Evidence for base-pairing between mammalian U2 and...
12
Evidence for base-pairing between mammalian U2 and U6 small nuclear ribonucleoprotein particles Thomas-Peter Hausner,^'^ Linda M. Giglio,*'^ and Alan M. Weiner* 'Department of Molecular Biophysics and Biochemistry, Yale University School of Medicine, New Haven, Connecticut 06510-8024 USA Intramolecular and intermolecular snRNA cross-links were generated by irradiating HeLa nuclear extract with 365 nm light in the presence of the psoralen derivative AMT. After deproteinization, cross-linked RNAs were resolved by gel electrophoresis and identified as anomalously migrating species by Northern blotting. In addition to the U4/U6 snRNA cross-link, we detected an intermolecular U2/U6 cross-link, as well as several apparently intramolecular Ul, U2, and U5 cross-links. Photoreversal of the U2/U6 cross-link with 254 nm irradiation released stoichiometric amounts of U2 and U6 snRNA. To localize the U2/U6 cross-link, the 3' end of U2 in the purified U2/U6 complex was labeled selectively using a novel oligonucleotide "splint" technique. The labeled U2/U6 complex was then subjected to rapid enzymatic RNA sequencing or to targeted digestion of the U2 and U6 components of the complex by RNase H and a panel of complementary oligonucleotides. The U2/U6 cross-link is located upstream of nucleotide 15 in U2 and downstream of nucleotide 85 in U6, suggesting that the phylogenetically conserved base-pairing between these regions (6 consecutive base pairs in human, Drosophila melanogaster, and Caenorhabditis elegans, 7 in Schizosaccharomyces pombe and Jiypanasoma brucei, 8 in Pisutn sativum, 11 in Saccharomyces cerevisiae) is significant. [Key Words: snRNA; snRNP; mRNA splicing; psoralen; cross-linking] Received August 1, 1990; revised version accepted October 1, 1990. In eukaryotes, introns are removed from nuclear mRNA precursors by a multicomponent ribonucleoprotein complex called the spliceosome (for review, see Steitz et al. 1988). The fully assembled spliceosome consists of four separate small ribonucleoprotein particles (the Ul, U2, U5, and U4/U6 snRNPs) and numerous auxiliary protein factors. The Ul, U2, and U5 snRNPs each con- tain a single small nuclear RNA (snRNA), whereas the U4/U6 snRNP consists of two snRNAs base-paired to each other (Bringmann et al. 1984; Hashimoto and Steitz 1984; Rinke et al. 1985; Brow and Guthrie 1988). All four spliceosomal snRNPs share a core of six common proteins; other proteins are unique to a single kind of snRNP (Liihrmann 1988). During spliceosome assembly, the Ul snRNP base- pairs with the 5'-splice site (Zhuang and Weiner 1986; Legrain et al. 1988; Ruby and Abelson 1988; Seraphin et al. 1988; Siliciano and Guthrie 1988; Seraphin and Ros- bash 1989) and the U2 snRNP interacts with the 3' re- gion of the intron (Black et al. 1985; Chabot et al. 1985; Ruskin et al. 1988) initially through the protein factor U2AF (Ruskin et al. 1988) and later by base-pairing with Present addresses: ^Bayer AG, Zentrale Forschung und Entwicklung, 5090-Leverkusen, Germany; ^Department of Cellular and Molecular Bi- ology, Bristol Meyers-Squibb, Wallingford, Connecticut 06492 USA. the branch site (Parker et al. 1987; Nelson and Green 1989; Wu and Manley 1989; Zhuang and Weiner 1989; Zhuang et al. 1989). In contrast, only protein factors are required for initial recognition of the 3'-splice site (Gerke and Steitz 1986; Tazi et al. 1986; Ruskin et al. 1988; Swanson and Dreyfuss 1988) and the polypyrimi- dine tract (Ruskin et al. 1988; Garcia-Blanco et al. 1989). The U4/U6 and U5 snRNPs are known to associate with each other before assembling into the spliceosome (Cheng and Abelson 1987; Konarska and Sharp 1987; Lossky et al. 1987; Lamond et al. 1988), but the role of these three snRNAs in mRNA splicing remains elusive. The spliceosome undergoes dynamic changes in con- formation and composition during mRNA splicing. The strongest evidence is that extensive base-pairing be- tween U4 and U6 snRNA in the U4/U6 snRNP is labi- lized or lost when the first phosphoester bond transfer occurs (Pikielny et al. 1986; Cheng and Abelson 1987; Lamond et al. 1988; Blencowe et al. 1989). Thus, it is possible that base pairs between spliceosomal snRNAs may be formed or broken in the course of assembly, ca- talysis, and disassembly of the spliceosome. In an effort to detect such base-pairing interactions, we have in- duced intermolecular and intramolecular RNA cross- links in HeLa nuclear extract by 365 nm irradiation in the presence of the psoralen derivative (4'-amino- methyl-4,5',8-trimethylpsoralen (AMT; for review, see 2146 GENES & DEVELOPMENT 4:2146-2156 © 1990 by Cold Spring Harbor Laboratory Press ISSN 0890-9369/90 $1.00 Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.org Downloaded from
Transcript of Evidence for base-pairing between mammalian U2 and...

Evidence for base-pairing between mammalian U2 and U6 small nuclear ribonucleoprotein particles Thomas-Peter Hausner,^'^ Linda M. Giglio,*'^ and Alan M. Weiner*
'Department of Molecular Biophysics and Biochemistry, Yale University School of Medicine, New Haven, Connecticut 06510-8024 USA
Intramolecular and intermolecular snRNA cross-links were generated by irradiating HeLa nuclear extract with 365 nm light in the presence of the psoralen derivative AMT. After deproteinization, cross-linked RNAs were resolved by gel electrophoresis and identified as anomalously migrating species by Northern blotting. In addition to the U4/U6 snRNA cross-link, we detected an intermolecular U2/U6 cross-link, as well as several apparently intramolecular Ul, U2, and U5 cross-links. Photoreversal of the U2/U6 cross-link with 254 nm irradiation released stoichiometric amounts of U2 and U6 snRNA. To localize the U2/U6 cross-link, the 3' end of U2 in the purified U2/U6 complex was labeled selectively using a novel oligonucleotide "splint" technique. The labeled U2/U6 complex was then subjected to rapid enzymatic RNA sequencing or to targeted digestion of the U2 and U6 components of the complex by RNase H and a panel of complementary oligonucleotides. The U2/U6 cross-link is located upstream of nucleotide 15 in U2 and downstream of nucleotide 85 in U6, suggesting that the phylogenetically conserved base-pairing between these regions (6 consecutive base pairs in human, Drosophila melanogaster, and Caenorhabditis elegans, 7 in Schizosaccharomyces pombe and Jiypanasoma brucei, 8 in Pisutn sativum, 11 in Saccharomyces cerevisiae) is significant.
[Key Words: snRNA; snRNP; mRNA splicing; psoralen; cross-linking]
Received August 1, 1990; revised version accepted October 1, 1990.
In eukaryotes, introns are removed from nuclear mRNA precursors by a multicomponent ribonucleoprotein complex called the spliceosome (for review, see Steitz et al. 1988). The fully assembled spliceosome consists of four separate small ribonucleoprotein particles (the Ul, U2, U5, and U4/U6 snRNPs) and numerous auxiliary protein factors. The Ul, U2, and U5 snRNPs each contain a single small nuclear RNA (snRNA), whereas the U4/U6 snRNP consists of two snRNAs base-paired to each other (Bringmann et al. 1984; Hashimoto and Steitz 1984; Rinke et al. 1985; Brow and Guthrie 1988). All four spliceosomal snRNPs share a core of six common proteins; other proteins are unique to a single kind of snRNP (Liihrmann 1988).
During spliceosome assembly, the Ul snRNP base-pairs with the 5'-splice site (Zhuang and Weiner 1986; Legrain et al. 1988; Ruby and Abelson 1988; Seraphin et al. 1988; Siliciano and Guthrie 1988; Seraphin and Ros-bash 1989) and the U2 snRNP interacts with the 3' region of the intron (Black et al. 1985; Chabot et al. 1985; Ruskin et al. 1988) initially through the protein factor U2AF (Ruskin et al. 1988) and later by base-pairing with
Present addresses: ^Bayer AG, Zentrale Forschung und Entwicklung, 5090-Leverkusen, Germany; ^Department of Cellular and Molecular Biology, Bristol Meyers-Squibb, Wallingford, Connecticut 06492 USA.
the branch site (Parker et al. 1987; Nelson and Green 1989; Wu and Manley 1989; Zhuang and Weiner 1989; Zhuang et al. 1989). In contrast, only protein factors are required for initial recognition of the 3'-splice site (Gerke and Steitz 1986; Tazi et al. 1986; Ruskin et al. 1988; Swanson and Dreyfuss 1988) and the polypyrimi-dine tract (Ruskin et al. 1988; Garcia-Blanco et al. 1989). The U4/U6 and U5 snRNPs are known to associate with each other before assembling into the spliceosome (Cheng and Abelson 1987; Konarska and Sharp 1987; Lossky et al. 1987; Lamond et al. 1988), but the role of these three snRNAs in mRNA splicing remains elusive.
The spliceosome undergoes dynamic changes in conformation and composition during mRNA splicing. The strongest evidence is that extensive base-pairing between U4 and U6 snRNA in the U4/U6 snRNP is labi-lized or lost when the first phosphoester bond transfer occurs (Pikielny et al. 1986; Cheng and Abelson 1987; Lamond et al. 1988; Blencowe et al. 1989). Thus, it is possible that base pairs between spliceosomal snRNAs may be formed or broken in the course of assembly, catalysis, and disassembly of the spliceosome. In an effort to detect such base-pairing interactions, we have induced intermolecular and intramolecular RNA crosslinks in HeLa nuclear extract by 365 nm irradiation in the presence of the psoralen derivative (4'-amino-methyl-4,5',8-trimethylpsoralen (AMT; for review, see
2146 GENES & DEVELOPMENT 4:2146-2156 © 1990 by Cold Spring Harbor Laboratory Press ISSN 0890-9369/90 $1.00
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Base-pairing between U2 and U6 snRNA
Cimino et al. 1985). Psoralen cross-linking appears to be specific for base-paired regions of RNA and DNA and has been used previously to demonstrate base-pairing between Ul and U2 snRNA and nuclear RNA (Calvet and Pederson 1981; Calvet et al. 1982), between U4 and U6 snRNA (Rinke et al. 1985), within Ul snRNA (Rinke et al. 1985; Calvet and Myers 1987), and probably between U3 and rRNA precursor sequences (Maser and Calvet 1989; Stroke and Weiner 1989). Here we report the identification and characterization of a novel, phylogenetic-ally conserved base-pairing interaction between the 5' end of U2 and the 3 ' end of U6 snRNA.
Results
Initial characterization of the covalent U2/U6 crosslink
We generated intermolecular and intramolecular RNA cross-links in HeLa nuclear extract by 365 nm irradiation in the presence of the psoralen derivative (AMT). After cross-linking, the extract was deproteinized and the snRNAs were identified by Northern blotting. Cross-linking in the presence of AMT generated two slowly migrating bands of comparable abundance, labeled U4/U6 and U2/U6, which were not observed after irradiation in the absence of AMT (Fig. lA, lanes 1 and 2). We attempted to increase the yield of U2/U6 (estimated to be < 5 % of total U2) by incubating a human 3-globin mRNA precursor in nuclear extract pretreated with EDTA to block cleavage at the 5'-splice site and cause accumulation of spliceosomes (Abmayr et al.
A B probed for: Ul U2 U4 U5 U6
1
U4/U6 U2/U6
U 2 t
uif| U 4 *
usi U 6 |
2 3 4 ^
*IS
J I U U C U lUr : VJi v."^
U4/U6 ~ ~ U2/U6 - -
^ ai
U2 5.i|f
U 4 ^ - -
U5 U6 "1
Figure 1. Northern blot of cross-linked snRNAs generated by psoralen cross-linking of nuclear extract. [A] Irradiation of nuclear extract in the absence (lane 1] or presence (lane 2) of AMT psoralen; irradiation in the presence of AMT psoralen after pre-treatment of nuclear extract with 4 mM EDTA for 30 min at 0°C followed by incubation under splicing conditions without (lane 3) or with (lane 4) 10 ng Hp6 mRNA precursor (Abmayr et al. 1988). This blot was probed for Ul, U2, U4, U5, and U6 simultaneously. [B] Irradiation in the presence of AMT psoralen but without mRNA precursor. Identical lanes were probed separately for Ul, U2, U4, U5, or U6 snRNA to identify cross-linked species. A labeled HpaU digest of pBR322 was used to facilitate alignment.
1988); however, the yield of U2/U6 was unaffected (Fig. lA, cf. lanes 3 and 4 with lane 2).
We tentatively identified the two major slowly migrating bands as cross-linked U4/U6 and U2/U6 complexes by probing identical Northern blots of cross-linked RNA for each spliceosomal snRNA separately (Fig. IB). The slowest migrating band that reacts with the U4 and U6 probes almost certainly corresponds to the U4/U6 complex (Bringmann et al. 1984; Hashimoto and Steitz 1984; Brow and Guthrie 1988), which is known to be susceptible to psoralen cross-linking (Rinke et al. 1985). The faster migrating band reacted with U2 and U6 probes. We also detected a band, migrating slightly above Ul , which probably corresponds to the intramolecular U l cross-link described previously (Rinke et al. 1985; Calvet and Myers 1987). Separation of the cross-linked RNAs on higher percentage polyacrylamide gels revealed additional cross-linked products that probably represent intramolecular U l , U2, and U5 crosslinks (data not shown). (In Fig. IB, the U4 probe was slightly contaminated with the U2 probe.)
That the U2/U6 cross-link is as abundant as the previously characterized U4/U6 cross-link (Rinke et al. 1985) can be seen when the blots are probed for all five snRNAs simultaneously (Fig. lA, lanes 2-4) . Although a third slowly migrating band can be seen between U4/U6 and U2/U6 (Fig. IB, lane U6), this species is at least 10-fold less abundant than U2/U6 and would not be detected in our subsequent analysis of the U2/U6 cross-link. The third band is seen only when a blot is probed with oligonucleotide U69o_io5 and overexposed to compensate for the inefficiency of this probe, which reacts well with uncross-linked U6 but very poorly with U2/U6 (Fig. IB, lane U6) because the oligonucleotide lies close to the U2/U6 cross-link (see below).
We then used targeted digestion of U2 to demonstrate that the U2/U6 band is an intermolecular cross-link between U2 and U6, not comigrating intramolecularly cross-linked U2 and U6 RNAs (Fig. 2). Cross-linked RNA was digested with RNase H and oligonucleotide U2i54_i72 (complementary to U2 nucleotides 154-172), and a Northern blot of the digestion products was probed for U2 or U6 RNA. As expected for a covalent intermolecular U2/U6 cross-link, RNase H cleavage of U2 increased the mobility of the U2/U6 complex to the same extent regardless of whether the Northern blot was probed for U2 (Fig. 2, cf. lanes 1 and 2) or U6 (cf. lanes 3 and 4). Targeted digestion of U2 did not affect the mobility of the U4/U6 complex. Similar results were obtained when U6 was digested with RNase H and a complementary oligonucleotide (data not shown).
To determine the stoichiometry of U2 and U6 in the U2/U6 complex, we analyzed the cross-linked complex from cells that had been metabolically labeled with P^PJphosphate for a generation (Fig. 3). Nuclear extract from labeled cells was cross-linked, and spliceosomal snRNPs were immunoprecipitated with anti-Sm antibodies. The U2/U6 and U4/U6 complexes are precipitated efficiently under these conditions (data not shown). The immunoprecipitated RNAs were separated
GENES & DEVELOPMENT 2147
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Hausner and Weiner
probed for:U2
U2/U6 ,
U6 U4/U6 * i ' U2/U6 m
U2
%
t 12
U6l
Figure 2. RNase H digestion of the apparent U2/U6 complex confirms that U2 and U6 are covalently cross-linked. Cross-linked RNAs were annealed with a DNA oligonucleotide complementary to U2i54_i72 and incubated with (lanes 2 and 4) or without (lanes 1 and 3) RNase H. RNAs were identified by Northern blotting with a U279_98 probe (lanes 1 and 2) or a U643_6o probe (lanes 3 and 4).
by two-dimensional denaturing polyacrylamide gel electrophoresis (5% first dimension, 9% second dimension) with or without photoreversal of the cross-links between dimensions by 254 nm light (Rabin and Crothers 1979). Two pairs of discrete spots can be seen (U4 and U6, U2 and U6) that comigrate in the first dimension but separate in the second dimension of the photore-versed gel (Fig. 3, left); without photoreversal, the U4/U6 and U2/U6 complexes are obscured by background labeling (Fig. 3, right). The snRNAs were cut out of the gel, and Cerenkov radiation was measured with a scintillation counter. When the counts per minute were corrected for background (using a gel fragment adjacent to each measured spot) and for the size of each snRNA, the ratio of U2 to U6 was 1.1 and the ratio of U4 to U6 was 1.0. We conclude that the U2/U6 complex contains equimolar amounts of U2 and U6.
Although we have not strictly excluded the existence of an additional undetected RNA component in the U2/U6 complex, this seems improbable because (1) the U2/U6 complex migrates faster than U4/U6 in all gel systems tested, (2) inefficient cross-linking is unlikely to produce a ternary cross-link, and (3) no evidence for a third RNA was found in experiments designed to localize the U2/U6 cross-link (see Figs. 5-7). We note, however, that the U2/U6 band was occasionally diffuse (cf. Fig. 2 with Fig. 1), perhaps reflecting minor variations in the cross-linking protocol, gel electrophoresis, or the precise site of U2/U6 cross-linking.
U2/U6 complex using RNA ligase and ^^P-labeled pCp, perhaps because psoralen monoadducts or the conformation of the U2/U6 complex rendered both 3 ' ends inaccessible. Nor were we able to identify the site of cross-linking by primer extension with reverse transcriptase, perhaps because psoralen monoadducts blocked efficient extension. We therefore developed a new technique for selectively labeling the 3 ' end of either U2 or U6 RNA in total deproteinized nuclear extract. An snRNA/DNA hybrid with a recessed RNA 3 ' end is created by annealing an appropriate oligonucleotide to the 3 ' end of the snRNA (see diagram in Materials and methods). Extension of the RNA 3 ' end with T7 DNA polymerase and a labeled deoxynucleoside triphosphate generates a full-length RNA bearing a unique, labeled, covalent DNA tail (Fig. 4A). The labeled products ( - 1 0 ^ cpm/ band) were eluted from the gel shown in Figure 4A and identified as U2 and U6 by rapid enzymatic sequencing (Donis-Keller et al. 1977; data not shown, but see Fig. 5, U2, lanes 1-4). Using the same labeling protocol we were able to label U2 but not U6 in the U2/U6 complex (Fig. 4B); however, U6 was labeled in the U4/U6 complex, albeit inefficiently. The identity of the tailed cross-linked complexes in Figure 4B was subsequently confirmed by Northern blotting (data not shown). (Trace labeling of U2 in Fig. 4B, lane 3, is due to slight contamination of the U6 oligonucleotide with the U2 oligonucleotide purified on the same gel.)
Localization of the U2/U6 cross-link by rapid enzymatic RNA sequencing
Labeled U2/U6 complex was eluted from the gel shown in Figure 4B and subjected to rapid enzymatic se-
first dimension then photoreversal
M
first dimension without photoreversal
U2
U4
U6
Oligonucleotide splint technique for selective 3'-end labeling of RNA
Initially, we had hoped to identify the exact site of cross-linking between U2 and U6 by the rapid RNA sequencing strategy of Calvet and Myers (1987). However, we were imable to label the 3 ' end of either RNA in the
Figure. 3. Photoreversal releases stoichiometric amounts of intact U2 and U6 from the U2/U6 complex. Nuclear extract from HeLa cells metabolically labeled with [^^P]phosphate was cross-linked, immunoprecipitated with Y12 antibodies, and separated by two-dimensional gel electrophoresis with {left) or without [right] photoreversal between dimensions. Autoradio-grams were quantified as described in the text.
2148 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Base-pairing between U2 and U6 snRNA
A B
U2
U6
M 1 ? ^
ii m U
U4/U6
U2/U6
U2
U6
Figure 4. Oligonucleotide splint technique for selective labeling of snRNAs in total deproteinized nuclear extract. [A] U6 or U2 was labeled separately in 2 (xl deproteinized nuclear extract. [B] U2 (lane 2) or U6 (lane 3) was labeled separately in 15 M-l deproteinized cross-linked extract. As a control, U2 and U6 were labeled simultaneously in 2 |uil deproteinized extract (lane I). Labeled products were fractionated on a 7% denaturing poly acryamide gel and autoradiographed for 15 min with Kodak X-Omat AR film. Markers (lane M) are a Hpall digest of pBR322.
quencing with RNase T l (Fig. 5, lanes 9 and 10). As controls, labeled uncross-linked U2 snRNA (lanes 1-4) and labeled cross-linked U2 containing psoralen monoad-ducts (lanes 5-8) were sequenced in parallel. Any crosslink involving the labeled RNA (whether intramolecular or intermolecular) would be expected to generate a gap in the sequencing ladder (Calvet and Myers 1983). Because we were able to identify a continuous T l ladder from nucleotide 183 to 31 for U2 in the U2/U6 complex (Fig. 5, cf. lane 10 with lanes 2-4), the cross-link must he 5' to G31 in U2. The T l ladders of the U2/U6 complex (lane 10) and cross-linked U2 (lanes 6-8) are less distinct than that of imcross-linked U2 (lanes 2-4), presumably because random psoralen monoadducts cause band broadening.
Mapping the U2/U6 cross-link by targeted RNase H digestion
Knowing the the U2/U6 cross-link lies upstream of U2 nucleotide 31, we used the strategy shown in Figure 6A to localize the cross-link more precisely on U2. The 3 ' end of U2 in the U2/U6 complex was labeled by the oligonucleotide splint technique, and the labeled U2/U6 complex was digested with RNase H in the presence of one of three complementary oligonucleotides spanning U2 nucleotides 1-42 (see Fig. 8, below). As controls, uncross-linked U2 and U6 were labeled and digested in parallel. We conclude that the cross-link must be located near the 5' end of U2 within oligonucleotide A (nucleotides 1-15) because RNase H digestion with oligonucleotides B (nucleotides 16-30) and C (nucleotides 28-42) yields the same size U2 fragment with both the cross-linked U2/U6 complex and the uncross-linked U2 control (Fig. 6B) but does not digest the uncross-linked
U6 control (data not shown). Note that poly(U) must be added to the RNase H digestion to prevent self-cleavage caused by annealing of the poly(dA) tail with internal U-rich RNA sequences (Fig. 6B, cf. first three lanes).
We used the same experimental strategy to localize the U2/U6 cross-link on U6 (Fig. 7A). The 3 ' end of U2 snRNA in the U2/U6 complex was labeled as in Figure 6, and the labeled complex was digested with RNase H in the presence of one of eight complementary oligonucleotides spanning the entire length of U6 (see Fig. 8). As controls, uncross-linked U2 and U6 were labeled by the oligonucleotide splint technique and digested in parallel; all eight oligonucleotides digested tailed U6 RNA (Fig. 7B,C, left) but not tailed U2 RNA (data not shown). We deduce that the cross-link must be located near the 3 ' end of U6, downstream of nucleotide 85, because (1) digestion with oligonucleotides A, B, C, D, and E yields progressively faster migrating species, all of which migrate more slowly than intact tailed U2 snRNA (Fig. 7B,C); (2) the smallest major U6 digestion product ob-
U2 U2AMT U2/U6 1234 5678 910
31 '33.36
52
63
68
97.98
10! 106 108 111-113
11S.118.H9 125.124
12? 131 136
Wi
147
152
157 158
163
166
17."i 176
180 181
Zs:
•••
Figure 5. Localization of the U2/U6 cross-link by rapid enzymatic RNA sequencing. The 3' end of U2 in the U2/U6 complex was labeled selectively by the oligonucleotide splint technique. After purification by gel electrophoresis, the labeled complex was sequenced enzymatically with Tl ribonuclease (lanes 9 and 10]. As controls, uncross-linked unirradiated U2 (lanes 1-4] and uncross-linked irradiated U2 containing psoralen monoadducts (lanes 5-8] were labeled and sequenced in parallel. No Tl ribonuclease (lanes 1, 5, and 9], 0.1 jjig (lanes 2 and 6), 0.05 |xg (lanes 3, 7, and 10], or 0.02 jxg (lanes 4 and 8] Tl ribonuclease. Numbers [left] indicate positions of guanosines in human U2 sequence (for references, see Reddy 1986).
GENES & DEVELOPMENT 2149
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Hausnei and Weinet
B U2 M U2AJ6 W
A U2 5-
X Y Z
U6 , ^»*»« U2 5" -^v*-**
X Y Z
PAGE
RNase H digestion with X, Y.Z or no
oligonucleotide
U2/U6
PAGE
U2 oligo poly(U)
RNase H
Figure 6. Mapping the U2/U6 cross-link by targeted digestion of U2 with RNase H and complementary oligonucleotides. {A) Deletion mapping strategy. Asterisks (*) indicate 3'-end label; (small open boxes) oligonucleotides; (PAGE) polyacrylamide gel electrophoresis. (B) The 3' end of U2 in the U2/U6 complex was labeled selectively, and the complex was digested by RNase H in the presence of oligonucleotide A (U2i_i5), B {Uli^
-3o)/ ^^ C (U228_42)/ ^s shown in Fig. 8. As a control, labeled uncross-linked U2 snRNA was digested with RNase H and the same oligonucleotides. RNase H digestions contained 1 jjig poly(U), where indicated. Markers (lane M) are as in Fig. 4.
tained with oligonucleotides E, E', and F corresponds to cleavage at the region expected for oligonucleotide F, based on comparison to size markers (Fig. 7C, left); and (3) digestion with oligonucleotide G does not affect the size of U2/U6, but digestion with oligonucleotides E, E', and F does.
Having localized the cross-link between nucleotides 1 and 15 at the 5' end of U2 and nucleotides 85 and 106 at the 3 ' end of U6, we are left with the paradox that U2 oligonucleotide A (directed against one side of the crosslink) cleaves both U2 and U2/U6, while U6 oligonucleotide G (directed against the other side of the cross-link) cleaves U6 but not U2/U6. Apparently, RNA structure (which is altered by the cross-link) affects the ability of a complementary oligonucleotide to form a cleavable duplex, and even a short RNA/DNA duplex is sufficient to assure cleavage by RNAse H.
Discussion
We have demonstrated that irradiation of HeLa nuclear extract with 365 nm light in the presence of the psoralen derivative AMT induces a covalent cross-link between the 5' end of U2 (nucleotides 1-15) and the 3 ' end of U6 (nucleotides 85-106). Psoralen cross-linking is thought to be specific for adjacent pyrimidines on opposite strands, either within or at the end of a base-paired duplex (Calvet and Pederson 1981; Calvet et al. 1982; Rinke et al. 1985; Calvet and Myers 1987; Stroke and Weiner 1989; Teare and Wollenzien 1990; for review, see Cimino et al. 1985). We therefore looked for potential base-pairing interactions in the cross-linked region
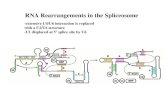
between homologous pairs of U2 and U6 snRNAs and were able to discern a phylogenetically conserved interaction consisting of 6 consecutive base pairs in human, Diosophia melanogastei, and Caenorhabidites elegans, 7 in Schizosacchawmyces pombe and Trypanosoma brucei, 8 in Pisum sativum, and 11 in Saccharomyces ceievisiae (Table 1). The position of the proposed base-pairing region within the 3'-terminal domain of U6 is well conserved, — 9 - 1 3 nucleotides downstream from the nearly invariant 5'-AAGGAUGA-3' sequence that spans the end of stem II in the U4/U6 interaction domain (Brow and Guthrie 1988). The highly conserved U6 sequence 5'-GNGA-3' always lies within the proposed base-pairing; the highly conserved U6 sequence 5'-CAAA-3' lies within the base-paired region of S. ceievisiae only. The position of the proposed base-pairing within the 5'-terminal domain of U2 appears to be more variable, ranging from U2 positions 1-7 in T. biucei to positions 6 -13 in P. sativum. No consensus for the proposed base-pairing interaction is obvious, although an oligopurine tract in U6 tends to pair with the oligopyri-midine tract in U2. Here we emphasize that the relaxed helix geometry available at the end of a duplex favors psoralen cross-linking (Cimino et al. 1985), so the actual cross-link could involve one or more unpaired bases (U97, U96, or Ugp in U6 with U2, C3, or Cg, or Cio in U2, respectively). Admittedly, the proposed base-pairing between U2 and U6 would be the first instance in which the site of conserved base-pairing varies in position on both pairing partners; however. Shine and Dalgamo (1974) noted considerable variation in the position of the eponymous sequence element with respect to the trans-
2150 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Base-paiiing between U2 and U6 snRNA
A U6 5-
X Y Z
U6 -^#»»# U2 5'
X Y Z B i ^ « » * «
U6
PAGE
RNase H digestion with X, Y,Z or no
oligonucleotide
U2/U6
PAGE
U2/U6
a I CD CO CD
z cc
^#iliiN»<lii«HW • » «
U2/U6 U2
8 - A B C D E F G
• *SL , j .
"'"•*»^-.:, »i« i>w«i i« i
.Jii>:«^|JM^>i4-4iJh*.
Figure 7. Mapping the U2/U6 cross-link by targeteci digestion of U6 with RNase H anci complementary oligonucleotides. [A] Deletion mapping strategy (for explanation, see Fig. 6A). [B] (Lanes U2/U6) The 3' end of U2 in the U2/U6 complex was labeled selectively, and the complex was digested by RNase H in the presence of oligonucleotides A (U6i_i7), B (U6i8_2<,), C (U63o_42), D (U643_6o), E {Ue^i^y^), F (U67s_ gp), and G (U69o_io5). (Lane U2) The 3' end of uncross-linked U2 was labeled selectively to determine the mobility of intact U2 snRNA. (Lanes U6) Labeled uncross-linked U6 snRNA was digested with RNase H and the same panel of oligonucleotides as a control. All RNase H digestions contained 1 ixg poly(U). (C) The experiment in B was repeated with oligonucleotides D, E, F, and an additional oligonucleotide, E' {U6^5_g4]. hi both B and C, oligonucleotide E is slightly contaminated with F. Markers (lane M) are as in Fig. 4.
lational initiation codon, and the work of Been and Cech (1987) suggests that conformational flexibility may actually be characteristic of RNA-catalyzed reactions.
Our physical evidence for a U2/U6 cross-link, combined with phylogenetic conservation of the potential for base-pairing in the region of the cross-link, strongly suggests that U2 and U6 form base pairs at some step in mRNA spHcing. What might this step be? In principle, base-pairing between U2 and U6 could occur during spliceosome assembly, sequential phosphoester bond transfer, spliceosome disassembly, or recycling of splice-osomal snRNPs. Distinguishing between these possibilities may be difficult. We found that the U2/U6 crosslink could be generated in nuclear extract prepared according to Heintz and Roeder (1984) but not otherwise specially treated; however, the yield of cross-link was quite variable (data not shown) and was not affected by the addition of all four nucleoside triphosphates or by treatment of the extract with alkaline phosphatase to
deplete endogenous nucleoside triphosphates (data not shown). Nor was the yield of cross-link affected by incubating an mRNA precursor under splicing conditions with untreated extract or with extract pretreated with EDTA (Abmayr et al. 1988) to block cleavage at the 5'-splice site (Fig. 1 and data not shown). Although cross-linking between U2 and U6 might thus appear to be independent of mRNA splicing, no conclusions can be drawn because snRNPs in the extract are present in vast excess over the added mRNA precursor, and endogenous U2/U6 complexes generated in vivo might outnumber those generated during active mRNA splicing in vitro.
The spliceosome is dynamic
Our evidence for base-pairing between U2 and U6 is consistent with the idea that the spliceosome is a dynamic structure in which RNA folds and refolds in the course of the reaction. For example, the proposed U2/U6 base-
GENES & DEVELOPMENT 2151
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Hausnei and Weiner
U2
I G- C CAAUAUAUUAAAUGGAUUUUUGGAGCAG ^
GCAUCe CCUGG
CGUOGC 6GACC,
"CCA"
UGACACGCAAAUU|:GUGAAGCGUU)CCAUAUUUU(U)^ V I I
A cap
90 100
Figure 8. Location of cross-link on secondary structure models of U2 and U4/U6 snRNAs. Oligonucleotides used in the targeted RNase H digestion of U2 and U6 snRNAs (Figs. 6 and 7) are indicated. The cross-linked regions, as defined by experiments shown in Figs. 6 and 1, are boxed and include the proposed U2/U6 base pairs (Table 1). U2 is drawn according to Ares and Igel (this issue), and U4/U6, according to Brow and Guthrie (1988). For clarity, the maximally base-paired U4/U6 structure is shown, although the ability of U6 to base-pair with U4 in the U4/U6 complex and with U2 in the U2/U6 complex could be mutually exclusive (see text).
pairing would disrupt much of phylogenetically conserved U2 stem I (particularly in S. cerevisiae-, Guthrie and Patterson 1988). This suggests that intramolecular U2 base-pairing and intermolecular U2/U6 pairing are mutually exclusive and occur at different steps in the splicing reaction. A related issue concerns U4 and U6, which are extensively base-paired in the U4/U6 snRNP prior to spliceosome assembly, as documented by melting studies (Bringmann et al. 1984; Hashimoto and Steitz 1984; Brow and Guthrie 1988), by psoralen cross-linking (Rinke et al. 1985), and by phylogenetic comparisons (Brow and Guthrie 1988). Yet U4/U6 base-pairing is either lost (Pikielny et al. 1986; Cheng and Abelson 1987; Lamond et al. 1988) or partially unwound after lariat formation (Blencowe et al. 1989), possibly by the action of an RNA helicase (Burgess et al. 1990). Can U6 form base pairs simultaneously with U4 and U2, or must the U4/U6 interaction be partially or completely disrupted before the U2/U6 interaction can take place? We are currently addressing this question experimentally. Finally, given the many compelling arguments that mRNA splicing is mechanistically (and perhaps ancestrally) related to group II self-splicing (Sharp 1985; Cech 1986; Bruzik and Steitz 1990), it is tempting to
speculate that the proposed U2/U6 base-pairing plays a role in catalysis itself; however, the absence of a consensus sequence for the proposed U2/U6 interaction suggests that this role is more likely to be structural than enzymatic.
Published data regarding the function of the 5' end of U2 and the 3 ' end of U6 are difficult to interpret unambiguously. Consistent with our proposed U2/U6 interaction, mammalian splicing is inhibited by RNase H digestion of the 5' end of U2 RNA (Black et al. 1985; Krainer and Maniatis 1985; Frendewey et al. 1987; Zill-mann et al. 1988), by antisense oligonucleotides directed against the 3 ' end of U6 (Blencowe et al. 1989), and by mutations in U2 nucleotides 3 - 6 (J. Wu and J.L. Manley, pers. comm.). On the other hand, deletion of most of U2 stem I may have little effect on splicing in a Xenopus oocyte assay system (Hamm et al. 1989, but see Pan and Prives 1989), deletion or mutation of the 3 ' end of yeast U6 does not invariably abolish splicing (Frabizio et al. 1989), a hybrid human/yeast U2 functions in S. cerevisiae although the proposed U2/U6 base-pairing is severely affected (Shuster and Guthrie 1990; Ares and Igel, this issue), and a triple mutation within the proposed base-pairing region of U2 in S. cerevisiae has no apparent
2152 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Base-paiiing be tween U2 and U6 snRNA
Table 1. Phylogenetically conserved base-pairing between U2 and U6 snRNAs
Human U2 5' 1 10 AUCGCUUCUCG
U6 3' GCGAAGUGC- 10 -AGUAGGAA
Drosophila melanogaster
Schizosaccharomyces pombe
Saccharomyces cerevisiae
Caenorhabditis elegans
Trypanosoma brucei
Pisum sativum
U2 5'
U6 3' 95
U2 5'
U6 3'
U2 5'
U6 3'
U2 5'
U6 3'
U2 5'
U6 3'
U2 5'
U6 3'
95 87
1 10 AUCGCUUCUCGG
GCGAAGUGCU- 9 -AGUAGGAA 86
1 8 AUUCUCUC
AAGAGAG- 12 -AGUAGGAA 88 82
1 10 ACGAAUCUCUUUG
UUUAGAGAAAC- 9 -AGUAGGAA 102 ~9T
1 8 AUCGCUUC
GCGAAG- 13 -AGUAGGAA 90 85
1 7 AUAUCUU
UAUAGAG- 1 1 -AGUCGGAA 93 87
1 10 AUACCUUUCUCGG
AAAGAGCU- 10 -AGUAGGAA 86 79
For tabulation of all sequences except C. elegans U2 (Thomas et al. 1990) and P. sativum (B.A. Hanley and M.A. Schuler, in prep.), see Reddy (1986, 1990). Three additional base pairs can be drawn for P. sativum by bulging U o in U6. The distance between the proposed U2/U6 base-pairing and the conserved U6 sequence 5'-AAGGAUGA-3' spanning the end of stem II in the U4/U6 interaction domain (Brow and Guthrie 1988) is given in nucleotides. Two other conserved U6 sequences, 5'-GNGA-3' and 5'-CAAA-3', are underlined and doubly underlined, respectively.
phenotype (D.J. Field, D.M. McMaster, and J.D. Friesen, pers. comm.). How might these apparent contradictions and paradoxes be resolved? Although fundamentally similar, splicing in yeast and mammals differs significantly; the branch site, for example, is essential in one system (Parker et al. 1987) but almost dispensable in the other except under special circumstances (Wu and Manley 1989; Zhuang and Weiner 1989; Zhuang et al. 1989). Thus, evidence against the proposed U2/U6 base-pairing in yeast systems may simply indicate that this U2/U6 interaction is not rate limiting or is structurally redundant under the conditions tested; the proposed U2/U6 interaction might even be unnecessary for splicing but might rather function, for example, in efficient recycling of snRNPs that remain bound to the in-tron lariat after exon ligation. In any case, many highly conserved sequence elements, including the entire
s tem-loop I of U l (Liao et al. 1990) and much of the base-pairing between U4 and U6 (Fabrizio and Abelson 1990) appear to be dispensable in yeast, genetically or biochemically. The implication is that evolution provides a far more sensitive test of function than laboratory assays and that spliceosomal snRNA structures and sequences are both redundant and robust.
Clearly, additional experiments will be required to test the significance of the proposed U2/U6 base-pairing, but taken together, the biochemical and phylogenetic data build a strong case for the importance of this interaction between the two most highly conserved spliceosomal snRNAs. Compensatory base changes in Ul snRNA can suppress 5'-splice site mutations both in mammals (Zhuang and Weiner 1986) and in yeast [Sera-phin et al. 1988; Siliciano and Guthrie 1988; Seraphin and Rosbash 1989), and compensatory base changes in
GENES & DEVELOPMENT 2153
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Hausner and Weinei
U2 snRNA can suppress branch site mutations both in yeast (Parker et al. 1987) and in mammals (Wu and Manley 1989; Zhuang and Weiner 1989). We hope to test the significance of the proposed base-pairing between U2 and U6 by asking whether mutations in U2 can be suppressed by compensatory base changes in U6 in vivo. The proposed base-pairing could also be tested in vitro using the S. ceievisiae system in which functional yeast snRNPs are reconstituted on exogenous U2 and U6 transcripts (Fabrizio et al. 1989; McPheeters et al. 1989).
Material and methods
Analysis of metabolically labeled RNA
HeLa cells were maintained in suspension culture at 37°C and 5% CO2 at 2.5 X 105 cells/ml in RPMI 1640 (Flow and GIBCO laboratories) supplemented with 10% (vol/vol) heat-inactivated fetal bovine serum, 2 mM glutamine, 100 |xg streptomycin/ml and 60 (j,g penicillin/ml. Cells (150 ml) were collected for 2 min at 500 rpm at room temperature, washed twice in phosphate-free medium, resuspended in 15 ml of the same medium containing 10 mCi P^Pjphosphate (Amersham), and harvested 12 hr later. Nuclei were purified as described (Weiner 1980), resuspended in 1 ml buffer D [20 mM HEPES/KOH (pH 7.9) 100 mM KCl, 0.2 mM EDTA, 0.5 mM PMSF, 0.5 mM DTT, and 20% (vol/vol) glycerol] and broken by sonication on ice (three pulses of 10 sec each at 20-sec intervals with a Branson sonicator at setting 3). Equal aliquots of the suspension were cross-linked as described below, immunoprecipitated with Y12 anti-Sm antibodies (Steitz 1989), phenol-extracted, and fractionated by two-dimensional denaturing polyacrylamide gel electrophoresis (5% first dimension, 9% second dimension; 19 : 1, acrylamide/bisa-crylamide] with or without photoreversal of the cross-links between dimensions. The first-dimension gel strip was photore-versed according to Rabin and Crothers (1979) by irradiation in the dark with a short-wavelength UV source (Chromato-UVe Transilluminator, model C-61; 254 nm peak emission) at a distance of 2.5 cm for 20 min at 4°C.
Cross-linking of nuclear extracts and Northern blotting
HeLa nuclear extracts were prepared according to Heintz and Roeder (1984), except that the extract was dialyzed three times for 30 min in buffer D. Nuclear extract (15-150 |xl) was deposited on Parafilm (American National Can) in an open petri dish on ice, AMT (HRI Associates) was added to 40 iig/ml, and the solution was irradiated from above at 4°C for 1 hr at a distance of 5 cm from a long-wavelength light source (Chromato-UVe Transilluminator, model C-62; 365 nm peak emission, 6500 |xW/cm^ at source surface). A 2.5-mm glass plate was placed on the light source to block shortwave irradiation. Fresh AMT (40 |xg/ml) was added to the reaction at 10-min intervals during the irradiation process (Schoeniger and Jelinek 1986). After cross-linking, the mixture was treated with 1 mg/ml proteinase K and 0.5% SDS for 15 min at 37°C, phenol-extracted, and ethanol-precipitated. The RNA products were fractionated by denaturing gel electrophoresis through 5% polyacrylamide (19 : 1, acrylamide/bisacrylamide) in 7 M urea in TBE and then elec-troblotted (Bio-Rad Trans-Blot) to Hybond-N nylon membrane (Amersham) for 3 hr at 150 mA in TAE buffer [40 mM Tris/ acetic acid (pH 8.5), 2 rtiM EDTA] at room temperature. The RNAs were covalently bound to the membrane (wet or dry) by irradiation for 2 min on each side with 306 nm light (Chromato-UVe Transilluminator, model TM-40; 306 nm peak emis
sion, 8000 |xW/cm^j. The membrane was preincubated for 30 min at 37°C in hybridization buffer [20 mM Na2HP04/NaH2P04 (pH 8.0), 1% SDS, 5x SSC (750 mM NaCl, 75 mM Na citrate)] and then hybridized for >3 hr at 37°C in the same buffer with ^^P-labeled single-stranded DNA oligonucleotides complementary to one or all five spliceosomal snRNAs (sp. act >10* cpm/ ng, 0.5 X 10 to 2 X 10* cpm/ml each): Ul28_47 (or Ulpg.no)/ U228_42 (O^ U2i52_i72), U458_75 (or U4i26_i4i), U568_88/ ^nd U643_6o (or U69o_io5), where subscripted numbers indicate snRNA nucleotides to which the probe is complementary. The membrane was washed three times in 5 x SSC at room temperature and autoradiographed with Kodak X-Omat AR film overnight. In some experiments, cross-linking was performed after nuclear extract was pretreated with 4 mM EDTA for 30 min at 0°C to block exon ligation (Abmayr et al. 1988) and then incubated for 60 min at 30°C under splicing conditions with 10 ng human p-globin mRNA precursor obtained by runoff transcription (Melton et al. 1984) from pSP64-A6 (Krainer et al. 1984).
Targeted digestion of snRNAs by RNase H
Unlabeled RNA corresponding to 15 |xl of nuclear extract was annealed with 250 ng of single-stranded DNA oligonucleotide (12-18 nucleotides long) by heating for 2 min at 70°C in 25 |xl of reaction buffer (20 mM Tris/HCl at pH 7.8, 100 mM KCl, 10 mM MgClj; 0.1 mM EDTA, 0.1 mM DTT). RNase H (8 units, Pharmacia) was then added for 1 hr at 30°C, and the reaction was phenol-extracted and ethanol-precipitated before electrophoresis. For snRNAs labeled by the splint technique (see below), annealing was in 20 fil [20 mM HEPES/KOH (pH 7.9], 50 mM KCl, 10 mM MgClj, 1 mM DTT, 1 jig poly(U), and 100-200 ng oligonucleotide] and digestion was for 30 min. To prevent loss of small digestion products, the entire reaction was evaporated in a Speed Vac Concentrator (Savant Instruments) and the resulting pellet was redissolved by boiling in 85% formamide before electrophoresis.
Oligonucleotide splint technique for selective 3'-end labeling of RNA
The 3' end of an RNA can be labeled selectively and uniquely by primer extension on a synthetic DNA oligonucleotide template using T7 DNA polymerase and a single deoxynucleotide triphosphate precursor as shown below (uppercase denotes RNA; lowercase denotes DNA). Under the conditions used, the RNA primer is fully extended but the DNA primer cannot be extended in the absence of the other three precursors:
U2 snRNA 5'-AUC . . . C C U C C A G G A A C G G U G C A C C C O H - 3 '
g g t c c t t g c c a c g t g g gt t t t t t t t t t t tg-5 ' i T7 DNA polymerase + |a-32P]dATP
5'-AUC . . . CCUCCAGGAACGGUGCACCCaaaaaaaaaaaaoH-3'
Uncross-linked (2 |xl) or cross-linked nuclear extract (15 [x\] was deproteinized and annealed with 400 ng DNA oligonucleotide by heating for 2 min at 70°C in 50 mM Tris/HCl (pH 7.8), 10 mM MgCl2, and 1 mM DTT. For labeling U2 snRNA, the
TTCCTGG and for labeling U6 snRNA, 5'-GTTTTTTTTTT-TTAAAAATATGGAACGCTT. Labeling of U6 snRNA also required addition of oligonucleotide U643_6o, which probably renders the 3' end of U6 more accessible to the DNA primer. The annealing reaction was quenched on ice for 3 min, 20 fJiCi [a-32p]dATP (3000 Ci/mmole, Amersham) and 26 units of T7 DNA polymerase (Sequenase, v. 2.0, U.S. Biochemicals) were added, and the reaction was incubated for 30 min at 37°C. After
2154 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Base-pairing between U2 and U6 snRNA
the addition of oyster glycogen carrier (1 [xg), the RNA was de-proteinized, ethanol-precipitated, purified by denaturing gel electrophoresis, and eluted into 400 [xl of elution buffer [10 mM Tris-HCl (pH 7.8), 500 mM sodium acetate, 1 mM EDTA, 1% (wt/vol) SDS], and 400 JJLI phenol/chloroform/isoamyl alcohol (50 : 49 : 1) by gentle overnight agitation at room temperature on a Nutator (Adams). Carrier tRNA was added, and the eluted RNA was ethanol-precipitated and subjected to rapid enzymatic RNA sequencing with Tl ribonuclease as described by Donis-Keller et al. (1977).
Acknowledgments
We thank Joan Steitz for Y12 antibodies, Jim Bruzik for masterful artwork in Figure 8, and liana Stroke and Bansidhar Datta for advice. This work was supported by National Institutes of Health grant GM-31073. T.-P.H. was a postdoctoral fellow of the Deutsche Forschungsgemeinschaft.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 USC section 1734 solely to indicate this fact.
References
Abmayr, S.M., R. Reed, and T. Maniatis. 1988. Identification of a functional mammalian spliceosome containing unspliced pre-mRNA. Pioc. Natl. Acad. Sci. 85: 7216-7220.
Been, M.D. and T.R. Cech. 1987. Selection of circularization sites in a Group IIVS RNA requires multiple alignments of an internal template-like sequence. Cell 50: 951-961.
Black, D.L., B. Chabot, and J.A. Steitz 1985. U2 as well as UI small nuclear ribonucleoproteins are involved in pre-mRNA spUcing. Cell 42: 737-750.
Blencowe, B.J., B.S. Sproat, U. Ryder, S. Barabino, and A.I. La-mond. 1989. Antisense probing of the human U4/U6 snRNP with biotinylated 2'-OMe RNA oligonucleotides. Cell 59: 531-539.
Bringmann, P., B. Appel, J. Rinke, R. Reuter, H. Theissen, and R. Luhrmann. 1984. Evidence for the existence of snRNAs U4 and U6 in a single ribonucleoprotein complex and for their association by inter-molecular base-pairing. EMBO f. 3: 1357-1363.
Brow, D.A. and C. Guthrie. 1988. Sphceosomal RNA U6 is remarkably conserved from yeast to mammals. Nature 334:213-218.
Bruzik, J.P. and J.A. Steitz. 1990. Spliced leader RNA sequences can substitute for the essential 5' end of Ul RNA during splicing in a mammalian in vitro system. Cell 62: 888-899.
Burgess, S., J.R. Couto, and C. Guthrie. 1990. A putative ATP binding protein influences the fidelity of branchpoint recognition in yeast splicing. Cell 60: 705-717.
Calvet, J.P. and J.A. Myers. 1987. In vivo secondary structure analysis of the small nuclear RNA Ul using psoralen cross-linking. /. Mol. Biol. 197: 543-553.
Calvet, J.P. and T. Pederson. 1981. Base-pairing interactions between small nuclear RNAs and nuclear RNA precursors as revealed by psoralen cross-linking in vivo. Cell 26: 363-370.
Calvet, J.P., L.M. Meyer, and T. Pederson. 1982. Small nuclear RNA U2 is base-paired to heterogeneous nuclear RNA. Science 217: 456-458.
Cech, T.R. 1986. The generahty of self-splicing RNA: Relationship to nuclear mRNA splicing. Cell 44: 207-210.
Chabot, B., D.L. Black, D.M. LeMaster, and J.A. Steitz. 1985.
The 3' splice site of pre-messenger RNA is recognized by a small nuclear ribonucleoprotein. Science 230: 1344-1349.
Cheng, C.-C. and J. Abelson. 1987. Spliceosome assembly in yeast. Genes Dev. 1: 1014-1027.
Cimino, G.D., H.B. Gamper, S.T. Isaacs, and J.E. Hearst. 1985. Psoralens as photoactive probes of nucleic acid structure and function: Organic chemistry, photochemistry, and biochemistry. Annu. Rev. Biochem. 54: 1151-1193.
Donis-Keller, H., A.M. Maxam, and W. Gilbert. 1977. Tl mapping of adenines, guanines, and pyrimidines in RNA. Nucleic Acids Res. 4: 2527-2538.
Fabrizio, P. and J. Abelson. Two domains at yeast U6 small nuclear RNA required for both steps of nuclear precursor messenger RNA sphcing. Science 250: 404-409.
Fabrizio, P., D.S. McPheeters, and J. Abelson. 1989. In vitro assembly of yeast U6 snRNP: A functional assay. Genes Dev. 3:2137-2150.
Frendewey, D., A. Kramer, and W. Keller. 1987. Different small nuclear ribonucleoprotein particles are involved in different steps of splicing complex formation. Cold Spring Harbor Symp. Quant. Biol. 52: 287-298.
Garcia-Blanco, M.A., S.F. Jamison, and P.A. Sharp. 1989. Identification and purification of a 62,000-dalton protein that binds specifically to the polypyrimidine tract of introns. Genes Dev. 3: 1874-1886.
Gerke, V. and J.A. Steitz. 1986. A protein associated with small nuclear ribonucleoprotein particles recognizes the 3' splice site of premessenger RNA. Cell 47: 973-984.
Guthrie, C. and B. Patterson. 1988. Spliceosomal snRNAs. Annu. Rev. Genet. 22: 387-419.
Hamm, J., N.A. Dathan, and I.W. Mattaj. 1989. Functional analysis of mutant Xenopus U2 snRNAs. Cell 59: 159-169.
Hashimoto, C. and J.A. Steitz. 1984. U4 and U6 RNAs coexist in a single small ribonucleoprotein particle. Nucleic Acids Res. 12: 3283-3293.
Heintz, N. and R.G. Roeder. 1984. Transcription of human his-tone genes in extracts from synchronized HeLa cells. Proc. Natl. Acad. Sci. 81: 2713-2717.
Konarska, M.M. and P.A. Sharp. 1987. Interactions between small nuclear ribonucleoprotein particles in formation of spliceosomes. Cell 49: 763-774.
Krainer, A.R. and T. Maniatis. 1985. Multiple factors including the small nuclear ribonucleoproteins Ul and U2 are necessary for pre-mRNA splicing in vitro. Cell 42: 725-736.
Krainer, A.R., T. Maniatis, B. Ruskin, and M.R. Green. 1984. Normal and mutant human p-globin pre-mRNAs are faithfully and efficiently spHced in vitro. Cell 36: 993-1005.
Lamond, A., M.M. Konarska, P.J. Grabowski, and P.A. Sharp. 1988. Spliceosome assembly involves the binding and release of U4 small nuclear ribonucleoproteins. Proc. Natl. Acad. Sci. 85:411-415.
Legrain, P., B. Seraphin, and M. Rosbash. 1988. Early commitment of yeast pre-mRNA to the spliceosome pathway. Mol. Cell. Biol. 8: 3755-3760.
Liao, X., L. Kretzner, B. Seraphin, and M. Rosbash. Universally conserved and yeast-specific Ul snRNA sequences are important but not essential for Ul snRNP functions. Genes Dev. 4: 1766-1776.
Lossky, M., G.J. Anderson, S.P. Jackson, and J.D. Beggs. 1987. Identification of a yeast snRNP protein and detection of snRNP-snRNP interactions. Cell 51: 1019-1026.
Luhrmann, R. 1988. snRNP proteins. In Structure and function of major and minor snRNPs (ed. M.L. Bimstiel), pp. 71-99. Springer-Verlag, Heidelberg.
Maser, R.L. and J.P. Calvet. 1989. U3 small nuclear RNA can be psoralen-cross-linked in vivo to the 5' external transcribed
GENES & DEVELOPMENT 2155
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

Hausnet and Weinei
spacer of pre-ribosomal RNA. Proc. Natl. Acad. Sci. 86: 6523-6527.
McPheeters, D.S., P. Fabrizio, and J. Abelson. 1989. In vitro re-constitution of functional yeast U2 snRNPs. Genes Dev. 3: 2124-2136.
Melton, D.A., P.A. Krieg, M.R. Rebagliati, T. Maniatis, K. Zinn, and M.R. Green. 1984. Efficient in vitro synthesis of biologically active RNA and RNA hybridization probes from plasmids containing a bacteriophage SP6 promoter. Nucleic Acids Res. 12: 7035-7056.
Nelson, K.K. and M.R. Green. 1989. Mammalian U2 snRNP has a sequence-specific RNA-binding activity. Genes Dev. 3: 1562-1571.
Pan, Z.-Q. and C. Prives. 1989. U2 snRNA sequences that bind U2-specific proteins are dispensable for the function of U2 snRNP in splicing. Genes Dev. 3: 1887-1898.
Parker, R., P.G. Siliciano, and C. Guthrie. 1987. Recognition of the TACTAAC box during mRNA splicing in yeast involves base-pairing to the U2-like snRNA. Cell 49: 229-23^.
Pikielny, C.W., B.C. Rymond, and M. Rosbash. 1986. Electrophoresis of ribonucleoproteins reveals an ordered assembly pathway of yeast splicing complexes. Nature 324: 341-345.
Rabin, D. and D.M. Crothers. 1979. Analysis of RNA secondary structure by photochemical reversal of psoralen crosslinks. Nucleic Acids Res. 7: 689-703.
Reddy, R. 1986. Compilation of small RNA sequences. Nucleic Acids Res. (suppl.) 14: r61-r72.
. 1990. Compilation of small RNA sequences. Nucleic Acids Res. (suppl.) 18: 2231-2235.
Rinke, J., B. Appel, M. Digweed, and R. Liihrmann. 1985. Localization of a base-paired interaction between small nuclear RNAs U4 and U6 in intact U4/U6 small ribonucleoprotein particles by psoralen cross-linking. /. Mol. Biol. 185: 7 2 1 -731.
Ruby, S.W. and J. Abelson. 1988. An early hierarchic role of Ul small nuclear ribonucleoprotein in spliceosome assembly. Science 242: 1028-1035.
Ruskin, B., P.D. Zamore, and M.R. Green. 1988. A factor, U2AF, is required for U2 snRNP binding and splicing complex assembly. Cell 52: 207-219.
Schoeniger, L.O. and W.R. Jelinek. 1986. 4.5S RNA is encoded by hundreds of tandemly linked genes, has a short half-life, and is hydrogen bonded in vivo to poly(A)-terminated RNAs in the cytoplasm of cultured mouse cells. Mol. Cell. Biol. 6: 1508-1519.
Seraphin, B. and M. Rosbash. 1989. Mutational analysis of the interactions between Ul small nuclear RNA and pre-mRNA of yeast. Gene 82: 145-151.
Seraphin, B., L. Kretzner, and M. Rosbash. 1988. A U l snRNA: pre-mRNA base-pairing interaction is required early in yeast spliceosome assembly but does not uniquely define the 5' cleavage site. EMBO /. 7: 2533-2538.
Sharp, P.A. 1985. On the origin of RNA splicing and introns. Cell 42: 397-400.
Shine, J. and L. Dalgamo. 1974. The 3'-terminal sequence of E. coli 16S rRNA; Complementarity to nonsense triplets and ribosome binding sites. Pwc. Natl. Acad. Sci. 71: 1342-1346.
Shuster, E.O. and C. Guthrie. 1990. Human U2 snRNA can function in pre-mRNA splicing in yeast. 1990. Nature 345: 270-273.
Siliciano, P.G. and C. Guthrie. 1988. 5' splice site selection in yeast: Genetic alterations in basepairing with U l reveal additional requirements. Genes Dev. 2: 1258-1267.
Steitz, J.A. 1989. Immunoprecipitation of ribonucleoproteins using autoantibodies. Methods Enzymol. 180: 468-481 .
Steitz, J.A., D.L. Black, V. Gerke, K. Parker, A. Kramer, D. Fren-dewey, and W. Keller. 1988. Functions of abundant U-snRNPs. In Structure and function of major and minor snRNPs |ed. M.L. Birnstiel), pp. 115-154. Springer-Verlag, Heidelberg.
Stroke, I.L. and A.M. Weiner. 1989. The 5' end of U3 snRNA can be cross-linked in vivo to the external transcribed spacer of rat ribosomal RNA precursors. /. Mol. Biol. 210: 497-512.
Swanson, M.S. and G. Dreyfuss. 1988. RNA binding specificity of hnRNP proteins: A subset bind to 3 ' end of introns. EMBO]. 7 :3519-3529.
Tazi, J., C. Albert, J. Temsamani, I. Reveillaud, G. Cathala, C. Brunei, and P. Jeanteur. 1986. A protein that specifically recognizes the 3 ' splice site of mammalian pre-mRNA introns is associated with a small nuclear ribonucleoprotein. Cell 47: 755-766.
Teare, J. and P. Wollenzien. 1990. The structure of a pre-mRNA molecular in solution detected with a site directed cross-linking reagent. Nucleic Acids Res. 18: 855-864.
Thomas, J., K. Lea, E. Zucker-Aprison, and T. Blumenthal. 1990. The spliceosomal snRNAs of Caenorhabditis elegans. Nucleic Acids Res. 18: 2633-2642.
Weiner, A.M. 1980. An abundant cytoplasmic 7S RNA is complementary to the dominant interspersed middle repetitive DNA sequence family in the human genome. Cell 22: 2 0 9 -218.
Wu, J. and J.L. Manley. 1989. Mammalian pre-mRNA branch site selection by U2 snRNP involves base pairing. Genes Dev. 3: 1553-1561.
Zhuang, Y. and A.M. Weiner. 1986. A compensatory base change in Ul snRNA suppresses a 5' splice site mutation. Cell 46: 827-835.
. 1989. A compensatory base change in human U2 snRNA suppresses a globin branch site mutation. Genes Dev. 3: 1545-1552.
Zhuang, Y., A.M. Goldstein, and A.M. Weiner. 1989. UA-CUAAC is the preferred branch site for mammalian mRNA spHcing. Proc. Natl. Acad. Sci. 86: 2752-2756.
Zillman, M., M.L. Zapp, and S.M. Berget. 1988. Gel electropho-retic isolation of splicing complexes containing U l small nuclear ribonucleoprotein particles. Mol. Cell. Biol. 8: 8 1 4 -821.
2156 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from

10.1101/gad.4.12a.2146Access the most recent version at doi: 4:1990, Genes Dev.
T P Hausner, L M Giglio and A M Weiner nuclear ribonucleoprotein particles.Evidence for base-pairing between mammalian U2 and U6 small
References
http://genesdev.cshlp.org/content/4/12a/2146.full.html#ref-list-1
This article cites 62 articles, 22 of which can be accessed free at:
License
ServiceEmail Alerting
click here.right corner of the article or
Receive free email alerts when new articles cite this article - sign up in the box at the top
Copyright © Cold Spring Harbor Laboratory Press
Cold Spring Harbor Laboratory Press on August 27, 2018 - Published by genesdev.cshlp.orgDownloaded from