
Epistatic interaction andfunctional ...
5
Proc. Natl. Acad. Sci. USA Vol. 91, pp. 7917-7921, August 1994 Genetics Epistatic interaction and functional compensation between the two tissue- and cell-specific sucrose synthase genes in maize (isozymes/posttranscriptional control/cross-regulation) PREM S. CHOUREY AND EARL W. TALIERCIO U.S. Department of Agriculture, Agricultural Research Service, and Departments of Agronomy and Plant Pathology and Program in Plant Molecular and Cellular Biology, University of Florida, Gainesville, FL 32611-0680 Communicated by Oliver E. Nelson, Jr., May 9, 1994 ABSTRACT A tissue-specific epistatic mode of gene inter- action was observed between molecularly homologous genes Shl and Susi (hereafter, Sh and Sus), encoding the sucrose synthase (SS) isozymes, SS1 and SS2, respectively. In Sh Sus genotype, both SS genes were expressed simultaneously and approximately equally in young seedlings; however, only the Sus-encoded SS2 protein was seen in the developing embryos. By contrast, the mutant sass genotype, lacking detectable levels of the SS2 protein in various tissues tested, showed expression of the Sh locus as judged by the detection of the SS1 protein in such embryos. Ectopic expression in embryos was seen from two separate Sh alleles, Sh-W22 and Sh'-5 (a revertant allele derived upon Ds excision from sh-m5933). In each case, the Sh expression at the protein level in embryos was unique to genotypes with the mutant sus gene. Based on the observed lack of phenotypic change in the sus mutant, we suggest that the ectopic expression of the Sh in otherwise Sus-specific tissues leads to functional compensation. There was no epistatic in- teraction of Sh and Sus at the RNA level as SS1 transcripts were detectable in both Sus and sus embryos. Thus, embryo speci- ficity between the two SS genes was determined at posttran- scriptional or at translational level of control. We surmise on the basis of these data that metabolic regulatory controls seem to override the normal constraints of tissue and cell specificity of the nonallelic isozyme genes to maintain efficient use of the pathways. Genetic redundancy in higher eukaryotes is well docu- mented. Duplicate or molecularly homologous genes encod- ing biochemically similar gene products are now a common phenomenon in both of the model plant systems, Arabidop- sis and maize. Mutational inactivation of a gene invariably uncovers another nonallelic gene encoding a similar function. Maize, generally considered a true diploid, is well analyzed for duplicate genes (1) and molecularly homologous se- quences (2). Biochemical genetic studies on developing maize endosperms indicate that many starch mutants asso- ciated with the loss of a specific enzyme have unaltered levels of the corresponding isozyme expressed either in the same tissue or in another tissue such as embryo and other sporo- phytic organs (3-6). Recent analyses in Arabidopsis, partic- ularly in amino acid biosynthesis, have similarly shown that genetic redundancy is a rule rather than exception for several enzymes in this pathway (7). Duplicated genes usually have distinct, nonoverlapping physiological functions that are dif- ficult to resolve by biochemical approaches alone. The com- bined use of genetic and molecular approaches is of particular use in the elucidation of their functional significance. In this regard, the two sucrose synthase (SS) genes in maize provide a model system. The enzyme plays an impor- tant role in energy metabolism by mobilizing sucrose into diverse pathways relating to metabolic, structural, and stor- age functions of plant cells. The two biochemically similar isozymes, SS1 and SS2, are encoded by the shrunken (Sh) and sucrose synthase (Sus) loci, respectively (8, 9). Both Sh and Sus have been cloned and shown to be molecularly homologous (10-12). A similar two-gene system for the two SS isozymes is also described in wheat, sorghum, sugarcane, and several dicot species, including Arabidopsis (13). In maize, the SS1 isozyme is present in abundant levels in developing endosperm. The loss of this protein, as in the case of the shrunken (sh) mutant, is associated with several alterations in developing endosperm, including a reduction in the amount of starch (4), an early cell degeneration (14), and a reduction in the number of protein bodies (unpublished data). There is otherwise no phenotypic change in the sh plant, even though Sh expression is detected in several tissues and cells of young plants of the wild-type genotype (15-17). The Sus-encoded SS2 protein is also detected in many tissues, including developing embryos (8), metabolic sinks such as meristematic tips of young roots and shoots (17), and the aleurone cell layer and basal part of the developing endosperm (18, 19). In addition, there is a marked difference in the regulatory properties of the two genes; anaerobic induction is specific for the Sh locus (15); however, such induction is restricted to only the RNA level as there is no detectable change at the protein level (17). Conversely, the Sus locus is induced in dark-grown seedlings, whereas the Sh expression is relatively unchanged (20). Despite such dichotomy in tissue and cell specificities as well as in the inductional properties of Sh and Sus loci, it was intriguing that a mutant sus genotype lacking the SS2 protein showed no detectable phenotypic change in the plant (21). We demonstrate here, based on the combined use of appro- priate genetic strains, gene-specific cDNA probes, and isozyme-specific monoclonal antibodies, that genetically re- dundant Sh and Sus loci are cross-regulated and are also functionally redundant. In addition, we observe, in develop- ing embryos, an epistatic relationship between the two genes at the protein level but not at the RNA level; thus, a translational hierarchy appears to play a major role in the ultimate determination of embryo specificity of SS expres- sion. MATERIALS AND METHODS Plant Materials. The isolation of Sh'-5 sus and sh sus genotypes has been described previously (ref. 21; unpub- lished data). The new genetic stocks, Sh sus and Sh'-5 Sus, were obtained by crossing the standard W22 inbred strain, Sh Sus, with sh sus and the Sh'-5 sus with sh Sus (the sh bz-m4 stock) stocks, respectively. The F1 heterozygotes from each of the two crosses were selfed for several generations. Abbreviations: SS, sucrose synthase; DAP, day(s) after pollination; Sh, shrunken 1; Sus, sucrose synthase 1. 7917 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact. Downloaded by guest on January 20, 2022
Transcript of Epistatic interaction andfunctional ...

Proc. Natl. Acad. Sci. USAVol. 91, pp. 7917-7921, August 1994Genetics
Epistatic interaction and functional compensation between the twotissue- and cell-specific sucrose synthase genes in maize
(isozymes/posttranscriptional control/cross-regulation)
PREM S. CHOUREY AND EARL W. TALIERCIOU.S. Department of Agriculture, Agricultural Research Service, and Departments of Agronomy and Plant Pathology and Program in Plant Molecular andCellular Biology, University of Florida, Gainesville, FL 32611-0680
Communicated by Oliver E. Nelson, Jr., May 9, 1994
ABSTRACT A tissue-specific epistatic mode of gene inter-action was observed between molecularly homologous genesShl and Susi (hereafter, Sh and Sus), encoding the sucrosesynthase (SS) isozymes, SS1 and SS2, respectively. In Sh Susgenotype, both SS genes were expressed simultaneously andapproximately equally in young seedlings; however, only theSus-encoded SS2 protein was seen in the developing embryos.By contrast, the mutant sass genotype, lacking detectable levelsof the SS2 protein in various tissues tested, showed expressionof the Sh locus as judged by the detection of the SS1 protein insuch embryos. Ectopic expression in embryos was seen fromtwo separate Sh alleles, Sh-W22 and Sh'-5 (a revertant allelederived upon Ds excision from sh-m5933). In each case, the Shexpression at the protein level in embryos was unique togenotypes with the mutant sus gene. Based on the observed lackof phenotypic change in the sus mutant, we suggest that theectopic expression of the Sh in otherwise Sus-specific tissuesleads to functional compensation. There was no epistatic in-teraction ofSh and Sus at the RNA level as SS1 transcripts weredetectable in both Sus and sus embryos. Thus, embryo speci-ficity between the two SS genes was determined at posttran-scriptional or at translational level of control. We surmise onthe basis of these data that metabolic regulatory controls seemto override the normal constraints of tissue and cell specificityof the nonallelic isozyme genes to maintain efficient use of thepathways.
Genetic redundancy in higher eukaryotes is well docu-mented. Duplicate or molecularly homologous genes encod-ing biochemically similar gene products are now a commonphenomenon in both of the model plant systems, Arabidop-sis and maize. Mutational inactivation of a gene invariablyuncovers another nonallelic gene encoding a similar function.Maize, generally considered a true diploid, is well analyzedfor duplicate genes (1) and molecularly homologous se-quences (2). Biochemical genetic studies on developingmaize endosperms indicate that many starch mutants asso-ciated with the loss ofa specific enzyme have unaltered levelsof the corresponding isozyme expressed either in the sametissue or in another tissue such as embryo and other sporo-phytic organs (3-6). Recent analyses in Arabidopsis, partic-ularly in amino acid biosynthesis, have similarly shown thatgenetic redundancy is a rule rather than exception for severalenzymes in this pathway (7). Duplicated genes usually havedistinct, nonoverlapping physiological functions that are dif-ficult to resolve by biochemical approaches alone. The com-bined use ofgenetic and molecular approaches is ofparticularuse in the elucidation of their functional significance.
In this regard, the two sucrose synthase (SS) genes inmaize provide a model system. The enzyme plays an impor-tant role in energy metabolism by mobilizing sucrose into
diverse pathways relating to metabolic, structural, and stor-age functions of plant cells. The two biochemically similarisozymes, SS1 and SS2, are encoded by the shrunken (Sh)and sucrose synthase (Sus) loci, respectively (8, 9). Both Shand Sus have been cloned and shown to be molecularlyhomologous (10-12). A similar two-gene system for the twoSS isozymes is also described in wheat, sorghum, sugarcane,and several dicot species, including Arabidopsis (13). Inmaize, the SS1 isozyme is present in abundant levels indeveloping endosperm. The loss of this protein, as in the caseof the shrunken (sh) mutant, is associated with severalalterations in developing endosperm, including a reduction inthe amount of starch (4), an early cell degeneration (14), anda reduction in the number of protein bodies (unpublisheddata). There is otherwise no phenotypic change in the shplant, even though Sh expression is detected in severaltissues and cells of young plants of the wild-type genotype(15-17). The Sus-encoded SS2 protein is also detected inmany tissues, including developing embryos (8), metabolicsinks such as meristematic tips of young roots and shoots(17), and the aleurone cell layer and basal part of thedeveloping endosperm (18, 19). In addition, there is a markeddifference in the regulatory properties of the two genes;anaerobic induction is specific for the Sh locus (15); however,such induction is restricted to only the RNA level as there isno detectable change at the protein level (17). Conversely,the Sus locus is induced in dark-grown seedlings, whereas theSh expression is relatively unchanged (20).
Despite such dichotomy in tissue and cell specificities aswell as in the inductional properties of Sh and Sus loci, it wasintriguing that a mutant sus genotype lacking the SS2 proteinshowed no detectable phenotypic change in the plant (21).We demonstrate here, based on the combined use of appro-priate genetic strains, gene-specific cDNA probes, andisozyme-specific monoclonal antibodies, that genetically re-dundant Sh and Sus loci are cross-regulated and are alsofunctionally redundant. In addition, we observe, in develop-ing embryos, an epistatic relationship between the two genesat the protein level but not at the RNA level; thus, atranslational hierarchy appears to play a major role in theultimate determination of embryo specificity of SS expres-sion.
MATERIALS AND METHODSPlant Materials. The isolation of Sh'-5 sus and sh sus
genotypes has been described previously (ref. 21; unpub-lished data). The new genetic stocks, Sh sus and Sh'-5 Sus,were obtained by crossing the standard W22 inbred strain, ShSus, with sh sus and the Sh'-5 sus with sh Sus (the sh bz-m4stock) stocks, respectively. The F1 heterozygotes from eachof the two crosses were selfed for several generations.
Abbreviations: SS, sucrose synthase; DAP, day(s) after pollination;Sh, shrunken 1; Sus, sucrose synthase 1.
7917
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Dow
nloa
ded
by g
uest
on
Janu
ary
20, 2
022
7918 Genetics: Chourey and Taliercio
Homozygous Sh and Sh'-5 seedlings in the F3 generationwere tested for homozygosity at sus and Sus loci, respec-tively, by Western and Southern blot analyses.Immature kernels at 22 days after pollination (DAP) and 8-
to 10-day-old seedlings of various genotypes were harvested,quick frozen in liquid N2, and stored at -800C until ready touse.
Protein Analyses. Crude extracts from fresh roots of 8- to10-day-old etiolated seedlings were prepared as described(16). Embryos of22 DAP frozen kernels were dissected on iceand rinsed several times in ice-cold water to remove en-dosperm juices adsorbed to the surface. Excised embryoswere homogenized using 1.0 g of fresh weight to 5.0 ml ofgrinding buffer (0.01 M Tris-maleate, pH 7.0), strainedthrough glass wool, and centrifuged at 20,000 x g for 20 min(21). Samples were electrophoresed on a 4.5-7.0% lineargradient nondenaturing polyacrylamide gel, electroblotted tonitrocellulose, and immunostained using the appropriate pri-mary antibody as detailed previously (16, 21, 22).DNA Analyses. Genomic DNA was isolated from 7-day-old
dark-grown seedlings as described (23). Approximately 15 Hgof DNA was digested with EcoRI enzyme according tomanufacturer's specifications (GIBCO/BRL). The digestedsamples were fractionated on 0.8% agarose gels, transferredto Nytran membranes (Schleicher & Schuell), and hybridizedto 32P-labeled 2.5-kb cDNA probe corresponding to the Susgene (12). Blot hybridization and washes were done asdescribed (12).RNA Analyses. Total RNA was isolated from immature
embryos as described (24). Ten-microgram samples weredenatured and fractionated on 1% agarose/formaldehydegels as described (12). RNA was transferred to Nytranmembrane and prehybridized in 6x SSC/0.05x Blotto/50%formamide/100 pg of sheared and denatured salmon spermDNA per ml for 4 hr at 420C. The filters were hybridized withSh, Sus (12), or Em2 (25) cDNA inserts for 16-18 hr at 65°Cand then washed, the first two washes in 6x SSC/5% SDSfollowed by two more washes in 0.lx SSC/1% SDS at 65°C.
RESULTSIsolation and Characterization of the Specific Genotypes.
The sh mutant is readily identifiable due to a collapsed(shrunken) seed phenotype. Twenty sh alleles of independentorigins have been isolated and analyzed so far and all suchmutants lack the SS1 protein (ref. 8; unpublished data). Thesh mutation used in these studies, sh bz-m4, represents adeletion of the entire coding region and lacks both RNA andthe protein. By contrast, there is thus far only a single susmutation lacking the SS2 protein; however, no associatedphenotypic change is seen with the loss of this protein (21).Consequently, identifying for Sus or mutant sus segregants istedious and is dependent on Western and/or Southern blotanalyses. The two SS genes are about 40 map units apart onchromosome 9 (11, 12), and recombinants are easily obtainedin the progeny of the appropriate crosses and identified asdescribed below.
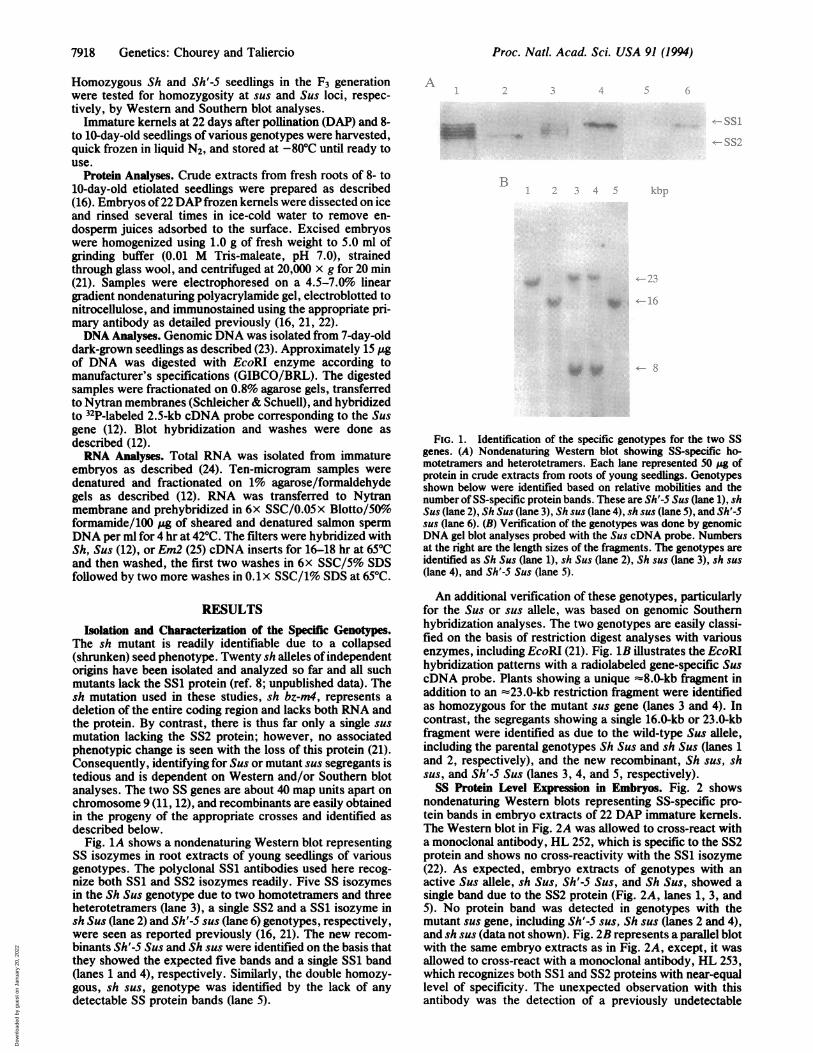
Fig. 1A shows a nondenaturing Western blot representingSS isozymes in root extracts of young seedlings of variousgenotypes. The polyclonal SS1 antibodies used here recog-nize both SS1 and SS2 isozymes readily. Five SS isozymesin the Sh Sus genotype due to two homotetramers and threeheterotetramers (lane 3), a single SS2 and a SS1 isozyme insh Sus (lane 2) and Sh'-5 sus (lane 6) genotypes, respectively,were seen as reported previously (16, 21). The new recom-binants Sh'-5 Sus and Sh sus were identified on the basis thatthey showed the expected five bands and a single SS1 band(lanes 1 and 4), respectively. Similarly, the double homozy-gous, sh sus, genotype was identified by the lack of anydetectable SS protein bands (lane 5).
FIG. 1. Identification of the specific genotypes for the two SSgenes. (A) Nondenaturing Western blot showing SS-specific ho-motetramers and heterotetramers. Each lane represented 50 .gofprotein in crude extracts from roots of young seedlings. Genotypesshown below were identified based on relative mobilities and thenumber of SS-specific protein bands. These are Sh'-S Sus (lane 1), shSus (lane 2), Sh Sus (lane 3), Sh sus (lane 4), sh sus (lane 5), and Sh'-Ssus (lane 6). (B) Verification of the genotypes was done by genomicDNA gel blot analyses probed with the Sus cDNA probe. Numbersat the right are the length sizes of the fragments. The genotypes areidentified as Sh Sus (lane 1), sh Sus (lane 2), S sus (lane 3), sh sus(lane 4), and Sh'-S Sus (lane 5).
An additional verification of these genotypes, particularlyfor the Sus or sus allele, was based on genomic Southernhybridization analyses. The two genotypes are easily classi-fied on the basis of restriction digest analyses with variousenzymes, including EcoRI (21). Fig. 1B illustrates the EcoRIhybridization patterns with a radiolabeled gene-specific SuscDNA probe. Plants showing a unique z8.0-kb fragment inaddition to an '=23.0-kb restriction fragment were identifiedas homozygous for the mutant sus gene (lanes 3 and 4). Incontrast, the segregants showing a single 16.0-kb or 23.0-kbfragment were identified as due to the wild-type Sus allele,including the parental genotypes Sh Sus and sh Sus (lanes 1and 2, respectively), and the new recombinant, Sh sus, shsus, and Sh'-5 Sus (lanes 3, 4, and 5, respectively).SS Protein Level Expression in Embryos. Fig. 2 shows
nondenaturing Western blots representing SS-specific pro-tein bands in embryo extracts of 22 DAP immature kernels.The Western blot in Fig. 2A was allowed to cross-react witha monoclonal antibody, HL 252, which is specific to the SS2protein and shows no cross-reactivity with the SS1 isozyme(22). As expected, embryo extracts of genotypes with anactive Sus allele, sh Sus, Sh'-5 Sus, and Sh Sus, showed asingle band due to the SS2 protein (Fig. 2A, lanes 1, 3, and5). No protein band was detected in genotypes with themutant sus gene, including Sh'-5 sus, Sh sus (lanes 2 and 4),and sh sus (data not shown). Fig. 2B represents a parallel blotwith the same embryo extracts as in Fig. 2A, except, it wasallowed to cross-react with a monoclonal antibody, HL 253,which recognizes both SS1 and SS2 proteins with near-equallevel of specificity. The unexpected observation with thisantibody was the detection of a previously undetectable
Proc. Natl. Acad. Sci. USA 91 (1994)
Dow
nloa
ded
by g
uest
on
Janu
ary
20, 2
022

Proc. Nat. Acad. Sci. USA 91 (1994) 7919
'L Z~~~~~~~~~~~~~~~~~~~~~~~r~Jrv>HA
Ss-)I
SSi.- i.S
B
.I.
,-SS'
B
, S.1
e-SS1
_mm <-SS2
4
FIG. 2. Nondenaturing Western blot showing SS1 and SS2 pro-teins and an epistatic interaction in developing embryos. (A) The blotwas allowed to cross-react with maize SS2-specific monoclonalantibody. Each lane represents 50 pg of protein in crude extractsfrom embryos of various genotypes. The SS2 protein was readilydetectable in Sus genotypes (lanes 1, 3, and 5) but was undetectablein mutant sus embryos (lanes 2 and 4). (B) A similar blot as in A wasallowed to cross-react with another SS-specific maize monoclonalantibody that recognizes both SS1 and SS2 isozymes. A slowmigrating band due to the SS1 protein, encoded by the Sh'-5 (lane 2)or the Sh (lane 4), was detected only in genotypic combinations withthe mutant sus gene and not the normal Sus gene (lanes 3 and 5).
protein in genotypes with the mutant sus gene, Sh'-5 sus andSh sus (lanes 2 and 4, respectively). This protein band was ofslightly slower mobility relative to the SS2 protein seen in Susgenotypes (lanes 1, 3, and 5). We conclude that this proteinwas the SS1 protein encoded by the Sh or the Sh'-5 allele. Inaddition to the characteristic slower mobility relative to theSS2 protein, it was cross-reactive with a monoclonal anti-body that shares epitope specificity with the SS1 protein andwas undetectable in the sh sus genotype (Fig. 1A, lane 5),including developing embryos (data not shown). It is note-worthy that although the SS1 protein band was detectable insus embryo extracts at loading levels as low as 20 pg (data notshown), it was undetectable in Sh Sus and Sh'-5 Sus geno-types at 50 pg (Fig. 2B, lanes 3 and 5) and up to 70 pg ofprotein tested for these two genotypes (data not shown). Theectopic levels ofSS1 protein in the mutant sus embryos were,however, slightly reduced as compared to SS2 protein in thewild-type Sus embryos, as judged by the intensity of the SSbands (Fig. 2B). Such a reduction in the level of SS1 proteinis also associated with a slight reduction in SS enzymeactivity in the mutant sus embryos, relative to Sus embryos(21).RNA Level Expression in Embryos. Fig. 3A illustrates a
RNA blot showing steady-state levels of the SS2 transcriptsin total RNAs of embryos of 22 DAP immature kernels usinga radiolabeled Sus cDNA as a hybridization probe. The threegenotypes with a wild-type Sus gene, including Sh'-5 Sus, shSus, and Sh Sus, showed abundant levels ofthe normal-sized2900-nt SS2 transcripts (lanes 1, 4, and 6). However, themutant sus genotypes, Sh'-S sus, sh sus, and Sh sus, showedno detectable levels of the normal-sized SS2 transcripts;instead, a slightly smaller sized, =2500-nt, transcript ofmuchreduced steady-state levels was observed in two of the threegenotypes. The detectability of this truncated transcript,SS2*, in the sh sus embryos (Fig. 3A, lanes 3 and 3a, showinga normal and a longer-duration exposure of the same blot to
C,*sk Ja.
FIG. 3. RNA gel blot showing SS-specific transcripts in immatureembryos. Ten micrograms of total RNA from various genotypes wasblotted onto Nytran membrane and hybridized with nearly full-lengthcDNA inserts. (A) Sus probe hybridized to a prominent 2900-nt SS2transcript in Sus genotypes (lanes 1, 4, and 6); no hybridization wasdetectable in the mutant sus embryos (lanes 2, 3, and 5) except atruncated transcript, SS2*, which was seen in sh sus (lanes 3 and 3a)and Sh sus (lane 5). (B) Sh probe hybridized to a 2750-nt SS1transcript that was undetectable in sh embryos (lanes 3 and 4) but wasdetectable in all Sh genotypes (lanes 1 and 2 and lanes 5 and 6). (C)Embryo-specific Em2 probe hybridized to -800-nt transcripts toshow an internal loading control of the RNAs of the genotypes, asshown.
the film, respectively) was of particular interest in the elu-cidation of its genetic basis. We attribute the truncatedtranscript to the mutant sus gene since there are only twoknown SS genes in the maize genome (8, 11, 12) and the entirecoding region of the sh locus is deleted in the sh sus doublerecessive combination. Moreover, there was very little or nocross-hybridization between the Sus cDNA probe and the2750-nt SS1 transcript under our hybridization and washconditions (ref. 12; see below). Although the SS2* transcriptwas undetectable in Sh'-5 sus (lane 2) but readily detectablein Sh sus embryos (lane 5), we believe such variabilitybetween the two genotypes was related to the highly labilenature of this truncated transcript during our extractionprocedures and handling. Our collective data from severalsimilar analyses have shown that, in general, the SS2*transcript was observed in both Sh sus and Sh'-5 sus geno-types (data not shown). The quality of RNAs in thesepreparations was comparable based on equal-level intensitiesofethidium bromide-stained rRNA bands (data not shown) aswell as an internal loading control (Fig. 3C), representingembryo-specific %800-nt transcripts visualized by hybridiza-tion to a labeled cDNA probe of wheat early methioninelabeled, Em2, gene (25).
Fig. 3B represents a parallel RNA blot, with the samesamples as in Fig. 3A, showing steady-state levels of SS1transcripts detectable by the radiolabeled Sh cDNA probe.As expected, no SS1 transcripts were detectable in embryosof the two sh genotypes, sh sus and sh Sus (Fig. 3B, lanes 3and 4, respectively). A single SS1 transcript of the knownsize, 2750 nt, was, however, readily detectable in all fourgenotypes with the Sh or Sh'-5 allele, independent ofwhetherit was in combination with the wild-type or the mutant susgene (lanes 1 and 2 and lanes 5 and 6). The detection of SS1
A
Genetics: Chourey and Taliercio
r,r
.-Zl
------I -:z -Z.
:. .. :. :.
Dow
nloa
ded
by g
uest
on
Janu
ary
20, 2
022

7920 Genetics: Chourey and Taliercio
transcripts in Sh'-5 Sus and Sh Sus embryos (lanes 1 and 6),which do not accumulate detectable levels of SS1 protein, asjudged by Western blot analysis (Fig. 2B, lanes 3 and 5), wasunexpected. In addition, there was a slight but reproducibilydetectable reduction in steady-state levels of SS1 transcriptsin Sus embryos relative to the mutant sus genotype (Fig. 3B,lanes 1 vs. 2 and 5 vs. 6). Because there were no correlateddifferences in the corresponding lanes of the EM2 transcripts(Fig. 3C), we believe these differences in SS1 transcriptswere not attributable to any loading differences.
DISCUSSIONEpistasis, Ectopic Activation, and Functional Compensa-
tion. We have used complementary molecular and geneticapproaches to show that tissue and cell specificity betweenthe two molecularly homologous SS genes is determined byan epistatic interaction. In developing embryos, the Sus locuswas epistatic to the Sh as evidenced by abundant levels of theSS2 protein and undetectable levels of the SS1 protein in anormal Sh Sus genotype. However, in the mutant sus geno-type, lacking the SS2 protein, SS1 protein was readilydetectable. Such an activation of the Sh locus was demon-strated for two separate Sh alleles, Sh-W22 (the W22 inbredbackground) and the Sh'-5 strain, which carries a Sh-revertant locus derived from an excision of the Ds elementfrom the sh-m5933 allele (26). The expression of the Sh-W22allele in embryos in this study is of particular importancebecause it was not clear from our previous study (21), limitedto the Sh'-5 allele, if the ectopic expression in embryos wasdue to an imprecise excision of the Ds element. The resultsfrom this study now clearly show that expression of the Shlocus at the protein level in developing embryos was attrib-utable to whether or not it was in combination with a mutantor a wild-type Sus gene.Although the data in this study are restricted to embryos,
we suggest that a similar interaction is also manifested indeveloping endosperm, including the aleurone cell layer. Wehave shown previously that although both SS1 and SS2isozymes are detectable in a Sh Sus endosperm, the twogenes are not expressed simultaneously in the same cell sincethe heteromeric isozymes are undetectable by Western blotanalyses (16). Such cell specificity was also seen at the RNAlevel of expression. RNA gel blots on RNAs prepared fromexcised basal parts and the upper parts (storage cells) of ShSus endosperms have shown the SS2 and SS1 transcripts,respectively (unpublished data). In addition, two indepen-dent immunohistological analyses have shown substantialspatial heterogeneity in the endosperm in terms of the ex-pression of SS genes (18, 19). The use of Sh Sus and sh Susgenotypes has shown that aleurone cells and basal endospermtransfer cells are Sus-specific, whereas the centrally locatedstorage cells engaged in starch biosynthesis are Sh-specific.However, if there is no Sh-encoded SS1 protein in theendosperm, as is the case ofthe sh mutation, the Sus-encodedSS2 protein is readily detectable in the central part of theendosperm (18, 19), except the cells lost due to cell degen-eration (14). Thus far, there has been no satisfactory expla-nation for the undetectable levels of the SS2 protein in thecentral region of the wild-type Sh Sus endosperm. Wespeculate, on the basis of the above embryo data, that ahighly active Sh locus is epistatic to Sus in this region of theendosperm cells. It should be noted, however, that the twoSS genes are not always in an epistatic mode. In youngseedlings both genes are expressed simultaneously in thesame cell as evidenced by the five isozymes, two homotet-ramers and three heterotetramers (16, 21). Thus, it appearsthat the degree of epistatic interaction is variable and inter-changeable in a tissue- and cell-specific fashion.
There is no phenotypic change in the sus mutant due to theloss of the SS2 protein, presumably due to ectopic expressionof the Sh in embryos and aleurone cells. We suggest that Shand Sus are functionally compensatory. Such a functionalduplication has been previously described between the mo-lecularly homologous regulatory genes, B and R, controllingthe anthocyanin pathway in maize (27). Functional similaritybetween Sh and Sus is compatible with much biochemicalsimilarity between the encoded isozymes (8,9), including the83% level of similarity in deduced amino acid sequences ofthe Sh and Sus cDNA clones (28). In addition, both isozymesare cytosolic in nature as judged by biochemical analyses (4,8,9, 16, 21). The functional compensation is however, in part,nonreciprocal in nature; in particular, the SS2 protein in shendosperm fails to substitute for the loss of the SS1 isozymeas evidenced by the mutant seed phenotype. It is presumablybecause the SS2 protein in the sh endosperm contributes upto only 5% of the total enzyme activity observed in thewild-type, Sh, genotype (8). This is obviously lower than thethreshold levels needed to restore the normal phenotype.
Possible Metabolic Controls and Posttranscriptional Regu-lation. Another important feature of the intergenic interactionbetween the SS genes was that posttranscriptional regulationappears to play an important role in the ultimate determina-tion of embryo specificity. Although there was a completeepistatic control of the Sh by the Sus at the protein level, onlya partial suppression of Sh expression was seen at the RNAlevel. The SS1 transcripts were readily detectable in Sh Susand Sh'-5 Sus embryos, which showed no detectable levels ofthe SS1 protein. These data are clearly indicative of, amongother possibilities, a certain rate-limiting step that may de-termine translational hierarchy among the two SS transcripts.In the embryo, SS2 transcripts may be preferred over SS1,and SS1 may be translated only when no normal SS2 tran-scripts are available. Such a translational preference may alsolead to greater stability of the SS2 transcripts in Sus embryos.Such a control may provide a possible basis for the observedgreater steady-state levels of the SS2 transcripts as comparedto the SS1 transcripts. Cotranslational regulation of stabilityof 3-tubulin mRNA in animal cells is well analyzed in termsof the molecular basis of such expression (29). A large arrayof mechanisms at posttranscriptional and translational levelsis also known to modulate gene expression (30-32). In plants,the translational elongation factor is known to be regulateddevelopmentally and may affect protein synthesis in a rate-limiting manner (33). Similarly, poly(A)-binding protein isalso implicated in the stability and translation of transcriptsleading to posttranscriptional control of gene expression (31).An alternative possibility is that SS1 protein is more labile inSus than the mutant sus embryos such that a detectableaccumulation of the protein fails to occur in the Sus genotype.Indeed, there was a slight reduction in the steady-state levelsof SS1 protein in the mutant sus genotype as compared to thelevels of SS2 protein in Sus embryos. Differential stabilitiesof these two proteins, among other factors, may also con-tribute to such an effect. The translational fate of the trun-cated SS2 RNA, which we know is polyadenylylated (un-published data), requires further investigation.
Posttranscriptional control of Sh expression may be amajor component of regulation. We have previously shown(17, 34) that anaerobically induced SS1 transcripts werepolyadenylylated, loaded onto polysomes, but led to noconcomitant increase in the levels of the SS1 protein relativeto the control seedlings. Furthermore, 35S uptake studiesshowed no evidence that SS1 protein is rapidly turned overin the anaerobic environment (17, 34). A similar posttran-scriptional modulation of anaerobic induction is also ob-served for the Sh homolog, Susi, in sorghum (22). In Ara-bidopsis, an -10-fold elevation in SS RNA due to anaerobic
Proc. Natl. Acad. Sci. USA 91 (1994)
Dow
nloa
ded
by g
uest
on
Janu
ary
20, 2
022

Proc. Natl. Acad. Sci. USA 91 (1994) 7921
induction ofthe Asusi gene is associated with only about 42%increase in enzyme activity (13).Any speculations concerning possible mechanisms on the
regulation of the Sh and Sus genes must take metabolicregulation by carbohydrates into consideration because SSprovides a critical link between the photosynthesized sucroseand its utilization in developing seed. In this context, elegantstudies on the regulation of seven photosynthetic genes (35)and the feedback regulation ofthe Sh promoter (36) by sugarsusing transient transformation assays in maize protoplastsare relevant. A 26-bp feedback element in the Sh promoter issufficient to elicit the metabolic response. Whether a similarelement is also present in the Sus promoter is not known.Another study reports a Sh- and Sus-specific induction indetached maize roots to increasing concentrations ofglucoseat RNA level; however, such gene specificity is not readilydetectable at the protein level (37). Elevated levels of tran-scripts homologous to maize Sus gene with increasing con-centrations of glucose in isolated rice embryos are alsodescribed (38). Collectively, these divergent studies showclear evidence of metabolic regulation of the SS genes at theRNA level. It is probable, based on functional compensatoryinteraction, that both Sh and Sus share common metabolicregulators. We suggest that Sh and Sus are in a competitivemode of expression and the hierarchical pattern may resultfrom the availability of tissue- and cell-specific trans-regulatory factors that may modulate the cross-regulatoryinteraction at posttranscriptional stages of gene expression.
Finally, it should be pointed out that the epistatic interac-tion between the two SS genes is similar to trans effectsbetween the duplicated sequences of transgenes broughttogether by transformation or by sexual crossing (39), at leastin terms of the end effect. As pointed out by Matzke andMatzke (39), the phenomenon of trans interactions is ratherwidespread and can vary significantly in its manner ofexpression. Whether such epistatic interactions, as describedhere, are a common occurrence among the endogenousisozyme genes remains to be seen.
This work was in part supported by U.S. Department of Agricul-ture-NRICGP Grant 91-37301-6343. We are grateful to W. R. Mar-cott, Jr., for Em2 plasmid and toO. E. Nelson, D. R. Pring, and L. J.Rowland for critical reading of the manuscript. This was a cooper-ative investigation of the U.S. Department of Agriculture-Agricultural Research Service and the Institute of Food and Agri-cultural Sciences, University of Florida (Agricultural ExperimentJournal Series No. R-03758).
1. Wendel, J. F., Stuber, C. W., Goodman, M. M. & Beckett,J. B. (1989) J. Hered. 80, 218-222.
2. Helentjaris, T., Weber, D. & Wright, S. (1988) Genetics 118,353-363.
3. Dickinson, D. B. & Preiss, J. (1969) Plant Physiol. 44, 1058-1062.
4. Chourey, P. S. & Nelson, 0. E. (1976) Biochem. Genet. 14,1041-1055.
5. Nelson, 0. E., Chourey, P. S. & Chang, M. T. (1978) PlantPhysiol. 62, 383-386.
6. Boyer, C. D. & Preiss, J. (1981) Plant Physiol. 67, 1141-1145.7. Niyogi, K. & Fink, G. R. (1992) Plant Cell 4, 721-733.8. Chourey, P. S. (1981) Mol. Gen. Genet. 184, 372-376.9. Echt, C. S. & Chourey, P. S. (1985) Plant Physiol. 79, 530-536.
10. Werr, W., Frommer, W. B., Mass, C. & Starlinger, P. (1985)EMBO J. 4, 1373-1380.
11. McCarty, D. R., Shaw, J. R. & Hannah, L. C. (1986) Proc.Natl. Acad. Sci. USA 83, 9099-9103.
12. Gupta, M., Chourey, P. S., Burr, B. & Still, P. E. (1988) PlantMol. Biol. 10, 215-224.
13. Martin, T., Frommer, W. B., Salanoubat, M. & Willmitzer, L.(1993) Plant J. 4, 367-377.
14. Chourey, P. S., Chen, Y.-C. & Miller, M. E. (1991) Maydica36, 141-146.
15. Springer, B., Werr, W., Starlinger, P., Zokolica, M. & Freeling,M. (1986) Mol. Gen. Genet. 205, 461-468.
16. Chourey, P. S., Latham, M. & Still, P. E. (1986) Mol. Gen.Genet. 203, 251-255.
17. Rowland, J. R., Chen, Y.-C. & Chourey, P. S. (1989) Mol.Gen. Genet. 218, 33-40.
18. Chen, Y.-C. & Chourey, P. S. (1989) Theor. Appl. Genet. 78,553-559.
19. Heinlein, M. & Starlinger, P. (1989) Mol. Gen. Genet. 215,441-446.
20. Nguyen-Quoc, B., Krivitzky, M., Huber, S. C. & Lecharny, A.(1990) Plant Physiol. 94, 516-523.
21. Chourey, P. S., DeRobertis, G. A. & Still, P. E. (1988) Mol.Gen. Genet. 214, 300-306.
22. Chourey, P. S., Taliercio, E. W. & Kane, E. J. (1991) PlantPhysiol. 96, 485-490.
23. Dellaporta, S. L., Wood, J. & Hicks, J. B. (1983) Plant Mol.Biol. Rep. 1, 19-21.
24. Wadsworth, G. J., Redinbaugh, M. G. & Scandalios, J. G.(1988) Anal. Biochem. 172, 279-283.
25. Litts, J. C., Colwell, G. W., Chakerian, R. L. & Quatrano,R. S. (1987) Nucleic Acids Res. 15, 3607-3618.
26. Courage-Tebbe, U., Doring, H. P., Federoff, N. & Starlinger,P. (1983) Cell 34, 383-393.
27. Chandler, V. L., Radicella, J. P., Robbins, T. P., Chen, J. &Turks, D. (1989) Plant Cell 1, 1175-1183.
28. Huang, X.-F., Nguyyen, Q.-B., Chourey, P. S. & Yelle, S.(1994) Plant Physiol. 104, 293-294.
29. Theodorakis, N. G. & Cleveland, D. W. (1992) Mol. Cell. Biol.12, 791-799.
30. Thatch, R. E. (1992) Cell 68, 177-180.31. Green, P. J. (1993) Plant Physiol. 102, 1065-1070.32. Gallie, D. R. (1993) Annu. Rev. Plant Physiol. Plant Mol. Biol.
44, 77-105.33. Ursin, V. M., Ironie, J. M., Hiatt, W. R. & Shewmaker, C. K.
(1991) Plant Cell 3, 583-591.34. Taliercio, E. W. & Chourey, P. S. (1989) Plant Physiol. 90,
1359-1364.35. Sheen, J. (1990) Plant Cell 2, 1027-1038.36. Mass, C., Schall, S. & Werr, W. (1990) EMBO J. 9, 3447-3452.37. Koch, K. E., Nolte, K. D., Duke, E. R., McCarty, D. R. &
Avigne, W. T. (1992) Plant Cell 4, 59-69.38. Karrer, E. K. & Rodriguez, R. L. (1992) Plant J. 2, 517-523.39. Matzke, M. A. & Matzke, A. J. M. (1993) Annu. Rev. Plant
Physiol. Plant Mol. Biol. 44, 53-76.
Genetics: Chourey and Taliercio
Dow
nloa
ded
by g
uest
on
Janu
ary
20, 2
022


















