
Expression of GARP selectively identifies activated human FOXP3 … · 2009-08-13 · vation (Fig....
6
Expression of GARP selectively identifies activated human FOXP3 regulatory T cells Rui Wang a , Lina Kozhaya a , Frances Mercer a , Alka Khaitan a,b , Hodaka Fujii c,d,e , and Derya Unutmaz a,c,d,1 Departments of a Microbiology, c Pathology, and b Pediatrics, d NYU Cancer Institute and New York University School of Medicine, New York, NY, 10016; and e Combined Program on Microbiology and Immunology, Research Institute for Microbial Diseases, Osaka University, Suita 565-0871, Japan Edited by Dan R. Littman, New York University Medical Center, New York, NY, and approved June 4, 2009 (received for review February 22, 2009) The molecules that define human regulatory T cells (Tregs) phe- notypically and functionally remain to be fully characterized. We recently showed that activated human Tregs express mRNA for a transmembrane protein called glycoprotein A repetitions predom- inant (GARP, or LRRC32). Here, using a GARP-specific mAb, we demonstrate that expression of GARP on activated Tregs correlates with their suppressive capacity. However, GARP was not induced on T cells activated in the presence of TGF, which expressed high levels of FOXP3 and lacked suppressive function. Ectopic expres- sion of FOXP3 in conventional T cells was also insufficient for induction of GARP expression in most donors. Functionally, silenc- ing GARP in Tregs only moderately attenuated their suppressive activity. CD25 T cells sorted for high GARP expression displayed more potent suppressive activity compared with CD25GARP cells. Remarkably, CD25GARP T cells expanded in culture con- tained 3–5 fold higher IL-17-secreting cells compared with either CD25GARP or CD25GARP cells, suggesting that high GARP expression can potentially discriminate Tregs from those that have switched to Th17 lineage. We also determined whether GARP expression correlates with FOXP3-expressing T cells in human immunodeficiency virus (HIV) infected subjects. A subset of HIV individuals with high percentages of FOXP3 T cells did not show proportionate increase in GARP T cells. This finding suggests that higher FOXP3 levels observed in these HIV individuals is possibly due to immune activation rather than to an increase in Tregs. Our findings highlight the significance of GARP both in dissecting duality of Treg/Th17 cell differentiation and as a marker to identify bona fide Tregs during diseases with chronic immune activation. Foxp3 HIV human TGFbeta Tregs R egulatory T cells (Tregs) play a key role in maintaining peripheral tolerance, preventing autoimmune diseases, and limiting chronic inflammatory diseases. Naturally occurring Tregs, among different subsets of suppressor T cells, are cur- rently defined by expression of the transcription factor FOXP3 and a variety of T-cell activation markers (1, 2). Functionally, Tregs respond poorly to TCR activation in terms of cytokine secretion and proliferation, and possess the ability to suppress the immune responses of effector cells (1, 2). In recent years, a myriad of mechanisms of Treg-mediated regulation have been identified (2). However, it is still unclear whether there are additional, yet-to-be-identified molecular pathways that can mediate Treg inhibitory activity. Naturally occurring Tregs express high levels of FOXP3, a master transcription factor known to be necessary for Treg development and function (1, 2). Ectopic overexpression of FOXP3 in naive T cells is able to partially endow these cells with Treg characteristics (35). TGF-treated naive T cells in both the human and murine systems can also up-regulate FOXP3 expression (68). Further- more, TGF was shown to induce contact-dependent suppressive activity in human naive T cells (9). However, in contrast to murine T cells, TGF-induced FOXP3 appears to be insufficient to confer a regulatory phenotype in human T cells (8). This finding renders FOXP3 an unreliable marker to define human T cells with sup- pressive ability. Other molecules that have been shown to be differentially expressed on Tregs, including CD25, CD62L, CTLA-4, and CD127, are modified during T-cell activation or differentiation, and under chronic immune activation conditions these may not adequately discriminate Tregs from recently acti- vated T cells (1, 2). We recently identified a leucine rich repeat (LRR)containing molecule called LRRC32 or GARP, which is highly expressed at the mRNA level in activated naturally occurring Tregs (10). Here we show that GARP identifies a subset of activated FOXP3 T cells with high suppressive capacity and discriminates FOXP3 non-suppressor TGF- treated T cells from bona fide Tregs. We also found that sorting for GARP expression excludes most IL-17–secreting cells within the CD25 T-cell subset. In addition, we show the major discordance between FOXP3 and GARP expression in HIV- infected individuals, where a portion of the FOXP3 T cells are possibly recently activated cells masquerading as Tregs. These findings reveal GARP as a highly specific molecule for activated Tregs, which could play an important role in their identification and function. Results Expression of GARP on Treg Cells. In the periphery, tolerance to self-antigens is partly mediated by naturally occurring Tregs. However, effector molecules expressed on the cell surface of activated Tregs are not fully determined. To search for novel molecules expressed on activated Tregs, we performed a mi- croarray analysis and identified an mRNA encoding a cell surface protein named GARP that is specifically expressed on Tregs post–TCR activation (10). Recently, a specific mAb against GARP was made available, which we positively con- firmed on GARP-transduced Jurkat T cells [supporting infor- mation (SI) Fig. S1 A]. Using this antibody, we confirmed our previous finding (10) that GARP is expressed at high levels specifically on human Tregs but not conventional T-cell subsets (Fig. 1A). Furthermore, GARP expression was restricted to the FOXP3 population (Fig. 1B). We next explored the kinetics of GARP expression in differ- ent T-cell subsets. In Tregs, GARP was up-regulated quickly after TCR stimulation and maintained on most Tregs for 2 days before the expression levels declined (Fig. 1C). In naive Tregs, defined as CD45RO-CD45RACD25FOXP3 T cells (TNregs), GARP expression was induced 1 day postactivation, and reached a level similar to mature Tregs on day 2 (Fig. 1C). We and others have also shown that after in vitro expansion, the TNregs differentiate into mature Tregs (11–14). GARP expres- sion was up-regulated on in vitro expanded Tregs upon reacti- Author contributions: D.U. designed research; R.W., L.K., and F.M. performed research; R.W., F.M., and D.U. analyzed data; A.K. and H.F. contributed new reagents/analytic tools; and R.W. and D.U. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/cgi/content/full/ 0901965106/DCSupplemental. www.pnas.orgcgidoi10.1073pnas.0901965106 PNAS August 11, 2009 vol. 106 no. 32 13439 –13444 IMMUNOLOGY Downloaded by guest on March 15, 2020
Transcript of Expression of GARP selectively identifies activated human FOXP3 … · 2009-08-13 · vation (Fig....

Expression of GARP selectively identifies activatedhuman FOXP3� regulatory T cellsRui Wanga, Lina Kozhayaa, Frances Mercera, Alka Khaitana,b, Hodaka Fujiic,d,e, and Derya Unutmaza,c,d,1
Departments of aMicrobiology, cPathology, and bPediatrics, dNYU Cancer Institute and New York University School of Medicine, New York, NY, 10016;and eCombined Program on Microbiology and Immunology, Research Institute for Microbial Diseases, Osaka University, Suita 565-0871, Japan
Edited by Dan R. Littman, New York University Medical Center, New York, NY, and approved June 4, 2009 (received for review February 22, 2009)
The molecules that define human regulatory T cells (Tregs) phe-notypically and functionally remain to be fully characterized. Werecently showed that activated human Tregs express mRNA for atransmembrane protein called glycoprotein A repetitions predom-inant (GARP, or LRRC32). Here, using a GARP-specific mAb, wedemonstrate that expression of GARP on activated Tregs correlateswith their suppressive capacity. However, GARP was not inducedon T cells activated in the presence of TGF�, which expressed highlevels of FOXP3 and lacked suppressive function. Ectopic expres-sion of FOXP3 in conventional T cells was also insufficient forinduction of GARP expression in most donors. Functionally, silenc-ing GARP in Tregs only moderately attenuated their suppressiveactivity. CD25� T cells sorted for high GARP expression displayedmore potent suppressive activity compared with CD25�GARP�cells. Remarkably, CD25�GARP� T cells expanded in culture con-tained 3–5 fold higher IL-17-secreting cells compared with eitherCD25�GARP� or CD25�GARP� cells, suggesting that high GARPexpression can potentially discriminate Tregs from those that haveswitched to Th17 lineage. We also determined whether GARPexpression correlates with FOXP3-expressing T cells in humanimmunodeficiency virus (HIV) �infected subjects. A subset of HIV�individuals with high percentages of FOXP3� T cells did not showproportionate increase in GARP� T cells. This finding suggests thathigher FOXP3 levels observed in these HIV� individuals is possiblydue to immune activation rather than to an increase in Tregs. Ourfindings highlight the significance of GARP both in dissectingduality of Treg/Th17 cell differentiation and as a marker to identifybona fide Tregs during diseases with chronic immune activation.
Foxp3 � HIV � human � TGFbeta � Tregs
Regulatory T cells (Tregs) play a key role in maintainingperipheral tolerance, preventing autoimmune diseases, and
limiting chronic inflammatory diseases. Naturally occurringTregs, among different subsets of suppressor T cells, are cur-rently defined by expression of the transcription factor FOXP3and a variety of T-cell activation markers (1, 2). Functionally,Tregs respond poorly to TCR activation in terms of cytokinesecretion and proliferation, and possess the ability to suppressthe immune responses of effector cells (1, 2). In recent years, amyriad of mechanisms of Treg-mediated regulation have beenidentified (2). However, it is still unclear whether there areadditional, yet-to-be-identified molecular pathways that canmediate Treg inhibitory activity.
Naturally occurring Tregs express high levels of FOXP3, a mastertranscription factor known to be necessary for Treg developmentand function (1, 2). Ectopic overexpression of FOXP3 in naive Tcells is able to partially endow these cells with Treg characteristics(3�5). TGF�-treated naive T cells in both the human and murinesystems can also up-regulate FOXP3 expression (6�8). Further-more, TGF� was shown to induce contact-dependent suppressiveactivity in human naive T cells (9). However, in contrast to murineT cells, TGF�-induced FOXP3 appears to be insufficient to confera regulatory phenotype in human T cells (8). This finding rendersFOXP3 an unreliable marker to define human T cells with sup-pressive ability. Other molecules that have been shown to be
differentially expressed on Tregs, including CD25, CD62L,CTLA-4, and CD127, are modified during T-cell activation ordifferentiation, and under chronic immune activation conditionsthese may not adequately discriminate Tregs from recently acti-vated T cells (1, 2).
We recently identified a leucine rich repeat(LRR)�containing molecule called LRRC32 or GARP, whichis highly expressed at the mRNA level in activated naturallyoccurring Tregs (10). Here we show that GARP identifies asubset of activated FOXP3� T cells with high suppressivecapacity and discriminates FOXP3� non-suppressor TGF�-treated T cells from bona fide Tregs. We also found that sortingfor GARP expression excludes most IL-17–secreting cells withinthe CD25� T-cell subset. In addition, we show the majordiscordance between FOXP3 and GARP expression in HIV-infected individuals, where a portion of the FOXP3� T cells arepossibly recently activated cells masquerading as Tregs. Thesefindings reveal GARP as a highly specific molecule for activatedTregs, which could play an important role in their identificationand function.
ResultsExpression of GARP on Treg Cells. In the periphery, tolerance toself-antigens is partly mediated by naturally occurring Tregs.However, effector molecules expressed on the cell surface ofactivated Tregs are not fully determined. To search for novelmolecules expressed on activated Tregs, we performed a mi-croarray analysis and identified an mRNA encoding a cellsurface protein named GARP that is specifically expressed onTregs post–TCR activation (10). Recently, a specific mAbagainst GARP was made available, which we positively con-firmed on GARP-transduced Jurkat T cells [supporting infor-mation (SI) Fig. S1 A]. Using this antibody, we confirmed ourprevious finding (10) that GARP is expressed at high levelsspecifically on human Tregs but not conventional T-cell subsets(Fig. 1A). Furthermore, GARP expression was restricted to theFOXP3� population (Fig. 1B).
We next explored the kinetics of GARP expression in differ-ent T-cell subsets. In Tregs, GARP was up-regulated quicklyafter TCR stimulation and maintained on most Tregs for 2 daysbefore the expression levels declined (Fig. 1C). In naive Tregs,defined as CD45RO-CD45RA�CD25�FOXP3� T cells(TNregs), GARP expression was induced 1 day postactivation,and reached a level similar to mature Tregs on day 2 (Fig. 1C).We and others have also shown that after in vitro expansion, theTNregs differentiate into mature Tregs (11–14). GARP expres-sion was up-regulated on in vitro expanded Tregs upon reacti-
Author contributions: D.U. designed research; R.W., L.K., and F.M. performed research;R.W., F.M., and D.U. analyzed data; A.K. and H.F. contributed new reagents/analytic tools;and R.W. and D.U. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0901965106/DCSupplemental.
www.pnas.org�cgi�doi�10.1073�pnas.0901965106 PNAS � August 11, 2009 � vol. 106 � no. 32 � 13439–13444
IMM
UN
OLO
GY
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

vation (Fig. 1C Lower), comparable to ex vivo analyzed Tregs(Fig. 1C Upper).
Together, these findings suggested that GARP is preferen-tially induced on T cells with Treg phenotype. Therefore, westudied the possibility of using GARP as a marker for activatedT cells with suppressor function. We first activated CD4� T cellsthrough their TCR and stained for GARP expression. Asexpected, most cells also up-regulated CD25 (Fig. 1D). A portionof CD25� T cells also expressed GARP after TCR-activation
(Fig. 1D). We then sorted these cells based on GARP and CD25expression (Fig. 1D). More than 90% of the GARP�CD25�sorted T cells also expressed FOXP3 (Fig. S1B). The sorted cellswere rested for 1 week in IL-2–containing media and thenreactivated to determine their suppressive activity. We foundthat the GARP�CD25� cells very strongly inhibited the pro-liferation of TCR-stimulated target cells at different target tosuppressor ratios (Fig. 1E). The GARP�CD25� cells alsosecreted significantly less IL-2 and IFN� as compared with theGARP-CD25� population, a characteristic of Tregs (Fig. S1C).Together, our data show that GARP identifies suppressor T cellswithin the activated CD4� T-cell subset.
GARP Plays a Role in Treg-Mediated Suppression. Because almost allGARP� T cells also express FOXP3, we hypothesized thatGARP is an effector molecule downstream of FOXP3 expres-sion and investigated whether FOXP3 is required for GARPinduction in Tregs. We expressed two different shRNAs againstFOXP3 in expanded Tregs, which have been shown to knockdown FOXP3 efficiently (Fig. 2A) (11). The FOXP3 shRNA-
0.4 1.3 24 72
TN TM TNreg Treg
GARP
#Cells
B
A
FOXP3
GARP
Teff Expanded Treg
Isotype
100 101 102 103 104100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104100
101
102
103
104
2.7
27
78
13
0.1
30
0.3
94
TNTMTNregTreg
Time (d): 0 1 2 3 4 6
60
40
20
0%GARP+cells
Time (d): 0 1 2 4 5 6 8
%GARP+cells
80
60
40
20
0
Teff
ExpandedTreg
C
100 101 102 103 104100
101
102
103
104
CD25
GARP
GARP+CD25+
D EE
Fig. 1. GARP is specifically expressed on activated Tregs and defines sup-pressor T cells. (A) Surface expression of GARP on TN (naive), TM (memory),TNreg, and Tregs. Different T-cell subsets were isolated based on CD25 andCD45RO expression as previously described (10). Cells were stimulated withanti-CD3 and anti-CD28 beads (TCR stimulation) overnight and stained forsurface GARP. (B) GARP expression restricted to FOXP3� cells. TN and TNregswere activated and expanded in vitro for 2 weeks to obtain Teff and expandedTregs. Cells were activated through TCR and stained for GARP/isotype andFOXP3. (C) Kinetics of GARP expression on T-cell subset postactivation. Datarepresent percentage of GARP� T cells at different time points postactivation.(D) Induction and sorting of GARP� T cells. CD4� T cells were stimulatedthrough TCR for 2 days and stained for GARP and CD25. (E) Suppressivefunction of GARP� T cells. GARP�CD25� and GARP-CD25� populations weresorted 2 days post–TCR activation, rested in culture for 8–10 days, and assayedfor suppressive activity using CFSE-labeled resting target T cells and differentratios of GARP� suppressor or GARP� effector T cells. The cells were thenstimulated with anti-CD3 antibody (50 ng/ml) in the presence of dendritic cellsfor 4 days, and proliferating cells were determined based on decrease in CFSEexpression as shown in Fig. S1D. Percent suppression was calculated based onCFSE dilution of target T cells in suppression culture as described before (10).Briefly, the percentage of target cells that undergo division in response to thestimuli alone, in the presence of expanded Tregs, or control Teff cells, wasdetermined. The percent suppression was then calculated by percent reduc-tion in proliferation of the target cells with suppressors as compared withtarget cells alone. All data shown are representative of at least three exper-iments performed with T cells from different healthy donors.
1.1 66 5.2 10
GARP
Control sh Control sh GARP sh1 GARP sh2
Teff Expanded Treg
#Cells
41 72 76 78
FOXP3
#Cells
%Suppression
80
60
40
20
0
< 0.001P < 0.001
Teff Expanded TregCont sh Cont sh GARPsh1 GARPsh2
A
B
C
2.7 68 45 30
#Cells
GARP
Control sh Control sh Foxp3 sh1 Foxp3 sh2
Teff Expanded Treg
43 71 27 5.5#Cells
FOXP3
Fig. 2. Silencing FOXP3 and GARP expression in Tregs. (A) Silencing FOXP3in Tregs through shRNAs. Purified TN or TNreg cells were stimulated andtransduced with control or FOXP3 shRNA encoding lentiviruses. Cells wereexpanded in vitro for 2 weeks, sorted based on GFP expression (expressed byvectors) and FOXP3 levels, and induction of GARP expression was determined2 days post–TCR stimulation. (B) Knockdown of GARP by shRNAs in expandedTregs. Cells were transduced with viruses encoding control or GARP shRNA,expanded for 2 weeks in vitro, and sorted for GFP expression encoded by thesame lentiviral vector. Cells were then stained for FOXP3. GARP was stained 24hours post–TCR reactivation. Data shown are representative of three healthydonors. (C) Suppressive activity of control or shRNA-expressing Teff and Tregswere assessed as described in Fig. 1. Representative data using a 1-to-4 ratioof Treg/Teff to target T cells are shown.
13440 � www.pnas.org�cgi�doi�10.1073�pnas.0901965106 Wang et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

expressing expanded Tregs showed a reduction of GARP up-regulation in comparison to control shRNA-expressing Tregs(Fig. 2 A). In contrast, knockdown of GARP did not changeFOXP3 expression in expanded Tregs (Fig. 2B).
We have shown before that overexpression of GARP in naivehuman T cells endows them with partial Treg phenotype (10). Ina reverse experiment, we found that silencing GARP inductionby shRNA expression in expanded Tregs resulted in modest butsignificant impairment in suppressive function (Fig. 2C). Thus,GARP could contribute to Treg-mediated suppression but doesnot account for the majority of their regulatory activity underour in vitro conditions. Silencing GARP in Tregs did not affecttheir low cytokine secretion characteristic (Fig. S2).
TGF�-Induced FOXP3� Human T Cells Do Not Express GARP. TGF�has been shown to induce FOXP3 expression in both human andmurine T cells (6–8). Although TGF� is shown to differentiatenaive T cells into functional Tregs in mice, in the human systemTGF�-induced FOXP3� cells do not display suppressive activity(8, 15). We showed that addition of TGF� during activation ofCD25� naive human T cells induces FOXP3 expression in amajority of these cells (Fig. 3A), which was maintained at highlevels for at least 1 week (Fig. S3). However, despite expressingFOXP3 at levels comparable to those of natural Tregs, theTGF�-treated cells did not express GARP protein on the cellsurface upon TCR activation (Fig. 3B) or GARP mRNA asdetermined by real-time polymerase chain reaction (PCR) anal-ysis. TGF�-treated human T cells also did not show suppressiveactivity in vitro (Fig. 3C), consistent with a previous report (8).We conclude that TGF�-induced FOXP3 is not sufficient forGARP expression. Consistent with this finding, we found that inT cells overexpressing FOXP3, at levels higher than those ofnatural Tregs (Fig. 3D), induction of GARP post–TCR activa-tion was either completely absent (Fig. 3D) or was present atmuch lower levels compared with Tregs (Fig. 3E). Together,these findings suggest that although FOXP3 is important, clearlyit is not sufficient to program the cells to induce GARPexpression.
GARP Expression Defines Suppressive Activated Tregs and ExcludesIL-17–Secreting CD25� T Cells. An important implication of theabove results is that TGF�-induced FOXP3 greatly confoundsanalysis of suppressive natural Tregs during disease conditions orinfections in humans, where T cells could be activated to induceFOXP3 (in the presence of TGF�) without differentiating intoTregs. We therefore further explored whether GARP can be amore reliable marker for determining Tregs after immuneactivation. To address this question, we first isolatedCD4�CD25� (Teff) and CD4�CD25� (canonical Tregs) Tcells, which were stimulated through TCR for 48 hours, and thenfurther sorted based on GARP expression (GARP� andGARP� respectively) (Fig. 4A). The CD25� Teff cells, which donot express GARP, were also mock sorted and used as controlcells. Because GARP-sorted T cells were activated to induceGARP expression, they were cultured and rested in IL-2–containing media for 10 days and then activated through theTCR to assess their suppressive capacity. CD25�GARP� weremore potent in suppressing the proliferation of the target T cellsas compared with the CD25�GARP� subset (Fig. 4B and Fig.S4). Teff control cells did not show any suppressive activity atdifferent suppressor-to-target ratios (Fig. 4B and Fig. S4)
In two recent reports, it was shown that a portion of humanCD25� or FOXP3� T cells contain high levels of IL-17–secreting Th17 cells (16, 17). Therefore we determined thepercentages of IL-17 and IFN�-secreting cells in GARP� andGARP� subsets, which were sorted from CD4�CD25� T cells(Fig. 4A). These sorted cells were activated with phorbol my-ristate acetate and ionomycin and stained with FOXP3, IL-17,
and IFN�. We found that most CD25�GARP� cells remainedFOXP3� and that a small portion (�1–2%) expressed IL-17 atlevels lower or comparable to those in CD25� Teff cells (Fig.4C). Remarkably, CD25�GARP� sorted cells contained muchgreater percentage of IL-17–secreting cells, which was 3–4-foldhigher compared with CD25� Teff cells (Fig. 4C).CD25�GARP� cells also expressed much lower levels ofIFN�� cells compared with CD25�GARP� cells (Fig. 4D). Incontrast to IL-17 expression, CD25�GARP� T cells also did notdisplay higher IFN�� cells compared with CD25�GARP� Teffcells (Fig. 4D). Together, these findings suggest that GARP notonly consistently defines T cells with the highest suppressive
26 79
FOXP3
#Cells
TGFβ: - +A B
#Cells
TGFβ: - +
CFSE
D
0.4 0.6
TGFβ: - +
#Cells
GARP
C
E
75
No Effector
72 70
FOXP3
#Cells Treg
FOXP3+Control
GARP
Fig. 3. Expression of GARP in TGF�-treated or FOXP3-overexpressing T cells.(A) Induction of FOXP3 by TGF� in TCR-stimulated T cells. TN cells wereactivated through TCR in the presence or absence of 20 ng/ml TGF�. FOXP3expression was determined by intracellular staining at different time pointspostactivation. Data shown are representative of day 6 postactivation. (B)Expression of GARP on TGF�-treated cells. TGF�-treated or untreated cellswere restimulated through TCR at different time points between 6 and 12 daysof culture post–original activation, and GARP expression was determinedafter 2 days. A representative GARP staining performed post–day 6 of firstactivation is shown. (C) Suppressive activity of TGF�-treated T cells. ActivatedT cells treated with or without TGF� were expanded in vitro, and theirsuppressive activity was assayed at different days postactivation. Representa-tive CFSE data are shown using a 1-to-4 suppressor-to-target ratio. (D) Expres-sion of FOXP3 on overexpressing T cells. Purified CD4� T cells were activatedand transduced with FOXP3 lentiviruses that also express RFP as a marker.After 12–14 days postculture, cells were sorted based on RFP expression,restimulated through TCR for 2 days, and expression of FOXP3 and GARPdetermined on FOXP3-overexpressing cells (FOXP3�), vector control-transduced cells (Control), or Tregs. GARP expression was determined forFOXP3� and Tregs after gating on FOXP3 expression. (E) Expression of GARPon FOXP3-overexpressing T cells analyzed from multiple donors.
Wang et al. PNAS � August 11, 2009 � vol. 106 � no. 32 � 13441
IMM
UN
OLO
GY
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020
activity but can also discriminate CD25� T cells that containhigh levels of IL-17–secreting cells.
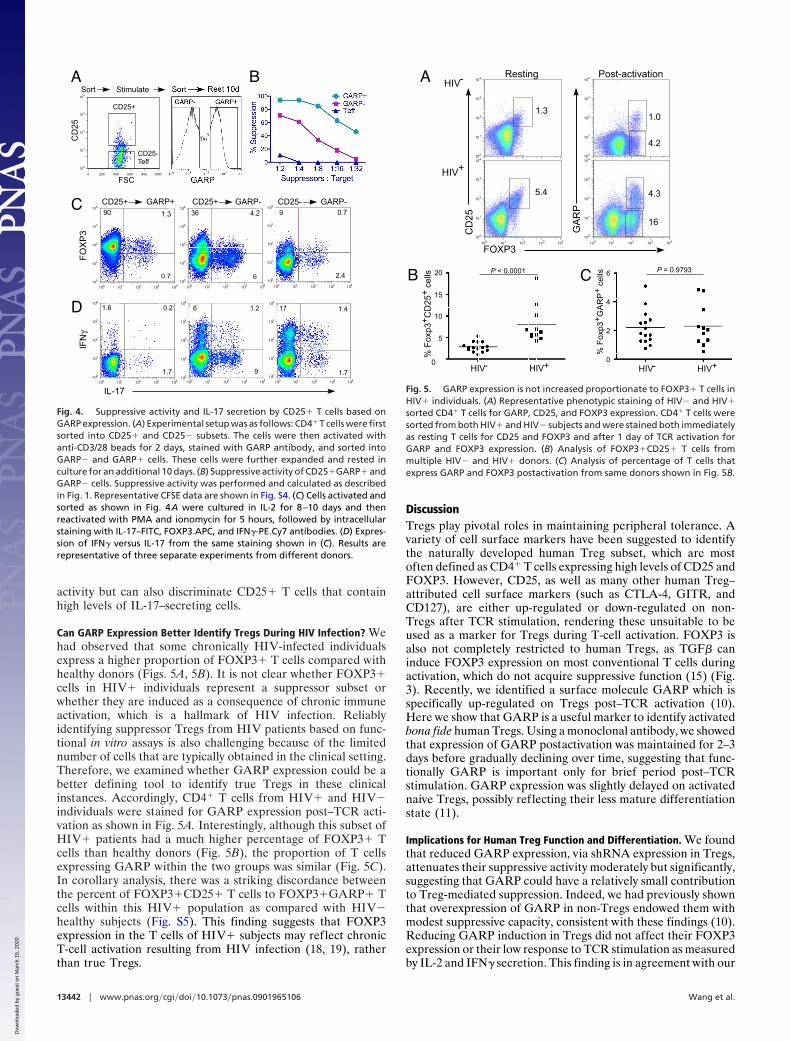
Can GARP Expression Better Identify Tregs During HIV Infection? Wehad observed that some chronically HIV-infected individualsexpress a higher proportion of FOXP3� T cells compared withhealthy donors (Figs. 5A, 5B). It is not clear whether FOXP3�cells in HIV� individuals represent a suppressor subset orwhether they are induced as a consequence of chronic immuneactivation, which is a hallmark of HIV infection. Reliablyidentifying suppressor Tregs from HIV patients based on func-tional in vitro assays is also challenging because of the limitednumber of cells that are typically obtained in the clinical setting.Therefore, we examined whether GARP expression could be abetter defining tool to identify true Tregs in these clinicalinstances. Accordingly, CD4� T cells from HIV� and HIV�individuals were stained for GARP expression post–TCR acti-vation as shown in Fig. 5A. Interestingly, although this subset ofHIV� patients had a much higher percentage of FOXP3� Tcells than healthy donors (Fig. 5B), the proportion of T cellsexpressing GARP within the two groups was similar (Fig. 5C).In corollary analysis, there was a striking discordance betweenthe percent of FOXP3�CD25� T cells to FOXP3�GARP� Tcells within this HIV� population as compared with HIV�healthy subjects (Fig. S5). This finding suggests that FOXP3expression in the T cells of HIV� subjects may reflect chronicT-cell activation resulting from HIV infection (18, 19), ratherthan true Tregs.
DiscussionTregs play pivotal roles in maintaining peripheral tolerance. Avariety of cell surface markers have been suggested to identifythe naturally developed human Treg subset, which are mostoften defined as CD4� T cells expressing high levels of CD25 andFOXP3. However, CD25, as well as many other human Treg–attributed cell surface markers (such as CTLA-4, GITR, andCD127), are either up-regulated or down-regulated on non-Tregs after TCR stimulation, rendering these unsuitable to beused as a marker for Tregs during T-cell activation. FOXP3 isalso not completely restricted to human Tregs, as TGF� caninduce FOXP3 expression on most conventional T cells duringactivation, which do not acquire suppressive function (15) (Fig.3). Recently, we identified a surface molecule GARP which isspecifically up-regulated on Tregs post–TCR activation (10).Here we show that GARP is a useful marker to identify activatedbona fide human Tregs. Using a monoclonal antibody, we showedthat expression of GARP postactivation was maintained for 2–3days before gradually declining over time, suggesting that func-tionally GARP is important only for brief period post–TCRstimulation. GARP expression was slightly delayed on activatednaive Tregs, possibly reflecting their less mature differentiationstate (11).
Implications for Human Treg Function and Differentiation. We foundthat reduced GARP expression, via shRNA expression in Tregs,attenuates their suppressive activity moderately but significantly,suggesting that GARP could have a relatively small contributionto Treg-mediated suppression. Indeed, we had previously shownthat overexpression of GARP in non-Tregs endowed them withmodest suppressive capacity, consistent with these findings (10).Reducing GARP induction in Tregs did not affect their FOXP3expression or their low response to TCR stimulation as measuredby IL-2 and IFN� secretion. This finding is in agreement with our
CD25
Stimulate Rest 10dSort SortA B
C
10
100
101
102
103
104
FSC
CD25+
CD25-Teff
0 200 400 600 800 1000
100
101
102
103
104
17 1.4
1.7100
101
102
103
104
1.6 0.2
1.7100
101
102
103
104
6 1.2
9
IL-17
IFN
γ
100
101
102
103
104
1.3
0.7
4.2
6
100 101 102 103 104
9 0.7
2.4
CD25-CD25+
FOXP3
90 36
GARP+ CD25+ GARP- GARP-
100
101
102
103
104
100
101
102
103
104
D
100 101 102 103 104100 101 102 103 104
100 101 102 103 104 100 101 102 103 104 100 101 102 103 104
Fig. 4. Suppressive activity and IL-17 secretion by CD25� T cells based onGARP expression. (A) Experimental setup was as follows: CD4� T cells were firstsorted into CD25� and CD25� subsets. The cells were then activated withanti-CD3/28 beads for 2 days, stained with GARP antibody, and sorted intoGARP� and GARP� cells. These cells were further expanded and rested inculture for an additional 10 days. (B) Suppressive activity of CD25�GARP� andGARP� cells. Suppressive activity was performed and calculated as describedin Fig. 1. Representative CFSE data are shown in Fig. S4. (C) Cells activated andsorted as shown in Fig. 4A were cultured in IL-2 for 8–10 days and thenreactivated with PMA and ionomycin for 5 hours, followed by intracellularstaining with IL-17–FITC, FOXP3.APC, and IFN�-PE.Cy7 antibodies. (D) Expres-sion of IFN� versus IL-17 from the same staining shown in (C). Results arerepresentative of three separate experiments from different donors.
P = 0.9793
HIV- HIV+
6
4
2
0
%Foxp3+GARP+c el lsP < 0.0001
HIV- HIV+
20
15
10
5
0
%Foxp3+CD25+cells
100
101
102
103
104
100
101
102
103
104
100 101 102 103 104100
101
102
103
104
100 101 102 103 104100
101
102
103
104
FOXP3
CD25
Resting Post-activation
GARP
5.4 4.3
16
1.0
4.2
1.3
HIV+
HIV-A
B C
Fig. 5. GARP expression is not increased proportionate to FOXP3� T cells inHIV� individuals. (A) Representative phenotypic staining of HIV� and HIV�sorted CD4� T cells for GARP, CD25, and FOXP3 expression. CD4� T cells weresorted from both HIV� and HIV� subjects and were stained both immediatelyas resting T cells for CD25 and FOXP3 and after 1 day of TCR activation forGARP and FOXP3 expression. (B) Analysis of FOXP3�CD25� T cells frommultiple HIV� and HIV� donors. (C) Analysis of percentage of T cells thatexpress GARP and FOXP3 postactivation from same donors shown in Fig. 5B.
13442 � www.pnas.org�cgi�doi�10.1073�pnas.0901965106 Wang et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

prior results that silencing FOXP3 in Tregs, which reducesGARP expression, also does not significantly affect their re-sponse to TCR signals (11). How GARP could be contributingto suppressive function of Tregs, and why these cells selectivelyexpress GARP upon activation remains unclear. An importantadvance in understanding GARP function was provided by Tranet al. (25), who discovered that GARP is a receptor for latency-associated peptide (LAP), which requires TGF� for binding toGARP. It is conceivable that expression of the LAP-TGF�complex on the cell surface of Tregs or GARP-overexpressingcells deliver a suppressive signal to target T cells that expressTGF� receptors. The association of LAP-TGF� with GARPwould also provide a mechanistic explanation for our previousfinding that ectopic expression of GARP induces FOXP3 inconventional T cells (10). Thus, we postulate that LAP-TGF� bound to GARP could induce expression of FOXP3through the signaling of TGF� receptors, as has been previouslysuggested (20).
We have shown that TCR stimulation is required for rapidinduction of GARP on resting human Tregs. Silencing FOXP3in Tregs reduced the level of GARP expression postactivation.Interestingly, GARP was only minimally up-regulated on cellsectopically expressing FOXP3. This finding, together with theresult that TGF�-induced FOXP3 is not sufficient for GARPinduction, suggests that there are other unknown factors re-quired for specific GARP expression. Because TGF� was shownrecently to contribute to Th17 differentiation in conjugation withIL-6 or IL-1� (21, 22), perhaps other cytokines or soluble factorscould also program GARP expression in conjunction withTGF�. It is also possible that the level and duration of FOXP3expression is important in determining whether a cell can expressGARP upon activation (23). However, we show that even whenectopic expression of FOXP3 is at higher levels than naturalTregs, GARP is not induced (Fig. 3D), indicating that the levelof FOXP3 is not a limiting factor for GARP induction. Alter-natively, FOXP3 expression during development of Tregs resultsin epigenetic modifications that may allow induction of GARPfrom these cells. It will also be of interest in future to determinewhether GARP expression in murine T cells is regulated simi-larly to that in humans, as TGF� signals are sufficient to conferTreg function in naive murine T cells (6, 7).
Implications for Human Th17 Cell Differentiation. Recently it hasbeen shown that CD25� Treg-like cells also secrete IL-17 (16,17). These could be either common Tregs that have switchedsides to Th17 lineage or cells with dual functionality. We foundthat CD25� T cells that are induced to express high GARP levelscontain few IL-17� cells, whereas the GARP-negative portionis greatly enriched in IL-17-secreting cells, regardless of FOXP3expression (Fig. 4C). Our findings are consistent with recentreports that a sizeable portion of human CD25�FOXP3� cellscontain IL-17–secreting cells (16, 17). Similar to these reports,we found that although CD25�GARP� cells that contain highIL-17 secretor also display suppressive function, this was signif-icantly lower compared with the suppressive capacity ofCD25�GARP� cells. Because it is difficult to separate IL-17�–secreting cells within this subset, it is not yet clear whetherCD25�GARP�IL-17� cells have suppressive capacity, as hasrecently been suggested (16). It also remains to be determinedwhether these CD25� T cells with dual Treg/Th17 cell charac-teristic are derived from naive T cells, or alternatively, whetherthey differentiate from Tregs that switch sides to Th17 lineagewhile retaining some Treg functionality. Voo et al. providedevidence for the latter model (17), which we also favor. In thisregard, a small percentage of IL-17� cells present in theGARP� cells could be transitional Th17-lineage committed Tcells, which eventually down-regulate FOXP3 expression andhave lost the capacity to up-regulate GARP expression. Indeed,
CD25�GARP� cells expressed relatively lower FOXP3 ascompared with CD25�GARP� cells after expansion in culture.Thus, expression of GARP could be an invaluable tool to furtherdissect the original duality or cross-differentiation pathways ofTreg and Th17 lineage T cells.
Implications for HIV Pathogenesis. Our results led us to explore thepossibility of using GARP as a marker to isolate Tregs from theactivated T-cell population. We showed that all GARP� T cellspossess Treg characteristics including FOXP3 expression, hypo-responsiveness, and suppression of T-cell activation. These find-ings predict that GARP expression would reliably identifysuppressive Tregs in ex vivo analysis of human T-cell subsets. Asa proof-of-principle experiment, we show that there is a verylarge discordance between GARP� and FOXP3� T cells insome HIV� individuals. This group of HIV� subjects had muchgreater levels of FOXP3� cells compared with GARP� cells.Based on our findings that GARP is preferentially expressed onhighly suppressor bona fide Tregs, we suggest that FOXP3expression in these patients is mostly caused by chronic immuneactivation, which can potentially result in FOXP3 inductionthrough TGF� (8). Thus, future studies using GARP as a markerto identify Tregs during HIV infection could resolve some of thecontradictory findings on the level of Tregs at various stages ofHIV disease (24). It will also be interesting to determinewhether, at mucosal tissues or sites of inflammation, where thereis continuous T-cell activation, GARP-expressing T cells can bedirectly identified, which would suggest ongoing activation ofTregs in these locations.
In summary, we demonstrated that surface expression ofGARP identifies both activated FOXP3� human T cells withhigh suppressive activity and excludes IL-17–secreting Treg-likecells. Thus, expression of GARP can be of great use in expandingbona fide human Tregs in vitro, both to dissect their biology andfor clinical in vivo use, for example to establish transplantationtolerance. Our findings also implicate additional factors otherthan FOXP3 or an epigenetic program for development orlineage commitment of true Tregs that express GARP. Expres-sion of GARP will also be useful in defining whether T cells withTreg phenotype that secrete IL-17 are derived from bona fideTregs that have switched to the Th17 side or whether these cellsrepresent a separate effector lineage. In addition, expression ofGARP can be a highly relevant biomarker to better identifyTregs in human diseases with high immune activation, such asHIV infection, or at sites of inflammation.
Materials and MethodsCell Purification and Culture. Human peripheral blood mononuclear cells(PBMCs) were isolated with Ficoll-Hypaque (Amersham Pharmacia), fromperipheral blood from normal donors and CD4� T cells were then separatedusing magnetic cell sorting (Dynabeads; Invitrogen). Purified CD4� T cells werethen stained with anti-CD25 and CD45RO antibodies (BD Biosciences), and thedifferent T-cell subsets, including naive and Tregs, were sorted using FACS Ariaflow cytometer (BD Biosciences) (10). Cells were cultured in RPMI (Life Tech-nologies), media containing FCS (Atlanta Biologicals), as described before (3).Monocyte-derived dendritic cells (DCs) were generated from CD14� cells fromnormal donors as previously described (3). The superantigen, staphylococcalenterotoxin B (SEB), was purchased from Sigma.
FACS Analysis. Cells were stained with relevant antibodies on ice for 30minutes in PBS buffer containing 2% FCS and 0.1% sodium azide. Cells werewashed twice before being analyzed by BD LSRII flow cytometer. Live cellswere gated based on forward and side scatter profiles or based on exclusionwith propidium iodide (PI) staining, and analysis was performed using FlowJosoftware (Tree Star). To stain for cytokines intracellularly, cells were activatedby PMA and ionomycin (Sigma) for 5 hours, followed by fixation and perme-abilization using a commercial kit (BD Biosciences), according to the manu-facturer’s instructions. The following antibodies were used for staining: IFN�
and IL-17 (eBioscience), CD45RO, CD25 (both from BD Biosciences). GARPstaining was performed 1–2 days post–TCR activation by first staining with
Wang et al. PNAS � August 11, 2009 � vol. 106 � no. 32 � 13443
IMM
UN
OLO
GY
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020

mouse anti-GARP (Alexis Biochemicals), followed by anti-mIgG2b bio (BDBiosciences), and finally with strep-APC (eBioscience). FOXP3 expression wasperformed via intracellular staining. Briefly, cells were washed and resus-pended in 1X Fixation/Permeabilization buffer (eBioscience) and incubated at4 °C for 30–60 minutes. Cells were then washed twice with 1X Permeabiliza-tion buffer and stained with anti-FOXP3 antibody (clone 259D, Biolegend), for30 minutes at 4 °C. Cells were washed twice with 1X Permeabilization bufferand analyzed by LSR II.
Cell Proliferation and Cytokine Assays. Cells were labeled with carboxyfluo-rescein succinimidyl ester (CFSE) (Molecular Probes), and activated by DC andeither anti-CD3 antibody (ATCC clone OKT3) or SEB (Sigma) at differentconcentrations as previously described (3). For GARP induction or cytokinesecretion, cells were activated with CD3/CD28 beads (Invitrogen). Superna-tants from activated T-cell cultures were collected 16 hours later; cytokineproduction was measured using cytometric bead array (CBA; BD Biosciences)and analyzed by FlowJo (Tree Star). Proliferation of the cells was assessed onday 4 or day 5 post–TCR activation by monitoring dilution of CFSE in the cells.Statistical analysis was performed with a paired t test for comparison betweentwo groups and one-way analysis of variance (ANOVA) for comparison withinthree or more data groups using Prism (GraphPad Software).
Suppression Assay. Resting CD4� T cells were sorted as described above,labeled with CFSE and used as targets as described (3). The CFS-labeled restingT cells and non-labeled ‘‘suppressor or effector’’ cells were mixed together atdifferent suppressor-to-target ratios. The cells were stimulated by solubleanti-CD3 antibody (OKT-3, 10–100 ng/ml) in the presence of DCs (1:10) oralternatively DCs pulsed with suboptimal concentrations of SEB (range, 0.1–0.001 ng/ml). Cells were harvested and analyzed by FACS (LSR-II, BD Bio-sciences) on day 4 or day 5 postactivation. The percentage of suppression was
calculated as previously described (10); data represent mean � SEM of dupli-cates or similar experiments from at least three different donors. Statisticalanalysis was performed with a paired t test for comparison between twogroups and one-way ANOVA for comparison within three or more datagroups.
Study Subjects and Statistical Analysis. Buffy coats from healthy adult subjects(n � 15) who were HIV negative and with no documentation of chronic viralinfections such as hepatitis B or C were obtained from NY Blood bank. Bloodsamples from adults with HIV infection (n � 11) were obtained during routinepatient care visits in Virology Clinic at Bellevue Hospital (New York, NY). Noselection criteria based on race or sex was included. All subjects providedwritten consent and the study was approved by the NYU Institutional ReviewBoard. Variables within healthy and HIV� donors were compared using apaired t test. Variables between healthy and infected individuals were com-pared by a Mann–Whitney test. Categorical variables were determined by a �2
test. All significance levels were based on two-tailed tests. Statistical analyseswere performed using Prism (GraphPad).
Quantitative PCR. Total RNA from cells was isolated and cDNA was synthesizedas previously described (3). Quantitative PCR was performed with the synthe-sized cDNA using Applied Biosystems 7300 apparatus. TaqMan primer andprobe mixes were purchased from Applied Biosystems; their IDs are as follows:�-actin (used as a reference gene): Hs99999903�ml; GARP: Hs00194136�ml;FOXP3 primer mix ID: Hs00203958�m1.
ACKNOWLEDGMENTS. We thank Dat Tran, Ethan Shevach, Aimee ElHed, andQi Wan for critical reading and suggestions for the manuscript. This study wassupported by National Institutes of Health (NIH) Grant R01 AI065303 (to D.U.),Centers for AIDS Research NIH Grant P30AI027742 (to A.K. and D.U.), R01AI059315 (to H.F.), and NIH Training Grant 2T32 AI007180–26A2 (to F.M.).
1. Sakaguchi S, Yamaguchi T, Nomura T, Ono M (2008) Regulatory T cells and immunetolerance. Cell 133:775–787.
2. Vignali DA, Collison LW, Workman CJ (2008) How regulatory T cells work. Nat RevImmunol 8:523–532.
3. Oswald-Richter K, et al. (2004) HIV infection of naturally occurring and geneticallyreprogrammed human regulatory T-cells. PLoS Biol 4:E198.
4. Yagi H, et al. (2004) Crucial role of FOXP3 in the development and function of humanCD25�CD4� regulatory T cells. Int Immunol 16:1643–1656.
5. Allan SE, et al. (2008) Generation of potent and stable human CD4� T regulatory cellsby activation-independent expression of FOXP3. Mol Ther 16:194–202.
6. Weber SE, et al. (2006) Adaptive islet-specific regulatory CD4 T cells control autoim-mune diabetes and mediate the disappearance of pathogenic Th1 cells in vivo.J Immunol 176:4730–4739.
7. Fantini MC, et al. (2006) Transforming growth factor beta induced FoxP3� regulatoryT cells suppress Th1 mediated experimental colitis. Gut 55:671–680.
8. Tran DQ, Ramsey H, Shevach EM (2007) Induction of FOXP3 expression in naivehuman CD4�FOXP3 T cells by T-cell receptor stimulation is transforming growthfactor-beta dependent but does not confer a regulatory phenotype. Blood110:2983–2990.
9. Yamagiwa S, Gray JD, Hashimoto S, Horwitz DA (2001) A role for TGF-beta in thegeneration and expansion of CD4�CD25� regulatory T cells from human peripheralblood. J Immunol 166:7282–7289.
10. Wang R, Wan Q, Kozhaya L, Fujii H, Unutmaz D (2008) Identification of a regulatory Tcell specific cell surface molecule that mediates suppressive signals and induces Foxp3expression. PLoS ONE 3:e2705.
11. Antons AK, et al. (2008) Naive precursors of human regulatory T cells require FoxP3 forsuppression and are susceptible to HIV infection. J Immunol 180:764–773.
12. Valmori D, Merlo A, Souleimanian NE, Hesdorffer CS, Ayyoub M (2005) A peripheralcirculating compartment of natural naive CD4 Tregs. J Clin Invest 115:1953–1962.
13. Seddiki N, et al. (2006) Persistence of naive CD45RA� regulatory T cells in adult life.Blood 107:2830–2838.
14. Sereti I, et al. (2005) In vivo expansion of CD4CD45RO-CD25 T cells expressing foxP3 inIL-2-treated HIV-infected patients. J Clin Invest 115:1839–1847.
15. Shevach EM, Tran DQ, Davidson TS, Andersson J (2008) The critical contribution ofTGF-beta to the induction of Foxp3 expression and regulatory T cell function. EurJ Immunol 38:915–917.
16. Beriou G, et al. (2009) IL-17-producing human peripheral regulatory T cells retainsuppressive function. Blood 113:4240–4249.
17. Voo KS, et al. (2009) Identification of IL-17-producing FOXP3� regulatory T cells inhumans. Proc Natl Acad Sci USA 106:4793–4798.
18. Appay V, Sauce D (2008) Immune activation and inflammation in HIV-1 infection:Causes and consequences. J Pathol 214:231–241.
19. Grossman Z, Meier-Schellersheim M, Paul WE, Picker LJ (2006) Pathogenesis of HIV infec-tion: What the virus spares is as important as what it destroys. Nat Med 12:289–295.
20. Andersson J, et al. (2008) CD4� FoxP3� regulatory T cells confer infectious tolerancein a TGF-beta-dependent manner. J Exp Med 205:1975–1981.
21. Awasthi A, Murugaiyan G, Kuchroo VK (2008) Interplay between effector th17 andregulatory T cells. J Clin Immunol 28:660–670.
22. Bettelli E, Korn T, Oukka M, Kuchroo VK (2008) Induction and effector functions ofT(H)17 cells. Nature 453:1051–1057.
23. Allan SE, Song-Zhao GX, Abraham T, McMurchy AN, Levings MK (2008) Induciblereprogramming of human T cells into Treg cells by a conditionally active form ofFOXP3. Eur J Immunol 38:3282–3289.
24. Holmes D, Jiang Q, Zhang L, Su L (2008) Foxp3 and Treg cells in HIV-1 infection andimmuno-pathogenesis. Immuol Res 41:248–266.
25. Tran, et al. (2009) GARP (LRRC32) is essential for the surface expression of latent TGF�
on platelets and activated FOXP3� regulating cells. Proc Natl Acad Sci USA 10.1073/PNAS.0901944106.
13444 � www.pnas.org�cgi�doi�10.1073�pnas.0901965106 Wang et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
5, 2
020


















