
Experimentele en numerieke studie van de sedimentatie van...
113
Faculteit Toegepaste Wetenschappen Vakgroep Civiele Techniek Laboratorium voor Hydraulica Directeur: Prof. Dr. Ir. R. Verhoeven Experimentele en numerieke studie van de sedimentatie van bloed in centrifuges Ben Verhoeven Promotor: Prof. Dr. Ir. P. Verdonck Begeleiders: Dr. Ir. S. Eloot Dr. F. De Somer Afstudeerwerk ingediend tot het behalen van de graad van burgerlijk scheikundig ingenieur Academiejaar 2005–2006
Transcript of Experimentele en numerieke studie van de sedimentatie van...

Faculteit Toegepaste Wetenschappen
Vakgroep Civiele Techniek
Laboratorium voor Hydraulica
Directeur: Prof. Dr. Ir. R. Verhoeven
Experimentele en numerieke studie
van de sedimentatie
van bloed in centrifuges
Ben Verhoeven
Promotor: Prof. Dr. Ir. P. Verdonck
Begeleiders: Dr. Ir. S. Eloot
Dr. F. De Somer
Afstudeerwerk ingediend tot het behalen van de graad van
burgerlijk scheikundig ingenieur
Academiejaar 2005–2006

De auteur geeft de toelating om dit afstudeerwerk beschikbaar te stellen voor consultatie
en om delen ervan te kopieren voor persoonlijk gebruik.
Elk ander gebruik valt onder de beperkingen van het auteursrecht, in het bijzonder met
betrekking tot de verplichting de bron te vermelden bij het aanhalen van resultaten uit dit
afstudeerwerk.
Zevergem, 16 juni 2006
Ben Verhoeven
i

Faculteit Toegepaste WetenschappenVakgroep Civiele Techniek
Laboratorium voor HydraulicaDirecteur: Prof. Dr. Ir. R. Verhoeven
Academiejaar 2005 – 2006
Experimentele en numerieke studie van de
sedimentatie van bloed in centrifuges
Ben Verhoeven
Promotor: Prof. Dr. Ir. P. Verdonck
Begeleiders: Dr. Ir. S. Eloot
Dr. F. De Somer
Overzicht
In dit afstudeerwerk worden de experimentele en numerieke resultaten van de sedimentatie vanbloedcellen in centrifuges beschreven. Het bloed wordt gescheiden in zijn verschillende bestand-delen: plasma, witte- en rode bloedcellen en de bloedplaatjes.
In het eerste hoofdstuk wordt de anatomie en de functie van het bloed besproken, gevolgd doorhet nut van de bloedproducten en een beschrijving van de verschillende types centrifuges.
Het tweede hoofdstuk handelt over belangrijke artikels omtrent dit onderwerp. Zoals enkelerecente uitgevoerde simulaties met Computational Fluid Dynamics.
In hoofdstuk 3 en hoofdstuk 4 wordt het experimenteel onderzoek van de sedimentatie van derode bloedcellen met runderbloed en humaanbloed beschreven.
Het volgende hoofdstuk maakt een vergelijking van de bekomen resultaten. Er wordt een verbandbeschreven tussen het resultaat van beide bloedsoorten. De resultaten verkregen met makkelijkverkrijgbaar runderbloed kunnen nu omgerekend worden naar resultaten met humaanbloed.
In hoofdstuk 6 wordt een numeriek model beschreven dat gevalideerd werd met behulp van de
resultaten van het experimenteel onderzoek.
Trefwoorden: bloedscheiding, sedimentatie, centrifugeren, runderbloed, humaanbloed, rou-
leaux, Fluent, simulatie, CFD.
ii

Voorwoord
Een afstudeerwerk is een heuse opdracht voor elke student. Een jaar lang wordt er intensief
gewerkt om een degelijk resultaat te bekomen. De weg naar dit resultaat kende goede
en slechte momenten. Extreem veel bloed en een beetje zweet en tranen heeft het mij
gekost om het einde te bereiken. De drive was gelukkig steeds aanwezig om dit thema te
onderzoeken. De mogelijkheid om op je eigen manier de zaak aan te pakken en problemen
op te lossen bleven mij motiveren.
Daarom wil ik Prof. Dr. Ir. P. Verdonck bedanken voor de mogelijkheid die hij me bood
om dit afstudeerwerk in zijn groep te verwezenlijken. De leuke activiteiten die beleefd
werden zoals na de halfwegpresentatie worden enorm geapprecieerd.
Mijn begeleidster Dr. Ir. S. Eloot wil ik hartelijk danken voor de deskundige begeleiding
die ze mij gaf. Ze was altijd paraat om mijn teksten met een kritisch oog te herlezen.
Bedankt Sunny!
De figuurlijke vader van dit onderzoek, Dr. F. De Somer, wens ik te bedanken voor zijn
deskundige uitleg en voor het opstarten van deze thesis. In het bijzonder wil ik nog Ir. G.
Mareels bedanken voor zijn hulp met de computers.
Verder wens ik nog mijn moeder te bedanken voor de vele computerhulp die ze me bood
tijdens het typwerk, natuurlijk Lore voor al het naleeswerk dat ze leverde en mijn vrienden
waarbij ik steeds terecht kon tijdens de iets hardere momenten.
iii

Inhoudsopgave
1 Het aferese proces van bloed 1
1 Anatomie en functie van het bloed . . . . . . . . . . . . . . . . . . . . . . 1
1.1 Bloedplasma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
1.2 Rode bloedcellen . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
1.3 Witte bloedcellen . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
1.4 Bloedplaatjes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
1.5 Productie en afbraak van bloedcellen . . . . . . . . . . . . . . . . . 7
1.6 Samenvatting . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
2 Toepassingen van de centrifuges . . . . . . . . . . . . . . . . . . . . . . . . 8
2.1 Therapeutische behandelingen . . . . . . . . . . . . . . . . . . . . . 8
2.2 Peri-operatieve transfusie . . . . . . . . . . . . . . . . . . . . . . . . 9
2.3 Toepassing in bloedtransfusiecentra . . . . . . . . . . . . . . . . . . 9
3 Types centrifuges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
3.1 Dideco . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
3.2 Volbloed verwerking . . . . . . . . . . . . . . . . . . . . . . . . . . 13
3.3 Aferese-apparatuur . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2 Modellen voor het simuleren van de sedimentatie van bloed 17
1 Sedimentatietheorie van het bloed . . . . . . . . . . . . . . . . . . . . . . . 17
1.1 Wet van Stokes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
1.2 Invoering van een vormfactor . . . . . . . . . . . . . . . . . . . . . 19
1.3 Correctiefactor voor de onderlinge impacteffecten . . . . . . . . . . 20
vi

INHOUDSOPGAVE vii
1.4 Invloed van de rouleaux vorming . . . . . . . . . . . . . . . . . . . 21
1.5 Besluit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
2 Verdere studie van de deeltjesinteractie bij de sedimentatie van bloedcellen 24
2.1 Invoering van een effectieve porositeit . . . . . . . . . . . . . . . . . 24
2.2 Model van Masliyah . . . . . . . . . . . . . . . . . . . . . . . . . . 25
2.3 Model van Barnea en Mizrahi . . . . . . . . . . . . . . . . . . . . . 25
2.4 Semi-empirisch model . . . . . . . . . . . . . . . . . . . . . . . . . 25
2.5 Resultaten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
2.6 Besluit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
3 Simulatie van het sedimentatieproces . . . . . . . . . . . . . . . . . . . . . 28
3.1 Het scheidingsproces . . . . . . . . . . . . . . . . . . . . . . . . . . 28
3.2 De transportverschijnselen . . . . . . . . . . . . . . . . . . . . . . . 28
3.3 Het simulatiemodel . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
3.4 Besluit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
4 Simulatie met behulp van Computational Fluid Dynamics . . . . . . . . . 32
4.1 Het scheidingsproces . . . . . . . . . . . . . . . . . . . . . . . . . . 32
4.2 Theorie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
4.3 Berekeningsmethode . . . . . . . . . . . . . . . . . . . . . . . . . . 37
4.4 Resultaten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
4.5 Besluit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
3 Experimentele studie met runderbloed 47
1 De metingen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
1.1 Voorbereiding van het bloed . . . . . . . . . . . . . . . . . . . . . . 47
1.2 De opstelling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
1.3 Meetcondities . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
1.4 Metingen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
2 De resultaten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51

INHOUDSOPGAVE viii
2.1 Tabel . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
2.2 Bespreking van de HemoCue resultaten . . . . . . . . . . . . . . . . 52
2.3 Bespreking van de volumes . . . . . . . . . . . . . . . . . . . . . . . 53
2.4 Bespreking van de hematocrieten . . . . . . . . . . . . . . . . . . . 55
4 Experimentele studie met humaanbloed 58
1 Studie van de data uit het Universitair Ziekenhuis Gent . . . . . . . . . . . 58
1.1 De parameters en metingen . . . . . . . . . . . . . . . . . . . . . . 58
1.2 Bespreking van de resultaten . . . . . . . . . . . . . . . . . . . . . . 61
1.3 Verband met runderbloed . . . . . . . . . . . . . . . . . . . . . . . 63
2 Experiment met humaanbloed . . . . . . . . . . . . . . . . . . . . . . . . . 65
2.1 De opstelling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
2.2 De parameters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
2.3 De resultaten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66
3 Besluit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
5 Verband tussen humaan en runderbloed 71
1 Analyse van de data van runderbloed . . . . . . . . . . . . . . . . . . . . . 71
2 Analyse van de data van humaanbloed . . . . . . . . . . . . . . . . . . . . 74
3 Verband tussen de uitlaathematocrieten van runderbloed en humaanbloed . 76
4 Besluit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77
6 Numerieke studie van de sedimentatie van bloedcellen 78
1 Probleem beschrijving . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 78
2 Het geometrisch model in Gambit . . . . . . . . . . . . . . . . . . . . . . . 79
2.1 Geometrie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
2.2 Zones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
2.3 Mesh . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82
3 Het numeriek model in Fluent . . . . . . . . . . . . . . . . . . . . . . . . . 82

INHOUDSOPGAVE ix
3.1 Inladen van de geometrie . . . . . . . . . . . . . . . . . . . . . . . . 82
3.2 Modelkeuze . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
3.3 Het Euler model . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84
3.4 Materialen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
3.5 Fasen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
3.6 De Randvoorwaarden . . . . . . . . . . . . . . . . . . . . . . . . . . 89
4 De simulaties . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
5 De resultaten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 93
5.1 Simulaties met runderbloed . . . . . . . . . . . . . . . . . . . . . . 93
5.2 Simulaties met humaanbloed . . . . . . . . . . . . . . . . . . . . . . 95
6 Tekortkomingen van het numeriek model . . . . . . . . . . . . . . . . . . . 96
6.1 Vorm van de bloedcellen . . . . . . . . . . . . . . . . . . . . . . . . 97
6.2 Botsingen tussen de bloedcellen . . . . . . . . . . . . . . . . . . . . 98
7 Toekomst perspectieven . . . . . . . . . . . . . . . . . . . . . . . . . . . . 98
8 Besluit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99
7 Algemeen besluit 100
1 Experimenteel onderzoek met runderbloed . . . . . . . . . . . . . . . . . . 100
2 Experimenteel onderzoek met humaanbloed . . . . . . . . . . . . . . . . . 100
3 Verband tussen humaan en runderbloed . . . . . . . . . . . . . . . . . . . . 101
4 Numeriek model . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 101

Symbolenlijst
Latijnse symbolen
a grote straal van een bloedcel m
b kleine straal of de dikte van een bloedcel m
C totale volumetrische deeltjesconcentratie mld/mlf
CN concentratie aan deeltjes van type N mol/m3
cin concentratie aan de inlaat mld/mlf
cuit concentratie aan de uitlaat mld/mlf
CD drag coefficient kg/s
D diffusiecoefficient m2/s
dε dikte van het lege volume rond een cel m
dgem gemiddelde deeltjes diameter m
dj afstand tussen de bloedcel en het naburig mesh element j m
d deeltjesdiameter m
eii restitutiecoefficient (-)~Fr resulterende kracht N~F uitwendige kracht N~FB opstuwingskracht N~FD wrijvingskracht N
g valversnelling m/s2
g0,ii radiale distributiefunctie (-)
Hct hematocriet of volumefractie rode bloedcellen (-)
I eenheidstensor (-)
I2D tweede invariant van de spanningstensor τi (-)
Kji momentum uitwisselingscoefficient (-)
k1 Einstein vormfactor (-)
k2 vormfactor van een botsingsdoublet (-)
mi massa van het deeltje kg
mf massa van het verplaatste fluıdum kg
x

N aantal fasen (-)
p totale druk Pa
pi partieeldruk van vaste fase i Pa
Q hydrodynamische interactieconstante (-)
q vulsnelheid ml/min
r straal van het deeltje m
rc afstand tot de rotatieas m
r2 botsingsconstante (-)
Re Reynolds getal (-)
S Stokes sedimentatiecoefficient s
T temperatuur �
v sedimentatiesnelheid m/s
~vi snelheid van fase i m/s
vr,i relatieve snelheid van fase i m/s
V volume m3
Griekse symbolen
αi volume fractie van fase i (-)
αi,max maximum pakkingsdichtheid van fase i (-)
η temperatuurscoefficient �−1
λi bulkviscositeit van i Pa/s
µ viscositeit van het fluıdum Pa/s
µr µsusp/µplasma (-)
ω hoeksnelheid rad/s
φ hoek van interne wrijving 0
ρi dichtheid van deeltje i kg/m3
ρf dichtheid van het fluıdum kg/m3
ρsusp dichtheid van de suspensie kg/m3
τi spanningstensor (-)
τi deeltjes relaxatie tijd s
Θi granulaire temperatuur �
xi

Hoofdstuk 1
Het aferese proces van bloed
Het woord aferese is afkomstig van het Grieks en betekent letterlijk ‘afnemen’. In deze
context gaat het over een medische technologie waarbij bloed van een donor of een patient
door een centrifuge stroomt om zo bepaalde bloedcellen te verwijderen. De overblijvende
bloedcellen kunnen eventueel terug in circulatie worden gebracht doorheen het menselijk
lichaam.
Bloed is een complexe vloeistof dat levensnoodzakelijke bestandsdelen bevat, deze staan
in voor allerhande functies. Indien iemand een tekort heeft aan bepaalde elementen in het
bloed kan dit nefaste gevolgen voor het lichaam hebben. Bepaalde aandoeningen kunnen
worden verholpen mits toevoeging van bepaalde bloedcellen. Het is duidelijk dat het proces
om donorbloed te scheiden in zijn belangrijkste bestanddelen uitermate interessant is voor
de medische wereld.
1 Anatomie en functie van het bloed
Bloed is een circulerend vloeibaar weefsel. Een gezond volwassen persoon heeft normaal 5
liter bloed wat 7% is van het lichaamsgewicht. Het bloed is samengesteld uit verscheidene
soorten bloedcellen, deze elementen vormen ongeveer 45% van het totaal volume aan bloed.
De andere 55% bestaat uit bloedplasma, een geelachtige vloeistof die het dragend medium
van het bloed is. Gesuspendeerd in het bloedplasma zijn er 3 soorten bloedcellen:
• Rode bloedcellen of erythrocyten
• Bloedplaatjes of trombocyten
• Witte bloedcellen of leukocyten
1

Hoofdstuk 1. Het aferese proces van bloed 2
Omdat het bloed bijna overal in het lichaam stroomt, dient het voor het transport van voe-
dingsstoffen, afvalstoffen, zuurstof, kooldioxide, hormonen en natuurlijk ook water. Ook
verdeelt het bloed de warmte over het lichaam, enerzijds om oververhitting van warmtepro-
ducerende organen (bijv. spieren) te voorkomen, maar ook om de lichaamstemperatuur op
peil te houden. Een heel belangrijke functie van het bloed is de afweer, het bloed verdedigt
het lichaam tegen schadelijke micro-organismen.
1.1 Bloedplasma
Het bloedplasma is het vloeibare gedeelte van het bloed waarin de bloedcellen en de bloed-
plaatjes gesuspendeerd zijn. Het bloedplasma bestaat hoofdzakelijk uit water maar bevat
ook nog andere componenten, zoals vermeld in tabel 1.1
Tabel 1.1: Samenstelling bloedplasma
Component Gewichtsfractie
Water 92 %
Proteınen 7 %
Zouten 0.8 %
Vetten 0.6 %
Glucose 0.1 %
Het bloedplasma dient hoofdzakelijk als medium voor de bloedcellen maar het is eveneens
van nut als vervoermiddel van glucose, lipiden, ionen, afval (ureum), kooldioxide en in
mindere mate (5%) van zuurstof. Deze materialen zijn afkomstig van uitwisselingsorganen
zoals de darmen en de lever en gaan naar plaatsen waar ze uit het bloed worden verwijderd,
zoals de nieren, de huid en bij de gewone stofwisseling in de cel.
1.2 Rode bloedcellen
De rode bloedcellen zijn het talrijkst aanwezig in het bloed. Per kubieke millimeter bezit
een vrouw 4.8 en een man 5.4 miljoen rode bloedcellen.

Hoofdstuk 1. Het aferese proces van bloed 3
Een rode bloedcel heeft een levensduur van ongeveer
120 dagen in het menselijk lichaam. Buiten het li-
chaam kunnen rode bloedcellen maximaal 42 dagen
overleven bij een temperatuur van 1 tot 6 �. In-
dien ze worden bevroren zijn ze voor 10 jaar bruik-
baar. Een eenheid die veel wordt gebruikt om de
hoeveelheid rode bloedcellen te definieren is het he-
matocriet, dit is het aandeel volume van rode bloed-
cellen in het bloed.Figuur 1.1: Rode bloedcellen
Soms wordt met het hematocriet ook het aandeel volume van alle bloedcellen genoemd. Een
andere eenheid noemt men het hemoglobine, dit is de massa van de hemoglobinemoleculen
per volume eenheid, meestal uitgedrukt in g/l. Rode bloedcellen zijn schijfvormige, kernloze
cellen en hebben een doorsnede van 7 tot 8 micrometer en een dikte van 2 micrometer. De
rode bloedcel heeft een dichtheid van 1,095 tot 1,100 g/cm3. Het midden van de cel is aan
beide kanten hol wat het oppervlak vergroot en voordelig is voor een betere uitwisseling
van zuurstof en kooldioxide. Deze celvorm wordt verzorgd door het cytoskelet. Dit bestaat
uit een stevig netwerk dat aan de binnenkant van het celmembraan ligt en zorgt voor zijn
vormvastheid. Omdat de rode bloedcellen door de kleinste haarvaten, die smaller zijn dan
hun diameter, moeten kunnen zijn ze vervormbaar of kunnen ze dubbelvouwen. Daarom
is hun cytoskelet erg elastisch.
In volgende paragrafen worden de functies van de rode bloedcellen, namelijk het transport
van zuurstof en van kooldioxide, besproken.
1.2.1 Transport van zuurstof
Rode bloedcellen bevatten geen kern, daarom spreekt men soms ook van bloedlichaampjes.
Zo is er meer plaats voor het eiwit hemoglobine (Hb) dat instaat voor het transport van
zuurstof en kooldioxide. Een rode bloedcel bevat ongeveer 2, 8 ·108 moleculen hemoglobine.
Aangezien zuurstof maar matig oplost in water kan het bloedplasma niet volledig de taak
van het zuurstoftransport op zich nemen. Door de rode bloedcellen kan het bloed veertig
keer zoveel zuurstof bevatten dan er alleen in plasma kan worden opgelost. Het hemoglobine
bestaat enerzijds uit een heemdeel, heem is een ijzerverbinding, en anderzijds uit een
eiwitdeel, globine. Het hemoglobine is opgebouwd uit vier ijzerbevattende heemgroepen.
Aan elke heemgroep is het mogelijk om een zuurstofmolecule te binden, zodat er dus vier
O2-moleculen op een hemoglobine-molecule passen. Als de eerste O2 is gebonden, treedt

Hoofdstuk 1. Het aferese proces van bloed 4
er een verandering op aan de structuur van het hemoglobine-molecule, zodat elke volgende
O2-molecule zich gemakkelijker bindt. Dit beinvloed sterk de bindingskinetiek van zuurstof
aan het bloed.
De reactie van het zuurstof met de ijzerhoudende heemgroep is omkeerbaar. In de longen
wordt het zuurstof opgenomen en in de cellen wordt het zuurstof afgestaan aan de cellen.
We krijgen volgende evenwichtsreactie:
Hb + 4O2 ⇐⇒ Hb(O2)4
Onder lage temperatuur, hoge pH, hoge O2concentratie zal de reactie overhellen naar rechts.
Het donkerrode hemoglobine wordt dan het lichtrode oxyhemoglobine. Deze condities
komen vooral voor in de longen, daar zal het hemoglobine zuurstof opnemen. In de rest
van het lichaam heersen hogere temperaturen, lagere pH en een lage O2concentratie. De
reactie zal nu overhellen naar links en zuurstof zal worden afgestaan.
1.2.2 Transport van kooldioxide
Kooldioxide reageert met water om koolzuur te vormen, deze wordt door het enzym
koolzuur-anhydrase gekatalyseerd. We krijgen volgende reactie:
CO2 + H2O ⇐⇒ HCO−3 + H+
Op plaatsen waar er weinig kooldioxide aanwezig is zal het bloed alkalischer zijn met als ge-
volg dat het hemoglobine de zuurstof sterker bindt en dus moeilijker afstaat. Omgekeerd,
waar er veel kooldioxide is zal het hemoglobine de zuurstof gemakkelijker loslaten. Het
kooldioxide en de vrijgekomen protonen bezetten een andere bindingsplaats op het hemo-
globine dan zuurstof maar beınvloeden de interne elektronen configuratie en de affiniteit
van het hemoglobine voor zuurstof zal dalen. Van het kooldioxide wordt 95% getranspor-
teerd door de rode bloedcellen, slechts 5% wordt via het bloedplasma getransporteerd.
1.3 Witte bloedcellen
De naam witte bloedcellen is een verzamelnaam voor bloedcellen met uiteenlopende oor-
sprong, een verschillende vorm en met andere functies. Ze zijn groter dan de rode bloedcel-
len (figuur 1.2) maar minder talrijk. Er komt een witte bloedcel op 1000 rode bloedcellen
voor. De witte bloedcellen ook leukocyten genoemd, spelen een belangrijke rol bij de be-
strijding van ziektekiemen omdat ze die kunnen onschadelijk maken. De witte bloedcellen
kunnen worden ingedeeld in twee grote groepen: de poly- en de mononucleaire leukocyten.

Hoofdstuk 1. Het aferese proces van bloed 5
1.3.1 Polynucleaire leukocyten
De polynucleaire leukocyten hebben een kern die verdeeld is in verscheidene segmenten,
zodat ze veelkernig lijken. Ze bevatten ook korrels of granulen in het cytoplasma, daarom
noemt men ze vaak granulocyten. Ze hebben een diameter tussen de 12 en de 14 micrometer.
Men onderscheidt drie soorten granulocyten: de neutrofielen, de eosinofielen, de basofielen.
De neutrofielen vertegenwoordigen meer dan de helft van de leukocytenpopulatie. Dringen
er bacterien of vreemde proteınen in het lichaam binnen, dan verlaten de neutrofielen de
bloedbaan doorheen de dunne wand van de haarvaten en verplaatsen ze zich naar de plaats
waar de ’indringer’ zich bevindt. Daar omsluiten ze de vreemde stof, nemen deze op en
verteren ze, dit is het verschijnsel dat fagocytose heet. Dit verteringsproces is mogelijk
dankzij de enzymen die in korrels worden afgescheiden. De eosinofielen zijn belangrijk bij
een parasitaire infectie of bij sommige allergische reacties. De basofielen spelen een rol bij
acute allergische reacties.
1.3.2 Mononucleaire leukocyten
Tot de mononucleairen behoren de lymfocyten en de monocyten. Beiden hebben slechts
een enkele grote kern.
De lymfocyten hebben een diameter van 6 tot 9 micrometer. Men onderscheidt twee typen
van lymfocyten: B- en T-lymfocyten. Elk op zich vervullen ze een bijzondere functie in
het afweersysteem van het organisme tegen ziekten. De B-lymfocyten vormen antistoffen,
terwijl de T-lymfocyten de vreemde proteınen bestrijden en vernietigen.
Figuur 1.2: Witte bloedcellen
De monocyten zijn eveneens in staat tot fagocytose, zo verlaten de monocyten de bloedbaan

Hoofdstuk 1. Het aferese proces van bloed 6
en begeven zich naar de infectiehaard, waar ze macrofagen worden genoemd. Ze nemen
ook dode cellen en andere afbraakstoffen op om ze te verteren. De monocyten zijn veel
groter dan de neutrofielen, hun diameter varieert tussen de 16 en de 20 micrometer. In
tabel 1.2 staat de relatieve verdeling van de verschillende witte bloedcellen vermeld.
Tabel 1.2: De relatieve verdeling van de verschillende witte bloedcellen
Polynucleairen
Neutrofielen 50− 65 %
Eosinofielenınen 1− 3 %
Basofielen 0− 1 %
Mononucleairen
Lymfocyten 25− 35 %
Monocyten 2− 8 %
1.4 Bloedplaatjes
De bloedplaatjes of trombocyten zijn eigenlijk geen cellen maar celfragmenten afkomstig
van hun moederstamcel. Hun diameter bedraagt slechts 2 micrometer en ze hebben een
willekeurige schijfvorm. Hun levensduur in de bloedbaan bedraagt ongeveer 10 dagen.
Buiten de bloedbaan moeten de plaatjes op kamertemperatuur worden gehouden en kunnen
ze 5 dagen overleven. De belangrijkste taken van de bloedplaatjes hebben betrekking op
de bloedstolling. Van zodra een bloedvat een defect vertoont, zal er zich onmiddellijk
een aggregaat van bloedplaatjes vasthechten aan de ruwe wanden van de wond, waardoor
in de meeste gevallen een volledige afsluiting van het lek tot stand komt. Wanneer het
echter om een grotere verwonding gaat, zal het bloedplaatjesaggregaat alleen niet volstaan
om de bloeding te stelpen. Dan worden er door de bloedplaatjes en het beschadigde
weefsel verscheidene factoren vrijgemaakt, die op hun beurt het ingewikkeld mechanisme
van de bloedstolling op gang brengen. De interactie van de bloedplaatjesfactoren en de
proteınefactoren van het stollingsmechanisme geven aanleiding tot de vorming van een
bloedklonter, waarmee het verwonde vat wordt gedicht.

Hoofdstuk 1. Het aferese proces van bloed 7
1.5 Productie en afbraak van bloedcellen
Al de verschillende soorten bloedcellen worden geproduceerd in het beenmerg dat zich
bevindt in de mergholte van bepaalde beenderen. Dagelijks worden er meer dan een biljoen
(1012) cellen gevormd. Deze cellen ontstaan allemaal uit een type cel, de hematopoetische
stamcellen. Figuur 1.3 beschrijft het schema dat aantoont hoe een cel kan leiden tot de
productie van alle bloedcellen.
Figuur 1.3: Hematopoese
Deze stamcel kan via mitosis ofwel zichzelf reproduceren ofwel meehelpen met de productie
van bloedcellen langs bovenstaande paden.
Oude en/of beschadigde cellen worden uiteindelijk in de lever en milt opgeruimd door ma-
crofagen. Aan de hand van hun elasticiteit wordt bepaald of de bloedcellen nog functioneel
zijn. De voedingsstoffen die daarbij vrijkomen worden weer afgegeven aan het bloed. Met
name het ijzer uit het hemoglobine wordt grotendeels weer herbruikt voor de aanmaak van
nieuw hemoglobine.
1.6 Samenvatting
Kort kunnen we de relevante fysische eigenschappen voor het modelleren van sedimentatie
samenvatten in tabel 1.3.

Hoofdstuk 1. Het aferese proces van bloed 8
Tabel 1.3: Fysische eigenschappen van de bloedcellen
Aantal Diameter Dichtheid
(l/µL) (µm) (kg/m3)
Rode bloedcellen 5 000 000 6 - 8 1 095 - 1 100
Witte bloedcellen 5 000 1 055 - 1 085
Granulocyten 3 000 12 - 14
Lymfocyten 1 500 6 - 9
Monocyten 500 16 - 20
Bloedplaatjes 200 000 2 - 4 1 050
2 Toepassingen van de centrifuges
Het aferese proces begint met de verwijdering van bloed van een patient of een donor.
Vervolgens passeert dit bloed in een centrifuge. Doordat de bloedcellen verschillende dicht-
heden hebben kunnen ze gemakkelijk worden gescheiden door de centrifugale kracht. De
snelle rotaties zorgen ervoor dat de bloedcellen aan een intense versnelling worden on-
derworpen zodat het sedimentatieproces versneld wordt. De gewenste component wordt
dan ontnomen en de resterende componenten kunnen worden teruggebracht door transfu-
sie in de patient of de donor. Deze scheiding kan worden uitgevoerd voor therapeutische
behandelingen, peri-operatieve transfusie en eveneens bij het stockeren van bloed voor
bloedbanken.
2.1 Therapeutische behandelingen
Een van de therapeutische scheidingsoperaties is het verzamelen van de bloedplaatjes. Zo
kan er een gel worden gemaakt die ervoor zorgt dat de herstelling van botfracturen en grote
wonden sneller en efficienter verloopt. Het proces om de bloedplaatjes uit het bloed te halen
is tot nu toe nog niet geperfectioneerd. De opbrengst van de moderne apparatuur bedraagt
slechts 70 %. Het is de bedoeling zoveel mogelijk plaatjes in een zo klein mogelijk volume
te krijgen, aangezien er een correlatie is tussen het aantal groeifactoren en de concentratie
aan bloedplaatjes. Dit afstudeerwerk start een onderzoekt met als doel de opbrengst te
verhogen.
Bij therapeutische aferese kan ook een component van het bloed worden verwijderd dat
niet goed functioneert of dat bijdraagt tot een ziekte. De verwijderde bloedcellen worden

Hoofdstuk 1. Het aferese proces van bloed 9
dan vervangen door gezonde cellen van een donor. Dit wordt veel toegepast bij leuke-
miepatienten. Zij worden behandeld met chemotherapie, waardoor veel beenmerg, dat
bloedplaatjes aanmaakt, wordt gedood. Hun beschadigde cellen worden zo vervangen, zij
krijgen dan regelmatig een donatie van bloedplaatjes. In andere gevallen van leukemie,
waarbij er teveel witte bloedcellen aanwezig zijn, kan de verwijdering van de witte bloed-
cellen helpen complicaties zoals trombose te vermijden.
2.2 Peri-operatieve transfusie
De centrifuges kunnen worden gebruikt voor donatie van plasma, rode bloedcellen of bloed-
plaatjes maar kunnen ook gebruikt worden voor bloedrecuperatie tijdens een operatie.
Wanneer een patient veel bloed verliest tijdens een operatie is dit bloed niet meer bruik-
baar, het stollingsmechanisme werd in gang gezet en maakt het onmogelijk om dit bloed
terug in de patient te brengen. Door de centrifuges kunnen echter de rode bloedcellen
gerecupereerd worden zodat de patient erna geen problemen zal hebben met slecht func-
tionerend bloed. Een andere manier die veel wordt toegepast, is bloed nemen van een
patient voor de operatie, dit weerlegt de risico’s van een dure transfusie van een derde.
2.3 Toepassing in bloedtransfusiecentra
In bloedtransfusiecentra wordt bloed afgenomen van een donor en eventueel verwerkt tot
verschillende eindproducten voor later gebruik. In de huidige geneeskunde krijgt elke
patient immers enkel het bloedbestanddeel dat hij nodig heeft. Het bloed wordt dan
in eerste instantie door een centrifuge gesplitst in een zakje rode bloedcellen en een zakje
plasma. In een aantal gevallen worden er nog bloedplaatjes van het bloed gescheiden.
De rode bloedcellen worden toegediend aan ziekenhuispatienten die een tekort hebben
aan bloed, zoals mensen met bloedarmoede of slachtoffers van verkeersongevallen. Op
deze manier kunnen ze worden gered want hun zuurstoftransport is terug op peil. Uit
plasma wordt een heel gamma van geneesmiddelen bereid voor tal van toepassingen. Zo
bieden stollingsfactoren hemofiliepatienten een quasi-normaal leven en kunnen vrouwen
met ernstige bloedingen na een bevalling geholpen worden. Antistoffen worden uit plasma
gehaald en gebruikt om ziekten te bestrijden, bijvoorbeeld tegen tetanus en geelzucht.
Eiwitpreparaten, zoals albumine, zijn onder meer nuttig voor brandwondenpatienten en
mensen die in shock toestand verkeren.

Hoofdstuk 1. Het aferese proces van bloed 10
3 Types centrifuges
In deze paragraaf worden de meest gebruikte centrifuges besproken. Er wordt uitgelegd
hoe ze functioneren en wat hun mogelijkheden zijn. In eerste instantie wordt de aandacht
besteed aan het materiaal van Dideco, aangezien het apparaat dat gemodelleerd wordt
van deze fabrikant is. Vervolgens wordt het materiaal besproken dat gebruikt wordt voor
de verwerking van volbloed en voor aferese van een bepaald bestanddeel. Het besproken
materiaal wordt gebruikt in het bloedtransfusiecentrum van Gent.
3.1 Dideco
Dideco is een Italiaans bedrijf en een van de marktleiders van bloedcentrifuges en hun
aanverwante apparaten. Ze bieden hun klanten drie soorten apparaten: Angel, Electa en
de Excel pro.
3.1.1 Angel
Het eenvoudigste model is de Angel (figuur 1.4).
Ze wordt gebruikt voor het aanmaken van de
plaatjesgel. Dit apparaat heeft de mogelijkheid
om verschillende hoeveelheden bloed te verwerken.
Op deze manier kan er een beperkt aantal bloed
worden afgenomen genoeg voor de aanmaak van
de gel. Figuur 1.4: Dideco Angel
3.1.2 Electa
De Electa (figuur 1.5) is een complexer apparaat
dat het bloed scheidt maar eveneens wast, zo-
dat niet gewenste stoffen zoals anticoagulenten,
fibronogenen en andere eiwitten uit het plasma
worden afgescheiden. Het geconcentreerde bloed
is dan ook bruikbaar voor een andere patient. Figuur 1.5: Dideco Electa
In de Electa worden centrifuges geplaatst met verschillende volumes, genaamd Latham
bowls (zie figuur 1.6).

Hoofdstuk 1. Het aferese proces van bloed 11
Hun kleinste model heeft een volume van 55ml,
wat goed kan gebruikt worden tijdens operaties
waar slechts een kleine nood is aan een bepaalde
hoeveelheid autoloog bloed. De andere centrifuges
hebben een inhoud van 125ml, 175ml en 225ml. Figuur 1.6: Dideco bowls
Eveneens beschikt de Electa over automatische sensoren die de buffy coat1 detecteren en
de kwaliteit van het bloed controleren. De Electa wordt gebruikt tijdens de experimentele
studie (zie hoofdstuk 3 en 4).
In figuur 1.7 is een schematische weergave te zien van een recuperatie van rode bloedcellen.
„ …
Figuur 1.7: Schematische weergave van de Electa
In eerste instantie wordt het bloed gefilterd om grove deeltjes te verwijderen. Vervolgens
kunnen we tijdens het gebruik van dit toestel drie fasen onderscheiden:
1. Het bloed wordt door een peristaltische pomp in de centrifuge gepompt met een
1De buffy coat is een algemene naam voor het laagje van bloedplaatjes en witte bloedcellen dat zich opde rode bloedcellen bevindt na sedimentatie.

Hoofdstuk 1. Het aferese proces van bloed 12
bepaalde vulsnelheid. De centrifuge wordt geleidelijk gevuld en er onstaat een duide-
lijke afscheiding van het plasma (zie figuur 1.8). Naarmate de centrifuge wordt gevuld
zal uiteindelijk het plasma de centrifuge langs boven verlaten.
2. Wanneer de sensor de buffy coat detecteert stopt de pomp en worden de rode bloedcel-
len gewassen door de wasvloeistof. Noteer dat er nu geen bloed meer wordt ingepompt
en dat er enkel gebruikt wasvloeistof de centrifuge verlaat.
3. Als het bloed voldoende gewassen is zal de peristaltische pomp terug gestart worden
in tegengestelde richting zodat de rode bloedcellen langs de inlaat naar buiten worden
gepompt.
Figuur 1.8: Centrifuge in werking
De Electa kan ook gebruikt worden voor het collecteren van bloedplaatjes. De sensor
detecteert de buffy coat wanneer die zich aan de uitgang bevindt en dan zal dit laagje
verzameld worden in een apart zakje. Indien de plaatjes niet bestemd zijn voor autologe
toepassingen, moeten de plaatjes nog gescheiden worden van de witte bloedcellen. Dit
wordt gedaan aan de hand van filters of een tweede operatie met een centrifuge.

Hoofdstuk 1. Het aferese proces van bloed 13
3.1.3 Excel pro
Dit toestel is een aferese apparaat dat het bloed
scheidt in al zijn componenten en eventueel be-
paalde bestandsdelen terugstuurt naar de donor.
Dit is het meest geavanceerde toestel van Dide-
co. Heel wat parameters worden live gemeten
zodat, indien er zich enig probleem voordoet, de
machine dit kan melden. Het toestel is eveneens
in staat om stamcellen te scheiden.
Figuur 1.9: Dideco Excel pro
3.2 Volbloed verwerking
In bloedtransfusiecentra wordt volbloed verwerkt tot verschillende eindproducten. In deze
paragraaf wordt het materiaal van Baxter besproken. Wanneer een bloedafname wordt
uitgevoerd, komt het bloed terecht in een steriel systeem van zakken. De zak, met daaraan
enkele satellietzakken, wordt in een centrifuge op hoge snelheid afgedraaid (zie figuur 1.10).
Figuur 1.10: Centrifuge voor volbloed
De cellen zullen zich ordenen in de plastiekzak, rode bloedcellen onderaan, daarop de
buffycoat en bovenaan bevindt zich het plasma. De zak bevat aan de boven en de onderzijde

Hoofdstuk 1. Het aferese proces van bloed 14
een slang die leidt naar de satelietzakjes. Nadat de zak werd gecentrifugeerd, wordt ze
in een pers geplaatst (zie figuur 1.11). De zak wordt dan langzaam leeggeperst waarbij
er automatisch voor gezorgd wordt dat de scheidingslijn in het midden blijft, zodat de
bloedplaatjes er niet kunnen uitgeperst worden.
Figuur 1.11: Pers voor volbloed
Wanneer de zak bijna leeg is wordt de pers gestopt en worden de slangen met een speciaal
seal-apparaat dichtgesmolten. Hetgeen nog overblijft zijn de bloedplaatjes, deze worden
gemengd met een bewaarvloeistof en dan nog lichtjes gecentrifugeerd om de resterende
rode bloedcellen te verwijderen. Zo wordt van een donor een zak rode bloedcellen, een zak
plasma en een zakje bloedplaatjes verkregen.
3.3 Aferese-apparatuur
3.3.1 Plaatjes aferese
Met behulp van aferese-apparatuur wordt van een donor slechts het bloedbestanddeel afge-
nomen waaraan op dat moment specifiek behoefte is. Door de aggressieve therapieen, zoals
chemotherapie, is de vraag naar bloedplaatjes zeer groot. Aan de hand van deze toestellen
is het mogelijk enkel de plaatjes af te nemen van de donor. Dit complex proces kan worden
uitgevoerd door het Excel apparaat van Dideco zoals hierboven al werd aangehaald. Een
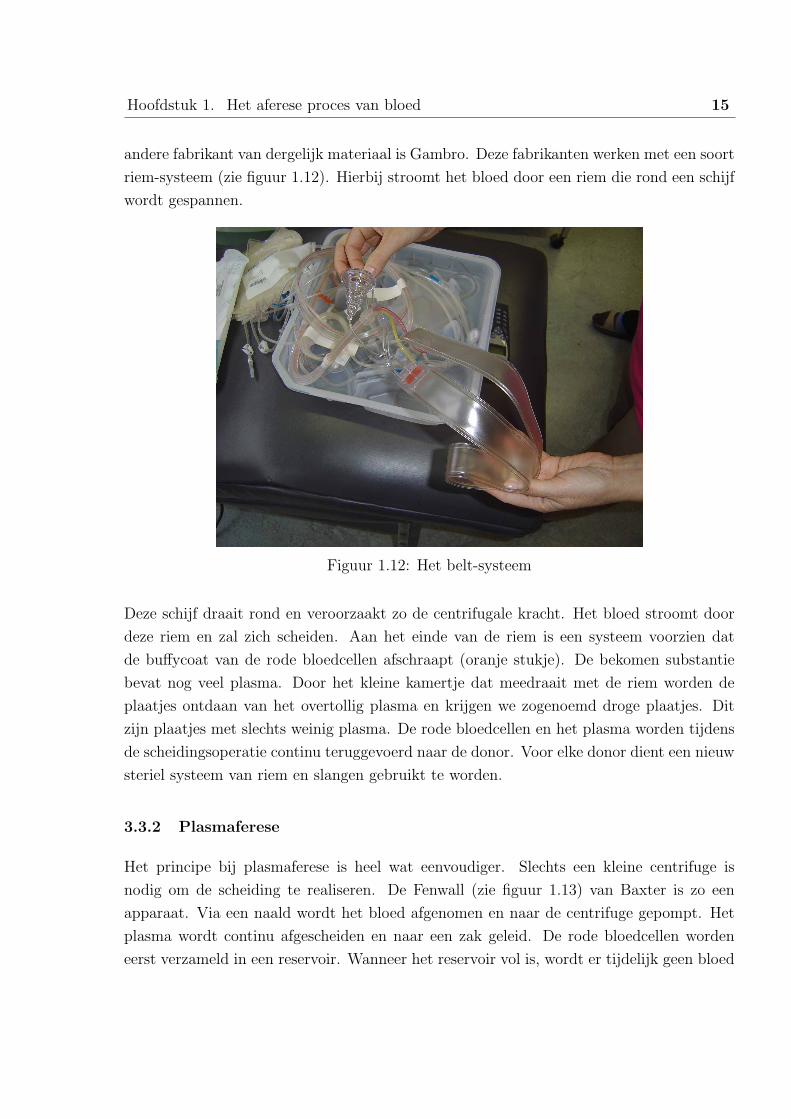
Hoofdstuk 1. Het aferese proces van bloed 15
andere fabrikant van dergelijk materiaal is Gambro. Deze fabrikanten werken met een soort
riem-systeem (zie figuur 1.12). Hierbij stroomt het bloed door een riem die rond een schijf
wordt gespannen.
Figuur 1.12: Het belt-systeem
Deze schijf draait rond en veroorzaakt zo de centrifugale kracht. Het bloed stroomt door
deze riem en zal zich scheiden. Aan het einde van de riem is een systeem voorzien dat
de buffycoat van de rode bloedcellen afschraapt (oranje stukje). De bekomen substantie
bevat nog veel plasma. Door het kleine kamertje dat meedraait met de riem worden de
plaatjes ontdaan van het overtollig plasma en krijgen we zogenoemd droge plaatjes. Dit
zijn plaatjes met slechts weinig plasma. De rode bloedcellen en het plasma worden tijdens
de scheidingsoperatie continu teruggevoerd naar de donor. Voor elke donor dient een nieuw
steriel systeem van riem en slangen gebruikt te worden.
3.3.2 Plasmaferese
Het principe bij plasmaferese is heel wat eenvoudiger. Slechts een kleine centrifuge is
nodig om de scheiding te realiseren. De Fenwall (zie figuur 1.13) van Baxter is zo een
apparaat. Via een naald wordt het bloed afgenomen en naar de centrifuge gepompt. Het
plasma wordt continu afgescheiden en naar een zak geleid. De rode bloedcellen worden
eerst verzameld in een reservoir. Wanneer het reservoir vol is, wordt er tijdelijk geen bloed
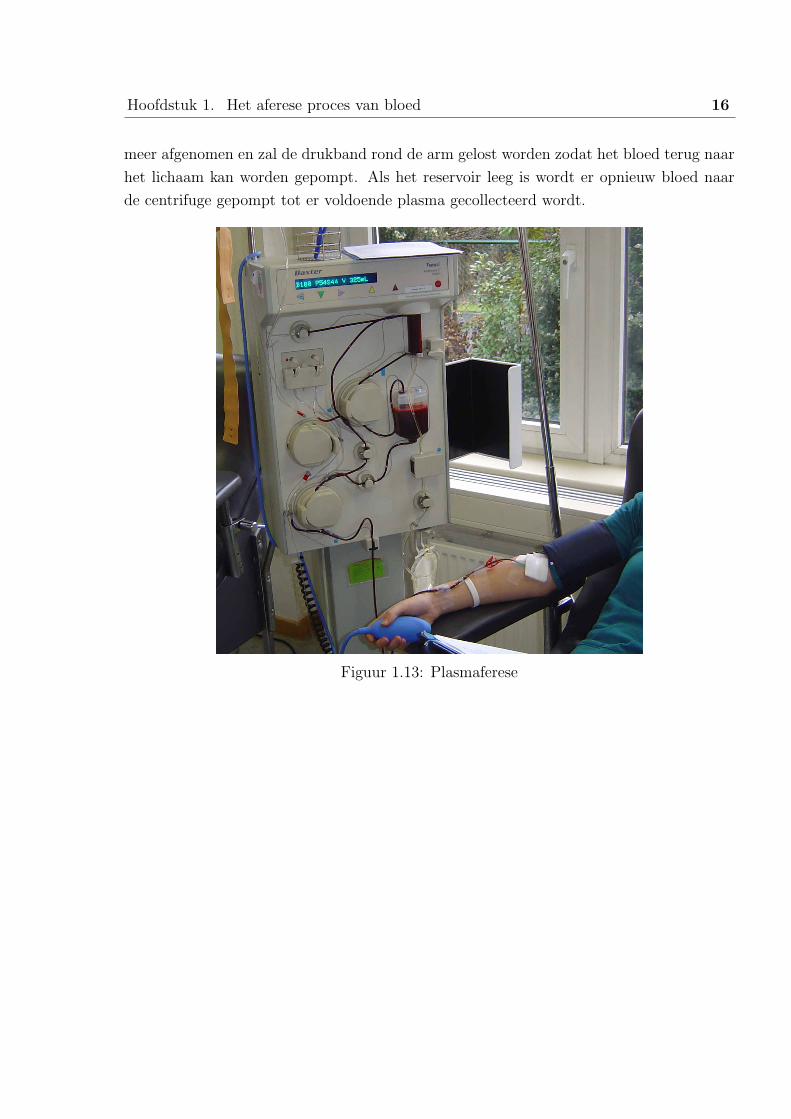
Hoofdstuk 1. Het aferese proces van bloed 16
meer afgenomen en zal de drukband rond de arm gelost worden zodat het bloed terug naar
het lichaam kan worden gepompt. Als het reservoir leeg is wordt er opnieuw bloed naar
de centrifuge gepompt tot er voldoende plasma gecollecteerd wordt.
Figuur 1.13: Plasmaferese

Hoofdstuk 2
Modellen voor het simuleren van de
sedimentatie van bloed
Gedurende voorbije 20 jaar zijn er al verschillende artikels verschenen die de sedimentatie
van bloed beschrijven. In dit hoofdstuk worden de belangrijkste modellen toegelicht die
gebruikt kunnen worden voor de modellering van een centrifuge. In de tijd dat er nog
geen goede computerpakketten waren, zoals Fluent, werd de sedimentatietheorie anders
bestudeerd dan nu. Heden kunnen vloeistofstromingen berekend worden door computers
terwijl vroeger wiskundige vergelijkingen werden opgesteld die dan getoetst werden door
experimentele studies. In dit hoofdstuk wordt eerst aandacht besteed aan deze oudere
modellen om dan recente manieren te bespreken voor het simuleren van de sedimentatie
van bloedcellen.
1 Sedimentatietheorie van het bloed
In 1984 beschreven Van Wie en Sofer [1] als eerste de sedimentatie van bloed. In volgende
paragrafen wordt beschreven hoe ze tot een wiskundige uitdrukking kwamen, vertrekkende
van de algemene sedimentatietheorie van Stokes.
1.1 Wet van Stokes
Midden de 19de eeuw bestudeerde Stokes de beweging van een deeltje in een fluıdum.
Vertrekkend van de krachten op een deeltje wordt volgende balans bekomen:
~Fr = ~F + ~FB + ~FD (2.1)
17

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 18
met:
~Fr resultante, nettokracht~F uitwendige krachten uitgeoefend op het deeltje~FB de opstuwingskracht, hier werkend in tegengestelde zin van de uitwendige kracht~FD wrijvingskracht
Deze krachtenbalans kan vereenvoudigd worden door te onderstellen dat het deeltje on-
middellijk bezinkt met zijn constante eindvalsnelheid, er is dus geen versnelling van het
deeltje. Deze eindvalsnelheid wordt bereikt als de resulterende kracht nul wordt, dus als de
wrijvingskracht de uitwendige- en de opstuwingskracht compenseert. De krachtenbalans
wordt dan:
F − FB = FD (2.2)
waarbij
FD = 6πµr v (2.3)
met:
µ de viscositeit van het fluıdum
r de straal van het deeltje
v de valsnelheid van het deeltje in het fluıdum
Vergelijking 2.3 is de wrijvingskracht volgens Stokes. Deze is alleen geldig in het laminaire
Stokes regime, wat betekent dat het Reynolds getal1. De krachtenbalans wordt dan:
(mi −mf ) g = 6πµri vi (2.4)
met:
mi de massa van het deeltje
mf de massa van het verplaatste fluıdum
g de valversnelling
De index i duid op het type bloedcel: rode of witte bloedcellen of de bloedplaatjes. Wanneer
vergelijking (2.4) gedeeld wordt door het volume van een deeltje en de versnelling vervangen
wordt door deze in een centrifugaalveld dan is de terminale valsnelheid volgens Stokes:
vi =2
9
r2i (ρi − ρf ) ω2 rc
µ(2.5)
1Het Reynolds getal is gedefinieerd als Re = D vν met D de pijpdiameter, ν de kinematische viscositeit
en v de vloeistofsnelheid ligt tussen 10−4 en 0, 1

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 19
met:
ρi de dichtheid van deeltje i
ρf de dichtheid van het fluıdum
ω de hoeksnelheid
rc de afstand tot de rotatieas
Bij het opstellen van de vergelijking van Stokes moet aan enkele belangrijke voorwaarden
worden voldaan. Deze zijn:
1. Het deeltje moet bolvormig zijn.
2. Het deeltje mag niet in de buurt komen van andere deeltjes die de sedimentatie
kunnen beınvloeden.
3. Het Stokes regime moet gelden, en het Reynolds getal gebaseerd op de deeltjesdia-
meter2 en de sedimentatiesnelheid, moet kleiner zijn dan een.
4. De Corioliseffecten moeten verwaarloosbaar zijn.
Bij bloedcellen zijn de deeltjes Reynolds getallen altijd kleiner dan een omdat de straal
van een bloedcel zeer klein is. Het Stokes regime is ook voldaan. De Corioliskrachten zijn
verwaarloosbaar omdat de centrifugaalkracht ongeveer 400-maal groter is. Aan voorwaarde
3 en 4 is dus voldaan. Aan voorwaarde 1 en 2 is niet altijd voldaan. De erythrocyten zijn
niet bolvormig en de cellen komen zeer dicht in elkaars buurt zodat ze elkaar zullen hinderen
in het sedimentatieproces.
1.2 Invoering van een vormfactor
Bovenstaand probleem kan worden verholpen door de Stokes vergelijking aan te passen.
Er kan bijvoorbeeld een vormfactor worden ingevoerd:
θ =f
f0
(2.6)
In vergelijking (2.6) stelt f de wrijvingscoefficient voor van een bloedcel en f0 is de wrij-
vingscoefficient van een sfeer met hetzelfde volume. Voor een rode bloedcel kan de vormfac-
tor benaderd worden door deze van een ellipsoıde, uitgedrukt in functie van de afmetingen
2Het Reynolds getal is hier anders gedefinieerd: Re = d vν met d de deeltjesdiameter, ν de kinematische
viscositeit en v de vloeistofsnelheid

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 20
van de kleine en de grote straal.
f
f0
=( b2
a2 − 1)12
ba
23
arctan( b2
a2 − 1)12
(2.7)
met:
a de grote straal van een bloedcel
b de kleine straal of de dikte van een bloedcel
De wet van Stokes voor de bezinking van een deeltje wordt dan:
vi =2
9
r2i (ρi − ρf ) ω2 rc
µθ(2.8)
of
vi = Si ω2rc (2.9)
met Si de sedimentatiecoefficient van deeltje i:
Si =2
9
r2i (ρi − ρf )
µθ(2.10)
Aan de hand van de ontwikkelde formules kan de sedimentatietheorie getoetst worden aan
experimenten uitgevoerd in sterk verdunde oplossingen. Helaas blijken er dan nog steeds
afwijkingen te zijn, die te wijten zijn aan de vormfactor die een benadering is van de echte
celvorm.
1.3 Correctiefactor voor de onderlinge impacteffecten
Sedimentatiesnelheden bij hogere concentraties kunnen worden bestudeerd door nog een
andere modificatie toe te passen op de stokesvergelijking. In studies van Vand en Hawksley[2]
wordt de vergelijking van Stokes hervormd door de viscositeit van de suspensie te berekenen
rekening houdende met volgende effecten:
1. Het effect dat onstaat door de aanwezigheid van vloeistofperturbaties, veroorzaakt
door naburige cellen.
2. Botsingen van de eerste orde met andere deeltjes.
3. De dichtheid van het fluıdim wordt vervangen door de dichtheid van de suspensie.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 21
De Vand-Hawksley [2] [3] theorie leidt dan tot volgende uitdrukking voor de sedimentatie-
snelheid:
vi = Si ω2 rc
ρi − ρsusp
ρi − ρf
exp
(− 4.1 Hct
1.64−Hct
)(2.11)
met:
vi de sedimentatiesnelheid voor component i
Si de Stokes sedimentatiecoefficient
Hct het hematocriet of volumefractie rode bloedcellen
ρsusp de dichtheid van de suspensie:
ρsusp = (Hct ρrbc + (1−Hct) ρplasma) (2.12)
1.4 Invloed van de rouleaux vorming
Aan de hand van vergelijking (2.11) kan de sedimentatiesnelheid van de rode bloedcellen
(Vrbc) de witte bloedcellen (Vwbc), en de bloedplaatjes (Vp) worden uitgezet in functie
van het hematocriet. De fysische karakteristieken die werden gebruikt kunnen worden
teruggevonden in tabel 2.1. De waarden van de Stokes sedimentatiecoefficient zijn degene
die worden gebruikt door Sartory [4]. Deze geschatte waarden zijn gebaseerd op zijn
ervaring en zijn heel wat betrouwbaarder dan de berekende waarden. Enerzijds is de
vormfactor niet correct te bepalen en anderzijds hebben rode bloedcellen de neiging om
samen te klonteren wat men rouleaux vorming noemt. Dit beınvloedt de straal van de
deeltjes en aangezien deze voorkomt onder een kwadraat in de vergelijking van Stokes
zouden de sedimentatiesnelheden kunnen worden onderschat.
Tabel 2.1: Karakteristieken van de componenten van het bloed
ρ (g/cm3) r (cm×104) S0 (sec×10) µ (mPa · s× 102)
RBC 1.093 3.75 12.0 —
WBC 1.066 5 – 7.5 1.2 —
Plaatjes 1.053 1.19 0.032 —
Plasma 1.026 — — 2.6
Aan de hand van figuur 2.1 kunnen we onderstellen dat de sedimentatiesnelheid sterk stijgt
bij lage hematocrieten. Sartory [4] heeft eveneens experimenteel aangetoond dat de opti-
male scheiding zich voordoet bij een hematocriet van 15%, er kan dan worden afgevraagd
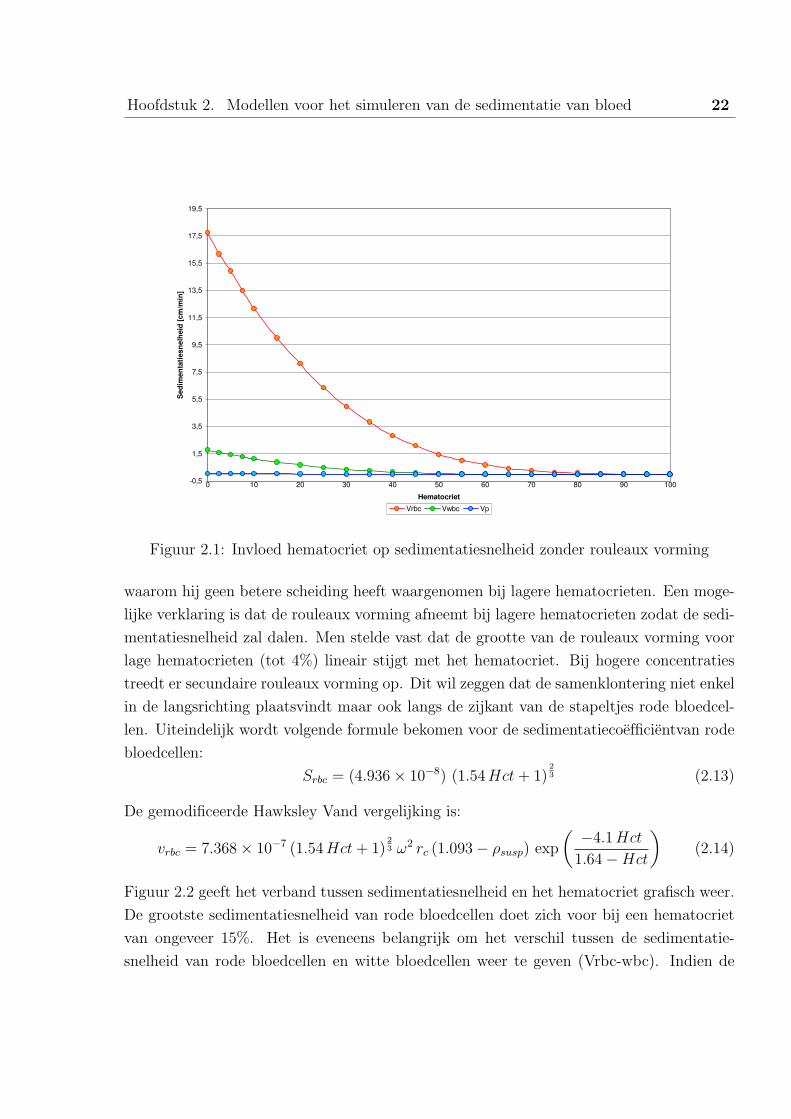
Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 22
-0,5
1,5
3,5
5,5
7,5
9,5
11,5
13,5
15,5
17,5
19,5
0 10 20 30 40 50 60 70 80 90 100
Hematocriet
Sed
imen
tati
esn
elh
eid
[cm
/min
]
Vrbc Vwbc Vp
Figuur 2.1: Invloed hematocriet op sedimentatiesnelheid zonder rouleaux vorming
waarom hij geen betere scheiding heeft waargenomen bij lagere hematocrieten. Een moge-
lijke verklaring is dat de rouleaux vorming afneemt bij lagere hematocrieten zodat de sedi-
mentatiesnelheid zal dalen. Men stelde vast dat de grootte van de rouleaux vorming voor
lage hematocrieten (tot 4%) lineair stijgt met het hematocriet. Bij hogere concentraties
treedt er secundaire rouleaux vorming op. Dit wil zeggen dat de samenklontering niet enkel
in de langsrichting plaatsvindt maar ook langs de zijkant van de stapeltjes rode bloedcel-
len. Uiteindelijk wordt volgende formule bekomen voor de sedimentatiecoefficientvan rode
bloedcellen:
Srbc = (4.936× 10−8) (1.54 Hct + 1)23 (2.13)
De gemodificeerde Hawksley Vand vergelijking is:
vrbc = 7.368× 10−7 (1.54 Hct + 1)23 ω2 rc (1.093− ρsusp) exp
(−4.1 Hct
1.64−Hct
)(2.14)
Figuur 2.2 geeft het verband tussen sedimentatiesnelheid en het hematocriet grafisch weer.
De grootste sedimentatiesnelheid van rode bloedcellen doet zich voor bij een hematocriet
van ongeveer 15%. Het is eveneens belangrijk om het verschil tussen de sedimentatie-
snelheid van rode bloedcellen en witte bloedcellen weer te geven (Vrbc-wbc). Indien de
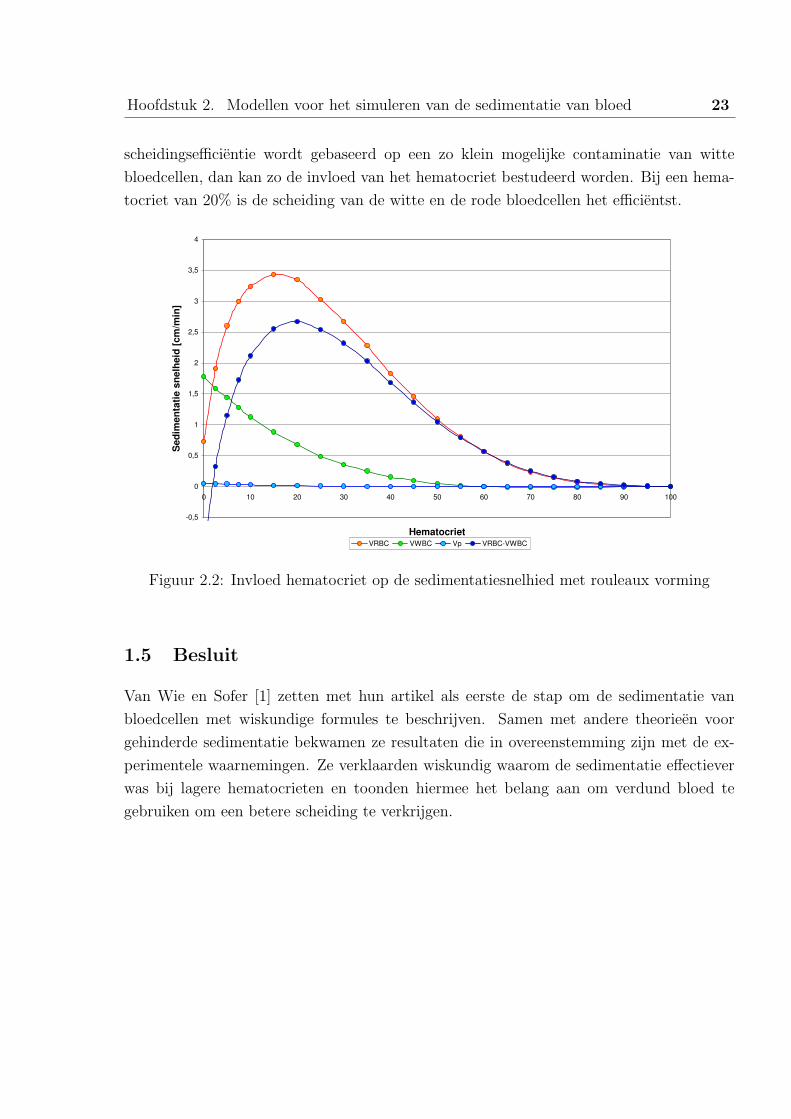
Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 23
scheidingsefficientie wordt gebaseerd op een zo klein mogelijke contaminatie van witte
bloedcellen, dan kan zo de invloed van het hematocriet bestudeerd worden. Bij een hema-
tocriet van 20% is de scheiding van de witte en de rode bloedcellen het efficientst.
-0,5
0
0,5
1
1,5
2
2,5
3
3,5
4
0 10 20 30 40 50 60 70 80 90 100
Hematocriet
Sed
imen
tati
e s
nelh
eid
[cm
/min
]
VRBC VWBC Vp VRBC-VWBC
Figuur 2.2: Invloed hematocriet op de sedimentatiesnelhied met rouleaux vorming
1.5 Besluit
Van Wie en Sofer [1] zetten met hun artikel als eerste de stap om de sedimentatie van
bloedcellen met wiskundige formules te beschrijven. Samen met andere theorieen voor
gehinderde sedimentatie bekwamen ze resultaten die in overeenstemming zijn met de ex-
perimentele waarnemingen. Ze verklaarden wiskundig waarom de sedimentatie effectiever
was bij lagere hematocrieten en toonden hiermee het belang aan om verdund bloed te
gebruiken om een betere scheiding te verkrijgen.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 24
2 Verdere studie van de deeltjesinteractie bij de sedi-
mentatie van bloedcellen
Enkele jaren later in 1988 beschreven Van Wie en Hustvedt [5] het sedimentatiemodel
opnieuw met meer aandacht voor de onderlinge deeltjesinteractie. Ondertussen werden er
nieuwe modellen ontwikkeld voor de sedimentatie van deeltjes. Deze kunnen dan worden
toegepast op de sedimentatie van bloedcellen. In het vorig model werd er geen reke-
ning gehouden met de verschillende deeltjesgrootte, de 2de orde effecten bij botsingen,
de verschillen in dichtheden, de vervormbaarheid en de elektrische lading van het mem-
braanoppervlak van de cellen. Om de invloed van deze factoren te onderzoeken werden de
resultaten vergeleken met experimentele data van runderbloed, dat geen rouleaux vorming
heeft.
2.1 Invoering van een effectieve porositeit
Om aan de eis van de verschillende deeltjesgrootte te voldoen, wordt een effectieve po-
rositeit gedefinieerd zoals beschreven door Patwardhan en Tien [6]. Aan de hand van de
effectieve porositeit kan de sedimentatiesnelheid worden berekend zoals deze van individu-
ele deeltjes in de wet van Stokes. In hun werk wordt de lege vloeistoflaag dat een deeltje
omsluit beschouwd. Ze stellen dat deze laag wordt gereduceerd voor grotere deeltjes zodat
er een schijnbare porositeit optreedt. De gemiddelde diameter van alle deeltjes wordt gede-
finieerd zodat deze kan gebruikt worden om de dikte van de vloeistoflaag rond een deeltje
te bepalen.
dε = dgem
([1− ρrbc − ρsusp
ρrbc − ρplasma
]− 13
− 1
)(2.15)
met:
dε de dikte van het lege volume
dgem de gemiddelde deeltjes diameter
Deze waarden worden dan gebruikt om de effectieve porositeit εi te berekenen:
εi = 1−(
1 +dε
2ri
)−3
(2.16)

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 25
2.2 Model van Masliyah
Patwardhan en Tien opteerden voor het model van Masliya [7] (2.17) om de effectieve
porositeit te introduceren. Dit model is handig omdat de effectieve porositeit tot een
macht verheven wordt en deze macht kan aangepast worden om experimentele trends te
benaderen.
vi = Si ω2 rc
(ρi − ρsusp
ρi − ρf
)ε2.7i (2.17)
2.3 Model van Barnea en Mizrahi
Dit model [8] is gebaseerd op de verdere uitbreiding van de wrijvingscoefficient en het deel-
tjes Reynolds getal. Deze vergelijking stemde goed overeen met verscheidene experimentele
data.
vi = Si ω2 rc
(ρi − ρsusp
ρi − ρf
)exp(−5(1−εi)
3εi)
1 + (1− εi)13
(2.18)
2.4 Semi-empirisch model
Zoals in vorige paragraaf aangehaald leiden studies de van Vand [2] er toe om in de be-
rekening van de sedimentatiesnelheid rekening te houden met eerste orde botsingen en
stromingsfluctuaties van naburige deeltjes. Vand werkte verder op het model van Einstein
[9] en bekwam volgende uitdrukking voor de viscositeit van een suspensie:
ln(µr) =k1C + r2(k2 − k1)C
2 + . . .
1.0−QC(2.19)
met:
µr µsusp/µplasma
k1 de Einstein vormfactor
k2 de vormfactor van een botsingsdoublet
r2 botsingstijdconstante
Q hydrodynamische interactieconstante
C de totale volumetrische deeltjesconcentratie
De kracht van deze vergelijking zit hem in de parameters die het deeltje beschrijven. De
waarde van k1 bijvoorbeeld is een functie van de vorm, starheid, orientatie en Brownse
beweging. Waarden voor k2 kunnen theoretisch bepaald worden voor sferen maar niet
voor complexe vormen zoals bloedcellen. Hawksley gebruikte Vand’s viscositeitscorrelatie
Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 26
voor de bestudering van deeltjes in een gefluıdiseerd bed, maar hield geen rekening met
hogere orde interacties tussen de deeltjes. Na invoering van hogere orde reacties en gebruik
van een vormfactor θ van 2.5, 3.175 voor k2, 4 voor r2 en 0.609 voor de hydrodynamische
interactieconstante wordt volgende vergelijking bekomen:
vi = Si ω2 rc
(ρi − ρsusp
ρi − ρf
)exp
(−2.5(1.0− εi) + 4.0(3.175− 2.5)(1.0− εi)
2 + . . .
1.0− 0.609(1.0− εi)
)(2.20)
2.5 Resultaten
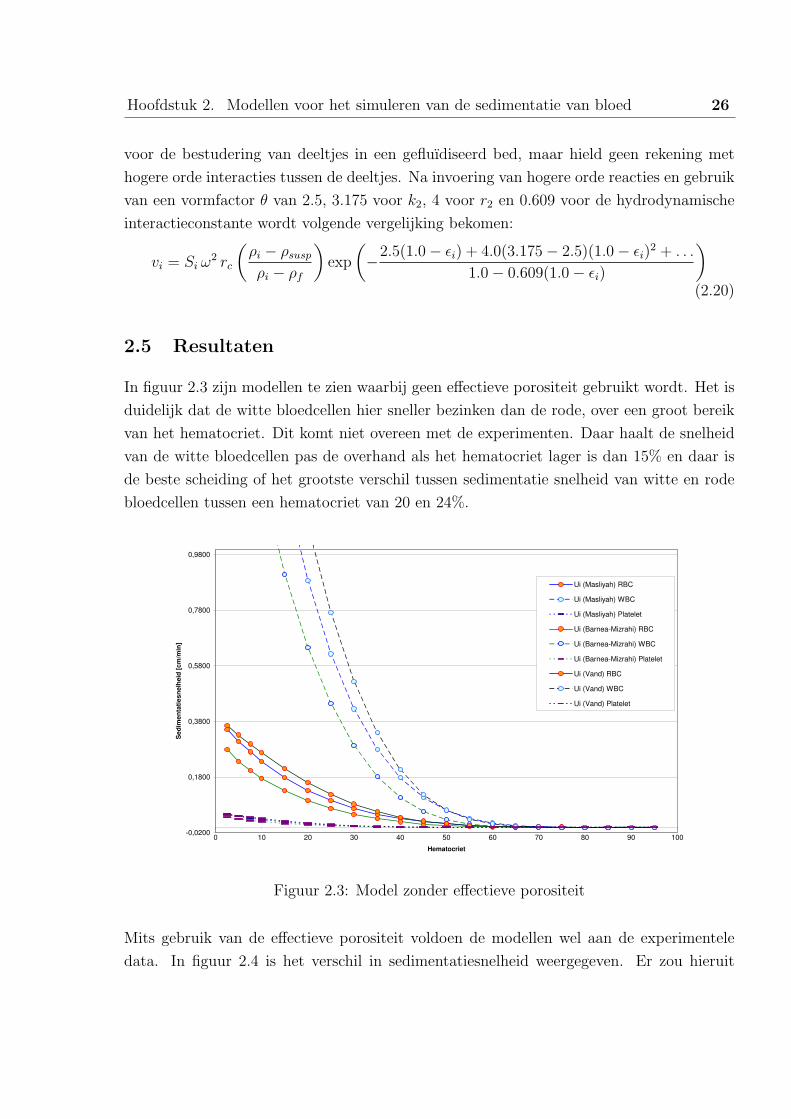
In figuur 2.3 zijn modellen te zien waarbij geen effectieve porositeit gebruikt wordt. Het is
duidelijk dat de witte bloedcellen hier sneller bezinken dan de rode, over een groot bereik
van het hematocriet. Dit komt niet overeen met de experimenten. Daar haalt de snelheid
van de witte bloedcellen pas de overhand als het hematocriet lager is dan 15% en daar is
de beste scheiding of het grootste verschil tussen sedimentatie snelheid van witte en rode
bloedcellen tussen een hematocriet van 20 en 24%.
-0,0200
0,1800
0,3800
0,5800
0,7800
0,9800
0 10 20 30 40 50 60 70 80 90 100
Hematocriet
Se
dim
en
tati
es
ne
lhe
id [
cm
/min
]
Ui (Masliyah) RBC
Ui (Masliyah) WBC
Ui (Masliyah) Platelet
Ui (Barnea-Mizrahi) RBC
Ui (Barnea-Mizrahi) WBC
Ui (Barnea-Mizrahi) Platelet
Ui (Vand) RBC
Ui (Vand) WBC
Ui (Vand) Platelet
Figuur 2.3: Model zonder effectieve porositeit
Mits gebruik van de effectieve porositeit voldoen de modellen wel aan de experimentele
data. In figuur 2.4 is het verschil in sedimentatiesnelheid weergegeven. Er zou hieruit
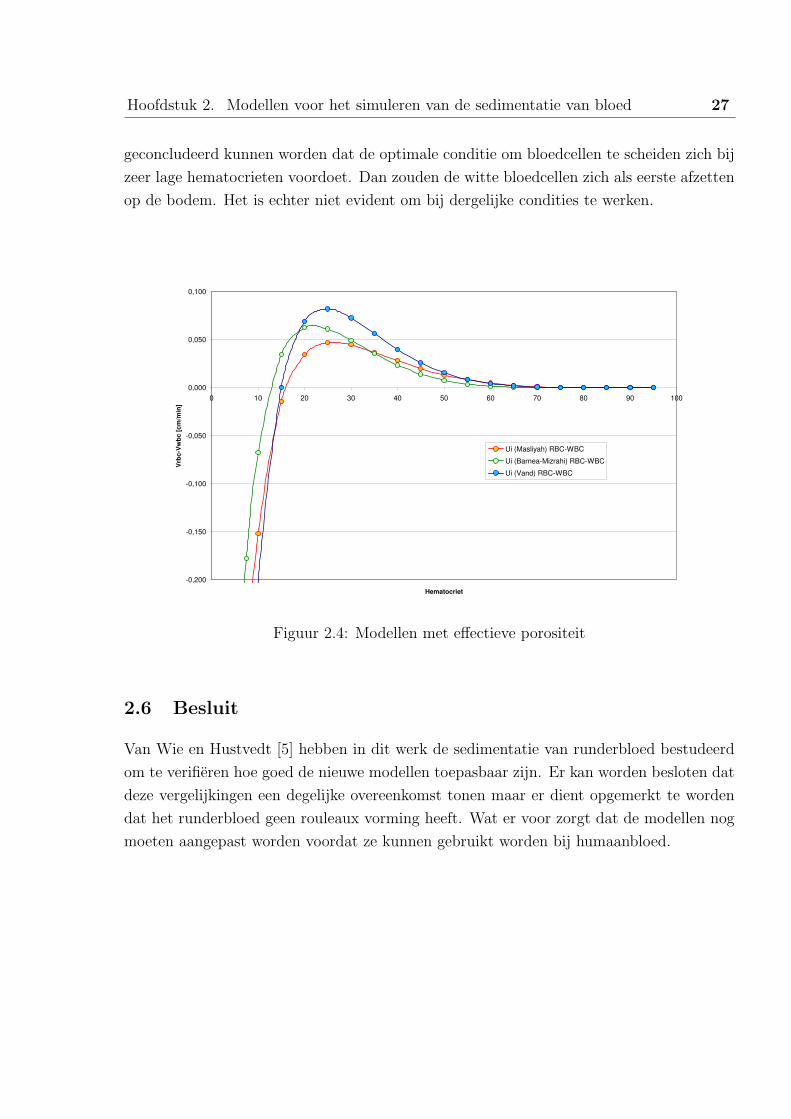
Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 27
geconcludeerd kunnen worden dat de optimale conditie om bloedcellen te scheiden zich bij
zeer lage hematocrieten voordoet. Dan zouden de witte bloedcellen zich als eerste afzetten
op de bodem. Het is echter niet evident om bij dergelijke condities te werken.
-0,200
-0,150
-0,100
-0,050
0,000
0,050
0,100
0 10 20 30 40 50 60 70 80 90 100
Hematocriet
Vrb
c-V
wb
c [
cm
/min
]
Ui (Masliyah) RBC-WBC
Ui (Barnea-Mizrahi) RBC-WBC
Ui (Vand) RBC-WBC
Figuur 2.4: Modellen met effectieve porositeit
2.6 Besluit
Van Wie en Hustvedt [5] hebben in dit werk de sedimentatie van runderbloed bestudeerd
om te verifieren hoe goed de nieuwe modellen toepasbaar zijn. Er kan worden besloten dat
deze vergelijkingen een degelijke overeenkomst tonen maar er dient opgemerkt te worden
dat het runderbloed geen rouleaux vorming heeft. Wat er voor zorgt dat de modellen nog
moeten aangepast worden voordat ze kunnen gebruikt worden bij humaanbloed.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 28
3 Simulatie van het sedimentatieproces
Met behulp van bovenstaande vergelijkingen kan een mathematisch model worden opge-
steld die de sedimentatie simuleert. Aan de Universiteit van Kaiserslautern werd door
Diehl [10] een dergelijk model opgesteld voor een centrifuge.
3.1 Het scheidingsproces
Dielh’s simulatie werd uitgevoerd op een spiraalvormige centrifuge. Aan het centrum
stroomt het bloed van de patient binnen, de bloedcellen gaan dan sedimenteren tegen de
buitenwand en zullen zo verder zakken tot het einde van de spiraal waar de rode bloedcellen
zich zullen ophopen. In het centrum van de spiraal wordt een wasvloeistof toegevoegd. De
hoogte van de gepakte laag rode bloedcellen aan het einde van de spiraal wordt continu
gemeten door een sensor. De kwaliteit van het instromend bloed hangt af van verschillende
parameters. De belangrijkste regelbare parameters van het proces zijn de vulsnelheid, de
rotatiesnelheid en het debiet van de wasvloeistof. Een ongekende parameter is het hema-
tocriet van het instromende bloed. Het vulniveau van de gepakte laag rode bloedcellen
dient als meetbare parameter om het proces te optimaliseren. Aan de hand van het vul-
niveau moeten de regelbare parameters worden aangepast zodat een optimale kwaliteit
van infuseerbaar bloed wordt bekomen. Hiervoor is een model nodig dat de invloed van
de parameters bestudeert, wat interessant is voor dit afstudeerwerk. De invloed van de
wasvloeistof is hier niet van belang en zal niet besproken worden.
3.2 De transportverschijnselen
Zoals bij de meeste modellen van roterende systemen worden ze getransformeerd naar niet
roterende systemen in een vlak. Zodoende wordt de spiraalvorm geprojecteerd naar een
schuin vlak (zie figuur 2.5).
De snelheid waarmee het concentraat aan de buitenkant van de kamer stroomt, is bedui-
dend lager dan de snelheid van het plasma bovenaan. Dit verschil is te wijten aan de
viscositeit van het bloed. Het functioneel verband tussen de relatieve viscositeit van bloed
en het hematocriet kan worden benaderd door een derde orde polynoom [10]. De relatieve
viscositeit van bloed wordt gerefereerd t.o.v. water(µwater = 1)).
µbloed,rel = 2.11912 + 4.37449 Hct− 8.65869 (Hct)2 + 23.3786 (Hct)3 (2.21)

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 29
Figuur 2.5: Schematische illustratie van de projectie
De stroming in de kamer kan worden geınterpreteerd als laminaire stroming van lagen
met verschillende viscositeit. De snelheid van zo een laag is omgekeerd evenredig met de
viscositeit:
vlaag ≈1
µlaag
(2.22)
Door deze manier van stroming zal het snelheidsprofiel niet parabolisch zijn zoals beschre-
ven voor laminaire stroming in een pijp.
De bezinkingssnelheid van een bloedcel wordt door Dhiel eenvoudigweg genomen door een
correctiefactor afhankelijk van het hematocriet toe te voegen bij de wet van Stokes:
fCorrectie(Hct) = (1−Hct)4 (2.23)
Uiteindelijk bekomt hij voor de wet van Stokes:
v = 4r2 ρ− ρ0
18µbloed
ω2 rc (1−Hct)4 (2.24)
Het gedrag van het systeem wordt beschreven door twee transportsystemen, de stroming
van het fluıdum in tangentiele of x-richting en de bezinking van de cellen in radiale of y-
richting. Door beide systemen te superponeren kan een model worden opgesteld dat de
sedimentatie simuleert.
3.3 Het simulatiemodel
Om een model te construeren wordt de kamer verdeeld in n equidistante segmenten en m
equidistante lagen. We verkrijgen zo in totaal n ·m gelijke volume elementen. De waarde

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 30
van de gravitationele kracht is gelijk aan de centripetale kracht in de kamer. Om gi,j te
bepalen wordt de afstand, ri,j, tot het centrum van het volume element ∆Vi,j berekend.
Dan is gi,j voor elk volume element, met ω als hoeksnelheid:
gi,j = ri,j ω2 (2.25)
Dit is een benadering aangezien de versnelling niet constant is in een volume element.
Samen met gi,j wordt er ook een Hcti,j(t) toegekend aan elk volume element.
Het algoritme om een simalutie uit te voeren bestaat uit vier stappen:
1. Berekening van het debiet van het fluıdum in de x-richting
2. Berekening van de nieuwe distributie van Hcti,j(t)
3. Berekening van de sedimentatiesnelheid volgens Stokes’ vergelijking
4. Berekening van de nieuwe distributie van Hcti,j(t)
3.3.1 Transport van het fluıdum
De debieten in en uit een volume element in de x-richting worden bepaald op basis van het
debiet gegenereerd door de peristaltische pomp (figuur 2.6). Het debiet dat instroomt in
een segment i, Vi, is wegens stationaire stroming gelijk aan het debiet van de pomp. De
som van de debieten in de lagen van een segement is dus gelijk aan het instromend debiet:
Vi =m∑
j=1
Vi,j (2.26)
Figuur 2.6: Schematische illustratie van de stromen

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 31
Zoals eerder vermeld is de snelheid of het debiet omgekeerd evenredig met de viscositeit
(2.22). De viscositeit kan berekend worden doordat de concentratie Hcti,j(t) van elk ele-
ment gekend is (2.21). Met (2.26) wordt bekomen dat:
Vi = stromingsfactori ·(
1
µi,1
+1
µi,2
+ · · ·+ 1
µi,m−1
+1
µi,m
)(2.27)
Omdat Vi en µi,j gekend zijn kan hieruit de stromingsfactor bepaald worden. Aan de hand
van die factor kan het debiet bepaald worden van elke laag in elk segment:
Vi,j = stromingsfactori ·1
µi,j
(2.28)
Voor elk volume element wordt deze berekening uitgevoerd zodat aan elk element ∆Vi,j
een viscositeit µi,j en een debiet Vi,j kan worden toegekend. Het getransporteerde volume
kan berekend worden uit het debiet Vi,j en de procestijd van een stap, ∆t. Het volume
wordt getransporteerd van een element ∆Vi−1,j naar het volgend element ∆Vi,j. Indien
het element dat volume niet kan ontvangen, wordt het overschot getransporteerd naar het
bovenliggend volume ∆Vi,j+1. Dit zal het geval zijn onderaan de kamer waar de cellen
bezinken. Deze lagen zullen vertragen doordat de viscositeit alsmaar zal stijgen door de
cellen die zich daar opstapelen. Bovenaan zal het omgekeerde zich voordoen, de cellen
zullen langzamerhand uit de laag bezinken waardoor deze zal versnellen. Het hematocriet
kan dan worden berekend uit:
Hct′i,j =
(∆Vi,j − Vi,j ·∆t
)·Hcti,j + Vi−1,j ·∆t ·Hcti−1,j
∆Vi,j
(2.29)
3.3.2 Sedimentatie van de bloedcellen
Voor elk element ∆Vi,j wordt de sedimentatiesnelheid vs(i,j) bepaald. Gebaseerd op deze
snelheid kan het aantal bloedcellen worden berekend dat zinkt naar het lager element:
Ri,j = 4 ·Hct′i,j · vs(i,j) ·∆t
dz(2.30)
Met Ri,j het aantal rode bloedcellen. Na deze berekening is het mogelijk om de nieuwe
concentraties te berekenen voor elke cel:
Hct′′i,j = Hct′i,j + Ri,j+1 −Ri,j (2.31)
Met deze als laatste stap is een cyclus doorlopen. De waarde Hct′′i,j(t) is de beginwaarde
van de volgende cyclus. Deze simulatie geeft de distributie van rode bloedcellen weer in de
centrifuge op elk tijdstip.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 32
3.4 Besluit
Dhiel [10] stelde een betrekkelijk goed model op om sedimentatie te simuleren. Alhoewel
de sedimentatiesnelheid niet accuraat beschreven wordt zoals bij de modellen van Van
Wie. Maar het is perfect mogelijk om de bezinkingssnelheid vooropgesteld door Van Wie
te gebruiken in dit model van Diehl. In formule (2.29) wordt het hematocriet niet exact
berekend, want de stroom die afkomstig is van het onderliggend elementje wordt niet
meegerekend.
4 Simulatie van het sedimentatieproces met behulp
van Computational Fluid Dynamics
In Augustus 2005 verscheen een artikel van De Gruttola [11] waarin een simulatie werd
uitgevoerd van de sedimentatie van bloed in een centrifuge met Computational Fluid Dy-
namics (CFD)3. Naderhand, in december 2005, verscheen een nieuwe versie van het artikel
waarbij er rekening gehouden werd met het niet-Newtoniaans zijn van het bloed. Dit arti-
kel is, aangezien zijn gelijkaardig doel, een relevante bron voor dit afstudeerwerk. In wat
volgt wordt het scheidingsproces en de sedimentatietheorie toegelicht. Vervolgens wordt
de berekeningsmethode aangehaald en als laatste worden de resultaten besproken.
4.1 Het scheidingsproces
In bovenvermeld artikel wordt in tegenstelling tot dit afstudeerwerk een continue centrifu-
ge gesimuleerd. Een cilindervormige kamer wordt geroteerd zodat de cellen afscheiden op
de buitenste wand (figuur 2.7). Dit drie dimensionale ontwerp wordt geprojecteerd naar
het vlak zodat de berekeningen eenvoudiger verlopen, de cellen zullen nu bezinken op de
onderste wand. Aan de linkerkant van de projectie (figuur 2.7) stroomt het bloed continu
in de centrifuge binnen.
3Computation Fluid Dynamics is een berekeningsmethode die stromingen, warmte, tweefase, turbu-lentie of stoftransport modelleert. Hierbij wordt eerst een bepaalde geometrie ingegeven die vervolgensverdeeld wordt in een groot aantal kleine elementen. Deze elementen worden doorgaans meshelementengenoemd. Vervolgens worden de stromingsvergelijkingen iteratief berekend voor elke meshelement zodatna convergentie een oplossing wordt gevonden.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 33
De rode bloedcellen zullen zich stapelen en een gepakt bed vormen op de wand. Het gepakt
bed zal blijven groeien terwijl er continu bloed wordt gepompt in de centrifuge. Wanneer
de centrifuge gevuld is, stroomt er langs de onderste uitgang geconcentreerd bloed naar
buiten. Het plasma verlaat langs de bovenste uitgang de centrifuge.
Blood
Plasma
centrifugalacceleration
Packed cell bed
Rotation axis
BloodPlasma
Packed cell bed
centrifugalacceleration
Figuur 2.7: De projectie van de drie dimensionale centrifuge
4.2 Theorie
4.2.1 Viscositeitsmodel van het bloed
In een centrifuge heersen grote hematocrietverschillen en grote snelheidsgradienten die niet
voorkomen in normale omstandigheden. Het bloed moet dus goed gemodelleerd worden
om nauwkeurige simulaties uit te kunnen voeren. Het is immers een vloeistof die een niet-
Newtoniaans gedrag vertoont. Het bloed wordt gemodelleerd als een suspensie van deeltjes
in een fluıdum. Er werden modellen opgesteld voor de viscositeit van suspensies maar deze
bevatten geen afhankelijkheid van de afschuifsnelheden. Het suspenderende fluıdum is het
bloedplasma en kan beschouwd worden als een onsamendrukbare Newtoniaanse vloeistof.
De viscositeit van het plasma is in functie van de temperatuur:
µplasma = µ37 exp[η(37− T )] (2.32)
met:
µplasma viscositeit van het plasma
µ37 viscositeit van het plasma bij 37 �(1.4 mPa s)
η temperatuurscoefficient (0.021 �−1)
T De temperatuur in �
Zoals de studies van Vand [2] hierboven aangetoond hebben is er volgend theoretisch model
opgesteld voor de viscositeit van een suspensie, hier uitgedrukt voor bloed:

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 34
µbloed = µplasma exp
(4.1 Hct
1.64−Hct
)(2.33)
Hoewel de theorie van Vand’s model kan worden toegepast op bloed, komen de resultaten
niet overeen met experimentele data [15]. Er is geen afhankelijkheid van de afschuifsnelheid
in formule (2.33), het model van Vand is dus Newtoniaans, wat de reden is van de slech-
te overeenstemming met de experimentele data. Uit deze data kan volgende empirische
correlatie worden opgesteld:
µbloed = µplasmaβαHct
(1−Hct)k(2.34)
met α en k constanten en β een factor afhankelijk van de afschuifsnelheid:
β = 1 +b
γn(2.35)
met b en n constanten en γ de afschuifsnelheid. Beide uitdrukkingen kunnen worden
gecombineerd tot een meer algemene vorm:
µbloed = µplasmaβ(1−Hct) exp
(4.1 Hct
1.64− Hct
)(2.36)
Deze vergelijking bevat de afschuifsnelheid, dit model stelt dus een niet-Newtoniaanse
vloeistof voor. Wanneer deze vergelijkingen vergeleken worden met de experimentele data
wordt vastgesteld dat vergelijking (2.36) beter overeenstemt voor een hematocriet lager dan
70 % en dat vergelijking (2.34) een betere gelijkenis toont voor een hematocriet hoger dan
70 %. In dit simulatiemodel worden beide vergelijkingen gebruikt voor hun respectievelijk
gebied. De constanten in de vergelijkingen (2.34) - (2.36) worden bepaald uit experimentele
data en zijn: b = 6.0s−1, α = 1, n = 0.75 en k = 0.5.
4.2.2 Stromingsdynamica
Om het stromingspatroon van het fluıdum in de centrifuge te bepalen wordt een Euleriaan-
se methode gebruikt. Dit wil zeggen dat de bewegingstoestand van de vloeistof berekend
wordt op een bepaalde plaats en tijd; x, y, z en t zijn onafhankelijke variabelen. Door het
oplossen van de Navier-Stokes vergelijkingen voor behoud van massa en momentum en re-
kening houdend met de eigenschappen van het bloed wordt het stromingspatroon bekomen.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 35
Deze berekening wordt op verschillende tijdstappen uitgerekend omdat de eigenschappen
van het bloed lokaal zullen veranderen in de tijd.
De beweging van de cellen wordt bepaald door de Langrangiaanse methode toe te passen.
Dit wil zeggen dat de bewegingen in ruimte en tijd worden gevolgd van elke cel terwijl
ze meestroomt met de vloeistof. De coordinaten zijn afhankelijke variabelen, zo zal de
sedimentatiesnelheid van een cel geschreven worden in een functionele vorm van de aard:
v = v(a, b, c, t) (2.37)
Met a, b, c de coordinaten van de initiele positie van de cel. Om die snelheid te berekenen
moeten voor elke cel de inwerkende krachten worden bepaald. Door de som van deze
krachten uit te rekenen kan de versnelling van een cel worden bepaald door middel van
Newton’s tweede wet van beweging.
4.2.3 Krachten op een bloedcel
De inwerkende krachten op een cel zijn: de centrifugale kracht vanwege de rotaties, de op-
stuwingskracht of Archimedeskracht, de wrijvingskracht, de Corioliskracht door verplaat-
singen in een draaiend referentiestelsel, de diffusiekracht, de Magnuskracht veroorzaakt
door de draaıng van een deeltje, de drukgradient, de Saffmankracht en de Bassetkracht.
In deze paragraaf worden bovenstaande krachten besproken.
De overheersende component van deze krachten is de som van de centrifugaalkracht en de
Archimedeskracht (zie linkerlid van (2.4)) of:
Vcel (ρcel − ρf ) ω2 rc (2.38)
Door de rotationele aard van het stelsel zal een deeltje dat beweegt in radiale richting een
bijkomende kracht ondervinden om de verandering in lineaire snelheid te compenseren; dit
is de Corioliskracht. Deze kracht wordt uitgedrukt door het vectorieel product:
~FCoriolis = 2(~ω × ~v) (2.39)
Een vlugge berekening van deze bijkomende kracht leert dat deze 3 grootte orden kleiner
is dan de centrifugaalkracht, bijgevolg is deze verwaarloosbaar.
Wanneer een cel roteert en zinkt in een fluıdum creeert het een draaiende stroming rond
zich. Langs de ene kant zal deze stroming in dezelfde zin zijn als de snelheid van de
zinkende cel, langs de andere kant zal deze in tegengestelde zin zijn. Zo onstaat er een

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 36
verschil in snelheden van het fluıdum rond de cel. Vanwege het Bernoulli effect onstaat
er dan ook een drukverschil en zal er een kracht worden uitgeoefend loodrecht op de
richting van de snelheid van de bloedcel. Dit is de Magnuskracht, deze kracht vind men
bijvoorbeeld veel terug te vinden bij balsporten. Wanneer deze kracht wordt uitgerekend bij
simulatiecondities blijkt dat deze kracht verwaarloosbaar is ten opzichte van de centrifugale
kracht.
Elk deeltje zal ook een effect ervaren afkomstig van de drukgradient. Deze wordt opgelegd
door de inlaat- en uitlaatcondities van de stroming. Maar aangezien de deeltjes een zeer
kleine oppervlakte hebben is de invloed van de andere krachten minstens 100 maal groter.
Dit effect is dus verwaarloosbaar.
De schuifspanningsgradienten in de vloeistof kunnen leiden tot een liftkracht, de Saffman
kracht. Deze kan worden verwaarloosd bij lage Reynolds getallen wat in deze simulatie het
geval is.
De Bassetkracht [13] onstaat wanneer een deeltje zich in een draaikolk bevindt. Dit kan
gebeuren bij dit simulatieproces. Deze kracht is afhankelijk van de geschiedenis van de
beweging van een deeltje, er bevindt zich een integraal over de tijd in de uitdrukking.
Studies van Michaelides en Vojir[14] tonen aan dat de invloed van deze kracht significant
is voor verhoudingen van vloeistof- tot deeltjesdichtheden tussen 0.002 en 0.7. In het
afereseproces is deze verhouding 0.93, de invloed van deze kracht kan dus verwaarloosd
worden.
De wrijvingskracht is dezelfde als de hier boven besproken vergelijking (2.3):
FD = 6πµr v (2.40)
Diffusie is het transport van deeltjes van een hogere concentratie naar een lagere concen-
tratie. Dit verband wordt gegeven door de wet van Fick:
~j = −D∇CN (2.41)
met:
~j de flux vector
D de diffusiecoefficient
CN de concentratie aan deeltjes van type N
Bij de bezinking van bloedcellen bij zwaartekracht is aangetoond door sartory[15] dat de
diffusiekracht verwaarloosbaar is in vergelijking met de bezinking van bloedcellen.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 37
De krachtenbalans is mits het weglaten van de verwaarloosbare krachten identiek aan
vergelijking (2.1). We bekomen voor de resulterende kracht op een deeltje:
Fr = mcel ·∂v
∂t= Vcel (ρcel − ρf ) ω2 rc + 6πµr v (2.42)
4.3 Berekeningsmethode
Zoals hierboven al vermeld, wordt er tijdens de simulatie op 2 berekeningsperspectieven
gesteund. In deze paragraaf worden deze samen met de geometrie en de simulatiecondities
besproken.
4.3.1 Euleriaans perspectief
Met behulp van een commerciele CFD-solver (CFD-ACE+ van ESI CFD, Huntsville, AL,
U.S.A.) worden de vergelijkingen voor het behoud van massa en momentum opgelost (Ver-
gelijking (2.43) en (6.1)). De vergelijking voor het behoud van momentum kan uitgedrukt
worden door een transportvergelijking van de hoeveelheid Φ:
∂(ρΦ)
∂t+∇ · (ρ~vΦ) = ∇ · (Γ∇Φ) + SΦ (2.43)
De eerste term is de tijdsafhankelijke term, de tweede beschrijft de convectie, de derde de
diffusie en de laatste term is de bronterm. De vergelijking voor behoud van massa wordt
genoteerd als:
∂ρ
∂t+∇ · (ρ~v) = 0 (2.44)
Het CFD-programma integreert deze vergelijkingen zodat het stromingsprofiel wordt be-
komen.
4.3.2 Lagrangiaans perspectief
De afgelegde weg van een bloedcel kan verkregen worden door de krachtenbalans (vergelij-
king (2.42)) te integreren over een bepaalde tijdstap. Dit wordt verricht door de numerieke
Euler methode toe te passen op deze gewone differentiaalvergelijking:

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 38
Φn+1 = Φn + f(tn+1, Φn+1)∆t (2.45)
Door de krachtenbalans op te lossen naar ∂v∂t
, wordt de functie van f(tn+1, Φn+1) bekomen.
Zo kan voor elke bloedcel de snelheid verkregen worden. De verplaatsing van elke bloedcel
wordt dan gevonden door de uitdrukking van de snelheid nogmaals te integreren.
Als van alle bloedcellen hun nieuwe posities berekend zijn, kunnen de lokale waarden van
het hematocriet worden aangepast. Dit wordt gedaan door in elk computationeel volume
element het aantal bloedcellen te tellen en zo het totale volume van de cellen in het volume
element te bepalen. Met behulp van de dichtheid en de oppervlakte van de doorsnede van
elk type bloedcel kan de dichtheid van de suspensie in elk meshelement worden berekend.
Zo wordt het volumetrisch gemiddelde van de bloedcellen en het plasma:
ρvolumeelement =
∑(ρiNiAi) + ρplasma · Aplasma
Avolumeelement
(2.46)
Eens de dichtheidsverdeling is bepaald kan de viscositeit worden berekend. Rekening hou-
dend met het hematocriet van elk volume element kan de viscositeit bepaald worden aan
de hand van formules (2.34) en (2.36). Nu de nieuwe dichtheden en de viscositeiten van
elk meshelement gekend zijn kan het CFD-programma het nieuwe stromingspatroon be-
rekenen. De software geeft enkel de snelheid van de stroming in het centrum van het
meshelement weer, maar de cellen kunnen zich op verschillende plaatsen in dit meshele-
ment bevinden. De snelheid op de plaats van een bloedcel wordt bekomen door interpolatie
van de snelheid van de naburige mesh elementen:
~vp =
∑(1dj
)2
~vj∑(1dj
)2 (2.47)
~vp de snelheid op de plaats van de bloedcel
~vj de snelheid van het naburig mesh element j
dj de afstand tussen de bloedcel en het centrum van het naburig mesh element j
4.3.3 Geometrie en mesh
De resultaten van de simulatie worden gecontroleerd met de door Brown [16] uitgevoerde
experimenten. Het is daarom noodzakelijk dat de simulatie zich bij dezelfde omstandighe-
den voordoet. Zo wordt dezelfde geometrie gebruikt als in het onderzoek van Brown. De

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 39
experimenten werden uitgevoerd in een centrifuge met een dikte van 4mm en twee verschil-
lende lengtes; 13cm en 43cm (zie figuur 2.7). De simulaties werden uitgevoerd bij lengtes
van 13cm, 16cm en 19cm. De hoogte van de centrifuge bedraagt 5cm en de binnenwand
bevindt zich op 6.82cm van de rotatie as.
Er werd gekozen voor een driehoekige ongestructureerde mesh met in de kortste configuratie
van de centrifuge 54700 mesh elementen. De keuze van een ongestructureerde mesh ver-
groot de rekensnelheid van het probleem wanneer er samen met een subroutine, geschreven
voor de Lagrangiaanse berekening van de beweging van de bloedcellen, gerekend wordt.
Een gridafhankelijkheidsstudie werd uitgevoerd met een fijne en een grovere mesh. De
grootte van een mesh element in de fijne mesh bedraagt tussen 1nm en 5nm, bij de grove
mesh ligt de grootte tussen 5nm en 30nm. Bloedcellen worden samen gegroepeerd per
tien cellen tot een groep, met als gevolg dat de oppervlakte van de groep voor witte
bloedcellen groter kunnen worden dan sommige mesh elementen in de fijne structuur. Dit
kan fouten opleveren maar na een berekening blijkt er maar een afwijking van 1% te zijn op
de sedimentatiesnelheid. Aangezien beide meshen dezelfde resultaten gaven, kon er voor
de grovere mesh worden geopteerd zodat de nodige rekenkracht minimaal blijft.
4.3.4 Simulatiecondities
Brown [16] voerde de experimenten uit met een rotatiesnelheid die varieerde van 1425
tot 3000 tpm, waarbij de meeste experimenten werden uitgevoerd met de kortere kamer
van 13cm en bij 3000 tpm. Het debiet aan bloed varieerde van 15 tot 130 mL/min, het
hematocriet van het instromende bloed varieerde van 26 tot 52%. Deze gegevens werden
gebruikt om de simulatiecondities te bepalen.
Voor de viscositeit van het bloedplasma werd een waarde van 1.65 mPa.s genomen. De
tijdstap werd gekozen op 0.1 s zodat de sedimentatie nauwkeurig gesimuleerd kon worden.
Deeltjes worden gegroepeerd per tien zodat de berekeningen sneller verlopen. De simula-
ties werden uitgevoerd bij 3000 tpm en bij een volumetrisch debiet van 30 ml/min. Drie
simulaties werden uitgevoerd met de kortste configuratie van de centrifuge, telkens met
een inlaathematocriet van 30, 35 en 40%. Twee andere simulaties werden uitgevoerd bij
de andere configuraties, 16cm en 19cm, bij een inlaathematocriet van 30%. De fysische
eigenschappen van de bloedcellen tijdens de simulaties worden weergegeven in tabel 2.2.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 40
Tabel 2.2: Fysische eigenschappen van de bloedcellen tijdens de simulaties
ρ (g/cm3) r (cm×104)
RBC 1.100 4
WBC 1.075 6
Plaatjes 1.040 1.5
Plasma 1.026 —
4.4 Resultaten
4.4.1 Concentratie aan rode bloedcellen
De rode bloedcellen hebben de grootste dichtheid, daarom zullen de cellen zich snel afzet-
ten aan de buitenste wand van de centrifuge. Daar vormen ze een gepakt bed van rode
bloedcellen met een hematocriet van boven de 80%. In deze laag zijn geen andere cellen
aanwezig. Voor de quasi stationaire fase wordt het gepakt bed voor de drie inlaathemato-
crieten weergegeven op figuur 2.8. Het gepakt bed wordt snel opgebouwd en blijft groeien
totdat het voor de rode bloedcellen niet meer mogelijk is om te bezinken. Dit gebeurt bij
een inlaathematocriet van 30% na 8 cm, bij 35% is er 9 cm nodig en bij 40% is er meer
dan 10 cm nodig om al de cellen te laten bezinken.
Figuur 2.8: Concentratie van de rode bloedcellen bij quasi stationaire toestand

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 41
Een dunne overgangslaag met een lagere concentratie aan rode bloedcellen bevindt zich
op het gepakt bed. In deze laag varieert het hematocriet van 10% tot 80%. Een hoger
inlaathematocriet komt overeen met een dikkere overgangslaag. Bij een inlaathematocriet
van 0.30 is deze laag bijna afwezig. Wanneer de simulaties worden uitgevoerd bij de langere
centrifuges zal de verblijftijd van een cel stijgen. Dit impliceert dat het gepakt bed langer
onderhevig is aan de centrifugale kracht zodat de overgangslaag eveneens zal verdwijnen
bij een inlaathematocriet van 35% en 40%.
4.4.2 Concentratie aan witte bloedcellen
Wanneer de witte bloedcellen de centrifuge binnen komen gaan ze zich hierin verspreiden
(zie figuur 2.9). Vervolgens gaan ze zich gaan ordenen boven op het gepakt bed en onder
de overgangslaag. In deze zone is het hematocriet ongeveer 80% wat op deze plaats zorgt
voor een evenwicht tussen de centrifugaalkracht en de opstuwingskracht. Wanneer het
inlaathematocriet stijgt zullen de zinkende witte bloedcellen meer weerstand ondervinden
van de dikkere overgangslaag waardoor ze meer tijd nodig zullen hebben om te bezinken.
Een grotere lengte van de centrifuge leidt tot een dunnere laag van witte bloedcellen op
het gepakt bed. Dit komt doordat de cellen meer tijd hebben om te bezinken. Het gepakt
bed zal bij langere centrifuges meer worden samengedrukt zodat de laag witte bloedcellen
de centrifuge verlaat langs de onderste uitgang.
Figuur 2.9: Concentratie van de witte bloedcellen bij quasi stationaire toestand

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 42
4.4.3 Concentratie aan bloedplaatjes
Bloedplaatjes zijn de kleinste en de lichtste cellen in het bloed. Hun stromingsgedrag zoals
getoond op figuur 2.10 toont aan dat de plaatjes hoofdzakelijk worden meegevoerd door
het bloedplasma. Nadat ze zich verspreid hebben over de centrifuge zullen de plaatjes die
zich aan de buitenwand bevinden, verdrongen worden door de rode bloedcellen. In het
midden is er een hogere concentratie aan plaatjes doordat naarmate de verblijftijd vordert
meer plaatjes uit het gepakt bed worden geperst en dat er meer plaatjes zullen bezinken
vanuit het plasma.
Figuur 2.10: Concentratie van de bloedplaatjes bij quasi stationaire toestand
4.4.4 Scheidingsefficientie
Om de prestatie van een centrifuge te onderzoeken wordt de scheidingsefficientie van het
bloedplasma, bloedplaatjes en witte bloedcellen geevalueerd. De scheidingsefficientie wordt
gedefinieerd als een graad van eliminatie van een bepaalde cel uit de instroom:
effscheiding = 1− cuit
cin
(2.48)
met:
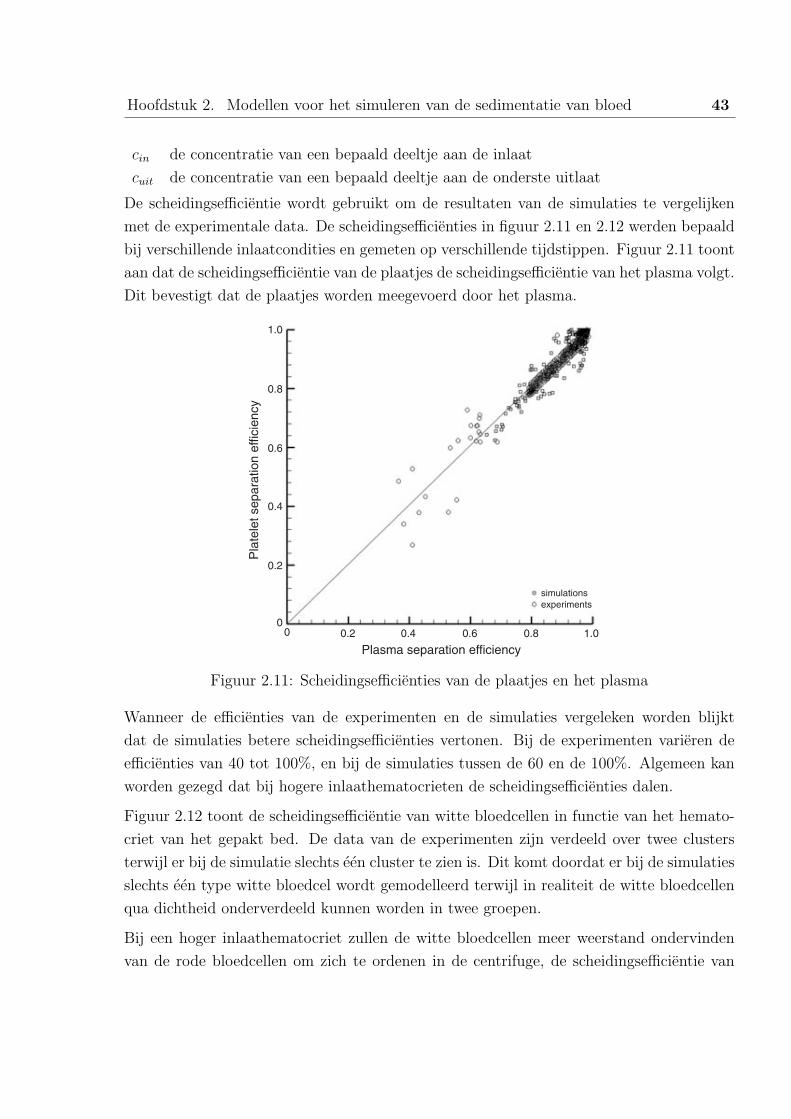
Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 43
cin de concentratie van een bepaald deeltje aan de inlaat
cuit de concentratie van een bepaald deeltje aan de onderste uitlaat
De scheidingsefficientie wordt gebruikt om de resultaten van de simulaties te vergelijken
met de experimentale data. De scheidingsefficienties in figuur 2.11 en 2.12 werden bepaald
bij verschillende inlaatcondities en gemeten op verschillende tijdstippen. Figuur 2.11 toont
aan dat de scheidingsefficientie van de plaatjes de scheidingsefficientie van het plasma volgt.
Dit bevestigt dat de plaatjes worden meegevoerd door het plasma.
0.8
1.0
Pla
tele
t se
pa
ratio
n e
ffic
ien
cy
Plasma separation efficiency
0.6
0.4
0.2
0.2 0.4 0.6 0.8
simulations
experiments
1.00
0
Figuur 2.11: Scheidingsefficienties van de plaatjes en het plasma
Wanneer de efficienties van de experimenten en de simulaties vergeleken worden blijkt
dat de simulaties betere scheidingsefficienties vertonen. Bij de experimenten varieren de
efficienties van 40 tot 100%, en bij de simulaties tussen de 60 en de 100%. Algemeen kan
worden gezegd dat bij hogere inlaathematocrieten de scheidingsefficienties dalen.
Figuur 2.12 toont de scheidingsefficientie van witte bloedcellen in functie van het hemato-
criet van het gepakt bed. De data van de experimenten zijn verdeeld over twee clusters
terwijl er bij de simulatie slechts een cluster te zien is. Dit komt doordat er bij de simulaties
slechts een type witte bloedcel wordt gemodelleerd terwijl in realiteit de witte bloedcellen
qua dichtheid onderverdeeld kunnen worden in twee groepen.
Bij een hoger inlaathematocriet zullen de witte bloedcellen meer weerstand ondervinden
van de rode bloedcellen om zich te ordenen in de centrifuge, de scheidingsefficientie van
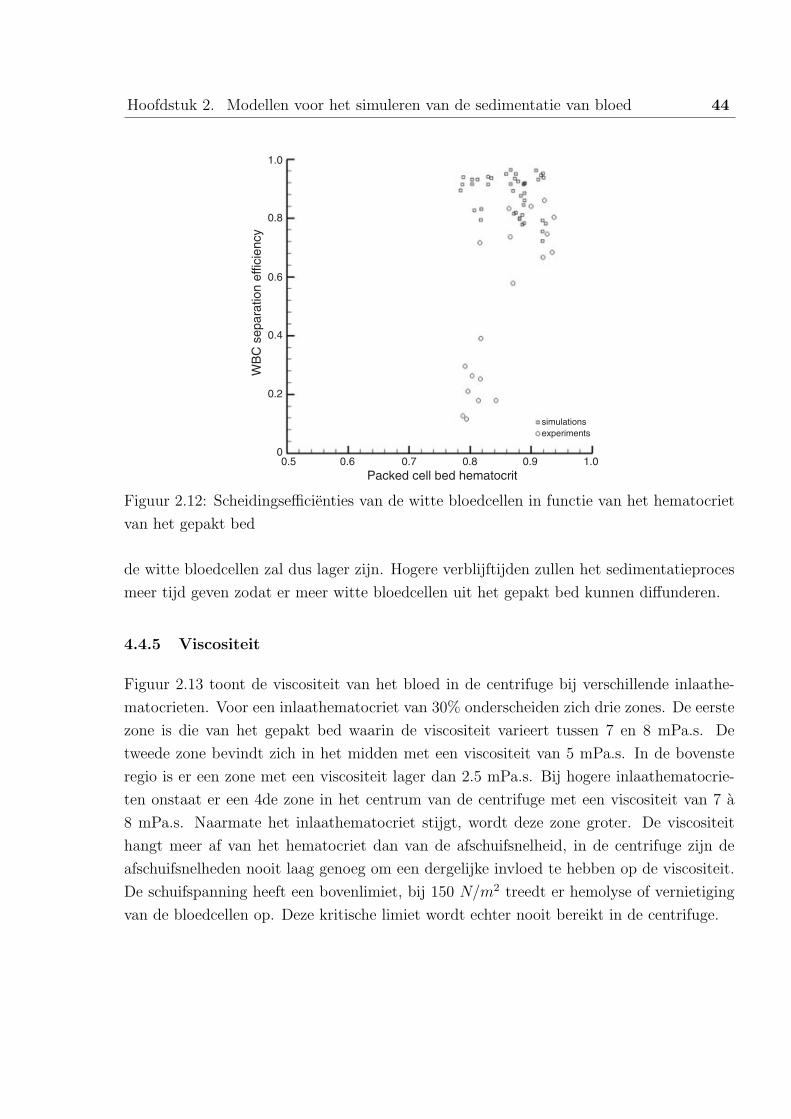
Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 44
1.0
WB
C s
epara
tion e
ffic
iency
Packed cell bed hematocrit
0.8
0.6
0.4
0.5 0.6 0.7 0.8 0.9 1.0
simulations
experiments
0.2
0
Figuur 2.12: Scheidingsefficienties van de witte bloedcellen in functie van het hematocriet
van het gepakt bed
de witte bloedcellen zal dus lager zijn. Hogere verblijftijden zullen het sedimentatieproces
meer tijd geven zodat er meer witte bloedcellen uit het gepakt bed kunnen diffunderen.
4.4.5 Viscositeit
Figuur 2.13 toont de viscositeit van het bloed in de centrifuge bij verschillende inlaathe-
matocrieten. Voor een inlaathematocriet van 30% onderscheiden zich drie zones. De eerste
zone is die van het gepakt bed waarin de viscositeit varieert tussen 7 en 8 mPa.s. De
tweede zone bevindt zich in het midden met een viscositeit van 5 mPa.s. In de bovenste
regio is er een zone met een viscositeit lager dan 2.5 mPa.s. Bij hogere inlaathematocrie-
ten onstaat er een 4de zone in het centrum van de centrifuge met een viscositeit van 7 a
8 mPa.s. Naarmate het inlaathematocriet stijgt, wordt deze zone groter. De viscositeit
hangt meer af van het hematocriet dan van de afschuifsnelheid, in de centrifuge zijn de
afschuifsnelheden nooit laag genoeg om een dergelijke invloed te hebben op de viscositeit.
De schuifspanning heeft een bovenlimiet, bij 150 N/m2 treedt er hemolyse of vernietiging
van de bloedcellen op. Deze kritische limiet wordt echter nooit bereikt in de centrifuge.

Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 45
Figuur 2.13: Viscositeit in de centrifuge
4.4.6 Rekenkracht
De simulaties werden uitgevoerd met een DEC-Alpha cluster bestaande uit acht 833MKz
en zes 667MHz 64-bit CPU’s met een totaal van 20 Gb aan RAM. Elke simulatie werd
sequentieel op een van de processors uitgevoerd. De snelste simulatie duurde 3 weken.
4.5 Besluit
Het sedimentatiemodel hier beschreven verschilt met de theorie van de sedimentatiesnel-
heden beschreven in de eerste paragrafen. Hier worden stromingen ook bestudeerd met als
gevolg dat het model moet worden aangepast. Bloed is een niet-Newtoniaanse vloeistof
zodat het viscositeitsmodel van Vand moet worden gewijzigd. Dit gebeurt met behulp van
experimentele data.
De dichtheid in een computationele cel wordt bepaald door vergelijking (2.46), hier wordt
gerekend met de oppervlakte van de doorsnede van een bloedcel. In drie dimensies kunnen
de cellen evenwel verder bezinken dan in twee dimensies kan worden voorgesteld. In dit
model wordt geen rekening gehouden met de rouleaux vorming van de rode bloedcellen,
ze worden beschouwd als individuele cellen. De botsingen tussen de deeltjes werden hier
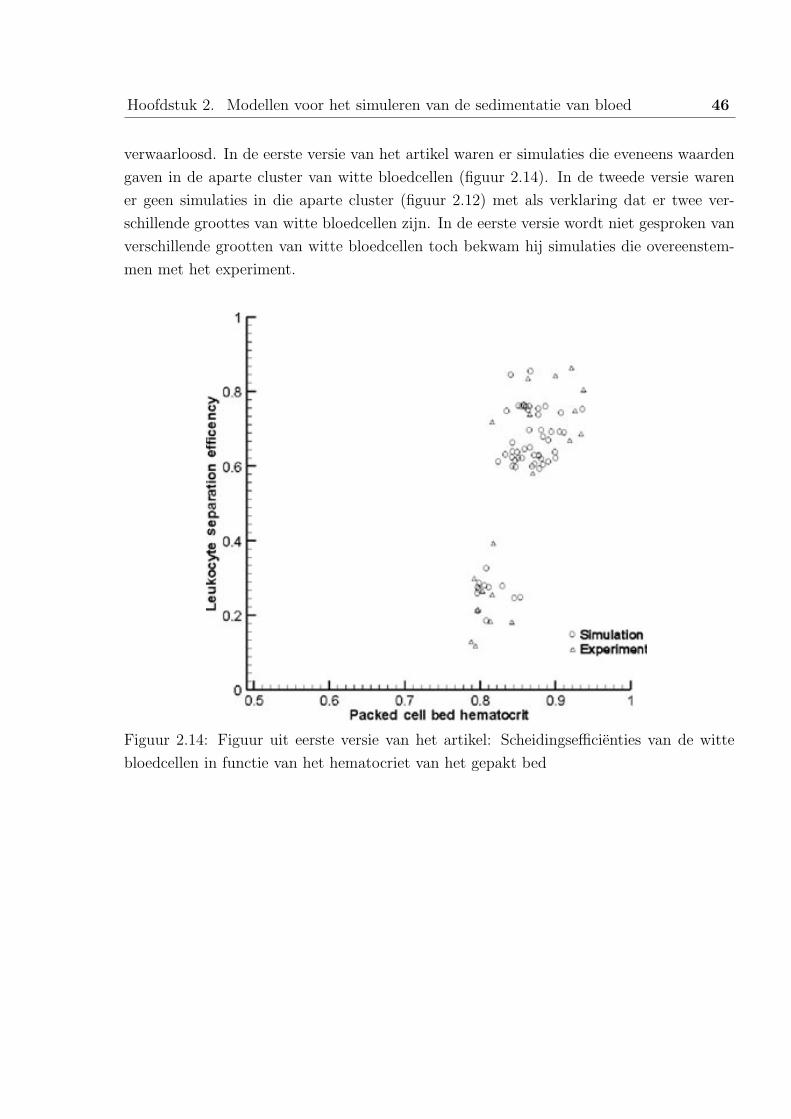
Hoofdstuk 2. Modellen voor het simuleren van de sedimentatie van bloed 46
verwaarloosd. In de eerste versie van het artikel waren er simulaties die eveneens waarden
gaven in de aparte cluster van witte bloedcellen (figuur 2.14). In de tweede versie waren
er geen simulaties in die aparte cluster (figuur 2.12) met als verklaring dat er twee ver-
schillende groottes van witte bloedcellen zijn. In de eerste versie wordt niet gesproken van
verschillende grootten van witte bloedcellen toch bekwam hij simulaties die overeenstem-
men met het experiment.
Figuur 2.14: Figuur uit eerste versie van het artikel: Scheidingsefficienties van de witte
bloedcellen in functie van het hematocriet van het gepakt bed

Hoofdstuk 3
Experimentele studie van de
sedimentatie van runderbloed in een
centrifuge
Deze studie onderzoekt het sedimentatiegedrag van runderbloed in een Dideco centrifuge.
In hoofdstuk 4 wordt een analoog experiment uitgevoerd met humaanbloed zodat de resul-
taten van beide proeven kunnen worden vergeleken. De data kunnen ook gebruikt worden
om een validatie uit te voeren van het numeriek model beschreven in hoofstuk 6. In dit
hoofdstuk wordt in detail besproken hoe het experiment met runderbloed werd uitgevoerd
en hoe de resultaten geınterpreteerd moeten worden.
1 De metingen
1.1 Voorbereiding van het bloed
Het runderbloed lijkt qua anatomie sterk op het menselijk bloed. In tabel 3.1 worden de
verschillende waarden gegeven.
Tabel 3.1: Verschil tussen humaan en runderbloed
Bloed HCT RBC [×106/µl] Diameter RBC [µm] WBC (×103/µl) Hb [g/dl]
Humaan 36-51 4.8-5.4 7-8 4-11 14-16
Rund 24-46 5-10 4-8 4-12 8-15
47

Hoofdstuk 3. Experimentele studie met runderbloed 48
Aan de hand van tabel 3.1 kan besloten worden dat het runderbloed meer rode bloedcellen
bevat maar dat deze kleiner kunnen zijn dan bij het humaanbloed. Om aan runderbloed
te geraken moest er naar het slachthuis worden gegaan (slachthuis van Zele). Omdat er
nog steeds een klein risico is op BSE, is het verplicht het bloed te vernietigen na gebruik.
De nodige documenten dienen hiervoor te worden ondertekend door de veearts van het
slachthuis. Nadat de koe of stier, een kogel door zijn schedel gekregen heeft wordt het
dier omgekeerd opgehangen en de halsslagader wordt opengesneden. De koe bloedt nu leeg
en het bloed kan opgevangen worden in een emmer. De binnenwand van de emmer werd
bevochtigd met heparine zodat het bloed niet begint te stollen. Het eerste dier was een
stier en gaf een 8-tal liter bloed, het volgende exemplaar, een koe, gaf nog eens 2 liter. Het
bloed wordt dan getransporteerd naar het labo van experimentele heelkunde in het UZ
Gent. Bloed is een levend weefsel en zal blijven zuurstof opnemen en kooldioxide afstaan.
Het bloed kan verzuren door de opstapeling van kooldioxide in het bloed. Een lage pH
waarde vernietigt het bloed en maakt het onbruikbaar om experimenten uit te voeren. Het
is belangrijk dat het bloed in contact blijft met lucht en dat het goed geroerd wordt zodat
het stoftransport van kooldioxide wordt bevorderd. Als het bloed in het labo is, wordt het
gefilterd om stukjes weefsel te verwijderen. Het bloed is nu klaar om gedilueerd te worden
met een fysiologische oplossing om op die manier andere hematocrieten te bereiken.
1.2 De opstelling
In dit experiment wordt het bloed gecentrifugeerd met een Dideco bowl waarvan de inhoud
125ml is. De bowl werd geplaatst in de Electa. Op figuur 3.1 is de opstelling afgebeeld. Er
zijn drie stromen; de instroom van het bloed, de uitstroom van het geconcentreerd bloed
en de uitstroom van het plasma.
Tijdens een experiment kunnen we volgende stappen onderscheiden:
1. De Electa doet een self-test om na te gaan of alles functioneert. Het toestel meldt
dat er geen wasvloeistof aanwezig is, maar deze opmerking wordt genegeerd omdat
in dit experiment de cellen niet worden gewassen.
2. De pomp start en het bloed wordt aangezogen met een bepaalde vulsnelheid in de
draaiende centrifuge. Er onstaat een scheiding van de componenten, het plasma
bevindt zich boven de rode bloedcellen. Eens de centrifuge gevuld is zal er plasma
langs de bovenzijde uitstromen. Deze stap is de vulfase.

Hoofdstuk 3. Experimentele studie met runderbloed 49
Figuur 3.1: Schematische opstelling van het experiment
3. Wanneer de detector de rode bloedcellen detecteert bovenaan in de centrifuge, wordt
de centrifuge gestopt en draait de pomp in de andere richting zodat de centrifuge
wordt leeggezogen. Dit is de uitstroomfase.
1.3 Meetcondities
Om de theoretische modellen goed te kunnen controleren is het belangrijk om veel parame-
ters die het sedimentatieproces beınvloeden aan te kunnen passen tijdens het experiment.
Er zal bijgevolg gewerkt worden met vier verschillende bloedconcentraties. De bedoeling is
om bloed te centrifugeren met een hematocriet van ongeveer 10, 20, 30 en 40%. Er wordt
een reeks metingen gedaan bij 5600 tpm en bij 2500 tpm. Bij de lage rotatiesnelheid blij-
ken de condities echter onvoldoende om bloed met een hoog hematocriet te scheiden. Een
laatste aanpasbare parameter is de vulsnelheid, die belangrijk is voor een goede scheiding.
De vulsnelheid bepaalt immers de verblijftijd in de centrifuge. Bij rotaties van 5600 tpm
wordt er gekozen voor een vulsnelheid van 100, 200 en 300 ml/min. Bij 2500 tpm wordt
gewerkt met een vulsnelheid van 100, 150 en 200 ml/min. Het heeft geen nut om de traag
draaiende centrifuge te bedrijven met de hoge vulsnelheid van 300 ml/min. Er zijn drie
parameters, zoals aangegeven in Tabel 3.2, het experiment wordt 24 keer uitgevoerd.

Hoofdstuk 3. Experimentele studie met runderbloed 50
Tabel 3.2: Parameters
Parameter
Rotatiesnelheid (tpm) 2 500, 5 600
Vulsnelheid (ml/min) 100, 200, 300*
Inlaat hematocriet (%) ≈ 10, 20, 30, 40 *
* Bij 2 500 tpm is de vulsnelheid 150 in plaats van 300 en wordt er niet met een inlaathematocriet van 40
gewerkt.
1.4 Metingen
Telkens wanneer er een nieuwe oplossing wordt gemaakt of wanneer het bloed uit de cen-
trifuge wordt gepompt, wordt het hemoglobine gemeten met een HemoCue (HemoCue AB,
Zweden). Dit apparaat bepaalt het hemoglobine van humaanbloed binnen het bereik van
0 tot 16 g/l. Het meetprincipe steunt op het absorptievermogen van azidemethemoglobine.
Dit complex wordt gevormd in de HemoCue door de reactie van natriumdeoxycholaat en
de erytrocyten. Het absorptievermogen wordt gemeten door twee golflengtes (570 en 880
nm) om voor mogelijke troebeling van diverse oorsprong te kunnen compenseren.
Omdat er gewerkt wordt met runderbloed in hogere concentraties is het nodig stalen te
nemen en deze te laten controleren door een gespecialiseerd labo. Dankzij deze dubbele
meting kunnen er eventuele afwijkingen van de HemoCue worden vastgesteld. In het labo
meet de Coulter Counter het hematocriet. Dit apparaat registreert het aantal deeltjes
in een bepaald volume. Het hart van het apparaat bestaat uit een kleine ruimte, met
daarin een membraan waarin zich een kleine porie bevindt. Over het membraan heerst een
potentiaalverschil dat continu gemeten wordt, de waarde is te zien op een beeldscherm.
Men laat vervolgens een monster passeren, dit monster bevat de te tellen cellen verdund
in een geleidende vloeistof. De deeltjes of cellen passeren een voor een het membraan door
de porie. Bevindt zich een deeltje in de porie dan wordt het potentiaalverschil heel even
verhoogd; er is een pulsje te zien op het beeldscherm, daarnaast telt het apparaat ook de
hoeveelheid pulsjes. De oorzaak van een pulsje is het feit dat een cel een hogere weerstand
heeft dan de vloeistof waarin het zich bevindt. Een hogere weerstand leidt tot een hoger
potentiaalverschil. Zo is er zelfs een verband tussen de grootte van het deeltje en de hoogte
van het pulsje. Daardoor is het mogelijk door het instellen van een benedengrens en een
bovengrens om alleen de cellen van een bepaalde grootte te tellen. Het is een zeer snelle,
nauwkeurige methode.

Hoofdstuk 3. Experimentele studie met runderbloed 51
Buiten het hematocriet worden ook de volumes van de drie stromen gemeten. Ze worden
afgelezen door een maatglas maar ook door de Electa. Door de lange leidingen kunnen
de volumes afgelezen van het maatglas afwijken van deze geregistreerd door het toestel.
De viscositeit van het bloed hangt af van de temperatuur. Het vers bloed koelt af tot
kamertemperatuur zodat de temperatuur ook dient gemeten te worden. Verder wordt van
de leidingen hun lengte en volume gemeten.
Tabel 3.3: Metingen
Metingen
Volume verwerkt bloed
Volume geconcentreerd bloed*
Volume plasma
Hematocriet instromend bloed**
Hematocriet geconcentreerd bloed**
Temperatuur
*Wordt gemeten door de machine en met behulp van een maatglas
**Wordt gemeten met de HemoCue en met de Coulter Counter
2 De resultaten
2.1 Tabel
Tabel 3.4 toont de meetresultaten. Meting 4 werd slecht uitgevoerd. Meting 13 en 14 zijn
onbruikbaar omdat er zich geen scheiding voordeed.

Hoofdstuk 3. Experimentele studie met runderbloed 52
Tabel 3.4: Meetresultaten
Num
mer
met
ing
Toe
rent
al[t
/min
]
Vul
snel
heid
[ml/
min
]
Vol
ume
in[m
l](E
lect
a)
Hem
oglo
bine
in[g
/l](H
emoc
ue)
Hem
oglo
bine
in[g
/l](C
oult
er)
Hem
atoc
riet
in%
(Cou
lter
)
Vol
.pl
asm
aui
t[m
l](g
emet
en)
Vol
.rb
cui
t[m
l](E
lect
a)
Vol
.rb
cui
t[m
l]ge
met
en
Hem
oglo
bine
uit
[g/l
](H
emoc
ue)
Hem
oglo
bine
uit
[g/l
]C
oult
er
Hem
atoc
riet
uit
%(C
oult
er)
Tem
pera
tuur
[�]
Ver
blijf
tijd
[s]
123456
4bis789101112
1314151617181920212223
5600560056005600560056005600560056005600560056005600
25002500250025002500250025002500250025002500
100200300100200300100100200300100200300
100200100200150100200150100200150
218195174307279255281381343305688653597
113115240222297180142163132123132
14.814.7
10.6
10.6
6.5
3.63.3
11.7
3.5
6.8
9.4
11.7
8.54
8.64
6.86
3.853.47
3.62
7.01
9.72
33.4
24.3
24.3
19.2
10.69.5
10
19.6
27.2
826646210145122170275205167597500420
00
146891586530331701
134142131132133133133136134134143142134
126110124124138136123122122118122
134120120121134133135136136134137136134
126120134135135132134133132127131
15.922
20.3hhhhhh19.522.217.816.414.616.514.814
12.211.79.26.37.910.18.69.111.110.201.1
19.418.316.624.217.615.918.317.916.615.016.415.014.1
9.486.497.9110.28.849.3411.110.310.4
55.151.646.968.149.344.651.850.946.942.746.942.340.2
26.818.322.429.125.126.531.229.229.2
30
26
25
23
22.5
22.5
130.858.534.8184.283.751
168.6228.6102.961
412.8195.9119.4
67.834.514466.6118.810842.665.279.236.952.8
2.2 Bespreking van de HemoCue resultaten
Het tijdens de experimenten gebruikte meettoestel, de HemoCue, toont een afwijking ten
opzichte van de nauwkeurige laboresultaten. Het verschil tussen beide wordt weergegeven
op figuur 3.2. De HemoCue heeft voor veel waarden slechts een kleine afwijking, maar bij
8 metingen is er een sterke afwijking van ongeveer 25%. Eigenaardig is dat deze 8 zwaar
foutieve metingen zich voordoen bij de eerste 10 metingen. Er zijn verschillende verkla-
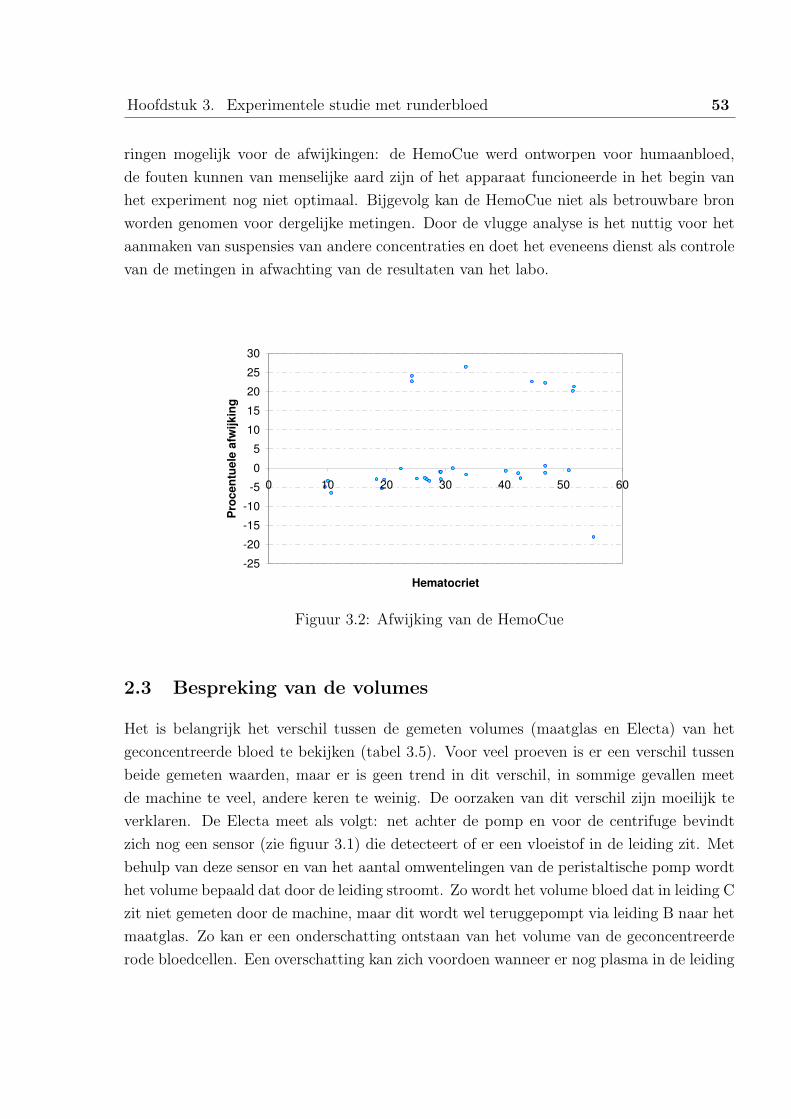
Hoofdstuk 3. Experimentele studie met runderbloed 53
ringen mogelijk voor de afwijkingen: de HemoCue werd ontworpen voor humaanbloed,
de fouten kunnen van menselijke aard zijn of het apparaat funcioneerde in het begin van
het experiment nog niet optimaal. Bijgevolg kan de HemoCue niet als betrouwbare bron
worden genomen voor dergelijke metingen. Door de vlugge analyse is het nuttig voor het
aanmaken van suspensies van andere concentraties en doet het eveneens dienst als controle
van de metingen in afwachting van de resultaten van het labo.
-25
-20
-15
-10
-5
0
5
10
15
20
25
30
0 10 20 30 40 50 60
Hematocriet
Pro
ce
ntu
ele
afw
ijk
ing
Figuur 3.2: Afwijking van de HemoCue
2.3 Bespreking van de volumes
Het is belangrijk het verschil tussen de gemeten volumes (maatglas en Electa) van het
geconcentreerde bloed te bekijken (tabel 3.5). Voor veel proeven is er een verschil tussen
beide gemeten waarden, maar er is geen trend in dit verschil, in sommige gevallen meet
de machine te veel, andere keren te weinig. De oorzaken van dit verschil zijn moeilijk te
verklaren. De Electa meet als volgt: net achter de pomp en voor de centrifuge bevindt
zich nog een sensor (zie figuur 3.1) die detecteert of er een vloeistof in de leiding zit. Met
behulp van deze sensor en van het aantal omwentelingen van de peristaltische pomp wordt
het volume bepaald dat door de leiding stroomt. Zo wordt het volume bloed dat in leiding C
zit niet gemeten door de machine, maar dit wordt wel teruggepompt via leiding B naar het
maatglas. Zo kan er een onderschatting ontstaan van het volume van de geconcentreerde
rode bloedcellen. Een overschatting kan zich voordoen wanneer er nog plasma in de leiding

Hoofdstuk 3. Experimentele studie met runderbloed 54
D zit dat niet in het maatglas werd gepompt. Het is moeilijk om te vertrouwen op een
van de resultaten, aangezien er altijd wel nog wat resterend bloed in de leiding of in het
pompsegment zal overblijven.
Tabel 3.5: Meetresultaten
Num
mer
met
ing
Toe
rent
al[t
/min
]
Vul
snel
heid
[ml/
min
]
Vol
ume
in[m
l]
Vol
.pl
asm
aui
t[m
l]
Vol
.rb
c[m
l](E
lect
a)
Vol
.rb
c[m
l]ge
met
en
Ver
schi
lrb
cV
elec−
Vge
m
Bal
ans
Ele
cta
Vin−
Vpl−
Vrb
c
Bal
ans
gem
eten
Vin−
Vpl−
Vrb
c
12356
4bis78910111215
1617181920212223
5600560056005600560056005600560056005600560056002500
25002500250025002500250025002500
100200300200300100100200300100200300100
200150100200150100200150
218195174279255281381343305688653597240
222297180142163132123132
826646145122170275205167597500420146
891586530331701
134142131133133133136134134143142134124
124138136123122122118122
134120120134133135136136134137136134134
135135132134133132127131
02211−10−20−20660−10
−1134−11−11−10−9−9
2−13−310−22−3044−521143−30
91−21−118−759
29800−24−3024−461743−40
−24−17−22−3−17−40
Een groot probleem is dat tijdens de uitstroomfase niet behandeld bloed in het pompseg-
ment zal gemengd worden met geconcentreerd bloed. Eveneens wordt er opgemerkt dat
tijdens de uitstroomfase het plasma dat zich nog in de leiding bevond ook zal worden te-
ruggezogen met het geconcentreerd bloed. Deze verschijnselen dienen in acht te worden
genomen bij de validatie van de modellen. Zo zal het hematocriet van de uitstroom la-
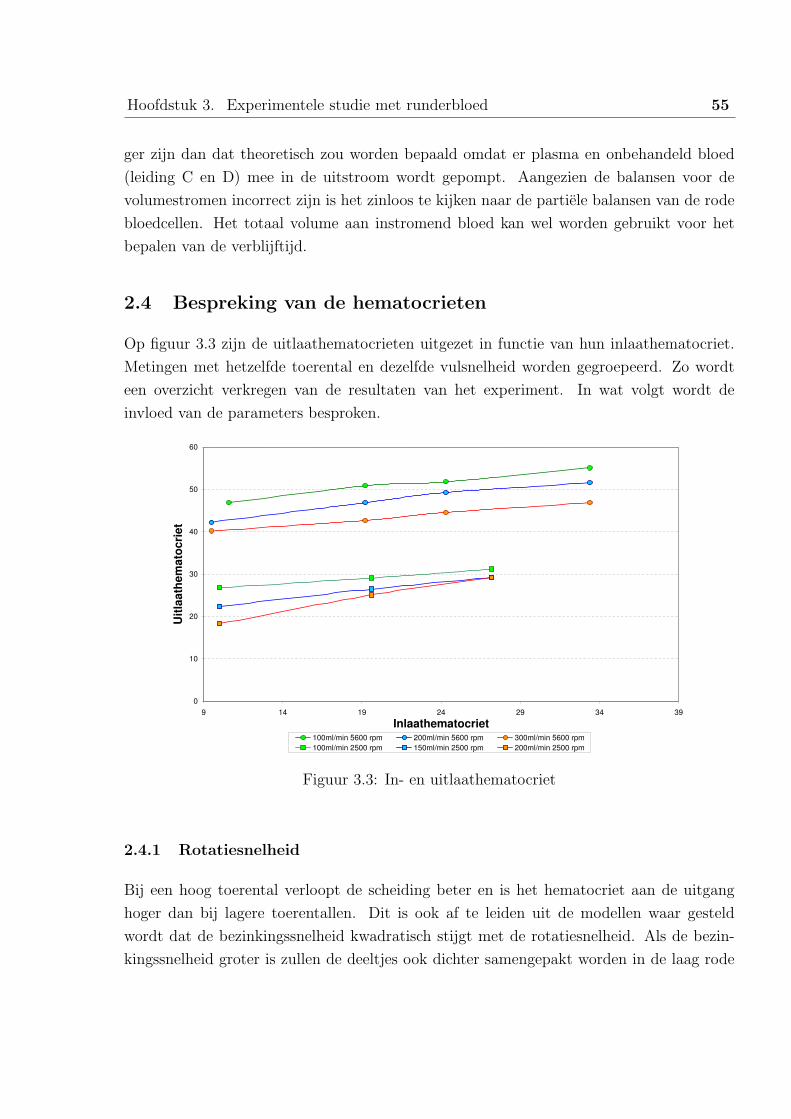
Hoofdstuk 3. Experimentele studie met runderbloed 55
ger zijn dan dat theoretisch zou worden bepaald omdat er plasma en onbehandeld bloed
(leiding C en D) mee in de uitstroom wordt gepompt. Aangezien de balansen voor de
volumestromen incorrect zijn is het zinloos te kijken naar de partiele balansen van de rode
bloedcellen. Het totaal volume aan instromend bloed kan wel worden gebruikt voor het
bepalen van de verblijftijd.
2.4 Bespreking van de hematocrieten
Op figuur 3.3 zijn de uitlaathematocrieten uitgezet in functie van hun inlaathematocriet.
Metingen met hetzelfde toerental en dezelfde vulsnelheid worden gegroepeerd. Zo wordt
een overzicht verkregen van de resultaten van het experiment. In wat volgt wordt de
invloed van de parameters besproken.
0
10
20
30
40
50
60
9 14 19 24 29 34 39
Inlaathematocriet
Uit
laa
the
ma
toc
rie
t
100ml/min 5600 rpm 200ml/min 5600 rpm 300ml/min 5600 rpm
100ml/min 2500 rpm 150ml/min 2500 rpm 200ml/min 2500 rpm
Figuur 3.3: In- en uitlaathematocriet
2.4.1 Rotatiesnelheid
Bij een hoog toerental verloopt de scheiding beter en is het hematocriet aan de uitgang
hoger dan bij lagere toerentallen. Dit is ook af te leiden uit de modellen waar gesteld
wordt dat de bezinkingssnelheid kwadratisch stijgt met de rotatiesnelheid. Als de bezin-
kingssnelheid groter is zullen de deeltjes ook dichter samengepakt worden in de laag rode
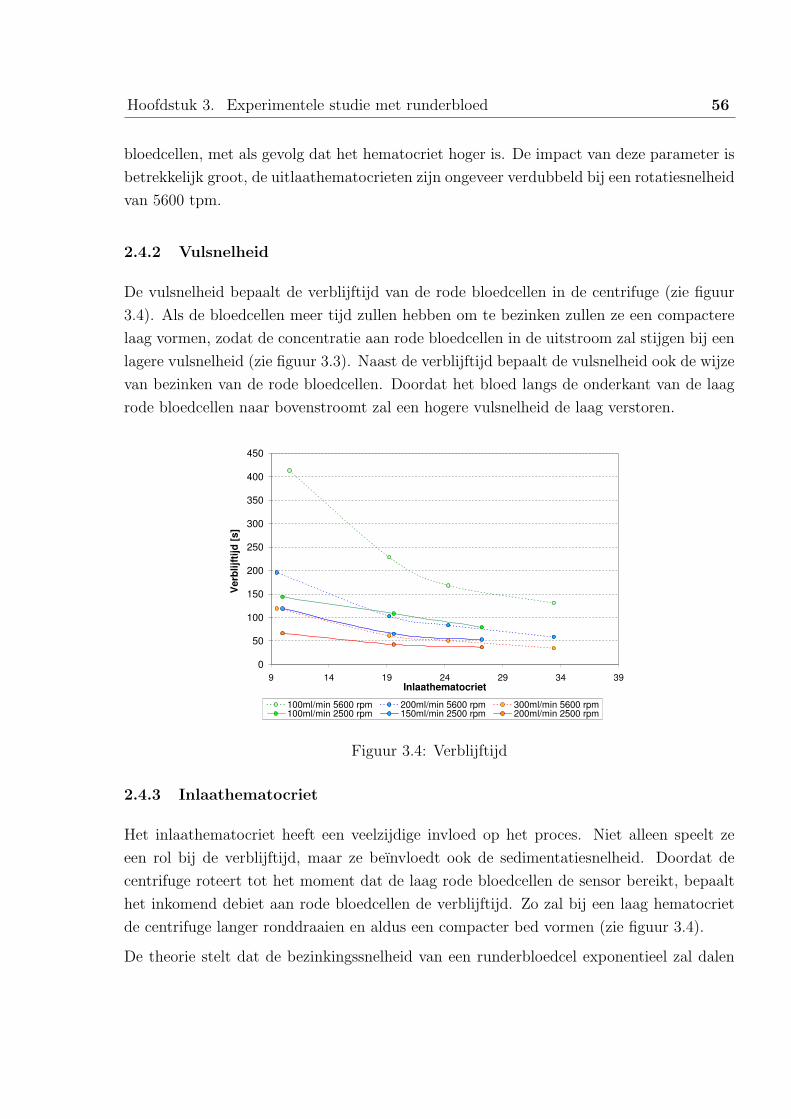
Hoofdstuk 3. Experimentele studie met runderbloed 56
bloedcellen, met als gevolg dat het hematocriet hoger is. De impact van deze parameter is
betrekkelijk groot, de uitlaathematocrieten zijn ongeveer verdubbeld bij een rotatiesnelheid
van 5600 tpm.
2.4.2 Vulsnelheid
De vulsnelheid bepaalt de verblijftijd van de rode bloedcellen in de centrifuge (zie figuur
3.4). Als de bloedcellen meer tijd zullen hebben om te bezinken zullen ze een compactere
laag vormen, zodat de concentratie aan rode bloedcellen in de uitstroom zal stijgen bij een
lagere vulsnelheid (zie figuur 3.3). Naast de verblijftijd bepaalt de vulsnelheid ook de wijze
van bezinken van de rode bloedcellen. Doordat het bloed langs de onderkant van de laag
rode bloedcellen naar bovenstroomt zal een hogere vulsnelheid de laag verstoren.
0
50
100
150
200
250
300
350
400
450
9 14 19 24 29 34 39Inlaathematocriet
Verb
lijf
tijd
[s]
100ml/min 5600 rpm 200ml/min 5600 rpm 300ml/min 5600 rpm100ml/min 2500 rpm 150ml/min 2500 rpm 200ml/min 2500 rpm
Figuur 3.4: Verblijftijd
2.4.3 Inlaathematocriet
Het inlaathematocriet heeft een veelzijdige invloed op het proces. Niet alleen speelt ze
een rol bij de verblijftijd, maar ze beınvloedt ook de sedimentatiesnelheid. Doordat de
centrifuge roteert tot het moment dat de laag rode bloedcellen de sensor bereikt, bepaalt
het inkomend debiet aan rode bloedcellen de verblijftijd. Zo zal bij een laag hematocriet
de centrifuge langer ronddraaien en aldus een compacter bed vormen (zie figuur 3.4).
De theorie stelt dat de bezinkingssnelheid van een runderbloedcel exponentieel zal dalen

Hoofdstuk 3. Experimentele studie met runderbloed 57
naarmate het meer naburen tegenkomt. Bij een laag inlaathematocriet zullen de cellen
initieel sneller bezinken, maar naarmate de laag rode bloedcellen compacter wordt verkleint
deze invloed.
Een hoog inlaathematocriet betekent ook simpelweg dat er initieel al een hoge concentratie
heerst aan bloedcellen. Daardoor vormen de cellen vlugger een gepakt bed en zal het
uitlaathematocriet groter zijn.
De invloed van de parameters op het proces wordt samengevat in tabel 3.6. Deze trends
zijn ook terug te vinden op figuur 3.3. Daar het verloop van het uitlaathematocriet stijgend
is, kan worden besloten dat het initieel aantal rode bloedcellen het meest doorweegt op
het uitlaathematocriet. Dit wil zeggen dat de invloed van het inlaathematocriet op de
verblijftijd en de bezinkingssnelheid niet opweegt tegen het voordeel van een compacte
inlaat.
Tabel 3.6: Invloed van de parameters
Parameter Procesinvloed Uitlaathematocriet
Verblijftijd ↘ ↘Inlaathematocriet ↗ Initiele concentratie RBC ↗ ↗
Initiele bezinkingingssnelheid ↘ ↘Vulsnelheid ↗ Verblijftijd ↘ ↘
Pakkingsverstoring ↗ ↘Rotatiesnelheid ↗ Bezinkingingssnelheid ↗ ↗
2.4.4 Besluit
De resultaten van het experiment zijn op het eerste zicht bevredigend met de theorie.
Ondanks de aanwezigheid van zekere fouten in het proces is het verklaren van de trends
van de hematocrieten toch mogelijk. Mits rekeninghoudend met de fouten zijn de resultaten
nuttig voor verder onderzoek en voor een validatie van het CFD model.

Hoofdstuk 4
Experimentele studie van de
sedimentatie van humaanbloed in een
centrifuge
Het experiment besproken in het vorige hoofdstuk werd uitgevoerd met runderbloed. Veel
experimenten voor het testen van centrifuges worden uitgevoerd met runderbloed omdat
het eenvoudiger te verkrijgen is. Het zou interessant zijn het verband tussen het gedrag van
runderbloed en humaanbloed in centrifuges te kennen. In het eerste deel van dit hoofdstuk
wordt aan de hand van in vivo data afkomstig van het Universitair Ziekenhuis van Gent
onderzocht of het mogelijk is om een verband of een correlatie op te stellen. In een tweede
deel werd geopteerd om een in vitro experiment uit te voeren met humaanbloed.
1 Studie van de data uit het Universitair Ziekenhuis
Gent
De data uit het Universitair Ziekenhuis Gent werd niet zelf opgemeten maar werd verkregen
door metingen tijdens in vivo scheidingsoperaties. In deze paragraaf worden deze in vivo
data besproken en vergeleken met de data van het experiment met runderbloed.
1.1 De parameters en metingen
De in vivo data werden bekomen bij andere procescondities dan het hier boven beschreven
experiment. In tabel 4.1 en tabel 4.2 worden de procesparameters en de metingen weerge-
58

Hoofdstuk 4. Experimentele studie met humaanbloed 59
geven van het experiment met ruderbloed en de in vivo data. Tussen haakjes staat het
aantal metingen bij die parameter.
Tabel 4.1: Gegevens runderbloed
Parameters
Rotatiesnelheid (tpm) 2 500 (12), 5 600 (12)
Vulsnelheid (ml/min) 100 (8), 200 (8), 300 (8)*
Uitstroomsnelheid (ml/min) 50 (24)
Metingen
Volume in, uit, waste en was
In- en uitlaathematocriet Hemocue, Coulter
In- en uitlaathemoglobine Coulter
Temperatuur
* Bij 2 500 tpm is de vulsnelheid 150 in plaats van 300.
Tabel 4.2: Gegevens humaanbloed
Parameters
Rotatiesnelheid (tpm) 5 600 (57)
Vulsnelheid (ml/min) 250 (1), 300 (39), 450 (14), 500 (3)
Wassnelheid (ml/min) 200 (2), 250 (52), 800 (3)
Uitstroomsnelheid (ml/min) 200 (1), 250 (39), 300 (14), 500 (3)
Metingen
Volume in, uit, waste en was
Inlaathematocriet Electa (57)
Uitlaathemoglobine Coulter (57)
Uitlaathematocriet Radiometer (29), Coulter (57) en Electa (57)
De in vivo data worden geordend in groepjes per vulsnelheid en per inlaathematocriet.
De Electa biedt de gebruiker de mogelijkheid om een aantal programma’s te selecteren,
deze programma’s hebben elk hun eigen parameters. Een drietal gegevens werden bepaald
tijdens een noodprocedure, vandaar de drie metingen bij een hoge vul-, was- en uitstroom-
snelheid. De 14 metingen bij een vulsnelheid van 450 ml/min hebben eveneens een uit-
stroomsnelheid van 300 ml/min. Een enkele meting werd uitgevoerd met een vulsnelheid,
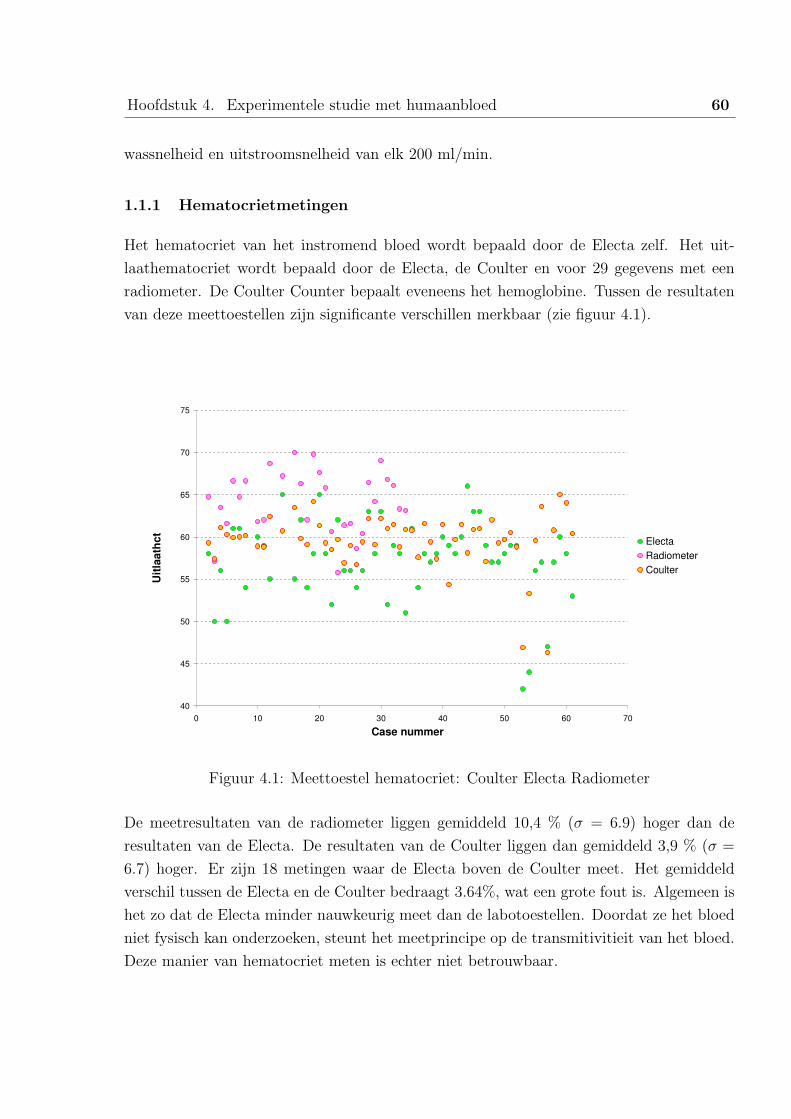
Hoofdstuk 4. Experimentele studie met humaanbloed 60
wassnelheid en uitstroomsnelheid van elk 200 ml/min.
1.1.1 Hematocrietmetingen
Het hematocriet van het instromend bloed wordt bepaald door de Electa zelf. Het uit-
laathematocriet wordt bepaald door de Electa, de Coulter en voor 29 gegevens met een
radiometer. De Coulter Counter bepaalt eveneens het hemoglobine. Tussen de resultaten
van deze meettoestellen zijn significante verschillen merkbaar (zie figuur 4.1).
40
45
50
55
60
65
70
75
0 10 20 30 40 50 60 70
Case nummer
Uit
laath
ct
Electa
Radiometer
Coulter
Figuur 4.1: Meettoestel hematocriet: Coulter Electa Radiometer
De meetresultaten van de radiometer liggen gemiddeld 10,4 % (σ = 6.9) hoger dan de
resultaten van de Electa. De resultaten van de Coulter liggen dan gemiddeld 3,9 % (σ =
6.7) hoger. Er zijn 18 metingen waar de Electa boven de Coulter meet. Het gemiddeld
verschil tussen de Electa en de Coulter bedraagt 3.64%, wat een grote fout is. Algemeen is
het zo dat de Electa minder nauwkeurig meet dan de labotoestellen. Doordat ze het bloed
niet fysisch kan onderzoeken, steunt het meetprincipe op de transmitivitieit van het bloed.
Deze manier van hematocriet meten is echter niet betrouwbaar.
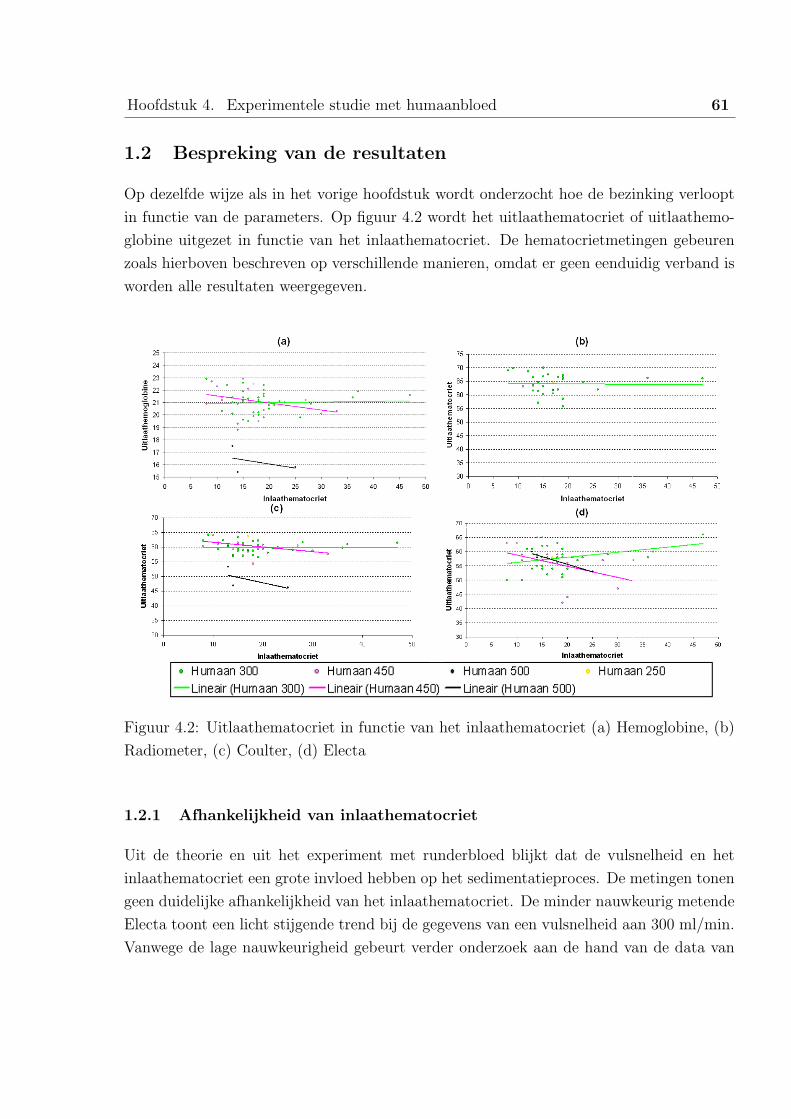
Hoofdstuk 4. Experimentele studie met humaanbloed 61
1.2 Bespreking van de resultaten
Op dezelfde wijze als in het vorige hoofdstuk wordt onderzocht hoe de bezinking verloopt
in functie van de parameters. Op figuur 4.2 wordt het uitlaathematocriet of uitlaathemo-
globine uitgezet in functie van het inlaathematocriet. De hematocrietmetingen gebeuren
zoals hierboven beschreven op verschillende manieren, omdat er geen eenduidig verband is
worden alle resultaten weergegeven.
Figuur 4.2: Uitlaathematocriet in functie van het inlaathematocriet (a) Hemoglobine, (b)
Radiometer, (c) Coulter, (d) Electa
1.2.1 Afhankelijkheid van inlaathematocriet
Uit de theorie en uit het experiment met runderbloed blijkt dat de vulsnelheid en het
inlaathematocriet een grote invloed hebben op het sedimentatieproces. De metingen tonen
geen duidelijke afhankelijkheid van het inlaathematocriet. De minder nauwkeurig metende
Electa toont een licht stijgende trend bij de gegevens van een vulsnelheid aan 300 ml/min.
Vanwege de lage nauwkeurigheid gebeurt verder onderzoek aan de hand van de data van
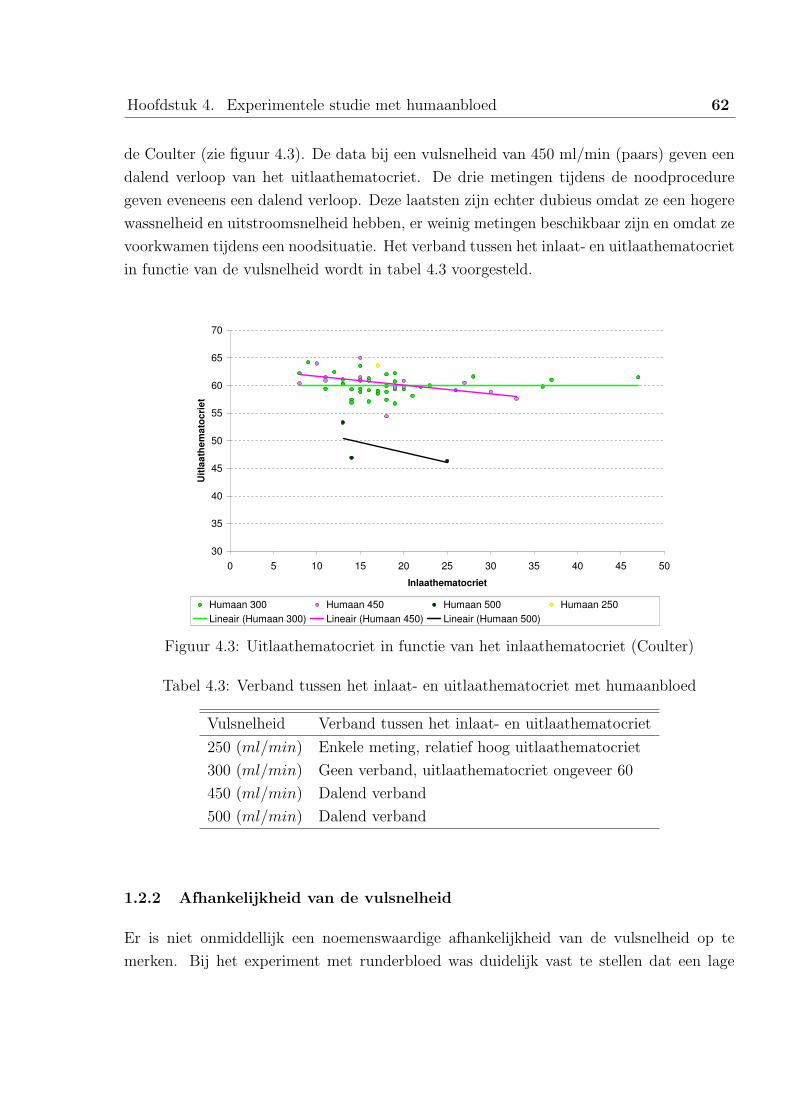
Hoofdstuk 4. Experimentele studie met humaanbloed 62
de Coulter (zie figuur 4.3). De data bij een vulsnelheid van 450 ml/min (paars) geven een
dalend verloop van het uitlaathematocriet. De drie metingen tijdens de noodprocedure
geven eveneens een dalend verloop. Deze laatsten zijn echter dubieus omdat ze een hogere
wassnelheid en uitstroomsnelheid hebben, er weinig metingen beschikbaar zijn en omdat ze
voorkwamen tijdens een noodsituatie. Het verband tussen het inlaat- en uitlaathematocriet
in functie van de vulsnelheid wordt in tabel 4.3 voorgesteld.
30
35
40
45
50
55
60
65
70
0 5 10 15 20 25 30 35 40 45 50
Inlaathematocriet
Uit
laa
the
ma
toc
rie
t
Humaan 300 Humaan 450 Humaan 500 Humaan 250
Lineair (Humaan 300) Lineair (Humaan 450) Lineair (Humaan 500)
Figuur 4.3: Uitlaathematocriet in functie van het inlaathematocriet (Coulter)
Tabel 4.3: Verband tussen het inlaat- en uitlaathematocriet met humaanbloed
Vulsnelheid Verband tussen het inlaat- en uitlaathematocriet
250 (ml/min) Enkele meting, relatief hoog uitlaathematocriet
300 (ml/min) Geen verband, uitlaathematocriet ongeveer 60
450 (ml/min) Dalend verband
500 (ml/min) Dalend verband
1.2.2 Afhankelijkheid van de vulsnelheid
Er is niet onmiddellijk een noemenswaardige afhankelijkheid van de vulsnelheid op te
merken. Bij het experiment met runderbloed was duidelijk vast te stellen dat een lage

Hoofdstuk 4. Experimentele studie met humaanbloed 63
vulsnelheid in verband staat met hogere uitlaathematocrieten. Op figuur 4.3 is dit niet
op te merken, de groep data van 300 ml/min en 450 ml/min zijn niet duidelijk te onder-
scheiden. De enige meting bij 250 ml/min toont wel een betere scheiding. De data van de
noodprocedure bij 500 ml/min tonen een reeks van drie lagere uitlaathematocrieten. Het
uniek zijn van de ene meting en het dubieus zijn van de andere maken echter dat deze
metingen te dubbelzinnig zijn om een afhankelijkheid van de vulsnelheid te stellen.
1.2.3 Afhankelijkheid van andere parameters
Naast de vulsnelheid en het inlaathematocriet zijn er nog andere data zoals het volume aan
wasvloeistof, het totaal gebruikt volume, het volume aan geconcentreerde rode bloedcellen,
het wastevolume, de wassnelheid, de uitstroomsnelheid en de procestijd. Aan de hand
van de volumemetingen kan een volumebalans worden opgesteld en gecontroleerd worden.
Deze klopt net als bij het experiment met runderbloed niet. Zo tonen de data van de
noodprocedure daar grote afwijkingen. De wassnelheid en uitstroomsnelheid hangen vast
aan het programma en dus de vulsnelheid dat gekozen werd door de gebruiker. Doordat
de drie parameters gelijktijdig veranderen kan de invloed ervan niet worden achterhaald.
Er zijn eveneens te weinig verschillende waarden om een verband te zoeken, zie tabel 4.2.
Er zijn data gemeten bij hetzelfde inlaathematocriet en dezelfde vulsnelheid. Verder kan
nog onderzocht worden naar de oorzaak van het verschil in uitlaathematocriet bij die data,
zie figuur 4.3 (inlaathematocriet: 15, 16, 18 en 19). Er is echter geen enkel verband
met bovenvermelde parameters. Mogelijke redenen voor de afwezigheid van verbanden
zijn niet eenvoudig te achterhalen. De oorzaak kan liggen bij de wijze van meten, het
wasproces of aan het bloed. Het bloed is immers telkens van een andere patient waardoor
de fysische eigenschappen van de vloeistof kunnen verschillen. Maar de belangrijkste reden
is waarschijnlijk de onnauwkeurige meting van het inlaathematocriet. Deze wordt enkel
gemeten door de Electa en is aldus niet betrouwbaar. Omdat er veel afhangt van het
inlaathematocriet is het proces moeilijk te doorgronden.
1.3 Verband met runderbloed
Het is duidelijk dat beide gegevens bij verschillende omstandigheden werden gemeten.
Buiten het verschil in parameters vermeld in tabellen 4.1 en 4.2 is er nog het verschil in
temperatuur van het bloed. Het humaanbloed wordt gecentrifugeerd op lichaamstempe-
ratuur terwijl het bloed van het rund gedurende het experiment afkoelde van 30 naar

Hoofdstuk 4. Experimentele studie met humaanbloed 64
22,5�. De temperatuur is een belangrijke factor omdat het de viscositeit van het plasma
beınvloedt.
y = 0,0021x + 59,902
R2 = 8E-05
35
40
45
50
55
60
65
70
0 5 10 15 20 25 30 35 40 45 50
Inlaathematocriet
Uit
laath
em
ato
crie
t Humaan 300
Humaan 450
Humaan 500
Humaan 250
100ml/min 5600 rpm
200ml/min 5600 rpm
300ml/min 5600 rpm
Lineair (Humaan 300)
Lineair (Humaan 450)
Figuur 4.4: Data van het experiment met runderbloed en vanuit het UZ
Om een verband te zoeken tussen het gedrag van humaan en runderbloed moeten de ge-
gevens bij dezelfde omstandigheden worden vergeleken. Op deze manier kan men er zeker
van zijn dat de onderzochte parameter het proces dusdanig beınvloedt. Zo kunnen de
experimentele gegevens bij 2500 tpm niet gebruikt worden. Perfect samenvallende pro-
cescondities zijn er niet, het humaanbloed wordt immers gewassen en heeft een grotere
uitstroomsnelheid. De enige reeks metingen die overeenstemt met beide data is bij 300
ml/min en 5600 tpm. Omdat er geen metingen zijn bij hogere vulsnelheden met runder-
bloed en een meting bij een vulsnelheid van 250 ml/min niet representatief is, kan er geen
correlatie worden opgesteld in functie van de vulsnelheid. In figuur 4.4 worden beide data
grafisch weergegeven. Wanneer de uitlaathematocrieten bij een vulsnelheid van 300 ml/min
worden vergeleken is er bij het runderbloed een duidelijke afhankelijkheid van het inlaathe-
matocriet. Het humaanbloed toont echter geen afhankelijkheid van het inlaathematocriet:
het uitlaathematocriet toont een vaste waarde van 60 % (σ = 3.30). Doordat de in vivo
data van het UZ Gent geen functioneel verband vertonen met het inlaathematocriet kan
er geen correlatie worden opgesteld.

Hoofdstuk 4. Experimentele studie met humaanbloed 65
2 Experiment met humaanbloed
De in vivo data beschreven in vorige paragraaf geven geen duidelijke verbanden weer. Het
is eveneens niet mogelijk om een correlatie op te stellen tussen het uitlaathematocriet van
runderbloed en humaanbloed. Om deze problemen te verhelpen werd besloten om een
analoog experiment uit te voeren als in hoofdstuk 3 maar dan met humaanbloed. Met als
gevolg dat de parameters nu zelf kunnen worden ingesteld zodat de resultaten beter kunnen
worden vergeleken met die van het experiment met runderbloed. De metingen worden zelf
uitgevoerd zodat fouten in de resultaten snel te achterhalen zijn.
Er werd 450 ml bloed afgenomen van een donor, dit kleine volume maakt het noodzakelijk
om het bloed na elke operatie terug te mengen. Doordat het bloed slechts van een donor
werd afgenomen is er geen afhankelijkheid van het bloed.
2.1 De opstelling
De opstelling van de proeven is analoog als beschreven in hoofdstuk 3 (zie figuur 3.1). Zo
wordt er eveneens gebruik gemaakt van de Electa met een bowl van 125ml. Het bloed werd
tijdens de eerste metingen opgevangen in zakken. Dit systeem maakt het moeilijk om een
representatief staal te nemen, daarom werd na twee operaties overgeschakeld naar een open
systeem met maatglazen in plaats van zakken zoals bij het experiment met runderbloed.
Enkel de zak voor het plasma werd behouden.
2.2 De parameters
De parameters worden op dezelfde manier als het experiment met runderbloed gekozen (zie
tabel 4.4). Omdat scheidingsoperaties bij een toerental van 2500 niet worden uitgevoerd
voor de recuperatie van rode bloedcellen worden hier enkel experimenten uitgevoerd bij
5600 tpm. De vulsnelheid varieert voor elk inlaathematocriet van 100 ml/min naar 200
ml/min tot 300 ml/min. Zo worden er dus 12 metingen gedaan. Het bloed wordt ver-
dund met een fysiologische oplossing zodat de concentraties ongeveer gelijk zijn als bij het
runderbloed.

Hoofdstuk 4. Experimentele studie met humaanbloed 66
Tabel 4.4: Parameters
Parameter
Rotatiesnelheid (tpm) 5 600
Vulsnelheid (ml/min) 100, 200, 300
Inlaathematocriet (%) ≈ 10, 20, 30, 40
2.3 De resultaten
2.3.1 Tabel
De resultaten van het experiment worden weergegeven in tabel 4.5. Bij aanvangen van
meting drie werd het zakkensysteem verwijderd. Tijdens de uitstroomfase van meting
vier en vijf werd de plasmazak niet omgedraaid zodat er plasma terug in het residu werd
gepompt. Het staal van meting zes werd fout opgenomen. Bovenstaande metingen worden
niet in rekening gebracht bij de verdere analyse.
2.3.2 Bespreking van de volumes
Op dezelfde manier als bij het experiment met runderbloed worden hier de volumes gemeten
van de drie stromen. Daarom zullen dezelfde fouten optreden zoals het resterend bloed
dat achterblijft in de pomp en de leidingen. De volumemetingen kunnen derhalve niet
gebruikt worden voor het bepalen van balansen maar uit het instromend volume kan wel
de verblijftijd berekend worden.
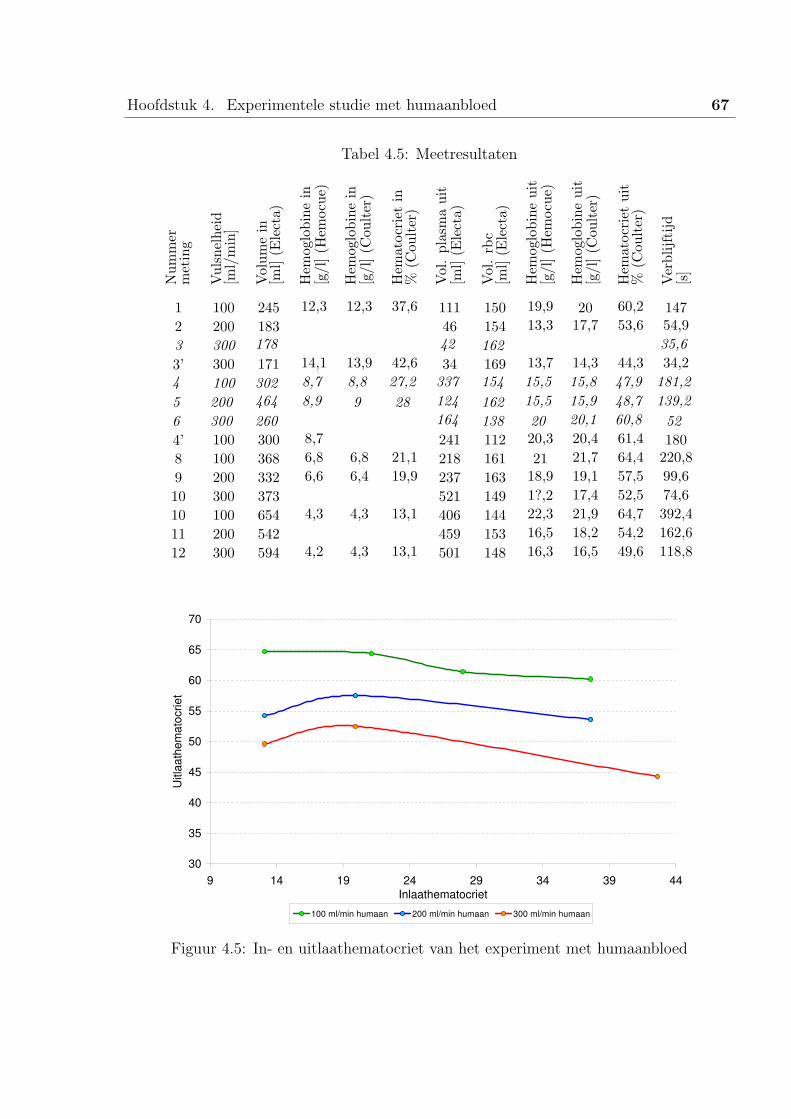
Hoofdstuk 4. Experimentele studie met humaanbloed 67
Tabel 4.5: MeetresultatenN
umm
erm
etin
g
Vul
snel
heid
[ml/
min
]
Vol
ume
in[m
l](E
lect
a)
Hem
oglo
bine
in[g
/l](H
emoc
ue)
Hem
oglo
bine
in[g
/l](C
oult
er)
Hem
atoc
riet
in%
(Cou
lter
)
Vol
.pl
asm
aui
t[m
l](E
lect
a)
Vol
.rb
c[m
l](E
lect
a)
Hem
oglo
bine
uit
[g/l
](H
emoc
ue)
Hem
oglo
bine
uit
[g/l
](C
oult
er)
Hem
atoc
riet
uit
%(C
oult
er)
Ver
blijf
tijd
[s]
1233’4564’8910101112
100200300300100200300100100200300100200300
245183178171302464260300368332373654542594
12,3
14,18,78,9
8,76,86,6
4,3
4,2
12,3
13,98,89
6,86,4
4,3
4,3
37,6
42,627,228
21,119,9
13,1
13,1
111464234
337124164241218237521406459501
150154162169154162138112161163149144153148
19,913,3
13,715,515,52020,321
18,91?,222,316,516,3
2017,7
14,315,815,920,120,421,719,117,421,918,216,5
60,253,6
44,347,948,760,861,464,457,552,564,754,249,6
14754,935,634,2
181,2139,252180
220,899,674,6392,4162,6118,8
30
35
40
45
50
55
60
65
70
9 14 19 24 29 34 39 44
Inlaathematocriet
Uitla
ath
em
ato
cri
et
100 ml/min humaan 200 ml/min humaan 300 ml/min humaan
Figuur 4.5: In- en uitlaathematocriet van het experiment met humaanbloed
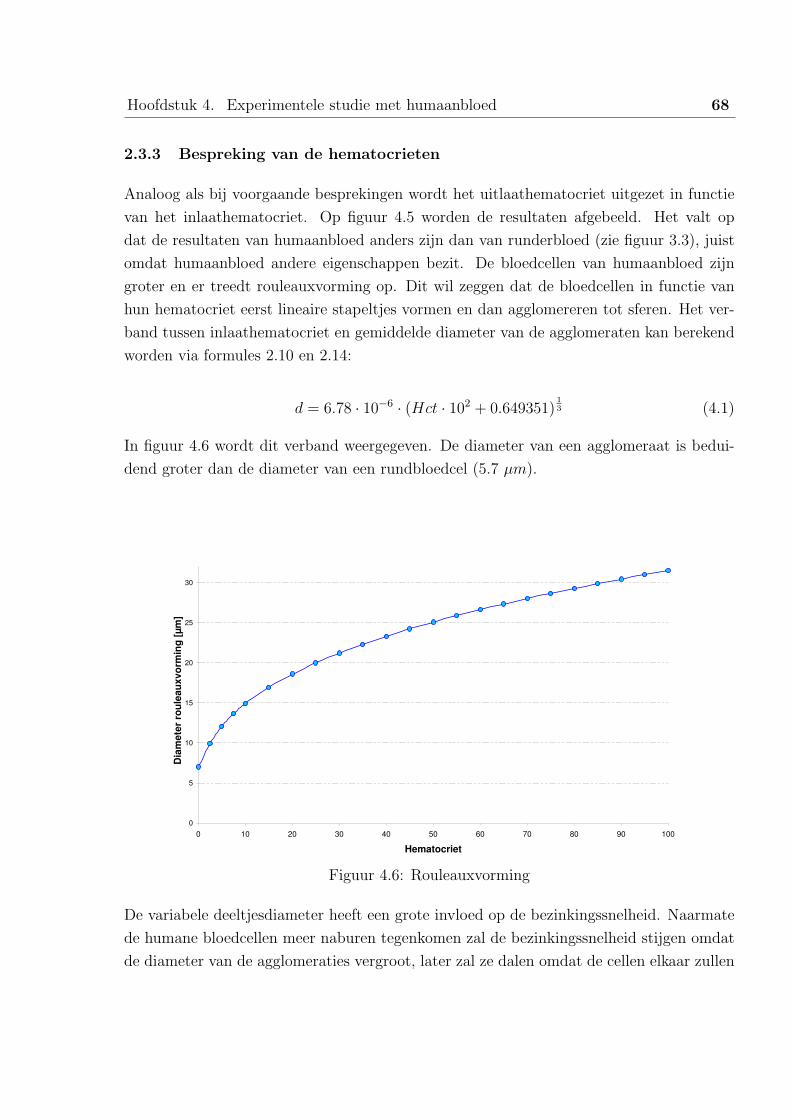
Hoofdstuk 4. Experimentele studie met humaanbloed 68
2.3.3 Bespreking van de hematocrieten
Analoog als bij voorgaande besprekingen wordt het uitlaathematocriet uitgezet in functie
van het inlaathematocriet. Op figuur 4.5 worden de resultaten afgebeeld. Het valt op
dat de resultaten van humaanbloed anders zijn dan van runderbloed (zie figuur 3.3), juist
omdat humaanbloed andere eigenschappen bezit. De bloedcellen van humaanbloed zijn
groter en er treedt rouleauxvorming op. Dit wil zeggen dat de bloedcellen in functie van
hun hematocriet eerst lineaire stapeltjes vormen en dan agglomereren tot sferen. Het ver-
band tussen inlaathematocriet en gemiddelde diameter van de agglomeraten kan berekend
worden via formules 2.10 en 2.14:
d = 6.78 · 10−6 · (Hct · 102 + 0.649351)13 (4.1)
In figuur 4.6 wordt dit verband weergegeven. De diameter van een agglomeraat is bedui-
dend groter dan de diameter van een rundbloedcel (5.7 µm).
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80 90 100
Hematocriet
Dia
me
ter
rou
lea
ux
vo
rmin
g [
µm
]
Figuur 4.6: Rouleauxvorming
De variabele deeltjesdiameter heeft een grote invloed op de bezinkingssnelheid. Naarmate
de humane bloedcellen meer naburen tegenkomen zal de bezinkingssnelheid stijgen omdat
de diameter van de agglomeraties vergroot, later zal ze dalen omdat de cellen elkaar zullen
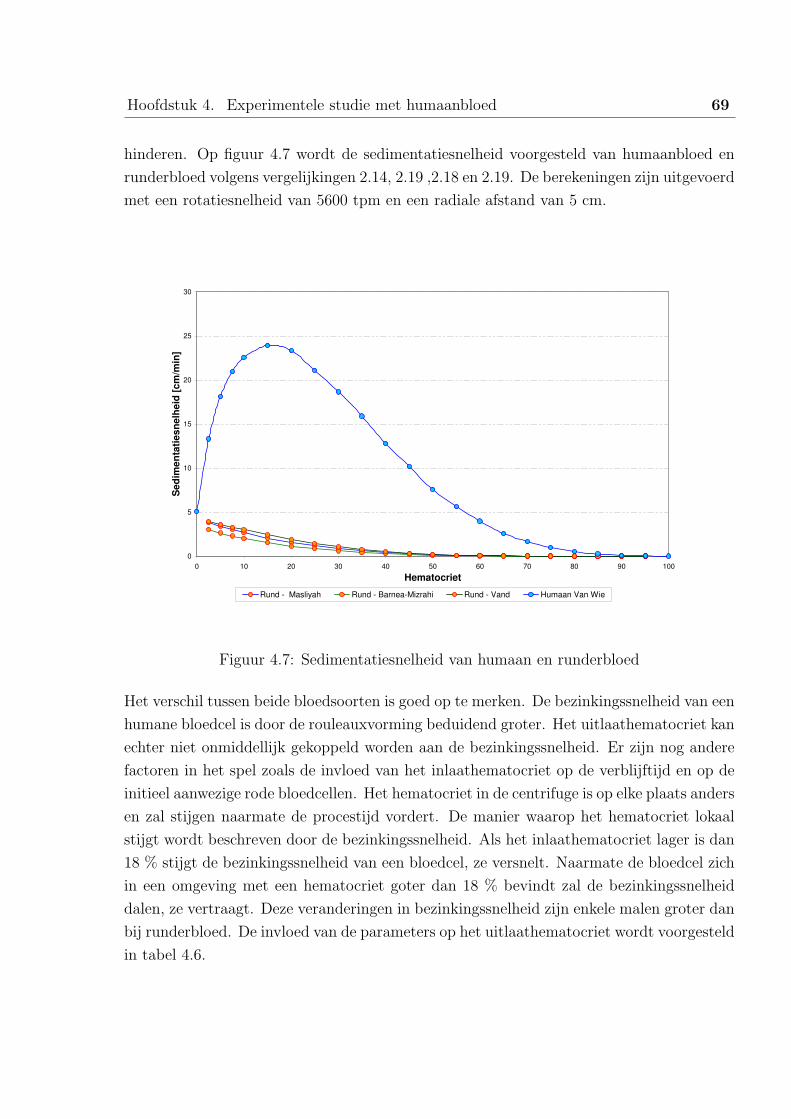
Hoofdstuk 4. Experimentele studie met humaanbloed 69
hinderen. Op figuur 4.7 wordt de sedimentatiesnelheid voorgesteld van humaanbloed en
runderbloed volgens vergelijkingen 2.14, 2.19 ,2.18 en 2.19. De berekeningen zijn uitgevoerd
met een rotatiesnelheid van 5600 tpm en een radiale afstand van 5 cm.
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80 90 100
Hematocriet
Se
dim
en
tati
es
ne
lhe
id [
cm
/min
]
Rund - Masliyah Rund - Barnea-Mizrahi Rund - Vand Humaan Van Wie
Figuur 4.7: Sedimentatiesnelheid van humaan en runderbloed
Het verschil tussen beide bloedsoorten is goed op te merken. De bezinkingssnelheid van een
humane bloedcel is door de rouleauxvorming beduidend groter. Het uitlaathematocriet kan
echter niet onmiddellijk gekoppeld worden aan de bezinkingssnelheid. Er zijn nog andere
factoren in het spel zoals de invloed van het inlaathematocriet op de verblijftijd en op de
initieel aanwezige rode bloedcellen. Het hematocriet in de centrifuge is op elke plaats anders
en zal stijgen naarmate de procestijd vordert. De manier waarop het hematocriet lokaal
stijgt wordt beschreven door de bezinkingssnelheid. Als het inlaathematocriet lager is dan
18 % stijgt de bezinkingssnelheid van een bloedcel, ze versnelt. Naarmate de bloedcel zich
in een omgeving met een hematocriet goter dan 18 % bevindt zal de bezinkingssnelheid
dalen, ze vertraagt. Deze veranderingen in bezinkingssnelheid zijn enkele malen groter dan
bij runderbloed. De invloed van de parameters op het uitlaathematocriet wordt voorgesteld
in tabel 4.6.

Hoofdstuk 4. Experimentele studie met humaanbloed 70
Tabel 4.6: Invloed van de parameters
Parameter Procesinvloed Uitlaathematocriet
Verblijftijd ↘ ↘Inlaathematocriet ↗ Initiele concentratie RBC ↗ ↗
< 18% Initiele bezinkingingssnelheid ↗ ↗> 18% Initiele bezinkingingssnelheid ↘ ↘
Vulsnelheid ↗ Verblijftijd ↘ ↘Pakkingsverstoring ↗ ↘
Voor een vulsnelheid van 200 ml/min en 300 ml/min toont figuur 4.5 een analoog verloop
aan als deze van de bezinkingssnelheid. Dit wil zeggen dat de invloed van de bezinkingssnel-
heid op het uitlaathematocriet groter is dan de initiele concentratie aan rode bloedcellen.
Bij runderbloed was dit anders, maar door de grote verschillen in sedimentatiesnelheid
neemt deze de overhand. De metingen bij 100 ml/min volgen de trend van figuur 4.5 niet,
de verblijftijd bij een inlaathematocriet van 13.1 % is zeer hoog waardoor de cellen veel
tijd hebben om een gepakter bed te vormen. Door de hogere bezinkingssnelheid van de
humane bloedcellen is de invloed van de verblijftijd hier ongeveer dubbel zo groot als bij
runderbloed.
3 Besluit
In het eerst deel van dit hoofstuk werd in vivo data beschreven afkomstig van het Universi-
tair Ziekenhuis Gent. Omdat het inlaathematocriet te rudimentair gemeten werd kon deze
data geen zinnige betekenis leveren. Daarom was het beter om zelf een experiment uit te
voeren met humaanbloed. De resultaten van dit experiment vallen goed te verklaren met
de theorie. Nu dat van runderbloed en humaanbloed een experiment werd uitgevoerd kan
een onderzoek gestart worden naar het verband tussen deze resultaten.

Hoofdstuk 5
Verband tussen humaan en
runderbloed
In dit hoofdstuk worden de resultaten van beide experimenten iets dieper bestudeerd. Het
is de bedoeling de data te interpoleren en om te vormen in een formule. Het uitlaathema-
tocriet wordt dan gegeven in functie van het inlaathematocriet en de vulsnelheid. Het gaat
hier om ruwe interpolaties die een beduidend grote fout kunnen geven. In de wereld van de
biologische wetenschappen is het immers moeilijk om consistente correlaties te bekomen
omdat er heel wat meer invloedsfactoren zijn. Zo bezit het bloed nog andere eigenschappen
die de sedimentatie in kleine mate kunnen beınvloeden zoals de celvorm, het aantal witte
bloedcellen of de dichtheid van de rode bloedcellen.
1 Analyse van de data van runderbloed
In deze paragraaf wordt een formule vooropgesteld die het uitlaathematocriet weergeeft in
functie van het inlaathematocriet en de vulsnelheid.
Hctuit R = f(Hctin R, q) (5.1)
met:
q vulsnelheid in ml/min
Hct hematocriet in %
Omdat het verband tussen inlaat- en uitlaathematocriet lineair is, wordt formule 5.1:
Hctuit R = a ·Hctin R + b · q + c (5.2)
71

Hoofdstuk 5. Verband tussen humaan en runderbloed 72
Voor elke vulsnelheid liggen de resultaten van het experiment ongeveer op drie evenwijdige
rechten (zie figuur 5.1). Voor elke reeks metingen wordt een trendlijn toegevoegd zodat
er een functioneel verband ontstaat tussen het uitlaat- en inlaathematocriet. Dit verband
wordt weergegeven in tabel 5.1
30
35
40
45
50
55
60
9 14 19 24 29 34
Inlaathematocriet
Uitla
ath
em
ato
cri
et
100 ml/min rund 200 ml/min rund 300 ml/min rund
Figuur 5.1: Trendlijnen
Tabel 5.1: Functies van de trendlijnen
Vulsnelheid y = f(x) R2
100ml/min y = 0.3505 x + 43.50 0.9882
200ml/min y = 0.3937 x + 39.02 0.9758
300ml/min y = 0.2844 x + 37.45 0.9956
Om een formule te bekomen die voor elke vulsnelheid geldt worden de rechten als even-
wijdigen benaderd. De richtingscoefficienten verschillen weinig, het gemiddelde geeft de
richtingscoefficient a van formule (5.2). Omdat de kleine rotaties van de rechten niet al te
veel invloed zouden hebben worden ze geroteerd op ongeveer het middelpunt van de trend-
lijnen. Hier wordt geroteerd bij het punt op een inlaathematocriet van 21. De functies van
de nieuwe rechten en de fout van de richtingscoefficientworden vermeld in tabel 5.2:
Nu worden de constante termen van de vergelijkingen in tabel 5.2 onderzocht zodat ze
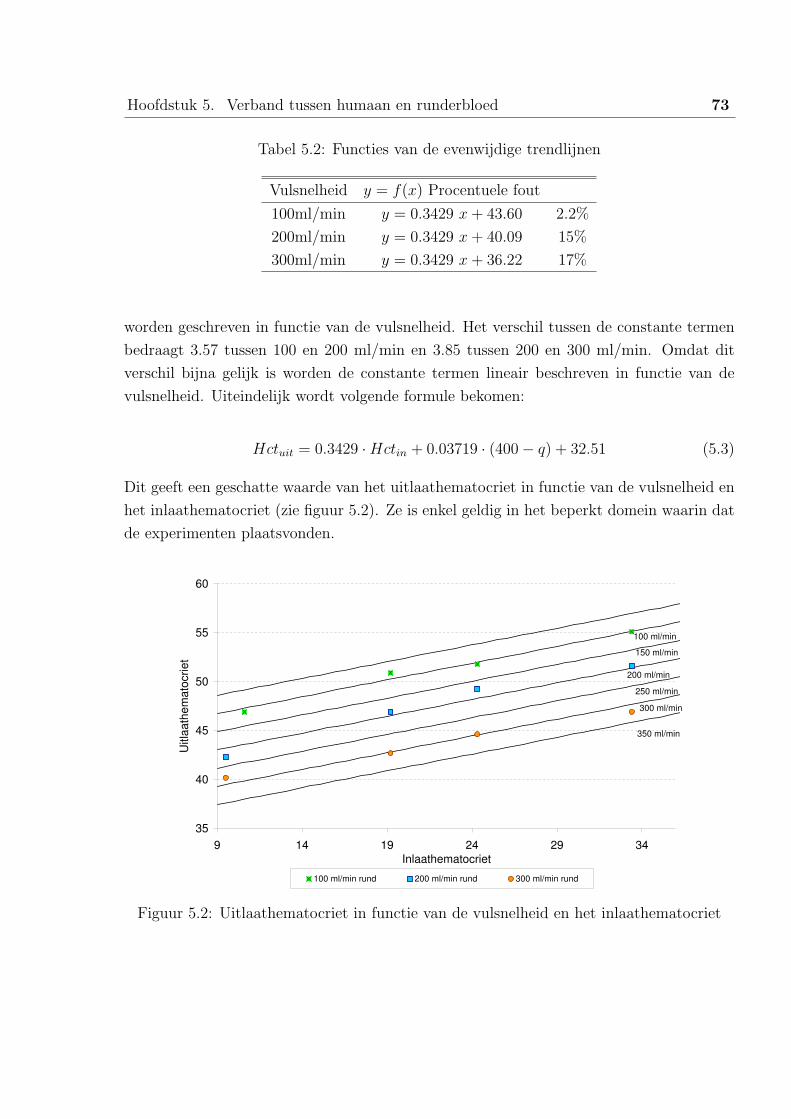
Hoofdstuk 5. Verband tussen humaan en runderbloed 73
Tabel 5.2: Functies van de evenwijdige trendlijnen
Vulsnelheid y = f(x) Procentuele fout
100ml/min y = 0.3429 x + 43.60 2.2%
200ml/min y = 0.3429 x + 40.09 15%
300ml/min y = 0.3429 x + 36.22 17%
worden geschreven in functie van de vulsnelheid. Het verschil tussen de constante termen
bedraagt 3.57 tussen 100 en 200 ml/min en 3.85 tussen 200 en 300 ml/min. Omdat dit
verschil bijna gelijk is worden de constante termen lineair beschreven in functie van de
vulsnelheid. Uiteindelijk wordt volgende formule bekomen:
Hctuit = 0.3429 ·Hctin + 0.03719 · (400− q) + 32.51 (5.3)
Dit geeft een geschatte waarde van het uitlaathematocriet in functie van de vulsnelheid en
het inlaathematocriet (zie figuur 5.2). Ze is enkel geldig in het beperkt domein waarin dat
de experimenten plaatsvonden.
200 ml/min
300 ml/min
350 ml/min
150 ml/min
100 ml/min
250 ml/min
35
40
45
50
55
60
9 14 19 24 29 34
Inlaathematocriet
Uitla
ath
em
ato
cri
et
100 ml/min rund 200 ml/min rund 300 ml/min rund
Figuur 5.2: Uitlaathematocriet in functie van de vulsnelheid en het inlaathematocriet
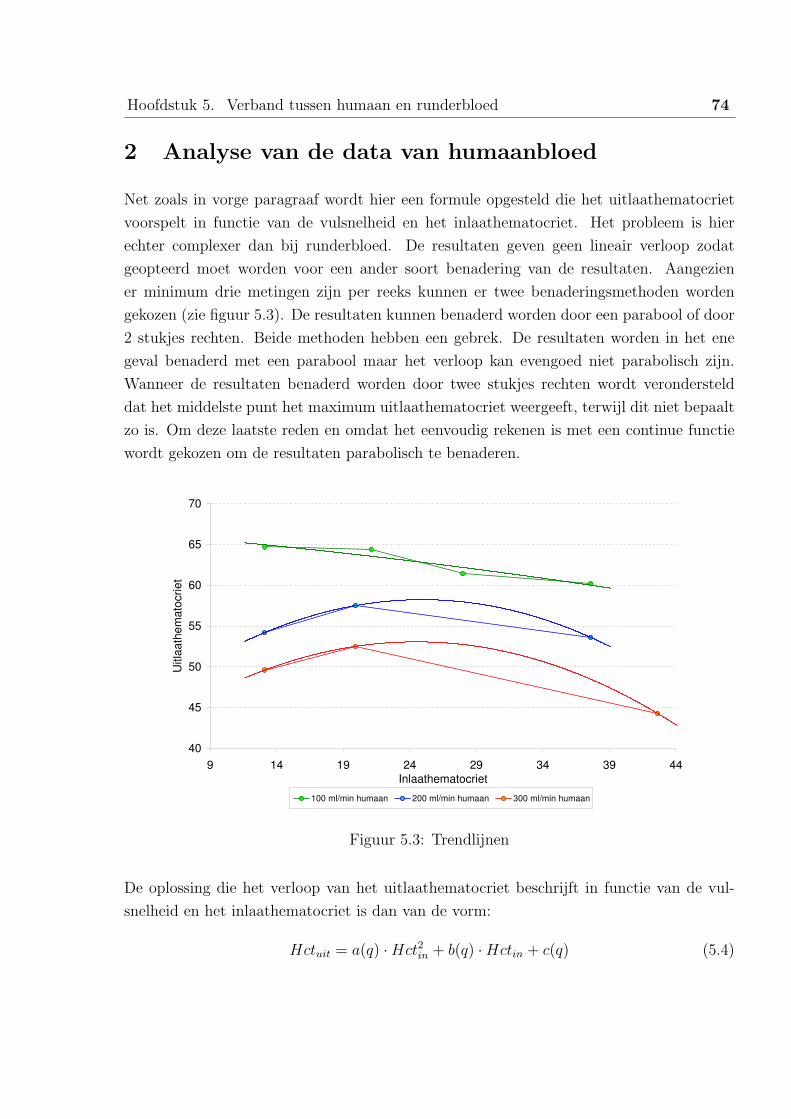
Hoofdstuk 5. Verband tussen humaan en runderbloed 74
2 Analyse van de data van humaanbloed
Net zoals in vorge paragraaf wordt hier een formule opgesteld die het uitlaathematocriet
voorspelt in functie van de vulsnelheid en het inlaathematocriet. Het probleem is hier
echter complexer dan bij runderbloed. De resultaten geven geen lineair verloop zodat
geopteerd moet worden voor een ander soort benadering van de resultaten. Aangezien
er minimum drie metingen zijn per reeks kunnen er twee benaderingsmethoden worden
gekozen (zie figuur 5.3). De resultaten kunnen benaderd worden door een parabool of door
2 stukjes rechten. Beide methoden hebben een gebrek. De resultaten worden in het ene
geval benaderd met een parabool maar het verloop kan evengoed niet parabolisch zijn.
Wanneer de resultaten benaderd worden door twee stukjes rechten wordt verondersteld
dat het middelste punt het maximum uitlaathematocriet weergeeft, terwijl dit niet bepaalt
zo is. Om deze laatste reden en omdat het eenvoudig rekenen is met een continue functie
wordt gekozen om de resultaten parabolisch te benaderen.
40
45
50
55
60
65
70
9 14 19 24 29 34 39 44
Inlaathematocriet
Uitla
ath
em
ato
cri
et
100 ml/min humaan 200 ml/min humaan 300 ml/min humaan
Figuur 5.3: Trendlijnen
De oplossing die het verloop van het uitlaathematocriet beschrijft in functie van de vul-
snelheid en het inlaathematocriet is dan van de vorm:
Hctuit = a(q) ·Hct2in + b(q) ·Hctin + c(q) (5.4)

Hoofdstuk 5. Verband tussen humaan en runderbloed 75
met:
q vulsnelheid in ml/min
a(q), b(q), c(q) functies van q
Net zoals in de vorige paragraaf worden nu de functies a(q), b(q) en c(q) bepaald. Het
verband tussen inlaathematocriet en uitlaathematocriet per vulsnelheid wordt weergegeven
in tabel 5.3.
Tabel 5.3: Functies van de parabolen
Vulsnelheid y = f(x) R2
100ml/min y = −0.0016 x2 − 0.1203 x + 66.81 0.9040
200ml/min y = −0.0288 x2 + 1.436 x + 40.33 1
300ml/min y = −0.0267 x2 + 1.308 x + 37.05 1
Het is nu de bedoeling dat de coefficienten van deze parabolen geschreven worden in functie
van de vulsnelheid. Omdat deze duidelijk niet lineair veranderen worden ze eveneens
benaderd door een parabolische functie. De functies a(q), b(q) en c(q) worden zo eenvoudig
berekend en weergegeven in tabel 5.4.
Tabel 5.4: De functies a(q), b(q) en c(q)
f(q)
a(q) = 1.463 · 10−6 q2 −7.110 · 10−4 q +0.05483
b(q) = −8.421 · 10−5 q2 +4.082 · 10−2 q −3.360
c(q) = 1.160 · 10−3 q2 −0.6127 q +116.5
Deze functies worden nu ingevuld in formule (5.4), zodat er een algemene vergelijking wordt
bekomen die het uitlaathematocriet voorspelt in functie van het inlaathematocriet en de
vulsnelheid.
Hctuit =(1.385 · 10−6 q2 − 6.795 · 10−4 q + 0.0525·
)Hct2in
+(−8.421 · 10−5 q2 + 4.082 · 10−2 q − 3.360·
)Hctin (5.5)
+ 1.160 · 10−3 q2 − 0.6127 q + 116.5
Om deze oplossing te verduidelijken wordt ze weergegeven voor enkele waarden van q op
grafiek 5.4.
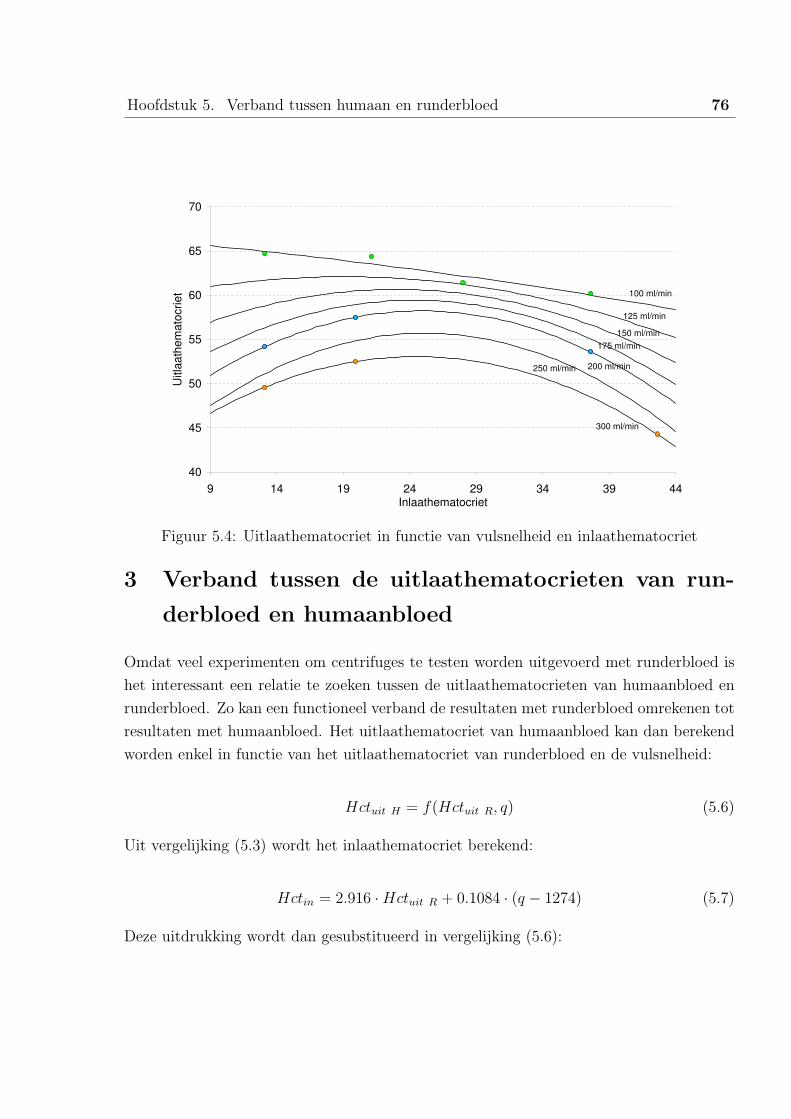
Hoofdstuk 5. Verband tussen humaan en runderbloed 76
100 ml/min
200 ml/min
300 ml/min
125 ml/min
150 ml/min
175 ml/min
250 ml/min
40
45
50
55
60
65
70
9 14 19 24 29 34 39 44
Inlaathematocriet
Uitla
ath
em
ato
cri
et
Figuur 5.4: Uitlaathematocriet in functie van vulsnelheid en inlaathematocriet
3 Verband tussen de uitlaathematocrieten van run-
derbloed en humaanbloed
Omdat veel experimenten om centrifuges te testen worden uitgevoerd met runderbloed is
het interessant een relatie te zoeken tussen de uitlaathematocrieten van humaanbloed en
runderbloed. Zo kan een functioneel verband de resultaten met runderbloed omrekenen tot
resultaten met humaanbloed. Het uitlaathematocriet van humaanbloed kan dan berekend
worden enkel in functie van het uitlaathematocriet van runderbloed en de vulsnelheid:
Hctuit H = f(Hctuit R, q) (5.6)
Uit vergelijking (5.3) wordt het inlaathematocriet berekend:
Hctin = 2.916 ·Hctuit R + 0.1084 · (q − 1274) (5.7)
Deze uitdrukking wordt dan gesubstitueerd in vergelijking (5.6):
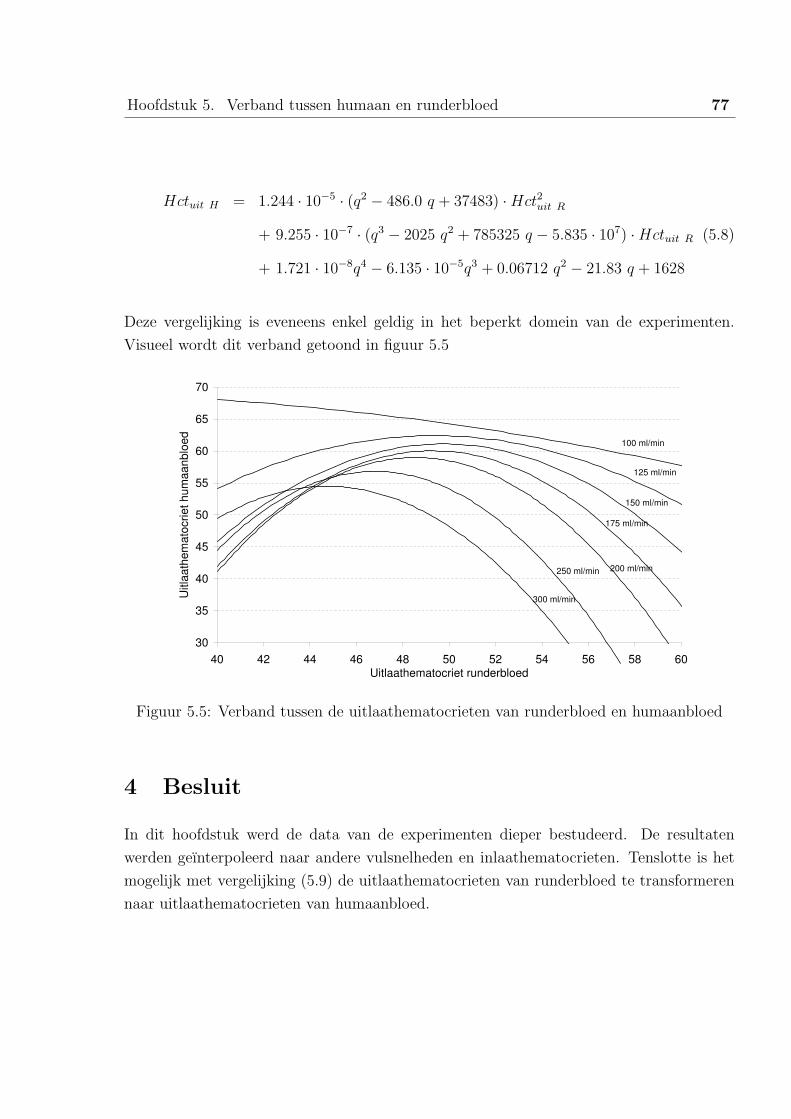
Hoofdstuk 5. Verband tussen humaan en runderbloed 77
Hctuit H = 1.244 · 10−5 · (q2 − 486.0 q + 37483) ·Hct2uit R
+ 9.255 · 10−7 · (q3 − 2025 q2 + 785325 q − 5.835 · 107) ·Hctuit R (5.8)
+ 1.721 · 10−8q4 − 6.135 · 10−5q3 + 0.06712 q2 − 21.83 q + 1628
Deze vergelijking is eveneens enkel geldig in het beperkt domein van de experimenten.
Visueel wordt dit verband getoond in figuur 5.5
100 ml/min
200 ml/min
300 ml/min
125 ml/min
150 ml/min
175 ml/min
250 ml/min
30
35
40
45
50
55
60
65
70
40 42 44 46 48 50 52 54 56 58 60
Uitlaathematocriet runderbloed
Uitla
ath
em
ato
cri
et
hu
ma
an
blo
ed
Figuur 5.5: Verband tussen de uitlaathematocrieten van runderbloed en humaanbloed
4 Besluit
In dit hoofdstuk werd de data van de experimenten dieper bestudeerd. De resultaten
werden geınterpoleerd naar andere vulsnelheden en inlaathematocrieten. Tenslotte is het
mogelijk met vergelijking (5.9) de uitlaathematocrieten van runderbloed te transformeren
naar uitlaathematocrieten van humaanbloed.

Hoofdstuk 6
Numerieke studie van de
sedimentatie van bloedcellen
In dit hoofdstuk wordt beschreven hoe een numeriek model geconstrueerd wordt dat de
sedimentatie van bloedcellen in de centrifuge simuleert. Aan de hand van de resultaten
van het vorig hoofdstuk kan het model getest worden op zijn accuraatheid. Eens er een
degelijk model voor handen is voor de sedimentatie van rode bloedcellen kunnen de witte
bloedcellen en de bloedplaatjes worden toegevoegd aan het model.
Tijdens de modelbeschrijving worden de verschillende stappen toegelicht in de ontwikkeling
van het numeriek model. Het model wordt ontworpen in Gambit, terwijl het programma
Fluent 6.2 de iteratieve berekeningen van de verschillende vergelijkingen op zich neemt.
Gambit [17] is een grafische gebruikersomgeving die de pre-processing mogelijkheden voor
Fluent bundelt. In dit programma wordt de geometrie en het grid voor het model gecreeerd.
Fluent 6.2 [18] is een Computational Fluid Dynamics programma (CFD) gebaseerd op
de eindige volume methode dat een uitgebreid gamma aan mogelijkheden biedt voor de
fysische modellering van stromingen (warmtetransport, tweefase, turbulentie, akoestiek) en
is tevens een hulpmiddel in uiteenlopende wetenschappelijke en industriele vakgebieden.
1 Probleem beschrijving
Het probleem kan ontbonden worden in drie delen:
• De centrifuge stroomt vol
• De centrifuge roteert
78

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 79
• De sedimentatie van de cellen in het plasma
In een eerste fase wordt de roterende centrifuge gevuld met bloed. Dit wil zeggen dat er
initieel lucht in de centrifuge zit. Ondertussen draait de centrifuge rond aan een snelheid
van 2500 of 5600 tpm en bewegen de cellen onder invloed van verschillende krachten. Deze
zaken dienen tegelijkertijd gemodelleerd te worden wat tot enige complicaties leidt.
Het volstromen of vullen van een geometrie gebeurt in Fluent met het Volume of Fluid
model (VOF). Het VOF model is ontworpen om fluıda die niet interageren te modelleren,
zoals bijvoorbeeld water en lucht. De sedimentatie wordt met een ander type model bere-
kend waar de fasen wel interageren. Er kan slechts een model gekozen worden. Doordat
de sedimentatie fundamenteel belangrijk is wordt het volstromen niet gemodelleerd. De
geometrie wordt dan initieel gevuld met plasma terwijl er plasma en bloedcellen langs de
inlaat naar binnen stromen. Dit heeft enige invloed op de geometrie. Lege ruimtes die
tijdens het proces niet gevuld worden met bloed worden verwijderd.
Het roteren van een geometrie in Fluent gebeurt door gebruik te maken van een rotating
reference frame (draaiend referentie assenstelsel). Hierbij worden extra termen toegevoegd
in de momentum vergelijking, zoals de Coriolis kracht. De gebruiker heeft de keuze om
de snelheid relatief of absoluut te beschouwen. Doordat in dit geval de centrifuge volledig
draait wordt de snelheid relatief of ten opzichte van het draaiend referentiekader geformu-
leerd. De snelle rotaties van de centrifuge zorgen voor extreme gradienten in de vloeistof.
Om dit in een stap uit te rekenen geraakt Fluent in moeilijkheden. Om dit te verhelpen
wordt de rotatiesnelheid van de centrifuge in opeenvolgende stappen opgevoerd. Het pro-
bleem wordt dus eerst stationair beschreven totdat het gewenste toerental bereikt wordt.
Dan wordt het probleem niet-stationair beschreven en wordt het bloed in de centrifuge
gepompt.
2 Het geometrisch model in Gambit
Het programma Gambit dient om de geometrie van het model te maken. Als de geometrie
geconstrueerd is, wordt hierop een mesh gelegd. Een mesh verdeelt het geconstrueerd
volume in kleine cellen waarop Fluent de eindige-volume methode toepast. De fijnheid
van de mesh kan worden ingesteld. Hoe fijner de mesh, hoe nauwkeuriger de berekeningen
in Fluent, maar hoe meer rekentijd het vergt om een convergentie van de oplossing te
bekomen. Hoe groter de mesh-cellen, hoe onnauwkeuriger de oplossing, maar hoe sneller

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 80
elke iteratiestap1 wordt berekend.
2.1 Geometrie
Figuur 6.1 toont de geometrie en de afmetingen van de centrifuge zoals die werd ontvan-
gen van de fabrikant Dideco. Niet alle afmetingen worden weergegeven maar met een
computeranalyse van de figuur kunnen al de coordinaten van de punten worden bepaald.
Figuur 6.1: Afmetignen van de Dideco centrifuge, 125ml.
Om de rekenkracht te beperken wordt het driedimensioneel ontwerp gemodelleerd in een
symmetrisch tweedimensioneel model. De inlaat en de uitlaat worden niet volledig getekend
in Gambit zodat enkel de relevante delen worden gemodelleerd. De lege ruimtes, die
niet worden opgevuld met bloed, worden uit de geometrie gesneden. Het resultaat wordt
weergegeven in figuur 6.2.
1De term iteratiestap wordt hier gebruikt als een uitgevoerde berekeningstap, de term iteratie duidt opde reeks berekeningen die uitgevoerd worden voor een bepaalde tijdsintervalstap

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 81
Figuur 6.2: Grid van de geroteerde centrifuge
a Axis
b Velocity inlet
c Outflow
d Internal
Overige Wall
F1-F6 Zone 1-6
2.2 Zones
In Gambit worden verschillende zones gedefinieerd. Aan elke ingegeven lijn en oppervlak
wordt een bepaalde functie toegewezen. In Fluent wordt deze die functie gedefinieerd. Zo
worden de lijnstukken, aangeduid met de kleine letters in figuur 6.2, als volgt gedefinieerd
(Engelse termen zoals in Gambit).
a Axis Definieert de rotatieas
b Velocity inlet Inlaat, debiet bepaald door de snelheid van het fluıdum
c Outflow Definieert de uitlaat zonder enige voorwaarde
d Internal Virtuele lijnen die de zones onderscheiden van elkaar
Overige Wall Muur waarlangs geen slip optreedt

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 82
2.3 Mesh
Nu de centrifuge opgesplitst is in zones kan er op verschillende manieren een mesh opgelegd
worden, zodat bepaalde delen nauwkeuriger berekend worden. Er wordt een quadrangulair
grid gebruikt. In een eerste fase van het modelleren werd geexperimenteerd met een grove
mesh om de rekentijd te doen dalen maar deze gaf divergerende resultaten. Het uiteindelijk
aantal gekozen meshelementen per zone wordt weergegeven in tabel 6.1 (ref. figuur 6.2).
Alle zones samen worden gedefinieerd als fluıdum.
Tabel 6.1: Aantal meshelementen
Zone Aantal
F1 3405
F2 5245
F3 2831
F4 11456
F5 4517
F6 699
3 Het numeriek model in Fluent
In deze paragraaf wordt de lezer geleid door de verschillende stappen die nodig zijn voor
de modelopbouw in Fluent. Waar nodig wordt er dieper ingegaan op de theoretische
achtergrond van het model.
3.1 Inladen van de geometrie
In Fluent wordt de geometrie en de mesh ingeladen. Deze mesh wordt eerst op schaal
gebracht en vervolgens geroteerd omdat Fluent eist dat de rotatieas op de X-as ligt. In
figuur [?] werd de geroteerde centrifuge afgebeeld. Een automatische controle van de mesh
bepaalt tenslotte dat er geen fouten zijn en dat er kan overgegaan worden naar de volgende
stap.

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 83
3.2 Modelkeuze
In het define menu wordt het rekenkundig model gedefinieerd. Hierin bevindt zich het
solver submenu, die de manier van uitrekenen bepaalt, en het multiphase submenu, die de
interactie tussen de fasen omschrijft.
3.2.1 Solver
In dit submenu wordt gedefinieerd op welke wijze er gerekend moet worden. Zo wordt
er aangeduid dat het probleem axisymmetrisch is zodat de vergelijkingen hieraan worden
aangepast. Ook de keuze dat de snelheid relatief, dus t.o.v. het referentie assenstelsel
wordt geformuleerd wordt hier vastgelegd. De tijdsafhankelijkheid van het probleem wordt
gespecifieerd. Tijdens het stelselmatig opdrijven van de rotaties van de centrifuge wordt
het probleem stationair omschreven. Eenmaal het vereiste toerental bereikt is, wordt het
probleem niet-stationair omschreven.
3.2.2 Multiphase
Dit submenu laat de gebruiker toe om een meerfase model te kiezen. In Fluent zijn er
drie mogelijkheden om een meerfase stroming te modelleren, het VOF model, het mixture
model en het Euler model.
• Het VOF (Volume of Fluid) model wordt gebruikt om niet interagerende fluıda te
modelleren. Het scheidingsoppervlak tussen deze vloeistoffen is van belang. De fluıda
worden beschreven als een enkel fluıdum. De berekeningen gebeuren dus met een set
van vergelijkingen waarbij de variabelen per meshelement berekend worden in functie
van de volume fracties.
• Het mixture model modelleert interagerende fasen. Deze fasen kunnen vloeistoffen
of deeltjes zijn. Net zoals het VOF model beschrijft het de meerfase stroming als
een eenfasestroming. Zo worden de behoudsvergelijkingen van massa en momentum
opgelost van het mengsel. Dit model kan worden toegepast voor stromingen geladen
met een klein aantal deeltjes.
• Het Euler model is het meest complexe model om meerfasestroming te modelleren.
Deze fasen kunnen eveneens vloeistoffen of deeltjes zijn. Het lost dan voor elke fase

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 84
de behoudsvergelijkingen op. De interactie tussen de fasen wordt aan de hand van
coefficienten verwezenlijkt.
Al deze modellen worden berekend op de Euleriaanse methode, maar het laatste model
is het volledigst en wordt volgens de nomenclatuur van Fluent het Euler model genoemd.
Fluent beschikt naast deze modellen ook over een Langrangiaanse module die de beweging
van deeltjes in een vloeistof kan berekenen, genaamd discrete phase model. Dit model is
echter niet toereikend om stromingen met een volumefractie groter den 10% te modelleren.
3.3 Het Euler model
Van bovenstaande modellen is slechts het Euler model in staat om de sedimentatie van
deeltjes in hoge concentraties te beschrijven. Door de complexiteit van het granulair model
wordt de berekeningsmethode die gebruikt wordt in dit model iets nauwer toegelicht. Voor
een complete voorstelling van het rekenkundig model wordt de lezer verwezen naar de
handleiding van Fluent [18].
3.3.1 Behoudswetten
De vergelijking voor behoud van massa zoals die wordt uitgerekend door Fluent is:
∂αiρi
∂t+∇ · (αiρi~vi) = 0 (6.1)
met:
αi de volume fractie van fase i
ρi de dichtheid van fase i
~vi de snelheid van fase i
De volumefractie van de primaire fase wordt niet expliciet op deze manier uitgerekend.
Doordat de som van volumefracties een bedraagt kan door het oplossen van deze vergelij-
king voor elke secundaire fase de volumefractie van de primaire fase bepaald worden.
Gebaseerd op verscheidene studies [19] [20] [21] [22] gebruikt Fluent een speciaal granulair
model om het gedrag van een mengsel bestaande uit een vloeistof en deeltjes te beschrijven.
De vergelijking die Fluent uitrekent voor behoud van momentum is:

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 85
∂(αiρi~vi)
∂t+∇ · (αiρi~vi~vi) = −αi∇p−∇pi +∇ · τi + αiρi~g +
N∑j
(Kji(~vj − ~vi)) (6.2)
met:
p de totale druk gedeeld door alle fasen
pi de partieeldruk van vaste fase i
Kji de momentum uitwisselingscoefficienten
N aantal fasen
In volgende paragrafen wordt uitgelegd hoe de momentum uitwisselingscoefficienten, de
partieeldruk van de vaste fase en de spanningsrektensor berekend worden.
3.3.2 De momentum uitwisselingscoefficienten
De momentum uitwisselingscoefficienten bepalen de overdracht van momentum tussen de
fasen. Voor vloeistof-vloeistof zijn deze uitwisselingscoefficienten anders dan voor vloeistof-
vast. De vloeistof-vast coefficient die in dit numeriek model gebruikt wordt is gedefinieerd
als:
Kji =αiρif
τi
(6.3)
waarbij τi de deeltjes relaxatie tijd voorstelt:
τi =ρid
2i
18µl
(6.4)
met:
di de diameter van de deeltjes van fase i
µl de viscositeit van het fluıdum
De parameter f in formule 6.3 wordt berekend door verscheidene correlaties naargelang
het probleem. Alle definities van f bezitten de wrijvingscoefficient die gebaseerd is op het
Reynolds getal. De correlatie die de gebruiker wenst wordt gekozen in het Phases scherm.
In dit numeriek model wordt gekozen voor de correlatie van Syamal et al [24]:
f =CDReiαi
24v2r,i
(6.5)

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 86
met CD:
CD =
(0.63 +
4.8√Rei − vr,i
)2
(6.6)
met Rei het relatief Reynolds getal:
Rei =ρldivr,i
µl
(6.7)
met vr,i de relatieve snelheid van i:
vr,i = |~vi − ~vl| (6.8)
3.3.3 De partieeldruk van de vaste fase
Voor granulaire stromen wordt onafhankelijk een partieeldruk van de vaste fase berekend.
Deze wordt gebruikt om de gradient te berekenen in vergelijking 6.2. Doordat een Maxwell
snelheidsverdeling gebruikt wordt om de deeltjes te beschrijven voert Fluent de granulaire
temperatuur in. Deze granulaire temperatuur is proportioneel met de kinetische energie
van de random bewegingen van de deeltjes. Ze komt voor in de uitdrukkingen van de
viscositeit en van de partieeldruk van de vaste fase. Zo bevat de uitdrukking van de
partieeldruk volgens Syamlal et al. [24] een kinetische term en een term te wijten aan de
botsingen:
pi = αiρiΘi + 2ρi(1 + eii)α2i g0,iiΘi (6.9)
met:
Θi de granulaire temperatuur
eii de restitutiecoefficient of het aandeel aan snelheid dat behouden blijft na botsing
g0,ii de radiale distributiefunctie
De restitutiecoefficient is een belangrijke factor in de berekeningen. Standaard heeft ze de
waarde van 0.9. Voor bloedcellen is deze theoretische waarde echter niet bekend. De radiale
distributiefunctie is een correctiefactor die de kans op een botsing voorstelt. Ze bepaalt de
overgang van de samendrukbare toestand, waarbij het volume tussen de deeltjes kan blijven
dalen, naar de niet samendrukbare toestand, waar de deeltjes niet meer kunnen bewegen.
Verscheidene uitdrukkingen zijn beschikbaar, hier werd gekozen voor de uitdrukking van
Arastoopour et al.[23]:

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 87
g0,ii =1
(1− αi
αi,max)
+3
2di
N∑k=1
αk
dk
(6.10)
met:
αi,max de maximum pakkingsdichtheid van fase i
N aantal fasen
3.3.4 De spanningstensor
De spanningstensor τi van fase i wordt gedefinieerd als:
τi = αiµi(∇~vi +∇~vTi ) + αi(λi −
2
3µi)∇ · ~viI (6.11)
met:
µi de viscositeit van i
λi de bulkviscositeit van i
I eenheidstensor
De bulkviscositeit bepaalt de bijdrage van de normaalspanningen tot de spannigstensor.
Ze beschrijft de weerstand van de deeltjes tegen samendrukbaarheid en expansie. Lun et
al. [20] stelden volgende functie voor:
λi =4
3αiρidig0,ii(1 + eii)
(Θi
pi
)1/2
(6.12)
De viscositeit bepaalt de bijdrage van de schuifspanningen tot de spannigstensor. Ze is het
gevolg van de uitwisseling van momentum door translatie en botsingen. Ze kan worden
ontbonden in drie componenten: een botsingscomponent µi,col, een kinetische component
µi,kin en een wrijvingscomponent µi,fr.
µi = µi,col + µi,kin + µi,fr (6.13)
Het botsingsgedeelte van de viscositeit wordt gemodelleerd als:
µi,col =4
5αiρidig0,ii(1 + eii)
(Θi
π
)1/2
(6.14)
De kinetische component kan berekend worden volgens Gidaspow [22] of volgens Syamlal
et al. [24]. De uitdrukking van Syamal et al. past beter met de andere gekozen modellen:

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 88
µi,kin =αidiρi
√Θiπ
6(3− eii)
[1 +
2
5(1 + eii) (3eii − 1) αsg0,ii
](6.15)
De wrijvingscomponent wordt berekend volgens Schaeffer et al. [25]:
µi,rmfr =pi sin φ
2√
I2D
(6.16)
met:
φ de hoek van interne wrijving hier 300
I2D de tweede invariant van de spanningstensor τi
3.4 Materialen
Terug in Fluent worden nu in het materials menu de materialen gedefinieerd. Hier worden
de dichtheden van het plasma en de rode bloedcellen ingegeven en eveneens de viscositeit
van het plasma. De dichtheid van plasma bedraagt 1026 kg/m3, die van rode bloedcellen
1100 kg/m3. Uit formule 2.32 wordt de viscositeit van het plasma berekend bij 22 �: 1.76
mPa s.
3.5 Fasen
Het phases menu laat de gebruiker toe om de parameters van het granulair model in te geven
en de interactie tussen de fasen te definieren. Het plasma krijgt het label phase 1 toegewezen
en de rode bloedcellen phase 2. De eerste fase stelt altijd het suspenderend fluıdum voor.
De tweede fase wordt gemodelleerd als granulaire substantie. Heel wat gegevens worden in
dit menu verzameld zodat het Euler model het probleem correct berekent. De gebruiker kan
kiezen tussen enkele modellen of kan bepaalde variabelen vervangen door zelf geschreven
functies.
In het fase menu wordt eveneens de diameter van de granulaire fase ingegeven. Bij de
simulaties met runderbloed is de diameter 5.7 µm. Bij humaanbloed is de diameter van
het zinkend agglomeraat lokaal afhankelijk van het hematocriet. Daarom wordt er gebruik
gemaakt van een user defined function (UDF). Een UDF is een geprogrammeerde functie
dat dynamisch geladen wordt in Fluent om de standaard code te verrijken. Deze UDFs
worden in de programmeertaal C geschreven. UDFs kunnen een verscheidenheid aan ta-
ken invullen in Fluent. Ze kunnen een waarde terugsturen, of een argument of variabele

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 89
veranderen, en input/output taken uitvoeren. Om een functie te definieren worden DEFI-
NE macro’s gebruikt. Om deze macro’s te kunnen gebruiken moet elke UDF starten met
include udf.h. De UDF die hier gebruikt wordt ziet er als volgt uit:
#include ′′udf.h′′
DEFINE PROPERTY(diameter,c,t)
{real diam;
real vol = C VOF(c,t);
diam=3.39067e-6*2*pow(vol+0.649351,1./3.);
return diam;
}
De macro DEFINE PROPERTY wordt gebruikt om een bepaalde materiaal eigenschap te
veranderen. Het eerste argument van deze macro is de naam van de UDF, c en t stellen
pointers voor naar een bepaalde cel. Fluent loopt over al de computationele cellen in de
geometrie via deze pointers. De macro C VOF(c,t) haalt de waarde van de volumefractie
van de granulaire fase. Voor elke waarde van c en t wordt de diameter gedefinieerd in functie
van de volume fractie. De diameter wordt dan bepaald door (zie vergelijking (4.1). Nadat
Fluent deze UDF gecompileerd heeft, kan deze geselecteerd worden in het keuzepaneel voor
de diameter van de granulaire fase.
In tabel 6.2 staan de andere gegevens die in dit paneel worden ingegeven. De keuzes van de
correlaties werden gebaseerd op basis van de informatie beschikbaar in de handleiding van
Fluent. De maximum pakkingsdichtheid voor sferische deeltjes is 0.63 maar bij bloedcellen
kunnen de concentraties veel hoger oplopen omdat de cellen zich efficient stapelen en bijna
geen vrije ruimte toelaten.
Verder, in het interactiepaneel, wordt voor f die voorkomt in de uitdrukking van de mo-
mentum uitwisselingscoefficient(vergelijking (6.3))de correlatie van Syamal [24] gekozen.
De waarde van de restitutiecoefficient bedraagt 0.9.
3.6 De Randvoorwaarden
Nu dat het model en de parameters gekozen zijn, kunnen de randvoorwaarden of boundary
conditions worden opgelegd. De functie die in gambit werd gegeven aan een bepaald lijnstuk

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 90
Tabel 6.2: Parameters van het granulair model
Eigenschap (E) Eigenschap (Nl) Keuze Formule
Diameter Diameter UDF 3.5
Granulair viscosity Kinetische component Syamlal et al.[24] 6.15
Bulk viscosity Bulkviscositeit Lun et al.[20] 6.13
Frictional viscosity Wrijvingscomponent Schaeffer [25] 6.16
Angular of friction Interne wrijvingshoek 30 6.16
Solids pressure Partieeldruk van de vaste fase Syamlal et al. [24] 6.9
Radial distribution Radiale distributiefunctie Arastoopour et al. [23] 6.10
Packing limit Maximum pakkingsdichtheid 0.90 6.10
of volume wordt hier gedefinieerd. Het referentie assenstelsel draait rond en aan de inlaat
stroomt het bloed binnen met een bepaald hematocriet en snelheid. Beide condities worden
in dit paneel aan het model toegevoegd.
3.6.1 De inlaat
Tijdens de experimenten was de vulsnelheid 100, 200 en 300 ml/min. In Fluent moet het
debiet worden ingegeven als snelheid loodrecht op het doorstromend oppervlak. Aan de
hand van de diameter van de inlaat (4.5 mm) wordt de snelheid berekend:
Tabel 6.3: Snelheid aan de inlaat
Vulsnelheid [ml/min] Snelheid [m/s]
100 0.10479
200 0.20959
300 0.31438
Deze waarden moeten ingegeven worden voor beide fasen. De volumefractie is tijdens het
stelselmatig opvoeren van de rotatiesnelheid nul. Eens de centrifuge op volle toeren draait
kan de volumefractie worden aangepast en kunnen de eigenlijke simulaties starten.
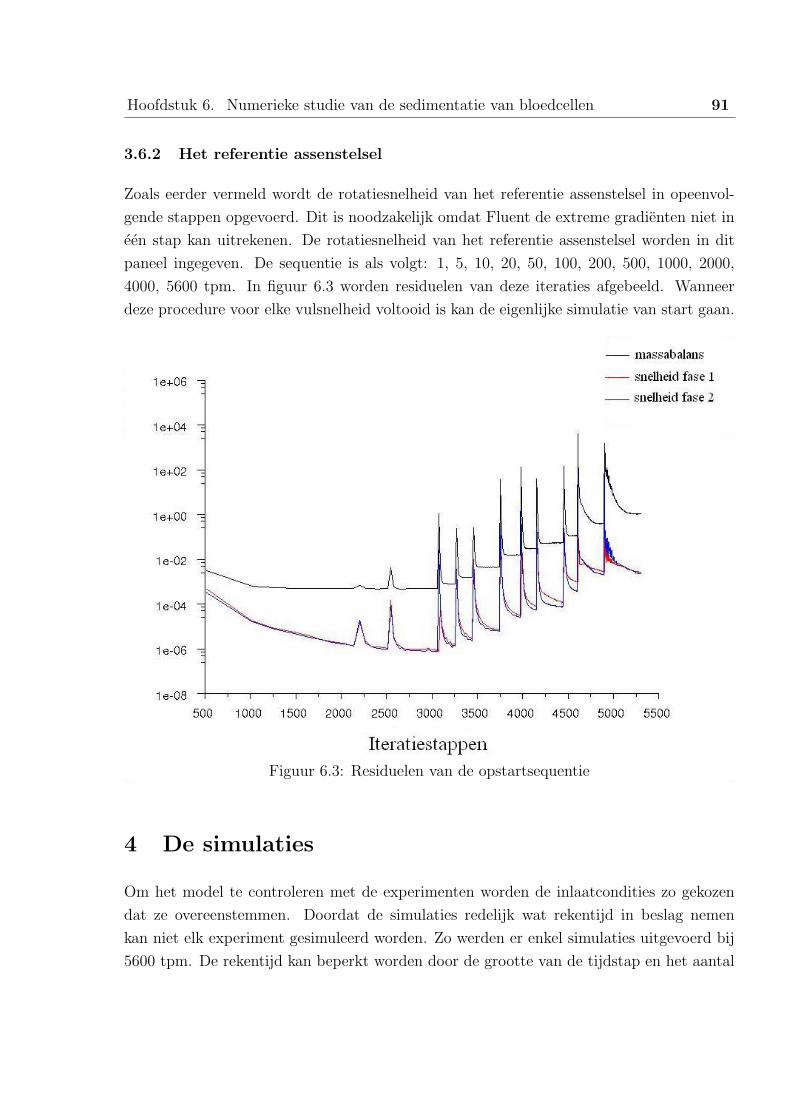
Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 91
3.6.2 Het referentie assenstelsel
Zoals eerder vermeld wordt de rotatiesnelheid van het referentie assenstelsel in opeenvol-
gende stappen opgevoerd. Dit is noodzakelijk omdat Fluent de extreme gradienten niet in
een stap kan uitrekenen. De rotatiesnelheid van het referentie assenstelsel worden in dit
paneel ingegeven. De sequentie is als volgt: 1, 5, 10, 20, 50, 100, 200, 500, 1000, 2000,
4000, 5600 tpm. In figuur 6.3 worden residuelen van deze iteraties afgebeeld. Wanneer
deze procedure voor elke vulsnelheid voltooid is kan de eigenlijke simulatie van start gaan.
Figuur 6.3: Residuelen van de opstartsequentie
4 De simulaties
Om het model te controleren met de experimenten worden de inlaatcondities zo gekozen
dat ze overeenstemmen. Doordat de simulaties redelijk wat rekentijd in beslag nemen
kan niet elk experiment gesimuleerd worden. Zo werden er enkel simulaties uitgevoerd bij
5600 tpm. De rekentijd kan beperkt worden door de grootte van de tijdstap en het aantal
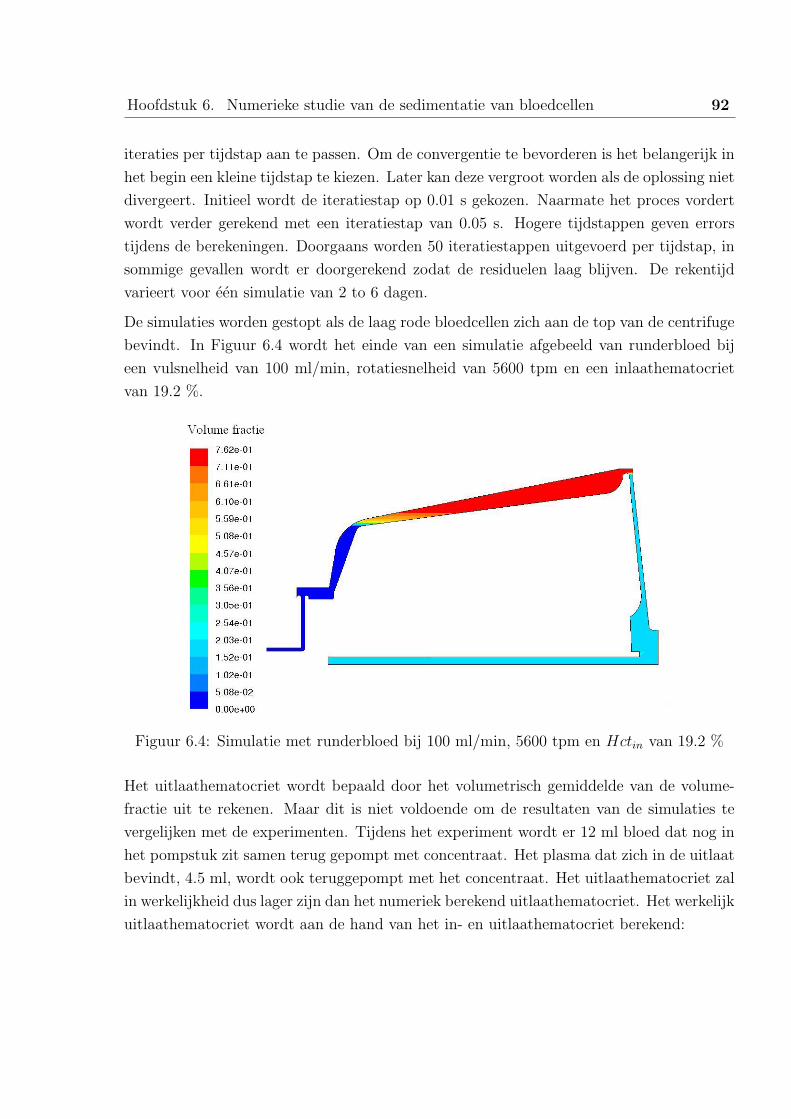
Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 92
iteraties per tijdstap aan te passen. Om de convergentie te bevorderen is het belangerijk in
het begin een kleine tijdstap te kiezen. Later kan deze vergroot worden als de oplossing niet
divergeert. Initieel wordt de iteratiestap op 0.01 s gekozen. Naarmate het proces vordert
wordt verder gerekend met een iteratiestap van 0.05 s. Hogere tijdstappen geven errors
tijdens de berekeningen. Doorgaans worden 50 iteratiestappen uitgevoerd per tijdstap, in
sommige gevallen wordt er doorgerekend zodat de residuelen laag blijven. De rekentijd
varieert voor een simulatie van 2 to 6 dagen.
De simulaties worden gestopt als de laag rode bloedcellen zich aan de top van de centrifuge
bevindt. In Figuur 6.4 wordt het einde van een simulatie afgebeeld van runderbloed bij
een vulsnelheid van 100 ml/min, rotatiesnelheid van 5600 tpm en een inlaathematocriet
van 19.2 %.
Figuur 6.4: Simulatie met runderbloed bij 100 ml/min, 5600 tpm en Hctin van 19.2 %
Het uitlaathematocriet wordt bepaald door het volumetrisch gemiddelde van de volume-
fractie uit te rekenen. Maar dit is niet voldoende om de resultaten van de simulaties te
vergelijken met de experimenten. Tijdens het experiment wordt er 12 ml bloed dat nog in
het pompstuk zit samen terug gepompt met concentraat. Het plasma dat zich in de uitlaat
bevindt, 4.5 ml, wordt ook teruggepompt met het concentraat. Het uitlaathematocriet zal
in werkelijkheid dus lager zijn dan het numeriek berekend uitlaathematocriet. Het werkelijk
uitlaathematocriet wordt aan de hand van het in- en uitlaathematocriet berekend:

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 93
Hctuit werkelijk =Hctuit numeriek · 133ml + Hctin · 12ml
133ml + 12ml + 4.5ml(6.17)
5 De resultaten
In deze paragraaf worden de resultaten van de simulaties besproken. In een eerste luik wor-
den de simulaties uitgevoerd met runderbloed terwijl in het tweede luik het humaanbloed
gesimuleerd wordt.
5.1 Simulaties met runderbloed
De resultaten van de simulaties en de experimenten worden weergegeven in tabel 6.4,
grafisch in figuur 6.5.
Tabel 6.4: Resultaten van de simulaties met runderbloed
Vulsnelheid [ml/min] Hctin Hctuit numeriek Hctuit werkelijk Hctuit experimenteel
300 33.4 40.3 38.5 46.9
300 20 28.3 26.8 42.7
300 9.5 21.8 20.1 40.2
200 33.4 44.1 41.9 51.6
100 33.4 60.2 56.1 55.1
100 19.2 55.9 51.3 50.9
100 9.5 52.7 47.7 46.9
De simulaties bij 100 ml/min tonen een degelijke overeenkomst. Bij andere vulsnelheden
onderschat het model de sedimentatie van rode bloedcellen. Dit wijst erop dat bij hogere
vulsnelheden Fluent de interactie tussen de cellen en het fluıdum niet juist modelleert. Dit
kan zijn omdat de weerstand die de bloedcellen ondervinden te groot berekend wordt. Dit
is mogelijk te verklaren door de niet sferische vorm van een runderbloedcel. Een andere
mogelijkheid is dat de cellen in realiteit een sterke cohesie hebben waardoor ze moeilijk
door de stroming worden beınvloed. Ze plakken als het ware beter tegen elkaar waardoor
ze zich beter kunnen verzetten tegen de stroming die ze wilt meesleuren naar de uitlaat. In
de simulaties is het duidelijk dat de bloedcellen sterker onderhevig zijn aan de stromingen
in de centrifuge. Bij hoge vulsnelheden is de afscheidingslijn tussen de rode bloedcellen
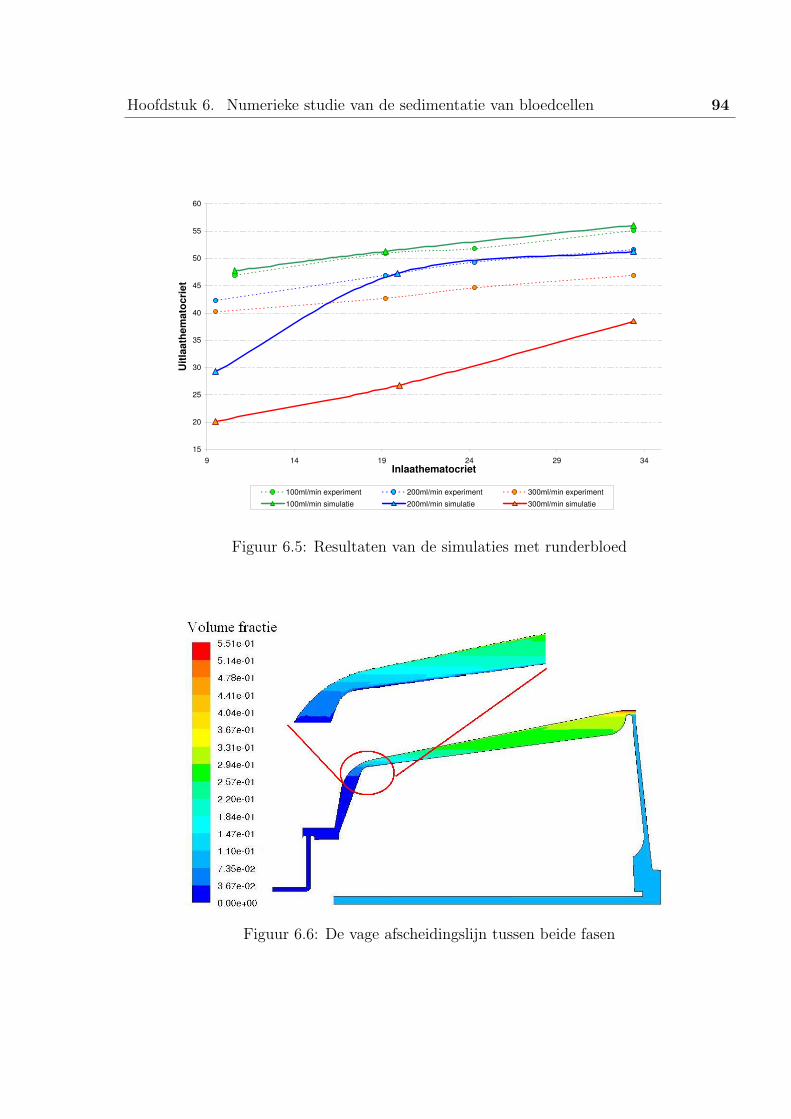
Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 94
15
20
25
30
35
40
45
50
55
60
9 14 19 24 29 34
Inlaathematocriet
Uit
laath
em
ato
crie
t
100ml/min experiment 200ml/min experiment 300ml/min experiment
100ml/min simulatie 200ml/min simulatie 300ml/min simulatie
Figuur 6.5: Resultaten van de simulaties met runderbloed
Figuur 6.6: De vage afscheidingslijn tussen beide fasen
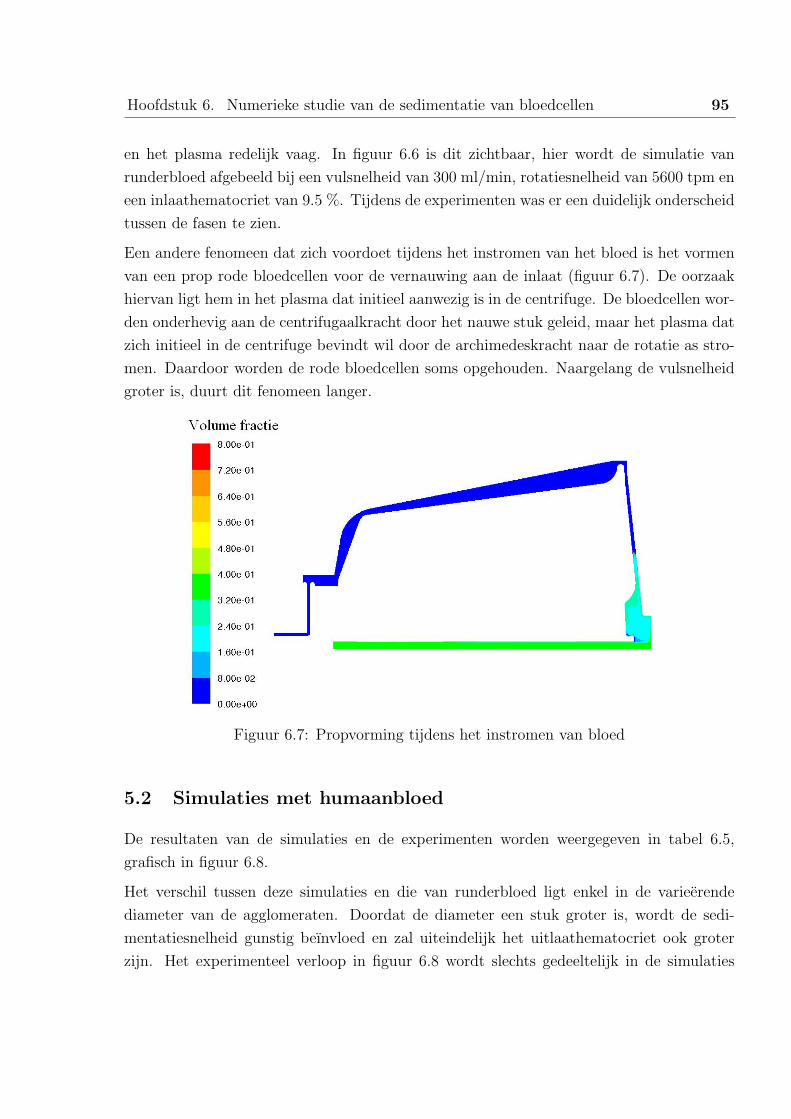
Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 95
en het plasma redelijk vaag. In figuur 6.6 is dit zichtbaar, hier wordt de simulatie van
runderbloed afgebeeld bij een vulsnelheid van 300 ml/min, rotatiesnelheid van 5600 tpm en
een inlaathematocriet van 9.5 %. Tijdens de experimenten was er een duidelijk onderscheid
tussen de fasen te zien.
Een andere fenomeen dat zich voordoet tijdens het instromen van het bloed is het vormen
van een prop rode bloedcellen voor de vernauwing aan de inlaat (figuur 6.7). De oorzaak
hiervan ligt hem in het plasma dat initieel aanwezig is in de centrifuge. De bloedcellen wor-
den onderhevig aan de centrifugaalkracht door het nauwe stuk geleid, maar het plasma dat
zich initieel in de centrifuge bevindt wil door de archimedeskracht naar de rotatie as stro-
men. Daardoor worden de rode bloedcellen soms opgehouden. Naargelang de vulsnelheid
groter is, duurt dit fenomeen langer.
Figuur 6.7: Propvorming tijdens het instromen van bloed
5.2 Simulaties met humaanbloed
De resultaten van de simulaties en de experimenten worden weergegeven in tabel 6.5,
grafisch in figuur 6.8.
Het verschil tussen deze simulaties en die van runderbloed ligt enkel in de varieerende
diameter van de agglomeraten. Doordat de diameter een stuk groter is, wordt de sedi-
mentatiesnelheid gunstig beınvloed en zal uiteindelijk het uitlaathematocriet ook groter
zijn. Het experimenteel verloop in figuur 6.8 wordt slechts gedeeltelijk in de simulaties
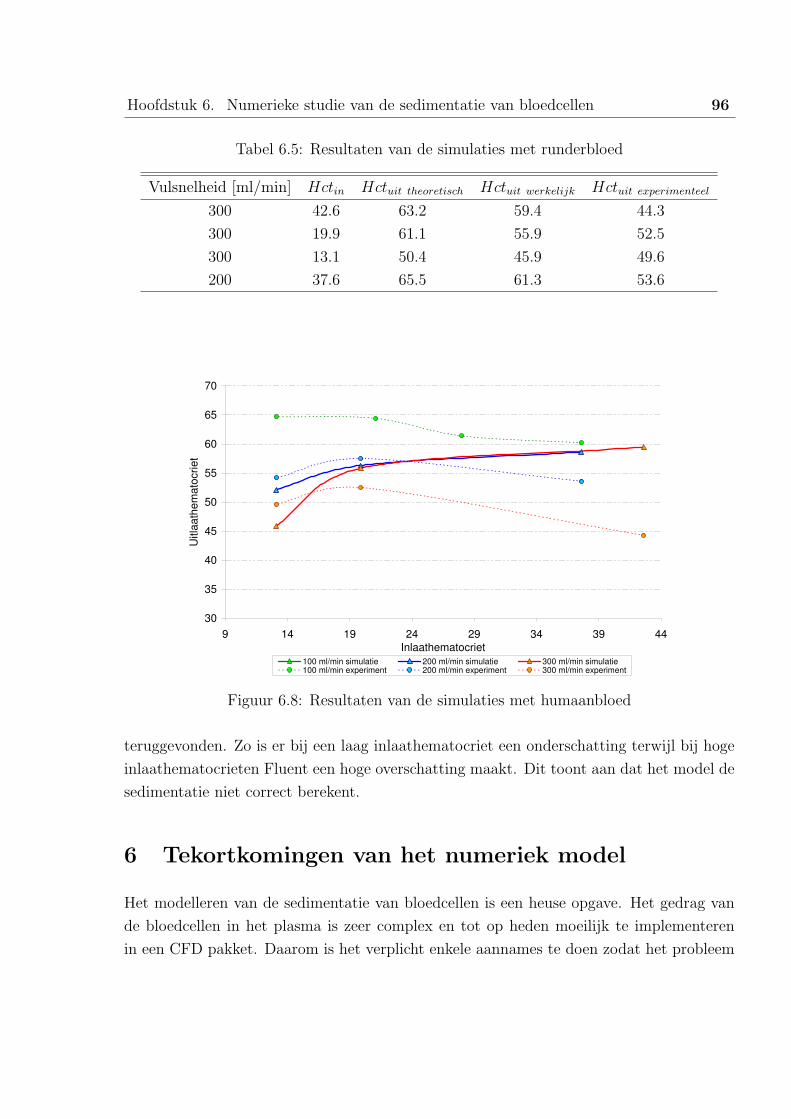
Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 96
Tabel 6.5: Resultaten van de simulaties met runderbloed
Vulsnelheid [ml/min] Hctin Hctuit theoretisch Hctuit werkelijk Hctuit experimenteel
300 42.6 63.2 59.4 44.3
300 19.9 61.1 55.9 52.5
300 13.1 50.4 45.9 49.6
200 37.6 65.5 61.3 53.6
30
35
40
45
50
55
60
65
70
9 14 19 24 29 34 39 44
Inlaathematocriet
Uitla
ath
em
ato
cri
et
100 ml/min simulatie 200 ml/min simulatie 300 ml/min simulatie100 ml/min experiment 200 ml/min experiment 300 ml/min experiment
Figuur 6.8: Resultaten van de simulaties met humaanbloed
teruggevonden. Zo is er bij een laag inlaathematocriet een onderschatting terwijl bij hoge
inlaathematocrieten Fluent een hoge overschatting maakt. Dit toont aan dat het model de
sedimentatie niet correct berekent.
6 Tekortkomingen van het numeriek model
Het modelleren van de sedimentatie van bloedcellen is een heuse opgave. Het gedrag van
de bloedcellen in het plasma is zeer complex en tot op heden moeilijk te implementeren
in een CFD pakket. Daarom is het verplicht enkele aannames te doen zodat het probleem

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 97
gesimuleerd kan worden in Fluent. Deze aannames hebben echter een grote invloed op het
proces waardoor de resultaten van de simulaties niet overeenkomen met de experimentele
resultaten. Men kan zich afvragen waarom de theorien beschreven in hoofdstuk 2 niet
gebruikt worden bij dit numeriek model. De expliciete uitdrukkingen van de sedimentatie-
snelheid kunnen echter niet gecombineerd worden met de behoudsvergelijkingen (6.2) van
het Euler model van Fluent.
6.1 Vorm van de bloedcellen
De vorm van een bloedcel wordt tijdens de simulaties sferisch verondersteld want het
Euler model is enkel instaat om het gedrag van sferische deeltjes te beschrijven. Het
Langragiaans model kan niet-sferische deeltjes modelleren maar is dan weer niet in staat
om hoge volumefracties te simuleren.
Figuur 6.9: Agglutinatie (links) en rouleaux vorming (rechts)
Humane bloedcellen zijn niet sferisch maar schijfvormig. Doordat ze identiek zijn kunnen
ze zich stapelen en agglomereren. Normaal hebben de cellen een lage affiniteit voor elkaar
omdat ze negatieve ladingen op het celoppervlak bezitten. Maar bepaalde proteınen in het
plasma gaan zich aan het oppervlak vasthechten zodat de negatieve ladingen verminderen.
De cellen stapelen zich dan lineair, er treedt primaire rouleaux vorming op. Secundaire
rouleauxvorming of agglutinatie treedt op vanaf een bepaald hematocriet. De bloedcellen
klonteren samen in plaats van zich op elkaar te stapelen. Het onderscheid tussen deze
twee fenomenen wordt weergegeven in figuur 6.9. Bij lage hematocrieten doet zich eerst de
lineaire of primaire rouleaux vorming voor, bij hogere hematocrieten de secundaire rouleaux

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 98
vorming. De grens tussen deze twee fenomenen is vaag. Naar gelang de aanwezigheid van
bepaalde proteınen en antilichamen worden de agglomeratie fenomenen beınvloed.
De complexe vormen van de agglomeraten maken het zeer moeilijk om het sedimentatie-
proces correct te simuleren. In dit numeriek model groeien de agglomeraten sferisch aan,
terwijl in werkelijkheid ze eerst lineair aangroeien. De bezinkingssnelheid is voor een lineair
agglomeraat groter dan voor een sfeer omdat een lineair stapeltje een kleinere wrijvings-
kracht ondervindt. De bezinkingssnelheid wordt dus onderschat bij lage hematocrieten.
De sferische benadering gaat ook niet op voor de agglomeraten, in werkelijkheid zijn dit
hoopjes van willeukeurige vorm. De wrijvingskracht van een willekeurig hoopje is groter
dan die van een sfeer, de bezinkingssnelheid wordt dus overschat bij hogere hematocrieten.
6.2 Botsingen tussen de bloedcellen
Naast de vorm van de bloedcellen is de interactie tussen de cellen ook belangrijk. De
bloedcellen kunnen op twee manieren reageren op botsingen, ze kunnen weerkaatsen of
ze kunnen agglomereren. In dit model wordt enkel ondersteld dat bij elke botsing een
bepaald aandeel aan kinetische energie verloren gaat. Het model in Fluent benadert het
probleem omdat het de mogelijkheid uitsluit dat cellen gaan agglomereren in plaats van te
weerkaatsen.
Door het kiezen van de restitutiecoefficient, dit is het aandeel van de snelheid dat behouden
blijft na een botsing, kan het aandeel aan kinetische energie dat verloren gaat worden
ingesteld. Standaard heeft deze een waarde van 0.9. Twee simulaties van humaanbloed,
uitgevoerd bij een vulsnelheid van 300 ml/min en een inlaathematocriet van 33.4 %, werden
berekend met een restitutiecoefficient van 0.99 en 0.80. Het verschil in uitlaathematocriet
blijf echter gering, respectivelijk 55.2% en 56.6%.
7 Toekomstperspectieven
De simulatie van het complexe sedimentatieproces is een grote uitdaging. De sleutel tot
een goede oplossing is de kennis van de agglomeratiefenomenen en hun invloed op de
wrijvingskracht. Deze complexe fenomenen zijn moeilijk te simuleren in een commercieel
beschikbaar pakket zoals Fluent. Het pad naar een succesrijk model moet dus niet langs
een puur theoretische weg gezocht worden.

Hoofdstuk 6. Numerieke studie van de sedimentatie van bloedcellen 99
Een van de mogelijke wegen is een trial and error2 methode toepassen op bepaalde pa-
rameters of correlaties. Er werd al gevonden dat de restitutiecoefficient niet bepaald veel
invloed heeft. Wat wel een grote invloed heeft is de correlatie van f die voorkomt in de
uitdrukking van de momentum uitwisselingscoefficient (vergelijking (6.3)). Doordat deze
de overdracht van momentum bepaalt tussen de twee fasen is het deze correlatie die de
sedimentatiesnelheid bepaalt. Ze werd gekozen volgens een correlatie van Syamal [24],
maar de resultaten tonen aan dat hier nog aan gesleuteld moet worden. Mits gebruik te
maken van de experimentele resultaten en vele test simulaties zou er een betere correlatie
vooropgesteld kunnen worden die de sedimentatie juister beschrijft. Het probleem is echter
dat de simulaties veel rekentijd vragen en dat de correlatie juiste resultaten moet geven op
alle inlaatcondities.
Eens dit model gevonden, kan het gebruikt worden voor tal van toepassingen. Het model
kan evenwel nog uitgebreid worden zodat de sedimentatie van de witte bloedcellen en de
bloedplaatjes ook gesimuleerd wordt. Aan de hand van een parameterstudie kan onderzocht
worden met welke parameters de beste afscheiding van de bloedplaatjes verkregen wordt.
Indien het inlaathematocriet gekend is kan de centrifuge zijn parameters aanpassen zodat
de afscheiding optimaal is. Het ultieme doel is aan de hand van een numeriek model
een centrifuge te ontwerpen die de afscheiding van de rode bloedcellen of bloedplaatjes
maximaliseert.
8 Besluit
In dit hoofdstuk werd een numeriek model beschreven dat de sedimentatie van bloedcellen
simuleert. De resultaten komen slechts voor enkele simulaties overeen met het experiment.
Het model is dus niet in staat om de sedimentatie correct te beschrijven. De oorzaak
ligt in de hoge complexiteit van bloedstroming. Het gedrag van de bloedcellen is moeilijk
te omschrijven, zeker in een krachtig centrifugaal veld met complexe stromingen. Door
verder onderzoek uit te voeren naar de parameters van het nummeriek model kan een
model gevonden worden dat de sedimentatie correct beschrijft.
2De trial and error methode is een manier van kennis verwerven over een bepaald proces. Hierbij wordteen optie geprobeerd en gecontroleerd. Als deze goed is wordt deze aanvaard, als ze fout is wordt de foutonderzocht zodat een beter optie gekozen kan worden. Dit proces herhaalt zich tot de gekozen optie eenaanvaardbaar resultaat levert.

Hoofdstuk 7
Algemeen besluit
In dit afstudeerwerk werd de sedimentatie van bloedcellen in centrifuges zowel experimen-
teel als numeriek onderzocht. Deze centrifuges scheiden het bloed in zijn verschillende
bestanddelen: plasma, witte- en rode bloedcellen en de bloedplaatjes. Hoofdstuk een be-
licht de anatomie en de functie van het bloed gevolgd door het nut van de bloedproducten
en een beschrijving van de verschillende types centrifuges. Belangrijke artikels en enkele
uitgevoerde simulaties worden in hoofdstuk 2 besproken en bekritiseerd. Het experimen-
teel onderzoek van de sedimentatie van de rode bloedcellen in runderbloed en humaanbloed
staan respectievelijk in hoofdstuk 3 en hoofdstuk 4 beschreven. Het volgende hoofdstuk
vergelijkt de bekomen resultaten zodat er een verband kan worden opgesteld tussen het
resultaat van beide bloedsoorten. In hoofdstuk 6 wordt een numeriek model opgebouwd
en scherp gecontroleerd op zijn accuraatheid.
1 Experimenteel onderzoek met runderbloed
De resultaten van dit onderzoek zijn goed te verklaren met de theorie. Ondanks de aan-
wezigheid van zekere fouten in het proces is het verklaren van de trends toch mogelijk. De
resultaten worden vergeleken met die van humaanbloed en worden gebruikt als controle
van het numeriek model.
2 Experimenteel onderzoek met humaanbloed
In een eerst deel werd in vivo data beschreven afkomstig van het Universitair Ziekenhuis
Gent. Omdat het inlaathematocriet te rudimentair gemeten werd kon deze data geen
100

Hoofdstuk 7. Algemeen besluit 101
zinnige betekenis leveren. Daarom was het beter om zelf een experiment uit te voeren met
humaanbloed. De resultaten van dit experiment vallen goed te verklaren met de theorie en
worden eveneens gebruikt voor de controle van het numeriek model en voor de vergelijking
met runderbloed.
3 Verband tussen humaan en runderbloed
Nadat van runderbloed en humaanbloed een experiment werd uitgevoerd werd een onder-
zoek gestart naar het verband tussen deze resultaten. De resultaten werden geextrapoleerd
naar andere vulsnelheden en inlaathematocrieten zodat het uitlaathematocriet voorspeld
kan worden in functie van de inlaatcondities. Tenslotte is het mogelijk de uitlaathema-
tocrieten van runderbloed te transformeren naar uitlaathematocrieten van humaanbloed.
Op die manier kunnen resultaten verkregen met makkelijk verkrijgbaar runderbloed om-
gerekend worden naar resultaten met humaanbloed.
4 Numeriek model
Als laatste hoofdstuk werd een numeriek model beschreven dat de sedimentatie van bloed-
cellen simuleert. De resultaten komen slechts voor enkele simulaties overeen met het ex-
periment. Het model is dus niet in staat om de sedimentatie correct te beschrijven. De
oorzaak ligt in de hoge complexiteit van het agglomeratiegedrag van bloed. Door verder
onderzoek uit te voeren naar bepaalde parameters van het nummeriek model kan het model
verbeterd worden.

Bibliografie
[1] Van Wie B.J. en Sofer S.S., Sedimentation theory and practical considerations for
the design of centrifugal blood cell processes, The International Journal of Artificial
Organs, 7, 215-222, 1984.
[2] Vand V.J., Viscosity of solutions and suspensions, I. Theory. The Journal of Physical
and Colloid Chemistry 52, 277-299, 1948.
[3] Hawskley, P.G., Some Aspects of Fluid Flow, Paper No. 7. Arnold, London, 1951
[4] Sartory W.K., Centrifugal partical elutriator and method of use, US patent No.3, 825,
175, 1974.
[5] Van Wie B.J., Hustvedt E.L., Particle interaction effects on blood cell sedimentation
and seperations, Biorheology, 25, 651-662,1988.
[6] Patwardhan V.J., Tien C., Sedimentation and liquid fluidization of solid particles of
different sizes and densities, Chemical Engeneering Science 40, 1051-1060, 1985.
[7] Masiliyah, J.H., Hindered settling in a mulit-species particle system, Chemical Enge-
neering Science 34, 1166-1169, 1979.
[8] Barnea, E en Mizrahi J., Generalized approach to the fluid dynamics of particulate
systems part I. General correlation of fluidization and sedimentation, The Chemical
Engeneering Journal 5, 171-189, 1973.
[9] Einstein, H. A. . The bed-load function for sediment transportation in open channel
flows Technical Bulletin 1026, 1950
[10] Dhiel A, Optimisation of a blood seperation process based on simulation, International
Conference on Simulation - Innovation Through Simulation ,19-26, York, 1998.
[11] De Gruttola S., Boomsma K. en Poulikakos D., Computational simulation of a non-
Newtonian Model of the blood seperation process, Artificial Organs 29, 949-959, 2005.
102

BIBLIOGRAFIE 103
[12] Longest P.W., Kleinstreuer C., Efficient computation of micro-particle dynamics in-
cluding wall effects, Comput Fluids 33, 577-601, 2004.
[13] Basset A.B., On motion of sphere in viscous liquid, Philos Trans R Soc Lond 179,
43-63,1988
[14] Vojir D.J., Michaelides E.E., Effect of the history term on the motion of rigid spheres
in a viscous fluid, Multhiphas Flow 20,253-64, 1974.
[15] Sartory W.K., Prediction of concentration profiles during erythrocytes sedimentation
by a hindered settling model. Biorheology 11, 253-64, 1974.
[16] Brown R., The physics of continuous-flow centrifugal cellseparation, Artificial Organs
13, 4- 12, 1989
[17] http://www.fluent.com/software/gambit/index.htm (2006), Algemene website voor
Gambit van het bedrijf Fluent inc.
[18] http://www.fluent.com/software/fluent/index.htm (2006), Algemene website voor
Fluent van het bedrijf Fluent inc.
[19] S. Ogawa, A. Umemura en N. Oshima. On the Equation of Fully Fluidized Granular
Materials, J. Appl. Math. Phys. 31, 483, 1980.
[20] C. K. K. Lun, S. B. Savage, D. J. Jeffrey, and N. Chepurniy. Kinetic Theories for
Granular Flow: Inelastic Particles in Couette Flow and Slightly Inelastic Particles in
a General Flow Field., J. Fluid Mech. 140,223-256, 1984
[21] J. L. Lebowitz. Exact Solution of Generalized Percus-Yevick Equation for a Mixture
of Hard Spheres, The Phy. Rev. 133(4A), A895-A899, 1964.
[22] J. Ding and D. Gidaspow. A Bubbling Fluidization Model Using Kinetic Theory of
Granular Flow, AIChE J. 36(4), 523-538, 1990.
[23] Hadjira Ibdir and Hamid Arastoopour. Modeling of multi-type particle flow using ki-
netic approach, AICHE Journal, May 2005.
[24] M. Syamlal, W. Rogers, and O’Brien T. J. MFIX Documentation: Volume 1, Theory
Guide, National Technical Information Service, Springfield, VA, 1993.
[25] D. G. Schaeffer. Instability in the Evolution Equations Describing Incompressible Gra-
nular Flow J. Diff. Eq., 66:19-50, 1987.


















