
Enzyme Catalysis Bill Royer Office: LRB 921 Phone: x6-6912 I. Transition state theory II. Mechanisms...
22
Enzyme Catalysis Bill Royer Office: LRB 921 Phone: x6-6912 . Transition state theory I. Mechanisms of catalysis Acid-base catalysis - Ribonuclease A Metal ion catalysis - Hammerhead Catalytic RNA Covalent catalysis - Chymotrypsin Enzymes have spectacular abilities to accelerate chemical reactions – often by factors of 10 6 -10 14 over non-catalyzed reactions. In this lecture, we will briefly discuss some of the strategies used by enzymes to achieve such remarkable rate increases.
-
Upload
justin-harrison -
Category
Documents
-
view
240 -
download
0
Transcript of Enzyme Catalysis Bill Royer Office: LRB 921 Phone: x6-6912 I. Transition state theory II. Mechanisms...

Enzyme CatalysisBill Royer
Office: LRB 921Phone: x6-6912
I. Transition state theory II. Mechanisms of catalysis
Acid-base catalysis - Ribonuclease A
Metal ion catalysis - Hammerhead Catalytic RNA
Covalent catalysis - Chymotrypsin
Enzymes have spectacular abilities to accelerate chemical reactions – often by factors of 106-1014 over non-catalyzed reactions. In this lecture, we will briefly discuss some of the strategies used by enzymes to achieve such remarkable rate increases.

I. Transition state theory
Consider the reaction A + B P + Q
where A + B react through transition state, X‡, to form products P + Q. K‡ is the equilibrium constant between A + B and X‡ and k' is the rate constant for conversion of X‡ to P + Q.
A + B P + Q‡K ‡ k'
A + B
P + Q
G reaction
G
G
Reaction coordinate
The minimum energy pathway of the reactionis shown in the reaction coordinate, ortransition state diagram, at left. Chemicalconversion of A + B to P + Q proceedsthrough a transition state ‡ which is theleast stable (least probable, highest freeenergy) species along the pathway. Molecules that achieve the activation energy,G‡, can go on to react while molecules thatfail to achieve the transition state fall back tothe ground state.
‡
‡
The transition state, X‡, is metastable. (Unlike a reaction intermediate, the transition state has only a transient existence, like a pebble balanced on a pin. By definition, a transition state cannot be isolated.) The transition state can be thought of as sharing some features of the reactants and some features of the products. That is, some bonds in the substrate are on their way to being broken and some bonds in the product are partially formed.

The transition state, X‡, is in rapid equilibrium with reactants with equilibrium constant K‡. K ‡ ‡
[A] [ B]
-RT lnK ‡ = G‡
G‡, the activation energy, is the difference in Gibbs free energy between the transition state, X‡, and the reactants. Since K‡ is an equilibrium constant, the now familiar
equation applies:
where T is the absolute temperature in degrees Kelvin (°C + 273) and R is the gas constant (1.98 cal / mol / degree). In other words, the frequency with which reactants achieve the transition state is inversely proportional to the activation energy barrier between the two.
The observed rate of the reaction, kobs, will be a function of the concentration of the reactants, the rate of conversion of X‡ to P + Q, k', and will decrease exponentially with an increase in G‡.
k = k' e obs-G‡ / RT
[A][B]

Thus, the smaller the difference in free energy of the reactants and the transition state, the faster the reaction proceeds. Enzymatic rate accelerations are achieved by lowering the activation barrier between reactants and the transition state, thereby increasing the fraction of reactants able to achieve the transition state. Enzymes reduce the activation barrier by destabilizing the ground state of enzyme-bound substrates and products, by stabilizing the transition state, and/or by introducing a new reaction pathway with a different transition state that has a lower free energy.
A + B
P + Q
G
Reaction coordinate
‡
Gcat‡
A+B P+Q
Uncatalyzed
Catalyzed
Enzymes accelerate reactions by lowering the energy barrier between reactants and products. G = G - G Although less energy is required to form the transition state in the catalyzed reaction, the ground states of the free substrates and products remain the same. The kinetic barrier is lowered by the same extent for the forward and reverse reactions. Consequently, a catalyst accelerates the reaction without affecting its equilibrium .
‡ ‡ ‡uncatalyzed catalyzed
If a catalyst lowers the activation barrier by G‡, the rate of the reaction is enhanced
by the factor e G‡/RT. Consequently, a ten-fold rate enhancement requires that G‡ = 1.36 kcal/mole, less than the energy of a single hydrogen bond. (G‡ = RTln10 = 1.98 x 10-3 kcal/mol*K x 298K*ln(10) = 1.36 kcal/mol)

(Nelson & Cox, Lehninger Principles of Biochemistry, 3rd ed., 2000)
Imaginary enzyme ("stickase") designed to catalyze "cleavage" (breaking) of a metal stick

A
P
G
Reaction coordinate
A‡ I ‡
I
k1A Pk2I
k1 k2<k1 k2>
If the formation of I, an intermediate, from A is slower than the formation of P from I (k < k ) the activation barrier for the first step must be higher than the activation barrier for the second step (thick line). If k is much slower than k , conversion of A to I is the rate-determining step for the reaction. That is, the overall reaction proceeds at a rate that can be no faster than k . Conversely, if formation of P from I is much slower than formation of I from A (k < k ), the activation barrier for the second step is higher (thin line) and formation of P from I is rate-determining.
1 2
2 1
1 2
1
A k1 k2
I P
For a reaction that involves several steps, each step will have a corresponding transition state.
k1 < k2
k1 > k2

II. Mechanisms of catalysis
A. Acid-base catalysis
Specific acid or base catalysis - Reaction rate is directly proportional to [H+] or [OH-].
Example: Alkaline hydrolysis of RNA
General acid or base catalysis - Reaction rate is proportional to [Bronsted acid] or [Bronsted base]
Bronsted acid - species that can donate protons Bronsted base - species that can combine with a proton
[Imidazole buffer]
Ra
te
[Imidazole buffer]
Ra
te
Specific Base Catalysis General Base Catalysis
pH 7.3
pH 7.0
pH 7.3
pH 7.0
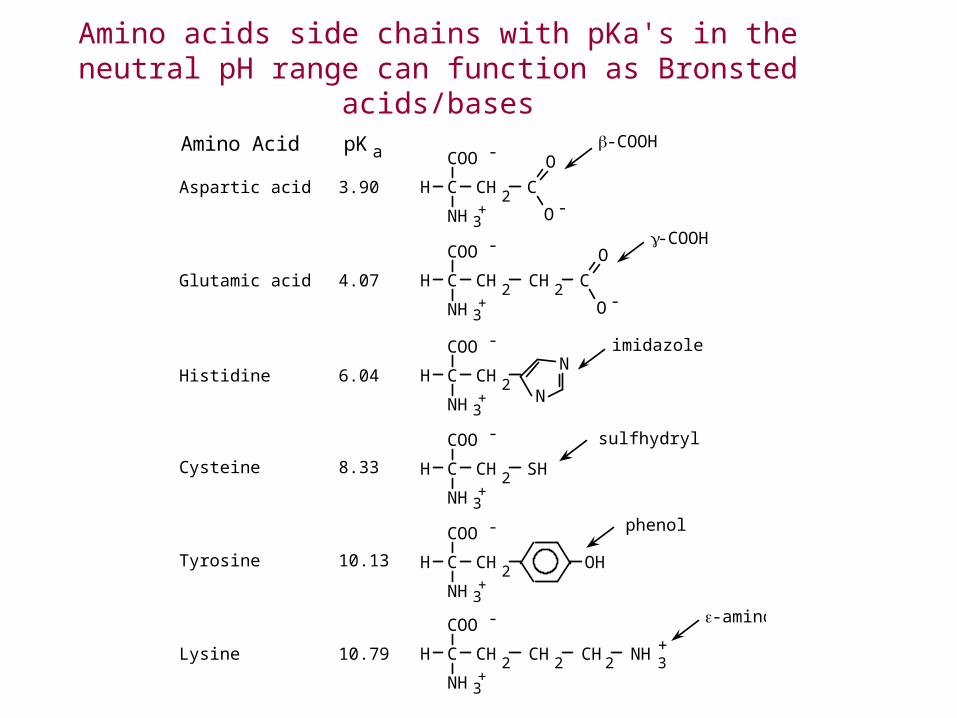
H C
NH
COO
CH C
O
O-2
-
+3
Aspartic acid
Amino Acid pK a
3.90
-COOH
H C
NH
COO
CH C
O
O-2
-
+3
Glutamic acid 4.07
-COOH
CH2
H C
NH
COO
CH2
-
+3
Histidine 6.04
imidazole
N
N
H C
NH
COO
CH SH2
-
+3
Cysteine 8.33
sulfhydryl
H C
NH
COO
CH2
-
+3
Tyrosine 10.13
phenol
OH
H C
NH
COO
CH CH2
-
+3
Lysine 10.79
-amino
CH2 2 NH 3+
Amino acids side chains with pKa's in the neutral pH range can function as Bronsted acids/bases

Biologically important nucleophilic groups:
Hydroxyl group R-OH
:: R-O:
:: + H+
-
Sulfhydryl group R-SH
::
Adapted from Voet & Voet, Biochemistry
R-S:
::
- + H+
R-NH3+ R-NH2
:
+ H+Amino group
HN NH+
R
HN N:
R
+ H+Imidazole group
Nucleophilicform
:
R-NH2 + C=O
Biologically important electrophiles:
C=OH+ Mn+
Protons Metal Ions Carbonyl carbon
O
OHOH
Base
O
O O
O5'...
P
Pyrimidine
O
HO
Base
3'...
O O-2',3'-Cyclic phosphate
O
O OH
P OOO
Pyrimidine
O5'...
HH O2
O
O OH
P OOO
Pyrimidine
O5'...
3' phosphate
O OH
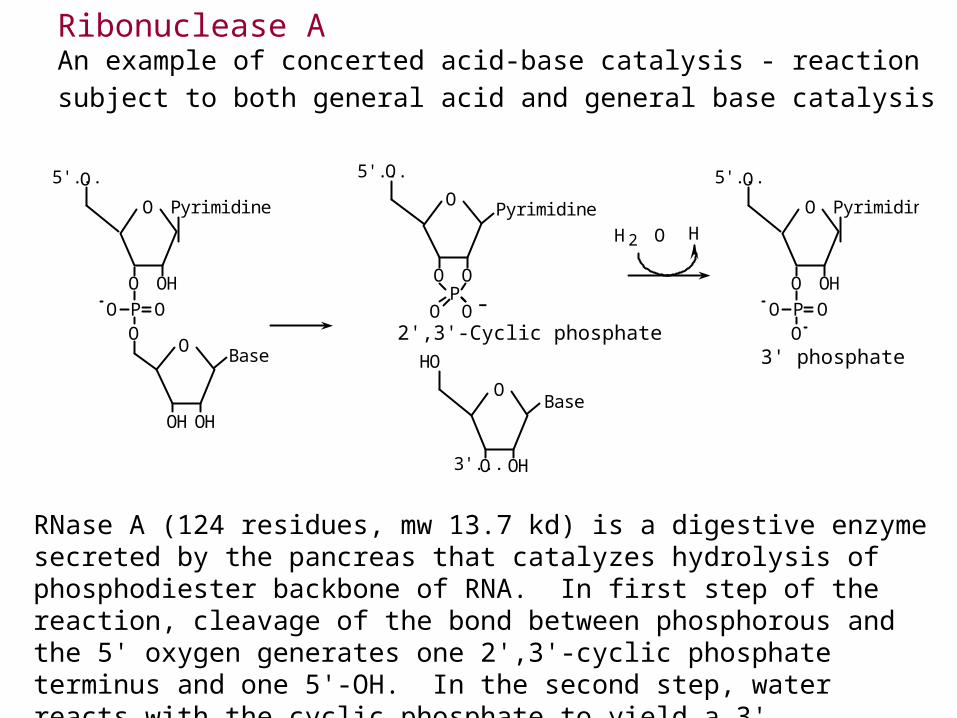
Ribonuclease AAn example of concerted acid-base catalysis - reaction subject to both general acid and general base catalysis
RNase A (124 residues, mw 13.7 kd) is a digestive enzyme secreted by the pancreas that catalyzes hydrolysis of phosphodiester backbone of RNA. In first step of the reaction, cleavage of the bond between phosphorous and the 5' oxygen generates one 2',3'-cyclic phosphate terminus and one 5'-OH. In the second step, water reacts with the cyclic phosphate to yield a 3' phosphate. The 2',3' cyclic phosphate can be isolated because it forms more rapidly than it hydrolyzes.

O
O
O
O5'...A
P
O
O
O
O OH
Base
3'...
O
O
O O
O5'...
P OOO
Pyrimidine
O
O OH
Base
3'...
H Base abstracts proton from 2' OH
NHHN +
His 119
Acid protonates 5' leaving group
NH:N +
His 12
Charge stabilization
Trigonal bipyramidal transition state
O
O O
O5'...
P
Pyrimidine
O
O OH
HO
Base
3'...
O O-
Intermediate
H N3
Lys 41
+
Transesterification
Nucleophilic attack of 2' O on phosphate
First Step: 2’3’ cyclic nucleotide produced. His 12 is general base, His 119 is general acid
Hydrolysis of 2',3' cyclic phosphate intermediate
O
O O
O5'...
P
Pyrimidine
O O- HO
HNH:N +
His 119
Base abstracts proton from H O2
Nucleophilic attack of H O on phosphate2
NHHN +
His 12
Acid protonates 2' OH leaving group
O
O OH
P OOO
Pyrimidine
O5'...
3' phosphate
Second Step:
His 12 is general acid, His 119 is general base

Geometry of the pentacovalent transition state. The central phosphorus atom is transiently bonded to 5 oxygen atoms. Three oxygens are coplanar with the phosphorus. The oxygen atoms of the leaving group is at one apex, and the oxygen atom of the attacking group is at the other apex of the trigonal bipyramid (in-line attack).
Proposed mechanism of RNase A catalysis. The unionized form of His 12 accepts a proton from the 2' OH which enhances its nucleophilicity. The protonated form of His 119 begins to donate its proton to the 5' O, and the 2'O begins to form a bond with P to form a pentacoordinate transition state. The negative charge that develops is stabilized electrostatically by the nearby positively charged side chain of lysine 41. The bond between P and the 5'-O breaks when the proton from histidine 119 is completely transferred. At the same time, a bond between P and the 2'-O becomes fully formed, producing the 2',3'-cyclic intermediate. Hydrolysis of the cyclic intermediate is a reversal of the first stage with H2O replacing the 5'-O component that was removed. Histidine 12 is now the proton donor and histidine 119 is the proton acceptor.
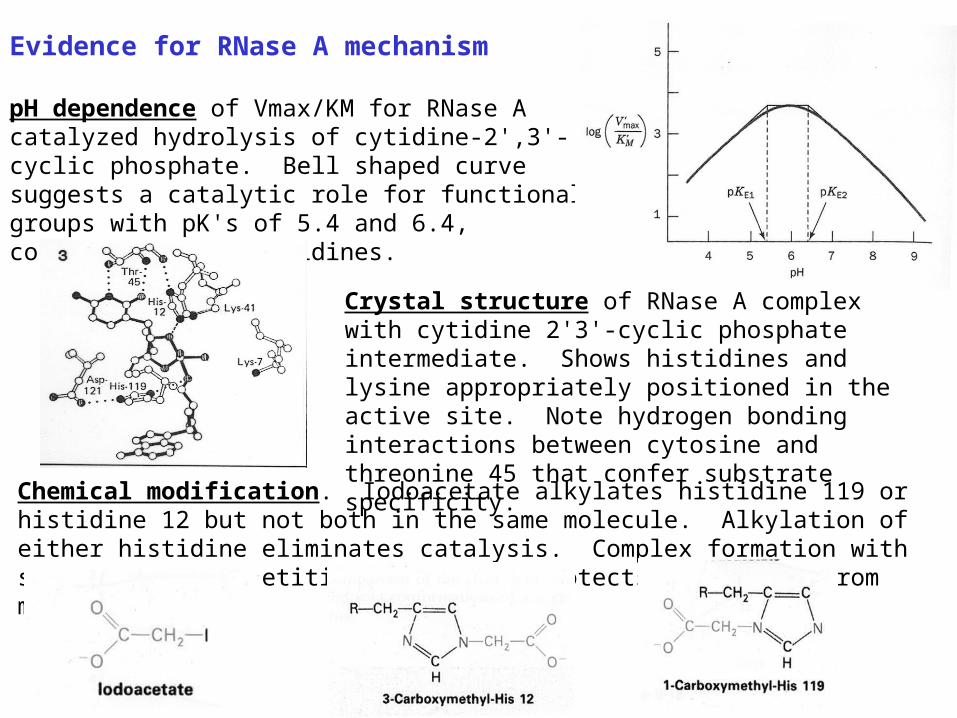
Evidence for RNase A mechanism pH dependence of Vmax/KM for RNase A catalyzed hydrolysis of cytidine-2',3'-cyclic phosphate. Bell shaped curve suggests a catalytic role for functional groups with pK's of 5.4 and 6.4, consistent with histidines.
Crystal structure of RNase A complex with cytidine 2'3'-cyclic phosphate intermediate. Shows histidines and lysine appropriately positioned in the active site. Note hydrogen bonding interactions between cytosine and threonine 45 that confer substrate specificity.
Chemical modification. Iodoacetate alkylates histidine 119 or histidine 12 but not both in the same molecule. Alkylation of either histidine eliminates catalysis. Complex formation with substrate or competitive inhibitors protects histidines from modification.

H O22+Mg 2+Mg OH - + H+ pKa = 11.4
B. Metal ion catalysis1. Water ionization. A metal ion's charge makes its bound water molecules more acidic than free H2O and therefore a source of OH- ions even below neutral pH (Metal ions have been called "Super acids").
2. Charge shielding - metal ions can have charge > +1.3. Oxidation-Reduction
GGACA
UCCUG
GGCC
CCGGGA
A
AGU AGU
C
AUU G GG5'
5'
Ribozyme
CCUGUCAGGAU
Substrate
cleavage site
Hammerhead Catalytic RNA
The Hammerhead Catalytic RNAThe hammerhead ribozyme, like RNase A, catalyzes a transesterification reaction to cleave the phosphodiester backbone of substrate RNAs yielding products with 5' hydroxyl and 2'3'cyclic phosphate termini. Unlike the RNase A-catalyzed reaction, the hammerhead reaction does not proceed through hydrolysis of the 2',3' cyclic phosphate.
The hammerhead ribozyme obviously has no amino acid side chains to carry out proton transfer and charge-shielding functions. RNAs are, however, capable of binding metal ions with high specificity and affinity and the hammerhead ribozyme appears to make use of metal ions to carry out both charge shielding and proton transfer functions.

C. Covalent catalysis - Transient formation of a catalyst-substrate covalent bond-Provides an alternative reaction pathway, with two lower energy transition states
1. A nucleophile (electron-rich group with a strong tendency to donate electrons to an electron-deficient nucleus) on the enzyme displaces a leaving group on the substrate, forming a covalent bond.
2. The enzyme substrate bond decomposes to form product and free enzyme. -Covalent catalyst must be a good nucleophile and a good leaving group - highly mobile electrons (imidazole of His, thiol of Cys, carboxyl of Asp, hydroxyl of Ser).
Chymotrypsin, 25 kd serine protease, catalyzes hydrolysis of proteins in the small intestine. Chymotrypsin catalyzes hydrolysis of esters as well as peptide bonds which has been useful for analysis of the catalytic mechanism, although not physiologically relevant.

[p-N
itro
ph
en
yla
te], M
Time (min)
10
20
30
2 4 6 8 10 12
chymotrypsin32M
24M
16M
8M
The plot at left shows the concentration of p-nitrophenol produced as a function of time in reactions containing different concentrations of chymotrypsin and a large excess of p-nirophenylacetate. An initial rapid phase ("burst") is followed by a slower phase. The size of the initial burst is proportional to the enzyme concentration. "Burst" kinetics provide evidence for a stable, enzyme-linked intermediate.
CCH 3 NO
O
2+ Chymotrypsin
p-Nitrophenylacetate
fastO NO2
-
p-Nitrophenylate
+ CCH3
O
chymotrypsin
Acyl-enzyme intermediate
slowH O2
H+
CCH3
O
-O
Acetate
+ chymotrypsin
Model reaction in which hydrolysis of acyl-enzyme intermediate is slow
Formation of the acyl-enzyme intermediate occurs during the initial rapid phase and slower hydrolysis (deacylation) of the acyl-enzyme intermediate occurs during the second, slower phase.
o

First stage in peptide bond hydrolysis: acylation. Hydrolysis of the peptide bond starts with an attack by the oxygen atom of the Ser195 hydroxyl group on the carbonyl carbon atom of the susceptible bond. The carbon-oxygen bond of this carbonyl group becomes a single bond, and the oxygen atom acquires a net negative charge. The four atoms now bonded to the carbonyl carbon are arranged as a tetrahedron. Transfer of a proton from Ser195 to His57 is facilitated by Asp102 which (i) precisely orients the imidazole ring of His57 and (ii) partly neutralizes the positive charge that develops on His57 during the transition state. The proton held by the protonated form of His57 is then donated to the nitrogen atom of the peptide bond that is cleaved. At this stage, the amine component is hydrogen bonded to His57, and the acid component of the substrate is esterified to Ser195. The amine component diffuses away.
Oxyanionhole

Second stage in peptide hydrolysis: deacylation. The acyl-enzyme intermediate is hydrolyzed by water. Deacylation is essentially the reverse of acylation with water playing the role as the attacking nucleophile, similar to Ser195 in the first step. First, a proton is drawn away from water. The resulting OH- attacks the carbonyl carbon of the acyl group that is attached to Ser195. As in acylation, a transient tetrahedral intermediate is formed. His57 then donates a proton to the oxygen atom of Ser195, which then releases the acid component of the substrate, completing the reaction.
Oxyanionhole
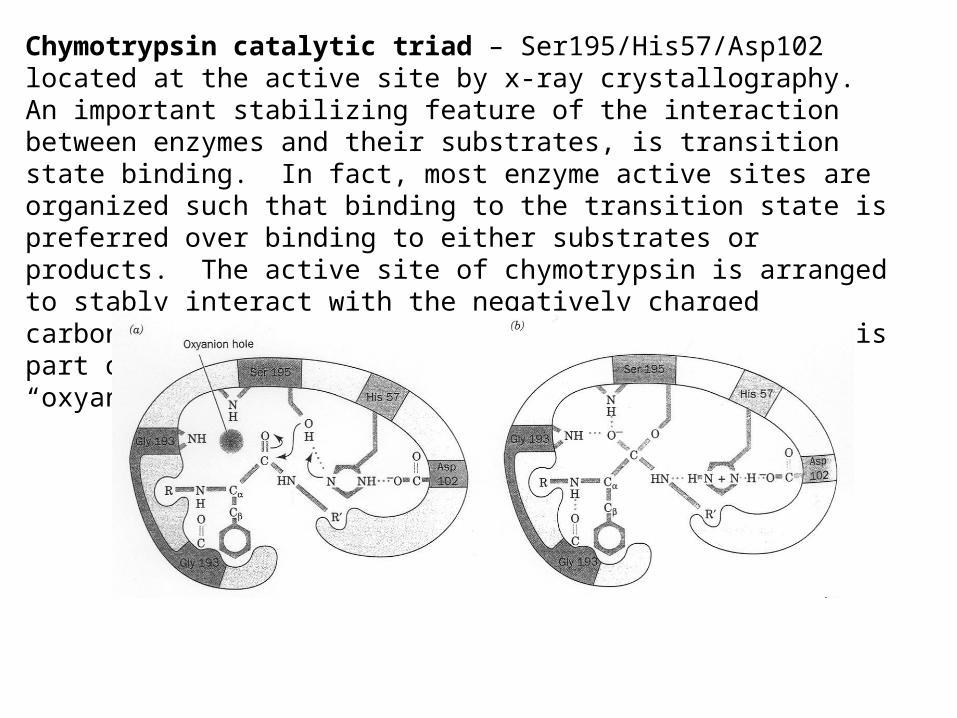
Chymotrypsin catalytic triad – Ser195/His57/Asp102 located at the active site by x-ray crystallography.An important stabilizing feature of the interaction between enzymes and their substrates, is transition state binding. In fact, most enzyme active sites are organized such that binding to the transition state is preferred over binding to either substrates or products. The active site of chymotrypsin is arranged to stably interact with the negatively charged carbonyl oxygen of the tetrahedral intermediate – this part of the active site is referred to as the “oxyanion hole”.
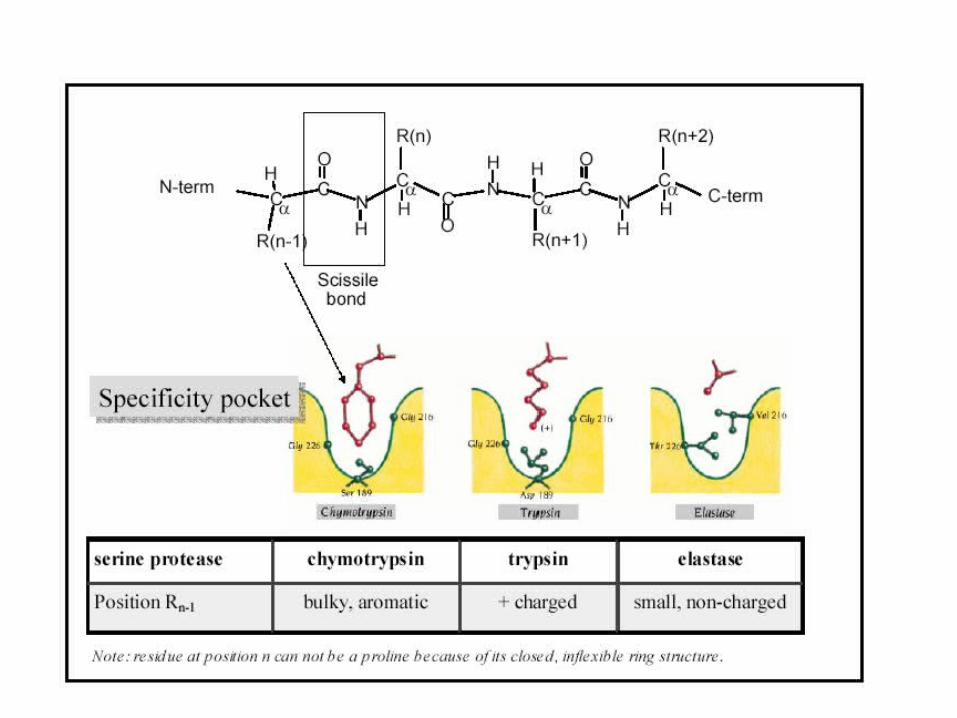

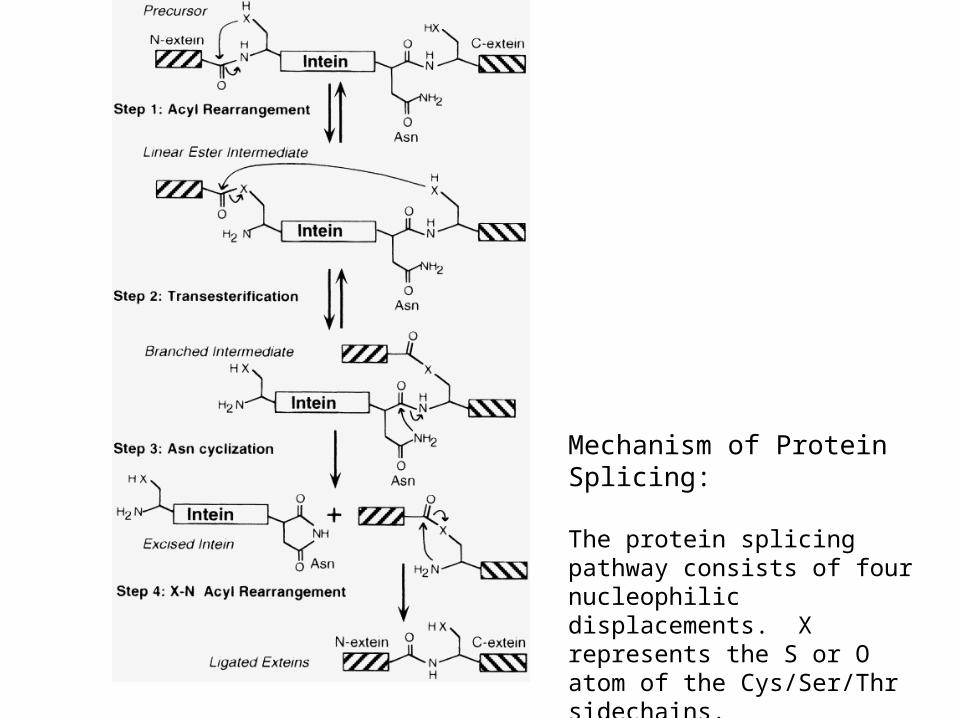
Mechanism of Protein Splicing:
The protein splicing pathway consists of four nucleophilic displacements. X represents the S or O atom of the Cys/Ser/Thr sidechains.
From: Perler, FB (1998) Cell 92, 1-4


















