
Environmental Microbiology Laboratory Department of - Repositorio
167
Environmental Microbiology Laboratory Department of Biology University of Puerto Rico at Rio Piedras Enterococcus faecalis-infecting phages (enterophages) as markers of human fecal pollution and as reservoirs of antibiotic-resistance and virulence genes Tasha M. Santiago Rodriguez A dissertation submitted to the Biology Intercampus doctoral program in Partial fulfillment of the requirements for the degree of Doctor of Philosophy April, 2013
Transcript of Environmental Microbiology Laboratory Department of - Repositorio

Environmental Microbiology Laboratory
Department of Biology
University of Puerto Rico at Rio Piedras
Enterococcus faecalis-infecting phages (enterophages) as markers of human fecal pollution
and as reservoirs of antibiotic-resistance and virulence genes
Tasha M. Santiago Rodriguez
A dissertation submitted to the Biology Intercampus doctoral program in
Partial fulfillment of the requirements for the degree of
Doctor of Philosophy
April, 2013

2
This dissertation has been accepted by the faculty of the
Biology Intercampus Doctoral Program
University of Puerto Rico
Rio Piedras Campus and Medical Sciences Campus
In partial fulfillment of the requirements for the degree of
DOCTOR OF PHILOSOPHY
In the subject of BIOLOGY
Gary A. Toranzos, Ph.D.
Thesis Advisor
Elvira Cuevas, Ph.D.
Committee Member
Maite Muniesa, Ph.D.
Committee Member
Carlos Gonzalez, Ph.D.
Committee Member
Steven E. Massey, Ph.D.
Committee Member
San Juan, Puerto Rico. April 2013.

3
TABLE OF CONTENTS
List of Figures 8-9
List of Tables 10-11
Abbreviations 12
Author’s biography 13
Thesis abstract 14
Dedication 15
Acknowledgments 16-17
Chapter 1: Introduction
General introduction 18
Bacterial and viral indicators
Total and thermotolerant coliforms 19
Enterococci 19-20
Bacteriophages 20-21
Molecular techniques 21-22
Ribotyping and Pulsed –Field gel electrophoresis (PFGE) 22
Denaturing Gradient Gel Electrophoresis (DGGE) and Terminal 22-23
Restriction Fragment Length Polymorphism (T-RFLP)
PCR-based techniques 23-24
Correlations of indicators with pathogens and illness 24-25
Antibiotic-resistance and virulence genes in enterococci 25-26
Tetracycline mode of action and resistance genes 26-27
Vancomycin mode of action and resistance-genes 28-29
Enterococcal surface protein 29-30
Horizontal Transfer of antibiotic-resistance and virulence genes 30-31
Thesis direction and general objectives 32
Novel microbial indicator of human fecal pollution 32
Enterophages as vectors of antibiotic resistance and virulence genes 32-33
Literature cited 34-43

4
Chapter 2: Enterophages, a group of phages infecting Enterococcus faecalis, and their
potential as alternate indicators of human faecal contamination (as published in Water
Science and Technology).
Abstract 44
Introduction 45
Alternate microbial indicators of fecal water contamination 46-47
Materials and Methods
Host strains 47-48
Host bacteria 48
Samples 48
Optimal conditions for viral replication 48-49
Single Layer Plaque assays 49
Plaque isolation 49
Transmission Electron Microscopy 50
Determination of Burst Sizes 50
Survival 51
Prevalence 51
Results and Discussion 51-52
Viral morphology 52
Burst sizes 53
Survival 53-54
Prevalence 54-55
Conclusions 55-56
Acknowledgements 56
Literature cited 57-59
Chapter 3: Characterization of Enterococcus faecalis-infecting phages (enterophages) as
markers of human fecal pollution in recreational waters (as published in Water Reseach).
Abstract 60-61

5
Introduction 61-63
Materials and Methods
Detection of enterophages in animal and human feces 63
Enterophages isolation and purification 64
Morphological characterization of enterophages 64-65
Nucleic acid analyses 65
Prevalence in raw sewage 65
Detection of enterophages and other viral and bacterial indicators 65-66
in a large watershed
Survival of enterophages and coliphages in waters and sand 67
Results and Discussion
Enterophages and coliphages in animal and human feces 67-68
Differences in enterophage isolates according to morphology, genetic 68-69
material and ability to replicate at different temperatures
Prevalence of enterophages in sewage and in a large watershed in Puerto 70-74
Rico
Survival of enterophages in fresh waters 74-75
Survival of enterophages in marine recreational water and sand 75-78
Conclusions 78-79
Acknowledgments 79
Literature cited 80-84
Chapter 4: Microbial quality of tropical inland waters and effects of rainfall events (as
published in Applied and Environmental Microbiology).
Abstract 85
Introduction 86-87
Materials and Methods
Sample collection and sampling sites 87-89
Enumeration of indicators by culture methods 90
DNA extraction and PCR conditions 90-92
USGS precipitation data 92

6
Statistical analyses 92-93
Results
Detection of indicators by culture-based methods 93-94
Detection of host-specific Bacteroides by PCR and enterococci by 94-96
qPCR
Correlations of indicators with rainfall and detection methods 96-98
Discussion
Prevalence of the bacterial and viral indicators detected by culture 98-100
methods
Monitoring indicator bacteria by molecular-based techniques 100-101
Conclusions 102
Acknowledgments 102
Supplementary Information 103-108
Literature cited 109-112
Chapter 5: Evaluation of Enterococcus phages as indeces of fecal pollution (as published in
Journal of Water and Health).
Abstract 113
Introduction 114-115
Materials and Methods
Detection of Enterococcus phages in feces and sewage 115-116
Isolation and characterization of Enterococcus phages 116
Inactivation rates and survival studies 116-117
Statistical analyses 117
Results
Enterococcus phages in animal feces and domestic sewage 117-121
Replication and morphology of Enterococcus-infecting phages 121-122
Inactivation rates and survival of Enterococcus phages 123
Survival of enterophages and coliphages in fresh waters 123-126
Enterophages in chlorinated and dechlorinated tap water and sterile 126-127
distilled, tap and wastewater

7
Discussion
Enterococcus-infecting phages in feces and domestic sewage 127-128
Inactivation rates and survival of Enterococcus phages 128-130
Conclusions 130-131
Acknowledgments 131
Supplementary information 131-138
Literature cited 139-143
Chapter 6: Antibiotic-resistance and virulence genes in tropical environmental
Enterococcus spp. (as accepted in Journal of Water and Health).
Abstract 144
Introduction 145-146
Materials and Methods
Study sites 146
DNA extraction of the bacterial fraction and enterococci 146-148
Virus concentration and DNA extraction 148
Lysogen induction 149
PCR amplification conditions 149-151
Statistical analyses 151
Results
Bacterial fraction and enterococci isolates 151-153
Viral fraction and induced phages 153-154
Discussion 154-156
Conclusions 156-157
Acknowledgments 157
Literature cited 158-162
Chapter 7: General Conclusions and Future Directions
General conclusions 163-165
Future directions 165-166
Literature cited 167

8
LIST OF FIGURES____________________________________________________________
Chapter 1
Figure 1: General mode of action of vancomycin.
Figure 2: Integration of a bacteriophage genome into a bacterial genome.
Chapter 2
Figure 1: Transmission Electron Microscopy image (TEM) of enterophages in this study.
Figure 2: Survival of enterophages at 22 °C (A) and at 37 °C (B).
Chapter 3
Figure 1: Shows study site. Sample points are ordered according to their position in the
watershed, localized in the central region of Puerto Rico.
Figure 2: Transmission Electron Microscopy image (TEM) of an enterophage isolate in this
study.
Figure 3: The genetic material of several enterophage isolates in a 0.7% agarose gel.
Figure 4: Enterophage concentration/mL of four different enterophage isolates among four
different temperatures.
Figure 5: Concentrations of enterophages (A), coliphages (B), enterococci (C) and
thermotolerant coliforms (D) at ten different fresh water sample points with different impacts.
Figure 6: Survival of enterophages (A) and coliphages (B) in fresh water.
Figure 7: Survival of enterophages (A) and coliphages (B) in marine water.
Chapter 4
Figure 1: Rio Grande de Arecibo watershed in Puerto Rico.
Figure 2: Credible intervals (CIs) for thermotolerant coliforms (A), enterococci (B), coliphages
(C), and enterophages (D) in the Rio Grande de Arecibo watershed.
Figure 3: Presence of human-, cattle-, and chicken-specific markers in the Rio Grande de
Arecibo watershed.
Figure 4: qPCR for enterococci and correlation between qPCR and culture-based techniques for
enterococci.

9
Figure 5: Correlation between rainfall and microbial indicators.
Supplementary Figure 1: Enterococci and thermotolerant coliforms CFU/100mL in the Rio
Grande de Arecibo watershed period of high (November to January) and low rainfall events
(February to April).
Supplementary Figure 2: Enterophages in the Rio Grande de Arecibo watershed according to the
incubation temperature and rainfall period.
Supplementary Figure 3: Coliphages in the Rio Grande de Arecibo watershed according to the
incubation temperature and rainfall period.
Chapter 5
Figure 1: Prevalence of Enterococcus phages in raw domestic sewage in three wastewater
treatment plants (WTP) in Puerto Rico.
Figure 2: Enterococcus faecalis-infecting phages isolated from domestic sewage.
Figure 3: Survival of enterophages and coliphages across a tropical watershed in Puerto Rico.
Figure 4: Survival of enterophages in various sterile water types at 37 ºC.
Chapter 6
Figure 1: Sampled sites in this study.

10
LIST OF TABLES_____________________________________________________________
Chapter 1
Table 1: Host specificity of different groups of F + RNA coliphages.
Table 2: Classification of most of the tet determinants according to their mode of action.
Table 3: Vancomycin-resistance in enterococci. Five phenotypes have been identified so far and
enterococci are classified according to the levels of vancomycin and teicoplanin it exhibit
resistant.
Chapter 2
Table 1: Concentrations of coliphages in sewage treatment plants in Puerto Rico (ND-Not
detected).
Table 2: Concentrations of enterophages detected at sewage treatment plants in Puerto Rico.
Chapter 3
Table 1: Prevalence of enterophages in raw and treated sewage at different domestic wastewater
treatment plants in Puerto Rico (PR) and Portugal (Port).
Chapter 4
Table 1: Summary of oligonucleotide primers and probes for PCR and TaqMan qPCR.
Table 2: Reported rainfall during the sampling period (November 2009 to April 2010).
Chapter 5
Table 1: Enterococcus and E. coli-infecting phages per gram of feces in chicken (n=30), cattle
(n=30), dogs (n=12) and pigs (n=10).
Table 2: Mean removal percents of Enterococcus phages in primary effluent from three WTP
(PR-A, PR-B and PR-C) in Puerto Rico.
Table 3: Host range of Enterococcus-infecting phages at different incubation temperatures.
Table 4: T90 values for enterophages and coliphages in three fresh water samples.
Table 5: T90 values (days) for enterophages and coliphages in chlorinated and dechlorinated
drinking water.
Supplementary Table 1: Average of the decay percent day -1
and T90 values for phages infecting
E. faecalis, E. faecium, E. casseliflavus and E. coli in raw sewage at 4°C.

11
Supplementary Table 2: Decay values (log·day-1
) for enterophages and coliphages in three fresh
water samples.
Supplementary Figure 1: Survival of enterophages and coliphages in three sites in a tropical
watershed during a period of high rainfall events.
Supplementary Figure 2: Survival of enterophages and coliphages in three sites in a tropical
watershed during a period of low rainfall events.
Chapter 6
Table 1: Primers in this study.
Table 2: Prevalence of antibiotic-resistance and virulence-encoding genes in the bacterial
fractions of tropical marine (OP, CBB) and fresh waters (LC, RP) and wet and dry sands (OP,
CBB).
Table 3: Prevalence of the esp variants present in E. faecalis and E. faecium and int in tropical
marine (OP, CBB) and fresh waters (LC, RP) and wet and dry beach sands in the Enterococcus
isolates. Percents are presented in parenthesis.
Table 4: Enterococcus isolates tested for tetracycline (16 μg/mL) and vancomycin (20 μg/mL)
resistance isolated from tropical samples.

12
Abbreviations
RNA: Ribonucleic Acid
DNA: Deoxyribonucleic Acid
PCR: Polymerase Chain Reaction
CFU: colony forming units
PFU: plaque forming units
ºC: degree Celsius
mL: milliliters
qPCR: quantitative Polymerase Chain Reaction
SD: standard deviation
μg: micrograms
BLAST: Basic Local Alignment Search Tool

13
Author’s Biography
Tasha M. Santiago-Rodriguez was born on September 4, 1985 in Cayey, Puerto Rico. She
attended the public schools Salvador Brau (elementary), Ramon E. Betances (junior high) and
Miguel Melendez Muñoz (high school) in Cayey. As a junior high school student, Tasha always
had a special interest in science and developed a fascination for microbes. Her academic
achievements and interest in science gave her the opportunity to study Natural Sciences in the
University of Puerto Rico, making Tasha the first of her family to pursue a college education. As
an undergraduate, she had an interest to in medicine. As one requirement for Medical School was
experience in research, she had the opportunity to do summer research at the University of
Medicine and Dentistry of New Jersey during the summer of 2005. She worked under the
supervision of Dr. Vincianne Gaussin and Dr. Boudewijn Kruithof, experts in the field of cardiac
valve development. In 2007, Tasha graduated Magna-Cum Laude, but was not accepted into
Medical School. During this time, Tasha strongly considered graduate school as an alternative to
pursue a career in one of her fields of interest, microbiology. In 2008, Tasha was accepted into
the graduate Biology Department at the University of Puerto Rico at Rio Piedras. Tasha has
presented in both local and international meetings including the Health-Related Water
Microbiology and the American Society of Microbiology meetings. This has given her the
opportunity to visit countries such as Greece and New Zealand. She has published 5 articles in
important journals related to water quality including Water Research and Applied and
Environmental Microbiology. Her fields of interest are not limited to environmental
microbiology or the microbial quality of waters, as other interests include ancient DNA and
mechanisms of microbial communication. She has contributed in the characterization of the
microbial communities of coprolites from Pre-Columbian cultures as a way to elucidate dietary
habits and cultural traditions. Data from this project have been accepted for publication in
PLosOne. Tasha has also contributed in the characterization of amber bacteria dating to millions
of years in collaboration with Dr. Raul Cano. She also has an interest in identifying genes
involved in ligninocellulose degradation as alternative sources of energy, resulting in at least 2
manuscripts. Her academic achievements in graduate school make her the student with the
highest number of publications in the history of the Biology Program at the UPR. She has been
accepted as a postdoctoral fellow at the University of California at San Diego, School of
Medicine.

14
THESIS ABSTRACT
The introduction of human fecal material into water sources represents a concern to public health
since pathogens could be introduced. Not all laboratories possess the facilities to detect human
enteric pathogens and thus microbial indicators are used. An ideal microbial indicator of human
fecal pollution should be: (i) enumerated using simple laboratory methods, (ii) present in fecally
polluted waters and absent in pristine waters, (iii) associated with the source of the
contamination, (iv) able to survive and inactivate similarly to the pathogen of concern and (v) be
detected in various geographical regions. No microbial indicator of fecal pollution satisfies all
these characteristics. Therefore, we have developed a relatively simple culture technique to
characterized bacteriophages that infect a specific type strain of Enterococcus faecalis, which we
call enterophages, as markers of human fecal pollution. Enterophages were detected exclusively
in domestic wastewaters in Puerto Rico and Portugal, tropical recreational waters and possess a
survival and inactivation rates similar to human pathogenic enteric viruses. Enterophages possess
non-contractile tails of 60 to 200nm, icosahedral capsids of 12 to 80nm and dsDNA genomes of
30 to 40kb. Given that their role as vectors of genes conferring antibiotic-resistance and
virulence to the bacterial host remains largely unknown, the complete genomes of two phages
infecting E. faecalis (one lytic and one lysogenic) are currently being sequenced. Lysogenic
Enterococcus phages, as well as their host isolated from marine and fresh waters, dry and wet
sands in Puerto Rico, were tested for tetracycline (tetM) and vancomycin-resistance (vanA and
vanB) genes. The prevalence of the enterococcal surface protein (Esp), a virulence factor, was
also determined given that it has received great attention as a marker of human fecal pollution.
The prevalence of an integrase-encoding gene (int) specific for E. faecalis phages was
determined since integrases are markers of lysogeny and are responsible for the integration of the
phage genome into the bacterial host. tetM, vanA, vanB, esp and int were only detected in the
bacterial fraction and enterococci, with the exception of int, which was detected in the viral
fractions and lysogenic enterococci phages.

15
DEDICATION
This thesis is dedicated to my mother and grandmother, for their unconditional support to pursue
this journey.

16
ACKNOWLEDGMENTS
Special thanks to my advisor Dr. Gary A. Toranzos and my thesis committee, Dr. Elvira Cuevas,
Dr. Maite Muniesa, Dr. Carlos Gonzalez and Dr. Steve Massey, for their guidance, support,
inputs, comments and suggestions for the present dissertation.
To all the coauthors of the publications resulting from the present project: Natasha Bonilla,
Catalina Davila, Dr. Patricia Marcos, Manuela Cadete, Sylvia Monteiro, Jessica Rivera, Mariel
Coradin, Joel Gonzalez, Dr. Carlos Toledo, Dr. Raymond Tremblay, Dr. Jorge Santo-Domingo,
Miguel Urdaneta, Dr. Hodon Ryu and Dr. Ricardo Santos.
To all the undergraduate students of the Environmental Microbiology Laboratory (2008-2013),
of which I had the opportunity to teach something and learn from: Gwendolyn Arguello, Jean F.
Ruiz, Gabriela Tirado, Alfredo Gonzalez, Dashari Colon, Jose Soto and Alex Vermont.
To the Autoridad de Acueductos y Alcantarillados for collecting sewage samples and Marisol
Rodriguez (my mom) for collecting fresh water samples from el Rio La Plata. To the US
Geological Survey and the Department of Natural Resources of Puerto Rico for data used in the
present project.
I would like to thank Dr. Pablo A. Ortiz Pineda who taught me several of the bioinformatic
programs and molecular techniques, and for support to pursue this project. I would also like to
thank Natasha Bonilla who taught me the culture techniques for the detection of enterophages
and Ana Rita Patricio and Silvia Planas who taught me DNA extraction, PCR and
bioinformatics.
Special thanks to the following personnel of the Biology Department: Dr. Garcia-Arrarás, Millie
Viera, Aidamarie Perez, Diana Rosario, Gladys Ramos and Jose Fontánez for their excellent
work.
I would like to give special thanks to Dr. Raul Cano for support, guidance, and for giving me the
opportunity to work in other projects (amber bacteria, El Yunque and the coprolites) which have

17
given me the opportunity to learn many cool things. Special thanks for teaching me lessons
which I know are very valuable as a scientist and as a person.
I would like to thank the Environmental Protection Agency, DEGI, the Biology graduate
program and the RISE program for financial support. Without this support, I would not have had
the opportunity to travel to present, network and learn new techniques.
To my brother Pedro Santiago and my aunts: Evelyn and Sonia Rodriguez, Johanna and Pichi
Vazquez, for their unconditional support. Most importantly, I would like to thank my mom,
Marisol Rodriguez and grandmother, Enriqueta Marrero, to who I owe this and at least 10 more
theses.

18
CHAPTER 1
General Introduction
Clean water sources are vital for drinking and recreational purposes, but many can get
contaminated by fecal material. Direct contact with and ingestion of waters contaminated by
fecal material can affect millions of people every year as a result of the gastrointestinal and
respiratory illnesses and eye and skin infections caused by enteric pathogens. Among the
possible types of fecal contamination, that from a human source may represent a major concern
to our health given that human enteric pathogenic bacteria, protozoans and enteric viruses are
among those that can cause most of the mentioned diseases. Thus, the identification of the fecal
contamination in waters is necessary to eliminate the source and to minimize the potential risk
that enteric pathogens represent to public health. Other concerns with human enteric pathogens is
that these can cause illness with infectious doses as low as 1 to 10 [3, 4], can exhibit resistance to
removal and inactivation treatments (e.g. chlorination) [5-7] and can be reintroduced from
sediments to surface waters after disturbances (e.g. precipitation events) [8-11].
To remediate the source of the fecal contamination and avoid disease, enteric pathogens can be
detected using culture and/or molecular methods. However, culture methods may be expensive
and time-consuming, and not all enteric pathogens are cultivable. Molecular methods may
circumvent culturing microorganisms, and although current water quality guideline standards are
implementing molecular methods, several drawbacks have been identified, such as the detection
of noninfectious and unviable enteric pathogens. In addition, detection of human enteric
pathogens using culture or molecular methods may be relatively expensive, require the right
personnel and equipment. Moreover, given that many different human enteric pathogens can be
introduced into water sources, it would be impractical trying to detect them simultaneously [3].
In order to infer fecal contamination and the possible presence of enteric pathogens in water
sources, other enteric microorganisms (e.g. enterococci, thermotolerant coliforms and
bacteriophages) are used, and these are known as microbial indicators of fecal pollution [3, 12-
14]. These microorganisms should satisfy several characteristics in order to be considered
reliable indicators of fecal contamination: (i) be detected using simple laboratory methods, (ii) be
present in fecally polluted waters and absent in pristine waters, (iii) be associated with the source

19
of the contamination, (iv) possess a survival similar to that of the pathogen of concern and (v) be
unable to replicate outside the host. However, as discussed below, currently used microbial
indicators fail to fulfill many of these characteristics and thus studies are currently being
conducted in order to identify and characterize novel markers of fecal pollution [7].
Bacterial and viral indicators
Total and thermotolerant coliforms
Total coliforms have long been used to assess the microbial quality of water sources since they
are present in the intestinal tract of animals. These bacteria can ferment lactose and produce gas
within 24 to 48 h at 35 ºC and include the genera Escherichia, Klebsiella, Enterobacter and
Citrobacter. Similarly, thermotolerant coliforms are part of the intestinal microbiota of warm-
blooded animals and fecal discharges contain large numbers of bacteria belonging to this group,
but unlike total coliforms, can grow at 45 °C [15]. Thermotolerant coliforms are currently being
used as an index of fecal contamination in many water sources; however, total and
thermotolerant coliforms may also originate from non-fecal sources such as run-off [16]. In
addition, some members of the coliform group can replicate in subtropical and tropical waters
and have been found to be part of the environmental microbiota of many of these water sources
[17, 18]. Those members that have been linked to fecal pollution possess survival times and
inactivation rates that cannot be correlated with those of many human enteric pathogens under
similar conditions. In addition, coliforms have been detected in the absence of enteric pathogens
and viceversa, and thus there is a lack of correlation between the number of these indicator
bacteria and enteric pathogens [13, 19].
Enterococci
Enterococci are gram-positive bacteria that include members of the genus Enterococcus, which
can grow at a wide temperature range (10 to 45 ºC), basic pH (9.6) and high salinities (6.5 %
NaCl). These have been accepted as indicators of fecal pollution since are commonly found in
the feces of warm blooded animals and their prevalence seems to be similar to that of many
bacterial pathogens [20]. However, confirmation methods are often needed when detecting
enterococci and these can be relatively laborious and results require are obtained within 24 h
[21]. Enterococci may not be used to track the source of the fecal contamination given that these

20
are present in the intestinal tracts of different warm-blooded animals and interestingly, these
bacteria have also been detected in the intestines of pigeons and in flies [22, 23]. In addition,
enterococci have been detected in pristine waters and have been detected when pathogens are
absent and viceversa [13, 24].
Bacteriophages
Bacteriophages are viruses that infect bacteria and are present in the intestinal tract of warm-
blooded animals. Phages are very specific in terms of the bacteria they infect and source (i.e.
those isolated from the feces of humans have not been isolated from the feces of other animals)
[25]. They have been specifically proposed and used as indicators of the virological quality of
waters since the 1970’s [26]. Because of this source specificity, the use of phages as indicators of
fecal pollution arose. Other reasons for considering phages as indicators of fecal contamination
included the need to find models of human enteric viruses and a reliable method to assess the
virological quality of waters. The similar morphology, structure and “behavior” of
bacteriophages to that of many human enteric viruses, suggests that they should be more reliable
indicators of the virological quality of water sources than indicator bacteria. In addition, the
bacteriophage method possesses several advantages compared to the bacteriological methods: are
relatively less expensive, results are obtained in less time (4 to 6 h after incubation) and do not
require laborious confirmation methods [24, 27]. In addition, bacteriophages are as or more
resistant to removal and water disinfection processes compared to many enteric viruses [28-30].
Among the bacteriophages proposed as indicators of fecal contamination are those infecting
Bacteroides fragilis. Bacteroides fragilis phages have been linked to human fecal pollution [31-
33], since they are absent when enteric viruses are absent, do not replicate under environmental
conditions, seem to be more resistant to various water treatments than human enteric viruses and
have been proposed as a tool for viral waterborne disease control [34]. However, they have been
detected only at certain geographical regions of USA and Europe. In addition, the techniques
used to detect B. fragilis bacteriophages are difficult since the conditions must be anaerobic [35,
36].

21
Table 1: Host specificity of different groups of F + RNA coliphages. Modified from [1].
Somatic and F (male)-specific coliphages have also been proposed and used as indicators of fecal
pollution [35, 37, 38]. Coliphages infect bacteria belonging to the Enterobacteriaceae family,
being E. coli the most widely host used for their detection. Somatic coliphages, like other
somatic phages, infect bacteria by attaching to specific surface receptors. In contrast, F(male)-
specific coliphages only infect conjugating bacteria by recognizing receptors in the pilli. F
(male)-specific coliphages possess genomes of + RNA or + DNA and belong to the Leviviridae
and Inoviridae family, respectively. F + RNA coliphages have been well characterized as
markers of fecal pollution and the possible sources, but less is known about F + DNA coliphages
[39]. However, the use of coliphages as indicators of specific sources of fecal contamination may
also have several disadvantages. The reason for this is that, even thought it has been suggested
that serotypes II and III of F (male)-specific coliphages could be used to infer human fecal
contamination [1, 40] (Table 1), other studies have found that the same serotypes can also be
found in animal feces [41]. In addition, coliphages are rarely detected in human feces, their
concentrations and survival cannot be correlated with that of many enteric viruses and certain
studies have found that somatic coliphages can replicate in the environment [35, 42].
Molecular techniques
Over the past years, molecular methods have been developed and tested to identify possible fecal
sources [43-45]. Currently developing methods generally involved the amplification of the
microbial indicator’s nucleic acids by using specific primers, although other methods have been
tested and will be discussed below. For some researchers, molecular methods offer the advantage
of obviating the need for cultivation. Culturing microorganisms is considered for some a
laborious procedure, which involves selective media and enrichment in order to isolate a specific
microbial indicator. Culturing microbial indicators may underestimate their numbers as some
Group Host
I Non-human animals.
II Humans and occasionally pigs.
III Exclusively human.
IV Non-human origin with rare human associations.

22
may be unviable and uncultivable, and thus molecular methods may circumvent this. However,
molecular methods may not always be as ideal as expected. In this section, the advantages and
disadvantages of several molecular methods that have been used and are currently accepted to
infer fecal pollution are discussed.
Ribotyping and Pulsed-Field Gel Electrophoresis (PFGE)
Ribotyping consists of a fingerprint pattern resulting from differences in the size of DNA
fragments. Total genomic DNA is extracted from pure cultures and is treated with enzymes,
resulting in digested DNA fragments. The fragments are separated using agarose gel
electrophoresis and then transferred to nylon membranes. Southern blot hybridization is then
performed using rDNA probes, which results in a pattern composed of several bands. Various
restriction enzymes can be used in further analyses if one wants to increase the specificity of the
results [46]. The technique has been used to distinguish E. coli of human and non-human origins.
However, the technique is poor when analyzing samples containing multiple sources of fecal
contamination [47, 48]. Similarly, in the PFGE technique, pure bacterial cultures are placed in
agarose plugs and the DNA is digested using restriction enzymes. The digested plugs are
embedded into electrophoresis gels with alternating currents. Although PFGE has been used for
the characterization of environmental E. coli and enterococci, specialized equipment is
necessary, the technique may be time-consuming and the number of isolates which can be
processed simultaneously is limited [49].
Denaturing Gradient Gel Electrophoresis (DGGE) and Terminal-Restriction Fragment
Length Polymorphism (T-RFLP)
The DGGE is a technique able to differentiate between PCR products having similar sizes and
this is due to the melting properties of the DNA fragments and their mobility in the gel. DGGE
has been used to characterize fecal bacteria populations of both animals and humans. Yet, one
disadvantage of this technique is that the gene targeted must possess enough sequence variability
among the bacteria in order to identify the possible fecal source [50]. PCR products, like those of
the 16S rRNA gene, can also be detected using a DNA sequencer when fluorescently labeled, a
technique known as T-RFLP. The method is used to determine differences in the lengths of gene
fragments and offers the advantage of obviating culturing bacteria from the environmental

23
samples. It has been tested in the feces of farm animals and humans, but a more studies are
needed to determine if T-RFLP is suitable for other types of fecal contamination [51, 52].
PCR-based techniques
PCR-based methods have acquired great interest as these have shown to identify many sources of
fecal contamination. Moreover, water quality guidelines may incorporate PCR-based techniques
to routinely assess the microbial quality of waters since these are relatively simple and results are
obtained in less time compared to most culturing methods. Among the most accepted PCR-based
methods are the host-specific. Bernhard and Field developed PCR primers for the amplification
of conserved 16S rRNA gene sequences present in Bacteroides species specifically present in
cattle and humans. Detection of these genera possess several advantages: (i) in phylogenetic
analyses, members have shown to group according to the host, (ii) are restricted to warm-
blooded animals, (iii) are among the most numerous microorganisms in feces and (iv) possess
short survival times once introduced into water sources, thus replication in the environment is
not likely. There was the need to design PCR primers for the detection of Bacteroides because
these microorganisms are difficult to culture due to their strictly anaerobic nature. Interestingly,
differences were noted in the 16S rRNA gene sequences of bacteria present in human and cattle
fecal material and accordingly, primers targeting these sequences can be used to identify possible
sources of fecal contamination [53].
Studies by Bernhard and Field are among the pioneer studies about primer design for the
detection of host-specific bacteria. However, the presence/absence of indicator bacteria may not
reflect the extent of the fecal contamination. For this reason, most PCR-based techniques aim to
quantify the nucleic acids of indicator bacteria, being quantitave PCR (qPCR) one of the most
accepted and currently tested in human and animal fecal materials and water sources. qPCR
assays for the identification of human fecal contamination have shown to be specific and
sensitive, as in the case of the human Bacteroides markers HF-183 and BacHum-UCD. Markers
of animal fecal contamination, such as BacCow-UCD (cattle) and BacCan-UCD (dog) have
shown to be specific, but not necessarily sensitive since these amplify a fraction of the
corresponding fecal material and in some cases, cross reactions with horse fecal matter (as in the
case of BacCow-UCD) and human feces (as in the case of BacCan-UCD) can occur [54]. In

24
addition, it has been suggested that qPCR may overestimate the concentrations of indicator
bacteria in different water sources, and thus many studies have correlated qPCR with culture-
based methods.
Correlation results between qPCR and culture methods depend in the water type tested. For
instance, in temperate marine waters, positive correlations between the two methods have been
noticed [55]. In subtropical marine waters, the detection of E. coli using uidA and enterococci
using the 23S rRNA gene have positively correlated with their respective culture methods.
Interestingly, in subtropical climates, the strength of the correlations also depends in the sample
site and the time at which the samples are collected. Accordingly, a stronger correlation between
qPCR and culture methods for enterococci are noticed when samples are collected in the
morning [56]. In the present thesis, the correlation between qPCR and culture methods for the
detection of enterococci in tropical inland waters is presented in Chapter 4. Correlations
between molecular and culture-based methods may suggest that either technique could be used.
This represents, for some researchers, an opportunity to substitute culturing microorganisms with
molecular techniques. Results are often obtained in less time and thus immediate actions can be
taken to remediate the source of the fecal contamination and minimize the possible risks to
public health. However, for some researchers, correlations between both molecular and culture
techniques may represent a disadvantage. The reason for this is that it has been suggested that a
toolbox of methods may be more appropriate to infer fecal pollution.
Correlations of indicators with pathogens and illness
Ideally, the characterization of indicators of fecal pollution should include correlation analyses
with enteric pathogens. Studies of this type are limited since not all laboratories possess the
facilities to detect enteric pathogens [57]. The presence of specific microbial indicators has
positively correlated with the presence of bacterial pathogens, as in the case of total coliforms
and Clostridium perfringes with Salmonella, but in other cases, coliforms do not correlate with
enteric pathogens [58, 59]. Possible reasons for this include differences in the prevalence and
survival times, and the ability of coliforms to become part of the environmental microbiota of
waters [60]. In terms of the enterococci, their prevalence has positively correlated with human
enteric viruses [61]. Other studies, however, have found no correlation between enterococci

25
detected by molecular or culture methods and Bacteroides spp. with Campylobacter spp. (a
pathogenic bacteria that causes gastroenteritis) [62]. Differences in the correlation analyses
between indicators and pathogens may be due to the water types tested, as aquatic ecosystems
may be differently impacted by ecological factors (e.g. salinity, transport of microbes from other
sites, turbidity and rainfall). Other influential variables may include differences in the type and
numbers of indicators and pathogens (as the latter are often detected in lower concentrations),
pathogen source, sample size and statistical methods. Statistical methods may have a great
influence on the results as these depend on the sample size and recent studies have suggested that
discrepancies between correlations between indicators and pathogens may be due to the
insufficient data for assessing these correlations [63]. It has also been suggested that microbial
indicators of fecal pollution may not necessarily infer the presence of pathogens, rather, it is a
probability of their co-occurrence [64].
Correlations studies also involve the epidemiological aspects of the health effects associated with
microbial indicators of fecal pollution. These studies aim to determine symptoms associated with
gastrointestinal and respiratory illness and ear, eye and skin infections after swimming in
possibly fecally contaminated waters and the correlation with the presence of microbial
indicators. Reports often involve threshold values of indicator-bacteria and symptoms associated
with the mentioned illnesses and how variations in the severity of the health effects correlate
with the extent of the fecal contamination [65]. Most of the positive correlations are observed
between gastrointestinal illness and enterococci, thermotolerant coliforms and E. coli and
interestingly, most of the thresholds are lower to those of current water quality guidelines [66].
Interestingly, other studies have reported a correlation between enterococci and skin illness [67],
but these variations may be due to differences in the experimental designs (e.g. the indicators
tested, participants and water type).
ANTIBIOTIC-RESISTANCE AND VIRULENCE GENES IN ENTEROCOCCI
Fecal contamination of water sources can also result in the introduction of bacteria harboring
antibiotic-resistance and virulence genes, but the possible risks to public health remain largely
unknown [68-70]. Specific antibiotic resistance phenotypes have been proposed as a tool for
microbial source tracking (MST) [71]. This is based on the hypothesis that bacteria exposed to

26
antibiotics will develop resistance and this selective pressure can be a mechanism for
discriminating fecal microorganisms from different sources. The potential problem when using
antibiotic resistance as a tool for MST is that bacteria can transfer the resistance genes to other
bacteria as these can be found on a variety of mobile genetic elements (e.g. plasmids and
transposons) [72-74]. Similarly, virulence factors have also been proposed as a means to infer
specific sources of fecal pollution. Virulence factors are ways that bacteria have developed to
circumvent the host defenses. However, as discussed below, many of these studies focus on the
presence/absence of the genes conferring virulence and most ignore the ecology of the
microorganisms harboring these genes.
One of the most studied bacteria harboring antibiotic-resistance and virulence genes is
Enterococcus spp., which can carry genes encoding for resistance to tetracycline and
vancomycin, a last resource drug, and the enterococcal surface protein (Esp), a virulence factor
[75, 76]. Tetracycline and vancomycin have been prescribed to treat infections caused by
Enterococcus [77], but the excessive use of these antibiotics has lead to an increase in
Enterococcus isolates exhibiting resistance to both antibiotics. Many of these antibiotic-resistant
enterococci can also harbor Esp, which enables bacteria to form biofilms, often difficult to treat
with antibiotics. In the next sections, the molecular aspects of tetracycline and vancomycin-
resistance genes and esp are discussed. Chapter 6 presents the prevalence of these gene in the
environment.
Tetracycline mode of action and resistance genes
Tetracyclines are among the wide-spectrum antibiotics commonly used for treating infections.
These belong to a family of antibiotics which inhibit protein synthesis by binding to the
ribosomal acceptor site (A-site) and thus inhibit the association of the tRNA with the bacterial
ribosome. Tetracycline-resistant bacteria can carry at least one of the 38 known tetracycline-
resistant genes. Most of these genes encode for the formation of efflux pumps, which are
membrane-associated proteins that move tetracycline out of the bacterial cell. Genes encoding
for efflux pumps can be found in both gram-positive and gram-negative bacteria. This efflux
activity in bacteria is the result of the expression of two genes: one that encodes for the
membrane-associated protein and another that encodes for a repressor protein. In the absence of

27
tetracyclines, the repressor protein inhibits the expression of the gene encoding for the formation
of the efflux pumps. Other tetracycline resistance genes encode for ribosomal protection
proteins, which attach for a short time to the tetracycline-binding site, inhibiting the association
of tetracycline with the A-site. Other data have suggested that ribosomal protection proteins
bind to the ribosomes and change their conformation [75, 76, 78]. Tetracycline resistance genes
also include those encoding for inactivating enzymes, which inactivate tetracycline (tet(X)) and
tet(U), whose function remains unknown (Table 2).
Table 2: Classification of most of the tet determinants according to their mode of action.
Modified from [75].
Prevalence of tetracycline-resistance genes has been determined in the feces of humans and
animals, clinical settings and waters in contact with swine-production facilities [79, 80]. These
have been identified in mobile elements, such as plasmids and transposons and thus their lateral
transmission has been reported, even between bacteria of different genera [81-83]. Most of the
tetracycline-resistance genes have been identified in gram-negative bacteria, but others are
present in gram-positive bacteria (e.g. tet(M), tet(K), tet(L)) and are not structurally similar to
those of gram-negative bacteria. Among the tetracycline-resistance genes found in gram-positive
bacteria, tet(M) is the most disperse and is found in many Enterococcus spp. (E. faecalis, E.
faecium and E. gallinarum) and has received greater attention [75, 83, 84]. For this reason, its
prevalence has been determined in clinical settings, sewage and environmental settings [79, 80,
85]
Efflux proteins Ribosomal protection Inactivating enzymes Unknown
tet(A), tet(B), tet(C),
tet(D), tet(E), tet(G),
tet(H), tet(J), tet(L),
tet(V), tet(Y), tet(Z)
tet(M), tet(O), tet(S),
tet(W), tet(Q), tet(T)
tet(X) tet(U)

28
Vancomycin mode of action and resistance-genes
Vancomycin is used to treat infections caused by gram-positive bacteria and it is often used to
treat Enterococcus infections exhibiting resistance to other antibiotics. It is, therefore, a last-
resource drug as it possesses hazardous side effects. It acts by inhibiting the synthesis of the cell
wall and thus bacterial replication. Briefly, vancomycin binds to the ᴅ-Ala-ᴅ-Ala moieties of the
N-acetylmuramic acid (NAM) and N-acetylglucosamine (NAG) peptides, preventing the
backbone polymers to form, resulting in the destabilization of the cell wall (Figure 1). However,
the use of this antibiotic has resulted in the isolation of vancomycin-resistant enterococci from
clinical settings and from the environment to a lesser extent [78, 86-88].
Figure 1: General mode of action of vancomycin. Vancomycin attacks the ᴅ-Ala-ᴅ-Ala moieties
of the cell wall, causing instability and cell death.
Five vancomycin-resistance phenotypes have been identified in enterococci so far: VanA, VanB,
VanC, VanD and VanD. Vancomycin-resistant enterococci are classified according to the
exhibited resistance to various concentrations of vancomycin and teicoplanin, an antibiotic
mainly used in Europe. Bacteria exhibiting the VanA phenotype are resistant to a wide
concentration of vancomycin and teicoplanin. The VanB phenotype is characterized by bacteria
showing resistance to different concentrations of vancomycin, but unlike the VanA phenotype,
bacteria exhibiting the VanB phenotype, are resistant to lower levels of teicoplanin. Enterococci
exhibiting the VanC phenotype are resistant to lower levels of vancomycin compared to the
VanA and VanB phenotypes. The VanD phenotype is characterized by bacteria resistant to
intermediate levels of vancomycin and has been described in E. faecium. The VanE phenotype is
characterized by E. faecalis strains exhibiting resistance to lower doses of vancomycin and
teicoplanin compared to the VanA phenotype (Table 3).
NAM/NAG
peptides
D-Ala-D-Ala
Moieties
Vancomycin

29
Among the vancomycin-resistance phenotypes in enterococci, the VanA and VanB have received
greater attention. These phenotypes are the result of the expression of operons which include
vanA or vanB, vanRAB, vanSAB, vanHAB, vanXAB and vanZAB. VanA and VanB are ligases which
produce ᴅ-Ala-ᴅ-Lac instead of ᴅ-Ala-ᴅ-Ala. VanH is a ᴅ-hydroxy acid dehydrogenase which
creates a pool of ᴅ-lactate for use in the previous reaction. VanX is a ᴅ, ᴅ-dipeptidase lacking
activity against ᴅ-Ala-ᴅ-Lac and reduces the availability of ᴅ-Ala- ᴅ-Ala produced by the
enterococcal ligase, thereby minimizing the competing synthesis of normal pentapeptide [89].
Among the genes discussed, those encoding for the ligases, vanA and vanB, are considered when
detecting specific vancomycin-resistance phenotypes and may be present in the bacterial
chromosome, plasmids or transposons [89]. The prevalence of vancomycin-resistant genes have
been mainly determined in hospitals, sewage and in aquatic ecosystems [90, 91].
Characteristic Phenotype
VanA VanB VanC VanD VanE
Vancomycin MIC
(mg/mL)
64->1000 4-1024 2-32 128 16
Teicoplanin MIC
(mg/mL)
16-512 < 0.5 < 0.5 4 0.5
Most frequent
enterococcal species
E. faecalis, E.
faecium
E. faecalis, E.
faecium
E. gallinarum, E. casseliflavus,
E. flavescens
E.
faecium
E.
faecalis
Table 3: Vancomycin-resistance in enterococci. Five phenotypes have been identified so far and
enterococci are classified according to the levels of vancomycin and teicoplanin it exhibit
resistant. Modified from [89].
Enterococcal surface protein
Enterococci exhibiting resistance to antibiotics may also harbor virulence factors [92]. Virulence
factors are mechanisms that bacteria have evolved to avoid the host defenses and thus can result
in pathogenicity. Among the virulence factors present in enterococci is the enterococcal surface
protein, encoded by esp. This gene has acquired great attention since nosocomial infections,
including urinary tract infections, endocarditis and bacteremia, are caused by enterococci
harboring esp [93, 94]. The major problem with enterococci harboring esp is that these can form

30
aggregates or biofilms which exhibit resistance to antimicrobial treatments [95]. The gene was
first discovered in E. faecalis strains, but a variant of the gene was later discovered in E. faecium.
Further studies detected the esp variant present in E. faecalis in both human and animal feces,
and the E. faecium variant was detected uniquely in human fecal material [96]. This resulted in
the characterization of esp as a tool for MST purposes.
Conflicting MST studies have tried to determine the specificity of esp. In these studies, PCR
primers were designed based on available sequences, and their ability to amplify enterococci
DNA in fecal matter, sewage and surface waters impacted by fecal contamination. Many studies
have been successful in the amplification of the gene from human feces and these same studies
have determined the absence of the gene in animal fecal material [97, 98]. However, other
studies have amplified the gene in environmental enterococci and thus the specificity of the gene
has been questioned. This is the case of studies performed by Byappanahalli and colleagues,
which have detected the gene in enterococci isolated from sources with no apparent input of
fecal material. The same studies have detected esp in Cladophora, one main source of
enterococci in water sources. In addition, environmental factor can affect the detection of esp,
including rainfall. It has been found that the prevalence of esp is higher after precipitation events
[99].
HORIZONTAL TRANSFER OF ANTIBIOTIC-RESISTANCE AND VIRULENCE
GENES
It is well known that tetracycline and vancomycin resistance genes are present in mobile
elements, and can be transferred between bacteria [76]. These genes can also be present in the
bacterial chromosome, and this is also the case of esp, but few studies have determined the
transferability of esp to other bacterial species [94, 100]. Among the possible vehicles mediating
the transmission of antibiotic-resistance and virulence genes, bacteriophages are often not
considered. Phages can harbor genes which are not indispensable for their “life cycles”, but
could be transferred to the recipient bacteria and result in an increased fitness. Such is the case of
the lateral transmission of antibiotic-resistance and virulence genes mediated by temperate
phages, but few studies are still available [101]. Lysogenic phages may have a greater impact in
the evolution of bacteria than what is believed.

31
Phages are classified as strictly virulent (lytic) or temperate (lysogenic). Lytic bacteriophages
lyse their host bacterium, within a couple of minutes to hours after infection, by producing
hundreds or thousands of phages. On the other hand, a temperate phage may not lyse its host
immediately. Rather, it integrates its genome into the bacterial chromosome, until environmental
conditions are unfavorable, triggering the expression of the phage lytic genes [102, 103]. The
state of a phage to remain “dormant” in a bacterial chromosome is known as lysogeny, and it is
governed by the expression or repression of the phage genes. Specifically, the protein known as
CI, which is codified by the cI gene is involved in the repression of the expression of the phage
genes involved in the lytic cycle. Expression of the cI gene is in turn regulated by the CII and
CIII proteins, encoded by the cII and cIII genes. It is possible that CI-like proteins in E. faecalis-
infecting phages possess the same function as those found in the λ bacteriophage. Lysogenic
phages also harbor anti-repressor genes, which encode for proteins that interfere with the
function of repressors, promoting the expression of the phage genes and thus inducing the lytic
cycle [104].
The lysogeny state in λ and lysogenic E. faecalis phages is governed by a module of genes which
include those encoding for integrases [104]. Integrases are responsible for the integration of the
phage genome into the bacterial genome and this is due by the recognition of specific nucleotide
sequences in both the phage and bacterial genome (Figure 2). The recognition sites in the phage
and bacterial genomes are known as the attP and attB sites, respectively [2, 105].
Figure 2: Integration of a bacteriophage genome into a bacterial genome. Integrases recognize
specific nucleotide sequences in the phage and bacterial genomes (attP and attB sites,
respectively). Modified from [2].

32
THESIS DIRECTION AND GENERAL OBJECTIVES
Novel microbial indicator of human fecal pollution
Given that no microbial indicator of human fecal pollution satisfies most of the characteristics
mentioned previously, there is a need of identifying and characterizing novel indicators of this
type of contamination. Therefore, we have proposed a group of phages that infect a specific type
strain of Enterococcus faecalis, which we call enterophages, as indicators of human fecal
pollution. The present project presents data of the characterization of enterophages as markers of
human fecal pollution. Although results are promising, there is still the need of testing
enterophages in other geographical areas as only domestic wastewaters, marine and fresh waters
in Puerto Rico and domestic wastewaters in Portugal have been tested.
Enterophages as vectors of antibiotic resistance and virulence genes
Since tetracycline and vancomycin-resistance and esp are present in enterococci, it is fair to
believe that lysogenic phages infecting Enterococcus possess genes similar to those found in the
bacterial hosts. This has been previously found in coliphages, which possess a module of genes
known as R factors. R factors can be transferred and confer tetracycline and streptomycin-
resistance to the host bacteria [106]. Other studies have found that the viral DNA fraction of
sewage and environmental samples harbor methicillin-resistance [107]. Similarly, certain
coliphages can harbor Shiga toxins-encoding genes, a virulence factor, which can be transferred
to specific E. coli strains and cause serious risks to health [108-110]. It was unknown if phages
infecting Enterococcus harbor the mentioned genes, and although none were detected in
Enterococcus phages or the viral DNA fractions of environmental samples, tetracycline and
vancomycin resistance genes, and esp, were detected in the bacterial fractions and enterococci
isolates. However, it should not be ignored that Enterococcus phages can harbor other antibiotic-
resistance and virulence genes, and thus future studies are still needed.
It was also unknown if enterococci phage isolates harbored integrase-encoding genes. One
importance of this gene is that it enables one to determine if a phage is temperate and thus
amplification of this gene in Enterococcus-infecting bacteriophages could be used as a marker of
lysogeny [104]. This in turn represents an opportunity to focus on those phages harboring
integrases to further detect antibiotic-resistance and virulence-encoding genes. Another

33
importance of detecting integrases, although this hypothesis needs to be further tested, is that
integrases may be used to determine the presence of mobile elements in samples and therefore,
by determining the prevalence of microorganisms harboring integrases in the environment, future
studies could determine the possible increased risks to public health as a result of lateral gene
transfer in the environment.
Finally, the complete genomes of two Enterococcus phages are currently being determined: one
strictly lytic to E. faecalis strain ATCC 19433 and the other was induced from an environmental
enterococci isolate. Sequencing of these two phage types will enable us to describe the genomes
of novel Enterococcus phages and may open the opportunity to further develop the molecular
techniques for their detection in sewage and fecally contaminated waters.

34
LITERATURE CITED
1. Long, S.C. and M.D. Sobsey, A comparison of the survival of F+RNA and F+DNA
coliphages in lake water microcosms. J Water Health, 2004. 2(1): p. 15-22.
2. Groth, A.C. and M.P. Calos, Phage integrases: biology and applications. J Mol Biol,
2004. 335(3): p. 667-78.
3. Moe, C.L., Waterborne Transmission of Infectious Agents in Manual of Environmental
Microbiology, R.L. Crawford, Editor. 1996, ASM: Washington, D.C. p. 22-240.
4. Hurst, C.J., Overview of water microbiology as it relates to public health in Manual of
Environmental Microbiology, R.L. Crawford, Editor. 1996, ASM: Washington, D.C. p.
219-222.
5. Payment, P. and E. Franco, Clostridium perfringens and somatic coliphages as indicators
of the efficiency of drinking water treatment for viruses and protozoan cysts Applied and
Environmental Microbiology, 1993. 59(8): p. 2418-24.
6. Keswick, B.H., Detection of enteric viruses in treated drinking water Applied and
Environmental Microbiology, 1984. 47(6): p. 1290-4.
7. Bosch, A., Human enteric viruses in the water environment: a minireview. Int Microbiol,
1998. 1(3): p. 191-6.
8. Bosch, A., et al., Occurrence of enteroviruses in marine sediment along the coast of
Barcelona, Spain. Can J Microbiol, 1988. 34(7): p. 921-4.
9. Rao, V.C., et al., Isolation of enteroviruses from water, suspended solids, and sediments
from Galveston Bay: survival of poliovirus and rotavirus adsorbed to sediments. Appl
Environ Microbiol, 1984. 48(2): p. 404-9.
10. Rose, J.B., et al., Climate variability and change in the United States: potential impacts
on water- and foodborne diseases caused by microbiologic agents. Environ Health
Perspect, 2001. 109 Suppl 2: p. 211-21.
11. Lipp, E.K., et al., The Effects of Seasonal Variability and Weather on Microbial Fecal
Pollution and Enteric Pathogens in a Subtropical Estuary. Estuaries, 2001. 24(2): p. 266-
276.

35
12. Ashbolt, N.J., et al., Blooming E. coli, what do they mean?, in Coliforms and E. coli,
problem or solution? , D.K.a.C.F. (ed.), Editor. 1997, The Royal Society of Chemistry:
Cambridge, England. p. 78-85.
13. Toranzos, G.A., et al., Detection of microorganisms in environmental freshwaters and
drinking waters., in Manual of Environmental Microbiology, R.L. Crawford, Editor.
1996, ASM Press: Washington, D.C. p. 249-260.
14. Berg, G., et al., Validity of fecal coliforms, total coliforms, and fecal streptococci as
indicators of viruses in chlorinated primary sewage effluents. Appl Environ Microbiol,
1978. 36(6): p. 880-4.
15. Golomidova, A., et al., The diversity of coliphages and coliforms in horse feces reveals a
complex pattern of ecological interactions. Appl Environ Microbiol, 2007. 73(19): p.
5975-81.
16. Van Donsel, D.J., E.E. Geldreich, and N.A. Clarke, Seasonal Variations in Survival of
Indicator Bacteria in Soil and Their Contribution to Storm-water Pollution. Appl
Microbiol, 1967. 15(6): p. 1362-70.
17. Rivera, S.C., T.C. Hazen, and G.A. Toranzos, Isolation of fecal coliforms from pristine
sites in a tropical rain forest. Appl Environ Microbiol, 1988. 54(2): p. 513-517.
18. Byappanahalli, M.N., et al., Growth and survival of Escherichia coli and enterococci
populations in the macro-alga Cladophora (Chlorophyta). FEMS Microbiol Ecol, 2003.
46(2): p. 203-11.
19. Fong, T.T. and E.K. Lipp, Enteric viruses of humans and animals in aquatic
environments: health risks, detection, and potential water quality assessment tools.
Microbiol Mol Biol Rev, 2005. 69(2): p. 357-71.
20. Wheeler, A.L., et al., Potential of Enterococcus faecalis as a human fecal indicator for
microbial source tracking. J Environ Qual, 2002. 31(4): p. 1286-93.
21. USEPA, Method 1600: Membrane Filter Test Method for Enterococci in Water, in EPA-
821-R-02-022. 2002, Office of Water: Washington, D.C.
22. Martin, J.D. and J.O. Mundt, Enterococci in insects. Appl Microbiol, 1972. 24(4): p. 575-
80.
23. Baele, M., Composition of enterococcal and streptococcal flora from pigeon intestines
Applied and Environmental Microbiology, 2002. 92(2): p. 348-51.

36
24. Bonilla, N., et al., Enterophages, a group of phages infecting Enterococcus faecalis, and
their potential as alternate indicators of human faecal contamination. Water Sci Technol,
2010. 61(2): p. 293-300.
25. Ogilvie, L.A., et al., Comparative (meta)genomic analysis and ecological profiling of
human gut-specific bacteriophage phiB124-14. PLoS One, 2012. 7(4): p. e35053.
26. Hilton, M.C. and G. Stotzky, Use of coliphages as indicators of water pollution. Can J
Microbiol, 1973. 19(6): p. 747-51.
27. Santiago-Rodriguez, T.M., et al., Characterization of Enterococcus faecalis-infecting
phages (enterophages) as markers of human fecal pollution in recreational waters. Water
Res, 2010. 44(16): p. 4716-25.
28. Grabow, W.O. and P. Coubrough, Practical direct plaque assay for coliphages in 100-ml
samples of drinking water. Appl Environ Microbiol, 1986. 52(3): p. 430-3.
29. Simkova, A. and J. Cervenka, Coliphages as ecological indicators of enteroviruses in
various water systems. Bull World Health Organ, 1981. 59(4): p. 611-8.
30. Yates, M.V., C.P. Gerba, and L.M. Kelley, Virus persistence in groundwater. Appl
Environ Microbiol, 1985. 49(4): p. 778-81.
31. Payan, A., et al., Method for isolation of Bacteroides bacteriophage host strains suitable
for tracking sources of fecal pollution in water. Appl Environ Microbiol, 2005. 71(9): p.
5659-62.
32. Gantzer, C., J. Henny, and L. Schwartzbrod, Bacteroides fragilis and Escherichia coli
bacteriophages in human faeces. Int J Hyg Environ Health, 2002. 205(4): p. 325-8.
33. Jofre, J., et al., Potential usefulness of bacteriophages that infect Bacteroides fragilis as
model organisms for monitoring virus removal in drinking water treatment plants. Appl
Environ Microbiol, 1995. 61(9): p. 3227-31.
34. Ebdon, J.E., et al., Phages of Bacteroides (GB-124): a novel tool for viral waterborne
disease control? Environ Sci Technol, 2012. 46(2): p. 1163-9.
35. Gantzer, C., et al., Detection of infectious enteroviruses, enterovirus genomes, somatic
coliphages, and Bacteroides fragilis phages in treated wastewater. Appl Environ
Microbiol, 1998. 64(11): p. 4307-12.

37
36. Weisberg, S.B., R.T. Noble, and J.F. Griffith, Microbial Indicators of Marine
Recreational Water Quality in Manual of Environmental Microbiology, R.L. Crawford,
Editor. 1996, ASM: Washington, D.C. p. 280-287.
37. Havelaar, A.H. and W.M. Hogeboom, A method for the enumeration of male-specific
bacteriophages in sewage. J Appl Bacteriol, 1984. 56(3): p. 439-47.
38. Hernandez-Delgado, E.A., M.L. Sierra, and G.A. Toranzos, Coliphages as alternate
indicators of fecal contamination in tropical waters. Environ. Toxicol, 1991. 6: p. 131-
143.
39. Vinje, J., et al., Molecular detection and genotyping of male-specific coliphages by
reverse transcription-PCR and reverse line blot hybridization. Appl Environ Microbiol,
2004. 70(10): p. 5996-6004.
40. Furuse, K., et al., Bacteriophage distribution in human faeces: continuous survey of
healthy subjects and patients with internal and leukaemic diseases. J Gen Virol, 1983. 64
(Pt 9): p. 2039-43.
41. Cole, D., S.C. Long, and M.D. Sobsey, Evaluation of F+ RNA and DNA coliphages as
source-specific indicators of fecal contamination in surface waters. Appl Environ
Microbiol, 2003. 69(11): p. 6507-14.
42. Allwood, P.B., et al., Survival of F-specific RNA coliphage, feline calicivirus, and
Escherichia coli in water: a comparative study. Appl Environ Microbiol, 2003. 69(9): p.
5707-10.
43. Dorai-Raj, S., O.G. J, and E. Colleran, Specificity and sensitivity evaluation of novel and
existing Bacteroidales and Bifidobacteria-specific PCR assays on feces and sewage
samples and their application for microbial source tracking in Ireland. Water Res, 2009.
43(19): p. 4980-8.
44. Hundesa, A., et al., Development of a quantitative PCR assay for the quantitation of
bovine polyomavirus as a microbial source-tracking tool. J Virol Methods, 2010. 163(2):
p. 385-9.
45. Gomez-Donate, M., et al., New molecular quantitative PCR assay for detection of host-
specific bifidobacteriaceae suitable for microbial source tracking. Appl Environ
Microbiol, 2012. 78(16): p. 5788-95.

38
46. Tarkka, E., H. Ahman, and A. Siitonen, Ribotyping as an epidemiologic tool for
Escherichia coli. Epidemiol Infect, 1994. 112(2): p. 263-74.
47. Carson, C.A., et al., Identification of fecal Escherichia coli from humans and animals by
ribotyping. Appl Environ Microbiol, 2001. 67(4): p. 1503-7.
48. Nelson, M., et al., Characterization of Escherichia coli populations from gulls, landfill
trash, and wastewater using ribotyping. Dis Aquat Organ, 2008. 81(1): p. 53-63.
49. Furukawa, T., et al., Genotypic analysis of Enterococci isolated from fecal-polluted water
from different sources by pulsed-field gel electrophoresis (PFGE) for application to
microbial source tracking. Microbes Environ, 2011. 26(2): p. 181-3.
50. Esseili, M.A., Kassem, II, and V. Sigler, Optimization of DGGE community
fingerprinting for characterizing Escherichia coli communities associated with fecal
pollution. Water Res, 2008. 42(17): p. 4467-76.
51. Hayashi, H., et al., Molecular analysis of fecal microbiota in elderly individuals using
16S rDNA library and T-RFLP. Microbiol Immunol, 2003. 47(8): p. 557-70.
52. Savichtcheva, O. and S. Okabe, Qualitative and quantitative estimation of host-specific
fecal pollution using Bacteroides-Prevotella 16S rRNA genetic markers by T-RFLP and
real-time PCR analyses. Water Sci Technol, 2009. 59(9): p. 1831-40.
53. Bernhard, A.E. and K.G. Field, A PCR assay To discriminate human and ruminant feces
on the basis of host differences in Bacteroides-Prevotella genes encoding 16S rRNA.
Appl Environ Microbiol, 2000. 66(10): p. 4571-4.
54. Kildare, B.J., et al., 16S rRNA-based assays for quantitative detection of universal,
human-, cow-, and dog-specific fecal Bacteroidales: a Bayesian approach. Water Res,
2007. 41(16): p. 3701-15.
55. Byappanahalli, M.N., et al., Linking non-culturable (qPCR) and culturable enterococci
densities with hydrometeorological conditions. Sci Total Environ, 2010. 408(16): p.
3096-101.
56. Noble, R.T., et al., Comparison of rapid quantitative PCR-based and conventional
culture-based methods for enumeration of Enterococcus spp. and Escherichia coli in
recreational waters. Appl Environ Microbiol, 2010. 76(22): p. 7437-43.

39
57. McQuaig, S.M., et al., Quantification of human polyomaviruses JC Virus and BK Virus
by TaqMan quantitative PCR and comparison to other water quality indicators in water
and fecal samples. Appl Environ Microbiol, 2009. 75(11): p. 3379-88.
58. Polo, F., et al., Relationship between presence of Salmonella and indicators of faecal
pollution in aquatic habitats. FEMS Microbiol Lett, 1998. 160(2): p. 253-6.
59. Morinigo, M.A., et al., Viability of Salmonella spp and indicator microorganisms in
seawater using membrane diffusion chambers. Antonie Van Leeuwenhoek, 1990. 57(2):
p. 109-17.
60. Savichtcheva, O. and S. Okabe, Alternative indicators of fecal pollution: relations with
pathogens and conventional indicators, current methodologies for direct pathogen
monitoring and future application perspectives. Water Res, 2006. 40(13): p. 2463-76.
61. Ottoson, J. and T.A. Stenstrom, Faecal contamination of greywater and associated
microbial risks. Water Res, 2003. 37(3): p. 645-55.
62. Hellein, K.N., et al., Culture-based indicators of fecal contamination and molecular
microbial indicators rarely correlate with Campylobacter spp. in recreational waters. J
Water Health, 2011. 9(4): p. 695-707.
63. Wu, J., et al., Are microbial indicators and pathogens correlated? A statistical analysis of
40 years of research. J Water Health, 2011. 9(2): p. 265-78.
64. Payment, P. and A. Locas, Pathogens in water: value and limits of correlation with
microbial indicators. Ground Water, 2011. 49(1): p. 4-11.
65. Pruss, A., Review of epidemiological studies on health effects from exposure to
recreational water. Int J Epidemiol, 1998. 27(1): p. 1-9.
66. Mujeriego, R., J.M. Bravo, and M.T. Feliu, Recreation in Coastal Waters, Public Health
Implications. VIemes Joumies Etud Pollutions Cannes CIESM, 1982: p. 585-594.
67. Fleisher, J.M., et al., The BEACHES Study: health effects and exposures from non-point
source microbial contaminants in subtropical recreational marine waters. Int J
Epidemiol, 2010. 39(5): p. 1291-8.
68. Novais, C., et al., Local genetic patterns within a vancomycin-resistant Enterococcus
faecalis clone isolated in three hospitals in Portugal. Antimicrob Agents Chemother,
2004. 48(9): p. 3613-7.

40
69. Whitman, R.L., et al., Incidence of the enterococcal surface protein (esp) gene in human
and animal fecal sources. Environ Sci Technol, 2007. 41(17): p. 6090-5.
70. Schwartz, T., et al., Detection of antibiotic-resistant bacteria and their resistance genes
in wastewater, surface water, and drinking water biofilms. FEMS Microbiol Ecol, 2003.
43(3): p. 325-35.
71. USEPA, Microbial Source Tracking Guide Document. EPA/600-R-05-064 2005:
Cincinnati, OH.
72. Kruse, H. and H. Sorum, Transfer of multiple drug resistance plasmids between bacteria
of diverse origins in natural microenvironments. Appl Environ Microbiol, 1994. 60(11):
p. 4015-21.
73. Ohlsen, K., et al., Impact of antibiotics on conjugational resistance gene transfer in
Staphylococcus aureus in sewage. Environ Microbiol, 2003. 5(8): p. 711-6.
74. Salyers, A.A., et al., Conjugative transposons: an unusual and diverse set of integrated
gene transfer elements. Microbiol Rev, 1995. 59(4): p. 579-90.
75. Roberts, M.C., Update on acquired tetracycline resistance genes. FEMS Microbiol Lett,
2005. 245(2): p. 195-203.
76. Chopra, I. and M. Roberts, Tetracycline antibiotics: mode of action, applications,
molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev,
2001. 65(2): p. 232-60 ; second page, table of contents.
77. Roberts, M.C., Tetracycline therapy: update. Clin Infect Dis, 2003. 36(4): p. 462-7.
78. Roberts, M.C., Antibiotic resistance mechanisms in bacteria of oral and upper
respiratory origin. Int J Antimicrob Agents, 1998. 9(4): p. 255-67.
79. Poyart-Salmeron, C., et al., Genetic basis of tetracycline resistance in clinical isolates of
Listeria monocytogenes. Antimicrob Agents Chemother, 1992. 36(2): p. 463-6.
80. Chee-Sanford, J.C., et al., Occurrence and diversity of tetracycline resistance genes in
lagoons and groundwater underlying two swine production facilities. Appl Environ
Microbiol, 2001. 67(4): p. 1494-502.
81. Speer, B.S., N.B. Shoemaker, and A.A. Salyers, Bacterial resistance to tetracycline:
mechanisms, transfer, and clinical significance. Clin Microbiol Rev, 1992. 5(4): p. 387-
99.

41
82. Agerso, Y., A.G. Pedersen, and F.M. Aarestrup, Identification of Tn5397-like and Tn916-
like transposons and diversity of the tetracycline resistance gene tet(M) in enterococci
from humans, pigs and poultry. J Antimicrob Chemother, 2006. 57(5): p. 832-9.
83. Charpentier, E., G. Gerbaud, and P. Courvalin, Presence of the Listeria tetracycline
resistance gene tet(S) in Enterococcus faecalis. Antimicrob Agents Chemother, 1994.
38(10): p. 2330-5.
84. Villedieu, A., et al., Prevalence of tetracycline resistance genes in oral bacteria.
Antimicrob Agents Chemother, 2003. 47(3): p. 878-82.
85. Auerbach, E.A., E.E. Seyfried, and K.D. McMahon, Tetracycline resistance genes in
activated sludge wastewater treatment plants. Water Res, 2007. 41(5): p. 1143-51.
86. Courvalin, P., Transfer of antibiotic resistance genes between gram-positive and gram-
negative bacteria. Antimicrob Agents Chemother, 1994. 38(7): p. 1447-51.
87. Ochman, H., J.G. Lawrence, and E.A. Groisman, Lateral gene transfer and the nature of
bacterial innovation. Nature, 2000. 405(6784): p. 299-304.
88. Hinnebusch, B.J., et al., High-frequency conjugative transfer of antibiotic resistance
genes to Yersinia pestis in the flea midgut. Mol Microbiol, 2002. 46(2): p. 349-54.
89. Cetinkaya, Y., P. Falk, and C.G. Mayhall, Vancomycin-resistant enterococci. Clin
Microbiol Rev, 2000. 13(4): p. 686-707.
90. Roberts, M.C., et al., Vancomycin-resistant Enterococcus spp. in marine environments
from the West Coast of the USA. J Appl Microbiol, 2009. 107(1): p. 300-7.
91. Torres, C., et al., vanA-mediated vancomycin-resistant Enterococcus spp. in sewage. J
Antimicrob Chemother, 1994. 33(3): p. 553-61.
92. Jett, B.D., M.M. Huycke, and M.S. Gilmore, Virulence of enterococci. Clin Microbiol
Rev, 1994. 7(4): p. 462-78.
93. Shankar, N., et al., Role of Enterococcus faecalis surface protein Esp in the pathogenesis
of ascending urinary tract infection. Infect Immun, 2001. 69(7): p. 4366-72.
94. Toledo-Arana, A., et al., The enterococcal surface protein, Esp, is involved in
Enterococcus faecalis biofilm formation. Appl Environ Microbiol, 2001. 67(10): p. 4538-
45.

42
95. Tendolkar, P.M., A.S. Baghdayan, and N. Shankar, The N-terminal domain of
enterococcal surface protein, Esp, is sufficient for Esp-mediated biofilm enhancement in
Enterococcus faecalis. J Bacteriol, 2005. 187(17): p. 6213-22.
96. Hammerum, A.M. and L.B. Jensen, Prevalence of esp, encoding the enterococcal surface
protein, in Enterococcus faecalis and Enterococcus faecium isolates from hospital
patients, poultry, and pigs in Denmark. J Clin Microbiol, 2002. 40(11): p. 4396.
97. Scott, T.M., et al., Potential use of a host associated molecular marker in Enterococcus
faecium as an index of human fecal pollution. Environ Sci Technol, 2005. 39(1): p. 283-
7.
98. Ahmed, W., et al., Evaluation of the host-specificity and prevalence of enterococci
surface protein (esp) marker in sewage and its application for sourcing human fecal
pollution. J Environ Qual, 2008. 37(4): p. 1583-8.
99. Byappanahalli, M.N., et al., Environmental occurrence of the enterococcal surface
protein (esp) gene is an unreliable indicator of human fecal contamination. Environ Sci
Technol, 2008. 42(21): p. 8014-20.
100. Lund, B., H. Billstrom, and C. Edlund, Increased conjugation frequencies in clinical
Enterococcus faecium strains harbouring the enterococcal surface protein gene esp. Clin
Microbiol Infect, 2006. 12(6): p. 588-91.
101. Canchaya, C., et al., Phage as agents of lateral gene transfer. Curr Opin Microbiol, 2003.
6(4): p. 417-24.
102. Brussow, H., C. Canchaya, and W.D. Hardt, Phages and the evolution of bacterial
pathogens: from genomic rearrangements to lysogenic conversion. Microbiol Mol Biol
Rev, 2004. 68(3): p. 560-602.
103. Brussow, H. and E. Kutter, Genomics and evolution of tailed phages, in Bacteriophages:
Biology and Applications, E. Kutter and A. Sulakvelidzae, Editors. 2005, CRC Press:
Washington, D.C. p. 91-129.
104. Yasmin, A., et al., Comparative genomics and transduction potential of Enterococcus
faecalis temperate bacteriophages. J Bacteriol, 2010. 192(4): p. 1122-30.
105. Hendrix, R.W., Bacteriophage genomics. Curr Opin Microbiol, 2003. 6(5): p. 506-11.

43
106. Drabble, W.T. and B.A. Stocker, R (transmissible drug-resistance) factors in Salmonella
typhimurium: pattern of transduction by phage P22 and ultraviolet-protection effect. J
Gen Microbiol, 1968. 53(1): p. 109-23.
107. Colomer-Lluch, M., J. Jofre, and M. Muniesa, Antibiotic resistance genes in the
bacteriophage DNA fraction of environmental samples. PLoS One, 2011. 6(3): p. e17549.
108. Muniesa, M., et al., Shiga toxin 2-converting bacteriophages associated with clonal
variability in Escherichia coli O157:H7 strains of human origin isolated from a single
outbreak. Infect Immun, 2003. 71(8): p. 4554-62.
109. Muniesa, M. and J. Jofre, Abundance in sewage of bacteriophages that infect Escherichia
coli O157:H7 and that carry the Shiga toxin 2 gene. Appl Environ Microbiol, 1998.
64(7): p. 2443-8.
110. Muniesa, M., et al., Characterization of a shiga toxin 2e-converting bacteriophage from
an Escherichia coli strain of human origin. Infect Immun, 2000. 68(9): p. 4850-5.

44
CHAPTER 2
Enterophages, a group of phages infecting Enterococcus faecalis and their potential as
alternate indicators of human fecal contamination
N. Bonilla1, T. M. Santiago-Rodriguez
1, P.
Marcos
2, M. Urdaneta
1, J. W. Santo Domingo
3 and G.
A. Toranzos1
1Environmental Microbiology Laboratory, Department of Biology, University of Puerto Rico, P.O.
Box 23360, San Juan, PR 00931-3360, 2Biological Sciences Department, General Studies,
University of Puerto Rico, San Juan, PR 00931, 3U.S. Environmental Protection Agency,
Cincinnati, Ohio
ABSTRACT
We have developed a method for the detection of viruses in environmental samples that we have
called enterophages, which specifically infect Enterococcus faecalis. This method has allowed us
to determine the prevalence and to study the ecology of this group of phages. The enterophages
replicate at 35ºC, and at 41ºC. The presence of NaN3 in the media inhibits the growth of
background microbiota and allow for an accurate, specific and rapid detection of these viruses.
Enterophages were present in raw domestic sewage at lower concentrations (average 1.8 x 102
PFU/100ml) than those of coliphages (average 1.7 x 105
PFU/100ml). Phages were characterized
by transmission electron microscopy showing icosahedral capsids, some with non-contractile
tails as well as icosahedral non-tailed capsids. Different isolates had capsid sizes ranging from 20
nm to about 75 nm in diameter. These data describe a new group of phages that may serve as
alternate indicators of human fecal pollution, especially in recreational waters. The ecology of
these enterophages indicates that these may be strictly of human origin.
Keywords: enterococci; enterophages; faecal contamination; indicators; recreational waters
Reference: Bonilla, N., et al., Enterophages, a group of phages infecting Enterococcus faecalis,
and their potential as alternate indicators of human faecal contamination. Water Sci Technol,
2010. 61(2): p. 293-300.

45
INTRODUCTION
Pathogenic microorganisms are present in recreational waters and other water sources as a result
of the presence of feces of warm-blooded animal. Some examples of these pathogens are
Salmonella, Shigella, Campylobacter spp., as well as several different types of enteric viruses
and pathogenic protozoa. The low infectious dose (as low as 1 to 50 in some cases) of many of
these pathogens is a cause for concern if they are released into the water environment, since
humans who are in contact with these contaminated waters, are bound to ingest the contaminated
water. Human rotaviruses, astroviruses and noroviruses viruses may be found in sewage-
contaminated waters and in fact, among the waterborne pathogens, these are of greatest concern,
since they may be more resistant to environmental conditions than pathogenic bacteria and they
may be more resistant to chlorination and UV disinfection [1], and have perhaps the lowest
infectious dose of any of the waterborne pathogens.
Indicator organisms have been used for the last century as surrogates for the presence of
pathogenic microorganisms in waters. In the past, bacterial indicators such as the thermotolerant
coliform group, Escherichia coli or the Enterococcus group to determine the microbiological
safety of waters have been tremendously useful. The levels of these indicators are monitored in
treated drinking waters as well as recreational waters and the concentrations at which they are
found enables (e.g.) beach managers in making decisions that involve closing beaches or keeping
them open to the public.
Escherichia coli and Enterococcus spp. have been used as indicators of the presence of
pathogenic microorganisms in different types of water. High concentrations of these indicators
have previously been found to correlate with risk to bathers; concentrations of E. coli have been
correlated with risk to bathers in freshwater beaches [2, 3], whereas concentrations of
Enterococcus spp. have been correlated with risk in marine beaches. As such, they are now
considered the most useful indicators of risk to bathers as a result of primary contact exposure to
bathing waters. However, bacterial indicators (E. coli and Enterococcus spp.) have been isolated
from pristine sites in a tropical rain forest [4] and are routinely detected as part of the
environmental microbiota in Puerto Rico and other tropical areas. Fecal indicators such as E. coli
and enterococci may also be able to multiply and become a significant environmental source in

46
tropical regions such as Hawaii, Guam, Puerto Rico and South Florida [5]. Thus their role as
microbial indicators of fecal contamination is not entirely clear and alternative indicators of
waterborne enteric viruses are needed.
Alternate microbial indicators of fecal water contamination
Thermotolerant and total coliforms tend to be poor indicator microorganisms because of their
short survival and susceptibility to water treatments [1]. Moreover, these microbial indicators
have the ability to grow in natural waters and there is a lack of correlation between the number of
coliforms and that of infectious microorganisms [6]. Also, pathogenic viruses have been found in
waters where the number of coliforms had not exceeded the standards [7].
Bacteriophages, especially coliphages, have also been used as model viruses for water quality
control [8]. Coliphages have been proposed as indicators of fecal water contamination [9-12], as
have those phages that infect Bacteroides fragilis [13], since many have similar morphological
characteristics to the enteric viruses as well as similar survival times in aquatic environments.
However, their usefulness has been questioned, since the group is made up of different viruses
belonging to several different families, each having different ecological and biological
characteristics. A more specific group seems to be the male-specific RNA coliphages, but their
usefulness is still being debated.
Out of all methods available for the determination of the microbiological water quality, the
bacteriophage methods take the least amount of time, and are the most reliable methods for the
detection of the target since they may not require confirmation procedures. Additionally, they
may be amongst the least costly methods once they are implemented in an environmental
monitoring laboratory.
A useful bacterial indicator is Enterococcus faecalis, because of its apparent inability to multiply
in sewage contaminated waters and their survival time in the environment seems to be greater
than E. coli [14]. E. faecalis shows a closer correlation with risk to bathers in point-source
impacted recreational waters [2]; however, we know very little about the ecology of this group of

47
microorganisms. Although E. faecalis has been described as having a limited host range, the
genus may be more cosmopolitan than previously thought. The usual hosts include humans,
chickens and dogs [3], but they are also present in the intestines of other animals, such as
pigeons [15] and have also been found as part of the microbiota of insects, including flies [16-
18]. In Puerto Rico we have detected Enterococcus spp. as part of the microbiota in sand flies,
and even in laboratory strains of Drosophila spp. (in preparation).
The detection of pathogenic microorganisms or reliable indicator microorganisms is important
for the determination of the microbiological quality of recreational waters. However, currently
used detection methods have several disadvantages: they can be expensive or the detection time
can take too long, taking on the average 24 to 48 hours before results can be obtained. This is
certainly not the best manner to manage possible risk in recreational waters. Additionally,
bacterial indicators are not reliable indicators of the possible presence of enteric viruses. We
need a method that will give reliable results in less than 12 to 18 hours, and the only non-
molecular method that seems to lend itself for this time interval is one that detects infectious
bacteriophages.
Molecular methods are fast and reliable, but dead bacteria, naked DNA or non-infectious viruses
may be detected, so the question of viability (in the case of bacteria) and infectivity (in the case
of viruses) come into play. Thus, the detection of infectious bacteriophages, when using the
plaque assay, if they can be correlated to risk, would perhaps be the most appropriate manner of
managing risk in recreational waters.
In the present paper we describe a method we developed for the detection of a group of phages
that infect specifically E. faecalis. Our goal is to eventually determine if this group of viruses can
be used as a reliable alternative indicator of risk to bathers as a result of the presence of human
pathogens in recreational waters.
MATERIALS AND METHODS
Host strains

48
Several isolates were obtained from raw domestic sewage from several sewage treatment plants
in Puerto Rico. These were obtained using membrane filtration using mEnterococcus media
prepared as indicated by the manufacturer. Isolated colonies were purified and isolated in pure
culture using the same media. Over 100 individual isolates were tested using the single and
double layer methods [19] for the detection of phages. Additionally, several Enterococcus spp.
type strains were tested as possible hosts, these were: E. pseudoavium, E. faecium, E. durans, E.
casseli, E. hirae, E. dispar, E. gallinarum and E. faecalis.
Host bacteria
Once the best host was determined to be E. faecalis, it was kept streaked for isolation in
mEnterococcus agar plates and incubated overnight at 35 ºC. The Petri dishes were kept at 4-7 ºC
for up to one week. To prepare the host bacteria for plaque assays, individual colonies were
picked and inoculated into sterile (Difco) Dextrose Azide Broth medium and incubated overnight
at 35 ºC. Five-mL of this inoculated medium was used for every 50 mL of sample added to
50mL of 1X regular-strength (Difco) Tryptic Soy Broth (TSB).
Samples
Sewage and recreational water samples were collected in sterile plastic bottles. Samples were
shipped to the laboratory at 2 to 8ºC using cold packs and stored at 4 ºC. Additionally, bird, and
several mammal fresh fecal samples were tested for the presence of enterophages.
Optimal conditions for viral replication
Several concentrations of calcium chloride (CaCl2·2H20 Fisher Scientific Co., NJ, USA) and
sodium azide (NaN3 MCB, OH) were tested, using as the base those concentrations used by other
authors [19] for the detection of coliphages (in the case of calcium chloride) and commercially
available mEnterococcus medium (in the case of sodium azide).
To determine the optimal concentration of medium, several media were tested, such as: Nutrient
Broth, Azide Dextrose and mEnterocuccus, at different concentrations. Similarly, 2X
concentration of TSB and 1.5 % agar was tested as was 1X TSB containing different
concentrations of agar. Agar concentrations ranging from 0.375 to 1.5 % were tested. Phage

49
plates were incubated at 22, 37, 41 and 45°C. Viral plaques were counted at 6, 18, 24 and 48
hours.
Single Layer Plaque Assays
Once optimal conditions were determined for enterophage plaque formation, a 50 mL volume of
1X TSB was prepared and agar added to a final concentration of 0.375 or 0. 75 % where mixed
with an equal volume of the sample. The medium was autoclaved and kept at 50 ºC. To a 50
mL-volume of sample, CaCl2 was added to a final concentration of 5.2 mg/mL, as was NaN3 to a
final concentration of 0.4 mg/mL plus 5ml of a freshly grown culture (overnight culture) of E.
faecalis. Solution was mixed carefully with the liquefied 50 mL volume of 1X TSB, and poured
into four sterile 100-mm diameter Petri dishes. After the agar solidified, the Petri dishes were
incubated at 22, 35 or 41 °C, in order to determine if different phage populations that replicate at
different temperatures are present. Viral plaques were counted after 3, 8, 24 and 48 hours of
incubation.
Plaque isolation
Discrete viral plaques were plucked using standard procedures. Briefly, sterile glass Pasteur
pipettes were used to obtain a plug which was then placed in an Eppendorf tube containing 500
μL of sterile 1X Phosphate Buffer Solution (PBS). Many of these phages are tailed and rough
treatment may break the tails rendering the viruses non-infective. The plug was gently dislodged
and broken by pipetting up and down. The tubes were centrifuged at 14,000 rpm for 10 min at 10
ºC. The supernatant was transferred into a sterile Eppendorf tube, and the supernatant tittered
using serial dilutions using the double layer method using the same media as above. Briefly, the
dilutions were mixed with 4 mL of 1X TSB containing the same concentrations of CaCl2 and
NaN3 as above and 0.75 % agar and then poured into Petri dishes containing 20 mL of bottom
agar media containing CaCl2 and NaN3 and 1.5 % agar. Plates were incubated at 35ºC and 5mL
of PBS or physiological saline (0.85 % NaCl) was added to those plates showing complete viral
lysis and slowly agitated rotationally for 10 min. The top agar was transferred to a sterile
Oakridge tube, pipetted up and down to break down the agar, and then centrifuged at 14,000 rpm
for 10 min at 7 ºC. The supernatant containing the viruses were kept in a sterile tube at 4-7 °C.

50
Transmission Electron Microscopy
The resulting supernatant from above was loaded into dialysis tubes (12,000-14,000 MW cutoff,
Spectrapor, Los Angeles, CA, USA) for hydroextraction and covered with crystalline
polyethylene glycol (PEG, Mol.wt. 8,000,Sigma Chem. Co. MO; APHA, 1989) clamped and
placed at 4-7 °C overnight. The hydroextracted solution was recovered, the inside of the dialysis
tubing washed with 100 μL of sterile 0.85 % NaCl transferred into a sterile Eppendorf tube and
kept at 4 °C till electron-microscopic analyses.
An aliquot of the concentrated portion was placed on carbon Type-B 200 mesh copper grids or
ultra thin carbon film/holey carbon 400 mesh copper grids and stained with uranyl acetate (UA)
2%, pH 4.5 or potassium phosphotungstate (PTA) 2%, pH 7.2. Alternatively, 30 μL of each
phage concentrate was placed on parafilm and the grids floated on top for 15 minutes at room
temperature prior to staining. All specimens were examined using a Karl Zeiss Leo 922 energy
filtered transmission electron microscope operated at 200 KV. At least 5 phage particles of each
type observed were measured. The phages were measured directly on the images, which had the
magnifications previously calibrated. Capsid sizes were measured between opposite apices.
Determination of Burst Sizes
To determine the burst sizes of each of phage isolate, E. faecalis was grown to mid-log phase in
Azide Dextrose Broth and the concentration determined. One-mL of the proper enterophage
dilution was added in order to have a Multiplicity of Infection (MOI) of 1.0. The phage was
allowed to come in contact with the bacteria by incubating at 35 °C for 5 minutes. The solution
was then centrifuged at 14,000rpm at 10°C for 10 minutes. The supernatant was eliminated, and
10 mL of Azide Dextrose were added to the tube and the pellet resuspended to the original
volume. Aliquots were obtained every 15 min for 3 hours, and the aliquots serially diluted with
1X TSB containing the previously indicated concentrations of NaN3 and CaCl2. A volume of the
resulting dilution was transferred into 4 mL of top agar (0.375 %) and poured into agar plates
containing the bottom agar described above. The plates were incubated at 35 °C for 24 hours.
Plaques were counted and burst sizes determined using the point in the graph where the values
remained constant.

51
Survival
A sewage sample containing about 103 enterophages/100ml was inoculated into 1L of unsterile
sea water. Samples were incubated at 23, 35 and 41 ºC and aliquots obtained twice of
thrice/week and analyzed for the presence of enterophages using the single-layer method as
described above.
Prevalence
Grab samples were obtained from the influent sites at several different domestic sewage
treatment plants in Puerto Rico. Samples were kept refrigerated till processing. Fifty-mL
volumes were analyzed as described above using the single-layer method.
RESULTS AND DISCUSSION
Sewage and surface water grab samples were originally analyzed using commercially available
enterococci media (mEnterococcus, mE, mEI, Azide Dextrose), which did not result in any
visible plaques using either the single or double layer method. This was surprising, since the
media mention are standard for the isolation and detection of Enterococcus spp., but none were
useful for the detection of viral plaques (data not shown). We also tested different concentrations
of the TSB, and though we got some plaques, the growth of the host was not very conducive to
the detection of viral plaques. When testing different concentrations of the media, we started out
with 2X concentrations, since that is the standard procedure for the detection of coliphages;
however, we were surprised to see that E. faecalis seems to be susceptible to the osmotic
pressures created by even the final 1X resulting concentration after mixing with an equal volume
of the sample. This phenomenon was even more obvious when testing marine waters, possibly as
a result of the increased osmotic pressure; this opens up other questions as to the survivability of
Enterococcus spp. in marine waters. Thus we decided to use a final concentration of 0.5 % of
TSB. Though viral plaques were visible when using only 0.5 % TSB under laboratory
conditions, when processing environmental samples, it was impossible to see any plaques in the
absence of NaN3. Of several concentrations of CaCl2, the optimal concentration was shown to be
5.2 mg/mL and the optimal concentration of sodium azide 0.4 mg/mL. Viral plaques were visible
at 0.75 % agar at 37 °C, but somehow clearer at 0.375 % agar at 41 °C (data not shown).

52
We tested fecal samples from several different animals, including birds, dogs and cats as well as
sand flies for the presence of enterophages and none were positive; though all of them were
positive for the presence of Enterococcus spp. (data not shown)
Viral Morphology
The morphology of the virions (Figure 1) show tailed phages and they are different from the
round capsid morphology observed by others [20] in enterophages isolated from human saliva;
though we also detected some isolates that were non-tailed, with icosahedral capsids about 20-25
nm in diameter (data not shown). Our best studied phage isolate to date showed an icosahedral
75 nm diameter capsid with a long non-contractile tail measuring 240 nm long. This morphology
is similar to the classical Bradley’s basic morphology Group B belonging to the Siphoviridae
family [21]. Further studies are needed to characterize other groups of enterophage isolates. It
should be noted that viruses infecting E. faecalis have been described previously, but not those
from environmental samples. Previous isolation and use of these phages was for typing of E.
faecalis clinical isolates [22] going back several decades. The fact that enterophages are also
found in human saliva [20] may indicate that they are in fact unique to humans, and as humans
usually spit when they immerse their heads into the water while swimming, the concentrations of
these enterophages may show to be a good indicator of risk to bathers.
(A) (B)
Figure 1: Enterophage (A) with an icosahedral head of 13 nm and a 56 nm non-contractile tail
(Bar = 20 nm). Enterophage group (B) with an icosahedral head of 75 nm with a non- contractile
240 nm length (Bar = 200 nm).

53
Burst Sizes
When viruses successfully infect cells, results in a certain number of progeny virions called the
burst size. It would seem apparent that the phages most likely to be found in sewage and fecally-
contaminated waters are those that have a high burst size. Our data indicated that the burst size
varies depending on the isolate and the numbers go from 102 to 10
5 virions/cell (data not shown).
We will focus our future characterization efforts on those enterophages that have the highest
burst size, since they may be more likely to be detected in low volumes of water samples.
Survival
Any indicator of risk should be at least as reliable as those currently in use and should meet
several criteria, and among these the ability to survive for at least as long as the pathogens under
the environmental conditions is a must. Our data (Figure 2) show that there may be different
populations of enterophages present in raw sewage, as indicated by the different numbers
obtained in these experiments in plates incubated at three different temperatures. However, all
three populations survived similarly and their numbers decreased by over two log10 over a
period of 7 days, which is similar to the survival times of enteric viruses as reported by others
[23]. Figure 2 (A) shows that although the enterophages survived better at ambient temperature
where they showed about a 0.5 log10 drop after 3 days, whereas there was at least 1-1.5 log10 drop
in concentrations at 37 ºC after the same length of time. It should be noted that there was no
replication of the enterophages at ambient temperature in spite of having native E. faecalis
present in the sewage inoculum which would have been shown by an increase in the number of
viruses. It is also noteworthy to see different groups of enterophages capable of replicating at
different temperatures (namely 22, 37 and 41 ºC). These data will guide future studies as to the
types of enterophages to focus on as the possible best indicators of human fecal contamination.
We are in the process of carrying out similar survival experiments in marine waters, freshwaters,
beach sand and soils in the presence of the E. faecalis host we have been using.

54
0
0.5
1
1.5
2
2.5
0 2 4 6 8 10 12
Nu
mb
er o
f p
ha
ges
/10
0m
L (
Lo
g)
Time (Days)
22°C
37°C
41 C
(A)
(B)
Figure 2: Survival of enterophages at 22 °C (A) and at 37 °C (B). Aliquots were tested and Petri
dishes were incubated at the three different temperatures shown.
Prevalence
When measuring coliphages and enterophages in treated and untreated sewage from Puerto Rico
at concentrations ranging from 1.4 x 105 to 2.6 x 10
5 and from 55 to 363 Plaque Forming Units
(PFU)/100ml for coliphages and enterophages, respectively (Tables 1 and 2). It is also
noteworthy that the enterophage plaques could be easily detected even in raw sewage samples
after 8 hours. Coliphage analyses of the same samples showed luxuriant growth of background
microbiota which masked the viral plaques; even in the presence of antibiotics. These data also
showed that sewage treatment plants are able of removing/inactivating from 87 to >99.9 % of
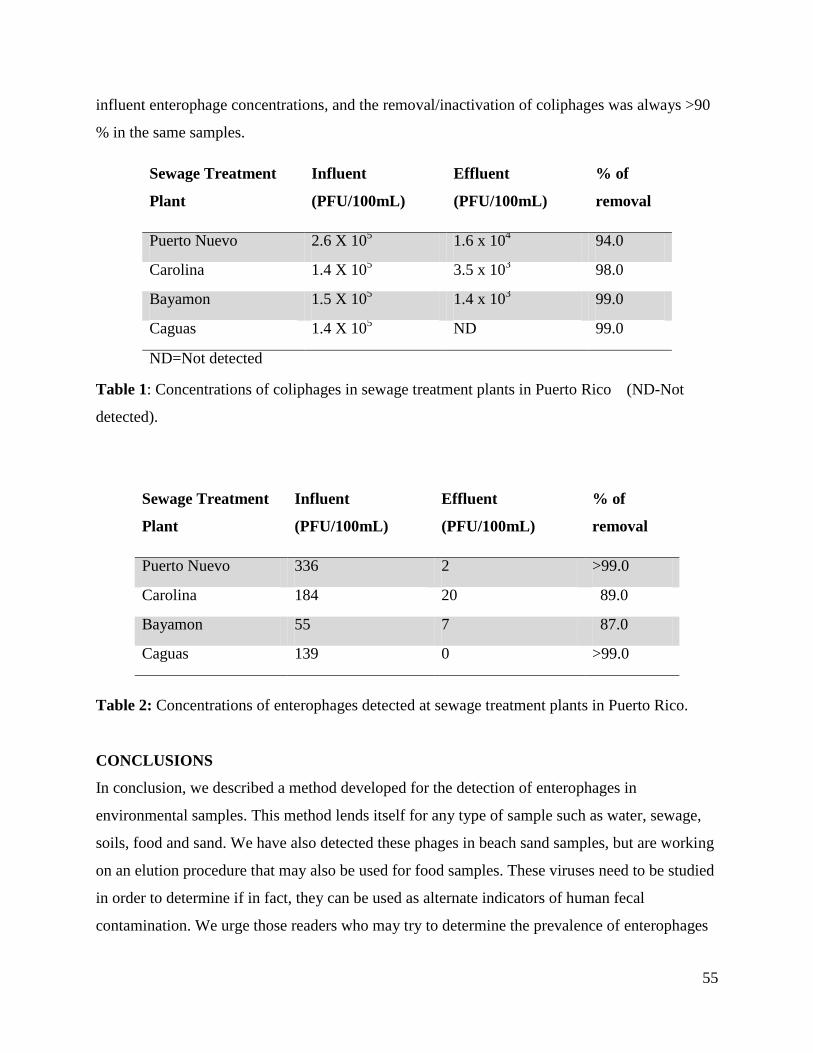
55
influent enterophage concentrations, and the removal/inactivation of coliphages was always >90
% in the same samples.
Sewage Treatment
Plant
Influent
(PFU/100mL)
Effluent
(PFU/100mL)
% of
removal
Puerto Nuevo 2.6 X 105 1.6 x 10
4 94.0
Carolina 1.4 X 105 3.5 x 10
3 98.0
Bayamon 1.5 X 105 1.4 x 10
3 99.0
Caguas 1.4 X 105 ND 99.0
ND=Not detected
Table 1: Concentrations of coliphages in sewage treatment plants in Puerto Rico (ND-Not
detected).
Sewage Treatment
Plant
Influent
(PFU/100mL)
Effluent
(PFU/100mL)
% of
removal
Puerto Nuevo 336 2 >99.0
Carolina 184 20 89.0
Bayamon 55 7 87.0
Caguas 139 0 >99.0
Table 2: Concentrations of enterophages detected at sewage treatment plants in Puerto Rico.
CONCLUSIONS
In conclusion, we described a method developed for the detection of enterophages in
environmental samples. This method lends itself for any type of sample such as water, sewage,
soils, food and sand. We have also detected these phages in beach sand samples, but are working
on an elution procedure that may also be used for food samples. These viruses need to be studied
in order to determine if in fact, they can be used as alternate indicators of human fecal
contamination. We urge those readers who may try to determine the prevalence of enterophages

56
under their own conditions to try to use the same procedures as well as the same host in order to
be able to compare results in the future. One of the problems we have encountered with
coliphages is that many different hosts were used over the years, making it difficult to determine
if in fact the same coliphages are being detected. We hope to avoid this problem with this group
of viruses. We have started an international collaboration with colleagues from several different
geographical areas encompassing several continents, to determine the prevalence of
enterophages in raw sewage and recreational waters. This will let us know if these phages can be
used as alternate indicators of human fecal contamination in waters and other media at a global
scale. Further work also needs to be done to further characterize enterophages not only
morphologically, but also molecularly before they are taken into consideration as indicators of
human fecal contamination.
ACKNOWLEDGMENTS
We thank Ing. Capeles and Ms. Cruz Minerva Ortiz from PRASA for their help with access to
sewage treatment plant samples. We also acknowledge the help of Margerie Rivera, Nataly
Montes, Veronica Marcantoni, and Leiribel Tavarez for their help in the analyses of the samples.
We also thank Camilo and French guy for their help with the TEM analyses.

57
LITERATURE CITED
1. Moe, C.L., Waterborne Transmission of Infectious Agents in Manual of Environmental
Microbiology, R.L. Crawford, Editor. 1996, ASM: Washington, D.C. p. 22-240.
2. Dufour, A.P., Bacterial indicators of recreational water quality. Can J Public Health,
1984. 75(1): p. 49-56.
3. Wheeler, A.L., et al., Potential of Enterococcus faecalis as a human fecal indicator for
microbial source tracking. J Environ Qual, 2002. 31(4): p. 1286-93.
4. Rivera, S.C., T.C. Hazen, and G.A. Toranzos, Isolation of fecal coliforms from pristine
sites in a tropical rain forest. Appl Environ Microbiol, 1988. 54(2): p. 513-517.
5. Byappanahalli, M. and R. Fujioka, Indigenous soil bacteria and low moisture may limit
but allow faecal bacteria to multiply and become a minor population in tropical soils.
Water Sci Technol, 2004. 50(1): p. 27-32.
6. Toranzos, G.A., et al., Detection of microorganisms in environmental freshwaters and
drinking waters., in Manual of Environmental Microbiology, R.L. Crawford, Editor.
1996, ASM Press: Washington, D.C. p. 249-260.
7. Fong, T.T. and E.K. Lipp, Enteric viruses of humans and animals in aquatic
environments: health risks, detection, and potential water quality assessment tools.
Microbiol Mol Biol Rev, 2005. 69(2): p. 357-71.
8. Stetler, R.E., Coliphages as indicators of enteroviruses Appl Environ Microbiol, 1984.
48(3): p. 668-670.
9. Havelaar, A.H. and W.M. Hogeboom, A method for the enumeration of male-specific
bacteriophages in sewage. J Appl Bacteriol, 1984. 56(3): p. 439-47.
10. Loh, C.L., et al., Development of a field test kit for coliphage detection in natural waters
Tox. Assess, 1988. 5: p. 451-460.

58
11. Hernandez-Delgado, E.A., M.L. Sierra, and G.A. Toranzos, Coliphages as alternate
indicators of fecal contamination in tropical waters. Environ. Toxicol, 1991. 6: p. 131-
143.
12. Moce-Llivina, L., F. Lucena, and J. Jofre, Enteroviruses and bacteriophages in bathing
waters. Appl Environ Microbiol, 2005. 71(11): p. 6838-44.
13. Tartera, C. and J. Jofre, Bacteriophages active against Bacteroides fragilis in sewage-
polluted waters. Appl Environ Microbiol, 1987. 53(7): p. 1632-7.
14. Muñiz, I., et al., Survival and activity of Streptococcus faecalis and Escherichia coli in
tropical fresh water. Microbial Ecology, 1989. 18: p. 125-134.
15. Baele, M., Composition of enterococcal and streptococcal flora from pigeon intestines Applied
and Environmental Microbiology, 2002. 92(2): p. 348-51.
16. Martin, J.D. and J.O. Mundt, Enterococci in insects. Appl Microbiol, 1972. 24(4): p. 575-
80.
17. Macovei, L. and L. Zurek, Ecology of antibiotic resistance genes: characterization of
enterococci from houseflies collected in food settings. Appl Environ Microbiol, 2006.
72(6): p. 4028-35.
18. Cox, C.R. and M.S. Gilmore, Native microbial colonization of Drosophila melanogaster
and its use as a model of Enterococcus faecalis pathogenesis. Infect Immun, 2007. 75(4):
p. 1565-76.
19. Grabow, W.O. and P. Coubrough, Practical direct plaque assay for coliphages in 100-ml
samples of drinking water. Appl Environ Microbiol, 1986. 52(3): p. 430-3.
20. Bachrach, G., et al., Bacteriophage isolation from human saliva. Lett Appl Microbiol,
2003. 36(1): p. 50-3.
21. Classification and nomenclature of viruses. Fourth report of the International Committee
on Taxonomy of Viruses. Intervirology, 1982. 17(1-3): p. 1-199.

59
22. Pleceas, P. and H. Brandis, Rapid group and species identification of enterococci by
means of tests with pooled phages. J Med Microbiol, 1974. 7(4): p. 529-33.
23. Ward, R.L., D.R. Knowlton, and P.E. Winston, Mechanism of inactivation of enteric
viruses in fresh water. Appl Environ Microbiol, 1986. 52(3): p. 450-9.

60
CHAPTER 3
Characterization of Enterococcus faecalis-infecting phages (enterophages) as markers of
human fecal pollution in recreational waters
T.M. Santiago-Rodríguez1, C. Dávila
1, J. González
1, N. Bonilla
1, P. Marcos
2, M. Urdaneta
1,
M. Cadete 3, S. Monteiro
3, R. Santos
3, J.W. Santo-Domingo
4 and G. A. Toranzos
1
1 Environmental Microbiology Laboratory, Department of Biology, University of Puerto Rico,
San Juan, PR, 2
Biological Sciences Department, General Studies, University of Puerto Rico, San
Juan, PR, 3
Instituto Superior Tecnico, Laboratorio de Analises, Lisboa, Portugal, 4
US
Environmental Protection Agency, Cincinnati, Ohio, USA
ABSTRACT
Enterophages are a novel group of phages that specifically infect Enterococcus faecalis and have
been recently isolated from environmental water samples. Although enterophages have not been
conclusively linked to human fecal pollution, we are currently characterizing enterophages to
propose them as viral indicators and possible surrogates of enteric viruses in recreational waters.
Little is known about the morphological or genetic diversity which will have an impact on their
potential as markers of human fecal contamination. In the present study we are determining if
enterophages can be grouped by their ability to replicate at different temperatures, and if
different groups are present in the feces of different animals. As one of the main objectives is to
determine if these phages can be used as indicators of the presence of enteric viruses, the
survival rate under different conditions was also determined as was their prevalence in sewage
and a large watershed. Coliphages were used as a means of comparison in the prevalence and
survival studies. Results indicated that the isolates are mainly DNA viruses. Their morphology as
well as their ability to form viral plaques at different temperatures indicates that several groups
of enterophages are present in the environment. Coliphage and enterophage concentrations
throughout the watershed were lower than those of thermotolerant coliforms and enterococci.
Enterophage concentrations were lower than coliphages at all sampling points. Enterophages
showed diverse inactivation rates and T90 values across different incubation temperatures in both

61
fresh and marine waters and sand. Further molecular characterization of enterophages may allow
us to develop probes for the real-time detection of these alternative indicators of human fecal
pollution.
Keywords: enterococci; enterophages; fecal pollution; indicators
Reference: Santiago-Rodriguez, T.M., et al., Characterization of Enterococcus faecalis-infecting
phages (enterophages) as markers of human fecal pollution in recreational waters. Water Res,
2010. 44(16): p. 4716-25.
INTRODUCTION
Contamination of water sources by sewage is a health-related risk because of the possible
presence of pathogenic microorganisms such as Salmonella, Campylobacter, enteric viruses as
well as pathogenic protozoa [1-4]. Thermotolerant coliforms, enterococci [5, 6], Bacteroides
fragilis phages [7, 8] and somatic and F(male)-specific coliphages [9-13] are used and have been
proposed to infer the presence of these pathogens in water sources. However, these
microorganisms fail to fulfill the criteria of an ideal microbial indicator. For instance, the
prevalence and survival of thermotolerant coliforms is short and cannot be correlated with that of
pathogens [14, 15], human pathogens are not always accompanied by enterococci and vice versa
[16-18] and Bacteroides fragilis phages are found only in some geographical areas [16]. The
survival of somatic and F+ RNA and F+ DNA coliphages is approximately 20-100 days at 20 °C
in fresh water, depending on the group, which cannot be correlated with the survival of some
enteric viruses [10, 19]. Yet, other studies have found that F+ RNA coliphages could be active as
long as 3 days and somatic coliphages as long as 7 days [20]. Even though serotypes II and III of
F+ RNA coliphages have been correlated with human fecal contamination [19, 21, 22], their
presence in cow, pig, horse and bird feces makes them unreliable markers of human fecal
pollution [23, 24]. Also, currently used indicators are very susceptible to standard chlorination
treatments [24], while enteric viruses like Coxsackie virus, Rotavirus and Norwalk viruses are
susceptible to higher chlorine concentrations and have been found in treated sewage [25].
Because the presence of enteric viruses in water sources is of great concern due to their
resistance to removal treatments, new and reliable indicators of human fecal pollution and the

62
possible presence of these pathogens are still needed. Therefore, we are proposing
bacteriophages that specifically infect Enterococcus faecalis, which we call enterophages, as
viral indicators of human fecal contamination. Enterophages are a new group of phages that have
been recently isolated from recreational waters. Most isolates are tailed-phages, with icosahedral
capsids of 80 nm in diameter and tails of 200 nm in length [15], while others have smaller
capsids (12 nm) and shorter tails (60 nm) (this study), which accordingly could belong to the
Siphoviridae family [15]. These isolates differ in size from other previously characterized
Enterococcus faecalis-infecting phages, which have bigger capsids and tails (93 nm and 204 nm,
respectively) [26]; other isolates do not possess tails and have sphere-shaped, spiked structures of
approximately 70 nm [26, 27]. We have previously developed the methods for their detection in
environmental samples and results showed that these are promising markers of human fecal
pollution. The survival of enterophages in marine recreational waters was seen to be
approximately 7-10 days [15], similar to that of enteric viruses under similar environmental
conditions [15, 28] and unlike bacterial fecal indicators, enterophages have not been detected in
pristine aquatic ecosystems [15].
Even though results show that enterophages possess some of the characteristics of an ideal viral
indicator of water contamination by human feces, further characterization is still needed. It still
remains unknown how many groups of enterophages can be present in waters contaminated with
feces, though it has been suggested that at least three groups exist [15]. Previous studies have
found that different groups of coliphages exist and can be grouped according to their ability to
replicate at specific temperatures [29]. Consequently, the aims of the present study are to
determine if enterophages are present in animal and human fecal samples, to characterize
enterophages by means of their morphology, composition of their genetic material and their
ability to replicate at some temperatures and not others; to determine the prevalence of
enterophages in a fresh water gradient in the central region of Puerto Rico as well as to
determine if there are temporal variations in sewage in Puerto Rico and Portugal, and to
determine their survival in fresh and marine waters. Because coliphages have been used and
proposed as water quality controls of fecal contamination, their detection in both animal and
human feces, as well as the prevalence and survival studies were done in parallel with that of
enterophages as a means of comparison. The survival of enterophages in sand was also

63
performed because of the possible introduction of pathogens by feces. Most bacteria are not
reliable indicators of fecal contamination in beach sand because these are part of the
environmental microbiota [30].
MATERIALS AND METHODS
Detection of enterophages in animal and human feces
Cattle were selected as a study model to detect enterophages because E. faecalis is the most
commonly enterococci found in their feces [31]. Eight grams of cattle feces from 15 different
animals were collected from 4 different farms and stored at -20 °C until processed. To detect
enterophages, 1 g of each sample, 0.1 mL of Tween-20 [32], were added to 50 mL of sterile
distilled water and processed using the single layer method. The suspension was added to an
equal volume of liquefied regular strength (1X) Tryptic Soy Broth (TSB) (Difco) + agar (0.75 %
w/v) (Difco) which had CaCl2·2H20 (Fisher Scientific Co. NJ, USA) and NaN3 (MCB, OH,
USA) (final concentration of 2.6 mg/mL and 0.4 mg/mL, respectively). Additionally, 5 mL of an
overnight culture of E. faecalis grown in Azide Dextrose Broth (Difco) were added to the
mixture. This method was similarly conducted for the detection of coliphages. Briefly, the
suspension was added to molten 2X-strenght TSB and agar (1.5 % w/v). One mL of a fresh
culture of Escherichia coli in TSB was used for their detection. To test the presence of different
groups of coliphages and enterophages (according to their ability to replicate at different
temperatures), the mixtures were poured into sterile Petri dishes and incubated at 22, 37 or 41 °C
for 24 h [15].
To detect enterophages and coliphages in humans, fresh feces from 5 healthy individuals were
processed as follows: 0.1-0.5 g of fresh feces were suspended in 3 mL of a sterile 0.85 % NaCl
solution. 0.1 and 1 mL of the suspension were separately processed using the double layer
method. Briefly, the volumes were added to 4 mL of molten 1X TSB and agar (final 0.75 %
w/v), which had CaCl2 and NaN3 as described. Three-mL of an overnight culture of E. faecalis
were added to the mixture and poured into Petri dishes containing 20 mL of 1X TSB and agar
(final 1.5 % w/v). For the detection of coliphages, 1 mL of an overnight culture of E. coli was
added to the mixture. Plates were incubated at 37 °C for 24 h.

64
Enterophage Isolation and Purification
To isolate enterophages for further characterization, a 50 mL volume sample of raw domestic
sewage was processed as described [15]. The solution was mixed, poured into sterile Petri dishes
and plates were incubated at different temperatures, namely 22, 37, 41 and 45 °C for 24 h.
Individual viral plaques detected at each of the incubation temperatures were isolated. Different
plaque size and translucency were used as criteria for the isolation. Plaques were plucked using
sterile Pasteur pipettes and the plug was placed in a sterile Eppendorf tube containing 500 μL of
sterile Phosphate Buffer Solution (PBS). The plugs were dislodged and emulsified using the
same pipette. The tubes were centrifuged at 14,000 rpm for 10 min at 10 ºC to remove cellular
debris and agar and the supernatant transferred into a sterile tube. The isolates were then
propagated using 100 μL of the supernatant and processed using the double layer method as
described. Then, for those plates showing complete viral lysis, 5 mL of PBS was added and
slowly agitated rotationally for 20 min. The top agar was transferred to a sterile centrifuge tube,
broken down and then centrifuged at 14,000 rpm for 10 min at 10 ºC. The supernatant containing
the viruses was kept in a sterile tube at 4-7 °C for further use. To determine if the isolates were
capable of replicating at other temperatures, an E. faecalis inoculum was spread throughout a
Petri dish containing 1X TSB, 0.75 % agar, CaCl2 and NaN3 using a sterile swab. An aliquot of
10 μL of the purified supernatant was placed on top of the Petri dishes and incubated at 22, 37,
41 and 45 °C for 24 h. Further titration of each isolate was done using the double layer method
as described above.
Morphological Characterization of Enterophages
Prior to the examination of the morphology of the enterophage isolates, hydroextraction was
used to concentrate the viruses; briefly, a dialysis tube (12,000-14,000 MW cutoff, Spectrapor,
Los Angeles, CA) was loaded with 5 mL of the enterophage isolate, covered with crystalline
polyethylene glycol (PEG, Mol.wt. 8, 000, Sigma Chem. Co. MO; APHA, 1989), clamped and
placed at 4-7 °C overnight. The dialysis tube was washed with 100 μL of sterile 0.85 % NaCl.
The resulting solution was transferred into a sterile Eppendorf tube and kept at 4 °C until
analysis. An aliquot of the final solution was placed on carbon Type-B 200 mesh copper grids or
ultra thin carbon film/holey carbon 400 mesh copper grids and stained with uranyl acetate 2 %,
pH 4.5. All specimens were examined using a Karl Zeiss Leo 922 energy filtered transmission

65
electron microscope operated at 200 KV. At least 5 phage particles of each type were observed
[15].
Nucleic Acid Analyses
Extraction of enterophages nucleic acids was conducted using standard techniques of proteinase
K and phenol:chloroform treatments, followed by precipitation with ethanol [33]. To determine
if the enterophage isolates genetic material was DNA or RNA, aliquots of the isolated nucleic
acids were treated with either 1 U/μl DNase (Promega, Madison, WI, USA) [34] or 10 μg/μl
RNase (Sigma-Aldrich, Co. St. Louis, MO, USA). Agarose gel electrophoresis was performed as
previously described [33], using 0.7 % of agarose (Sigma-Aldrich, Co. St. Louis, MO, USA) gels
in TAE buffer. Bands were visualized after staining with an ethidium bromide solution (final
concentration of 0.5 μg/mL).
Prevalence in raw sewage
To determine the prevalence of enterophages in raw and treated domestic sewage from Puerto
Rico and Portugal, samples were collected monthly for 6 months and processed using the single
layer method as described [15]. For the detection of coliphages, serial dilutions were done and
processed using the double layer method as described. Plates were incubated at 37 °C for 24 h.
Detection of enterophages and other viral and bacterial indicators in a large watershed
Ten sampling sites subjected to different environmental conditions in the Rio Grande de Arecibo
watershed, localized in the central region of Puerto Rico, were selected to determine the
prevalence of enterophages in a natural setting (Figure 1). One liter samples from each point
were collected in sterile plastic bottles every week for 2.5 months and kept at 8 °C until
processing. Samples were analyzed for thermotolerant coliforms, enterococci, coliphages and
enterophages [15, 35]. For bacteria enumeration, 100 mL of water per sample site were filtered
using 47 mm polycarbonate membrane filters (GE Water & Process Technologies, pore size
0.45μm). This was done separately for the enumeration of both bacterial indicators. For the
enumeration of thermotolerant coliforms, filters were placed on Difco m-FC agar and incubated
at 45 °C for 24 h. Enumeration of enterococci was done by placing the membrane filter on Difco
m-Enterococcus agar and incubated at 37 °C for 48 h. Coliphage and enterophage were

66
enumerated by processing 100 mL of the water samples using the single layer method as
described in section 2.1 [15]. The solution was poured into Petri dishes and incubated at 22, 37,
41 or 45 °C for 24 h to test for the possible presence of different groups of enterophages across
the watershed.
Figure 1: Shows study site. Sample points are ordered according to their position in the
watershed, localized in the central region of Puerto Rico. Rio Grande de Arecibo watershed
localized in the central region of Puerto Rico. Sample points in this study are ordered according
to their position in the watershed (See legend).

67
Survival of enterophages and coliphages in waters and sand
To determine the survival of enterophages and coliphages in fresh and marine waters as well as
in beach sand, 2 L of fresh or marine water samples and 2 kg of sand were collected in sterile
plastic bottles and kept at 8 °C till used. At the laboratory, the water and sand samples were
placed in a sterile covered glass beaker. In order to simulate the die-off rate of these
bacteriophages under natural conditions [36, 37], raw domestic sewage with a coliphage
concentration of 104/100 mL
was added to the samples; the same concentration of an
enterophage isolate from a laboratory stock was also added to the samples. The samples were
kept in the dark at 22 °C for up to 12 days. Every two days, 50 mL aliquots were separately
processed for the detection of enterophages and coliphages using the single layer method as
described. Similarly, 50 g of sand and 0.1 mL of Tween-20 were added to 50 mL of sterile
distilled water and vigorously shake. The suspensions were processed as previously described
[15].
In order to make comparisons, decay constants (kd) from both enterophages and coliphages in
fresh and marine waters and sand were determined using the slopes of linear regressions made on
the semilog plots (PFU versus time). Decay rates (percent/ 2 days) were calculated by
multiplying the decay constants by 100. The time to reach a 90 % reduction in PFU densities
(T90) was also determined by dividing ln (0.1)/kd [36, 38-40].
RESULTS AND DISCUSSION
Enterophages and coliphages in animal and human feces
Coliphages have been used as viral indicators of fecal pollution, but their host specificity
includes cows, swine, humans, etc. In addition, those groups found in humans can also be found
in animal feces [23]. Similarly to these studies, we found coliphages in 12 of the 15 cattle feces
samples at an incubation temperature of 22 °C, but not at other incubation temperatures. Their
concentrations ranged from 8 Plaque Forming Units (PFU) to 326 PFU/g (data not shown). On
the other hand, enterophages were not detected in any of the cattle feces. Even though
enterophages were not detected, additional fecal samples from different animal species must be
processed to confirm enterophages specificity. Coliphages were detected in 3 out of the 5 human
fecal samples (approximately 80 PFU/g; data not shown), but enterophages were not detected.

68
These results could be due to the low concentrations of enterophages found in human feces.
Also, other incubation temperatures must be tested for their detection in fecal samples (namely
22 and/or 41 °C). Regarding the method used, the single layer method could be more appropriate
for their detection in human feces. These results are similar to those found in other studies which
showed that bacteriophages may be present in human feces at concentrations ranging from 0 to
105
CFU/g [41-44]. These studies have found that many of the bacteriophages in the human
intestine could be those infecting gram-positive bacteria, as in the case of enterophages [41]. The
absence of enterophages in cattle feces and other animals is encouraging, since it suggests that
these viruses are restricted to human hosts.
Differences in enterophage isolates according to morphology, genetic material and ability
to replicate at different temperatures
Both the morphology and genetic material of the enterophage isolates in this study correspond to
those viruses belonging to the Siphoviridae family. The morphology of the enterophage isolates
described here indicates that these are tailed phages, with icosahedral capsids of 12 nm in
diameter and non-contractile tails of 60 nm long (Figure 2). This morphology is different from
other virions we have described recently [15], which showed a bigger capsid and a longer tail,
similar to the classical Bradley’s basic morphology Group B belonging to the Siphoviridae
family [45].
Figure 2: Transmission Electron Microscopy image (TEM) of an enterophage isolate in this
study. The image shows that the isolate possesses an icosahedral head of 12 nm and a 60 nm
non-contractile tail (Bar =20 nm).

69
Figure 3: The genetic material of several enterophage isolates in a 0.7% agarose gel. The
molecular weight marker is shown in lane M (Lambda DNA HindIII digest, SigmaAldrich Co.
St. Louis, MO, USA). Lanes 1 and 4 contain untreated nucleic acids from two different isolates.
Lanes 2 and 5 contain DNase treated samples and lanes 3 and 6 contain RNase-treated samples.
In terms of the genetic material of enterophages, high molecular weight bands were observed in
the Rnase-treated samples. Enterophage nucleic acid molecular weight appears to be more than
23 kb and be in a super coiled conformation. Fragments in lanes 2 and 5 were degradated by
DNase, showing that the genetic material of the enterophage isolates described here is composed
of DNA (Figure 3). However, it still remains the possibility that the morphology and genomes of
other isolates could be different from those described. It also remains unknown if the isolates in
this study are double stranded or single stranded DNA viruses.
Enterophages isolated at 22 °C were capable of replicating at 22, 37, 41 and 45 °C. However,
those enterophages that were isolated at 37 and 41 °C replicated at 22, 37 and 41 °C, but not at
45 °C (Figure 4). Though further characterization of enterophages is still needed, these results
suggest that at least two groups of enterophages exist. Because enterophages can be isolated at
environmental temperatures, they can be detected in places without the facility of an incubator
by incubating the plates at ambient temperatures.

70
0
2
4
6
8
10
12
14
1 2 3 4
En
tero
ph
ag
e c
on
cen
tra
tio
n/m
L (
Lo
g)
Phage groups
22°C
37°C
41°C
45°C
Figure 4: Enterophage concentration/mL of four different enterophage isolates among four
different temperatures. Group 1 and 2 represent phages isolated from 22 ºC, group 3 represent an
isolate from 37 ºC and group 4 represents the isolate from 41 ºC. Results represent the geometric
mean of three replicas and standard deviations are represented by error bars.
Prevalence of enterophages in sewage and in a large watershed in Puerto Rico
Although enterophages were isolated at lower concentrations than coliphages, they were detected
in raw and treated domestic sewage in this study (Table 1). Interestingly, even though the same
host strain was used to detect enterophages in both Portugal (Port) and Puerto Rico (PR)
treatment plants, they were found in higher concentrations in raw domestic sewage from
Portugal, but coliphage concentrations were lower than those found in Puerto Rico (data not
shown). Prevalence studies in Portugal indicate that these viruses are not restricted to particular
geographical areas and could be tested as viral indicators in different waters types. Both
enterophages and coliphages (data not shown) were also detected in treated sewage in Puerto
Rico. Treated sewage was not processed for the enumeration of coliphages from Portuguese
treatment plants. Primary treatment is performed on raw sewage from domestic wastewater
treatment plants PR-A, PR-B and PR-C, while raw sewage from PR-D receives tertiary
treatment. In raw sewage receiving primary treatment, approximately 56 to 97 % of enterophages
were removed (Table 1), while a 91 % removal was seen in sewage from wastewater treatment

71
plant PR-D. Coliphages, on the other hand, were removed from a 95 to a 98 % in PR-A, PR-B
and PR-C, whereas more than 99 % of coliphages were removed from raw sewage in PR-D (data
not shown). Approximately 60 to > 99 % of enterophages were removed from raw sewage in
Portuguese treatment plants. Prevalence results from Puerto Rican treatment plants suggest that
enterophages may be more resistant to removal treatments than coliphages and could be
potentially more useful as indicators of the presence of enteric viruses even in treated sewage.
Water Treatment
Plant
Enterophages in inffluent
(PFU/100 mL)
Enterophages in effluent
(PFU/100 mL)
PR-A 82 ± 51.5 56 ± 55.6
PR-B 60 ± 46.1 14 ± 14.5
PR-C 115 ± 140.1 18 ± 1.2
PR-D 53 ± 53.5 6 ± 10.7
Port-A 17 ± 3.2 1 ± 0.9
Port-B 634 ± 15.7 253 ± 4.6
Port-C 695 ± 21.6 3 ± 0.9
Port-D 22 ± 2.8 2 ± 0.6
Port-E 575 ± 22.9 5 ± 1.3
Port-F 212 ± 7.0 12 ± 1.3
Port-G 405 ± 6.6 11 ± 1.4
Port-H 173 ± 10.9 21 ± 1.7
Port-I 774 ± 11.2 3 ± 0.9
Port-J 487 ± 6.5 4 ± 0.9
Table 1: Prevalence of enterophages in raw and treated sewage at different domestic wastewater
treatment plants in Puerto Rico (PR) and Portugal (Port). All data represent the average
concentration and standard deviation of viral plaques over a six-month period (n = 6).
In the large watershed, coliphages were found at higher concentrations than enterophages in all
sample points, but both bacteriophages were found at lower concentrations than thermotolerant
coliforms and enterococci (Figure 5). The high concentrations of bacterial indicators in the
watershed suggest that these may also be from animal origin, consequently, using these to track
the source of the contamination may not be reliable [46, 47]. No fluctuations in enterococci and
thermotolerant coliforms concentrations were found throughout the sampling period (Figure 5C
and 5D, respectively). Coliphages were detected at all incubation temperatures in most of the
sample points. These results suggest that different groups of coliphages were introduced into the

72
water sources, which may not necessarily be of human origin. Coliphage concentrations were
found to fluctuate throughout the sampling period; however, fluctuations were not as prominent
as those of enterophages (Data not shown).
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
4.50
5.00
1 2 3 4 5 6 7 8 9 10
En
tero
ph
ag
es
PF
U/1
00
mL
(L
og
10)
Sample sites
22°C
37°C
41°C
45°C
(A)
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
4.50
5.00
1 2 3 4 5 6 7 8 9 10
Co
lip
ha
ges
PF
U/1
00
mL
(L
og
10)
Sample sites
22°C
37°C
41°C
45°C
(B)

73
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
4.50
5.00
1 2 3 4 5 6 7 8 9 10
En
tero
cocc
us
CF
U/1
00
mL
(L
og
10)
Sample sites
(C)
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
4.50
5.00
1 2 3 4 5 6 7 8 9 10
Th
erm
oto
lera
nt c
oli
form
s
CF
U/1
00
mL
(L
og
10)
Sample sites
(D)
Figure 5: Concentrations of enterophages (A), coliphages (B), enterococci (C) and
thermotolerant coliforms (D) at ten different fresh water sample points with different impacts.
Sample points are numbered according to their position in the watershed: (1) Garza lake, (2)

74
Vaca river, (3) Before WTP Adjuntas, (4) After WTP Adjuntas, (5) Rio Grande de Arecibo, (6)
Before WTP Utuado, (7) After WTP Utuado, (8) Mouth of Rio Grande de Arecibo, (9)
Criminales river and (10) Caguana river at Jayuya. Numbers represent the geometric mean of a
2.5 month sampling period. Standard deviation is represented by error bars.
We are in the process of further characterizing the enterophage isolates molecularly, including
the sequencing of the viral genomes, which may allow us to determine unique sequences to be
used for the development of real-time methods for their detection in recreational and other water
sources. Also, future epidemiological studies should be focused on determining the relationship
between the prevalence of enterophages and enteric viruses in recreational waters and the
symptoms associated with ingesting water testing positive for the presence of enterophages.
Survival of enterophages in fresh waters
Enterophages were detected at 37 and 41 °C at 4 h after the incubation of the Petri dishes, but not
at 22 °C (data not shown). This indicates that these temperatures can be used to detect
enterophages in water samples in less time than bacterial indicators at similar temperatures.
Enterophages have a survival similar to that of human enteric viruses in fresh waters (Figure
6A) [28] and are able of replicating at 22, 37 and 41 °C. On the other hand, the survival of
coliphages in this study was more than 12 days, which cannot be correlated with that of certain
enteric viruses (Figure 6B). Even though temperature does not affect the formation of viral
plaques, it seems to affect the decay rates of enterophages. At 22 and 37 °C, the decay rates were
33.7 and 38.5 % every 2 days, respectively and at 41 °C the inactivation rate was 35.9 % every 2
days. T90 values also differed among the different temperatures. At 22 and 41 °C, 90 % of the
initial inoculum was inactivated at approximately 6.8 and 6.6 days, respectively and 6.0 days at
37 °C. Although it is not new that temperature does affect the decay rates of microorganisms [19,
36], our experiments on enterophages decay rates are part of their characterization as markers of
human fecal pollution and further experiments are still needed. Temperature also affected the
inactivation rates of coliphages. At 22 °C, approximately 11.6 % of the initial inoculum was
inactivated every 2 days. At 37 °C, coliphages had an inactivation rate of 9.9 % every 2 days; but
the highest inactivation rate was seen at 41 °C (26.2 % every 2 days). The time required to
inactivate 90 % of the initial coliphage concentration at 22 °C was 20.0 days, 23.3 days at 37 °C

75
and 9.0 days at 41 °C (Figure 6B). The long die-off rate of coliphages indicates that some
groups may not be used to track recent fecal contamination in water sources. Future survival
studies of enterophages in fresh waters will be done in order to determine the survival of
enterophages and coliphages in different points across the watershed described here and those
factors that could potentially affect their survival.
Survival of enterophages in marine recreational water and sand
As in the survival study of enterophages in fresh waters, plaques were detected at 37 and 41 °C
at 4 h after the incubation, but not at 22 °C, which suggests that these temperatures are favorable
to detect enterophages in short incubation periods. This could be due to diminished adsorption
rates of enterophages at low temperatures possibly because of a reduced affinity between phages
and bacterial surface receptors. Also, the adsorption rate is usually higher at the optimum
temperature for the growth of the host bacteria, suggesting that actively functioning bacteria are
required for a successful infection [48]. Nevertheless, after 11 days viral plaques were not
detected at 4 hr of incubation to any of the temperatures, but they were still detected after 24
hours. This may indicate that the adsorption rates were reduced with time.
The enterophage isolate used in this study has a survival time in both marine waters and sand of
11 to 13 days (Figure 7A and 8A), which is less than coliphages (Figure 7B and 8B). The decay
rates were 27, 29.3 and 30.7 % every 2 days at 22, 37 and 41 °C, respectively. The calculated T90
values for each temperature were 8.5, 7.9 and 7.5 days correspondingly. In the study of
enterophage survival in sand, the decay rates were 17, 20.6 and 21.4 and the calculated T90
values for each temperature, were 13.5, 11.2 and 10.8 respectively. Regarding coliphages, the
decay rates in marine water were 5, 4.5 and 7.7 % every 2 days. As in the study of coliphages in
fresh water, the higher decay rate occurred at 41 °C. The decay rates for coliphages in sand were
3.6, 4.1 and 6.2 and the T90 values were 64, 56.2 and 37.1.
Given that the isolate used in both survival studies (fresh water and marine water and sand) was
the same, it is interesting that the observed survival time was higher in marine water and sand
environments. Factors that could cause these longer survival periods include the high

76
concentration of inorganic salts, due to the fact that phage adsorption is strongly dependent on
salt concentrations [49].
These results and previous survival experiments using coliphages have found that it would be
impossible to correlate their survival with recent fecal contamination [50]. The survival of
enterophages in fresh and marine waters could indicate the recent introduction of human feces
into water sources. Molecular methods, like those currently tested on other microorganisms [51],
could also be developed to determine the die-off rate of enterophages in recreational waters. In
addition, because the effect of sunlight has been extensively studied on bacteria and
bacteriophages [36, 38], the inactivation of enterophages by sunlight has to be tested in future
studies.
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
0 1 2 3 4 5 6 7 8 9 10 11 12 13Coli
ph
age
PF
U/1
00
mL
(L
og
10)
Time (days)
22°C
37°C
41°C
A
B
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
0 1 2 3 4 5 6 7 8 9 10 11 12 13En
tero
ph
ag
e P
FU
/10
0m
L (L
og
10)
Time (days)
22°C
37°C
41°C
A

77
Figure 6: Survival of enterophages (A) and coliphages (B) in fresh water. Numbers represent the
arithmetic mean of three replicas. Standard deviations are represented by error bars.
Figure 7: Survival of enterophages (A) and coliphages (B) in marine water. Numbers represent
the average of three replicas. Standard deviations are represented by error bars.

78
Figure 8: Survival of enterophages (A) and coliphages (B) in sand. Numbers represent the
average of three replicas. Standard deviations are represented by error bars.
CONCLUSIONS
The absence of enterophages in cattle feces could indicate their specificity to the human colon
and could be potentially used to determine the presence of human enteric viruses in recreational
waters. Differences in morphology, genome and replication temperatures are factors that should
be taken in consideration when characterizing a new viral indicator, as in the case of
enterophages. These characteristics should be taken in consideration when choosing which

79
enterophage isolates have the potential of being molecularly characterized for their detection in
water sources. Enterophages could be used as surrogates of enteric viruses in recreational waters
due to their resistance to primary and tertiary treatments and similarity in die-off rates in fresh
and marine waters. Unlike proposed viral indicators, enterophages are not constrained to specific
regions and could be used to infer human fecal pollution in different water sources.
Few indicators have been proposed to infer the introduction of pathogens into beach sand. The
die-off rate of enterophages could also indicate recent fecal contamination, not only in water
sources, but also in beach sand.
ACKNOWLEDGMENTS
We thank Carlos Toledo for reviewing the manuscript at its early and late stages, Marisol
Rodriguez for collecting samples, Jean Frances Ruiz and Gwendolyn Argüello for processing the
samples.

80
LITERATURE CITED
1. Hurst, C.J., Overview of Water Microbiology as it Relates to Public Health, in Manual of
Environmental Microbiology, R.L. Crawford, Editor. 1996, ASM Press. p. 219-222.
2. Fong, T.T. and E.K. Lipp, Enteric viruses of humans and animals in aquatic
environments: Health risks, detection, and potential water quality assessment tools. Microbiol
Mol Biol Rev, 2005. 69(2): p. 357-371.
3. Moe, C.L., Waterborne Transmission of Infectious Agents, in Manual of Environmental
Microbiology, R.L. Crawford, Editor. 1996, ASM Press. p. 222-240.
4. Toranzos, G.A., McFeters, G. A., Borrego, J.J. and Savill, M., Detection of
Microorganisms in Environmental Freshwaters and Drinking Waters, in Manual of
Environmental Microbiology, R.L. Crawford, Editor. 1996, ASM Press. p. 249-260.
5. Ashbolt, N.J., Grabow, W.O.K. and Snozzi, M., Indicators of microbial water quality, in
World Health Organization (WHO). Water Quality: Guidelines, Standards and Health., L.A.B.
Fewtrell, J., Editor. 2001, IWA Publishing: London, UK. p. 289-316.
6. Wolf, H.W., The coliform count as a measure of water quality, in Water Pollution
Microbiology, T. Mitchell, Editor. 1972, Wiley-Interscience: New York. p. 333-345.
7. Jofre, J., et al., Potential usefulness of bacteriophages that infect Bacteroides fragilis as
model organisms for monitoring virus removal in drinking water treatment plants. Appl Environ
Microbiol, 1995. 61(9): p. 3227-31.
8. Gantzer, C., et al., Detection of infectious enteroviruses, enterovirus genomes, somatic
coliphages, and Bacteroides fragilis phages in treated wastewater. Appl Environ Microbiol,
1998. 64(11): p. 4307-12.
9. Grabow and Coubrough, Practical direct plaque assay for coliphages in 100 mL samples
of drinking water. Appl Environ Microbiol, 1986. 52: p. 430-433.
10. Allwood, P.B., et al., Survival of F-specific RNA coliphage, feline calicivirus, and
Escherichia coli in water: a comparative study. Appl Environ Microbiol, 2003. 69(9): p. 5707-
10.

81
11. Gantzer, C., et al., Adsorption and survival of faecal coliforms, somatic coliphages and
F-specific RNA phages in soil irrigated with wastewater. Water Sci Technol, 2001. 43(12): p.
117-24.
12. Hernandez-Delgado, E.A., Sierra, M.L. and Toranzos, G.A. , Coliphages as alternate
indicators of fecal contamination in tropical waters. Environ. Toxicol. Water Qual., 1991. 6: p.
131-143.
13. Stetler, R.E., Coliphages as indicators of enteroviruses. Appl Environ Microbiol, 1984.
48(3): p. 668-70.
14. Toranzos, G.A., McFeters, G. A., Borrego, J. J. and Savill, M., Detection of
microorganisms in environmental freshwaters and drinking waters, in Manual of Environmental
Microbiology, C.J. Hurst, Editor. 2007, ASM Press: Washington, DC. p. 249-264.
15. Bonilla, N., Santiago, T., Marcos, P., Urdaneta, M., Santo Domingo, J. and Toranzos, G.,
Enterophages, a group of phages infecting Enterococcus faecalis and their potential as alternate
indicators of human fecal contamination. Water Sci Technol, 2010 61: p. 293-300.
16. Weisberg, S.B., Noble, R.T. and Griffith, J.F., Microbial Indicators of Marine
Recreational Water Quality, in Manual of Environmental Microbiology, D.L. Crawford, Editor.
1996, ASM Press: Washington, D. C. p. 280-287.
17. Baele, M., et al., Composition of enterococcal and streptococcal flora from pigeon
intestines. J Appl Microbiol, 2002. 92(2): p. 348-51.
18. Cox, C.R., Gilmore, M. S., Native microbial colonization of Drosophila and its use as a
model of Enterococcus faecalis pathogenesis. Infect. Immun., 2007. 75(4): p. 1565-1576.
19. Long, S.C. and M.D. Sobsey, A comparison of the survival of F+RNA and F+DNA
coliphages in lake water microcosms. J Water Health, 2004. 2(1): p. 15-22.
20. Moce-Llivina, L., F. Lucena, and J. Jofre, Enteroviruses and bacteriophages in bathing
waters. Appl Environ Microbiol, 2005. 71(11): p. 6838-44.

82
21. Furuse, K., et al., Distribution of ribonucleic acid coliphages in raw sewage from
treatment plants in Japan. Appl Environ Microbiol, 1981. 41(5): p. 1139-43.
22. Scott, T.M., et al., Microbial source tracking: current methodology and future directions.
Appl Environ Microbiol, 2002. 68(12): p. 5796-803.
23. Cole, D., S.C. Long, and M.D. Sobsey, Evaluation of F+ RNA and DNA coliphages as
source-specific indicators of fecal contamination in surface waters. Appl Environ Microbiol,
2003. 69(11): p. 6507-14.
24. Tavakoli, A., et al., Chlorine residual efficiency in inactivating bacteria from secondary
contamination in Isfahan, 2002. East Mediterr Health J, 2005. 11(3): p. 425-34.
25. Kelly, S.M. and W.W. Sanderson, The effect of chlorine in water on enteric viruses. II.
The effect of combined chlorine on poliomyelitis and Coxsackie viruses. Am J Public Health
Nations Health 1960. 50: p. 14-20.
26. Bachrach, G., et al., Bacteriophage isolation from human saliva. Lett Appl Microbiol,
2003. 36(1): p. 50-3.
27. Uchiyama, J., et al., Isolation and characterization of a novel Enterococcus faecalis
bacteriophage phiEF24C as a therapeutic candidate. FEMS Microbiol Lett, 2008. 278(2): p.
200-6.
28. Ward, R.L., D.R. Knowlton, and P.E. Winston, Mechanism of inactivation of enteric
viruses in fresh water. Appl Environ Microbiol, 1986. 52(3): p. 450-9.
29. Seeley, N.D. and Primrose, The effect of temperature on the ecology of aquatic
bacteriophages. J. gen. Virol., 1980. 46: p. 87-95.
30. Khiyama, H.M. and J.C. Makemson, Sand beach bacteria: enumeration and
characterization. Appl Microbiol, 1973. 26(3): p. 293-7.
31. Devriese, L.A., et al., Enterococcal and streptococcal species isolated from faeces of
calves, young cattle and dairy cows. J Appl Bacteriol, 1992. 72(1): p. 29-31.

83
32. Golomidova, A., et al., The diversity of coliphages and coliforms in horse feces reveals a
complex pattern of ecological interactions. Appl Environ Microbiol, 2007. 73(19): p. 5975-81.
33. Sambrook, J., E. F. Fritsch, and T. Maniatis, RNase that is free of DNase, in Molecular
cloning: a laboratory manual. 1989, Cold Spring Harbor Laboratory Press: Cold Spring Harbor,
N.Y.
34. Huang, Z., M.J. Fasco, and L.S. Kaminsky, Optimization of Dnase I removal of
contaminating DNA from RNA for use in quantitative RNA-PCR. Biotechniques, 1996. 20(6): p.
1012-4, 1016, 1018-20.
35. Method 1600: Membrane Filter Test Method for Enterococci in Water, in EPA-821-R-
97-004. 1997, Office of Water, Washington D.C. p. .
36. Noble, R.T., I.M. Lee, and K.C. Schiff, Inactivation of indicator micro-organisms from
various sources of faecal contamination in seawater and freshwater. J Appl Microbiol, 2003. 96:
p. 464-472.
37. Duran, A.E., et al., Removal and inactivation of indicator bacteriophages in fresh waters.
J Appl Microbiol, 2001. 92: p. 338-347.
38. Noble, R.T. and J.A. Fuhrman, Virus decay and its causes in coastal waters. Appl
Environ Microbiol, 1996. 63(1): p. 77-83.
39. Sinton, L.W., R.J. Davies-Colley, and R.G. Bell, Inactivation of enterococci and fecal
coliforms from sewage and meatworks effluents in seawater chambers. Appl Environ Microbiol,
1994. 60(6): p. 2040-8.
40. Sinton, L., C. Hall, and R. Braithwaite, Sunlight inactivation of Campylobacter jejuni and
Salmonella enterica, compared with Escherichia coli, in seawater and river water. J Water
Health, 2007. 5(3): p. 357-65.
41. Breitbart, M., et al., Metagenomic analyses of an uncultured viral community from
human feces. J Bacteriol, 2003. 185(20): p. 6220-3.

84
42. Cornax, R., et al., Bacteriophages presence in human faeces of healthy subjects and
patients with gastrointestinal disturbances. Zentralbl Bakteriol, 1994. 281(2): p. 214-24.
43. Furuse, K., et al., Bacteriophage distribution in human faeces: continuous survey of
healthy subjects and patients with internal and leukaemic diseases. J Gen Virol, 1983. 64 (Pt 9):
p. 2039-43.
44. Gantzer, C., J. Henny, and L. Schwartzbrod, Bacteroides fragilis and Escherichia coli
bacteriophages in human faeces. Int J Hyg Environ Health, 2002. 205(4): p. 325-8.
45. Classification and nomenclature of viruses. Fourth report of the International Committee
on Taxonomy of Viruses. Intervirology, 1982. 17(1-3): p. 1-199.
46. Field, K.G. and M. Samadpour, Fecal source tracking, the indicator paradigm, and
managing water quality. Water Res, 2007. 41(16): p. 3517-38.
47. Bae, S. and S. Wuertz, Discrimination of viable and dead fecal Bacteroidales bacteria by
quantitative PCR with propidium monoazide. Appl Environ Microbiol, 2009. 75(9): p. 2940-4.
48. Binetti, A.G., Quiberoni A., Reinheimer, J.A. , Phage adsorption to Streptococcus
thermophilus: Influence of environmental factors and characterization of cell-receptors. Food
Research International, 2002. 35: p. 73-83.
49. Moldovan, R., E. Chapman-McQuiston, and X.L. Wu, On kinetics of phage adsorption.
Biophys J, 2007. 93(1): p. 303-15.
50. Hernandez-Delgado, E.A. and Toranzos, G.A., In-situ replication studies of Somatic and
Male-specific coliphages in a tropical pristine river. Wat. Sci. Technol., 1995. 31: p. 247-250.
51. Bae, S. and S. Wuertz, Rapid decay of host-specific fecal Bacteroidales cells in seawater
as measured by quantitative PCR with propidium monoazide. Water Res, 2009. 43(19): p. 4850-
9.

85
CHAPTER 4
Microbial quality of tropical inland waters and effects of rainfall events
T.M. Santiago-Rodriguez1, R.L. Tremblay
2, 3, C.Toledo-Hernandez
4, J.E.Gonzalez-Nieves
4, H.
Ryu4, J.W. Santo-Domingo
4 and G.A. Toranzos
1
1Environmental Microbiology Laboratory, Department of Biology, University of Puerto Rico,
Rico, San Juan, Puerto Rico 00931, 2Department of Biology, Call Box 860, University of Puerto
Rico, Humacao, Puerto Rico 00792, 3Crest-Catec, Center for Applied Tropical Ecology and
Conservation, PO Box 23341, University of Puerto Rico, Rio Piedras, Puerto Rico, USA 00931,
4US Environmental Protection Agency, Cincinnati, OH, USA
ABSTRACT
Novel markers of fecal pollution in tropical waters are needed since conventional methods
recommended for other geographical regions may not apply. To address this, the prevalence of
thermotolerant coliforms, enterococci, coliphages and enterophages was determined by culture
methods across a watershed. Additionally, human-, chicken- and cattle-specific PCR assays were
used to identify potential fecal pollution sources in this watershed. An enterococci quantitative
PCR (qPCR) assay was tested and correlated with culture methods at three sites since water
quality guidelines could incorporate this technique as a rapid detection method. Various rainfall
events reported prior to sample collection at three sites, were considered in the data analyses.
Thermotolerant coliforms, enterococci, coliphages and enterophages were detected across the
watershed. Human-specific Bacteroides, unlike the cattle and chicken-specific bacteria, were
detected mostly at sites with the corresponding fecal impact. Enterococci were detected by
qPCR as well, but positive correlations with the culture method were noted at two sites,
suggesting that either technique could be used. However, no positive correlations were noted for
an inland lake tested, suggesting that qPCR may not be suitable for all water bodies.
Concentrations of thermotolerant coliforms and bacteriophages were consistently lower after
rainfall events, pointing to a possible dilution effect. Rainfall positively correlated with
enterococci detected by culturing and qPCR, but this was not the case for the inland lake. The
toolbox of methods and correlations presented here could be potentially applied to assess the
microbial quality of various water types.

86
Keywords: bacterial indicators, bacteriophages, enterophages Microbial Source Tracking,
rainfall
Reference: Santiago-Rodriguez, T.M., et al., Microbial quality of tropical inland waters and
effects of rainfall events. Appl Environ Microbiol, 2012. 78(15): p. 5160-9.
INTRODUCTION
Monitoring microbial indicators of fecal pollution in tropical waters remains an issue of concern
since these may not accurately indicate the presence of microbes associated with fecal matter.
Indicators of fecal contamination used in different geographical areas include the thermotolerant
coliforms and enterococci [1]. These indicator bacteria are present in the intestinal tract of warm-
blooded animals and therefore, their presence in waters may indicate fecal pollution. However, it
has been shown that thermotolerant coliforms and enterococci may be part of the environmental
microbiota of tropical waters [2]. In addition, many bacterial indicators cannot be used to
indicate the time and source of the fecal contamination since they can replicate outside their host
and are present in the feces of different animals [3-8]. These shortcomings with regard to
indicator bacteria have prompted the use of alternate indicators, such as bacteriophages, which
show promising characteristics. For instance, coliphages, which are normally isolated from feces
and fecally-contaminated waters, do not appear to replicate outside their host and certain groups
have similar survival characteristics to enteric viruses in waters [9-12]. However, certain
coliphage groups are present in the feces of different warm-blooded animals and therefore, may
not be used to discriminate the source of the fecal pollution [13].
Recently, isolated phages that infect a specific Enterococcus faecalis type strain (enterophages)
have been proposed as good indicators of human-specific fecal contamination. Enterophages
have a similar survival time as human enteric viruses in marine and fresh waters and sand [14-
17], have been detected in raw and treated domestic sewage in Puerto Rico and Portugal and
have been detected neither in pristine waters nor in animal feces (e.g. pigs, dogs, cattle and
chickens) [16, 17]. However, more data are needed in order to accept enterophages as alternate
indicators of human fecal contamination. One way to further test enterophages as indicators of

87
fecal pollution is to compare them with currently used indicators and emerging molecular
methods in various water types.
Microbial Source Tracking (MST) methods can complement traditional methods used to assess
microbial water quality [18]. Specifically, host-specific assays have been tested in various water
sources and several have been successfully used to discriminate among the sources of fecal
pollution. Many Bacteroides species make up the intestinal microbiota of warm-blooded animals
their primary habitat and some species have shown high levels of host-specificity. In addition,
Bacteroides cannot replicate outside the intestinal tract as most species are strictly anaerobic
bacteria [19, 20]. Quantitative PCR (qPCR) is also perhaps one of the most promising methods
that indicate the levels of the target contaminant. However, as with culture-based methods,
molecular techniques suffer from shortcomings, such as the inability to distinguish between
viable and dead cells or the infectivity status of the target microorganism.
Detection of bacterial and viral indicators by culture or molecular-based techniques may be
influenced by rainfall events. It has been shown that rainfall may lead to higher numbers of
indicators and pathogens into surface waters and thus may represent an increased risk to human
health [21, 22]. Runoff, resuspension of sediments and sewage overflows, resulting from rainfall
events, may contribute to an increment of indicators and pathogens in surface waters [23, 24].
Nonetheless, rainfall is often not considered when monitoring the microbial quality of waters.
One main question to address is whether sampling under dry conditions alone is sufficient to
infer the presence of fecal indicators and pathogens or if results differ under wet conditions.
However, it is relatively difficult to confirm that an increase in microbial indicators and
pathogens after precipitation events truly represents a recent input of fecal matter [23]. In the
present study, we assessed the microbial quality of a tropical watershed using currently used
(thermotolerant coliforms, enterococci and coliphages) and proposed indicators (enterophages),
as well as molecular methods (host-specific assays and qPCR), considering rainfall as a possible
influential variable.
MATERIALS AND METHODS
Sample collection and sampling sites

88
In all cases, single one-liter grab samples were collected in sterile plastic bottles with sodium
thiosulfate (final concentration of approximately 10.0 mg/L) [25], up to four times per month
from November 2009 to April 2010. A total of 13 samples per site were collected and tested,
kept at 6-8 °C and processed within 4 to 6 h. Samples were collected from the Rio Grande de
Arecibo watershed in Puerto Rico, characterized by sporadic rainfall throughout the year.
Watersheds, unlike those found in many regions, are composed of small creeks. One of the
interesting characteristics of the Rio Grande de Arecibo watershed is that it has very distinct land
uses that contribute to human and/or animal fecal matter loadings. The watershed possesses both
urban and rural areas inhabited by more than 100,000 people, with waters that are used for
recreational activities and as a drinking water source. The watershed is linked to two of the
biggest dams on the island, providing approximately 380 million liters of drinking water daily to
the north coast [26].
Ten sampling sites were selected based on the potential different fecal pollution sources
associated with them. Site 1 (Largo Garza) is bordered at the southwestern part by the Reserva
Forestal de Toro Negro, thus the number of households are limited and farm animals, such as
poultry and cattle, are scarce or absent. Site 2 (Rio Vaca) is located downstream from site 1 and
residencies, poultry and cattle are also limited. Site 3 and 4 (Río Cidra) are located downstream
from the urban nucleus of the municipality of Adjuntas. Therefore, both sites are presumably
influence by urban runoff and local poultry. Site 3 is located upstream of the domestic
wastewater treatment plant (WTP) at Adjuntas and site 4 is located downstream from this WTP.
Site 5 (Río Grande de Arecibo) is lined by houses that drain their wastewaters directly into the
river and poultry and horses were also frequently observed nearby or in the river. Site 6 is
located at the southern entrance of the urban nucleus of the municipality of Utuado. It is
bordered by houses that manage their wastewater through septic tanks and by grass-fields, where
cattle were frequently observed. Site 7 is located 200 m downstream from the domestic WTP at
Utuado and poultry and cattle were frequently observed. Site 8 is located where the Rio Grande
de Arecibo discharges into the Atlantic Ocean, is impacted by cattle grazing and used for
recreational activities. The Río Criminales (site 9) is located between a fenced-farm with more
than 100 cows and human residences with septic tanks. Site 10 is located at the Rio Caguana and
receives the input of a WTP. This place is also impacted by cattle and horses (Figure 1).

89
Figure 1: Rio Grande de Arecibo watershed in Puerto Rico. Water samples were collected from
Lago Garza (site 1), Rio Vaca (2), upstream from the WTP at Adjuntas (3), downstream from the
WTP at Adjuntas (4), Rio Grande de Arecibo (5), upstream from the WTP at Utuado (6),
downstream from the WTP at Utuado (7), the estuary (8), Rio Criminales (9), and Rio Caguana
(10). Reproduced from reference 13 with permission.

90
Enumeration of indicators by culture methods
Thermotolerant coliforms were enumerated using m-FC agar incubated at 45 °C for 24 h and
enterococci using m-Enterococcus agar incubated at 37 °C for up to 48 h [27]. Coliphages and
enterophages were quantified using the single layer method as described previously [16]. The
type strains used for phage enumeration were Escherichia coli (ATCC 15597) and E. faecalis
(ATCC 19433). Plates were incubated at 22, 37, 41 and 45 °C to detect bacteriophages that
replicate at different temperatures. However, data presented in this study correspond to those
phages that replicate at 22 ºC since preliminary analyses suggested that this may be the optimal
temperature.
DNA extraction and PCR conditions
Water samples (100mL) were filtered through polycarbonate membranes (0.4 μm pore size, 47
mm diameter) (GE Water and Process Technologies, Trevose, PA). Total DNA was extracted
from the membranes using Mo Bio PowerSoil kits (MO BIO Laboratories, Carlsbad, CA)
according to the manufacturer’s protocol. DNA concentration was estimated using a NanoDrop
ND-1000 UV spectrophotometer (NanoDrop Technologies, Wilmington, DE). DNA extracts
were stored at -20 °C until further processing and analyzed using host specific PCR assays
commonly used in fecal source tracking studies and qPCR assay for general enterococci [28]
(Table 1). PCR was performed using cattle and human specific Bacteroides assays targeting 16S
rRNA genes and two chicken-specific assays targeting functional genes [29-31]. For
convenience, cattle and human-specific Bacteroides will be referred as CSB and HSB,
respectively. PCR amplifications were performed in 25 μL using the polymerase TaKaRa Ex
TaqTM (Takara Bio Inc.) in a Bio-Rad Tetrad2 Peltier Thermal Cycler (Bio-Rad, Hercules, CA)
under the following cycling conditions: an initial denaturation step at 95 °C for 5 min, followed
by 35 cycles of 1 min at 95 °C, 1 min at optimum annealing temperature, and 1 min at 72 °C.
PCR products were visualized in 1.5 % agarose gels using GelStar Nucleic Acid gel stain
(Lonza, Rockland, ME, USA).
The Taqman qPCR assay targeting the 23S rRNA gene of Enterococcus spp. (Entero1) was
performed in 25 μL reaction mixtures containing 1× TaqMan universal PCR master mix with
AmpErase uracil-N-glycosylase (Applied Biosystems, Foster City, CA), 0.2 mg/mL bovine

91
serum albumin (Sigma), 200 nM of each primer and FAM-labeled TaqMan probe. The qPCR
assays were performed using a 7900 HT Fast Real-Time Sequence Detector (Applied
Biosystems). All reaction mixtures were prepared in triplicate in MicroAmp Optical 96-well
reaction plates with MicroAmp Optical Caps (Applied Biosystems). The amplification protocol
involved incubation at 50 °C for 2 min to activate uracil-N-glycosylase, followed by 10 min of
incubation at 95 °C, and 40 cycles of 95 °C for 15 s and 60 °C for 1 min. PCR data were
analyzed using ABI’s Sequence Detector software (version 2.2.2). Duplicate serial dilutions of E.
faecalis genomic DNA (10-8
to 10-12
g/reaction) were used to generate standard curves. Percent
amplification efficiencies were calculated following the instrument’s manufacturer instructions
(Applied Biosystems). No-template controls were used to check for cross-contamination (two per
PCR plate). Assays were performed with 2 µL DNA extracts in a total volume of 100 µL, and
ten-fold dilutions of each DNA extract were used to test for PCR inhibition. Based on the
standard curve, qPCR intensities (QI) were expressed as a unit of pg/2 µL (i.e., genomic DNA
mass/reaction volume). Subsequently, the concentrations of the target gene (C) in water samples
were calculated by the following equation C (pg/100 mL) = QI (pg/2 µL) × CF × DF, where CF
(µl/mL) is a conversion factor of 1 and DF is a dilution factor of 50 [32].
Assay Primer and probe sequences (5’ to 3’)* Ta
(°C)**
Size
(bp)
Reference
General Bacteroides Bac32F: AACGCTAGCTACAGGCTT 53 694 Bernhard &
Bac708R: CAATCGGAGTTCTTCGTG Field (2000a)
Human-specific
Bacteroides (HSB)
HF183: ATCATGAGTTCACATGTCCG 63 543 Bernhard &
Field (2000b)
Cattle-specific
Bacteroides (CSB)
CF128: CCAACYTTCCCGWTACTC 62 598 Bernhard &
Field (2000b)
Chicken-specific
Bacteroides
CP2-9F: GTAAGACAGCAACCCCATGTA 56 245 Lu et al. (2007)
CP2-9R: ACCTATGGTTCAACACGCTTTA
Chicken-specific
Clostridium
CP3-49F: GTCCAGCGCCTCATTGAT 57 329 Lu et al. (2007)
CP3-49R: TGGTGATCGACTTTTCCAAT
General Enterococcus
qPCR (Entero1)
ECST748F: AGAAATTCCAAACGAACTTG 60 92 Ludwig &
Schleifer (2000) ENC854R: CAGTGCTCTACCTCCATCATT

92
GPL813:6FAM-
TGGTTCTCTCCGAAATAGCTTTAGGGCTA-
TAMRA
GPL813TQ:6FAM-
TGGTTCTCTCCGAAATAGCTTTAGGGCTA-
TAMRA
Table 1: Summary of oligonucleotide primers and probes for PCR and TaqMan qPCR.
USGS precipitation data
Precipitation data reported 24 and 48 h and one week prior to sample collection were obtained
from the US Geological Survey (USGS) Caribbean Water Science Center
(http://pr.water.usgs.gov/) for sites 1, 6 and 7 (USGS stations 50020100, 50021700 and
50024950, respectively) (Table 2). Precipitation data was collected from these sites due to their
proximity to a USGS station.
Site USGS station 24 h 48 h 1 week
Lago Garza (1) 50020100 1.6±0.4 5.7±0.4 64.3±40.3
Before WTP Utuado (6) 50021700 13.3±15.3 18.5±15.7 60.2±35.6
After WTP Utuado (7) 50024950 8.3±10.2 8.6±10.3 40.6±25.6
Table 2: Reported rainfall (mm; mean ± SD) for indicated period (November 2009 to April
2010) before sample collection. Rainfall data was collected from the USGS stations indicated.
Statistical analyses
The mean concentrations of cultivable thermotolerant coliforms, enterococci, coliphages and
enterophages and enterococci detected by qPCR were calculated using a Bayesian approach as
described previously [33]. Estimates were calculated assuming normal distribution of response.
Analysis of comparison among concentration at different temperatures, sample dates and sample
sites were analyzed using a one-way or two-way ANOVA [33]. Interaction effects were not
considered in these analyses and in all Bayesian analyses, credible intervals (CI) were calculated.
In addition, to determine correlations between rainfall and the concentration of the indicators (24
h, 48 h and one week) prior to collecting the sample, a Cross-correlation analysis was used.

93
However, prediction of the response or effect of rainfall to microbial concentrations was
estimated using non-linear regression as described by [33-37]. In all cases, the error around the
point estimate, the 95% CI, is calculated. The 95% CI is the area under the curve of the posterior
distribution, as compared to the 95% confidence interval, that is the probability that the point
estimate would be between the lower and upper bound [36]. All analyses were performed with
the software package IBM SPSStatistics v.19.
RESULTS
Detection of indicators by culture-based methods
Thermotolerant coliforms showed the lowest counts at sites 1 and 9, with mean concentrations of
0.0 and 11.6 CFU/100 mL, respectively. The highest concentrations of thermotolerant coliforms
correspond to sites 4 and 7, with mean concentrations of 817.8 and 1053.0 CFU/100 mL,
respectively (Figure 2A). The variance of thermotolerant coliforms did not differ across the
sampled points (354.0-356.0 CFU/100 mL), except for site 10, in which the SD was 32.3
CFU/100 mL. Enterococci exhibited lower counts at sites 1 and 10, with mean concentrations of
0.0 and 9.6 CFU/100 mL, respectively, and higher counts at site 9 (499.2 CFU/100 mL) (Figure
2B). The variance of enterococci did not differ across most of the sampled points, exhibiting a
standard deviation (SD) of 99.23-99.67 CFU/100 mL, except for site 10, which showed a SD of
21.2 CFU/100 mL. In terms of the phages, these were detected in lower concentrations
compared to the bacterial indicators. The lowest and highest means for coliphages correspond to
sites 1 and 9 (32.7 PFU/100 mL and 314.0 PFU/100 mL, respectively) (Figure 2C). The
variance for the coliphages differed across the sampling sites, in which the lowest and highest
SD correspond to sites 1 (6.3 PFU/100 mL) and 9 (139.1 PFU/100 mL), respectively. In terms of
the enterophages, the highest mean concentrations correspond to sites 3 and 7 (Figure 2D) and
variances did not differ across the sampling sites (SD = 4.9). CI for the bacterial and viral
indicators are represented in Figure 2.

94
Figure 2: Credible intervals (CIs) for thermotolerant coliforms (A), enterococci (B), coliphages
(C), and enterophages (D) in the Rio Grande de Arecibo watershed. Results represent the means
of the sampling period (n=13), and credible intervals, representing positive values, were drawn
as well. Except for enterophages, CIs could not be calculated for site 10 since most data are
zeros.
Detection of host-specific Bacteroides by PCR and enterococci by qPCR
Of the 13 samples tested per site, HSB were not detected at sites 1 and 2, were detected once at
sites 3, 6, 8 and 9, twice at sites 4 and 5 and three times at sites 7 and 10. CSB were detected
once at sites 1 and 9 and twice at sites 7 and 8, but were not detected in samples collected from
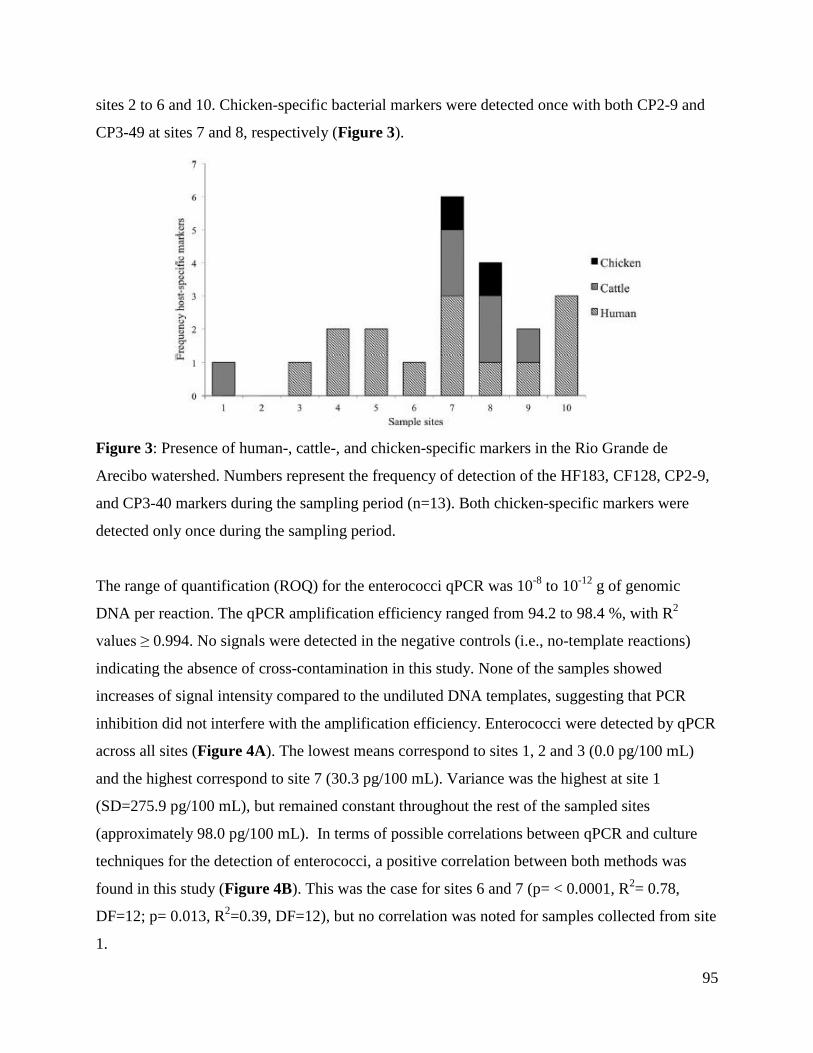
95
sites 2 to 6 and 10. Chicken-specific bacterial markers were detected once with both CP2-9 and
CP3-49 at sites 7 and 8, respectively (Figure 3).
Figure 3: Presence of human-, cattle-, and chicken-specific markers in the Rio Grande de
Arecibo watershed. Numbers represent the frequency of detection of the HF183, CF128, CP2-9,
and CP3-40 markers during the sampling period (n=13). Both chicken-specific markers were
detected only once during the sampling period.
The range of quantification (ROQ) for the enterococci qPCR was 10-8
to 10-12
g of genomic
DNA per reaction. The qPCR amplification efficiency ranged from 94.2 to 98.4 %, with R2
values ≥ 0.994. No signals were detected in the negative controls (i.e., no-template reactions)
indicating the absence of cross-contamination in this study. None of the samples showed
increases of signal intensity compared to the undiluted DNA templates, suggesting that PCR
inhibition did not interfere with the amplification efficiency. Enterococci were detected by qPCR
across all sites (Figure 4A). The lowest means correspond to sites 1, 2 and 3 (0.0 pg/100 mL)
and the highest correspond to site 7 (30.3 pg/100 mL). Variance was the highest at site 1
(SD=275.9 pg/100 mL), but remained constant throughout the rest of the sampled sites
(approximately 98.0 pg/100 mL). In terms of possible correlations between qPCR and culture
techniques for the detection of enterococci, a positive correlation between both methods was
found in this study (Figure 4B). This was the case for sites 6 and 7 (p= < 0.0001, R2= 0.78,
DF=12; p= 0.013, R2=0.39, DF=12), but no correlation was noted for samples collected from site
1.

96
Figure 4: qPCR for enterococci and correlation between qPCR and culture-based techniques for
enterococci. (A) Results show the mean genomic mass (pg)/100 mL (n=13) for enterococci
across the Rio Grande de Arecibo watershed and the corresponding credible intervals. (B)
Correlation between qPCR and culture- based methods for the detection of enterococci (site 7).
Correlations of indicators with rainfall and detection methods
At site 1, a positive correlation was noted with enterococci detected by culture methods and
rainfall reported 24 h, for 48 h, but not for one week prior sample collection. At sites 6 and 7, a
positive correlation was noted with the precipitation reported 24 h (Figure 5A), for 48 h and one
week prior sample collection. For enterococci qPCR data from sites 6 and 7, a positive
correlation was found at 24 h (Figure 5B), for 48 h and one prior sample collection, but this was
not the case for site 1 (Figure 5C). Results from the correlation analyses between enterococci
detected by culture methods or qPCR are shown in Table 3. No correlation was found with
thermotolerant coliforms and precipitation in any of the study sites (Figure 5D). Similarly, no
correlations were found with coliphages or enterophages and precipitation (Figure 5E and F). In

97
addition, positive correlations between thermotolerant coliforms and coliphages were found at
sites 1 and 7 (Nonparametric Spearman’s test: ρ= 0.45; p=0.014 and ρ= 0.85; p=0.049,
respectively). No other correlations between any of the microbial indicators detected by culture
methods were noted.

98
Figure 5: Correlation between rainfall and microbial indicators. Precipitation data were collected
from sites 1, 6, and 7 due to the proximity of a USGS station. Results show the possible effect of
precipitation reported 24 h before collection of the samples on enterococci detected by culture
methods (site 7) (A), 24 h before collection of the samples on enterococci detected by qPCR (site
7) (B), 24 h before collection of the samples on enterococci detected by qPCR (site 1) (C), 48 h
before collection of the samples on thermotolerant coliforms (site 1) (D), 24 h before collection
of the samples on coliphages (site 7) (E), and 24 h before collection of the samples on
enterophages (site 7) (F).
DISCUSSION
Prevalence of the bacterial and viral indicators detected by culture methods
Detection of thermotolerant coliforms throughout the watershed suggests an input of fecal
matter, although it is relatively difficult to identify the possible sources. High concentrations of
these indicator bacteria at sites 4 and 7 may suggest an inefficient removal by sewage treatment
or an additional input of fecal matter, but future studies are needed to confirm this. Moreover,
rainfall may play an important role in the loading dynamics. For instance, when no precipitation
was reported at site 1, thermotolerant coliforms ranged between 0 to ~40 CFU/100mL and 1
CFU/100mL was detected when 10.2 mm of rain were reported 24 h prior to collecting the
samples (data not shown). This may suggest that rainfall may have a possible dilution effect on
the thermotolerant coliforms. Interestingly, 0 to ~ 35 CFU/100mL were detected when no
precipitation was reported, but ~ 40 CFU/100mL were detected when >25.4 mm of rain were
reported for 48 h prior to sample collection at site 1. This may suggest that tropical sediments
may be a source of thermotolerant coliforms and these may have been resuspended due to the
rainfall reported in the previous 48 h. However, future studies are still needed to determine the
prevalence of these indicator bacteria in tropical sediments and other possible loading
mechanisms after rainfall events. A similar pattern was noted with thermotolerant coliforms and
rainfall reported for 24 and 48 h prior to collecting the samples at sites 6 and 7, but unlike site 1,
high numbers of these bacteria could also be associated with their transport from higher sites of
the watershed. Additional studies are needed to determine the possible transport mechanisms of
these indicator bacteria in tropical watersheds. No differences in the concentrations of
thermotolerant coliforms were noted when correlated with the rainfall reported for one week

99
prior sample collection. This suggests that different periods of precipitation may differently
influence the numbers of these bacteria when detected by culture methods.
It has been suggested that some enterococci are naturally-occurring in tropical waters [2]. Results
presented here suggest that many Enterococcus spp detected by culture methods are from a fecal
origin. The reason for this is that site 1 is one of the highest sites of the watershed, possibly one
of the less impacted by human or animal activities and very low numbers were detected. These
results are consistent with previous studies in which lower numbers of indicator bacteria have
been detected in inland lakes [38]. Higher enterococci numbers at sites 4 and 7 suggests that
these sites are point-sources of fecal pollution. Interestingly, site 10 represents a point-source of
fecal contamination as well, but low numbers may be due to an efficient removal by sewage
treatment. Also, the possibility remains that rainfall may also contribute to the input of
enterococci into tropical surface waters. Even though rainfall did not correlate with enterococci
detected at site 1, a positive correlation was noted with rainfall reported for one week prior to
sample collection at site 6. In addition, a positive correlation between enterococci and rainfall
reported 24 and for 48 h and one week prior to sample collection at site 7 suggests that loading
of enterococci into surface waters may be influenced by precipitation events. Based on previous
studies, it is possible that sediments and runoff could contribute to the input of these bacteria into
surface waters, but future studies are still needed in order to confirm if this is the case for waters
in Puerto Rico. Results presented here are consistent with previous studies in which higher
numbers of enterococci are detected after rainfall events in Hawaiian marine waters [39].
However, previous studies have not considered the effect of various precipitation periods prior to
sample collection. The present study suggests that various rainfall episodes may be considered
prior to inferring a fecal input in tropical surface waters. However, the possibility remains that,
although positive correlations were noted, rainfall may not always result in an increase in
enterococci numbers.
Both coliphages and enterophages were detected throughout the watershed, indicating that the
sampled sites may be impacted by human or animal fecal matter at some extent. The high
concentrations of coliphages (compared to enterophages) suggest that more than one source
could be contributing to their input and/or that they are present in higher numbers in feces. The

100
low concentrations of enterophages throughout the watershed may suggest that: (i) these are
present in low concentrations in human feces, (ii) few sources of human fecal pollution are
contributing to the input of these bacteriophages (e.g. septic tanks or domestic WTP), (iii)
enterophages are able to survive for short periods in tropical fresh waters and/or (iv) may be
diluted as a result of precipitation events. Even though enterophage concentrations were
relatively low, further studies need to compare their numbers with those of enteric viruses under
similar conditions since a reliable indicator should be at least as abundant as the pathogen of
concern. Also, the possibility remains that rainfall may have a possible dilution effect on both
phages. The reason for this is that up to 1000 and 50 coliphage and enterophage PFU/100mL,
respectively, were detected when no rainfall was reported and approximately 300 and 0 PFU/100
mL, respectively, were detected when >38.1 mm of rain were reported.
Monitoring indicator bacteria by molecular-based techniques
Human and animal fecal contaminations are among the major concerns to public health since
pathogens could be present [40-42]. Therefore, it is important to validate source-specific
markers, particularly in the tropics, as most studies were done in temperate regions. One
promising tool for indicating human fecal contamination is the Bacteroides marker HF183 [43,
44]. In the present study, HSB were mostly detected in samples collected downstream of all three
domestic WTP, suggesting that their presence is the result of the input of human fecal matter.
This supports the specificity of the HF183 marker in tropical inland waters with human fecal
impact. In terms of the cattle-specific markers, these have been shown to be a promising assay to
detect ruminant pollution in environmental waters. However, some studies have shown that CF
128 cross reacts with DNA extracts from various farm animals [45]. In the present study, CSB
were detected at sites 1, 2 and 3, in which cattle is not present, but it was expected to detect CSB
at site 6 since cattle grazing takes place. Site 5, which could be impacted by horse fecal matter,
tested positive for CSB by using the CF128 marker and this is comparable with previous studies
in which CF128 amplified CSB DNA in horse fecal matter [20]. Similarly, chicken-specific
Bacteroides were not detected using CP2-9 or CP3-49 in waters impacted by chicken fecal
matter. Sample sites 4, 5 and 7 were considered to be impacted by chicken-fecal matter, but
chicken-specific Bacteroides were detected once at site 7 during the sampling period and
chicken-specific Clostridium were detected once in samples collected from site 8. However, it

101
should be noted that the number of chickens in these sites are relatively low, suggesting that the
assays may not be sensitive enough to detect low levels of poultry pollution.
qPCR is currently being considered as a possible rapid detection method to be incorporated into
USEPA guidelines; therefore, it is important to test this technique in different water types prior
to implementation. Positive correlations between qPCR and culture methods in temperate waters
suggest that detection of enterococci by qPCR could also be used as a tool for determining
health-related risks and that both techniques may be reliable for the detection of enterococci [46].
In the present study, enterococci genomic mass detected by qPCR was found to be higher at sites
with known point-sources of fecal pollution. However, detection of the multiple copies of the
23S rRNA gene present in the bacterial genome, viable-non-cultivable bacteria, as well as DNA
from dead cells may add to the apparent increase in enterococci genomic mass [47]. Rainfall may
also contribute to the increment of enterococci detected by qPCR. At sites 6 and 7, a positive
correlation with rainfall may suggest that it may have an indirect contribution to the input of
Enterococcus into tropical surface waters (e.g. septic tanks overflows and animal droppings).
However, as with culture methods, it is difficult to predict that rainfall may always have a
positive correlation with enterococci detected by qPCR.
Correlations between culture-based and qPCR methods in this study may suggest that either of
the techniques may be used to detect enterococci in tropical inland waters. Correlation results in
this study are among the few performed for tropical inland waters, but similar outcomes have
been reported across the US [38]. Specifically, both methods were positively correlated in
Hawaiian marine waters, but this correlation was not noted for fresh waters and estuaries [39].
Results suggest that qPCR may not be suitable for all water types since different outcomes are
obtained from both methods and a correlation may not be expected in various water types at all
times. Future studies need to determine if correlations between qPCR and culture methods in
tropical inland waters exist throughout the year. Correlations between molecular and culture
techniques and between thermotolerant coliforms and coliphages suggest that the bacteriological
and virological quality of tropical inland waters should be evaluated using diverse MST tools.
This opens up the possibility of using a toolbox whenever determining the microbiological
quality of tropical inland waters.

102
CONCLUSIONS
Few studies have evaluated the microbial quality of tropical inland waters. One problem is the
need to develop appropriate water quality standards for tropical waters since guidelines used in
other areas may not be applicable. The present study tested combinations of markers to assess the
microbial quality of a tropical watershed in Puerto Rico, but results presented here need to be
further tested in other tropical watersheds. Even though a different combination of MST
techniques was tested in this study, there is still a need of developing a more robust toolbox to
infer fecal pollution and the possible sources in various tropical water types. In terms of the
bacterial indicators, these alone may not be suitable when inferring fecal contamination or its
source in tropical inland waters. Additional studies are needed to further examine the loading
dynamics of indicator bacteria into tropical fresh waters as a result of rainfall events.
Enumeration of coliphages and enterophages may be a more appropriate way to infer fecal
contamination since these are not ubiquitous in tropical waters and their numbers do not increase
after rainfall events. In terms of the host-specific markers in this study, only HF183 seemed a
reliable means of inferring the targeted source of fecal contamination in tropical waters. Cattle
and chicken-specific Bacteroides and Clostridium markers may not distinguish the source of the
fecal pollution in tropical waters; therefore, additional markers that could more reliably detect
cattle and chicken-specific sources of fecal pollution in Puerto Rico remain an important need.
Positive correlations between culture and molecular methods in this study should be considered
prior to implementing qPCR as a rapid method for inferring fecal pollution in the tropics since it
may not apply to all water bodies, specifically to inland lakes. qPCR assays targeting enterococci
in tropical waters must be carefully considered and further tested in other tropical areas since
rainfall may overestimate the numbers of enterococci detected by this method. The methods and
results presented here could be potentially useful in other water types.
ACKNOWLEDGMENTS
We thank the USGS for the precipitation data, the Department of Natural Resources of Puerto
Rico for information about the Rio Grande de Arecibo watershed and Gwendolyn Argüello and
Jean F. Ruiz for processing the samples. We also thank Dr. Pablo A. Ortiz-Pineda for managing
the figures at the modification stage. This research was partially supported by MBRS-RISE (NIH
Grant Number 2R25GM061151-09) and the US Environmental Protection Agency.

103
SUPPLEMENTARY INFORMATION
(A)
(B)
0
500
1000
1500
2000
2500
3000
1 2 3 4 5 6 7 8 9 10
En
tero
co
cci C
FU
/10
0m
L
Sample sites
Nov-Jan
0
500
1000
1500
2000
2500
3000
1 2 3 4 5 6 7 8 9 10
En
tero
co
cci C
FU
/10
0m
L
Sample sites
Feb-Apr

104
(C)
(D)
Supplementary Figure 1: Enterococci and thermotolerant coliforms CFU/100mL in the Rio
Grande de Arecibo watershed. Panels (A) and (B) represent the concentration of enterococci
during a period of high and low rainfall events, respectively. Panels (C) and (D) represent the
concentration of thermotolerant coliforms during the same periods of rainfall. Minimum and
maximum values are represented by error bars.
0
1000
2000
3000
4000
5000
6000
7000
8000
1 2 3 4 5 6 7 8 9 10
Th
erm
oto
lera
nt co
lifo
rms
CF
U/1
00
mL
Sample sites
Nov-Jan
0
1000
2000
3000
4000
5000
6000
7000
8000
1 2 3 4 5 6 7 8 9 10
Th
erm
oto
lera
nt co
lifo
rms
CF
U/1
00
mL
Sample sites
Feb-Apr

105
(A)
(B)
-0.20
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
1.80
2.00
1 2 3 4 5 6 7 8 9 10
En
tero
ph
age
PF
U/1
00
mL
(L
og
10)
Sample sites
Nov-Jan
Feb-Apr
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
1.80
2.00
1 2 3 4 5 6 7 8 9 10
En
tero
ph
age P
FU
/10
0m
L (L
og
10)
Sample sites
Nov-Jan
Feb-Apr
22ºC
37ºC

106
(C)
(D)
Supplementary Figure 2: Enterophages in the Rio Grande de Arecibo watershed according to
the incubation temperature and rainfall period. Figure shows enterophages that replicate at 22
(A), 37 (B), 41 (C) and 45°C during the periods of November to January and February to April.
Standard deviations are shown by error bars.
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
1.80
2.00
1 2 3 4 5 6 7 8 9 10
En
tero
ph
age P
FU
/10
0m
L (L
og
10)
Sample sites
Nov-Jan
Feb-Apr
41ºC
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
1.80
2.00
1 2 3 4 5 6 7 8 9 10
En
tero
ph
age P
FU
/10
0m
L (L
og
10)
Sample sites
Nov-Jan
Feb-Apr
45ºC

107
(A)
(B)
0.00
0.50
1.00
1.50
2.00
2.50
3.00
1 2 3 4 5 6 7 8 9 10
Co
lip
ha
ge P
FU
/10
0m
L (
Lo
g1
0)
Sample sites
Nov-Jan
Feb-Apr
22ºC
0.00
0.50
1.00
1.50
2.00
2.50
3.00
1 2 3 4 5 6 7 8 9 10
Co
lip
ha
ge P
FU
/10
0m
L (
Lo
g 10)
Sample sites
Nov-Jan
Feb-Apr
37ºC

108
(C)
(D)
Supplementary Figure 3: Coliphages in the Rio Grande de Arecibo watershed according to the
incubation temperature and rainfall period. Figure shows enterophages that replicate at 22 (A),
37 (B), 41 (C) and 45°C during the periods of November to January and February to April.
Standard deviations are shown by error bars.
0.00
0.50
1.00
1.50
2.00
2.50
3.00
1 2 3 4 5 6 7 8 9 10
Co
lip
ha
ges
PF
U/1
00
mL
(L
og
10)
Sample sites
Nov-Jan
Feb-Apr
41ºC
0.00
0.50
1.00
1.50
2.00
2.50
3.00
1 2 3 4 5 6 7 8 9 10
Co
lip
ha
ge P
FU
/10
0m
L (
Lo
g 10)
Sample sites
Nov-Jan
Feb-Apr
45ºC

109
LITERATURE CITED
1. Muñiz, I., et al., Survival and activity of Streptococcus faecalis and Escherichia coli in
tropical fresh water. Microbial Ecology, 1989. 18: p. 125-134.
2. Rivera, S.C., T.C. Hazen, and G.A. Toranzos, Isolation of fecal coliforms from pristine
sites in a tropical rain forest. Appl Environ Microbiol, 1988. 54(2): p. 513-517.
3. Byappanahalli, M.N., et al., Growth and survival of Escherichia coli and enterococci
populations in the macro-alga Cladophora (Chlorophyta). FEMS Microbiol Ecol, 2003.
46(2): p. 203-11.
4. Hardina, C.M. and R.S. Fujioka, Soil: The environmental source of Escherichia coli and
enterococci in Hawaii's streams. Environmental Toxicology 1991. 6(2): p. 185-195.
5. Whitman, R.L., et al., Occurrence of Escherichia coli and enterococci in Cladophora
(Chlorophyta) in nearshore water and beach sand of Lake Michigan. Appl Environ
Microbiol, 2003. 69(8): p. 4714-9.
6. Devriese, L.A., et al., Characterization and Identification of Enterococcus species
isolated from the Intestines of Animals. International Journal of Systematic Bacteriology,
1987. 37(3): p. 257-259.
7. Bahirathan, M., L. Puente, and P. Seyfried, Use of yellow-pigmented enterococci as a
specific indicator of human and nonhuman sources of fecal pollution. Can J Microbiol,
1998. 44: p. 1066-1071.
8. Whitlock, J.E., D.T. Jones, and V.J. Harwood, Identification of the sources of fecal
coliforms in an urban watershed using antibiotic resistance analysis. Water Res, 2002.
36(17): p. 4273-82.
9. Hernandez-Delgado, E.A. and G.A. Toranzos, In situ replication studies of somatic and
male-specific coliphages in a tropical pristine river. Wat Sci Tech, 1995. 31(5-6): p. 247-
250.
10. Cole, D., S.C. Long, and M.D. Sobsey, Evaluation of F+ RNA and DNA coliphages as
source-specific indicators of fecal contamination in surface waters. Appl Environ
Microbiol, 2003. 69(11): p. 6507-14.
11. Furuse, K., et al., Bacteriophage distribution in human faeces: continuous survey of
healthy subjects and patients with internal and leukaemic diseases. J Gen Virol, 1983. 64
(Pt 9): p. 2039-43.

110
12. Gantzer, C., J. Henny, and L. Schwartzbrod, Bacteroides fragilis and Escherichia coli
bacteriophages in human faeces. Int J Hyg Environ Health, 2002. 205(4): p. 325-8.
13. Calci, K.R., et al., Occurrence of male-specific bacteriophage in feral and domestic
animal wastes, human feces, and human-associated wastewaters. Appl Environ
Microbiol, 1998. 64(12): p. 5027-9.
14. Ward, R.L., D.R. Knowlton, and P.E. Winston, Mechanism of inactivation of enteric
viruses in fresh water. Appl Environ Microbiol, 1986. 52(3): p. 450-9.
15. Rao, V.C., et al., Isolation of enteroviruses from water, suspended solids, and sediments
from Galveston Bay: survival of poliovirus and rotavirus adsorbed to sediments. Appl
Environ Microbiol, 1984. 48(2): p. 404-9.
16. Bonilla, N., et al., Enterophages, a group of phages infecting Enterococcus faecalis, and
their potential as alternate indicators of human faecal contamination. Water Sci Technol,
2010. 61(2): p. 293-300.
17. Santiago-Rodriguez, T.M., et al., Characterization of Enterococcus faecalis-infecting
phages (enterophages) as markers of human fecal pollution in recreational waters. Water
Res, 2010. 44(16): p. 4716-25.
18. Santo-Domingo, J.W., et al., Quo vadis source tracking? Towards a strategic framework
for environmental monitoring of fecal pollution. Wat Res, 2007. 41: p. 3539-3552.
19. Allsop, K. and D.J. Stickler, An assessment of Bacteroides fragilis group organisms as
indicators of human faecal pollution. J Appl Bacteriol, 1985. 58(1): p. 95-9.
20. Lamendella, R., et al., Assessment of fecal pollution sources in a small northern-plains
watershed using PCR and phylogenetic analyses of Bacteroidetes 16S rRNA gene. FEMS
Microbiol Ecol, 2007. 59(3): p. 651-60.
21. Rose, J.B., et al., Climate variability and change in the United States: potential impacts
on water- and foodborne diseases caused by microbiologic agents. Environ Health
Perspect, 2001. 109 Suppl 2: p. 211-21.
22. Lipp, E.K., et al., The Effects of Seasonal Variability and Weather on Microbial Fecal
Pollution and Enteric Pathogens in a Subtropical Estuary. Estuaries, 2001. 24(2): p. 266-
276.
23. Kistemann, T., et al., Microbial load of drinking water reservoir tributaries during
extreme rainfall and runoff. Appl Environ Microbiol, 2002. 68(5): p. 2188-97.

111
24. Arnone, R.D. and J.P. Walling, Waterborne pathogens in urban watersheds. J Water
Health, 2007. 5(1): p. 149-62.
25. USEPA, Microbiological Methods for Monitoring the Environment: Water and Wastes
1978.
26. DRNA, Inventario de Recursos de Agua de Puerto Rico (Draft). 2004, Department of
Natural Resources San Juan, PR
27. USEPA, Method 1600: Membrane Filter Test Method for Enterococci in Water, in EPA-
821-R-02-022. 2002, Office of Water: Washington, D.C.
28. Ludwig, W. and K.H. Schleifer, How quantitative is quantitative PCR with respect to cell
counts? Syst Appl Microbiol, 2000. 23(4): p. 556-62.
29. Bernhard, A.E. and K.G. Field, A PCR assay To discriminate human and ruminant feces
on the basis of host differences in Bacteroides-Prevotella genes encoding 16S rRNA.
Appl Environ Microbiol, 2000. 66(10): p. 4571-4.
30. Bernhard, A.E. and K.G. Field, Identification of nonpoint sources of fecal pollution in
coastal waters by using host-specific 16S ribosomal DNA genetic markers from fecal
anaerobes. Appl Environ Microbiol, 2000. 66(4): p. 1587-94.
31. Lu, J., J. Santo Domingo, and O.C. Shanks, Identification of chicken-specific fecal
microbial sequences using a metagenomic approach. Water Res, 2007. 41(16): p. 3561-
74.
32. Ryu, H., et al., Development and evaluation of a qPCR assay targeting fecal pollution of
sandhill cranes (Grus canadensis). Appl Environ Microbiol, 2012.
33. McCarthy, M.A., Bayesian Methods for Ecology. 2007, Cambridge University Press:
Cambridge, UK.
34. Hoff, P.D., A first course in Bayesian Statistical Methods. 2010, Springer Science
+Business Media: New York, USA.
35. Sivia, D.S. and J. Skilling, Data Analysis: A Bayesian Tutorial. 2011, Oxford University
Press: Oxford, England, UK.
36. Kéry, M., Introduction to WinBugs for ecologist: A bayesian approach to regression,
ANOVA, mixed models and related analysis. 2010, Academic Press: London, U.K.
37. Link, W.A. and R.J. Barker, Bayesian Inference with ecological application. 2010,
Academic Press: London, U.K.

112
38. Kinzelman, J.L., et al., Comparative evaluation of molecular and culture methods for
fecal indicator bacteria for use in inland recreational waters. 2011, Water Environment
Research Foundation. p. 1-360.
39. Yan, T., D.K. Goto, and F. Feng, Concentration dynamics of fecal indicators in Hawaii's
coastal and inland sand , soil, and water during rainfall events. PATH6R09, 2011: p. 1-
76.
40. Walters, S.P., V.P. Gannon, and K.G. Field, Detection of Bacteroidales fecal indicators
and the zoonotic pathogens E. coli 0157:H7, Salmonella, and Campylobacter in river
water. Environ Sci Technol, 2007. 41(6): p. 1856-62.
41. Yoder, J.S., et al., Surveillance for waterborne-disease outbreaks associated with
recreational water--United States, 2001-2002. MMWR Surveill Summ, 2004. 53(8): p.
1-22.
42. Licence, K., et al., An outbreak of E. coli O157 infection with evidence of spread from
animals to man through contamination of a private water supply. Epidemiol Infect, 2001.
126(1): p. 135-8.
43. Sercu, B., et al., Storm drains are sources of human fecal pollution during dry weather in
three urban southern California watersheds. Environ Sci Technol, 2009. 43(2): p. 293-8.
44. Ahmed, W., et al., Evaluation of multiple sewage-associated Bacteroides PCR markers
for sewage pollution tracking. Water Res, 2009. 43(19): p. 4872-7.
45. Ahmed, W., et al., Detection and source identification of faecal pollution in non-sewered
catchment by means of host-specific molecular markers. Water Sci Technol, 2008. 58(3):
p. 579-86.
46. Haugland, R.A., et al., Comparison of Enterococcus measurements in freshwater at two
recreational beaches by quantitative polymerase chain reaction and membrane filter
culture analysis. Water Res, 2005. 39(4): p. 559-68.
47. Harwood, V.J., K.V. Gordon, and C. Staley, Validation of rapid methods for enumeration
of markers for human sewage contamination in recreational waters. 2011.

113
CHAPTER 5
Evaluation of Enterococcus-infecting phages as indeces of fecal pollution
T. M. Santiago-Rodriguez1, P. Marcos
2, S. Monteiro
3, M.Urdaneta
1, R. Santos
3 and G. A.
Toranzos1
1Environmental Microbiology Laboratory, Department of Biology, University of Puerto Rico,
Rico, San Juan, Puerto Rico, 2Biological Sciences Department, General Studies, University of
Puerto Rico, San Juan, Puerto Rico, 3Laboratório de Análises, Instituto Superior Técnico, Lisboa,
Portugal
ABSTRACT
No microbial source tracking tool satisfies all the characteristics of an ideal indicator of human
fecal pollution. For this reason, the potential of Enterococcus faecalis phages (enterophages) as
markers of this type of contamination was tested by using eight Enterococcus type strains as the
possible hosts. The prevalence of enterophages in animal feces and domestic sewage were
determined, as were the inactivation rates in raw sewage at 4 ºC and surface and tap waters at 22
ºC. Enterophages were exclusively detected in raw sewage (up to 66.0 PFU/100 mL), suggesting
a strictly human origin; and exhibited inactivation rates of approximately 0.002 to 0.05, 0.3 to
0.5 and 0.4 to 1.4 log/day in raw sewage and surface and tap waters, respectively, similar to
those of previous reports on human enteric viruses under similar conditions. Interestingly, phages
infecting other Enterococcus type strains were detected in both animal feces and domestic
sewage in concentrations of up to 335.8 and PFU/g and 96.0 PFU/100 mL, and certain phage
isolates infected several of the strains tested. This clearly indicates the possible promiscuous
nature of some Enterococcus phages and thus opens up the opportunity to further characterize
these as indices of specific fecal sources.
Keywords: enterococci, enterophages, fecal pollution, microbial indicators, Microbial Source
Tracking
Reference: Santiago-Rodriguez TM, et al. Evaluation of Enterococcus phages as indeces of
fecal pollution. Journal of Water and Health. 2013. 11(1): p. 51-63.

114
INTRODUCTION
Human pathogens can be introduced into water sources as a result of fecal pollution. Among
these, enteric viruses are of great concern since they: (i) can cause disease with infectious doses
as low as 1 to 10 particles, (ii) affect 5 to 18 million people every year and (iii) can be
resuspended from sediments to surface waters after disturbances [1-4]. Enteric viruses are often
detected by culture and molecular techniques [5-8]. However, culture methods are notoriously
expensive and laborious and noninfectious viral particles or naked viral nucleic acids can be
detected by molecular techniques [9, 10]. Since not all laboratories have the facilities for enteric
virus detection, microbial indicators have been used to infer the presence of these pathogens in
waters and sewage; however, many microbial indicators fail to fulfill the characteristics of a
reliable marker of fecal pollution [11]. For instance, relatively large numbers of both enterococci
and thermotolerant coliforms have been detected in pristine waters and are able to replicate in
these environments [12-14]. Algae and sediments could also be sources of indicator bacteria and
although certain type strains have been linked to fecal pollution, their presence in the feces of
different warm-blooded animals makes them unreliable for microbial source tracking (MST)
purposes [15]. In addition, enteric viruses can be present in waters that meet bacteriological
standards and absent from waters failing to meet this criteria [1].
Bacteriophages have also been proposed as markers of fecal contamination and as models of
human enteric viruses. Such is the case of phages infecting Bacteroides fragilis and Escherichia
coli (coliphages). However, B. fragilis phages have been detected only at certain geographical
regions [16-19] and the techniques used for their detection are relatively difficult due to the strict
anaerobic nature of the bacterial host [20, 21]. Coliphages may not discriminate the source of the
fecal contamination since these have been detected in the feces of different animals [22, 23]. In
addition, survival experiments have been performed with these bacteriophages [20, 24-26].
Coliphages can survive for long periods in pristine waters and may not necessarily indicate
recent fecal pollution [24, 26, 27].
There is a lack of microbial indicators of human fecal pollution. For this reason, we have
proposed phages that infect a specific type strain of Enterococcus faecalis (enterophages) as
markers of human fecal contamination. Most enterophage isolates characterized in our laboratory

115
are tailed phages, with icosahedral capsids measuring between 13 to 80 nm and DNA genomes
>23 kbp [28, 29]. In terms of their reliability as indicators of human fecal pollution,
enterophages: (i) have been detected in raw and treated domestic sewage [28, 29], (ii) possess a
survival similar to that of many enteric viruses in fresh waters [2, 30] and (iii) have not been
detected in cattle feces. Even though previous results have been promising, more data are needed
to fully determine the potential of enterophages as indicators of human fecal contamination. The
present study focused on the use of various Enterococcus species (spp.) as the bacterial hosts,
and fecal material from different animals and domestic sewage were tested for the presence and
prevalence of enterophages. We also determined the inactivation rates and survival times of
enterophage isolates under various conditions, and compared results to those studies using
human enteric viruses. Therefore, the aims of the present study were to: determine the presence
of enterophages in animal feces, their prevalence in raw and treated domestic sewage,
inactivation rates and survival times under various conditions, including raw sewage at 4 ºC,
fresh and tap waters at 22 ºC and various sterile water types at 37 ºC.
MATERIALS AND METHODS
Detection of Enterococcus phages in feces and sewage
Poultry, cattle, pigs and humans are among the most common sources of fecal pollution [31, 32].
For this reason, the presence of enterophages in animal feces and domestic sewage was tested by
using Enterococcus type strains from the American Type Culture Collection (ATCC) and
included: E. faecalis (ATCC 19433), E. faecium, E. gallinarum, E. durans (ATCC 19432), E.
dispar (ATCC 51266), E. hirae (ATCC 8043), E. casseliflavus (ATCC 25788) and E.
pseudoavium (ATCC 49372) as the possible bacterial hosts. In addition, the presence of
Enterococcus phages in dog feces was tested since close contact with humans and lateral
transmission of microorganisms can occur [33]. Samples were also processed for coliphages
using E. coli C3000 (ATCC 15597) as the bacterial host as a means of comparison. Composites
of the fecal samples were processed by using the single layer method: Briefly, one gram of
chicken (n=30), cattle (n=30), pig (n=10) and dog (n=12) feces were eluted in 50 mL of a saline
solution (0.85% w/v) per gram of feces and mixed with an equal amount of liquefied media as
described previously [29]. In order to detect the possible presence of different bacteriophage
thermal groups, the mixture was poured into four Petri dishes and incubated at 22, 37 and 41 °C

116
and viral plaques were enumerated at 24 h. The presence of Enterococcus phages was tested in
raw sewage and primary effluent from three wastewater treatment plants (WTP) in Puerto Rico.
Samples were collected and processed monthly from August 2010 to August 2011 using the
single layer method with a modification [28]. Sixty-mL aliquots were added to equal amounts of
molten media and processed individually by using the Enterococcus type strains described above
as the bacterial hosts. Petri dishes were incubated at 22, 37 and 41 °C, viral plaques were
enumerated at 24 h and the percent removal of each phage group was calculated.
Isolation and characterization of Enterococcus phages
Individual viral plaques were isolated as described previously [28]. Several of the isolates were
tested against different Enterococcus type strains using a spot test to determine the host
specificity and replication temperature. Briefly, 1 μL of the bacteriophage isolate was placed on
top of a Petri dish containing Trypticase Soy Broth (TSB) (Difco), agar (1.5% w/v), CaCl2·2H2O
(Fisher Scientific Co. NJ, USA) and NaN3 (MCB, OH, USA) (final concentration of 2.6 mg/mL
and 0.4 mg/mL, respectively) and a lawn of the bacteria tested. A total of three dishes were
incubated at 22, 37 or 41 °C for 24h. In addition, the morphology of two E. faecalis phages was
determined by using Transmission Electron Microscopy, as described previously [28].
Inactivation rates and survival studies
Current regulations require that water and sewage samples be processed within 6 h and stored at
4-7 °C prior to analysis. To test if storage time could affect the detection of Enterococcus
phages, we determined the inactivation rates of phages infecting E. faecalis, E. casseliflavus, E.
faecium and E. coli in raw sewage. One-liter of raw domestic sewage was seeded with a
concentration of 104
plaque forming units (PFU)/100 mL from prepared laboratory stocks and
kept at 4-7 ºC. Aliquots were obtained daily and processed using the single layer method as
indicated above. To simulate the survival of enterophages and coliphages under environmental
conditions, raw domestic sewage was added to three fresh water samples collected from the Rio
Grande de Arecibo watershed in Puerto Rico [34, 35]. Experiments were performed during a
period of low and higher rainfall events and included: (i) the highest and thus one of the less
polluted sites of the watershed (site 1), (ii) immediately after a WTP (site 2), since it represents a
point-source of fecal contamination and (iii) the estuary (site 3). The survival experiments were

117
conducted as described above and seeded samples were kept at 22 °C, average temperature of
waters in Puerto Rico [29]. All precipitation data was obtained from the US Geological Survey
(USGS) Caribbean Water Science Center, stations 50020100, 50024950 and 50021700
(http://pr.water.usgs.gov/). Similarly, for the survival of enterophages and coliphages in tap
water, 2 L were collected in a sterile container and seeded with enterophages and coliphages and
aliquots were processed as described [28]. In order to determine the possible effect of chlorine in
the survival of enterophages and coliphages, one of the samples was dechlorinated using sodium
thiosulfate (final concentration of 10.0 mg/L) and kept at 22 °C. Similarly, survival experiments
with an enterophage isolate were performed at 37 °C (the optimal growth temperature of the host
and the isolation temperature of the enterophage tested) in sterile tap, distilled and wastewater, as
described previously.
Results for all inactivation rates and survival experiments were plotted as semi-log plots of time
(days) versus bacteriophage PFU/100 mL. Decay constants (kd) were calculated by using the
slopes of the linear regressions of the semi-log plots (-log10 PFU·day-1
) [29, 36]. T90 values, or
the time to reach a 90 % reduction in PFU densities, were calculated as ln (0.1)/kd [34, 37, 38].
Statistical analyses
Non-parametric one-way analyses of variances (Kruskal-Wallis) were used to determine
differences in the prevalence of Enterococcus phages in raw sewage as influenced by the
bacterial host tested and incubation temperature. The same analyses were used to determine
differences in the inactivation rates of the Enterococcus phages (E. faecalis, E. faecium and E.
casseliflavus) and coliphages in raw sewage at 4 ºC [23, 39]. For the survival experiments in
fresh waters, the analyses were used to determine differences in the inactivation rate of
enterophages and coliphages as influenced by the incubation temperature and sampled site.
Statistical analyses were performed using the R statistical software (version 2.11.1) [40].
RESULTS
Enterococcus phages in animal feces and domestic sewage
Interestingly, phages infecting E. faecium, E. casseliflavus and E. pseudoavium were detected in
chicken feces at 37 °C (Table 1). Phages infecting E. gallinarum, E. durans, E. dispar and E.

118
hirae were not detected in chicken, cattle or pig feces. Coliphages were detected in chicken
(1.0±0.0 PFU/g) and cattle feces at 22 and 37 °C (40.3±28.0 and 335.81±4.0 PFU/g,
respectively) and in dog feces at 41 °C (120.79±29.45 PFU/g). Neither coliphages nor
Enterococcus phages were detected in pig feces. Enterophages were detected at all temperatures
tested in raw sewage collected from all WTP and at 22 and 37 °C in the primary effluents from
two WTP. Interestingly, E. faecium-infecting phages were also detected in raw and treated
sewage from two WTP at all temperatures tested.
Host
Source E. coli E. faecalis E. faecium E. casseliflavus E. pseudoavium
Chicken 1.0±0.0a ND 107.5±10.6
b 65.0±0.0
b 3.8±1.8
b
Cattle 40.3±28.0a ND ND ND ND
Dogs
335.81±4.0b,
120.79.±29.45c ND ND ND ND
Pigs ND ND ND ND ND
ND=Not detected
a=22 ºC
b=37 ºC
c=41 ºC
Table 1: Enterococcus and E. coli-infecting phages per gram of feces in chicken (n=30), cattle
(n=30), dogs (n=12) and pigs (n=10). Enterococcus spp, included: E. faecalis, E. faecium, E.
gallinarum, E. hirae, E. durans, E. dispar, E. casseliflavus and E. pseudoavium. Only hosts
exhibiting positive results in at least one fecal source were included in the table.
Higher concentrations of both E. faecalis and E. faecium phages were detected in domestic
sewage compared to other enterococci phages and exhibited removal percents of 37.0 to ≥ 99.0
% and 47.0 to ≥ 99.0 %, respectively. Other Enterococcus phages exhibited various removal
percents depending on the WTP and incubation temperature (Table 2). Phages infecting E.
casseliflavus were detected at 22 and 37 °C in raw sewage collected from all WTP and at 22 and
37 °C in the primary effluents. Enterococcus pseudoavium-infecting phages were also detected at
22 °C in raw sewage from all WTP and at 37 °C in two WTP, but were not detected in the
primary effluents.

119
PR-A PR-B
Host 22ºC 37 ºC 41 ºC 22 ºC 37 ºC 41 ºC
E. faecalis 37.0±44.0 44.0±33.0 ≥99.0±0.9 83.0±16.8 80.0±15.9 ≥99.0±0.09
E. faecium 63.0±16.0 86.0±5.8 ND 47.0±42.0 60.0±37.0 ≥99.0±0.09
E. gallinarum ND ND ND ND ND ND
E. hirae ND ND ND ≥99.0±0.09 ≥99.0±0.9 ND
E. durans ND ND ND ≥99.0±0.09 ≥99.0±0.09 ND
E. dispar ≥99.0±0.09 ND ND 48.0±25.2 ≥99.0±0.45 ≥99.0±0.09
E. casseliflavus 27.0±32.4 ≥99.0±0.9 ND 81.0±12.1 ≥99.0±0.09 ≥99.0±0.09
E. pseudoavium ≥99.0±0.09 ≥99.0±0.9 ND 87.0±1.0 ≥99.0±0.09 ≥99.0±0.09
PR-C
Host 22 ºC 37 ºC 41 ºC
E. faecalis ≥99.0±0.09 ≥99.0±0.09 ≥99.0±0.09
E. faecium ≥99.0±0.09 ≥99.0±0.09 ≥99.0±0.09
E. gallinarum ND ND ND
E. hirae ≥99.0±0.09 ND ND
E. durans ND ND ND
E. dispar ≥99.0±0.09 ND ND
E. casseliflavus 73.0±20.4 ≥99.0±0.09 ND
E. pseudoavium ≥99.0±0.09 ND ND
Table 2: Mean removal percents of Enterococcus phages in primary effluent from three WTP
(PR-A, PR-B and PR-C) in Puerto Rico. Numbers were calculated for different incubation
temperatures, namely 22, 37 and 41 °C. Phages infecting several Enterococcus were not detected
(ND).
Other phages detected in raw sewage were those infecting E. hirae, E. durans and E. dispar, but
these were not constantly detected in primary effluent (Figure 1).

120
(A)
(B)
-1.00
-0.50
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
faecalis faecium gallinarum hirae durans dispar casseliflavus pseudoaviumBacte
rio
phage P
FU
/10
0m
L(L
og
10)
Enterococcus host
22ºC
-1.00
-0.50
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
faecalis faecium gallinarum hirae durans dispar casseliflavus pseudoaviumBacte
rio
phage P
FU
/10
0m
L(L
og
10)
Enterococcus host
37ºC

121
(C)
Figure 1: Prevalence of Enterococcus phages in raw domestic sewage in three wastewater
treatment plants (WTP) in Puerto Rico. Petri dishes were incubated at 22 (A), 37 (B) and 41°C
(C) and each column represent a WTP. Enterococcus spp included E. faecalis, E. faecium, E.
gallinarum, E. hirae, E. durans, E. dispar, E. casseliflavus and E. pseudoavium. Results
represent the log transformed geometric mean of n=12 samples and standard deviations are
represented by error bars.
Replication and morphology of Enterococcus-infecting phages
Most of the Enterococcus phage isolates in this study did not infect a specific bacterial host, with
the exception of those infecting E. faecalis, which replicated at all three replication temperatures
tested as shown previously [29]. Table 3 shows the host range and replication temperature of the
tested phages. In terms of the morphology of the E. faecalis phages in this study, these exhibited
long non-contractile tails of approximately 180 nm and icosahedral capsids of approximately 80
nm (Figure 2). This morphology is similar to the enterophages characterized previously [29].
-1.50
-1.00
-0.50
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
faecalis faecium gallinarum hirae durans dispar casseliflavus pseudoavium
Bact
erio
phag
e P
FU
/10
0m
L (L
og
10)
Enterococcus host
41ºC

122
a=22 ºC
b=37 ºC
c=41 ºC
Table 3: Host range of Enterococcus-infecting phages at different incubation temperatures.
Table shows the source of the isolated phages and the Enterococcus type strains used. Stocks
tested contained only one phage type and were tested against other Enterococcus spp., as shown
in the following columns.
Figure 2: Enterococcus faecalis-infecting phages isolated from domestic sewage. Isolates
exhibited long non-contractile tails of approximately 200nm and icosahedral capsids of 80nm.
Bar=100nm.
Host
Range
Source
Host for
Isolation
E.
faecalis
E.
faecium
E.
gallinarum
E.
hirae
E.
durans
E.
dispar
E.
casseliflavus
E.
pdeudoavium
Sewage E. faecalis + a,b,c
- - - - - - -
E. faecium + a,b,c
+ a,b
- - - + a,b
- + a,b
E. hirae - - - + a,b,c
- + c - +
b,c
E. dispar + a,b,c
- - - - + b,c
- + a,b
E. casseliflavus - - - + b,c
- + b +
a +
a,b,c
E. pseudoavium + b - - +
c - +
b - +
a,b
Fresh
water E. faecalis + a,b,c
- - - - - - -
E. faecium - + b - - - - - +
a,b
Poultry E. faecium - + b - - - +
a,b - +
a,b
E. casseliflavus - + a,b
+ a,b,c
+ a,b,c
+ a,b,c
+ a,b,c
+ a,b,c
+ a,b,c
E. pseudoavium - + a,b,c
+ a,b,c
+ a,b,c
+ a +
a,b,c - +
a,b,c

123
Inactivation rates and survival of Enterococcus phages
The initial titer of E. faecalis, E. faecium, E. casseliflavus and E. coli phages in raw sewage at 4
ºC remained constant for 6 days. Calculated decays for E. faecalis phages were 0.050 ± 0.030,
0.030 ± 0.030 and 0.080 ± 0.030 log·day-1
at 22, 37 and 41 °C, respectively. Enterococcus
faecium phages exhibited a decay of 0.0040 ± 0.0020 and 0.0020 ± 0.0020 log·day-1
at 22 and 37
°C, respectively. For this specific E. faecium phage isolate, 41 °C was inhibitory. Similarly, the
E. casseliflavus phage isolate in these experiments was able to replicate only at 22 °C and
exhibited a decay of 0.020 ± 0.030 log·day-1
. Coliphages exhibited decays of 0.010 ±0.020,
0.040 ±0.020 and 0.020 ± 0.050 log·day-1
at 22, 37 and 41 °C, respectively.
Survival of enterophages and coliphages in fresh waters
During the period of low rainfall events, up to 1.01 mm of rain was reported at site 1and no
precipitation was reported at sites 2 or 3 24 h prior to collecting the samples. During this period,
enterophages exhibited an average decay of 0.47 ± 0.03 log·day-1
at all sites. Coliphages
exhibited an average decay of 0.47 ± 0.03 log·day-1
at sites 1 and 2 and 0.33 ± 0.07 log·day-1
at
site 3. During the period of higher rainfall events, up to 2.54 mm of rain were reported at sites 1
and 3 and 0.20 mm were reported at site 2. During this period, enterophages exhibited an average
decay of 0.40 ± 0.00 and 0.30 ± 0.00 log·day-1
at sites 1 and 2 and 0.37 ± 0.03 log·day-1
at site 3.
Coliphages exhibited and average decay of 0.19 ± 0.12, 0.08 ± 0.03 and 0.20 ± 0.10 log·day-1
at
sites 1, 2 and 3, respectively. Calculated T90 values for enterophages and coliphages differed
during the period of low and high rainfall and across the incubation temperatures as well (Table
4). The survival of enterophages and coliphages during both rainfall periods are shown in Figure
3.

124
Table 4: T90 values for enterophages and coliphages in three fresh water samples. Samples were
collected from the Rio Grande de Arecibo watershed in Puerto Rico during a period of low and
higher rainfall events. Sites 1, 2 and 3 correspond to the unpolluted site, the point-source of fecal
pollution and the estuary, respectively.
(A)
Enterophages
Low rainfall High rainfall
Site 22°C 37°C 41°C 22°C 37°C 41°C
1 4.8±0.8 4.8±0.8 3.7±1.0 4.0±1.0 3.1±0.7 2.9±0.5
2 4.6±0.3 4.6±0.2 5.2±0.4 7.2±2.3 7.2±2.2 6.6±1.7
3 3.9±1.4 3.2±0.6 3.2±0.9 5.3±0.9 5.3±1.0 4.8±1.5
Coliphages
Low rainfall High rainfall
Site 22°C 37°C 41°C 22°C 37°C 41°C
1 5.2±0.5 4.6±1.4 4.7±0.9 32.2±2.6 10.3±1.3 6.9±0.9
2 5.1±0.002 5.0±1.4 4.6±0.7 45.3±9.2 22.2±13.0 25.0±5.0
3 8.1±4.1 7.0±2.2 5.8±3.3 24.0±15.2 12.2±5.0 6.5±2.0
-1.50
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
Enterophages: None or low rainfall

125
(B)
(C)
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
Enterophages: Higher rainfall
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og 1
0)
Time (days)
1
2
3
Coliphages: None or low rainfall

126
(D)
Figure 3: Survival of enterophages and coliphages across a tropical watershed in Puerto Rico.
Sampled sites included the highest site (1), after a waste treatment plant (2) and the estuary (3).
Samples were collected during a period of low rainfall events and processed for the detection of
enterophages (A) and coliphages (B). The survival of enterophages (C) and coliphages (D)
during a period of higher rainfall events was also determined. Data represent those phages that
replicated at 37 ºC.
Enterophages in chlorinated and dechlorinated tap water and sterile distilled, tap and
wastewater
The enterophage isolate in this study survived approximately 12 days in chlorinated tap water
with an initial free chlorine concentration of 0.13 ± 0.02 ppm and more than 12 days in
dechlorinated tap water (data not shown). These exhibited decays of 0.23 ± 0.16, 1.05 ± 0.72 and
1.42 ± 0.75 log·day-1
in chlorinated tap water and 0.14 ± 0.045, 0.70 ± 0.38 and 1.02 ± 0.12
log·day-1
in dechlorinated tap water at 22, 37 and 41 ºC, respectively. Coliphages survived for
more than 12 days in both chlorinated and dechlorinated tap water (data not shown) and
exhibited a decay of 0.60 ± 0.0062, 0.83 ± 0.26 and 1.10 ± 0.20 log·day-1
in chlorinated tap water
at 22, 37 and 41 ºC, respectively. In dechlorinated tap water, coliphages showed a decay of 0.13
± 0.045, 0.70 ± 0.38 and 1.02 ± 0.12 log·day-1
at 22, 37 and 41 ºC. Calculated T90 values for
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
Coliphages: Higher rainfall

127
enterophages and coliphages in both chlorinated and dechlorinated tap water are described in
Table 5. In terms of the survival experiments with the enterococci phage isolate at 37 ºC, kd
values for wastewater, tap and distilled water were 0.038, 0.36 and 0.37 log·day-1
, respectively.
A decay of approximately 2 log10 was observed when testing sterile wastewater during 43 days
(Figure 4A). On the other hand, when testing sterile tap and distilled water, enterophages
exhibited a notable decay, surviving approximately 11 days (Figure 4B and C). T90 values were
approximately 6 days for both tap and distilled water and 60 days for wastewater.
Table 5: T90 values (days) for enterophages and coliphages in chlorinated and dechlorinated
drinking water.
Figure 4: Survival of enterophages in various sterile water types at 37 ºC. Samples include (A)
wastewater, (B) tap (chlorinated) and (C) distilled waters and standard deviation is represented
by error bars.
Enterophages Coliphages
Temperature
(°C) Chlorinated Dechlorinated Chlorinated Dechlorinated
22 9.4±7.0 18.1±7.7 18.1±7.7 39.1±4.3
37 8.9±7.0 28.3±6.0 32.1±6.0 43.3±8.3
41 6.9±1.7 28.3±3.7 28.3±3.7 33.0±3.9
(A) (B)
(C)

128
DISCUSSION
Enterococcus-infecting phages in feces and domestic sewage
The source (human) and host-specificity exhibited by enterophages suggest that the E. faecalis
strain in this study may detect human-specific phages. The relative wide range of Enterococcus
spp. infected by specific phages showed that these have a promiscuous nature, as shown with the
coliphages elsewhere [41]. This in turn suggests that future studies are needed in order to
confirm if Enterococcus phages may recognize receptors which are shared by the enterococci
tested in the present study. Several of the Enterococcus strains tested also exhibited immunity to
certain phages, suggesting the presence of prophages, prophage remnants or Clustered Regularly
Interspaced Short Palindromic Repeats (CRISPR) [42, 43]. It is also possible that results may be
influenced by the culture media. Host range and replication temperature may be used to group
enterococci phages as those of an animal or human origin; this in turn suggests their potential for
MST purposes [44]. Results also suggest that humans are reservoirs of various enterococci phage
groups, as seen by the influence of the bacterial host used (x2 = 27.50, df = 6, p = 0.00012) and
temperature tested (x2 = 24.27, df = 2, p = 5.37 e-06). In addition, given that there is the need of
characterizing markers of chicken-fecal pollution, enterococci phages detected in chicken fecal
matter could be further characterized and tested in various water types impacted by this source of
fecal contamination. Future studies need to determine potential hosts for the detection of
Enterococcus phages in other animal feces.
In terms of the coliphages, previous studies have shown that their concentrations vary according
to each individual and can range from 100 to 10
7 PFU/g in cows, pigs and humans and < 10
PFU/g in dog feces [45, 46]. In the present study, coliphage concentrations in cows and humans
are within the range reported previously, but higher concentrations were detected in dog feces.
Although coliphages were not detected in pig feces in the present study, 0 to < 10 PFU/g have
been reported elsewhere [45].
Inactivation rates and survival of Enterococcus phages
Our results showed that titers of E. faecalis, E. faecium, E. casseliflavus and E. coli phages do
not decrease significantly at 4 °C (~0.002 to 0.05 log·day-1
, depending on the bacterial host),

129
suggesting that samples may be stored longer at refrigeration temperatures prior to analyses.
Similar outcomes have been reported for adenovirus 40 and 41 and poliovirus 1 in sewage at 4
ºC, which have exhibited a decrease of 2.5, 2.0 and 2.2 log after 50 days [47]. In fresh waters, the
similar replication of enterophages at all temperatures tested suggests that a group predominated
in the analyses. Unlike enterophages, coliphages exhibited differences in their replication at
different temperatures (x2 = 11.46, df = 2, p = 0.0033), suggesting that various groups were
present, as previously suggested [48]. In terms of the inactivation rates, enterophages did not
exhibit differences across the sampled sites during the period of low or high rainfall events. This
is comparable with the inactivation rates of poliovirus, coxsackievirus and rotaviruses SA11,
which exhibit similar inactivation rates in unpolluted and polluted sites (~ 0.5 to 1.0 log·day-1
)
[27]. Similarly, coliphages did not show differences in their inactivation rates across the sampled
sites during the period of low rainfall, but this was not the case for the period of higher rainfall
events (x2
= 10.11, df = 2, p = 0.0064). This is comparable with echovirus 7, which exhibits
different inactivation rates in unpolluted (1.0 log·day-1
) and polluted sites (0.5 log·day-1
) [27].
Although there are no hard data that may explain this, several factors may be involved in these
differences in survival times during both rainfall periods including pH changes, dilution of
proteolytic enzymes and antiviral chemicals, as described elsewhere [49-51]. Another possibility
is that due to the relatively long survival times of coliphages in surface waters (20 to 160 days)
and sediments (30 days) at 20 ºC, it is reasonable to believe that the opportunity to attach to
sediment particles is greater [26, 35]. Once attached, coliphages may be desorbed after rainfall
events, and thus their numbers may be higher. In terms of the estuary, polioviruses,
coxsackieviruses, rotaviruses SA11 and echoviruses can survive approximately 3 days in
estuaries, similar to enterophages during the period of low rainfall events in the present study,
but exhibit a more rapid decay compared to both enterophages and coliphages (~ 0.5 to 2.5
log·day-1
) [27].
Survival of the tested bacteriophages may also be influenced by chlorine and the environmental
microbiota. Few studies have determined the survival of enteric viruses in waters with chlorine
levels similar to those found in tap water [52]. Therefore, comparing the survival of enterophages
and coliphages with that of enteric viruses in chlorinated tap water is relatively difficult. Certain
enteric viruses can survive up to 30 minutes while others can survive 16 h and exhibit decays of

130
~ 1 log·h-1
in waters with free chlorine levels of 0.10 ppm at 22 ºC [53]. This suggests that
enteric viruses are more susceptible to chlorine than both enterophages and coliphages, which
exhibited T90 values of ~7 to 9 and18 to 32 days, and decays of ~ 0.20 to 1.40 and 0.60 to 1.10
log·day-1
, respectively. This in turn suggests that chlorine may have a more visible effect on the
survival of enterophages compared to coliphages. In dechlorinated tap water, T90 values for
enterophages detected at 37 and 41 ºC were similar to that of Poliovirus 1 at 23 °C
(approximately 30 days); however, neither the enterophages nor the coliphages possess a
survival similar to that of adenoviruses 40 and 41, under similar conditions [47]. These studies,
however, did not consider the effect of the environmental microbiota. Results from using
different sterile water types in the present study suggest that the absence of other microorganisms
enhances the survival of enterophages; therefore, future studies should consider the
environmental microbiota in the survival of enteric viruses and bacteriophages in various water
types.
CONCLUSIONS
This is one of the few studies in which the potential of enterophages as markers of human fecal
pollution has been further tested. The presence of enterophages in chicken, cattle, dog and pig
fecal materials and their prevalence in domestic sewage were determined by testing various
Enterococcus spp. as the bacterial hosts. This, however, may represent one limitation to the
present study as phages infecting other Enterococcus spp. may also be present in fecal material
and fecally contaminated waters and thus future studies are needed in order to determine this.
Humans and animals are reservoirs of Enterococcus phages and those specifically infecting E.
faecalis seem promising indeces of human fecal pollution. The present study also determined the
inactivation rates and survival times of enterophages and compared results with those of human
enteric viruses reported elsewhere under similar conditions; however, the present studies must
also be performed in situ. Enterophages could be considered models of certain human enteric
viruses in various water types since their inactivation rates and survival times are similar to those
reported previously. Phages infecting E. faecium, E. casseliflavus and E. pseudoavium
replicating at 37 °C may be used to infer the presence of chicken-fecal matter in fresh water
sources, but more studies need to determine if these phages are present in waters impacted by
chicken-fecal matter. The present study opens the possibility to further characterize

131
Enterococcus phages as indeces of specific fecal sources, especially in various geographical
regions and to predict their behavior in different areas by using specific mathematical models,
for example. Also, future studies need to develop the techniques for the detection of
enterophages using molecular methods and compare results with culture techniques.
ACKNOWLEDGMENTS
We thank Carlos Toledo-Hernandez, Cruz Minerva Ortiz of the Autoridad de Acueductos y
Alcantarillados for collecting the fresh water and sewage samples. We also thank Jorge W.
Santo-Domingo for providing the Enterococcus spp. type strains and the USGS for the
precipitation data. This research was partially supported by MBRS-RISE, NIH Grant Number
2R25GM061151-09, and the Center for Renewable Energy and Sustainability with a grant from
the Department of Defense to the University of Puerto Rico.
SUPPLEMENTARY INFORMATION
Decay (%)·day
-1 T90 (days)
Host 22°C 37°C 41°C 22°C 37°C 41°C
E. faecalis 4.7±3.1 3.4±2.7 8.3±3.6 49.0±29.7 67.1±154.3 27.8±13.8
E. faecium 0.4±0.2 0.2±0.2 N/D 579.0±300.5 1158.8±712.0 N/D
E. casseliflavus 1.9±2.7 N/D N/D 121.8±59.3 N/D N/D
E. coli 1.0±1.5 4.1±2.0 2.1±5.3 224.9±120.8 55.6±49.1 111.0±82.4
Supplementary Table 1: Average of the decay percent day -1
and T90 values for phages infecting
E. faecalis, E. faecium, E. casseliflavus and E. coli in raw sewage at 4°C. Both values were
calculated for phages detected at 22, 37 and 41 °C. Not all phages were detected at all incubation
temperatures and thus this is represented by N/D (not detected).

132
Enterophages
Low rainfall High rainfall
Site 22°C 37°C 41°C 22°C 37°C 41°C
1 0.5±0.04 0.5±0.04 0.4±0.06 0.4±0.1 0.4±0.1 0.4±0.1
2 0.5±0.03 0.5±0.02 0.5±0.02 0.3±0.1 0.3±0.1 0.3±0.1
3 0.4±0.006 0.5±0.02 0.5±0.02 0.4±0.1 0.4±0.2 0.3±0.1
Coliphages
Low rainfall High rainfall
Site 22°C 37°C 41°C 22°C 37°C 41°C
1 0.4±0.07 0.5±0.1 0.5±0.1 0.07±0.004 0.2±0.03 0.3±0.03
2 0.5±0.02 0.5±0.1 0.5±0.04 0.05±0.01 0.1±0.04 0.09±0.02
3 0.3±0.2 0.3±0.1 0.4±0.1 0.1±0.04 0.2±0.06 0.3±0.05
Supplementary Table 2: Decay values (log·day-1
) for enterophages and coliphages in three
fresh water samples. Samples were collected from the Rio Grande de Arecibo watershed in
Puerto Rico during a period of low and higher rainfall events. Sites 1, 2 and 3 correspond to the
unpolluted site, the point-source of fecal pollution and the estuary, respectively.
(A)
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
22ºC

133
(B)
(C)
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
37ºC
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
41ºC
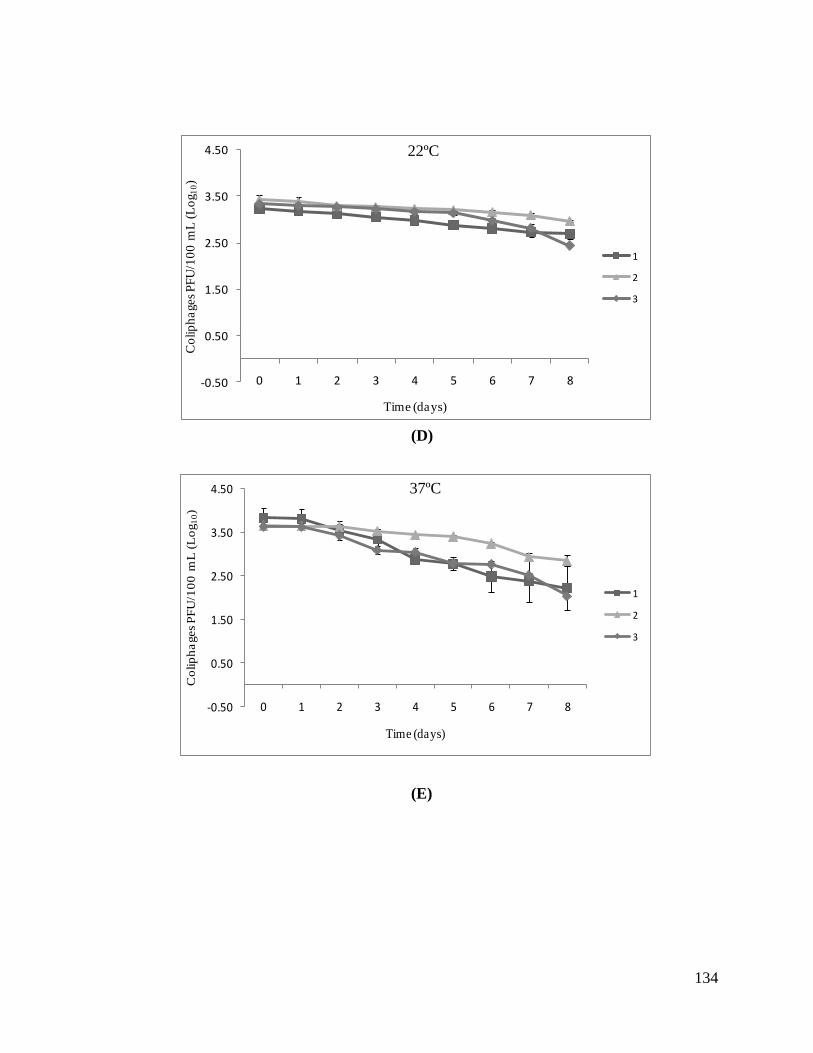
134
(D)
(E)
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og 1
0)
Time (days)
1
2
3
22ºC
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
37ºC

135
(F)
Supplementary Figure 1: Survival of enterophages and coliphages in three sites in a watershed.
Samples were collected during a period of high rainfall events and included: a relatively
unpolluted site (1), after a WTP (2) and the estuary (3). Panels A, B and C correspond to the
survival of enterophages detected at 22, 37 and 41°C, respectively. Panels D, E and F correspond
to the coliphages detected at 22, 37 and 41°C, respectively. Results represent the average of three
replicas and standard deviations are represented by error bars.
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og 1
0)
Time (days)
1
2
3
41ºC
-1.50
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
22ºC

136
(A)
(B)
(C)
-1.50
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
37ºC
-1.50
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8En
tero
ph
ages
PF
U/1
00
mL
(L
og
10)
Time (days)
1
2
3
41ºC

137
(D)
(E)
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og 1
0)
Time (days)
1
2
3
22ºC
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og 1
0)
Time (days)
1
2
3
37ºC

138
(F)
Supplementary Figure 2: Survival of enterophages and coliphages in three sites in a tropical
watershed. Samples were collected during a period of low rainfall and include a relatively
unpolluted site (1), after a WTP (2) and the estuary (3). Panels A, B and C correspond to the
survival of enterophages detected at 22, 37 and 41°C, respectively. Panels D, E and F correspond
to the coliphages detected at 22, 37 and 41°C, respectively. Results represent the average of three
replicas and standard deviations are represented by error bars.
-0.50
0.50
1.50
2.50
3.50
4.50
0 1 2 3 4 5 6 7 8
Co
lip
ha
ges
PF
U/1
00
mL
(L
og 1
0)
Time (days)
1
2
3
41ºC

139
LITERATURE CITED
1. Bosch, A., Human enteric viruses in the water environment: a minireview. Int Microbiol,
1998. 1(3): p. 191-6.
2. Rao, V.C., et al., Isolation of enteroviruses from water, suspended solids, and sediments
from Galveston Bay: survival of poliovirus and rotavirus adsorbed to sediments. Appl
Environ Microbiol, 1984. 48(2): p. 404-9.
3. Rose, J.B., et al., Climate variability and change in the United States: potential impacts
on water- and foodborne diseases caused by microbiologic agents. Environ Health
Perspect, 2001. 109 Suppl 2: p. 211-21.
4. Lipp, E.K., et al., The Effects of Seasonal Variability and Weather on Microbial Fecal
Pollution and Enteric Pathogens in a Subtropical Estuary. Estuaries, 2001. 24(2): p. 266-
276.
5. Fout, G.S., et al., A multiplex reverse transcription-PCR method for detection of human
enteric viruses in groundwater. Appl Environ Microbiol, 2003. 69(6): p. 3158-64.
6. Jaykus, L.A., R. De Leon, and M.D. Sobsey, A virion concentration method for detection
of human enteric viruses in oysters by PCR and oligoprobe hybridization. Appl Environ
Microbiol, 1996. 62(6): p. 2074-80.
7. Schwab, K.J., R. De Leon, and M.D. Sobsey, Immunoaffinity concentration and
purification of waterborne enteric viruses for detection by reverse transcriptase PCR.
Appl Environ Microbiol, 1996. 62(6): p. 2086-94.
8. Greening, G.E., L. Woodfield, and G.D. Lewis, RT-PCR and chemiluminescent ELISA
for detection of enteroviruses. J Virol Methods, 1999. 82(2): p. 157-66.
9. Schwab, K.J., et al., Distribution of Norwalk virus within shellfish following
bioaccumulation and subsequent depuration by detection using RT-PCR. J Food Prot,
1998. 61(12): p. 1674-80.
10. Leclerc, H., et al., Bacteriophages as indicators of enteric viruses and public health risk
in groundwaters. J Appl Microbiol, 2000. 88(1): p. 5-21.
11. Havelaar, A.H., M. van Olphen, and Y.C. Drost, F-specific RNA bacteriophages are
adequate model organisms for enteric viruses in fresh water. Appl Environ Microbiol,
1993. 59(9): p. 2956-62.

140
12. Byappanahalli, M.N., et al., Growth and survival of Escherichia coli and enterococci
populations in the macro-alga Cladophora (Chlorophyta). FEMS Microbiol Ecol, 2003.
46(2): p. 203-11.
13. Toranzos, G.A., et al., Detection of microorganisms in environmental freshwaters and
drinking waters, in Manual of Environmental Microbiology, R.L. Crawford, Editor.
1996, ASM Press: Washington, D.C. p. 249-260.
14. Rivera, S.C., T.C. Hazen, and G.A. Toranzos, Isolation of fecal coliforms from pristine
sites in a tropical rain forest. Appl Environ Microbiol, 1988. 54(2): p. 513-517.
15. Baele, M., Composition of enterococcal and streptococcal flora from pigeon intestines.
Applied and Environmental Microbiology, 2002. 92(2): p. 348-51.
16. Scott, T.M., et al., Microbial source tracking: current methodology and future directions.
Appl Environ Microbiol, 2002. 68(12): p. 5796-803.
17. Allsop, K. and D.J. Stickler, An assessment of Bacteroides fragilis group organisms as
indicators of human faecal pollution. J Appl Bacteriol, 1985. 58(1): p. 95-9.
18. Gantzer, C., J. Henny, and L. Schwartzbrod, Bacteroides fragilis and Escherichia coli
bacteriophages in human faeces. Int J Hyg Environ Health, 2002. 205(4): p. 325-8.
19. Puig, A., J. Jofre, and R. Araujo, Bacteriophages infecting various Bacteroides fragilis
strains differ in their capacity to distinguish human from fecal pollution. Water Sci
Technol, 1997. 35(11-12): p. 359-362.
20. Gantzer, C., et al., Detection of infectious enteroviruses, enterovirus genomes, somatic
coliphages, and Bacteroides fragilis phages in treated wastewater. Appl Environ
Microbiol, 1998. 64(11): p. 4307-12.
21. Weisberg, S.B., R.T. Noble, and J.F. Griffith, Microbial Indicators of Marine
Recreational Water Quality, in Manual of Environmental Microbiology, R.L. Crawford,
Editor. 1996, ASM: Washington, D.C. p. 280-287.
22. Cole, D., S.C. Long, and M.D. Sobsey, Evaluation of F+ RNA and DNA coliphages as
source-specific indicators of fecal contamination in surface waters. Appl Environ
Microbiol, 2003. 69(11): p. 6507-14.
23. Brion, G.M., J.S. Meschke, and M.D. Sobsey, F-specific RNA coliphages: occurrence,
types, and survival in natural waters. Water Res, 2002. 36(9): p. 2419-25.

141
24. Hernandez-Delgado, E.A. and G.A. Toranzos, In situ replication studies of somatic and
male-specific coliphages in a tropical pristine river. Wat Sci Tech, 1995. 31(5-6): p. 247-
250.
25. Grabow, W.O. and P. Coubrough, Practical direct plaque assay for coliphages in 100-ml
samples of drinking water. Appl Environ Microbiol, 1986. 52(3): p. 430-3.
26. Long, S.C. and M.D. Sobsey, A comparison of the survival of F+RNA and F+DNA
coliphages in lake water microcosms. J Water Health, 2004. 2(1): p. 15-22.
27. Hurst, C.J. and C.P. Gerba, Stability of Simian Rotavirus in Fresh and Estuarine Water.
Appl Environ Microbiol, 1980. 39(1): p. 1-5.
28. Bonilla, N., et al., Enterophages, a group of phages infecting Enterococcus faecalis, and
their potential as alternate indicators of human faecal contamination. Water Sci Technol,
2010. 61(2): p. 293-300.
29. Santiago-Rodriguez, T.M., et al., Characterization of Enterococcus faecalis-infecting
phages (enterophages) as markers of human fecal pollution in recreational waters. Water
Res, 2010. 44(16): p. 4716-25.
30. Ward, R.L., D.R. Knowlton, and P.E. Winston, Mechanism of inactivation of enteric
viruses in fresh water. Appl Environ Microbiol, 1986. 52(3): p. 450-9.
31. USEPA, Microbial Source Tracking Guide Document in EPA/600-R-05-064 2005:
Cincinnati, OH.
32. Ahmed, W., et al., Real-time PCR detection of pathogenic microorganisms in roof-
harvested rainwater in Southeast Queensland, Australia. Appl Environ Microbiol, 2008.
74(17): p. 5490-6.
33. Guardabassi, L., S. Schwarz, and D.H. Lloyd, Pet animals as reservoirs of antimicrobial-
resistant bacteria. Journal of Antimicrobial Chemotherapy, 2004. 54: p. 321-332.
34. Noble, R.T., I.M. Lee, and K.C. Schiff, Inactivation of indicator micro-organisms from
various sources of faecal contamination in seawater and freshwater. J Appl Microbiol,
2003. 96: p. 464-472.
35. Duran, A.E., et al., Removal and inactivation of indicator bacteriophages in fresh waters.
J Appl Microbiol, 2001. 92: p. 338-347.
36. Yates, M.V., C.P. Gerba, and L.M. Kelley, Virus persistence in groundwater. Appl
Environ Microbiol, 1985. 49(4): p. 778-81.

142
37. Noble, R.T. and J.A. Fuhrman, Virus decay and its causes in coastal waters. Appl
Environ Microbiol, 1996. 63(1): p. 77-83.
38. Sinton, L.W., R.J. Davies-Colley, and R.G. Bell, Inactivation of enterococci and fecal
coliforms from sewage and meatworks effluents in seawater chambers. Appl Environ
Microbiol, 1994. 60(6): p. 2040-8.
39. Allwood, P.B., et al., Survival of F-specific RNA coliphage, feline calicivirus, and
Escherichia coli in water: a comparative study. Appl Environ Microbiol, 2003. 69(9): p.
5707-10.
40. Team, R.D.C., R: A Language and Environmeny for Statistical Computing. 2010, R
Foundation for Statistical Computing: Vienna, Austria.
41. Goodridge, L., A. Gallaccio, and M.W. Griffiths, Morphological, host range, and genetic
characterization of two coliphages. Appl Environ Microbiol, 2003. 69(9): p. 5364-71.
42. Canchaya, C., et al., Prophage genomics. Microbiol Mol Biol Rev, 2003. 67(2): p. 238-
76, table of contents.
43. Palmer, K.L. and M.S. Gilmore, Multidrug-resistant enterococci lack CRISPR-cas.
MBio, 2010. 1(4).
44. Purnell, S.E., J.E. Ebdon, and H.D. Taylor, Bacteriophage lysis of Enterococcus host
strains: a tool for microbial source tracking? Environ Sci Technol, 2011. 45(24): p.
10699-705.
45. Calci, K.R., et al., Occurrence of male-specific bacteriophage in feral and domestic
animal wastes, human feces, and human-associated wastewaters. Appl Environ
Microbiol, 1998. 64(12): p. 5027-9.
46. Dhillon, T.S., et al., Studies on bacteriophage distribution: virulent and temperate
bacteriophage content of mammalian feces. Appl Environ Microbiol, 1976. 32(1): p. 68-
74.
47. Enriquez, C.E., C.J. Hurst, and C.P. Gerba, Survival of enteric adenovirus 40 and 41 in
tap, sea and waste water. Wat Res, 1995. 29(11): p. 2548-2553.
48. Osawa, S., K. Furuse, and I. Watanabe, Distribution of ribonucleic acid coliphages in
animals. Appl Environ Microbiol, 1981. 41(1): p. 164-8.
49. Ohgaki, S., et al., Adsorption of Coliphages to Particulates. Water Sci Technol, 1986.
18(7): p. 267-275.

143
50. Sidwell, R.W., G.J. Dixon, and E. McNeil, Quantitative studies on fabrics as
disseminators of viruses: Persistence of vaccinia virus on fabrics impregnated with a
virucidal agent. Appl Microbiol, 1967. 15(4): p. 921-7.
51. Sobsey, M.D. and R.C. Cooper, Enteric virus survival in algal-bacterial wastewater
treatment systems—I: Laboratory studies. Water Res, 1973. 7(5): p. 669-685.
52. Payment, P., et al., Virological examination of drinking water: a Canadian collaborative
study. Can J Microbiol, 1984. 30(1): p. 105-12.
53. Kelly, S.M. and W.W. Sanderson, The effect of chlorine in water on enteric viruses. II.
The effect of combined chlorine on poliomyelitis and Coxsackie viruses. Am J Public
Health Nations Health 1960. 50: p. 14-20.

144
CHAPTER 6
Antibiotic-resistance and virulence genes in Enterococcus isolated from tropical
recreational waters and sand
T. M. Santiago-Rodriguez1, J. I. Rivera
1, M. Coradin
1 and G. A. Toranzos
1
1Environmental Microbiology Laboratory, Department of Biology, University of Puerto Rico,
Rico, San Juan, Puerto Rico 00931-3360
ABSTRACT
The prevalence of enterococci harboring tetracycline and vancomycin-resistance genes, as well
as the enterococcal surface protein (esp) has mostly been determined in clinical settings, but their
prevalence in tropical recreational waters remains largely unknown. The present study
determined the prevalence of tetM (tetracycline-resistance), vanA and vanB (vancomycin-
resistance) in the bacterial and viral fractions, enterococci and their induced phages isolated from
tropical recreational marine and fresh waters, dry and wet sands. Since lysogenic phages can act
as vectors for antibiotic-resistance and virulence factors, the prevalence of the mentioned genes,
as well as that of an integrase-encoding gene (int) specific for Enterococcus faecalis phages was
determined. Up to 60 % and 54 % of the bacterial fractions and enterococci harbored at least one
of the tested genes, respectively, suggesting that bacteria in tropical environments may be
reservoirs of antibiotic-resistance and virulence genes. int was detected in the viral fractions and
in one Enterococcus isolate after induction. This study opens the opportunity to determine if the
presence of bacteria harboring antibiotic-resistance and virulence genes in tropical recreational
waters represents a threat to public health.
Keywords: antibiotic-resistance, bacteriophages, enterococci, enterococcal surface protein
Reference: Santiago-Rodriguez, et al. Antibiotic-resistance and virulence genes in Enterococcus
isolated from tropical recreational waters and sand. 2013. Journal of Water and Health.
Accepted.

145
INTRODUCTION
There is an increasing concern towards bacteria harboring antibiotic-resistance and virulence
genes in recreational waters. One of the main reasons is that it remains unknown if
microorganisms harboring these genes represent a risk to bathers. Among these, enterococci have
received great attention since these are ubiquitous to many aquatic ecosystems [1-5]. However,
certain Enterococcus spp. are also important opportunistic pathogens and infections have been
treated with tetracyclines. This has resulted in tetracycline-resistant enterococci and these may
harbor at least one of the tet determinants, being tetM one of the most studied in gram-positive
bacteria [6-8]. Resistant Enterococcus infections can be treated with vancomycin, but only as a
last resort due to the hazardous side effects [9]. However, enterococci have also acquired
resistance to vancomycin due to genes such as vanA and vanB, commonly described in E.
faecalis and E. faecium. In addition, enterococci exhibiting resistance to antibiotics can harbor
genes encoding for virulence factors, as in the case of the enterococcal surface protein (Esp),
described in E. faecalis and E. faecium [10-13]. The esp variant present in E. faecium was
proposed as a marker of human fecal contamination and there have been conflicting studies on
the usefulness of this gene as a species-specific marker [14-17].
Another concern is the transmission of antibiotic-resistant and virulence genes by mobile
elements, such as plasmids and transposons, but bacteriophages can also act as vectors of these
genes [18, 19]. Lysogenic phages can carry additional bacterial genes which could promote
fitness and virulence in the recipient bacteria [20-22]. These genes are often encoded by the host
and may be accidentally packaged into the phage capsid. It has been shown that the viral DNA
fraction of sewage and environmental waters harbor β-lactamase-encoding genes and that
lysogenic phages infecting E. faecalis may harbor genes that influence virulence in the bacterial
host [23, 24]. Most lysogenic phages infecting enterococci described so far belong to the
Siphoviridae family (dsDNA) and one possible approach to identify these is by amplifying
integrase-encoding genes (int). Integrases are considered markers of lysogeny and are involved
in the integration of the phage genome into that of the bacterial host [25]. Detection of phage
integrases within an ecosystem may provide insights as to what possible risks exist within a
specific ecosystem [26].

146
The prevalence of tetM, and vanA and vanB has mostly been determined in animal husbandry
facilities, and temperate and subtropical recreational waters, respectively [5, 27]; and although
the prevalence of esp has been determined in tropical regions, few studies are still available [28].
In addition, the presence of antibiotic-resistance and virulence genes in the bacteriophage
fraction of recreational waters, and integrases as a way to identify these, remains largely
unknown. Therefore, the main aim of the present study was to determine the prevalence of tetM,
vanA, vanB, and esp variants present in E. faecalis and E. faecium, as well as int, in the bacterial
and DNA viral fractions, enterococci and their lysogenic phages in tropical recreational waters.
MATERIALS AND METHODS
Study sites
Samples were collected from OP, CBB, LC and RP in Puerto Rico (Figure 1). OP and CBB are
beaches located in the Atlantic Ocean, alongside the northern area of the island and are heavily
used by bathers year-round. LC is a man-made lake whose influents are three of the island’s
major rivers, serves as a major water reservoir for the San Juan metropolitan area and is also
used for recreational purposes. RP receives the input of minor rivers and streams used for
recreational activities. One-liter grab samples of marine (OP, n=35; and CBB, n=13), and fresh
(LC, n=8; RP, n=10) waters were collected in sterile plastic bottles from July 2011 to May 2012.
In addition, 100 g of wet and dry sands from OP (n=16 and n=15, respectively) and CBB (n=14
and n=11, respectively) were collected since sand may act as a reservoir of bacteria harboring
antibiotic-resistance and virulence genes [29, 30]. All water and sand samples were stored at 6-8
ºC and processed within 6 h.
DNA extraction of the bacterial fraction and enterococci
To determine the presence of the mentioned genes in the bacterial fraction, 100 mL were
processed by membrane filtration (0.4 μm pore size, 47 mm diameter) (GE Water and Process
Technologies, Trevose, PA). The filter was placed in 25 mL of Azide Dextrose Broth (Difco)
and incubated at 37 ºC for 24 h. DNA was extracted from 1 mL by using the Fermentas GeneJet
Genomic DNA Purification Kit following the manufacturer’s instructions. Alternatively, filters
were placed in tubes containing 0.3 g of acid-wash glass beads (Sigma) and stored at -20 ºC until
processed. For this, 600 μL of AE buffer were added, placed on a bead-beater for 60 s at

147
maximum speed and centrifuged for 1 min at 14,000 rpm. The supernatant was collected, re-
centrifuged for 5 min at 14,000 rpm to remove any remaining debris and stored at -20 ºC until
processed [31]. For the sand samples, 10 g were directly added into 20 mL of Azide Dextrose
Broth, incubated at 37 °C for 24 h and 1 mL was used for DNA extraction using the Fermentas
GeneJet Genomic DNA Purification Kit. Alternatively, 10 g were eluted in 120 mL of 0.1%
Tween20, 100 mL of the elution were filtered and DNA from the membranes was extracted as
described above.
Figure 1: Sampled sites in this study. Sites included CBB ( ), OP ( ), RP ( a), and LC ( ) in
Puerto Rico. Marine water and dry and wet sands were collected from CBB and OPB. Fresh
water was collected from RP and LC.
Atlantic Ocean

148
Enumeration of enterococci was performed by using membrane filtration and m-Enterococcus
agar (Difco) as previously described [32, 33]. For the detection of enterococci in sand, 10 g were
eluted in 0.1% Tween20 followed by filtration of the resulting solution and membranes were
placed on m-Enterococcus agar as described. After 24-48h incubation at 37 ºC, individual
colonies were picked and transferred onto m-Enterococcus plates containing tetracycline (final
concentration 16 μg/mL) or vancomycin (final concentration 20 μg/mL). Colonies growing in the
presence of these antibiotics were picked and enriched in 1 mL of Azide Dextrose Broth for up
to 48 h at 37 ºC and DNA was extracted as described above. DNA quality and concentration was
estimated by using a NanoDrop® (ND-1000) spectrophotometer.
Virus concentration and DNA extraction
Filtrates from above were recovered, concentrated and purified for the detection of the
mentioned genes in the viral DNA fractions. Briefly, a final concentration of 0.2 % chloroform
(v/v) was added to the viral suspensions and kept at room temperature for 30 min to eliminate
any remaining viable bacteria. Solid NaCl (Fisher Scientific, enzyme grade ≥ 99.9 %, NJ, USA)
was added to the suspensions at a final concentration of 0.5 M and stored at 4 °C for 1 h. Any
remaining bacterial debris was removed by centrifugation at 8,500 rpm for 10 min at 4 °C and
the supernatant was transferred to sterile Oakridge tubes. PEG 8000 (Promega) was dissolved in
the phage suspension in a final concentration of 10 % and stored at 4 °C for 24 h. Bacteriophages
were sedimented at 14,000 rpm at 4 °C for 15 min and the supernatant was discarded. The phage
containing pellet was resuspended in 0.5 mL of PBS and left overnight at 4 °C. The residual PEG
and bacterial debris were removed by adding an equal volume of chloroform to a Phase Lock Gel
tube (5 Prime Inc., MD, USA) and centrifuging at 9500 rpm for 15 min [34]. Prior to viral DNA
extraction, samples were treated using DNAse (Sigma) (final concentration 1U/μL). Viral DNA
was extracted by using phenol:chloroform followed by precipitation with 95 % ethanol or the
Wizard DNA Clean up system following the manufacturer’s instructions. In order to confirm the
size and purity of the viral DNA, 4 μL were visualized in 0.7 % agarose gels using GelStar
Nucleic Acid Gel Stain (Lonza, Rockland, ME, USA).
Lysogen induction

149
One-hundred-μL of an overnight culture of 50 randomly selected enterococci isolates harboring
one or more of the genes of interest, were added to 5 mL of Azide Dextrose (Difco) containing
mitomycin C (Sigma) (final concentration of 1 μg/mL). Samples were incubated in a water bath
at 37 °C for 4-6 h and centrifuged at 14,000 rpm for 15 min at 4 ºC [35]. Supernatants were
tested for the presence of lysogenic phages exhibiting the formation of viral plaques by using the
double layer method and E. faecalis (ATCC 19433), E. faecium, E. gallinarum, E. hirae (ATCC
8043), E. durans (ATCC 19432), E. dispar (ATCC 51266), E. casseliflavus (ATCC 25788), E.
pseudoavium (ATCC 49372) and Staphylococcus aureus (ATCC 25923) as the bacterial hosts.
Briefly, 100 μL of the possible phage lysates and 1 mL of a fresh culture of each bacterial host
were added to 4 mL containing Trypticase Soy Broth (TSB) (Difco) and agar (0.75 % w/v) and
poured onto a Petri dish containing TSB, agar (1.5 % w/v), CaCl2·2H2O (Fisher Scientific Co.
NJ, USA) and NaN3 (MCB, OH, USA) (final concentration of 2.6 mg/mL and 0.4 mg/mL,
respectively). Two temperatures were tested for formation of viral plaques (22 and 37 ºC) and
enumerated after 24 h. Phages exhibiting the formation of viral plaques were isolated,
propagated using E. faecalis (ATCC 19433) as the bacterial host and concentrated as described
[36]. A composite of the phage lysates lacking the formation of viral plaques using the
mentioned bacterial strains was concentrated as described. DNA was extracted from phages
exhibiting and lacking the formation of viral plaques as described above. One of the induced
phage isolates was characterized morphologically by using type-B 200 mesh copper grids placed
on top of individual viral plaques and stained with uranyl acetate (UA) 2%, pH 4.5.
Bacteriophages were visualized by using a Karl Zeiss Leo 922 energy filtered transmission
electron microscope operated at 200 KV.
PCR amplification conditions
All primers used in the present study are described in Table 1. Primer design in this study was
performed using Primer 3 (http://frodo.wi.mit.edu/primer3/) and verified for the formation of
secondary structures using NetPrimer (http://www.premierbiosoft.com/netprimer/). Detection of tetM
was performed by designing primers targeting the tetM sequence found in Gene Bank (accession
number AY304474.1). Primers used for the detection of vanA and vanB were those previously
described [5, 37]. For the detection of the esp variant of E. faecalis, primers were designed using
the gene sequence found in Gene Bank (accession number DQ845099.1). Detection of the esp

150
variant of E. faecium was performed using primers described elsewhere [14]. For the detection of
lysogenic phages, primers were designed using the integrase gene sequence of the Enterococcus
phage phiFL1B found in Gene Bank (accession number GQ478082.1).
Target gene Primer sequence Direction Reference
tetM GGAAAATACGAAGGTGAACA Forward This study
GAATCCCCATTTTCCTAAGT Reverse This study
vanA GGGAAAACGACAATTGC Forward Roberts et. al, 2009
GTACAATGCGGCCGTTA Reverse Roberts et. al, 2009
vanB TTGCATGGACAAATCACTGC Forward Roberts et. al, 2009
GCTCGTTTTCCTGATGGATG Reverse Roberts et. al, 2009
esp E. faecalis CACAAATGGGTGAAGGAAGA Forward This study
AGACGAATTTCCCAGTTTGC Reverse This study
esp E. faecium TATGAAAGCAACAGCACAAGTT Forward Scott et. al, 2005
ACGTCGAAAGTTCGATTTCC Reverse Scott et. al, 2005
int TATTAGGAAAACCTCCGTCA Forward This study
ATATCTTGGGCGTAAGTGAA Reverse This study
16S rRNA AGAGTTTGATCCTGGCTCAG Forward Amann et al., 1995
ACGGGCGGTGTGTRC Reverse Amann et al., 1995
Table 1: Primers in this study. Target genes include those encoding for resistance to tetracycline
(tetM) and vancomycin (vanA and vanB), the enterococcal surface protein (esp) variants found in
E. faecalis and E. faecium and an integrase-encoding gene specific for E. faecalis phages (int).
The 16S rRNA gene amplification was used as a control in the viral fractions and lysogenic
Enterococcus phages.
Amplifications were performed in a total volume of 8μL per reaction, including 1 μL of genomic
DNA at a concentration of approximately 10 ng/μL, 4 μL of Fermentas DreamTaq PCR Master
Mix, 2.2 μL of milliQ water and 0.4μL of each primer at 10 μM. PCR conditions for tetM were:
an initial denaturation of 94 ºC for 5 min, followed by 25 cycles of 94 ºC for 30 s, annealing of
55 ºC for 30 s, followed by an extension of 72 ºC for 30 s and a final extension of 72 ºC for 7
min. Both vanA and vanB PCR conditions were: an initial denaturation of 96 ºC for 3 min,
followed by 35 cycles of 96 ºC for 30 s, annealing of 60 ºC for 1 min, extension of 72 ºC for 2
min and a final extension of 72 ºC for 10 min. For the detection of the esp variant found in E.
faecalis the following conditions were used: an initial denaturation of 95 ºC for 2 min, followed

151
by 30 cycles of 95 ºC for 45 s, annealing of 57 ºC for 45 s and an extension of 72 ºC for 1 min.
Detection of the esp variant found in E. faecium was done by using an initial denaturation of 95
ºC for 3 min, followed by 35 cycles of 94 ºC for 1 min, annealing of 58 ºC for 1 min and an
extension of 72 ºC for 1 min. int PCR parameters were: an initial denaturation of 95 ºC for 2 min,
followed by 35 cycles of 94 ºC for 45 s, annealing of 55 ºC for 45 s, followed by an extension of
72 ºC for 1 min and a final extension of 72 ºC for 7 min. To ensure that no bacterial DNA was
present in the viral DNA, amplification of the 16S rRNA gene was performed using the
following PCR conditions: an initial denaturation of 95 ºC for 3 min, followed by 30 cycles of 95
ºC for 30 s, annealing of 52 ºC for 30 s, followed by an extension of 72 ºC for 30 s and a final
extension of 72 ºC for 10 min. PCR products were visualized in 1.0 % agarose gels using GelStar
Nucleic Acid gel stain. Positive controls were included for tetM and both esp variants and
potential int products were sequenced using an ABI 3130xl Genetic Analyzer.
Statistical analyses
Fisher’s exact tests were used to determined differences in the prevalence of the mentioned genes
between sample types within the sample sites. The same analysis aimed to determine significant
differences in the prevalence of the esp variants between sample types. All analyses were
performed with the R statistical software (v.2.11.1) [38].
RESULTS
Bacterial fraction and enterococci isolates
Table 2 shows the prevalence of the tested genes in the bacterial fractions of the water and sand
samples. A significant difference was noted in the prevalence of the esp variants of water
samples collected from CBB (p = 0.030). Table 3 shows the prevalence of the esp variants and
int in the enterococci isolates. Table 4 shows the prevalence of tetracycline and vancomycin-
resistant enterococci and tetM, vanA and vanB. Not all enterococci exhibiting resistance to either
antibiotic harbored tetM or vanA and vanB. Interestingly, several of these isolates harbored two
of the genes of interest. In OP waters, 2 of the isolates were positive for both tetM and vanA, 8
harbored tetM and the esp variant present in E. faecalis and 1 harbored both vanA and the esp
variant found in E. faecalis. In wet sands collected from OP and CBB, 2 and 1 of the isolates
were positive for tetM and the esp variant present in E. faecium, respectively. In dry sands

152
collected from CBB, 3 of the enterococci isolated were positive for tetM and the esp variant
present in E. faecium. Similarly, 1 of the isolates from LC was positive for the latter combination
of genes (data not shown). Significant differences were noted for the prevalence of tetM in CBB
waters and dry sand (p = 0.0015) and CBB wet and dry sands (p = 0.0038). The prevalence of
both esp variants in waters collected from OP was significantly different as well (p = 0.033).
OP
water
(n=35)
OP wet
sand
(n=16)
OP dry
sand
(n=15)
CBB
water
(n=13)
CBB wet
sand
(n=14)
CBB dry
sand
(n=11)
LC
(n=8)
RP
(n=10)
tetM 6 (17) 2 (13) 2 (13) 4 (31) 1 (7) 1 (9) 2 (25) 2 (20)
vanA 7 (20) 4 (25) 2 (13) ND* 1 (7) 2 (18) 2 (25) ND*
vanB ND* ND* ND* ND* ND* ND* ND* ND*
esp E. faecalis 2 (5) 2 (13) 2 (13) 7 (54) 7 (50) 4 (36) 2 (25) 6 (60)
esp E. faecium 6 (17) 4 (25) 5 (33) 1 (8) ND* ND* ND* ND*
int 4 (11) 1 (6) 1 (7) 2 (15) 1 (7) ND* 3 (38) 2 (20)
ND*=not detected
Table 2: Prevalence of antibiotic-resistance and virulence-encoding genes in the bacterial
fractions of tropical marine (OP, CBB) and fresh waters (LC, RP) and wet and dry sands (OP,
CBB). Genes included those conferring resistance to tetracycline (tetM) and vancomycin (vanA
and vanB), the enterococcal surface protein (esp) variants present in E. faecalis and E. faecium
and an integrase-encoding gene specific for E. faecalis phages (int). Percents are presented in
parenthesis.
OP
water
(n=99)
OP wet
sand
(n=55)
OP dry
sand
(n=46)
CBB
water
(n=25)
CBB wet
sand
(n=21)
CBB dry
sand
(n=25)
LC
(n=19)
RP
(n=66)
esp E.
faecalis
10 (10) 1 (2) 1 (2) 2 (8) 1 (5) ND* ND* 6 (9)
esp E.
faecium
2 (2) 6 (11) 3 (7) 2 (8) 2 (10) 5 (20) ND* 5 (8)
int 3 (3) ND* ND* ND* ND* ND* 3 (16) ND*
ND*=not detected

153
Table 3: Prevalence of the esp variants present in E. faecalis and E. faecium and int in tropical
marine (OP, CBB) and fresh waters (LC, RP) and wet and dry beach sands in the Enterococcus
isolates. Percents are presented in parenthesis.
ND*=not detected
Table 4: Enterococci isolates tested for tetracycline (16 μg/mL) and vancomycin (20 μg/mL)
resistance isolated from tropical samples. The presence of tetracycline (tetM) and vancomycin
(vanA and vanB) resistance genes was also tested in the isolates.
Viral fraction and induced phages
None of the antibiotic-resistance or virulence genes were detected in the viral DNA fractions.
None of the samples were positive for the 16S rRNA gene and only int was detected in one of
the filtrates. Similarly, the antibiotic-resistance and virulence genes tested were not detected in
the induced enterococci phages. Not all induced enterococci phages were able to infect most of
the Enterococcus or S. aureus type strains in this study, but those that formed viral plaques were
able to replicate at 22 and 37ºC in E. faecalis. In addition, enterococci phages induced from an
enterococci isolate exhibited two different viral plaque morphologies (Figure 2A). Some plaques
were turbid on the edges and translucent in the center, while other plaques were completely
translucent, and these phages were not positive for the integrase-encoding gene tested in this
study. On the other hand, one of the induced enterococci phages that did not form viral plaques
using the mentioned enterococci type strains was positive for int. Sequencing of int showed a 99
% similarity (e value=4 e-81) with int of E. faecalis phages phiFL1A, 1B, 1C, 2A, 2B, 3A and
Antibiotic
Gene
Sample Tested Tetracycline Vancomycin Tested tetM vanA vanB
OP water 188 26(14) 18(10) 99 13(13) 2 ND*
OP wet sand 75 14(19) 10(13) 55 9(16) ND* ND*
OP dry sand 136 31(23) 31(23) 46 9(20) ND* ND*
CBB water 65 5(8) 3(5) 25 2(8) ND* ND*
CBB wet sand 75 23(31) 8(11) 21 2(10) ND* ND*
CBB dry sand 130 74(57) 16(12) 25 13(52) ND* ND*
LC 173 50(29) 70(40) 19 3(16) ND* ND*
RP 87 22(29) 24(28) 66 3(5) ND* ND*

154
3B. In terms of the morphology of one of the induced enterococci phage, this exhibited an
icosahedral capsid and a flexible tail of approximately 80 and 200nm, respectively (Figure 2B).
(A) (B)
Figure 2: Characterization of induced enterococci phages. Panel (A) shows the viral plaque
morphologies of an enterococci isolate. Panel (B) shows the morphology of an induced
enterococci phage (Bar=200 nm). Phage exhibited an icosahedral capsid of approximately 80 nm
and a flexible tail of approximately 200 nm.
DISCUSSION
Many studies focusing on the prevalence of tet determinants in the environment have been done
in waters associated with swine production facilities and livestock [27, 39, 40]. However, few
studies that focus on the prevalence of tetracycline-resistance genes in tropical recreational
waters are available. In the present study, tetM was detected in both the bacterial fractions and
enterococci isolated from waters and wet and dry sands. Interestingly, significant differences
were noted in the prevalence of tetM in waters and wet and dry sands collected from CBB. This
may suggest that the prevalence of tetM may be independent of the sample type, but this was not
the case for all the sampled sites. The detection of tetracycline resistant bacteria in the sites
tested may suggest that these are being introduced via human fecal material (as there are no
animal husbandry facilities in proximity). Another possibility is that the presence of tetracycline
in waters is selecting for resistant bacteria. This is reasonable since it has been shown that
tetracycline can be introduced via feces and urine and detected in concentrations of
approximately 0.10 μg/L [41, 42]. The horizontal transfer of genes conferring tetracycline-

155
resistance via mobile elements between autochthonous and exogenous bacteria is likely, but in
tropical recreational waters it may be difficult to identify enterococci from a fecal source and
those from the environment. Interestingly, enterococci that exhibited resistance to tetracycline
but were negative for tetM suggests that these may harbor other tet determinants.
This is also one of the few reports on the detection of vanA and vanB in tropical recreational
waters. Results suggest that several bacterial species in tropical environments may harbor vanA
or vanA-like genes (e.g. S. aureus) [43]. The present study is comparable with previous results in
which enterococci harboring vanA have been isolated from marine waters [5, 44]. In contrast,
vanB was not detected in any of the samples tested, but this is similar to results presented
elsewhere [45]. Although the reasons for this remain to be determined, it is possible that
harboring vanA, instead of vanB, may be more beneficial since harboring vanA confers resistance
to vancomycin and teicoplanin [46]. It should be noted that most of the enterococci isolates in
the present study that exhibited resistance to vancomycin lacked vanA or vanB. It is possible that
the enterococci isolates tested harbored other genes conferring resistance to vancomycin (e.g.
vanC, vanD and vanE); although it is also feasible that genes conferring resistance to other
antibiotics may also be conferring resistance to vancomycin. Similar outcomes have been
reported in Europe in which vancomycin-resistant enterococci have been associated with the use
of avoparcin [47]. The source of vanA in tropical waters and sand remains to be determined,
although the input of fecal matter remains a possibility since vancomycin-resistant bacteria have
been detected in sewage effluents [44]. As with tetracycline, future studies need to determine if
vancomycin is being introduced into the waters tested and if it is a selective force in tropical
environments.
Detection of both esp variants in the bacterial fractions and enterococci isolates of several of the
samples may suggest an input of human and animal fecal material [13]. The absence of the E.
faecium variant in the bacterial fractions and in enterococci of several of the samples evaluated
was not expected since these sites are visited throughout the year and may not be free from fecal
contamination from non-point sources. This may suggest that the E. faecium variant may not
necessarily indicate human fecal pollution in all water types, especially in the tropics, in which
enterococci are naturally found in many water bodies [48]. Only the bacterial fraction of waters

156
collected from CBB and the enterococci isolated from OP water exhibited a difference in the
prevalence of both esp variants. This may suggest that, in tropical environments, similar results
are obtained when detecting either esp variant.
In the present study, none of the tested genes were detected in the viral DNA fraction or the
induced enterococci phages. However, the presence of other antibiotic-resistance and virulence-
encoding genes in bacteriophages should not be ignored. In terms of int, although it was detected
in the enterococci isolates and one of the induced phages, it is possible that other Enterococcus
phages may harbor int sequences different from the variant tested in the present study. Phages
infecting enterococci have exhibited differences in their lysogeny modules and int sequences [24,
49]. The present study also suggests that Enterococcus isolates may harbor more than one
inducible prophage, but the function of various prophages in the genomes of enterococci remains
to be determined [50, 51].
CONCLUSIONS
This is one of the first reports which aimed to identify a battery of antibiotic-resistance and
virulence genes in both the bacterial and viral fractions of tropical recreational waters and sand.
The presence of antibiotic-resistance and virulence genes in the environmental microbiota of
tropical samples suggests that the transmission of these genes is not restricted to strains of the
clinical setting. The exact mechanisms of horizontal transfer of antibiotic-resistance and
virulence genes under these environmental conditions need to be determined, as do the role of
environmental microbiota as reservoirs of these genes. By detecting int in lysogenic phages,
further studies could determine the presence of antibiotic-resistance and virulence factors in their
genomes. Our data also point to the need to take into consideration lysogenic phages when
determining the presence or absence of bacterial viruses in the environment.
Most epidemiological studies on recreational waters have focused on gastrointestinal illness;
though a few recent ones have also included and eye, skin and ear infections. The present study
showed a relatively high prevalence of Enterococcus spp. exhibiting resistance to antibiotics
which also harbored virulence genes. It remains to be determined if these bacteria represent a
risk to bathers and if these are specifically involved in skin infections. However, it is worrying

157
that opportunistic pathogens involved in skin infections and harboring antibiotic resistance are
present in recreational waters and sands, where bathers may be exposed to them.
ACKNOWLEDGEMENTS
We thank M.G. Dominguez-Bello, N. Shankar and M.N. Byappanahalli for sharing the positive
controls used in this study; Miguel Urdaneta for the phage microscopy and Rita Patricio, Jean M.
Carrasquillo, Roberto Lorenzini, Gabriel Vázquez and Gabriela Tirado for sample collection and
processing. This research was partially supported by MBRS-RISE (NIH Grant Number
2R25GM061151-09).

158
LITERATURE CITED
1. Byappanahalli, M.N., et al., Growth and survival of Escherichia coli and enterococci
populations in the macro-alga Cladophora (Chlorophyta). FEMS Microbiol Ecol, 2003.
46(2): p. 203-11.
2. Devriese, L.A., et al., Enterococcal and streptococcal species isolated from faeces of
calves, young cattle and dairy cows. J Appl Bacteriol, 1992. 72(1): p. 29-31.
3. Jett, B.D., M.M. Huycke, and M.S. Gilmore, Virulence of enterococci. Clin Microbiol
Rev, 1994. 7(4): p. 462-78.
4. Rivera, S.C., T.C. Hazen, and G.A. Toranzos, Isolation of fecal coliforms from pristine
sites in a tropical rain forest. Appl Environ Microbiol, 1988. 54(2): p. 513-517.
5. Roberts, M.C., et al., Vancomycin-resistant Enterococcus spp. in marine environments
from the West Coast of the USA. J Appl Microbiol, 2009. 107(1): p. 300-7.
6. Tannock, G.W., The normal microflora as a reservoir of antibiotic resistance-
determinants, in Medical Importance of the Normal Microflora. 1999, Kluwer Academic
Publishers: Dordrecht, The Netherlands. p. 388-404.
7. Roberts, M.C., Update on acquired tetracycline resistance genes. FEMS Microbiol Lett,
2005. 245(2): p. 195-203.
8. Murray, B.E., Diversity among multidrug-resistant enterococci. Emerg Infect Dis, 1998.
4(1): p. 37-47.
9. Rocha, J.L., et al., Uncommon vancomycin-induced side effects. Braz J Infect Dis, 2002.
6(4): p. 196-200.
10. Klare, I., et al., Spread of ampicillin/vancomycin-resistant Enterococcus faecium of the
epidemic-virulent clonal complex-17 carrying the genes esp and hyl in German hospitals.
Eur J Clin Microbiol Infect Dis, 2005. 24(12): p. 815-25.
11. Toledo-Arana, A., et al., The enterococcal surface protein, Esp, is involved in
Enterococcus faecalis biofilm formation. Appl Environ Microbiol, 2001. 67(10): p. 4538-
45.
12. Shankar, N., et al., Role of Enterococcus faecalis surface protein Esp in the pathogenesis
of ascending urinary tract infection. Infect Immun, 2001. 69(7): p. 4366-72.

159
13. Hammerum, A.M. and L.B. Jensen, Prevalence of esp, encoding the enterococcal surface
protein, in Enterococcus faecalis and Enterococcus faecium isolates from hospital
patients, poultry, and pigs in Denmark. J Clin Microbiol, 2002. 40(11): p. 4396.
14. Scott, T.M., et al., Potential use of a host associated molecular marker in Enterococcus
faecium as an index of human fecal pollution. Environ Sci Technol, 2005. 39(1): p. 283-
7.
15. Ahmed, W., et al., Evaluation of the host-specificity and prevalence of enterococci
surface protein (esp) marker in sewage and its application for sourcing human fecal
pollution. J Environ Qual, 2008. 37(4): p. 1583-8.
16. Layton, B.A., S.P. Walters, and A.B. Boehm, Distribution and diversity of the
enterococcal surface protein (esp) gene in animal hosts and the Pacific coast
environment. J Appl Microbiol, 2009. 106(5): p. 1521-31.
17. Byappanahalli, M.N., et al., Environmental occurrence of the enterococcal surface
protein (esp) gene is an unreliable indicator of human fecal contamination. Environ Sci
Technol, 2008. 42(21): p. 8014-20.
18. Oancea, C., et al., Conjugative transfer of the virulence gene, esp, among isolates of
Enterococcus faecium and Enterococcus faecalis. J Antimicrob Chemother, 2004. 54(1):
p. 232-5.
19. Garcia-Vallve, S., A. Romeu, and J. Palau, Horizontal gene transfer in bacterial and
archaeal complete genomes. Genome Res, 2000. 10(11): p. 1719-25.
20. Hendrix, R.W., Bacteriophage genomics. Curr Opin Microbiol, 2003. 6(5): p. 506-11.
21. Hendrix, R.W., et al., The origins and ongoing evolution of viruses. Trends Microbiol,
2000. 8(11): p. 504-8.
22. Hendrix, R.W., et al., Evolutionary relationships among diverse bacteriophages and
prophages: all the world's a phage. Proc Natl Acad Sci U S A, 1999. 96(5): p. 2192-7.
23. Colomer-Lluch, M., J. Jofre, and M. Muniesa, Antibiotic resistance genes in the
bacteriophage DNA fraction of environmental samples. PLoS One, 2011. 6(3): p. e17549.
24. Yasmin, A., et al., Comparative genomics and transduction potential of Enterococcus
faecalis temperate bacteriophages. J Bacteriol, 2010. 192(4): p. 1122-30.
25. Groth, A.C. and M.P. Calos, Phage integrases: biology and applications. J Mol Biol,
2004. 335(3): p. 667-78.

160
26. Heinemann, J.A., et al., Environment arrays: a possible approach for predicting changes
in waterborne bacterial disease potential. Environ Sci Technol, 2006. 40(23): p. 7150-6.
27. Agerso, Y., A.G. Pedersen, and F.M. Aarestrup, Identification of Tn5397-like and Tn916-
like transposons and diversity of the tetracycline resistance gene tet(M) in enterococci
from humans, pigs and poultry. J Antimicrob Chemother, 2006. 57(5): p. 832-9.
28. Betancourt, W.Q. and R.S. Fujioka, Evaluation of enterococcal surface protein genes as
markers of sewage contamination in tropical recreational waters. Water Sci Technol,
2009. 60(1): p. 261-6.
29. Hartz, A., et al., Survival potential of Escherichia coli and Enterococci in subtropical
beach sand: implications for water quality managers. J Environ Qual, 2008. 37(3): p.
898-905.
30. Fernandes Cardoso de Oliveira, A.J., P.T. Ranzani de Franca, and A.B. Pinto,
Antimicrobial resistance of heterotrophic marine bacteria isolated from seawater and
sands of recreational beaches with different organic pollution levels in southeastern
Brazil: evidences of resistance dissemination. Environ Monit Assess, 2010. 169(1-4): p.
375-84.
31. USEPA, Method A: Enterococci in Water by TaqMan® Quantitative Polymerase Chain
Reaction (qPCR) Assay, in EPA-821-R-10-004. 2010: Washington, D.C.
32. Santiago-Rodriguez, T.M., Davila, C., Gonzalez, J., Bonilla, N., Marcos, P., Urdaneta,
M., Cadete, M., Monteiro, S., Santos, R., Santo-Domingo, J. and Toranzos, G.,
Characterization of Enterococcus faecalis-infecting phages (enterophages) as markers of
human fecal pollution in recreational waters. Water Research, 2010. 44(16): p. 4716-
4725.
33. USEPA, Method 1600: Membrane Filter Test Method for Enterococci in Water, in EPA-
821-R-02-022. 2002, Office of Water: Washington, D.C.
34. Boulanger, P., Purification of Bacteriophages and SDS-PAGE Analysis of Phage
Structural Proteins from Ghost Particles, in Bacteriophages: Methods and Protocols,
M.R.J. Clokie and A.M. Kropinski, Editors. 2009, Humana Press: NewYork, New York.
p. 227-238.

161
35. Raya, R.R. and E.M. Hebert, Isolation of Phage via Induction of Lysogens, in
Bacteriophages: Methods and Protocols, M.R.J. Clokie and A.M. Kropinski, Editors.
2009, Humana Press: New York, NY. p. 23-33.
36. Bonilla, N., et al., Enterophages, a group of phages infecting Enterococcus faecalis, and
their potential as alternate indicators of human faecal contamination. Water Sci Technol,
2010. 61(2): p. 293-300.
37. Soge, O.O., et al., Characterization of methicillin-resistant Staphylococcus aureus and
methicillin-resistant coagulase-negative Staphylococcus spp. isolated from US West
Coast public marine beaches. J Antimicrob Chemother, 2009. 64(6): p. 1148-55.
38. Team, R.D.C., R: A Language and Environmeny for Statistical Computing. 2010, R
Foundation for Statistical Computing: Vienna, Austria.
39. Aminov, R.I., N. Garrigues-Jeanjean, and R.I. Mackie, Molecular ecology of tetracycline
resistance: development and validation of primers for detection of tetracycline resistance
genes encoding ribosomal protection proteins. Appl Environ Microbiol, 2001. 67(1): p.
22-32.
40. Chee-Sanford, J.C., et al., Occurrence and diversity of tetracycline resistance genes in
lagoons and groundwater underlying two swine production facilities. Appl Environ
Microbiol, 2001. 67(4): p. 1494-502.
41. Zhu, J., et al., Analysis of oxytetracycline, tetracycline, and chlortetracycline in water
using solid-phase extraction and liquid chromatography-tandem mass spectrometry. J
Chromatogr A, 2001. 928(2): p. 177-86.
42. Lindsey, M.E., T.M. Meyer, and E.M. Thurman, Analysis of trace levels of sulfonamide
and tetracycline antimicrobials in groundwater and surface water using solid-phase
extraction and liquid chromatography/mass spectrometry. Anal Chem, 2001. 73(19): p.
4640-6.
43. Zhu, W., et al., Vancomycin-resistant Staphylococcus aureus isolates associated with
Inc18-like vanA plasmids in Michigan. Antimicrob Agents Chemother, 2008. 52(2): p.
452-7.
44. Novais, C., et al., Environmental contamination with vancomycin-resistant enterococci
from hospital sewage in Portugal. Appl Environ Microbiol, 2005. 71(6): p. 3364-8.

162
45. Schwartz, T., et al., Detection of antibiotic-resistant bacteria and their resistance genes
in wastewater, surface water, and drinking water biofilms. FEMS Microbiol Ecol, 2003.
43(3): p. 325-35.
46. Cetinkaya, Y., P. Falk, and C.G. Mayhall, Vancomycin-resistant enterococci. Clin
Microbiol Rev, 2000. 13(4): p. 686-707.
47. Bager, F., et al., Avoparcin used as a growth promoter is associated with the occurrence
of vancomycin-resistant Enterococcus faecium on Danish poultry and pig farms. Prev
Vet Med, 1997. 31(1-2): p. 95-112.
48. Muñiz, I., et al., Survival and activity of Streptococcus faecalis and Escherichia coli in
tropical fresh water. Microbial Ecology, 1989. 18: p. 125-134.
49. Stevens, R.H., M.R. Ektefaie, and D.E. Fouts, The annotated complete DNA sequence of
Enterococcus faecalis bacteriophage phiEf11 and its comparison with all available
phage and predicted prophage genomes. FEMS Microbiol Lett, 2011. 317(1): p. 9-26.
50. Brussow, H., C. Canchaya, and W.D. Hardt, Phages and the evolution of bacterial
pathogens: from genomic rearrangements to lysogenic conversion. Microbiol Mol Biol
Rev, 2004. 68(3): p. 560-602.
51. Banks, D.J., B. Lei, and J.M. Musser, Prophage induction and expression of prophage-
encoded virulence factors in group A Streptococcus serotype M3 strain MGAS315. Infect
Immun, 2003. 71(12): p. 7079-86.

163
CHAPTER 7
General conclusions and future directions
GENERAL CONCLUSIONS
A culture method was developed for the detection of Enterococcus faecalis phages
(enterophages) in recreational waters, sewage, sand and fecal samples. Our method is the most
recent approach to infer the virological quality of water sources. This method can also be tested
for the detection of enterophages in other sample types such as soils and food. It should be noted
that if the enterophage method is to be tested in other laboratories, a consistency should be kept
(reagents and the bacterial host) in order to compare results. The method enabled us to isolate
these viruses to further characterize them ecologically, morphologically and genetically.
Enterophages possess many of the characteristics of an indicator of fecal pollution and
specifically that from a human source. Enterophages seem to be restricted to human fecal
material, as these were not detected in pig, chicken, cow or dog feces. The present study
determined the inactivation rates and survival times of enterophages under various conditions.
Results were compared to those of human enteric viruses reported elsewhere under similar
conditions. Moreover, the survival of enterophages in beach sand was determined, and their short
survival rates, compared to coliphages, suggest recent fecal pollution. Enterophages could be
considered surrogates of certain human enteric viruses since their inactivation rates and survival
times are similar to those reported. Enterophages exhibit resistances to removal and inactivation
methods, similarly to human enteric pathogens.
Enterophages were tested as indicators of fecal pollution in tropical inland waters as few studies
have evaluated the microbial quality of these waters. One problem is the need to develop
appropriate water quality standards for tropical waters since those used in other geographical
regions may not be applicable. We tested various Microbial Source Tracking (MST) methods
(enerococci, thermotolerant coliforms, enterophages, coliphages, human- and cattle-specific
Bacteroides, chicken-specific Bacteroides and Clostridium, and enterococci by qPCR) to assess
the microbial quality of a tropical watershed in Puerto Rico. Enumeration of both coliphages and
enterophages may be a more appropriate to infer fecal contamination, compared to the

164
bacteriological method. The reason is that these bacteriophages are not ubiquitous in tropical
waters. The human Bacteroides seemed a reliable means of inferring human fecal contamination
in tropical waters, unlike the cattle and chicken-specific Bacteroides and Clostridium markers.
Interestingly, positive correlations between the culture and molecular methods for the detection
of enterococci should be considered prior to implementing qPCR as a rapid method for inferring
fecal pollution in the tropics. The qPCR method for enterococci may not apply to all water
bodies, specifically to tropical inland lakes. qPCR assays targeting enterococci in tropical waters
may overestimate the numbers of enterococci as many molecular methods may also amplify
naked DNA.
The latter study is one of the few (if not the only) that has considered the effect of rainfall events
in the detection of a battery of microbial indicators, coliphages and enterophages in tropical
waters. Rainfall is often not considered when determining the prevalence of microbial indicators
in surface waters. Rainfall events may result in the resuspension of sediments into surface
waters. Sediments often harbor enteric microbes, specifically those that can replicate and/or
persist for long periods in the environment. This represents a concern to public health since many
waterborne outbreaks occur after precipitation events, but can also represent a disadvantage for
traditional indicators. The reason is that if the numbers of a microbial indicator in surface waters
increase after rainfall events may suggest that their input may not be recent. This is turn may
suggest that these microbes have replicated under environmental conditions.
The presence of enterophages in chicken, cattle, dog and pig feces, as well as their prevalence in
domestic sewage were determined using E. faecalis, E. faecium, E. gallinarum, E. hirae, E.
durans, E. dispar, E. casseliflavus and E. pseudoavium as the bacterial hosts. Results showed
that enterophages infect E. faecalis exclusively, and that both humans and animals harbor
Enterococcus phages. Interestingly, phages infecting E. faecium, E. casseliflavus and E.
pseudoavium replicating at 37 °C may be used to infer the presence of chicken-fecal matter in
fresh water sources.
The present project determined the prevalence of tetracycline (tetM) and vancomycin-resistance
(vanA and vanB) and the enterococcal surface protein genes (esp) of E. faecalis and E. faecium in

165
both the bacterial and viral fractions of tropical environmental samples. Detection of the
mentioned genes in the environmental microbiota of tropical samples suggests that the
transmission of these genes is not restricted to strains of the clinical setting. The present study
showed a relatively high prevalence of Enterococcus spp. exhibiting resistance to antibiotics,
which also harbor the respective antibiotic-resistance genes and esp. Although phages did not
harbor the genes tested in the present study, the presence of other genes should not be ignored, as
well as their role as possible vectors. Detection of an integrase-encoding gene in lysogenic
phages will enable the detection of specific antibiotic-resistance and virulence-encoding genes in
their genomes. Importantly, results suggest that lysogenic phages should be considered when
determining the prevalence of bacteriophages in the environment.
FUTURE DIRECTIONS
Future studies need to simultaneously determine the prevalence of enterophages and enteric
viruses in various water types. This will enable direct correlations between indicators and enteric
viruses, and thus could further support the potential of enterophages as surrogates of these
pathogens. In addition, there is still the need to correlate the presence of enterophages with
gastrointestinal illnesses, eye, ear and skin infections.
The detection of enterophages by molecular methods needs to be developed and optimized. For
this reason, the complete genomes of lytic and lysogenic phages infecting E. faecalis are
currently being sequenced in collaboration with the Human Microbiome Project. This will not
only enable to develop the molecular techniques for the detection of these phages in the
environment, but will also represent a means to understand the genomes of lytic and lysogenic
phages infecting E. faecalis. The role of bacteriophages in biogeochemical processes, bacterial
pathogenicity and evolution has been a subject of increasing interest [1]. Many studies have
focused on the effects of strictly lytic phages on bacterial populations and as a possible tool to
treat bacterial infections [2, 3]. Lytic phages may be less genetically diverse (compared to
lysogenic phages) since DNA exchange seems to be more restricted due to their strictly virulent
nature [4]. Lysogenic phages, on the other hand, are the main focus of many genomic studies
since these may help shape bacterial genomes [5, 6]. Lysogenic phages are important vehicles for
DNA segments, which can make the bacterial host more competitive in a current ecosystem or

166
allow it to adapt to a new one [2, 7, 8]. Sequencing lytic and lysogenic E. faecalis phages from
humans may open up the opportunity to further study their possible roles and how these may
affect Enterococcus populations in the intestine. Although many recent studies have focused on
lysogenic phages, understanding the genomic structures of both strictly virulent and lysogenic
phages is important since both phage types are involved in shaping bacterial populations.

167
LITERATURE CITED
1. Brussow, H., C. Canchaya, and W.D. Hardt, Phages and the evolution of bacterial
pathogens: from genomic rearrangements to lysogenic conversion. Microbiol Mol Biol
Rev, 2004. 68(3): p. 560-602.
2. Rohwer, F. and R.V. Thurber, Viruses manipulate the marine environment. Nature, 2009.
459(7244): p. 207-12.
3. Zhang, Q.G. and A. Buckling, Phages limit the evolution of bacterial antibiotic
resistance in experimental microcosms. Evolutionary Applications, 2012.
4. Casjens, S.R., Comparative genomics and evolution of the tailed-bacteriophages. Curr
Opin Microbiol, 2005. 8(4): p. 451-8.
5. Dinsdale, E.A., et al., Functional metagenomic profiling of nine biomes. Nature, 2008.
452(7187): p. 629-32.
6. Muniesa, M., et al., Shiga toxin 2-converting bacteriophages associated with clonal
variability in Escherichia coli O157:H7 strains of human origin isolated from a single
outbreak. Infect Immun, 2003. 71(8): p. 4554-62.
7. Canchaya, C., et al., Phage as agents of lateral gene transfer. Curr Opin Microbiol, 2003.
6(4): p. 417-24.
8. Hendrix, R.W., et al., The origins and ongoing evolution of viruses. Trends Microbiol,
2000. 8(11): p. 504-8.


















