
Expression Cloning of the STRL33/BONZO/TYMSTR … · Expression Cloning of the STRL33/BONZO/TYMSTR...
11
of September 17, 2018. This information is current as Chemokines Elements of CC, CXC, and CX3C STRL33/BONZO/TYMSTR Ligand Reveals Expression Cloning of the Andrew, Lijun Wu and Michael Briskin Simona Mak, Dulce Soler, Patricia Langdon, David P. Alyson Wilbanks, Susan Carr Zondlo, Kristine Murphy, http://www.jimmunol.org/content/166/8/5145 doi: 10.4049/jimmunol.166.8.5145 2001; 166:5145-5154; ; J Immunol References http://www.jimmunol.org/content/166/8/5145.full#ref-list-1 , 23 of which you can access for free at: cites 43 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2001 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on September 17, 2018 http://www.jimmunol.org/ Downloaded from by guest on September 17, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of Expression Cloning of the STRL33/BONZO/TYMSTR … · Expression Cloning of the STRL33/BONZO/TYMSTR...

of September 17, 2018.This information is current as
ChemokinesElements of CC, CXC, and CX3CSTRL33/BONZO/TYMSTR Ligand Reveals Expression Cloning of the
Andrew, Lijun Wu and Michael BriskinSimona Mak, Dulce Soler, Patricia Langdon, David P. Alyson Wilbanks, Susan Carr Zondlo, Kristine Murphy,
http://www.jimmunol.org/content/166/8/5145doi: 10.4049/jimmunol.166.8.5145
2001; 166:5145-5154; ;J Immunol
Referenceshttp://www.jimmunol.org/content/166/8/5145.full#ref-list-1
, 23 of which you can access for free at: cites 43 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2001 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Expression Cloning of the STRL33/BONZO/TYMSTR LigandReveals Elements of CC, CXC, and CX3C Chemokines
Alyson Wilbanks,1 Susan Carr Zondlo,1 Kristine Murphy, 1 Simona Mak, Dulce Soler,Patricia Langdon, David P. Andrew,2 Lijun Wu, and Michael Briskin 3
STRL33/BONZO/TYMSTR is an orphan chemokine and HIV/SIV coreceptor receptor that is expressed on activated T lympho-cytes. We describe an expression cloning strategy whereby we isolated a novel chemokine, which we name CXCL16. CXCL16 isan a (CXC) chemokine but also has characteristics of CC chemokines and a structure similar to fractalkine (neurotactin) in havinga transmembrane region and a chemokine domain suspended by a mucin-like stalk. A recombinant version of CXCL16 fails tomediate chemotaxis to all known chemokine receptor transfectants tested but does mediate robust chemotaxis, high affinitybinding, and calcium mobilization to Bonzo receptor transfectants, indicating that this is a unique receptor ligand interaction. Invitro polarized T cell subsets including Th1, Th2, and Tr1 cells express functional Bonzo, suggesting expression of this receptorin chronic inflammation, which we further verified by demonstration of CXCL16-mediated migration of tonsil-derived CD41 Tlymphocytes. CXCL16 is expressed on the surface of APCs including subsets of CD191 B cells and CD141 monocyte/macrophages,and functional CXCL16 is also shed from macrophages. The combination of unique structural features of both Bonzo andCXCL16 suggest that this interaction may represent a new class of ligands for this receptor family. Additionally, this chemokinemight play a unique dual role of attracting activated lymphocyte subsets during inflammation as well as facilitating immuneresponses via cell-cell contact.The Journal of Immunology,2001, 166: 5145–5154.
T he chemokines comprise an ever growing family of smallsecreted proteins involved in a number of inflammatoryand immunological processes including lymphocyte hom-
ing, suppression and stimulation of angiogenesis, suppressionof HIV infection, and enhancement of CTL responses (1–3). Allchemokines to date (with one exception noted) exhibit a con-served structure that is predominated by conserved disulfidebonds, Cys1-Cys3 and Cys2-Cys4 (1, 3). To date there are foursubfamilies, based on the arrangement of N-terminal cysteineresidues including the CC, CXC, and one member each of theC and the CX3C whereby, in each case, X represents any res-idue other than cysteine (1, 3). The selective binding of thechemokine subfamily members to G protein-coupled receptors(GPCRs) results in the subclassification of chemokine receptorsas CCR1–11, CXC chemokine receptor (CXCR)41–5, XCR1,and CX3CR1 (1, 2, 4 – 8). The selectivity of these receptor li-gand interactions will, in part, contribute to the specificity bywhich leukocyte subsets might preferentially localize to differ-ent extravascular sites (4, 9, 10). However, in many instances,as multiple receptors appear to be expressed on individual sub-
sets it is likely that overlapping gradients of different chemo-kines and combinations of receptor ligand interactions mightdetermine directionality of leukocyte trafficking from the cir-culation into tissues (9, 11).
Recently, several GPCRs have been cloned with no known nat-ural ligands and are thus referred to as “orphan GPCRs” (12). Oneorphan receptor referred to as STRL33/BONZO/TYMSTR (re-ferred to herein as Bonzo), simultaneously identified in threegroups, was shown to be expressed on activated T cells, and canserve as an HIV/SIV coreceptor. Subsequent studies have shownBonzo to be a receptor more frequently expressed on memory Tcells with a preferential expression on CD8-positive T cells (but isexpressed on CD4-positive cells as well) (13–15). As no knownchemokines have been shown to interact with Bonzo, it has re-mained an orphan receptor, while sequence comparisons stronglysuggest that it is a CCR. Here, we use a novel expression cloningstrategy to isolate a cDNA that upon transfection into 293 cellrecipients mediates a robust chemotactic response to Bonzo recep-tor transfectants. This cDNA encoded a novel chemokine, whichwe term CXCL16.
Although it is, in the strictest definition, a member of theCXC family, CXCL16 is distantly related to all known chemo-kines and might be phylogenetically closer to certainb chemo-kines thana chemokines. Additionally, it has a novel featurethat is shared with the only known CX3C chemokine, neuro-tactin/fractalkine, in that sequence predicts that it is membranebound and suspended by a heavily glycosylated mucin stalk (16,17). We show that Bonzo/CXCL16 interactions define a uniquereceptor ligand pair as CXCL16 fails to bind all known che-mokine receptors. We also show in this study that this ligandfunctionally interacts with activated lymphocytes and describenovel features of expression on leukocyte subsets. These data,along with the structural features of both Bonzo and CXCL16,indicate that this receptor ligand pair is unique among all mem-bers of this family known to date.
Millennium Pharmaceuticals, Cambridge, MA 02139
Received for publication November 27, 2000. Accepted for publication February5, 2001.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 A.W., S.C.Z., and K.M. contributed equally to this work.2 Current address: CuraGen Corporation, 322 East Main Street, Branford, CT 06405.3 Address correspondence and reprint requests to Dr. Michael J. Briskin, MillenniumPharmaceuticals, Sidney Street, Cambridge, MA 02139. E-mail address:[email protected] Abbreviations used in this paper: CXCR, CXC chemokine receptor; GPCR, Gprotein-coupled receptor; BAB, binding assay buffer; MDC, macrophage-derivedchemokine; MIP, macrophage-inflammatory protein; SDF, stromal cell-derivedfactor; SLC, secondary lymphoid chemokine; EF1, elongation factor-1.
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Materials and MethodsmAbs, isolation of primary cells, and cell lines
Medium for transfectants was standard DMEM with 10% FCS (Life Tech-nologies, Gaithersburg, MD). L1-2 cells, a murine B cell lymphoma, wereobtained from E. Butcher at Stanford University (Stanford, CA). Receptortransfectants expressing Bonzo were generated as previously described (18,19). Chemokine receptor transfectants encoding CXCR5, CCR1–9, andCX3CR1 were maintained in RPMI 1649 medium (Life Technologies)with 0.88 g/L gentamicin (G418), 10% HyClone serum, 10 nM HEPES,1% penicillin/streptomycin, 1%L-glutamine, 1 mM sodium pyruvate, and55 nM 2-ME.
Chronically activated Th1, Tr1, and Th2 lymphocytes were prepared aspreviously described (20, 21) with the addition of generation of Tr1 sub-sets. For Tr1 lymphocytes IL-10 was used at 10 ng/ml. After initial acti-vation in the presence of anti-CD28 and anti-CD3 mAbs Th1, Tr1, and Th2lymphocytes were restimulated for 5 days with specific cytokines, but withthe addition of anti-CD95 ligand (1mg/ml) to prevent apoptosis. ActivatedTh1, Tr1, and Th2 lymphocytes were maintained in this way for a maxi-mum of three cycles.
Tonsils were obtained from Massachusetts Eye and Ear (Cambridge,MA). Tissue was macerated with surgical scissors, mixed with DMEM(Life Technologies) and passed through a cell strainer (Becton Dickinson,Franklin Lakes, NJ). Cells were washed three times with PBS before stain-ing. CD41 T cells were isolated by positive magnetic selection using CD4microbeads (Miltenyi Biotec, Auburn, CA) and the manufacturer’sinstructions.
PBMC were isolated by density gradient centrifugation using Lym-phoprep (Nycomed, Oslo, Norway). Monocytes were seeded into T75flasks, allowed to adhere, and cultured for 10 days in RPM1 1640 supple-mented with 2.5 mM HEPES, 20mg/ml gentamicin, 2 mML-glutamine,1% penicillin/streptomycin, 2% nonessential amino acids, and 1 mM so-dium pyruvate (all obtained from Life Technologies). On the 10th day ofculture, cells were incubated with either 50 ng/ml LPS (Sigma, St. Louis,MO) or 10 ng/ml TNF-a (R&D Systems, Minneapolis, MN) for 4 and 24 h.Supernatants were drawn off the cells and used in chemotaxis experimentswith Bonzo/L1.2 transfectants and GusB (CCR11)/L1.2 transfectants as anegative control.
MAbs reactive with Bonzo were generated by immunizing mice withL1.2 cells expressing high levels of transfected Bonzo, as previously de-scribed (19). These mAbs were screened to ensure selectivity on numerousL1.2 transfectants expressing chemokine receptors (CCR1–CCR8,CXCR1–CXCR5, GPR5, V28, and GPR9-6) or orphan GPCRs (Bob,LyGPR, AF, APJ, and RDC). Chemokines were obtained from R&D Sys-tems, PeproTech (Rocky Hill, NJ), or were synthesized using an automatedsolid-phase peptide synthesizer using previously described methods (22).MAbs to CXCL16 were generated by immunization of BALB/C mice withthe synthetic chemokine and screening for reactivity by ELISA.
FACS staining
Before the addition of primary Ab, PBMCs or tonsil cells were incubatedin PBS with 5% True Clot human serum (Scantibodies, Santee, CA) andmouse IgG (Sigma) to prevent nonspecific staining. Cells were incubatedfor 30 min with specific mAbs or isotype controls (Sigma). FITC- or Cy5-goat anti-mouse IgG Abs (Jackson ImmunoResearch, West Grove, PA)were used as the secondary Abs at 1/200 dilutions. Abs against all surfaceAgs including CD4, CD14, and CD19 directly conjugated to PE were ob-tained from PharMingen (San Diego, CA). Cell staining was analyzed ona FACScan (Becton Dickinson) using the CellQuest program.
cDNA library expression cloning
An expression library was made from spleen mRNA purchased from Clon-tech Laboratories (Palo Alto, CA). cDNA was prepared as previously de-scribed (23) using synthesis reagents from Life Technologies with the ex-ception that an ECOR1 adapter (Pharmacia, Piscataway, NJ) was ligated tothe 59 end to facilitate directional cloning into the elongation factor-1(EF1)-based vector pcDEF3. cDNA was partitioned into 96 pools of 1000colonies each, and plasmid DNAs were prepared as previously described(23).
Transfections were performed into 293T cells as described previously(23) with the exception that 60,000 cells/well were plated in collagen-coated plates (Becton Dickinson). Approximately 18 h after transfection,medium was changed to 0.5 ml/well of standard DMEM/10% FCS. Forty-eight hours after changing medium, supernatant was harvested, cell debriswas removed by microcentrifugation, and the medium was then used inchemotaxis assays.
For the chemotaxis assays in the expression-cloning screen (and withmacrophage supernatants), exponentially growing Bonzo transfectantswere resuspended at a density of 13 107/ml in an assay buffer that con-sisted of DMEM supplemented with 10% bovine calf serum. The cell sus-pension (100ml) was placed in the upper chamber of a 24-well chemotaxisplate (Costar; Corning Glass, Corning, NY), and 0.5 ml of the supernatantfrom each transfected well was placed in the lower chamber. The plateswere then incubated for 6–24 h at 37°C. Numbers of migrating cells werequantitated on a FACScan (Becton Dickinson) using the acquisition phaseat 30 s. cDNA pools that, upon transfection, mediated chemotaxis abovebackground were subsequently enriched as previously described (23).
Chemotaxis with purified CXCL16 (described below) and L1-2 receptortransfectants was identical with the set-up for expression cloning with theexception that defined concentrations of chemokines were used and thebuffer was changed to 50% RPMI, 50% M199 medium, and 0.5% BSA.Chemotaxis with in vitro-derived effector cells and isolated CD41 tonsillymphocytes was performed using 3-mm pore diameter gelatin-coatedtranswell inserts followed by growth of 23 105 ECV304 cells as previ-ously described (20). An aliquot of 200ml of cell suspension (input of 83105 cells) was added to each insert. After 2 h, the inserts were removed andthe number of cells that had migrated through the ECV304 monolayer tothe lower well was counted for 30 s on a Becton Dickinson FACScan withthe gates set to acquire the cells of interest. Using this technique 100%migration would be 25,000 cells for Th1/Th2 cells, where this numberrepresents the cells in the lower well counted on the FACScan for 1 min.In all cases the data points were the result of duplicate wells, with the meanvalue shown and the error bars representing the sample SD.
Sequencing and sequence analysis
Sequencing of the entire cDNA insert was accomplished in conjunctionwith Seqwrite (Houston, TX) and the Tufts University sequencing corefacility. Overlapping primers (originally using an SP6 primer for the 39endand a primer from the EF1 promoter for the 59sequence) made to bothstrands as sequence information was gathered, resulting in complete un-ambiguous sequence of both strands of the insert. Sequence comparisonwith known chemokines was performed with the Lasergene system(DNAstar, Madison, WI), using the Clustal method with a gap penalty of10 and a gap length penalty of 10. Pairwise alignment parameters were:ktuple 5 2, gap penalty-5, window5 4, and diagonals saved5 4.
Construction and purification of recombinant CXCL16 His-tagged protein
Fusion proteins consisting of regions of CXCL16 fused to a C-terminal 53histidine (His) were made in the pEF1/V5-His vector from Invitrogen(Carlsbad, CA) by fusion with a PCR-generated fragment containing theentire extracellular domain of CXCL16. A 59primer starting at the initi-ation methionine with aBamHI site and a 39primer ending at Val155 withan XbaI site were used in the PCR. Primer sequences are available uponrequest. PCR inserts were purified from agarose gels along with vectorDNA digested withBamHI andXbaI and recombinants isolated by stan-dard procedures. Maxiprep (Qiagen, Chatsworth, CA) DNA was used intransient transfections of 10-cm plates using Lipofectamine (Life Technol-ogies). Three days after transfection, supernatant was collected and cen-trifuged, and the clarified supernatant was then run over 5 ml wheat germagglutinin conjugated to agarose in a column (Vector Laboratories, Bur-lingame, CA). PBS was used as the wash buffer and eluted with 100 mMacetic acid (pH 2.8). The 10-ml elution was brought to pH neutrality with5 ml 1 M Tris base (pH 10.5) and run over an nickel-nitrilotriacetic acid-agarose column (Qiagen) washed with 50 mM NaH2PO4 (pH 8), 300 mMNaCl, and 20 mM imidazole. Elution was performed with 50 mMNaH2PO4 (pH 8), 300 mM NaCl, and 250 mM imidazole in 53 1-mlaliquots. Samples were dialyzed using a 3-ml 10,000 m.w. cutoff Slide-a-lyzer (Pierce, Rockford, IL) into PBS overnight. OD280 readings weretaken to determine the protein concentration.
A Glycoprotein Deglycosylation Kit (Calbiochem, San Diego, CA) wasused to deglycosylate 2mg of CXCL16 protein. The standard denaturingprotocol was followed, and 150 ng each of untreated and deglycosylase-treated protein was run on a 4–20% Tris-glycine gel (Novex, San Diego,CA). Protein was transferred onto a nitrocellulose membrane (Novex) andhybridized with a CXCL16 mAb (SD7). The NEN (Boston, MA) Renais-sance system was used for detection.
Receptor binding assays
Exponentially growing L1.2 Bonzo transfectants were counted on the dayof the assay and resuspended in binding assay buffer (BAB; 10 mMHEPES/1 mM CaCl2/5 mM MgCl2/0.5% BSA/0.05% sodium azide at
5146 EXPRESSION CLONING OF THE STRL33/TYMSTR/BONZO LIGAND
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

2.5 3 106/ml) at a density of 2.53 106/ml. Purified CXCL16 was labeledwith 125I using a sodium iodide method (Amersham, Arlington Heights, IL)and diluted in BAB to 4 nM. Cold CXCL16 (in PBS at 5.3mM) was dilutedfor competition in BAB to 0.4, 2, 4, 20, 40, and 400 nM. Reactions con-sisted, in triplicate (for a final volume of 100ml) of 50 ml of cells (1.253105 total), 25ml 125I-CXCL16 (final concentration5 1 nM), and 25ml coldCXCL16 serially diluted from 100 to 0 nM. The specific activity of thelabeled ligand was 7.83 1010 nmol/cpm, total counts bound were 813.67,nonspecific counts were 222.33 resulting in 591.34 specific counts binding.For calculation of total125I input, 50 ml cells was added to 25ml 125I-CXCL16 and 25ml BAB. All tubes were incubated for 1 h at room tem-perature. Cells were spun down at 3500 rpm and washed five times in BAB1 0.5 M NaCl. After the final wash, the cell pellet was resuspended in 100ml wash buffer, and the125I counts were calculated by a Cobra II Auto-Gamma scintillation counter (Packard, Meriden, CT) along with the totalinput sample. Binding data were calculated using a program written inExcel (L. Wu, unpublished observation).
Calcium flux assays
Bonzo/L1.2 transfectants or parental cells were washed once in PBS andresuspended in load buffer (HBSS, 20 mM HEPES, 2.5 mM probenecid,0.1% BSA, and 1% FBS). Fluo-3 (Molecular Probes, Eugene, OR) wasdissolved in 50% DMSO/50% pluronic acid and added to the cells at a finalconcentration of 4mM. Cells were incubated for 1 h at 37°C. Then, cellswere washed twice in load buffer and plated into 96-well assay plates at300,000 cells per well. The plate was spun for 5 min at 1200 rpm to pelletcells on the bottom of the well. Chemokine (50ml) was added to a separate96-well plate at varying concentrations to achieve final concentrations asindicated in the figure. Ca21 mobilization was then measured on a 96-wellFLIPR System (Molecular Devices, Sunnyvale, CA).
Northern and Southern blots
Human multiple tissue mRNA blots (Human I, Immune System II) fromClontech Laboratories were used for Northern blot analysis. A 400-bpcDNA fragment representing the chemokine domain from the 59EcoRI siteto a EcoRV site was used as the probe template and primed with randomhexamers to produce an [a-32P]dCTP-labeled probe. Hybridization wasperformed according to the manufacturer’s instructions with four addi-tional high stringency washes at 65°C in 0.13SSC and 0.1% SDS, andthen exposed to Kodak (Rochester, NY) XAR film with an intensifyingscreen. Blots were stripped and reprobed with ab-actin probe, the templateof which was provided by the blot manufacturer.
For Southern blots, human genomic DNA (15mg/reaction; ClontechLaboratories) was digested byBamHI, EcoRI, andHindIII and run on a1.4% agarose gel. DNA was transferred to a Protran nitrocellulose mem-brane (Schleicher & Schuell, Keene, NH) and cross-linked with a Strata-linker. The probe template was the same as for the Northern blots. The blotwas washed three times for 30 min at 60°C in 13 SSC and 0.1% SDS, andwas exposed to film for 2 days at room temperature.
ResultsExpression cloning reveals a novel chemokine
L1.2 Bonzo transfectants were tested in chemotaxis assays againsta panel of all known chemokine receptor ligands (along with or-phan chemokines), and no response was noted in any physiologicaldose range (data not shown). As this data suggested that Bonzolikely binds an unknown ligand, we subsequently devised a strat-egy whereby we could transfect pools of cDNAs, harvest super-natants, and ask whether we could detect a chemotactic response.As Bonzo had previously been shown to be highly expressed inspleen (13, 14), an expression library was constructed from spleenmRNA. Transient transfection of 96 pools (representing;1000independent clones/pool) revealed one pool with a response abovebackground. Subsequent rounds of enrichment increased the ac-tivity, and eventual deconvolution resulted in a single cDNA clonethat, upon transfection, mediated a robust chemotactic response toBonzo receptor transfectants (data not shown).
Sequence analysis reveals a novel chemokine
The isolated cDNA revealed a large open reading frame of 254 aawith an N-terminal hydrophobic predicted signal peptide of 27 aa(Fig. 1A). The signal peptide is followed by 90 residues with a total
of six cysteines, with an N-terminal CYC motif consistent withmembers of thea chemokine family (1, 3). The predicted chemo-kine domain is followed by a region that is largely composed ofS/T/G/P residues (37/84 aa or 44%), which is the hallmark of amucin domain (23), followed by a hydrophobic membrane span-ning a region of 25 residues and a short 28-residue cytoplasmictail, containing two additional cysteines in each of these regions.This overall structure is similar to the one CX3C chemokine, frac-talkine/neurotactin (16, 17), while a C-terminal mucin-like regionhas also been observed for JE (murine monocyte chemoattractantprotein-1) (24). Although this protein must be shed or proteolyti-cally cleaved for chemotactic activity, there are no known dibasiccleavage sites before the transmembrane region.
Alignment of this new sequence (using only the chemokine do-main) with a number of othera chemokines shows it is distantlyrelated to all CXC chemokines with the best homology with stro-mal cell-derived factor (SDF)-1 at 18.8% (Fig. 1B). Additionalsearches with different regions of the protein sequence reveal nohomology to any proteins in the database. Although the predictedprotein has characteristics of both CXC and CX3C chemokines,similarities to CC chemokines exist as well, including 1) homology(closer than to any CXC chemokines) to macrophage-inflamma-tory protein (MIP)-1b at 22%; 2) similarity to Eskine/CTAK andTeck in having 27 residues between Cys 2 and 3, while this loopis generally no longer than 24 residues in all other chemokines(25); and 3) a relationship to secondary lymphoid chemokine(SLC)/6-C-Kine in having six cysteine residues in its chemokinedomain (26, 27). Due to the CYC motif, we provisionally refer tothis protein as a novel CXC chemokine termed CXCL16 (28) andwe propose to rename STRL33/BONZO/TYMSTR as CXCR6.
FIGURE 1. Sequence of phylogenetic analysis of CXCL16.A, Pre-dicted signal peptide and transmembrane regions are indicated by a boldunderline. Past the signal peptide and before the line demarcating the mu-cin domain is the predicted chemokine domain. All cysteines are indicatedin bold. B, Dendrogram showing phylogenetic relationship of CXCL16with selecteda chemokines as well as MIP-1a and MIP-1b. These se-quence data are available from GenBank under accession numberAF337812.
5147The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
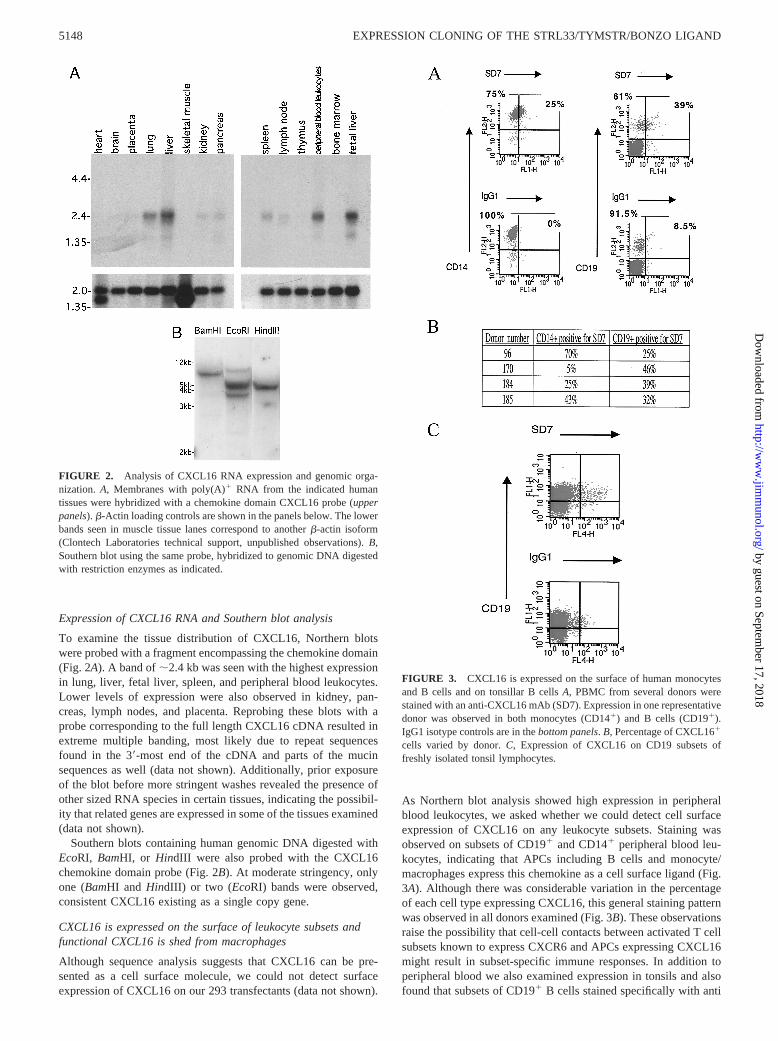
Expression of CXCL16 RNA and Southern blot analysis
To examine the tissue distribution of CXCL16, Northern blotswere probed with a fragment encompassing the chemokine domain(Fig. 2A). A band of;2.4 kb was seen with the highest expressionin lung, liver, fetal liver, spleen, and peripheral blood leukocytes.Lower levels of expression were also observed in kidney, pan-creas, lymph nodes, and placenta. Reprobing these blots with aprobe corresponding to the full length CXCL16 cDNA resulted inextreme multiple banding, most likely due to repeat sequencesfound in the 39-most end of the cDNA and parts of the mucinsequences as well (data not shown). Additionally, prior exposureof the blot before more stringent washes revealed the presence ofother sized RNA species in certain tissues, indicating the possibil-ity that related genes are expressed in some of the tissues examined(data not shown).
Southern blots containing human genomic DNA digested withEcoRI, BamHI, or HindIII were also probed with the CXCL16chemokine domain probe (Fig. 2B). At moderate stringency, onlyone (BamHI andHindIII) or two (EcoRI) bands were observed,consistent CXCL16 existing as a single copy gene.
CXCL16 is expressed on the surface of leukocyte subsets andfunctional CXCL16 is shed from macrophages
Although sequence analysis suggests that CXCL16 can be pre-sented as a cell surface molecule, we could not detect surfaceexpression of CXCL16 on our 293 transfectants (data not shown).
As Northern blot analysis showed high expression in peripheralblood leukocytes, we asked whether we could detect cell surfaceexpression of CXCL16 on any leukocyte subsets. Staining wasobserved on subsets of CD191 and CD141 peripheral blood leu-kocytes, indicating that APCs including B cells and monocyte/macrophages express this chemokine as a cell surface ligand (Fig.3A). Although there was considerable variation in the percentageof each cell type expressing CXCL16, this general staining patternwas observed in all donors examined (Fig. 3B). These observationsraise the possibility that cell-cell contacts between activated T cellsubsets known to express CXCR6 and APCs expressing CXCL16might result in subset-specific immune responses. In addition toperipheral blood we also examined expression in tonsils and alsofound that subsets of CD191 B cells stained specifically with anti
FIGURE 2. Analysis of CXCL16 RNA expression and genomic orga-nization. A, Membranes with poly(A)1 RNA from the indicated humantissues were hybridized with a chemokine domain CXCL16 probe (upperpanels).b-Actin loading controls are shown in the panels below. The lowerbands seen in muscle tissue lanes correspond to anotherb-actin isoform(Clontech Laboratories technical support, unpublished observations).B,Southern blot using the same probe, hybridized to genomic DNA digestedwith restriction enzymes as indicated.
FIGURE 3. CXCL16 is expressed on the surface of human monocytesand B cells and on tonsillar B cellsA, PBMC from several donors werestained with an anti-CXCL16 mAb (SD7). Expression in one representativedonor was observed in both monocytes (CD141) and B cells (CD191).IgG1 isotype controls are in thebottom panels.B, Percentage of CXCL161
cells varied by donor.C, Expression of CXCL16 on CD19 subsets offreshly isolated tonsil lymphocytes.
5148 EXPRESSION CLONING OF THE STRL33/TYMSTR/BONZO LIGAND
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

CXCL16 mAbs (Fig. 3C), suggesting that in settings of chronicinflammation CXCL16 might participate in cell-cell interactionsas well.
We also asked whether functional chemokine could be shedfrom the surface of the leukocyte subsets expressing surfaceCXCL16. Cultured macrophages were propagated by adherence toplastic, and CXCL16 expression was still observed after severaldays in culture (data not shown). After replacement of medium,supernatants were collected in the presence or absence of inflam-matory mediators and examined for chemotactic activity. No in-crease of cell surface staining was observed after 4- or 24-h incu-bation with LPS or TNF-a (data not shown). After 4 h, minimalchemotactic activity was seen above background, with no discern-able difference between supernatants from unstimulated and stim-ulated cells. However, after 24 h, significant increase in activitywas seen that was moderately increased by LPS and increased by;2-fold in the presence of TNF-a, whereas control orphan GPCR(GusB/CCR11) transfectants fail to chemotax (Fig. 4). These dataindicate that functional chemokine was shed into the medium, sug-gesting that either increases in gene expression or processingmight contribute to observed increase in biological activity.
Activated T lymphocyte subsets express CXCR6 and functionallyrespond to CXCL16
CXCR6 was originally cloned from activated T lymphocytes, andrecent reports have demonstrated the expression of CXCR6 onactivated CD41 and CD81 T lymphocyte subsets along with NKcells (13, 14, 29–31). To look at expression and function of Bonzoin activated T cells, we generated T helper subsets by polarizationin the presence of specific Th1, Th2, and Tr1 cytokines in condi-tions of repeated rounds of cytokine stimulation to mimic settingsof chronic inflammation (21). Upon initial expansion in the pres-ence of Th1, Th2, or Tr1 cytokines, we observed expression ofCCR7 on all subsets, whereas CCR4 (as previously demonstrated)is only expressed on Th2 cells, and CXCL16 was low or undetec-ted on all subsets (Fig. 5A). Upon multiple rounds of cytokinestimulation, levels of CCR7 were reduced in all subsets, suggest-ing a transition to a memory phenotype, whereas CCR4 expressionwas greatly increased on Th2 cells. CXCR6 expression was also
greatly increased on all three subsets, indicating that CXCR6 ex-pression might mark effector cells in settings of chronic inflam-mation (Fig. 5A). Upon examination of chemokine responsiveness,we observed that all three subsets subjected to multiple rounds ofcytokine stimulation respond to RANTES, a ligand for both CCR5and CCR1 (which marks all subsets), whereas only Th2 cells mi-grate in response to the CCR4 ligand macrophage-derived chemo-kine (MDC) (Fig. 5B). All three subsets migrate with similar ef-ficiency to CXCL16, showing that CXCR6 is functional oneffector T cell subsets.
We next asked whether activated cells from a chronically in-flamed tissue also express CXCR6 and respond to CXCL16. Stain-ing of tonsil-derived lymphocytes demonstrates that a subpopula-tion of CD41 T cells expresses CXCR6 (Fig. 6A). Additionally,CXCL16 also mediates chemotaxis of the isolated CD4-positivecells. The observed chemotaxis is selectively blocked by anti-CXCR6 mAb 7F3, which indicates that this is a unique receptorligand interaction (Fig. 6B).FIGURE 4. Chemotaxis induced by CXCL16 secreted from human
macrophages. Supernatants taken from 4- and 24-h cultures of monocyte-derived macrophages induce chemotaxis of CXCR6/L1-2 transfectants.Levels of secreted CXCL16, as measured by chemotaxis, increased after24-h stimulation with LPS and TNF-a, whereas control GusB (CCR11)transfectants fail to chemotax.
FIGURE 5. Expression and function of CXCR6 on effector cell subsets.A, Th1, Th2, and Tr1 effector cells were generated from umbilical bloodCD41 lymphocytes and stained after 1 cycle (primary stimulation) of cy-tokine stimulation with anti CCR7 mAb 7H12, anti CCR4 mAb 1G1, andanti-CXCR6 mAb 7F3.B, After two rounds of cytokine stimulation (sec-ondary stimulation) mAb staining was repeated.C, After repeated roundsof stimulation, only Th2 cells migrate to MDC, whereas all three subsetsshow similar response to both RANTES and CXCL16. Chemokine con-centrations were 100 nM for CXCL16 and 100 ng/ml of MDC andRANTES.
5149The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Recombinant CXCL16 is heavily glycosylated
To further examine chemokine binding and receptor specificity, arecombinant form of CXCL16 was constructed into a fusion pro-tein, which encompassed the entire predicted extracellular domainfused to a C-terminal polyhistidine sequence. Although CXCL16has only one predictedN-linked glycosylation site (NETT at re-sides 168–171), we do predict a heavily glycosylated mucin se-quence that is likely to be rich inO-linked glycans. The expressionconstruct encoding recombinant CXCL16 was transfected into293T cells and purified over successive wheat germ agglutinin andnickel-nitrilotriacetic acid columns. Western blotting with a poly-clonal Ab to the chemokine domain detects a prominent species ofMr of ;40 kDa, which is twice the size of the predicted proteinbackbone ofMr 19 kDa. Treatment of the recombinant protein witha mixture of deglycosidases includingN-glycosidase F, endo-a-N-acetylgalactosaminidase,a2–3,6,8,9-neuraminidase, B1,4-ga-lactosidase, andb-N-acetylglucosaminidase resulted in a signifi-cant increase in mobility and a shift inMr from 40 to 23 kDa (Fig.7). This verifies the prediction that CXCL16, similar to fractalkine,is highly glycosylated (16).
Only CXCR6 functionally responds to CXCL16
Purified CXCL16 was tested in a chemotaxis assay and exhibiteda robust response to CXCR6-L1-2 transfectants, showing a typicalbell-shaped response with peak activity ranging from 10 to 50 nM(Fig. 8A). CXCL16 was then tested in a chemotaxis assay againsta panel of all known chemokine receptors including CCR1-9,CXCR1-5, and CX3CR1. Although known ligands for these re-ceptors exhibited a robust response in this assay (data not shown),recombinant CXCL16 failed to mediate a response to any receptorother than CXCR6 at all concentrations tested (Fig. 8B). Althoughthis experiment does not exclude the possibility that CXCL16
might bind to these (and other nonchemokine-binding GPCRs),these data clearly illustrate that, for known chemokine receptors,only CXCR6 functionally responds to CXCL16. We next askedwhether CXCL16 could specifically signal a rise in intracellularcalcium in the CXCR6-transfected cell line. CXCL16 mediates adose-dependent rise in intracellular calcium that is similar to thatseen for SDF-1a, the ligand for CXCR4, which is expressed on
FIGURE 6. Expression and function ofCXCR6 on tonsil CD4-positive lymphocytesA, CD41 T cells were isolated from fresh ton-sils and stained with anti-CXCR6 mAb 7F3 oran isotype control.B, Tonsil-derived CD41 Tcells chemotax to the indicated concentrationsof CXCL16. Anti-CXCR6 mAb 7F3 reducedmigration to background levels. CXCR6 L1.2transfectants were simultaneously run in thesame assay as a control.
FIGURE 7. Analysis of purified CXCL16 protein. Purified protein (150ng) was resolved on a 4–20% Tris-glycine gel, transferred to nitrocellulose,and probed with a 1/5 dilution of a supernatant containing an anti-CXCL16mAb. Mobility is decreased to a size roughly corresponding to the proteinbackbone after incubation in the presence of a glycosidase mixture includingN-glycosidase F, endo-a-N-acetylgalactosaminidase,a2–3,6,8,9-neuromini-dase,b1,4-galactosidase, andb-N-acetylglucosaminidase.
5150 EXPRESSION CLONING OF THE STRL33/TYMSTR/BONZO LIGAND
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
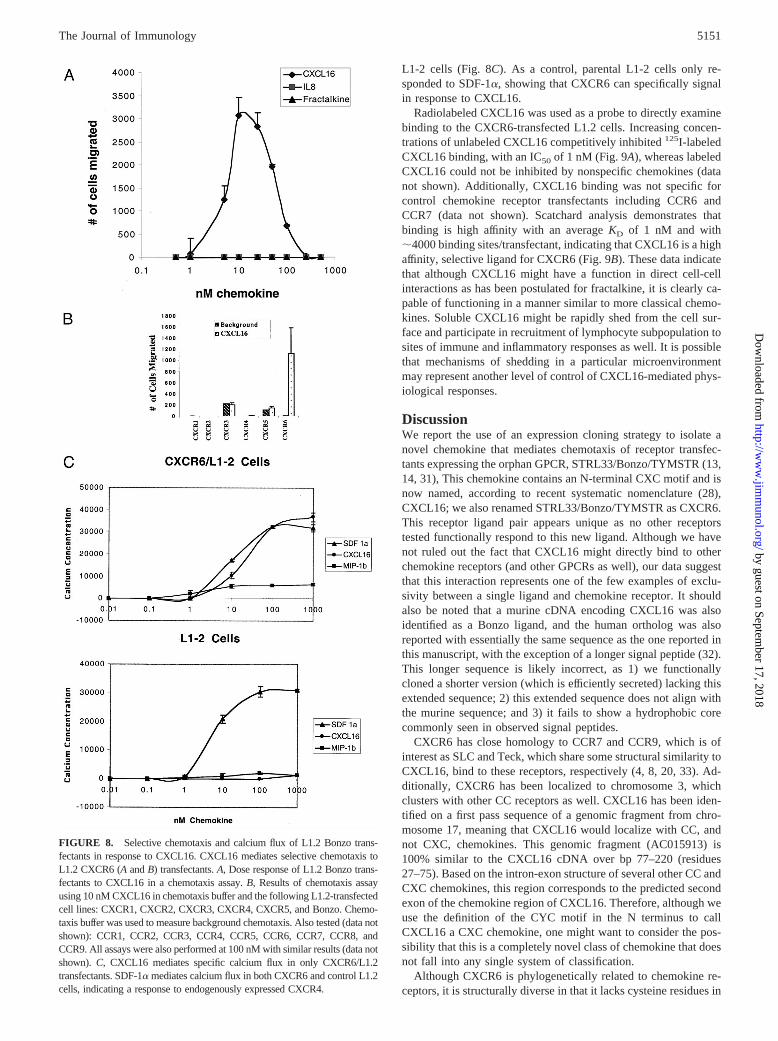
L1-2 cells (Fig. 8C). As a control, parental L1-2 cells only re-sponded to SDF-1a, showing that CXCR6 can specifically signalin response to CXCL16.
Radiolabeled CXCL16 was used as a probe to directly examinebinding to the CXCR6-transfected L1.2 cells. Increasing concen-trations of unlabeled CXCL16 competitively inhibited125I-labeledCXCL16 binding, with an IC50 of 1 nM (Fig. 9A), whereas labeledCXCL16 could not be inhibited by nonspecific chemokines (datanot shown). Additionally, CXCL16 binding was not specific forcontrol chemokine receptor transfectants including CCR6 andCCR7 (data not shown). Scatchard analysis demonstrates thatbinding is high affinity with an averageKD of 1 nM and with;4000 binding sites/transfectant, indicating that CXCL16 is a highaffinity, selective ligand for CXCR6 (Fig. 9B). These data indicatethat although CXCL16 might have a function in direct cell-cellinteractions as has been postulated for fractalkine, it is clearly ca-pable of functioning in a manner similar to more classical chemo-kines. Soluble CXCL16 might be rapidly shed from the cell sur-face and participate in recruitment of lymphocyte subpopulation tosites of immune and inflammatory responses as well. It is possiblethat mechanisms of shedding in a particular microenvironmentmay represent another level of control of CXCL16-mediated phys-iological responses.
DiscussionWe report the use of an expression cloning strategy to isolate anovel chemokine that mediates chemotaxis of receptor transfec-tants expressing the orphan GPCR, STRL33/Bonzo/TYMSTR (13,14, 31), This chemokine contains an N-terminal CXC motif and isnow named, according to recent systematic nomenclature (28),CXCL16; we also renamed STRL33/Bonzo/TYMSTR as CXCR6.This receptor ligand pair appears unique as no other receptorstested functionally respond to this new ligand. Although we havenot ruled out the fact that CXCL16 might directly bind to otherchemokine receptors (and other GPCRs as well), our data suggestthat this interaction represents one of the few examples of exclu-sivity between a single ligand and chemokine receptor. It shouldalso be noted that a murine cDNA encoding CXCL16 was alsoidentified as a Bonzo ligand, and the human ortholog was alsoreported with essentially the same sequence as the one reported inthis manuscript, with the exception of a longer signal peptide (32).This longer sequence is likely incorrect, as 1) we functionallycloned a shorter version (which is efficiently secreted) lacking thisextended sequence; 2) this extended sequence does not align withthe murine sequence; and 3) it fails to show a hydrophobic corecommonly seen in observed signal peptides.
CXCR6 has close homology to CCR7 and CCR9, which is ofinterest as SLC and Teck, which share some structural similarity toCXCL16, bind to these receptors, respectively (4, 8, 20, 33). Ad-ditionally, CXCR6 has been localized to chromosome 3, whichclusters with other CC receptors as well. CXCL16 has been iden-tified on a first pass sequence of a genomic fragment from chro-mosome 17, meaning that CXCL16 would localize with CC, andnot CXC, chemokines. This genomic fragment (AC015913) is100% similar to the CXCL16 cDNA over bp 77–220 (residues27–75). Based on the intron-exon structure of several other CC andCXC chemokines, this region corresponds to the predicted secondexon of the chemokine region of CXCL16. Therefore, although weuse the definition of the CYC motif in the N terminus to callCXCL16 a CXC chemokine, one might want to consider the pos-sibility that this is a completely novel class of chemokine that doesnot fall into any single system of classification.
Although CXCR6 is phylogenetically related to chemokine re-ceptors, it is structurally diverse in that it lacks cysteine residues in
FIGURE 8. Selective chemotaxis and calcium flux of L1.2 Bonzo trans-fectants in response to CXCL16. CXCL16 mediates selective chemotaxis toL1.2 CXCR6 (AandB) transfectants.A, Dose response of L1.2 Bonzo trans-fectants to CXCL16 in a chemotaxis assay.B, Results of chemotaxis assayusing 10 nM CXCL16 in chemotaxis buffer and the following L1.2-transfectedcell lines: CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, and Bonzo. Chemo-taxis buffer was used to measure background chemotaxis. Also tested (data notshown): CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, andCCR9. All assays were also performed at 100 nM with similar results (data notshown).C, CXCL16 mediates specific calcium flux in only CXCR6/L1.2transfectants. SDF-1a mediates calcium flux in both CXCR6 and control L1.2cells, indicating a response to endogenously expressed CXCR4.
5151The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

the N terminus and the third extracellular loop that are conservedin all other receptors to date (14). One might postulate thatCXCR6, due to the loss of a critical disulfide bond, has a uniquemembrane topology and therefore might bind to a structurallyunique chemokine. Interestingly, a shorter synthetic versionCXCL16, truncated at S95 before the fifth and sixth cysteine res-idues, lacks the potency of the entire extracellular region and ap-pears to be unstable at higher temperatures or pH (data not shown).In contrast, other chemokines that have been similarly truncated,such as Teck and fractalkine, retain activity (D. Soler, unpublisheddata), suggesting the possibility that the extra cysteine residues inthe chemokine domain are critical for overall structure and bindingto this novel receptor. We have shown by HPLC analysis thatsynthetic truncated CXCL16 shows deficiencies in efficient folding(data not shown), which might require these residues as well. Al-though these extra cysteine residues might be critical for efficientfolding, it is also possible that, due to the unique features ofCXCR6, this receptor ligand interaction might be completelyunique and require residues that have not been required for otherchemokines studied thus far.
CXCL16 is the second example of a chemokine that has thesequence of a transmembrane protein and would be predicted to beexpressed on the cell surface. Interestingly, our attempts to showexpression on the 293 cells (and Chinese hamster ovary cell trans-fectants) failed to demonstrate cell surface expression of CXCL16and likely explains our success in the identification of a solubleactive form derived from a membrane-bound chemokine. This sug-gests that in certain settings, the chemokine can be efficiently shedinto the surrounding medium; accordingly, we did observe func-tional activity from supernatants derived from cultured macro-phages. This activity was increased in the presence of LPS andTNF-a, indicating that CXCL16 may be rapidly shed from the cellsurface and participate in recruitment of lymphocyte subpopula-tion to sites of both immune and inflammatory responses. It ispossible that mechanisms of shedding in a particular microenvi-ronment may represent a level of control of CXCL16-mediatedphysiological responses.
Additionally, we observed cell surface expression on subsets ofperipheral blood leukocytes including CD191 B cells and CD141
monocytes. This expression pattern suggests that novel interac-
tions might occur between APCs and CXCR6 expressed on acti-vated T cell subsets, thus raising the possibility that immune re-sponses may be facilitated by direct interactions mediated betweencell types expressing this receptor ligand pair. This expression pat-tern also raises the possibility that other CD14-derived cells, suchas monocyte-derived dendritic cells, might express this ligand aswell. Along these lines, we have observed expression of CXCL16in subsets (with a similar staining pattern to what we observe inmacrophages) of monocyte-derived dendritic cells, cultured in thepresence of IL-4 and GM-CSF (data not shown). This, to someextent, parallels observations showing expression of murineCXCL16 in spleen- and lymph node-derived dendritic cells andindicates similarity in observations between these studies (32).Conversely, expression of murine CXCL16 in B cells was notreported in that study although this does not preclude the possi-bility that expression might be seen there as well, as it may haveyet to be examined.
Expression of the other known membrane-bound chemokine,CX3CL1 (fractalkine/neurotactin), has also been shown on den-dritic cells in epidermis and lymphoid tissues (34, 35). CX3CL1 isalso chemotactic for activated T cells and may play a role in re-cruitment and adhesion of T cells to secondary lymphoid tissuesand directly interact with T cells in immune responses. Therefore,CXCL16 may represent a second member of the chemokine familythat can play a dual role in recruitment and retention of lympho-cyte subsets. Other chemokines, such as MDC, have been shownto be expressed by dendritic cells and play a role in attractingactivated T cell subsets (36). MDC and fractalkine both map tochromosome 16q13 (35). Although CXCL16 might share somefunctional relationship with these chemokines, it appears to resideon chromosome 17 and, therefore, does not map to a chromosomeencoding either these chemokines or CXC chemokines as well,further illustrating the unique nature of this new ligand.
We have observed CXCL16 expression on tonsil-derivedCD191s and CXCR6 expression on tonsil CD41s. This observa-tion raises the possibility that, within secondary lymphoid tissue, Bcells expressing CXCL16 might be activated by a T cell-dependentprocess. MDC and fractalkine have also been shown to be ex-pressed by activated B cells as well (36, 37), although expressionof fractalkine was observed only at the RNA level. This is the first
FIGURE 9. Binding of CXCL16 to the CXCR6 chemokine receptor in L1.2-transfected cells.A, 125I-CXCL16 is specific for L1.2 CXCR6 transfectants.Panel shows binding activity of L1.2 CXCR6 transfectants with 1 nM125I-CXCL16 with or without increasing concentrations of cold competitor. Thecalculated value of 50% inhibition is 1 nM CXCL16.B, Scatchard analysis of binding data. Results shown are the average of two experiments. ThecalculatedKd is 1 nM.
5152 EXPRESSION CLONING OF THE STRL33/TYMSTR/BONZO LIGAND
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

report of surface expression of a chemokine on a B cell suggestingthat binding between this surface-encoded chemokine and its cog-nate receptor on T cells might contribute to T cell activation. TheseB cell/T cell interactions may occur after CXCR61 T cell/dendriticcell interactions have been initiated, and it is possible thatCXCL16 may play a role in both of these steps. As the cytoplasmictail of CXCL16 has a number of potential phosphorylation sites, itwill also be of interest to see whether interactions with CXCR6result in signals transmitted through this chemokine, thus mediat-ing signals in APCs in a novel manner.
Although chemokines have been classified on the basis of thestructure surrounding the N-terminal cysteine residues, anotherclassification has emerged as of late, subdividing chemokines intoeither homeostatic (lymphoid) or inflammatory, based on their siteof production and cell types that respond to them. Certain chemo-kines are produced in lymphoid tissues such as BCA-1 (in stromalcells in B cell follicles) and SLC (in high endothelial venules andby stromal cells in T cells) (26, 27, 38). Others are more known toattract activated leukocytes and are induced by inflammatory stim-uli including IL-8, RANTES, monocyte chemoattractant protein-1,MIP-1a, and eotaxin (among others) (39–42), which are allknown to be more highly expressed at sites of inflammation. Al-though many chemokines fall into these categories, some do notclearly fit either paradigm. MDC, in addition to having a role insecondary lymphoid tissue, has also been shown to be produced ininflamed lungs (43, 44). CXL16 might prove to be a dual-functionchemokine as well, as its expression is shown here to be in sec-ondary lymphoid tissues and in addition in nonlymphoid tissues,most notably in liver and lung tissues. Additionally, we show thatactivated T cell subsets including chronic Th1, Th2, Tr1, andCD41 T cells derived from tonsil functionally respond to this che-mokine. The observations of surface expression of CXCL16 onAPCs suggest a potential homeostatic role in lymphoid tissue,whereas the functional interaction with polarized effector cells sug-gests a role in chronic inflammation as well.
As the sequencing of the genome becomes complete, the con-cept of “novel sequence identification” is rapidly becoming a thingof the past. There is now a new challenge to assign function ofthousands of expressed sequenced clones where classification viaconventional means is not possible. CXCL16 was represented inthe database as an expressed sequence tag for 5 years, but due tolack of homology could not be identified by searching with relatedsequences. Although newer searching algorithms that use second-ary structure and pattern predictions might solve some of this di-lemma, the cloning of this chemokine raises the possibility thatseveral other functionally relevant proteins of this class might alsobe identified in this manner. The isolation of this structurallyunique chemokine and demonstration of binding to a novel che-mokine receptor (the first to lack cysteine residues previously con-sidered essential for function) breaks all known paradigms forknown chemokine receptor/ligand interactions. Additionally, theobservation of both surface expression of CXCL16 on APCs andshedding of active soluble material raises the possibility that thischemokine might play dual roles in inflammation and homeostasis.
Note added in proof. While in revision it came to our attentionthat in an attempt to use expression cloning from monocytes toisolate novel receptors binding to OxLDL, a cDNA encoding areceptor termed SR-PSOX (scavenger receptor that binds phos-phatidylserine and oxidized lipoprotein) was identified. ThiscDNA turns out to encode CXCL16, further illustrating the pos-sibility of additional functional roles of this protein.
AcknowledgmentsWe thank Nasim Kassam for technical assistance in generation of hybrid-omas and Jose-Carlos Gutierrez-Ramos and Craig Gerard for critical read-ing of this manuscript. We also thank Keith Robison for initial identifica-tion of the identity of CXCL16 to SR-PSOX (seeNote added in proof).
References1. Baggiolini, M., B. Dewald, and B. Moser. 1997. Human chemokines: an update.
Annu. Rev. Immunol. 15:675.2. Baggiolini, M. 1998. Chemokines and leukocyte traffic.Nature 392:565.3. Ward, S. G., K. Bacon, and J. Westwick. 1998. Chemokines and T lymphocytes:
more than an attraction.Immunity 9:1.4. Gunn, M. D., K. Tangemann, C. Tam, J. G. Cyster, S. D. Rosen, and
L. T. Williams. 1998. A chemokine expressed in lymphoid high endothelialvenules promotes the adhesion and chemotaxis of naive T lymphocytes.Proc.Natl. Acad. Sci. USA 95:258.
5. Imai, T., M. Baba, M. Nishimura, M. Kakizaki, S. Takagi, and O. Yoshie. 1997.The T cell-directed CC chemokine TARC is a highly specific biological ligandfor CC chemokine receptor 4.J. Biol. Chem. 272:15036.
6. Jarmin, D. I., M. Rits, D. Bota, N. P. Gerard, G. J. Graham, I. Clark-Lewis, andC. Gerard. 2000. Identification of the orphan G-protein-coupled receptor 2 asCCR10, a specific receptor for the chemokine ESkine.J. Immunol. 164:3460.
7. Yoshida, T., D. Izawa, T. Nakayama, K. Nakahara, M. Kakizaki, T. Imai,R. Suzuki, M. Miyasaka, and O. Yoshie. 1999. Molecular cloning of mXCR1, themurine SCM-1/lymphotactin receptor.FEBS Lett. 458:37.
8. Yoshida, R., T. Imai, K. Hieshima, J. Kusuda, M. Baba, M. Kitaura,M. Nishimura, M. Kakizaki, H. Nomiyama, and O. Yoshie. 1997. Molecularcloning of a novel human CC chemokine EBI1-ligand chemokine that is a spe-cific functional ligand for EBI1, CCR7.J. Biol. Chem. 272:13803.
9. Campbell, J. J., and E. C. Butcher. 2000. Chemokines in tissue-specific andmicroenvironment-specific lymphocyte homing.Curr. Opin. Immunol. 12:336.
10. Gunn, M. D., V. N. Ngo, K. M. Ansel, E. H. Ekland, J. G. Cyster, andL. T. Williams. 1998. A B-cell-homing chemokine made in lymphoid folliclesactivates Burkitt’s lymphoma receptor-1.Nature 391:799.
11. Foxman, E. F., J. J. Campbell, and E. C. Butcher. 1997. Multistep navigation andthe combinatorial control of leukocyte chemotaxis.J. Cell Biol. 139:1349.
12. Heesen, M., M. A. Berman, A. Charest, D. Housman, C. Gerard, and M. E. Dorf.1998. Cloning and chromosomal mapping of an orphan chemokine receptor:mouse RDC1.Immunogenetics 47:364.
13. Deng, H. K., D. Unutmaz, V. N. KewalRamani, and D. R. Littman. 1997. Ex-pression cloning of new receptors used by simian and human immunodeficiencyviruses.Nature 388:296.
14. Liao, F., G. Alkhatib, K. W. Peden, G. Sharma, E. A. Berger, and J. M. Farber.1997. STRL33, a novel chemokine receptor-like protein, functions as a fusioncofactor for both macrophage-tropic and T cell line-tropic HIV-1.J. Exp. Med.185:2015.
15. Loetscher, P., M. Seitz, M. Baggiolini, and B. Moser. 1996. Interleukin-2 regu-lates CC chemokine receptor expression and chemotactic responsiveness in Tlymphocytes.J. Exp. Med. 184:569.
16. Bazan, J. F., K. B. Bacon, G. Hardiman, W. Wang, K. Soo, D. Rossi,D. R. Greaves, A. Zlotnik, and T. J. Schall. 1997. A new class of membrane-bound chemokine with a CX3C motif.Nature 385:640.
17. Pan, Y., C. Lloyd, H. Zhou, S. Dolich, J. Deeds, J. A. Gonzalo, J. Vath,M. Gosselin, J. Ma, B. Dussault, et al. 1997. Neurotactin, a membrane-anchoredchemokine upregulated in brain inflammation. [Published erratum appears in1997 Nature 389:100.]Nature 387:611.
18. Wu, L., N. Gerard, R. Wyatt, H. Choe, C. Parolin, N. Ruffing, A. Borsetti,A. A. Cardoso, E. Desjardin, W. Newman, et al. 1996. CD4-induced interactionof primary HIV-1 gp120 glycoproteins with the chemokine receptor CCR-5.Na-ture 384:179.
19. Ponath, P. D., S. Qin, T. W. Post, J. Wang, L. Wu, N. P. Gerard, W. Newman,C. Gerard, and C. R. Mackay. 1996. Molecular cloning and characterization of ahuman eotaxin receptor expressed selectively on eosinophils.J. Exp. Med. 183:2437.
20. Zabel, B. A., W. W. Agace, J. J. Campbell, H. M. Heath, D. Parent, A. I. Roberts,E. C. Ebert, N. Kassam, S. Qin, M. Zovko, et al. 1999. Human G protein-coupledreceptor GPR-9–6/CC chemokine receptor 9 is selectively expressed on intesti-nal homing T lymphocytes, mucosal lymphocytes, and thymocytes and is re-quired for thymus-expressed chemokine-mediated chemotaxis.J. Exp. Med. 190:1241.
21. Murphy, E., K. Shibuya, N. Hosken, P. Openshaw, V. Maino, K. Davis,K. Murphy, and A. O’Garra. 1996. Reversibility of T helper 1 and 2 populationsis lost after long-term stimulation.J. Exp. Med. 183:901.
22. Topham, P. S., V. Csizmadia, D. Soler, D. Hines, C. J. Gerard, D. J. Salant, andW. W. Hancock. 1999. Lack of chemokine receptor CCR1 enhances Th1 re-sponses and glomerular injury during nephrotoxic nephritis.J. Clin. Invest. 104:1549.
23. Shyjan, A. M., M. Bertagnolli, C. J. Kenney, and M. J. Briskin. 1996. Humanmucosal addressin cell adhesion molecule-1 (MAdCAM-1) demonstrates struc-tural and functional similarities to thea4b7-integrin binding domains of murineMAdCAM-1, but extreme divergence of mucin-like sequences.J. Immunol. 156:2851.
24. Kawahara, R. S., and T. F. Deuel. 1989. Platelet-derived growth factor-induciblegene JE is a member of a family of small inducible genes related to platelet factor4. J. Biol. Chem. 264:679.
5153The Journal of Immunology
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

25. Baird, J. W., R. J. B. Nibbs, M. Komai-Koma, J. A. Connolly, K. Ottersbach,I. Clark-Lewis, F. L. Liew, and G. J. Graham. 1999. ESkine, a novelb-chemo-kine is differentially spliced to produce secretable and nuclear targeted isoforms.J. Biol. Chem.
26. Hedrick, J. A., and A. Zlotnik. 1997. Identification and characterization of a novelb chemokine containing six conserved cysteines.J. Immunol. 159:1589.
27. Hromas, R., C. H. Kim, M. Klemsz, M. Krathwohl, K. Fife, S. Cooper,C. Schnizlein-Bick, and H. E. Broxmeyer. 1997. Isolation and characterization ofExodus-2, a novel C-C chemokine with a unique 37-amino acid carboxyl-termi-nal extension.J. Immunol. 159:2554.
28. Zlotnik, A., and O. Yoshie. 2000. Chemokines: a new classification system andtheir role in immunity.Immunity 12:121.
29. Sharron, M., S. Pohlmann, K. Price, E. Lolis, M. Tsang, F. Kirchhoff,R. W. Doms, and B. Lee. 2000. Expression and coreceptor activity of STRL33/Bonzo on primary peripheral blood lymphocytes.Blood 96:41.
30. Unutmaz, D., W. Xiang, M. J. Sunshine, J. Campbell, E. Butcher, andD. R. Littman. 2000. The primate lentiviral receptor Bonzo/STRL33 is coordi-nately regulated with CCR5 and its expression pattern is conserved betweenhuman and mouse.J. Immunol. 165:3284.
31. Loetscher, M., A. Amara, E. Oberlin, N. Brass, D. Legler, P. Loetscher, M.D’Apuzzo, E. Meese, D. Rousset, J. L. Virelizier, et al. 1997. TYMSTR, a pu-tative chemokine receptor selectively expressed in activated T cells, exhibitsHIV-1 coreceptor function.Curr. Biol. 7:652.
32. Matloubian, M., A. David, S. Engel, J. E. Ryan, and J. G. Cyster. 2000. Atransmembrane CXC chemokine is a ligand for HIV-coreceptor Bonzo.Nat. Im-mun. 1:298.
33. Zaballos, A., J. Gutierrez, R. Varona, C. Ardavin, and G. Marquez. 1999. Cuttingedge: identification of the orphan chemokine receptor GPR-9-6 as CCR9, thereceptor for the chemokine TECK.J. Immunol. 162:5671.
34. Papadopoulos, E. J., C. Sassetti, H. Saeki, N. Yamada, T. Kawamura,D. J. Fitzhugh, M. A. Saraf, T. Schall, A. Blauvelt, S. D. Rosen, and S. T. Hwang.1999. Fractalkine, a CX3C chemokine, is expressed by dendritic cells and isup-regulated upon dendritic cell maturation.Eur. J. Immunol. 29:2551.
35. Kanazawa, N., T. Nakamura, K. Tashiro, M. Muramatsu, K. Morita, K. Yoneda,K. Inaba, S. Imamura, and T. Honjo. 1999. Fractalkine and macrophage-derivedchemokine: T cell-attracting chemokines expressed in T cell area dendritic cells.Eur. J. Immunol. 29:1925.
36. Schaniel, C., E. Pardali, F. Sallusto, M. Speletas, C. Ruedl, T. Shimizu, T. Seidl,J. Andersson, F. Melchers, A. G. Rolink, and P. Sideras. 1998. Activated murineB lymphocytes and dendritic cells produce a novel CC chemokine which actsselectively on activated T cells.J. Exp. Med. 188:451.
37. Foussat, A., A. Coulomb-L’Hermine, J. Gosling, R. Krzysiek,I. Durand-Gasselin, T. Schall, A. Balian, Y. Richard, P. Galanaud, and D. Emilie.2000. Fractalkine receptor expression by T lymphocyte subpopulations and invivo production of fractalkine in human.Eur. J. Immunol. 30:87.
38. Nagira, M., T. Imai, K. Hieshima, J. Kusuda, M. Ridanpaa, S. Takagi,M. Nishimura, M. Kakizaki, H. Nomiyama, and O. Yoshie. 1997. Molecularcloning of a novel human CC chemokine secondary lymphoid-tissue chemokinethat is a potent chemoattractant for lymphocytes and mapped to chromosome9p13.J. Biol. Chem. 272:19518.
39. Bazzoni, F., M. A. Cassatella, F. Rossi, M. Ceska, B. Dewald, and M. Baggiolini.1991. Phagocytosing neutrophils produce and release high amounts of the neu-trophil-activating peptide 1/interleukin 8.J. Exp. Med. 173:771.
40. Ponath, P. D., S. Qin, D. J. Ringler, I. Clark-Lewis, J. Wang, N. Kassam,H. Smith, X. Shi, J.-A. Gonzalo, W. Newman, et al. 1996. Cloning of the humaneosinophil chemoattractant, eotaxin: expression, receptor binding and functionalproperties provide a mechanism for the selective recruitment of eosinophils.J. Clin. Invest. 97:604.
41. Schall, T. J., K. Bacon, K. J. Toy, and D. V. Goeddel. 1990. Selective attractionof monocytes and T lymphocytes of the memory phenotype by cytokine RAN-TES.Nature 347:669.
42. Sherry, B., P. Tekamp-Olson, C. Gallegos, D. Bauer, G. Davatelis, S. D. Wolpe,F. Masiarz, D. Coit, and A. Cerami. 1988. Resolution of the two components ofmacrophage inflammatory protein 1, and cloning and characterization of one ofthose components, macrophage inflammatory protein 1b. J. Exp. Med. 168:2251.
43. Godiska, R., D. Chantry, C. J. Raport, S. Sozzani, P. Allavena, D. Leviten,A. Mantovani, and P. W. Gray. 1997. Human macrophage-derived chemokine(MDC), a novel chemoattractant for monocytes, monocyte-derived dendriticcells, and natural killer cells.J. Exp. Med. 185:1595.
44. Gonzalo, J. A., Y. Pan, C. M. Lloyd, G. Q. Jia, G. Yu, B. Dussault, C. A. Powers,A. E. Proudfoot, A. J. Coyle, D. Gearing, and J. C. Gutierrez-Ramos. 1999.Mouse monocyte-derived chemokine is involved in airway hyperreactivity andlung inflammation.J. Immunol. 163:403.
5154 EXPRESSION CLONING OF THE STRL33/TYMSTR/BONZO LIGAND
by guest on September 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from


















