
Enhancement of Oxidative Stress Tolerance in - Plant Physiology
12
Plant Physiol. (1 996) 11 2: 1703-1 71 4 Enhancement of Oxidative Stress Tolerance in Transgenic - Tobacco Plants Overproducing Fe-Superoxide Dismutase in Chloroplasts’ Wim Van Camp, Katelijne Capiau, Marc Van Montagu, Dirk Inzé, and Luit Slooten* Laboratorium voor Genetika, Department of Genetics, Flanders lnteruniversity lnstitute for Biotechnology, Universiteit Gent, K.L. Ledeganckstraat 35, 8-9000 Gent, Belgium (W.V.C., M.V.M., D.I.); Laboratoire Associé de I’lnstitut National de Ia Recherche Agronomique, France (D.I.); and Laboratorium voor Biofysica, Faculteit Wetenschappen, Vrije Universiteit Brussel, Pleinlaan 2, 1050 Brussels, Belgium (K.C., L.S.) A chimeric gene consisting of the coding sequence for chloro- plastic Fe superoxide dismutase (FeSOD) from Arabidopsis thaliana, coupled to the chloroplast targeting sequence from the pea ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit, was expressed in Nicofiana fabacum cv Petit Havana SR1. Expres- sion of the transgenic FeSOD protected both the plasmalemma and photosystem I1 against superoxide generated during illumination of leaf discs impregnated with methyl viologen. By contrast, overpro- duction of a mitochondrial MnSOD from Nicotiana plumbaginifolia in the chloroplasts of cv SR1 protected only the plasmalemma, but not photosystem 11, against methyl viologen (L. Slooten, K. Capiau, W. Van Camp, M. Van Montagu, C. Sybesma, D. lnzé [19951 Plant Physiol 107: 737-750). The difference in effectiveness correlates with different membrane affinities of the transgenic FeSOD and MnSOD. Overproduction of FeSOD does not confer tolerance to HzO2, singlet oxygen, chilling-induced photoinhibition in leaf disc assays, or to salt stress at the whole plant level. In nontransgenic plants, salt stress led to a 2- to 3-fold increase in activity, on a protein basis, of FeSOD, cytosolic and chloroplastic Cu/ZnSOD, ascorbate peroxidase, dehydroascorbate reductase, and glutathione reductase. In FeSOD-overproducing plants under salt stress, the induction of cytosolic and chloroplastic Cu/ZnSOD was suppressed, whereas induction of a water-soluble chloroplastic ascorbate per- oxidase isozyme was promoted. ~ The photosynthetic electron transport chain contains at the acceptor side of PSI a number of autooxidizable en- zymes that reduce oxygen to superoxide (reviewed by Badger, 1985; Asada and Takahashi, 1987; Asada, 1994). These include Fd (Furbank and Badger, 1982; Hosein and Palmer, 1983) and the Fe-S centers X or A/B in the aprotic This work was supported by grants from the Vlaams Actie- programma Biotechnologie (no. 067), the Belgian National Fund for Scientific Research (no. 3.0104.90), the Belgian Programme on Interuniversity Poles of Attraction (Prime Minister’s Office, Sci- ence Policy Programming, no. 38), the International Atomic Agency (no. 5285), and the International Human Frontier Science Program (IHFSP RG 434/94M). W.V.C. is a postdoctoral fellow of the Belgian National Fund for Scientific Research (Belgium). D.I. is a Research Director of the Institut National de Ia Recherche Agronomique (France). * Corresponding author; e-mail Islooten8vnet3.vub.ac.be; fax 32-2-6293389. membrane interior of the PSI reaction center complex (Ta- kahashi and Asada, 1988). The superoxide may mediate cyclic electron flow around PSI (Asada, 1994) or it may diffuse it to the stromal membrane surface, where it is dismutated to oxygen and H,O, in nonenzymic and enzy- mic reactions (see below). Recent evidence suggests that superoxide and H,O, can also be produced by PSII during high-light treatment of thylakoids or intact chloroplasts (Landgraf et al., 1995).H,O, at 10 PM inhibits CO, fixation (Kaiser, 1979); at concentrations as low as 1 PM it causes a substantial inactivation of thiol-modulated Calvin cycle enzymes (Buchanan, 1980, 1991; Takeda et al., 1995). Su- peroxide can inactivate some metal-containing enzymes such as the Fd-linked nitrate reductase, catalase, and per- oxidases (for review, see Asada and Takahashi, 1987).But the chief danger is that H,O, can react with reduced metal ions, especially Fe, resulting in the formation of the hy- droxyl radical. The hydroxyl radical initiates self- propagating reactions leading to peroxidation of mem- brane lipids, base mutations, breakage of DNA strands, and destruction of proteins (Asada and Takahashi, 1987; Halliwell, 1987; Bowler et al., 1992). In addition, there is some evidence that part of the superoxide generated in illuminated chloroplasts diffuses toward the thylakoid lu- men. Because of the low pH in the lumen during illumina- tion, the superoxide can be protonated in that compart- ment, yielding the perhydroxyl radical, which, unlike the superoxide anion, can initiate lipid peroxidation directly (for review, see Asada and Takahashi, 1987). The condi- tions leading to damage caused by active oxygen species will be referred to as oxidative stress. Formation of active oxygen species in the chloroplasts is enhanced when carbon assimilation is inhibited. Oxidative Abbreviations: APx, ascorbate peroxidase (EC 1.11.1.11); AT, 3-amino-1,2,4-triazole; DHAR, dehydroascorbate reductase (EC 1.8.5.1); eosin, 2,4,5,7-tetrabromo-fluorescein; F,/ F, quantum efficiency for exciton trapping by the PSII reaction center in dark- adapted material; GA3P-DH, NADP-dependent glyceraldehyde-3- phosphate dehydrogenase (EC 1.2.1.13); GR, glutathione reductase (EC 1.6.4.2); KCN, potassium cyanide; MDHAR, monodehy- droascorbate reductase (EC 1.6.5.4); MV, methyl viologen; SOD, superoxide dismutase (EC 1.15.1.1); trFeSOD, transgenic (overpro- duced) FeSOD; trMnSOD, transgenic (overproduced) MnSOD. 1703 www.plantphysiol.org on December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.
Transcript of Enhancement of Oxidative Stress Tolerance in - Plant Physiology

Plant Physiol. (1 996) 11 2: 1703-1 71 4
Enhancement of Oxidative Stress Tolerance in Transgenic - Tobacco Plants Overproducing Fe-Superoxide
Dismutase in Chloroplasts’
Wim Van Camp, Katelijne Capiau, Marc Van Montagu, Dirk Inzé, and Luit Slooten*
Laboratorium voor Genetika, Department of Genetics, Flanders lnteruniversity lnstitute for Biotechnology, Universiteit Gent, K.L. Ledeganckstraat 35, 8-9000 Gent, Belgium (W.V.C., M.V.M., D.I.); Laboratoire Associé de I’lnstitut National de Ia Recherche Agronomique, France (D.I.); and Laboratorium voor Biofysica, Faculteit
Wetenschappen, Vrije Universiteit Brussel, Pleinlaan 2, 1050 Brussels, Belgium (K.C., L.S.)
A chimeric gene consisting of the coding sequence for chloro- plastic Fe superoxide dismutase (FeSOD) from Arabidopsis thaliana, coupled to the chloroplast targeting sequence from the pea ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit, was expressed in Nicofiana fabacum cv Petit Havana SR1. Expres- sion of the transgenic FeSOD protected both the plasmalemma and photosystem I1 against superoxide generated during illumination of leaf discs impregnated with methyl viologen. By contrast, overpro- duction of a mitochondrial MnSOD from Nicotiana plumbaginifolia in the chloroplasts of cv SR1 protected only the plasmalemma, but not photosystem 11, against methyl viologen (L. Slooten, K. Capiau, W. Van Camp, M. Van Montagu, C. Sybesma, D. lnzé [19951 Plant Physiol 107: 737-750). The difference in effectiveness correlates with different membrane affinities of the transgenic FeSOD and MnSOD. Overproduction of FeSOD does not confer tolerance to HzO2, singlet oxygen, chilling-induced photoinhibition i n leaf disc assays, or to salt stress at the whole plant level. In nontransgenic plants, salt stress led to a 2- to 3-fold increase in activity, on a protein basis, of FeSOD, cytosolic and chloroplastic Cu/ZnSOD, ascorbate peroxidase, dehydroascorbate reductase, and glutathione reductase. I n FeSOD-overproducing plants under salt stress, the induction of cytosolic and chloroplastic Cu/ZnSOD was suppressed, whereas induction of a water-soluble chloroplastic ascorbate per- oxidase isozyme was promoted.
~
The photosynthetic electron transport chain contains at the acceptor side of PSI a number of autooxidizable en- zymes that reduce oxygen to superoxide (reviewed by Badger, 1985; Asada and Takahashi, 1987; Asada, 1994). These include Fd (Furbank and Badger, 1982; Hosein and Palmer, 1983) and the Fe-S centers X or A/B in the aprotic
This work was supported by grants from the Vlaams Actie- programma Biotechnologie (no. 067), the Belgian National Fund for Scientific Research (no. 3.0104.90), the Belgian Programme on Interuniversity Poles of Attraction (Prime Minister’s Office, Sci- ence Policy Programming, no. 38), the International Atomic Agency (no. 5285), and the International Human Frontier Science Program (IHFSP RG 434/94M). W.V.C. is a postdoctoral fellow of the Belgian National Fund for Scientific Research (Belgium). D.I. is a Research Director of the Institut National de Ia Recherche Agronomique (France).
* Corresponding author; e-mail Islooten8vnet3.vub.ac.be; fax 32-2-6293389.
membrane interior of the PSI reaction center complex (Ta- kahashi and Asada, 1988). The superoxide may mediate cyclic electron flow around PSI (Asada, 1994) or it may diffuse it to the stromal membrane surface, where it is dismutated to oxygen and H,O, in nonenzymic and enzy- mic reactions (see below). Recent evidence suggests that superoxide and H,O, can also be produced by PSII during high-light treatment of thylakoids or intact chloroplasts (Landgraf et al., 1995). H,O, at 10 PM inhibits CO, fixation (Kaiser, 1979); at concentrations as low as 1 PM it causes a substantial inactivation of thiol-modulated Calvin cycle enzymes (Buchanan, 1980, 1991; Takeda et al., 1995). Su- peroxide can inactivate some metal-containing enzymes such as the Fd-linked nitrate reductase, catalase, and per- oxidases (for review, see Asada and Takahashi, 1987). But the chief danger is that H,O, can react with reduced metal ions, especially Fe, resulting in the formation of the hy- droxyl radical. The hydroxyl radical initiates self- propagating reactions leading to peroxidation of mem- brane lipids, base mutations, breakage of DNA strands, and destruction of proteins (Asada and Takahashi, 1987; Halliwell, 1987; Bowler et al., 1992). In addition, there is some evidence that part of the superoxide generated in illuminated chloroplasts diffuses toward the thylakoid lu- men. Because of the low pH in the lumen during illumina- tion, the superoxide can be protonated in that compart- ment, yielding the perhydroxyl radical, which, unlike the superoxide anion, can initiate lipid peroxidation directly (for review, see Asada and Takahashi, 1987). The condi- tions leading to damage caused by active oxygen species will be referred to as oxidative stress.
Formation of active oxygen species in the chloroplasts is enhanced when carbon assimilation is inhibited. Oxidative
Abbreviations: APx, ascorbate peroxidase (EC 1.11.1.11); AT, 3-amino-1,2,4-triazole; DHAR, dehydroascorbate reductase (EC 1.8.5.1); eosin, 2,4,5,7-tetrabromo-fluorescein; F,/ F,,,, quantum efficiency for exciton trapping by the PSII reaction center in dark- adapted material; GA3P-DH, NADP-dependent glyceraldehyde-3- phosphate dehydrogenase (EC 1.2.1.13); GR, glutathione reductase (EC 1.6.4.2); KCN, potassium cyanide; MDHAR, monodehy- droascorbate reductase (EC 1.6.5.4); MV, methyl viologen; SOD, superoxide dismutase (EC 1.15.1.1); trFeSOD, transgenic (overpro- duced) FeSOD; trMnSOD, transgenic (overproduced) MnSOD.
1703 www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from
Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1704 Van Camp et al. Plant Physiol. Vol. 11 2, 1996
stress arises, for example, when high irradiance is com- bined with chilling temperatures, drought, or heat (Bowler et al., 1992). Salt stress is another condition that may give rise to oxidative stress, as suggested by the increase in activities of antioxidant enzymes in response to high salin- ity, and by the correlation of salt tolerance with antioxidant enzyme levels (Gossett et al., 1994; Olmos et al., 1994; Hernandez et al., 1995; Sehmer et al., 1995).
An elaborate antioxidant system, a defense against oxi- dative stress, is present in the chloroplasts. SOD catalyzes the dismutation of superoxide into oxygen and H,O,. SODs are classified, according to their metal cofactor, as FeSOD, MnSOD, or Cu / ZnSOD. Chloroplasts generally contain Cu/ZnSOD and, in a number of plant species, FeSOD (Van Camp et al., 1994a). APx reduces H,O, to water, with ascorbate as the electron donor. Chloroplastic APx was recently found to occur in a stromal and a membrane-bound form (Miyake and Asada, 1992). The re-reduction of the reaction product of APx, monodehy- droascorbate, proceeds along different pathways, depend- ing on the type of APx involved. The reaction product of stromal APx is reduced by NADPH, either directly or via glutathione. The enzymes taking part in this so-called ascorbate-glutathione cycle (Foyer and Halliwell, 1976; Na- kano and Asada, 1981) are GR, DHAR, and MDHAR (for reviews, see Asada and Takahashi, 1987; Halliwell, 1987). In contrast, monodehydroascorbate produced by membrane-bound APx is re-reduced directly by Fd (Mi- yake and Asada, 1994).
The overproduction of antioxidant enzymes provides a way to study the role of these enzymes in the antioxidant system, and to study the contribution of oxidative stress tolerance to tolerance to physiological types of stress. Over- production of SOD in the chloroplasts has been found to result in enhanced oxidative stress tolerance in transgenic tobacco (Bowler et al., 1991; Sen Gupta et al., 1993a; Foyer et al., 1994; Van Camp et al., 1994b; Slooten et al., 1995), alfalfa (McKersie et al., 1993), potato (Perl et al., 1993), and, according to preliminary data, in cotton (Allen, 1995). Sim- ilar results were obtained with overproduction of SOD in the mitochondria of alfalfa (McKersie et al., 1993) and in the cytosol of potato (Perl et al., 1993). In many of these studies, oxidative stress tolerance was assessed in assays based on the use of MV. This herbicide passes electrons from various electron transport chains to oxygen, generat- ing superoxide. During illumination, MV generates super- oxide primarily in the chloroplasts (Halliwell, 1984; Slooten et al., 1995) and thus simulates the oxidative stress compo- nent of the environmental stresses. In addition, an en- hanced tolerance to freezing stress in transgenic alfalfa overproducing SOD in the chloroplasts has been reported (McKersie et al., 1993), and transgenic tobacco overproduc- ing SOD in the chloroplasts exhibits an enhanced tolerance to chilling in the dark (Foyer et al., 1994) or in the light (Sen Gupta et al., 1993a).
We have shown that expression of plant mitochondrial MnSOD in the chloroplasts of transgenic Nicofiana fabacum cv SR1 and cv PBD6 reduces cellular damage generated by treatment with MV (Bowler et al., 1991; Slooten et al., 1995)
or ozone (Van Camp et al., 1994b). In PBD6, overproduc- tion of MnSOD had a clear protective effect on MV-induced ion leakage and, albeit to a lesser extent, also on MV- induced inactivation of the PSII reaction center. In SR1, protection was observed with regard to ion leakage, but not with regard to the PSII reaction center (Slooten et al., 1995). Apparently, improvement in the antioxidant defense was more pronounced at the plasmalemma than at PSII, in spite of the fact that the overproduced SOD is located in the chloroplasts. It is possible that the pathway from MV- mediated superoxide generation at PSI to damage at PSII is poorly accessible to plant mitochondrial MnSOD expressed in the chloroplasts. This lack of protection of PSII might be specific for overproduced plant mitochondrial MnSOD, since chloroplastic overproduction of Cu/ ZnSOD in potato (Perl et al., 1993) and of Escherickia coli MnSOD in tobacco (Foyer et al., 1994) does alleviate the inhibition of photo- synthesis by MV.
Here we report that overproduction of FeSOD in the chloroplasts of N. tabacum cv SR1 protects both the plas- malemma and PSII from MV-induced damage. This dem- onstrates that FeSOD provides better protection of chloro- plasts than MnSOD. We argue that this is because FeSOD, in contrast to MnSOD, is indigenous to the chloroplasts. Functional differences between the FeSOD and MnSOD enzymes of E. coli have been reported with regard to pro- tection of DNA and proteins (Hopkin et al., 1992). The data presented in this study, to our knowledge, are the first to demonstrate that in plant chloroplasts, FeSOD and MnSOD have different protective properties that may be related to their suborganellar location. We also show that overpro- duction of FeSOD interferes with signal pathways, leading to induction of cytosolic Cu/ZnSOD during salt stress. In addition, overproduction of FeSOD leads to induction of chloroplastic APx during salt stress.
MATERIALS AND METHODS
Ceneration of FeSOD-Overproducing Plants
To construct pEXSOD10, a PCR product was generated with the 5' primer TCAAGTGCTGTAGATCTAAAC- TACGTCCTC (positions 54-83 of pSOD10, BglII site intro- duced) and the 3' primer ACACACAAAACGGATCCA- CACTCAGAAAAG (complementary to positions 842-871 of pSOD10, BamHI site introduced) using pSODlO as a template. pSODlO contains an FeSOD cDNA from Arabi- dopsis fkaliana (Van Camp et al., 1990). The amplified frag- ment was cloned in a dT-tailed, HincII-digested pUC18. The fragment was re-excised by a BamHI-BglII digest and cloned into the BamHI site of pKAHl (Bowler et al., 1991). This generates an in-frame fusion of the chloroplast transit peptide derived from the small subunit of Rubisco from pea with the FeSOD mature protein. Compared with the mature FeSOD expressed in A. fkaliana, the mature fusion protein contains an additional Met after processing. The chimeric construct was cloned as a ClaI-BamHI fragment into the ClaI-BamHI-digested binary vector pGSJ780A (Bowler et al., 1991). This generated the cassette for chlo- roplastic FeSOD overproduction under control of the cau-
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Tobacco Overproducing Fe-Superoxide Dismutase 1705
liflower mosaic virus 35s promoter. Transformation of Nicotiana tabacum var Petit Havana SR1 with this cassette, which was named pEXSOD10, was according to the meth- ods of De Block et al. (1987).
Plant Material
Plants were grown in pots on peat-based compost con- taining fertilizer and were watered with demineralized water to which, after bolting, 1 vol % of a commercial fertilizer (NPK 6-3-6) was added. Unless otherwise indi- cated, the plants were grown in a 12-h light/12-h dark cycle, with day and night temperatures of about 22 and 16"C, respectively. The light, provided by mercury-halogen vapor lamps with a daylight spectrum (HQI-T, Osram, Munich, Germany), had an intensity of approximately 135 pmol m-' s-*. Unless indicated otherwise, the experiments were carried out on the seventh to ninth leaves from the top at 10 to 12 weeks after sowing.
In some experiments the plants were transferred to a growth chamber at 1 month after sowing (i.e. at the fifth- leaf stage), at which time they were grown in a 12-h light / 12-h dark cycle at 22/15"C. In the course of 8 d the light intensity was gradually raised from 180 to 650 pmol m-' s-*. The light intensity remained at this leve1 during the next 14 d, and the experiments were performed at the end of this period.
In the salt-stress experiments, plants were grown at an intensity of 65 pmol m-'s-', with day and night temper- atures of about 25 and 20"C, respectively. The plants were watered with demineralized water, to which 60 mM NaCl was added from d 12 after sowing.
Assessment of Oxidative Stress Tolerance
Leaf discs of approximately 1.5 cm2 were preincubated overnight with reagents that mediate the formation of toxic oxygen species during subsequent illumination of the leaf discs: MV generates superoxide (Bowler et al., 1991), AT generates H,O, (see "Results"), and eosin generates singlet oxygen (Knox and Dodge, 1985). The MV assays were carried out as described by Slooten et al. (1995). In the case of AT or eosin treatment, the leaf discs were placed in Petri dishes containing 3 mL of aqueous solutions of these com- pounds. The Petri dishes were put in a gas-tight, thermo- statted container with a glass lid. The leaf discs were illu- minated with white light from mercury-halogen vapor lamps, filtered by 8 cm of water. The light intensity was 360 pmol m-' s-I. The temperature in the Petri dishes during illumination was maintained at 18°C. After illumination the leaf discs were incubated in the dark for at least 2 h to ensure complete dark adaptation prior to measuring F,/ F,,, as a measure of the activity of the PSII reaction cen- ters. This ratio gives the exciton trapping efficiency when a11 photochemical traps are open (Genty et al., 1989). The fluorescence measurements were made as described by Slooten et al. (1995). The conductance of the floating solu- tions was determined as a measure of ion leakage from the leaf discs, which was due to lipid peroxidation of the cell membranes (Slooten et al., 1995).
For the photoinhibition experiments, the leaf discs were floated overnight on an aqueous solution of 75 p~ of the chloroplast translation inhibitor chloramphenicol prior to illumination. At this concentration chloramphenicol does not act as a PSI electron acceptor (Okada et al., 1991).
Biochemical Assays
The basal medium for the preparation of leaf disc ex- tracts for enzyme assays contained 50 mM potassium phos- phate, 0.1% Triton X-100, 20% (w/v) SUC, 2% polyvinyl- polypyrrolidone, 1 mM EDTA, 1 mM EGTA, 1 mM PMSF, 4 mM benzamidine, 4 mM caproic acid, and KOH to pH 7.6. SUC and polyvinylpolypyrrolidone were omitted from the basal medium used for the preparation of extracts for SOD determinations. Whole-leaf extracts were prepared by grinding leaf discs (0.2 g) in an ice-cold mortar with 1.2 mL of basal medium supplemented with the following addi- tions: for APx, GR, and MDHAR, 30 mM sodium ascorbate; for GA3P-DH, 30 mM sodium ascorbate and 0.05% (w/v) P-mercaptoethanol; for DHAR, 2 mM sodium ascorbate; and for SOD, 0.05% (w/v) P-mercaptoethanol and 0.1 mg/mL BSA. The extracts were centrifuged for 30 min at 40,OOOg. The supernatants were stored on ice and used the same day. The assays of APx, DHAR, GR, and MDHAR were carried out as described previously (Slooten et al., 1995). GA3P-DH was assayed as described by Stitt et al. (1989). For these assays, 1 unit equals 1 pmol of substrate converted per min.
APx activity on nondenaturing polyacrylamide gels was detected as described by Mittler and Zilinskas (1993). SOD activity was determined first in a solution assay, and then after an activity staining following electrophoresis on non- denaturing polyacrylamide gels (Bowler et al., 1991). The gels were stained as described previously (Slooten et al., 1995) and then scanned with a one-dimensional scanning densitometer. The relative contribution of each of the SOD isoforms to the overall activity was determined from the contribution of the area under the corresponding peak of the densitogram to the total area. The overall SOD activity in leaf disc extracts was determined in the solution assay with the same assay mixture. In the solution assay one unit of SOD activity causes a half-maximal inhibition of the rate of light-induced, riboflavin-mediated reduction of ni- troblue tetrazolium. In both assays the activity was mea- sured with and without 2 mM KCN, which causes a virtu- ally complete inhibition of Cu/ZnSOD (Geller and Winge, 1984).
For the preparation and washing of chloroplasts we used the low-salt grinding and resuspension media de- scribed by Cerovic and Plesnicar (1984), except that P-mercaptoethanol was added to both media at 0.05% (v/ v). Chloroplasts were prepared by grinding 4 g of leaf material for 3 s in 40 mL of ice-cold grinding medium in a blender (Waring). The homogenate was filtered through four layers of Miracloth (Calbiochem) and centrifuged for 1.5 min at 600g at 4°C. The sedimented chloroplasts were washed twice in 20 mL of resuspension medium, followed by centrifugation as before.
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1706 Van Camp et al. Plant Physiol. Vol. 11 2, 1996
Chlorophyll in whole-leaf extracts was determined as described previously (Slooten et al., 1995). Protein was estimated according to Bradford (1976) using BSA as a standard. A11 absorbance measurements were carried out on a spectrophotometer (Cary 2300, Varian Techron, Mul- grave, Victoria, Australia).
RESULTS
SOD Activity Levels in Nonstressed Transgenic and Control Plants
To overexpress chloroplastic FeSOD from A. thaliana in tobacco chloroplasts, we made a chimeric gene in which the FeSOD coding region was fused in frame with the coding sequence for the chloroplast transit peptide of the Rubisco small subunit. The expression of this gene fusion was driven by the 35s promoter from the cauliflower mo- saic virus. Relatively high levels of FeSOD activity were observed in leaf extracts of seven primary transformants. Six of these lines, each containing a single copy of the transgene, were made homozygous and selected for further study. Most of the data shown below were obtained with line SRl-14, but the results were confirmed with severa1 other lines.
Figure 1 shows the banding patterns observed after non- denaturing electrophoresis of leaf extracts or chloroplasts from mature leaves, followed by SOD activity staining. The bands are numbered in order of increasing mobility. In leaf extracts from nontransgenic plants (Fig. lA, dashed line), three bands were observed after staining without KCN:
1
1
Figure 1 . Densitogram of the SOD-banding pattern of leaf extracts (A and B) and chloroplasts (C and D) from FeSOD-overproducing plants of line SR1-14 (solid lines) and control plants (dashed lines). The gels were stained in the absence (A and C) or presence (B and D) of 2 mM KCN. Migration i s from left to right. Bands 1 and 2, Engineered FeSOD; band 3, a mixture of cytosolic Cu/ZnSOD and endogenous FeSOD; band 3’, endogenous FeSOD; bands 4 and 5, chloroplastic Cu/ZnSOD.
Table 1. SOD activities in leaf extracts from FeSOD-overproducing and control plants
Mean 2 SE (13 plants). Activity
SR1 SR1-14 SOD Species
unitshg chlorophyll MnSOD 0.62 ? 0.20 0.52 ? 0.1 7 FeSOD 1.88 2 0.1 9 13.85 2 0.63 Chloroplastic Cu/ZnSOD 5.23 k 1.33 3.67 k 0.79 Cytosolic Cu/ZnSOD 6.04 2 0.64 6.70 Ifr 0.80
band 3 contains both chloroplastic FeSOD and cytosolic Cu/ZnSOD, and bands 4 and 5 represent chloroplastic Cu/ZnSOD (Slooten et al., 1995). Preincubation with KCN prior to staining caused a complete inactivation of Cu/ ZnSOD; the remaining activity represents only FeSOD (Fig. lB, dashed line, band 3’). In chloroplast extracts from nontransgenic plants (Fig. lC, dashed line), cytosolic Cu/ ZnSOD was absent, and band 3’ again corresponds to chloroplastic FeSOD. Pretreatment with KCN abolished the chloroplastic Cu/ ZnSOD activity, but had little effect on the activity of chloroplastic FeSOD (Fig. lD, dashed line). Extracts from immature leaves also contain a small, slow- moving band corresponding to mitochondrial MnSOD (not shown) (Slooten et al., 1995).
Leaf and chloroplast extracts from FeSOD-overproducing plants (line SR1-14) exhibited two extra bands (nos. 1 and 2) representing trFeSOD (Fig. 1, solid lines). Similar results were obtained with other transgenic lines (not shown). In the KCN-treated lanes, the endogenous FeSOD shows up as a small shoulder at the fast-moving side of band 2 (Fig. 1, B and D, solid lines). A comparison with the corresponding traces from nontransgenic plants indicates that FeSOD- overproducing plants contained about one-half of the en- dogenous FeSOD activity that was found in nontransgenic plants. Thus, either trFeSOD leads to a suppression of the endogenous FeSOD activity, or some of the endogenous FeSOD forms heterodimers with trFeSOD.
We used the SOD activity gels to estimate the activities of the different SOD isoforms, as described in ”Materials and Methods.” The staining intensities of the bands on the activity gels indicate that KCN, which was used to inacti- vate Cu/ZnSOD, also caused an inhibition of both endog- enous and overproduced FeSOD. This can be seen most clearly in extracts prepared from isolated chloroplasts, which do not contain cytosolic Cu/ZnSOD (Fig. 1, C and D). The KCN inhibition amounted to 20 to 30% for both endogenous and overproduced FeSOD. In the gels without KCN, the contribution of cytosolic Cu/ZnSOD to band 3 was obtained by subtracting the contribution of FeSOD. This contribution was estimated from the KCN-treated gels, taking the observed KCN inhibition by FeSOD into account. The results for line SR1-14 are shown in Table I. In leaf extracts from SR1-14, we found an average of 13.8 units/mg chlorophyll of FeSOD, most of which represents transgenic FeSOD (cf. Fig. 1). There was no significant difference in activity of mitochondrial MnSOD and cytoso- lic and chloroplastic Cu/ZnSOD between transgenic and control plants.
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Tobacco Overproducing Fe-Superoxide Dismutase 1707
Table II. KCN-insensitive SOD activity in leaf extracts from differ- ent transgenic lines overproducing FeSOD
Mean t SE (four plants, unless indicated otherwise). Line Activity
SR1 (control) SR1-23-1 SR1-36-7 SR1-42-8 SR1-14 SR1-32-2 SR1-21-1
u n i t s h g chlorophyll 2.88 2 0.43
10.56 2 0.71 10.84 i 0.39 12.34 2 0.73 14.37 i 0.65" 17.21 ? 0.85 17.32 i 1 .O4
a 13 plants.
The data shown in Table I indicate that the total activity of chloroplastic SOD species (FeSOD and chloroplastic Cu/ZnSOD) was, on a chlorophyll basis, in line SR1-14 approximately 2.5 times higher than in control plants. However, chloroplastic Cu/ ZnSOD is quite variable in activity; it occurs only in young, expanding leaves, and previous experiments yielded no indication that it pro- vides tolerance against light-dependent, MV-mediated oxidative stress (Slooten et al., 1995). Therefore, the ratio of FeSOD contents may be more relevant in this respect. The chloroplastic FeSOD activity was, on a chlorophyll basis, in line SR1-14 approximately 7.4 times higher than in control plants.
The SOD activity observed in the presence of KCN rep- resented mainly FeSOD (cf. Fig. l), sometimes with a minor contribution by MnSOD (less than 10% of the total KCN- insensitive activity, as indicated by the banding patterns on the SOD activity gels). Table I1 shows the KCN-insensitive activities observed in the different transgenic lines. The difference in KCN-insensitive SOD activity between the highest and the lowest expressor amounted to only approx- imately 60%.
-A---- - &
10
5 --o Control i -0- SRI -1 4
o 0. 0.0 0.5 1.0 1.5 2.0
Oxidative Stress Tolerance in Transgenic and Control Plants
Plants overproducing FeSOD were clearly more tolerant to MV than control plants. This is shown in Figure 2 for young plants (9 weeks) and older plants (13 weeks) of line SR1-14. These plants were grown at a light intensity of 135 pmol m-' s-'. Older plants are considerably more tolerant to MV than young plants, as indicated by the difference in scales along the x axis. We reported previously that in var SR1, overproduction of MnSOD provided protection against MV-induced ion leakage (due to cell membrane deterioration), but not against MV-induced inactivation of PSII (Slooten et al., 1995). In contrast, overproduction of FeSOD resulted in protection against both types of damage. This is indicated by the fact that the extent of MV-induced ion leakage (Fig. 2, A and B), as well as the extent of the MV-induced decrease in F,/F,,, (Fig. 2, C and D), were lower in transgenic plants than in control plants. Similar results were obtained with other transgenic lines (Fig. 3). Furthermore, similar results were obtained with plants grown at a light intensity of 650 pmol m-' s-' (Table 111), although these plants were 15 to 30 times less sensitive to MV than plants grown at 135 pmol m-' s-'.
AT inhibits catalase (Margoliash et al., 1960; Havir, 1992) and causes an increase in the H,O, content of tobacco leaves during illumination (Chen et al., 1993); at the same time, it causes inactivation of endogenous, chloroplastic APx. Apparently as a consequence of the resulting increase in the concentration of H,O, in the chloroplasts, the PSII reaction center is inactivated. This is not accompanied by any significant ion leakage from the leaf discs (L. Slooten, K. Capiau, S. Kushnir, M. Van Montagu, and D. Inzé, unpublished data). Plants overproducing FeSOD were not more tolerant to AT than control plants (Table IV).
Leaf discs impregnated with eosin generate singlet oxy- gen during illumination (Knox and Dodge, 1985). Plants overproducing FeSOD were not more tolerant to eosin than
Figure 2. MV-induced conductance increase (A and B) and MV-induced decrease in FJF,,, (ex- pressed as a percentage of no MV) (C and D) in leaf discs from transgenic and control plants at 9 weeks (A and C) or 14 weeks (B and D) after sowing. Conductance increase was expressed in microsiemens cm-' 35 mg-' fresh weight. Error bars indicate SD for f o u r plants. Open symbols, Control plants; solid symbols, SR1-14.
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1708 Van Camp et al. Plant Physiol. Vol. 1 1 2, 1996
Figure 3. MV-induced conductance increase (A), and MV-induced decrease in FJF,,,,, (ex- pressed as a percentage of no MV) (B) in leaf discs from transgenic and control plants at 1 1 weeks after sowing. The M V concentration was 2 p ~ . Conductance increase was expressed in microsiemens cm-’ 35 mg-’ fresh weight. Error bars indicate SE for four plants. Numbers in the figures indicate the significance of the difference in mean values between transgenic and control plants, as established with a two-sided t test.
control plants. It made no difference in this respect whether the damage was assessed from the decrease in activity of the PSII reaction center (Table IV), or from the increase in the conductance of the floating solution (not shown).
In addition, plants overproducing FeSOD were not more tolerant to chilling-induced photoinhibition than control plants (Table V). It made no difference in this respect whether the photoinhibitory treatment was carried out for 4 h at 360 pmol m-’s-’ or for 30 h at 30 pmol m-’s-*.
Membrane Affinity of trFeSOD and trMnSOD
We reported previously that in var SR1, overproduction of MnSOD provided protection against MV-induced ion leakage (due to cell membrane deterioration), but not against MV-induced inactivation of PSII (Slooten et al., 1995). In contrast, overproduction of FeSOD resulted in protection against both types of damage. Jt seemed possi- ble that this might be due to a different suborganellar location of the overproduced MnSOD and FeSOD. Specif- ically, these enzymes might have a different membrane affinity. To test this hypothesis, we determined the activity of the overproduced SODs and of a stromal marker en- zyme, NADP-dependent GABP-DH, in washed chloro- plasts from MnSOD-overproducing plants and from FeSOD-overproducing plants. We made use of the obser- vation that a considerable proportion of the chloroplasts became leaky during isolation and lost their stromal con- tent before washing. One would expect that electrostati- cally membrane-associated enzymes would be washed out to a lesser extent than stromal enzymes. Prior to assay, the suspension was sonicated to the extent that the chloro- plasts that until then had remained intact were disrupted in a medium in which the enzymes were quantitatively released. In addition, we measured the activity of
Table 111. Enhancement of MV tolerance in FeSOD-overproducing plants
MV-induced conductance increase (in microsiemens cm-’ 35 mg-’ fresh weight) and MV-induced decrease in FJF,,,,, (in percent- age of no MV) in leaf discs from transgenic and control plants grown a ta light intensity of 650 pmol m-’s-’. The M V concentration was 14 p ~ . Mean -+ SE for six plants.
Plants Conductance lncrease Decrease in FJF,,,,,
SRI-14 1 .I6 2 0.49 10.12 rt 2.02 SR1 4.02 rt 1.41 24.15 -+ 2.02
0.99
23-1
A
> 0.995
36-7
B 1
0.97 0.98 0.94 E Y 5 ‘= 20
g 10
i
.- a? VI
* control 21-1 23-1 36-7
GA3P-DH and of the overproduced SODs in leaf extracts prepared from the same leaf as the chloroplasts. A11 activ- ities were expressed on a chlorophyll basis. For each en- zyme, the recovery in the chloroplasts was calculated as the ratio of activities in sonicated chloroplasts to that in leaf extracts. The recovery of GA3P-DH was approximately 35%. We then calculated the ratio of recoveries of GA3P-DH over the overproduced SOD. This ratio is ex- pected to be 1 for a stromal enzyme and lower than 1 for a membrane-associated enzyme. Finally, we calculated the ratio of recoveries of FeSOD and MnSOD (R,,,,,/R,,,,,) as the ratio between RCA3P.DH/RMnSOD in MnSOD- overproducing plants, and RCA3P.DH / RFeSOD in FeSOD- overproducing plants, where R is the recovery of the en- zyme indicated in the subscript. This method allowed us to compare recoveries of FeSOD and MnSOD corrected for differences in quality of the chloroplast preparation. This procedure was repeated five times in independent experi- ments. The results are shown in Table VI. Overproduced MnSOD exhibited about the same recovery as GA3P-DH, indicating that it behaves like a stromal enzyme. Overpro- duced FeSOD exhibited higher recoveries, indicating elec- trostatic binding to chloroplast membranes. This was espe- cially clear from the average ratio of recoveries of FeSOD and MnSOD, calculated as indicated above.
Effect of trFeSOD on lnduction of Antioxidant Enzymes during Salt Stress
Plants respond to salt stress with increases in antioxidant enzyme activities, indicating that oxidative stress is in- volved in salt stress (Gossett et al., 1994; Hernandez et al., 1995; Sehmer et al., 1995). To study the effect of FeSOD overproduction on salt tolerance, we grew transgenic and control plants under salt stress (see “Materials and Meth-
Table IV. Lack of enhancement of A J and eosin tolerance in FeSO D-o verproducing pla n ts
Leaf discs were illuminated for 16 h in the presence of 5 mM AT, or for 2 h in the presence of 0.1 mM eosin. For additional details, see ”Materials and Methods.” Mean 2 SE (number of plants).
Decrease in FJF,,,
AT Eosin Plants
% of dark controls SRI 31 2 1.4 (5) 44.1 2 2.5 (6) SRI-14 30 t 2.6 (5) 47.7 2 3.0 (6)
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Tobacco Overproducing Fe-Superoxide Dismutase 1709
Table V. Lack of enhancement of tolerance to chilling-induced photoinhibition in FeSOD- overproducing plants
Leaf discs floating on water containing 75 WM chloramphenicol were illuminated as indicated in closed Petri dishes. FJF,,,,, was measured 2 h after cessation of illumination. Mean t SE (number of plants).
Photoinhibitory Treatment Decrease in FJF,,,
Liaht intensitv Duration Temoerature SR1 SR1-14
pmol m-z 5 - l % of dark control 39.1 t 1.3 (6)
30 31 h 4 "C 24.8 t 2.7 (4) 28.6 t 2.1 (4) 360 4 h 7 "C 36.5 t 1.5 (6)
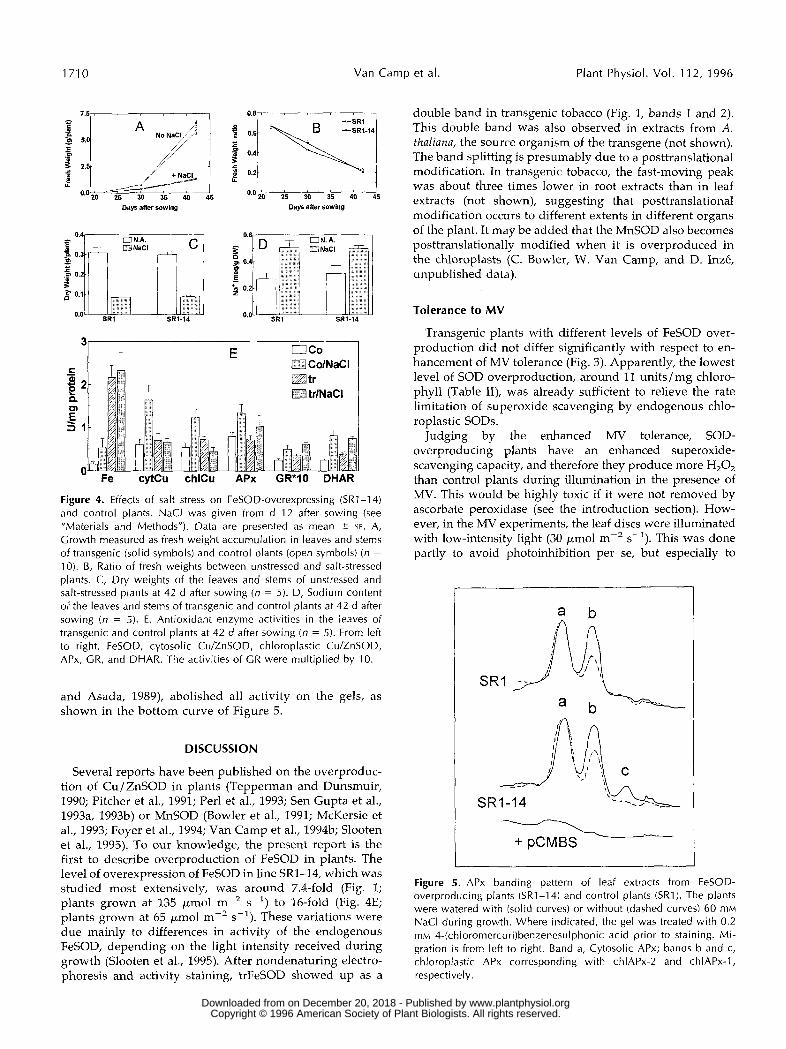
ods"). Salt-stressed plants accumulated considerably less biomass, in terms of fresh weight, than control plants (Fig. 4A). The salt concentration in the soil increased gradually with time, so that growth became increasingly inhibited (not shown). As a consequence, the ratio of fresh weights in salt-stressed plants over fresh weights in control plants decreased steadily with time (Fig. 4B). There was no dif- ference in this respect between FeSOD-overproducing and control plants. In addition, there was no difference in dry matter accumulation between FeSOD-overproducing and control plants at the end of the experiment (Fig. 4C). At the end of the experiment, the concentration of dissolved so- dium in the soil was 120 mM in the salt-treated pots, compared with 5 mM in the controls (not shown), and the sodium content of salt-stressed plants was approximately twice that of the control plants (Fig. 4D).
Salt stress was accompanied by oxidative stress, as indi- cated by increases in activities of a11 tested antioxidant enzymes in nontransgenic plants. Specifically, the activities of chloroplastic FeSOD, cytosolic Cu / ZnSOD, chloroplastic Cu / ZnSOD, APx, GR, and DHAR were approximately two to three times higher, on a protein basis, in salt-stressed nontransgenic plants than in unstressed nontransgenic plants at the end of the experiment (Fig. 4E). FeSOD- overproducing plants exhibited a similar increase in over- a11 activity of APx, GR, and DHAR during salt stress. However the induction of cytosolic and chloroplastic Cu /
ZnSOD, observed in control plants under salt stress, was suppressed in FeSOD-overproducing plants (Fig. 4E).
To discriminate between chloroplastic and cytosolic APx isozymes, we electrophoresed leaf extracts on non- denaturing polyacrylamide gels, and stained the gels for APx activity (Fig. 5). Three bands of APx activity can be distinguished after nondenaturing electrophoresis of leaf extracts from bolting tobacco plants: one band corre- sponding to cytosolic APx, and two bands corresponding to chloroplastic APx (L. Slooten, K. Capiau, S. Kushnir, M. Van Montagu, and D. Inzé, unpublished data). The latter two will be denoted as chlAPx-1 and chlAPx-2 for ease of reference. The plants used in the present experiments were still in the rosette stage, and were grown at a rela- tively low light intensity. Under those circumstances, only the cytosolic APx and chlAPx-2 were observed in extracts from unstressed, nontransgenic plants. Under salt stress, nontransgenic plants usually exhibited an increase in ac- tivity of chlAPx-2 in comparison with nonstressed plants. Unstressed, FeSOD-overproducing plants exhibited a sim- ilar isozyme pattern as unstressed nontransgenic plants. However, during salt stress, the FeSOD-overproducing plants exhibited an increase in activity of both chloroplas- tic APx isozymes. Thus, the induction of chlAPx-1 was accelerated specifically in FeSOD-overproducing plants under salt stress. 4-(Chloromercuri)benzenesulphonic acid, a specific inhibitor of ascorbate peroxidase (Chen
Table VI. Comparison of the recoveries of overproduced SODs in isolated chloroplasts Washed chloroplasts were resuspended in 2.4 mL of the extraction medium used for determination
of GA3P-DH in whole leaf extracts (see "Materials and Methods"), except that polyvinylpolypyrroli- done, Triton, and ascorbate were omitted. The chloroplasts were broken by sonication. Part of the sonicate was used for chlorophyll determination. The remainder was centrifuged for 1 h at 45,OOOg. The supernatants were used for assay of SOD (spectrophotometric, as well as after nondenaturing PACE) and of GA3P-DH. All data are presented as mean t SD from five independent experiments. The activities of trMnSOD and trFeSOD in leaf extracts from transgenic plants were 11.8 2 1.7 and 13.8 t 2.3 units/mg chlorophyll, respectively. The activities of GA3P-DH in leaf extracts from MnSOD- and FeSOD- overproducing plants were 10.3 t 0.9 and 10.4 t 2.4 units/mg chlorophyll, respectively. R is the recovery in isolated chloroplasts of the enzyme indicated in the subscript. For other details, see text.
FeSOD-Overproducing- Plants ~
~
MnSOD-Overproducing Plants
RM",OV 39.9 t i 4 . a ~ ~ RkSOD 50.4 t 18.4% RCA3PDH 38.8 t 11.4% RGA3P.DH 35.5 t 18.8% R,,,,,$LSOD 1 .O3 t 0.41 0.69 t 0.25 RGA3P-DH/RFeSOV
RF~SOD ~- - 1.50 2 0.12 RMnSOD
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.
1710 Van Camp et al. Plant Physiol. Vol. 11 2, 1996
e!
Days after sowing Days after sowing
0.4 E
% 0 3
E D 0 2
a -
6 0.1
SRI SR1-14 SRI-14 0.0
E c7 co
C .- $ 2 n
3 1 E"
o Fe cytCu chlCu APx GR*10 DHAR
Figure 4. Effects of salt stress on FeSOD-overexpressing (SR1-14) and control plants. NaCl was given from d 12 after sowing (see "Materials and Methods"). Data are presented as mean i- SE. A, Growth measured as fresh weight accumulation in leaves and stems of transgenic (solid symbols) and control plants (open symbols) (n =
10). B, Ratio of fresh weights between unstressed and salt-stressed plants. C, Dry weights of the leaves and stems of unstressed and salt-stressed plants at 42 d after sowing (n = 5). D, Sodium content of the leaves and stems of transgenic and control plants at 42 d after sowing (n = 5). E, Antioxidant enzyme activities in the leaves of transgenic and control plants at 42 d after sowing (n = 5). From left to right, FeSOD, cytosolic Cu/ZnSOD, chloroplastic CulZnSOD, APx, GR, and DHAR. The activities of GR were multiplied by 10.
and Asada, 1989), abolished all activity on the gels, as shown in the bottom curve of Figure 5.
DISCUSSION
Severa1 reports have been published on the overproduc- tion of Cu/ZnSOD in plants (Tepperman and Dunsmuir, 1990; Pitcher et al., 1991; Perl et al., 1993; Sen Gupta et al., 1993a, 1993b) or MnSOD (Bowler et al., 1991; McKersie et al., 1993; Foyer et al., 1994; Van Camp et al., 1994b; Slooten et al., 1995). To our knowledge, the present report is the first to describe overproduction of FeSOD in plants. The level of overexpression of FeSOD in line SR1-14, which was studied most extensively, was around 7.4-fold (Fig. 1; plants grown at 135 pmol m-' sK1) to 16-fold (Fig. 4E; plants grown at 65 Fmol m-' sK1). These variations were due mainly to differences in activity of the endogenous FeSOD, depending on the light intensity received during growth (Slooten et al., 1995). After nondenaturing electro- phoresis and activity staining, trFeSOD showed up as a
double band in transgenic tobacco (Fig. 1, bands 1 and 2). This double band was also observed in extracts from A. tkaliana, the source organism of the transgene (not shown). The band splitting is presumably due to a posttranslational modification. In transgenic tobacco, the fast-moving peak was about three times lower in root extracts than in leaf extracts (not shown), suggesting that posttranslational modification occurs to different extents in different organs of the plant. It may be added that the MnSOD also becomes posttranslationally modified when it is overproduced in the chloroplasts (C. Bowler, W. Van Camp, and D. Inzé, unpublished data).
Tolerance to MV
Transgenic plants with different levels of FeSOD over- production did not differ significantly with respect to en- hancement of MV tolerance (Fig. 3). Apparently, the lowest level of SOD overproduction, around 11 units / mg chloro- phyll (Table 11), was already sufficient to relieve the rate limitation of superoxide scavenging by endogenous chlo- roplastic SODs.
Judging by the enhanced MV tolerance, SOD- overproducing plants have an enhanced superoxide- scavenging capacity, and therefore they produce more H202 than control plants during illumination in the presence of MV. This would be highly toxic if it were not removed by ascorbate peroxidase (see the introduction section). How- ever, in the MV experiments, the leaf discs were illuminated with low-intensity light (30 pmol mK2 s-'). This was done partly to avoid photoinhibition per se, but especially to
Figure 5. APx banding pattern of leaf extracts from FeSOD- overproducing plants (SR1-14) and control plants (SR1). The plants were watered with (solid curves) or without (dashed curves) 60 m M NaCl during growth. Where indicated, the gel was treated with 0.2 mM 4-(chloromercuri)benzenesulphonic acid prior to staining. Mi- gration is from left to right. Band a, Cytosolic APx; bands b and c, chloroplastic APx corresponding with chlAPx-2 and chlAPx-1, respectively.
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Tobacco Overproducing Fe-Superoxide Dismutase 1711
avoid a situation in which the H,O,-scavenging capacity of endogenous APx would be overwhelmed by a high rate of superoxide production and scavenging. In addition, we have evidence that in tobacco leaf discs, H,O, by itself does not readily inactivate either the PSII reaction center or the plasmalemma (L. Slooten, K. Capiau, S. Kushnir, M. Van Montagu, and D. Inzé, unpublished data). Presumably, both of these factors contributed to the clear-cut protection by trFeSOD of the PSII reaction center against damage induced by superoxide.
We reported previously that in var SR1, overproduction of MnSOD provided protection against MV-induced ion leakage (due to cell membrane deterioration), but not against MV-induced inactivation of PSII (Slooten et al., 1995). In contrast, overproduction of FeSOD resulted in protection against both types of damage. This difference cannot be attributed to differences in activity levels of overproduced SOD, since plants overproducing MnSOD contained on average 13 units/mg chlorophyll of the overproduced activity (Slooten et al., 1995), which is sim- ilar to that for FeSOD-overproducing plants. However, trFeSOD is by origin a chloroplastic enzyme, whereas trMnSOD is by origin a mitochondrial enzyme. During illumination, the major site of electron donation to MV has been shown to be the Fe-S center B in PSI (Fujii et al., 1990). From there the oxygen radicals (produced by reoxi- dation of reduced MV) must propagate out of the chloro- plasts and to the cell membrane, resulting in ion leakage, and to the reaction center of PSII, resulting in the loss of F , / F,,,. To prevent a11 types of superoxide-induced dam- age, the overproduced SOD should scavenge the super- oxide at the site of its formation, i.e. at the acceptor side of PSI. To achieve this the SOD probably has to bind elec- trostatically to a specific domain in the stromal membrane in which PSI is embedded. The charge distribution on the SODs may be such that trFeSOD is better able to recognize these binding domains than trMnSOD. The results pre- sented in Table VI are consistent with this hypothesis: trFeSOD is at least partially membrane-bound, whereas trMnSOD behaves like a stromal enzyme. The protection provided by trFeSOD against inactivation of the PSII re- action center may thus be due to trFeSOD binding in the vicinity of the PSI reaction center, enabling it to scavenge superoxide at the site of its formation. By contrast, trMn- SOD, behaving like a stromal enzyme, may only be capa- ble of providing protection against damage spreading through the stroma toward the cell membrane and lead- ing to ion leakage (Slooten et al., 1995).
It will certainly be necessary to confirm this hypothesis by determining the suborganellar localization of trFeSOD in the chloroplasts. But it may be noted that in spinach, both chloroplastic (Ogawa et al., 1995b) and cytosolic Cu/ ZnSOD (Ogawa et al., 1995a) are preferentially membrane- associated; the chloroplastic Cu/ ZnSOD is associated with the stromal membranes and with stroma-facing thylakoid membranes (Ogawa et al., 199513). Furthermore, functional differences between FeSOD and MnSOD were also ob- served in E. coli, in which MnSOD is more effective than FeSOD in protecting DNA and FeSOD is more effective
than MnSOD in protecting a cytosolic enzyme against MV- mediated oxygen radical damage. Here, too, the differences were attributed to differences in charge and/or charge distribution between MnSOD and FeSOD (Hopkin et al., 1992).
Tolerance to Other Types of Oxidative Stress
The lack of enhancement of tolerance to systems gener- ating eosin or H,O, (Table IV) is in accordance with the fact that these toxic oxygen species are not utilized by SOD. Overproduction of FeSOD did not enhance the tolerance to photoinhibitory conditions either (Table V). Mainly from in vitro studies with various PSII preparations, it was con- cluded that the primary event in photoinhibition is inhibi- tion of whole-chain electron transport, either at the accep- tor side or at the donor side of the PSII reaction center (for review, see Prasil et al., 1992; Aro et al., 1993; Barber, 1994). During acceptor-side photoinhibition, active oxygen spe- cies formed by the still-functional primary radical pair initiate the degradation of the D1 protein of the reaction center. Most of the attention has been focused on singlet oxygen (Vass et al., 1992; Hideg et al., 1994), but superoxide has been implicated as well (Miyao, 1994). During donor- side photoinhibition, degradation of the D1 protein is not strictly dependent on oxygen, yet the degradation is strongly accelerated by superoxide (Chen et al., 1995), which can be generated by the still-functional primary radical pair. Superoxide production becomes manifest only after complete removal of Mn, which is somehow involved in a SOD-like activity displayed by the reaction center itself (Ananyev et al., 1994; Chen et al., 1995). Donor-side pho- toinhibition can occur under conditions that destabilize the water-splítting complex, such as exposure to low temper- ature (Wang et al., 1992). In contrast to acceptor-side pho- toinhibition, donor-side photoinhibition can occur at low light intensities (Eckert et al., 1991).
In the present study, overproduction of FeSOD did not enhance the tolerance to chilling-induced photoinhibition, either at low or high light intensities (Table V). This is in agreement with the results obtained with MnSOD- overproducing plants (Slooten et al., 1995). There are sev- era1 possible explanations of this result: (a) photoinhibition was not due to superoxide production in the reaction cen- ter; (b) photoinhibition was due to superoxide production in the reaction center, but the superoxide brought about oxidative damage without becoming accessible to overex- pressed SOD; and (c) the reaction centers did produce superoxide accessible to transgenic SOD, but this was then converted to H,O,, which can be equally damaging (Miyao-Tokutomi et al., 1995). In light of the above discus- sion, we assume that explanations b and c are more likely than explanation a. In view of the results discussed in the previous section (indicating that trFeSOD does protect the PSII reaction center against superoxide produced by PSI), it would seem that explanation b is more likely than expla- nation c.
Sen Gupta et al. (1993a, 199313) found that transgenic tobacco (N. tabacum cv Xanthi) overproducing pea Cu/ ZnSOD in the chloroplasts exhibited an enhanced tolerance
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1712 Van Camp et al. Plant Physiol. Vol. 11 2, 1996
to inhibition of CO, fixation that was induced by high light at low temperatures. This indicates that in the experiments of Sen Gupta et al. (1993a, 1993b), inhibition of CO, fixation was caused by the production of superoxide at a rate exceeding the endogenous scavenging capacity. Sen Gupta et al. (1993a, 1993b) used lower temperatures and higher light intensities during the stress treatment than we did. Hence, the rate of superoxide production may have been higher in their experiments than in ours. However, in the experiments of Sen Gupta al. (1993a), the recovery from chilling-induced inhibition of CO, fixation was virtually complete within about 20 min after return to 25°C. Recov- ery from oxidative damage to the PSII reaction center generally requires severa1 hours (Thiele et al., 1995; cf. Greer et al., 1986). Therefore, it seems unlikely that in the experiments of Sen Gupta et al. (1993a), the inhibition of CO, fixation was due to oxidative damage to the PSII reaction center.
Effect of trFeSOD on lnduction of Antioxidant Enzymes during Salt Stress
We observed two chloroplastic APx isozymes on APx activity gels (Fig. 5) (L. Slooten, K. Capiau, S. Kushnir, M. Van Montagu, and D. Inzé, unpublished data). These are water-soluble enzymes and are therefore different from the intrinsically membrane-bound APx first found in spinach (Miyake and Asada, 1992). We found a similar membrane- bound APx activity in cuvette assays in membrane prepa- rations from tobacco chloroplasts, but could not visualize this activity on APx activity gels (L. Slooten, K. Capiau, S. Kushnir, M. Van Montagu, and D. Inzé, unpublished data).
In nonstressed tobacco plants, overproduction of FeSOD had no effect on activity levels of other antioxidant en- zymes, including APx. This was also the case when the plants were grown at a light intensity of 650 pmol m-'s-' (see "Materials and Methods") (not shown). This agrees with earlier data obtained with transgenic plants overpro- ducing MnSOD in the chloroplasts (Slooten et al., 1995). However, when we applied salt stress, only the transgenic, FeSOD-overproducing plants exhibited at least a 3-fold increase in activity of chlAPx-1. In contrast, chlAPx-2 was enhanced in both the transgenic and the nontransgenic plants in response to salt stress (Fig. 5). This suggests that at least chlAPx-1 is induced by H,O,. During salt stress, H,O, is probably generated at higher rates in transgenic, FeSOD-overproducing plants than in nontransgenic plants. The differential response of the two chloroplastic APx isozymes to salt stress indicates some functional differen- tiation of these isozymes. In var SR1, chlAPx-1 represents only a minor fraction of the total chloroplastic APx. We noted that leaf extracts from cv PBDG contained higher activities of chlAPx-1 than leaf extracts from cv SR1 after electrophoresis on nondenaturing gels (L. Slooten, K. Ca- piau, S. Kushnir, M. Van Montagu, and D. Inzé, unpub- lished data). Sen Gupta et al. (1993b) reported that trans- genic tobacco (N. tabacum cv Xanthi) overproducing pea Cu / ZnSOD in the chloroplasts exhibited a 3-fold increase in APx activity. The difference between the results of their experiments and ours may be due to differences in the APx
isozyme distribution pattern between cv Xanthi and cv SR1.
In accordance with data from other groups, we found that salt stress leads to increases in the activity of many antioxidant enzymes (chloroplastic FeSOD, cytosolic Cu / ZnSOD, chloroplastic Cu/ZnSOD, APx, GR, and DHAR) in nontransgenic plants, indicating that oxidative stress is one of the components of salt stress (Fig. 4E). In the absence of stress, the activities of a11 of these enzymes except FeSOD were the same in nontransgenic and FeSOD-overproducing plants. However, the increase in activity of cytosolic and chloroplastic Cu/ ZnSOD, observed in control plants under salt stress, was suppressed in FeSOD-overproducing plants (Fig. 4E). Cytosolic Cu/ZnSOD and trFeSOD are located in different cellular compartments, indicating that trFeSOD interferes with a signal pathway leading to induction of cytosolic Cu/ ZnSOD, and probably also of chloroplastic Cu/ZnSOD. This interference is probably due to the fact that trFeSOD lowers the concentration of superoxide in the chloroplasts. Similar results were obtained previously with plants overproducing a mitochondrial MnSOD in the chlo- roplasts (Slooten et al., 1995). It remains to be determined whether superoxide itself or a molecule derived from a reaction with superoxide constitutes the signal for induc- tion of cytosolic Cu/ZnSOD.
Salt stress is complex, imposing a water deficit because of osmotic effects and exerting ion-specific toxic effects on a wide variety of metabolic activities (Greenway and Munns, 1980; Cheeseman, 1988). The water deficit might be expected to cause oxidative stress, which is similar to drought stress (Bowler et al., 1992). Indeed, Singha and Choudhuri (1990) reported that superoxide and H,O, could be important in the mechanism of salt injury. This notion was corroborated by subsequent reports indicating an increase in the activities of antioxidant enzymes in response to high salinity, and by correlations of salt tolerance with antioxidant enzyme levels (Gossett et al., 1994; Olmos et al., 1994; Hernandez et al., 1995; Sehmer et al., 1995). In our experiments the chloro- plasts of salt-stressed transgenic plants exhibited elevated activities not only of SOD but also of APx in comparison with salt-stressed control plants (see above). Nevertheless, FeSOD-overproducing plants were no more tolerant to salt stress (as indicated by biomass accumulation; Fig. 4, A-C) than were control plants. This suggests that in the present experiments, growth of salt-stressed plants was not limited by the capacity of the chloroplasts to scavenge superoxide and H,O,. It cannot be excluded that the lack of induction of cytosolic Cu / ZnSOD in transgenic plants overproducing FeSOD contributed to the lack of enhancement of salt toler- ante in these plants.
CONCLUSIONS AND PROSPECTS
The results obtained with transgenic plants overproduc- ing either FeSOD or MnSOD in the chloroplasts indicate that FeSOD provides better protection against MV-dependent oxidative stress than does MnSOD. We attribute this tenta- tively to the higher membrane affinity of trFeSOD, allowing this enzyme to scavenge superoxide radicals at the site of their formation, i.e. near PSI. This difference between Mn-
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Tobacco Overproducing Fe-Superoxide Dismutase 1713
SOD and FeSOD is probably connected to the original sub- cellular localization of these enzymes. From a11 this it may be anticipated that FeSOD-overproducing plants will be more tolerant to physiological stresses entailing enhancement of light-induced superoxide formation than MnSOD- overproducing plants. FeSOD-overproducing plants were not more tolerant to salt stress than control plants, indicating that, a t least under the present assay conditions, the superoxide-scavenging capacity w a s not a limiting factor for growth under salt stress. However in salt-stressed plants the overproduced enzyme interfered with signal pathways for induction of antioxidant enzymes i n such a manner that induction of one chloroplastic APx isozyme was promoted and induction of cytosolic and chloroplastic Cu/ ZnSOD w a s inhibited. Thus, these plants may provide clues for the elu- cidation of signal pathways involved i n induction of other antioxidant enzymes. Furthermore, it seems likely that FeSOD-overproducing plants can provide interesting mate- rial to assess the importance of oxidative stress i n various physiological types of stress, and in addition, provide a good starting point for studying cooperative effects between different overproduced antioxidant enzymes in the scaveng- ing of toxic oxygen species.
Received May 13, 1996; accepted September 9, 1996. Copyright Clearance Center: 0032-0889/96/ 112/ 1703/12.
LITERATURE CITED
Allen RD (1995) Dissection of oxidative stress tolerance using transgenic plants. Plant Physiol107 1049-1054
Ananyev G, Renger G, Wacker U, Klimov V (1994) The produc- tion of superoxide radicals and the superoxide dismutase activ- ity of photosystem 11. The possible involvement of cytochrome b559. Photosynth Res 41: 327-338
Aro E-M, Virgin I, Andersson B (1993) Photoinhibition of photo- system 11: inactivation, protein damage and turnover. Biochim Biophys Acta 1143: 113-134
Asada K (1994) Production and action of active oxygen species in photosynthetic tissues. In CH Foyer, PM Mullineaux, eds, Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants. CRC Press, Boca Raton, FL, pp 77-104
Asada K, Takahashi M (1987) Production and scavenging of active oxygen in photosynthesis. In DJ Kyle, CB Osmond, CJ Arntzen, eds, Photoinhibition (Topics in Photosynthesis, Vol9). Elsevier, Amsterdam, pp 227-287
Badger MR (1985) Photosynthetic oxygen exchange. Annu Rev Plant Physiol 36: 27-53
Barber J (1994) Molecular basis of the vulnerability of photosystem I1 to damage by light. Aust J Plant Physiol 22: 201-208
Bowler C, Slooten L, Vandenbranden S, De Rycke R, Botterman J, Sybesma C, Van Montagu M, Inzé D (1991) Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J 10: 1723-1732
Bowler C, Van Montagu M, Inzé D (1992) Superoxide dismutase and stress tolerance. Annu Rev Plant Physiol Plant Mo1 Biol 43:
Bradford MM (1976) A rapid and sensitive method for the quan- tification of microgram quantities of protein utilizing the prin- ciple of protein-dye binding. Ana1 Biochem 72: 248-254
Buchanan BB (1980) Role of light in the regulation of chloroplast enzymes. Annu Rev Plant Physiol31: 341-374
Buchanan BB (1991) Regulation of CO, assimilation in oxygenic photosynthesis: the ferredoxin/ thioredoxin system. Arch Bio- chem Biophys 288: 1-9
83-116
Cerovic ZG, Plesnicar M (1984) An improved method for the isolation of intact chloroplasts of high photosynthetic capacity. Biochem J 223: 543-545
Cheeseman JM (1988) Mechanism of salt tolerance in plants. Plant Physiol87 547-550
Chen G-X, Asada K (1989) Ascorbate peroxidase in tea leaves: occurrence of two isozymes and the differences in their enzy- matic and molecular properties. Plant Cell Physiol 3 0 987-998
Chen G-X, Blubaugh DJ, Homann PH, Golbeck JH, Cheniae GM (1995) Superoxide contributes to the rapid inactivation of spe- cific secondary donors of the photosystem I1 reaction center during photodamage of manganese-depleted photosystem I1 membranes. Biochemistry 34: 2317-2332
Chen Z, Silva H, Klessig DF (1993) Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 262 1883-1886
De Block M, Botterman J, Vandewiele M, Dockx J, Thoen C, Gosselé V, Movva R, Thompson C, Van Montagu M, Leemans J (1987) Engineering herbicide resistance in plants by expression of a detoxifying enzyme. EMBO J 6: 2513-2518
Eckert H-J, Geiken B, Bernarding J, Napiwotzki A, Eichler H-J, Renger G (1991) Two sites of photoinhibition of the electron transfer in oxygen evolving and Tris-treated PSII membranes from spinach. Photosynth Res 2 7 97-108
Foyer CH, Descourvières P, Kunert KJ (1994) Protection against oxygen radicals: an important defense mechanism studied in transgenic plants. Plant Cell Environ 17: 507-523
Foyer CH, Halliwell B (1976) The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascor- bic acid metabolism. Planta 133: 21-25
Fujii T, Yokoyama E-i, Inoue K, Sakurai H (1990) The sites of electron donation of photosystem 1 to methyl viologen. Biochim Biophys Acta 1015: 41-48
Furbank RT, Badger MA (1982) Oxygen exchange associated with electron transport and photophosphorylation in spinach chloro- plasts. Biochim Biophys Acta 723: 400-409
Geller BL, Winge DR (1984) Subcellular distribution of superox- ide dismutases in rat liver. Methods Enzymol 105: 105-114
Genty B, Briantais J-M, Baker NR (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta
Gossett DG, Millhollon EP, Lucas MC (1994) Antioxidant re- sponse to NaCl stress in salt-tolerant and salt-sensitive cultivars of cotton. Crop Sci 3 4 706-714
Greenway H, Munns R (1980) Mechanism of salt tolerance in non-halophytes. Annu Rev Plant Physiol 31: 149-190
Greer DH, Berry JA, Bjorkman O (1986) Photoinhibition of pho- tosynthesis in intact bean leaves: role of light and temperature, and requirement for chloroplast-protein synthesis during recov- ery. Planta 168: 253-260
Halliwell B (1984) Chloroplast Metabolism: The Structure and Function of Chloroplasts in Green Leaf Cells. Clarendon Press, Oxford, UK
Halliwell B (1987) Oxidative damage, lipid peroxidation and anti- oxidant protection in chloroplasts. Chem Phys Lipids 44: 327-340
Havir EA (1992) The in vivo and in vitro inhibition of catalase from leaves of Nicotiana sylvestris by 3-amino-1,2,4-triazole. Plant Physiol 99: 533-537
Hernandez JA, Olmos E, Corpas FJ, Sevilla F, De1 Rio LA (1995) Salt-induced oxidative stress in chloroplasts of pea plants. Plant Sci 105: 151-167
Hideg E, Spetea C, Vass I (1994) Singlet oxygen production in thylakoid membranes during photoinhibition as detected by EPR spectroscopy. Photosynth Res 39: 191-199
Hopkin KA, Papazian MA, Steinman HM (1992) Functional dif- ferences between manganese and iron superoxide dismutases in Escherichia coli K-12. J Biol Chem 267: 24253-24258
Hosein B, Palmer G (1983) The kinetics and mechanism of oxida- tion of reduced spinach ferredoxin by molecular oxygen and its reduced products. Biochim Biophys Acta 723: 383-390
990: 87-92
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1714 Van Camp et al. Plant Physiol. Vol. 11 2 , 1996
Kaiser WM (1979) Reversible inhibition of the Calvin cycle and activation of oxidative pentose phosphate cycle in isolated intact chloroplasts by hydrogen peroxide. Planta 145: 377-382
Knox JP, Dodge AD (1985) The photodynamic action of eosin, a singlet-oxygen generator: some effects on leaf tissue of Pisum sativum L. Planta 164: 22-29
Landgraf P, Doege M, Ohmann E, Tschiersch H (1995) Light stress and reactive oxygen species: consequences for photosyn- thesis in Euglena gradis . In P Mathis, ed, Photosynthesis: From Light to Biosphere, Vol IV. Kluwer Academic Publishers, Dor- drecht, The Netherlands, pp 465-468
Margoliash E, Novogrodsky A, Schejter A (1960) Irreversible reaction of 3-amino-l:2:4-triazole and related inhibitors with the protein of catalase. Biochem J 74: 339-350
McKersie BD, Chen Y, de Beus M, Bowley SR, Bowler C, Inzé D, DHalluin K, Botterman J (1993) Superoxide dismutase en- hances tolerance of freezing stress in transgenic alalfa (Medicago sativa L.). Plant Physiol 103: 1155-1163
Mittler R, Zilinskas BA (1993) Detection of ascorbate peroxidase activity in native gels by inhibition of ascorbate-dependent re- duction of nitroblue tetrazolium. Ana1 Biochem 212: 540-546
Miyake C, Asada K (1992) Thylakoid-bound ascorbate peroxidase in spinach chloroplasts and photoreduction of its primary oxi- dation product monodehydroascorbate radicals in thylakoids. Plant Cell Physiol 33: 541-553
Miyake C, Asada K (1994) Ferredoxin-dependent photoreducion of the monodehydroascorbate radical in spinach thylakoids. Plant Cell Physiol 35: 539-549
Miyao M (1994) Involvement of active oxygen species in degra- dation of the D1 protein under strong illumination in isolated subcomplexes of photosystem 11. Biochemistry 33: 9722-9730
Miyao-Tokutomi M, Okada K, Yamamoto N (1995) Specific cleav- age of the D1 protein of PSII by exogenous active oxygen spe- cies. In P Mathis, ed, Photosynthesis: From Light to Biosphere, Vol IV. Kluwer Academic Publishers, Dordrecht, The Nether- lands, pp 243-246
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22: 867-880
Ogawa K, Kanematsu S , Asada K (1995a) Spinach chloroplastic and cytosolic CuZnSODs are localized at the sites of superoxide generation. In P Mathis, ed, Photosynthesis: From Light to Bio- sphere, Vol IV. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 339-342
Ogawa K, Kanematsu S , Takabe K, Asada K (199513) Attachment of CuZn-superoxide dismutase to thylakoid membranes at the site of superoxide generation (PSI) in spinach chloroplasts: de- tection by immunogold labeling after rapid freezing and substi- tution method. Plant Cell Physiol 3 6 565-573
Okada K, Satoh K, Katoh S (1991) Chloramphenicol is an inhibitor of photosynthesis. FEBS Lett 295: 155-158
Olmos E, Hernandez JA, Sevilla F, Hellin E (1994) Induction of severa1 antioxidant enzymes in the selection of a salt-tolerant cell line of Pisum sativum. J Plant Physiol 144 594-598
Perl A, Perl-Treves R, Galili S , Aviv D, Shalgi E, Malkin S , Galun E (1993) Enhanced oxidative-stress defense in transgenic potato overexpressing tomato Cu,Zn superoxide dismutases. Theor Appl Genet 85: 568-576
Pitcher LH, Brennan E, Hurley A, Dunsmuir P, Tepperman JM, Zilinskas BA (1991) Overproduction of petunia chloroplastic copper / zinc superoxide dismutase does not confer ozone toler- ante in transgenic tobacco. Plant Physiol97: 452455
Prasil O, Adir N, Ohad I (1992) Dynamics of photosystem 11: mechanism of photoinhibition and recovery processes. In J Bar- ber, ed, Topics in Photosynthesis, Vol 11. The Photosystems: Structure, Function and Molecular Biology. Elsevier, Amster- dam, pp 295-348
Sehmer L, Alaoui-Sosse B, Dizengremel P (1995) Effect of salt stress on growth and on the detoxifying pathway of pedunculate oak seedlings (Quercus robur L.). J Plant Physiol 147: 144-151
Sen Gupta A, Heinen JL, Holaday AS, Burke JJ, Allen RD (1993a) Increased resistance to oxidative stress in transgenic plants that overexpress chloroplastic Cu / Zn superoxide dismutase. Proc Natl Acad Sci USA 90: 1629-1633
Sen Gupta A, Webb RP, Holaday AS, Allen RD (1993b) Overex- pression of superoxide dimutase protects plants from oxidative stress. Plant Physiol 103: 1067-1073
Singha S, Choudhuri MA (1990) Effect of salinity (NaCl) on H,02 metabolism in Vigna and Oryza seedlings. Biochem Physiol Pflanz 186: 69-74
Slooten L, Capiau K, Van Camp W, Van Montagu M, Sybesma C, Inzé D (1995) Factors affecting the enhancement of oxidative stress tolerance in transgenic tobacco overexpressing manganese suyeroxide dismutase in the chloroplasts. Plant Physiol 107:
Stitt M, McC Lilley R, Gerhardt R, Heldt HW (1989) Metabolite levels in specific cells and subcellular compartments of plant leaves. Methods Enzymol 174: 518-552
Takahashi M, Asada K (1988) Superoxide production in the aprotic interior of thylakoid membranes. Arch Biochem Biophys
Takeda T, Yokota A, Shigeoka S (1995) Resistance of photosyn- thesis to hydrogen peroxide in algae. Plant Cell Physiol 3 6
Tepperman JM, Dunsmuir P (1990) Transformed plants with el- evated levels of chloroplastic SOD are not more resistant to superoxide toxicity. Plant Mo1 Biol 14 501-511
Thiele A, Winter K, Krause GH (1995) Photoinhibition of photo- synthesis related to xanthophyll cycle and D1 protein turnover in higher plants. In P Mathis, ed, Photosynthesis: From Light to Biosphere, Vol IV. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 437-440
Van Camp W, Bowler C, Villarroel R, Tsang EWT, Van Montagu M, Inzé D (1990) Characterization of iron superoxide dismutase cDNAs from plants obtained by genetic complementation in Escherichia coli. Proc Natl Acad Sci USA 87: 9903-9907
Van Camp W, Van Montagu M, Inzé D (1994a) Superoxide dis- mutases. In CH Foyer, PM Mullineaux, eds, Causes of Photooxi- dative Stress in Plants and Amelioration of Defense Systems. CRC Press, Boca Raton, FL, pp 317-341
Van Camp W, Willekens H, Bowler C, Van Montagu M, Inzé D, Reupold-Popp R, Sandermann H Jr, Langebartels C (199413) Elevated levels of superoxide dismutase protect transgenic plants against ozone damage. Bio/ Technology 12: 165-168
Vass I, Styring S, Hundal T, Koivuniemi A, Aro E-M, Andersson B (1992) Reversible and irreversible intermediates during pho- toinhibition of photosystem 11: stable reduced Q, species pro- mote chlorophyll triplet formation. Proc Natl Acad Sci USA 89:
Wang WS, Chapman DJ, Barber J (1992) Effect of cold treatments on the binding stability of photosystem two extrinsic proteins and associated increase in susceptibility to photoinhibition. Plant Physiol 99: 21-25
737-750
267: 714-722
1089-1095
1408-1412
www.plantphysiol.orgon December 20, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.


















