ESTABLISHING POSITIONAL INFORMATION IN DROSOPHILA … · 1 ESTABLISHING POSITIONAL INFORMATION IN...
5
1 ESTABLISHING POSITIONAL INFORMATION IN DROSOPHILA EARLY EMBRYOGENESIS: THE ANTERIOR-POSTERIOR AXIS Mutational analysis of early Drosophila development: The initial insights into the control of pattern formation emerged from studies of the fruit fly led primarily by Christiane Nüsslein-Volhard, Eric Wieschaus and their colleagues. These researchers performed extensive genetic screens, essentially saturating the genome for mutations that alter the anterior-posterior (a-p) and dorsal-ventral (d-v) patterning of the embryo. Their assay was to examine the larval exoskeleton in the Drosophila embryo for any relevant changes. Their reasoning was that any abnormality in the body plan would be identified in the larval exoskeleton. The larval exoskeleton is a non-cellular structure made of a polysaccharide polymer named chitin that is produced as a secretion of the epidermal cells of the embryo. Each structure of the exoskeleton is made by the epidermal cells immediately underlying that structure. With its intricate pattern of hairs, indentations and other structures the exoskeleton provides numerous landmarks to serve as indicators of the fates assigned to the many epidermal cells. In particular there are many distinct anatomical characteristics along the a-p or the d-v axes. Furthermore, because all the nutrients necessary to develop to the larval stage are pre-packed in the egg, mutant embryos in which a-p or d-v cells fates are drastically altered can nonetheless develop to the end of embryogenesis and produce a mutant larva. The exoskeleton of such a larva mirrors the mutant fates assigned to subsets of the epidermal cells and thus can identify genes worthy of detailed analysis. An example is the phenotype derived from the mutant bicoid (bcd) below left as opposed to the wild type exoskeleton secreted from a normal embryo (below right) ANTERIOR (HEAD) POSTERIOR (TAIL)
Transcript of ESTABLISHING POSITIONAL INFORMATION IN DROSOPHILA … · 1 ESTABLISHING POSITIONAL INFORMATION IN...

1
ESTABLISHING POSITIONAL INFORMATION IN DROSOPHILA EARLY EMBRYOGENESIS: THE ANTERIOR-POSTERIOR AXIS
Mutational analysis of early Drosophila development: The initial insights into the control of pattern formation emerged from studies of the fruit fly led primarily by Christiane Nüsslein-Volhard, Eric Wieschaus and their colleagues. These researchers performed extensive genetic screens, essentially saturating the genome for mutations that alter the anterior-posterior (a-p) and dorsal-ventral (d-v) patterning of the embryo. Their assay was to examine the larval exoskeleton in the Drosophila embryo for any relevant changes. Their reasoning was that any abnormality in the body plan would be identified in the larval exoskeleton. The larval exoskeleton is a non-cellular structure made of a polysaccharide polymer named chitin that is produced as a secretion of the epidermal cells of the embryo. Each structure of the exoskeleton is made by the epidermal cells immediately underlying that structure. With its intricate pattern of hairs, indentations and other structures the exoskeleton provides numerous landmarks to serve as indicators of the fates assigned to the many epidermal cells. In particular there are many distinct anatomical characteristics along the a-p or the d-v axes. Furthermore, because all the nutrients necessary to develop to the larval stage are pre-packed in the egg, mutant embryos in which a-p or d-v cells fates are drastically altered can nonetheless develop to the end of embryogenesis and produce a mutant larva. The exoskeleton of such a larva mirrors the mutant fates assigned to subsets of the epidermal cells and thus can identify genes worthy of detailed analysis. An example is the phenotype derived from the mutant bicoid (bcd) below left as opposed to the wild type exoskeleton secreted from a normal embryo (below right)
ANTERIOR (HEAD) POSTERIOR (TAIL)

2
The screen identified two types of mutations that affect the body plan: the zygotically acting genes and the maternal-effect genes. The former are genes, whose expression in the zygote is the sole source of their gene products. The latter are genes, which their mRNAs are provided by the mother in the ovary. Thus the maternal effect proteins are already present in the oocyte at fertilisation. In maternal-effect mutations, the phenotype of the offspring depends on the genotype of the mother, not of the offspring because the source of the genes is the mother’s genome. Hence, if we label a recessive maternal-effect mutation m, in a cross of an m/m female with a +/+ male, all the embryos will be abnormal although they are genetically normal (m/+).
The Drosophila a-p axis: Positional information along the a-p axis of the Drosophila embryo is initially established through the creation of concentration gradients by two transcription factors: the BCD and HB-M proteins, which are products of two maternal effect genes. These factors interact to generate different patterns of gene expression along the axis. The BCD protein, encoded by the bcd gene, is distributed in a steeper gradient in the early embryo, whereas the HB-M protein encoded by one of the mRNAs of the hunchback (hb) gene, is distributed in a shallower but longer gradient. Both concentration gradients have their high points at the anterior pole. In somewhat different ways, the gradients of both these proteins depend on the diffusion of protein from a localised origin: each protein is produced through localised translation of an mRNA species, one tethered to microtubules at the anterior pole of the syncitial embryo and the other tethered at the posterior pole.
The origin of the BCD gradient is quite straightforward. The maternal bcd mRNA packaged during oogenesis into the developing oocyte is tethered to (the minus end of) microtubules, which are located at the anterior pole. Translation of BCD protein begins midway through the early nuclear divisions of the embryo. The protein diffuses in the common cytoplasm of the syncitium. Because the protein is a transcription factor, it contains signals that direct it to the nuclei. Those nuclei nearer to the anterior pole incorporate a higher concentration of the diffusing BCD protein that the nuclei further away. This difference creates the steep of the BCD gradient. The origin of the HB-M gradient is more complex. Unlike the bcd mRNA, the hb mRNA is uniformly distributed throughout the oocyte and the syncitial embryo. However, translation of hb-m mRNA is blocked by a repressor protein the NANOS (NOS) protein. In the mother nos mRNA is deposited at the posterior end in the oocyte through its association with the plus (+) ends of microtubules. At the time of its translation, NOS becomes distributed by diffusion in a gradient opposite that of BCD. The NOS gradient has its high point at the posterior pole and drops down to background levels in the middle of the a-p axis. NOS protein inhibits translation of hb-m mRNA the level of inhibition being proportional to the amount of NOS protein. This produces the shallow gradient of HB-M from the anterior to the posterior of the embryo.
3
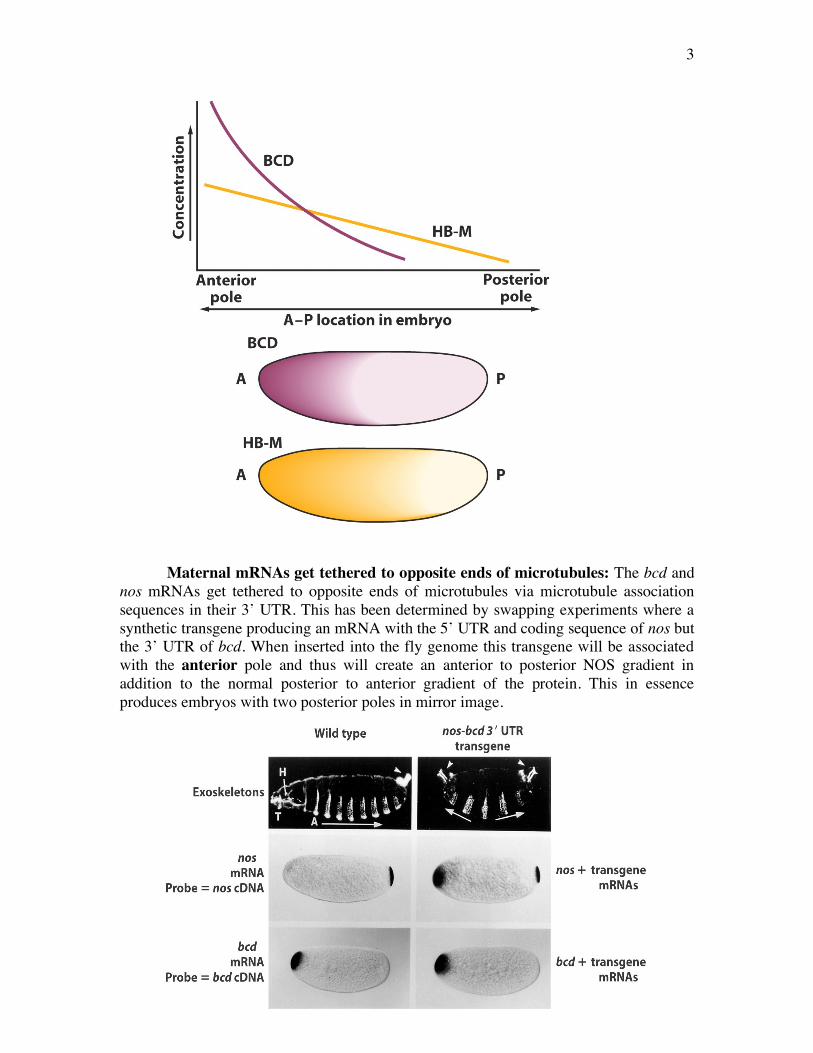
Maternal mRNAs get tethered to opposite ends of microtubules: The bcd and
nos mRNAs get tethered to opposite ends of microtubules via microtubule association sequences in their 3’ UTR. This has been determined by swapping experiments where a synthetic transgene producing an mRNA with the 5’ UTR and coding sequence of nos but the 3’ UTR of bcd. When inserted into the fly genome this transgene will be associated with the anterior pole and thus will create an anterior to posterior NOS gradient in addition to the normal posterior to anterior gradient of the protein. This in essence produces embryos with two posterior poles in mirror image.
4
This double abdomen embryo arises because the NOS protein is now able to repress HB-M throughout the a-p axis.
Studying the BCD gradient: In order to determine the effect of BCD gradient in a-p positional information there are a few genetic experiments one can do.
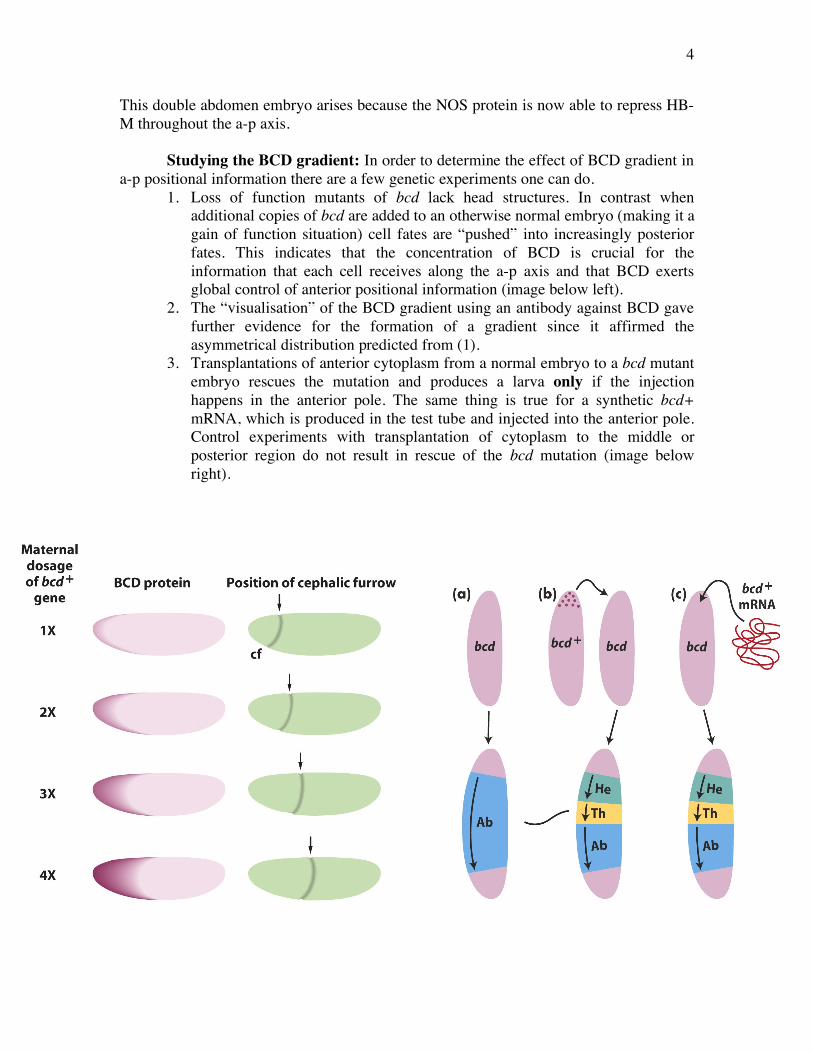
1. Loss of function mutants of bcd lack head structures. In contrast when additional copies of bcd are added to an otherwise normal embryo (making it a gain of function situation) cell fates are “pushed” into increasingly posterior fates. This indicates that the concentration of BCD is crucial for the information that each cell receives along the a-p axis and that BCD exerts global control of anterior positional information (image below left).
2. The “visualisation” of the BCD gradient using an antibody against BCD gave further evidence for the formation of a gradient since it affirmed the asymmetrical distribution predicted from (1).
3. Transplantations of anterior cytoplasm from a normal embryo to a bcd mutant embryo rescues the mutation and produces a larva only if the injection happens in the anterior pole. The same thing is true for a synthetic bcd+ mRNA, which is produced in the test tube and injected into the anterior pole. Control experiments with transplantation of cytoplasm to the middle or posterior region do not result in rescue of the bcd mutation (image below right).

5
A forth, crucial member: the mRNA of the gene caudal is distributed like hb-m uniformly across the embryo. A posterior to anterior gradient of the Caudal protein is established by inhibition of caudal protein synthesis by BCD. Since BCD concentration is low at the posterior of the embryo, Caudal protein concentration is highest there. Mutations in the gene caudal, result in abnormal development of abdominal segments.
Selected reading: a) Wolpert L et al, Principles of Development (Oxford University Press) chapter
5 pp145-150. b) St Johnston D and Nüsslein-Volhard C. The origin of pattern and polarity in
the Drosophila embryo. Cell 68: 201-219 (1992). c) St Johnston D. The intracellular localisation of messenger RNAs. Cell 81:
161-170 (1995). d) Driever W and Nüsslein-Volhard C. The bicoid protein determines position in
the Drosophila embryo in a concentration dependent manner. Cell 54: 95-104 (1988).
e) For further reading on the generality of RNA localization: Meignin C and Davis I (2009). Transmitting the message: intracellular mRNA localization Curr Op Cell Biol 22, 112-119.