
Evoluzione molecolare -...
79
Evoluzione molecolare
Transcript of Evoluzione molecolare -...

Evoluzione molecolare

Beta -lattamasi
Selezione della resistenza agli antibiotici

Charles Darwin, 1859
”On the origin of species by means of natural selection”
1) Forme di vita diverse originano da antenati comuni per discendenza con modificazioni
2) La selezione naturale è il motore dell'evoluzione
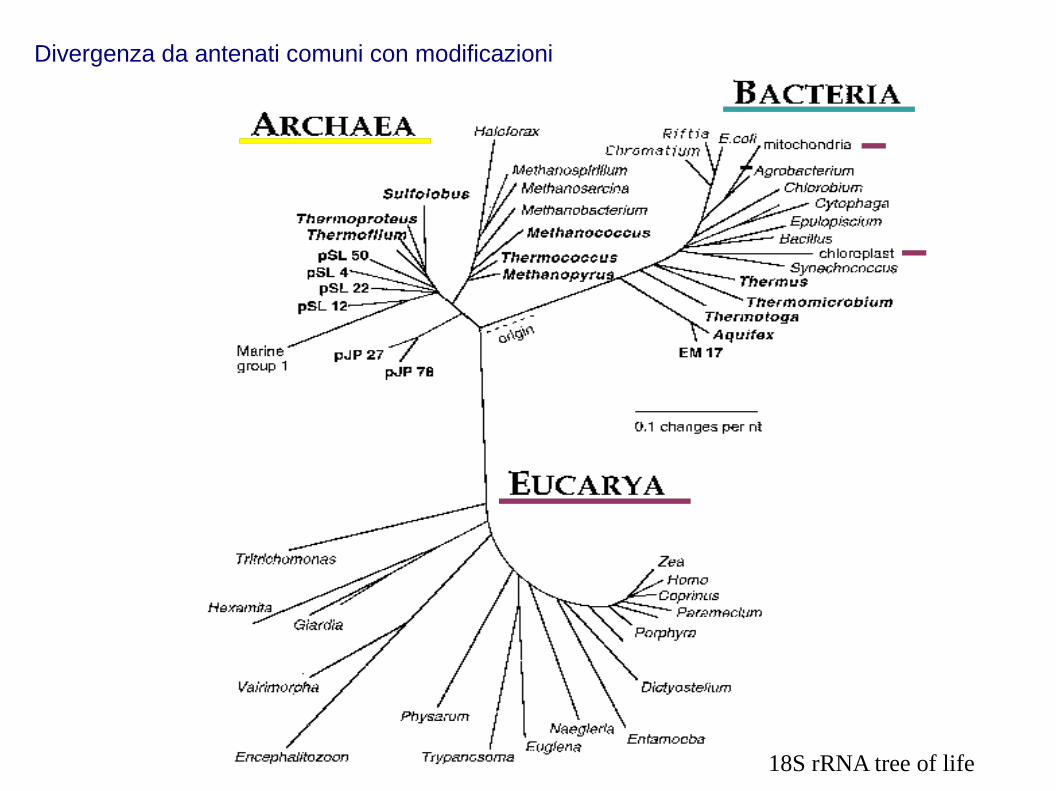
18S rRNA tree of life
Divergenza da antenati comuni con modificazioni

Science 2006
Pnas 2006
“Descent with modifications is only one of the evolutionary mechanisms”
Ricostruzione “automatica” dell’albero della vita...
Visione critica dell’albero della vita.

Sintesi moderna della teoria evolutiva
Unione di idee e osservazioni di diverse discipline scientifiche: genetica, genetica di popolazioni, paleontologia, biochimica, matematica, statistica.
Le mutazioni costituiscono la sorgente di variazione. Possono essere sfavorevoli, favorevoli, o neutrali.
●Le variazioni di frequenze di alleli in una popolazione sono determinanti per l'accettazione delle mutazioni. La deriva genetica e la selezione naturale, contribuiscono alla variazione delle frequenze alleliche.
●La ricombinazione genetica ha un ruolo fondamentale per l'evoluzione dei geni.
>1937, Fisher, J. Huxley, Haldane, Simpson, Dobzhansky
“Nulla in biologia ha senso se non alla luce dell'evoluzione”T. Dobzhanski
I fenomeni evolutivi sono spiegati attraverso meccanismi biologici conosciuti:
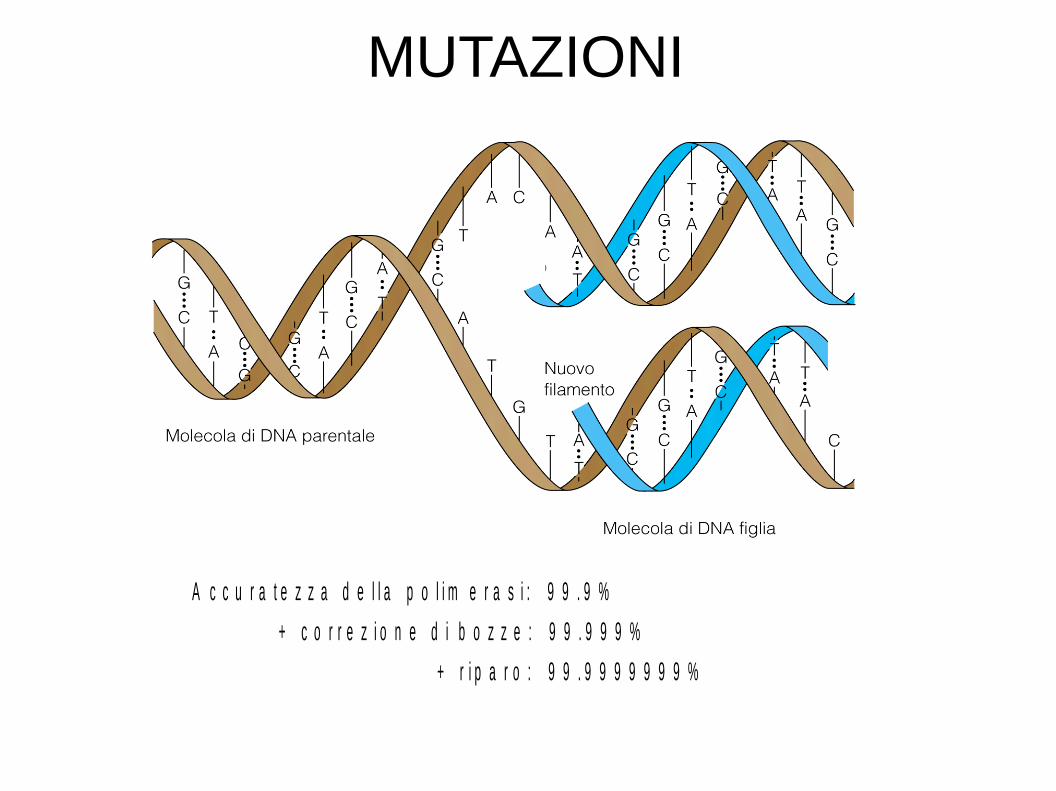
A c c u r a t e z z a d e l l a p o l i m e r a s i : 9 9 . 9 %
+ c o r r e z i o n e d i b o z z e : 9 9 . 9 9 9 %
+ r i p a r o : 9 9 . 9 9 9 9 9 9 9 %
MUTAZIONI

Mutazioni: alterazioni dell'informazione codificata nel DNA
Sostituzioni: cambiamenti basi
-Transizioni: cambiamenti Purina/Purina o Pirimidina/Pirimidina
-Transversioni: cambiamenti Purina/Pirimidina o viceversa
Inserzioni: aggiunte di nucleotidi
Delezioni: rimozioni di nucleotidi

Mutazioni in sequenze codificanti
● Sostituzioni
♦ sinonime: non modificano l'amino acido♦ di senso: cambiano un amino acido in uno diverso♦ non-senso: cambiano un amino acido in un codone di
stop
● Inserzioni/Delezioni
♦ Con cornice di lettura mantenuta (multipli di tre)
♦ Frameshift

Bias mutazionale (esempio)
G:U
G:T
Duplex
G:C
G:C or A:T
Negli organismi in cui si ha metiliazione delle citosine le mutazioni G:C → A:T sono più probabili delle mutazioni A:T → G:C
riparo
riparo
Duplex

Comparsa e esito delle mutazioni
comparsa della mutazione(caso)
accettazione della mutazione(caso, selezione naturale?)
• mutazione neutrale• mutazione svantaggiosa • mutazione vantaggiosa
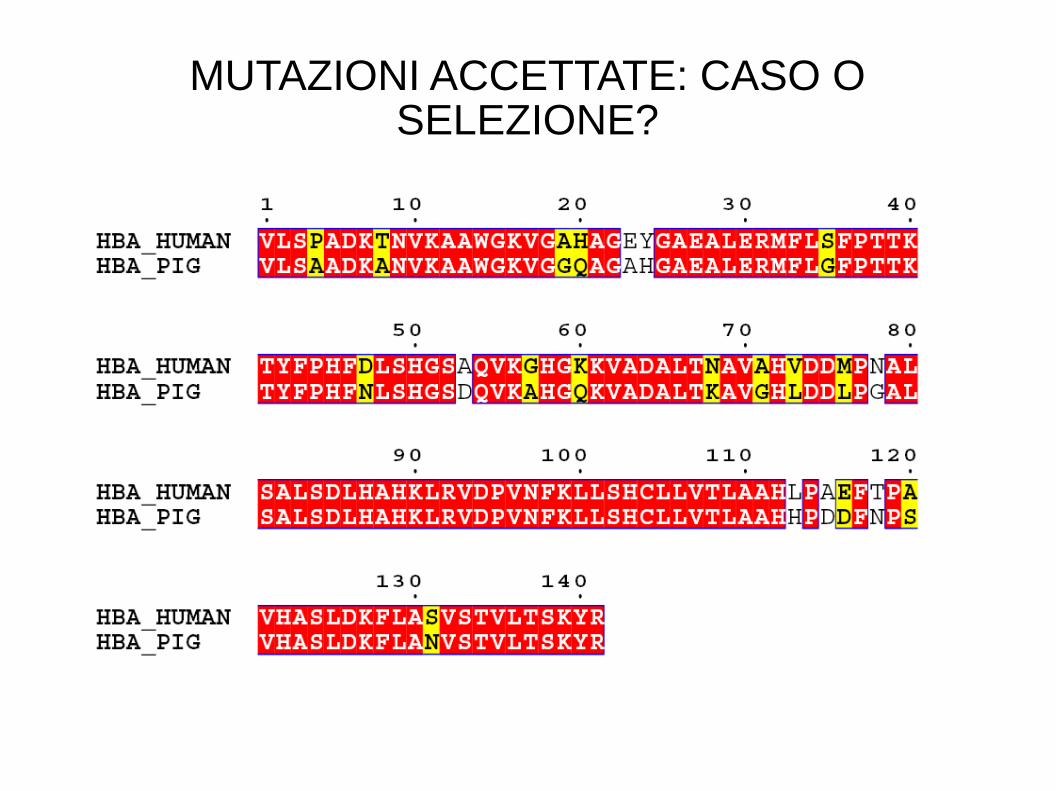
MUTAZIONI ACCETTATE: CASO O SELEZIONE?

La maggior parte delle mutazioni osservate sono neutrali
Kimura, M. Evolutionary rate at the molecular level, Nature (1968)

Teoria neutrale dell'evoluzione molecolare
Kimura, M. Evolutionary rate at the molecular level, Nature (1968)Kimura, M. The neutral theory of molecular evolution (1983)
Le mutazioni neutrali (o debolmente svantaggiose) possono essere accettate per effetto del caso
Le mutazioni vantaggiose sono favorite ma possono non essere accettate per effetto del caso
La comparsa delle mutazioni neutrali (o svantaggiose) è molto più probabile della comparsa delle mutazioni vantaggiose
Il caso (oltre alla selezione naturale) ha grande importanza nell'accettazione delle mutazioni.

Predizioni della teoria neutrale
• Regioni “più importanti” mutano meno rapidamente (o sono immutabili)
• residui critici per la struttura• residui critici per la catalisi
• Regioni “meno importanti” mutano più rapidamente
• introni• pseudogeni• terza posizione dei codoni• regioni “periferiche” delle proteine

Accumulo di mutazioni nelle diverse regioni dei geni codificanti per proteine

Genetic drift (deriva genetica)A
llele
freq
uen
cy
Alle
le fr
equ
enc
y
Generations Generations
Small population N = 20
Large populationN = 2000
:= Fissazione o scomparsa di una mutazione attraverso fluttuazioni casuali delle frequenze alleliche
p = 1 / 2N
la probabilità (p) di fissazione è inversamente proporzionale alla dimensione della popolazione (2N in una popolazione diploide)
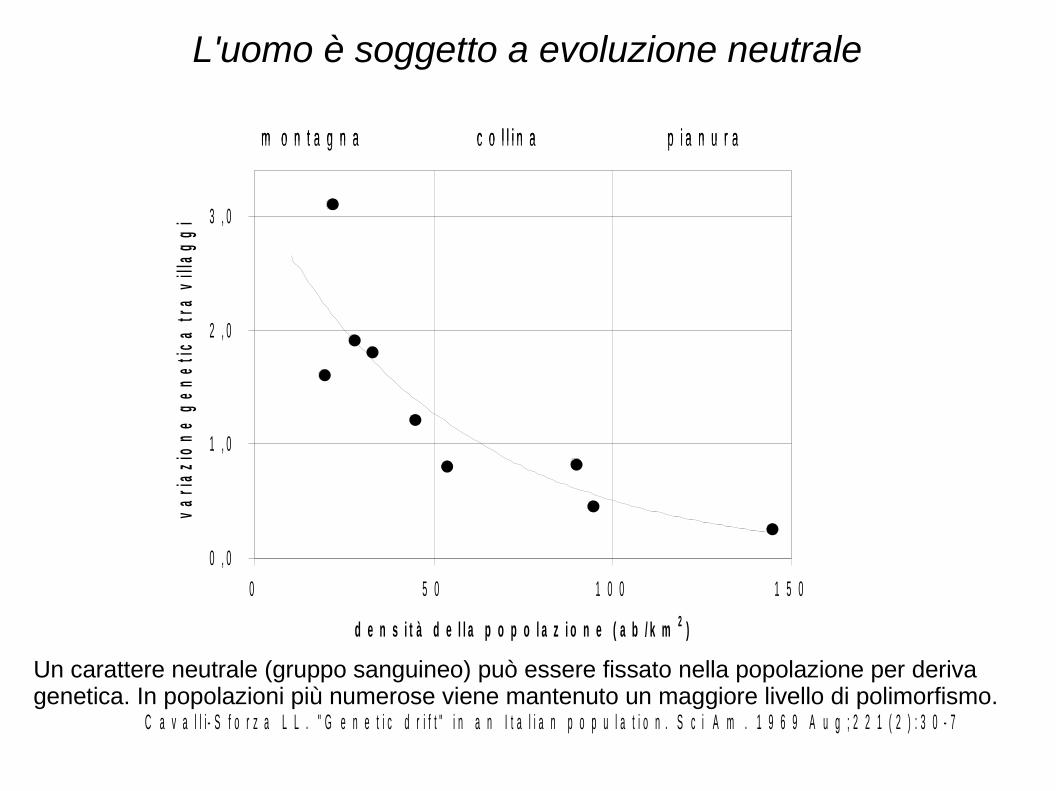
L'uomo è soggetto a evoluzione neutrale
0 , 0
1 , 0
2 , 0
3 , 0
0 5 0 1 0 0 1 5 0
d e n s i t à d e l l a p o p o l a z i o n e ( a b / k m 2 )
va
ria
zio
ne
ge
ne
tic
a t
ra v
illa
gg
i
m o n t a g n a c o l l i n a p i a n u r a
0 , 0
1 , 0
2 , 0
3 , 0
0 5 0 1 0 0 1 5 0
d e n s i t à d e l l a p o p o l a z i o n e ( a b / k m 2 )
va
ria
zio
ne
ge
ne
tic
a t
ra v
illa
gg
i
m o n t a g n a c o l l i n a p i a n u r a
C a v a l l i - S f o r z a L L . " G e n e t i c d r i f t " i n a n I t a l i a n p o p u l a t i o n . S c i A m . 1 9 6 9 A u g ; 2 2 1 ( 2 ) : 3 0 - 7
Un carattere neutrale (gruppo sanguineo) può essere fissato nella popolazione per deriva genetica. In popolazioni più numerose viene mantenuto un maggiore livello di polimorfismo.
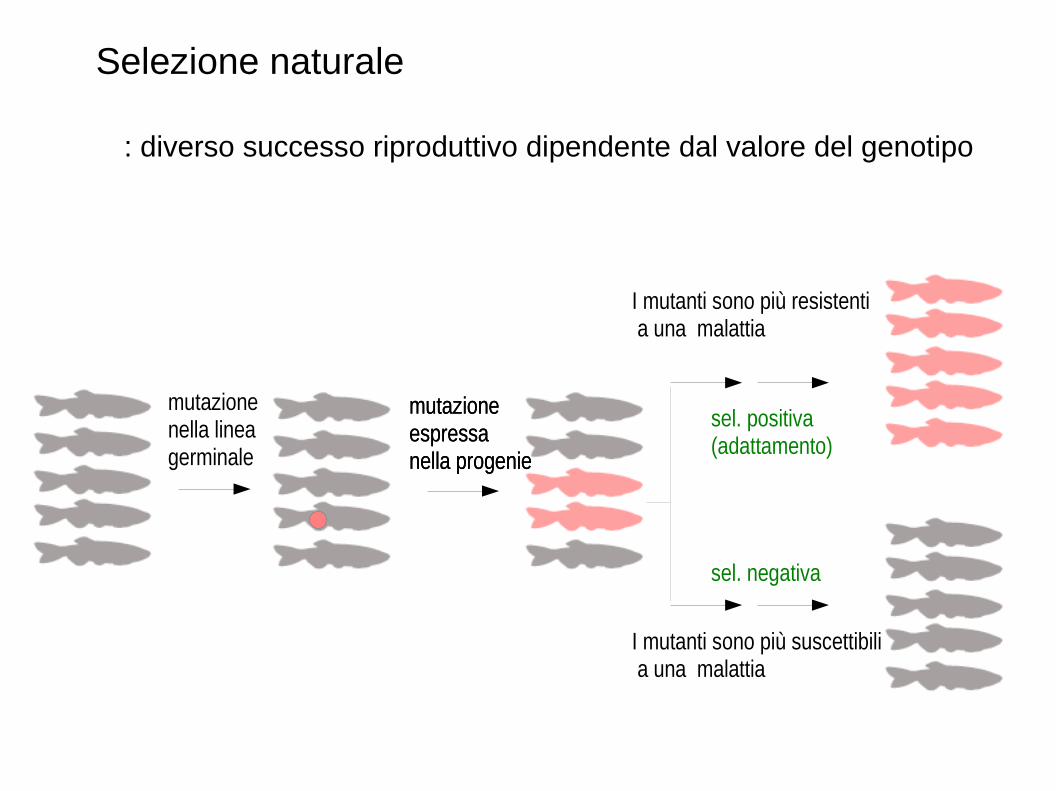
Selezione naturale
: diverso successo riproduttivo dipendente dal valore del genotipo
I mutanti sono più resistenti a una malattia
mutazioneespressa nella progenie
mutazioneespressa nella progenie
mutazionenella linea germinale
I mutanti sono più suscettibili a una malattia
sel. positiva (adattamento)
sel. negativa
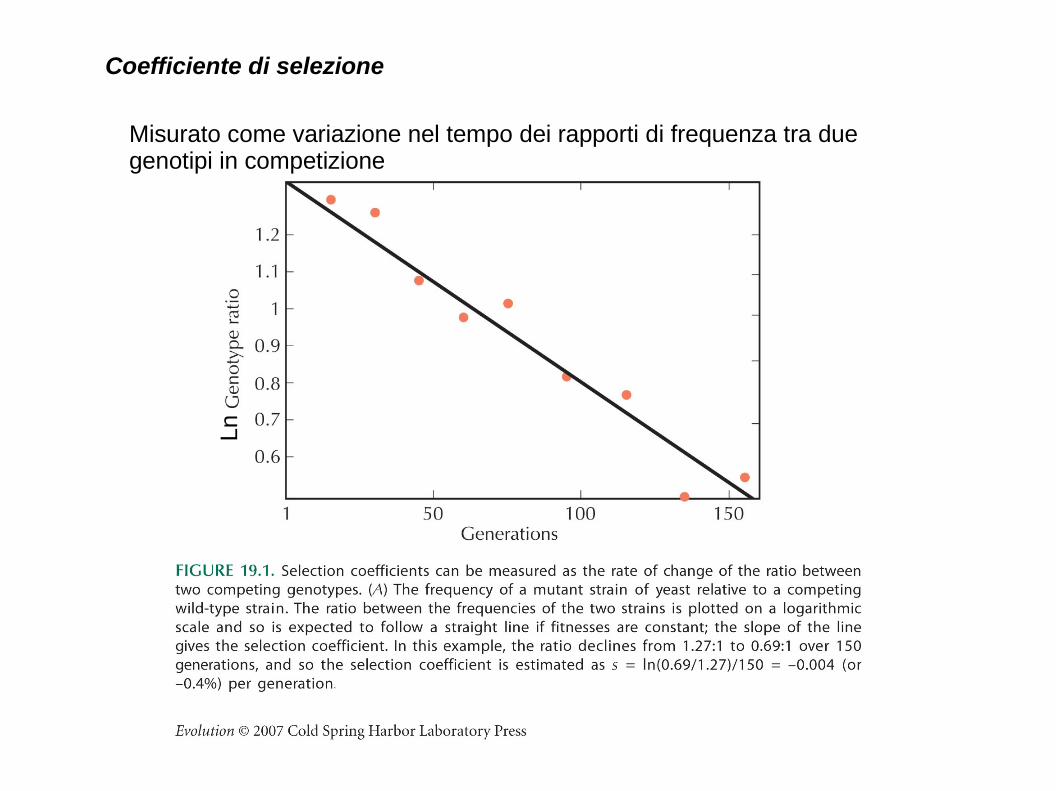
Coefficiente di selezione
Misurato come variazione nel tempo dei rapporti di frequenza tra due genotipi in competizione
Ln
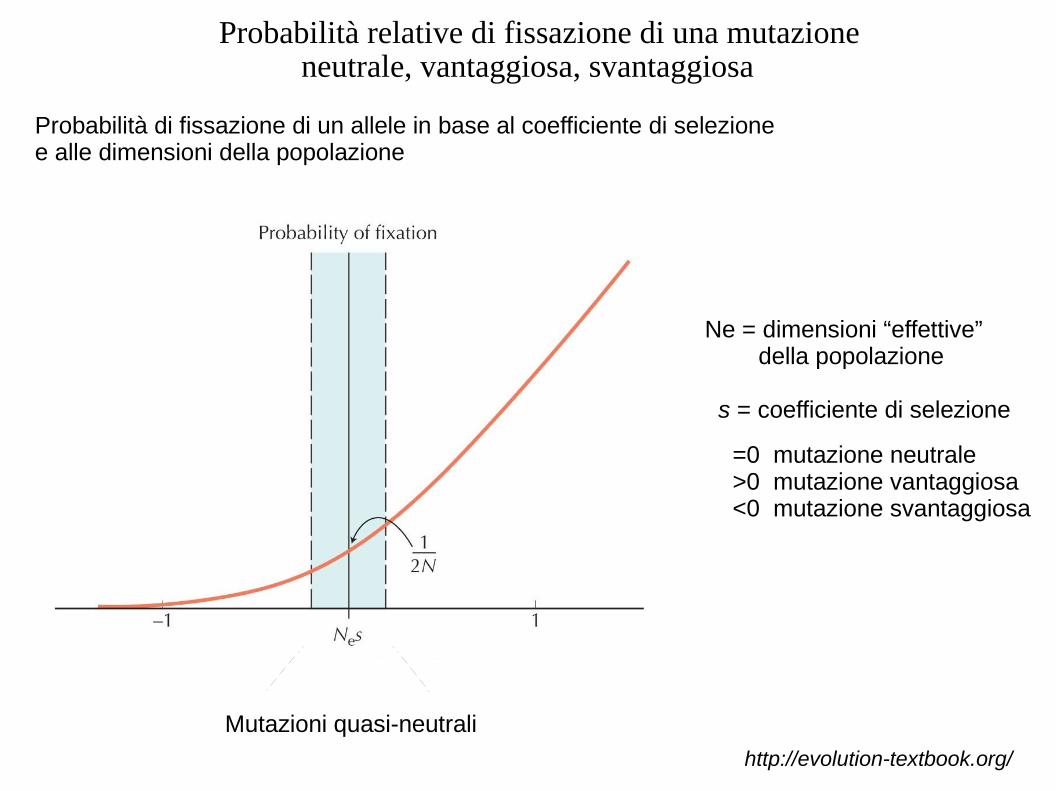
Probabilità relative di fissazione di una mutazione neutrale, vantaggiosa, svantaggiosa
Ne = dimensioni “effettive” della popolazione
s = coefficiente di selezione
=0 mutazione neutrale>0 mutazione vantaggiosa<0 mutazione svantaggiosa
Probabilità di fissazione di un allele in base al coefficiente di selezione e alle dimensioni della popolazione
Mutazioni quasi-neutrali
http://evolution-textbook.org/

L'importanza della ricombinazione
A) Singolo gruppo di linkage B) Tre gruppi distinti
vantaggiosa
deb. vantaggiosa
svantaggiosa
neutrale
Effetto del linkage (assenza di ricombinazione) sul destino di alleli alternativi. A) i tre loci sotto selezione sono completamente legati. Si assume che il vantaggio complessivo dell'allele azzurro (vantaggioso) e dell'allele nero (svantaggioso) sia maggiore dell'allele verde (deb. vantaggioso). Le mutazioni vantaggiose, svantaggiose e neutrali sono fissate inseme (effetto hitchhiking). La mutazione vantaggiosa in verde è eliminata. B) i tre loci sono indipendenti. Entrambe le mutazioni vantaggiose sono fissate, la mutazione svantaggiosa è eliminata. Tratta da Lynch, M. The origin of genome architecture.

Bannasch et al. Mutations in the SLC2A9 Gene Cause Hyperuricosuria and Hyperuricemia in the Dog. PLoS Genet. 2008
Kidney Liver
Cys188Phe
Mutazione in GLUT9 causa iperuricemia nel cane DalmataGLUT9 è in linkage con un gene che determina la grandezza delle macchie del manto.
Selezione per linkage di mutazioni patologiche nelle razze canine

Dimensioni del genoma e complessità degli organismi

C-value paradox
Per ragioni storiche il contenuto di DNA nel genoma di un organismo è riferito come C-value, ovvero come constant-value, dall'osservazione che il peso in pg del DNA è costante negli individui di una stessa specie.
1 Gbp ≈ 1 pg.
(109) * (660) * (1,66 · 10-24) = 1,02 · 10-12 g
Il C-value del DNA diploide umano (2C) è c.a 6.3 pg nei maschi e 6.4 pg nelle femmine
Il “C-value paradox” (Thomas 1971), deriva dall'osservazione che
1) il contenuto di DNA aploide può avere grandi variazioni anche nello stesso gruppo di organismi (es. nelle angiosperme varia tra 0.1 pg e 125 pg)
2) il contenuto di DNA aploide può non avere una relazione diretta con la “complessità” di un organismo (es alcuni protisti hanno C-values 100 volte superiori a quello dell'uomo).
Thomas, C.A. Jr. (1971) The genetic organization of chromosomes. Annu. Rev. Genet.

Mutation load e Mutation rate
Il mutational load (U) è il tasso di mutazioni deleterie per genoma per generazione.
Nell'uomo si stima U ≈ 2
Il mutation rate () è il numero di mutazioni totali per sito per generazione.
Nell'uomo si calcola = 1,1 · 10-8
Se tutte queste mutazioni fossero deleterie, il mutational load umano sarebbe
U = (1,1 · 10-8 )* (3 ·109 bp) * (2) = 66
Ovvero ciacuno riceve dai propri genitori circa 66 mutazioni, ma solo una piccola percentuale di queste (<5%) sono deleterie.
Analysis of genetic inheritance in a family quartet by whole-genome sequencing. Roach J. C.. et al., Science 2010
A resolution of the mutation load paradox in humans. Lesecque Y, Keightley PD, Eyre-
Walker A. Genetics. 2012

Negli anni '70, per spiegare il paradosso del C-value e l'incompaibilità tra mutation rate and mutation load, venne proposto che vari genomi fossereo costituiti in grande parte da DNA privo di una funzione. Susumu Ohno chiamò questo DNA “Junk DNA”.
Junk: Discarded material, such as glass, paper, or metal, some of which may be reused in some form.
Ohno, S. (1972) So much “junk” DNA in our genome. Brookhaven Symp. Biol. 23, 366–370

The ENCODE Project: ENCyclopedia Of DNA Elements
The human genome encodes the blueprint of life, but the function of the vast majority of its nearly three billion bases is unknown. The Encyclopedia of DNA Elements (ENCODE) project has systematically mapped regions of transcription, transcription factor association, chromatin structure and histone modification. These data enabled us to assign biochemical functions for 80% of the genome, in particular outside of the well-studied protein-coding regions.
Encode criticism
The C-value paradox, junk DNA and ENCODE. Eddy SR. Curr Biol. 2012
On the immortality of television sets: "function" in the human genome according to the evolution-free gospel of ENCODE. Graur D, et al. Genome Biol Evol. 2013
Is junk DNA bunk? A critique of ENCODE. Doolittle WF. Proc Natl Acad Sci U S A. 2013

Neutral evolution of genome complexity
● La selezione naturale domina nelle popolazioni grandi
● La deriva genetica domina nelle popolazioni piccole
● La complessità genomica si origina passivamente per azione della deriva genetica
● La complessità genomica è occasione di innovazioni adattative

OMOLOGIA
Emoglobina alfa uomo - Emoglobina alfa maiale

ATCGGCCACTTTCGCGATCA
Sequenza ancestrale
ATAGGCCACTTTCGCGATCA
ATAGGCCACTTTCGCGATTA
ATAGGGCACTTTCGCGATTA
ATAGGGCACTTT-GCGATTA
ATAGGGCACTTT-GCGATGA
ATCGGCCACTTTCGCGATCG
ATCGGCCACTTTCGTGATCG
ATCGGCCACGTTCGTGATCG
ATCGCCCACGTTCGCGATCG
ATTGCCCACGTTCGCGATCG
Sequenze omologhe
ATCGGCCACGTTCGCGATCG
Evoluzione nel tempo dell'informazione biologica
Omologia = condivisione di un ancestore comune
Evento di separazione
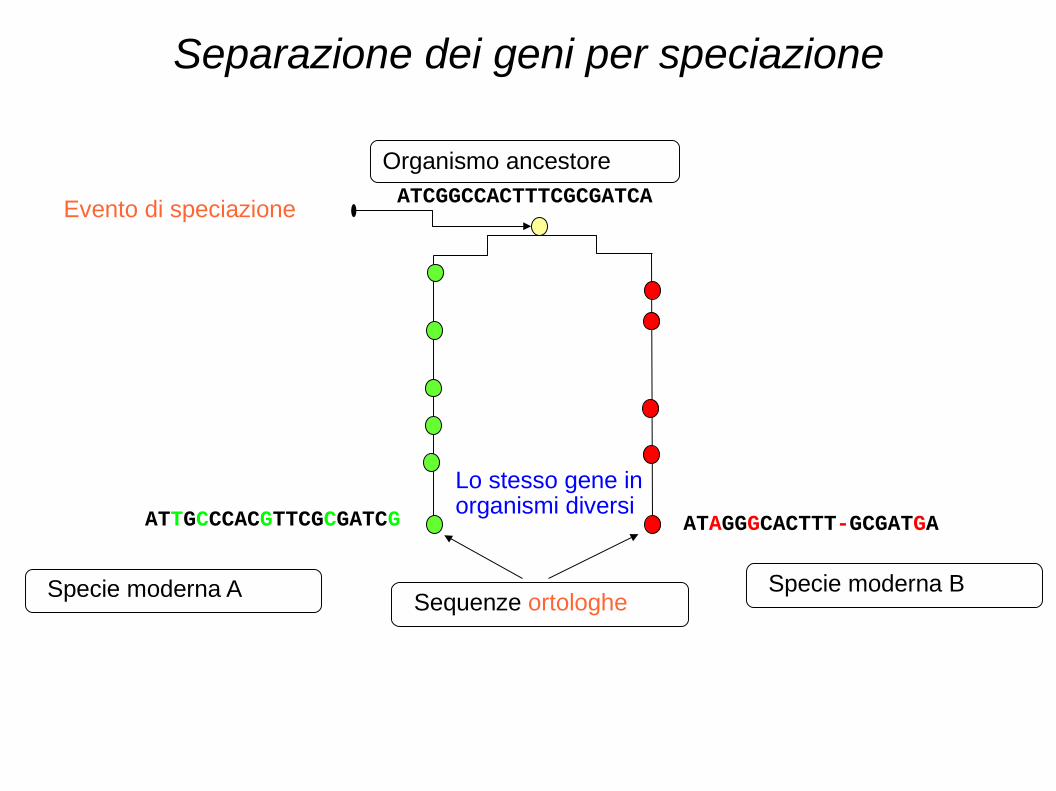
ATCGGCCACTTTCGCGATCA
Organismo ancestore
ATAGGGCACTTT-GCGATGA ATTGCCCACGTTCGCGATCG
Sequenze ortologhe
Separazione dei geni per speciazione
Specie moderna A Specie moderna B
Evento di speciazione
Lo stesso gene in organismi diversi

Separazione per speciazione
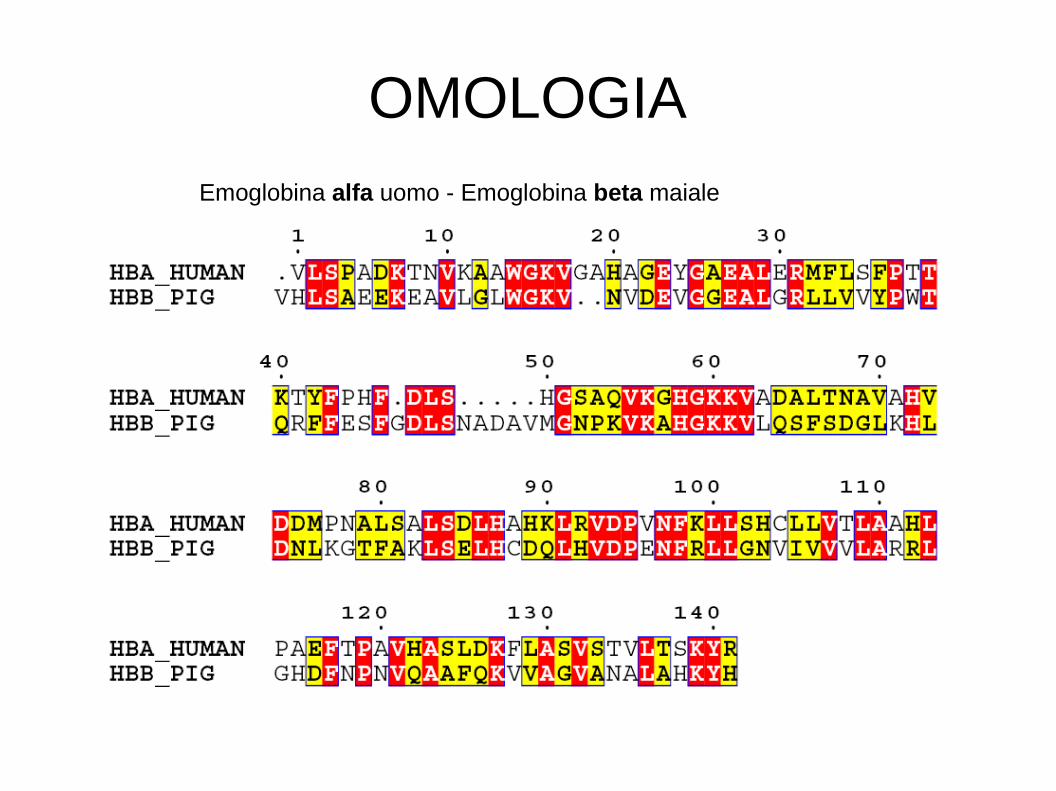
OMOLOGIA
Emoglobina alfa uomo - Emoglobina beta maiale
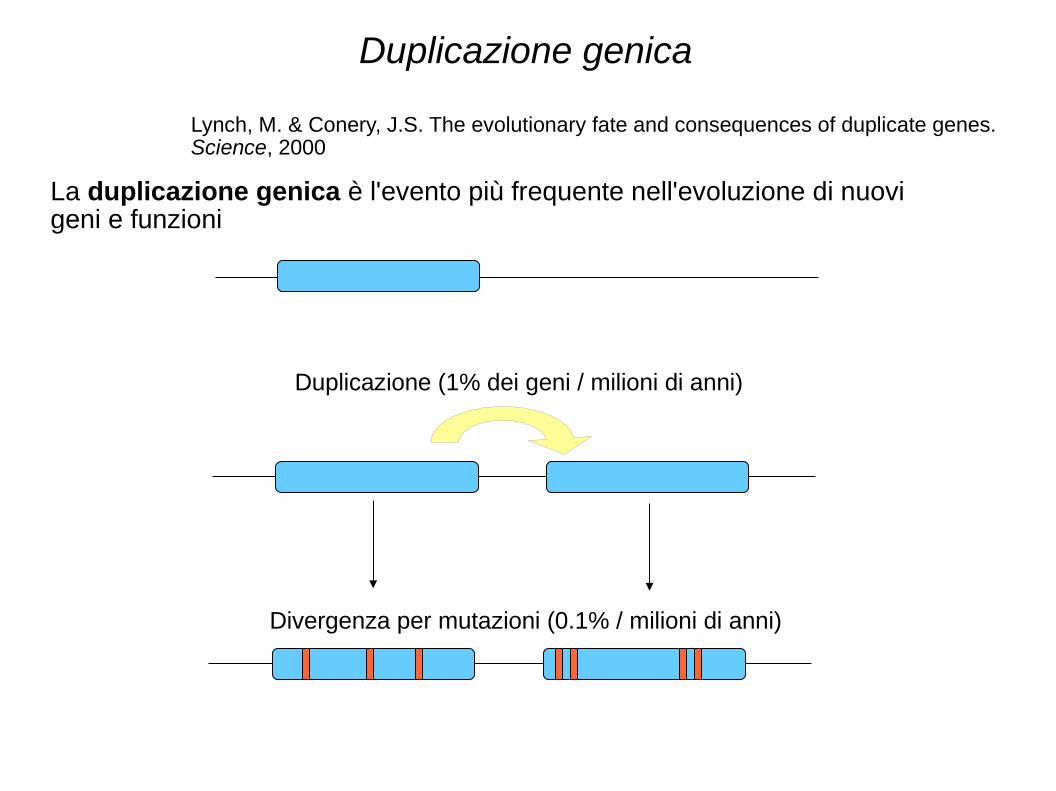
Duplicazione genica
Duplicazione (1% dei geni / milioni di anni)
Divergenza per mutazioni (0.1% / milioni di anni)
La duplicazione genica è l'evento più frequente nell'evoluzione di nuovi geni e funzioni
Lynch, M. & Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science, 2000
Demografia delle popolazioni umane

Demografia dei geni duplicati
Distribuzione dell'età di coppie di geni duplicati in sei genomi eucariotici. Dalla distribuzionesi possono ricavare informazioni sui tassi di nascita e morte dei geni duplicati. Da Lynch and Connery, 2003 “The evolutionary demografy of duplicated genes”
Recent duplications
Oldduplications
Baby-boom

Esiti della duplicazione genica
Silenziamento di una copia (pseudogene)
Divergenza funzionale
X
Mantenimento della funzione nelle due copie
(Vita media di un gene duplicato: 3-8 My)
(Es. Proteine ribosomali, tRNA)
(Funzione simile, oppure drasticamente diversa dalla funzione originaria)
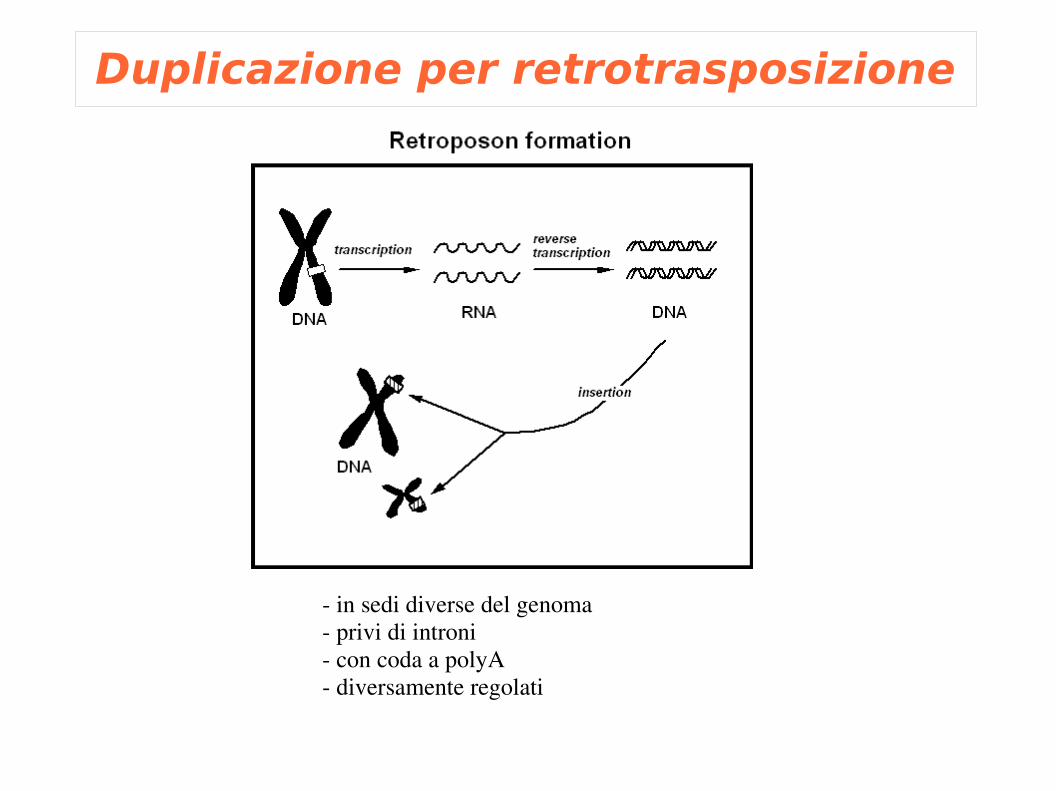
Duplicazione per retrotrasposizione
in sedi diverse del genoma privi di introni con coda a polyA diversamente regolati

Esiti della duplicazione per retroposizione
Silenziamento di una copia (pseudogene processato)
Acquisizione di una nuova funzione
XMantenimento della funzione nelle due copie
(meno probabile)
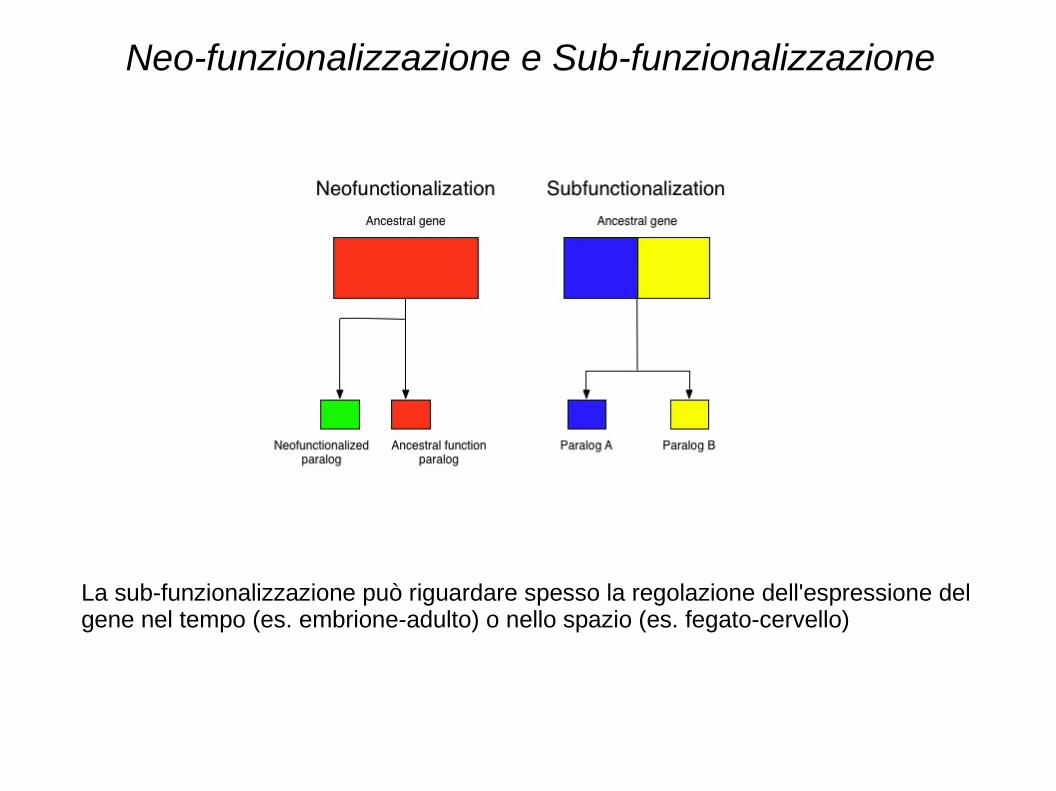
Neo-funzionalizzazione e Sub-funzionalizzazione
La sub-funzionalizzazione può riguardare spesso la regolazione dell'espressione del gene nel tempo (es. embrione-adulto) o nello spazio (es. fegato-cervello)
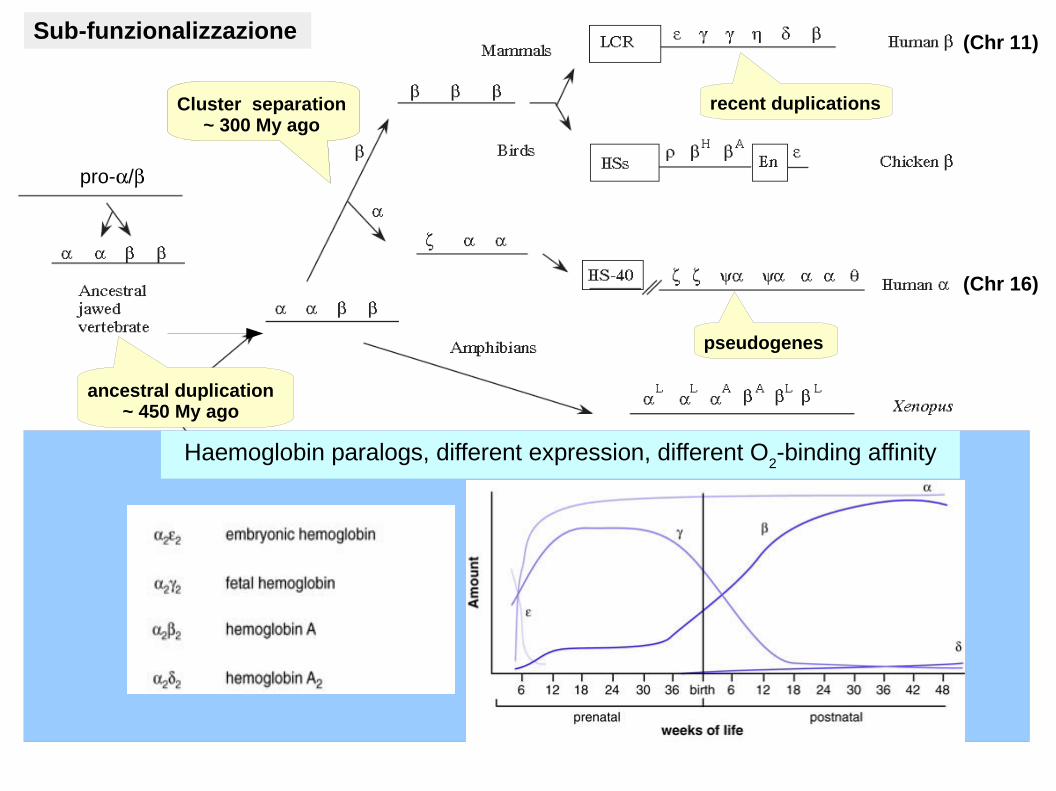
pro-/
ancestral duplication~ 450 My ago
Cluster separation~ 300 My ago
recent duplications
pseudogenes
(Chr 11)
(Chr 16)
Cluster separation~ 300 My ago
Haemoglobin paralogs, different expression, different O2-binding affinity
Sub-funzionalizzazione

H I U a s e - T T R : a n e x t r e m e c a s e o f d i v e r g e n t e v o lu t i o n
D i s t r i b u t i o n : v e r t e b r a t e s
F u n c t i o n : T h y r o x i n e t r a n s p o r t e r“ t r a n s t h y r e t i n ( T T R ) ”
D i s t r i b u t i o n : v e r t e b r a t e s , o t h e r o r g a n i s m s
F u n c t i o n : h y d r o l y s i s o f5 - h y d r o x y i s o u r a t e
Bacteria
Plants
Fungi
Inverteb.
Verteb.
Verteb.
Gene duplication> 500 My ago
Hydroxyisourate hydrolase and transthyretin: extreme functional divergence
Neo-funzionalizzazione

La duplicazione può interessare interi genomi
Duplicazionegenomica
Wolfe, KH, curr. biol
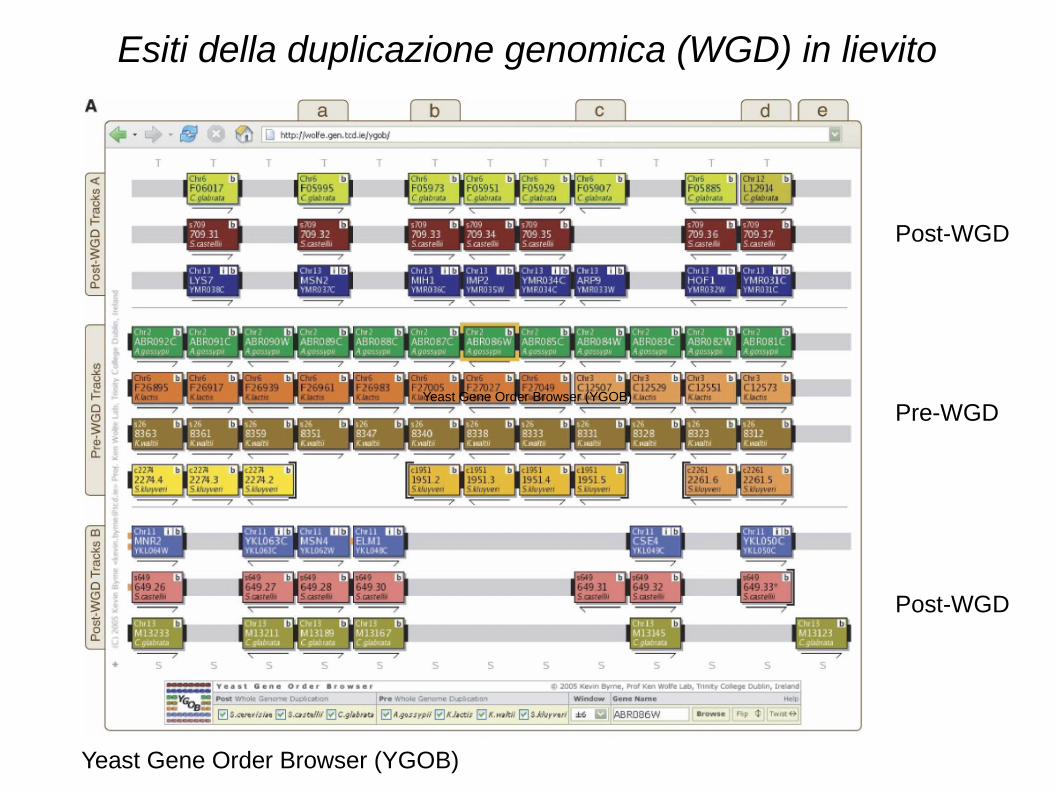
Esiti della duplicazione genomica (WGD) in lievito
Post-WGD
Pre-WGD
Post-WGD
Yeast Gene Order Browser (YGOB)
Yeast Gene Order Browser (YGOB)

Due duplicazioni genomiche nell'evoluzione dei vertebrati(2R hypothesis)
Ohno S (1970). Evolution by Gene Duplication.Dehal, et al (2005). "Two Rounds of Whole Genome Duplication in the Ancestral Vertebrate".
Time
1R
2R
550 Mya
450 Mya
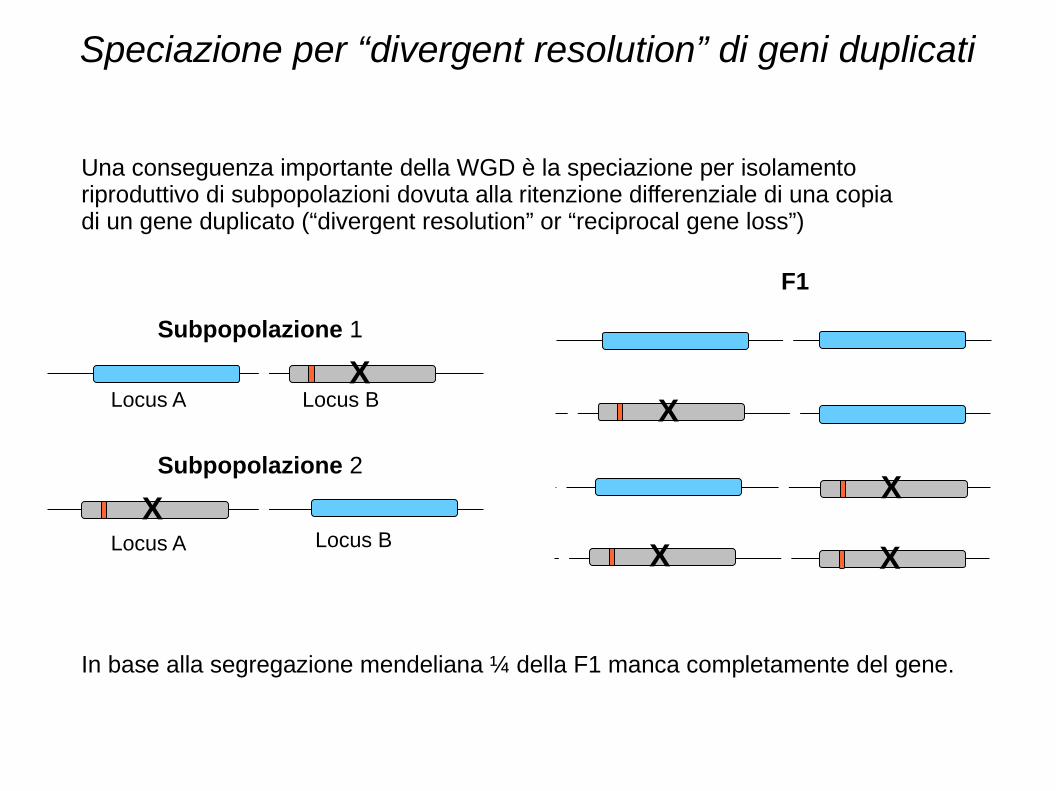
Speciazione per “divergent resolution” di geni duplicati
Una conseguenza importante della WGD è la speciazione per isolamento riproduttivo di subpopolazioni dovuta alla ritenzione differenziale di una copia di un gene duplicato (“divergent resolution” or “reciprocal gene loss”)
X
Subpopolazione 1
X
Subpopolazione 2
F1
Locus A Locus B
Locus A Locus B
X
X X
X
In base alla segregazione mendeliana ¼ della F1 manca completamente del gene.

ATCGGCCACTTTCGCGATCA
gene ancestore
ATAGGGCACTTT-GCGATGA ATTGCCCACGTTCGCGATCG
Sequenze paraloghe
Separazione per duplicazione genica
gene moderno A gene moderno B
Evento di duplicazione
Geni originati per duplicazione in uno stesso genoma

Rapporti evolutivi tra geni
Ortologia: I geni si separano per speciazione. La filogenesi dei geni riflette la storia degli organismi
Paralogia: I geni si separano per duplicazione all’interno di uno stesso organismo. La filogenesi riflette la storia dei geni
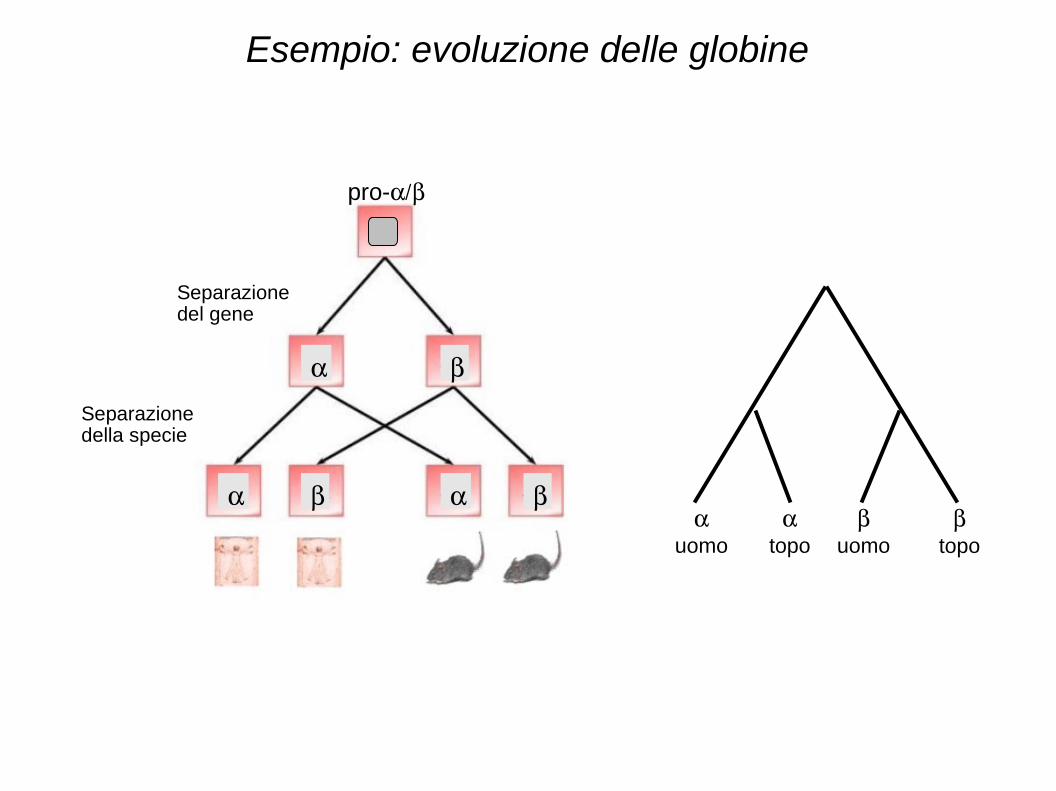
Esempio: evoluzione delle globine
Separazionedel gene
Separazione della specie
pro-
uomo
topo
uomo
topo

Diversi rapporti di parentela tra sequenze
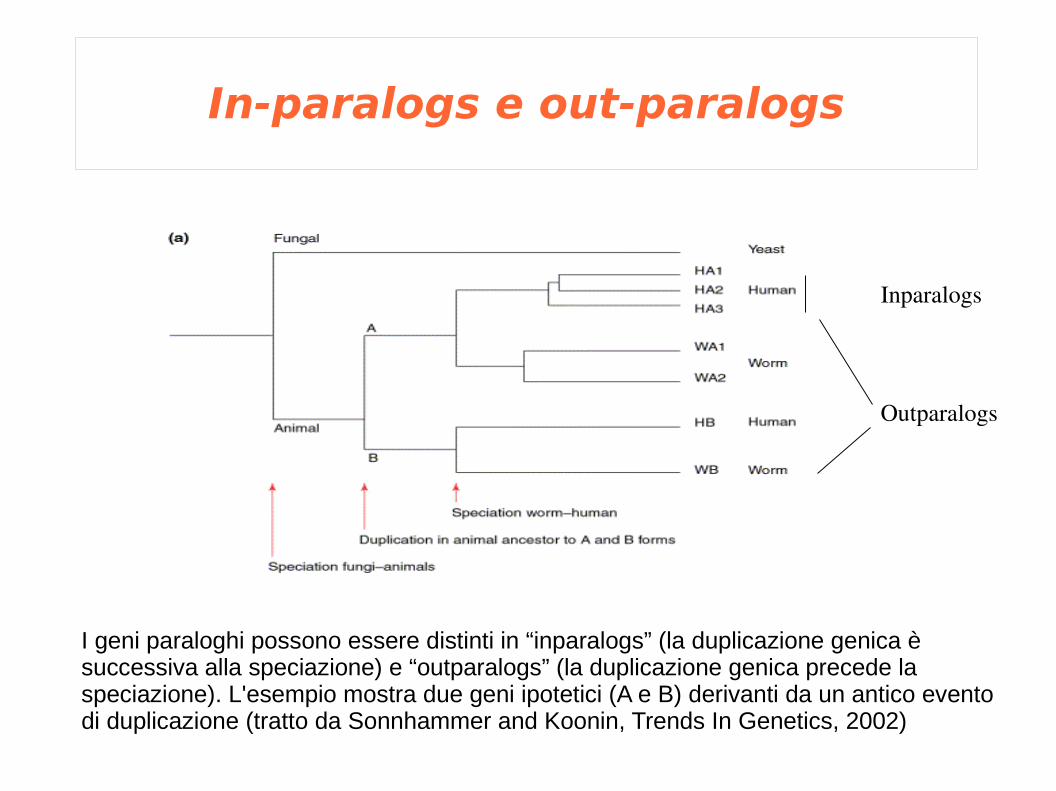
In-paralogs e out-paralogs
Outparalogs
Inparalogs
I geni paraloghi possono essere distinti in “inparalogs” (la duplicazione genica è successiva alla speciazione) e “outparalogs” (la duplicazione genica precede la speciazione). L'esempio mostra due geni ipotetici (A e B) derivanti da un antico evento di duplicazione (tratto da Sonnhammer and Koonin, Trends In Genetics, 2002)

Omologia Identità di funzione

Caratteristiche dei geni omologhi
- Proteine derivanti da geni omologhi hanno struttura 3D simile
- Proteine derivanti da geni paraloghi possono avere una funzione uguale o simile
- Proteine derivanti da geni ortologhi hanno probabilmente una funzione uguale o simile

Metodi per la distinzione di tipi diversi omologia
1) Ricostruzione filogenetica. Confronto tra “gene trees” e “species trees”
2) Best reciprocal hit Ricerca di omologia in genomi completi

Best Reciprocal Hit (BRH)
il gene a della specie x ha come miglior mach il gene b della specie y.
Il gene b della specie y ha come miglior match il gene a nella specie x.
a
b
Best hit
Best hit
a' Best hit
a e b : BRHa' e b : non BRH

Gene trees e Species trees
frog
mouse
human
frog
mouse
human
frog
mouse
human
mouse
frog
human
Species tree
Gene tree(complete set) Gene tree
(uncomplete set)

Xenologia: trasferimento orizzontale di geni
Albero delle specie Albero dei geni
trasferimentoorizzontale
Albero dei geni inconciliabile con l'albero delle specie
- intra-specie e inter-specie (frequente nei procarioti)
- endosinbionte → ospite (eucarioti)
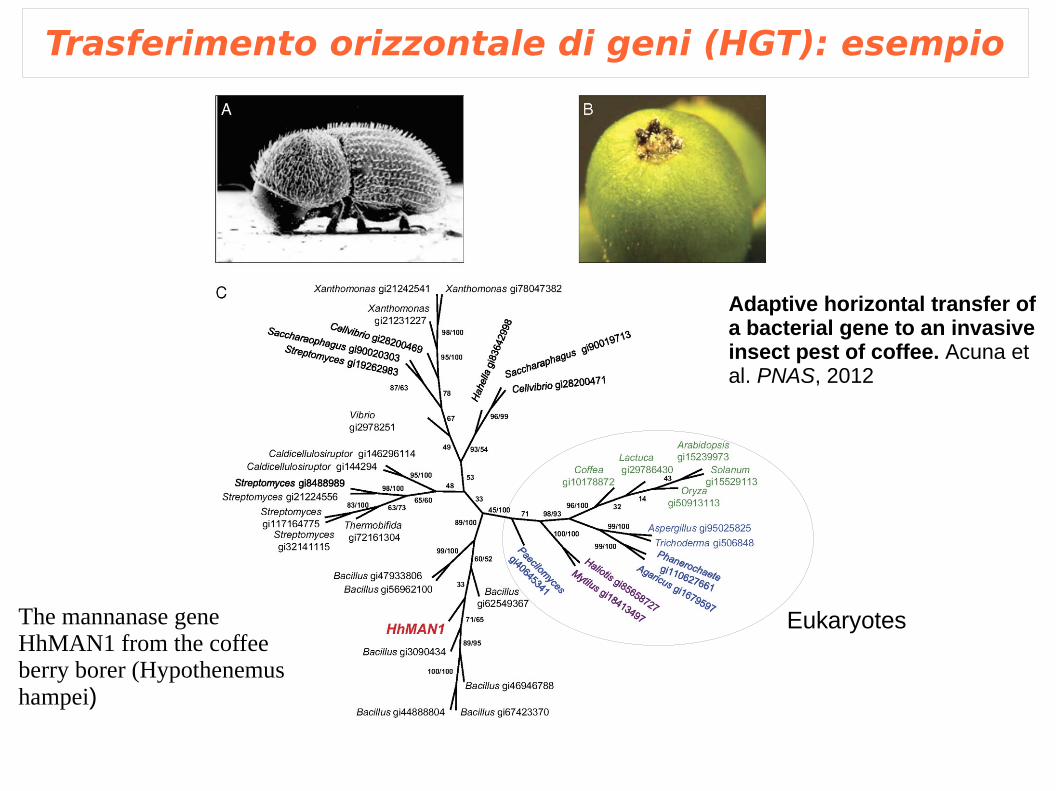
Trasferimento orizzontale di geni (HGT): esempio
The mannanase gene HhMAN1 from the coffee berry borer (Hypothenemus hampei)
Adaptive horizontal transfer of a bacterial gene to an invasive insect pest of coffee. Acuna et al. PNAS, 2012
Eukaryotes

Esiti possibili del trasferimento orizzontale
Donor
recipient
Acquisizione di un nuovo gene
Acquisizione di un analogo
Acquisizione di un paralogo
Mantenimento / silenziamento
Mantenimento / silenziamento/ gene displacement
Mantenimento / silenziamento/ gene displacement
“non orthologous gene displacements”: sostituizione di un gene originario ad opera di un gene trasferito orizzontalmente

Mutazioni come orologio molecolare?
Mutazioni ~ Tempo

Orologio molcolare
Ipotesi di una relazione proporzionale tra tempo evolutivo e numero di mutazioni accumulate
L'orologio è stocastico
L'orologio necessita di una calibrazione attraverso un riferimento indipendente (es. Reperti fossili). Una volta conosciuta la velocità dell'orologio, la divergenza tra sequenze può essere usata per stimare il tempo di separazione

Orologio molecolare nell'emoglobina alfa
Calcolo il numerodi mutazioni...
...Trovo il tempo di separazione
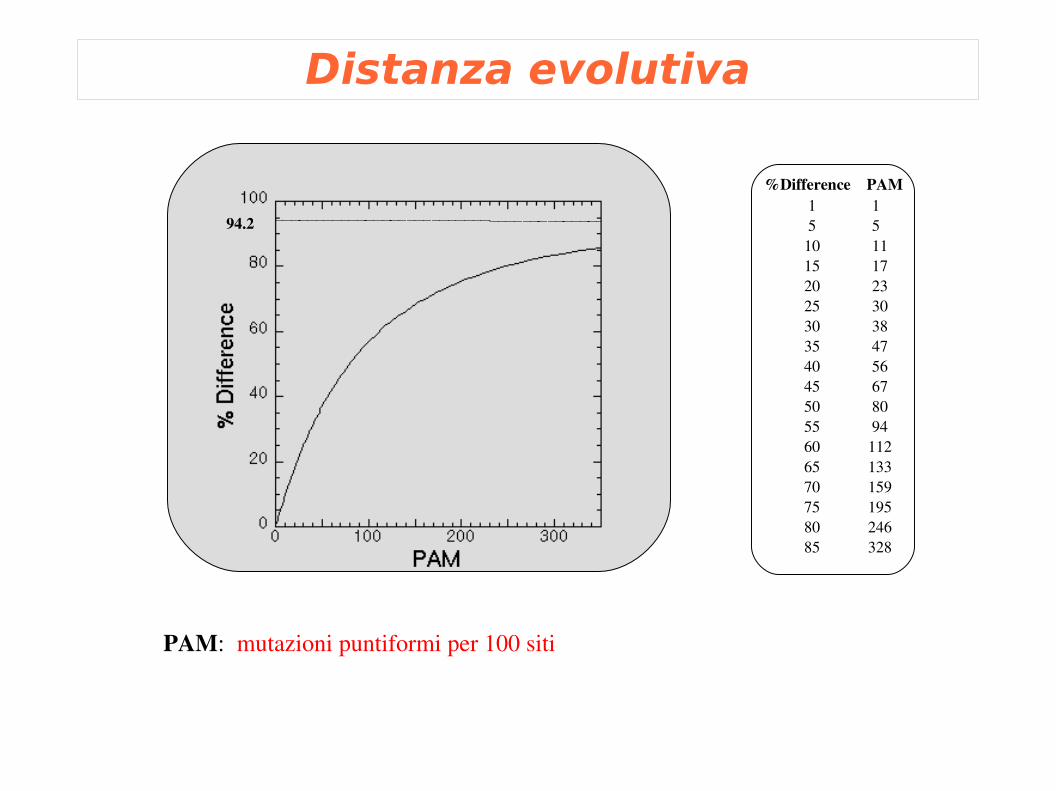
Distanza evolutiva
94.2
%Difference PAM 1 1 5 5 10 11 15 17 20 23 25 30 30 38 35 47 40 56 45 67 50 80 55 94 60 112 65 133 70 159 75 195 80 246 85 328
PAM: mutazioni puntiformi per 100 siti
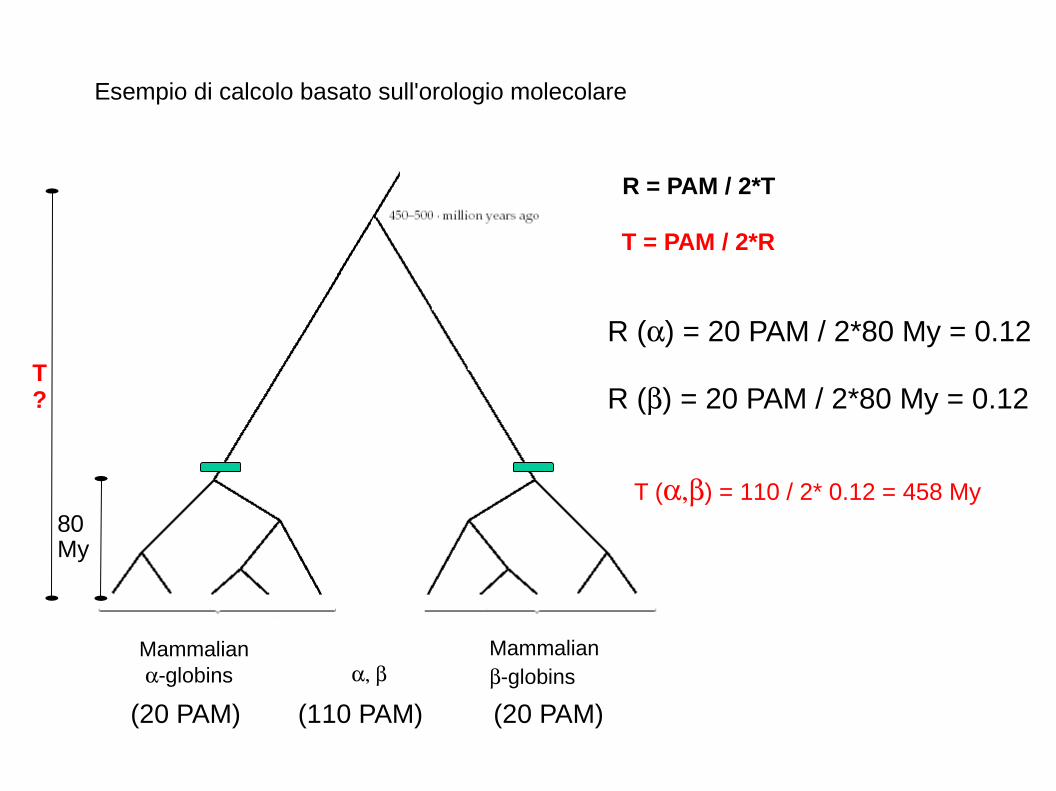
Mammalian -globins
Mammalian -globins
Esempio di calcolo basato sull'orologio molecolare
R () = 20 PAM / 2*80 My = 0.12
R () = 20 PAM / 2*80 My = 0.12
R = PAM / 2*T
T = PAM / 2*R
T () = 110 / 2* 0.12 = 458 My
T?
80My
(20 PAM) (20 PAM)(110 PAM)

Proteine diverse, velocità di evoluzione diverse
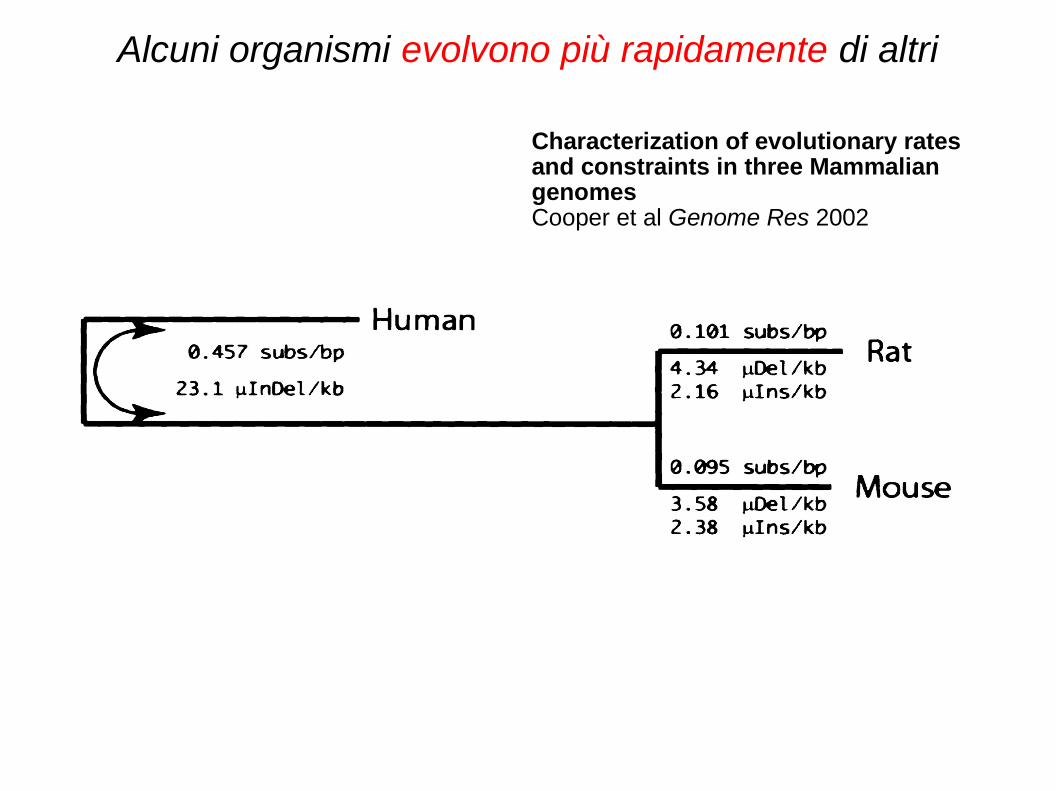
Alcuni organismi evolvono più rapidamente di altri
Characterization of evolutionary rates and constraints in three Mammalian genomesCooper et al Genome Res 2002

Gli uomini evolvono (forse) più rapidamente delle donne...
Strong male-driven evolution of DNA sequences in humans and apesMAKOVA AND LI, Nature 2002
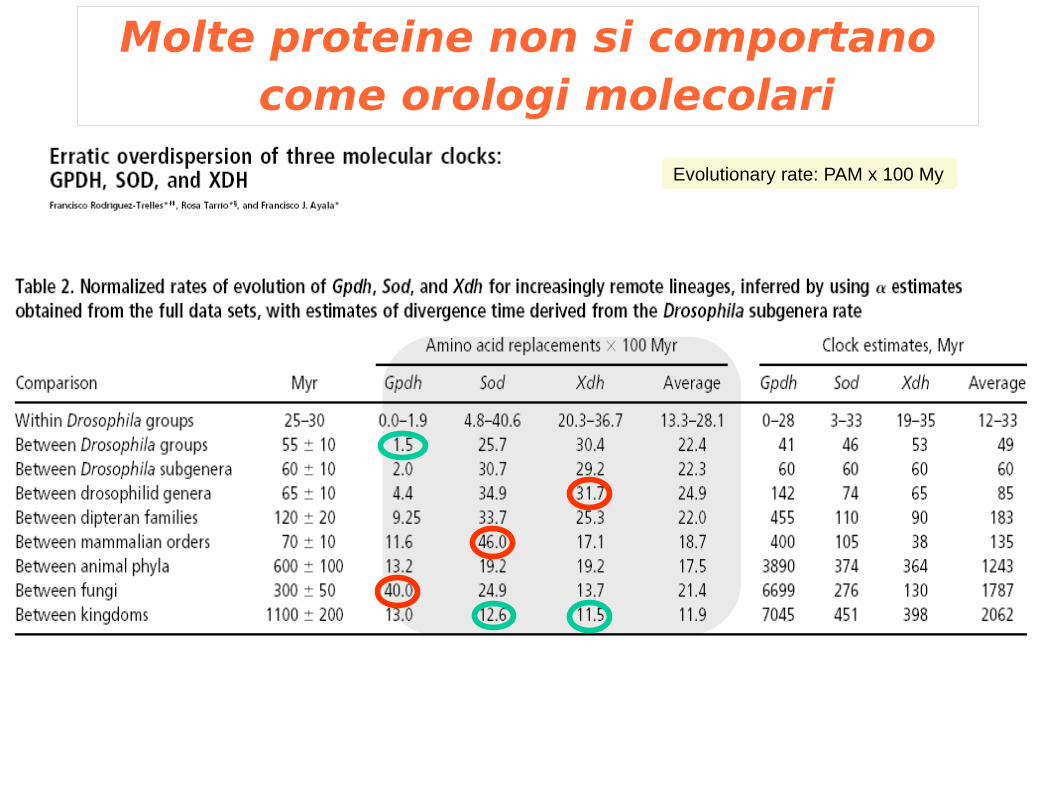
Molte proteine non si comportano come orologi molecolari
Evolutionary rate: PAM x 100 My

Cause delle deviazioni dall'orologio molecolare
1. Tasso di mutazione: Caratteristiche specie-specifiche negli enzimi di replicazione e riparo del DNA influiscono sulla velocità di mutazione
2. Tempo di generazione: Tempi di generazione differenti determinano una diversa velocità di fissazione delle mutazioni
3. Selezione naturale: Gli organismi possono essere soggetti in modo differente alla selezione naturale. Variazioni ambientali possono avere significato differente per i diversi organismi.
4. Funzione del gene: Cambiamenti funzionali di un gene nel corso dell'evoluzione possono determinare variazioni nella velocità di mutazione
Ayala, F. Bioessays 1999 Jan;21(1):71-5

Analisi dei cambiamenti evolutivi in relazione alla funzione
- Tipo e sede delle mutazioni
- Velocità di accumulo delle mutazioni
Determinazione delle pressione selettiva:
- Selezione purificatrice
- Neutralità
- Selezione positiva (adattamento)
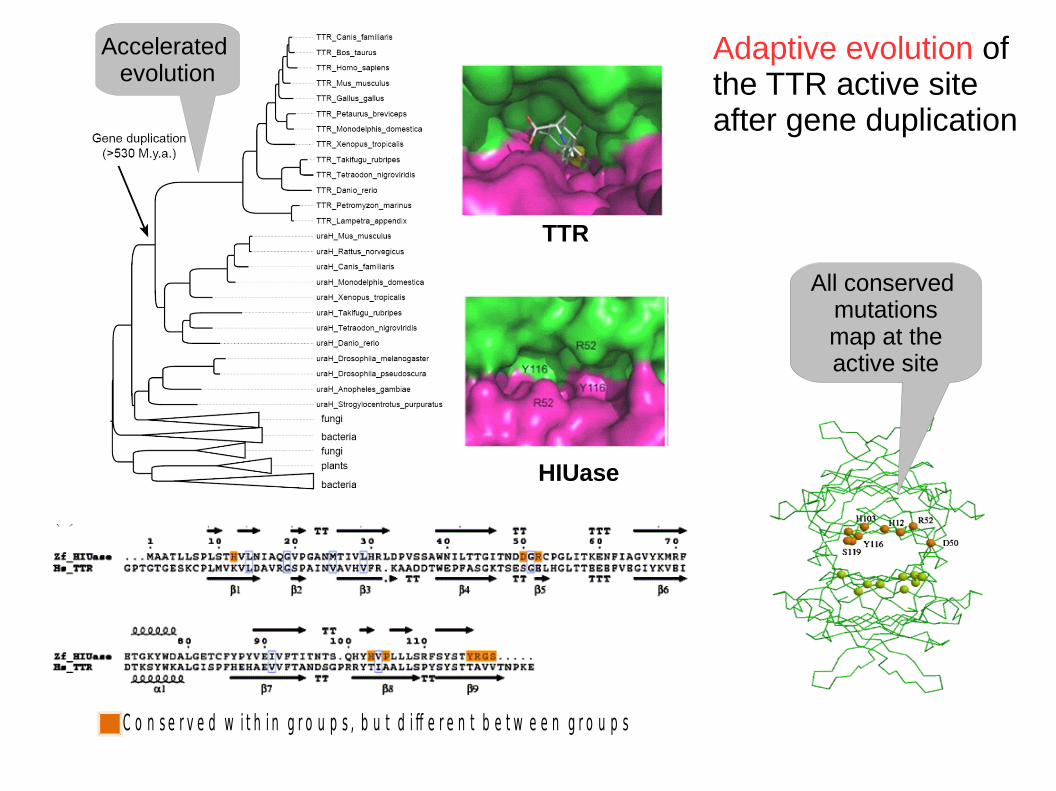
Adaptive evolution of the TTR active site after gene duplication
H IU a se T T R
C o n s e r v e d w i t h i n g r o u p s , b u t d i ff e r e n t b e t w e e n g r o u p s
HIUase
TTR
H IU a se T T R
Accelerated evolution
All conserved mutations
map at the active site
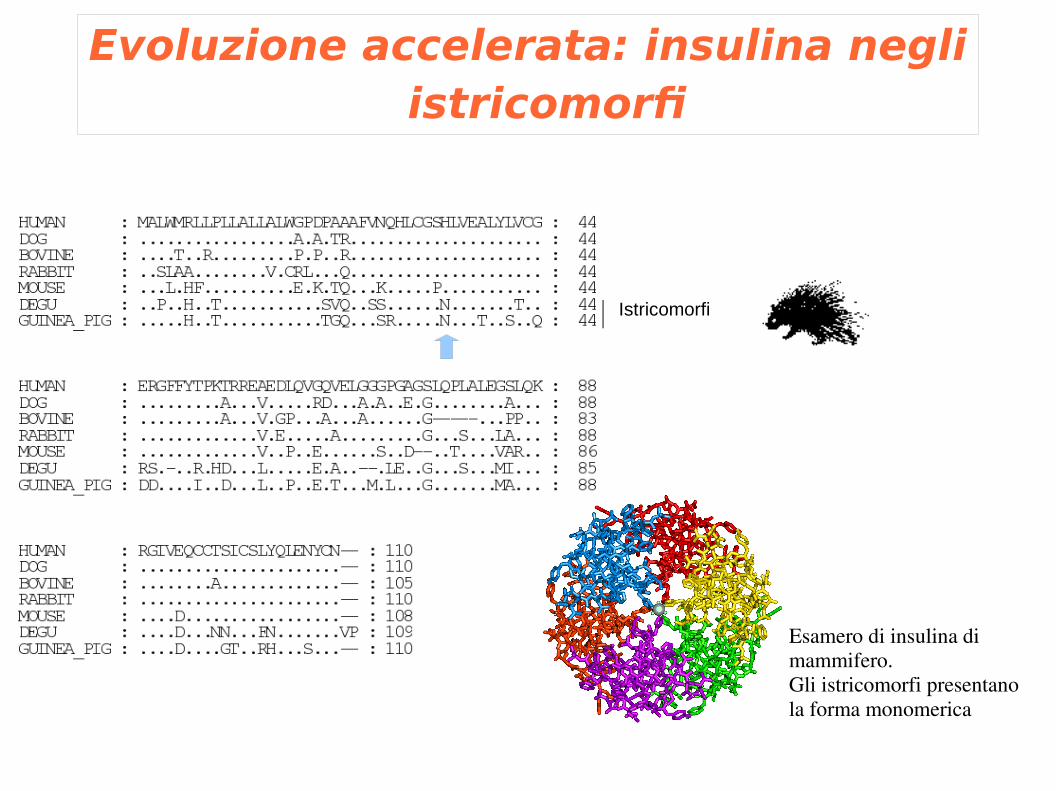
Evoluzione accelerata: insulina negli istricomorfi
Istricomorfi
Esamero di insulina di mammifero.Gli istricomorfi presentano la forma monomerica
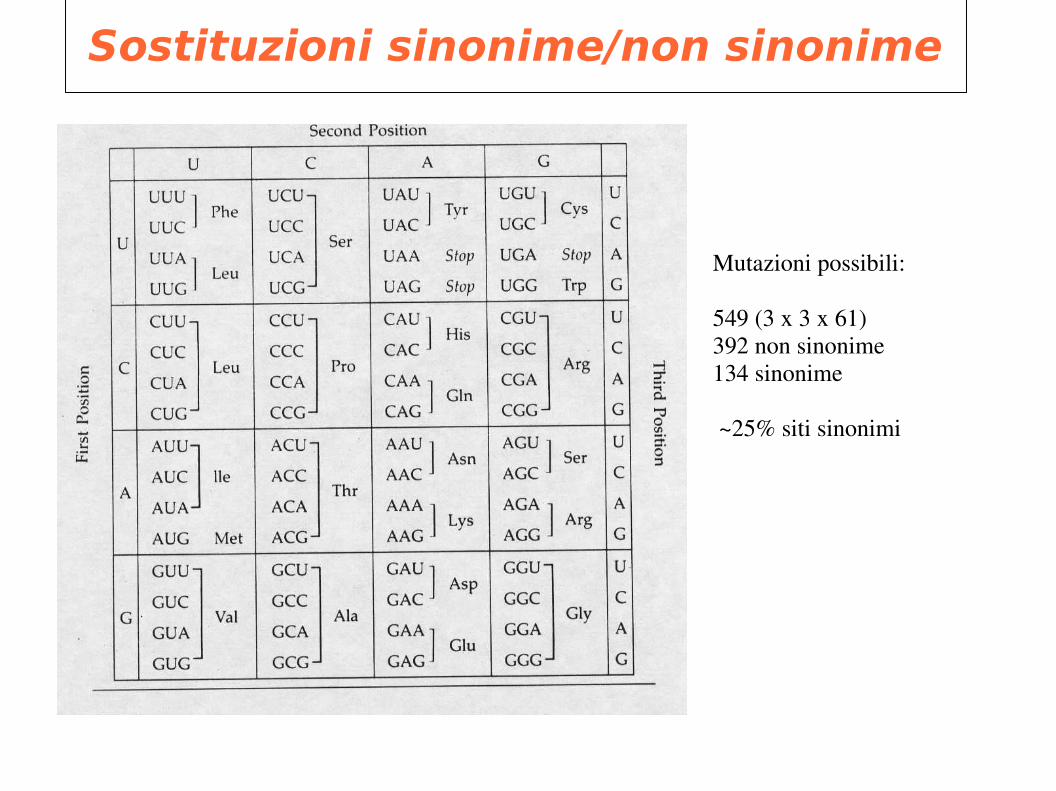
Sostituzioni sinonime/non sinonime
Mutazioni possibili: 549 (3 x 3 x 61) 392 non sinonime134 sinonime
~25% siti sinonimi

Test Ka/Ks
KN numero di sostituzioni / totale siti
KA numero di sostituzioni aminoacidiche / totale siti non sinonimi
KS numero di sostituzioni sinonime / totale siti sinonimi
KA/KS < 1 selezione purificatrice
KA/KS ~ 1 neutralità (pseudogeni)
KA/KS >1 selezione positiva (adattamento)
KA KS KA/KS
Istone 3 0 6.38 0Istone 4 0 6.12 0Actina alpha 0.01 3.68 0.002Insulina 0.13 4.02 0.010Mioglobina 0.56 4.44 0.126Urochinasi 1.28 3.92 0.326Ig K 1.87 5.90 0.316

Adattamento evolutivo nel lisozimaMessier et al. Nature, 1997

Pseudogenizzazione della miosina 16 e cambiamenti anatomici nell'evoluzione umana
Inattivazione della miosina 16 (MYH16) negli ominidi per mutazione di frameshift nell'esone 18
Data stimata inattivazione di MYH16: 2.4 Myr
STEDMAN et al, NATURE 2004
dN:
sost. non sinonime; ds: sost. sinonime

Calcolo del tempo di inattivazione del gene CHOU et al. PNAS 2002; STEDMAN et al, NATURE 2004
dN mutaz. non sinonime; d
s: mutaz sinonime
w1 = d
N1/d
s (pseudogene);
w = dN/d
s (altre specie)
dN1
mutaz. non sinonime nello pseudogene
[NB: dN
equivalente a Ka; ds equivalente a Ks]
T 1=Tw1−w
1−w
X
t1
t-t1
X


















