
Epigenetic modifications and human disease Nat Biotech review 2010... · tone modification profiles...
12
NATURE BIOTECHNOLOGY VOLUME 28 NUMBER 10 OCTOBER 2010 1057 generated from the same donor DNA are not identical to, and develop diseases with different penetrance from, their donor 1,3 . Human clones that arise spontaneously—monozygotic twins—are identical at the DNA sequence level, but have different DNA methylation 4,5 and his- tone modification profiles 4 that might affect the penetrance of several diseases, such as cancer 4 or autoimmune disorders 6 . But this phenom- enon is also observed at a single cell level: how can stem cells develop into any type of cell and how does a liver cell always give rise to two new liver cells after cell division? Again, epigenetics seems to be part of the answer as it has been described as one of the key factors in cellular differentiation 7,8 (see the review by Meissner 9 in this issue). The importance of epigenetics in maintaining normal development and biology is reflected by the observation that many diseases develop when the wrong type of epigenetic marks are introduced or are added at the wrong time or at the wrong place 10 . For instance, a clear causality role for DNA methylation in cancer is suggested by hypermethylation of some genes (e.g., p16 INK4a , p14 ARF and MGMT) as an early event in tumorigenesis, as well as by tumor type-specific methylation landscape 11 . Here we summarize recent progress in the field of epigenetic research and its role in disease, preparing ourselves for the surprises that epigenetics might hold in the future. Epigenetic modifications and their machineries For didactic purposes, epigenetic modifications can be grouped into three main categories: DNA methylation, histone modifications and nucleosome positioning. It is important to keep in mind the interplay between epige- netic factors—as the observed outcome is always the sum of their interac- tions—and the many positive and negative feedback mechanisms. DNA methylation. The most widely studied epigenetic modification in humans is cytosine methylation. DNA methylation occurs almost exclusively in the context of CpG dinucleotides. The CpG dinucleotides tend to cluster in regions called CpG islands 1 , defined as regions of Even before DNA was identified as the molecule of inheritance, scientists knew that not every gene in an organism can be active in each cell at all times. Even so, all cells in an organism share the same genetic information. Conrad Waddington coined the term ‘epigenetic landscape’ 1,2 for the molecular mechanisms that convert this genetic information into observable traits or phenotypes. In many instances, epigenetic gene expression patterns and associated phenotypes persist through mitosis or even meiosis, although no change in the primary DNA sequence has occurred. Consequently, epigenetics is generally understood to be the study of mechanisms that control gene expression in a potentially heritable way. Recent breakthroughs in the understanding of the mechanisms under- lying epigenetic phenomena and their prevalence as contributors to the development of human disease have led to a greatly enhanced interest in epigenetic research. On a molecular level, covalent modifications of cytosine bases and histones, and changes in the positioning of nucleosomes are commonly regarded as the driving epigenetic mechanisms. They are fundamental to the regulation of many cellular processes, including gene and microRNA expression, DNA-protein interactions, suppression of transposable ele- ment mobility, cellular differentiation, embryogenesis, X-chromosome inactivation and genomic imprinting. In multicellular organisms, the ability of epigenetic marks to persist during development and potentially be transmitted to offspring may be necessary for generating the large range of different phenotypes that arise from the same genotype 1,3–5 . For instance, cloned animals Epigenetic modifications and human disease Anna Portela 1 & Manel Esteller 1,2 Epigenetics is one of the most rapidly expanding fields in biology. The recent characterization of a human DNA methylome at single nucleotide resolution, the discovery of the CpG island shores, the finding of new histone variants and modifications, and the unveiling of genome-wide nucleosome positioning maps highlight the accelerating speed of discovery over the past two years. Increasing interest in epigenetics has been accompanied by technological breakthroughs that now make it possible to undertake large-scale epigenomic studies. These allow the mapping of epigenetic marks, such as DNA methylation, histone modifications and nucleosome positioning, which are critical for regulating gene and noncoding RNA expression. In turn, we are learning how aberrant placement of these epigenetic marks and mutations in the epigenetic machinery is involved in disease. Thus, a comprehensive understanding of epigenetic mechanisms, their interactions and alterations in health and disease, has become a priority in biomedical research. 1 Cancer Epigenetics and Biology Program (PEBC), Bellvitge Biomedical Research Institute (IDIBELL), Barcelona, Catalonia, Spain. 2 Institucio Catalana de Recerca i Estudis Avançats (ICREA), Barcelona, Catalonia, Spain. Correspondence should be addressed to M.E. ([email protected]). Published online 13 October 2010; doi:10.1038/nbt.1685 REVIEW © 2010 Nature America, Inc. All rights reserved.
Transcript of Epigenetic modifications and human disease Nat Biotech review 2010... · tone modification profiles...

nature biotechnology volume 28 number 10 oCTober 2010 1057
generated from the same donor DNA are not identical to, and develop diseases with different penetrance from, their donor1,3. Human clones that arise spontaneously—monozygotic twins—are identical at the DNA sequence level, but have different DNA methylation4,5 and his-tone modification profiles4 that might affect the penetrance of several diseases, such as cancer4 or autoimmune disorders6. But this phenom-enon is also observed at a single cell level: how can stem cells develop into any type of cell and how does a liver cell always give rise to two new liver cells after cell division? Again, epigenetics seems to be part of the answer as it has been described as one of the key factors in cellular differentiation7,8 (see the review by Meissner9 in this issue).
The importance of epigenetics in maintaining normal development and biology is reflected by the observation that many diseases develop when the wrong type of epigenetic marks are introduced or are added at the wrong time or at the wrong place10. For instance, a clear causality role for DNA methylation in cancer is suggested by hypermethylation of some genes (e.g., p16INK4a, p14ARF and MGMT) as an early event in tumorigenesis, as well as by tumor type-specific methylation landscape11. Here we summarize recent progress in the field of epigenetic research and its role in disease, preparing ourselves for the surprises that epigenetics might hold in the future.
Epigenetic modifications and their machineriesFor didactic purposes, epigenetic modifications can be grouped into three main categories: DNA methylation, histone modifications and nucleosome positioning. It is important to keep in mind the interplay between epige-netic factors—as the observed outcome is always the sum of their interac-tions—and the many positive and negative feedback mechanisms.
DNA methylation. The most widely studied epigenetic modification in humans is cytosine methylation. DNA methylation occurs almost exclusively in the context of CpG dinucleotides. The CpG dinucleotides tend to cluster in regions called CpG islands1, defined as regions of
Even before DNA was identified as the molecule of inheritance, scientists knew that not every gene in an organism can be active in each cell at all times. Even so, all cells in an organism share the same genetic information. Conrad Waddington coined the term ‘epigenetic landscape’1,2 for the molecular mechanisms that convert this genetic information into observable traits or phenotypes. In many instances, epigenetic gene expression patterns and associated phenotypes persist through mitosis or even meiosis, although no change in the primary DNA sequence has occurred. Consequently, epigenetics is generally understood to be the study of mechanisms that control gene expression in a potentially heritable way.
Recent breakthroughs in the understanding of the mechanisms under-lying epigenetic phenomena and their prevalence as contributors to the development of human disease have led to a greatly enhanced interest in epigenetic research.
On a molecular level, covalent modifications of cytosine bases and histones, and changes in the positioning of nucleosomes are commonly regarded as the driving epigenetic mechanisms. They are fundamental to the regulation of many cellular processes, including gene and microRNA expression, DNA-protein interactions, suppression of transposable ele-ment mobility, cellular differentiation, embryogenesis, X-chromosome inactivation and genomic imprinting.
In multicellular organisms, the ability of epigenetic marks to persist during development and potentially be transmitted to offspring may be necessary for generating the large range of different phenotypes that arise from the same genotype1,3–5. For instance, cloned animals
Epigenetic modifications and human diseaseAnna Portela1 & Manel Esteller1,2
Epigenetics is one of the most rapidly expanding fields in biology. The recent characterization of a human DNA methylome at single nucleotide resolution, the discovery of the CpG island shores, the finding of new histone variants and modifications, and the unveiling of genome-wide nucleosome positioning maps highlight the accelerating speed of discovery over the past two years. Increasing interest in epigenetics has been accompanied by technological breakthroughs that now make it possible to undertake large-scale epigenomic studies. These allow the mapping of epigenetic marks, such as DNA methylation, histone modifications and nucleosome positioning, which are critical for regulating gene and noncoding RNA expression. In turn, we are learning how aberrant placement of these epigenetic marks and mutations in the epigenetic machinery is involved in disease. Thus, a comprehensive understanding of epigenetic mechanisms, their interactions and alterations in health and disease, has become a priority in biomedical research.
1Cancer Epigenetics and Biology Program (PEBC), Bellvitge Biomedical Research Institute (IDIBELL), Barcelona, Catalonia, Spain. 2Institucio Catalana de Recerca i Estudis Avançats (ICREA), Barcelona, Catalonia, Spain. Correspondence should be addressed to M.E. ([email protected]).
Published online 13 October 2010; doi:10.1038/nbt.1685
rev iew©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

1058 volume 28 number 10 oCTober 2010 nature biotechnology
DNA methylation does not occur exclusively at CpG islands. The term CpG island shores, referring to regions of lower CpG density that lie in close proximity (~2 kb) of CpG islands, has recently been coined. The methylation of these CpG island shores is closely associated with tran-scriptional inactivation (Fig. 1b). Most of the tissue-specific DNA methy-lation seems to occur not at CpG islands but at CpG island shores19,20. Differentially methylated CpG island shores are sufficient to distinguish between specific tissues and are conserved between human and mouse. Moreover, 70% of the differentially methylated regions in reprogramming are associated with CpG island shores20,21.
DNA methylation is less frequently coupled with transcriptional acti-vation, as when, for instance, it occurs at gene bodies (Fig. 1c). Gene body methylation is common in ubiquitously expressed genes and is positively correlated with gene expression22. It has been proposed that it might be related to elongation efficiency and prevention of spurious initiations of transcription23.
DNA methylation and DNA methylation–associated proteins not only participate in gene transcription regulation in cis, but also act in trans, being involved in nuclear organization and in the establish-ment of specific chromosomal territories. An imprinted region can physically interact with sequences distant in the primary sequence or on different chromosomes. These physical interactions in trans can
more than 200 bases with a G+C content of at least 50% and a ratio of observed to statistically expected CpG frequencies of at least 0.6. CpG dinucleotides are usually quite rare in mammalian genomes (~1%). About 60% of human gene promoters are associated with CpG islands and are usually unmethylated in normal cells, although some of them (~6%) become methylated in a tissue-specific manner during early development or in differentiated tissues12 (Fig. 1a).
In general, CpG-island methylation is associated with gene silenc-ing. DNA methylation plays a key role in genomic imprinting, where hypermethylation at one of the two parental alleles leads to monoal-lelic expression13. A similar gene-dosage reduction is observed in X-chromosome inactivation in females14.
DNA methylation can inhibit gene expression by various mechanisms. Methylated DNA can promote the recruitment of methyl-CpG-binding domain (MBD) proteins. MBD family members in turn recruit histone-modifying and chromatin-remodeling complexes to methylated sites15,16. DNA methylation can also directly inhibit transcription by precluding the recruitment of DNA binding proteins from their target sites17. In contrast, unmethylated CpG islands generate a chromatin structure favorable for gene expression by recruiting Cfp1, which associates with histone methyl-transferase Setd1, creating domains rich in the histone methylation mark H3K4 trimethylation (H3K4me3; see below)18.
E2 E3
TF TF
RNA pol
E2E1
~
E4
E2 E3
MBDx MBDMBDMBD
DNMT
x
E1 E2E1
E2
E3 E4E3xxx
Repetitive sequenceRepetitive sequence
x
DNMT
E1 E1RNA pol
a
b
c
d
Unmethylated CpG island
Unmethylated CpG island shore
Methylated gene body
Methylated repetitive sequence
Methylated CpG island
Methylated CpG island shore
Unmethylated gene body
Unmethylated repetitive sequenceTranspositionRecombinationGenome instability
2kb
RNA pol
RNA pol
RNA pol
RNA pol RNA pol
RNA pol
RNA pol
RNA pol
TF TF
TF TF TF TF TF TF
~2kb E2E1
Figure 1 DNA methylation patterns. DNA methylation can occur in different regions of the genome. The alteration of these patterns leads to disease in the cells. The normal scenario is depicted in the left column and alterations of this pattern are shown on the right. (a) CpG islands at promoters of genes are normally unmethylated, allowing transcription. Aberrant hypermethylation leads to transcriptional inactivation. (b) The same pattern is observed when studying island shores, which are located up to 2 kb upstream of the CpG island. (c) However, when methylation occurs at the gene body, it facilitates transcription, preventing spurious transcription initiations. In disease, the gene body tends to demethylate, allowing transcription to be initiated at several incorrect sites. (d) Finally, repetitive sequences appear to be hypermethylated, preventing chromosomal instability, translocations and gene disruption through the reactivation of endoparasitic sequences. This pattern is also altered in disease.
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.
nature biotechnology volume 28 number 10 oCTober 2010 1059
are restored in induced pluripotent stem cells, suggesting a key role in origin and maintenance of pluripotent state28,29. Mechanisms of no-CpG methylation remain unclear29.
In addition to 5-methylcytosines, 5-hydroxymethyl-2′-deoxycytidine has also been observed. So far, 5-hydroxymethyl-2′-deoxycytidine has been reported in Purkinje cells (constituting 0.6% of total nucleotides) and in granule cells (constituting 0.2% of total nucleotides), but it seems not to be present in cancer cell lines30. These new DNA modifications need to be further studied to determine their implications for normal and diseased epigenetic regulation.
More work is also required in the development of new technological approaches31,32 and powerful analytical tools33, which have proven to be crucial for the progress of the field34. Massive parallel sequencing is providing lots of data, but its accurate analysis and interpretation, and its price remain as the last drawbacks to work with DNA methylomes at base resolution. Beyond sequencing-based technologies, the recently released, refined methylation arrays are worth considering for certain genomic questions.
DNA methylation is mediated by the DNMT family of enzymes that catalyze the transfer of a methyl group from S-adenosyl methionine to DNA. In mammals, five members of the DNMT family have been reported: DNMT1, DNMT2, DNMT3a, DNMT3b and DNMT3L, but only DNMT1, DNMT3a and DNMT3b possess methyltransferase activity.
regulate transcription, as shown for the H19 imprinting control region and the Osbpl1a/Impact loci24. Other examples of epigenetic players that cause three-dimensional (3D) rearrangements of the genome to regulate gene expression are the DNA methylation enzyme (DNA meth-yltransferase 1; DNMT1) that participates in the maintenance of the nucleolar compartment architecture25 and the methyl-CpG-binding domain (MBD) protein MeCP2, which is required for the formation of a silent chromatin loop at the Dlx5-Dlx6 locus26 (the 3D organization of the genome is discussed in more detail in the review by van Steensel and Dekker27 in this issue).
DNA methylation is not only linked to gene transcription regulation. A significant fraction of deeply methylated CpGs is found in repeti-tive elements (Fig. 1d). This DNA methylation is needed to protect chromosomal integrity, which is achieved by preventing reactivation of endoparasitic sequences that cause chromosomal instability, transloca-tions and gene disruption11.
Although DNA methylation mainly occurs in the CpG dinucle-otide context in mammals, non-CG methylation has recently been described in humans at CHG and CHH sites (where H is A, C or T). CHG and CHH methylation has been found in stem cells and seems to be enriched in gene bodies directly correlated with gene expres-sion and to be depleted in protein binding sites and enhancers28. The levels of non-CpG methylation decrease during differentiation and
b Histone modifications
c Chromatin remodeling
DNMT1
DNMT3A
DNMT3B
HAT
among others…
HDAC
HMT
HDM
Kinase
Phosph.
a DNA methylation Maintenance DNMT de novo DNMT
Hemimethylated DNA
It is recruited to methylated DNA by URHF1
Indirectly repressed by miR-29b, through SP1
Recruited by EZH2 and G9A (HMTs)
Interaction with nucleosomescontaining methylated DNA
Directly repressed by miR-29b
DNMT3A is recruited by HRR3me
Acetylation Methylation Phosphorylation
SWI/SNF ISWI Mi-2 INO80
HDAC1 and 2 can berecruited by MeCP2
mir-449a targets HDAC1
SET7 (HMT) regulatesDNMT1 stability
SETDB1 and Suv39h(HMTs) are recruitedby MBD1
KDM1B (HDM) is requiredto establish maternalgenomic imprint
LSD1 is a subunit of theNuRD complex
H3S10ph blocks H3K9me
H3S10ph facilitates H3recognition by GCN5 (HAT)
JAK2 phosphorylates H3,releasing HP1α
miR-9* and miR-124 mediate the BAF to npBAF switch
BRM is recruited by MeCP2
ISW2 excludes SWI/SNFfrom promoters bypostioning nucleosomes
NURF recognizes theH3K4me3
H4K16ac inhibits chromatinremodeling by ISWI
SET domains (HMT)recognize ISWI-remodelednucleosomal species
CHD5 expression isrepressed by CpG islandmethylation
MBD3 is an integralsubunit of Mi-2/NurD
HDAC and 2 are integralcomponents of Mi-2/NuRD
SWR1 removes the H2A-H2Bdimmers and replaces themwith H2A.Z-H2B dimmers
p400 has HAT activity
H2Aph enhances INO80recruitment
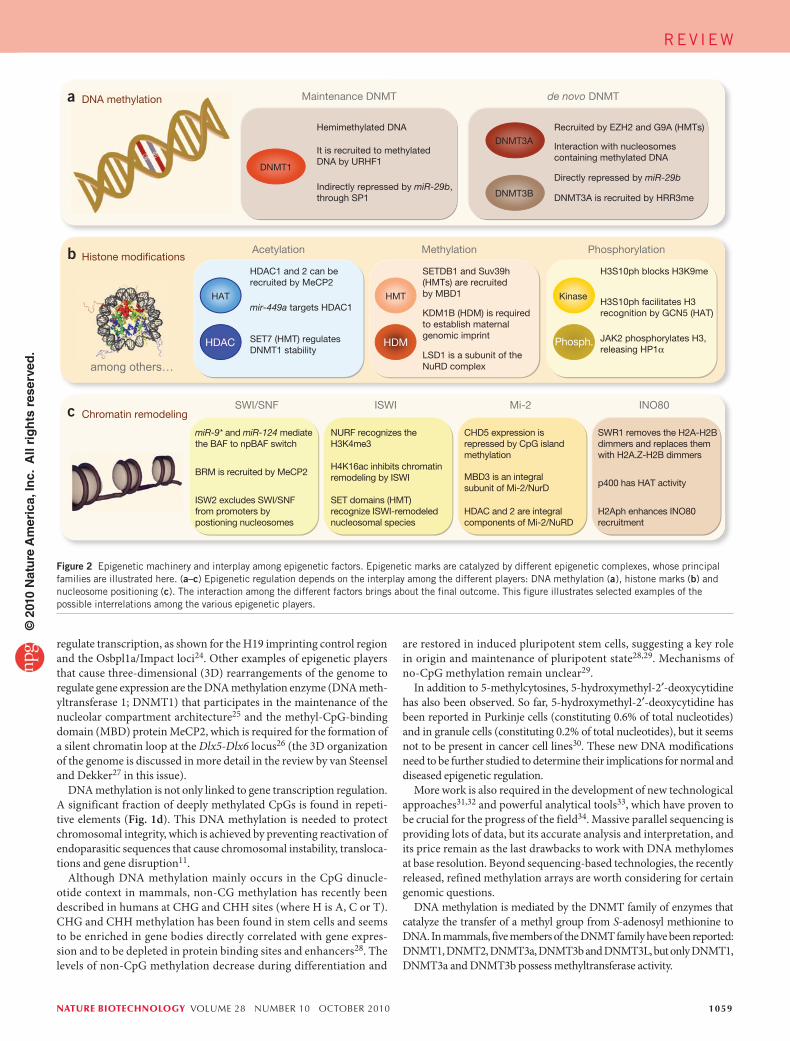
Figure 2 Epigenetic machinery and interplay among epigenetic factors. Epigenetic marks are catalyzed by different epigenetic complexes, whose principal families are illustrated here. (a–c) Epigenetic regulation depends on the interplay among the different players: DNA methylation (a), histone marks (b) and nucleosome positioning (c). The interaction among the different factors brings about the final outcome. This figure illustrates selected examples of the possible interrelations among the various epigenetic players.
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

1060 volume 28 number 10 oCTober 2010 nature biotechnology
All histones are subject to post-transcriptional modification. Several post-transcriptional modifications occur in histone tails: acetylation, methylation, phosphorylation, ubiquitination, SUMOylation and ADP-ribosylation52,54, among others (Fig. 3). Histone modifications have important roles in transcriptional regulation, DNA repair55, DNA repli-cation, alternative splicing56 and chromosome condensation52.
In relation to its transcriptional state, the human genome can be roughly divided into actively transcribed euchromatin and transcriptionally inac-tive heterochromatin. Euchromatin is characterized by high levels of acetylation and trimethylated H3K4, H3K36 and H3K79. On the other hand, heterochromatin is characterized by low levels of acetylation and high levels of H3K9, H3K27 and H4K20 methylation57. Recent studies have demonstrated that histone modification levels are predictive for gene expression. Actively transcribed genes are characterized by high levels of H3K4me3, H3K27ac, H2BK5ac and H4K20me1 in the promoter and H3K79me1 and H4K20me1 along the gene body58 (Fig. 4).
However, the notion of heterochromatin as a transcriptionally inactive region has been challenged by the discovery of numerous noncoding RNAs (ncRNAs) derived from heterochromatic loci51. For instance, Schizosaccharomyces pombe centromeric regions express siRNAs that bind to the RNA-induced transcriptional silenc-ing complex and provide sequence specificity to the complex. The RNA-induced transcriptional silencing complex is required for H3K9 methylation at centromeric repeats and for the recruitment of the histone methylation enzyme Clr4, which is essential for the spread-ing of heterochromatic domains51,59,60. But centromeric siRNAs are not the only ncRNAs that are capable of directing histone modifica-tions61. Well-known examples of this phenomenon in humans are the ncRNAs XIST and HOTAIR. XIST is involved in the silencing of the inactive X chromosome in females, through the recruitment of Polycomb-repressing complexes (PRC) with methyltransferase and histone ubiquitinase activity62,63. HOTAIR is a lincRNA transcribed from the HOXC cluster that represses genes in the HOXD cluster by recruiting the histone methyltransferase PRC2 (ref. 64).
All the modifications described so far are covalent post-transcrip-tional modifications. However, a new type of modification has recently been described. The histone H3 tail is clipped after the Ala21 residue, cutting off the N-terminal 21 residues and associated post-transcrip-tional modifications. This modification represents the first massive clearing of histone marks to be reported. Histone H3 clipping seems to be inhibited by H3K4me65.
Histones can be modified at different sites simultaneously. The core histones forming the nucleosome can each have several modifications, giving rise to cross-talk among the different marks. Communication among histone modifications can occur within the same site66, in the same histone tail67 and among different histone tails68 (Fig. 2b). Thus, a single histone mark does not determine outcome alone; instead, it is the combination of all marks in a nucleosome or region that specifies outcome. A recent paper has described the existence of up to 51 distinct ‘chromatin states’ based on the enrichment of specific combinations of histone modifications. Distinct biological roles are suggested for the different chromatin states69. An interesting case of co-existing histone modifications is found in ES cells within the ‘bivalent domains’, where the H3K4me3 active mark is found together with the H3K27me3 repres-sive mark at promoters of developmentally important genes. Bivalent domains enable ES cells to tightly regulate and rapidly activate gene expression during different developmental processes, but are lost with cell commitment70,71.
As mentioned before, all the epigenetic players interact with each other. An interesting example of the interplay between histone modi-fications and DNA methylation is the relationship between DNMT3L
The catalytic members of the DNMT family are customarily classi-fied into de novo DNMTs (DNMT3A and DNMT3B) and maintenance DNMTs (DNMT1). DNMT3A and DNMT3B are thought to be respon-sible for establishing the pattern of methylation during embryonic devel-opment. The de novo DNMTs are highly expressed in embryonic stem (ES) cells and downregulated in differentiated cells15. The DNMT3 family con-tains a third member, DNMT3L, which is required for establishing mater-nal genomic imprinting, despite being catalytically inactive35. DNMT3L is expressed during gametogenesis when genomic imprinting takes place. It acts as a general stimulatory factor for DNMT3a and DNMT3b and interacts and co-localizes with them in the nucleus36,37.
The maintenance DNMT, DNMT1, has a 30- to 40-fold preference for hemimethylated DNA, and also has de novo DNMT activity. DNMT1 is the most abundant DNMT in the cell and is transcribed mostly during the S phase of the cell cycle. It is most often needed to methylate hemimethy-lated sites that are generated during semi-conservative DNA replication (Fig. 2). In a cellular context the affinity of DNMT1 to newly synthesized DNA is increased by its interaction with the DNA polymerase processing factor proliferating cell nuclear antigen (PCNA), ensuring localization to the replication fork38. The ubiquitin-like plant homeodomain and RING finger domain-containing protein 1 (UHRF1) could perform a similar function, tethering DNMT1 to hemimethylated DNA, thanks to its SET and RING associated–domain, that shows strong preferential binding to hemimethylated CpGs39 (Fig. 2a).
However, the division of labor between de novo and maintenance methylation is not always so clear, and a revised model has recently been proposed by Jones and Liang40. The updated model still supports the idea that the bulk of DNA methylation in dividing cells would be maintained by DNMT1 in conjunction with UHRF1 and PCNA. But it also proposes that DNMT3A and DNMT3B, which have been shown to anchor strongly to nucleosomes containing methylated DNA41 (Fig. 2a), are compartmentalized in methylated regions, methylating the sites missed by DNMT1 at the replication fork. Finally, DNMT2, despite containing all the catalytic signature motifs of conventional DNMTs, has almost no DNMT activity. However, it has been reported that DNMT2 methylates tRNAAsp (ref. 42).
One of the most intriguing questions in the DNA methylation field is how the DNA methylation machinery is directed to specific sequences in the genome. Several mechanisms have been proposed, mainly suggest-ing interaction of DNMTs with other epigenetic factors41,43–47 (Fig. 2). More recently, small inhibitory (si)RNA-mediated, RNA-directed DNA methylation have also been described. In plants, RNA-directed DNA methylation is a stepwise process initiated by double-stranded RNAs that recruit DNMTs to catalyze de novo DNA methylation of specific regions including not only gene promoters but also repetitive sequences48–51. Although the process is well studied in plants and some of the RNA-directed DNA methylation components are conserved in mammals, it is still unclear if similar processes are involved in regulating DNA methyla-tion in animals. There are no reports suggesting the involvement of long intergenic ncRNA (lincRNAs) in DNA methylation.
Histone modifications. Histones are key players in epigenetics. The core histones H2A, H2B, H3 and H4 group into two H2.A-H2.B dimers and one H3-H4 tetramer to form the nucleosome. A 147-bp segment of DNA wrapped in 1.65 turns around the histone octamer and neighbor-ing nucleosomes are separated by, on average, ~50 bp of free DNA. The core histones are predominantly globular except for their N-terminal tails, which are unstructured52. Histone H1 is called the linker histone. It does not form part of the nucleosome but binds to the linker DNA (that is, the DNA separating two histone complexes), sealing off the nucleosome at the location where DNA enters and leaves53.
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature biotechnology volume 28 number 10 oCTober 2010 1061
Nucleosome positioning. Nucleosomes are a barrier to transcrip-tion that blocks access of activators and transcription factors to their sites on DNA, at the same time they inhibit the elongation of the transcripts by engaged polymerases. The packaging of DNA into nucleosomes appears to affect all stages of transcription, thereby regulating gene expression.
In particular, the precise position of nucleosomes around the tran-scription start sites (TSSs) has an important influence on the initia-tion of transcription. A preferential positioning of nucleosomes can be described at any given genomic locus. Nucleosome displacements of as few as 30 bp at TSS have been implicated in changes in the activity of RNA polymerase II. Moreover, the 5′ and 3′ ends of genes possess nucleosome-free regions needed to provide space for the assembly and disassembly of the transcription machinery. The loss of a nucleosome directly upstream of the TSS is tightly correlated with gene activation, whereas the occlusion of the TSS by a nucleosome is associated with gene repression77,78 (Fig. 4).
Nucleosome positioning not only determines accessibility of the tran-scription factors to their target DNA sequence but has also been reported to play an important role in shaping the methylation landscape79 (Fig. 4). Besides transcription regulation, nucleosome occupancy also participates in directing meiotic recombination events80.
The precise function of nucleosomes is influenced by the incorpora-tion of different histone variants. Histone variants are distinguished from core histones by the fact that they are expressed outside of S phase and are incorporated into chromatin independently from DNA replication. They differ from core histones in their tails, in their domain structure and in a few key amino acids57. Histone variants regulate nucleosome positioning and gene expression23. For example, the incorporation of the histone variant H2A.Z protects genes against DNA methylation81. Thus, the interplay among different epigenetic partners becomes evident once
and H3K4. DNMT3L specifically interacts with histone H3 tails, inducing de novo DNA methylation by recruitment of DNMT3A; however, this interaction is strongly inhibited by H3K4me43. Furthermore, several histone methyltransferases have also been reported to direct DNA methylation to specific genomic targets by recruiting DNMTs44,45, helping in this way to set the silenced state established by the repressive histone marks. Moreover, histone methyltransferases and demethylases can also modulate the stability of DNMT proteins, thereby regulating DNA methylation levels46,47 (Fig. 2b). On the other hand, DNA methylation can also direct his-tone modifications. For instance, methylated DNA mediates H3K9me through MeCP2 recruitment72.
Many enzymes that catalyze covalent post-transcriptional modi-fications have been described52,73. Because the modifications are dynamic, enzymes to remove these post-transcriptional modifications have also been reported52,73,74. However, the list of histone modifi-cations, its writers and erasers, might not yet be completed. Of the enzymes that modify histones, methyltransferases, histone demethy-lases and kinases are the most specific to individual histone subunits and residues52,75. Conversely, most of the histone acetyltransferases (HATs) and histone deacetylases (HDACs) are not highly specific and modify more than one residue.
Many transcriptional co-activators (e.g., GCN5, PCAF, CBP, p300, Tip60 and MOF) have been reported to possess intrinsic HAT activity, whereas many transcriptional co-repressor complexes (e.g., mSin3a, NCoR/SMRT and Mi-2/NuRD) contain subunits with HDAC activity66. Surprisingly, it has recently been reported that HDACs and HATs are both targeted to transcribed regions of active genes by phosphorylated RNA polymerase II. Thus, most HDACs in the human genome function to reset chromatin by removing acetylation at active genes, whereas HATs, by contrast, are mainly linked to transcriptional activation76.
N-S R G K Q G G K A R A K A K S… …L R K G N… …L G K V T… …L P K K T E S H…-C9 151
P A
5
A
13
A A
36
A M
99 120119
MU P
H2A
14
P
N-…P A K S A... …K G S K K A V T K… …V Y K V L… …Y N K R S… …L A K H A… …K A V T K…-C5
A
12
A
15 20
M
43
A
85
A
108
A
116 120
UAMA
UA
H2B
MMMM
N-A R T K Q T A R K S T G G K A P R K Q L A T K A A R K S A P A T G G V K K P H R Y R P G T V… 2 3
P
4 8 9
MA
10
P
11
P
17
A
18 23
MA
2627
MA
28
P
36
M
37
A
14
MA
MA
41
P
…Y Q K S T… …D F K T D…-C56
M
79
45
P
MAH3.1
N-S G R G K G G K G L G K G G A K R H R K... …R I S G L… …V L K V F… …K R K… …L K R Q…-C1
P M
3
A
5
A
8 12
MA A
16 20
MA
47
P M
59
A
77
A
79
A
91
M
92
H4
H1.4
N-S E T A P… …A E K T P V… …K S A G A A K R K A S… …K A V A A S K E R… …A L K K A L...26
M
273 3617
A
3418 46 52
…K S L V S K G T L V Q T K… …S F K L N… …K S A K K T… …K K A K S… …P K S P A… -C
P P P A P UA A
64
M
85
A
90
A
MA
97
MA
106
M M
149 154
P
63
168
MA
172
P
186
P
Figure 3 Histone modifications. All histones are subject to post-transcriptional modifications, which mainly occur in histone tails. The main post-transcriptional modifications are depicted in this figure: acetylation (blue), methylation (red), phosphorylation (yellow) and ubiquitination (green). The number in gray under each amino acid represents its position in the sequence.
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

1062 volume 28 number 10 oCTober 2010 nature biotechnology
peptidase 6 (DPP6) in melanomas are other well-studied examples of hypomethylated genes in cancer19,92. The most common LOI event due to hypomethylation is insulin-like growth factor 2 (IGF2), which has been reported in a wide range of tumor types, including breast, liver, lung and colon cancer96.
In contrast to global DNA hypomethylation, hypermethylation is observed at specific CpG islands (Fig. 1a). The transcriptional inactiva-tion caused by promoter hypermethylation affects genes involved in the main cellular pathways: DNA repair (hMLH1, MGMT, WRN, BRCA1), vitamin response (RARB2, CRBP1), Ras signaling (RASSFIA, NOREIA), cell cycle control (p16INK4a, p15INK4b, RB), p53 network (p14ARF, p73 (also known as TP73), HIC-1) and apoptosis (TMS1, DAPK1, WIF-1, SFRP1), among others15. Hypermethylated promoters have been pro-posed as a new generation of biomarkers and hold great diagnostic and prognostic promise for clinicians97 (reviewed in more detail by Jones and colleagues98 in this issue).
However, even though the focus of most studies is on CpG islands located in promoters, recent findings suggest that most of the aberrant DNA meth-ylation in cancer occurs in CpG island shores (e.g., in HOXA2 and GATA2) (Fig. 1b). Notably, most changes in CpG island shores (45–65%) seem to be associated with regions that become hypermethylated during normal tissue differentiation (e.g., in TGFB1 and PAX5)19,20. Differential DNA methylation seems to correlate with gene expression at CpG island shores just as it does with CpG islands21.
Human tumors are also characterized by an overall miRNA downregu-lation99 often caused by hypermethylation at the miRNA promoters100. For example, miR-124a is repressed by hypermethylation, mediating CDK6 activation and Rb phosphorylation101. Interestingly, inactivation of miRNA expression by hypermethylation is not only linked to cancer but also to metastasis development. Silencing of miR-148, miR-34b/c and miR-9 by promoter hypermethylation favors tumor dissemination from the original location102.
Hypermethylation patterns are tumor-type specific and it is still unclear why certain regions become hypermethylated, whereas others remain unmethylated. One possibility is that inactivation of particular genes confers a growth advantage, resulting in clonal selection15. In some cases, it has also been proposed that aberrant CpG-island meth-ylation could be due to the recruitment of DNMTs and HDACs to specific target genes mediated by fusion proteins, such as the promyelo-cytic leukemia–retinoic acid receptor-α (PML–RARA) fusion protein, expressed in some leukemias103. Another possibility is the spreading of methylation from highly methylated sequences to their surroundings is more pronounced in cancer. It has been reported that epigenetic silencing by DNA methylation can span 1-Mb-long regions of a chro-mosome104, resembling the loss of heterozygosity often observed in human tumors. This global distortion of the DNA methylation pattern could also be mediated by dysregulation of DNMT expression. DNMT1 and DNMT3b are overexpressed in many tumor types105. Moreover, DNMT expression can also be regulated by miRNAs. The miR-29 family is known to directly target and downregulate DNMT3A and DNMT3B, and indirectly target DNMT1 (ref. 106) (Fig. 2a).
Histone modifications. The most prominent alteration in histone modifi-cation in cancer cells is a global reduction of monoacetylated H4K16 (ref. 107). Loss of acetylation is mediated by HDACs, which have been found to be overexpressed108 or mutated109 in different tumor types. The main class of HDACs implicated in this process is the Sirtuin family of proteins110. Gene expression and deacetylase activity of SirT1 is upregulated in several cancer types. Moreover, SirT1 interacts with DNMT1, thus affecting DNA methylation patterns25. HDAC expression can be regulated by miRNAs, such as miR-449a, which, by repressing the expression of HDAC-1 in
more. The nucleosome remodeling machinery is influenced by DNA methylation82 and has been linked with specific histone modifications83 (Fig. 2c). MicroRNAs (miRNAs) can also regulate histone variant replace-ment84 or interact with chromatin remodeling complexes mediating the exchange of specific subunits85.
Several groups of large macromolecular complexes are known to move, destabilize, eject or restructure nucleosomes in an ATP hydrolysis–depen-dent manner. These complexes, known as chromatin remodeling com-plexes, can be classified into four families (SWI/SNF, ISWI, CHD and INO80) that share similar ATPase domains but differ in the composition of their unique subunits86.
In the first of these families, the SWI/SNF family, members have as a catalytic unit either Brahma (BRM) or BRG1, which share ~75% of iden-tity but differ in their first 60 amino acids. SWI/SNF family complexes are master regulators of gene expression, regulating expression of—among others—FOS, CSF-1, CRYAB, MIM-1, p21 (also known as CDKN1A), HSP70, VIM and CCNA2. Moreover, SWI/SNF has also been reported to modulate alternative splicing87.
Many members of the second class, the ISWI family, such as ACF and CHRAC, have been reported to promote chromatin assembly and to repress transcription. However, NURF, another complex of this family, is capable of activating RNA polymerase II thus participating in transcrip-tional activation88.
In the CHD family, some members participate in the sliding and ejec-tion of nucleosomes, promoting transcription; however, others, such as the Mi-2/NuRD complex, have repressive roles and contain HDAC activity and MBD proteins88 (Fig. 2c).
Members of the last group, the INO80 family, have been reported to participate in multiple cellular processes: transcriptional activation, DNA repair, telomere regulation, chromosome segregation and DNA replication among others86. However, the SWR1 member has the unique ability to restructure the nucleosome, removing the H2A-H2B dimers and replacing them with H2A.Z-H2B dimers88 (Fig. 2c).
Epigenetic modifications in cancerIn addition to featuring classic genetic mutations, cancer cells present a profoundly distorted epigenetic landscape (Table 1). The cancer epigenome is characterized by global changes in DNA methylation, histone modification patterns and chromatin-modifying enzyme-expression profiles11,89, which play important roles in cancer initia-tion and progression.
DNA methylation. Cancer cells are characterized by a massive global loss of DNA methylation90 (20–60% less overall 5-methyl-cytosine). At the same time, the acquisition of specific patterns of hypermethylation at the CpG islands of certain promoters is frequently observed (Fig. 1a).
Global hypomethylation occurs mainly at repetitive sequences, promoting chromosomal instability, translocations, gene disruption and reactivation of endoparasitic sequences90,91 (Fig. 1d). A clear case is the LINE family member L1, which has been shown to be hypomethylated in a wide range of cancers, including breast, lung, bladder and liver tumors92.
Hypomethylation at specific promoters can activate the aber-rant expression of oncogenes and induce loss of imprinting (LOI) in some loci. For instance, MASPIN (also known as SERPINB5), a tumor suppressor gene that becomes hypermethylated in breast and prostate epithelial cells93, appears to be hypomethylated in other tumor types. MASPIN hypomethylation, and therefore its expres-sion, increases with the degree of dedifferentiation of some types of cancer cells94,95. S100P in pancreatic cancer, SNCG in breast and ovar-ian cancers and melanoma-associated gene (MAGE) and dipeptidyl
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature biotechnology volume 28 number 10 oCTober 2010 1063
by cytokine signaling. H3K41ph prevents the binding of heterochromatin protein1α (HP1α) to this region of H3, increasing the expression of the genes located there, as it was reported in the lmo2 promoter. JAK2 is frequently activated by chromosomal translocations or point mutations in hematological malignancies126.
Nucleosome positioning. All families of chromatin remodelers have been tied to cancer, although in most cases the molecular mechanisms underlying their function remain unclear. For instance, BRG1 and BRM, the ATPase subunits of SWI/SNF complexes, have been characterized as tumor suppressors and are silenced in about 15–20% of primary non-small-cell lung cancers127. Surprisingly, an oncogenic role for BRG1 as a p53 destabilizer has also been proposed128. Mutations in SNF5, a subunit of the SWI/SNF remodeling complex, have been observed in sporadic renal rhabdoid tumors and in choroid plexus carcinomas, meduloblasto-mas and central primitive neuroectodermal tumors129.
Nucleosome remodeling is also involved in the transcriptional repres-sion by promoter hypermethylation (Fig. 4). Promoter hypermethylation results in the occupation of the TSS by a nucleosome, as has been reported for MLH1 in colon cancer130. The genes encoding subunits of the chro-matin remodeling complexes (e.g., CHD5 (ref. 131)) themselves are also targets of CpG island hypermethylation in cancer, thereby downregu-lating its expression and impairing the normal chromatin remodeling processes (Fig. 2c).
In addition to nucleosome positioning, histone variants have also been related to cancer. For example, increased expression of MacroH2A is involved in senescence. Thus, lung tumors with highly expressed MacroH2A have a better prognosis, with lower proliferation rates and less frequent recurrence132.
Epigenetic modifications in neurodevelopmental disordersThe central nervous system is one of the most complex systems in humans. Not only do the different regions of an organ present different expression patterns, but the same cell type has different transcriptional regulation depending on its localization in the organ133. The mitotic exit, when neu-ral cells lose their multipotency, is a key step in nervous system develop-ment85,134, requiring a very precise tuning of the transcriptional program. Epigenetic factors are key players in this regulation. Genetic mutations in epigenetic genes cause dysfunctions that lead to certain neurodevel-opmental disorders. Here, we classify them according to the epigenetic machinery that becomes mutated.
prostate cancer cells, regulates cell growth and viability111 (Fig. 2b). In addition to alteration in HDAC expression, several cancer types (e.g., colon, uterus, lung and leukemia) also bear translocations leading to the formation of aberrant fusion proteins, mutations or deletions in HATs and HAT-related genes112,113, thus contributing to the global imbalance of histone acetylation.
Besides the global loss of H4K16ac, cancer cells suffer a global loss of the active mark H3K4me3 (ref. 114) and the repressive mark H4K20me3 (ref. 107), and a gain in the repressive marks H3K9me (ref. 115) and H3K27me3 (ref. 116). Altered distribution of the histone methyl marks in cancer cells is mainly due to the aberrant expression of both histone methyltransferases and histone demethy-lases75. A recent publication has described inactivating mutations in the histone methyltransferase SETD2 and in the histone demethylase UTX and JARID1C in renal carcinomas117. Another example is the histone methyltransferase EZH2—a subunit of the PRC2 and PRC3 complexes—which enhances proliferation and neoplastic transfor-mation and is overexpressed in several cancer types. Overexpression of the lincRNA HOTAIR in breast tumors and metastases retargets PRC2 and alters H3K27me3 landscape118. Moreover, EZH2 expres-sion is upregulated in many cancer tumors due to the genomic loss of miR-101 (ref. 119). In addition to its histone methyltransferase activ-ity, EZH2 interacts with DNMTs directly controlling DNA methyla-tion116. NSD1, another histone methyltransferase, has been reported to undergo promoter DNA methylation-dependent silencing in neu-roblastomas120. DOT1L, the major H3K79 histone methyltransferase, is essential for the establishment of a euchromatic state that allows the expression of tumor suppressor genes121,122.
In leukemias, the presence of mixed lineage leukemia (MLL) fusion oncoproteins leads to aberrant patterns of H3K79 and H3K4 methylation, resulting in altered gene expression of MLL targets123,124. Some histone demethylases (e.g., GASC1, LSD1, JmjC and UTRX) have also been shown to be upregulated or amplified in several cancers, including prostate cancer and squamous cell carcinomas125.
Although further studies are needed, histone phosphorylation also seems to be relevant in cancer. Histone phosphorylation plays a role in DNA damage-repair response, chromosome stability and apopto-sis. Recently JAK2, a nonreceptor tyrosine kinase that regulates several cellular processes by inducing cytoplasmic signaling cascade, has been reported also to be present in the nucleus, directly phosphorylating H3Y41 (Fig. 2b). Phosphorylated H3Y41 (H3Y41ph) levels are regulated
H3K9 H3K9 H3K9 H3K9H3K27
H4K20 H4K20 H4K20H3K27 H3K27
H3K4
H3K36H3K79
M
M
M
MM
MM
MMM
MM
M
A AA
Figure 4 Nucleosome positioning patterns. Nucleosome positioning plays an important role in transcriptional regulation. Transcriptionally active gene promoters possess a nucleosome-free region at the 5′ and 3′ untranslated region, providing space for the assembly and disassembly of the transcription machinery. The loss of a nucleosome directly upstream of the TSS is also necessary for gene activation, whereas the occlusion of this position leads to transcription repression. DNA methylation regulates transcription, and thus interferes with nucleosome positioning. Methylated DNA seems to be associated with ‘closed’ chromatin domains, where DNA is condensed into strictly positioned nucleosomes, thereby impeding transcription. Conversely, unmethylated DNA is associated with ‘opened’ chromatin domains, which allow transcription.
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

1064 volume 28 number 10 oCTober 2010 nature biotechnology
in the cAMP-response element binding protein (CBP), in another 3% of cases with mutations in EP300 and in ~42% of cases with an unidentified cause. CBP and EP300 function as transcriptional co-activators in addition to their HAT activity135. In Cbp+/− mice H2B acetylation is reduced by more than 30%, suggesting that the failure in long-term memory forma-tion could be explained by chromatin changes in one or several loci that control memory storage141.
The neurodevelopmental disease Coffin-Lowry syndrome is a rare X-linked disorder caused by loss-of-function mutations in RSK2, a serine/threonine protein kinase. RSK2 participates in the MAP kinase pathway, inducing the transient transcription of a set of genes. RSK2 mediates H3S10ph directly, changing chromatin structure and facilitating the binding of CBP, which acetylates H3 residues. Thus, RSK2 promotes gene transcription through chromatin opening142.
Nucleosome positioning. ATRX syndrome is an X-linked disorder caused by mutations in ATRX, a member of the Snf2 family of chromatin
DNA methylation. Rett syndrome is an X-linked neurological disease caused by point mutations in the MBD protein MeCP2. Both upregu-lation and downregulation of MeCP2 in the brain are associated with neurodevelopmental defects. Customarily, MeCP2 has been considered to function as a gene silencer, mediating the recruitment of HDACs to methylated DNA (Fig. 2b). Recently, new data have highlighted impor-tant roles for MeCP2 in chromatin architecture, regulation of mRNA splicing135,136 and active transcription of genes (e.g., Sst, Gprin1)137. Although transcriptional alterations have been described in some genes (e.g., Fkbp5, Mobp, Ddc and S100a9)138, imprinted regions (e.g., DLX5) and miRNAs (e.g., miR-184)139,140, MeCP2 deficiency does not result in high levels of genome-wide transcriptional alteration. It stills remains unknown whether or not the described alterations are causative.
Histone modifications. Rubinstein-Taybi syndrome is an autosomal dominant disorder associated with the dysfunction of a HAT. It is a geneti-cally heterogeneous disease associated in ~55% of cases with mutations
Table 1 Epigenetic modifications in human diseases
Aberrant epigenetic mark Alteration ConsequencesExamples of genes affected and/or resulting disease
Cancer
DNA methylation CpG island hypermethylation Transcription repression MLH1 (colon, endometrium, stomach11), BRCA1 (breast, ovary11), MGMT (several tumor types11), p16INK4a (colon11)
CpG island hypomethylation Transcription activation MASPIN (pancreas92), S100P (pancreas92), SNCG (breast and ovary92), MAGE (melanomas92)
CpG island shore hypermethylation Transcription repression HOXA2 (colon20),GATA2 (colon20)
Repetitive sequences hypomethylation Transposition, recombination genomic instability
L1 (ref. 11), IAP11, Sat2 (ref. 107)
Histone modification Loss of H3 and H4 acetylation Transcription repression p21WAF1 (also known as CDKN1A)11
Loss of H3K4me3 Transcription repression HOX genes
Loss of H4K20me3 Loss of heterochromatic structure Sat2, D4Z4 (ref. 107)
Gain of H3K9me and H3K27me3 Transcription repression CDKN2A, RASSF1 (refs. 115–116)
Nucleosome positioning Silencing and/or mutation of remodeler subunits
Diverse, leading to oncogenic transformation BRG1, CHD5 (refs. 127–131)
Aberrant recruitment of remodelers Transcription repression PLM-RARa103 recruits NuRD
Histone variants replacement Diverse (promotion cell cycle/destabilization of chromosomal boundaries)
H2A.Z overexpression/loss
Neurological disorders
DNA methylation CpG island hypermethylation Transcription repression Alzheimer’s disease (NEP)135
CpG island hypomethylation Transcription activation Multiple sclerosis (PADI2)135
Repetitive sequences aberrant methylation Transposition, recombination genomic instability
ATRX syndrome (subtelomeric repeats)135,143
Histone modification Aberrant acetylation Diverse Parkinson’s and Huntington’s diseases135
Aberrant methylation Diverse Huntington’s disease and Friedreich’s ataxia135
Aberrant phosphorylation Diverse Alzheimer’s disease135
Nucleosome positioning Misposition in trinucleotide repeats Creation of a ‘closed’ chromatin domain Congenital myotonic dystrophy151
Autoimmune diseases
DNA methylation CpG island hypermethylation Transcription repression Rheumatoid arthritis (DR3)154,155
CpG island hypomethylation Transcription activation SLE (PRF1, CD70, CD154, AIM2)6
Repetitive sequences aberrant methylation Transposition, recombination genomic instability
ICF (Sat2, Sat3), rheumatoid arthritis (L1)152,155
Histone modification Aberrant acetylation Diverse SLE (CD154, IL10, IFN-γ)6
Aberrant methylation Diverse Diabetes type 1 (CLTA4, IL6)159
Aberrant phosphorylation Diverse SLE (NF-κB targets)
Nucleosome positioning SNPs in the 17q12-q21 region Allele-specific differences in nucleosome distribution
Diabetes type 1 (CLTA4, IL6)
Histone variants replacement Interferes with proper remodeling Rheumatoid arthritis (histone variant macroH2A at NF-κB targets)157
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature biotechnology volume 28 number 10 oCTober 2010 1065
Nucleosome positioning. It has been suggested that the amplifica-tion of CTG repeats in congenital myotonic dystrophy is a very strong nucleosome positioning signal that mediates the creation of a closed chro-matin domain151. Despite this fact, which needs further investigation, little is known about the possible implications of nucleosome positioning or histone variants in neuronal malignancies.
Epigenetic modifications in autoimmune diseasesAutoimmune diseases are characterized by the breakdown of immune tolerance to specific self-antigens. Different types of epigenetic alterations have been reported in this type of disorder (Table 1).
DNA methylation. Most of the research relating autoimmunity disorders and epigenetic changes has focused on DNA methylation alterations. In fact, one of the best known autoimmune diseases, the ICF (immunodefi-ciency, centromeric instability and facial anomalies) syndrome, is caused by heterozygous mutations in DNMT3B. ICF patients show marked DNA hypomethylation in the pericentromeric satellite 2 and 3 repeats, alpha satellite sequences, Alu sequences and the D4Z4 and NBL2 repeats. Conversely, ICF patients have almost unchanged global DNA methylation levels143,147, although several genes regulating development, neurogenesis and immune function have aberrant expression152.
Other autoimmune diseases, unrelated to mutations in the DNA methylation machinery, also present global hypomethylation, as is the case for systemic lupus erythematosus (SLE) and rheumatoid arthritis. The hypomethylated regions are not yet well defined, although some hypomethylated sites have been reported. SLE patients have DNA hypomethylation in PRF1, CD70, CD154, IFGNR2, MMP14, LCN2, CSF3R and AIM2 among other genes, and also in the ribosomal RNA gene promoter, 18S and 28S (ref. 6). The mechanisms responsible for this widespread hypomethylation are beginning to be revealed. It has been recently reported that hypomethylation in SLE is partially mediated by miR-21 and miR-148a that directly and indirectly target DNMT1 (ref. 153). In rheumatoid arthritis, not only hypomethylated sites (e.g., in L1 and IL6) but also hypermethylated sites (e.g., in DR3) have been described154,155.
Histone modifications. Little is known about the role of histone modi-fications in autoimmune diseases, although initial studies are begin-ning to shed some light in this area. In human SLE T-cells, the HDAC inhibitor trichostatin A reverses the aberrant expression of CD154, IL10 and interferon (IFN)-γ products156. A role for histone modifications in rheumatoid arthritis has also been described. Because the transcrip-tion factor NF-κB—a key regulator inflammatory—binds very poorly to nucleosomal DNA, histone modifications are needed to allow effi-cient NF-κB binding to its targets: histone H3K9 and S10 (also known as PSMD6) phosphoacetylation, reduction in H3K9me and increase in H3/H4 acetylation157. Thus, in rheumatoid arthritis, the reduced activity of HDACs plays a key role in regulating NF-κB–mediated gene expression158. Patients with type 1 diabetes also present a characteristic pattern of histone marks, showing lymphocytes but not monocytes with increased H3K9me2 in a subset of genes associated with autoimmune and inflammatory pathways (e.g., CLTA4, IL6)159.
However, histone modifications have a role not only in transcription regulation. Nucleosomes are key autoantigens in SLE, being present in the circulation because of increased apoptosis and/or insufficient clearance. In apoptosis, histone modifications occur, such as H2BS14 phosphorylation160, H3T45 phosphorylation161, H3K4 trimethyla-tion162, H4 triacetylation at K8, K12 and K16 (ref. 163) as well as H2BK12 acetylation164. It has been suggested that histone modifica-tions arising during apoptosis make released apoptotic nucleosomes
remodelers. The ATRX protein interacts with the SET domain of the histone methyltransferase EZH2, the Daxx transcriptional cofactor, MeCP2 and the chromoshadow domain of HP1 proteins. It partici-pates among other cellular processes in heterochromatin formation, chromosome alignment at the meiotic spindle, chromosome cohesion in somatic cells and maintenance of X-chromosome inactivation in women. Because no DNA repair defects or genomic instability occurs in ATRX patients, it has been suggested that ATRX may regulate the transcription of a specific set of target genes. Although global DNA methylation is unchanged in ATRX patients, aberrant DNA methyla-tion in some repetitive sequences has been reported135,143.
Epigenetic modifications in neurodegenerative and neurological diseasesRecent studies have also shed some light on the relationship between epigenetic alterations and neurodegenerative and/or neurological diseases. The majority of the evidence centers on DNA methylation and histone modification (Table 1).
DNA methylation. DNA methylation patterns appear to be distorted in a great deal of neurological diseases, giving rise to hyper- and hypomethylated sites. For instance, FMR1 promoter hypermethyla-tion has been described in Fragile X syndrome patients. Fragile X syndrome is caused by a CGG trinucleotide repeat expansion in the 5′-untranslated region of FMR1. Expansion of the CGG trinucleotide repeats to >200 copies induces methylation of FMR1, leading to its transcriptional silencing144. Other reported cases of hypermethy-lated promoters include neprilysin (NEP, also known as MME) in Alzheimer’s disease, FXN in Friedreich’s ataxia and SMN2 in spinal muscular atrophy135.
Conversely, hypomethylated sites have also been reported. For example, the substantia nigra of Parkinson’s patients overexpresses tumor necrosis factor alpha (TNFα) due to its promoter hypomethy-lation, thereby inducing apoptosis of neuronal cells145. Other cases of hypomethylation were reported in the promoter region of PADI2 for multiple sclerosis patients135 and in the Avp enhancer for mice subjected to early-life stress146. Alterations in DNA methylation pat-terns not only affect gene promoters but may also lead to LOI. Classic examples of LOI are the Prader-Willi and the Angelman syndromes. Both diseases involve aberrant DNA methylation in the imprint-ing controlled region at 15q11-q13. Prader-Willi syndrome arises from the loss of paternally expressed genes in this region, whereas Angelman syndrome arises from the loss of the maternally expressed UBE3A gene147.
Histone modifications. The pattern of histone marks is also altered in neurological diseases, histone hypoacetylation being the most fre-quently observed change. A good example of histone hypoacetylation is amyotrophic lateral sclerosis (ALS). ALS patients have aggregates of the protein FUS in cytoplasmic deposits of misfolded proteins. FUS is able to bind CBP, strongly inhibiting its HAT activity and to negatively regulate specific CREB target genes. Thus, overexpression of FUS induces histone hypoacetylation135. Other cases of hypoacetylation in neurological diseases are found in Parkinson’s and Huntington’s disease135 and Friedreich’s ataxia148. Despite histone hypoacetylation, more changes relating neurological diseases and histone marks have been reported. For example, histone acetylation and phosphoryla-tion alterations are typical in Alzheimer’s disease and epilepsy, H3K9 hypertrimethylation has been described in Huntington’s disease135 and Friedreich’s ataxia149 and the histone demethylase PHF8 has been involved in X-linked mental retardation150.
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

1066 volume 28 number 10 oCTober 2010 nature biotechnology
would be of enormous use in basic and applied research and would be relevant for focusing pharmacological research on the most promising epigenetic targets. A key topic for future research is the implementation of mechanisms for the release of whole genome methylation and histone modification maps into public databases.
COMPETING FINANCIAL INTERESTSThe authors declare no competing financial interests.
Published online at http://www.nature.com/naturebiotechnology/. Reprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions/.
1. Esteller, M. Epigenetics in evolution and disease. Lancet 372, S90–S96 (2008).2. waddington, C.H. Introduction to Modern Genetics (Macmillan, 1939).3. Rideout, w.M., III, Eggan, K. & Jaenisch, R. Nuclear cloning and epigenetic reprogram-
ming of the genome. Science 293, 1093–1098 (2001).4. Fraga, M.F. et al. Epigenetic differences arise during the lifetime of monozygotic twins.
Proc. Natl. Acad. Sci. USA 102, 10604–10609 (2005).5. Kaminsky, Z.A. et al. DNA methylation profiles in monozygotic and dizygotic twins.
Nat. Genet. 41, 240–245 (2009).6. Javierre, B.M. et al. Changes in the pattern of DNA methylation associate with twin
discordance in systemic lupus erythematosus. Genome Res. 20, 170–179 (2010).7. Chi, A.S. & Bernstein, B.E. Developmental biology. Pluripotent chromatin state. Science
323, 220–221 (2009).8. Meissner, A. et al. Genome-scale DNA methylation maps of pluripotent and differenti-
ated cells. Nature 454, 766–770 (2008).9. Meissner, A. Epigenetic modifications in pluripotent and differentiated cells. Nat.
Biotechnol. 28, 1079–1088 (2010).10. Esteller, M. CpG island hypermethylation and tumor suppressor genes: a booming
present, a brighter future. Oncogene 21, 5427–5440 (2002).11. Esteller, M. Cancer epigenomics: DNA methylomes and histone-modification maps.
Nat. Rev. Genet. 8, 286–298 (2007).12. Straussman, R. et al. Developmental programming of CpG island methylation profiles
in the human genome. Nat. Struct. Mol. Biol. 16, 564–571 (2009).13. Kacem, S. & Feil, R. Chromatin mechanisms in genomic imprinting. Mamm. Genome
20, 544–556 (2009).14. Reik, w. & Lewis, A. Co-evolution of X-chromosome inactivation and imprinting in
mammals. Nat. Rev. Genet. 6, 403–410 (2005).15. Esteller, M. Epigenetic gene silencing in cancer: the DNA hypermethylome. Hum. Mol.
Genet. 16 Spec No 1, R50–R59 (2007).16. Lopez-Serra, L. & Esteller, M. Proteins that bind methylated DNA and human cancer:
reading the wrong words. Br. J. Cancer 98, 1881–1885 (2008).17. Kuroda, A. et al. Insulin gene expression is regulated by DNA methylation. PLoS ONE
4, e6953 (2009).18. Thomson, J.P. et al. CpG islands influence chromatin structure via the CpG-binding
protein Cfp1. Nature 464, 1082–1086 (2010).19. Irizarry, R.A. et al. The human colon cancer methylome shows similar hypo- and hyper-
methylation at conserved tissue-specific CpG island shores. Nat. Genet. 41, 178–186 (2009).
20. Doi, A. et al. Differential methylation of tissue- and cancer-specific CpG island shores distinguishes human induced pluripotent stem cells, embryonic stem cells and fibro-blasts. Nat. Genet. 41, 1350–1353 (2009).
21. Ji, H. et al. Comprehensive methylome map of lineage commitment from haematopoi-etic progenitors. Nature 467, 338–342 (2010).
22. Hellman, A. & Chess, A. Gene body-specific methylation on the active X chromosome. Science 315, 1141–1143 (2007).
23. Zilberman, D., Gehring, M., Tran, R.K., Ballinger, T. & Henikoff, S. Genome-wide analy-sis of Arabidopsis thaliana DNA methylation uncovers an interdependence between methylation and transcription. Nat. Genet. 39, 61–69 (2007).
24. Zhao, Z. et al. Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nat. Genet. 38, 1341–1347 (2006).
25. Espada, J. et al. Epigenetic disruption of ribosomal RNA genes and nucleolar archi-tecture in DNA methyltransferase 1 (Dnmt1) deficient cells. Nucleic Acids Res. 35, 2191–2198 (2007).
26. Horike, S., Cai, S., Miyano, M., Cheng, J.F. & Kohwi-Shigematsu, T. Loss of silent-chromatin looping and impaired imprinting of DLX5 in Rett syndrome. Nat. Genet. 37, 31–40 (2005).
27. van Steensel, B. & Dekker, J. Genomics tools for unraveling chromosome architecture. Nat. Biotechnol. 28, 1089–1095 (2010).
28. Lister, R. et al. Human DNA methylomes at base resolution show widespread epig-enomic differences. Nature 462, 315–322 (2009).
29. Laurent, L. et al. Dynamic changes in the human methylome during differentiation. Genome Res. 20, 320–331 (2010).
30. Kriaucionis, S. & Heintz, N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science 324, 929–930 (2009).
31. Berman, B.P., weisenberger, D.J. & Laird, P.w. Locking in on the human methylome. Nat. Biotechnol. 27, 341–342 (2009).
32. weisenberger, D.J. et al. DNA methylation analysis by digital bisulfite genomic sequenc-ing and digital MethyLight. Nucleic Acids Res. 36, 4689–4698 (2008).
more immunogenic, leading to activation of antigen-presenting cells, which could result in autoantibody production162.
Nucleosome positioning. No studies have yet made a connection between nucleosome positioning and autoimmune diseases. Notably, it has recently been reported that single-nucleotide polymorphisms in the 17q12-q21 region, which have been associated with a higher risk of asthma, type 1 diabetes, primary biliary cirrhosis and Crohn’s disease, lead to allele-spe-cific differences in nucleosome distribution165. Moreover, in rheumatoid arthritis, the incorporation of the histone variant macroH2A interferes with the binding of the transcription factor NF-κB and impedes SWI/SNF-dependent remodeling157.
Conclusions and perspectivesIn the past decade the fast-evolving field of epigenetics has taken center stage, as shown by the results of a simple PubMed search of the term ‘epi-genetic’: there were around 200 papers published in 1999, but more than 2,500 in 2009. Such startling growth in the number of publications attests to the intense research activity being undertaken in the field.
Great progress has been made in the description of epigenetic modi-fications in normal and diseased tissues. Thus far, efforts in epigenetic research have mainly focused on cancer, but as the field has grown, it has provided new insights into other types of diseases, particularly neu-rological and autoimmune diseases. Epigenetic alterations are likely to be found in other disorders; indeed, they have already been described in cardiovascular diseases166–168, metabolic diseases169, myopathies170 and children born from assisted reproductive treatments171.
In the past months, we have witnessed a flood of new discoveries: the description of comprehensive DNA methylomes of humans22 and viruses146, the putative identification of non-CpG methylation28, the definition of CpG island shores19, the involvement of aberrant DNA methylation in other diseases besides cancer6,135, the description of new histone modifications and histone variants and their roles45,126,161, the report of new epigenetic machinery such as the DNA demethylase Tet1 (refs. 172,173) and the histone kinase JAK2 (ref. 126), the description of new mutations in the epigenetic machinery99 and the flurry of ncRNA studies that highlight the importance of RNA-mediated regulation in epigentics174,175.
Many key questions remain unanswered: what are the functions of non-CpG methylation and 5-hydroxymethylcytosine in human cells? Are there new DNA or histone modifications yet to be discovered? What are the rules of the so-called histone code? What are the roles and function of ncRNAs and how many more ncRNAs are yet to be described? How is the placement of epigenetic marks and its specificity regulated? How are caus-ative epigenetic changes going to be distinguished from mere bystander alterations? Is it always clear whether a specific epigenetic modification is a cause or a consequence of a certain process? One of the most intrigu-ing questions is how do the various epigenetic players interact and what mechanisms convey sequence specificity to the enzymes involved? Further research is needed and efforts focused on such questions will be key in our progress toward a complete map of epigenetic regulation.
Advances in technological development are enabling epigenomic analysis on a large scale. The first whole-genome, high-resolution maps for epigenetic modifications are appearing, but we should not stop here. Detailed human DNA methylomes, histone modification and nucleosome positioning maps in healthy and diseased tissues are needed. In this regard several international projects and initiatives have been established: the NIH Roadmap Epigenomics Program, the ENCODE Project, the AHEAD Project and the Epigenomics NCBI browser, among others (see the commentaries by Bernstein and colleagues176 and Satterlee and colleagues177 in this issue). The detailed study of the epigenetic maps
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

nature biotechnology volume 28 number 10 oCTober 2010 1067
70. Mikkelsen, T.S. et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448, 553–560 (2007).
71. Bernstein, B.E. et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 125, 315–326 (2006).
72. Fuks, F. et al. The methyl-CpG-binding protein MeCP2 links DNA methylation to histone methylation. J. Biol. Chem. 278, 4035–4040 (2003).
73. Bhaumik, S.R., Smith, E. & Shilatifard, A. Covalent modifications of histones dur-ing development and disease pathogenesis. Nat. Struct. Mol. Biol. 14, 1008–1016 (2007).
74. Chang, B., Chen, Y., Zhao, Y. & Bruick, R.K. JMJD6 is a histone arginine demethylase. Science 318, 444–447 (2007).
75. Chi, P., Allis, C.D. & wang, G.G. Covalent histone modifications–miswritten, misinter-preted and mis-erased in human cancers. Nat. Rev. Cancer 10, 457–469 (2010).
76. wang, Z. et al. Genome-wide mapping of HATs and HDACs reveals distinct functions in active and inactive genes. Cell 138, 1019–1031 (2009).
77. Schones, D.E. et al. Dynamic regulation of nucleosome positioning in the human genome. Cell 132, 887–898 (2008).
78. Cairns, B.R. The logic of chromatin architecture and remodelling at promoters. Nature 461, 193–198 (2009).
79. Chodavarapu, R.K. et al. Relationship between nucleosome positioning and DNA methylation. Nature 466, 388–392 (2010).
80. Getun, I.v., wu, Z.K., Khalil, A.M. & Bois, P.R. Nucleosome occupancy landscape and dynamics at mouse recombination hotspots. EMBO Rep. 11, 555–560 (2010).
81. Zilberman, D., Coleman-Derr, D., Ballinger, T. & Henikoff, S. Histone H2A.Z and DNA methylation are mutually antagonistic chromatin marks. Nature 456, 125–129 (2008).
82. Harikrishnan, K.N. et al. Brahma links the SwI/SNF chromatin-remodeling complex with MeCP2-dependent transcriptional silencing. Nat. Genet. 37, 254–264 (2005).
83. wysocka, J. et al. A PHD finger of NURF couples histone H3 lysine 4 trimethylation with chromatin remodelling. Nature 442, 86–90 (2006).
84. Lal, A. et al. miR-24-mediated downregulation of H2AX suppresses DNA repair in terminally differentiated blood cells. Nat. Struct. Mol. Biol. 16, 492–498 (2009).
85. Yoo, A.S., Staahl, B.T., Chen, L. & Crabtree, G.R. MicroRNA-mediated switching of chromatin-remodelling complexes in neural development. Nature 460, 642–646 (2009).
86. Ho, L. & Crabtree, G.R. Chromatin remodelling during development. Nature 463, 474–484 (2010).
87. Reisman, D., Glaros, S. & Thompson, E.A. The SwI/SNF complex and cancer. Oncogene 28, 1653–1668 (2009).
88. Clapier, C.R. & Cairns, B.R. The biology of chromatin remodeling complexes. Annu. Rev. Biochem. 78, 273–304 (2009).
89. Sharma, S., Kelly, T.K. & Jones, P.A. Epigenetics in cancer. Carcinogenesis 31, 27–36 (2009).
90. Goelz, S.E., vogelstein, B., Hamilton, S.R. & Feinberg, A.P. Hypomethylation of DNA from benign and malignant human colon neoplasms. Science 228, 187–190 (1985).
91. Gaudet, F. et al. Induction of tumors in mice by genomic hypomethylation. Science 300, 489–492 (2003).
92. wilson, A.S., Power, B.E. & Molloy, P.L. DNA hypomethylation and human diseases. Biochim. Biophys. Acta 1775, 138–162 (2007).
93. Futscher, B.w. et al. Aberrant methylation of the maspin promoter is an early event in human breast cancer. Neoplasia 6, 380–389 (2004).
94. Futscher, B.w. et al. Role for DNA methylation in the control of cell type specific maspin expression. Nat. Genet. 31, 175–179 (2002).
95. Bettstetter, M. et al. Elevated nuclear maspin expression is associated with microsatel-lite instability and high tumour grade in colorectal cancer. J. Pathol. 205, 606–614 (2005).
96. Ito, Y. et al. Somatically acquired hypomethylation of IGF2 in breast and colorectal cancer. Hum. Mol. Genet. 17, 2633–2643 (2008).
97. Li, M. et al. Sensitive digital quantification of DNA methylation in clinical samples. Nat. Biotechnol. 27, 858–863 (2009).
98. Kelly, T.K., De Carvalho, D.D. & Peter A Jones, P.A. Epigenetic modifications as thera-peutic targets. Nat. Biotechnol. 28, 1069–1078 (2010).
99. Melo, S.A. et al. A TARBP2 mutation in human cancer impairs microRNA processing and DICER1 function. Nat. Genet. 41, 365–370 (2009).
100. Saito, Y. et al. Specific activation of microRNA-127 with downregulation of the proto-oncogene BCL6 by chromatin-modifying drugs in human cancer cells. Cancer Cell 9, 435–443 (2006).
101. Lujambio, A. et al. Genetic unmasking of an epigenetically silenced microRNA in human cancer cells. Cancer Res. 67, 1424–1429 (2007).
102. Lujambio, A. et al. A microRNA DNA methylation signature for human cancer metas-tasis. Proc. Natl. Acad. Sci. USA 105, 13556–13561 (2008).
103. Di Croce, L. et al. Methyltransferase recruitment and DNA hypermethylation of target promoters by an oncogenic transcription factor. Science 295, 1079–1082 (2002).
104. Frigola, J. et al. Epigenetic remodeling in colorectal cancer results in coordinate gene suppression across an entire chromosome band. Nat. Genet. 38, 540–549 (2006).
105. Miremadi, A., Oestergaard, M.Z., Pharoah, P.D. & Caldas, C. Cancer genetics of epi-genetic genes. Hum. Mol. Genet. 16 Spec No 1, R28–R49 (2007).
106. Garzon, R. et al. MicroRNA-29b induces global DNA hypomethylation and tumor sup-pressor gene reexpression in acute myeloid leukemia by targeting directly DNMT3A and 3B and indirectly DNMT1. Blood 113, 6411–6418 (2009).
107. Fraga, M.F. et al. Loss of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark of human cancer. Nat. Genet. 37, 391–400 (2005).
33. Down, T.A. et al. A Bayesian deconvolution strategy for immunoprecipitation-based DNA methylome analysis. Nat. Biotechnol. 26, 779–785 (2008).
34. Laird, P.w. Principles and challenges of genome-wide DNA methylation analysis. Nat. Rev. Genet. 11, 191–203 (2010).
35. Bourc’his, D., Xu, G.L., Lin, C.S., Bollman, B. & Bestor, T.H. Dnmt3L and the establish-ment of maternal genomic imprints. Science 294, 2536–2539 (2001).
36. Chen, Z.X., Mann, J.R., Hsieh, C.L., Riggs, A.D. & Chedin, F. Physical and functional interactions between the human DNMT3L protein and members of the de novo meth-yltransferase family. J. Cell. Biochem. 95, 902–917 (2005).
37. Holz-Schietinger, C. & Reich, N.O. The inherent processivity of the human de novo DNA methyltransferase 3A (DNMT3A) is enhanced by DNMT3L. J. Biol. Chem. 285, 29091–29100 (2010).
38. Chuang, L.S. et al. Human DNA-(cytosine-5) methyltransferase-PCNA complex as a target for p21wAF1. Science 277, 1996–2000 (1997).
39. Bostick, M. et al. UHRF1 plays a role in maintaining DNA methylation in mammalian cells. Science 317, 1760–1764 (2007).
40. Jones, P.A. & Liang, G. Rethinking how DNA methylation patterns are maintained. Nat. Rev. Genet. 10, 805–811 (2009).
41. Jeong, S. et al. Selective anchoring of DNA methyltransferases 3A and 3B to nucleosomes containing methylated DNA. Mol. Cell. Biol. 29, 5366–5376 (2009).
42. Goll, M.G. et al. Methylation of tRNAAsp by the DNA methyltransferase homolog Dnmt2. Science 311, 395–398 (2006).
43. Ooi, S.K. et al. DNMT3L connects unmethylated lysine 4 of histone H3 to de novo methylation of DNA. Nature 448, 714–717 (2007).
44. Tachibana, M., Matsumura, Y., Fukuda, M., Kimura, H. & Shinkai, Y. G9a/GLP com-plexes independently mediate H3K9 and DNA methylation to silence transcription. EMBO J. 27, 2681–2690 (2008).
45. Zhao, Q. et al. PRMT5-mediated methylation of histone H4R3 recruits DNMT3A, coupling histone and DNA methylation in gene silencing. Nat. Struct. Mol. Biol. 16, 304–311 (2009).
46. Esteve, P.O. et al. Regulation of DNMT1 stability through SET7-mediated lysine methylation in mammalian cells. Proc. Natl. Acad. Sci. USA 106, 5076–5081 (2009).
47. wang, J. et al. The lysine demethylase LSD1 (KDM1) is required for maintenance of global DNA methylation. Nat. Genet. 41, 125–129 (2009).
48. Mosher, R.A. & Melnyk, C.w. siRNAs and DNA methylation: seedy epigenetics. Trends Plant Sci. 15, 204–210 (2010).
49. Matzke, M.A. & Birchler, J.A. RNAi-mediated pathways in the nucleus. Nat. Rev. Genet. 6, 24–35 (2005).
50. vrbsky, J. et al. siRNA-mediated methylation of Arabidopsis telomeres. PLoS Genet. 6, e1000986 (2010).
51. Zaratiegui, M., Irvine, D.v. & Martienssen, R.A. Noncoding RNAs and gene silencing. Cell 128, 763–776 (2007).
52. Kouzarides, T. Chromatin modifications and their function. Cell 128, 693–705 (2007).
53. Daujat, S., Zeissler, U., waldmann, T., Happel, N. & Schneider, R. HP1 binds specifi-cally to Lys26-methylated histone H1.4, whereas simultaneous Ser27 phosphorylation blocks HP1 binding. J. Biol. Chem. 280, 38090–38095 (2005).
54. Rando, O.J. & Chang, H.Y. Genome-wide views of chromatin structure. Annu. Rev. Biochem. 78, 245–271 (2009).
55. Huertas, D., Sendra, R. & Munoz, P. Chromatin dynamics coupled to DNA repair. Epigenetics 4, 31–42 (2009).
56. Luco, R.F. et al. Regulation of alternative splicing by histone modifications. Science 327, 996–1000 (2010).
57. Li, B., Carey, M. & workman, J.L. The role of chromatin during transcription. Cell 128, 707–719 (2007).
58. Karlic, R., Chung, H.R., Lasserre, J., vlahovicek, K. & vingron, M. Histone modification levels are predictive for gene expression. Proc. Natl. Acad. Sci. USA 107, 2926–2931 (2010).
59. Moazed, D. Small RNAs in transcriptional gene silencing and genome defence. Nature 457, 413–420 (2009).
60. Grewal, S.I. & Jia, S. Heterochromatin revisited. Nat. Rev. Genet. 8, 35–46 (2007).61. Khalil, A.M. et al. Many human large intergenic noncoding RNAs associate with chro-
matin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 106, 11667–11672 (2009).
62. Chow, J. & Heard, E. X inactivation and the complexities of silencing a sex chromosome. Curr. Opin. Cell Biol. 21, 359–366 (2009).
63. Agrelo, R. & wutz, A. X inactivation and disease. Semin. Cell Dev. Biol. 21, 194–200 (2010).
64. Rinn, J.L. et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 129, 1311–1323 (2007).
65. Santos-Rosa, H. et al. Histone H3 tail clipping regulates gene expression. Nat. Struct. Mol. Biol. 16, 17–22 (2009).
66. wang, Z. et al. Combinatorial patterns of histone acetylations and methylations in the human genome. Nat. Genet. 40, 897–903 (2008).
67. Duan, Q., Chen, H., Costa, M. & Dai, w. Phosphorylation of H3S10 blocks the access of H3K9 by specific antibodies and histone methyltransferase. Implication in regulating chromatin dynamics and epigenetic inheritance during mitosis. J. Biol. Chem. 283, 33585–33590 (2008).
68. Nakanishi, S. et al. Histone H2BK123 monoubiquitination is the critical determi-nant for H3K4 and H3K79 trimethylation by COMPASS and Dot1. J. Cell Biol. 186, 371–377 (2009).
69. Ernst, J. & Kellis, M. Discovery and characterization of chromatin states for systematic annotation of the human genome. Nat. Biotechnol. 28, 817–825 (2010).
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.

1068 volume 28 number 10 oCTober 2010 nature biotechnology
144. Gheldof, N., Tabuchi, T.M. & Dekker, J. The active FMR1 promoter is associated with a large domain of altered chromatin conformation with embedded local histone modifications. Proc. Natl. Acad. Sci. USA 103, 12463–12468 (2006).
145. Pieper, H.C. et al. Different methylation of the TNF-alpha promoter in cortex and substantia nigra: Implications for selective neuronal vulnerability. Neurobiol. Dis. 32, 521–527 (2008).
146. Murgatroyd, C. et al. Dynamic DNA methylation programs persistent adverse effects of early-life stress. Nat. Neurosci. 12, 1559–1566 (2009).
147. Robertson, K.D. DNA methylation and human disease. Nat. Rev. Genet. 6, 597–610 (2005).
148. Herman, D. et al. Histone deacetylase inhibitors reverse gene silencing in Friedreich’s ataxia. Nat. Chem. Biol. 2, 551–558 (2006).
149. Al-Mahdawi, S. et al. The Friedreich ataxia GAA repeat expansion mutation induces comparable epigenetic changes in human and transgenic mouse brain and heart tis-sues. Hum. Mol. Genet. 17, 735–746 (2008).
150. Kleine-Kohlbrecher, D. et al. A functional link between the histone demethylase PHF8 and the transcription factor ZNF711 in X-linked mental retardation. Mol. Cell 38, 165–178 (2010).
151. Kumari, D. & Usdin, K. Chromatin remodeling in the noncoding repeat expansion diseases. J. Biol. Chem. 284, 7413–7417 (2009).
152. Jin, B. et al. DNA methyltransferase 3B (DNMT3B) mutations in ICF syndrome lead to altered epigenetic modifications and aberrant expression of genes regulating develop-ment, neurogenesis and immune function. Hum. Mol. Genet. 17, 690–709 (2008).
153. Pan, w. et al. MicroRNA-21 and microRNA-148a contribute to DNA hypomethylation in lupus CD4+ T cells by directly and indirectly targeting DNA methyltransferase 1. J. Immunol. 184, 6773–6781 (2010).
154. Javierre, B.M., Esteller, M. & Ballestar, E. Epigenetic connections between autoimmune disorders and haematological malignancies. Trends Immunol. 29, 616–623 (2008).
155. Karouzakis, E., Gay, R.E., Gay, S. & Neidhart, M. Epigenetic control in rheumatoid arthritis synovial fibroblasts. Nat. Rev. Rheumatol. 5, 266–272 (2009).
156. Mishra, N., Brown, D.R., Olorenshaw, I.M. & Kammer, G.M. Trichostatin A reverses skewed expression of CD154, interleukin-10, and interferon-gamma gene and protein expression in lupus T cells. Proc. Natl. Acad. Sci. USA 98, 2628–2633 (2001).
157. vanden Berghe, w. et al. Keeping up NF-kappaB appearances: epigenetic control of immunity or inflammation-triggered epigenetics. Biochem. Pharmacol. 72, 1114–1131 (2006).
158. Huber, L.C., Stanczyk, J., Jungel, A. & Gay, S. Epigenetics in inflammatory rheumatic diseases. Arthritis Rheum. 56, 3523–3531 (2007).
159. Miao, F. et al. Lymphocytes from patients with type 1 diabetes display a distinct pro-file of chromatin histone H3 lysine 9 dimethylation: an epigenetic study in diabetes. Diabetes 57, 3189–3198 (2008).
160. Ajiro, K. Histone H2B phosphorylation in mammalian apoptotic cells. An association with DNA fragmentation. J. Biol. Chem. 275, 439–443 (2000).
161. Hurd, P.J. et al. Phosphorylation of histone H3 Thr-45 is linked to apoptosis. J. Biol. Chem. 284, 16575–16583 (2009).
162. van Bavel, C.C. et al. Apoptosis-induced histone H3 methylation is targeted by autoantibodies in systemic lupus erythematosus. Ann. Rheum. Dis. published online doi:10.1136/ard.2010.129320 (10 August 2010).
163. Dieker, J.w. et al. Apoptosis-induced acetylation of histones is pathogenic in systemic lupus erythematosus. Arthritis Rheum. 56, 1921–1933 (2007).
164. van Bavel, J.J. & Cunningham, w.A. Self-categorization with a novel mixed-race group moderates automatic social and racial biases. Pers. Soc. Psychol. Bull. 35, 321–335 (2009).
165. verlaan, D.J. et al. Allele-specific chromatin remodeling in the ZPBP2/GSDMB/ORMDL3 locus associated with the risk of asthma and autoimmune disease. Am. J. Hum. Genet. 85, 377–393 (2009).
166. Turunen, M.P., Aavik, E. & Yla-Herttuala, S. Epigenetics and atherosclerosis. Biochim. Biophys. Acta 1790, 886–891 (2009).
167. Movassagh, M. et al. Differential DNA methylation correlates with differential expres-sion of angiogenic factors in human heart failure. PLoS ONE 5, e8564 (2010).
168. Hang, C.T. et al. Chromatin regulation by Brg1 underlies heart muscle development and disease. Nature 466, 62–67 (2010).
169. Symonds, M.E., Sebert, S.P., Hyatt, M.A. & Budge, H. Nutritional programming of the metabolic syndrome. Nat. Rev. Endocrinol. 5, 604–610 (2009).
170. Zeng, w. et al. Specific loss of histone H3 lysine 9 trimethylation and HP1gamma/cohesin binding at D4Z4 repeats is associated with facioscapulohumeral dystrophy (FSHD). PLoS Genet. 5, e1000559 (2009).
171. wilkins-Haug, L. Epigenetics and assisted reproduction. Curr. Opin. Obstet. Gynecol. 21, 201–206 (2009).
172. Tahiliani, M. et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mam-malian DNA by MLL partner TET1. Science 324, 930–935 (2009).
173. Ito, S. et al. Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature 466, 1129–1133 (2010).
174. Ghildiyal, M. & Zamore, P.D. Small silencing RNAs: an expanding universe. Nat. Rev. Genet. 10, 94–108 (2009).
175. Mattick, J.S. The genetic signatures of noncoding RNAs. PLoS Genet. 5, e1000459 (2009).
176. Bernstein, B. The NIH Roadmap Epigenome Mapping Consortium. Nat. Biotechnol. 28, 1045–1048 (2010).
177. Satterlee, J. Tackling the epigenome: challenges and opportunities for collaborative efforts. Nat. Biotechnol. 28, 1039–1044 (2010).
108. Zhu, P. et al. Induction of HDAC2 expression upon loss of APC in colorectal tumori-genesis. Cancer Cell 5, 455–463 (2004).
109. Ropero, S. et al. A truncating mutation of HDAC2 in human cancers confers resistance to histone deacetylase inhibition. Nat. Genet. 38, 566–569 (2006).
110. vaquero, A., Sternglanz, R. & Reinberg, D. NAD+-dependent deacetylation of H4 lysine 16 by class III HDACs. Oncogene 26, 5505–5520 (2007).
111. Noonan, E.J. et al. miR-449a targets HDAC-1 and induces growth arrest in prostate cancer. Oncogene 28, 1714–1724 (2009).
112. Moore, S.D. et al. Uterine leiomyomata with t(10;17) disrupt the histone acetyltrans-ferase MORF. Cancer Res. 64, 5570–5577 (2004).
113. Bryan, E.J. et al. Mutation analysis of EP300 in colon, breast and ovarian carcinomas. Int. J. Cancer 102, 137–141 (2002).
114. Hamamoto, R. et al. SMYD3 encodes a histone methyltransferase involved in the proliferation of cancer cells. Nat. Cell Biol. 6, 731–740 (2004).
115. Kondo, Y. et al. Alterations of DNA methylation and histone modifications contribute to gene silencing in hepatocellular carcinomas. Hepatol. Res. 37, 974–983 (2007).
116. vire, E. et al. The Polycomb group protein EZH2 directly controls DNA methylation. Nature 439, 871–874 (2006).
117. Dalgliesh, G.L. et al. Systematic sequencing of renal carcinoma reveals inactivation of histone modifying genes. Nature 463, 360–363 (2010).
118. Gupta, R.A. et al. Long non-coding RNA HOTAIR reprograms chromatin state to pro-mote cancer metastasis. Nature 464, 1071–1076 (2010).
119. varambally, S. et al. Genomic loss of microRNA-101 leads to overexpression of histone methyltransferase EZH2 in cancer. Science 322, 1695–1699 (2008).
120. Berdasco, M. et al. Epigenetic inactivation of the Sotos overgrowth syndrome gene histone methyltransferase NSD1 in human neuroblastoma and glioma. Proc. Natl. Acad. Sci. USA 106, 21830–21835 (2009).
121. Jones, B. et al. The histone H3K79 methyltransferase Dot1L is essential for mammalian development and heterochromatin structure. PLoS Genet. 4, e1000190 (2008).
122. Jacinto, F.v., Ballestar, E. & Esteller, M. Impaired recruitment of the histone meth-yltransferase DOT1L contributes to the incomplete reactivation of tumor suppressor genes upon DNA demethylation. Oncogene 28, 4212–4224 (2009).
123. Krivtsov, A.v. et al. H3K79 methylation profiles define murine and human MLL-AF4 leukemias. Cancer Cell 14, 355–368 (2008).
124. wang, P. et al. Global analysis of H3K4 methylation defines MLL family member targets and points to a role for MLL1-mediated H3K4 methylation in the regulation of transcriptional initiation by RNA polymerase II. Mol. Cell. Biol. 29, 6074–6085 (2009).
125. Shi, Y. Histone lysine demethylases: emerging roles in development, physiology and disease. Nat. Rev. Genet. 8, 829–833 (2007).
126. Dawson, M.A. et al. JAK2 phosphorylates histone H3Y41 and excludes HP1alpha from chromatin. Nature 461, 819–822 (2009).
127. Medina, P.P. & Sanchez-Cespedes, M. Involvement of the chromatin-remodeling factor BRG1/SMARCA4 in human cancer. Epigenetics 3, 64–68 (2008).
128. Naidu, S.R., Love, I.M., Imbalzano, A.N., Grossman, S.R. & Androphy, E.J. The SwI/SNF chromatin remodeling subunit BRG1 is a critical regulator of p53 necessary for proliferation of malignant cells. Oncogene 28, 2492–2501 (2009).
129. Roberts, C.w. & Orkin, S.H. The SwI/SNF complex–chromatin and cancer. Nat. Rev. Cancer 4, 133–142 (2004).
130. Lin, J.C. et al. Role of nucleosomal occupancy in the epigenetic silencing of the MLH1 CpG island. Cancer Cell 12, 432–444 (2007).
131. Mulero-Navarro, S. & Esteller, M. Chromatin remodeling factor CHD5 is silenced by promoter CpG island hypermethylation in human cancer. Epigenetics 3, 210–215 (2008).
132. Sporn, J.C. et al. Histone macroH2A isoforms predict the risk of lung cancer recurrence. Oncogene 28, 3423–3428 (2009).
133. Gibbs, J.R. et al. Abundant quantitative trait Loci exist for DNA methylation and gene expression in human brain. PLoS Genet. 6, e1000952 (2010).
134. wynder, C., Hakimi, M.A., Epstein, J.A., Shilatifard, A. & Shiekhattar, R. Recruitment of MLL by HMG-domain protein iBRAF promotes neural differentiation. Nat. Cell Biol. 7, 1113–1117 (2005).
135. Urdinguio, R.G., Sanchez-Mut, J.v. & Esteller, M. Epigenetic mechanisms in neuro-logical diseases: genes, syndromes, and therapies. Lancet Neurol. 8, 1056–1072 (2009).
136. Hite, K.C., Adams, v.H. & Hansen, J.C. Recent advances in MeCP2 structure and function. Biochem. Cell Biol. 87, 219–227 (2009).
137. Chahrour, M. et al. MeCP2, a key contributor to neurological disease, activates and represses transcription. Science 320, 1224–1229 (2008).
138. Urdinguio, R.G. et al. Mecp2-null mice provide new neuronal targets for Rett syndrome. PLoS ONE 3, e3669 (2008).
139. Nomura, T. et al. MeCP2-dependent repression of an imprinted miR-184 released by depolarization. Hum. Mol. Genet. 17, 1192–1199 (2008).
140. Urdinguio, R.G. et al. Disrupted microRNA expression caused by Mecp2 loss in a mouse model of Rett syndrome. Epigenetics 5, 656–663 (2010).
141. Alarcon, J.M. et al. Chromatin acetylation, memory, and LTP are impaired in CBP+/− mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its ameliora-tion. Neuron 42, 947–959 (2004).
142. Clayton, A.L., Rose, S., Barratt, M.J. & Mahadevan, L.C. Phosphoacetylation of histone H3 on c-fos- and c-jun-associated nucleosomes upon gene activation. EMBO J. 19, 3714–3726 (2000).
143. De Sario, A. Clinical and molecular overview of inherited disorders resulting from epigenomic dysregulation. Eur. J. Med. Genet. 52, 363–372 (2009).
RE v IE w©
201
0 N
atu
re A
mer
ica,
Inc.
All
rig
hts
res
erve
d.


















