid proteins and a single strand of a positive-sense RNA genome. Figure adapted from Cheng et al., Cell, 1995 (Cheng RH 1995).

14
Figure 2.2 Alphaviral genome
An alphavirus has a single-stranded, positive-sense RNA genome. The nonstructural proteins are translated from the genomic RNA and are involved in replication of the genome, whereas the structural proteins are translated from the subgenomic RNA, with a promoter as shown in the figure.

15
Figure 2.3 Alphaviral Structural Proteins
Alphaviral structural proteins are expressed as polyproteins. The Capsid protein © cleaves itself after getting translated (Arrow 1). The N terminus of the remaining portion of polyprotein (envelope glycoproteins) has a signal sequence due to which it translocates into the endoplasmic reticulum, while another signal sequence anchors it in the membrane. A proteolytic cleavage after this signal peptide releases E3-E2 (pE2) (Arrow 3) (Garoff H 1990). Another proteolytic cleavage at C terminal of heavily palmitoylated 6kD segment (Arrow 4), releases E1 which is then anchored by stop-transfer signal. Cleavage between E3 and E2 takes place in trans-golgi and is mediated by furin-like protease (Arrow 2)
2.3 Replication cycle of Alphaviruses
The virus enters the cell via receptor-mediated endocytosis (Figure 2.4) (DeTulleo L
1998). The presence of a cholesterol and sphingolipid rich bilayer has been shown to be
required for entry of SINV and SFV (H. A. Kielian MC 1984, Phalen T 1991, Lu YE
1999). Once inside the endosome, the acidic pH leads to dissociation of the E1-E2
heterodimer complex exposing the hidden fusion loop of the E1 glycoprotein. Insertion of
the fusion loop into the endosomal membrane is followed by formation of E1-homotrimer
complex (H. A. Kielian MC 1985, Boggs WM 1989, Justman J 1993, Li L 2010). This

16
fusion process releases nucleocapsid core into the cytoplasm. Disassembly of the capsid
releases he RNA genome, which is translated from two ORFs to form (I) the non-
structural polyprotein (P1234), which is required for the replication of the genome (early
phase) and (II) the structural polyprotein (late phase). The positive strand is first
converted to a negative strand of RNA, which is predominant during early phase. This
negative strand, during the late phase of replication cycle is either replicated into full-
length positive genomes, which are incorporated into new virion or it is spliced into
subgenomic positive-sense RNA. The subgenomic RNA is translated into structural
polyprotein (C-pE2-6K-E1) (Jose J 2009) (Leung JY 2011). Upon cleavage, Capsid
protein starts oligomerizing and then associates with the single-stranded RNA genome to
form the nucleocapsid (Tellinghuisen TL 1999). The cleavage of Capsid protein is
followed by the processing of the remaining polypeptide, which now consists of envelope
glycoproteins. The N-terminus of this polypeptide has signal sequence which helps in
translocation of the polypetide into the ER. A 30 residue-long signal sequence anchors C-
terminus of the polypeptide. A proteolytic cleavage after this signal peptide releases E3-
E2 (pE2) (Garoff H 1990). Another proteolytic cleavage takes place in remaining the
polypeptide between the hydrophobic 6K and E1 protein (Liljeström P 1991). The E1 and
pE2 undergo complex folding with the help of chaperones. Disulfide bond formation
takes place to form the pE2-E1 heterodimer in the ER (Anthony RP 1992). Post-
translational modifications like glycosylation, palmitoylation in case of E2 follow this
(Sefton 1977). Once the heterodimer reaches trans-golgi, furin-like protease cleaves E3
from E2 (Gaedigk-Nitschko K 1990). This cleavage is essential for entry of virus
(Salminen A 1992 ). The glycoproteins are transported to the plasma membrane where

17
the virions are assembled. Interaction between nucleocapsid unit and E2 takes place and
budding occurs (Cheng RH 1995).
Figure 2.4 Replication cycle of alphaviruses
Alphaviruses enter the cell via receptor mediated endocytosis. The process of fusion is triggered by the acidic pH in the endosomes. The RNA gnome is released in the cytoplasm and translated into non-structural polyproteins, which are required for replication of the genomic RNA, and structural polyproteins. The capsid proteins, which are derived from the structural polyproteins and the genomic RNA form nucleocapsid cores, which are transported to the plasma membrane. Envelope glycoproteins, which are derived from the structural polyprotein, reach the plasma membrane through secretory pathway. The assembly of the virus particle takes place at the plasma membrane, which is followed by budding. Figure adapted from Schwartz et al., Nature Reviews Microbiology, 2010 (Schwartz O 2010).

18
2.4 Pseudotyping
Viruses are commonly used as vectors to deliver a desired gene to target cells for
the purpose of gene therapy or gene transfer. For gene therapy/transfer, one either needs
to expand the host range of the gene delivery vector or narrow it down to a specific
target. The recognition and infection of host cells by viruses depends on their envelope
proteins. Hence by incorporating the envelope proteins of other viruses, the ability to
enter cells other than those a virus normally infects can be conveyed to this parent virus.
The chimeric virus having the desired host range/tissue tropism thus formed is called a
pseudotyped virus (Sanders 2002).
Pseudotyped viruses have many experimental and clinical applications. Gene
therapy/transfer is one of the important applications of these pseudotyped viruses. Based
on the therapeutic need, the range of cells targeted by the viral vector for gene
therapy/transfer can be modified by pseudotyping the virus with the envelope of another
virus (Sanders 2002). Apart from their utility as gene-delivery vectors, pseudotyped
viruses can also be used as tools to study the entry of enveloped viruses.
Retroviruses, such as human immunodeficiency virus (HIV), feline
immunodeficiency virus (FIV), and Moloney murine leukemia virus (MoMuLV) are used
as gene-delivery vectors, because they allow fast and stable transfer of genes under the
influence of strong promoters to cells (Naldini L 1996, Kafri T 1997, Poeschla EM 1998,
Noh MJ 2010). However, they have limited host range and tissue tropism. For the
treatment of systemic genetic disorders a viral vector needs to be pseudotyped with an
envelope of a virus that has a broad tissue tropism. Since RRV, an alphavirus, has wide
host range and tissue tropism, it was incorporated in ØNX pseudotyped system to

19
produce RRV-MoMuLV pseudotyped virus (Sharkey CM 2001). Recombinant
retroviruses and lentiviruses bearing alphavirus glycoproteins have been shown to be
efficient gene transfer/therapy vectors (Sharkey CM 2001, Kang Y 2002). Previous
studies showed that pseudotyping with RRV was far less cytotoxic but equally efficient
as pseudotypes incorporating envelope glycoproteins of Vesicular Stomatitis virus (VSV-
G), a prototype pseudotyped virus (Kang Y 2002). Retroviruses pseudotyped with RRV
envelope glycoproteins were successfully used for in vivo gene-transfer studies in
hepatocytes, glial cells, muscle tissue and airway epithelial tissue (Kang Y 2002).
2.5 Generation of pseudotyped viruses
Producer ØNXnlslacZ cells (a second generation 293-T based retroviral packaging cell
line that is stably transfected with a MoMuLV gag-pol plasmid and the MFG.S-nlslacZ
plasmid, which produces envelope-protein-deficient, replication-incompetent MoMuLV
particles and encodes nuclear-localized β-galactosidase) are transfected with plasmid
encoding for alphaviral glycoproteins to generate alphaviral-MoMuLV pseudotypes
(Figure 2.5) (Sharkey CM 2001). The gag-pol plasmid expresses the core proteins of
MoMuLV under the influence of RSV promoter. The MFG.S-nlslacZ plasmid has the
gene of choice, which for experimental purposes is lacZ, along with the nuclear
localizing sequence (nls), LTR and psi (Ψ) sequence. The Ψ sequence helps in packaging
of LacZ into the budding pseudotyped virus. These naked nucleocapsid cores however,
are incapable of transducing the cells and require an envelope to do so. Hence these
producer ØNXnlslacZ cells are transfected with plasmid encoding the envelope protein
of other viruses, such as alphaviruses. The plasmid encoding for alphaviral glycoproteins

20
is expressed under the influence of CMV promoter. The retroviral promoter in the LTR
region is required for in expression of LacZ gene into β-galactosidase. β-galactosidase
then gets translocated to the nucleus under the effect of nuclear localizing sequence.
The pseudotyped viruses generated using this method are non-infectious and
replication incompetent. By replacing the MFG.S-nlslacZ gene with a vector-expressing
gene they can be used as efficient gene-delivery agents. Since the three plasmids involved
in formation of these pseudotypes have three different promoters, the chances of
recombination are very minimal. This makes the retroviral pseudotypes incorporating
alphaviral glycoproteins ideal candidates as gene-therapy/ gene-transfer vectors.

21
Figure 2.5 Pseudotyping
Alphaviral-MoMuLV pseudotypes are generated by transfecting producer ØNXnlslacZ cells with a plasmid encoding alphaviral glycoproteins (figure shows RRV-envelope plasmid). The ØNXnlslacZ cell-line is a second generation 293-T based retroviral packaging cell line that is stably transfected with the MoMuLV gag-pol and MFG.S-nlslacZ plasmids. The pseudotyped viruses generated using this method are replication incompetent and function as gene-delivery agents.

22
CHAPTER 3. MATERIALS AND METHODS
3.1 Cell lines and cell culture
Producer ØNXnlslacZ cells (a second generation 293-T based retroviral packaging cell
line that is stably transfected with MoMuLV gag-pol plasmid and MFG.S-nlslacZ
plasmid, which produces envelope-protein-deficient, replication-incompetent MoMuLV
particles that encode nuclear-localized β-galactosidase) (Sharkey CM 2001); BHK;
CHO22; CHO18.4 (a mutant cell line derived from CHO22 not expressing HS) (Heil ML
2001, Jan J 1999) [generously donated by Dr. Richard Kuhn’s laboratory] were grown in
Dulbecco’s Modified Eagle’s Medium (DMEM) with 10% heat-inactivated fetal bovine
serum (FBS) and penicillin (10 U/mL ) at 37°C in 5% CO2. NIH 3T3 cells were grown in
DMEM with 10% calf serum (CS) and penicillin (10 U/mL) at 37°C in 5% CO2. Sf9
insect cells were grown in Sf9/III medium with 3% FBS at 28 °C without CO2. C6/36
mosquito cells were grown in DMEM with 10% heat-inactivated-FBS at 28 °C in 5%
CO2, as well as in DMEM with de-lipidated heat-inactivated-FBS at 28 °C in 5% CO2.
De-lipidated serum was obtained by centrifugation of serum in presence of Cab-O-SiL
(Cabot Co.). Cab-O-SiL acts by adsorbing lipids in the serum. ASMase-/- mutant mouse
skin fibroblast cells and ASMase+/+ wild-type mouse skin fibroblast cells (kindly)

23
donated by Dr. Glyn Dawson’s Laboratory) (Qin J 2012), were grown in DMEM with
10% heat-inactivated-FBS at 37 °C in 5% CO2.
3.2 Plasmids
Regions of the RRV cDNA containing amino acid substitutions T216R, T216V, N218R
or N218V (Heil ML 2001) [kind gifts from Dr. Richard Kuhn’s laboratory] were
removed through restriction endonuclease cutting with ScaI and RsrII. These enzymes cut
the genomic RRV clone at nucleotides 9201 and 9568 respectively. Each of these
fragments was individually ligated with ScaI/BglII fragment spanning nucleotides 12-
1766 and the large RsrII/BglII fragment (resulting from a partial RsrII digest) from
nucleotide 2133 to nucleotide 12. These plasmids are designated pRRV-E2E1A-T216R,
pRRV-E1E2A-T216V, pRRV-E2E1A-N218R and pRRV-E2E1A-N218V respectively
according to the single amino acid change in each glycoprotein coding sequence. The
integrity of each cloned plasmid was confirmed by sequence analysis.
Plasmids expressing envelope glycoproteins of alphaviruses namely pRRV-
E1E2A (Sharkey CM 2001), pVEEV-E1E2, pSINV-E1E2 and pSFV-RRV-E1E2 were
constructed by digesting the cDNA of viruses (RRVand SFV- BamHI and XbaI, VEEV-
HindIII and XbaI, SINV-BamHI and XhoI) and ligating the fragment containing E3-E2-
6K-E1 coding region into pcDNA3.1/zeo (+) cut with same enzymes. VSV-G envelope
plasmid, pCMV-G, (Kang Y 2002) and penv1min (Taylor GM 1999) were used for
expression of Vesicular Stomatitis virus envelope glycoproteins and MoMuLV envelope
glycoproteins respectively. pBACgus1-RRV-E1E2 was constructed by digesting pRRV-
E1E2A with BglII and AvrII and inserting the fragment containing the E3-E2-6K-E1

24
coding region into pBACgus1 plasmid cut with the same enzymes. Bacmid bMW033,
which expresses replication-competent baculovirus with intact envelope glycoprotein,
gp64, and bacmid expressing baculovirus cores deficient in gp64, bMW024, were kindly
gifted by Dr. Colin T. Dolphin’s laboratory (Westenberg M 2013).
3.3 Production of virus by transient expression of envelope glycoproteins
4,000,000 producer ØNXnlslacZ cells were grown overnight on 10 cm plates. The
following day they were washed with phosphate-buffered-saline solution (PBS), and 10
mL of fresh growth medium was added to them. Plasmids expressing RRV envelope
glycoproteins: pRRV-E1E2A, pRRV-E1E2A-T216R, pRRV-E1E2A-N218R, pRRV-
E1E2A-T216V and pRRV-E1E2A-N218V were incubated with 12 µL PLUS reagent
(Life Technologies) in DMEM for 15 minutes followed by incubation with 18 µL
Lipofectamine (Life Technologies) for 15 minutes in DMEM at room temperature. Each
of these plasmids was then added to the respective plates containing the producer cells
grown overnight, and the plates were incubated at 37 °C in 5% CO2. 3 hours after
incubation the liquid from each plate was replaced with 10 mL of fresh growth medium.
This was further incubated for 48 hours at 37 °C in 5% CO2 after which the supernatant
medium containing RRV-MoMuLV pseudotyped virus was collected from each of these
plates.
4,000,000 producer ØNXnlslacZ cells were grown overnight in 10 cm plates. The
following day, they were washed with phoshate buffered saline solution (PBS) and 10
mL fresh medium was added to them. Envelope glycoproteins expressing plasmids
pRRV-E1E2A, pVEEV-E1E2, pSINV-E1E2 and pSFV-E1E2 were incubated with 18 µL

25
of Lipofectamine 2000 (Life Technologies) for 15 minutes in DMEM at room
temperature. Each transfection mix containing the envelope plasmids was then added to
the respective plates containing the producer ØNXnlslacZ cells grown overnight, and
then the plates were incubated at 37 °C in 5% CO2. 4 hours after incubation the medium
from each plate was replaced with 10 mL fresh medium and this was further incubated
for 48 hours at 37 °C in 5% CO2. The supernatant media overlying these cells contained
pseudotyped virus. MoMuLV pseudotyped with MoMuLV glycoproteins and MoMuLV
pseudotyped with VSV-G were produced similarly.
3.4 Transduction assay
10 mL of supernatant media overlying producer ØNXnlslacZ cells were collected 48
hours after transfection and passed through a 0.45 µm filter. Hexadimethrine bromide (5
µg/mL) was added to the filtered supernatant media. This media was used to transduce
target cell-lines, NIH 3T3, BHKs, CHO22 and CHO18.4 (a mutant cell line derived from
the parental CHO22 that does not express HS). After 4 hours of incubation, the media
were replaced with fresh media. 48 hours after transduction, cells were stained with X-gal
(Taylor GM 1999).Transduction units were then calculated by the formula: Transduction
Units/ml (TU/mL) = (No. of blue cells/Total number of cells)*(No. of cells
plated/Volume of media added).
3.5 Immunoblot assay
10 mL of supernatant media from producer cells transfected with different plasmids were
passed through a 0.45 µm filter. The virus particles were collected by ultra-centrifugation
of the supernatant media through a 30% sucrose cushion in Beckman 50.2 Titanium rotor

26
at 28000 rpm. The pellets of virus particles accumulated at the bottom were suspended in
SDS-PAGE buffer containing 2% β-mercaptoethanol. The viral pellets were analyzed by
SDS-PAGE gel and then transferred to nitrocellulose paper. Presence of the envelope
glycoproteins RRV-E2 and RRV-E1 along with the MoMuLV-Capsid (p30) proteins
was detected by incubating with rabbit anti-E2 (1:5000), rabbit anti-E1 (1:5000) and goat
anti-Raucher Leukemia virus-Capsid (p30) polyclonal primary antibodies (1:1000)
respectively. Anti-rabbit antibodies (Chemicon) and anti-goat antibodies coupled with
horseradish peroxidase were used as secondary antibodies (1:5000) to detect RRV
envelope glycoproteins and MoMuLV capsid proteins respectively. The amount of
protein present was measured based on chemiluminescence using FluorChem E imaging
system. The lysate and cell-debris samples were obtained by lysing the producer
ØNXnlslacZ cells transfected with different RRV envelope plasmids, using lysis buffer
containing 1% Triton-X 100, 50 mM Tris, 5 mM EDTA and 150 mM NaCl. Immunoblot
analysis of these preparations was also done as above.
3.6 Immunofluorescence assay
48 hours after transfection of CHO22 and CHO18.4 cell lines with RRV-envelope
encoding plasmids, cells were washed with PBS, fixed with 2% formaldehyde and
permeabilized with 0.1% Triton-X 100. This was followed by blocking them with 1%
BSA solution after which they were incubated overnight at 4°C with (1:100) primary
Anti-E2 (rabbit) antibodies. After washing off the excess primary antibodies, cells were
incubated with (1:500) Alexa Fluor 488-coupled anti-rabbit antibodies for 1-2 hours at

27
room temperature followed by removal of excess antibodies by washing. The cells were
visualized by fluorescence microscopy (Olympus IX) using MetaMorph software.
3.7 Addition of Heparinase I to Producer cells
44 hours after transfecting producer ØNXnlslacZ cells with RRV envelope plasmids, the
supernatant media overlying the cells were replaced with fresh media, and the cells were
incubated with 6 µg/mL Heparinase I at 37°C. After 4 hours the supernatant media were
collected and passed through a 0.45 µm filter. Hexadimethrine bromide (5 µg/mL) was
added to the filtered supernatant media. These media were used to transduce target cell
lines (NIH 3T3, BHKs, CHO22 and CHO18.4)
3.8 Addition of Heparinase I to target cells
Target cells (BHK, CHO22 and CHO18.4) grown overnight were incubated with
increasing concentrations (0, 2, 4, 6 µg/mL) of Heparinase I, an enzyme that cleaves HS
from the cell surface, for one hour at 37°C. After being washed with PBS, these cells
were transduced using supernatant media from producer ØNXnlslacZ cells as explained
earlier.
3.9 Normalization following quantitative analysis of the immunoblot assay
10 mL supernatant media from producer cells transfected with different plasmids were
collected and replaced with 10 mL fresh media. Collected media were passed through a
0.45 µm filter and subjected to ultracentrifugation in a Beckman 50.2 Titanium rotor at
28000 rpm with a 30% sucrose cushion to obtain pellets of viral particles. An
immunoblot assay of virus pellets using anti-E2 antibodies was performed. Using Image J
software, the amount of RRV-E2 glycoprotein in each virus pellet was quantified. Based

28
on these values the media overlying the producer cells was normalized across all the
samples and used for transduction of target cells (NIH 3T3, BHK, CHO22 and
CHO18.4). Normalization was performed by appropriate dilutions of the media
containing WT-RRV, T216R-RRV, T216V-RRV, N218R-RRV and N218V-RRV
pseudotyped viruses.
3.10 Transduction assays in cells treated with lipid-lowering drugs
The target cells (BHK and NIH-3T3) were incubated with 4 µg/mL of lovastatin (Sigma-
Aldrich) for 12 hours followed by another 12 hours of incubation with 2.5 mM of methyl-
beta-cyclodextrin (ß-mCD) (Sigma-Aldrich) 4 µg/mL of lovastatin, after which they were
washed with phosphate-buffered-saline solution (PBS). Supernatant media overlying the
producer ØNXnlslacZ cells were replaced with serum-free media, and the cells were
incubated for 5 hours. These serum-free media containing the pseudotyped viruses were
collected after passing them through a 0.45 µm filter. Hexadimethrine bromide (5 µg/mL)
was added to filtered supernatant media. These media were added to the target cells,
NIH-3T3 and BHK for transduction. After 24 hours of incubation, the media were
replaced with fresh media. 48 hours after transduction, cells were stained with X-gal.
Transduction units were then calculated by the formula (TU/mL) = (No. of blue cells/
Total number of cells)*No. of cells plated/ Volume of media added. These experiments
were also performed by replenishing the cholesterol content of cholesterol-depleted cells.
Repletion of cholesterol was done by incubating cholesterol-depleted cells in media
containing 50 µg/mL of cholesterol for 1 hour.

29
3.11 Production of baculovirus virus pseudotyped with alphavirus envelope
glycoproteins
500,000 Sf9 cells were grown for an hour on 6 cm plates. Following that the media were
replaced with 6 mL of fresh Sf9/III + 3% FBS media. Plasmid pBACgus1-RRV-E1E2
was coincubated with bMW024 and 18 µL of Cellfectin (Life technologies) for 30
minutes in 500 µL Sf9/III medium at room temperature. The transfection mix was added
to the plate containing Sf9 cells. 24 hours after incubation the medium from the plate was
replaced with 6mL fresh medium and this was further incubated for 4 days at 28 °C. The
supernatant medium containing baculovirus pseudotypes was passed through a 0.45 µm
filter and used for the transduction assay. Baculovirus incorporating VSV-G was
produced by co-transfection of pBACgus1-VSV-G and bMW024. Replication-competent
baculovirus was produced by transfection of cells with bMW033.
3.12 Transduction assay in C6/36 cells grown in sterol-free conditions
5 µg/mL of hexadimethrine bromide was added to filtered supernatant media containing
the baculovirus pseudotypes. These media were added to the C6/36 cells grown in 6 well
plates. After 12 hours of incubation, the media were replaced with fresh media. 72 hours
after transduction, cells were visualized under a fluorescent microscope (Olympus IX)
using MetaMorph software. Transduction units were calculated using the formula stated
above after counting the fluorescent cells.
3.13 Transduction of ASMase-/- cells
MoMuLV pseudotypes with alphavirus glycoproteins were produced as mentioned
above. The supernatent media overlying producer ØNXnlslacZ cells were used to

30
transduce target ASMase-/- and ASMase+/+ cells. The media were replaced with fresh
media after 24 hours. 48 hours after transduction, cells were stained with X-gal and
counted. Transduction units were then calculated.
3.14 Verification of cholesterol concentrations in ASMase-/- cells and ASMase+/+
cells
The difference in the concentration of cholesterol in ASMase-/- and ASMase+/+ cells
was verified by a fluorometric cholesterol assay. Cholesterol was extracted from one
million cells by lysing them using 200 µL of chloroform: methanol solution (v/v 2:1).
These samples were dried by allowing the solvent to evaporate at room temperature and
resolubilized using cholesterol-assay buffer. The concentrations of cholesterol in these
cells were determined using fluorometric cholesterol assay kit (Caymen Chemicals) and
96-well FLX microplate reader.

31
CHAPTER 4. ROLE OF HEPARAN SULFATE IN ENTRY AND EXIT OF ROSS
RIVER VIRUS GLYCOPROTEIN-PSEUDOTYPED RETROVIRAL VECTORS
4.1 Summary
The role of heparan sulfate (HS) in alphavirus entry is a topic of considerable ongoing
investigation. Variants of Ross River virus (RRV) that bind to HS have previously been
selected by serial passaging in cell culture. To explore the effects of mutations that
convey HS utilization specifically upon the process of entry, we pseudotyped Moloney
Murine Leukemia virus (MoMuLV), with the envelope of RRV. We substituted amino-
acid residues 216 and 218 on RRV-E2-envelope glycoprotein with basic amino-acid
residues, because these mutations were previously shown to confer upon the virus an
ability to bind HS. However, pseudotyped virus incorporating the basic amino-acid
substitutions possessed lower transduction titers. Using immunoblot and
immunofluorescence assays, we demonstrate that the affinity towards HS impeded
release of pseudotyped virus from producer cells. Addition of heparinase to the HS-
expressing target cells reduces the transduction efficiency of the virus carrying the basic
amino-acid substitutions, whereas no such effect is seen in cells lacking HS. These virus
particles had enhanced transduction capacity that was dependent upon utilization of HS
as an attachment factor. However, increased affinity towards HS also affected viral egress

32
negatively. This is reminiscent of the interaction between influenza virus and sialic acid,
in which the former utilizes the latter as an attachment factor for its entry, but the same
sialic acid, in the absence of neuraminidase, hinders viral release. General principles
concerning viral adaptation to the use of attachment factors and improving pseudotyped
virus titers through modifying cell membrane components can be derived from these
results.
4.2 Introduction
Viruses are commonly used as vectors to deliver a desired gene to the target cells
for the purpose of gene therapy or gene transfer. Retroviruses, such as human
immunodeficiency virus (HIV), feline immunodeficiency virus (FIV), and Moloney
murine leukemia virus (MoMuLV) may be useful as gene-delivery vectors, because they
allow stable transfer of genes under the influence of strong promoters to cells (Naldini L
1996, Kafri T 1997, Poeschla EM 1998, Noh MJ 2010). Whereas the recognition and
infection of host cells by viruses depends on their envelope proteins, incorporating the
envelope proteins from other viruses can convey to the parent virus the ability to enter
different cells than those it normally infects. These chimeric viruses are called
pseudotyped viruses. Recombinant retroviruses and lentiviruses bearing alphavirus
glycoproteins have been created and have been shown to be efficient gene
transfer/therapy vectors (Sharkey CM 2001, Kang Y 2002). Apart from their utility as
gene transfer/therapy vectors, alphavirus pseudotypes have been used as tools to study
the entry of alphaviruses.

33
Alphaviruses are arthropod-borne viruses that cause severe infections in a wide
range of vertebrates. The clinical manifestations in infected humans include fever, rash,
arthralgia, myalgia, headaches and polyarthritis (Strauss J H 1994). Alphaviruses are
classified into two categories: (i) Old World viruses, including Ross River virus (RRV),
Semliki Forest virus (SFV), Chikungunya virus (CHKV) and Sindbis virus (SINV) and
(ii) New World viruses, Venezuelan equine encephalitis virus (VEEV) and Eastern
equine encephalitis virus (EEEV) (Strauss J H 1994).
An alphavirus is an enveloped, single-stranded, positive-sense RNA virus
(11.5kb) (Strauss J H 1994). The RNA genome is surrounded by the capsid shell, which
is made up of 240 units of the capsid protein. This in turn is surrounded by an envelope
consisting of the glycoproteins E1, E2, and E3 embedded in a cell-membrane-derived
lipid bilayer. Each spike is a trimer of an E1/E2 heterodimer (Cheng RH 1995), and E3 is
non-stoichiometrically abundant. The envelope and the inner nucleocapsid core share
icosahedral symmetries with triangulation number T=4 (Strauss J H 1994, Cheng RH
1995).
RRV, like other alphaviruses, has a wide host range and tissue tropism, as their
infectious cycle requires infecting both mammals and insects (Jose J 2009, C.-V. C.
Kielian MC 2010, Kahl CA 2004). Alphaviruses enter the cells via receptor-mediated
endocytosis (DeTulleo L 1998, Sharkey CM 2001, C.-V. C. Kielian MC 2010). Binding
to receptors and attachment factors is mediated by E2 glycoproteins, whereas E1
glycoproteins mediate fusion of the viral and endosomal membranes (Strauss J H 1994,
Smith TJ 1995). Many proteinaceous receptors are thought to be responsible for
internalization of alphaviruses particles. However, these receptors have not been

34
characterized completely (C.-V. C. Kielian MC 2010, Jose J 2009). Cell-surface
attachment factors, which facilitate the initial binding of alphaviruses and thus increase
the concentration of virus particles on the cell surface, such as DC-SIGN and L-SIGN
have been suggested to play a role in alphaviral entry (N. E. Klimstra WB 2003).
Heparan sulfate (HS), an attachment factor used by a number of viruses, such as human
papilloma virus, hepatitis C virus, most herpes viruses, adeno-associated virus, dengue
virus, vaccinia virus, yellow fever virus and human respiratory syncytial virus (Giroglou
T 2001, Barth H 2003, S. P. Shukla D 2001, Summerford C 1998, Chen Y 1997, Chung
CS 1998, Germi R 2002, Feldman SA 2000), has also been proposed to be involved in
alphaviral entry (G. D. Byrnes AP 1998, Zhu W 2010, C.-N. J. Gardner CL 2013). In
particular, it has been proposed that capacity to use HS as an attachment factor plays a
role in the tissue tropism of Eastern Equine Encephalitis virus (E. G. Gardner CL 2011).
HS is ubiquitously found in the human body on the cell surfaces and in the
extracellular matrix. It is a glycosaminoglycan made up of repeating units of uronic acid
and glucosamine with varying degrees of sulfation (Figure 4.1), which confers a negative
charge (Lopes CC 2006). This negative charge allows ionic interaction with cationic
molecules.

35
Figure 4.1 Structure of Heparan Sulfate
HS is glycosaminoglycan made up of repeating units of uronic acid and glucosamine with varying degrees of sulfation.
Sindbis virus has been shown to use HS as an attachment factor (G. D. Byrnes AP
1998). It was later shown that serial passaging in cell culture led to adaptive mutations in
Sindbis virus, which led to its selection of HS as an attachment factor (R. K. Klimstra
WB 1998). Similar studies done in RRV revealed that serial passaging in chick embryo
fibroblasts led to mutations at position 218 on the E2 envelope glycoprotein that resulted
in replacement of the original Asparagine residue with Lysine (Kerr PJ 1993).
Incorporation of these N218R and N218K mutations in RRV was shown to permit RRV
to utilize HS as an attachment factor (Heil ML 2001). Similarly, basic amino-acid
substitutions of residue 216 of RRV leads to utilization of HS as attachment factor (ML
2001). It was also shown that even though substitutions did not result in formation of the
HS-binding motifs XBBXBX or XBXBBBX (Figure 4.2), the electrostatic interaction
between negatively charged HS and basic amino-acids residues clustered around the
receptor-binding region of the E2 glycoprotein (Figure 4.3 A and B) made utilization of
HS as an attachment factor possible (Heil ML 2001, Zhang W 2005).

36
Figure 4.2 RRV-E2 Envelope glycoprotein amino-acid sequence
Amino acid residue T216 and N218 (arrows) of RRV-E2 envelope glycoprotein were substituted with basic amino-acid residues, namely arginine. The amino-acid sequence of the RRV-E2 envelope glycoprotein (Figure 4.2A) contains other basic amino-acid residues in the vicinity of these two amino-acid residues (asterisk).

37
Figure 4.3 RRV-E2 Envelope glycoprotein and HS
We have previously described pseudotyping of recombinant retroviruses and
lentiviruses with alphaviral glycoproteins, including those of RRV (Sharkey CM 2001,
Kang Y 2002, Kahl CA 2004). In vivo gene transfer to hepatocytes and neuroglial cells
was successfully achieved by pseudotyping lentivirus with RRV envelope glycoproteins
Substitution of amino-acid residues T216 and N218 with basic amino-acid residues does not result in formation of either of the HS binding motifs, XBBXBX or XBXBBBX. However, these residues along with other basic amino-acid residues cluster around the peak of the RRV-E2 envelope glycoprotein (Figure 4.3A) (Zhang W 2005). Thus, the receptor-binding region of RRV-E2 glycoprotein develops electropositive charge around itself. This results in increased affinity of RRV-E2 glycoprotein towards negatively charged heparan sulfate. Cryo-electron microscopy imaging shows HS bound to N218R RRV-E2 glycoprotein (Figure 4.3B). Figure from Zhang et al., Virology, 2005 (Zhang W 2005).
A
B

38
(Kang Y 2002). In order to explore additional potential advantages of alterations to the
viral glycoproteins for transduction by RRV-MoMuLV pseudotyped virus, we created
RRV-envelope glycoproteins with the substitutions of basic amino-acid residues that had
been previously demonstrated to promote utilization of HS as an attachment factor for
RRV. We demonstrate that these substitutions promote the utilization of HS by the RRV
glycoprotein-pseudotyped retrovirus. Interestingly, these substitutions also appear to
reduce the release of pseudotyped virus from producer cells. Incorporation of
substitutions therefore has both advantages and disadvantages for gene transduction by
alphavirus-pseudotyped retroviruses and lentiviruses.
4.3 Results
4.4.1 Transduction by T216R-RRV and N218R-RRV pseudotyped viruses is lower
in both HS expressing as well as non-expressing cell lines
To determine whether substituting amino-acid residues of RRV-envelope glycoproteins
with basic amino-acid residues made the virus utilize HS and thereby increase its
transduction efficiency, T216R-RRV and N218R-RRV pseudotyped viruses were
studied. Pseudotypes with no substitutions in envelope glycoproteins, WT-RRV, and
pseudotypes with envelope glycoproteins containing substitutions of residues 216 or 218
with Valine, T216V-RRV and N218V-RRV, were used as controls. Transduction assays
were performed using these pseudotyped viruses. The data show that the transduction
titers of WT-RRV, T216V-RRV, N218V-RRV pseudotyped viruses were higher than
those of pseudotyped viruses with basic amino-acid substitutions (T216R-RRV and
N218R-RRV) in both HS expressing as well as non-expressing cell lines (Figure 4.4).

39
Figure 4.4 T216R-RRV and N218R-RRV pseudotypes have lower transduction titers
Supernatant media overlying producer ØNXnlslacZ cells producing WT-RRV, T216R-RRV, T216V-RRV, N218R-RRV and N218V-RRV pseudotyped viruses respectively were collected and passed through a 0.45 µm filter. Hexadimethrine bromide (5 µg/mL) was added to the filtered supernatant media. This media was used to transduce target cell-lines (A) NIH 3T3, (B) BHKs, (C) CHO22 (parent cell-line) and (D) CHO18.4 (cell-line not expressing HS). The transduced target cells were stained with X-gal and transduction titers were calculated. The experiments were repeated thrice. The values shown by the graph represent the mean TU/mL values (± SD) calculated from these experiments.
0
5000
10000
15000
20000
Tran
sdu
ctio
n (
TU/m
L)
NIH- 3T3
01000200030004000500060007000
Tran
sdu
ctio
n (
TU/m
L)
BHK
01000200030004000500060007000
Tran
sdu
ctio
n (
TU/m
L)
CHO18.4
01000200030004000500060007000
Tran
sdu
ctio
n (
TU/m
L)
CHO22
A B
C D

40
4.3.2 T216R-RRV and N218R-RRV envelope glycoproteins are retained in the
producer cells
To investigate the reason for lower transduction titers in T216R-RRV and N218R-RRV,
immunoblot assays were carried out. Immunoblot analysis (Figure 4.5) of the pellets
containing the viral particles that were collected after ultra-centrifugation showed that
there was very little incorporation of the T216R-RRV and N216R-RRV-envelope
glycoproteins as compared to that of the WT-RRV, T216V-RRV and N218V-RRV-
envelope glycoproteins. The amount of capsid protein in the viral pellets observed
however was equal across all the samples. Thus the virus particles from T216R-RRV and
N218R-RRV samples were lacking the envelope glycoprotein around the retroviral cores.
This explains the reason for lower TU/mL in these samples. Further, the amounts of
envelope glycoproteins and capsid proteins in the lysates obtained from producer
ØNXnlslacZ cells were the same across in all samples. This shows that the mutations did
not affect the production of the T216R-RRV and N218R-RRV envelope glycoprotein.
The cell debris, which is the insoluble part of producer cell (parts of the cells resistant to
1% Triton-X 100 lysis solution), however, contained higher levels of the glycoproteins in
the T216R-RRV and N218R-RRV expressing cells as compared to wild-type, T216V-
RRV and N218V-RRV expressing cells. The quantities of the capsid proteins were same
across all the samples. This could suggest that the envelope glycoproteins of T216R-
RRV, N218R-RRV are being retained within the producer cells. Thus even though
envelope and capsid proteins were produced in equal quantities in all samples, greatly
reduced quantities of complete pseudotyped virus particles were not formed in T216R-
RRVand N218R-RRV samples. Envelope glycoproteins transfected with plasmids

41
encoding envelopes with these mutations, were retained within producer ØNXnlslacZ
cells and only naked MoMuLV cores lacking the envelope were released.
Figure 4.5 T216R-RRV and N218R-RRV envelope glycoproteins are inefficiently
incorporated in pseudotyped virus
Virus pellets (A) were spun down from supernatant media overlying producer ØNXnlslacZ cells transfected with respective RRV-envelope encoding plasmids, while the lysate (B) and the cell debris (C) were obtained by lysing the same producer ØNXnlslacZ cells. These preparations were run on SDS-PAGE gels and transferred on to nitrocellulose paper. RRV-envelope glycoproteins (E1 and E2) and MoMuLV Capsid proteins in these samples were detected by immunoblotting with rabbit anti-E1, rabbit anti-E2 and goat anti-Capsid (p30) polyclonal primary antibodies (1:5000) respectively, followed by secondary antibodies coupled with horseradish peroxidase. These experiments were performed thrice.

42
4.4.3 Attachment of T216R-RRV and N218R-RRV envelope glycoproteins to HS
leads to retention of these glycoproteins within the cells producing them.
Immunoblot assays showed that the envelope glycoproteins of T216R-RRV and N218R-
RRV were retained within the producer ØNXnlslacZ cells. To show that the retention of
glycoproteins could be due to their affinity towards HS expressed by the producer cells,
an immunofluorescence assay was performed. Since virus particles released from
producer ØNXnlslacZ can reenter those same cells, the study of this exit process cannot
be isolated from the entry process using immunofluorescence assay in these cells. Hence
to study the role of HS in the retention of the envelope glycoproteins, the glycoprotein
expressing plasmids were transfected in to CHO22 and CHO18.4 cell-lines. Results
showed that there was excess retention of T216R-RRV and N218R-RRV envelope
proteins in CHO22 cells along the secretory pathway during transport to the plasma
membrane as compared to WT-RRV, T216V-RRV and N218V-RRV envelope
glycoproteins (Fig. 3 A and Supplemental Fig. 1). In CHO 18.4, a cell line that does not
express HS, no excess retention of T216R-RRV and N218R-RRV envelope glycoproteins
was seen in comparison to rest of the samples (Fig. 3 B). It can be concluded that the
affinity of T216R-RRV and N218R-RRV glycoproteins towards HS leads to their
intracellular retention in virus-producing cells, and predominantly naked MoMuLV
cores, incapable of transducing target cells, are released from cells producing these
mutant envelope glycoproteins.

43
Figure 4.6 Intracellular retention of T216R-RRV and N218R-RRV in HS-expressing
cells
(A) CHO22 and (B) CHO18.4 were transfected with respective RRV-envelope plasmids. After fixing and permeabilizing the cells, they were incubated with primary Anti-E2 (rabbit) antibodies. After washing off the excess primary antibodies, cells were incubated with Alexa Fluor 488-coupled anti-rabbit antibodies and visualized under fluorescent microscope (rendered red color). (C) CHO22 cells expressing T216R and N218R RRV E2-glycoproteins (green fluorescence) were probed with Golgi-specific marker BODIPY® TR-C5-ceramide complexed to BSA (red fluorescence)

44
CHO22
CHO18.4
CHO22 with golgi marker
Figure 4.6
T216R
A
B
T216V
N218R
N218V WT
Control
Control T216R N218R
WT T216V N218V
C
T216R N218R

45
4.3.4 Attachment of T216R-RRV and N218R-RRV Envelope Glycoproteins to HS
hinders the release of virus particles
To determine whether the affinity of RRV envelope glycoproteins towards HS expressed
on the surface of the producer cells affects the release of virus particles Heparinase I was
added to the producer ØNXnlslacZ cells. The aim of this experiment was to see whether
cleaving of HS from the surface of these producer cells facilitates the release of T216R-
RRV and N218R-RRV pseudotyped viruses and thus increases their transduction
efficiency. T216R-RRV and N218R-RRV pseudotyped viruses collected from producer
ØNXnlslacZ cells treated with Heparinase I have higher transduction titers as compared
to the ones not treated with Heparinase I (Figure 4.7). Addition of Heparinase I to
producer ØNXnlslacZ cells producing WT-RRV, T216V-RRV and N218V-RRV
pseudotyped viruses shows no effect on their respective transduction efficiency. Thus HS
expressed on surface of producer ØNXnlslacZ cells hinders the process of release of
T216R-RRV and N218R-RRV pseudotyped viruses.

46
Table 4.1 Paired T-test
Figure 4.7 Addition of Heparinase I to Producer ØNXnlslacZ cells releases T216R-RRV
and N218R-RRV pseudotyped viruses
0
20
40
60
80
100
120
140
160
Control WildType
T216R T216V N218R N218V
% T
ran
sdu
ctio
n (
% T
U/m
L)
No Heparinase
Heparinase
Sample T-value P-value
WT 1.24 0.342
T216R -10.55 0.009 *
T216V 0.38 0.741
N218R -4.82 0.040*
N218V -0.69 0.561
Producer ØNXnlslacZ cells transfected with respective RRV-envelope plasmids are treated with Heparinase I. Supernatant media were collected from these cells and used for transduction of target cells (BHK) as described earlier. Transduction titers were calculated for each experiment which was repeated thrice. The graph represents transduction titers, where 100% TU/mL for WT, T216R, T216V, N218R and N218V are 6090, 1650, 3860, 1760 and 4510 TU/mL respectively. The effect of addition of Heparinase I to producer cells is shown statistically using the paired T-Test in the table alongside (Table 4.1).

47
4.3.5 T216R-RRV and N218R-RRV pseudotyped viruses utilize HS as an
attachment factor
To investigate whether T216R-RRV and N218R-RRV pseudotyped viruses utilize HA as
an attachment factor, Heparinase I was added to the target cells. Heparinase I cleaves HS
from the cell surface by breaking the α (1-4) glycosidic bond of HS. The data
demonstrate (Figure 4.8) that in BHK and CHO22 cells, as the concentration of
Heparinase I increases, the transduction titers of T216R-RRV and N218R-RRV
pseudotyped viruses decrease, whereas transduction titers of WT-RRV, T216V-RRV and
T218V-RRV pseudotyped viruses do not change drastically. However, in CHO18.4 the
transduction titers remains equal across all samples irrespective of Heparinase I
concentration. In addition, whereas transduction by WT-RRV, T216V-RRV and T218V-
RRV pseudotyped viruses in CHO22 and CHO18.4 cells is approximately equivalent,
transduction by T216R-RRV and N218R-RRV pseudotyped viruses is higher in CHO22
cells than in CHO18.4 cells. Furthermore, treatment of the CHO22 cells with heparinase
reduces transduction by the T216R-RRV and N218R-RRV pseudotyped viruses
approximately to the level seen in CHO18.4 cells. Therefore in cell lines expressing HS,
the transduction efficiency of T216R-RRV and N218R-RRV pseudotyped viruses
decreases, as the amount of cell surface HS decreases, whereas no such effect is seen in
cell lines not expressing HS. Thus, these data demonstrate that T216R-RRV and N218R-
RRV pseudotyped viruses utilize HS as an attachment factor.

48
Figure 4.8 Heparinase treatment of target cells reduces transduction by T216R-RRV and
N218R-RRV pseudotyped viruses
Target cells (A) BHK, (B) CHO22 and (C) CHO18.4 were treated with increasing concentrations of Heparinase I for 1 hour before transducing them with WT-RRV and mutant pseudotyped virus, respectively. The experiments were performed in triplicate. The correlation between concentrations of Heparinase I and transduction titers for each sample was determined by calculating Pearson coefficients using Minitab software in (Table 4.2) BHK, (Table 4.3) CHO22 and (Table 4.4) CHO18.4 cell lines.

49
Figure 4.8
01000200030004000500060007000
Tran
sdu
ctio
n (
TU/m
L)
Heparinase Concentration
BHK
Control
Wild Type
T216R
T216V
N218R
N218V
0
1000
2000
3000
4000
5000
6000
7000
0µg/ml 2µg/ml 4µg/ml 6µg/ml
Tran
sdu
ctio
n (
TU/m
L)
Heparinase Concentration
CHO22
Control
Wild Type
T216R
T216V
N218R
N218V
Type Pearson
Coefficient P value
WT 0.008 0.976
T216R -0.842 *0.001
T216V 0.085 0.792
N218R -0.896 *0.0001
N218V -0.052 0.865
Type Pearson
Coefficient P value
WT -0.159 0.707
T216R -0.737 *0.037
T216V -0.105 0.805
N218R -0.848 *0.008
N218V -0.055 0.898
Type Pearson
Coefficient P value
WT 0.365 0.374
T216R 0.132 0.755
T216V -0.129 0.761
N218R 0.143 0.763
N218V 0.257 0.539
Table 4.2 Pearson Coefficient BHK
0
1000
2000
3000
4000
5000
6000
7000
0µg/ml 2µg/ml 4µg/ml 6µg/ml
Tran
sdu
ctio
n (
TU/m
L)
Heparinase Concentration
CHO 18.4
Control
Wild Type
T216R
T216V
N218R
N218V
Table 4.2 Pearson Coefficient CHO22
Table 4.2 Pearson Coefficient CHO18.4

50
4.3.6 Transduction efficiency of T216R-RRV and N218R-RRV is higher in cell lines
expressing HS
Intracellular retention of the T216R-RRV and N218R-RRV glycoproteins results in
markedly reduced transduction efficiencies by the pseudotyped virus recovered from the
supernatant medium of producer cells. It may be that there are fewer virus particles that
bear alphavirus glycoproteins and/or that virus particles bear fewer glycoproteins. To
investigate whether attachment to HS expressed on target cells increases the transduction
efficiency by the virus particles bearing the low quantities of T216R-RRV and N218R-
RRV glycoproteins that are released, normalization of virus concentration by
glycoprotein content was performed. The RRV-E2 envelope glycoprotein content of the
virus particles collected by ultracentrifugation of the supernatant media of each of the
pseudotyped-virus producing cells was measured by immunoblotting. The data show
(Figure 4.9) that after normalization based on the amount of RRV-E2 glycoprotein, the
transduction efficiency for T216R-RRV and N218R-RRV pseudotyped viruses were
significantly higher than that of wild type, T216V-RRV and T218V-RRV pseudotyped
viruses in NIH 3T3, BHK and CHO22 cells. In CHO18.4, however, there was no
significant difference in the transduction efficiency across these samples. Thus, in cell
lines expressing HS, when the amount of RRV-E2 glycoprotein protein is equal across all
the samples, T216R-RRV and N218R-RRV pseudotyped viruses have higher
transduction efficiency than wild-type, T216V-RRV and N218V-RRV pseudotyped
viruses.

51
Figure 4.9 T216R-RRV and N218R-RRV pseudotyped viruses possess higher
transduction efficiencies when transductions by viruses are normalized by glycoprotein
content
Virus was collected by ultracentrifugation from supernatant media overlying producer ØNXnlslacZ cells transfected with respective RRV-envelope encoding plasmids. The virus-containing samples were analyzed by SDS-PAGE and transferred onto nitrocellulose paper. RRV-E2 glycoprotein was detected by immunoblotting with rabbit anti-E2 primary and anti-rabbit secondary antibodies coupled with horseradish peroxidase. Using Image J software, (A) the amount of RRV-E2 glycoprotein in each virus sample was quantified. A representative image is shown. The amount of media added to target cells (B) NIH-3T3, (C) BHK, (D) CHO22 and (E) CHO18.4, for transduction was normalized against the amount of RRV-E2 glycoprotein present in the given sample. The transduction of the CHO22 and CHO18.4 cells was performed with different starting samples of the five viruses, so the transduction efficiencies measured in this experiment between the two cell lines cannot be directly compared. The experiments were repeated thrice. The values shown by the graph represent the mean TU/mL values (± SD) calculated from these experiments.

52
Figure 4.9
A
0
2000
4000
6000
8000
Tran
sdu
ctio
n (
TU/m
L)
NIH-3T3
0
500
1000
1500
2000
2500
Tran
sdu
ctio
n (
TU/m
L)
CHO22
0
500
1000
1500
2000
Tran
sdu
ctio
n (
TU/m
L)
CHO18.4
0
500
1000
1500
2000
2500
Tran
sdu
ctio
n (
TU/m
L)
BHK B C
D E

53
4.4 Discussion
Pseudotyped viruses have many experimental and clinical applications. Gene
therapy/transfer is one of the important applications of these pseudotyped viruses. Based
on the therapeutic need, the range of cells targeted by the viral vector for gene
therapy/transfer can be either expanded or narrowed by pseudotyping the virus with the
envelope of another virus (Sanders 2002). For the treatment of systemic genetic disorders
a viral vector needs to be pseudotyped with an envelope of a virus that has a broad tissue
tropism. Since RRV, an alphavirus, has wide host range and tissue tropism, it was
incorporated in ØNX pseudotyped system to produce RRV-MoMuLV pseudotyped virus
(Sharkey CM 2001). Previous studies showed that pseudotyping with RRV was far less
cytotoxic but equally efficient as VSV-G (Kang Y 2002). Retrovirus pseudotyped with
RRV envelope glycoproteins was successfully used for in vivo gene transfer in
hepatocytes, glial cells, muscle tissue and airway epithelial tissue (Kang Y 2002). To
further enhance the transduction efficiency and further broaden the range of cells that can
be targeted by the pseudotyped viral vector additional modifications of these envelope
glycoproteins are needed.
Earlier studies in RRV show that amino acid T216 on RRV-E2 glycoprotein is the
site involved in interaction with the receptors (Smith TJ 1995). The RRV48 strain of
RRV normally does not utilize HS as an attachment factor. Serial passaging of this strain
led to mutations on the E2 envelope glycoprotein in which the original amino-acid
residues were replaced with basic amino-acid residues (Kerr PJ 1993). Further studies
showed that site-directed mutagenesis involving replacement of threonine residue at 216
or the neighboring asparagine residue at 218 with a basic amino acid residue confers

54
upon the virus an affinity towards a negatively charged molecule like HS (Heil ML
2001). Cryo-electron microscopy and image reconstruction studies of RRV-E2
glycoprotein bound to heparin further confirmed this (Zhang W 2005).
Substituting the amino-acid residues at 216 and 218 sites on RRV-E2 glycoprotein with
basic amino acid residues in the RRV-MoMuLV pseudotyped virus, allows the
pseudotyped virus to use HS, an ubiquitous molecule present on most of the cell surfaces,
as an attachment factor. Using HS as an attachment factor increases the concentration of
the virus on the cell surface, which can later be internalized by cell surface receptors via
endocytosis. Thus the three-dimensional search for the receptor is reduced to a two-
dimensional search, which increases the efficiency of cell entry, thereby enhancing the
transduction efficiency of the pseudotyped vector. HS binding ability could also
potentially alter the tissue tropism of this viral vector (Kang Y 2002, E. G. Gardner CL
2011).
Ability to use HS as an attachment factor however affects the assembly and
release process of the virus particles. Due to the increased affinity of RRV-E2
glycoproteins towards HS, envelope glycoproteins get retained along the secretory
pathway, especially in the Golgi, since HS based glycosylation of other proteins takes
place in the Golgi (DK 2011). Thus the envelope glycoproteins with basic amino-acid
mutations are prevented from reaching the plasma membrane. Hence they are not
efficiently incorporated into the budding pseudotyped virus. Secondly, even if the
glycoproteins reach the plasma membrane and get incorporated into budding pseudotyped
virus, the data indicate that their affinity towards HS prevents the release of virus particle
from the cell surface. Studies done in Sindbis virus have shown that virus particles

55
utilizing HS had lesser viremia resulting in lower pathogenicity. This led to the reversion
of virus particles adapted to utilize HS to that wild-type form that had reduced affinity
towards HS. It was proposed that one reason for lesser viremia seen in Sindbis virus
utilizing HS was decreased dissemination of the virus (G. D. Byrnes AP 2000). Similar
studies in Venezuela equine encephalitis virus and Chikungunya virus show attenuation
of HS-binding mutant viruses due to rapid clearance and decrease in the spread of virus
particles (Bernard KA 2000, H. J. Gardner CL 2014).
This interaction between the HS-utilizing viruses and HS is reminiscent of the
interaction between influenza virus and its attachment factor, sialic acid. Sialic acid
facilitates the entry of influenza virus by attaching to the viral and increases their
concentration on the cell surface (Skehel JJ 2000, Wagner R 2002). However, the same
attachment factors, when expressed on the cells producing the virus particles, hinder the
release of the viral particles from the cell surface (Wagner R 2002). The release of
influenza virus from cell surfaces is facilitated by the Neuraminidase protein intrinsic to
the virus, which cleaves the sialic acid (Air GM 1989, Bucher D 1975). Because such an
intrinsic process for facilitating the release of virus particles is not present in our
pseudotyped viral vector, one of our future goals is to develop a producer cell line that
does not express HS for the production of this pseudotyped viral vector that utilizes HS
as an attachment factor. A general principle arising from these studies is that altering
producer cell membrane components may improve pseudotyped virus production through
enhancing viral particle release.
The attachment of viruses to cell-surface heparan sulfate has been proposed both
to be an inherent component of viral entry in vivo and an adaptation to propagation in cell

56
culture. An important consequence of our studies is the conclusion that utilization of
heparan sulfate as an attachment factor can have deleterious effects on viral release either
through intracellular retention of the heparan-sulfate-binding viral proteins or adsorption
of the viral particles to producer cells. HSV-1 utilizes heparan sulfate as an attachment
factor for infection of host cells (S. P. Shukla D 2001, L. J. Shukla D 1999). Recently it
has been demonstrated that increased host heparinase expression is a consequence of
herpes simplex virus-1 infection and that augmentation of heparinase expression
correlates with enhanced HSV-1 release (Hadigal SR 2015). These data reinforce our
conclusion that if a virus utilizes an ubiquitous and abundant cell component for entry
then there must be a mechanism for release of the virus from an infected cell if efficient
infection is to occur. This issue is likely to be particularly critical when the utilization of
the attachment factor appears to arise from an adaptation to growth in cell culture

57
CHAPTER 5. ROLE OF CHOLESTEROL IN ENTRY OF PSEUDOTYPES
INCORPORATING ALPHAVIRAL GLYCOPROTEINS AND NIEMANN PICK’S
DISEASE
5.1 Summary
The tissue tropism of a virus can be modified by replacing the envelope-glycoproteins of
that virus with those of other viruses, through the process of pseudotyping. Such
pseudotyped viruses are good candidates as vectors for gene therapy/ transfer. In this
study, we propose that retroviruses incorporating alphaviral glycoproteins will be suitable
gene delivery vectors in Niemann Pick’s disease, type A (NPD-A). NPD-A is a genetic
lipid-storage disorder characterized by severe deficiency of acid sphingomyelinase
(ASMase), which results in excess accumulation of cholesterol and sphingolipids in the
lysosomes and late endosomes. Previously it has been shown that cholesterol is required
by alphaviruses for fusion process during the entry. We show that alphavirus-
glycoprotein retrovirus pseudotypes also require cholesterol for entry into the cells.
Furthermore, excess cholesterol in cells facilitates the entry of alphavirus-glycoprotein
retroviral pseudotypes. Alphavirus-glycoprotein pseudotypes transduce the acid
sphingomyelinase deficient (ASMase-/-) cells derived from Niemann Pick’s disease-
model mice more efficiently than the cells from healthy mice (ASMase+/+). Thus

58
alphavirus-glycoprotein pseudotypes hold a potential as suitable vector for gene therapy/
transfer in NPD-A.
5.2 Introduction
Neimann Pick’ disease (NPD) is a lysosomal storage disease. Types A and B of
this disease are caused by mutations in the SMPD1 gene, leading to severe deficiency of
ASMase activity. Deficiency in this enzyme leads to excess accumulation of
sphingolipids in lysosomes and late endosomes (Schuchman EH 2001). Accumulation of
sphingolipids is coupled with the accumulation of cholesterol in lysosomes and
endosomes (Puri V 1999, Choudhury A 2004). NPD-A is a more severe form this
disease, than the other three types NPD-B, NPD-C and NPD-D. Clinical manifestations
in NPD-A include persistent early jaundice, hepatosplenomegaly, enlarged abdomen,
delay in developmental milestones, intellectual developmental disorders, stunted growth,
hypotonia and rigidity (Crocker AC 1958). NPD-A has poor prognosis leading to an early
death, usually by 2 to 4 years of age. Currently there is no cure available for this disease.
However, gene transfer studies in cultured NPD cells as well as diseased-mouse models
have shown to correct the metabolic defect (Suchi M 1992, Dodge JC 2005).
Viruses are frequently used as gene therapy/ gene transfer vectors to introduce the
gene of choice into cells. Since entry of an enveloped virus depends on its envelope
proteins, the range of target cells that a virus can enter into can be modified by replacing
the envelope glycoproteins of that virus with those of other viruses having the desired
range of tissue tropism, through the process of pseudotyping (Sanders 2002). Such
pseudotyped viruses are good candidates as vectors for gene therapy/transfer. Therapeutic

59
need would determine the choice of virus whose envelope glycoproteins need to be
incorporated in the pseudotyped virus (Sanders 2002).
Retroviruses are commonly used as gene delivery vectors because they allow
stable transfer and efficient expression of the genetic material under the influence of their
strong promoter. However, they have limited host range and tissue tropism. With the
purpose of broadening the tissue tropism and further enhancing the transduction
efficiency of this virus, we pseudotyped Moloney Murine Leukemia Virus (MoMuLV), a
model retrovirus, with envelope glycoproteins of Ross River Virus (RRV), an alphavirus
(Sharkey CM 2001). Successful in vivo gene transfer studies have been conducted
previously using lentiviral pseudotypes incorporating alphaviral envelope glycoproteins
(Kang Y 2002).
Alphaviruses are mosquito-borne viruses and are classified into Old World
viruses, which include Ross River Virus (RRV), Sindbis Virus (SINV), Semliki Forest
virus (SFV) and New World viruses, which include Venezuelan Equine Encephalitis
virus (VEEV), Eastern Equine Encephalitis virus (EEEV) and Western Equine
Encephalitis virus (WEEV). Alphaviruses have wide host-range and tissue tropism.
Alphaviruses enter the cells via receptor-mediated endocytosis. Two of the alphaviral
glycoproteins, E1 and E2 are required for this entry process (C.-V. C. Kielian MC 2010).
The E2-glycoprotein is involved in the initial receptor-binding phase, whereas the E1
glycoprotein is responsible for fusion of viral and cellular membranes (C.-V. C. Kielian
MC 2010, Jose J 2009). Once the virus gets internalized, the acidic pH of endosomes
leads to dissociation of the E1-E2 heterodimer exposing the hidden fusion loop of E1 (C.-
V. C. Kielian MC 2010). The insertion of fusion loop into the endosomal membrane

60
initiates the fusion process (C.-V. C. Kielian MC 2010). Studies in alphaviruses like
Sindbis Virus (SINV) and Semliki Forest Virus (SFV) using liposomes show that
cholesterol and sphingolipids are required for this step of fusion of the viral membrane
with the cellular membrane (Helenius A 1984) (Nieva JL 1994). Micro-domains of
cholesterol and sphingolipids are called lipid-rafts (Figure 5.1). These cholesterol-rich
lipid rafts have also been shown to be required for glycoprotein transport and the exit
process (G. D. Ng CG 2006, C. I. Ng CG 2008). Furthermore, studies have shown that
SINV causes fatal encephalitis in knock-out mice deficient in acid sphingomyelinase
(ASMase) as compared to healthy mice (G. D. Ng CG 2006).

61
Figure 5.1 Cholesterol and Sphingolipid rich lipid rafts
Compartmentalization of lipid bilayer forms micro-domains called as ‘lipid rafts’ which are rich in cholesterol and sphingolipids content. The factors contributing to formation and stability of lipid rafts are hydrogen bonding between polar heads, interaction between long hydrophobic chains and complementarity between the shapes of lipid molecules. Various proteins are embedded within these lipid-rafts.

62
Alphaviruses mainly infect brain, liver, bones, muscles, lungs and reticulo-
endothelial tissue and these sites can be targeted using pseudotyped viruses incorporating
alphaviral glycoproteins (Kang Y 2002). An overlap exists between the organs infected
by alphaviruses and the organs that get affected in Niemann Pick’s Disease. At the
cellular level, the fusion process of alphaviruses, which requires cholesterol, takes place
in endosomes. In NPD-A, accumulation of sphingolipids and cholesterol also takes place
in endosomes and lysosomes (Puri V 1999, Choudhury A 2004). By incorporating the
alphaviral glycoproteins in pseudotype system, we are trying to investigate if cholesterol
can have an effect on transduction efficiency of MoMuLV pseudotyped with alphavirus
envelope glycoproteins. We demonstrate that these pseudotyped viruses have higher
transduction efficiency in Niemann Pick-Type A model cells. These pseudotypes thus
hold a potential as vectors for gene transfer/therapy studies in Niemann Pick’s disease.
5.3 Results
5.3.1 Alphavirus-MoMuLV pseudotypes require cholesterol-rich lipid rafts for entry
in mammalian cells
Since the entry process of envelope forming virus in a pseudotyped virus can be studied
in isolation, alphavirus-MoMuLV pseudotypes were used to determine if cholesterol-rich
lipid rafts are required for cell entry. To show that lipid rafts are required for entry, the
lipid-rafts in target cells (BHK and NIH3T3) were disrupted using cholesterol lowering
drugs, namely lovastatin and ß-mCD. The experiment was also performed on cholesterol-
depleted cells, which were replenished with cholesterol. MoMuLV pseudotyped with
MoMuLV glycoproteins was used as positive control (S. J. Lu X 2000), whereas

63
MoMuLV pseudotyped with Vesicular stomatitis virus glycoprotein was used as negative
control (Yonezawa A 2005).
The data shown in Figure 5.2A and Figure 5.2B suggest that, the transduction
titers for pseudotypes with alphaviral glycoproteins are lower in cells treated with lipid-
lowering drugs as compared to non-treated cells. On replenishing the cells with
cholesterol, higher values of transduction units/mL are obtained. No such difference is
noted in transduction efficiency of pseudotypes incorporating VSV glycoproteins. Thus,
cholesterol-laden lipid rafts are essential for entry of alphavirus-MoMuLV pseudotyped
virus.

64
Figure 5.2 Alphavirus-MoMuLV pseudotypes require cholesterol for entry in mammalian
cells
0
20
40
60
80
100
120
VSV MoMuLV RRV SFV SINV VEEV
% T
ran
sdu
ctio
n
NIH-3T3
Untreated cells
Cells treated with Lovastatin andmßCD
Repletion with Cholesterol
0
20
40
60
80
100
120
VSV MoMuLV RRV SFV SINV VEEV
% T
ran
sdu
ctio
n
BHK
Untreated cells
Cells treated with Lovastatin andmßCD
Repletion with Cholesterol
Supernatant media overlying producer ØNXnlslacZ cells containing the pseudotyped virus particles were passed through a 0.45 µm filter and 5 µg/mL of hexadimethrine bromide was added to these filtered media. These media were used to transduce target cells, NIH-3T3 (Figure 5.1A) and BHK (Figure 5.1B). Cholesterol depletion was achieved in target cells by incubating the target cells with cholesterol-lowering drugs, namely lovastatin and methyl-beta-cyclodextrin (ß-mCD). The transduced target cells were stained with X-gal and transduction titers were calculated. Results shown are averages ± SD of three independent assays. The values shown in the graph represent transduction titers in percentages, where 100% TU/mL for VSV, MoMuLV, RRV, SFV, SINV and VEEV pseudotypes are 15000, 15000, 15000, 13000, 16000, 11000 and 15000 in NIH-3T3 cells, and 4000, 0, 5500, 5000,3500, 7000 in BHK cells respectively. The experiment was also performed following repletion of the cholesterol-depleted cells with 50 µg/mL cholesterol (β-sterol).
A
B

65
5.3.2 Alphavirus-baculovirus pseudotypes require cholesterol for entry into insect
cells
To rule out any direct effect of lipid-lowering drugs on the transduction values,
transduction assays were carried out in mosquito-derived C6/36 cells. The cholesterol
content of these cells can be reduced to as low as 1- 2% of their actual value, when grown
in sterol-free conditions (X. Y. Lu X 2002). Since pseudotyped viruses grown from
ØNXnlslacZ cells cannot transduce insect cells, baculovirus pseudotypes were produced.
Baculovirus pseudotypes incorporating baculovirus glycoprotein, gp64 was used as
positive control (Kataoka C 2001).The data show that transduction titers of these
baculovirus pseudotyped viruses incorporating alphaviral glycoproteins are lower in cells
grown in sterol-free condition as compared to cells not grown in sterol restricted
conditions (Figure 5.3). Repletion of cells grown in sterol-free conditions with
cholesterol leads to increase in the transduction efficiency of alphavirus-baculovirus
pseudotypes.

66
Figure 5.3 Alphavirus-baculovirus pseudotypes require cholesterol for entry in insect
cells
5 µg/mL of hexadimethrine bromide was added to the filtered supernatant media overlying SF9 cells. These media containing the baculovirus pseudotypes were used to transduce target C6/36 cells grown in DMEM+10% FBS (a), cholesterol depleted media (b) and media replenished with cholesterol (c). Cholesterol depletion was achieved in C6/36 cells by growing these cells in sterol-free condition. Experiments were also performed in these cells after replenishing them with β-sterol by adding 50 µg/mL cholesterol. The transduced target cells were identified using the fluorescent microscopy and transduction titers were calculated. The experiments were repeated thrice and atleast 10 fields (one field represented) were inspected per plate for each sample (Figure 5.2A). The graph shows averages±SD of transduction titers in percentages from three independent assays (Figure 5.2B), where100% TU/mL for Baculovirus, VSV and RRV pseudotypes are 1800, 450 and 450 respectively.
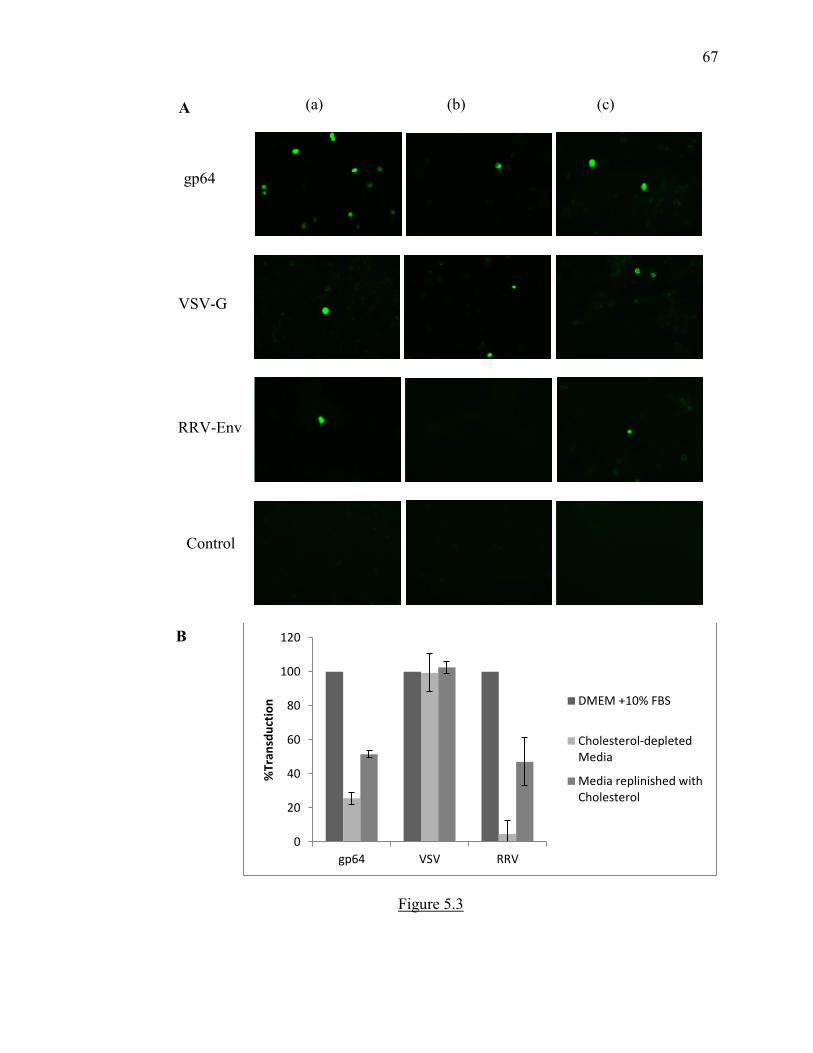
67
Figure 5.3
0
20
40
60
80
100
120
gp64 VSV RRV
%Tr
ansd
uct
ion
DMEM +10% FBS
Cholesterol-depletedMedia
Media replinished withCholesterol
gp64
VSV-G
RRV-Env
(a) (b) (c) A
Control
B

68
5.3.3 Alphavirus-MoMuLV pseudotypes transduce ASMase-/- cells more efficiently
than ASMase+/+ cells
To see whether excess cholesterol facilitates the entry of these pseudotyped viruses into
the cells, transduction assay was carried out in ASMase-/- cells and ASMase+/+ cells.
Absence of acid sphingomyelinase leads to excess accumulating of sphingomyelin and
cholesterol in the cells. Fluorometric assay confirmed that the cholesterol levels in
ASMase-/- cells were higher than ASM+/+ cells (Figure 5.4B). The data show (Figure
5.4A) that transduction efficiency of pseudotypes with alphaviral glycoproteins is higher
in ASMase-/- cells than in ASMase+/+ cells while not much difference was noted in
transduction efficiency of pseudotypes with VSV glycoproteins across these two cell-
lines.

69
Figure 5.4 Transduction efficiency of alphaviral pseudotypes is higher in ASMase-/- cells
0
100
200
300
400
500
600
VSV MoMuLV RRV VEEV SFV SINV
% T
ran
sdu
ctio
n
ASMase+/+ vs. ASMase-/- cells
ASMase+/+
ASMase-/-
0
20
40
60
80
100
120
Asmase+/+ Asmase-/-
% C
ho
lest
ero
l C
on
cen
trat
ion
Supernatant media overlying producer ØNXnlslacZ cells containing the pseudotyped virus particles were passed through a 0.45 µm filter. Hexadimethrine bromide (5 µg/mL) was added to these filtered media. These media were used to transduce target ASMase+/+ and ASMase-/- cells. The transduced target cells were stained with X-gal and transduction titers were calculated. Results shown are averages±SD of three independent assays represented in percentages (Figure 5.3A). The difference in the concentration of cholesterol in ASMase-/- and ASMase+/+ cells was verified using a fluorometric cholesterol assay kit (Figure 5.3B).
A
B

70
5.4 Discussion
NPD-Type A is an autosomal recessive genetic disorder caused by mutation in
SMPD1 gene and is characterized by excess accumulation of sphingolipids and
cholesterol in the cells. The treatment is mainly symptomatic and there is no cure
available for this disease. However, gene transfer studies performed using viral vectors
have shown to correct the metabolic defect in cultured cells as well as mouse models
(Suchi M 1992, Dodge JC 2005).
In our earlier study, we produced a pseudotyped virus by replacing the envelope
of Moloney Murine Leukemia Virus (MoMuLV) with the envelope of Ross River Virus
(RRV). Pseudotyping helps us in combining features of retrovirus, that are ideal for gene
transfer such as ability to allow fast and stable transfer of transgene to cells and efficient
expression of the transgene for prolonged period of time under the influence of its strong
promoter system, along with the features of alphaviruses which is to allow entry into
wide range of cells. This makes these pseudotyped viruses a potential gene
therapy/transfer vector in systemic genetic disorders. Successful in vivo gene transfer
studies have been conducted using lentiviral pseudotypes incorporating alphaviral
glycoproteins (Kang Y 2002). The cells specifically targeted by pseudotypes
incorporating alphaviral glycoproteins are neuronal cells, neuroglia, hepatocytes, Kupffer
cells and reticulo-endothelial cells (Kang Y 2002). Modest transductions are also seen in
myocytes and airway epithelial cell (Kang Y 2002). Interestingly, the tissues targeted by
the pseudotypes with alphaviral glycoproteins are similar to the ones affected in NPD-A
(Hua G 2013).

71
Since the entry of the virus depends on the envelope of the virus, the pseudotyped
virus has the entry mechanism similar to that of envelope forming virus and this entry
mechanism can be studied in isolation using the pseudotyped system. Using lipid-
lowering drugs and sterol-free conditions to grow cells, we show that pseudotypes
incorporating alphaviral glycoproteins require cholesterol for entry into the cells just like
wild-type alphaviruses. Studies in alphaviruses have shown that ASMase-/- mice were
more susceptible to fatal encephalitis (G. D. Ng CG 2006). Studies done in human NPD
cells which lacked ASMase enzyme attributed the higher virulence to rapid replication of
the virus in ASMase deficient cells and to production of more infectious viral particles
from these cells (C. I. Ng CG 2008). Here, using pseudotyped viruses, in which entry of
the envelope forming virus can be exclusively studied in isolation, we show that the
excess cholesterol in these cells facilitates the entry of pseudotyped viruses incorporating
alphaviral glycoproteins. Thus, efficient entry in ASMases deficient cells could be
another factor responsible for higher virulence and pathogenicity of alphaviruses in these
cells.
These characteristics make pseudotyped viruses incorporating alphaviral
glycoproteins suitable vectors for gene transfer/therapy in NPD-A. Further studies using
animal disease-models need to be done in order to validate their application in clinical
field.

72
REFERENCES
Air GM, Laver WG. "The neuraminidase of influenza virus." Proteins 6:341–356 (1989).
Anson, DS. "The use of retroviral vectors for gene therapy-what are the risks? A review
of retroviral pathogenesis and its relevance to retroviral vector-mediated gene
delivery." Genetic vaccines and Therapy 2(1):9 (2004).
Anthony RP, Paredes AM, Brown DT. "Disulfide bonds are essential for the stability of
the Sindbis virus envelope." Virology 90(1):330-336 (1992).
Barth H, Schafer C, Adah MI, Zhang F, Linhardt RJ, Toyoda H, Kinoshita-Toyoda A,
Toida T, Van Kuppevelt TH, Depla E, Von Weizsacker F, Blum HE, Baumert TF.
"Cellular binding of hepatitis C virus envelope glycoprotein E2 requires cell
surface heparan sulfate." The Journal of Biological Chemistry 278(42):41003-
41012 (2003).
Bernard KA, Klimstra WB, Johnston RE. "Mutations in the E2 glycoprotein of
Venezuelan equine encephalitis virus confer heparan sulfate interaction, low
morbidity, and rapid clearance from blood of mice." Virology 276(1):93-103
(2000).

73
Blaese R, Culver K, Miller A, Carter C. "T lymphocyte-directed gene therapy for
ADAnegative SCID: Initial trial results after 4 years." Science .270(5235):475
(1995).
Boggs WM, Hahn CS, Strauss EG, Strauss JH, Griffin DE. "Low pH-dependent Sindbis
virus-induced fusion of BHK cells: differences between strains correlate with
amino acid changes in the E1 glycoprotein." Virology 169(2):485-488 (1989).
Bucher D, Palese P. "The biologically active proteins of influenza virus: neuraminidase."
In The influenza viruses and influenza, by Editor- Kilbourne ED., 83-123.
Academic Press, New York, N.Y., 1975.
Byrnes AP, Griffin DE. "Binding of Sindbis Virus to Cell Surface Heparan Sulfate."
Journal of Virology 72(9):7349-7356 (1998).
Byrnes AP, Griffin DE. "Large-plaque mutants of Sindbis virus show reduced binding to
heparan sulfate, heightened viremia, and slower clearance from the circulation."
Journal of Virology 74(2):644-651 (2000).
Chen Y, Maguire T, Hileman RE, Fromm JR, Esko JD, Linhardt RJ, Marks RM.
"Dengue virus infectivity depends on envelope protein binding to target cell
heparan sulfate." Nature Medicine 3(8):866-871 (1997).
Cheng RH, Kuhn RJ, Olson N H, Rossmann MG, Choi HK, Smith TJ, Baker T S.
"Nucleocapsid and glycoprotein organization in an enveloped virus." Cell
80(4):621-30 (1995).
Choudhury A, Sharma DK, Marks DL, Pagano RE. "Elevated Endosomal Cholesterol
Levels in Niemann-Pick Cells Inhibit Rab4 and Perturb Membrane Recycling."
Molecular Biology of the Cell 15(10): 4500–4511 (2004).

74
Chung CS, Hsiao JC, Chang YS, Chang W. "A27L protein mediates vaccinia virus
interaction with cell surface heparan sulfate." Journal of Virology 72(2):1577-
1585 (1998).
Coffin JM, Hughes SH, Varmus HE. Retroviruses. Cold Spring Harbor Laboratory Press,
1997.
Crocker AC, Farber S. "Niemann-Pick disease: a review of eighteen patients." Medicine
(Baltimore) 37(1):1-95 (1958).
DeTulleo L, Kirchhausen T. "The clathrin endocytic pathway in viral infection." The
EMBO journal 17(16): 4585–4593 (1998).
DK, Banfield. "Mechanisms of Protein Retention in the Golgi." Cold Spring Harbor
Perspectives in Biology 3(8):a005264 (2011).
Dodge JC, Clarke J, Song A, Bu J, Yang W, Taksir TV, Griffiths D, Zhao MA,
Schuchman EH, Cheng SH, O'Riordan CR, Shihabuddin LS, Passini MA, Stewart
GR. "Gene transfer of human acid sphingomyelinase corrects neuropathology and
motor deficits in a mouse model of Niemann-Pick type A disease." PNAS
102(49):17822-7 (2005).
Feldman SA, Audet S, Beeler JA. "The fusion glycoprotein of human respiratory
syncytial virus facilitates virus attachment and infectivity via an interaction with
cellular heparan sulfate." Journal of Virology 74(14):6442-6447 (2000).
Freed EO, Risser R. "The role of envelope glycoprotein processing in murine leukemia
virus infection." Journal of Virology 61(9):2852-2856 (1987).
Gaedigk-Nitschko K, Schlesinger MJ. "The Sindbis virus 6K protein can be detected in
virions and is acylated with fatty acids." Virology 175(1):274-281 (1990).

75
Gardner CL, Choi-Nurvitadhi J, Sun C, Bayer A, Hritz J, Ryman KD, Klimstra WB.
"Natural Variation in the Heparan Sulfate Binding Domain of the Eastern Equine
Encephalitis Virus E2 Glycoprotein Alters Interactions with Cell Surfaces and
Virulence in Mice." Journal of Virology 87(15):8582-8590 (2013).
Gardner CL, Ebel GD, Ryman KD, Klimstra WB. "Heparan sulfate binding by natural
eastern equine encephalitis viruses promotes neurovirulence." Proceedings of the
National Academy of Science 108(38):16026-16031 (2011).
Gardner CL, Hritz J, Sun C, Vanlandingham DL, Song TY, Ghedin E, Higgs S, Klimstra
WB, Ryman KD. "Deliberate attenuation of chikungunya virus by adaptation to
heparan sulfate-dependent infectivity: a model for rational arboviral vaccine
design." PLOS Neglected tropical diseases 8(2):e2719 (2014).
Garoff H, Huylebroeck D, Robinson A, Tillman U, Liljestrom P. "The signal sequence of
the p62 protein of Semliki Forest virus is involved in initiation but not in
completing chain translocation." Journal of Cell Biology 111(3):867-76 (1990).
Germi R, Crance JM, Garin D, Guimet J, Lortat-Jacob H, Ruigrok RW, Zarski JP, Drouet
E. "Heparan sulfate-mediated binding of infectious dengue virus type 2 and
yellow fever virus." Virology 292(1):162-168 (2002).
Giroglou T, Florin L, Schäfer F, Streeck RE, Sapp M. "Human papillomavirus infection
requires cell surface heparan sulfate." Journal of Virology 75(3):1565-1570
(2001).

76
Hacein-Bey-Abina S, Hauer J, Lim A, Picard C, Wang GP, Berry CC, Martinache C,
Rieux-Laucat F, Latour S, Belohradsky BH, Leiva L, Sorensen R, Debre M,
Casanova JJ, Blanche S, Durandy A, Bushman FD, Fischer A, Cavazzana-Calvo
M. "Efficacy of gene therapy for X-linked severe combined immunodeficiency."
The New England Journal of Medicine 363 (2010), 355–364 (2010).
Hadigal SR, Agelidis AM, Karasneh GA, Antoine TE, Yakoub AM, Ramani VC,
Djalilian AR, Sanderson RD, Shukla D. "Heparanase is a host enzyme required
for herpes simplex virus-1 release from cells." Nature communications 6:6985
(2015).
Heil ML, Albee A,Strauss JH, Kuhn RJ. "An Amino Acid Substitution in the Coding
Region of the E2 Glycoprotein Adapts Ross River Virus To Utilize Heparan
Sulfate as an Attachment Moiety." Journal of Virology 75(14): 6303–6309
(2001).
Helenius A, Kielian MC. "Role of cholesterol in fusion of Semliki Forest virus with
membranes." Journal of Virology 52(1): 281–283. (1984).
Hua G, Kolesnick R. "Using ASMase knockout mice to model human diseases."
Handbook of Experimental Pharmacology 216:29-54 (2013).
Jan J, Byrnes AP, Griffin DE. "Characterization of a Chinese Hamster Ovary Cell Line
Developed by Retroviral Insertional Mutagenesis That Is Resistant to Sindbis
Virus Infection." Journal of Virology 73(6):4919–4924 (1999).
Jose J, Snyder JE, Kuhn RJ. "A structural and functional perspective of alphavirus
replication and assembly." Future Microbiology 4: 837–856. (2009).

77
Justman J, Klimjack MR, Kielian M. "Role of spike protein conformational changes in
fusion of Semliki Forest virus." Journal of Virology 67(12):7597-7607 (1993).
Kafri T, Blomer U, Peterson DA, Gage FH, and Verma IM. "Sustained expression of
genes delivered directly into liver and muscle by lentiviral vectors." Nature
Genetics 17:314 - 317 (1997).
Kahl CA, Marsh J, Fyffe J, Sanders DA, Cornetta K. "Human immunodeficiency virus
type 1-derived lentivirus vectors with envelope glycoproteins derived from Ross
River virus and Semliki Forest virus." Journal of Virology 78(3):1421-1430
(2004).
Kang Y, Stein CS, Heth JA, Sinn PL, Penisten AK, Staber PD, Ratliff KL, Shen H,
Barker CK, Martins I, Sharkey M, Sanders DA, McCray Jr. PB, Davidson BL. "In
Vivo Gene Transfer Using a Nonprimate Lentiviral Vector Pseudotyped with
Ross River Virus Glycoproteins." Journal of Virology 76(18):9378-9388 (2002).
Kataoka C, Y Kaname, Taguwa S, Abe T, Fukuhara T, Tani H, Moriishi K, Matsuura Y.
"Baculovirus GP64-Mediated Entry into Mammalian Cells." Journal of Virology
86: (5 )2610-2620 (2001).
Kerr PJ, Weir R C, Dalgarno L. "Ross River virus variants selected during passage in
chick embryo fibroblasts: serological, genetic, and biological changes." Virology
193:446–449 (1993).
Kielian MC, Chanel-Vos C, Liao M. "Alphavirus Entry and Membrane Fusion." Viruses
2(4): 796–825 (2010).
Kielian MC, Helenius A,. "Role of cholesterol in fusion of Semliki Forest virus with
membranes." Journal of Virology 52(1): 281–283. (1984).

78
Kielian MC, Helenius A. "pH-Iinduced alterations in the fusogenic spike protein of
Semliki Forest virus." Journal of Cell Biology 101(6):2284-2291 (1985).
Klimstra WB, Nangle EM, Smith MS, Yurochko AD, Ryman KD. "DC-SIGN and L-
SIGN can act as attachment receptors for alphaviruses and distinguish between
mosquito cell- and mammalian cell-derived viruses." Journal of Virology
77:12022–12032 (2003).
Klimstra WB, Ryman KD, Johnston RE. "Adaptation of Sindbis Virus to BHK Cells
Selects for Use of Heparan Sulfate as an Attachment Receptor." Journal of
Virology 72(9): 7357–7366 (1998).
Leung JY, Ng MM, Chu JJ. "Replication of Alphaviruses: A Review on the Entry
Process of Alphaviruses into Cells." Advances in Virology 2011(249640) (2011).
Li L, Jose J, Xiang Y, Kuhn RJ, Rossmann MG. "Structural changes of envelope proteins
during alphavirus fusion." Nature 468(7324):705-708 (2010).
Li SD, Huang L. "Gene therapy progress and prospects: non-viral gene therapy by
systemic delivery." Gene Therapy 13:1313–1319 (2006).
Liljeström P, Garoff H. "Internally located cleavable signal sequences direct the
formation of Semliki Forest virus membrane proteins from a polyprotein
precursor." Journal of Virology 65(1):147-154 (1991).
Lopes CC, Dietrich CP, Nader HB. "Specific structural features of syndecans and
heparan sulfate chains are needed for cell signaling." Brazilian Journal of
Medical and Biological Research 39: 157-167 (2006).
Lu X, Silver J. "Ecotropic Murine Leukemia Virus Receptor Is Physically Associated
with Caveolin and Membrane Rafts." Virology 276(2):251-258 (2000).

79
Lu X, Xiong Y, Silver J. "Asymmetric Requirement for Cholesterol in Receptor-Bearing
but Not Envelope-Bearing Membranes for Fusion Mediated by Ecotropic Murine
Leukemia Virus." Journal of Virology 76(13):6701-6709. (2002).
Lu YE, Cassese T, Kielian M. "The Cholesterol Requirement for Sindbis Virus Entry and
Exit and Characterization of a Spike Protein Region Involved in Cholesterol
Dependence." Journal of Virology 73(5):4272-4278 (1999).
ML, Heil. PhD Thesis. 2001.
Naldini L, Blomer U, Gallay P, Ory D, Mulligan R, Gage FH, Verma IM, Trono D. "In
vivo gene delivery and stable transduction of nondividing cells by a lentiviral
vector." Science 272(5259): 263-267 (1996).
Ng CG, Coppens I, Govindarajan D, Pisciotta J, Shulaev V, Griffin DE. "Effect of host
cell lipid metabolism on alphavirus replication, virion morphogenesis, and
infectivity." PNAS 105(42):16326–16331 (2008).
Ng CG, Griffin DE. "Acid Sphingomyelinase Deficiency Increases Susceptibility to Fatal
Alphavirus Encephalomyelitis." Jornal of Virology 80(22):10989-10999 (2006).
Nieva JL, Bron R, Corver J, Wilschut J. "Membrane fusion of Semliki Forest virus
requires sphingolipids in the target membrane." The EMBO Journal 13(12):
2797–2804 (1994).
Nitta T, Tam R, Kim JW, Fan H. "The cellular protein La functions in enhancement of
virus release through lipid rafts facilitated by murine leukemia virus glycosylated
Gag." mBio 2(1):e00341-10 (2011).

80
Noh MJ, Copeland RO, Yi Y, Choi KB, Meschter C, Hwang S, Lim CL, Yip V, Hyun JP,
Lee HY, Lee KH. "Pre-clinical studies of retrovirally transduced human
chondrocytes expressing transforming growth factor-beta-1 (TG-C)." Cytotherapy
12(3):384-393 (2010).
Periasamy M, Kalyanasundaram A. "SERCA2a Gene Therapy for Heart Failure: Ready
for Primetime?" Molecular Therapy 16 6, 1002–1004 (2008).
Phalen T, Kielien M. "Cholesterol is required for infection by Semliki Forest virus."
Journal of Cell Biology 112(4):615-623 (1991).
Pinter A, Kopelman R, Li Z, Kayman SC, Sanders, DA. "Localization of the Labile
Disulfide Bond between SU and TM of the Murine Leukemia Virus Envelope
Protein Complex to a Highly Conserved CWLC Motif in SU That Resembles the
Active-Site Sequence of Thiol-Disulfide Exchange Enzymes." Journal of
Virology 71(10): 8073–8077 (1997).
Poeschla EM, Wong-Staal F, Looney DJ. "Efficient transduction of nondividing human
cells by feline immunodeficiency virus lentiviral vectors." Nature Medicine
4(3):354-357 (1998).
Pornillos O, Garrus JE, Sundquist WI. "Mechanisms of enveloped RNA virus budding."
Trends in Cell Biology 12(12):569-579 (2002).
Puri V, Watanabe R, Dominguez M., Sun X, Wheatley CL, Marks DL, Pagano RE.
"Cholesterol modulates membrane traffic along the endocytic pathway in
sphingolipid storage diseases." Nature Cell Biology 1(6):386-8 (1999).

81
Qin J, Berdyshev E, Poirer C, Schwartz NB, Dawson G. "Neutral Sphingomyelinase 2
Deficiency Increases Hyaluronan Synthesis by Up-regulation of Hyaluronan
Synthase 2 through Decreased Ceramide Production and Activation of Akt."
Journal of Biological Chemistry 287(17):13620-13632 (2012).
Rein A, McClure MR, Rice NR, Luftig RB, Schultz AM. "Myristylation site in Pr65(gag)
is essential for virus particle formation by Moloney murine leukemia virus."
Proceedings of the National Academy of Sciences 83(19): 7246–7250 (1986).
Rein, A. "Murine Leukemia Viruses: Objects and Organisms." Advances in Virology
2011 (2011).
Salminen A, Wahlberg JM, Lobigs M, Liljeström P, Garoff H. "Membrane fusion process
of Semliki Forest virus. II: Cleavage-dependent reorganization of the spike
protein complex controls virus entry." Jornal of Cell Biology 116(2): 349–357
(1992 ).
Sanders, DA. "No false start for novel pseudotyped vectors." Current Opinion in
Biotechnology 13(5):437-42 (2002).
Schuchman EH, Desnick RJ. Niemann-Pick disease types A and B: acid
sphingomyelinase deficiencies. In: The Metabolic and Molecular Bases of
Inherited Disease. McGraw-Hill, 3589-3610, 2001.
Schwartz O, Albert ML. "Biology and pathogenesis of chikungunya virus." Nature
Reviews Microbiology 8, 491-500 (2010).
Sefton, BM. "Immediate glycosylation of Sindbis virus membrane proteins." Cell
10(4):659-668 (1977).

82
Sharkey CM, North CL, Kuhn RJ, Sanders DA. "Ross River Virus Glycoprotein-
Pseudotyped Retroviruses and Stable Cell Lines for Their Production." Journal of
Virology 75(6): 2653–2659. (2001).
Shinnick TM, Lerner RA, Sutcliffe JG. "Nucleotide sequence of Moloney murine
leukaemia virus." Nature 293: 543 - 548 (1981).
Shukla D, Liu J, Blaiklock P, Shworak NW, Bai X, Esko JD, Cohen GH, Eisenberg RJ,
Rosenberg RD, Spear PG. "A novel role for 3-O-sulfated heparan sulfate in
herpes simplex virus 1 entry." Cell 99(1):13-22 (1999).
Shukla D, Spear PG. "Herpesviruses and heparan sulfate: an intimate relationship in aid
of viral entry." The journal of Clinical investigation 108(4): 503–510 (2001).
Silverman RH, Nguyen C, Weight CJ, Klein EA. "The human retrovirus XMRV in
prostate cancer and chronic fatigue syndrome." Nature Reviews Urology 7, 392–
402 (2010).
Skehel JJ, Wiley DC. "Receptor binding and membrane fusion in virus entry: the
influenza hemagglutinin." Annual Review of Biochemistry 69:531-569 (2000).
Smith TJ, Cheng RH, Olson NH, Peterson P, Chase E, Kuhn RJ. "Putative receptor
binding sites on alphaviruses as visualized by cryoelectron microscopy."
Proceedings of the National Academy of Science 92:10648-10652 (1995).
Strauss J H, Strauss EG. "The Alphaviruses: Gene Expression, Replication, and
Evolution." Microbiolgy and Molecular Bioliology Reviews 58(3): 491–562
(1994).

83
Suchi M, Dinur T, Desnick RJ, Gatt S, Pereira L, Gilboa E, Schuchman EH. "Retroviral-
mediated transfer of the human acid sphingomyelinase cDNA: correction of the
metabolic defect in cultured Niemann-Pick disease cells." PNAS 89(8):3227-31
(1992).
Summerford C, Samulski RJ. "Membrane-associated heparan sulfate proteoglycan is a
receptor for adeno-associated virus type 2 virions." Journal of Virology
72(2):1438-45 (1998).
Taylor GM, Sanders DA. "The Role of the Membrane-spanning Domain Sequence in
Glycoprotein-mediated Membrane Fusion." Molecular Biology of the Cell 10(9):
2803–2815 (1999).
Tellinghuisen TL, Hamburger AE, Fisher BR, Ostendorp R, Kuhn RJ. "In vitro assembly
of alphavirus cores by using nucleocapsid protein expressed in Escherichia coli."
Journal of Virology 73(7):5309-5319 (1999).
Wagner R, Matrosovich M, Klenk H. "Functional balance between haemagglutinin and
neuraminidase in influenza virus infections." Reviews in Medical Virology
12(3):159-166 (2002).
Westenberg M, Soedling HM, Hirani N, Nicholson LJ, Mann DA, Dolphin CT.
"Seamless replacement of Autographa californica multiple nucleopolyhedrovirus
gp64 with each of five novel type II alphabaculovirus fusion sequences generates
pseudotyped virus that fails to transduce mammalian cells." Journal of General
Virology 93, 1583–1590 (2013).
Wirth T, Parkerb N, Ylä-Herttuala S. "History of gene therapy." Gene 525(2):162–169
(2013).

84
Wirth T, Samaranayake H, Pikkarainen J, Määttä AM, Ylä-Herttuala S. "Clinical trials
for glioblastoma multiforme using adenoviral vectors." Current Opinion in
Molecular Therapeutics 11(5):485-492. (2009).
www.genetherapynet.com/viral-vectors.html. n.d.
Yeager M, Wilson-Kubalek EM, Weiner SG, Brown PO, Rein A. "Supramolecular
organization of immature and mature murine leukemia virus revealed by electron
cryo-microscopy: implications for retroviral assembly mechanisms." Proceedings
of the National Academy of Sciences 95(13):7299-7304 (1998).
Yonezawa A, Cavrois M, Greene WC. "Studies of Ebola Virus Glycoprotein-Mediated
Entry and Fusion by Using Pseudotyped Human Immunodeficiency Virus Type 1
Virions: Involvement of Cytoskeletal Proteins and Enhancement by Tumor
Necrosis Factor Alpha." Journal of Virology 79 (2) 918-926 (2005).
Zhang W, Heil M, Kuhn RJ, Baker TS. "Heparin binding sites on Ross River virus
revealed by electron cryo-microscopy." Virology 332(2): 511–518. (2005).
Zhu W, Wang L, Yang Y, Jia J, Fu S, Feng Y, He Y, Li J, Liang G. "Interaction of E2
Glycoprotein with Heparan Sulfate Is Crucial for Cellular Infection of Sindbis
Virus." PLoS ONE 5(3): e9656 (2010).

85
VITA
Dr. Aditi Kesari was born in Ahmednagar, India on October 4, 1984. Within a
couple of years, Aditi, her parents, Mr. Shashikant Kesari and Mrs. Shubhada Kesari, and
her elder brother, Mr. Amit Kesari, moved to Pune. She did her schooling at Modern
High School. Aditi graduated from High School by standing 20th in the Merit List
amongst 200,000 students in Secondary School Board Certificate Examination, India.
She then went to Fergusson College for Higher Secondary education. Aditi appeared for
the State-level Common Entrance Test for Medicine in 2002, through which she secured
admission into B.J. Medical College, one of India’s top Medical Colleges. She received
Bachelor of Medicine and Bachelor of Surgery (MBBS) degree in 2008. She worked at
Jehangir hospital in the Cardiology department before coming to Purdue to pursue PhD.
At Purdue, Aditi joined Dr. David Sanders’ laboratory in August 2010. Her work
involved application of virology in the field of gene therapy. She studied entry
mechanism of Alphaviral glycoprotein pseudotyped viruses, which can be used as
potential gene therapy vectors. Aditi presented her work with a lecture at European
Molecular Biology Laboratory Symposium held at Germany in 2013, and at American
Society of Virology conference held at Canada in 2015. She also presented a poster at
American Society of Gene and Cell Therapy Conference in 2014.

86
Aditi met her husband, Aditya Josyula while at Purdue. They have recently
become parents to their son, Aariv and are enjoying their parenthood.






![[VII]. Regulation of Gene Expression Via Signal Transduction Reading List VII: Signal transduction Signal transduction in biological systems.](https://static.fdocuments.net/doc/165x107/56649e385503460f94b28319/vii-regulation-of-gene-expression-via-signal-transduction-reading-list-vii.jpg)











![Advanced Clinical Research Center Division of Molecular ... · VSV-pseudotyped HIV [HIV(VSV)] vectors was re-ported as a powerful tool for gene transduction into human blood cells,](https://static.fdocuments.net/doc/165x107/5ec158c5d1b32422f346db7f/advanced-clinical-research-center-division-of-molecular-vsv-pseudotyped-hiv.jpg)