Evidence that the limb bud ectoderm is required for ... · PDF fileEvidence that the limb bud...
12
Evidence that the limb bud ectoderm is required for survival of the underlying mesoderm Marian Fernandez-Teran a , Maria A. Ros a,b , Francesca V. Mariani c,n a Departamento de Anatomía y Biología Celular, Facultad de Medicina, Universidad de Cantabria, 39011 Santander, Spain b Instituto de Biomedicina y Biotecnología de Cantabria, CSIC–SODERCAN–Universidad de Cantabria, 39011 Santander, Spain c Department of Cell and Neurobiology, Broad CIRM Center for Regenerative Medicine & Stem Cell Research, Keck School of Medicine, University of Southern California, Los Angeles, CA 90089, United States article info Article history: Received 5 May 2013 Received in revised form 21 June 2013 Accepted 25 June 2013 Available online 9 July 2013 Keywords: Limb development Pattern formation AER Dorsal ectoderm Cell death abstract The limb forms from a bud of mesoderm encased in a hull of ectoderm that grows out from the flank of the embryo. Coordinated signaling between the limb mesoderm and ectoderm is critical for normal limb outgrowth and patterning. The apical ectodermal ridge (AER), found at the distal tip, is a rich source of signaling molecules and has been proposed to specify distal structures and maintain the survival of cells in the underlying distal mesoderm. The dorsal and ventral non-AER ectoderm is also a source of signaling molecules and is important for dorsal–ventral patterning of the limb bud. Here we determine if this ectoderm provides cell survival signals by surgically removing the dorsal or ventral ectoderm during early chicken limb bud development and assaying for programmed cell death. We find that, similar to the AER, removal of the dorsal or ventral non-AER ectoderm results in massive cell death in the underlying mesoderm. In addition, although a re-epithelialization occurs, we find perturbations in the timing of Shh expression and, for the case of the dorsal ectoderm removal, defects in soft tissue and skeletal development along the proximal–distal axis. Furthermore, ectoderm substitution experiments show that the survival signal produced by the dorsal limb ectoderm is specific. Thus, our results argue that the non- AER ectoderm, like the AER, provides a specific survival signal to the underlying mesoderm that is necessary for normal limb development and conclusions drawn from experiments in which the non-AER ectoderm is removed, need to take into consideration this observation. & 2013 Elsevier Inc. All rights reserved. Introduction Vertebrate limb development is considered an excellent model for developmental biology studies. The first morphological evi- dence of limb formation is the outgrowth of a bud from the lateral flank of the embryo. This bud consists of an accumulation of mesodermally-derived cells encased in an ectoderm hull. Critical cell–cell signaling interactions occur between these mesodermal and ectodermal components to direct outgrowth and patterning (Benazet and Zeller, 2009). In amniotes, the distal ectoderm of the limb bud adopts a thickened morphology at the dorso-ventral boundary called the apical ectodermal ridge (AER). This specia- lized region of ectoderm expresses a number of important signal- ing molecules that control limb patterning but also provides survival signals for the underlying distal mesoderm (as reviewed in Fernandez-Teran and Ros, 2008). Experiments carried out by John Saunders in the late 1940s showed that removal of the AER in chicken embryos causes limb truncations (Saunders, 1948). Early removal of the AER (stage 17–18HH) resulted in the most severe truncations at a proximal level, while late removal of the AER (stage 25 and later) resulted in progressively less severe distal truncations. The outcomes of this experiment were used by Wolpert and colleagues to propose the Progress Zone Model of limb patterning (Summerbell et al., 1973). Later, this model was questioned as it was shown that the AER is critical first for survival, and later for the proliferation of the subjacent mesoderm which provided an alternative explanation for the limb truncation phenotypes if proximal–distal specification occurs early (Dudley et al., 2002; Rowe et al., 1982). Thus, the AER clearly has an important role in limb development by promoting proliferation and survival of the underlying limb mesoderm. Several members of the fibroblast growth factor (FGF) family are specifically expressed in the AER (Martin, 1998). Based on this observation, it was shown that the truncation phenotype after AER removal can be rescued by the exogenous administration of FGFs (Fallon et al., 1994; Niswander et al., 1994) suggesting that FGFs are the factors from the AER that control cell survival. Four members of the Fgf gene family, Fgf8, Fgf4, Fgf9 and Fgf17, are expressed in Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/developmentalbiology Developmental Biology 0012-1606/$ - see front matter & 2013 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.ydbio.2013.06.032 n Corresponding author. E-mail addresses: [email protected] (M.A. Ros), [email protected] (F.V. Mariani). Developmental Biology 381 (2013) 341–352
-
Upload
nguyenkhanh -
Category
Documents
-
view
216 -
download
2
Transcript of Evidence that the limb bud ectoderm is required for ... · PDF fileEvidence that the limb bud...

Developmental Biology 381 (2013) 341–352
Contents lists available at ScienceDirect
Developmental Biology
0012-16http://d
n CorrE-m
journal homepage: www.elsevier.com/locate/developmentalbiology
Evidence that the limb bud ectoderm is required for survivalof the underlying mesoderm
Marian Fernandez-Teran a, Maria A. Ros a,b, Francesca V. Mariani c,n
a Departamento de Anatomía y Biología Celular, Facultad de Medicina, Universidad de Cantabria, 39011 Santander, Spainb Instituto de Biomedicina y Biotecnología de Cantabria, CSIC–SODERCAN–Universidad de Cantabria, 39011 Santander, Spainc Department of Cell and Neurobiology, Broad CIRM Center for Regenerative Medicine & Stem Cell Research, Keck School of Medicine,University of Southern California, Los Angeles, CA 90089, United States
a r t i c l e i n f o
Article history:Received 5 May 2013Received in revised form21 June 2013Accepted 25 June 2013Available online 9 July 2013
Keywords:Limb developmentPattern formationAERDorsal ectodermCell death
06/$ - see front matter & 2013 Elsevier Inc. Ax.doi.org/10.1016/j.ydbio.2013.06.032
esponding author.ail addresses: [email protected] (M.A. Ros), fma
a b s t r a c t
The limb forms from a bud of mesoderm encased in a hull of ectoderm that grows out from the flank ofthe embryo. Coordinated signaling between the limb mesoderm and ectoderm is critical for normal limboutgrowth and patterning. The apical ectodermal ridge (AER), found at the distal tip, is a rich source ofsignaling molecules and has been proposed to specify distal structures and maintain the survival of cellsin the underlying distal mesoderm. The dorsal and ventral non-AER ectoderm is also a source of signalingmolecules and is important for dorsal–ventral patterning of the limb bud. Here we determine if thisectoderm provides cell survival signals by surgically removing the dorsal or ventral ectoderm duringearly chicken limb bud development and assaying for programmed cell death. We find that, similar to theAER, removal of the dorsal or ventral non-AER ectoderm results in massive cell death in the underlyingmesoderm. In addition, although a re-epithelialization occurs, we find perturbations in the timing ofShh expression and, for the case of the dorsal ectoderm removal, defects in soft tissue and skeletaldevelopment along the proximal–distal axis. Furthermore, ectoderm substitution experiments show thatthe survival signal produced by the dorsal limb ectoderm is specific. Thus, our results argue that the non-AER ectoderm, like the AER, provides a specific survival signal to the underlying mesoderm that isnecessary for normal limb development and conclusions drawn from experiments in which the non-AERectoderm is removed, need to take into consideration this observation.
& 2013 Elsevier Inc. All rights reserved.
Introduction
Vertebrate limb development is considered an excellent modelfor developmental biology studies. The first morphological evi-dence of limb formation is the outgrowth of a bud from the lateralflank of the embryo. This bud consists of an accumulation ofmesodermally-derived cells encased in an ectoderm hull. Criticalcell–cell signaling interactions occur between these mesodermaland ectodermal components to direct outgrowth and patterning(Benazet and Zeller, 2009). In amniotes, the distal ectoderm of thelimb bud adopts a thickened morphology at the dorso-ventralboundary called the apical ectodermal ridge (AER). This specia-lized region of ectoderm expresses a number of important signal-ing molecules that control limb patterning but also providessurvival signals for the underlying distal mesoderm (as reviewedin Fernandez-Teran and Ros, 2008). Experiments carried out byJohn Saunders in the late 1940s showed that removal of the AER in
ll rights reserved.
[email protected] (F.V. Mariani).
chicken embryos causes limb truncations (Saunders, 1948). Earlyremoval of the AER (stage 17–18HH) resulted in the most severetruncations at a proximal level, while late removal of the AER(stage 25 and later) resulted in progressively less severe distaltruncations. The outcomes of this experiment were used byWolpert and colleagues to propose the Progress Zone Model oflimb patterning (Summerbell et al., 1973). Later, this model wasquestioned as it was shown that the AER is critical first forsurvival, and later for the proliferation of the subjacent mesodermwhich provided an alternative explanation for the limb truncationphenotypes if proximal–distal specification occurs early (Dudleyet al., 2002; Rowe et al., 1982). Thus, the AER clearly has animportant role in limb development by promoting proliferationand survival of the underlying limb mesoderm.
Several members of the fibroblast growth factor (FGF) familyare specifically expressed in the AER (Martin, 1998). Based on thisobservation, it was shown that the truncation phenotype after AERremoval can be rescued by the exogenous administration of FGFs(Fallon et al., 1994; Niswander et al., 1994) suggesting that FGFs arethe factors from the AER that control cell survival. Four membersof the Fgf gene family, Fgf8, Fgf4, Fgf9 and Fgf17, are expressed in

M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352342
the AER in both mouse and chicken embryos. To study AER–FGFfunction, these genes have been inactivated in the mouse singlyand in combination and analyzed for their contribution to limbbud development and skeletal patterning (Mariani et al., 2008;Mariani and Martin, 2003). The individual inactivation of Fgf4, Fgf9or Fgf17 has no consequence on overall limb patterning (Colvinet al., 2001; Moon et al., 2000; Sun et al., 2000; Xu et al., 2000).However, the inactivation of Fgf8 or, Fgf8 in combination with anyof the other AER–FGFs, results in alterations in limb pattern thatcan range in severity with the most extreme being a complete lossof the limb (Boulet et al., 2004; Lewandoski et al., 2000; Marianiet al., 2008; Moon and Capecchi, 2000; Sun et al., 2002). Analysisof limb bud development in different AER–FGF combinationknock-out lines showed changes in limb bud size, cell survival,gene expression, and skeletal pattern that correlated with thestrength of the FGF signal (Mariani et al., 2008; Mariani andMartin, 2003). Thus, in addition to providing survival and prolif-erative signals, the AER controls limb skeletal patterning and theAER-FGFs are critical mediators of this activity.
Besides the signaling interactions between the AER and meso-derm, the non-AER ectoderm has a role in limb patterning. Inparticular, the ectoderm on the dorsal and ventral sides of the limbbud is known to influence dorsal and ventral patterning. Expres-sion of the En1 transcription factor in the ventral ectodermrestricts Wnt7a to the dorsal ectoderm (Cygan et al., 1997;Loomis et al., 1998). Wnt7a from the dorsal ectoderm inducesthe expression of the homeobox gene Lmx1b, a homeobox-containing transcription factor responsible for establishing dorsalidentity in the subjacent mesoderm (Loomis et al., 1998; Riddleet al., 1995).Wnt7a also influences anterior/posterior patterning bymaintaining normal levels of Shh expression and loss of Wnt7a inthe mouse results in dorsal to ventral transformations and in avariable loss or malformation of posterior structures, mostly digit5 and the ulna (Parr and McMahon, 1995; Yang and Niswander,1995). A number of studies also show that the limb ectoderm is anegative regulator of chondrogenic differentiation, a function thatappears to be mediated by canonical Wnt signaling (Hartmann,2006; ten Berge et al., 2008).
In analyzing the patterns of programmed cell death in AER–FGFmutants we noticed that not only was there cell death in theproximal mesoderm of the limb bud in the AER–FGF mutants butalso cell death in the proximal dorsal ectoderm (Boulet et al.,2004; Mariani et al., 2008; Sun et al., 2002) and wondered if theremight be a relationship between the death of the ectoderm andthe survival of the underlying mesoderm. We therefore decided todetermine if, like the AER, the non-AER ectoderm also has asurvival function in addition to a role in patterning, and if so, ifremoval of the ectoderm would impact limb bud morphologicaldevelopment, gene expression, and skeletal patterning.
Materials and methods
Embryos
Fertilized chicken eggs were obtained from local sources. Eggswere incubated, opened, and embryos staged following standardprotocols (Hamburger and Hamilton, 1992; Ros et al., 2000).GFP-transgenic chicken embryos (McGrew et al., 2004) werelocally produced at the Servicio de Estabulación y Experimenta-ción Animal of the University of Cantabria. Mouse embryosdeficient for Fgf8 specifically in the AER were generated employingan Msx2-Cre transgene as described previously (Lewandoski et al.,2000; Mariani et al., 2008; Sun et al., 2002).
Removal of limb ectoderm
Right wing buds of stage 19–21HH embryos were exposed andthe dorsal or ventral ectoderm was surgically removed with theaid of Nile blue sulfate (NBS) application (about one microliter of0.15% NBS in distilled water applied over the limb surface). NBSstaining not only allowed easy visualization of the ectoderm butalso loosened up the ectoderm all the way to the lateral border ofthe somites. After a few seconds, the NBS was washed away with1� PBS to avoid toxicity. Then, using a fine sharpened tungstenneedle, a superficial cut was made all along the junction betweenthe dorsal or ventral ectoderm and the AER. This microsurgicaltechnique was not used in previous studies (Yang and Niswander,1995) and was introduced here to prevent undesired damageor detachment of the AER when peeling off the ectoderm.To compare with previous reports, the removal of the dorsalectoderm was done without the previous cut. Removal of justthe AER was done with the aid of a fine tungsten needle.
In situ hybridization, histology and skeletal preparations
Digoxigenin-labeled antisense riboprobes were prepared, andwhole mount in situ hybridization performed according to stan-dard procedures (Nieto et al., 1996). The probes used were Wnt7aand Lmx1b (Riddle et al., 1995), Shh (Roelink et al., 1994) and Wnt6(ARK genomics, ChEST972J11).
For histology, samples were routinely embedded in paraffin,sectioned and stained with Hematoxylin–Eosin. Some sampleswere embedded in araldite for semi-thin (1 μm thick) sections andstained with Toluidine blue according to standard protocols.
Scanning electron microscopy
Experimental and control wing buds were fixed in 2.5%Glutaraldehyde in 0.1 M-cacodylate buffer (pH 7.2), dehydratedin acetone, dried by the critical point method and then coatedwith gold and observed with an Inspect S microscope (FEICompany).
Cell death analysis
Cell death was detected by in situ detection of DNA fragmenta-tion using terminal deoxynucleotidyl transferase (TdT) mediateddeoxyuridine-triphosphate (dUTP) nick end-labeling (TUNEL)with the In Situ Cell Death Detection Kit, Fluorescein (Roche cat#S7110) on embryo sections or by staining whole embryos withLysoTracker Red-DND99 (Life Technologies, cat #L-7528) as pre-viously described (Fogel et al., 2012) and subsequently sectioningthem at 25 μm on a Leica vibrating microtome.
Recombinant limb experiments
Recombinant limbs were prepared by assembling operatedwing buds inside limb ectodermal jackets (Ros et al., 2000). Toobtain ectodermal jackets, forelimb or hindlimb buds wereremoved from stage 21–22HH chicken embryos and incubated at4 1C for 60 min in 0.5% trypsin. Trypsin was inactivated byincubation in 10% chicken serum for 5 min on ice before gentlyseparating the ectoderm from the mesoderm. In some cases, therecombinant limbs were generated by wrapping the operated limbbuds with back ectoderm obtained from the dorsal flank region ofstage 21–22 embryos as devised by Errick and Saunders (1976).The back ectoderm was obtained following the same procedure asfor the ectodermal jackets. The recombinant limbs were grafted tothe paraxial mesoderm of stage HH20–22 host embryos.

+20h +18h-AER
+18h
DER at Stage 20-21HH
DER at Stage 23HH
DER at Stage 24HH
NBS at Stage 20-21HH
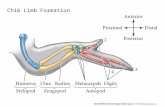
Fig. 1. Pattern of cell death after removal of the dorsal ectoderm. (A)–(D) The dorsal limb ectoderm was removed at Stage 20–21HH and the embryos re-incubated after theoperation for the indicated times. Cell death was observed as soon as 4 h after surgery and was quite extensive by 20 h. (E) When the dorsal ectoderm (DE) is removed alongwith the AER, by 18 h, the limb is much reduced in size and contains large domains of cell death. (F) The contra-lateral limb of B is shown as a control. (G), (H) Show theresults of removing the DE at stage 23HH, 8 h later. (I) (J) Show the results of removing the DE at stage 24HH, 8 h later. At both time points cell death is observed in the dorsalmesoderm when the ectoderm is removed but a lower levels than when the removal is done at 20–21HH. Note in I and J the cell death in the middle of the budcorresponding to the ‘opaque patch’ (Fernandez-Teran et al., 2006). (K) To control for NBS toxicity, the dorsal limb ectoderm on one side (arrow) was treated with NBS inexcess at 20–21HH and then cultured for 18 h. No ectopic cell death was seen. (L) Skeletal preparation of these samples showed no gross defects in skeletal patterning.Unmanipulated (top), NBS treated (bottom).
M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352 343
M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352344
Results
Removal of the dorsal ectoderm leads to massive cell death in theunderlying mesoderm
To determine if the ectoderm has a survival role, we firstdecided to remove the dorsal ectoderm of stage 20–21HH wingbuds with and without simultaneous removal of the AER andevaluate the consequences on the survival of the mesoderm(Fig. 1). Our results showed that the removal of the dorsalectoderm was followed by massive cell death mainly located inthe underlying dorsal mesoderm. This cell death was clearlyvisible 3–4 h after the operation (Fig. 1A) and persisted up to20 h, after which, cell death tapered off (Fig. 1B–D). In another
h r
u
EMR EDC EMU
EMLANC
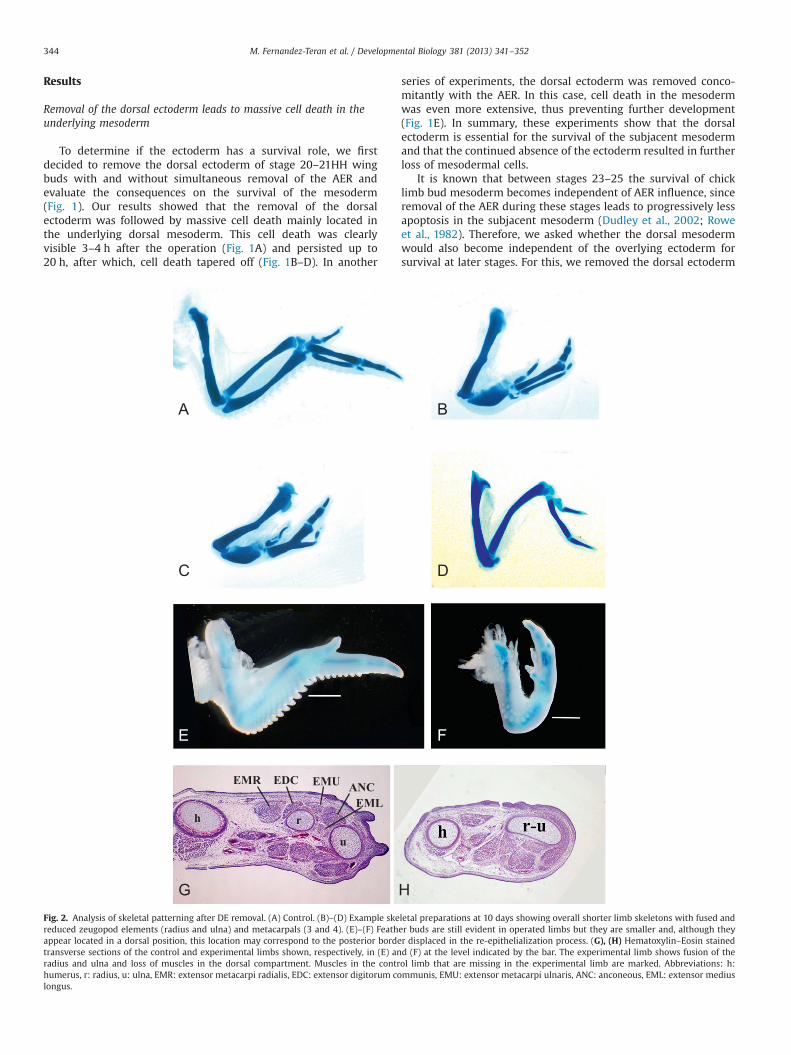
Fig. 2. Analysis of skeletal patterning after DE removal. (A) Control. (B)–(D) Example skereduced zeugopod elements (radius and ulna) and metacarpals (3 and 4). (E)–(F) Feathappear located in a dorsal position, this location may correspond to the posterior bordetransverse sections of the control and experimental limbs shown, respectively, in (E) anradius and ulna and loss of muscles in the dorsal compartment. Muscles in the contrhumerus, r: radius, u: ulna, EMR: extensor metacarpi radialis, EDC: extensor digitorum colongus.
series of experiments, the dorsal ectoderm was removed conco-mitantly with the AER. In this case, cell death in the mesodermwas even more extensive, thus preventing further development(Fig. 1E). In summary, these experiments show that the dorsalectoderm is essential for the survival of the subjacent mesodermand that the continued absence of the ectoderm resulted in furtherloss of mesodermal cells.
It is known that between stages 23–25 the survival of chicklimb bud mesoderm becomes independent of AER influence, sinceremoval of the AER during these stages leads to progressively lessapoptosis in the subjacent mesoderm (Dudley et al., 2002; Roweet al., 1982). Therefore, we asked whether the dorsal mesodermwould also become independent of the overlying ectoderm forsurvival at later stages. For this, we removed the dorsal ectoderm
letal preparations at 10 days showing overall shorter limb skeletons with fused ander buds are still evident in operated limbs but they are smaller and, although theyr displaced in the re-epithelialization process. (G), (H) Hematoxylin–Eosin stainedd (F) at the level indicated by the bar. The experimental limb shows fusion of theol limb that are missing in the experimental limb are marked. Abbreviations: h:mmunis, EMU: extensor metacarpi ulnaris, ANC: anconeous, EML: extensor medius
M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352 345
in stage 23 and 24 wing buds and analyzed cell death after theoperation at several time-points. Similar to what has beenreported for the AER-removal experiments, our results showedprogressively less apoptosis as the ectoderm is removed at laterstages, indicating that from stage 23HH onwards the survival ofthe dorsal mesoderm becomes progressively less sensitive to theremoval of the dorsal ectoderm (Fig. 1G–J). AER removal at laterstages leads to a reduction in the proliferation of the subjacentmesoderm and subsequent distal truncations (Dudley et al., 2002).Whether the dorsal and ventral ectoderm may have an effect onthe proliferation rate of the peripheral mesoderm is yet to bedetermined.
Nile blue sulfate (NBS) was used to remove the ectoderm (seethe Materials and methods section) but is known to have a toxiceffect if a high amount (dosage) is applied. Application of NBS inexcess of what we used (3 ml instead of 1) failed to cause obvious
DER t0 DER+10
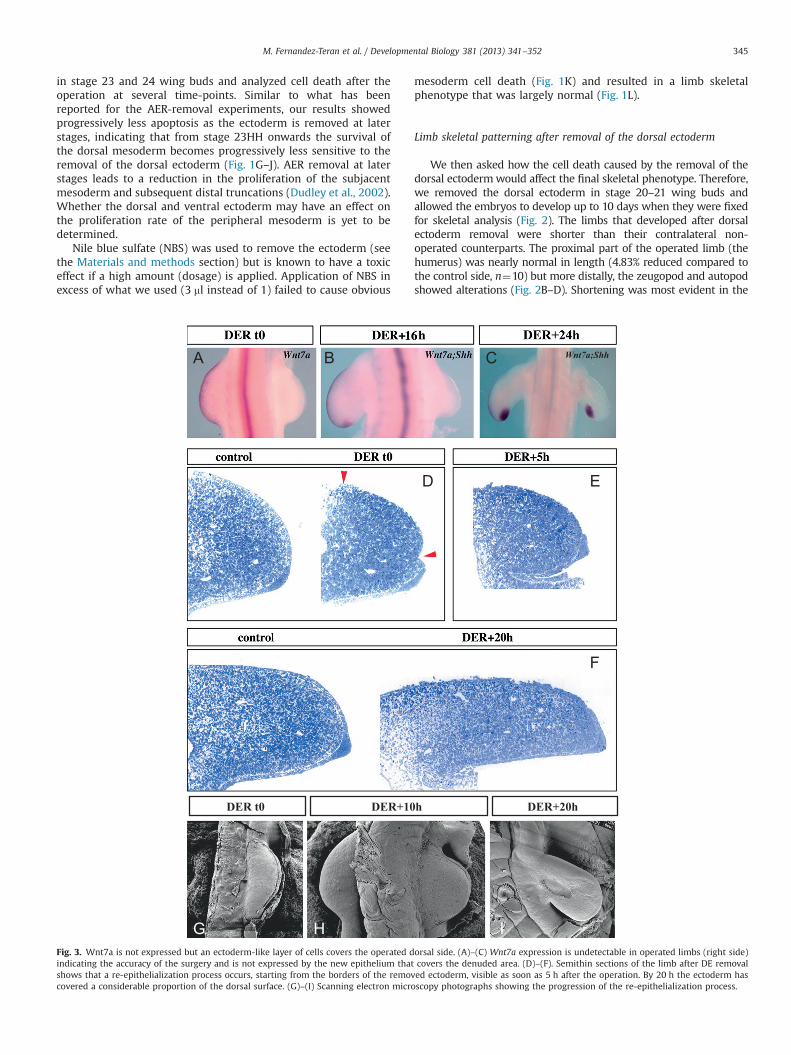
Fig. 3. Wnt7a is not expressed but an ectoderm-like layer of cells covers the operated dindicating the accuracy of the surgery and is not expressed by the new epithelium thatshows that a re-epithelialization process occurs, starting from the borders of the removcovered a considerable proportion of the dorsal surface. (G)–(I) Scanning electron micro
mesoderm cell death (Fig. 1K) and resulted in a limb skeletalphenotype that was largely normal (Fig. 1L).
Limb skeletal patterning after removal of the dorsal ectoderm
We then asked how the cell death caused by the removal of thedorsal ectodermwould affect the final skeletal phenotype. Therefore,we removed the dorsal ectoderm in stage 20–21 wing buds andallowed the embryos to develop up to 10 days when they were fixedfor skeletal analysis (Fig. 2). The limbs that developed after dorsalectoderm removal were shorter than their contralateral non-operated counterparts. The proximal part of the operated limb (thehumerus) was nearly normal in length (4.83% reduced compared tothe control side, n¼10) but more distally, the zeugopod and autopodshowed alterations (Fig. 2B–D). Shortening was most evident in the
Wnt7a;Shh
h DER+20h
orsal side. (A)–(C) Wnt7a expression is undetectable in operated limbs (right side)covers the denuded area. (D)–(F). Semithin sections of the limb after DE removaled ectoderm, visible as soon as 5 h after the operation. By 20 h the ectoderm hasscopy photographs showing the progression of the re-epithelialization process.

M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352346
zeugopod where the operated side was 40.4% reduced compared tothe control side (n¼10). In half the cases, the radius and ulnaappeared to be fused into a single thick misshaped element(Fig. 2B–C; 8 out of 16), while in the remaining of the cases the ulnaand radius, although shorter and closer together than normal, werestill distinguishable as individual elements (not shown). In theautopod, the reduction in size of the skeletal elements was moremoderate although the third and fourth metacarpals were frequentlyfused but not always (Fig. 2B–C) (24.3% reduced compared to thecontrol side, n¼10). In 2 cases out of 16 (12.5%), digit 4 was missing.We found these results surprising because previous studies in whichthe dorsal ectoderm had been removed, resulted in the loss of themost posterior digit, digit 4 in the chick wing, and occasionally alsothe ulna (Yang and Niswander, 1995) in a phenotype similar to that ofthe Wnt7a mutant limb (Parr and McMahon, 1995). However, in onlya minority of cases (2 out of 16) did we observe loss of digit 4 in ouroperated limbs.
One possible explanation for the difference in the outcomebetween the experiments presented here and those previouslyreported (Yang and Niswander, 1995) may reside in the procedureused to remove the dorsal ectoderm. As indicated in the Materialsand methods section, we used the classical procedure based on thetransient application of a concentrated solution of Nile blue sulfate(NBS) over the ectodermal surface. The NBS makes the ectodermblister and facilitates its separation from the subjacent mesoderm.However, it can be difficult to control the area in which the NBSacts and frequently, particularly at earlier stages, the AER may alsobe affected. Therefore, we introduced a modification of theprotocol in which we separated the AER from the dorsal ectodermwith a cut at the junction between the dorsal ectoderm and theAER. This cut prevents the AER from being pulled away when theectoderm is peeled off. To compare both procedures, we per-formed a series of experiments with and without this modifica-tion. We found that when the dorsal ectoderm was not previouslyseparated from the AER, the percentage of cases in which theposterior digit 4 was lost increased significantly (50%, 4 out of8 cases Fig. 2D). Thus the loss of digit 4 is possibly due to theremoval of the dorsal ectoderm without a superficial cut leadingeasily to a loosening of the AER at a posterior level. Interestingly,removal of the posterior AER, parallel to somite levels mid-18 tomid-19 results in the deletion of digit 4 in 71% of cases (Rowe andFallon, 1981). Therefore, we conclude that the loss of posteriorstructures after removal of the dorsal ectoderm is likely caused bythe damage to the posterior AER.
Besides the reduction in size and the skeletal abnormalities, thelimb that develops after removal of the dorsal ectoderm changesshape and became flat dorsally in stark contrast to the roundedshape of the normal dorsal limb bud. At least two factors likelycontributed to this change in shape—the contraction of the bordersof the ectoderm wound and the loss of dorsal tissue due to celldeath. The operated limb lacked muscles in the dorsal compart-ment as well as the muscles between the two zeugopod elements(Fig. 2G–H).
After the operation the ectoderm heals over the dorsal mesoderm butdoes not re-express Wnt7a
To evaluate whether some dorsal ectoderm may have been leftin place after the experiment, we assessed Wnt7a expression, amarker of the dorsal ectoderm. Removal of the dorsal ectodermresulted in the total absence of Wnt7a expression in the denudedregion when evaluated immediately after the operation (100%n¼4; Fig. 3A) indicating that the removal procedure was complete.We also looked at time points after the procedure to see if Wnt7aexpression ever recovered. We found that Wnt7a expression was
not restored in the experimental limb at least up to 24 h after theoperation, the period analyzed (n¼6, Fig. 3B–C).
Even though Wnt7a expression is not detectable, the ectodermcould still have healed over the denuded mesoderm. To investigatethis point we analyzed semi-thin sections of experimental limbsfixed at different time-points after surgical manipulation at stage20HH (Fig. 3D–F). Our analysis showed that a re-epithelializationoccurred, starting from the borders of the removed ectoderm, andclearly visible by 5 h after the operation (Fig. 3E). By 20 h after theoperation, the ectoderm, although somewhat different morpholo-gically compared to a normal limb bud, had covered most of thedorsal surface of the limb (Fig. 3F). Thus, although Wnt7a was notdetected, the ectoderm did heal over the exposed mesoderm.Identical results were found when the re-epithelialization wasexamined by scanning electron microscopy (Fig. 3G–I).
Downregulation of Shh expression after removal of the dorsalectoderm
It has been shown that Shh expression in the ZPA depends onsignals from the dorsal ectoderm, specifically Wnt7a (Parr andMcMahon, 1995; Yang and Niswander, 1995). Therefore, wedecided to investigate Shh expression in our operated limb budsin which Wnt7a expression was lost, although the posterior digits,which are thought to depend on Shh expression were still presentin the majority of cases. Analysis of Shh expression after removal ofthe dorsal ectoderm showed a dramatic down-regulation occur-ring between 1 and 3 h after the operation (n¼7, Fig. 4A and B).In the interval between 3 and 6 h after the operation Shh expres-sion was strongly down-regulated (n¼7) and in some specimensbarely detectable (4 out of 7; Fig. 4B–C). Later, from 16 h after theoperation, Shh expression was generally detected (8 out of 11) inthe posterior limb bud border however at a lower level and in areduced domain when compared to the normal contralateral limb(Fig. 4D–G). It is important to note that residual Shh expression inthe posterior mesoderm was not accompanied by significantWnt7a re-expression since the ectoderm that overlies the Shh-expressing mesoderm did not express Wnt7a at detectable levelswhen analyzing tissue sections simultaneously for Shh and Wnt7aexpression (n¼2; Fig. 4H–J).
Another question we asked was whether the reduction instaining with the Shh RNA in situ hybridization probe mightsimply be a reflection of the loss of cells during the period ofmassive cell death after the operation. However, the observationthat Shh was rapidly down-regulated after the surgery (before thepeak of cell death) and that another gene, Lmx1b, was less rapidlyaffected (n¼2, Fig. 4K and L) suggested that Shh expression isparticularly sensitive to ectoderm influence in accordance with theknown requirement of Wnt7a for Shh expression (Parr andMcMahon, 1995).
Our results also indicate that a normal set of digits, includingposterior digits can still form even after a substantial reduction inthe level of Shh expression between stages 21HH to 25HH.
Tests for the specificity of the ectodermal signal
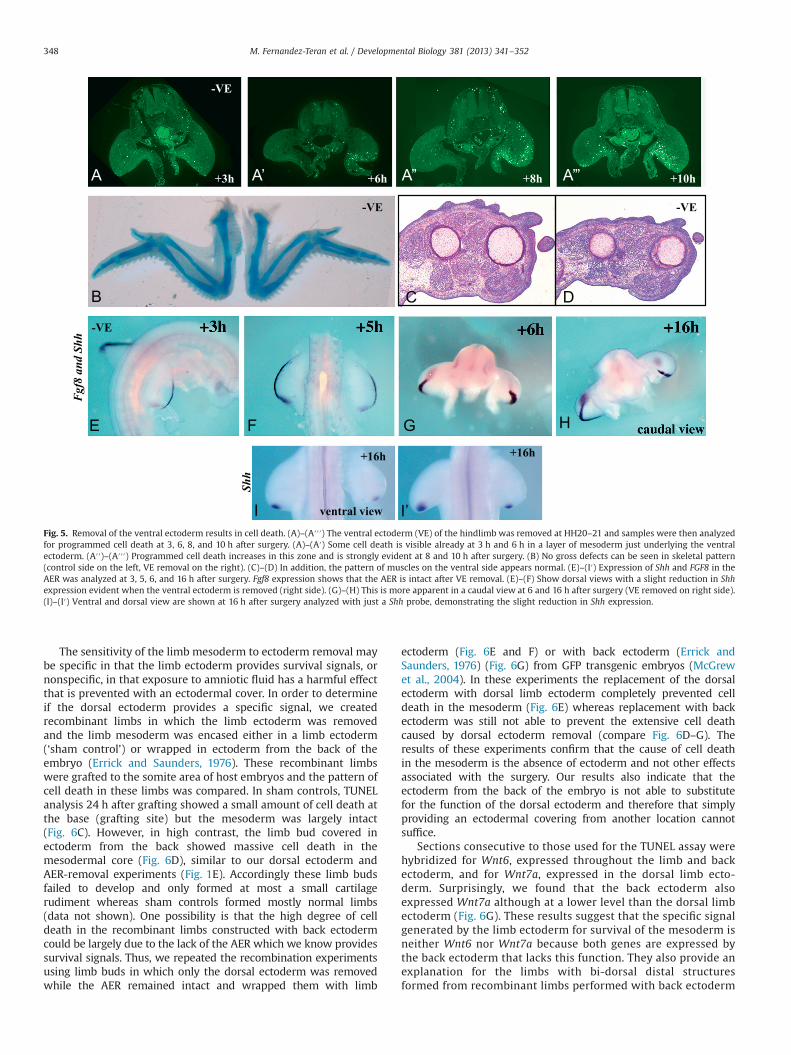
Our results so far indicate that the dorsal mesoderm requiresthe overlying ectoderm for survival. We also found that the ventralmesoderm is dependent on the overlying ectoderm for survivalsignals (Fig. 5). Ventral ectoderm removals at HH20–21 alsoresulted in TUNEL positivity in the ventral mesoderm visible from3 to 10 h after the operation (Fig. 5A′–A″′). The amount of celldeath was not as extensive as when the dorsal ectoderm isremoved and indeed both skeletal (Fig. 5B) and musclepatterning (Fig. 5C and D) were grossly normal. The effect onShh expression was still apparent but much milder than after

Lmx1
b
Fig. 4. Shh and Lmx1b expression after removal of the dorsal ectoderm. (A)–(C) Shh expressionwas dramatically reduced at 1 to 6 h after the removal of the dorsal ectoderm.(D)–(G) After this time point, Shh was detected but at lower levels than normal even 40 h after surgery. (H)–(J) Double RNA in situ hybridization for Wnt7a and Shh showingthat Wnt7a is not detectable in the operated limb in the ectoderm overlying Shh-expressing mesoderm 24 h after the surgery. (K)–(L) Lmx1b expression was still presentalthough notably decreased on the operated side at 6 h after surgery. Its expression is mostly gone by 16 h.
M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352 347
removal of the dorsal ectoderm (Fig. 5E–I′). Thus, removal of largeportions of either the dorsal or ventral ectoderm leads to death inthe subjacent mesoderm.
Previous studies have shown that removal of smaller patches ofdorsal ectoderm did not result in subjacent cell death (Rowe et al.,1982). To check the influence of the size of the ectoderm removed,we removed the proximal or distal half of the dorsal ectoderm at20–21HH and assayed for cell death after surgery. Proximal ecto-derm removal resulted in TUNEL positivity that was somewhat
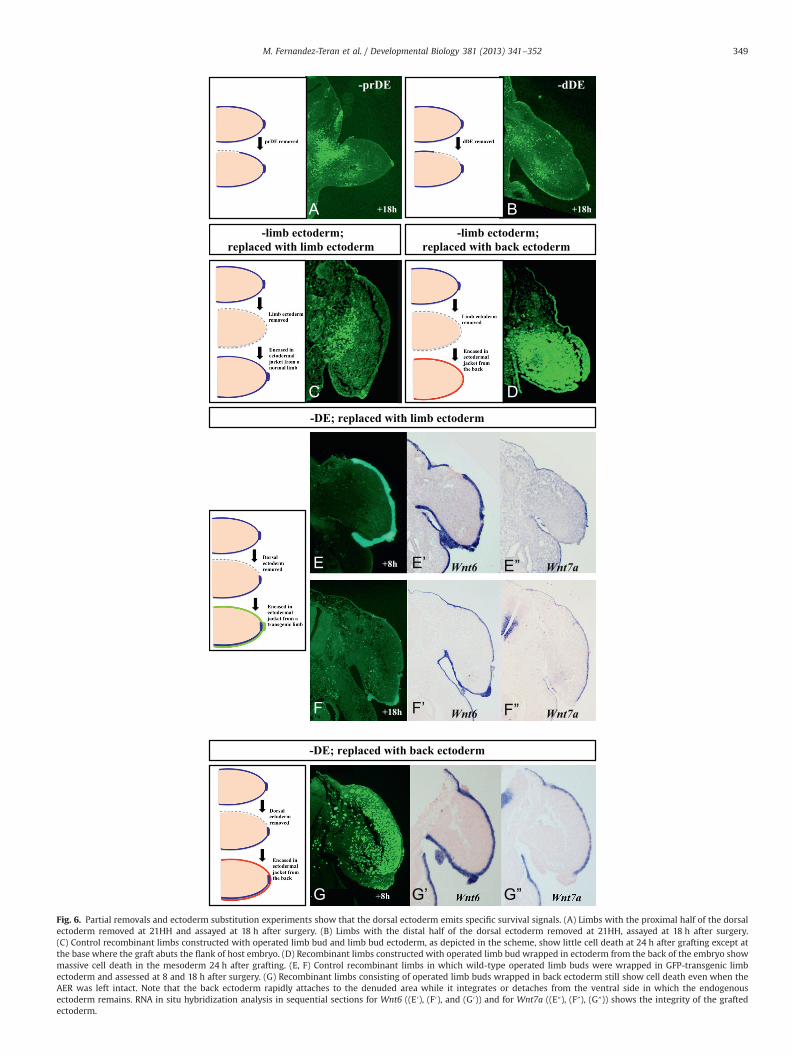
reduced but similar in location to full dorsal ectoderm removals(Fig. 6A; 18 h after surgery). Like full dorsal ectoderm removals, celldeath only occurred in a window of time after the surgery since celldeath was no longer evident at 24 h after surgery (data not shown).Interestingly distal dorsal ectoderm removals failed to result in celldeath (Fig. 6B; 18 h after the surgery) suggesting that distal cellsstill receive enough survival signals from the AER or possibly theremaining proximal dorsal ectoderm. Both proximal and distalectoderm removal had very mild skeletal defects (data not shown).
+3h +6h +8h +10h
+16h +16h
Fgf8
and
Shh
Shh
-VE
-VE
-VE -VE
ventral view
Fig. 5. Removal of the ventral ectoderm results in cell death. (A)–(A′′′) The ventral ectoderm (VE) of the hindlimb was removed at HH20–21 and samples were then analyzedfor programmed cell death at 3, 6, 8, and 10 h after surgery. (A)–(A′) Some cell death is visible already at 3 h and 6 h in a layer of mesoderm just underlying the ventralectoderm. (A′′)–(A′′′) Programmed cell death increases in this zone and is strongly evident at 8 and 10 h after surgery. (B) No gross defects can be seen in skeletal pattern(control side on the left, VE removal on the right). (C)–(D) In addition, the pattern of muscles on the ventral side appears normal. (E)–(I′) Expression of Shh and FGF8 in theAER was analyzed at 3, 5, 6, and 16 h after surgery. Fgf8 expression shows that the AER is intact after VE removal. (E)–(F) Show dorsal views with a slight reduction in Shhexpression evident when the ventral ectoderm is removed (right side). (G)–(H) This is more apparent in a caudal view at 6 and 16 h after surgery (VE removed on right side).(I)–(I′) Ventral and dorsal view are shown at 16 h after surgery analyzed with just a Shh probe, demonstrating the slight reduction in Shh expression.
M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352348
The sensitivity of the limb mesoderm to ectoderm removal maybe specific in that the limb ectoderm provides survival signals, ornonspecific, in that exposure to amniotic fluid has a harmful effectthat is prevented with an ectodermal cover. In order to determineif the dorsal ectoderm provides a specific signal, we createdrecombinant limbs in which the limb ectoderm was removedand the limb mesoderm was encased either in a limb ectoderm(‘sham control’) or wrapped in ectoderm from the back of theembryo (Errick and Saunders, 1976). These recombinant limbswere grafted to the somite area of host embryos and the pattern ofcell death in these limbs was compared. In sham controls, TUNELanalysis 24 h after grafting showed a small amount of cell death atthe base (grafting site) but the mesoderm was largely intact(Fig. 6C). However, in high contrast, the limb bud covered inectoderm from the back showed massive cell death in themesodermal core (Fig. 6D), similar to our dorsal ectoderm andAER-removal experiments (Fig. 1E). Accordingly these limb budsfailed to develop and only formed at most a small cartilagerudiment whereas sham controls formed mostly normal limbs(data not shown). One possibility is that the high degree of celldeath in the recombinant limbs constructed with back ectodermcould be largely due to the lack of the AER which we know providessurvival signals. Thus, we repeated the recombination experimentsusing limb buds in which only the dorsal ectoderm was removedwhile the AER remained intact and wrapped them with limb
ectoderm (Fig. 6E and F) or with back ectoderm (Errick andSaunders, 1976) (Fig. 6G) from GFP transgenic embryos (McGrewet al., 2004). In these experiments the replacement of the dorsalectoderm with dorsal limb ectoderm completely prevented celldeath in the mesoderm (Fig. 6E) whereas replacement with backectoderm was still not able to prevent the extensive cell deathcaused by dorsal ectoderm removal (compare Fig. 6D–G). Theresults of these experiments confirm that the cause of cell deathin the mesoderm is the absence of ectoderm and not other effectsassociated with the surgery. Our results also indicate that theectoderm from the back of the embryo is not able to substitutefor the function of the dorsal ectoderm and therefore that simplyproviding an ectodermal covering from another location cannotsuffice.
Sections consecutive to those used for the TUNEL assay werehybridized for Wnt6, expressed throughout the limb and backectoderm, and for Wnt7a, expressed in the dorsal limb ecto-derm. Surprisingly, we found that the back ectoderm alsoexpressed Wnt7a although at a lower level than the dorsal limbectoderm (Fig. 6G). These results suggest that the specific signalgenerated by the limb ectoderm for survival of the mesoderm isneither Wnt6 nor Wnt7a because both genes are expressed bythe back ectoderm that lacks this function. They also provide anexplanation for the limbs with bi-dorsal distal structuresformed from recombinant limbs performed with back ectoderm
-prDE -dDE
-DE; replaced with limb ectoderm
-DE; replaced with back ectoderm
-limb ectoderm; replaced with limb ectoderm
-limb ectoderm;replaced with back ectoderm
+8h Wnt6 Wnt7a
+18h Wnt6 Wnt7a
+18h +18h
Fig. 6. Partial removals and ectoderm substitution experiments show that the dorsal ectoderm emits specific survival signals. (A) Limbs with the proximal half of the dorsalectoderm removed at 21HH and assayed at 18 h after surgery. (B) Limbs with the distal half of the dorsal ectoderm removed at 21HH, assayed at 18 h after surgery.(C) Control recombinant limbs constructed with operated limb bud and limb bud ectoderm, as depicted in the scheme, show little cell death at 24 h after grafting except atthe base where the graft abuts the flank of host embryo. (D) Recombinant limbs constructed with operated limb bud wrapped in ectoderm from the back of the embryo showmassive cell death in the mesoderm 24 h after grafting. (E, F) Control recombinant limbs in which wild-type operated limb buds were wrapped in GFP-transgenic limbectoderm and assessed at 8 and 18 h after surgery. (G) Recombinant limbs consisting of operated limb buds wrapped in back ectoderm still show cell death even when theAER was left intact. Note that the back ectoderm rapidly attaches to the denuded area while it integrates or detaches from the ventral side in which the endogenousectoderm remains. RNA in situ hybridization analysis in sequential sections for Wnt6 ((E′), (F′), and (G′)) and for Wnt7a ((E″), (F″), (G″)) shows the integrity of the graftedectoderm.
M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352 349

M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352350
and a stripe of dissociated and re-aggregated AER cells at the tip(Errick and Saunders, 1976).
Discussion
The analysis presented here provides compelling evidence thatthe non-AER limb ectoderm is required during limb bud develop-ment for the survival of the subjacent mesoderm. We show thatthe surgical removal of the dorsal ectoderm is followed by massivecell death in the subjacent dorsal mesoderm, drastic loss of dorsalsoft tissues and skeletal defects including reduction in size andfusion of elements. Comparable but less strong effects areobserved when the ventral ectoderm is removed (Fig. 5). Partialectodermal removals indicate that the severity of the defect isproportional to the extent of the area denuded of ectoderm and tothe proximity of the AER. Furthermore, experiments in which thelimb ectoderm is replaced by non-limb ectoderm show that theprotective factor(s) from the limb ectoderm is specific.
The limb ectoderm provides specific survival signals to the underlyingmesoderm
Our experiments clearly show that the non-AER ectoderm has asurvival role, as cell death in the subjacent mesoderm occurs whenit is removed. The extent of cell death depends on the portion ofthe ectoderm removed and also on the proximity of the AER,which may be able to substitute for the distal dorsal ectoderm. Itwas previously shown that the removal of patches of dorsalectoderm was not followed by cell death (Rowe et al., 1982) andtherefore it was considered that the survival effect was specific tothe AER. However, our study suggests that a considerable portionof the ectoderm needs to be removed in order to appreciate thesurvival role. In contrast, the nearby dorsal and ventral ectoderm isnot sufficient to prevent distal cell death after AER-removalsuggesting that either the survival signals from the dorsal andventral ectoderm are not potent enough or they involve a differentsignaling pathway than the survival signals from the AER.
The results from Gasseling and Saunders (1961) are similarto the results presented here. For example, they showed that whenleg buds are stripped of ectoderm the remaining mesodermsuffered massive apoptosis and failed to develop whereas thesame ectoderm-deprived leg buds formed terminal leg parts ifsupplied with an AER. As we have seen with the distal ectodermremoval experiments, these results underscore the strong survivalfunction of the AER and suggest that it is capable of substituting atleast partially for the dorsal and ventral ectoderm particularly inearly limb buds with reduced proximo-distal extension.
The molecular nature of the survival signal(s) from the ecto-derm remains to be determined but our recombination experi-ments show that they are specific to the limb ectoderm as theectoderm from the back of the embryo is unable to prevent thedeath of the subjacent mesoderm. Our experiments also rule outthe involvement of Wnt7a in the survival ectodermal function, asWnt7a is expressed by the back ectoderm (which lacks thisfunction) and furthermore, it is not expressed by the ventralectoderm (which has this function). In addition, the phenotypesof theWnt7a and Lmx1b-null mice, where a ventral transformationis observed rather than loss of the dorsal limb compartment, alsosupports this conclusion (Chen et al., 1998; Parr and McMahon,1995).
Role of the limb ectoderm in skeletal patterning and chondrogenesis
The removal of the dorsal limb ectoderm has been performedpreviously with different results (Martin and Lewis, 1986; Yang
and Niswander, 1995). Martin and Lewis (1986) used ultravioletirradiation to destroy the dorsal ectoderm and reported a reduc-tion in size of the skeleton but normal skeletal pattern in limbs.Their results are fully consistent with our work including the highfrequency of ulna/radius fusions in their irradiated limbs. How-ever, Yang and Niswander (1995) who used a surgical techniquereported defects in skeletal patterning: the loss of posteriorskeletal elements, mainly the ulna and digit 4. Because theseauthors focused on Shh expression we interpret that their interestin a complete removal of the dorsal ectoderm overlying the Shhexpression domain may have slightly damaged the AER at thislevel. This interpretation is supported by the frequent reproduc-tion of their phenotypes when we performed the dorsal ectodermremovals without the previous separation of the AER from thedorsal ectoderm. Also, it is supported by the similarity in pheno-types with those obtained after the removal of the posterior partof the AER (Rowe and Fallon, 1981).
A more general role of the ectoderm in blocking cartilagedifferentiation superficially and limiting chondrogenesis to thecore of the limb bud has been established (Cooper et al., 2011;Martin and Lewis, 1986; Solursh, 1984; Solursh et al., 1981). Forexample, the removal of the ectoderm from limb bud explantscaused the mesoderm to differentiate as cartilage (Kosher et al.,1979). Conversely, limb ectoderm can inhibit chondrogenesis oflimb mesoderm in culture. More recently this effect has beenshown to be mediated by Wnt/βCatenin signaling which blocksdifferentiation of the chondrogenic lineage (Geetha-Loganathanet al., 2010; Hartmann, 2006; ten Berge et al., 2008). Interestingly,this effect on blocking chondrogenesis is not shared by theectoderm that re-epithelializes the denuded ectoderm area, asthe skeletal elements of the operated limbs form much closer tothe ectoderm than normal.
Normal limb development and patterning requires constantfeedback between the ectoderm and the underlying mesoderm. Inorder to dissect the relationship between these two tissue types,ectoderm-removal experiments have been used to determine theconsequences of loss of the ectoderm on limb patterning. Theobserved defects can then be used to argue that the ectodermprovides signals to specify the structures that are perturbed ormissing. In the case of AER-removal experiments the truncated limbskeleton observed supported the notion that the AER was notproviding distalizing signals directly but rather maintained thedistalizing function of the proposed underlying progress zone(Summerbell et al., 1973). Dudley et al., 2002, subsequently proposedthat the truncations could be explained by the loss or reduction inthe progenitors that give rise to the different proximal–distal skeletalelements. Similarly, we find that removal of the non-AER dorsalectoderm causes the underlying mesoderm to undergo cell death.This cell death likely removes skeletal progenitors for all the skeletalelements resulting in limbs that are smaller and shorter than normal(see Fig. 2B and C). Cell death can be seen along the entire proximal–distal extent of the limb bud 4–10 h after surgery, however, the celldeath pattern 20 h after surgery is particularly concentrated in thedorsal proximal region (Fig. 1D). This cell death may particularlyremove zeugopod progenitors and therefore account for the moredramatic reduction in these specific elements. Removal of the dorsalectoderm in the hindlimb bud at HH20 also causes reduction in thezeugopod and autopod length and fusion of the tarsal elements (datanot shown). Without knowing the effect of dorsal ectoderm removalon cell survival, one might conclude from these experiments that thedorsal ectoderm provides signals that specify the zeugopod elementor pattern it into two elements, however, given the location of thecell death and likely fate of these cells (Dudley et al., 2002; Vargessonet al., 1997), this is unlikely to be the case. Therefore, it seemsreasonable to propose that the skeletal patterning defects observedafter removal of the dorsal ectoderm in the chicken embryo are

M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352 351
fundamentally the consequence of the massive cell death in thedorsal limb compartment rather than the loss of a specification signalfrom the dorsal ectoderm.
The Wnt7a null mouse shows a variable loss of posteriorelements most frequently the ulna and digit 5. The variability inthe loss of posterior skeletal elements has been correlated withthe degree of Shh downregulation in this mutant (Parr andMcMahon, 1995). Thus, another possibility is that the variabilityin Shh recovery after the deprival of the ectoderm in our experi-ments could also contribute to the variability of the skeletalpatterning and the loss of digit 4.
Proximal cell death may also be removing other cells importantfor proper limb formation. Limb muscle cells migrate in dorsal andventral streams from the somites (Christ and Brand-Saberi, 2002;Mok and Sweetman, 2011; Vasyutina and Birchmeier, 2006) andgiven our observation that the dorsal muscle compartment fails toform when the dorsal ectoderm is removed, the dorsal stream hasbeen very likely affected. One possibility is that muscle progenitorsare specifically dependent on survival signals from the dorsalectoderm. Another possibility is that the cell death or failure tospecify other soft tissue progenitors perturbs muscle progenitormigration, patterning, and/or differentiation. In any case, it is clearthat any conclusions drawn about the role of the dorsal ectodermduring limb patterning using the type of experiment shown here,needs to take into consideration the critical survival signalprovided by the ectoderm.
Non-AER ectoderm and Shh
It is well known that ectodermal signals, predominantly fromthe dorsal ectoderm, are required for the proper expression of Shh(Parr and McMahon, 1995; Yang and Niswander, 1995). Our resultsare fully consistent with these previous studies in that there is aclose ectoderm/Shh expression relationship. Indeed the timing ofShh down-regulation after removal of the AER (Niswander et al.,1994) and after removal of the dorsal or ventral ectoderm is verysimilar.
Since the downregulation in Shh occurs very rapidly after theremoval of the ectoderm, and the role of Shh in regulating celldeath in the limb bud is well-documented (Bastida et al., 2009;Fogel et al., 2012; Ros et al., 2003; Sanz-Ezquerro and Tickle,2003), we have also considered the possibility that the mesoder-mal cell death was mediated by the loss of Shh. However, theobservation that the cell death pattern in the absence of Shh isvery different from the cell death pattern after removal of thedorsal ectoderm, does not support this hypothesis. Indeed, thepredominantly anterior and central pattern of cell death in thechick ozd mutant limb bud which lacks Shh expression, and in Shhnull mouse limb buds (Fogel et al., 2012; Ros et al., 2003) is in starkcontrast with the superficial band of cell death that follows theremoval of the dorsal ectoderm. Nevertheless, Shh may contributeto the survival and specification of muscle progenitors (Hu et al.,2012).
Despite severe reduction in Shh expression, anterior-posteriorskeletal patterning is mostly normal, except for the above-described alterations that we interpret are predominantly due tothe loss of muscle and skeletal precursors by cell death. Thus,normal Shh expression transiently during early developmentmight suffice to generate normal anterior-posterior patterning,supporting studies with similar conclusions in the mouse (Zhu andMackem, 2011; Zhu et al., 2008).
Proximal mesoderm cell death in AER–FGF mutants
When the function of multiple AER–FGFs is disrupted, or evenwhen the function of just Fgf8 is eliminated in the hindlimb
(Fig. 1SB), cell death is located in the dorsal proximal mesodermat distance from the AER, in a region where some cell death occursnormally (Boulet et al., 2004; Mariani et al., 2008; Moon andCapecchi, 2000; Sun et al., 2002). A simple explanation for whycell death occurs at a location distant from the AER is thatmesoderm survival is sensitive to the potency of AER–FGFs andwhen this signaling is reduced, cells nearby are sufficientlyprotected, while cells at a distance are now subject to cell death(Sun et al., 2002). Interestingly, we have noticed that regardless ofwhich Cre transgene is used to reduce AER–FGF signaling theproximal dorsal ectoderm overlying the cells dying in the meso-derm undergoes cell death in the AER–FGF mutants (Fig. 1SB and(Boulet et al., 2004; Mariani et al., 2008; Sun et al., 2002)). Thus,given the results of this current study, one possibility is that theproximal dorsal ectoderm is particularly sensitive to FGF survivalsignals and that loss of these cells subsequently causes the massivecell death in the underlying proximal mesoderm. According to thisinterpretation, the ventral ectoderm might be less affected by areduction in AER–FGF signals because of its reduced extensioncompared to the dorsal ectoderm.
Given the pattern of cell death in the AER–FGF mutants, AERremoval might also be expected to result in proximal ectodermand mesodermal cell death, however this has never been reported.Indeed, we find that although, as reported, AER-removal at HH20–21 results in a rapid wave of cell death distally (Dudley et al., 2002;Rowe et al., 1982), to our surprise, there is no associated proximalcell death even at time points long after the surgery (up to 36 hafter AER-removal, Fig. 1SC). This result correlates with thecompletely normal morphology of the proximal skeletal elementsafter AER removal at this early stage (Rowe et al., 1982; Rowe andFallon, 1981; Summerbell et al., 1973). The reason for the differ-ence in cell death pattern when comparing surgical vs. geneticmanipulation of the AER is not clear. Species-specific differences,could provide an explanation but also, the two experiments differbecause surgical removal of the AER results in a sudden loss of allsignaling from the AER (other factors are lost besides the AER–FGFs) as well as in dramatic changes in the distal mesoderm due todistal cell death. Additional studies will be needed to more fullyunderstand how AER signaling impacts survival of both themesoderm and ectoderm.
Conclusions
Several specific roles for the limb ectoderm are well establishedincluding an anti-chondrogenic effect. In addition specific roles forthe dorsal or ventral ectoderm, through the production of Wnt7aand En1, respectively, are also well-known to control of D–Vpatterning. Here we identify a new and critical function of thelimb ectoderm: its requirement for the survival of the subjacentlimb mesoderm employing a signal that is not shared by non-limbectoderm. Future work would be needed to identify the survivalsignal(s). This could be done by identifying signaling moleculespresent in the dorsal ectoderm but not found expressed in backectoderm.
Acknowledgments
We are very grateful to Dr. Gail Martin for the insightfuldiscussions that helped initiate this study and for her criticalcomments on the manuscript. In addition, we thank Marisa Juncofor excellent technical assistance and The Roslin Institute,University of Edinburg for providing transgenic animals. This workwas supported by grant BFU2011-24972 from the Spanish Ministryof Science (M.R.) and by the University of Southern California (F.M.).

M. Fernandez-Teran et al. / Developmental Biology 381 (2013) 341–352352
Appendix A. Supporting information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.ydbio.2013.06.032.
References
Bastida, M.F., Sheth, R., Ros, M.A., 2009. A BMP-Shh negative-feedback loop restrictsShh expression during limb development. Development 136, 3779–3789.
Benazet, J.D., Zeller, R., 2009. Vertebrate limb development: moving from classicalmorphogen gradients to an integrated 4-dimensional patterning system. ColdSpring Harbor Symp. Quant. Biol. 1, a001339.
Boulet, A.M., Moon, A.M., Arenkiel, B.R., Capecchi, M.R., 2004. The roles of Fgf4 andFgf8 in limb bud initiation and outgrowth. Dev. Biol. 273, 361–372.
Chen, H., Lun, Y., Ovchinnikov, D., Kokubo, H., Oberg, K.C., Pepicelli, C.V., Gan, L.,Lee, B., Johnson, R.L., 1998. Limb and kidney defects in Lmx1b mutant micesuggest an involvement of LMX1B in human nail patella syndrome. Nat. Genet.19, 51–55.
Christ, B., Brand-Saberi, B., 2002. Limb muscle development. Int. J. Dev. Biol. 46,905–914.
Colvin, J.S., Green, R.P., Schmahl, J., Capel, B., Ornitz, D.M., 2001. Male-to-female sexreversal in mice lacking fibroblast growth factor 9. Cell 104, 875–889.
Cooper, K.L., Hu, J.K., ten Berge, D., Fernandez-Teran, M., Ros, M.A., Tabin, C.J., 2011.Initiation of proximal–distal patterning in the vertebrate limb by signals andgrowth. Science 332, 1083–1086.
Cygan, J.A., Johnson, R.L., McMahon, A.P., 1997. Novel regulatory interactionsrevealed by studies of murine limb pattern in Wnt-7a and En-1 mutants.Development 124, 5021–5032.
Dudley, A.T., Ros, M.A., Tabin, C.J., 2002. A re-examination of proximodistalpatterning during vertebrate limb development. Nature 418, 539–544.
Errick, J.E., Saunders Jr., J.W., 1976. Limb outgrowth in the chick embryo induced bydissociated and reaggregated cells of the apical ectodermal ridge. Dev. Biol. 50,26–34.
Fallon, J.F., Lopez, A., Ros, M.A., Savage, M.P., Olwin, B.B., Simandl, B.K., 1994. FGF-2:apical ectodermal ridge growth signal for chick limb development. Science 264,104–107.
Fernandez-Teran, M., Ros, M.A., 2008. The Apical Ectodermal Ridge: morphologicalaspects and signaling pathways. Int. J. Dev. Biol. 52, 857–871.
Fernandez-Teran, M.A., Hinchliffe, J.R., Ros, M.A., 2006. Birth and death of cells inlimb development: a mapping study. Dev. Dyn. 235, 2521–2537.
Fogel, J.L., Thein, T.Z., Mariani, F.V., 2012. Use of LysoTracker to detect programmedcell death in embryos and differentiating embryonic stem cells. J. Vis. Exp..
Gasseling, M.T., Saunders Jr., J.W., 1961. Effects of the apical ectodermal ridge ongrowth of the versene-stripped chick limb bud. Dev. Biol. 3, 1–25.
Geetha-Loganathan, P., Nimmagadda, S., Christ, B., Huang, R., Scaal, M., 2010.Ectodermal Wnt6 is an early negative regulator of limb chondrogenesis inthe chicken embryo. BMC Dev. Biol. 10, 32.
Hamburger, V., Hamilton, H.L., 1992. A series of normal stages in the developmentof the chick embryo. 1951. Dev. Dyn. 195, 231–272.
Hartmann, C., 2006. AWnt canon orchestrating osteoblastogenesis. Trends Cell Biol.16, 151–158.
Hu, J.K., McGlinn, E., Harfe, B.D., Kardon, G., Tabin, C.J., 2012. Autonomous andnonautonomous roles of Hedgehog signaling in regulating limb muscle forma-tion. Genes Dev. 26, 2088–2102.
Kosher, R.A., Savage, M.P., Chan, S.C., 1979. In vitro studies on the morphogenesisand differentiation of the mesoderm subjacent to the apical ectodermal ridge ofthe embryonic chick limb-bud. J. Embryol. Exp. Morphol. 50, 75–97.
Lewandoski, M., Sun, X., Martin, G.R., 2000. Fgf8 signalling from the AER is essentialfor normal limb development. Nat. Genet. 26, 460–463.
Loomis, C.A., Kimmel, R.A., Tong, C.X., Michaud, J., Joyner, A.L., 1998. Analysis of thegenetic pathway leading to formation of ectopic apical ectodermal ridges inmouse Engrailed-1 mutant limbs. Development 125, 1137–1148.
Mariani, F.V., Ahn, C.P., Martin, G.R., 2008. Genetic evidence that FGFs have aninstructive role in limb proximal–distal patterning. Nature 453, 401–405.
Mariani, F.V., Martin, G.R., 2003. Deciphering skeletal patterning: clues from thelimb. Nature 423, 319–325.
Martin, G.R., 1998. The roles of FGFs in the early development of vertebrate limbs.Genes Dev. 12, 1571–1586.
Martin, P., Lewis, J., 1986. Normal development of the skeleton in chick limb budsdevoid of dorsal ectoderm. Dev. Biol. 118, 233–246.
McGrew, M.J., Sherman, A., Ellard, F.M., Lillico, S.G., Gilhooley, H.J., Kingsman, A.J.,Mitrophanous, K.A., Sang, H., 2004. Efficient production of germline transgenicchickens using lentiviral vectors. EMBO Rep. 5, 728–733.
Mok, G.F., Sweetman, D., 2011. Many routes to the same destination: lessons fromskeletal muscle development. Reproduction 141, 301–312.
Moon, A.M., Boulet, A.M., Capecchi, M.R., 2000. Normal limb development inconditional mutants of Fgf4. Development 127, 989–996.
Moon, A.M., Capecchi, M.R., 2000. Fgf8 is required for outgrowth and patterning ofthe limbs. Nat. Genet. 26, 455–459.
Nieto, M.A., Patel, K., Wilkinson, D.G., 1996. In situ analysis of chick embryos inwhole mount and tissue sections. In: Bronner-Fraser, M. (Ed.), Methods in CellBiology. Academic Press, New York, pp. 219–235.
Niswander, L., Jeffrey, S., Martin, G.R., Tickle, C., 1994. A positive feedback loopcoordinates growth and patterning in the vertebrate limb. Nature 371, 609–612.
Parr, B.A., McMahon, A.P., 1995. Dorsalizing signal Wnt-7a required for normalpolarity of D–V and A–P axes of mouse limb. Nature 374, 350–353.
Riddle, R.D., Ensini, M., Nelson, C., Tsuchida, T., Jessell, T.M., Tabin, C., 1995.Induction of the LIM homeobox gene Lmx1 by WNT7a establishes dorsoventralpattern in the vertebrate limb. Cell 83, 631–640.
Roelink, H., Augsburger, A., Heemskerk, J., Korzh, V., Norlin, S., Ruiz i Altaba, A.,Tanabe, Y., Placzek, M., Edlund, T., Jessell, T.M., et al., 1994. Floor plate andmotor neuron induction by vhh-1, a vertebrate homolog of hedgehog expressedby the notochord. Cell 76, 761–775.
Ros, M.A., Dahn, R.D., Fernandez-Teran, M., Rashka, K., Caruccio, N.C., Hasso, S.M.,Bitgood, J.J., Lancman, J.J., Fallon, J.F., 2003. The chick oligozeugodactyly (ozd)mutant lacks sonic hedgehog function in the limb. Development 130, 527–537.
Ros, M.A., Simandl, B.K., Clark, A.W., Fallon, J.F., 2000. Methods for manipulating thechick limb bud to study gene expression, tissue interactions, and patterning.Methods Mol. Biol. 137, 245–266.
Rowe, D.A., Cairns, J.M., Fallon, J.F., 1982. Spatial and temporal patterns of cell deathin limb bud mesoderm after apical ectodermal ridge removal. Dev. Biol. 93,83–91.
Rowe, D.A., Fallon, J.F., 1981. The effect of removing posterior apical ectodermalridge of the chick wing and leg on pattern formation. J. Embryol. Exp. Morphol.65, 309–325, Suppl..
Sanz-Ezquerro, J.J., Tickle, C., 2003. Fgf signaling controls the number of phalangesand tip formation in developing digits. Curr. Biol. 13, 1830–1836.
Saunders Jr., J.W., 1948. The proximo-distal sequence of origin of the parts of thechick wing and the role of the ectoderm. J. Exp. Zool. 108, 363–403.
Solursh, M., 1984. Ectoderm as a determinant of early tissue pattern in the limbbud. Cell Differ. 15, 17–24.
Solursh, M., Singley, C.T., Reiter, R.S., 1981. The influence of epithelia on cartilageand loose connective tissue formation by limb mesenchyme cultures. Dev. Biol.86, 471–482.
Summerbell, D., Lewis, J.H., Wolpert, L., 1973. Positional information in chick limbmorphogenesis. Nature 244, 492–496.
Sun, X., Lewandoski, M., Meyers, E.N., Liu, Y.H., Maxson Jr., R.E., Martin, G.R., 2000.Conditional inactivation of Fgf4 reveals complexity of signalling during limbbud development. Nat. Genet. 25, 83–86.
Sun, X., Mariani, F.V., Martin, G.R., 2002. Functions of FGF signalling from the apicalectodermal ridge in limb development. Nature 418, 501–508.
ten Berge, D., Brugmann, S.A., Helms, J.A., Nusse, R., 2008. Wnt and FGF signalsinteract to coordinate growth with cell fate specification during limb develop-ment. Development 135, 3247–3257.
Vargesson, N., Clarke, J.D., Vincent, K., Coles, C., Wolpert, L., Tickle, C., 1997. Cell fatein the chick limb bud and relationship to gene expression. Development 124,1909–1918.
Vasyutina, E., Birchmeier, C., 2006. The development of migrating muscle precursorcells. Anat. Embryol. (Berl.) 211 (1), 37–41.
Xu, J., Liu, Z., Ornitz, D.M., 2000. Temporal and spatial gradients of Fgf8 and Fgf17regulate proliferation and differentiation of midline cerebellar structures.Development 127, 1833–1843.
Yang, Y., Niswander, L., 1995. Interaction between the signaling molecules WNT7aand SHH during vertebrate limb development: dorsal signals regulate ante-roposterior patterning. Cell 80, 939–947.
Zhu, J., Mackem, S., 2011. Analysis of mutants with altered shh activity andposterior digit loss supports a biphasic model for shh function as a morphogenand mitogen. Dev. Dyn. 240, 1303–1310.
Zhu, J., Nakamura, E., Nguyen, M.T., Bao, X., Akiyama, H., Mackem, S., 2008.Uncoupling Sonic hedgehog control of pattern and expansion of the developinglimb bud. Dev. Cell 14, 624–632.