EXTENT AND RATE OF CHROMOSOME SEGREGATION IN TWO ... · some number 4, were found in the hybrid...
8
J. Cell Set. 63, 215-221 (1979) 215 Printed in Great Britain © Company of Biologists Limited l<)7<) EXTENT AND RATE OF CHROMOSOME SEGREGATION IN TWO INTRASPECIFIC MOUSE CELL HYBRIDS: A^xDIPLOID FOETAL ERYTHROCYTE AND AgxBg, M. H. RUSSELL, B. J. McGEE AND E. ENGEL Departments of Anatomy and Medicine, School of Medicine, Vanderbilt University, Naslwille, Tennessee 37232, U.S.A. SUMMARY Patterns of chromosome segregation were studied in 2 different intraspecific mouse cell hybrids: (1) A 9 x B 82 , formed by fusing 2 cell lines of heteroploid fibroblasts, and (2) UWE, originating from the fusion of A, cells with euploid foetal erythrocytes. Detailed analyses of Giemsa (G)-banded chromosomes and chromosome arms of both parental and hybrid cells were made for each hybrid type, in order to determine the specificity of the losses and to assess the influence of ploidy and cell differentiation. Unlike the A, x B M hybrids, which revealed a significant chromosome loss under selective tissue culture pressures only after 9 months, the UWE hybrids showed a sharp reduction in the total chromosome number during the initial 2 months under similar pressures. However, with no additional cloning, UWE remained karyotypically stable after that time. This rapid chromo- somal segregation in UWE hybrids may be caused by properties of the parental foetal erythro- cytes. In UWE cells, the majority of the chromosome arms were retained or duplicated. Less than a quarter of the total number of chromosome arms were segregated or lost, and these were all chromosome arms with abnormal mouse G-banding patterns, present only in the heteroploid A 9 parental cells. In two of the four A 9 x B 82 hybrid lines, there was marked segregation of chromosome arms whose banding patterns were identical to those of wild type mouse telocentric chromosomes. For both types of intraspecific cell hybrids, two thirds or more of the chromo- some arms had banding patterns which were the same as those of the wild type genome. INTRODUCTION Since little is understood about the phenomenon of chromosome segregation in somatic cell hybrids, studies of 2 different intraspecific hybrids were undertaken to examine in detail chromosome loss as a function of specificity, ploidy, and cell dif- ferentiation. In intraspecific cell hybrids, usually only moderate losses of chromosomes occur over many generations (Littlefield, 1966; Yoshida & Ephrussi, 1967; Ruddle, Chen, Shows & Silagi, 1970). However, much greater chromosome losses may occur within a year in intraspecific cell hybrids giving rise to segregant cells that resemble the parental cells in total chromosome number, when the hybrid cells are placed under directed tissue culture pressures (Engel, McGee & Harris, 19696; Engel, Empson & Harris, 1971). Radically segregated hybrids arising several months after the initial cell fusion have also been described in intraspecific hepatoma crosses- subjected to selection in culture (Weiss & Chaplain, 1971).
Transcript of EXTENT AND RATE OF CHROMOSOME SEGREGATION IN TWO ... · some number 4, were found in the hybrid...

J. Cell Set. 63, 215-221 (1979) 215Printed in Great Britain © Company of Biologists Limited l<)7<)
EXTENT AND RATE OF CHROMOSOME
SEGREGATION IN TWO INTRASPECIFIC
MOUSE CELL HYBRIDS: A^xDIPLOID FOETAL
ERYTHROCYTE AND AgxBg,
M. H. RUSSELL, B. J. McGEE AND E. ENGELDepartments of Anatomy and Medicine, School of Medicine,Vanderbilt University, Naslwille, Tennessee 37232, U.S.A.
SUMMARY
Patterns of chromosome segregation were studied in 2 different intraspecific mouse cellhybrids: (1) A9 x B82, formed by fusing 2 cell lines of heteroploid fibroblasts, and (2) UWE,originating from the fusion of A, cells with euploid foetal erythrocytes. Detailed analyses ofGiemsa (G)-banded chromosomes and chromosome arms of both parental and hybrid cellswere made for each hybrid type, in order to determine the specificity of the losses and to assessthe influence of ploidy and cell differentiation.
Unlike the A, x BM hybrids, which revealed a significant chromosome loss under selectivetissue culture pressures only after 9 months, the UWE hybrids showed a sharp reduction in thetotal chromosome number during the initial 2 months under similar pressures. However, withno additional cloning, UWE remained karyotypically stable after that time. This rapid chromo-somal segregation in UWE hybrids may be caused by properties of the parental foetal erythro-cytes.
In UWE cells, the majority of the chromosome arms were retained or duplicated. Less thana quarter of the total number of chromosome arms were segregated or lost, and these were allchromosome arms with abnormal mouse G-banding patterns, present only in the heteroploidA9 parental cells. In two of the four A9 x B82 hybrid lines, there was marked segregation ofchromosome arms whose banding patterns were identical to those of wild type mouse telocentricchromosomes. For both types of intraspecific cell hybrids, two thirds or more of the chromo-some arms had banding patterns which were the same as those of the wild type genome.
INTRODUCTION
Since little is understood about the phenomenon of chromosome segregation insomatic cell hybrids, studies of 2 different intraspecific hybrids were undertaken toexamine in detail chromosome loss as a function of specificity, ploidy, and cell dif-ferentiation. In intraspecific cell hybrids, usually only moderate losses of chromosomesoccur over many generations (Littlefield, 1966; Yoshida & Ephrussi, 1967; Ruddle,Chen, Shows & Silagi, 1970). However, much greater chromosome losses may occurwithin a year in intraspecific cell hybrids giving rise to segregant cells that resemblethe parental cells in total chromosome number, when the hybrid cells are placedunder directed tissue culture pressures (Engel, McGee & Harris, 19696; Engel,Empson & Harris, 1971). Radically segregated hybrids arising several months afterthe initial cell fusion have also been described in intraspecific hepatoma crosses-subjected to selection in culture (Weiss & Chaplain, 1971).

2i6 M. H. Russell, B. J. McGee and E. Engel
Chromosome segregation was investigated in a mouse intraspecific cell hybrid(UWE), originating from the fusion of a heteroploid, differentiated fibroblast of theAB cell line with a euploid, mouse foetal erythrocyte. Patterns of chromosomesegregation for UWE were obtained by analyses of individual trypsin-Giemsa (G)-banded chromosomes and chromosome arms of both the parental and the hybridcells. The chromosome changes in UWE cells were compared to those seen in A8 x B82,another intraspecific mouse cell hybrid formed from the fusion of cells from twodifferent lines of heteroploid fibroblasts (Engel, McGee & Harris, 19690,6; Engelet al. 1971; Russell, Engel, Vaughn & McGee, 1977).
MATERIALS AND METHODS
Nucleated erythrocytes were obtained from C|H mouse embryos approximately 12 days old.The embryos were beheaded, and their blood was collected into Petri dishes containing Hanks'balanced salt solution and o-i fig/ml of heparin. To reduce contamination by other embryoniccells, the cell suspension was placed in a Falcon flask for 3 h at 37 °C, which allowed contami-nating cells to attach to the plastic substrate.
For cell fusion, i-6 x 10" nucleated erythrocytes were mixed with 5 x io6 A, cells, a derivativeof the L cell line (Littlefield, 1964), in the presence of 6000 haemagglutinating units of u.v.-inactivated Sendai virus at 4 °C for 5 min. The cell suspension was then incubated at 37 °Cfor 30 min. Three flasks, containing Minimum Essential Medium (MEM) with 15 % foetalcalf serum, were seeded. The medium was replaced by a selective medium containing hypoxan-thine, aminopterin, and thymidine (HAT) (Littlefield, 1964) with 15 % foetal calf serum after3 days. Hybrid colonies termed UWE appeared in all of the flasks; the first harvest andkaryotypic analysis by conventional staining procedures were done 3 weeks later. At that time,chromosome-banding techniques were not available. Six UWE clones were transferred fromthe primary flasks and harvested 7 weeks after the initial fusion. One clone was maintained forover 3 years in HAT before being frozen in liquid nitrogen. Two years and eight months afterthe original fusion this UWE clone and the parental Ae cell line, grown continuously in MEM,were studied by the G-banding technique as described in a previous paper (Russell et al. 1977).
Twenty high-quality G-banded metaphases were selected for both the A9 line and the UWEclone. The standardized guide for the Q- and G-bands of normal mouse telocentric chromo-somes (Committee on Standardized Genetic Nomenclature for Mice, 1972) and the characteriza-tion of the A, line by Allderdice et al. (1973) and Russell et al. (1977) were all used as referencesfor karyotyping the parental and hybrid chromosomes. The A9 cell line used in this fusion wasfound to differ significantly (P < 005) from the AB previously described (Russell et al. 1977)when the mean number of chromosomes of the groups and subgroups in each line were comparedby an analysis of variance of the completely randomized design. Although both A, cell lineshad a common origin, cultivation under different laboratory conditions appears to have resultedin different heteroploid genomes.
RESULTS
The hybrid cells
There was marked segregation of chromosomes in the UWE hybrid cells within3 to 7 weeks after the initial fusion. The sum of the 2 parental chromosome setscontributed to the UWE hybrid cell would be in the range of 90 chromosomes; but7 weeks after fusion, examination of 6 clones revealed that the number of chromo-somes per cell ranged from 63 to 72. The clone having the lowest number of

Chromosome loss in intraspecific hybrids 217
chromosomes per cell, which was approximately 63, was maintained and studied forover 3 years.
The first detailed study of this hybrid clone was done by routine staining 3 monthsafter the initial fusion. In 22 cells examined, the average total number of chromosomeswas 63-27 with a range of 44 to 77. Of the 3 major chromosome groups, the hybridcell line contained a mean number of 19-68 biarmed (B) chromosomes, 4-31 telo-centric (T) chromosomes, and 2-14 dot (D) chromosomes. There was also a fourthgroup, ring (r) chromosomes, which were found in this hybrid cell line only at thisstage; these were present at an average of 0-14 chromosomes per cell.
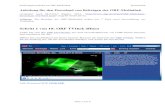
Ml >i i S it * i M • * «T t r i c M2 M3 M4 M5
9 ,0 II 12 13 m "" °
I ftII it t15 16 18 19
6/1 5/3 4/4 10/4 15/4
1.1.Bdic
Nh/Ni , M9/I4 Mh/17
f i e (I • •5/5 17/9* 16/10 Mc/Mc MIO/Md Ml/Mm Nj/Nk
Fig. 1. A G-banded karyotype of a UWE hybrid cell (chromosomes 9 and 13 havebeen combined as denoted by •). T chromosomes in the upper left-hand corner havethe same banding patterns as the wild type mouse T chromosomes. For the Bchromosomes the p arm was named first, followed by the q arm, e.g., 6/1.
This hybrid clone was further studied by the G-banding technique, 2 years and 8months after the original fusion (Fig. 1). The average total number of chromosomesfor the 20 cells examined was 63-60, with a range of 53 to 67. The mean number of Bchromosomes present was 20-15; T chromosomes, 42-05; and D chromosomes,1-40 (Table 1). The mean numbers of chromosomes of each major group found inthe previous conventional study were compared with those found in the G-banding

2i8 M. H. Russell, B. J. McGee and E. Engel
study; an analysis of variance of completely randomized design was used. There wasno significant difference in the total number of chromosomes in the B and T groups,but the D groups were significantly different (P < 0-05).
Comparison of the parental and hybrid cells
A G-banded karyotypic analysis of Ag, maintained under the same tissue cultureconditions as UWE, was made at approximately the same time as the G-bandingstudy of UWE. A9 averaged 52-80 chromosomes for 30 cells, with a range of 50 to 60chromosomes. These chromosomes were divided into 3 major groups as previouslydescribed for the hybrid cell line (Table 1). The other parental cell type, the mousefoetal erythrocyte, had a normal diploid number of 40 chromosomes, which were allT chromosomes.
Table 1. Average chromosome number in major groups and subgroupsfor parental and hybrid cells
Celltype
A.E*Foetal mouse
rbcA,E/foetal mouse
rbc#
TotalB
24-500
20-15
M
2 1 0
0
i-oo
>
5-250
5-00
SM
17-150
I4"i5
TotalT
26950
42-05
TotalD
i-350
1-40
Totalchrom.
52804 0
6360
• Twenty cells analysed for each cell type 2 years and 8 months after the initial fusion-B, biarmed; M, metacentric; i, isochromosome; SM, submetacentric; T, telocentric;
D, dot-like; chrom., chromosomes.
The mean numbers of chromosomes in the major groups and subgroups of AB
and of the hybrid clone were compared by the analysis of variance. The B and Tchromosome groups of A9 and of UWE were significantly different (P < 0-05), whilethere was no significant difference for the D chromosome group. The metacentric andsubmetacentric subgroups were significantly different (P< 0-05) for these 2 celllines. However, there was no significant difference for the isochromosomes.
All of the normal mouse T chromosomes of the diploid karyotype, except chromo-some number 4, were found in the hybrid cells as normal T chromosomes. Over 73 %of the total number of T chromosomes in UWE were normal mouse chromosomes;this figure was 100% for foetal mouse erythrocytes and 60% for A9. These percen-tages were similar for chromosome arms derived from normal T chromosomes.
To detect overall patterns of duplication, retention, segregation, or complete lossof redundant chromosome material in UWE hybrid cells, the sums of the parentalchromosome arms, representing the state of affairs immediately after cell fusion, werecompared with the corresponding number of chromosome arms in the hybrids. Thisprocedure has been previously described for Ae x B82 cell hybrids (Russell et al.
1977)-When the identifiable chromosome arms in the UWE hybrid cells were compared

Chromosome loss in intraspecific hybrids 219
with the sum of the identifiable arms in A9 and foetal erythrocytes, 60% were foundto be duplicated and 40% retained. Segregation occurred in 14% of the unidentifi-able chromosome arms. Also, 14% of the unidentifiable arms had new bandingpatterns not previously recognized in A .̂ Over one fifth of the unidentifiable armswere lost, one quarter retained, and an additional quarter duplicated. Replicationof the identifiable arms was much greater than that of the unidentifiable arms.
Over 75 % of the total number of chromosome arms, including both the identi-fiable and unidentifiable, in the hybrid cells were retained or duplicated. However,more than 20% of the total chromosome arms that were segregated or lost werechromosomes of unidentifiable origin present in the heteroploid AB parent cells.
DISCUSSION
Marked segregation of chromosomes appeared during the early phase of growthof intraspecific hybrid cells formed between heteroploid cells of a permanent mousecell line and euploid, nucleated foetal mouse blood cells. Approximately 20 to 30chromosomes were lost from these cells 7 weeks after the initial fusion. From theanalyses of the major chromosome groups, it appeared that the hybrids remainedkaryotypically stable for the majority of the B and T chromosomes for a period ofover two and a half years thereafter. When the hybrid chromosomes for the majorgroups and subgroups were compared to what would be expected for the sum of theparental chromosome groups, there was a loss of chromosomes in the B groups,including the metacentric and submetacentric subgroups, and in the T group, sothat the total number of chromosomes in the hybrid cell approached that of one of theparent cells.
In the UWE cells, duplication and retention of chromosome arms occurred in overthree quarters of the total number of identifiable and unidentifiable arms: less thana quarter of the total number of chromosome arms were segregated or lost. It isimportant to note that only unidentifiable chromosome arms were segregated orlost, and this loss was not balanced by the appearance of new chromosome materialin the hybrid cells. The data thus suggest that the losses mainly involved abnormalchromosomes derived from the A9 parent cell.
Hybrid cells of A9 x B82 showed little chromosome loss during the early phase ofgrowth in vitro (Engel et al. 1969 a), but under selective pressure in vitro over aprolonged period of time, highly significant losses occurred (Engel et al. 19696, 1971).
This slow but substantial, chromosome segregation in the heteroploid x heteroploidhybrid cells is in marked contrast to the rapid loss (within a 2-month period) ofapproximately one third of the chromosomes in the heteroploid x euploid cells ofUWE.
Over two thirds of the chromosome arms in both the parental Afl and B82 lines andin the four A9 x B82 hybrid lines were shown by G-banding analysis to be identicalin banding patterns to the normal mouse T chromosomes. Similarly, two thirds ofthe total chromosome arms in UWE cells were of normal mouse origin.
G-banding studies of four A^ x B82 hybrid lines revealed that the majority of theredundant chromosome arms were of identifiable, normal mouse origin; and these

220 M. H. Russell, B. J. McGee and E. Engel
tended to be segregated in two of the hybrid cells lines, but retained in the other two(Russell et al. 1977). This analysis of A9 x B82 demonstrated that segregation andchromosome recombination accounted for the evolution of the karyotype in the hybridcells. This could result in a reduction of the chromosome modes to those of the parentalcells, and in an increased heterogeneity in the distribution of chromosome classes.
Ploidy and cell differentiation both appear to be important factors in determiningchromosome loss in these intraspecific hybrids. According to Handmaker (1973), inintraspecific hybrids an unknown property that confers karyotypic stability upon aeuploid cell at mitosis is either lost or suppressed when that cell is fused with a hetero-ploid one. However, fusion of euploid, differentiated cells with heteroploid, dif-ferentiated cells of the same species can result in hybrids that appear karyotypicallystable (Hashmi & Miller, 1976; Szpirer, Szpirer & Wiener, 1976; le Borgne deKaoiiel, Billard & Macieira-Coelho, 1978). Generally, chromosome loss is small inintraspecific, differentiated euploid x euploid cell hybrids (Handmaker, 1973; Migeon,Norum & Corsaro, 1974; Hoehn et al. 1975); although, recently, recombinant euploidhybrids have been recovered from the fusion of two euploid Indian muntjac cell lines(Yoshida & Sasaki, 1978).
It has been observed in interspecific cells hybrids formed from the fusion of chickerythrocytes with both heteroploid mouse cells and Chinese hamster cells that thereactivated erythrocyte nucleus undergoes premature chromosome condensation,which may result in the loss of almost all the chick chromosome material in the newlyformed hybrid cell (Schwartz, Cook & Harris, 1971; Boyd & Harris, 1973). Similarresults were obtained when frog erythrocytes were fused with heteroploid mousecells (P. R. Cook, personal communication). Premature chromosome condensationof the foetal erythrocyte may also occur in intraspecific cell hybrids like UWE andgive rise to increased chromosome loss in the early phase of growth. Chromosomeloss, although not as great as that seen in UWE, has been reported in an intraspecifichybrid formed between an aneuploid murine foetal liver cell line and a mutant Lcell line, 5 months after initial fusion (Rintoul, Colofiore & Morrow, 1973).
Karyotypic analyses need to be done on intraspecific hybrids between diploid,undifferentiated cells and heteroploid, differentiated cells, and between diploid,undifferentiated cells crossed with themselves. Such studies might help to elucidatefurther the influence of ploidy and cell differentiation on chromosome segregation inintraspecific hybrid cells. It is clear, however, that some intraspecific hybrid combina-tions may be quite suitable for the purposes of segregation analysis and gene mapping.
This work was initiated by E. E. while on sabbatical leave at the department of Dr. HenryHarris at the Sir William Dunn School of Pathology, Oxford, England. The study was supportedby a grant from the United States Public Health Services (M. C. T.-000423-15). M. H. R.is a Fellow of the National Institute of Child Health and Human Development, N. I. H.Training Grant HD07043.

Chromosome loss in intraspecific hybrids 221
REFERENCES
ALLDERDICE, P. W., MILLER, O. J., MILLER, D. A., WARBURTON, D., PEARSON, P. L., KLEIN, G.& HARRIS, H. (1973). Chromosome analysis of two related heteroploid mouse cell lines byquinacrine fluorescence. J. Cell Set. 12, 263-274.
BOYD, Y. & HARRIS, H. (1973). Correction of genetic defects in mammalian cells by the inputof small amounts of foreign genetic material. J. Cell Set. 13, 841-861.
COMMITTEE ON STANDARDIZED GENETIC NOMENCLATURE FOR MICE (1972). Standard karyotypeof the mouse, Mus musculus. J. Hered. 63, 69-72.
ENGEL, E., EMPSON, J. & HARRIS, H. (1971). Isolation and karyotypic characterization ofsegregants of intraspecific hybrid somatic cells. Expl Cell Res. 68, 231-234.
ENGEL, E., MCGEE, B. J. & HARRIS, H. (1969a). Cytogenetic and nuclear studies on A, andB81 fused together by Sendai virus: the early phase. J. Cell Set. 5, 93-100.
ENGEL, E., MCGEE, B. ] . & HARRIS, H. (19696). Recombination and segregation in somaticcell hybrids. Nature, Land. 223, 152-155.
HANDMAKER, S. D. (1973). Hybridization of eukaryotic cells. A. Rev. Microbiol. 27, 189—204.HASHMI, S. & MILLER, O. J. (1976). Further evidence of X-linkage of hypoxanthine phos-
phoribosyl transferase in the mouse. Cytogenet. Cell Genet. 17, 35-41.HOEHN, H., BRYANT, E. M., JOHNSTON, P., NORWOOD, T. H. & MARTIN, G. M. (1975). Non-
selective isolation, stability and longevity of hybrids between normal human somatic cells.Nature, Lond. 258, 608-610.
DE KAOOEL, LE BORGNE, C, BILLARD, C. & MACIEIRA-COELHO, A. (1978). Growth characteris-tics in vitro of hybrids between normal and transformed cell lines. Int.J. Cancer 21, 338-347.
LlTTLEFIELD, J. W. (1964). Selection of hybrids from mating of fibroblasts in vitro and theirpresumed recombinants. Science, N. Y. 145, 709—710.
LlTTLEFIELD, J. W. (1966). The use of drug-resistant markers to study the hybridization ofmouse fibroblasts. Expl Cell Res. 41, 190-196.
MIGEON, B. R., NORUM, R. A. & CORSARO, C. M. (1974). Isolation and analysis of somatichybrids derived from two human diploid cells. Proc. natn. Acad. Set. U.S.A. 71, 937-941.
RlNTOUL, D., COLOFIORE, J. & MORROW, J. (1973). Expression of differentiated propertiesin fetal liver cells and their somatic cell hybrids. Expl Cell Res. 78, 414-422.
RUDDLE, F. H., CHEN, T., SHOWS, T. B. & SILAGI, S. (1970). Interstrain somatic cell hybridsin the mouse. Chromosome and enzyme analyses. Expl Cell Res. 60, 139-147.
RUSSELL, M. H., ENGEL, E., VAUGHN, W. K. & MCGEE, B. J. (1977). Karyotypic analyses ofparental and hybrid intraspecific mouse cells, A9/Be,, by Giemsa- and centromeric-banding.Jf. Cell Set. 25, 59-71.
SCHWARTZ, A. G., COOK, P. R. & HARRIS, H. (1971). Correction of a genetic defect in a mam-malian cell. Nature, New Biol. 230, 5-7.
SZPIRER, C , SZPIRER, J. & WIENER, F. (1976). The expression of differentiated functions insomatic cell hybrids: retention and activation of C, production. Differentiation 5, 139-149.
WEISS, M. C. & CHAPLAIN, M. (1971). Expression of differentiated functions in hepatoma cellhrbrids: reappearance of tyrosine aminotransferase inducibility after the loss of chromosomes.Proc. natn. Acad. Sci. U.S.A. 68, 3026-3030.
YOSHIDA, M. C. & EPHRUSSI, B. (1967). Isolation and karyological characteristics of sevenhybrids between somatic mouse cells in vitro. J. cell. Physiol. 69, 33-34.
YOSHIDA, M. C. & SASAKI, M. (1978). Euploid somatic recombinants with two active X orXYt Y, chromosomes isolated from cultured male Indian muntjac cells after HVJ virusfusion, and their use for gene assignment. Somat. Cell Genet. 4, 437-450.
{Received 1 September 1978 - Revised 12 October 1978)
IJ CKL 36