
Expression Mycobacterium Immunological Analysis …iai.asm.org/content/61/1/260.full.pdf262...
8
Vol. 61, No. 1 INFECTION AND IMMUNITY, Jan. 1993, p. 260-267 0019-9567/93/010260-08$02.00/0 Copyright X 1993, American Society for Microbiology Expression of the Mycobacterium tuberculosis 19-Kilodalton Antigen in Mycobacterium smegmatis: Immunological Analysis and Evidence of Glycosylation THOMAS GARBE,t* DAVID HARRIS, MARTIN VORDERMEIER, RAJU LATHIGRA,4 JURAJ IVANYI, AND DOUGLAS YOUNG Medical Research Council Tuberculosis and Related Infections Unit, Royal Postgraduate Medical School, Hammersmith Hospital, Ducane Road, London W12 OHS, United Kingdom Received 8 July 1992/Accepted 21 October 1992 The gene encoding a 19-kDa antigen from Mycobacterium tuberculosis was expressed as a recombinant protein in the rapid-growing species Mycobacterium smegmatis. The recombinant antigen was expressed at a level approximately ninefold higher than in M. tuberculosis and, like the native antigen, was found in the pellet fraction after high-speed centrifugation of bacterial extracts. The 19-kDa antigen in crude bacterial extracts, and the purified recombinant antigen, bound strongly to concanavalin A, indicating the possibility of posttranslational glycosylation. The recombinant antigen stimulated T-cell proliferation in vitro when added to assays either in the form of whole recombinant bacteria or as a purified protein. Homologous expression of mycobacterial antigens in a rapid-growing mycobacterial host may be particularly useful for the immunological characterization of proteins which are subject to posttranslational modification. An extensive panel of mycobacterial proteins involved in recognition by the host immune system has been identified by biochemical fractionation or by screening of recombinant DNA expression libraries (reviewed in reference 32). Sev- eral of these antigens have been proposed as potential targets for improved diagnostic tests or for incorporation into novel subunit vaccines, but difficulties in obtaining sufficient quantities of the purified reagents have impeded comparative experimental testing of such suggestions (13, 30, 32). In addition to the requirement for strict containment facilities, pathogenic mycobacteria grow very slowly (Myco- bacterium tuberculosis doubling time, 24 h) or not at all (Mycobacterium leprae) in laboratory culture, presenting significant practical obstacles to large-scale growth for bio- chemical fractionation. For some antigens, members of conserved heat shock protein families, for example, these problems have been overcome by high-level expression of the relevant genes in standard Escherichia coli recombinant DNA systems (18, 27). Several other antigens have been expressed as fusion proteins in E. coli but have proved difficult to overexpress as free proteins (9, 17, 29). This latter class includes proteins containing signal sequences or other features indicative of a requirement for posttranslational modification (6, 8, 17, 30, 31). It is attractive to speculate that a mycobacterial host may provide the optimal system for expression of such antigens. The recent development of techniques and vectors for transformation of mycobacteria (14, 21, 23, 24) has been stimulated by the goal of creating a new generation of recombinant Mycobacterium bovis BCG vaccines (1, 14, 25). * Corresponding author. t Present address: Department of Molecular Genetics, Biochem- istry and Microbiology, University of Cincinnati College of Medi- cine, 3110 Medical Sciences Building, 231 Bethesda Avenue, Cin- cinnati, OH 45267-0524. : Present address: MedImmune Inc, Gaithersburg, MD 20878. In addition to the utility of such systems for expressing a range of heterologous antigens from other unrelated patho- gens, the same approach is also convenient for transferring genes between mycobacterial species. It has been demon- strated, for example, that genes encoding the superoxide dismutase enzymes of M. leprae and M. tuberculosis are expressed from their own promoters in Mycobacterium smegmatis (a rapid-growing species suitable for laboratory manipulation), while provision of an exogenous promoter was essential for expression of the same genes in E. coli (26, 33). In addition, it was found that the mycobacterial system allowed expression of functionally active superoxide dismu- tase, in contrast to the enzymatically inactive recombinant product in E. coli (33). In the present study, we have examined the use of a mycobacterial expression system to facilitate the immunological characterization of a 19-kDa antigen from M. tuberculosis. The 19-kDa antigen was originally identified by using a set of murine monoclonal antibodies binding to M. tuberculosis and a limited number of nontuberculous mycobacteria (5) and was subsequently shown to elicit both humoral and cell-mediated immune responses in mice and patients with tuberculosis (7, 9, 13). Results from nucleotide sequence analysis (2) and biochem- ical characterization of the 19-kDa antigen (31) suggest that the mature protein is secreted across the cell membrane and is present as a lipoprotein in M. tuberculosis. Similarly, posttranslational acylation of the corresponding protein from Mycobacterium avium-intracellulare has been suggested (20), and analysis of the 19-kDa antigen purified from M. bovis has provided evidence of glycosylation (8). The 19-kDa antigen has no marked sequence homology with other known proteins, and its biochemical function has not yet been established. We report here on the application of an M. smegmatis expression system to study posttranslational modification and T-cell recognition of the 19-kDa antigen from M. tuber- culosis. 260 on July 3, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Expression Mycobacterium Immunological Analysis …iai.asm.org/content/61/1/260.full.pdf262...

Vol. 61, No. 1INFECTION AND IMMUNITY, Jan. 1993, p. 260-2670019-9567/93/010260-08$02.00/0Copyright X 1993, American Society for Microbiology
Expression of the Mycobacterium tuberculosis 19-KilodaltonAntigen in Mycobacterium smegmatis: Immunological
Analysis and Evidence of GlycosylationTHOMAS GARBE,t* DAVID HARRIS, MARTIN VORDERMEIER, RAJU LATHIGRA,4
JURAJ IVANYI, AND DOUGLAS YOUNGMedical Research Council Tuberculosis and Related Infections Unit, Royal Postgraduate
Medical School, Hammersmith Hospital, Ducane Road,London W12 OHS, United Kingdom
Received 8 July 1992/Accepted 21 October 1992
The gene encoding a 19-kDa antigen from Mycobacterium tuberculosis was expressed as a recombinantprotein in the rapid-growing species Mycobacterium smegmatis. The recombinant antigen was expressed at alevel approximately ninefold higher than in M. tuberculosis and, like the native antigen, was found in the pelletfraction after high-speed centrifugation of bacterial extracts. The 19-kDa antigen in crude bacterial extracts,and the purified recombinant antigen, bound strongly to concanavalin A, indicating the possibility ofposttranslational glycosylation. The recombinant antigen stimulated T-cell proliferation in vitro when added toassays either in the form of whole recombinant bacteria or as a purified protein. Homologous expression ofmycobacterial antigens in a rapid-growing mycobacterial host may be particularly useful for the immunologicalcharacterization of proteins which are subject to posttranslational modification.
An extensive panel of mycobacterial proteins involved inrecognition by the host immune system has been identifiedby biochemical fractionation or by screening of recombinantDNA expression libraries (reviewed in reference 32). Sev-eral of these antigens have been proposed as potentialtargets for improved diagnostic tests or for incorporationinto novel subunit vaccines, but difficulties in obtainingsufficient quantities of the purified reagents have impededcomparative experimental testing of such suggestions (13,30, 32). In addition to the requirement for strict containmentfacilities, pathogenic mycobacteria grow very slowly (Myco-bacterium tuberculosis doubling time, 24 h) or not at all(Mycobacterium leprae) in laboratory culture, presentingsignificant practical obstacles to large-scale growth for bio-chemical fractionation. For some antigens, members ofconserved heat shock protein families, for example, theseproblems have been overcome by high-level expression ofthe relevant genes in standard Escherichia coli recombinantDNA systems (18, 27). Several other antigens have beenexpressed as fusion proteins in E. coli but have proveddifficult to overexpress as free proteins (9, 17, 29). This latterclass includes proteins containing signal sequences or otherfeatures indicative of a requirement for posttranslationalmodification (6, 8, 17, 30, 31). It is attractive to speculatethat a mycobacterial host may provide the optimal systemfor expression of such antigens.The recent development of techniques and vectors for
transformation of mycobacteria (14, 21, 23, 24) has beenstimulated by the goal of creating a new generation ofrecombinant Mycobacterium bovis BCG vaccines (1, 14, 25).
* Corresponding author.t Present address: Department of Molecular Genetics, Biochem-
istry and Microbiology, University of Cincinnati College of Medi-cine, 3110 Medical Sciences Building, 231 Bethesda Avenue, Cin-cinnati, OH 45267-0524.
: Present address: MedImmune Inc, Gaithersburg, MD 20878.
In addition to the utility of such systems for expressing arange of heterologous antigens from other unrelated patho-gens, the same approach is also convenient for transferringgenes between mycobacterial species. It has been demon-strated, for example, that genes encoding the superoxidedismutase enzymes of M. leprae and M. tuberculosis areexpressed from their own promoters in Mycobacteriumsmegmatis (a rapid-growing species suitable for laboratorymanipulation), while provision of an exogenous promoterwas essential for expression of the same genes in E. coli (26,33). In addition, it was found that the mycobacterial systemallowed expression of functionally active superoxide dismu-tase, in contrast to the enzymatically inactive recombinantproduct in E. coli (33). In the present study, we haveexamined the use of a mycobacterial expression system tofacilitate the immunological characterization of a 19-kDaantigen from M. tuberculosis. The 19-kDa antigen wasoriginally identified by using a set of murine monoclonalantibodies binding to M. tuberculosis and a limited numberof nontuberculous mycobacteria (5) and was subsequentlyshown to elicit both humoral and cell-mediated immuneresponses in mice and patients with tuberculosis (7, 9, 13).Results from nucleotide sequence analysis (2) and biochem-ical characterization of the 19-kDa antigen (31) suggest thatthe mature protein is secreted across the cell membrane andis present as a lipoprotein in M. tuberculosis. Similarly,posttranslational acylation of the corresponding protein fromMycobacterium avium-intracellulare has been suggested(20), and analysis of the 19-kDa antigen purified from M.bovis has provided evidence of glycosylation (8). The 19-kDaantigen has no marked sequence homology with otherknown proteins, and its biochemical function has not yetbeen established.We report here on the application of an M. smegmatis
expression system to study posttranslational modificationand T-cell recognition of the 19-kDa antigen from M. tuber-culosis.
260
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

RECOMBINANT ANTIGENS IN M. SMEGMATIS 261
MATERIALS AND METHODS
Bacterial strains and plasmids. M. smegmatis 1-2c is aderivative of M. smegmatis mc26 (14), which shows highefficiency of transformation (33). M. smegmatis was grownin Middlebrook 7H9 medium (Difco Laboratories; Detroit,Mich.) supplemented with glucose (2%, wt/vol). Kanamycinsulfate was added at 50 ,ug/ml for culture of strains trans-formed with shuttle plasmids. M. tuberculosis H37Rv is avirulent strain originally isolated from a tuberculosis patientand was supplied by B. W. Allen (Royal PostgraduateMedical School, Hammersmith Hospital, London, UnitedKingdom). M. tuberculosis was grown on Middlebrook 7H11agar plates, supplemented with 0.05% Tween 80 (BDH) andOADC (oleic acid, albumin, dextrose, catalase [Difco]).Protein extracts were prepared from M. tuberculosis har-vested from plates and disrupted in distilled water by usingglass beads as described previously (13). The total proteinconcentration was estimated by using a protein assay systemsupplied by Bio-Rad Laboratories (Richmond, Calif.) withbovine serum albumin (BSA) as the standard. E. coli JM105(Pharmacia) and TG1 (22) were grown on Luria-Bertanimedium with 50 Wg of kanamycin sulfate per ml added asdescribed by Sambrook et al. (22). pBAK-7q is a derivativeof pBAK14, a shuttle plasmid capable of replicating inmycobacteria and in E. coli (33), containing a recombinant1.8-kb SmaI fragment which includes the structural geneencoding the M. tuberculosis 19-kDa antigen (2, 9).DNA manipulation. Plasmid DNA was prepared by stan-
dard procedures and analyzed by restriction enzyme diges-tion and agarose gel electrophoresis as described by Sam-brook et al. (22). Transformation of M. smegmatis withshuttle plasmids was carried out by electroporation as de-scribed previously (33). E. coli was transformed by usingstandard methods (22).
Gel electrophoresis and Western blotting (immunoblotting).Electrophoresis in polyacrylamide gels containing sodiumdodecyl sulfate (SDS-PAGE) and blotting onto nitrocellulosemembranes was carried out by using standard procedures(15, 28). Samples used for Western blot analysis contained0.3 to 3 ,ug of total protein. Antigens were stained onnitrocellulose membranes by using monoclonal antibodiesspecific for the 19-kDa antigen, namely, TB23 (3, 5), HYT6(5), and F29-47 (5), by using techniques described previously(33). Quantitative analysis of Western blots was carried outby using a Shimadzu CS-9000 dual-wavelength, flying-spotscanning densitometer at 550 nm. Results are expressed interms of peak area as relative absorbance units.The procedures for staining of nitrocellulose blots with
peroxidase-conjugated concanavalin A (ConA) were essen-tially identical to those used for antibody staining. Nonspe-cific binding was blocked by incubating blots for 1 h with 4%(wt/vol) BSA in phosphate-buffered saline (PBS) with TritonX-100 (0.2%, vol/vol). After repeated washes with PBS andPBS-Triton X-100, the blots were incubated with 20 ml ofConA-peroxidase conjugate (0.2 purpurogallin units per ml;Sigma) in 2% (wt/vol) BSA in PBS-Triton X-100 for 1 h.After further washing, blots were stained for peroxidaseactivity by adding 3,3'-diaminobenzidine HCI and hydrogenperoxide in PBS.
Purification of the recombinant 19-kDa antigen. The recom-binant 19-kDa antigen was purified by a novel procedurebased on exploitation of its relative insolubility in urea andsubunit molecular weight. M. smegmatis 1-2c transformedwith pBAK-7q was grown at 37°C for 4 days in six 2-literconical flasks containing 500 ml of Middlebrook 7H9-glucose
medium with 50 ,ug of kanamycin sulfate per ml in a rotaryincubator at 150 rpm. Bacteria were pelleted for 10 min at10,000 x g, resuspended in 50 ml of water, and sonicated fora total of 10 min (MSE Soniprep 150, 19-mm probe, atmaximum output). Cell debris was removed by centrifuga-tion for 10 min at 16,000 x g, and the supernatant wascentrifuged at 48,000 x g overnight at 20°C. The resultingpellet was resuspended in 10 ml of water, urea was added at480 mg/ml, and the suspension was rolled overnight at 4°C.After centrifugation at 230,000 x g for 3 h at 20°C, a turbidpellet was obtained on top of a much-larger, translucent,reddish-brown pellet. The upper, turbid, pellet was resus-pended in 8 M urea to a final volume of 25 ml and centrifugedfor a further 3 h at 230,000 x g. The turbid grey pellet wasresuspended in a minimum volume of 8 M urea and thendiluted 1:10 in water and centrifuged for 3 h at 13,000 rpm(Sorvall, GSA rotor). The resulting pellet was resuspendedovernight at 4°C in an equal volume of 50 mM Tris HCl (pH8.0) containing 2% (vol/vol) Triton X-100 and 5 mM EDTA.After centrifugation for 3 h at 27,000 x g, the resulting whitepellet was resuspended in 5 ml of 50 mM Tris HCl (pH 8.0)containing 5% SDS and 10% 3-mercaptoethanol and held ona boiling water bath for 5 min. The sample was centrifuged at10,000 x g for 10 min at 4°C, and the supernatant (with 10%sucrose added) was loaded onto a Biogel P-100 gel bed (2.6by 100 cm; Bio-Rad) equilibrated with 50 mM Tris HCl (pH8.0)-1% SDS-1% 3-mercaptoethanol. The 19-kDa antigeneluted between 137 and 153 ml.
Antigens and synthetic peptides for T-cell assays. A heat-killed preparation of M. tuberculosis H37Ra was obtainedfrom Difco. A recombinant protein consisting of the M.tuberculosis 19-kDa antigen fused to glutathione S-trans-ferase (rGST19) was expressed in E. coli and isolated asdescribed in detail elsewhere (9). A synthetic peptide, p19.7,corresponding to residues 61 to 80 of the M. tuberculosis19-kDa antigen (VTGSVVCITAAGNVNIAIGG), was syn-thesized by simultaneous solid-phase multiple-peptide tech-nology as previously described (9).Murine T-celi line. A CD4+ murine T-cell line specific for
the M. tuberculosis 19-kDa antigen was generated as fol-lows. C57BV10 mice were immunized in the hind footpadswith a total of 50 ,ug of rGST19 emulsified in incompleteFreund's adjuvant. Seven days later, the draining popliteallymph node cells were removed, and single-cell suspensionswere prepared in complete tissue culture medium (RPMI-1640 medium supplemented with 5% fetal calf serum[GIBCO, Paisley, Scotland], 5 x 10' M P-mercaptoetha-nol, 2mM L-glutamine, 100 U of penicillin per ml, and 100 ,gof streptomycin sulfate per ml). Primed lymph node cellswere cultured in 24-well plates (Nunc, Roskilde, Denmark)at a concentration of 4 x 106 cells per well in the presence of20 ,ug of rGST19 per ml. After 6 days, viable cells wererecovered by centrifugation over Ficoll gradients and recul-tured at a concentration of 0.5 x 106 cells per well togetherwith 3 x 106 irradiated syngeneic spleen cells as antigen-presenting cells. After 5 days of rest, 0.5 x 106 cells per wellwere restimulated in the presence of irradiated antigen-presenting cells and 20 ,ug of rGST19 per ml. A stable cellline was maintained by the same cycles of rest and restimu-lation for more than 6 months.
T-celi proliferation assays. Proliferation assays with therGST19 T-cell line were performed at the end of a restingcycle. T cells (2 x 104 cells per well) were added in triplicateto 96-well flat-bottom microtiter plates (Nunc) containingantigen diluted to the appropriate concentration and 3 x 101irradiated syngeneic spleen cells per well. Microcultures
VOL. 61, 1993
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
262 GARBE ET AL.
HindllI1'1XhoI-f
4t pBAK-7q10.0 kB BamHI
EcoRV
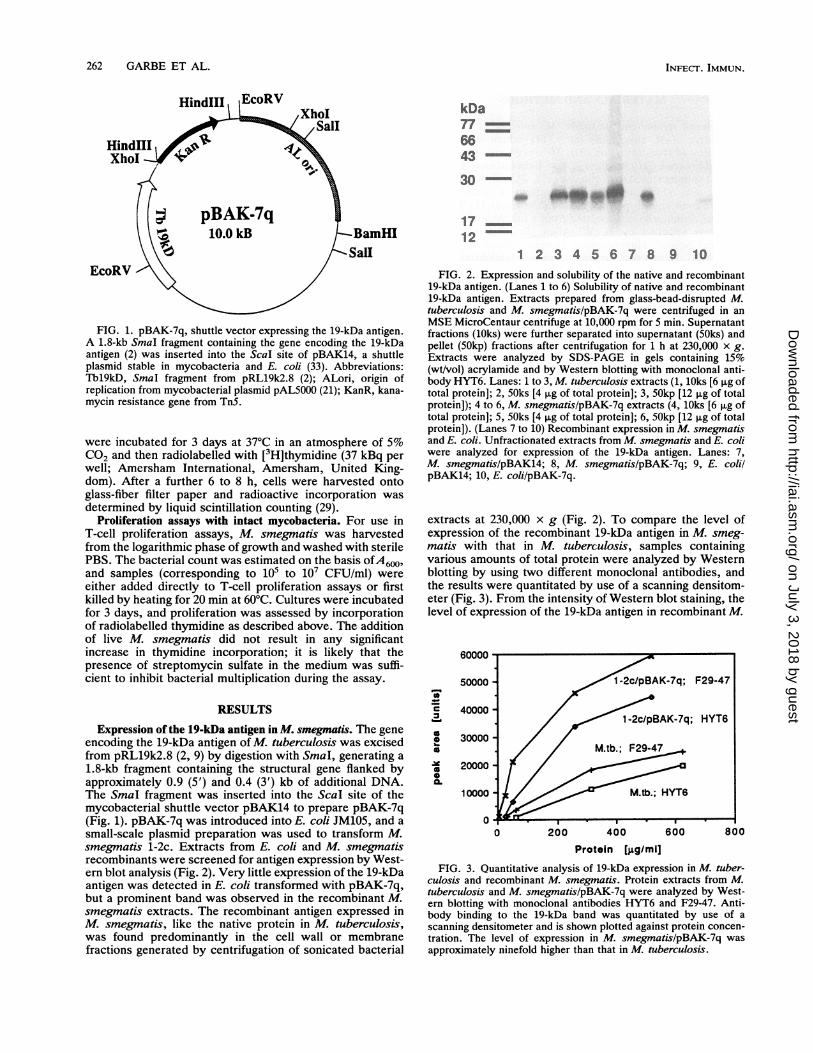
FIG. 1. pBAK-7q, shuttle vector expressing the 19-kDa antigen.A 1.8-kb SmaI fragment containing the gene encoding the 19-kDaantigen (2) was inserted into the Scal site of pBAK14, a shuttleplasmid stable in mycobacteria and E. coli (33). Abbreviations:Thl9kD, SmaI fragment from pRL19k2.8 (2); ALori, origin ofreplication from mycobacterial plasmid pAL5000 (21); KanR, kana-mycin resistance gene from TnS.
were incubated for 3 days at 37°C in an atmosphere of 5%CO2 and then radiolabelled with [3H]thymidine (37 kBq per
well; Amersham International, Amersham, United King-dom). After a further 6 to 8 h, cells were harvested ontoglass-fiber filter paper and radioactive incorporation was
determined by liquid scintillation counting (29).Proliferation assays with intact mycobacteria. For use in
T-cell proliferation assays, M. smegmatis was harvestedfrom the logarithmic phase of growth and washed with sterilePBS. The bacterial count was estimated on the basis ofA600,and samples (corresponding to 105 to 107 CFU/ml) wereeither added directly to T-cell proliferation assays or firstkilled by heating for 20 min at 60°C. Cultures were incubatedfor 3 days, and proliferation was assessed by incorporationof radiolabelled thymidine as described above. The additionof live M. smegmatis did not result in any significantincrease in thymidine incorporation; it is likely that thepresence of streptomycin sulfate in the medium was suffi-cient to inhibit bacterial multiplication during the assay.
RESULTS
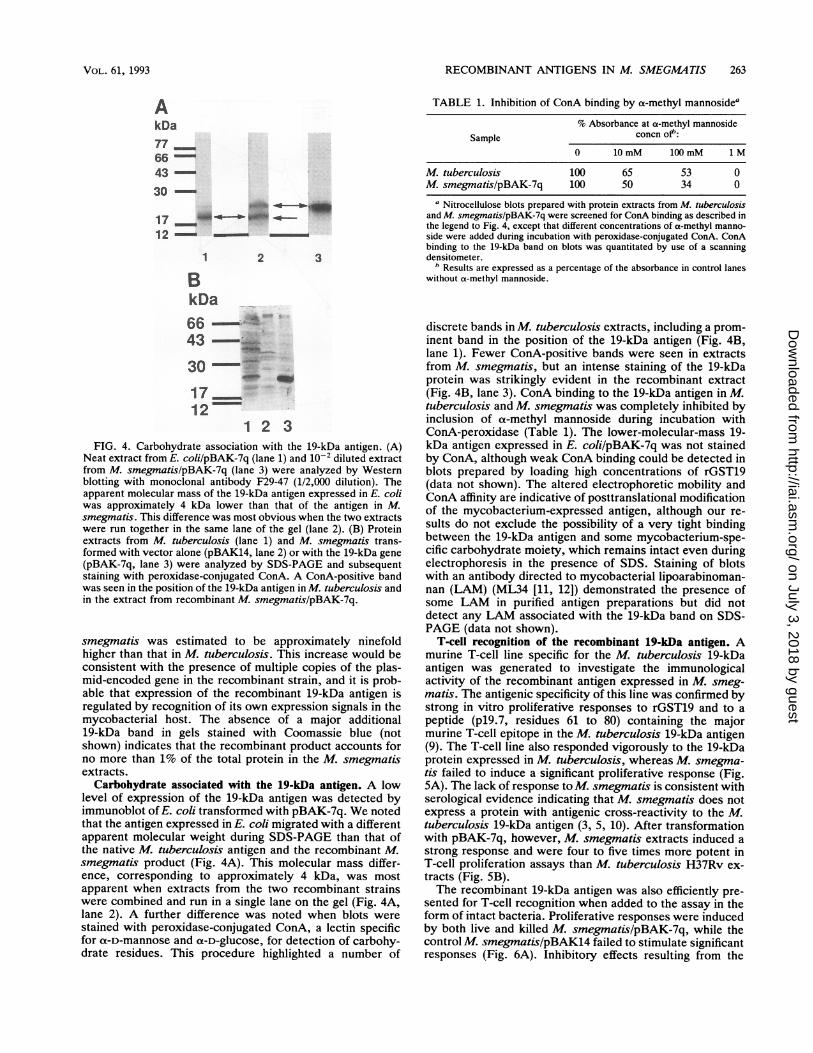
Expression of the 19-kDa antigen in M. smegmatis. The geneencoding the 19-kDa antigen of M. tuberculosis was excisedfrom pRL19k2.8 (2, 9) by digestion with SmaI, generating a1.8-kb fragment containing the structural gene flanked byapproximately 0.9 (5') and 0.4 (3') kb of additional DNA.The SmaI fragment was inserted into the ScaI site of themycobacterial shuttle vector pBAK14 to prepare pBAK-7q(Fig. 1). pBAK-7q was introduced into E. coli JM105, and asmall-scale plasmid preparation was used to transform M.smegmatis 1-2c. Extracts from E. coli and M. smegmatisrecombinants were screened for antigen expression by West-ern blot analysis (Fig. 2). Very little expression of the 19-kDaantigen was detected in E. coli transformed with pBAK-7q,but a prominent band was observed in the recombinant M.smegmatis extracts. The recombinant antigen expressed inM. smegmatis, like the native protein in M. tuberculosis,was found predominantly in the cell wall or membranefractions generated by centrifugation of sonicated bacterial
kDa776643
30 -0*W. otw ~d 0
1712
1 2 3 4 5 6 7 8 9 10FIG. 2. Expression and solubility of the native and recombinant
19-kDa antigen. (Lanes 1 to 6) Solubility of native and recombinant19-kDa antigen. Extracts prepared from glass-bead-disrupted M.tuberculosis and M. smegmatislpBAK-7q were centrifuged in anMSE MicroCentaur centrifuge at 10,000 rpm for 5 min. Supernatantfractions (lOks) were further separated into supernatant (SOks) andpellet (50kp) fractions after centrifugation for 1 h at 230,000 x g.Extracts were analyzed by SDS-PAGE in gels containing 15%(wt/vol) acrylamide and by Western blotting with monoclonal anti-body HYT6. Lanes: 1 to 3, M. tuberculosis extracts (1, lOks [6 pug oftotal protein]; 2, 50ks [4 ,ug of total protein]; 3, 50kp [12 p.g of totalprotein]); 4 to 6, M. smegmatis/pBAK-7q extracts (4, lOks [6 p,g oftotal protein]; 5, 50ks [4 p,g of total protein]; 6, 50kp [12 pg of totalprotein]). (Lanes 7 to 10) Recombinant expression in M. smegmatisand E. coli. Unfractionated extracts from M. smegmatis and E. coliwere analyzed for expression of the 19-kDa antigen. Lanes: 7,M. smegmatis/pBAK14; 8, M. smegmatislpBAK-7q; 9, E. colilpBAK14; 10, E. coli/pBAK-7q.
extracts at 230,000 x g (Fig. 2). To compare the level ofexpression of the recombinant 19-kDa antigen in M. smeg-matis with that in M. tuberculosis, samples containingvarious amounts of total protein were analyzed by Westernblotting by using two different monoclonal antibodies, andthe results were quantitated by use of a scanning densitom-eter (Fig. 3). From the intensity of Western blot staining, thelevel of expression of the 19-kDa antigen in recombinant M.
60000
I-
a
'19
a.
50000
40000
30000
20000
10000
08000 200 400 600
Protein [g.g/miJFIG. 3. Quantitative analysis of 19-kDa expression in M. tuber-
culosis and recombinant M. smegmatis. Protein extracts from M.tuberculosis and M. smegmatis/pBAK-7q were analyzed by West-ern blotting with monoclonal antibodies HYT6 and F29-47. Anti-body binding to the 19-kDa band was quantitated by use of ascanning densitometer and is shown plotted against protein concen-tration. The level of expression in M. smegmatislpBAK-7q wasapproximately ninefold higher than that in M. tuberculosis.
INFECT. IMMUN.
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
RECOMBINANT ANTIGENS IN M. SMEGMATIS 263
AkDa77664330
17 __u'-4b _-a-12 - -
1 2 3
BkDa66 I43
30 --1712
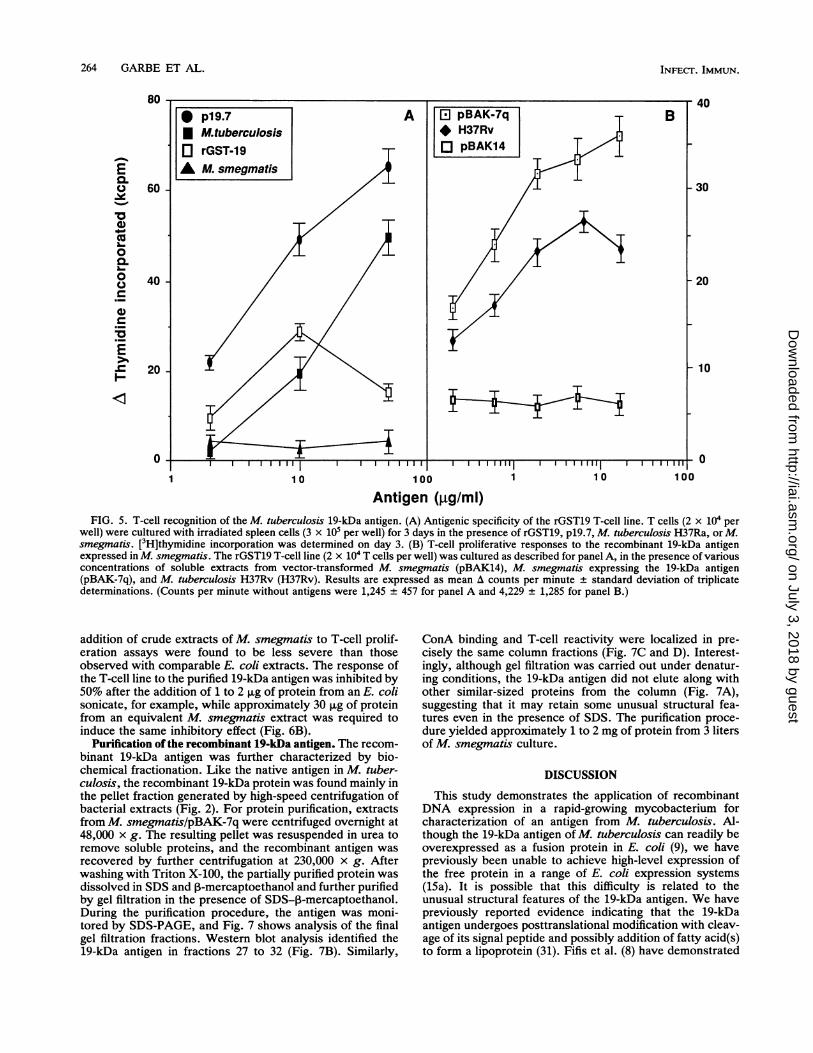
1 2 3FIG. 4. Carbohydrate association with the 19-kDa antigen. (A)
Neat extract from E. coli/pBAK-7q (lane 1) and 10-2 diluted extractfrom M. smegmatislpBAK-7q (lane 3) were analyzed by Westernblotting with monoclonal antibody F29-47 (1/2,000 dilution). Theapparent molecular mass of the 19-kDa antigen expressed in E. coliwas approximately 4 kDa lower than that of the antigen in M.smegmatis. This difference was most obvious when the two extractswere run together in the same lane of the gel (lane 2). (B) Proteinextracts from M. tuberculosis (lane 1) and M. smegmatis trans-formed with vector alone (pBAK14, lane 2) or with the 19-kDa gene(pBAK-7q, lane 3) were analyzed by SDS-PAGE and subsequentstaining with peroxidase-conjugated ConA. A ConA-positive bandwas seen in the position of the 19-kDa antigen in M. tuberculosis andin the extract from recombinant M. smegmatis/pBAK-7q.
smegmatis was estimated to be approximately ninefoldhigher than that in M. tuberculosis. This increase would beconsistent with the presence of multiple copies of the plas-mid-encoded gene in the recombinant strain, and it is prob-able that expression of the recombinant 19-kDa antigen isregulated by recognition of its own expression signals in themycobacterial host. The absence of a major additional19-kDa band in gels stained with Coomassie blue (notshown) indicates that the recombinant product accounts forno more than 1% of the total protein in the M. smegmatisextracts.Carbohydrate associated with the 19-kDa antigen. A low
level of expression of the 19-kDa antigen was detected byimmunoblot of E. coli transformed with pBAK-7q. We notedthat the antigen expressed in E. coli migrated with a differentapparent molecular weight during SDS-PAGE than that ofthe native M. tuberculosis antigen and the recombinant M.smegmatis product (Fig. 4A). This molecular mass differ-ence, corresponding to approximately 4 kDa, was mostapparent when extracts from the two recombinant strainswere combined and run in a single lane on the gel (Fig. 4A,lane 2). A further difference was noted when blots werestained with peroxidase-conjugated ConA, a lectin specificfor t-D-mannose and a-D-glucose, for detection of carbohy-drate residues. This procedure highlighted a number of
TABLE 1. Inhibition of ConA binding by a-methyl mannosidea
% Absorbance at a-methyl mannosideSample concn of":
0 10 mM 100 mM IM
M. tuberculosis 100 65 53 0M. smegmatis/pBAK-7q 100 50 34 0
a Nitrocellulose blots prepared with protein extracts from M. tuberculosisand M. smegmatis/pBAK-7q were screened for ConA binding as described inthe legend to Fig. 4, except that different concentrations of a-methyl manno-side were added during incubation with peroxidase-conjugated ConA. ConAbinding to the 19-kDa band on blots was quantitated by use of a scanningdensitometer.
b Results are expressed as a percentage of the absorbance in control laneswithout a-methyl mannoside.
discrete bands in M. tuberculosis extracts, including a prom-inent band in the position of the 19-kDa antigen (Fig. 4B,lane 1). Fewer ConA-positive bands were seen in extractsfrom M. smegmatis, but an intense staining of the 19-kDaprotein was strikingly evident in the recombinant extract(Fig. 4B, lane 3). ConA binding to the 19-kDa antigen in M.tuberculosis and M. smegmatis was completely inhibited byinclusion of a-methyl mannoside during incubation withConA-peroxidase (Table 1). The lower-molecular-mass 19-kDa antigen expressed in E. coli/pBAK-7q was not stainedby ConA, although weak ConA binding could be detected inblots prepared by loading high concentrations of rGST19(data not shown). The altered electrophoretic mobility andConA affinity are indicative of posttranslational modificationof the mycobacterium-expressed antigen, although our re-sults do not exclude the possibility of a very tight bindingbetween the 19-kDa antigen and some mycobacterium-spe-cific carbohydrate moiety, which remains intact even duringelectrophoresis in the presence of SDS. Staining of blotswith an antibody directed to mycobacterial lipoarabinoman-nan (LAM) (ML34 [11, 12]) demonstrated the presence ofsome LAM in purified antigen preparations but did notdetect any LAM associated with the 19-kDa band on SDS-PAGE (data not shown).
T-cell recognition of the recombinant 19-kDa antigen. Amurine T-cell line specific for the M. tuberculosis 19-kDaantigen was generated to investigate the immunologicalactivity of the recombinant antigen expressed in M. smeg-matis. The antigenic specificity of this line was confirmed bystrong in vitro proliferative responses to rGST19 and to apeptide (p19.7, residues 61 to 80) containing the majormurine T-cell epitope in the M. tuberculosis 19-kDa antigen(9). The T-cell line also responded vigorously to the 19-kDaprotein expressed in M. tuberculosis, whereas M. smegma-tis failed to induce a significant proliferative response (Fig.5A). The lack of response to M. smegmatis is consistent withserological evidence indicating that M. smegmatis does notexpress a protein with antigenic cross-reactivity to the M.tuberculosis 19-kDa antigen (3, 5, 10). After transformationwith pBAK-7q, however, M. smegmatis extracts induced astrong response and were four to five times more potent inT-cell proliferation assays than M. tuberculosis H37Rv ex-tracts (Fig. 5B).The recombinant 19-kDa antigen was also efficiently pre-
sented for T-cell recognition when added to the assay in theform of intact bacteria. Proliferative responses were inducedby both live and killed M. smegmatis/pBAK-7q, while thecontrol M. smegmatis/pBAK14 failed to stimulate significantresponses (Fig. 6A). Inhibitory effects resulting from the
VOL. 61, 1993
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
264 GARBE ET AL.
80
60
40
20
0
1 00
Antigen (gg/mi)
40
30
20
10
0
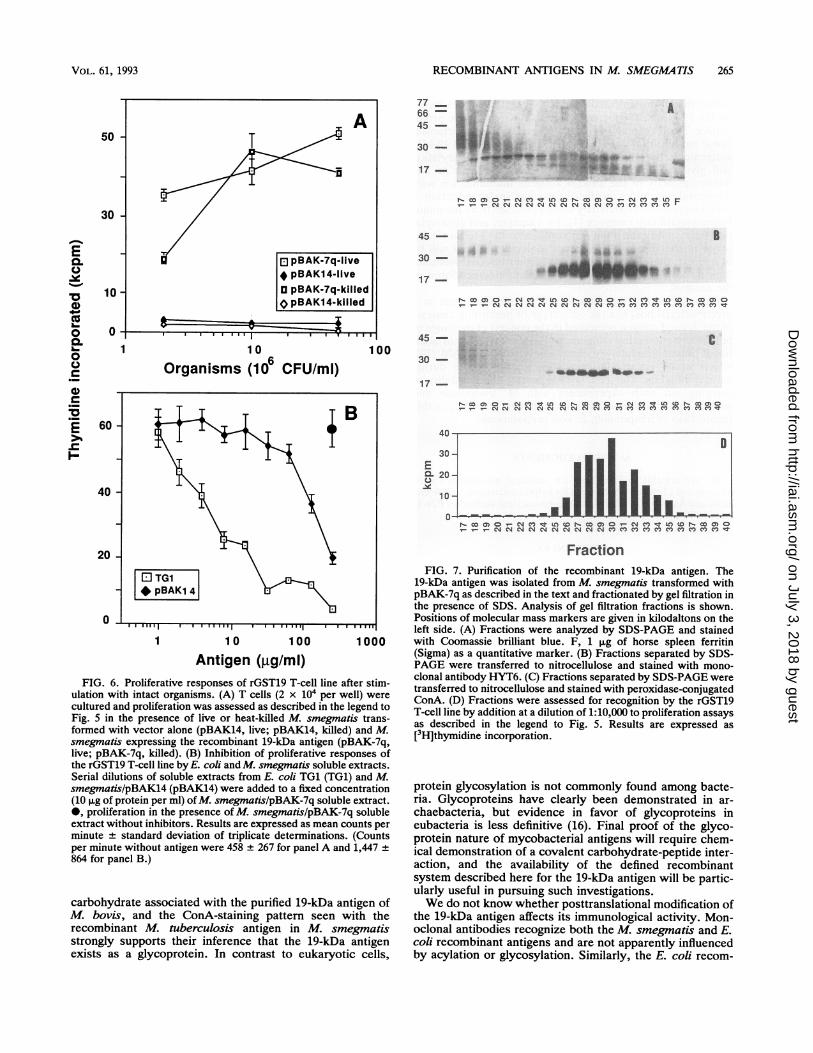
FIG. 5. T-cell recognition of the M. tuberculosis 19-kDa antigen. (A) Antigenic specificity of the rGST19 T-cell line. T cells (2 x 104 per
well) were cultured with irradiated spleen cells (3 x 105 per well) for 3 days in the presence of rGST19, p19.7, M. tuberculosis H37Ra, or M.smegmatis. [3H]thymidine incorporation was determined on day 3. (B) T-cell proliferative responses to the recombinant 19-kDa antigenexpressed in M. smegmatis. The rGST19 T-cell line (2 x 104 T cells per well) was cultured as described for panel A, in the presence of variousconcentrations of soluble extracts from vector-transformed M. smegmatis (pBAK14), M. smegmatis expressing the 19-kDa antigen(pBAK-7q), and M. tuberculosis H37Rv (H37Rv). Results are expressed as mean A counts per minute ± standard deviation of triplicatedeterminations. (Counts per minute without antigens were 1,245 ± 457 for panel A and 4,229 ± 1,285 for panel B.)
addition of crude extracts of M. smegmatis to T-cell prolif-eration assays were found to be less severe than thoseobserved with comparable E. coli extracts. The response ofthe T-cell line to the purified 19-kDa antigen was inhibited by50% after the addition of 1 to 2 ,ug of protein from an E. colisonicate, for example, while approximately 30 ,ug of proteinfrom an equivalent M. smegmatis extract was required toinduce the same inhibitory effect (Fig. 6B).
Purification ofthe recombinant 19-kDa antigen. The recom-binant 19-kDa antigen was further characterized by bio-chemical fractionation. Like the native antigen in M. tuber-culosis, the recombinant 19-kDa protein was found mainly inthe pellet fraction generated by high-speed centrifugation ofbacterial extracts (Fig. 2). For protein purification, extractsfrom M. smegmatislpBAK-7q were centrifuged overnight at48,000 x g. The resulting pellet was resuspended in urea toremove soluble proteins, and the recombinant antigen wasrecovered by further centrifugation at 230,000 x g. Afterwashing with Triton X-100, the partially purified protein wasdissolved in SDS and 3-mercaptoethanol and further purifiedby gel filtration in the presence of SDS-13-mercaptoethanol.During the purification procedure, the antigen was moni-tored by SDS-PAGE, and Fig. 7 shows analysis of the finalgel filtration fractions. Western blot analysis identified the19-kDa antigen in fractions 27 to 32 (Fig. 7B). Similarly,
ConA binding and T-cell reactivity were localized in pre-cisely the same column fractions (Fig. 7C and D). Interest-ingly, although gel filtration was carried out under denatur-ing conditions, the 19-kDa antigen did not elute along withother similar-sized proteins from the column (Fig. 7A),suggesting that it may retain some unusual structural fea-tures even in the presence of SDS. The purification proce-dure yielded approximately 1 to 2 mg of protein from 3 litersof M. smegmatis culture.
DISCUSSION
This study demonstrates the application of recombinantDNA expression in a rapid-growing mycobacterium forcharacterization of an antigen from M. tuberculosis. Al-though the 19-kDa antigen of M. tuberculosis can readily beoverexpressed as a fusion protein in E. coli (9), we havepreviously been unable to achieve high-level expression ofthe free protein in a range of E. coli expression systems(1Sa). It is possible that this difficulty is related to theunusual structural features of the 19-kDa antigen. We havepreviously reported evidence indicating that the 19-kDaantigen undergoes posttranslational modification with cleav-age of its signal peptide and possibly addition of fatty acid(s)to form a lipoprotein (31). Fifis et al. (8) have demonstrated
0.10-0
.1-*
-0
0.o0
:5._
E.--S
INFECT. IMMUN.
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
RECOMBINANT ANTIGENS IN M. SMEGAL4TIS 265
1 0
Organisms (106 CFU/mI)
10 100
77 _6645
30-17- *1M. *
17 - c
A
B45 -
30 -
17 -
N W 0 01Y N C) t4 CLO CD N C Q 0 r- N NN C<D N COC) 0
100
1000
Antigen (,g/mi)FIG. 6. Proliferative responses of rGST19 T-cell line after stim-
ulation with intact organisms. (A) T cells (2 x 104 per well) werecultured and proliferation was assessed as described in the legend toFig. 5 in the presence of live or heat-killed M. smegmatis trans-formed with vector alone (pBAK14, live; pBAK14, killed) and M.smegmatis expressing the recombinant 19-kDa antigen (pBAK-7q,live; pBAK-7q, killed). (B) Inhibition of proliferative responses ofthe rGST19 T-cell line by E. coli and M. smegmatis soluble extracts.Serial dilutions of soluble extracts from E. coli TG1 (TG1) and M.smegmatislpBAK14 (pBAK14) were added to a fixed concentration(10 p,g of protein per ml) ofM. smegmatis/pBAK-7q soluble extract.0, proliferation in the presence of M. smegmatis/pBAK-7q solubleextract without inhibitors. Results are expressed as mean counts perminute + standard deviation of triplicate determinations. (Countsper minute without antigen were 458 + 267 for panel A and 1,447864 for panel B.)
carbohydrate associated with the purified 19-kDa antigen ofM. bovis, and the ConA-staining pattern seen with therecombinant M. tuberculosis antigen in M. smegmatisstrongly supports their inference that the 19-kDa antigenexists as a glycoprotein. In contrast to eukaryotic cells,
C45 -
30 -
17 -
1N Co a) 0 Nces c e LO CD 9 co a 0 N XC IC') U) tD N co an 0_ _ _ eN N NJ N NN NN N NCOX C O C') Cl CO)CO
3CEQL 2Cl
ic
0
N CO 0 0 N C) R M C N CO ) 0 - N CO O O N seD 0a:0o
FractionFIG. 7. Purification of the recombinant 19-kDa antigen. The
19-kDa antigen was isolated from M. smegmatis transformed withpBAK-7q as described in the text and fractionated by gel filtration inthe presence of SDS. Analysis of gel filtration fractions is shown.Positions of molecular mass markers are given in kilodaltons on theleft side. (A) Fractions were analyzed by SDS-PAGE and stainedwith Coomassie brilliant blue. F, 1 ,ug of horse spleen ferritin(Sigma) as a quantitative marker. (B) Fractions separated by SDS-PAGE were transferred to nitrocellulose and stained with mono-clonal antibody HYT6. (C) Fractions separated by SDS-PAGE weretransferred to nitrocellulose and stained with peroxidase-conjugatedConA. (D) Fractions were assessed for recognition by the rGST19T-cell line by addition at a dilution of 1:10,000 to proliferation assaysas described in the legend to Fig. 5. Results are expressed as[3H]thymidine incorporation.
protein glycosylation is not commonly found among bacte-ria. Glycoproteins have clearly been demonstrated in ar-chaebacteria, but evidence in favor of glycoproteins ineubacteria is less definitive (16). Final proof of the glyco-protein nature of mycobacterial antigens will require chem-ical demonstration of a covalent carbohydrate-peptide inter-action, and the availability of the defined recombinantsystem described here for the 19-kDa antigen will be partic-ularly useful in pursuing such investigations.We do not know whether posttranslational modification of
the 19-kDa antigen affects its immunological activity. Mon-oclonal antibodies recognize both the M. smegmatis and E.coli recombinant antigens and are not apparently influencedby acylation or glycosylation. Similarly, the E. coli recom-
50
30
pBAK-7q-livepBAK14-live
o pBAK-7q-killed0 pBAK14-killed
-r
E0.0
._10
0s.0
L-
10 -
0 -
1
60 -B
40 -
20 -
1
VOL. 61, 1993
r- W M 0 r- N M I.P M W r- M M 0 r--V-- V-- v 0 m m m
!N 0
4. .. 41*
..., 04wi ip
0 TG1# pBAK1 4
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

266 GARBE ET AL.
binant fusion protein, and several synthetic peptides basedon the 19-kDa sequence, have been shown to induce prolif-erative T-cell responses (7, 9), indicating that posttransla-tional modification is not essential for expression of 19-kDaantigenicity. It remains possible, however, that acylationand glycosylation may have more subtle effects on theimmunogenicity of the 19-kDa antigen. Immunization withlipopeptides provides an efficient mechanism for induction ofT-cell responses restricted by class I proteins from the majorhistocompatibility complex (4), for example, and glycosyla-tion may modify the ability of peptides to bind to particularmajor histocompatibility complex molecules (10). It will beof interest, therefore, to carry out further detailed immuno-logical comparisons of the modified and unmodified forms ofthe antigen.Expression of a recombinant M. tuberculosis gene in a
mycobacterial host has the advantage of permitting analysisof the defined antigen presented in the context of the range ofpotentially competing antigens and in the presence of addi-tional immunomodulatory components, such as LAM (19),which would accompany exposure to the antigen duringnatural infection. Our results demonstrate the feasibility ofcarrying out such experiments with M. smegmatis andsuggest that mycobacterial extracts may be less toxic in thisregard than corresponding E. coli preparations.
ACKNOWLEDGMENTS
We are grateful to Dan Tang for technical assistance, CarlosMoreno and Christiane Abou-Zeid for helpful discussion, and ArendKolk and Ase Andersen for providing monoclonal antibodies F29-47and HYT6.
REFERENCES
1. Aldovini, A., and R. A. Young. 1991. Humoral and cell-mediatedimmune responses to live recombinant BCG-HIV vaccines.Nature (London) 351:479-482.
2. Ashbridge, K. R., R. J. Booth, J. D. Watson, and R. B. Lathigra.1989. Nucleotide sequence of the 19 kDa antigen gene fromMycobacterium tuberculosis. Nucleic Acids Res. 17:1249.
3. Coates, A. R. M., J. Hewitt, B. W. Allen, J. Ivanyi, and D. A.Mitchison. 1981. Antigenic diversity of Mycobacterium tuber-culosis and Mycobacterium bovis detected by means of mono-clonal antibodies. Lancet ii:167-169.
4. Deres, K., H. Schild, K.-H. Wiesmuller, G. Jung, and H.-J.Rammensee. 1989. In vivo priming of virus-specific, cytotoxic Tlymphocytes with synthetic lipopeptide vaccine. Nature (Lon-don) 342:561-564.
5. Engers, H. D., and Workshop Participants. 1986. Results of a
World Health Organization-sponsored workshop to character-ize antigens recognized by mycobacterium-specific monoclonalantibodies. Infect. Immun. 51:718-720. (Letter to the editor.)
6. Espitia, C., and R. Mancilla. 1989. Identification, isolation andpartial characterization of Mycobacterium tuberculosis glyco-protein antigens. Clin. Exp. Immunol. 77:378-383.
7. Faith, A., C. Moreno, R. Lathigra, E. Roman, M. Fernandez, S.Brett, D. M. Mitchell, J. Ivanyi, and A. D. M. Rees. 1991.Analysis of human T-cell epitopes in the 19,000 MW antigen ofMycobacterium tuberculosis: influence of HLA-DR. Immunol-ogy 74:1-7.
8. Fifis, T., C. Costopoulos, A. J. Radford, A. Bacic, and P. R.Wood. 1991. Purification and characterization of major antigensfrom a Mycobacterium bovis culture filtrate. Infect. Immun.59:800-807.
9. Harris, D. P., H. M. Vordermeier, E. Roman, R. Lathigra, S. J.Brett, C. Moreno, and J. Ivanyi. 1991. Murine T cell-stimulatorypeptides from the 19-kDa antigen of Mycobacterium tuberculo-sis: epitope restricted homology with the 28kDa protein of
Mycobacterium leprae. J. Immunol. 147:2706-2712.10. Ishioka, G. Y., A. G. Lamont, D. Thomson, A. Bulbow, F. C. A.
Gaeta, A. Sette, and H. M. Grey. 1992. MHC interaction and Tcell recognition of carbohydrate and glycopeptides. J. Immunol.148:2446-2451.
11. Ivanyi, J., K. Sharp, P. Jackett, and G. Bothamley. 1988.Immunological study of defined constituents of mycobacteria.Springer Semin. Immunopathol. 10:279-300.
12. Ivanyi, J., S. Sinha, R. Aston, D. Cussell, M. Keen, and U.Sengupta. 1983. Definition of species specific and cross-reactiveantigenic determinants of Mycobacterium leprae using mono-clonal antibodies. Clin. Exp. Immunol. 52:528-536.
13. Jackett, P. S., G. H. Bothamley, H. V. Bathra, A. Mistry, D. B.Young, and J. Ivanyi. 1988. Specificity of antibodies to immun-odominant mycobacterial antigens in pulmonary tuberculosis. J.Clin. Microbiol. 26:2313-2318.
14. Jacobs, W. R., M. Tuckman, and B. R. Bloom. 1987. Introduc-tion of foreign DNA into mycobacteria using a shuttle phasmid.Nature (London) 327:532-535.
15. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
15a.Lathigra, R. B. Unpublished data.16. Lechner, J., and F. Wieland. 1989. Structure and biosynthesis of
prokaryotic glycoproteins. Annu. Rev. Biochem. 58:173-194.17. Matsuo, K., R. Yamaguchi, A. Yamazaki, H. Tasaka, K.
Terasaka, and T. Yamada. 1990. Cloning and expression of thegene for the cross-reactive ot antigen of Mycobacterium kan-sasii. Infect. Immun. 58:550-556.
18. Mehlert, A., and D. B. Young. 1989. Biochemical and antigeniccharacterization of the Mycobacterium tuberculosis 71 kD an-tigen, a member of the 70 kD heat shock protein family. Mol.Microbiol. 3:125-130.
19. Moreno, C., J. Taverne, A. Mehlert, C. A. W. Bate, R. Brealey,A. Meager, G. A. W. Rook, and J. H. L. Playfair. 1989.Lipoarabinomannan from Mycobacterium tuberculosis inducesthe production of tumour necrosis factor from human andmurine macrophages. Clin. Exp. Immunol. 76:240-245.
20. Nair, J., D. A. Rouse, and S. L. Morris. 1992. Nucleotidesequence analysis and serologic characterization of the Myco-bacterium intracellulare homologue of the Mycobacterium tu-berculosis 19kDa antigen. Mol. Microbiol. 6:1431-1439.
21. Ranes, M. G., J. Rauzier, M. Lagranderie, M. Gheorghiu, andB. Gicquel. 1990. Functional analysis of pAL5000, a plasmidfrom Mycobacterium fortuitum: construction of a 'mini' Myco-bacterium-Escherichia coli shuttle vector. J. Bacteriol. 172:2793-2797.
22. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, N.Y.
23. Snapper, S. B., L. Lugosi, A. Jekkel, R. E. Melton, T. Kieser,B. R. Bloom, and W. R. Jacobs. 1988. Lysogeny and transfor-mation in mycobacteria: stable expression of foreign genes.Proc. Natl. Acad. Sci. USA 85:6987-6991.
24. Snapper, S. B., R. E. Melton, S. Mustafa, T. Kieser, and W. R.Jacobs, Jr. 1990. Isolation and characterization of efficientplasmid transformation mutants of Mycobacterium smegmatis.Mol. Microbiol. 4:1911-1919.
25. Stover, C. K., V. F. de la Cruz, T. R. Fuerst, J. E. Burlein, L. A.Benson, L. T. Bennett, G. P. Bansal, J. F. Young, M. H. Lee,G. F. Hatfull, S. B. Snapper, R. G. Barletta, W. R. Jacobs, Jr.,and B. R. Bloom. 1991. New use of BCG for recombinantvaccine. Nature (London) 351:456-460.
26. Thangaraj, H., F. I. Lamb, E. 0. Davis, P. J. Jenner, L. H.Jeyakumar, and M. J. Colston. 1990. Identification, sequencing,and expression of Mycobacterium leprae superoxide dismutase,a major antigen. Infect. Immun. 58:1937-1942.
27. Thole, J. E. R., W. J. Keulen, A. H. J. Kolk, D. G. Groothuis,L. G. Berwald, R. H. Thiejema, and J. D. A. van Embden. 1987.Characterization, sequence determination, and immunogenicityof a 64-kilodalton protein of Mycobacterium bovis BCG ex-
pressed in Escherichia coli K-12. Infect. Immun. 55:1466-1475.28. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretic
INFECT. IMMUN.
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

RECOMBINANT ANTIGENS IN M. SMEGMATIS 267
transfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 76:4350-4354.
29. Vordermeier, H. M., D. P. Harris, E. Roman, R. Lathigra, C.Moreno, and J. Ivanyi. 1991. Identification of T cell stimulatorypeptides from the 38-kDa protein of Mycobacterium tuberculo-sis. J. Immunol. 147:1023-1029.
30. Young, D., T. Garbe, R. Lathigra, and C. Abou-Zeid. 1990.Protein antigens: structure, function and regulation, p. 1-35. InJ. McFadden (ed.), Molecular biology of the mycobacteria.
Academic Press Ltd., London.31. Young, D. B., and T. R. Garbe. 1991. Lipoprotein antigens of
Mycobacterium tuberculosis. Res. Microbiol. 142:55-65.32. Young, D. B., S. H. E. Kaufmann, P. W. M. Hermans, and
J. E. R. Thole. 1992. Mycobacterial protein antigens: a compi-lation. Mol. Microbiol. 6:133-145.
33. Zhang, Y., R. Lathigra, T. Garbe, D. Catty, and D. Young. 1991.Genetic analysis of superoxide dismutase, the 23 kilodaltonantigen of Mycobacterium tuberculosis. Mol. Microbiol. 5:381-391.
VOL. 61, 1993
on July 3, 2018 by guesthttp://iai.asm
.org/D
ownloaded from















![PBAK [Autosaved]](https://static.fdocuments.net/doc/165x107/577c83eb1a28abe054b6d155/pbak-autosaved.jpg)


