Ermak et al Hp Gastritis Gastro 113 1118 1997
11
GASTROENTEROLOGY 1997;113:1118 – 1128 Gastritis in Urease-Immunized Mice After Helicobacter felis Challenge May Be Due to Residual Bacteria THOMAS H. ERMAK,* RU DING,* BRUCE EKSTEIN,* JOSEPH HILL, ‡ GWENDOLYN A. MYERS,* CYNTHIA K. LEE,* JACQUES PAPPO,* HAROLD K. KLEANTHOUS,* and THOMAS P. MONATH* *OraVax, Inc., Cambridge, Massachusetts; and ‡ Clemson Livestock Diagnostic Lab, Clemson University, Columbia, South Carolina Background & Aims: Oral immunization with recombi- in the gastric mucosa. Protection is associated with mild nant Helicobacter pylori urease (rUre) coadministered inflammation in the antrum characterized by the presence with a mucosal adjuvant protects mice against chal- of immunoglobulin A – secreting B cells and CD4 / /ab- lenge with Helicobacter felis. In this study, the duration TCR / T cells. 10 Several months after protection has been of protection and gastritis after challenge were charac- achieved, immunized mice may exhibit a lymphocytic terized at sequential time intervals up to 1 year. Meth- infiltration into the corpus, 7 raising questions as to ods: Outbred Swiss – Webster mice were orally immu- whether immunization was effective in resolving Helico- nized with rUre plus adjuvant and examined for the bacter-induced gastritis and if immunopathologic effects presence of H. felis infection and leukocyte infiltration were induced by the vaccine. into the gastric mucosa. Results: When defined by gas- H. pylori – induced gastritis in humans is characterized tric urease activity, 70% – 95% of rUre-immunized mice by infiltration of lymphocytes, plasma cells, and poly- were protected for between 2 and 57 weeks. Challenge morphonuclear leukocytes 11–13 and, in severe cases, is with H. felis increased the inflammatory response in the gastric mucosa of rUre-immunized mice, which also associated with epithelial changes consisting of parietal had elevated CD4 / and CD8 / T cells. The CD8 / cells cell loss, hyperplasia of the surface epithelium (foveolar represented a population of gastric intraepithelial cells, hyperplasia), and intestinal metaplasia. 14 Murine H. felis – which expressed the mucosal a E -integrin. Epithelial induced gastritis is characterized by a similar but primar- changes consisting of parietal cell loss and hyperplasia ily lymphocytic infiltrate as well as formation of microab- of the epithelium occurred in approximately 20% of the scesses and cystic glands. 4 mice. Antimicrobial triple therapy significantly de- In this study, the effect of rUre immunization on the creased the degree of gastritis and epithelial alteration duration of protection was examined for periods of up to in the stomach. Conclusions: These results indicate a year after challenge with H. felis, with special attention that oral immunization of mice with rUre produces a to the kinetics of antral and corpus gastritis, the pheno- long-lasting inhibition of H. felis infection but that resid- type of infiltrating T cells, and the epithelial cell compo- ual bacteria may produce a persistent lymphocytic in- filtration under these experimental conditions. sition of the gastric mucosa. We found that 70%–95% of mice were protected up to 57 weeks after challenge as defined by gastric urease activity. In addition, Ç50% H of rUre-immunized mice remained free of infection and elicobacter pylori is a gram-negative spiral bacterium that colonizes the gastric mucosa of humans, caus- had no gastritis. Approximately 10% of rUre-immunized ing chronic superficial gastritis and peptic ulcers; it has mice developed a chronic lymphocytic gastritis, charac- been implicated in adenocarcinoma and atrophy of the terized by both CD4 / and CD8 / T cells. However, clear- mucosa. 1,2 Helicobacter felis colonizes the gastric mucosa ance of H. felis by triple antimicrobial treatment after of mice and has been used as a convenient model for challenge prevented gastritis in all mice, implicating re- studying immunity in the development of oral vaccines sidual bacteria as the cause of lymphocytic infiltration against H. pylori. 3–6 Oral immunization with recombi- nant H. pylori urease (rUre) coadministered with a muco- after rUre immunization. sal adjuvant such as cholera toxin (CT) or Escherichia coli heat-labile enterotoxin (LT) induces a mucosal immune Abbreviations used in this paper: CT, cholera toxin; DAB, diamino- benzidine; HBSS, Hanks’ balanced salt solution; IEL, intraepithelial response, protects mice against challenge with H. felis, lymphocyte; LT, Escherichia coli heat-labile enterotoxin; rUre, recom- and clears bacteria from gastric tissue of previously in- binant Helicobacter pylori urease. fected mice. 7–10 Prophylactically immunized mice chal- q 1997 by the American Gastroenterological Association 0016-5085/97/$3.00 lenged by H. felis are protected from bacterial infection / 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro
-
Upload
thomas-ermak -
Category
Documents
-
view
383 -
download
2
Transcript of Ermak et al Hp Gastritis Gastro 113 1118 1997

GASTROENTEROLOGY 1997;113:1118–1128
Gastritis in Urease-Immunized Mice After Helicobacter felisChallenge May Be Due to Residual Bacteria
THOMAS H. ERMAK,* RU DING,* BRUCE EKSTEIN,* JOSEPH HILL,‡ GWENDOLYN A. MYERS,*CYNTHIA K. LEE,* JACQUES PAPPO,* HAROLD K. KLEANTHOUS,* and THOMAS P. MONATH**OraVax, Inc., Cambridge, Massachusetts; and ‡Clemson Livestock Diagnostic Lab, Clemson University, Columbia, South Carolina
Background & Aims: Oral immunization with recombi- in the gastric mucosa. Protection is associated with mildnant Helicobacter pylori urease (rUre) coadministered inflammation in the antrum characterized by the presencewith a mucosal adjuvant protects mice against chal- of immunoglobulin A–secreting B cells and CD4//ab-lenge with Helicobacter felis. In this study, the duration TCR/ T cells.10 Several months after protection has beenof protection and gastritis after challenge were charac- achieved, immunized mice may exhibit a lymphocyticterized at sequential time intervals up to 1 year. Meth- infiltration into the corpus,7 raising questions as toods: Outbred Swiss–Webster mice were orally immu- whether immunization was effective in resolving Helico-nized with rUre plus adjuvant and examined for the
bacter-induced gastritis and if immunopathologic effectspresence of H. felis infection and leukocyte infiltrationwere induced by the vaccine.into the gastric mucosa. Results: When defined by gas-
H. pylori–induced gastritis in humans is characterizedtric urease activity, 70%–95% of rUre-immunized miceby infiltration of lymphocytes, plasma cells, and poly-were protected for between 2 and 57 weeks. Challengemorphonuclear leukocytes11–13 and, in severe cases, iswith H. felis increased the inflammatory response in
the gastric mucosa of rUre-immunized mice, which also associated with epithelial changes consisting of parietalhad elevated CD4/ and CD8/ T cells. The CD8/ cells cell loss, hyperplasia of the surface epithelium (foveolarrepresented a population of gastric intraepithelial cells, hyperplasia), and intestinal metaplasia.14 Murine H. felis–which expressed the mucosal aE-integrin. Epithelial induced gastritis is characterized by a similar but primar-changes consisting of parietal cell loss and hyperplasia ily lymphocytic infiltrate as well as formation of microab-of the epithelium occurred in approximately 20% of the scesses and cystic glands.4mice. Antimicrobial triple therapy significantly de-
In this study, the effect of rUre immunization on thecreased the degree of gastritis and epithelial alterationduration of protection was examined for periods of up toin the stomach. Conclusions: These results indicatea year after challenge with H. felis, with special attentionthat oral immunization of mice with rUre produces ato the kinetics of antral and corpus gastritis, the pheno-long-lasting inhibition of H. felis infection but that resid-type of infiltrating T cells, and the epithelial cell compo-ual bacteria may produce a persistent lymphocytic in-
filtration under these experimental conditions. sition of the gastric mucosa. We found that 70%–95%of mice were protected up to 57 weeks after challengeas defined by gastric urease activity. In addition, Ç50%H of rUre-immunized mice remained free of infection and
elicobacter pylori is a gram-negative spiral bacteriumthat colonizes the gastric mucosa of humans, caus-
had no gastritis. Approximately 10% of rUre-immunizeding chronic superficial gastritis and peptic ulcers; it hasmice developed a chronic lymphocytic gastritis, charac-been implicated in adenocarcinoma and atrophy of theterized by both CD4/ and CD8/ T cells. However, clear-mucosa.1,2 Helicobacter felis colonizes the gastric mucosaance of H. felis by triple antimicrobial treatment afterof mice and has been used as a convenient model forchallenge prevented gastritis in all mice, implicating re-studying immunity in the development of oral vaccinessidual bacteria as the cause of lymphocytic infiltrationagainst H. pylori.3–6 Oral immunization with recombi-
nant H. pylori urease (rUre) coadministered with a muco- after rUre immunization.sal adjuvant such as cholera toxin (CT) or Escherichia coliheat-labile enterotoxin (LT) induces a mucosal immune Abbreviations used in this paper: CT, cholera toxin; DAB, diamino-
benzidine; HBSS, Hanks’ balanced salt solution; IEL, intraepithelialresponse, protects mice against challenge with H. felis,lymphocyte; LT, Escherichia coli heat-labile enterotoxin; rUre, recom-and clears bacteria from gastric tissue of previously in-binant Helicobacter pylori urease.
fected mice.7–10 Prophylactically immunized mice chal- q 1997 by the American Gastroenterological Association0016-5085/97/$3.00lenged by H. felis are protected from bacterial infection
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

ORAL IMMUNIZATION WITH RECOMBINANT UREASE 1119October 1997
pathologic, and immunohistochemical analyses. A longitudinalMaterials and Methodssegment including the antrum and corpus plus a piece of
Animals, Immunization, and Challenge attached intestine was fixed in 10% neutral buffered formalin,later routinely processed, embedded in paraffin, sectioned at 5Specific pathogen–free, 8-week-old outbred femalemm, and stained with H&E. For immunohistochemical analy-Swiss–Webster mice that were free of Helicobacter muridarumsis, gastric segments were mounted in O.C.T. embedding com-were obtained from Taconic Farms, Inc. (Germantown, NY).pound (Miles Scientific, Naperville, IL) and quick-frozen inAll procedures were conducted with approval of the OraVaxFreon 22 cooled to its freezing point by liquid nitrogen.Institutional Animal Care and Use Committee.
Mice were given four oral immunizations at weekly intervalsUrease Activityof 25–200 mg rUre (OraVax Development Section, OraVax,
Inc., Cambridge, MA) with either 10 mg CT or 5 mg LT The presence of H. felis in gastric tissue was assessedadjuvant in 0.2 mL phosphate-buffered saline (PBS) or adju- by urease activity measured spectrophotometrically using avant alone in PBS. rUre- and CT- or rUre- and LT-immunized colorimetric urease test. Antral segments (one quarter of themice not challenged with H. felis served as controls. Mice entire antrum) from each mouse were dissected free of corpuswere challenged 2–4 weeks after the last immunization with and intestine, placed in 0.5 mL of urea broth containing phenola single intragastric dose of 1 1 107 H. felis organisms (CS- red, and incubated as a whole piece of tissue for 4 hours at1 strain). This dose represents 103 times the 90%-infectious room temperature. Thereafter, samples were centrifuged, andintragastric dose.9 200 mL of the supernatant was used to determine the ab-
sorbance at 550 nm. The limit of sensitivity of this assay wasProtocols to Study Protection and previously determined to be Ç5 1 103 H. felis organisms.Gastritis Using CT or LT as Adjuvants
HistopathologyA 7-month study was performed to examine protectionBacterial colonization, gastritis, and epithelial changesand gastritis in groups of 10 rUre- and 10 CT-immunized
were evaluated on randomized, coded histological sections ofmice 2, 10, and 30 weeks after challenge. A larger, year-longgastric mucosa by an experienced veterinary pathologist (J.H.)study was also conducted using the more clinically relevantwho was blinded to the experimental design and number ofLT as a mucosal adjuvant. Groups of 25 rUre-immunized andanimal groups. H. felis organisms were visualized in gastric20 LT-immunized mice were challenged 4 weeks after immu-sections with a modified Steiner silver stain (Sigma).15 Thenization and examined 2, 10, 17, 34, and 57 weeks later.number of H. felis organisms per longitudinal section from the
Protocol to Study Gastric Intraepithelial intestine through the entire antrum and corpus was scored asLymphocytes follows: 0, no bacteria; 1, 1–20 bacteria; 2, 21–50 bacteria;
3, 51–100 bacteria; or 4, ú100 bacteria.Intraepithelial lymphocytes (IELs) were isolated fromFor evaluation of gastritis, H&E-stained sections weregastric tissue of mice immunized with rUre and either CT or
scored based on the intensity of the infiltration of lymphocytes,LT as an adjuvant and killed 4–6 months after challenge.plasma cells, and neutrophils.9 Grades were defined as follows:0, none; 1, a few leukocytes scattered in the deep mucosa; 2,Protocol to Examine the Role of Residualmoderate numbers of leukocytes in the deep to mid mucosaBacteria in Gastritisand occasional neutrophils in gastric glands (microabscesses);
Groups of 10 mice were immunized with 25 mg rUre 3, dense infiltrates in the mid to deep mucosa, a few microab-and 10 mg CT. Starting 2 weeks after challenge and continuing scesses, and one or two lymphoid aggregates; and 4, dense,through the 4th week after challenge, mice received 0.675 diffuse infiltrates throughout the lamina propria and into themg metronidazole, 1.5 mg tetracycline, and 0.185 mg Pepto- submucosa, with prominent lymphoid aggregates, and severalBismol (Procter and Gamble, Cincinnati, OH) daily for 14 microabscesses.days or no antimicrobial treatment at all. Control groups for The degree of epithelial change was scored as follows: 0,the experiment received (1) no immunization, challenge, and none; 1, small, focal areas of parietal cell loss in the corpus and/triple therapy, (2) no immunization, challenge, and no triple or hyperplasia of the surface epithelium; 2, epithelial changestherapy, or (3) CT immunization, challenge, and no triple throughout 75% of the mucosa; 3, epithelial changes through-therapy. Gastric tissue was sampled 10 and 20 weeks after out the mucosa plus one to three microabscesses or cysticchallenge and scored for gastritis and epithelial changes, and glands; or 4, epithelial changes throughout the mucosa plushalf the samples from rUre-immunized mice were processed four or more microabscesses or cystic glands.for immunohistochemical evaluation of T-cell subsets. Metaplasia of surface mucus cells was determined in histo-
logical sections stained with alcian blue. Control sections wereTissue Analysis stained with periodic acid–Schiff reagent. The degree of meta-
plasia was scored as follows: 0, none; 1, a few isolated cells orThe stomach was dissected along the lesser curvature,divided into quarters, and subjected to urease activity, histo- one focal area of alcian blue–positive mucous cells; 2, at least
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

1120 ERMAK ET AL. GASTROENTEROLOGY Vol. 113, No. 4
two focal areas of metaplasia in the surface epithelium or cystic Resultsglands; or 3, metaplasia throughout ú50% of the epithelium.
Duration of Protection, Histopathology,and T Cells Using CT as an AdjuvantAnalysis of T-Cell PhenotypeOral immunization with rUre and CT resulted inCryosections were blocked with avidin-biotin–
60% protection against H. felis infection through 30blocking reagents (Vector Laboratories, Burlingame, CA) andweeks after challenge, but 6 of 15 rUre-immunized micesequentially incubated with rat monoclonal antibodies againsthad gastritis scores of ¢3 (only 1 of 15 CT-immunizedmouse CD4 (clone RM4-5), CD8 (clone 53-6.7), aE-integrinmice had severe gastritis). Analysis of the CD4/ and(M290), b7 -integrin (clone M293), Vb2 (clone B20.6), Vb6
CD8/ cells from mice at 30 weeks after challenge showed(clone RR4-7), Vb7 (clone TR310), or Vb14 (clone 14-2) (Phar-mingen, San Diego, CA). Sections were then incubated with that rUre-immunized mice with gastritis scores of ¢3biotinylated rabbit anti-rat immunoglobulin G (Vector Labo- had CD4/ and CD8/ cells in the gastric mucosa, butratories), followed by horseradish peroxidase conjugated to an that CT-immunized mice had only CD4/ cells (data notavidin-biotin complex (Vector Laboratories), diaminobenzidine shown). Because of the small number of animals exam-(DAB), and methyl green. Control sections were incubated ined, a larger, quantitative experiment was performedwithout primary monoclonal antibody. The number of CD4/ using LT instead of CT as an adjuvant because LT hador CD8/ cells was counted by one of the authors (T.H.E.) in greater potential for eventual use in human vaccines.0.5-mm segment lengths of gastric mucosa and expressed persquare millimeter of field. Duration of Protection, Histopathology,
and T Cells Using LT as an AdjuvantTwo-Color Flow Cytometry of Gastric IELs
Duration of protection. The year-long study inGastric and intestinal tissues were incubated in Ca2/- rUre- plus LT-immunized mice was composed of four
and Mg2/-free Hanks’ balanced salt solution (HBSS) con- experimental groups: (1) rUre- plus LT-immunized andtaining ethylenediaminetetraacetic acid (EDTA), and the
H. felis–challenged; (2) LT-immunized and H. felis–dispersed IELs were purified by Percoll gradient centrifuga-
challenged; (3) rUre- plus LT-immunized but not chal-tion.16,17 Gastric and intestinal IEL were incubated simultane-lenged; and (4) LT-immunized but not challenged. Asously with fluorescein isothiocyanate–conjugated and biotin-determined by the absence of gastric urease activity,conjugated antibodies in V-bottomed microtiter plate wells70%–95% of mice immunized with rUre remained pro-(106 cells/well) and then with the appropriate dilution of strep-tected at various time intervals up to 57 weeks aftertavidin-conjugated phycoerythrin (Becton Dickinson, Moun-challenge (Figure 1A, rUre / LT group). In contrast,tain View, CA). The stained lymphocytes were fixed in 1%ú95% of LT-immunized mice remained infected at theparaformaldehyde and analyzed (10,000 cells) using an Epics
XL (Coulter Corp., Miami, FL), from which the percentage of maximal detectable level over this time interval (Figureeach lymphocyte subpopulation was determined. 1A, LT group). The kinetics of protection as defined
by histological examination for H. felis in gastric tissueDetection of Gastric Autoantibodies sections was similar (Figure 1B ). However, some mice
with negative gastric urease activity had low bacterialAcetone-fixed cryosections of gastric mucosa from nor-counts (õ20 bacteria per section compared with ú100mal mice were incubated with sera (1:100 dilution) from rUre-per section for infected controls) and, therefore, slightlyplus CT-immunized mice (n Å 8) killed 7 months after H.lower levels of full protection (30%–82%).felis challenge. Sections were subsequently incubated with
Histopathology. Unchallenged mice had only ahorseradish peroxidase–conjugated sheep anti-mouse immu-noglubulins (Amersham Corp., Arlington Heights, IL), fol- few infiltrating leukocytes in the gastric mucosa (Figurelowed by DAB and methyl green. Control sections were labeled 2). However, all mice challenged with H. felis had awithout the test sera. Unchallenged rUre- plus CT-immunized greater density of cells in the antrum, and rUre-immu-mice (n Å 4) and CT-immunized H. felis–challenged mice (n nized challenged mice had a greater infiltration in theÅ 4) at the same time interval served as experimental controls. corpus. Lymphocytes were the predominant cell type,
infiltrated the lamina propria from the muscularis mu-Statistics cosa to the surface epithelium, and sometimes formed
nodulelike aggregates in the deep mucosa and submu-Statistical analyses were performed with JMP softwarecosa. IELs were also found in the mucosa of rUre-immu-using Fisher’s Exact Test for dichotomous variables, the Wil-nized mice. Although infiltration of lymphocytescoxon’s/Kruskal–Wallis (rank sums) test for continuous or
ordinal variables, or linear regression. increased in response to challenge,Ç50% of rUre-immu-
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro
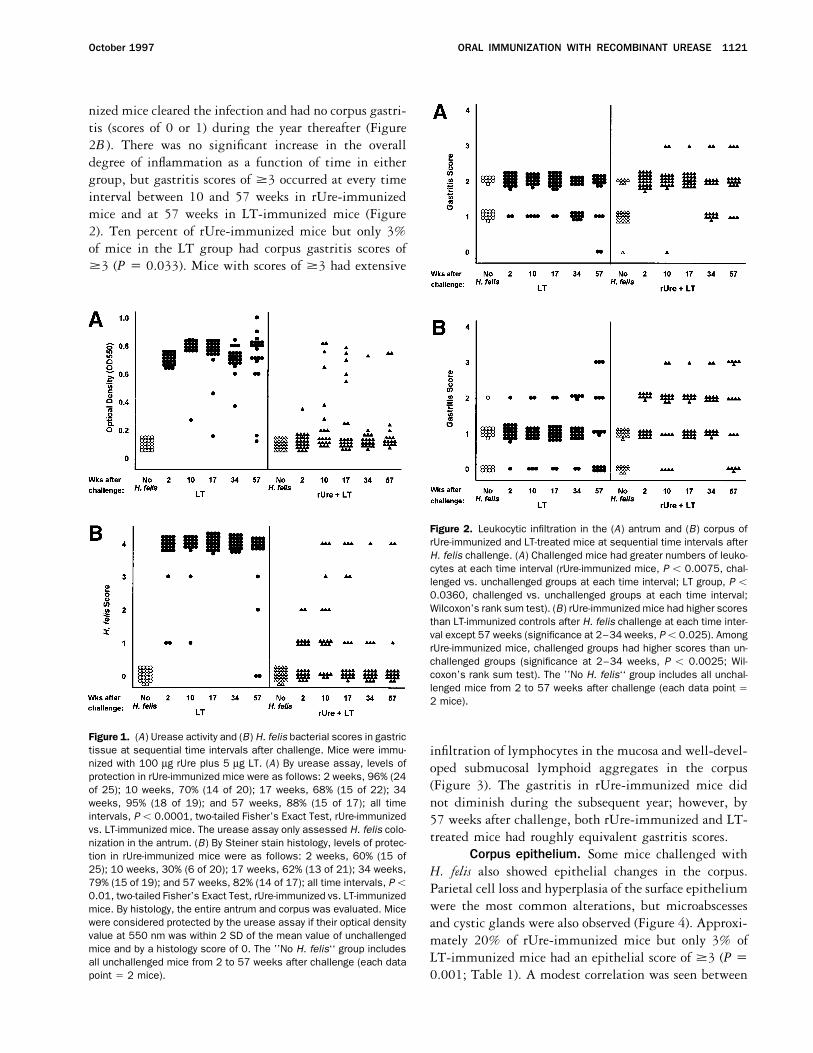
ORAL IMMUNIZATION WITH RECOMBINANT UREASE 1121October 1997
nized mice cleared the infection and had no corpus gastri-tis (scores of 0 or 1) during the year thereafter (Figure2B ). There was no significant increase in the overalldegree of inflammation as a function of time in eithergroup, but gastritis scores of ¢3 occurred at every timeinterval between 10 and 57 weeks in rUre-immunizedmice and at 57 weeks in LT-immunized mice (Figure2). Ten percent of rUre-immunized mice but only 3%of mice in the LT group had corpus gastritis scores of¢3 (P Å 0.033). Mice with scores of ¢3 had extensive
Figure 2. Leukocytic infiltration in the (A) antrum and (B) corpus ofrUre-immunized and LT-treated mice at sequential time intervals afterH. felis challenge. (A) Challenged mice had greater numbers of leuko-cytes at each time interval (rUre-immunized mice, P õ 0.0075, chal-lenged vs. unchallenged groups at each time interval; LT group, P õ0.0360, challenged vs. unchallenged groups at each time interval;Wilcoxon’s rank sum test). (B) rUre-immunized mice had higher scoresthan LT-immunized controls after H. felis challenge at each time inter-val except 57 weeks (significance at 2–34 weeks, Põ 0.025). AmongrUre-immunized mice, challenged groups had higher scores than un-challenged groups (significance at 2–34 weeks, P õ 0.0025; Wil-coxon’s rank sum test). The ’’No H. felis‘‘ group includes all unchal-lenged mice from 2 to 57 weeks after challenge (each data point Å2 mice).
Figure 1. (A) Urease activity and (B) H. felis bacterial scores in gastrictissue at sequential time intervals after challenge. Mice were immu- infiltration of lymphocytes in the mucosa and well-devel-nized with 100 mg rUre plus 5 mg LT. (A) By urease assay, levels of oped submucosal lymphoid aggregates in the corpusprotection in rUre-immunized mice were as follows: 2 weeks, 96% (24
(Figure 3). The gastritis in rUre-immunized mice didof 25); 10 weeks, 70% (14 of 20); 17 weeks, 68% (15 of 22); 34weeks, 95% (18 of 19); and 57 weeks, 88% (15 of 17); all time not diminish during the subsequent year; however, byintervals, P õ 0.0001, two-tailed Fisher’s Exact Test, rUre-immunized 57 weeks after challenge, both rUre-immunized and LT-vs. LT-immunized mice. The urease assay only assessed H. felis colo- treated mice had roughly equivalent gastritis scores.nization in the antrum. (B) By Steiner stain histology, levels of protec-
Corpus epithelium. Some mice challenged withtion in rUre-immunized mice were as follows: 2 weeks, 60% (15 of25); 10 weeks, 30% (6 of 20); 17 weeks, 62% (13 of 21); 34 weeks, H. felis also showed epithelial changes in the corpus.79% (15 of 19); and 57 weeks, 82% (14 of 17); all time intervals, Põ Parietal cell loss and hyperplasia of the surface epithelium0.01, two-tailed Fisher’s Exact Test, rUre-immunized vs. LT-immunized
were the most common alterations, but microabscessesmice. By histology, the entire antrum and corpus was evaluated. Micewere considered protected by the urease assay if their optical density and cystic glands were also observed (Figure 4). Approxi-value at 550 nm was within 2 SD of the mean value of unchallenged mately 20% of rUre-immunized mice but only 3% ofmice and by a histology score of 0. The ’’No H. felis‘‘ group includes
LT-immunized mice had an epithelial score of ¢3 (P Åall unchallenged mice from 2 to 57 weeks after challenge (each datapoint Å 2 mice). 0.001; Table 1). A modest correlation was seen between
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

1122 ERMAK ET AL. GASTROENTEROLOGY Vol. 113, No. 4
gastritis and epithelial scores in the corpus (r2 Å 0.528), areas of metaplasia only were found in the corpora ofmice that had been challenged with H. felis (Table 1).suggesting that perhaps the degree of leukocytic infiltra-
tion was a factor but not the only determinant of epithe- Metaplasia was found in surface epithelium (Figure 4C )and in epithelium lining cystic glands. Neither metapla-lial change.
Occasional metaplasia of surface mucous cells was sia in the antrum nor formation of intestinal-type gobletcells was evident in any of the mice.found in the corpora of all groups, but focal or extensive
CD4/ and CD8/ T cells. Only small numbers ofscattered CD4/ T cells were found in the mucosa of micenot challenged with H. felis (Figure 5). After challenge,however, larger numbers of CD4/ cells were apparent in
Figure 3. Lymphocytic infiltration in the corpus of H. felis–challengedmice. H&E-stained histological sections. (A) Unimmunized (LT) Figure 4. Epithelial alteration in the corpus of an H. felis–challenged,
rUre-immunized mouse (gastritis score, 3; epithelial score, 3). (A)mouse: gastritis score, 1; epithelial score, 0. Corpus is composed ofsurface epithelium (S), gastric pits lined by mucous and parietal cells H&E-stained histological section. Parietal and chief cells are de-
creased in cystic glands containing mucus and cell debris and lined(P), and chief cells (C) in the basal glands (original magnification 901).M, muscularis mucosa. (B and C) Gastritis, epithelial hyperplasia, and by hyperplastic epithelium (arrows) (original magnification 2101). (B)
Alcian blue–stained histological section. Mucous cell metaplasiadilated glands in rUre-immunized mouse: gastritis score, 3; epithelialscore, 3. The corpus shows extensive infiltration of lymphocytes in (arrows) in the corpus of rUre-immunized mice (metaplasia score, 2).
The surface epithelium contains alcian blue–reactive mucus (originallamina propria (asterisks), submucosal lymphoid aggregates (L), andparietal cell loss (original magnification 901). magnification 2101).
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

ORAL IMMUNIZATION WITH RECOMBINANT UREASE 1123October 1997
Table 1. Alteration of Gastric Epithelium in the Corpus of found in protected rUre-immunized mice that did notrUre- Plus LT-Immunized and LT-Immunized Mice at develop a lymphocytic gastritis and in unprotected LT-Sequential Time Intervals After H. felis Challenge immunized mice (Figures 2 and 5).
Weeks after challenge Expression of mucosal integrins. To ascertainwhether gastric T cells belong to a mucosal or a systemic
Group 2 10 17 34 57 lineage, gastric tissue was screened for the presence ofEpithelial score ¢3a mucosal integrins. The aE- and b7 -integrins are present
rUre / LT/H. felis challenge 6/25 3/20 3/22 3/19 6/17 as part of the aEb7 heterodimer on intestinal IELs18,19
LT/H. felis challenge 0/24 0/23 0/24 1/24 3/21and mediate adhesion with E-cadherin on epithelialrUre / LT 0/10 0/9 0/8 0/9 0/7
LT 0/10 0/10 0/7 0/8 0/7 cells.20,21 The b7 -integrin is also present as part of theMetaplasia score ¢2b
a4b7 Peyer’s patch homing receptor.22 In rUre-immu-rUre / LT/H. felis challenge 2/25 3/20 5/22 5/19 6/17
nized mice, b7 -integrin–positive cells were localizedLT/H. felis challenge 0/24 0/23 0/24 1/24 3/21throughout the mucosa and were more numerous than
aEpithelial score ¢3 defined as hyperplasia of surface epithelium oreither CD4/ or CD8/ cells, whereas aE-integrin–posi-parietal cell loss throughout the mucosa plus 1–3 microabscessestive cells had a similar distribution to, but a slightlyor cystic glands.
bMetaplasia ¢2 defined as alcian blue–positive cells in 2 or more higher density than, CD8/ cells (Table 2). Gastric aE-focal areas of surface epithelium or cystic glands. Unchallenged rUre integrin–positive cells were also found in LT-immunized/ LT and LT mice had no metaplasia scores ¢2.
mice challenged with H. felis, indicating that a popula-tion of gastric CD80 cells (20%–40%) also expressedaE-integrin (Table 2).
To examine whether CD8/ cells in gastric tissue repre-the gastric mucosa of rUre-immunized mice comparedsented a population of aE-integrin–positive IELs, iso-with the other experimental groups (Figure 5A and B ).lated gastric and intestinal CD8/ IELs were examinedCD4/ cells localized in the lamina propria between gas-by flow cytometry (Figure 8). Two-color plots of intesti-tric pits, in the subglandular regions of the deep mucosanal and gastric IELs were very similar, and Ç90% ofabove the muscularis mucosa, and in submucosalgastric CD8/ IELs expressed aE-integrin.lymphoid aggregates below the muscularis mucosa (Fig-
TCR b-chain usage. An analysis of TCR b-chainure 6A and B ). In the corpus, the highest number ofexpression by T cells in gastric tissue (Vb2 , Vb6 , Vb7 , andCD4/ cells occurred in rUre-immunized mice 17–57Vb14 ) revealed a significant infiltration of Vb6
/ cells in theweeks after challenge, with a maximum density moregastric mucosa of rUre-immunized but not LT-immunizedthan four times greater than that observed in the LT-mice. Vb6
/ cells had a similar distribution as CD8/ cells,immunized group (Figure 5).were on the average only half as numerous, and wereCD8/ cells were rare in the mucosa of all mice thatabsent if CD8/ cells were not present (Table 2).were not challenged and in LT-immunized mice chal-
lenged with H. felis (Figure 5C and D ). In rUre-immu-Role of Residual Bacteria in Production ofnized mice, however, CD8/ cells were noted in the an-Gastritistrum of Ç75% and in the corpus of ú85% after
challenge with H. felis. The highest numbers of CD8/ Tests were performed to examine whether autoim-munity or impurities in the urease might be responsiblecells were found between 10 and 57 weeks after chal-
lenge, and the density of CD8/ cells and CD4/ cells for the leukocytic infiltration. Analysis of sera from rUre-immunized mice at 7 months after challenge (8 micecorrelated in the corpus (r2 Å 0.681) but not the antrum
(r2 Å 0.116). CD8/ cells infiltrated the lamina propria, receiving CT as an adjuvant) did not reveal anti–parietalcell antibodies reactive with normal mouse gastric tissue.were prominent within the epithelium as IELs (Figure 6C
and D ), and were also present in submucosal lymphoid Likewise, immunization of mice with E. coli GroEL (apotential rUre contaminant) and CT was not protectiveaggregates.
Protected vs. unprotected rUre-immunized mice. and produced no greater gastritis than CT treatment overa 10-week observation period after challenge (data notAn analysis of unprotected vs. protected rUre-immunized
mice pooled from time intervals from 10 to 57 weeks shown).Because significant lymphocytic infiltration was ob-revealed that no unprotected mice had elevated gastritis
scores (no gastritis scores ¢3 or T cells in either the served only in mice with no or only a few H. felis organ-isms (õ20 per section), we examined whether residualantrum or corpus) (Figure 7). Thus, T-cell infiltration in
rUre-immunized unprotected mice was similar to that bacteria below the level of detection of the urease assay
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

1124 ERMAK ET AL. GASTROENTEROLOGY Vol. 113, No. 4
Figure 5. Kinetics of CD4/ cell and CD8/ cell infiltration in the (A and C) antrum and (B and D) corpus of H. felis–challenged mice. (A and B)rUre-immunized mice (protected only) had greater numbers of CD4/ T cells than LT-control mice (P õ 0.02; antrum CD4/ cells at 2, 10, and34 weeks and corpus at 34 weeks, rUre / LT vs. LT). (C and D) rUre-immunized mice had elevated CD8/ cell counts in gastric mucosa ateach time interval from 10 through 57 weeks (P õ 0.01; rUre / LT vs. LT). CD8/ cells were rare to absent in unchallenged rUre-immunizedmice and LT-treated mice challenged with H. felis. P values given in the figure are for all mice from 10 to 57 weeks after H. felis challenge(rUre / LT vs. LT).
or histological study might be responsible for gastritis. that were given either CT or no immunization, and hada higher score (P Å 0.0387) (only in the corpus) thanrUre-immunized mice that received CT as the adjuvant
were used in this study based on the observation that mice that received no immunization or adjuvant beforeantimicrobials (data not shown). Likewise, the densityCT produced slightly greater lymphocytic infiltration
than LT. For antimicrobial treatment, mice were given of CD4/ and CD8/ cells infiltrating the antrum wassignificantly reduced (P õ 0.01) by antimicrobial treat-14 twice-daily treatments of metronidazole, tetracycline,
and Pepto-Bismol between 2 and 4 weeks after H. felis ment, and the induction of CD4/ and CD8/ cells in thecorpus was prevented (0/9 in antimicrobial-treated micechallenge. This time interval was chosen to allow lym-
phocytes time to infiltrate the antrum and inhibit bac- vs. 3/11 in rUre-immunized mice that received no ther-apy; significance not achieved) (Table 3).terial colonization before antimicrobial treatment. By
10–20 weeks after challenge, mice that underwent anti-Discussionmicrobial therapy had significantly lower gastritis (P õ
0.01) and epithelial scores (P õ 0.05) than mice under- The present study indicates that immunization ofmice with rUre produces a long-lasting reduction in H.going no antimicrobial treatment (Table 3), had gastritis
or epithelial scores equivalent to those of challenged mice felis colonization of gastric mucosa up to 1 year after
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

ORAL IMMUNIZATION WITH RECOMBINANT UREASE 1125October 1997
Figure 6. Distribution of T cells in the gastric mucosa of rUre-immunized mouse with gastritis and epithelial scores of 3. Cryosection of thecorpus 57 weeks after challenge with H. felis. (A) CD4/ T cells localize in the lamina propria below surface epithelium, between gastric pits,adjacent to the muscularis mucosa (M), and in deep mucosal lymphoid aggregates (L). CD4/ cell counts in this mouse were 140 cells/mm2
field. (B) Higher-power view showing CD4/ T cells in the lamina propria (asterisk). (C) CD8/ T cells localize in the gastric epithelium and inthe lamina propria between gastric pits. CD8/ cell counts in this mouse were 160 cells/mm2 field. (D) Higher-power view showing CD8/ IELs(arrows) in the surface epithelium. (Original magnifications: A and C, 901; B, 1801; D, 3601.)
challenge. H. pylori urease, a major protein required for proportion of LT-immunized mice infected with H. felis,although with less T-cell infiltration. The gastritis pro-initial colonization of the gastric mucosa,23–26 has been
shown to be an active immunogen in protecting mice duced by H. felis infection was similar to that observedin humans infected with H. pylori in terms of infiltrationagainst H. felis infection.7–10 Approximately 50% of
rUre-immunized mice that were subsequently challenged of CD4/ and CD8/ lymphocytes, loss of parietal cells,and hyperplasia of the surface epithelium.27–29 However,with H. felis remained uninfected during the year thereaf-
ter and had no significant gastritis. Challenge with H. the infiltration of T cells was deeply mucosal rather thansurface localized. This difference may be related to thefelis increased lymphocyte counts in the gastric antrum,
which is the location in the stomach where the greatest panmucosal colonization by H. felis compared with thesurface epithelial colonization by H. pylori.3,5,13,30 How-H. felis colonization occurs in the absence of immuniza-ever, it may be a characteristic feature of murine gastritis,tion.3–6 After immunization, infiltration of lymphocytesbecause a similar deep mucosal to submucosal pattern isinto the antrum probably represents a desirable, vaccine-present in mice with non–Helicobacter-related gastritis.31induced response to infection.4,10 However, antral gastri-
tis did not resolve over time, and a marked lymphocytic Epithelial changes in the corpus consisted primarily ofparietal cell loss and hyperplasia of the surface epitheliumgastritis developed in the corpus during the 10 weeks
after challenge inÇ15% of the mice and persisted during (foveolar hyperplasia), but microabscesses and cysticglands were also observed. Similar changes were alsothe subsequent year. By the end of the year-long study,
a similar gastritis also developed in roughly the same found in chronically H. felis–infected mice from this as
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

1126 ERMAK ET AL. GASTROENTEROLOGY Vol. 113, No. 4
pus and in preventing epithelial changes in the corpus toa level comparable with that found in controls. Likewise,CD4/ and CD8/ cell counts were significantly reducedin the antrum. The few cells still remaining in the gastricmucosa of rUre-immunized mice after treatment mayhave been present because the antimicrobial regimen wasinitiated 2 weeks after challenge (a time when an immuneresponse against the organism was already established),and by the end of the study, the infiltration had notcompletely resolved.
Increased CD8/ cells in the gastric mucosa may berelated to the presence of residual bacteria at the apicalsurface or within epithelial cells. Gastric CD8/ cells per-sisted after challenge, indicating that the stimulus re-mained in the gastric tissue over time, but were reducedafter antimicrobial treatment. Intestinal IELs are believed
Figure 7. T-cell infiltration in protected vs. unprotected rUre-immu-to play a role in immune surveillance against epithelialnized H. felis–challenged mice. Unprotected mice were available at
10 (n Å 4), 17 (n Å 4), and 57 (n Å 2) weeks after challenge. injury by recognizing abnormal peptides derived fromCD4/ cell and CD8/ gastritis did not develop in the corpus of any infectious agents and expressed on epithelial cells.36 Per-unprotected mice (significant differences from protected mice were
haps gastric IELs function in a similar manner.not achieved). P values compare T-cell populations in Not Protectedvs. Protected groups. Protected mice were defined as having a urease Gastritis in the corpus did not appear to be the conse-assay optical density value at 550 nm within 2 SD of the value in quence of autoantibody production against parietal cells.control mice (õ0.2); Not Protected mice were defined as having a
No reactivity was observed in this study or another studyurease assay optical density value at 550 nm of ú0.5 or a bacterialin which mice were immunized with either urease A orscore of ¢3.
B subunits.37 The T-cell response was not limited to thecorpus but included the antrum as well. Furthermore,CD8/ IELs do not appear to be a component of murinewell as other studies.3,32 Mucous cell metaplasia in H.autoimmune gastritis, where anti–parietal cell autoanti-felis–challenged mice occurred in the surface epitheliumbodies can be detected in sera.31
of the corpus and appeared distinct from H. pylori–associ-It is not clear why corresponding levels of gastritisated metaplasia in humans, where the distribution is
were not observed in unprotected rUre-immunized micecharacteristically antral and mucous cells can undergoif residual bacteria per se are responsible for gastritis. Incomplete metaplasia into intestinal-type goblet cells.33
these mice, bacterial infection was high, but a significantGastritis in rUre-immunized mice was characterizedlymphocytic infiltration was not produced. One factor inby the presence of both CD4/ and CD8/ T cells, al-murine inflammation that may account in part for thisthough initially (at 2 weeks) it involved primarily CD4/
cells. In other studies, these CD4/ cells have been shownto primarily express a,b-TCR.10 T cells expressed the
Table 2. Relative Proportion of T Cells in Gastric Mucosa ofmucosal homing integrin b7 , which is present on a4b7/
a Subset of H. felis–Challenged Micelymphocytes homing to mucosal tissues.18,19,22,34 Virtu-rUre / LT LTally all CD8/ cells represented gastric aE-integrin–posi-
tive IELs. The aE-integrin is present on intestinal IELsT-cell antigen Antrum Corpus Antrum Corpus
as part of aEb7 , the ligand for E-cadherin on epithelialCD4 (RM4-5) 181 { 19 252 { 37 104 { 16 77 { 5cells, but not on peripheral T cells.18,19,22 A subset ofCD8a (53-6.7) 96 { 17 197 { 22 õ1 õ1gastric CD8/ cells also appeared to express Vb6 , whichb7-Integrin (M293) 269 { 33 406 { 54 85 { 16 44 { 8
is found at relatively high frequency on intestinal IELs aE-Integrin (M290)a 147 { 21 271 { 31 44 { 7 15 { 4Vb6 (RR4-7) 49 { 15 91 { 30 õ2 õ2bearing homodimeric CD8a chains.35 Together, these
data support the hypothesis that gastric CD8/ cells rep- NOTE. 8 rUre-immunized mice with the highest CD4/ cell and CD8/
cell counts and 8 LT-treated mice with the highest CD4/ cell countsresent a population of cells in the stomach correspondingwere analyzed for expression of b7 -integrin, aE-integrin, and Vb6. T-to or derived from intestinal IELs.cell counts made in serial sections.
Triple antimicrobial treatment was effective in reduc- aThe T-cell region of the spleen contained only scattered aE-integrin–positive cells, which represented õ10% of CD8/ cells.ing lymphocytic infiltration in both the antrum and cor-
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

ORAL IMMUNIZATION WITH RECOMBINANT UREASE 1127October 1997
Figure 8. Expression of aE-inte-grin on purified gastric CD8/
IELs as determined by two-colorflow cytometry. More than 90%of gastric CD8/ IELs expressedaE-integrin. (A) Gastric and (B)intestinal IELs were isolated byincubating tissue in Ca2/,Mg2/–free EDTA-HBSS me-dium and purified by Percoll gra-dient centrifugation. IELs werepooled from 10 rUre- plus CT-immunized mice.
observation is host responsiveness. It has been shown that A comparison of H. pylori vaccines in murine H. felismodels indicates that rUre is among the most efficacioussome mouse strains, such as BALB/c, develop low-grade
inflammation in response to H. felis, whereas other single Helicobacter protein examined to date.8,39 Oral im-munization with rUre produces a long-lasting reductionstrains, such as C57BL/6, develop high-grade inflamma-
tion.38 In our experimental model using outbred mice, in H. felis organisms, but sterilizing immunity may notbe achieved under these experimental conditions, re-bacterial density in itself does not appear to determine
the inflammatory response. In addition to host factors, sulting in the development of a persistent T-cell gastritisin Ç10%–20% of cases. This response does not seembacterial virulence factors, which are poorly understood
in H. felis, may also contribute to inflammation. In the to be unique to rUre, because a mixture of Helicobacterantigens may also produce a persistent inflammatory re-case of a chronic infection such as H. felis, genetic diversi-
fication or selection may occur over time, leading to sponse.8 Further studies are under way to determinewhether these phenomena occur in nonmurine hosts anddifferent levels of inflammation.to develop vaccines capable of eliminating bacteria fromthe gastric mucosa.
Table 3. Effect of Antimicrobial Triple Therapy onHistopathology in the Antrum and Corpus of rUre- ReferencesImmunized, H. felis–Challenged Mice
1. Blaser MJ. Helicobacter pylori and the pathogenesis of gastrodu-Parameter rUre / CT rUre / CT / Rx Pa
odenal inflammation. J Infect Dis 1990;161:626–633.2. Cover TL, Blaser MJ. Helicobacter pylori and gastroduodenal dis-
Antrumease. Annu Rev Med 1992;43:135–145.
Median gastritis score 2 (1–3) 1 (1–2)3. Fox JG, Lee A, Otto G, Taylor NS, Murphy JC. Helicobacter felis
(range) (n Å 20) (n Å 20) õ0.01gastritis in gnotobiotic rats: an animal model of Helicobacter
Median epithelial 1 (0–3) 0 (0–1)pylori gastritis. Infect Immun 1991;59:785–791.
score (range) (n Å 20) (n Å 20) õ0.014. Fox JG, Blanco M, Murphy JC, Taylor NS, Lee A, Kabok Z, PappoCD4/ cells 130 { 45 50 { 26
J. Local and systemic immune responses in murine Helicobacter(mean { SD) (n Å 11) (n Å 9) õ0.01felis active chronic gastritis. Infect Immun 1993;61:2309–CD8/ cells 78 { 65 3 { 32315.(mean { SD) (n Å 11) (n Å 9) õ0.01
5. Lee A, Fox JG, Otto G, Murphy J. A small animal model of humanCorpusHelicobacter pylori active chronic gastritis. GastroenterologyMedian gastritis score 2 (0–3) 0 (0–1)1990;99:1315–1323.(range) (n Å 20) (n Å 20) õ0.01
6. Chen M, Lee A, Hazell SL. Immunization against HelicobacterMedian epithelial 1 (0–3) 0 (0–1)infection in a mouse/Helicobacter felis model. Lancet 1992;score (range) (n Å 20) (n Å 20) 0.04339:1120–1121.CD4/ cells 122 { 116 54 { 29
7. Michetti P, Corthesy–Theulaz I, Davin C, Haas R, Vaney A–C,(mean { SD) (n Å 11) (n Å 9) NSHeitz M, Bille J, Kraehenbuhl J–P, Saraga E, Blum AL. Immuniza-CD8/ cells 135 { 228 7 { 3tion of BALB/c mice against Helicobacter felis infection with Heli-(mean { SD) (n Å 11) (n Å 9) NSb
cobacter pylori urease. Gastroenterology 1994;107:1002–1011.aWilcoxon’s rank sum test.
bP õ 0.07. 8. Ferrero RL, Thiberge JM, Huerre M, Labigne A. Recombinant anti-
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro

1128 ERMAK ET AL. GASTROENTEROLOGY Vol. 113, No. 4
gens prepared from the urease subunits of Helicobacter ssp.: colonization of gnotobiotic piglets by Helicobacter pylori. J MedMicrobiol 1992;37:123–127.evidence of protection in a mouse model of gastric infection.
Infect Immun 1994;62:4981–4989. 26. Hu L-T, Mobley HLT. Purification and N-terminal analysis of ureasefrom Helicobacter pylori. Infect Immun 1990;58:992–998.9. Lee CK, Weltzin R, Thomas WD, Jr., Kleanthous H, Ermak TH,
Soman G, Hill JE, Ackerman SK, Monath TP. Oral immunization 27. Bayerdorffer E, Oertel H, Lehn N, Kasper G, Mannes GA, Sauer-bruch T, Stolte M. Topographic association between active gastri-with recombinant Helicobacter pylori urease induces secretory
IgA antibodies and protects mice from challenge with Helico- tis and Campylobacter pylori colonization. J Clin Pathol 1989;42:834–839.bacter felis. J Infect Dis 1995;172:161–172.
10. Pappo J, Thomas WD, Jr., Kabok Z, Taylor NS, Murphy JC, Fox JG. 28. Scolnick JV, Tompkins LS. Helicobacter pylori and gastroduode-nal disease: pathogenesis and host-parasite interaction. InfectEffect of oral immunization with recombinant urease on murine
Helicobacter felis gastritis. Infect Immun 1995;63:1246–1252. Agents Dis 1993;1:294–309.29. Stolte M, Eidt S, Ohnsmann A. Differences in Helicobacter pylori11. Genta RM, Lew GM, Graham DY. Changes in the gastric mucosa
following eradication of Helicobacter pylori. Mod Pathol 1993;6: associated gastritis in the antrum and body of the stomach. ZGastroenterol 1990;28:229–233.281–289.
12. Carpenter HA, Talley NJ. Gastroscopy is incomplete without bi- 30. Dubois A, Fiala N, Heman-Ackah LM, Drazek ES, Tarnawski A,Fishbein WN, Perez-Perez GI, Blaser MJ. Natural gastric infectionopsy: clinical relevance of distinguishing gastropathy from gastri-
tis. Gastroenterology 1995;108:917–924. with Helicobacter pylori in monkeys: a model for spiral bacterialinfection in humans. Gastroenterology 1994;106:1405–1417.13. Sipponen P, Siurala M, Goodwin CS. Histology and ultrastructure
of Helicobacter pylori infections: gastritis, duodenitis, and peptic 31. Sakaguchi S, Ermak TH, Toda M, Berg LJ, Ho W, de St. Groth BF,Peterson PA, Sakaguchi N, Davis MM. Induction of autoimmuneulceration, and their relevance as precancerous conditions. In:
Goodwin CS, Worsley BW, eds. Helicobacter pylori: biology and disease in mice by germline alteration of the T cell receptor geneexpression. J Immunol 1994;152:1471–1484.clinical practice. Boca Raton, FL: CRC, 1993:37–62.
14. Owen DA, Kelley JK. Atlas of gastrointestinal pathology. Philadel- 32. Fox JG, Li X, Cahill RA, Andruitis K, Rustgi AK, Odze R, Wang TC.Hypertrophic gastropathy in Helicobacter felis–infected wild-typephia: Saunders, 1994.
15. Garvey W, Fathi A, Bigelow F. Modified Steiner for the demonstra- C57BL/6 mice and p53 hemizygous transgenic mice. Gastroen-terology 1996;110:155–166.tion of spirochetes. J Histotechnol 1985;8:15–17.
16. Lefrancois L, Lycke N. Isolation of mouse small intestinal intraep- 33. Jass JR, Strudley I, Faludy J. Histochemistry of epithelial metapla-sia and dysplasia in human stomach and colorectum. Scand Jithelial lymphocytes, Peyer’s patch, and lamina propria cells. In:
Coligan JE, Kruisbeek AM, Margulies DH, Shevach EM, Strober Gastroenterol Suppl 1984;104:109–130.34. Berlin, C, Berg EL, Briskin MJ, Andrew DP, Kilshaw PJ, HolzmannW, eds. Current protocols in immunology. New York: Wiley, 1995:
3.19.1–3.19.16. B, Weissman IL, Hamann A, Butcher EC. a4b7 integrin mediateslymphocyte binding to the mucosal vascular addressin MAdCAM-17. Harriman GR, Lycke NY, Elwood LJ, Strober W. T lymphocytes
that express CD4 and the alpha beta–T cell receptor but lack 1. Cell 1993;74:185–195.35. Rocha P, Vassalli, Guy-Grand D. The Vb repertoire of mouse gutThy-1. Preferential localization in Peyer’s patches. J Immunol
1990;145:2406–2414. homodimeric CD8/ intraepithelial T cell receptor a/b/ lympho-cytes reveals a major extrathymic pathway of T cell differentia-18. Kilshaw PJ, Murant SJ. A new surface antigen on intraepithelial
lymphocytes in the intestine. Eur J Immunol 1990;20:2201– tion. J Exp Med 1991;173:483–486.36. Blumberg RS, Balk SP. Recognition of intestinal epithelial cell2207.
19. Roberts K, Kilshaw PJ. The mucosal T cell integrin aM290b7 ligands by T-cells. Mucosal Immun Update 1994;2:3–5.37. Claeys D, Corthesy-Theulaz I, Gaudin M, Saraga E, Porta N,recognizes a ligand on mucosal epithelial cell lines. Eur J Immu-
nol 1993;23:1630–1635. Kraehenbuhl J-P, Blum AL, Michetti P. Immunization with H. pyloriurease do not induce serum auto-antibodies against the gastric20. Karecla PI, Bowden SJ, Green SJ, Kilshaw PJ. Recognition of e-
cadherin on epithelial cells by the mucosal T cell integrin mucosa in mice. Gut 1994;37(Suppl 1):A93.38. Sakagami T, Dixon M, O’Rourke J, Howlett R, Alderuccio F, VellaaM290b7 (aEb7). Eur J Immunol 1995;25:852–856.
21. Cepek KL, Shaw SK, Parker CM, Russell GJ, Morrow JS, Rimm J, Shimoyama T, Lee A. Atrophic gastric changes in both Helico-bacter felis and Helicobacter pylori infected mice are host depen-DL, Brenner MB. Adhesion between epithelial cells and T lympho-
cytes mediated by e-cadherin and the aEb7 integrin. Nature 1994; dent and separate from antral gastritis. Gut 1996;39:639–648.39. Marchetti M, Arico B, Burroni D, Figura N, Rappuoli R, Ghiara P.372:190–193.
22. Kilshaw PJ, Murant SJ. Expression and regulation of b7(bp) inte- Development of a mouse model of Helicobacter pylori infectionthat mimics human disease. Science 1995;267:1655–1658.grins on mouse lymphocytes: relevance to the mucosal immune
system. Eur J Immunol 1991;21:2591–2597.23. Tsuda M, Karita M, Morshed MG, Okita K, Nakazawa T. A urease-
negative mutant of Helicobacter pylori constructed by allelic ex- Received November 22, 1996. Accepted July 1, 1997.Address requests for reprints to: Thomas H. Ermak, Ph.D., OraVax,change mutagenesis lacks the ability to colonize the nude mouse
stomach. Infect Immun 1994;62:3586–3589. Inc., 38 Sidney Street, Cambridge, Massachusetts 02139. Fax:(617) 494-0927.24. Eaton, KA, Krakowka S. Effect of gastric pH on urease-dependent
colonization of gnotobiotic piglets by Helicobacter pylori. Infect The authors thank Timothy Tibbitts, Jennifer Bakios, KathleenGeorgakopoulos, and Heather Gray for excellent technical assis-Immun 1994;62:3604–3607.
25. Eaton, KA, Morgan DR, Krakowka S. Motility as a factor in the tance.
/ 5e21$$0043 09-09-97 13:00:34 gasas WBS-Gastro