
Effekte skrotaler Hyperthermie auf die Perfusion sowie auf ... · welche aus der Arteria spermatica...
133
Tierärztliche Hochschule Hannover Effekte skrotaler Hyperthermie auf die Perfusion sowie auf histologische und immunhistochemische Merkmale des caninen Hodens und Nebenhodens INAUGURAL – DISSERTATION zur Erlangung des Grades einer Doktorin der Veterinärmedizin - Doctor medicinae veterinariae - ( Dr. med. vet. ) vorgelegt von Annika Herr Hamburg Hannover 2010
Transcript of Effekte skrotaler Hyperthermie auf die Perfusion sowie auf ... · welche aus der Arteria spermatica...

Tierärztliche Hochschule Hannover
Effekte skrotaler Hyperthermie auf die Perfusion sowie auf histologische und immunhistochemische Merkmale
des caninen Hodens und Nebenhodens
INAUGURAL – DISSERTATION
zur Erlangung des Grades einer
Doktorin der Veterinärmedizin - Doctor medicinae veterinariae -
( Dr. med. vet. )
vorgelegt von
Annika Herr Hamburg
Hannover 2010

Wissenschaftliche Betreuung: Prof. Dr. A.-R. Günzel-Apel
Reproduktionsmedizinische Einheit der Kliniken
Klinik für Kleintiere
1. Gutachter: Prof. Dr. A.-R. Günzel-Apel
2. Gutachter: Prof. Dr. C. Pfarrer
Tag der mündlichen Prüfung: 20.05.2010

Meinen Eltern Meinem Freund
Frieda


1 Einleitung .............................................................................................9
2 Literaturübersicht..............................................................................11
2.1 Anatomie des Hundehodens.........................................................11
2.1.1 Morphologie..................................................................................................... 11
2.1.2 Blutgefäßversorgung der Hoden...................................................................... 13
2.1.3 Histologie des Hodens..................................................................................... 13
2.2 Spermatogenese, Spermiogenese und Keimepithelzyklus........16
2.3 Apoptose.........................................................................................17
2.3.1 Allgemein......................................................................................................... 17
2.3.2 Nachweisverfahren der Apoptose.................................................................... 20
2.3.3 Apoptose am Hoden........................................................................................ 21
2.4 Nekrose ...........................................................................................23
2.5 Degenerative Prozesse an den Hoden .........................................23
2.6 Proliferation am Hoden..................................................................24
2.6.1 Allgemein......................................................................................................... 24
2.6.2 Nachweisverfahren.......................................................................................... 24
2.7 Sonographie der Hoden.................................................................25
2.7.1 Technische Grundlagen................................................................................... 25
2.7.2 Dopplersonographie ........................................................................................ 25
2.7.3 Auswertung von Dopplerkurven....................................................................... 27
2.7.4 Zweidimensionale Sonographie der Hoden ..................................................... 28
2.7.5 Dopplersonographie der Hoden....................................................................... 29
2.7.6 Zweidimensionale Sonographie der Prostata .................................................. 29
2.8 Wärmeregulation am Hoden..........................................................30
2.9 Einfluss einer Hyperthermie auf das Hodengewebe...................31
3 Material und Methoden .....................................................................35
3.1 Tiere.................................................................................................35
3.2 Versuchsaufbau .............................................................................36
3.2.1 Vorlauf ............................................................................................................. 36

3.2.2 Wärmeapplikation............................................................................................ 37
3.2.3 Untersuchungen nach 48-stündiger Wärmeapplikation ................................... 38
3.3 Morphologische Untersuchung ....................................................39
3.4 Ultraschalluntersuchung ...............................................................40
3.5 Kastration und Aufbereitung der Hoden......................................40
3.6 Histologische und immunhistochemische Verfahren.................42
3.6.1 Hämalaun-Eosin Färbung................................................................................ 42
3.6.2 TUNEL - Verfahren.......................................................................................... 42
3.6.3 Immunhistologischer Nachweis der aktivierten Form der Caspase-3 .............. 42
3.6.4 Ki-67-Antigen................................................................................................... 43
3.7 Mikroskopische Auswertung ........................................................44
3.8 Statistische Auswertung ...............................................................45
4 Ergebnisse .........................................................................................46
4.1 Befunde der morphologischen Untersuchung............................46
4.2 Befunde der sonographischen Untersuchung ............................49
4.3 Histologische und immunhistochemische Befunde ...................56
4.3.1 Hoden.............................................................................................................. 56
4.3.2 Nebenhoden .................................................................................................... 73
5 Diskussion .........................................................................................81
5.1 Versuchsdurchführung..................................................................81
5.2 Morphologische Befunde ..............................................................82
5.3 Sonographische Befunde ................................................................................... 83
5.4 Histologische Befunde........................................................................................ 85
5.5 Schlussfolgerungen.......................................................................95
6 Zusammenfassung............................................................................96
7 Summary............................................................................................99
8 Literaturverzeichnis ........................................................................102
9 Anhang.............................................................................................114

9.1 Histologische und immunhistochemische Verfahren – Arbeitsprotokolle................................................................................114
9.2 Skrotale Wärmeapplikation und Aufbereitung der Hoden: Materialien, Chemikalien und Geräte ...............................................122
9.3 Verläufe der Skrotaltemperatur...................................................125
9.4 Alter und Gewicht der Rüden......................................................127

Abkürzungsverzeichnis
Ak = Antikörper
°C = Grad Celsius
c = Geschwindigkeit der Ultraschallwellen
ca. = circa
cm = Zentimeter
DNA = Desoxyribonucleic acid = Desoxyribonucleinsäure
DPV = diastolic peak velocity = diastolische Maximalgeschwindigkeit
EDV = enddiastolic velocity = enddiastolische Geschwindigkeit
evt. = eventuell
HE = Hämalaun-Eosin
kg = Kilogramm
m = Meter
min = Minute
ml = Milliliter
mm = Millimeter
MHz = Megahertz
μm = Mikrometer
PBS = phosphatgepufferte Natriumchlorid-Lösung
PCR = Polymerase Ketten Reaktion
PI = pulsatility index = Pulsatilitätsindex
RI = resistance index = Widerstandsindex
RNA = Ribonucleic acid
SPV = systolic peak velocity = systolische Maximalgeschwindigkeit
s = Sekunde
TAMAX = time averaged maximum velocity
= amplitudengewichtete mittlere Geschwindigkeit
TNF = Tumor Nekrose Faktor
TUNEL = TdT-mediated dUTP-biotin nick end labelling
v = relative Geschwindigkeit
z.B. = zum Beispiel

Einleitung
9
1 Einleitung
Die Bedeutung der Fruchtbarkeit ihrer Hunde rückt zunehmend in das Bewusstsein
der Hundezüchter. Züchten ist nicht mehr nur ein Hobby, sondern für viele Züchter
auch ein wirtschaftlicher Faktor. Aus diesem Grunde besteht zunehmendes Interesse
an einer umfassenden Fertilitätsdiagnostik auch bei Rüden.
Eine herabgesetzte Fruchtbarkeit kann unter Anderem auf einer zu geringen
Spermienproduktion in den Hoden bestehen. Degenerative Veränderungen des
spermienbildenen Hodenparenchyms entstehen in der Mehrzahl der Fälle ohne eine
akute klinische Symptomatik. Vielmehr kommt es häufig über eine aufsteigende
Infektion zu einer subakuten oder chronischen Orchitis mit schleichender,
fortschreitender Abnahme der Fruchtbarkeit bis zur Infertilität. Von anderen
Haustierarten ist bekannt, dass es aufgrund einer Hyperthermie zu einer akuten,
klinisch nicht apparenten Entzündung, jedoch in der Folge zu reversiblen oder
irreversiblen degenerativen Gewebeveränderungen kommen kann. So wurden bei
Ratten, Bullen, Hengsten und Lamas massive Effekte einer kurzzeitigen moderaten
Hyperthermie auf die Ejakulatbeschaffenheit sowie auf morphologische und
histologische Hodenparameter nachgewiesen (KHAN u. BROWN 2002; ALBRECHT
2006; SCHWEIZER 2006; SCHWALM et al. 2007).
Die Spermatogenese läuft beim Hund in 8 bis 10 Keimepithelzyklen ab (FOOTE et al.
1972; IBACH et al. 1976; SOARES et al. 2009). Es ist davon auszugehen, dass auch
unter physiologischen Bedingungen nicht alle Spermatogonien zu Spermien werden,
sondern viele dem programmierten Zelltod, der Apoptose unterworfen sind. Apoptose
kann durch verschiedene externe und interne Faktoren ausgelöst werden (MAJNO u.
JORIS 1995). So z.B. durch Erhöhung der Skrotaltemperatur. Eine weitere Form des
Zelluntergangs ist die Nekrose, die im Gegensatz zur Apoptose keinen genetisch
kontrollierten Prozess darstellt und häufig mit einer Entzündung einhergeht.

Einleitung
10
Ziel der vorliegenden Studie war es, den Einfluss einer kurzfristigen moderaten
skrotalen Hyperthermie auf Morphologie und gewebliche Beschaffenheit der Hoden
gesunder Beagle-Rüden zu untersuchen sowie deren Ausmaß und Reversibilität zu
erfassen.

Literaturübersicht
11
2 Literaturübersicht
2.1 Anatomie des Hundehodens
2.1.1 Morphologie
Die Hoden des Hundes liegen im Hodensack und sind kaudal im
Zwischenschenkelspalt sichtbar (GUTZSCHEBAUCH 1936). Üblicherweise befinden
sich die Hoden beim Hund nicht nebeneinander, sondern hintereinander
(GUTZSCHEBAUCH 1936) wobei sich der linke Hoden etwas weiter kaudal als der
rechte befindet (EVANS u. CHRISTENSEN 1993). Die Hoden werden auch als
samenbereitende Organe bezeichnet und damit funktionell von den Organen der
Spermienreifung und -speicherung unterschieden. Mit seiner ovalen Form besitzt der
Hoden eine Extremitas capitata (dem Nebenhodenkopf anliegend) sowie eine
Extremitas caudata (dem Nebenhodenschwanz anliegend). Der Bereich, der dem
Nebenhodenkörper anliegt wird als Margo epididymalis bezeichnet, die ihm
gegenüberliegende Seite als Margo liber. Die seitlichen Flächen des Hodens werden
als Facies lateralis und Facies medialis bezeichnet (GASSE 2004).
Die intraskrotale Lage der Hoden ist bei der überwiegenden Mehrzahl der Rüden
horizontal, wobei die Extremitas capitata leicht nach kranioventral geneigt und die
Extremitas caudata entsprechend nach kaudodorsal angehoben ist
(GUTZSCHEBAUCH 1936; GÜNZEL-APEL 1994).
Es besteht eine Korrelation zwischen der Hodengröße und dem Gewicht des Rüden
(GÜNZEL-APEL et al. 1994)
Die Nebenhoden werden unterteilt in Kopf, Körper und Schwanz (CORNWALL 2009).
Sie liegen den Hoden direkt an und sind am Margo epididymalis mit diesem
verwachsen (LIEBICH 2004). Der Nebenhodenkopf des Hundes besteht zum größten
Teil aus den 15-16 Ductuli efferentes (GASSE 2004), welche die Samenzellen aus
dem Rete testis in den Nebenhodenkopf befördern. Je nach Größe des Hundes sind
die Nebenhodenköpfe stecknadelkopf- bis erbsengroß. Der Nebenhodenkörper liegt

Literaturübersicht
12
dem Hoden strangförmig dorsolateral auf und mündet in den Nebenhodenschwanz.
Der den Nebenhodenkörper durchziehende Nebenhodenkanal liegt im Bereich des
Nebenhodenschwanzes in extrem stark aufgeknäuelter Form und dient als
Spermienreservoir, während Nebenhodenkopf und –körper primär der
Spermienreifung dienen (CORNWALL 2009). Die Größe des Nebenhodenschwanzes
variiert ebenfalls mit der Körpergröße des Hundes und ist linsen- bis haselnussgroß
(GÜNZEL-APEL et al. 1994). Die Form des Nebenhodenschwanzes kann kappen-,
halbkugel-, kugel- oder spitzkegelförmig sein. In der Regel ist der
Nebenhodenschwanz deutlich vom Hoden abgesetzt (GUTZSCHEBAUCH 1936).
Die Hoden sind von mehreren Häuten umgeben (GUTZSCHEBAUCH 1936). Ganz
außen befindet sich das Skrotum, welches anatomisch aus der Haut und der
Unterhaut (Tunica dartos), sowie der Fascia spermatica externa besteht. Die Tunica
dartos bildet ein kontraktiles Netz aus glatten Muskelzellen. Im Inneren wird das
Skrotum durch ein Septum unterteilt. In jedem dieser Hohlräume liegt ein Hoden,
umgeben vom Processus vaginalis. Dieser besteht aus der Fascia spermatica interna
und der Tunica vaginalis. Direkt dem Hoden anliegend hält die Tunica albuginea testis
durch ihre derbe Konsistenz den Hoden unter Druck. Die Tunica albuginea ist ca.
1 bis 2 mm dick, reich an Kollagenfasern und führt die Versorgungsgefäße in einer
Tunica vasculosa in sich. Außerhalb des Processus vaginalis liegt für jeden Hoden
separat der Musculus cremaster mit seiner Fascie. Außer dem Ligamentum scroti,
einem derben Bindegewebsband welches im distalen Bereich den Processus
vaginalis mit dem Skrotum verbindet, hat der Hoden keine feste Verbindung zum
Skrotum. Dieses wird lediglich durch lockeres Bindegewebe gefüllt. Die äußere Haut
des Skrotums ist dünn und wenig behaart, eine individuelle Pigmentierung wird
beobachtet. Auch sind zahlreiche Schweiß- und Talgdrüsen vorhanden
(GUTZSCHEBAUCH 1936; EVANS u. CHRISTENSEN 1993; GASSE 2004).

Literaturübersicht
13
2.1.2 Blutgefäßversorgung der Hoden
Die Blutgefäße im Hoden haben drei Hauptaufgaben, die Regulation der Temperatur
und des Blutdruckes und den Transport von Hormonen (SINOWATZ 2001). Der
Blutfluss zum Hoden erfolgt über die im Samenstrang verlaufende Arteria testicularis,
welche aus der Arteria spermatica interna entspringt (GUTZSCHEBAUCH 1936). Dort
ist sie, im Plexus pampiniformis, rankenförmig geschlängelt und tritt im weiteren
Verlauf an der Extremitas capitata in den Hoden ein. In diesem verläuft sie am Margo
epididymalis als so genannte Marginalarterie zur Extremitas caudata. An dieser
angekommen, teilt sie sich (als so genannte Zentripetalarterie) in einen medialen und
einen lateralen Ast auf, welche weitere Zweige in das zwischen den Lobuli des
Hodens befindliche Bindegewebe abgegeben. Im Mediastinum testis knäueln sich die
Arterien erneut auf und laufen als Zentrifugalarterien zurück in Richtung Hodenkapsel.
Der venöse Abfluss läuft zu einem geringen Teil über kleinere Venen, die dem Verlauf
der Ductuli efferentes folgen und schließlich die Extremitas capitata erreichen. Der
überwiegende Teil verläuft durch das Parenchym zur Hodenkapsel und bildet dort ein
Venennetz, welches an der Basis des Samenstrangs als Plexus pampiniformis vorliegt
(GUTZSCHEBAUCH 1936; SINOWATZ 2001). Durch diese speziell entwickelte Form
des Plexus wird eine große Austauschfläche zwischen Arterie und Vene gebildet.
Diese fördert den Wärmeaustausch und erleichtert die Diffusion (COULTER u.
KASTELIC 1994). Um die Fließgeschwindigkeit im Plexus herabzusetzen, sind
mehrere Anastomosen zwischen den einzelnen Venen ausgebildet (LIEBICH 2004).
2.1.3 Histologie des Hodens
Hoden Die dem Hoden direkt anliegende Tunica vasculosa liegt beim Hund und beim
Schafbock oberflächlich, bei Hengst und Eber in tieferen Schichten (SINOWATZ
2001; LIEBICH 2004). Zusätzlich enthält die Tunica albuginea, neben der Tunica
vasculosa, elastische und einzelne glatte Muskelzellen (MONTKOWSKI 1992;
LIEBICH 2004). Der Hoden wird komplett von ihr umschlossen und von aus ihr

Literaturübersicht
14
entlassenen Bindegewebstrabekeln durchzogen (MONTKOWSKI 1992; LIEBICH
2004). Durch diese Bindegewebssepten (Septula cortis) wird der Hoden in mehrere
Lobuli testis, mit den darin befindlichen stark geschlängelten Samenkanälchen (Tubuli
seminiferi convoluti oder contorti), unterteilt. Diese münden in einen gestreckten
Endabschnitt, die Tubuli seminiferi recti und vereinigen sich im Mediastinum zum Rete
testis (LIEBICH 2004). Das Mediastinum testis ist ein aus den Septula testis gebildeter
bindegewebiger Bereich, welcher beim Hund zentral im Hodenparenchym liegt und
den Hoden auf seiner gesamten Länge durchzieht (MONTKOWSKI 1992). Das Rete
testis liegt als Netzwerk im Mediastinum und ist von einer deutlichen Schicht
elastischer Fasern umgeben. In den Septula testis verlaufen größere Blut- und
Lymphgefäße sowie Nerven (MONTKOWSKI 1992). Am Hoden ist vor allem der
sympathische Anteil des vegetativen Nervensystems stark ausgebildet (SINOWATZ
2001). Die Tubuli seminiferi convoluti sind 50 bis 80 cm lang und im Durchschnitt 200
bis 300 μm breit (LIEBICH 2004). Sie bestehen aus Keim- und Stützzellen
(Sertolizellen). Beim Hund liegen die Sertolizellen direkt der Basalmembran der Tubuli
an, sowie vereinzelt zwischen den Spermatogonien (FORD 1969). Der Kern scheint
direkt an der Basalmembran zu kleben. Die Tubuli seminiferi convoluti werden außen
durch eine Basalmembran sowie durch lockeres Bindegewebe begrenzt, in welches
die hormonproduzierenden Leydigschen Zwischenzellen eingebettet sind (GASSE
2004). Die Leydigschen Zwischenzellen liegen sowohl solitär als auch in Gruppen im
Bereich des Rete testis oder an Lymphgefäßen (MONTKOWSKI 1992).
Die oben erwähnten Tubuli recti sind beim Hund, genau wie beim Bullen, noch mit
einem als Terminalsegment bezeichneten Abschnitt versehen. Hier ragen
Sertolizellgruppen keilförmig in das Lumen und behindern so ein Zurückfließen der
Samenzellen. Außerdem besitzen sie auch hier phagozytotische Fähigkeiten.
MONTKOWSKI (1992) unterteilt dieses Terminalsegment aufgrund der
Epithelbeschaffenheit in drei Abschnitte sowie eine endständige kelchartige
Erweiterung.
Das intertubuläre Segment besteht zum Einen aus den intertubulären Spalträumen
innerhalb eines Lobulus, welche beim Hund fast vollständig mit in Gruppen liegenden

Literaturübersicht
15
Leydigschen Zwischenzellen und von kleineren Blut- und Lymphgefäßen sowie von
elastischen Fasern ausgefüllt werden. Zum Anderen gibt es den Bereich, der aus der
Tunica albuginea einstrahlenden Bindegewebstrabekel. Hier befinden sich größere
Blut- und Lymphgefäße sowie Fibrozyten und mononukleäre Zellen und, wie in den
intertubulären Spalträumen, elastische Fasern und Leydigsche Zwischenzellen.
Nebenhoden Im Bereich des Nebenhodenkopfes befinden sich Kanälchen welche als Ductuli
efferentes testis, gesäumt mit glatten Muskelzellen zum Spermientransport, die Tunica
albuginea des Hodens durchbrechen.
Am Ende des Nebenhodenkopfes vereinigen sich diese, beim Hund nach LIEBICH
(2004) 15 bis 16 Kanälchen, zu dem Nebenhodenkanal, dem Ductus epididymidis
(GASSE 2004). Gesäumt ist dieser Gang von einem zweireihigen hochprismatischen
Epithel, welches ein ideales Milieu für die Spermienreifung schafft. Aus dem
Nebenhodenkanal entspringt der Samenleiter, Ductus deferens, in welchem die
Spermien während der Ejakulation durch die Kontraktion glatter Muskulatur zum
Beckenstück der Harnröhre befördert werden (GASSE 2004). Im Samenleiter befindet
sich zusätzlich ein mit Drüsengewebe ausgekleideter Abschnitt, die Pars glandularis
ductus deferentis.
REHM (2000) wertete HE-gefärbte präparierte Hoden von 50 einjährigen Beagle-
Rüden aus. Bei 30% der Rüden stellte er eine bilaterale, segmentale
Hypospermatogenese fest, welche sich in Form verkleinerter Keimzellen,
geschrumpfter Tubuli seminiferi contorti, Sertoli only, multinukleärer Riesenzellen und
geschwollener Spermatozyten manifestierte. Bei 20% der Beagle-Rüden im Alter von
6 bis 32 Monaten wiesen GOEDKEN et al. (2008) eine Hypospermie nach, davon bei
31,3% bilateral. Bei allen Rüden war ein multifokales Fehlen elongierter
Spermatozyten zu verzeichnen. Häufig traten Zelldebris, Riesenzellen und
geschwollene Spermatozyten auf. 75% der betroffenen Rüden waren sechs bis acht
Monate alt und demzufolge noch nicht geschlechtsreif. Über elf Monate alte Rüden
waren weniger als 10% betroffen. Bei 10% der Rüden wurden apoptotische Zellen in

Literaturübersicht
16
minimalem Umfang gefunden. Die multinukleären Riesenzellen waren bei 73,8% der
Rüden zu finden, davon 85% beidseits mit ein bis sechs Zellen pro Tubulus und
weniger als zehn Tubuli pro Hoden. Im Nebenhoden wurden nur eine multinukleäre
Riesenzelle und einige unreife Spermien gefunden.
2.2 Spermatogenese, Spermiogenese und Keimepithelzyklus
Im Rahmen der Spermatogenese entstehen aus den Spermatogonien haploide
Spermatiden, welche dann während der Spermiogenese zu morphologisch reifen
Spermien ausdifferenzieren. Dieser Prozess läuft zyklisch in den Tubuli seminiferi ab
(FORD 1969; SCHNORR u. KRESSIN 2001). Zunächst kommt es durch mitotische
Teilungen zur Vermehrung der Spermatogonien, gefolgt von einer Wachstumsphase,
die zur Entstehung von Spermatozyten 1. Ordnung führt. Durch Reduktion der
Chromosomenzahl entstehen Spermatozyten 2. Ordnung, welche ziemlich schnell
durch meiotische Teilung zu Spermatiden werden. Die haploiden Spermatiden gehen
nun zur Spermiogenese über, die in vier Schritten abläuft (FORD 1969;
MONTWKOWSKI 1992). In der Golgi-Phase wandern abgeschnürte Vesikel zum Kern
und bilden so eine akrosomale Vakuole. In der darauf folgenden Kappen-Phase
verdichtet sich die Vakuole zu einem Granulom, welches sich halbkugelförmig
ausbreitet und im weiteren Verlauf die Kopfkappe bildet. In der sich anschließenden
Akrosom-Phase kondensiert der Kern und nimmt eine längliche Gestalt an. Zusätzlich
verlagert sich das Zytoplasma hinter den Zellkern. Nachfolgend findet eine
Reifungsphase statt.
FOOTE und SWIERSTRA (1972) teilen den Keimepithelzyklus des Hundes in acht
Stadien ein, beginnend mit dem Zeitpunkt, an dem alle reifen Stadien in das Lumen
entlassen werden. Als Kriterien gelten sowohl die Kernform, als auch die Lage zur
Basalmembran. Insgesamt unterteilen sie A- und B-Spermatogonien, welche aber
nicht weiter klassifiziert werden. Die A-Spermatogonien zeigen sich in den Stadien
zwei bis fünf, in deren Verlauf sie zahlenmäßig zunehmen, die B-Spermatogonien in
den Stadien sechs bis acht. Anschließend folgt die Wandlung zu präleptotenen

Literaturübersicht
17
Spermatozyten 1. Ordnung, welche nach den vier Meiosestadien die zweite
Reifeteilung durchlaufen. In den ersten vier Stadien existieren zwei Zellgenerationen.
Die Dauer eines Keimepithelzyklus wird mit 13,6 Tagen angegeben. Insgesamt dauert
die Spermatogenese beim Hund acht bis neun Wochen und läuft dementsprechend in
vier bis fünf Zyklen ab.
Ebenfalls in acht Stadien, basierend auf dem Akrosomsystem, wird der
Keimepithelzyklus des Hundes von SOARES et al. (2009) eingeteilt. In dieser Studie
wurden zusätzlich Unterschiede zwischen einzelnen Rassen und Mischlingen
beleuchtet. Des Weiteren untersuchten sie anhand von intratestikulären Thymidin-
Injektionen die Dauer des Keimepithelzyklus. Die gefundenen Ergebnisse decken sich
mit denen von FOOTE und SWIERSTRA (1972). Die durchschnittliche Dauer eines
Zyklus wurde mit 13,7 Tagen angegeben. Einzig beim American Pitbull zeigte sich
eine durchschnittliche Dauer von 12,6 Tagen. Zwischen den anderen untersuchten
Rassen (Pinscher, Beagle, Pudel, Labrador und Mischlinge) wurden keine
Unterschiede entdeckt.
Dagegen teilen IBACH et al. (1976) den Keimepithelzyklus des Hundes in zehn
Stadien ein. Sie beginnen mit der Verlagerung der Kopfkappe in Richtung
Basalmembran und unterteilen die Zellgruppen in A- (mehrere Generationen), I- und
B-Spermatogonien sowie in primäre und sekundäre Spermatozyten. Im Anschluss
daran entstehen die Spermatiden, welche einen runden oder gestreckten Kern
aufweisen. Die Entwicklungsrichtung läuft von der Basalmembran zum Lumen hin ab
(Ford 1969).
2.3 Apoptose
2.3.1 Allgemein
Unter Apoptose ist der aktive, programmierte Tod einer einzelnen Zelle zu verstehen
(KATAOKA u. TSURUO 1996). Sie funktioniert als Gegenspieler zur Mitose und ist für
die Aufrechterhaltung der Homöostase im Körper unerlässlich (ELMORE 2007).

Literaturübersicht
18
Entscheidend ist immer, dass der apoptotische Prozess unter genetischer Kontrolle
steht (MAJNO u. JORIS 1995).
Es existieren zwei mögliche Ursachen, die einen apoptotischen Prozess auslösen
können. Zum einen der genetisch programmierte Zelltod, welcher bei der embryonalen
Entwicklung greift und zum anderen eine durch äußere Reize (z.B. Strahlung,
Medikamente und Wärme) ausgelöste Apoptose (MAJNO u. JORIS 1995).
Initiiert werden kann die Apoptose über extrinsische und intrinsische Stimuli (NAGATA
1997). Eine wichtige Rolle spielen hierbei Noxen wie radioaktive Strahlung oder
Medikamente, die die DNA schädigen (KATAOKA u. TSURUO 1996).
Der extrinsische Weg läuft über eine Bindung von TNF und Fas-Ligand, beide
gehören zu den Zytokinen, an ihre spezifischen Rezeptoren (NAGATA 1997). Hierbei
kommt es zur Aktivierung über die Procaspase-8 zur Caspase-8 (SCHULTZ u.
HARRINGTON 2003).
Bei dem intrinsischen Weg steigt die Permeabilität der Mitochondrienmembran infolge
dessen das Membranpotential abnimmt. Dies hat zur Folge, dass es zum Austritt
verschiedener Proteine kommt, wie z.B. Cytochrom C, welches die Procaspase-9
aktiviert (NAGATA 1997; SCHULTZ u. HARRINGTON 2003; SINHA HIKIM et al.
2003). Der intrinsische Weg wird zum einen durch positive Faktoren (Strahlung,
Toxine, freie Radikale, Hitze), zum Anderen durch negative Faktoren, die über das
eigene Fehlen apoptotische Prozesse starten können (Hormone, Wachstumsfaktoren,
Zytokine), ausgelöst (ELMORE 2007).
Beide Wege führen zur Aktivierung der Effektorcaspasen, allen voran der Caspase-3,
welche von allen Initiatorcaspasen aktiviert werden kann. Alle Effektorcaspasen
aktivieren Endonukleasen, welche dann DNA-Brüche hervorrufen. Caspase-3 speziell
bewirkt die Spaltung des ICAD (Inhibitor of Caspase Activating DNA-ase), eines
Proteins, welches die DNA schützt. Zusätzlich wird eine DNA-Helicase gespalten,
welche die Chromatinkondensation bewirkt. Ferner werden von den Caspasen
Cytoskelettbestandteile gespalten, so dass die für die Apoptose typische
Zellschrumpfung stattfinden kann (FISCHER et al. 2003).

Literaturübersicht
19
Der dritte, auch von ELMORE (2007) erwähnte, Weg entspricht der von
MARTINVALET et al. (2005) beschriebenen Caspase unabhängigen Apoptose. Dabei
spielt das Granzym A eine entscheidende Rolle. Es bewirkt einen intrazellulären
Anstieg reaktiver Sauerstoffverbindungen sowie ein Absinken des Membranpotentials
der Mitochrondrien. Eingebracht in die Zelle wird das Granzym A über zytotoxische T-
Zellen und natürliche Killerzellen. Im weiteren Verlauf zeigt die betroffene Zelle viele
Merkmale der Apoptose, lediglich die äußere Mitochondrienmembran bleibt intakt, so
dass kein Cytochrom C austreten kann, und somit keine Caspasen aktiviert werden.
Dagegen kommt es zu einer Spaltung des sogenannten Set-Komplex, einem aus
mehreren Proteinen bestehenden Komplex, dessen Aufgabe im Schutz der DNA
besteht. Wird dieser Komplex gespalten, kommt es zu einer Aktivierung von DNAsen
die die DNA spalten. Im Unterschied zu den intrinsischen und extrinsischen Wegen
wird in diesem Fall die DNA einzelsträngig gespalten (MARTINVALET et al. 2005).
Einen Caspase-3 unabhängigen Weg des Zelltodes zeigen JÄHNICKE et al. (1998)
auf. Hierbei wurden MCF-Brustkrebszellen, welche aufgrund eines Gendefektes keine
Caspase-3 beinhalten, mit Tumor Nekrose Faktor behandelt. Auch diese Zellen
zeigten eine apoptotische Reaktion, allerdings ohne DNA-Fragmentation,
Zellschrumpfung oder Ausbildung des typischen „blebbings“ (Vorgang der
Ausstülpung von Plasma- und Kernmembran, welche weiter zu Vesikeln abgeschnürt
werden).
Zu Beginn des apoptotischen Prozesses kommt es zu einer Verringerung des
Zellvolumens mit anschließender Kondensation des Zellkerns (MAJNO u. JORIS
1995; KATAOKA u. TSURUO 1996). Letztere erfolgt als halbmondförmige
Ablagerung, der Margination des Chromatins (MAJNO u. JORIS 1995). Andere
Zellorganellen sind von diesem Prozess nicht betroffen (KATAOKA u. TSURUO
1996). Nach MAJNO und JORIS (1995) kann jedoch eine geringe Schwellung der
Mitochondrien auftreten, welche jedoch für den Ablauf der Apoptose nicht notwendig
ist. Nachdem die Zellmembran Blasen geschlagen hat („blebbing“), zerbricht der
Zellkern und es entsteht der apoptotische Restkörper (MAJNO u. JORIS 1995;
KATAOKA u. TSURUO 1996). Dieser wird komplett von neutrophilen Granulozyten

Literaturübersicht
20
phagozytiert (KATAOKA u. TSURUO 1996). Dagegen schreiben MAJNO und JORIS
(1995) sowie JACOBSON et al. (1997) die Phagozytose den Makrophagen und
benachbarten Zellen zu. Andererseits kann der apoptotische Restkörper auch als
freier apoptotischer Körper bestehen bleiben (MAJNO u. JORIS 1995). Eine
Entzündungsreaktion bleibt aus, da apoptotische Zellen eine so geringe Menge an
Chemo-Attraktanzien produzieren, dass keine Leukozyten angelockt werden.
Apoptose kann innerhalb von Minuten ablaufen (MAJNO u. JORIS 1995). In der
Agonie der Apoptose entlassen die Zellen viele Pseudopodien, es kommt zum
„budding“ (KERR u. HARMON 1991). Bei der Nekrose zeigen sich dagegen auch
Veränderungen am Zytoplasma in Form von Eosinophilie, Zusammenbruch der
Zellstrukturen und Fragmentation (MAJNO u. JORIS 1995).
2.3.2 Nachweisverfahren der Apoptose
Die üblichsten Verfahren für den Nachweis apoptotischer Zellen ist das TUNEL-
Verfahren (TdT-mediated dUTP-biotin nick end labeling) und der Nachweis aktivierter
Caspase-3.
Bei dem TUNEL-Verfahren wird die Tatsache genutzt, dass es während der Apoptose
durch Endonukleasen zu einer DNA-Fragmentation kommt. Die frei werdenden Enden
werden im Laufe des Verfahrens durch das Enzym TdT (terminal deoxynucleotidyl
transferase) mit markierten Nukleotiden versehen, welche lichtmikroskopisch sichtbar
gemacht werden können (GAVRIELI et al. 1992). Apoptotische Zellen sind nur über
eine kurze Zeitspanne vorhanden. Von der Initiierung bis zur Elimination dauert es
maximal drei Stunden (GAVRIELI et al. 1992). Dabei ist nicht sicher, wie viele
Strangbrüche vorhanden sein müssen, um ein positives Ergebnis zu erzielen
(ELMORE 2007). Auch Zellen, welche nekrotisch sind, können sich gerade in der
DNA-Reparationsphase oder der Transkriptionsphase befinden und aufgrund dessen
ebenfalls positiv dargestellt werden.
Bei dem Caspase-3-Verfahren wird in einem immunhistochemischen Verfahren die
aktivierte Form der Caspase-3 im Gewebe nachgewiesen. Mittels eines spezifischen
Caspase-Markers können zytoplasmaständige Enzyme markiert und sichtbar gemacht

Literaturübersicht
21
werden. Allerdings ist eine Aktivierung von Caspasen nicht unbedingt mit Apoptose
gleichzusetzen. Hinzu kommt, dass die Caspasen oft nur in einem geringen
Zeitfenster darstellbar sind (ELMORE 2007).
Weitere mögliche Verfahren zur Erfassung apoptotischer Zellen stellen die
Elektronenmikroskopie und PCR-Assays dar.
2.3.3 Apoptose am Hoden
Am Hoden können folgende Faktoren Apoptose auslösen: Entzug oder Zugabe von
Wachstumsfaktoren oder Hormonen, eine veränderte Interaktion benachbarter Zellen
sowie DNA -Schädigung (zum Beispiel durch Strahlung) (DANSRANJAVIN 2000).
Beim Menschen zeigen ca. 8% der ausgewerteten Hodentubuli mit normaler
Spermatogenese mindestens eine apoptotische Zelle. Am häufigsten sind
Spermatozyten (63%) betroffen, doch können auch alle anderen
Spermatogenesestadien apoptotische Prozesse aufweisen. Im Keimepithelzyklus
dient die Apoptose dem Schutz vor geschädigten Zellen sowie fehlerhaften Mitose-
und Meiose-Produkten. Des weiteren kommt der Apoptose eine Bedeutung in der
Regulation der Zellhomöostase zu, da die Sertolizellen nur eine bestimmte Anzahl
Keimzellen versorgen können (DANSRANJAVIN 2000). 75% der Keimzellen sterben
während ihrer Entwicklung, vor allem in den frühen Stadien der Spermatogenese ab.
Bei Mäusen kam es nach vollständigem Testosteronentzug zu einer gesteigerten
Apoptoserate im Hoden (TROJANO et al. 1994). BILLIG et al. (1995) erzielten diesen
Effekt am Rattenhoden durch Entzug von Gonadotropinen. Betroffen waren jedoch
nur Tiere im Alter von 16 bis 32 Tagen. Bei erwachsenen Tieren zeigte sich,
unabhängig vom Gonadotropin-Entzug eine vermehrte DNA-Fragmentation in den
Stadien XII bis XIV des Keimepithelzyklus, während das Stadium VIII kaum betroffen
war. SINHA HIKIM et al. (2003) zeigten bei Ratten mit Hilfe eines
Testosteronimplantats eine Steigerung der Apoptose in den Stadien VII bis VIII,
welche in der Studie von BILLIG et al. (1995) eher die weniger betroffenen Stadien
darstellten.

Literaturübersicht
22
Nach 15-minütiger Erwärmung auf 42°C zeigten die Hoden adulter Ratten eine
erhöhte Apoptoserate. Diese trat in den Tagen 1 und 2 nach Wärmeapplikation vor
allem in den Stadien I bis IV und XII bis XIV auf. In den Stadien V bis VIII waren
dagegen nur geringe Veränderungen nachzuweisen. Am empfindlichsten zeigten sich
pachytäne Spermatozyten, frühe Spermatiden, diplotäne und sich teilende
Spermatozyten. Neun Tage nach der Wärmeapplikation waren schwere Schäden an
den Tubuli seminiferi contorti zu finden. Apoptotische Zellen wurden nur noch
vereinzelt nachgewiesen. 56 Tage nach der Wärmeapplikation hatte eine komplette
Regeneration stattgefunden (SINHA HIKIM et al. 2003). Durch Kombination des
Testosteronimplantats mit der kurzzeitigen Hitzeeinwirkung konnte nahezu eine
Azoospermie erzeugt werden. SINHA HIKIM et al. (2003) verdeutlichten durch ihre
Studie außerdem die Beteiligung von Mitochondrien durch den Nachweis von
ausgetretenem Cytochrom C und Caspasen. Zusätzlich zeigten sie anhand von
Knock-out Mäusen, dass der Weg der Apoptose über den FAS/FAS-Ligand Komplex
bei der hitzeinduzierten Apoptose am Hoden eine eher untergeordnete Rolle spielt.
Bei Lamas wurden nach einer vierwöchigen Erhöhung der Umgebungstemperatur auf
29°C 22 bis 30 TUNEL-positive Zellen pro 100 Tubuli festgestellt. Betroffen waren hier
vor allem primäre Spermatozyten. Diese Veränderungen waren im Vergleich zur
Kontrollgruppe allerdings nicht signifikant (SCHWALM et al. 2007).
KAWAKAMI et al. (2000) bestimmten die Anzahl apoptotischer Zellen bei vier Rüden
mit Azoospermie und bei drei Hunden mit normaler Ejakulatbeschaffenheit.
Durchschnittlich 3% der runden Spermatiden stellten sich mittels TUNEL-Verfahren
positiv dar. Drei Hunde mit Azoospermie wiesen in 3% der Leydigschen
Zwischenzellen DNA-Fragmentation auf. Sertolizellen konnten zu keinem Zeitpunkt
TUNEL-positiv dargestellt werden. In den Hoden der Hunde mit normaler
Ejakulatbeschaffenheit wurde keine Keimzellapoptose nachgewiesen.

Literaturübersicht
23
2.4 Nekrose
Im Gegensatz zur Apoptose ist die Nekrose kein genetisch kontrollierter sondern ein
energieunabhängiger Prozess, an dem die Zelle passiv beteiligt ist (ELMORE 2007).
Gekennzeichnet ist die Nekrose durch irreversible Veränderungen im Zellkern wie
Karyolysis, Pyknose und Karyorrhexis sowie durch Veränderungen im Zytoplasma in
Form von Kondensation, Eosinophilie und Fragmentation (MAJNO u. JORIS 1995).
Durch Störung der Ionenkanäle kommt es bei nekrotischen Prozessen zur
Verschiebung des osmotischen Gradienten und in der Folge dessen zu einem
Wassereinstrom in die Zelle. Die Zelle schwillt an und rupturiert. Dadurch werden
intrazelluläre Komponenten in die Umgebung freigesetzt und locken Immunzellen an,
die wiederum durch Zytokinproduktion einen entzündlichen Prozess bewirken
(SCHEIDEL 2000). Weiter treten vermehrt Zytoplasmavakuolen, zytoplasmatische
Bläschen sowie kondensierte, rupturierte oder geschwollene Mitochondrien auf
(ELMORE 2007). Eine Nekrose kann ebenfalls durch verschiedene intrinsische und
extrinsische Faktoren ausgelöst werden. Dauer und Stärke eines Stimulus sind
entscheidend dafür, ob die Zelle durch Nekrose oder Apoptose zu Grunde geht.
2.5 Degenerative Prozesse an den Hoden
Erkrankungen der Hoden, die mit Degenerationserscheinungen und verminderter
Ejakulatbeschaffenheit einhergehen können, haben verschiedenste Ursachen. In der
Bauchhöhle verbliebene Hoden sind gewöhnlich kleiner als skrotale Hoden und
weisen keine Spermatogenese auf. Solche Hoden besitzen eine erhöhte Neigung zur
tumorösen Entartung (WEISS u. KÄUFER-WEISS 2007). Akute degenerative und
atrophische Zustände sind häufig nur histologisch an hydropischer Degeneration,
Kernpyknosen, mehrkernigen Riesenzellen und Störungen der Samenzellbildung zu
erkennen. Als Ursachen kommen hier vor allem Traumata, akute oder chronische
entzündliche Prozesse, Infektionskrankheiten, klimatische Veränderungen und
Ernährungsfehler zum Tragen. Eine senile Hodenatrophie, also eine

Literaturübersicht
24
Altersdegeneration, geht in der Regel mit einer Lipofuszinablagerung einher, welche
den Hoden braun verfärbt.
2.6 Proliferation am Hoden
2.6.1 Allgemein
Aufgrund der verschiedenen gleichzeitig in den Tubuli ablaufenden
Spermatogenesestadien ist das Hodengewebe von ständigen Proliferationsprozessen
gekennzeichnet. Hierbei proliferieren in erster Linie die Spermatogonien welche sich
durch mitotische Teilung vermehren. In den anderen Stadien der Spermatogenese
erfolgen lediglich Umbauvorgänge bzw. meiotische Teilungen (MONTKOWSKI 1992).
Beim Bullen ist die Proliferation am stärksten in den Stadien fünf bis acht ausgeprägt,
wenn die B- Spermatogonien sich teilen. Sertolizellen können hingegen in keinem
Bereich mithilfe eines Proliferationsmarkers dargestellt werden (WROBEL et al. 1993).
Bei Lamas waren unter Kontrollbedingungen in 20,5% der Tubulusanschnitte keine
einzige positive Zelle zu finden. Durch vierwöchige Wärmeapplikation stieg der
Prozentsatz unmittelbar auf 54,3% an, um im Verlauf der darauf folgenden sechs
Wochen auf 11% abzufallen.
2.6.2 Nachweisverfahren
Proliferationsprozesse lassen sich mithilfe des Markers Ki-67 nachweisen
(SCHOLZEN u. GERDES 2000). Während der Ruhephase des Zellzyklus befindet
sich das Ki-67 Protein ausschließlich im Zellkern, weshalb es in diesem Stadium nicht
nachgewiesen werden kann.
Das Ki-67 Antigen eignet sich besonders gut zur Bestimmung der Proliferation, da es
ein größeres Zeitfenster bietet als andere immunhistochemische Verfahren wie z.B.
der Nachweis des PCNA (Proliferating cell nuclear antigen) (WROBEL et al. 1993).

Literaturübersicht
25
2.7 Sonographie der Hoden
2.7.1 Technische Grundlagen
Bei der Ultraschalltechnik werden durch piezoelektrische Kristalle im Schallkopf
impulsartige Schallwellen (durch Ionenverschiebung im Kristall) erzeugt. Diese
werden nach Reflexion des betroffenen Gewebes vom Schallkopf wieder empfangen.
Ultraschallwellen liegen im Bereich von 20 kHz bis zehn MHz und sind für den
Menschen nicht hörbar (POULSEN NAUTRUP 2007). Sie werden im Ultraschallgerät
in Spannungssignale umgewandelt und als zweidimensionales Bild dargestellt (GIESE
1997). Es können nur senkrecht getroffene Grenzflächen sicher beurteilt werden
(POULSEN NAUTRUP 2007). Zwischen Schallkopf und Körperoberfläche befindliche
Luft verhindert eine Ultraschalldarstellung oder führt zu Artefakten. Aus diesem
Grunde sind sowohl die Entfernung des Haarkleides und als auch die Verwendung
eines Ultraschallgels unverzichtbar.
2.7.2 Dopplersonographie
Der so genannte Dopplereffekt ist ein Effekt, welcher die Änderung einer Frequenz
beschreibt, wenn sich Sender und Empfänger relativ aufeinander zu bewegen (GIESE
1997). Wichtig ist hierbei die Geschwindigkeit der Wellen (c) und die
Relativgeschwindigkeit (v) von Sender und Empfänger. Mithilfe dieses Effektes
können Blutströmungsgeschwindigkeit und Blutflussrate in peripheren Arterien
bestimmt werden. Reflektiert werden die eintretenden Ultraschallwellen hierbei von
den sich bewegenden Erythrozyten (GIESE 1997). Fließt das Blut auf den Schallkopf
zu, erscheinen die Wellen gestaucht (f1 > f0 und fd > f0). Definitionsgemäß wird
dieses Blut oberhalb der Nulllinie und rot dargestellt. Bei vom Ultraschallkopf weg
fließendem Blut werden die Schallwellen gedehnt (f1 < f0 und fd < f0). Diese
Blutflüsse werden unterhalb der Nulllinie und blau dargestellt (POULSEN NAUTRUP
2007). Die Kombination aus Dopplerverfahren und der zweidimensionalen

Literaturübersicht
26
Sonographie wird auch als Duplexsonographie bezeichnet. Sie ermöglicht die exakte
Zuordnung des Blutflusses zu bestimmten Strukturen.
Beim farbcodierten Dopplerverfahren werden die Messvolumina eher flächenhaft
abgebildet. Dieses Verfahren ist schnell und bietet als Einziges eine wirkliche
Kombination aus zweidimensionalem Ultraschallbild und Dopplerfunktion (POULSEN
NAUTRUP 2007).
Unterschieden wird hierbei zwischen dem kontinuierlichem Dopplerverfahren (CW) mit
zwei Piezokristallen (einen zum Senden, einen zum Empfangen), und dem gepulsten
Dopplerverfahren (PW) mit nur einem Kristall. Bei der gepulsten Dopplersonographie
besteht die Möglichkeit einer Winkelkorrektur, so dass auch in nicht exakt parallel
verlaufenden Gefäßen der Blutfluss gemessen werden kann (POULSEN NAUTRUP
2007). Bei der gepulsten Duplexsonographie wird zuerst die richtige Platzierung im
zweidimensionalen Bild geprüft und anschließend auf den Dopplerbetrieb
umgeschaltet.
Fehlerquellen ergeben sich bei der gepulsten Dopplersonographie sowie der
farbkodierten Dopplersonographie aus dem Aliasing-Phänomen und der Nyquist-
Frequenz (maximal eindeutig messbare Doppler-Shift).
Das Aliasing-Phänomen zeigt die Grenzen einer gepulsten Messung auf. Eine hohe
Beobachtungsfrequenz erlaubt auch eine präzise Angabe der Geschwindigkeit. Dazu
muss der Dopplershift etwas kleiner als die maximale Pulsrepetitionsrate gewählt
werden (POULSEN NAUTRUP 2007). Wird eine Blutströmung oberhalb der Nyquist-
Frequenz gemessen kommt es zum Aliasing-Phänomen und der Blutfluss wird,
obwohl er auf den Schallkopf zufließt, unterhalb der Nulllinien angezeigt.
Um diesem Phänomen vorzubeugen, können die Eindringtiefe oder die
Dopplerfrequenz verringert werden. Dabei sind eventuelle Einbußen der Abbildungen
zu beachten. Durch Verschiebung der Nulllinie kann ein Messbereich zu Lasten des
anderen verdoppelt werden. Darüber hinaus besteht die Möglichkeit der Erhöhung der
Pulsrepetitionsrate mittels HPRF (high-puls-repetition-frequency) Dopplerverfahren.
Hierbei werden weitere Messvolumina eingerichtet, was allerdings wiederum zu einer
gewissen Ungenauigkeit führen kann (NEUERBURG-HEUSLER u. HENNERICI
1999).

Literaturübersicht
27
2.7.3 Auswertung von Dopplerkurven
Dopplerultraschallkurven können sowohl qualitativ als auch quantitativ ausgewertet
werden. Die qualitative Auswertung erfolgt über die Parameter Flussmuster,
Fließrichtung, Art der Strömung (laminar oder turbulent) sowie die
Geschwindigkeitsverteilung im Gefäß.
Zur quantitativen Auswertung werden verschiedene Geschwindigkeiten und Indizes
berechnet (TAYLOR u. HOLLAND 1990). Für die Bestimmung arterieller Blutflüsse
werden die systolische Maximalgeschwindigkeit (systolic peak velocity = SPV), die
diastolische Geschwindigkeit (diastolic peak velocity = DPV), die enddiastolische
Geschwindigkeit (enddiastolic peak velocity = EDV) und die amplitudengewichtete
mittlere Maximalgeschwindigkeit (time averaged maximum velocity = TAMAX)
gemessen. Als Indizes stehen der Pulsatilitätsindex (PI) und der Widerstandsindex
(RI) im Vordergrund.
Der PI ist unabhängig von dem Winkel der Arterie und von der Geschwindigkeit der
Trägerflüssigkeit (GOSLING u. KING 1974). Er wurde als Indikator für proximale
Stenosen etabliert.
Der Widerstandsindex (RI) (POURCELOT 1974) wurde zur Charakterisierung der
Arteria carotis communis etabliert. Er gibt eine Aussage über den Gefäßwiderstand
distal der Messstelle.
Ein weiterer Index, der A/B-Quotient (STUART et al. 1980) eignet sich besonders für
Arterien mit niedrigem peripheren Widerstand.

Literaturübersicht
28
Bei Arterien mit hohem peripheren Widerstand wird häufig der S/D-Quotient nach
DEEG u. WILD (1990) verwendet:
BIAGIOTTI et al. (2002) konnten bei Männern durch Messung von RI und SPV
zwischen obstruktiver und nicht obstruktiver Dysspermie unterscheiden. Hierbei
zeigten Männer mit nichtobstruktiver Dyspermie signifikant niedrigere Werte.
Bei Männern mit pathologischem Spermabefund fanden PINGGERA et al. (2008)
einen signifikanten Anstieg des RI im Vergleich zu einer gleichaltrigen Kontrollgruppe
mit Normospermie. Bei Vorliegen von Variozelen (Krampfadern im Venengeflecht des
Hodens) waren die Werte am höchsten. Dieses deckt sich mit den Untersuchungen
von BIAGIOTTI et al. (2002). Auch hier wiesen die Männer mit Variozelen die
höchsten Werte auf.
2.7.4 Zweidimensionale Sonographie der Hoden
Zur Sonographie der Hoden eignet sich besonders ein hochauflösender
Linearschallkopf (SEYREK-INTAS et al. 2010). Empfohlen wird die Verwendung einer
Vorlaufstrecke, z. B. der kontralaterale Hoden (LÜERSSEN u. JANTHUR 2007), sowie
das direkte Aufsetzen des Schallkopfes auf den zu untersuchenden Hoden (SEYREK-
INTAS et al. 2010). Die Sonographie der Hoden kann sowohl am stehenden, wie auch
am liegenden Tier durchgeführt werden (LÜERSSEN u. JANTHUR 2007). Der
physiologische Hoden ist gut umschrieben, oval, mit einer glatten Oberfläche. Eine
definierte Kapsel ist als dünne echoreiche Linie darstellbar (PUGH et al. 1990). Die
Echogenität sollte homogen, feinkörnig sein (BARR 1992; LÜERSSEN u. JANTHUR
2007; SEYREK-INTAS et al. 2010). Andere Quellen sprechen von grob-körnig (PUGH
et al. 1990). Im Zentrum des Hodens zeichnet sich eine echoreiche Linie, das
Mediastinum testis, ab (PUGH et al. 1990; BARR 1992; LÜERSSEN u. JANTHUR
2007). Der Nebenhoden stellt sich beim Hund etwas gröber als der Hoden dar und

Literaturübersicht
29
auch die Oberfläche kann etwas unregelmäßig erscheinen (BARR 1992; GÜNZEL-
APEL et al. 2001).
2.7.5 Dopplersonographie der Hoden
Am Hoden des Hundes zeigten sich im physiologischen Zustand ein monophasisches
Flussmuster sowie ein hoher diastolischer Blutfluss. Tumorös veränderte Hoden
weisen eine signifikante Zunahme der systolischen Maximalgeschwindigkeit und der
amplitudengewichteten mittleren Geschwindigkeit auf. Am Nebenhoden konnte kein
Blutfluss festgestellt werden (GÜNZEL-APEL et al. 2001). Eine Studie an 42 klinisch
geschlechtsgesunden Beagle-Rüden ergab in der Marginalarterie einen
durchschnittlichen PI von 0,78 und RI von 0,49 (GUMBSCH et al. 2002).
2.7.6 Zweidimensionale Sonographie der Prostata
Die Untersuchung der Prostata wird in Rückenlage oder seitlicher Lage durchgeführt.
Bei kurzen Untersuchungen ist auch eine Untersuchung am stehenden Tier möglich.
Der üblicherweise gewählte Konvexschallkopf (5 MHz) wird hierbei parapräputial,
leicht zum Becken hin gekippt, aufgesetzt (PRÜFER et al. 2007, SEYREK-INTAS et
al. 2010). Die Prostata befindet sich kaudal der Blase. Das Prostataparenchym zeigt
sich mittelechogen und ist beim jüngeren Hund fein gekörnt. Ältere Hunde zeigen eine
gröbere Körnung. Im Längsschnitt stellt sich die Prostata oval dar, im Querschnitt
zeigt sie aufgrund der Lappung eine Schmetterlingsform (GÜNZEL-APEL et al. 2001;
PRÜFER et al. 2007; SEYREK-INTAS 2010). Die Prostata sollte sich symmetrisch mit
weichen Grenzlinien darstellen (JOHNSTON et al. 1991). Die Länge der Prostata wird
definiert nach RUEL et al. (1998) als die längste die beiden Lappen teilende Strecke,
die Höhe als die maximale Strecke im 90° Winkel dazu. Eine Studie an 100 intakten
erwachsenen Hunden (die im Rahmen einer Impfung vorgestellt wurden) ergab eine
durchschnittliche Länge von 3,3 cm (± 0,9) sowie eine durchschnittliche Breite von
2,6 cm (± 0,7) (RUEL et al. 1998).

Literaturübersicht
30
2.8 Wärmeregulation am Hoden
Die Innentemperatur des Skrotums liegt bei Säugetieren 2 bis 7°C unter der
Körperkerntemperatur.
Bei adulten Ratten zeigte sich ein konstanter Temperaturgradient zwischen Hoden
und Rektum von 3 bis 5°C. Dieser wurde bei einem Anstieg der Rektaltemperatur
größer, da die Temperatur im Hoden sich nicht verändert (KORMANO 1967). Die
Adaptation männlicher Ratten an eine Umgebungstemperatur von 35°C führte zu
einer Herabsetzung der Befruchtungsraten, einhergehend mit Degeneration der
Hoden. Der Gradient zwischen Rektal- und Hodentemperatur blieb dabei aber
erhalten (SOD-MORIAH et al. 1974). Entscheidend ist also das Gleichgewicht
zwischen der über die Arteria testicularis eingebrachten Wärme und der durch
Stoffwechselvorgänge entstehenden Wärme sowie der über das Skrotum
abgegebenen Wärme.
An der Aufrechterhaltung der Skrotaltemperatur sind verschiedene Mechanismen
beteiligt (SETCHELL 1978a/b). Der Hauptmechanismus, die vom Körper produzierte
Wärme zu reduzieren, liegt im Gegenstromprinzip. Das Gegenstromprinzip funktioniert
aufgrund von Temperaturdifferenzen. Diese resultieren daraus, dass das venöse Blut
kälter ist als das arterielle Blut. Bestandteile des Gegenstromprinzips sind die stark
geknäuelte Arteria testicularis sowie der darum angeordnete Venenplexus, Plexus
pampiniformis, die zusammen als testicular vascular cone bezeichnet werden
(COULTER u. KASTELIC 1994). Da Arterie und Vene an vielen Stellen nur durch die
Gefäßwand begrenzt sind, kann ein effektiver Wärmeaustausch stattfinden. Ein
weiterer, wenn auch geringerer Anteil der Wärme wird über oberflächlich verlaufende
Arterien abgegeben (SETCHELL 1978a).
Die testikulären Blutgefäße sind sympathisch innerviert. Ein vermehrter Blutfluss
bewirkt immer auch, durch größere Abstrahlung der Wärme über oberflächliche
Arterien sowie eine verstärkte Evaporation, eine Kühlung des Hodengewebes
(SETCHELL 1978b). Zusätzlich kommt es durch den erhöhten Blutfluss auch zu

Literaturübersicht
31
einem stärkeren Verlust von Flüssigkeit, sowie vermehrter Schweißbildung am
Skrotum.
Durch Kontraktionen der Muskelanteile der Tunica dartos kann das Skrotum in Falten
gelegt und die Hoden hochgezogen und näher an die Bauchhöhle gebracht werden.
Neben dem Musculus cremaster, der als quergestreifter Muskel zu keiner lang
anhaltenden Kontraktion befähigt ist, sind die in die Skrotalhaut integrierten glatten
Muskelzellen beteiligt, welche über Katecholamine stimuliert werden (SETCHELL
1978a).
2.9 Einfluss einer Hyperthermie auf das Hodengewebe
Der Einfluss einer Hyperthermie auf den Hoden wurde bereits bei verschiedenen
Haustierarten untersucht. ALBRECHT (2006) erwärmte die Hoden von vier Bullen
über 48 Stunden um durchschnittlich 4,3°C durch Anbringen eines Suspensoriums.
24 Stunden nach Entfernung des Suspensoriums war eine Zunahme des Umfangs um
9%, des Volumens um 22% und der Länge und Breite um 7% zu verzeichnen. Auch
bei Hengsten, welche 48 Stunden ein Suspensorium trugen, führte der dadurch
erreichte Temperaturanstieg um 2 bis 3°C zu einer Verringerung der
Disulfidbindungen und der Chromatinstabilität, gemessen mittels sperm chromatin
structure assay (SCSA). Betroffen waren hier vor allem primäre Spermatozyten,
während sich bereits im Nebenhoden befindliche Spermien keinen Schaden nahmen
(LOVE u. KENNEY 1999). Dieses deckt sich mit Ergebnissen anderer Studien. So
bewirken die meisten Methoden zur Erwärmung der Hoden Schäden an den primären
Spermatozyten und den frühen Spermatiden. Einige Studien weisen aber auch Effekte
auf die Spermatogonien und die Sertolizellen nach (SETCHELL 2006). Ebenfalls bei
Bullen konnte nach 48 Stunden Erwärmung mittels Suspensorium Auswirkungen auf
die Samenqualität beobachtet werden. So kam es zwar nicht zu einer Veränderung
der Ejakulatmenge, wohl aber zu einer geringen Verschlechterung der Motilität sowie
zu massiven Schäden der Morphologie (VOGLER et al. 1993).

Literaturübersicht
32
Im Falle einer akuten Orchitis kommt es ebenfalls zu einer Hyperthermie am Hoden
(SINOWATZ 2001).
Diese Erkrankung führt innerhalb kürzester Zeit zu irreparablen Schäden am
Keimepithel mit erheblicher Beeinträchtigung der Samenbeschaffenheit. Auch
subakute und chronische Orchitiden können den Untergang des Keimepithels
bedingen. In experimentellen Studien zeigten sich die durch Wärmeapplikation
enstandenen Schäden oftmals als reversibel. In einigen Fällen aber zeigten sich nicht
zu unterschätzende Langzeiteffekte, welche die Fruchtbarkeit nachhaltig
einschränkten (SETCHELL 2006). Es existieren aber auch große individuelle
Schwankungen zwischen den einzelnen Rassen einer Tierart sowie einzelnen Tieren
einer Spezies.
Bei Lamas führte die vierwöchige Haltung in Wärmeställen bei 29°C zu einer
Abnahme der Spermienkonzentration, die ein bis vier Wochen nach Hitzeeinwirkung
am stärksten war. Gleichzeitig stieg der Anteil morphologisch abweichender Spermien
an. Ab der dritten Woche sank die Mortalität der Spermien bis zu einem Minimum von
20% in der achten Woche ab (SCHWALM et al. 2007).
Die mittlere Blutflussgeschwindigkeit fiel bei allen vier von ALBRECHT (2006)
untersuchten Bullen, z. T. biphasisch, ab. Die Werte normalisierten sich sehr
unterschiedlich. Nach initialem Abfall bei allen Bullen, zeigten zwei Bullen einen
weiteren kontinuierlichen Abfall bis Woche zwei mit anschließender Normalisierung in
Woche vier bzw. fünf. Bei einem Bullen normalisierte sich der Blutfluss zwischen
Woche zwei und vier mit leichten Schwankungen. Der letzte Bulle zeigte bis Woche
vier sehr niedrige Werte mit anschließendem Anstieg auf Ausgangsniveau in Woche
fünf. Keinen Einfluss auf den Blutfluss in der Arteria testicularis hat eine Kühlung des
Skrotums. Hier kam es lediglich zu einem reduzierten Blutfluss innerhalb der
Skrotalgefäße (GLODE et al. 1984).
Zur Darstellung der Ödematisierung eines einzelnen Hodens wurde sowohl von
ALBRECHT (2006) als auch SCHWEIZER (2006) die Ermittlung des Grauwertes
genutzt. So zeigten die vier Bullen von SCHWEIZER (2006) maximale Grauwerte in
Woche eins nach Wärmeapplikation. Ein Abfall auf Aufgangswerte erfolgte, mit einer
Ausnahme in Woche sechs, in den Wochen zwei bis vier. Bei drei Hengsten, welche

Literaturübersicht
33
eine Erwärmung der Hoden um durchschnittlich 2 bis 4°C mittels Suspensorium über
48 Stunden erfahren hatten, zeigten sich unterschiedliche Ergebnisse in der
Auswertung des mittleren Grauwertes. So zeigte ein Hengst kaum Veränderungen,
wohingegen ein anderer Hengst einen massiven Abfall direkt nach der
Wärmeapplikation mit anschließender Regeneration zeigte. Der dritte Hengst zeigte
ebenfalls einen massiven Abfall des Grauwertes direkt nach Wärmeapplikation mit
einem Anstieg auf Ausgangswert bis Woche sieben nach Wärmeapplikation.
HE gefärbte Hodengewebsschnitte von Lamas, nach vierwöchiger Haltung im
Wärmestall, wurden von SCHWALM et al. (2007) untersucht. Der Anteil vollständig
degenerierter Hodentubuli betrug in der Kontrollgruppe 5%, unmittelbar nach der
vierwöchigen Hyperthermie 40% und sank innerhalb der darauf folgenden zwei bis
sechs Wochen auf 10% ab. Der am häufigsten bei Hyperthermiestudien
dokumentierte Effekt des Zelluntergangs ist die Apoptose unter Beteiligung von
reaktiven Sauerstoffspezies, sowie dem Tumor supressor Gen p53 und der
Freisetzung von Cytochrom C (SETCHELL 2006).
So wurde auch die Apoptoserate von SCHWALM et al. (2007) mit Hilfe des TUNEL-
Verfahrens untersucht. In der Kontrollgruppe waren 22 bis 30 Zellen pro 100
Hodentubuli positiv. Es zeigten sich keine signifikanten Veränderungen gegenüber
den behandelten Tieren, welche drei, fünf, sieben und neun Wochen nach der
vierwöchigen Wärmephase kastriert wurden. Betroffen waren bei allen Tieren vor
allem primäre Spermatozyten. Insgesamt wurde postuliert, dass nicht eine
verminderte Apoptoserate sondern eine vermehrte Proliferation der Keimzellen wieder
eine Normalisierung der Spermaparameter bedingte.
Eine 45-minütige Erhöhung der Umgebungstemperatur auf 42°C führte bei Ratten zu
einem Anstieg TUNEL-positiver Zellen bis 15 Minuten nach Wärmeapplikation, gefolgt
von einer Abnahme. Betroffen waren vor allem die Typ A-Spermatogonien, während
die Spermatiden relativ resistent erschienen (KHAN u. BROWN 2002).
Mithilfe des Ki-67-Verfahrens stellten SCHWALM et al. (2007) die Proliferation im
Hoden dar. Hierbei wurde ein Quotient aus positiven Zellen gegenüber negativen
Zellen, bezogen auf zehn Hodentubuli aufgestellt. In 20,5% der Tubuli der
Kontrolltiere zeigten sich keine positiven Zellen. Dagegen stieg die Zahl auf 54,4%

Literaturübersicht
34
direkt nach der Hitzeeinwirkung, anschließend erfolgte ein langsamer Abfall auf 11,1%
sechs Wochen nach Hitzeeinwirkung.
Durch den Nachweis von Cytochrom C und Caspasen wurde eine Beteiligung der
Mitochrondrien an den apoptotischen Prozessen nachgewiesen (SINHA HIKIM et al.
2003). Auch zeigte sich in Hoden nach 20 Minuten bei 43°C mittels RNA-Extraktion
und PCR eine verminderte Anzahl DNA - reparierender Gene, sowie eine vermehrte
Anzahl apoptotischer Zellen und Hitzeschockproteine (ROCKETT et al. 2001).

Material und Methoden
35
3 Material und Methoden
Dieser Tierversuch wurde mit dem Aktenzeichen Tierversuchsnummer 08/1469
genehmigt.
3.1 Tiere
Für die Studie standen insgesamt zehn Beagle-Rüden im Alter von ein bis drei Jahren
(Durchschnitt zu Beginn der Studie 23,7 Monate) aus der
Reproduktionsmedizinischen Einheit der Kliniken, Klinik für Kleintiere mit einem
durchschnittlichen Gewicht von 14,8 kg zur Verfügung. Die Rüden wurden in Gruppen
von zwei bis drei Hunden in Außenzwingern mit Schutzhütten gehalten. Einmal täglich
wurden sie mit einer kommerziellen Mischung aus Feucht- und Trockenfutter gefüttert.
Wasser stand ad libitum zur Verfügung. Drei Tiere dienten als Kontrollgruppe; bei
sieben Hunden wurde ein- bzw. zweimalig eine skrotale Temperaturerhöhung
induziert. Alle Hunde befanden sich während des gesamten Studienzeitraumes in
einem guten Pflege- und Allgemeinzustand. Anhand andrologischer,
spermatologischer und mikroskopischer Parameter wurde bei allen Rüden klinische
und bakterielle Geschlechtsgesundheit sowie Normospermie diagnostiziert.
Material und Methoden
36
3.2 Versuchsaufbau
3.2.1 Vorlauf
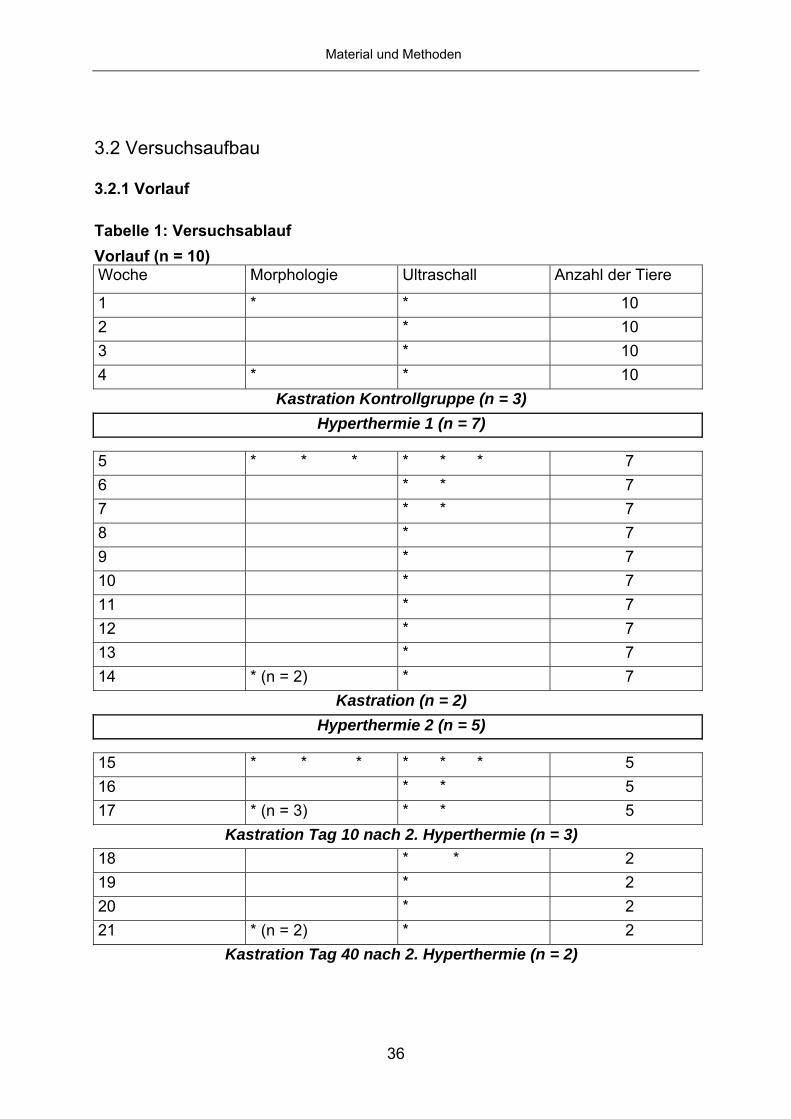
Tabelle 1: Versuchsablauf Vorlauf (n = 10) Woche Morphologie Ultraschall Anzahl der Tiere
1 * * 10 2 * 10 3 * 10 4 * * 10
Kastration Kontrollgruppe (n = 3) Hyperthermie 1 (n = 7)
5 * * * * * * 7 6 * * 7 7 * * 7 8 * 7 9 * 7 10 * 7 11 * 7 12 * 7 13 * 7 14 * (n = 2) * 7
Kastration (n = 2) Hyperthermie 2 (n = 5)
15 * * * * * * 5 16 * * 5 17 * (n = 3) * * 5
Kastration Tag 10 nach 2. Hyperthermie (n = 3) 18 * * 2 19 * 2 20 * 2 21 * (n = 2) * 2
Kastration Tag 40 nach 2. Hyperthermie (n = 2)

Material und Methoden
37
Zu Beginn und am Ende einer vierwöchigen Vorlaufzeit wurden Hoden, Nebenhoden
und Prostata aller zehn Hunde morphologisch untersucht. Außerdem fand einmal pro
Woche eine Ultraschalluntersuchung statt. Die Rüden der Kontrollgruppe (n = 3)
wurden anschließend kastriert.
3.2.2 Wärmeapplikation
Bei den verbleibenden sieben Rüden wurde über einen Zeitraum von 48 Stunden eine
Temperaturerhöhung im Bereich des Skrotums erzeugt. Alle sechs Stunden wurden
die Rüden an der Leine zum Harn- und Kotabsatz auf eine Wiese geführt. Wasser
stand ad libitum zur Verfügung, gefüttert wurden die Hunde wie gewohnt einmal
täglich. Um eine kontinuierliche Wärmeapplikation zu gewährleisten, wurde den
Hunden ein Suspensorium angelegt. Dieses bestand aus einer äußeren wasser- und
luftundurchlässigen Schicht aus thermoplastischem Kautschuk (Ethylen-Propylen-
Dien-Kautschuk und Polypropylen, Fa. Engineering Technology Marketing GmbH,
Saalburg-Eversdorf) wie er zur Abdeckung für Türfeststeller im Automobil
Verwendung findet. Die zweite Schicht bestand aus Rolta Polsterwatte (Fa.
Tierärztebedarf J. Lenecke, Schortens) um das Skrotum gegen mechanische
Schädigungen zu schützen. Direkt dem Hodensack anliegend befand sich ein weiches
Babysöckchen, bestehend aus 60% Baumwolle, 20% Nylon, 19% Polyester und 1%
Elastane-Lycra (Fa. KiK Textilien und Non-Food GmbH, Bönen).
Das Suspensorium wurde mit Hilfe von vier gepolsterten Bändern (Abb. 1) an einem
handelsüblichen Geschirr befestigt, so dass ein sicherer Sitz gewährleistet war.

Material und Methoden
38
Abbildung 1: Suspensorium zur Erhöhung der Skrotaltemperatur bei Rüden
In das Suspensorium wurde ein Temperaturfühler (Fa. TFA Klima Logger, Dostmann
GmbH, Wertheim-Reicholtsheim) so eingearbeitet, dass er dem Skrotum unmittelbar
auflag. Das zugehörige Kabel wurde so am Geschirr befestigt, dass kein Zug auf den
Fühler entstehen konnte und dann über die Abdeckung der Box zum Sender geleitet.
Die Messstation gab ein Alarmsignal ab, sobald sich die Temperatur aus dem
vorgegebenen Temperaturbereich (36 bis 42°C) entfernte. Auf diese Weise konnte ein
evtl. Verrutschen des Fühlers oder des Suspensoriums sofort bemerkt und behoben
werden. Alle 60 Minuten wurde die aktuelle Skrotaltemperatur aufgezeichnet.
Während des gesamten 48-stündigen Zeitraums der Wärmeapplikation wurden die
Rüden lückenlos im Wechsel von zwei Personen beaufsichtigt. Falls erforderlich
wurde zusätzlich ein Halskragen aufgelegt um die Tiere am Benagen des
Suspensoriums sowie der Haltekonstruktionen und Temperaturkabel zu hindern.
3.2.3 Untersuchungen nach 48-stündiger Wärmeapplikation
Im Anschluss an die Wärmeapplikation wurden alle sieben Rüden über neun Wochen
folgenden Untersuchungen unterworfen: unmittelbar und 24 Stunden nach Entfernen
des Suspensoriums morphologische Untersuchung sowie Sonographie von Hoden
und Prostata und dopplersonographische Untersuchung der Hodenperfusion.

Material und Methoden
39
In den darauf folgenden drei Wochen wurden eine zweimalige sonographische
Untersuchung von Hoden und Prostata sowie dopplersonographische
Untersuchungen der Hodenperfusion in drei- bis viertägigen Abständen durchgeführt,
danach wurden eine sonographische Untersuchung von Hoden und Prostata sowie
dopplersonographische Untersuchung der Hodenperfusion pro Woche durchgeführt.
Am Ende der neun Wochen zusätzlich eine morphologische Untersuchung von Hoden
und Prostata. Danach wurden zwei Rüden kastriert. Bei den fünf verbleibenden
intakten Rüden fand eine zweite Hyperthermiephase, identisch zur ersten statt. Die
daran anschließenden Untersuchungen entsprachen der oben beschriebenen
Vorgehensweise. Drei Rüden wurden zehn Tage und zwei Rüden 40 Tage nach der
zweiten Wärmeapplikation kastriert.
3.3 Morphologische Untersuchung
Die morphologische Untersuchung der Genitalorgane erfolgte in der von GÜNZEL-
APEL (1994) beschriebenen Vorgehensweise. Die äußere Haut des Skrotums wurde
adspektorisch auf ihre Beschaffenheit und eventuelle Veränderungen untersucht.
Palpatorisch wurde die Verschieblichkeit der einzelnen Schichten gegeneinander und
gegen den Hoden geprüft. Der Skrotalumfang wurde mit Hilfe eines Maßbandes
gemessen. An den Hoden wurden folgende Parameter erfasst: adspektorisch
geschätzte Größe, mittels Schieblehre gemessene Länge und Breite, Lage, Form und
Konsistenz der Hoden. Die Nebenhodenanteile, Kopf, Körper und Schwanz, wurden
auf ihre Abgrenzbarkeit untersucht. Am Nebenhodenschwanz wurde zusätzlich die
Größe geschätzt und Form, Lage und Konsistenz bestimmt. Ferner wurden die
Samenstränge bezüglich der Verschieblichkeit der Schichten und die
Hodensacklymphknoten bezüglich ihrer Palpierbarkeit untersucht.
Bei der rektalen Palpation der Prostata wurden die Größe sowie Oberfläche, Lappung,
Schmerzhaftigkeit und Konsistenz beurteilt.
Die Untersuchung von Penis und Präputium erfolgt durch Adspektion und Palpation.

Material und Methoden
40
3.4 Ultraschalluntersuchung
Alle Ultraschalluntersuchungen wurden mit einem Gerät des Typs Logiq 5 Pro
(General Electrics Medical Systems, Solingen) durchgeführt.
Die Hunde wurden auf einer gepolsterten Unterlage in Seitenlage verbracht. Aufgrund
der guten Kooperation der Hunde war eine Sedation in keinem Fall erforderlich. Zur
besseren Ankopplung wurden der Hodensack und ein kleiner Bereich paramedian ca.
3 cm vom Penis entfernt geschoren und handelsübliches Ultraschallgel aufgetragen.
Die Hoden wurden nacheinander mit einem 12-MHz-Linearschallkopf untersucht. Die
Darstellung erfolgte im Real-time-Modus im Längs- und Querschnitt. Es wurde die
Echogenität und Homogenität des Hodenparenchyms beurteilt. Länge und Breite der
Hoden wurden am jeweils größten Durchmesser gemessen. Anschließend wurde
mittels Farbdoppler die Arteria marginalis im kranialen Hodenabschnitt und der
Blutfluss durch dreimalige Messung der systolischen Maximalgeschwindigkeit (SPV)
in cm/s, der Widerstandsindex (RI) und der Pulsatilitätindex (PI) ermittelt. Alle Bilder
wurden zur Dokumentation und zur Auswertung abgespeichert.
Von der Prostata wurde im zweidimensionalen Bild (5 MHz Konvexschallkopf) die
Parenchymbeschaffenheit beurteilt und die maximale Länge und Höhe jeweils
zweimal gemessen.
3.5 Kastration und Aufbereitung der Hoden
Die Kastration der Hunde fand zu den im Versuchsablauf (Tabelle 1) angegebenen
Zeitpunkten in der Klinik für Kleintiere der Tierärztlichen Hochschule Hannover statt.
Nach Einleitung der Narkose wurde der Bereich zwischen Penis und Skrotum rasiert
und mehrfach nach Standard der Klinik gewaschen und desinfiziert. Die Operation
erfolgte nach der in der Klinik üblichen Vorgehensweise. Direkt nach Entnahme der
Hoden wurden diese mittels scharfer Mikrotomklinge in drei Teile geschnitten und in
entsprechend beschriftete (Name des Rüden, Tag der Kastration, Hodenabschnitt)
Behälter mit 4% Formalinlösung gelegt.

Material und Methoden
41
Nach 24 Stunden wurden die Hoden aus dem Formalin entnommen und im Institut für
Pathologie in jeweils eine Scheibe pro Abschnitt geschnitten. Diese Proben wurden
anschließend in einen Gewebeeinbettungsautomaten (Hypercenter II, Shandon,
Frankfurt/Main) überführt. In dem Gerät wurden die Proben nach einstündiger Fixation
in 10%igem Formalin und Spülung mit Leitungswasser in einer aufsteigenden
Alkoholreihe (70%, 85%, 96%) dehydriert. Anschließend erfolgte die Entfernung des
Alkohols mittels Isopropanol und Essigsäure-Butylester.
Am nächsten Tag erfolgte die Einbettung mit Hilfe des Einbettsystems Tissue Tek
(Sakura, AT Zoeterwounde, Niederlande). Hierbei wurden die Präparate in 64°C
warmes Paraplast Plus® (Shandon, Frankfurt/Main) eingebettet und auf 5°C gekühlt.
Aus den Blöcken wurden mittels Rotationsmikrotom (Microm International GmbH,
Walldorf) 2 μm dicke Schnitte angefertigt und auf Superfrost® Plus Objektträger (Fa.
Menzel Gläser, Braunschweig) gezogen. Um gleichmäßige Präparate zu erhalten
wurden die Blöcke um ca. ¼ herunter geschnitten. Die darauf folgenden Schnitte
wurden der Untersuchung zugeführt. Vor dem unmittelbaren Aufziehen wurden die
Schnitte in ein Wasserbad mit Raumtemperatur überführt, anschließend folgte ein
zweites Wasserbad bei 40°C, um gestreckte gleichmäßige Schnitte zu gewährleisten.
Im weiteren Verlauf wurden die Schnitte im Wärmeschrank (Heraeus, Hanau) bei
70°C getrocknet. Insgesamt wurden pro Hund sechs Schnitte angefertigt, pro Hoden
je ein Schnitt aus dem kranialen, mittleren und kaudalen Segment.

Material und Methoden
42
3.6 Histologische und immunhistochemische Verfahren
3.6.1 Hämalaun-Eosin Färbung
Für die HE-Färbung wurde ein Färbautomat Leica 4040 (Leica Microsystems GmbH,
Wetzlar) genutzt. Dieser arbeitet nach einem standardisierten Protokoll (siehe 9.1).
Die Schnitte wurden entparaffiniert und anschließend mit Hämalaun und Eosin gefärbt
und im weiteren Verlauf mittels eines Eindeckautomaten (Promounter RCM 2000,
Medite Medizintechnik Burgdorf) mit Deckgläsern versehen.
3.6.2 TUNEL - Verfahren
Zur Darstellung apoptotischer Zellen wurde zum einen das TUNEL-Verfahren
angewandt. Hierfür kam das ApopTag® Plus Kit (Fa. Chemicon® International, Ltd.,
Hampshire, United Kingdom) zum Einsatz, welches die Aufbereitung der Schnitte in
mehreren Schritten und enzymatische Reaktionen mittels TdT-Enzymen beinhaltet
(siehe 9.1). Hierbei werden alle Zellen mit fragmentierter DNA im mikroskopischen
Bild braun dargestellt. Jede dieser verfärbten Zellen wird zusätzlich auf ihre
Morphologie untersucht. Ausgezählt wurden nur Zellen mit typischer pyknotischer
Morphologie. Als Positivkontrolle diente ein Präparat der Milchdrüse einer Ratte.
Folgeschnitte der zu untersuchende Gewebeschnitte wurden mit PBS anstelle der
working strenghth Tdt solution inkubiert und dienten so der Negativkontrolle.
3.6.3 Immunhistologischer Nachweis der aktivierten Form der Caspase-3
Zum Nachweis der aktivierten Caspase-3 wurde nach der ABC-Methode (siehe 9.1)
für formalinfixierte Paraffinmaterialien verfahren. Als Blockserum wurde ein
Ziegenserum verwandt. Dieses dient der Blockierung unspezifischer Bindungsstellen.
Als primärer Antikörper diente ein polyklonaler, vom Kaninchen stammender
Antikörper. Dieser ist gegen die p17 Untereinheit der aktivierten Caspase-3 gerichtet.

Material und Methoden
43
Als sekundärer Antikörper kam ein GAR-b (Goat-anti Rabbit, Ziege anti Kaninchen
Antikörper) zum Einsatz.
Aufgrund der beschleunigten Umwandlung von DAB (3,3`-Diaminobenzidin-
tetrahydrochloridihydrat) durch das Enzym Peroxidase entsteht an der Reaktionsstelle
ein präzipitierendes Farbprodukt, welches sich ablagert und im mikroskopischen Bild
braun darstellt.
Auch hier lief eine Positivkontrolle der Methode in Form eines Präparates einer
Hundemilz mit. Als Negativkontrollen dienten parallele Schnitte des zu
untersuchenden Gewebes welche mit 1:3000 in PBS mit 1% BSA verdünntem
Kaninchenserum anstelle des Primärantikörpers (Kaninchen anti Caspase-3) inkubiert
wurden.
3.6.4 Ki-67-Antigen
Für den Nachweis des Ki-67-Antigens kam der monoklonale Anti-Human Ki-67-
Antikörper Clone MIB-1 zum Einsatz. Dieser monoklonale Mausantikörper gilt als
Referenzantikörper für formalinfixierte Präparate. Die Reaktion läuft ebenfalls mit Hilfe
der ABC – Methode ab. Als zweiter Antikörper fand ein GAM-b (Goat-anti Mouse)
Verwendung (siehe 9.1). Zunächst werden hierbei antigene Vernetzungen gelöst und
mittels NHS (Natural Horse Serum) blockiert. Anschließend kann der spezifische
primäre Antikörper binden. Als weitere Komponente wird das mit einer Peroxidase
konjungierte Streptavidin hinzugegeben. Dieses kann nun mit Hilfe eines
peroxidasespezifischen Chromogens sichtbar gemacht werden. So wird das Antigen
über zwei Antikörperbrücken indirekt lokalisiert und sichtbar gemacht. Bei jeder
immunhistochemischen Reaktion lief ein Schnittpräparat eines Hundedarms als
Positivkontrolle mit. Für die Negativkontrolle wurden Schnitte des zu untersuchenden
Gewebes durch Weglassen des Primärantikörper mit geführt.

Material und Methoden
44
3.7 Mikroskopische Auswertung
Die mikroskopische Auswertung erfolgte mittels eines Standard-Binokular-
Lichtmikroskops (Carl-Zeiss, Oberkochen; Okular x10, Objektiv x16/x40). Hierbei
wurden die Schnitte meanderförmig durchgemustert. Dabei wurde zunächst der
Hodenbereich, anschließend der Nebenhodenbereich untersucht.
In der HE-Färbung wurden 50 nahezu runde Tubuli bei 40-facher Vergrößerung
ausgewertet. Zwischen den erfassten Tubuli wurden jeweils vier Gesichtsfelder frei
gelassen. Nach Durchmusterung des gesamten Schnittes wurde der Vorgang von der
anderen Seite her wiederholt, bis 50 verschiedene Tubuli ausgewertet waren. Im
Bereich der Nebenhoden wurden immer zwei Gesichtsfelder übersprungen und 25
Anschnitte ausgewertet.
In den Hoden wurden folgende Degenerationsanzeichen quantitativ erfasst:
A: verkleinerte Keimzellen E: geschwollene Spermatozyten
B: geschrumpfte Tubuli F: Zelldebris
C: Sertoli only G: verdickte Basalmembran
D: multinukleäre Riesenzellen H: vakuolig aufgelockertes Zytoplasma
Als Sertoli only klassifizierte Hodentubuli zeigen keine Keimzellen, sondern
ausschließlich Sertolizellen.
An den Nebenhoden wurde auf folgende Veränderungen geachtet:
a) unreife Spermien c) Zelldebris
b) vakuolig aufgelockertes Epithel d) keine Spermien im Lumen
Die für das TUNEL-Verfahren, Caspase-3-Nachweis und den Ki-67 Nachweis
präparierten Schnitte wurden auf zwei verschiedene Arten ausgewertet. Zum einen
erfolgte eine Durchmusterung analog zu den HE-gefärbten Schnitten. Ausgewertet
wurden ebenfalls 50 quer angeschnittene Tubuli bei 40facher Vergrößerung.

Material und Methoden
45
Die zweite Auswertung richtete sich an den Sertolizellen aus. In jedem zehnten
Gesichtsfeld wurden die quer angeschnittenen Tubuli bei zehnfacher Vergrößerung
aufgesucht und bei 40facher Vergrößerung begutachtet. Gezählt wurden pro Tubulus
die Anzahl der Sertolizellen und die Anzahl der positiven Zellen. Wurden im letzten
Tubulus 100 Sertolizellen überschritten, wurde dieser trotzdem komplett ausgewertet.
Im Nebenhodenbereich wurden insgesamt 25 Querschnitte auf die Anzahl positiver
Zellen untersucht. Positive Zellen waren anhand ihrer Braunfärbung zu erkennen.
3.8 Statistische Auswertung
Aufgrund der geringen Tierzahl war eine statistische Auswertung nicht möglich. Die
Auswertung erfolgte daher rein deskriptiv. Die Berechnung der Mittelwerte,
Standardabweichungen sowie Minima und Maxima und die Erstellung der Graphiken
erfolgte mit dem Programm Microsoft Excel®.
Die Auswertung erfolgte vergleichend zwischen den behandelten Tieren und der
Kontrollgruppe sowie der einzelnen Tiere vor und nach der Wärmeapplikation, immer
unter Berücksichtigung der Kastrationszeitpunkte sowie der Anzahl der
Wärmeapplikationen.

Ergebnisse
46
4 Ergebnisse
Alle für die Studie verwendeten Rüden waren während des gesamten
Untersuchungszeitraumes klinisch allgemein- und geschlechtsgesund.
Die Skrotaltemperatur betrug vor der Wärmeapplikation durchschnittlich 32,1°C
(28,4 - 35,2°C). Während der ersten 48-stündigen Hyperthermiephase (n = 7) wurde
diese Basaltemperatur um durchschnittlich 3,7°C auf 35,8°C (34,8 bis 41,6°C)
erhöht. Im Zuge der zweiten Hyperthermiephase (n = 5) wurde eine
Temperaturerhöhung um 3,8°C auf 35,9°C (34,0 bis 38,8°C) erreicht.
4.1 Befunde der morphologischen Untersuchung
Bei allen sieben Hunden zeigte sich direkt nach Abnahme des Suspensoriums eine
Ödematisierung des Skrotums. Bei sechs Hunden kam es zusätzlich zu einer
Rötung. Zwei Hunde zeigten leichte Scheuerstellen am Skrotum. Bereits 24 Stunden
nach Abnahme des Suspensoriums war die Ödematisierung fast vollständig
abgeklungen. Zu diesem Zeitpunkt war bei drei Hunden noch eine leichte Rötung der
Skrotalhaut zu verzeichnen. Die Scheuerstellen heilten innerhalb einer Woche ab.
Nach der zweiten Wärmeapplikation waren bei drei Hunden umschriebene,
geringgradig verdickte Bereiche der Skrotalhaut direkt nach Abnahme des
Suspensoriums zu erkennen.
Eine Zu- oder Abnahme der Hodendimensionen und des skrotalen Umfangs konnte
nach den Hyperthermiephasen nicht festgestellt werden (Abb. 2 und 3)

Ergebnisse
47
10,0
11,0
12,0
13,0
14,0
15,0
16,0cm
Hyperthermie 1 Hyperthermie 2
Abbildung 2: Mittlerer (±SD) Skrotalumfang (cm), vor und nach der 1. und 2. skrotalen Hyperthermie; n = Anzahl der Rüden, T = Tag, SD = Standardabweichung
n=10 n=7 n=5 n=3 n=2
Vorlauf 9 Wochen 10. T 40. T

Ergebnisse
48
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0cm
Länge Breite
Hyperthermie 1 Hyperthermie 2
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5cm
Länge BreiteHyperterthermie 1 Hyperthermie 2
Abbildung 3: Mittlere (±SD) Länge und Breite des a) linken und b) rechten Hodens, gemessen mittels Schieblehre, vor und nach der 1. und 2. skrotalen Hyperthermie, n = Anzahl der Rüden, T = Tag, SD = Standardabweichung
n=10 n=7 n=5 n=3 n=2
Vorlauf 9 Wochen 10. T 40. T
n=10 n=7 n=5 n=3 n=2
Vorlauf 9 Wochen 10. T 40 T.
a)
b)

Ergebnisse
49
4.2 Befunde der sonographischen Untersuchung
Die im Ultraschall gemessenen Werte für die Hodenlänge waren im Mittel
geringgradig höher, die der Breite eher etwas niedriger als die mittels Schieblehre
erhobenen Befunde. Alle sonographisch beurteilten Parameter zeigten weder
innerhalb der Gruppen noch im Gruppenvergleich auffällige Unterschiede.
Nach der Wärmeapplikation waren keine Veränderungen der Hoden- und
Prostatadimensionen nachweisbar (Abb. 4).
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0cm Kontrollgruppe n=3
Vorlauf n=7Nach 1. Hyperthermie n=7Nach 2. Hyperthermie n=5
Abbildung 4: Mittlere (±SD) Länge (L) und Breite (B) des linken (li) und rechten (re) Hodens (H), sowie der Länge (L) und Höhe (H) der Prostata (P) in der Kontrollgruppe, vor und nach der 1. und 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Auch im zeitlichen Verlauf zeigten die Hoden- und Prostatadimensionen keine
deutlichen hyperthermiebedingten Schwankungen (Abb. 5 und 6). Alle messbaren
Abweichungen waren maximal im Millimeterbereich zu finden.
H li L H li B H re L H re B P L P H

Ergebnisse
50
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0cm
Länge BreiteHyperthermie 1 Hyperthermie 2
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0cm
Länge BreiteHyperthermie 1 Hyperthermie 2
Abbildung 5: Mittels Ultraschall gemessene mittlere (±SD) Länge und Breite des linken (a) und rechten (b) Hodens vor und nach der 1. und 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
a)
n=10 n=7 n=5 n=2
Vorlauf 9 Wochen 10 Tage 40 Tage
b)
n=10 n=7 n=5 n=2
Vorlauf 9 Wochen 10 Tage 40 Tage

Ergebnisse
51
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0cm
Länge HöheHyperthermie 1 Hyperthermie 2
Abbildung 6: Mittels Ultraschall gemessene mittlere (±SD) Länge und Höhe der Prostata vor und nach der 1. und 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Die Untersuchung der Hodenperfusion erbrachte für die systolische
Maximalgeschwindigkeit (SPV) eine sehr hohe Variationsbreite. Dies betraf sowohl
die Hoden eines Tieres (Vergleich links-rechts) als auch die einzelnen
Untersuchungstage. Lediglich ein leichter Anstieg nach der Wärmeapplikation auf die
winkelkorrigierten Werte war zu erkennen (Abb. 7). Die Kontrollgruppe zeigte
insgesamt niedrigere Werte.
Vorlauf 9 Wochen 10 Tage 40 Tage
n=10 n=7 n=5 n=2

Ergebnisse
52
0,0
5,0
10,0
15,0
20,0
25,0
30,0
35,0
m/s
Hyperthermie 1 Hyperthermie 2
0,0
5,0
10,0
15,0
20,0
25,0
30,0
35,0
m/s
Hyperthermie 1 Hyperthermie 2
Abbildung 7: Mittlere (±SD) winkelkorrigierte systolische Maximal-geschwindigkeit (SPV) der A. marginalis am linken (a) und rechten (b) Hoden vor und nach der 1. und 2. sktoralen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
n = 10 n = 7 n = 5 n = 2
n = 10 n = 7 n = 5 n = 2
Vorlauf 9 Wochen 10 Tage 40 Tage
a)
b)
Vorlauf 9 Wochen 10 Tage 40 Tage

Ergebnisse
53
Der Mittelwertvergleich zwischen den einzelnen Gruppen zeigt ebenfalls einen nur
geringgradigen Anstieg der systolischen Maximalgeschwindigkeit unmittelbar nach
den Hyperthermiephasen an (Abb. 8).
0,0
5,0
10,0
15,0
20,0
25,0
30,0
35,0m/s Kontrollgruppe n=3
Vorlauf n=7Nach 1. Hyperthermie n=7Nach 2. Hyperthermie n=5
Abbildung 8: Mittlere (±SD) winkelkorrigierte systolische Maximalgeschwindig-keit (SPV) der A. marginalis des rechten und linken Hodens vor und nach der 1. und 2. sktoralen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Der Pulsatilitätsindex (PI) und der Widerstandsindex (RI) variierten in ihrer
Ausprägung ebenfalls so stark, dass keine eindeutigen Auswirkungen der
Wärmeapplikation zu erkennen waren.
Dies wurde sowohl im zeitlichen Verlauf (Abb. 9) als auch im unmittelbaren Vergleich
nach den Wärmeapplikationen sichtbar (Abb. 10). Insgesamt waren beide Indizes in
der Kontrollgruppe niedriger als bei den Versuchstieren.
SPV links SPV rechts

Ergebnisse
54
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
PI RI
Hyperthermie 1 Hyperthermie 2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
PI RI
Hyperthermie 1 Hyperthermie 2
Abbildung 9: Mittlerer (±SD) Pulsatilitätsindex (PI) und Widerstandsindex (RI) am a) linken Hoden und b) rechten Hoden vor und nach der 1. und 2. sktoralen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
a)
b)
n = 10 n = 7 n = 5 n = 2
n = 10 n = 7 n = 5 n = 2
Vorlauf 9 Wochen 10 Tage 40 Tage
Vorlauf 9 Wochen 10 Tage 40 Tage

Ergebnisse
55
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8Kontrollgruppe n=3Vorlauf n=7Nach 1. Hyperthermie n=7Nach 2. Hyperthermie n=5
Abbildung 10: Mittlerer (±SD) Pulsatilitätsindex (PI) und Widerstandsindex (RI) am linken und rechten Hoden in der Kontrollgruppe, im Vorlauf sowie unmittelbar nach der 1. und 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
PI links PI rechts RI links RI rechts

Ergebnisse
56
4.3 Histologische und immunhistochemische Befunde
Insgesamt waren nach Wärmeapplikation nur sehr geringe histologische und
immunhistochemische Veränderungen feststellbar.
4.3.1 Hoden
HE-gefärbte Präparate
In den HE-gefärbten Hodenschnitten fiel vor allem eine vakuolige Auflockerung auf
(Abb. 11). In der Kontrollgruppe waren durchschnittlich 3,2 Hodentubuli (2-5)
betroffen, während es neun Wochen nach einmaliger Wärmeapplikation im Mittel 4,2
Hodentubuli (2-7) waren. Zehn Tage nach der zweiten Wärmeapplikation zeigten
durchschnittlich 4 Hodentubuli (1-8) vakuolige Veränderungen. 40 Tage nach zweiter
Wärmebehandlung waren mit 6,7 (3-11) die meisten Hodentubuli betroffen.

Ergebnisse
57
Abbildung 11: a) Vakuolig veränderte Zellen des Keimepithels eines Hundes 40 Tage nach der 2. skrotalen Hyperthermie (x200); b) unveränderte Hodentubuli eines Hundes der Kontrollgruppe (x100)
a)
b)

Ergebnisse
58
Auch das vermehrte Auftreten von Zelldebris in den Hodentubuli (Abb. 12) ließ einen
zeitlichen Zusammenhang mit der Wärmeapplikation erkennen. In der Kontrollgruppe
war der Anteil betroffener Tubuli mit durchschnittlich 1,9 (1-4) am geringsten. Neun
Wochen nach einmaliger Wärmeapplikation war durchschnittlich die größte Anzahl
Tubuli betroffen (x̄ = 3,1; 1-6). Zehn Tage nach zweiter Wärmeapplikation zeigten
sich mit 2,0 (0-3) nahezu identische Werte gegenüber der Kontrollgruppe. 40 Tage
nach der zweiten Wärmeapplikation zeigte sich die durchschnittliche Anzahl an
Tubuli mit Zelldebris (x̄ = 2,9; 1-5) ähnlich wie neun Wochen nach einmaliger
Wärmeapplikation.
Abbildung 12: Zelldebris im Lumen eines Hodentubulus eines Hundes neun
Wochen nach einmaliger skrotaler Hyperthermie (x400)
Die übrigen erfassten Degenerationserscheinungen waren in allen Versuchsgruppen
nur vereinzelt nachweisbar (Abb. 13).

Ergebnisse
59
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
9,0
10,0
Anzahl betroffener Tubuli pro 50 Tubuli
Sertoli only
Multinukleäre Riesenzellen
Geschwollene Spermatozyten
Zelldebris
Verdickte Basalmembran
Vakuolig aufgelockertes Epithel
Abbildung 13: Mittlere Anzahl (±SD) der von verschiedenen Degenerations-erscheinungen betroffenen Hodentubuli in der Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 (n = 2) Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Apoptose positive Zellen (TUNEL-Verfahren)
Bezogen auf 100 Sertolizellen zeigte sich eine deutliche Zunahme der TUNEL-
positiven Zellen (Abb. 14) zehn und 40 Tage nach der zweiten Wärmeapplikation.
Dagegen lag in der Kontrollgruppe mit durchschnittlich 2,9 (0-8) und neun Wochen
nach einmaliger Wärmeapplikation mit 2,0 (0-4) TUNEL-positiven Zellen pro 100
Sertolizellen ein ähnliches Niveau vor. Zehn Tage nach zweiter Wärmeapplikation
stellten sich mit durchschnittlich 5,7 (3-10) und 40 Tage nach zweiter
Wärmeapplikation mit 6,7 (3-17) nahezu doppelt so viele Zellen TUNEL-positiv dar
(Abb. 15).
Kontrollgruppe 9 Wochen 10 Tage 40 Tage

Ergebnisse
60
Abbildung 14: a) Hodentubuli eines Hundes der Kontrollgruppe, eine einzelne TUNEL - positive Zelle (↑) in einem Tubulus (x100); b) vier TUNEL - positive Zellen (↑) in einem Hodentubulus eines Hundes zehn Tage nach der 2. skrotalen Hyperthermie (x400)
a)
b)

Ergebnisse
61
0,0
2,0
4,0
6,0
8,0
10,0
12,0
Anzahl TUNEL-positiver Zellen pro 100 Sertolizellen
Kontrollgruppe n=3
9 Wochen n=2
10 Tage n=3
40 Tage n=2
Abbildung 15: Anzahl TUNEL - positiver Zellen pro 100 Sertolizellen in der Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40 Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Von dem Anstieg TUNEL-positiver Zellen war in erster Linie das Stadium vier des
Keimepithelzyklus nach FOOTE und SWIERSTRA (1972) betroffen (Abb. 16). Die
am meisten positiven Zellen waren primäre Spermatozyten und Spermatogonien.

Ergebnisse
62
0
5
10
15
20
25
30
1 2 3 4 5 6 7 8
%
Abbildung 16: Prozentualer Anteil aller Tubuli mit TUNEL - positiven Zellen pro 100 Sertolizellen aufgeteilt nach den Stadien des Keimepithelzyklus über alle Gruppen zusammengefasst (Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 (n = 2) Tage nach der 2. skrotalen Hyperthermie)
Auch die Auswertung pro 50 Hodentubuli erbrachte die höchsten durchschnittlichen
Anteile TUNEL – positiver Zellen zehn Tage und 40 Tage nach der zweiten
Wärmeapplikation. In der Kontrollgruppe fanden sich im Mittel 3,9 (0-8) TUNEL –
positive Zellen pro 50 Hodentubuli. Allerdings war unter dieser Voraussetzung auch
neun Wochen nach einmaliger Wärmeapplikation (x̄ = 6,9; 3-12) eine moderate
Zunahme zu verzeichnen. Zehn Tage (x̄ = 10; 4-16) und 40 Tage (x̄ = 8,8; 4-14) nach
zweiter Wärmeapplikation stellte sich ca. die doppelte Anzahl an Zellen TUNEL –
positiv dar (Abb. 17).

Ergebnisse
63
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
Anzahl TUNEL-positiver Zellen pro 50
Tubuli
Kontrollgruppe n=3
9 Wochen n=2
10 Tage n=3
40 Tage n=2
Abbildung 17: Anzahl TUNEL-positiver Zellen pro 50 Hodentubuli in der Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40 Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Apoptose positive Zellen (Caspase-3-Nachweis)
Insgesamt war die Anzahl positiver Zellen im Caspase-3-Nachweis deutlich geringer
als nach Auswertung mit dem TUNEL-Verfahren.
Bezogen auf 100 Sertolizellen zeigte die Kontrollgruppe durchschnittlich 0,5 (0-2)
Caspase-3-positive Zellen (Abb. 18), während neun Wochen nach einmaliger
Hyperthermie mit 2,9 (0-6) die meisten positiven Zellen pro 100 Sertolizellen
detektiert wurden. Zehn Tage nach zweiter Wärmeapplikation waren durchschnittlich
1,1 (0-5) Caspase – 3 – positive Zellen pro 100 Sertolizellen zu finden. 40 Tage nach
der zweiten Wärmeapplikation lag eine ähnliche Situation wie neun Wochen nach
einmaliger Wärmeapplikation vor (x̄ = 2,2; 0-9) (Abb. 20).

Ergebnisse
64
Abbildung 18: a) Hodentubuli eines Kontrolltieres, keine Caspase - 3 - positive Zelle (x100); b) Hodentubuli eines Rüden neun Wochen nach der 1. skrotalen Hyperthermie, drei Caspase - 3 - positive Zellen (↑) in einem Hodentubulus (x100)
a)
b)

Ergebnisse
65
Abbildung 19: Caspase-3-positive Zelle (↑) im Hodentubulus eines Hundes zehn Tage nach der 2. skrotalen Hyperthermie (x400)

Ergebnisse
66
0,0
1,0
2,0
3,0
4,0
5,0
6,0
Anzahl Caspase-3-positiver Zellen pro 100 Sertolizellen
Kontrollgruppe n=3
9 Wochen n=2
10 Tage n=3
40 Tage n=2
Abbildung 20: Anzahl Caspase-3-positiver Zellen pro 100 Sertolizellen in der Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40 Tage nach der 2. skrotalen Hyperthermie
Die größte Anzahl Caspase-3-positiver Zellen fanden sich in den Stadien vier und
sechs des Keimepithelzyklus nach FOOTE und SWIERSTRA (1972) (Abb. 21),
betroffen waren wiederum Spermatogonien und primäre Spermatozyten.

Ergebnisse
67
0
5
10
15
20
25
30
35
40
45
1 2 3 4 5 6 7 8
%
Abbildung 21: Prozentualer Anteil aller Tubuli mit Caspase-3-positiven Zellen pro 100 Sertolizellen aufgeteilt nach den Stadien des Keimepithelzyklus über alle Gruppen zusammengefasst (Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 (n = 2) Tage nach der 2. skrotalen Hyperthermie)
Bei der Auswertung pro 50 Hodentubuli war eine ähnliche Tendenz zu erkennen.
In der Kontrollgruppe war die Anzahl Caspase-3-positiver Zellen am geringsten
(x̄ = 0,9; 0-5). Neun Wochen nach einmaliger Wärmeapplikation waren mit mehr als
doppelt so vielen Caspase-3-positiven Zellen (x̄ = 3,9; 0-7) im Vergleich zur
Kontrollgruppe, die meisten apoptotischen Zellen zu beobachten.
Zwischen den Tieren, die zehn Tage nach zweiter Wärmeapplikation (x̄ = 2,2; 0-17)
und denjenigen, die 40 Tage nach zweiter Wärmeapplikation (x̄ = 2,3; 0-6) kastriert
wurden, zeigte sich nur ein geringer Unterschied (Abb. 22).

Ergebnisse
68
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
Anzahl Caspase-3-positiver Zellen pro 50
Tubuli
Kontrollgruppe n=3
9 Wochen n=2
10 Tage n=3
40 Tage n=2
Abbildung 22: Anzahl Caspase -3 - positiver Zellen pro 50 Hodentubuli in der Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40 Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung

Ergebnisse
69
Proliferierende Zellen im Ki-67-Antigen Nachweis
Einzig Ki-67 positive Spermatogonien wurden ausgezählt (Abb. 23).
In der Kontrollgruppe (x̄ = 118,6; 62-176) sowie 40 Tage nach zweiter
Wärmeapplikation (x̄ = 119,7; 74-172) zeigte sich die größte Anzahl an
proliferierenden Zellen pro 100 Sertolizellen (Abb. 24).
Zehn Tage nach der zweiten Wärmeapplikation (x̄ = 111,6; 38-228) war die
Proliferationsrate pro 100 Sertolizellen etwas geringer. Neun Wochen nach
einmaliger Wärmeapplikation (x̄ = 84,2; 56-120) hingegen war die geringste
Proliferation zu beobachten (Abb. 24).

Ergebnisse
70
Abbildung 23: a) Ki - 67 - positive Spermatogonien (↑) in den Hodentubuli eines Hundes zehn Tage nach der 2. skrotalen Hyperthermie (x100); b) einzelne Ki - 67 - positive Spermatogonien in dem Hodentubuli eines Hundes der Kontrollgruppe (x100); positiv erscheinende Spermatozyten wurden nicht mitgezählt
b)
a)

Ergebnisse
71
0,0
20,0
40,0
60,0
80,0
100,0
120,0
140,0
160,0
180,0
Anzahl Ki-67-positiver Zellen pro 100 Sertolizellen
Kontrollgruppe n=3
9 Wochen n=2
10 Tage n=3
40 Tage n=2
Abbildung 24: Anzahl Ki-67-positiver Zellen pro 100 Sertolizellen in der Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40 Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Bei der Auszählung pro 50 Hodentubuli zeigte sich in der Kontrollgruppe die
niedrigste Anzahl proliferierender Zellen mit durchschnittlich 199,6 (110-302). Neun
Wochen nach einmaliger Hyperthermie fanden sich 231,1 (134-310) positive Zellen
pro 50 Hodentubuli, zehn Tage nach zweiter Wärmeapplikation 239,0 (120-350). Die
40 Tage nach zweiter Wärmeapplikation gewonnenen Hoden zeigten mit 293,8 (110-
492) ungefähr die gleiche Proliferation wie die anderen Versuchsgruppen auf (Abb.
25).

Ergebnisse
72
0,0
50,0
100,0
150,0
200,0
250,0
300,0
350,0
400,0
Anzahl Ki-67-positiver Zellen pro 50 Tubuli Kontrollgruppe n=3
9 Wochen n=2
10 Tage n=3
40 Tage n=2
Abbildung 25: Anzahl Ki-67-positiver Zellen pro 50 Hodentubuli in der Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40 Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung

Ergebnisse
73
4.3.2 Nebenhoden
HE – gefärbte Präparate
Im Bereich der Nebenhoden lag die Anzahl der Tubulusanschnitte mit vakuolig
aufgelockerten Tubulusepithelzellen (Abb. 26) auf einem höherem Niveau als in den
Hoden. Bei allen wärmebehandelten Tieren war die mittlere Anzahl (x̄ ) betroffener
Tubulusanschnitte pro 25 ausgezählter Tubulusanschnitte gegenüber der
Kontrollgruppe (x̄ = 2,4; 1-5) deutlich erhöht. So zeigten sich neun Wochen nach
einmaliger Wärmeapplikation durchschnittlich 12,7 (1-25) Tubulusanschnitte mit
vakuolig aufgelockerten Epithelzellen. Zehn Tage nach zweiter Wärmeapplikation
waren es 16,2 (1-25) und 40 Tage nach zweiter Wärmeapplikation 11,3 (0-25)
Tubulusanschnitte pro 25 ausgezählter Tubulusanschnitte (Abb. 27).

Ergebnisse
74
Abbildung 26: a) Vakuolig aufgelockerte Epithelzellen in einem Nebenhodentubulus eines Hundes zehn Tage nach der 2. skrotalen Hyperthermie (x100); b) Nebenhodentubulusanschnitt ohne vakuolig verändertes Epithel eines Hundes der Kontrollgruppe (x100)
b)
a)
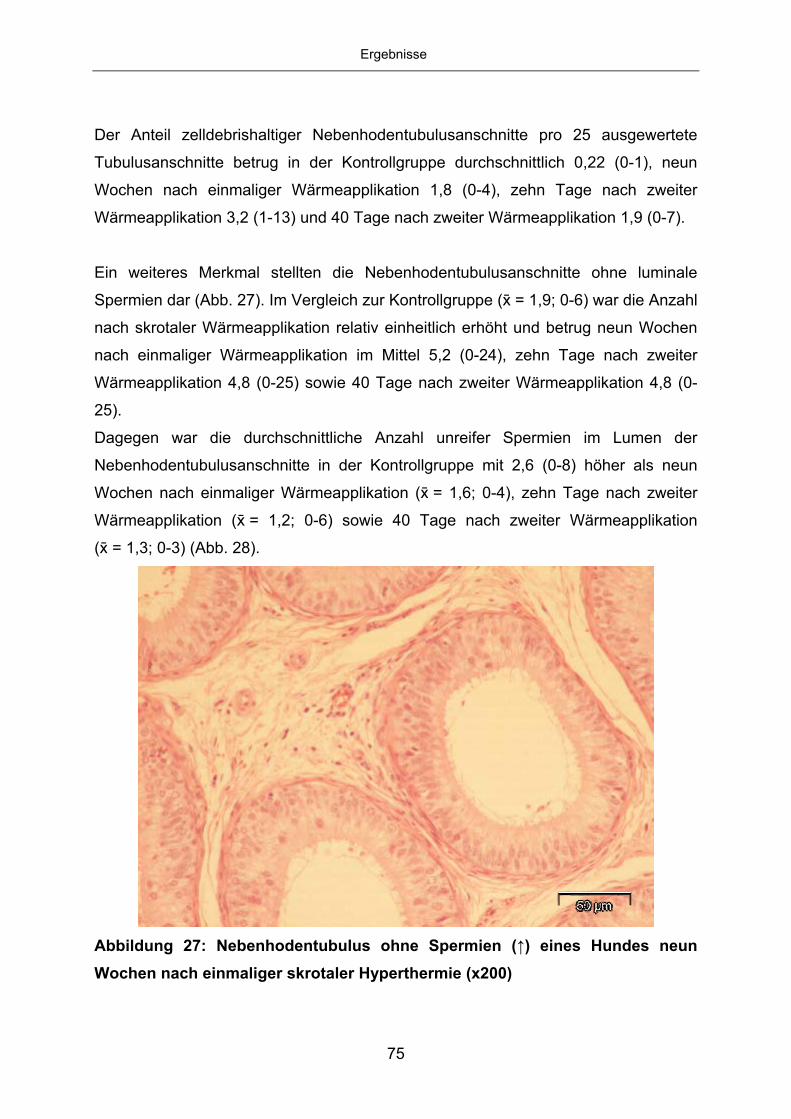
Ergebnisse
75
Der Anteil zelldebrishaltiger Nebenhodentubulusanschnitte pro 25 ausgewertete
Tubulusanschnitte betrug in der Kontrollgruppe durchschnittlich 0,22 (0-1), neun
Wochen nach einmaliger Wärmeapplikation 1,8 (0-4), zehn Tage nach zweiter
Wärmeapplikation 3,2 (1-13) und 40 Tage nach zweiter Wärmeapplikation 1,9 (0-7).
Ein weiteres Merkmal stellten die Nebenhodentubulusanschnitte ohne luminale
Spermien dar (Abb. 27). Im Vergleich zur Kontrollgruppe (x̄ = 1,9; 0-6) war die Anzahl
nach skrotaler Wärmeapplikation relativ einheitlich erhöht und betrug neun Wochen
nach einmaliger Wärmeapplikation im Mittel 5,2 (0-24), zehn Tage nach zweiter
Wärmeapplikation 4,8 (0-25) sowie 40 Tage nach zweiter Wärmeapplikation 4,8 (0-
25). Dagegen war die durchschnittliche Anzahl unreifer Spermien im Lumen der
Nebenhodentubulusanschnitte in der Kontrollgruppe mit 2,6 (0-8) höher als neun
Wochen nach einmaliger Wärmeapplikation (x̄ = 1,6; 0-4), zehn Tage nach zweiter
Wärmeapplikation (x̄ = 1,2; 0-6) sowie 40 Tage nach zweiter Wärmeapplikation
(x̄ = 1,3; 0-3) (Abb. 28).
Abbildung 27: Nebenhodentubulus ohne Spermien (↑) eines Hundes neun Wochen nach einmaliger skrotaler Hyperthermie (x200)

Ergebnisse
76
0,0
5,0
10,0
15,0
20,0
25,0
Anzahl betroffener Tubuli pro 25
Tubulusanschnitte
unreife Spermien
vakuolig aufgelockertes Epithel
Zelldebris
keine Spermien im Lumen
Abbildung 28: Mittlere Anzahl (±SD) der von verschiedenen Degenerationserscheinungen betroffenen Nebenhodentubulusanschnitte in der Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 (n = 2) Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Apoptose positive Zellen (TUNEL-Verfahren)
Im Nebenhodenepithel zeigten sich in der Kontrollgruppe durchschnittlich 0,4 (0-2)
TUNEL-positive Zellen pro 25 ausgewertete Tubulusanschnitte. Neun Wochen nach
einmaliger Wärmeapplikation (x̄ = 3,8; 1-9) sowie zehn Tage nach zweiter
Wärmeapplikation (x̄ = 4; 0-10) konnte eine ca. 8-fach höhere Anzahl TUNEL-
positiver Zellen im Vergleich zur Kontrollgruppe festgestellt werden. Dagegen
bestand zwischen der Kontrollgruppe und den Tieren, die 40 Tage nach zweiter
Wärmeapplikation (x̄ = 0,8; 0-4) kastriert wurden, kein nennenswerter Unterschied
(Abb. 29).
Kontrollgruppe 9 Wochen 10 Tage 40 Tage

Ergebnisse
77
Auch im Nebenhodenstroma war eine Zunahme der TUNEL-positiven Zellen (um das
ca. 3-4-Fache) vor allem neun Wochen nach einmaliger und zehn Tage nach zweiter
Wärmeapplikation zu beobachten.
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
Anzahl TUNEL-positiver Zellen pro 25
Tubulusanschnitte / Gesichtsfelder
Lumen Epithel
Stroma
Abbildung 29: Anzahl TUNEL - positiver Zellen pro 25 Nebenhodentubulus-anschnitte im Stroma, Epithel und Lumen in der Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2), sowie zehn (n = 3) und 40 Tage (n = 2) nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Apoptose positive Zellen (Caspase-3-Nachweis)
Bei der Auswertung der Nebenhodentubulusanschnitte zeigten sich vor allem im
Nebenhodenepithel Caspase-3-positive Zellen. In der Kontrollgruppe (x̄ = 0,2; 0-1)
Kontrollgruppe 9 Wochen 10 Tage 40 Tage

Ergebnisse
78
wurde nur eine geringe Anzahl Caspase-3-positiver Zellen gefunden. Auch neun
Wochen nach einmaliger Wärmeapplikation (x̄ = 0,6; 0-2) lagen die Caspase-3-
positiven Zellen auf einem niedrigen Niveau. Lediglich zehn Tage nach zweiter
Wärmeapplikation war ein Anstieg der Caspase-3-positiven Zellen um das 8-Fache
bzw. 3-Fache auf durchschnittlich 1,6 positive Zellen pro 25 ausgezählte
Nebenhodentubulusanschnitte (0-10) zu verzeichnen. 40 Tage nach zweiter
Wärmeapplikation (x̄ = 0,2; 0-1) zeigte sich kein Unterschied im Vergleich zur
Kontrollgruppe (Abb. 30).
0,0
0,5
1,0
1,5
2,0
2,5
Anzahl Caspase-3-positiver Zellen pro 25
Tubulusanschnitte / Gesichtsfelder
Lumen Epithel
Stroma
Abbildung 30: Anzahl Caspase-3-positiver Zellen pro 25 Nebenhoden-tubulusanschnitte im Stroma, Epithel und Lumen in der Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 Tage (n = 2) nach der 2. zweiten skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Kontrollgruppe 9 Wochen 10 Tage 40 Tage

Ergebnisse
79
Proliferierende Zellen im Ki – 67 – Antigen Nachweis
In den Nebenhoden wiesen in erster Linie die Epithelzellen eine verstärkte
Proliferation auf (Abb. 31). Die Proliferationsrate lag in der Kontrollgruppe bei
durchschnittlich 9,2 Zellen pro 25 ausgezählte Tubulusanschnitte und war neun
Wochen nach einmaliger Wärmeapplikation um ca. die Hälfte reduziert (x̄ = 5,8; 0-
16). Zehn Tage (x̄ = 11,0; 1-30) bzw. 40 Tage (x̄ = 11,3; 0-44) nach zweiter
Wärmeapplikation waren die Resultate annähernd identisch und auch in Bezug zur
Kontrollgruppe nicht deutlich abweichend (Abb. 32).
Abbildung 31: Ki-67 - positive Zellen (↑) im Nebenhodentubulusepithel eines Hundes neun Wochen nach einmaliger skrotaler Hyperthermie (x400)

Ergebnisse
80
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
16,0
18,0
20,0
Anzahl Ki-67-positiver Zellen pro 25
Tubulusanschnitten / Gesichtsfeldern Stroma
Epithelial
Luminal
Abbildung 32: Anzahl Ki-67 - positiver Zellen pro 25 Nebenhoden-tubulusanschnitte im Stroma, Epithel und Lumen in der Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 Tage (n = 2) nach der zweiten skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
Kontrollgruppe 9 Wochen 10 Tage 40 Tage

Diskussion
81
5 Diskussion
5.1 Versuchsdurchführung
Wie bei Bullen, Hengsten und Ratten gezeigt wurde, kann eine moderate
Hyperthermie über einen Zeitraum von nur 48 Stunden oder eine kurzfristige
Hyperthermie bei Temperaturen um 40°C pathologische Veränderungen der Hoden
erzeugen (COULTER u. KASTELIC 1994; LOVE u. KENNEY 1999; KHAN u. BROWN
2002; WU 2005; ALBRECHT 2006; SCHWEIZER 2006; SCHWALM et al. 2007).
Aufgrund dessen wurde in der vorliegenden Studie ein Zeitraum von 48 Stunden für
die skrotale Wärmeapplikation gewählt. Mit Hilfe eines Suspensoriums wurde eine
moderate Temperaturerhöhung von durchschnittlich 3,8°C erzielt. In der ersten
Hyperthermiephase betrug die Durchschnittstemperatur 35,8°C (34,8-41,6°C), in der
zweiten Hyperthermiephase wurde eine durchschnittliche Temperatur von 35,9°C
(34,0-38,8°C) erreicht. Ein vergleichbarer Anstieg wurde auch bei Hengsten (34,7 bis
35,3°C bei SCHWEIZER 2006 und eine Erhöhung um 2-3°C bei LOVE u. KENNEY
1999) erreicht. Bei Bullen wurde über 48 Stunden eine durchschnittliche
Temperaturerhöhung um 3,9°C; 5,5°C; 6,3°C und 6,6°C erreicht (ALBRECHT 2006).
Aufgrund der kontinuierlichen Überwachung der Tiere konnte die Hyperthermie über
den gesamten 48-stündigen Versuchszeitraum relativ konstant gehalten werden.
Zusätzlich erwies sich das verwendete Temperaturmesssystem für die stündliche
Temperaturkontrolle und -dokumentation als geeignet. Das integrierte Alarmsystem
ermöglichte die sofortige Kontrolle und Korrektur des Sitzes des Suspensoriums.
Aufgrund der kleinen Tierzahlen pro Gruppe konnte lediglich eine deskriptive
statistische Auswertung erfolgen. Dies ist auch in den Studien von WU (2005),
ALBRECHT (2006) und SCHWEIZER (2006) der Fall.
Da die hier verwendeten Rüden ein regelmäßiges Handling gewohnt waren, verhielten
sie sich bei allen Untersuchungsmaßnahmen äußert kooperativ, so dass keine
Sedation erforderlich war. Die morphologische Untersuchung erfolgte nach der von
GÜNZEL-APEL (1994) beschriebene Vorgehensweise.

Diskussion
82
Die chirurgische Entfernung der Hoden wurde unter sorgfältiger Schonung des
Hodengewebes vor Quetschungen und anderweitigen Schädigungen durchgeführt.
Gleiches galt auch für die Aufteilung der Hoden in jeweils drei Segmente (kranial,
Mitte, kaudal) unmittelbar nach der Gonadektomie. Diese Dreiteilung der Hoden
wurde gewählt, um den Nebenhoden in seinen funktionellen Abschnitten
Nebenhodenkopf, -körper und -schwanz zu erfassen. Aufgrund der geringen Anzahl
TUNEL- und Caspase-3-positiver Zellen erfolgte die Auswertung insgesamt für den
gesamten Nebenhoden. Die Aufbereitung der Organe für die histologischen und
immunhistochemischen Untersuchungen fand unter exakter Einhaltung der
erforderlichen Arbeitsschritte statt. Zum Nachweis apoptotischer Zellen kamen zwei
verschiedene Verfahren (TUNEL-Verfahren und Caspase-3-Nachweis) zur
Anwendung, so dass nach den Empfehlungen von ELMORE (2007) eine möglichst
große Genauigkeit gewährleistet werden konnte.
Der Nachweis des Ki-67-Antigens hat sich zur Feststellung der Zellproliferation
aufgrund des im Vergleich zu anderen Verfahren größeren Zeitfensters bewährt
(WROBEL et al. 1993) und wurde deswegen in dieser Studie eingesetzt.
Die mikroskopische Auswertung mittels meanderförmiger Durchmusterung der
Gewebeschnitte erlaubte eine möglichst objektive Beurteilung, da jeder Bereich des
Schnittes erreicht wurde. Auch konnten durch „Überspringen“ einer bestimmten
Anzahl an Gesichtsfeldern Hodentubuli und Nebenhodentubulusanschnitte nach dem
Zufallsprinzip ausgewählt werden.
5.2 Morphologische Befunde
Bei der morphologischen Untersuchung zeigten die Rüden unmittelbar nach
Beendigung der skrotalen Hyperthermie eine auch bei Bullen und Hengsten
(ALBRECHT 2006; SCHWEIZER 2006) beschriebene Ödematisierung des Skrotums.
Diese verschwand innerhalb von 24 Stunden. Die bei sechs Hunden beobachtete
zusätzliche Rötung der Skrotalhaut beruhte wahrscheinlich auf der nicht vollständig
vermeidbaren Reibung des Suspensoriums am Skrotum. Diese kann zu einer

Diskussion
83
zusätzlichen lokalen Temperaturerhöhung infolge einer Reizung des Gewebes und
der Blutgefäße geführt haben. Da diese leichten Irritationen bei drei Hunden (Rüde: 5,
Kastration neun Wochen nach einmaliger Wärmeapplikation; Rüde: 6, Kastration zehn
Tage nach zweiter Wärmeapplikation; Rüde: 10, Kastration 40 Tage nach zweiter
Wärmeapplikation) bereits nach 24 Stunden, bei den anderen drei Hunden (Rüde: 4,
Kastration neun Wochen nach einmaliger Wärmeapplikation; Rüde: 7, Kastration zehn
Tage nach zweiter Wärmeapplikation; Rüde: 8, Kastration 10 Tage nach zweiter
Wärmeapplikation) wenige Tage später vollständig abgeklungen waren, sind
allenfalls geringfügige Effekte auf das Hodenparenchym zu erwarten. Dies gilt auch
für die bei zwei Hunden (Rüde: 4, Kastration neun Wochen nach einmaliger
Wärmeapplikation; Rüde: 8, Kastration zehn Tage nach zweiter Wärmeapplikation)
zusätzlich festgestellten Scheuerstellen mit punktuell stärkerer Reizung der
Skrotalhaut durch das Suspensorium. Trotz der subjektiv feststellbaren
Ödematisierung des Skrotums war nur eine geringe Zunahme des Skrotalumfanges
von 0,2 cm nachzuweisen. Als Grund für eine solche Zunahme in Anschluss an einen
Temperaturstimulus sieht ALBRECHT (2006) eine provozierte aseptische
Entzündungsreaktion. In dem Falle dieser Studie scheint es sich allerdings eher um
einen Prozess des Skrotums zu handeln, welches als Reaktion auf den
Temperaturstimulus Flüssigkeit eingelagert hat. Aufgrund der Tatsache, dass diese
Ödematisierung bereits nach 24 Stunden wieder normalisiert wurde ist von keiner
Auswirkung auf den Hoden auszugehen.
Alle weiteren morphologischen Parameter wiesen weder tendenzielle noch
spezifische Veränderungen im Zusammenhang mit der Hyperthermie auf.
5.3 Sonographische Befunde
Für die Ultraschalluntersuchungen wurden die in der Literatur als ideal bezeichneten
Bedingungen geschaffen (BARR 1992; LÜERSSEN u. JANTHUR 2007). So wurde bei
allen Rüden das Skrotum sowie zusätzlich ein Bereich paramedian des Penis
geschoren. Des Weiteren wurden durch eine weiche Unterlage, Ruhe und einen

Diskussion
84
abgedunkelten Raum die Untersuchungsbedingungen optimiert. Aufgrund der
Kooperationsbereitschaft der Hunde konnten alle Messungen in Ruhe und mit Sorgfalt
durchgeführt werden.
Auch die im Rahmen der Ultraschalluntersuchung gemessenen Parameter ließen
keine Veränderung mit klarem Bezug zur Wärmeapplikation erkennen.
Schwankungen zeigten sich sowohl im Vergleich der einzelnen Gruppen zur
Kontrollgruppe als auch innerhalb der einzelnen Gruppen im Vergleich vor und direkt
nach Wärmeapplikation. Ebenso wiesen die Dimensionen der Prostata keine auf die
skrotale Hyperthermie zurückzuführenden Veränderungen auf.
Die systolische Maximalgeschwindigkeit (SPV) war in allen Versuchsphasen von
starken inter- und intraindividuellen Schwankungen geprägt. Lediglich unmittelbar
nach den Hyperthermiephasen stieg die mittlere systolische Maximalgeschwindigkeit
in allen Versuchsgruppen gegenüber der Kontrollgruppe. Anschließend lagen die
Schwankungen im Bereich der Kontrollgruppe. ALBRECHT (2006) dagegen konnte
bei 4 Bullen, welche ebenfalls eine skrotale Hyperthermie mittels Suspensorium
erfahren haben, initial einen Abfall der Blutflußgeschwindigkeit in der Arteria
testicularis ein bis zwei Wochen nach Abnahme des Suspensoriums feststellen,
postuliert aber eine entzündungsbedingte Hyperämie während und direkt nach
Hyperthermie mittels Suspensorium. Anschließend kam es auch in dieser Studie zu
hohen individuellen Schwankungen. Gemessen wurde 30 Stunden nach Abnahme
des Suspensoriums, sodass diese vermutete entzündungsbedingte Hyperämie nicht
ganz ausgeschlossen werden konnte. Aus diesem Grunde wurde bei den Rüden
dieser Studie direkt nach Abnahme des Suspensoriums sowie nach 24 Stunden eine
sonographische Untersuchung durchgeführt. Ein anschließender Abfall der mittleren
systolischen Maximalgeschwindigkeit konnte nicht nachgewiesen werden. Ursache
hierfür könnten etwas geringere Temperaturerhöhungen gewesen sein. Aber auch
eine geringere Empfindlichkeit der Rüden könnte dem zugrunde liegen.
Bei Ratten hatte eine skrotale Temperaturerhöhung keinen Einfluss auf den
intratestikulären Blutfluss (GALIL u. SETCHELL 1988). Dasselbe wird für eine skrotale
Temperaturerniedrigung bei Hunden geltend gemacht (GLODE et al. 1984).

Diskussion
85
5.4 Histologische Befunde
Hoden Die HE-Färbung wurde als Übersichtsfärbung genutzt. Die Gewebeschnitte wurden
auf die von GOEDKEN et al. (2008) beschriebenen Degenerationserscheinungen
(verkleinerte Keimzellen, geschrumpfte Tubuli, „Sertoli only“, multinukleäre
Riesenzellen, geschwollene Spermatozyten, Zelldebris und verdickte Basalmembran)
untersucht. Zusätzlich wurde das Merkmal „vakuolig aufgelockertes Zytolasma“
untersucht. Aufgrund des Alters der in dieser Studie verwendeten Beagle-Rüden von
durchschnittlich 23,7 Monaten zu Studienbeginn zeigten sich bei allen Tieren nur
geringe Degenerationserscheinungen. Alle in dieser Studie untersuchten Hunde
zeigten adulte Spermatogenese, wie sie bei FORD (1969) beschrieben ist. REHM
(2000) stellte bei 30% der untersuchten Beagle–Rüden im Alter von 13 Monaten eine
bilaterale segmentale Hypospermatogenese fest. GOEDKEN et al. (2008) dagegen
beobachteten bei 20% eine verringerte Spermatogenese, 75% dieser Tiere waren
sechs bis acht Monate alt. In diesem Alter sieht FORD (1969) erst eine beginnende
Spermatogenese, sodass derartige Befunde nicht auf Degeneration sondern auf das
junge Alter zurückzuführen sind. Befunde dieser Art waren bei keinem der in der
vorliegenden Studie verwendeten Beagle-Rüden festzustellen. Als Zeichen der
Degeneration wurden lediglich vereinzelte Hodentubuli mit Zelldebris und
multinukleären Riesenzellen nachgewiesen. GOEDKEN et al. (2008) nutze genau wie
REHM (2000) für toxikologische Studien gezüchtete Beagle-Rüden, allerdings aus
verschiedenen Laboren. Auch wenn diese bisher an keinem Versuche teilnahmen
scheint es doch in dieser Zuchtlinie, evt. erblich bedingte, zu stärkeren pathologischen
Veränderungen im Hodenepithel zu kommen, da es bei keinem in dieser Studie
verwendeten Rüden zu so starken histologischen Abweichungen kam. Zusätzlich zu
den oben beschriebenen Degenerationserscheinungen war in den
Hodentubulusepithelzellen eine vakuolige Auflockerung zu verzeichnen. Im Vergleich
zur Kontrollgruppe war die Anzahl der betroffenen Hodentubuli bei den 40 Tage nach
zweiter Wärmeapplikation kastrierten Tieren auf das Doppelte erhöht (Kontrollgruppe:
x̄ = 3,2 ± 0,9; 40 Tage nach zweiter Wärmeapplikation x̄ = 6,7 ± 2,2). Die vermehrte

Diskussion
86
Bildung zytoplasmatischer Bläschen kann für eine vakuolige Degeneration der Zellen
sprechen. Hierbei kommt es durch Verschiebung des Ionensystems der Zelle zu
einem vermehrten Flüssigkeitseinstrom. Diese vermehrte Flüssigkeit sammelt sich in
den erweiterten Zisternen des endoplasmatischen Retikulums oder des
Grundplasmas (SANDERSLEBEN et al. 1989). Der Prozess ist reversibel, kann aber
auch in nekrotische Prozesse übergehen. Eine vermehrte Bildung zytoplasmatischer
Bläschen wird grundsätzlich als Merkmal nekrotischer Prozesse angesehen
(ELMORE 2007). Zu bedenke wäre also, dass in der vorliegenden Studie bereits eine
einmalige Wärmeapplikation zu einer Verschiebung des Ionensystems führte, welche
noch nach neun Wochen feststellbar war. Die Bildung von Vakuolen wird aber auch
als charakteristisch für Hitzestress angesehen (MOORE 1924; DAVIS u. FIRLIT
1966). YIN et al. (1997) postuliert, dass Vakuolen sich immer im Anschluss an
apoptotische Prozesse bilden. So konnte bei experimentellem Kryptorchismus an
adulten Mäusen eine Korrelation zwischen Vakuolen und vermehrter DNA-
Fragmentation festgestellt werden (YIN et al. 1997). Auch zehn Tage nach einer
zweiten Wärmeapplikation zeigte sich eine ähnliche Anzahl Tubuli mit vakuoliger
Degeneration. 40 Tage nach der zweiten Wärmeapplikation erhöhte sich die Anzahl
der Tubuli mit vakuoliger Degeneration um ein Drittel. Ein gesetzter Wärmestimulus
wirkt sich also nachhaltig auf die Ionenbilanz der Hodentubuluszellen aus oder aber
bewirkt verstärkt Keimzellverluste über apoptotische Prozesse. Da auch in der
Kontrollgruppe eine geringe Anzahl an Hodentubuli mit vakuoliger Auflockerung zu
verzeichnen war, ist trotz größtmöglicher Sorgfalt im Umgang mit den Hoden eine
Artefaktbildung nicht vollständig auszuschließen. Aufgrund der Tatsache, dass bei
allen wärmebehandelten Tieren eine quantitative Zunahme der Hodentubuli mit
vakuoliger Degeneration eintrat, ist ein Effekt durch die Wärmeapplikation durchaus
zu vermuten. Diese Langzeiteffekte einer lokalen Hyperthermie sind möglicherweise
auch auf eine Schädigung der Sertolizellen zurückzuführen (SETCHELL 2006), oder
aber resultieren in einer verminderte Repopulation der apoptotischen Zellen.
Entsprechend waren bei den 40 Tage nach zweiter Wärmeapplikation kastrierten
Tieren mehr Hodentubuli mit Zelldebris feststellbar als bei den Kontrolltieren
(Kontrollgruppe x̄ = 1,9 ± 0,8; 40 Tage nach zweiter Wärmeapplikation x̄ = 2,9 ± 1,3),

Diskussion
87
was bedeuten würde, dass eine Zunahme vakuoliger, nekrotischer oder apoptotischer
Zellen zu einem Anstieg an Zelldebris führen kann. Hierbei ist zu bedenken, dass
gebildete apoptotische Restkörper phagozytiert werden oder im Gewebe bestehen
bleiben (MAJNO u. JORIS 1995). Dies könnten eine mögliche Ursache für das
vermehrte Auftreten von Zelledebris sein. Auch nekrotischen Zellen zerbrechen
(MAJNO u. JORIS 1995; ELMORE 2007) und könnten so ursächlich für den in dieser
Studie nachgewiesenen Zelldebris sein.
Bei der Auswertung der apoptotischen Zellen war die Anzahl der TUNEL-positiven
Zellen höher als die Anzahl aktivierter Caspase-3-positiver Zellen. Dies ist anhand der
Prinzipien der einzelnen Verfahren zu erklären. So wird im TUNEL-Verfahren
einzelsträngige DNA nachgewiesen, die auch bei nekrotischen Zellen vorkommen
kann. Mit dem Caspase-3-Verfahren wird spezifisch aktivierte Caspase-3
nachgewiesen, welche ausschließlich bei apoptotischen Prozessen auftritt (ELMORE
2007).
Im Einzelnen zeigten sich in der Auszählung pro 100 Sertolizellen im TUNEL-
Verfahren deutlich mehr positive Zellen bei allen wärmebehandelten Tieren als in der
Kontrollgruppe (x̄ = 2,9 0-8). Die größte Anzahl positiver Zellen wurde im basalen
Bereich der Hodentubuli gefunden und betraf somit die frühen
Spermatogenesestadien, vor allem primäre Spermatozyten und Spermatogonien.
Dies deckt sich mit den Ergebnissen anderer Studien (KHAN u. BRWON 2002;
SCHWALM et al. 2007). Das am häufigsten betroffene Stadium des Keimepithelzyklus
war das Stadium vier nach FOOTE und SWIERSTRA (1972), in welchem sich an der
Basalmembran Sertolizellen und Typ A Spermatogonien sowie weiter luminal
elongierte Spermatiden in Gruppen befinden. Neun Wochen nach einmaliger
Wärmeapplikation konnten durchschnittlich lediglich 2,0 (0-4) TUNEL-positive Zellen
pro 100 Sertolizellen, ähnlich der Kontrollgruppe, identifiziert werden. Nach der
zweiten Wärmeapplikation stieg die Anzahl der positiven Zellen auf einen fast dreifach
erhöhten Wert an (Tag 10: x̄ = 5,7 (3-10); Tag 40: x̄ = 6,7 (3-17)). Zu bedenken ist hier
jedoch, dass nicht nur eine zweite Hyperthermiephase stattgefunden hat, sondern
auch der Zeitraum zwischen Wärmeapplikation und Kastration deutlich geringer war.

Diskussion
88
Eine Regeneration auch nach zweiter Wärmeapplikation kann also mit den gewählten
Kastrationszeitpunkten nicht ausgeschlossen werden. Um eine Aussage
diesbezüglich treffen zu können, müsste die Anzahl TUNEL-positiver Zellen in einer
weiteren Gruppe mit Kastration neun Wochen nach zweiter Wärmeapplikation
bestimmt werden.
Mit dem Caspase-3-Nachweis wurden neun Wochen nach einmaliger
Wärmeapplikation (x̄ = 2,9; 0-6) fünfmal so viele positive Zellen pro 100 Sertolizellen
wie in der Kontrollgruppe (x̄ = 0,5; 0-2) identifiziert. Am häufigsten waren hier Tubuli
des Stadiums sechs des Keimepithelzyklus nach FOOTE und SWIERSTRA (1972)
betroffen, welches durch viele Gruppen von Typ B Spermatogonien und elongierten
Spermatiden gekennzeichnet ist. An der Basalmembran finden sich vor allem
Sertolizellen sowie Typ B Spermatogonien und vereinzelte Typ A Spermatogonien.
Auch in dem Caspase-3-Nachweis waren in dieser Studie vor allem primäre
Spermatozyten und Spermatogonien betroffen. Zehn Tage nach zweiter
Wärmeapplikation (x̄ = 1,1; 0-5) fanden sich mehr Hodentubuli mit Caspase-3 -
positiven Zellen als in der Kontrollgruppe, jedoch weniger als neun Wochen nach
einmaliger Wärmeapplikation. 40 Tage nach zweiter Wärmeapplikation (x̄ = 2,2; 0-9)
lag eine ähnliche Situation wie neun Wochen nach einmaliger Wärmeapplikation vor.
Der gesetzte Wärmestimulus erhöht demnach die Apoptoserate in den Zellen der
Hodentubuli so nachhaltig, dass eine vermehrte Aktivität der Caspase-3 auch zeitlich
versetzt nachzuweisen ist.
Bei der Auswertung pro 50 Hodentubuli war die gleiche Tendenz zu erkennen. Auch
hier wiesen alle Tiere, bei denen eine skrotale Hyperthermie erzeugt worden war, eine
erhöhte Anzahl apoptotischer Zellen auf. Im TUNEL-Verfahren trat dieser Effekt
zeitlich näher zum gesetzten Wärmestimulus auf. So betrug die Anzahl TUNEL-
positiver Zellen pro 50 Hodentubuli neun Wochen nach einmaliger Wärmeapplikation
mit 6,9 ± 2,8 etwa das Doppelte der Kontrollgruppe (3,9 ± 2,2). Zehn Tage (10,0 ± 3,3)
und 40 Tage nach der zweiten Hyperthermie (8,8 ± 5,5) waren die Werte sogar höher.

Diskussion
89
Auch die Anzahl Caspase-3-positiver Zellen pro 50 Hodentubuli stieg neun Wochen
nach der ersten Wärmeapplikation deutlich um das Vierfache der Kontrollgruppe (x̄ =
0,9 ± 1,3), auf 3,9 ± 2,5 an. Nach der zweiten Wärmeapplikation war die Anzahl
Caspase-3-positiver Zellen etwa doppelt so hoch wie in der Kontrollgruppe (Tag 10:
2,2 ± 4,0; Tag 40: 2,2 ± 1,6).
Demnach waren sowohl bei der Auswertung pro 100 Sertolizellen als auch pro 50
Hodentubuli jeweils zehn und 40 Tage nach zweiter Wärmeapplikation vermehrt
TUNEL-positive Zellen zu identifizieren, während neun Wochen nach einmaliger
Wärmeapplikation deutlich mehr Caspase-3-positive Zellen als in den anderen
Gruppen zu verzeichnen waren. Es ist davon auszugehen, dass dieser Befund auch
bei den Tieren vorlag die einer zweiten skrotalen Hyperthermie unterworfen wurden.
Eine einmalige Wärmeapplikation führte zu einer Erhöhung der Apoptoserate, welche
noch nach einem vollständigen Spermatogenesezyklus nachweisbar war. Dagegen
kam es nach der zweiten Wärmeapplikation möglicherweise aufgrund der
potenzierten Schädigung zu nekrotischer Prozessen und zu einem vermehrten
Untergang von Zellen. Die im Vergleich zu den Caspase-3-positiven Zellen absolut
größere Anzahl TUNEL-positiver Zellen schließt nekrotische Zellen mit ein
(MARTINVALET et al. 2005) und unterstützt hiermit die oben genannte These. Nicht
auszuschließen ist allerdings, dass generell kurz nach Wärmeapplikation eher
nekrotische Prozesse stattfinden. Da jedoch zehn bzw. 40 Tage nach erster
Wärmeapplikation keine Kastrationen durchgeführt wurden, kann diesbezüglich keine
Aussage getroffen werden. Auch denkbar wäre es, dass es aufgrund vermehrter
apoptotischer Prozesse zeitlich nah zu dem Temperaturstimulus zu einer vermehrten
Vakuolenbildung kommt. Diese entstehen in Korrelation zu apoptotischen Zellen (YIN
et al. 1997) und liegen in den Bereichen der Hodentubuli in denen vorher die
apoptotischen Zellen lagen. Neun Wochen nach einmaliger Wärmeapplikation kann
es bereits zu einer Rebesiedelung dieser Bereiche mit neuen Keimzellen gekommen
sein.
Beide Auswertungsverfahren (pro 100 Sertolizellen und pro 50 Hodentubuli) wurden
auch in anderen Studien angewandt (SINHA HIKIM et al. 2003; SCHWALM et al.

Diskussion
90
2007). Die Auswertung pro 50 Tubuli ist als genauere Methode zu beurteilen, da die
Auszählung apoptotischer Zellen pro 100 Sertolizellen, bedingt durch die variable
Anzahl an Sertolizellen pro Hodentubulus, eine sehr unterschiedliche Anzahl an
Hodentubuli berücksichtigt.
Bei Ratten kam es bereits nach 15-minütiger Erwärmung im Wasserbad bei 42°C zu
einem deutlicher Anstieg TUNEL-positiver Zellen im Hodengewebe (SINHA HIKIM et
al. 2003). Neun Tage nach diesem Hitzestimulus wiesen die Hodentubuli zwar
weiterhin schwere Schäden auf, die Anzahl TUNEL-positiver Zellen war jedoch
rückläufig. In der vorliegenden Studie waren dagegen auch neun Wochen nach einem
moderateren, jedoch länger andauernden Wärmestimulus deutlich mehr TUNEL-
positive Zellen nachweisbar als bei den Kontrolltieren die keine Hyperthermie erfahren
hatten. Eine vollständige Regeneration war bei den Ratten 56 Tage nach gesetztem
Wärmestimulus zu verzeichnen (SINHA HIKIM et al. 2003). Auch KHAN und BROWN
(2002) wiesen bei Ratten unter identischen Hyperthermiebedingungen einen Anstieg
TUNEL-positiver Zellen nach, der jedoch bereits 15 Stunden nach dem gesetzten
Hitzestimulus wieder absank. Bei Lamas, welche vier Wochen bei 29°C in einem
Klimastall gehalten wurden, waren trotz schlechterer Ejakulatbeschaffenheit keine
signifikanten Unterschiede in der Anzahl TUNEL-positiver Zellen drei Wochen nach
Ende der Wärmeapplikation nachweisbar. So scheinen die Veränderungen des
Spermas nicht die Folge einer erhöhten Apoptoserate gewesen zu sein. An
kryptorchiden Hunden im Alter von ein bis zwei Jahren konnte gezeigt werden, dass
die Spermatogenese umso schwerwiegender gestört ist, je weiter der betroffene
Hoden vom Skrotum entfernt liegt (inguinal, abdominal) und je höher demzufolge
seine Umgebungstemperatur ist (VERONESI et al. 2009). Auswirkungen auf den
kontralateralen skrotalen Hoden konnten bei keinem der zehn untersuchten Hunde
nachgewiesen werden.
Ein kurzfristiger, massiver Wärmestimulus scheint also einen größeren Effekt
auszulösen als eine moderate, über einen längeren, aber dennoch überschaubaren

Diskussion
91
Zeitraum bestehende Hyperthermie. Eine Regeneration des Keimepithels nach
Schädigung durch Wärmeeinwirkung scheint aber in allen Fällen möglich.
Die Auswertung der Proliferation im Hodengewebe brachte für beide
Auswertmethoden unterschiedliche Ergebnisse. So zeigte sich nach Auswertung von
50 Tubuli die geringste Proliferation in der Kontrollgruppe (x̄ = 199,6; 110-302)
wohingegen alle Versuchgruppen nahezu identische Ergebnisse brachten. So nahm
die Proliferation bei allen Versuchstieren um ca. 15% im Vergleich zur Kontrollgruppe
zu. Neun Wochen nach einmaliger Wärmeapplikation zeigten sich pro 50 Tubuli 199,6
Ki-67 positive Spermatogonien (110-302), zehn Tage nach zweiter Wärmeapplikation
239 (120-350) und 40 Tage nach zweiter Wärmeapplikation 239,8 (110-492).
Demnach scheint eine moderate Hyperthermie die Proliferation anzuregen. Spekulativ
bleibt, ob es unmittelbar im Anschluss an die Wärmeapplikation zu einem initialen
Abfall der Vermehrung von Spermatogonien kommt. Eine verstärkte Proliferation
sechs Wochen nach einem gesetzten Temperaturstimulus konnte auch bei Lamas
beobachtet werden (SCHWALM et al. 2007). Die verstärkt proliferierenden
Spermatogonien gleichen so die hauptsächlich geschädigten Spermatozyten aus und
bewirken eine Regeneration des Hodengewebes.
Nach Auszählung von 100 Sertolizellen zeigte sich ein anderes Ergebnis. Neun
Wochen nach einmaliger Wärmeapplikation war eine verminderte Proliferation (x̄ =
84,2; 56-120) zu beobachten, während die Kontrollgruppe (x̄ = 118,6; 62-176) eine
nahezu identische Anzahl Ki-67 positiver Spermatogonien wie die zehn Tage (x̄ =
111,6; 38-228) und 40 Tage nach zweiter Wärmeapplikation (x̄ = 119,7; 74-172)
kastrierten Tiere aufwies. Aufgrund der sehr variablen Anzahl an ausgezählter Tubuli
bei dieser Methode scheint es sich eher um einen Methodenfehler zu handeln. Es ist
davon auszugehen, dass die oben beschriebene verstärkte Proliferation der
Versuchsgruppen eher den durch den Temperaturstimulus gesetzte Effekt
widerspiegelt.

Diskussion
92
Nebenhoden In den Nebenhoden war bei allen wärmebehandelten Tieren die Anzahl der
Tubulusanschnitte mit aufgelockerten Epithelzellen um das Fünf- bis Siebenfache
gegenüber der Kontrollgruppe x̄ = 2,4 (1-5) erhöht. So zeigten die Tiere neun Wochen
nach einmaliger Wärmeapplikation im Durchschnitt 12,7 (1-25), zehn Tage nach
zweiter Wärmeapplikation 16,2 (1-25) und 40 Tage nach zweiter Wärmeapplikation
11,3 (0-25) Tubulusanschnitte mit vakuolig aufgelockertem Epithel. Demnach scheint
das Nebenhodenepithel sensibler auf einen Wärmestimulus zu reagieren als das
Hodentubulusepithel. Dabei kommt es offensichtlich innerhalb kurzer Zeit zur
Zellschädigung, welche dann über mehrere Wochen nachweisbar bleibt. Eine erhöhte
Anzahl vakuoliger Epithelzellen korrelierte auch im Bereich der Nebenhoden mit einer
vermehrten Anzahl von Tubulusanschnitten mit Zelldebris. Somit scheint der gesetzte
Wärmestimulus auch hier über eine vakuolige Degeneration zur Zellnekrose zu führen
(ELMORE 2007), oder eine vermehrte Anzahl apoptotischer Zellen war vorhanden
(YIN 1997), welche sich in der vermehrten Anzahl an zelldebrishaltigen
Tubulusanschnitten widerspiegelt. PERA et al. (1996) untersuchten die Auswirkung
einer Temperatur von 37°C auf spezische mRNA (CD52/CE5; codiert für ein
spezifisches spermienbindendes sekretorisches Protein) caniner
Nebenhodenepithelzellen in einer Zellkultur. Hierbei ließ sich nach fünf Tagen, im
Vergleich zur Kultur bei 33°C, keine spezifische mRNA mehr nachweisen. Auch bei
zwei unilateralen Kryptorchiden Hunden wurde im abdominalen Hoden kaum
spezifische mRNA nachgewiesen. Dies führt zu der Annahme, dass evt. noch im
Hoden gebildete Spermien nicht mehr im Nebenhoden überlebensfähig sind, da durch
den Temperaturstimulus die Proteinbildung der Tubulusepithelien gestört wurde. Auch
diese abgestorbenen Spermien können zu der vermehrten Bildung von Zelldebris
beitragen. 10 Tage nach zweiter Wärmeapplikation zeigten sich die meisten
zelldebrishaltigen Nebenhodentubulusanschnitte was die These der gestörten
Nebenhodenpassage stützt.
Insgesamt waren bei allen wärmebehandelten Tieren mehr spermienfreie
Tubulusanschnitte nachweisbar als bei den Kontrolltieren. Da jedoch keine
eindeutigen Hinweise auf eine Spermatogenesestörung in Form von verkleinerten

Diskussion
93
Keimzellen, geschrumpften Tubuli, Sertoli only, geschwollenen Spermatozyten und
verdickter Basalmembran in den Hodentubuli vorlagen, ist eine diesbezügliche
Ursache nicht eindeutig abzuleiten. Eine mögliche Ursache ist jedoch die, durch
apoptotische Prozesse, geringgradig verminderte Spermatogenese. VERONESI et al.
(2009) konnten bei keinem der zehn untersuchten kryptorchiden Rüden Spermien in
dem Nebenhoden des jeweils betroffenen Hodens nachweisen. In der Gruppe der 20
Kontrolltiere mit skrotalen Hoden zeigten insgesamt nur drei Nebenhoden diesen
Befund, sodass dieser Effekt durchaus als hyperthermiebedingte Veränderung zu
werten ist.
Apoptotische Zellen waren vor allem im Nebenhodenepithel nachweisbar. Es bestand
kein Unterschied zwischen Nebenhodenkopf, -körper und -schwanz. Neun Wochen
nach einmaliger Wärmeapplikation zeigte sich eine erhöhte Anzahl apoptotischer
Zellen sowohl mit dem TUNEL-Verfahren als auch dem Caspase-3-Nachweis. Mit
beiden Nachweisverfahren wurden zehn Tage nach der zweiten Hyperthermie mit
einer jeweiligen durchschnittlichen Erhöhung um das Achtfache im Vergleich zur
Kontrollgruppe die meisten positiven Zellen pro 25 ausgezählte Tubulusanschnitte
dargestellt. 40 Tage nach der zweiten Wärmeapplikation war eine Abnahme der
positiven Zellen zu verzeichnen. Auch im Bereich der Nebenhoden scheint es also
einen durch die Wärmeapplikation bedingten Effekt auf den Energiehaushalt der
Epithelzellen zu geben. Da die Auswirkung einer erhöhten Temperatur auf die
Apoptoserate des Nebenhodens bisher nicht an Hunden oder anderen Tierarten
untersucht wurde bleibt es zu vermuten, dass Epithel und Stroma getrennt von Lumen
betrachtet werden müssen. Im Lumen befinden sich die Spermien. Diese zeigten in
beiden Verfahren zur Bestimmung von Apoptosen eine erhöhte Rate, im Nachweis
aktivierter Caspase-3 noch deutlicher, als beim Nachweis von DNA-Brüchen. So
scheinen auch die Spermien im Nebenhoden nicht vor einer Hitzeeinwirkung
geschützt, was mit der These der gestörten Proteinsynthese im Nebenhodenepithel
korreliert (YIN 1997). Wärmeeinwirkung und andere Noxen, wie oxidativer Stress,
führen zu einer Fehlerhaften Proteinfaltung im Nebenhodenepithel (CORNWALL
2009). Diese fehlgefalteten Proteine können toxisch auf die Spermien einwirken.

Diskussion
94
SCHWEIZER (2006) bewirkte mittels moderaten skrotalen Hyperthermie über 48
Stunden an Hengsten eine verminderte mittlere Geschwindigkeit der Spermien. Diese
führt sie auf gestörte Entwicklungsprozesse während der Nebenhodenpassage
zurück. ALBRECHT (2006) postuliert ebenfalls eine Schädigung der Spermien
während der Nebenhodepassage bei Wärmeapplikation. Hier reduzierte sich die
Vorwärtsmotilität.
Auch das Nebenhodenstroma scheint von der Erwärmung betroffen zu sein, auch hier
konnte mit beiden Detektionsverfahren (TUNEL und Caspase-3-Nachweis) bei allen
wärmebehandelten Rüden mehr positive Zellen, wenn auch in geringer Anzahl, als bei
den Kontrolltieren (TUNEL-Verfahren: x̄ = 0,4 ± 0,6; Caspase-3-Nachweis: x̄ = 0)
identifiziert werden.
Die Proliferationsrate im Nebenhodenepithel neun Wochen nach einmaliger
Wärmeapplikation (= 5,8 ± 4,8) war um 36,7% geringer als in der Kontrollgruppe
(= 9,2 ± 5,8). Sowohl zehn (= 11,0 ± 8,3) als auch 40 Tage nach zweimaliger
Wärmeapplikation (= 11,3 ± 12,9) stieg die Proliferationsrate deutlich im Vergleich zu
neun Wochen nach einmaliger Wärmeapplikation an. Auch im Vergleich zur
Kontrollgruppe war ein geringer Anstieg zu verzeichnen. Das durch die
Wärmeapplikation geschädigte Epithel scheint also durch vermehrte Proliferation den
Effekt auszugleichen.
In den Nebenhodenlumen zeigte sich neun Wochen nach einmaliger
Wärmeapplikation (reduziert um 45,2%) sowie zehn Tage nach zweimaliger
Wärmeapplikation (reduziert um 58,2%) eine verminderte Proliferationsrate. 40 Tage
nach zweimaliger Wärmeapplikation war die Proliferationsrate lediglich um 19 %
reduziert. Fraglich bleibt, welche Zellen im Nebenhodenlumen proliferativen
Prozessen unterliegen können. Da hier hauptsächlich Spermien zu erwarten sind
kann eine Auszählung falsch positiver Zellen, welche eigentlich nicht mehr
proliferieren können, nicht ausgeschlossen werden.

Diskussion
95
Analog zu der Verteilung im Nebenhodenlumen zeigte sich auch im
Nebenhodenstroma eine im Vergleich zur Kontrollgruppe (= 3,2 ± 1,8) vermehrte
Proliferation zehn (= 4,2 ± 3,8) und 40 Tage nach zweimaliger Wärmeapplikation (=
3,8 ± 3,5), während neun Wochen nach einmaliger Wärmeapplikation (= 2,8 ± 1,6) die
Werte auf dem Niveau der Kontrollgruppe blieben. Auch die Zellen des
Nebenhodenstromas reagieren dementsprechend auf einen Hitzestimulus mit
kompensatorisch gesteigerter Proliferation.
5.5 Schlussfolgerungen
1. Die Hoden von Hunden scheinen im Vergleich zu den Gonaden von Hengsten und
Bullen weniger empfindlich auf einen moderaten Wärmestimulus zu reagieren.
2. Eine moderate Erwärmung der Hoden über 48 Stunden scheint keine irreversiblen
Schäden zu verursachen und demzufolge die Fruchtbarkeit nicht nachhaltig zu
beeinflussen.
3. Die Regeneration der durch skrotale Hyperthermie bedingten Schäden des
Hodengewebes ist nach einem Spermatogenesezyklus nicht vollständig
abgeschlossen.

Zusammenfassung
96
6 Zusammenfassung
Annika Herr: Effekte skrotaler Hyperthermie auf die Perfusion sowie auf histologische und immunhistochemische Merkmale des caninen Hodens und Nebenhodens
Ziel der vorliegenden Studie war es, die Empfindlichkeit des caninen Hodens und
Nebenhodens auf einen moderaten 48-stündigen Temperaturstimulus zu prüfen.
Hierzu wurden die Hoden von sieben Hunden mit Hilfe eines Suspensoriums,
bestehend aus einem äußeren Gehäuse aus thermoplastischem Kautschuk, einer
Babysocke und Watte, umgeben. Es wurde eine Temperaturerhöhung um
durchschnittlich 3,7°C auf 35,8°C in der ersten Hyperthermiephase sowie um 3,8°C
auf 35,9°C in der zweiten Hyperthermiephase erreicht.
Während einer vierwöchigen Kontrollperiode wurden zweimalig eine morphologische
Untersuchung sowie einmal wöchentlich eine farbkodierte dopplersonographische
Untersuchung (Messung der SPV, des RI und des PI) durchgeführt. Im Anschluss an
den Temperaturstimulus wurde die Untersuchungsfrequenz erhöht. Die Kontrolltiere
wurden nach vier Wochen kastriert. Bei 2 Tieren erfolgte die Kastration neun Wochen
nach einmaliger Wärmeapplikation, sowie bei 3 und 2 Rüden zehn bzw. 40 Tage nach
einer zweiten Wärmeapplikation. Drei weitere Hunde wurde als Kontrolltiere keinem
skrotalen Temperaturstimulus ausgesetzt. Bei diesen Tieren erfolgt eine Kastration
nach 4 Wochen.
Die Hoden und Nebenhoden wurden in drei Segmente (kranial, Mitte, kaudal)
unterteilt und in Formalin (4%) fixiert. Nach der histologischen Aufbereitung der
Organe in Paraffinblöcke wurden diese auf 2 μm Dicke geschnitten und für
histologische und immunhistochemische Untersuchungen gefärbt. Als Verfahren
kamen die HE-Färbung, das TUNEL-Verfahren, der Nachweis aktivierter Caspase-3
und der Nachweis des Ki-67 Antigens zur Anwendung.

Zusammenfassung
97
Unmittelbar nach Entfernung des Suspensoriums war über einen Zeitraum von
maximal 24 Stunden eine leichte Ödematisierung des Skrotums feststellbar. Einige
Hunde zeigten zusätzlich Rötung und Scheuerstellen. Alle morphologischen Hoden-
und Nebenhodenparameter wiesen in Bezug auf die Wärmeapplikation keine
Veränderungen auf.
Aufgrund der gemessenen physiologischen intra- und interindividuellen Variabilität
des Blutflusses mittels Dopplersonographie ließ sich keine dem Temperaturstimulus
zuzuordnende Veränderung nachweisen. Dies gilt sowohl für die SPV als auch für den
RI und PI.
In den HE-gefärbten Hodenpräparaten waren mit Ausnahme einzelner
Tubulusepithelzellen mit vakuoliger Degeneration und Hodentubuli mit Zelldebris
keine degenerativen Gewebeveränderungen mit Bezug zur skrotalen Hyperthermie
zu finden. Die vakuolige Degeneration stellt möglicherweise eine reversible
Schädigung des Keimepithels durch den Temperaturstimulus dar, welche aber auch
zur Nekrose einzelnen Zellen führen kann.
Die beiden zur Darstellung von apoptotischen Zellen verwendeten Verfahren, TUNEL
und Nachweis der aktivierten Caspase-3, lieferten unterschiedliche Ergebnisse mit
einer klaren Tendenz vermehrt positiver Zellen bei behandelten Tieren im Gegensatz
zur Kontrollgruppe. Auch die beiden verwendeten Auszählmethoden (pro 100
Sertolizellen und pro 50 Hodentubuli) lieferten nicht ganz übereinstimmende
Ergebnisse. Der Auswertung pro 50 Hodentubuli ist der Vorzug zu geben, da diese
genauer definierte Voraussetzungen bietet als die Auswertung pro 100 Sertolizellen.
Generell scheint es, dass ein einmaliger Temperaturstimulus neun Wochen nach
diesem immer noch eine vermehrte Anzahl apoptotischer Zellen hervorbringt, es aber
nach einem zweiten Stimulus eher zu nekrotischen Veränderungen im Keimepithel
des Hundehodens kommt. Auch im Nebenhodenbereich zeigten sich hauptsächlich
vakuolige Veränderungen, hier vor allem im Nebenhodentubulusepithel.

Zusammenfassung
98
Mithilfe der Ki-67-Methode konnte ebenfalls gezeigt werden, dass eine Auszählung
pro 50 Tubuli und pro 100 Sertolizellen unterschiedliche Ergebnisse liefert. So stieg in
der Auswertung pro 50 Tubuli die Proliferation kompensatorisch bei allen
wärmebehandelten Tieren im Gegensatz zur Kontrollgruppe an. Nach Auszählung von
100 Sertolizellen hingegen zeigte sich eine reduzierte Proliferation neun Wochen nach
einmaliger Wärmeappliaktion.
Zusammenfassend konnte festgestellt werden, dass die Hoden und Nebenhoden des
Hundes einen moderaten Temperaturstimulus während einer Periode von 48 Stunden
ohne makromorphologische Auffälligkeiten im Bereich der Hoden kompensieren. Es
kommt weder zu einer Schwellung noch zu von der Norm anweichenden Befunden
hinsichtlich Lage, Form und Konsistenz. Auf histologischer Ebene zeigten sich
reversiblen Veränderungen sowohl der Hoden, als auch der Nebenhoden.

Summary
99
7 Summary
Annika Herr: Effects of scrotal hyperthermia on the blood flow and on histological and immunohistochemical characteristics of the canine testis and epididymidis. The aim of the present study was to evaluate the effect of a mild hyperthermia
induction for a period of 48 hours on the canine testis and epididymidis. Therefore,
testes of seven dogs were heated using a jockstrap which resulted in an increase of
the temperature up to 35.8°C in the first period respectively up to 35.9°C in the second
period of hyperthermia.
For four weeks, all dogs underwent a control period in which ultrasound was
performed once a week. The parameters measured included systolic peak velocity
(SPV), resistance index (RI), and pulsatility index (PI). Morphological examinations of
the testes were performed twice within these four weeks. After this control period the
first hyperthermia of the testes was induced in the seven dogs. In the following nine
weeks ultrasound and morphological examinations were performed more frequently.
At the end of this period two of the seven dogs were castrated. The residual five dogs
underwent a second period of hyperthermia induction (48 hours) and were castrated
after ten (three dogs) and 40 days (two dogs), respectively.
Additionally, a control group of three dogs was castrated after four weeks without
undergoing any period of hyperthermia.
Immediately after castration the testes and epididymidis were dissected into three
pieces (cranial, midst, caudal) and fixed in formalin (4%). DNA fragmentation during
apoptosis was identified by TUNEL method and apoptotic cells were detected
immunohistochemically by activated caspase-3. Furthermore, H.E. staining was used
to identify signs of degeneration in the tissue. Proliferation was determined by the Ki-
67 method.

Summary
100
Immediately after removal of the jockstrap a mild scrotal edema was detected for a
time period of maximum 24 hours. Additionally, some dogs showed redness and
excoriation. Morphological parameters of testis and epididymidis did not reveal any
changes due to hyperthermia induction.
Based on the detection of physiological intra- and interindividual variability of blood
flow by dopplersonography a temperature-depending change in SPV, RI, and PI could
not be verified.
HE-staining of the testis revealed dead cell material and cells with vacuoles. All other
signs of degeneration were barely found. This leads to the assumption that those cells
underwent vacuolic degeneration which can finally end up in necrosis.
An increase of apoptotic cells was found in the testes of the seven dogs using TUNEL
and caspase-3. The highest amount of caspase-3 positive cells was found nine weeks
after the first period of hyperthermia, whereas more TUNEL-positive cells were found
ten and 40 days after the second period of hyperthermia. Thus, following the second
period of hyperthermia more cells died due to necrosis rather than apoptosis.
Furthermore, the counting of apoptotic cells per seminiferous tubules seemed to be
more reliable than counting per sertoli cells.
Using the Ki-67 method it could be shown that counting cells per 50 tubules
respectively per 100 sertoli cells revealed divergent results. Counting per 50 tubules
showed an increase in proliferating spermatogonia in all animals with induced
hyperthermia. After counting per 100 sertoli cells the control group and all animals
which underwent hyperthermia twice showed nearly identical results whereas the
dogs castrated nine weeks after one period of hyperthermia showed a decreased
proliferation.
The results of the present study demonstrate that dogs tolerate a mild heat stimulus
over a period of 48 hours without any changes in the scrotal morphology and
perfusion. In contrast to this, certain signs of histological changes including an

Summary
101
increase of apoptotic cells as well as a decrease of the proliferation rate were evident.
Those histological changes seem to be reversible.

Literaturverzeichnis
102
8 Literaturverzeichnis
ALBRECHT, P. N. (2006): Testikuläre Gewebetextur und Durchblutung sowie
Ejakulatbeschaffenheit von Bullen unter Einbeziehung der Analyse reaktiver
Sauerstoffspezies und des Spermienchromatins nach experimentaler skrotaler
Hyperthermie.
Hannover, Tierärztl. Hochsch., Diss.
BARR, F. (1992): Geschlechtsapparat. In: BARR, F. (Hrsg): Ultraschalldiagnostik bei
Hund und Katze.
Gustav Fischer Verlag, Jena, Stuttgart, New York 1992, S. 77-94.
BIAGIOTTI, G., G. CAVALLINI, F. MODENINI, G. VITALI u. L. GIANAROLI (2002):
Spermatogenesis and spectral echo-colour Doppler traces from the main testicular
artery.
BJU Int. 90, 903-908.
BILLIG, H., I. FURUTA, C. RIVIER, J. TAPANAINEN, M. PARVINEN u. A. J. HSUEH
(1995): Apoptosis in testis germ cells: developmental changes in gonadotropin
dependence and localization to selective tubule stages.
Endocrinology 136, 5-12.
CORNWALL, G. A. (2009): New insights into epididymal biology and function.
Hum Reprod Update. 15, 213-227
COULTER, G. H. u. J. P. KASTELIC (1994): Testicular thermoregulation in bulls.
in: Proceedings of the 15th Technical Conference on Artificial Insemination.
Nat. Assoc. Anim. Breed 1994; S. 28-34.

Literaturverzeichnis
103
DANSRANJAVIN, T. (2000): Untersuchung der Apoptose und der regulierenden
Faktoren FAS/FAS-Ligand, p 53 und bcl-2 in der normalen menschlichen
Spermatogenese und in der Genese von Keimzelltumoren.
Mathem.-Naturwissenschaftl. Diss., Martin-Luther-Universität Halle-Wittenberg
DAVIS, J. R. u. FIRLIT, C. F. (1966): The germinal epithelium of cryptorchid testes
experimentally induced in prepubertal and adult rats.
Fertil steril 17, 187-200
DEEG, K. H. u. F. WILD (1990): Abdominelle Dopplersonographie im Kindesalter.
Klin. Pädiatr. 202, 371-378.
ELMORE, S. (2007): Apoptosis: a review of programmed cell death.
Toxicol. Pathol. 35, 495-516.
EVANS, H. E., G. C. CHRISTENSEN (1993): The urogenital system.
in: EVANS, H. E. (Hrsg.): Miller´s anatomy of the dog.
Saunders, Philadelphia, S. 494-558.
FISCHER, U., R. U. JÄHNICKE u. K. SCHULZE-OSTHOFF (2003): Many cuts to ruin:
a comprehensive update of caspase subtrates.
Cell Death Diff. 10, 76-100.
FOOTE, R. H. u. E. E. SWIERSTRA (1972): Spermatogenesis in the dog.
Anat. Rec. 173, 341-352.
FORD, L. (1969): Testicular maturation in dogs.
Am. J. Vet. Res. 30, 331-336.

Literaturverzeichnis
104
GALIL, K. A. u. B. P. SETCHELL (1988): Effects of local heating of the testis on
testicular blood flow and testosterone secretion in the rat.
Int. J. Androl. 11, 73-85.
GASSE, H. (2004): Männliche Geschlechtsorgane, Organa genitalia masculina.
in: NICKEL, R., A. SCHUMMER u. E. SEIFERLE (Hrsg.): Lehrbuch der Anatomie der
Haustiere, 9. unveränderte Auflage, Band II Eingeweide.
Blackwell Wissenschafts-Verlag, S. 341-352.
GAVRIELI Y., Y. SHERMAN u. S. A. BEN-SASSON (1992): Indentification of
programmed call death in situ via specific labeling of nuclear DNA fragmentation.
Cell. Biol. 19, 493-501.
GIESE, W. (1997): Ultraschall.
in: GIESE, W. (Hrsg.): Kompendium der Physik für Veterinärmediziner.
Enke Verlag, Stuttgart, S. 89-99.
GLODE, L. M., J. ROBINSON u. L. D. HORWITZ (1984): Scrotal hypothermia and
testicular blood flow on the dog. Absence of thermal regulation.
J. Androl. 5, 227-229.
GOEDKEN, M. J.., R. L. KERLIN u. D. MORTON (2008): Spontaneous and age-
related testicular findings on beagle dogs.
Toxicol. Pathol. 36, 465-471.
GOSLING, R. G. u. D. H. KING (1974): Arterial assessment by Doppler shift
ultrasound.
Proc. R. Soc. Med. 67, 447-449.

Literaturverzeichnis
105
GÜNZEL-APEL, A. R. (1994): Der Zuchtrüde.
in: GÜNZEL-APEL, A. R. (Hrsg.): Fertilitätskontrolle und Samenübertragung beim
Hund.
Verlag Gustav Fischer, Jena, Stuttgart 1994, S. 11-48.
GÜNZEL-APEL, A. R., P. TERHAER u. D. WABERSKI (1994): Hodendimensionen
und Ejakulatbeschaffenheit fertiler Rüden unterschiedlicher Körpergewichte.
Kleintierpraxis 22, 483-486.
GÜNZEL-APEL, A. R., C. MÖHRKE u. C. POULSEN NAUTRUP (2001): Colour-coded
and pulsed Doppler sonography of the canine testis, epididymidis and prostate gland:
physiological and pathological findings.
Reprod. Domest. Anim. 36, 236-240.
GUMBSCH, P., C. GABLER u. A. HOLZMANN (2000): Colour-coded duplex
sonography of the testis of dogs.
Vet. Rec. 151, 140-144.
GUTZSCHEBAUCH, A. (1939): Der Hoden der Haussäugetiere und seine Hüllen in
biologischer und artdiagnostischer Hinsicht.
Zschr. Anat. Entw.gesch. 105, 434-458.
IBACH, B., L. WEISSBACH u. B. HILSCHER (1976): Stages of the cycle of the
seminiferous epithelium of the dog.
Andrologia 8, 297-307.
JACOBSON, M. D., M. WEIL u. M. C. RAFF (1997): Programmed cell death in animal
development.
Cell 88, 347-354.

Literaturverzeichnis
106
JÄNICKE, R. U., M. L. SPRENGART, M. R. WATI u. A. G. PORTER (1998):
Caspase – 3 is required for DNA fragmentation and morphological changes
associated with apoptosis.
J. Biol. Chem. 273, 9357-9360.
JOHNSTON, G. R., D. A. FEENEY, B. RIVERS u. P. A. WALTER (1991): Diagnostic
imaging of the male canine reproductive organs - Methods and limitations.
Vet Clin North Am Small Anim Pract. 21, 553-589.
KATAOKA, S. u. T. TSURUO (1996): Apoptosis.
The Oncologist 1, 399-401.
KAWAKAMI, E., T. HORI u. T. TSUTSUI (2000): Azoospermia of dogs with apoptotic
germ cells and leydig cells.
J. Vet. Med. Sci. 62, 529-531.
KERR, J. F. R. u. B. V. HARMIN (1991): Definition and incidence of apoptosis: an
historical perspective.
in: L.D. TOMEI u. F. O. COPE (Hrsg.): Apoptosis: The molecular Basis of Cell Death.
Cold Spring Harbor Laboratory Press, New York 1991, S. 5-29.
KHAN, V. R. u. I. R. BROWN (2002): The effect of hyperthermia on the induction of
cell death in brain, testis and thymus of the adult and developing rat.
Cell Stress and Chaperons 7, 73-90.
KORMANO, M. (1967): Development of the rectum-testis temperature difference in
the post-natal rat.
J. Reprod. Fertil. 14, 427-437.

Literaturverzeichnis
107
LIEBICH, H.-G. (2004): Männliche Geschlechtsorgane, Organa genitalia masculina.
in: LIEBICH, H.-G. (Hrgs.): Funktionelle Histologie der Haussägetiere. 3. Aufl.
Verlag Schattauer Stuttgart, New York 2004 , S. 271-284.
LOVE, C. C. u. R. M. KENNEY (1999): Scrotal heat stress induces altered sperm
chromatin structure associated with a decrease in protamine disulfide bonding in the
stallion.
Biol. Reprod. 60, 615-620.
LÜERSSEN, D. u. M. JANTHUR (2007): Skrotum, Hoden und Nebenhoden.
in: POULSEN NAUTRUP, C. u. R. TOBIAS (Hrsg.): Atlas und Lehrbuch der
Ultraschalldiagnostik bei Hund und Katze. 4. Aufl.,
Schlütersche Verlagsanstalt Hannover 2007, S. 273-286.
MAJNO, G. u. I. JORIS (1995): Apoptosis, oncosis and necrosis. An overview of cell
death.
Am. J. Pathol. 146, 3-15.
MARTINVALET, D., P. ZHU u. J. LIEBERMANN (2005): Granzyme A induces
caspase-independent mitochondrial damage, a required first step for apoptosis.
Immunity 22, 355-370.
MONTKOWSKI, A. (1992): Lichtmikroskopische, ultrastrukturelle und glykohisto-
chemische Untersuchungen am Hoden des Hundes (canis familiaris).
München, Ludwig-Maximilian Universität, Diss.
MOORE, C. R. (1924): Properties of the gonads as controllers of somatic and
psychical characteristics: VI. Testicular reactions in experimental cryptorchidism.
Am J Anat. 34, 269-316

Literaturverzeichnis
108
NAGATA, S. (1997): Apoptosis by death factor.
Cell 88, 355-365.
NEUERBURG-HEUSLER, D. u. M. HENNERICI (1999): Ultraschallverfahren.
in: NEUERBURG, D. u. M. HENNERICI (Hrsg.): Gefäßdiagnostik mit Ultraschall.
2. Aufl., Verlag Georg Thieme, Stuttgart 1999, S. 1-23.
PERA, I., R. IVELL u. C. KIRCHHOFF (1996): Body temperature (37) specifically
down-regulates the messenger ribonucleic acid for the major sperm surface antigen
CD52 in epididymal cell culture.
Endocrinology 137, 4451-4459
PINGGERA, G. M., M. MITTERBERGER, G. BARTSCH, H. STRASSER, J. GRADLE,
F. AIGNER, L. PALLWEIN u. F. FRAUSCHER (2008): Assessment of the
intratesticular resistive index by colour Doppler ultrasonography measurements as a
predictor of spermatogenesis.
BJU Int. 101, 722-726.
POULSEN NAUTRUP, C. (2007): Allgemeiner Teil.
in: POULSEN NAUTRUP, C. u. R. TOBIAS (Hrsg.): Atlas und Lehrbuch der
Ultraschalldiagnostik bei Hund und Katze. 4. Aufl.
Schlütersche Verlagsanstalt Hannover 2007, S. 18-82.
POURCELOT, L. (1974): Applications cliniques de l’examen Doppler transcutane.
in: PERONNEAU, P. (Hrsg.): Vlocimetric ultrasonore Doppler.
INSERM 34, 213-240.
PRÜFER, A., D. LÜERSSEN u. M. JANTHUR (2007): Prostata.
in: POULSEN NAUTRUP, C. u. R. TOBIAS (Hrsg.): Atlas und Lehrbuch der
Ultraschalldiagnostik bei Hund und Katze. 4. Aufl.
Schlütersche Verlagsanstalt, Hannover 2007, S. 282-289.

Literaturverzeichnis
109
PUGH, C. R., L. J. KONDE u. R. D. PARK (1990): Testicular ultrasound in the normal
dog.
Vet. Radio. 31, 195-199.
REHM, S. (2000): Spontaneous testicular lesions in purpose-bred beagle dogs.
Toxicol. Pathol. 28, 782-787.
RUEL, Y., P. Y. BARTHEZ, A. MAILLES u. D. BEGON (1998): Ultrasonographic
evaluation of the prostate in healthy intact dogs.
Vet. Radiol. 39, 212-216.
ROCKETT, J. C., F. L.MAPP, J. B. GARGES, J. C. LUFT, C. MORI u. D. J. DIX
(2001): Effects of hyperthermia on spermatogenesis, apoptosis, gene expression and
fertility in adult male mice.
Biol. Reprod. 1, 229-239.
SANDERSLEBEN v., J., K. DÄMMRICH u. E. DAHME (1989): Die Reaktion der Zelle.
in: SANDERSLEBEN v., J., K. DÄMMRICH u. E. DAHME (Hrsg.): Pathologische
Histologie der Haussäugetiere. 3. Aufl.
Gustav Fischer Verlag, Stuttgart, New York 1989, S. 15-18
SCHEIDEL, P. (2000): Apoptose? Welche Bedeutung hat sie beim Auftreten von
Autoimmun- und Tumorerkrankungen?
Frauenheilkunde Aktuell 9/3, 35-36
SCHNORR, B. u. M. KRESSIN (2001): Entwicklung und Bau der Samenzellen.
in: B. SCHNORR u. M. KRESSIN (Hrsg.): Embryologie der Haustiere. 3. Aufl.
Enke Verlag Stuttgart 2001, S 3-11.

Literaturverzeichnis
110
SCHOLZEN, T. u. J. GERDES (2000): The Ki-67 protein: from the known and the
unknown.
J. Cell Physiol. 182, 311-322.
SCHULTZ D. R. u. W. J. jr. HARRINGTON (2003): Apoptosis: programmed cell death
at a molecular level.
Semin. Arthritis Rheum. 32, 346-369.
SCHWALM, A., M. GANLY, G. ERHARDT u. M. BERGMANN (2007): Changes in
testicular histology and sperm quality in llamas (lama glama), following exposure to
high ambient temperature.
Theriogenology 67, 1316-1323.
SCHWEIZER, C. (2006): Diagnostik hyperthermiebedingter Veränderungen der
Samenparameter und des Hodengewebes beim Hengst mit Hilfe durchfluss-
zytometrischer Sperma-Assays und computergestützter Echostrukturanalyse
Hannover, Tierärztl. Hochsch., Diss.
SETCHELL, B. P. (1978a): Vascular supply and drainage of the testis.
in: B. P. Setchell (Hrsg.): The mammalian testis.
ELEK BOOKS LIMITED London 1978, S. 50-69.
SETCHELL, B. P. (1978b): The scrotum and thermoregulation.
in: B. P. Setchell (Hrsg.): The mammalian testis
ELEK BOOKS LIMITED London 1978, S. 90-103.
SETCHELL, B. P. (2006): The effects of heat on the testes of mammals.
Anim. Reprod. 3, 81-91

Literaturverzeichnis
111
SEYREK-INTAS, D., M. GERWING u. M. KRAMER (2010): Sonographie der Prostata
und Hoden; Lage, Standardschnittebenen, Normalanatomie, Pathologie.
Prakt. Tierarzt 91, 30-34.
SINHA HIKIM, A. P., Y. LUE, M. DIAZ-ROMERO, P.H. YEN, C. WANG u. R. S.
SCHWERDLOFF (2003): Deciphering the pathways of germ cell apoptosis in the
testis.
J. Steroid Biochem. Mol. Biol. 85, 175-182.
SINOWATZ, F. (2001): Morphologie und Histologie der männlichen Genitalorgane.
in: BUSCH, W. u. A. HOLZMANN (Hrsg.): Veterinärmedizinische Andrologie.
Verlag Schattauer, Stuttgart 2001, S. 1-18
SOARES, J. M., G. F. AVELAR u. L. R. FRANCA (2009): The seminiferous epithelium
cycle and its duration in different breeds of dog (Canis familiaris).
J. Anat. 215, 462-471.
SOD-MORIAH, U. A., G. M. GOLDBERG u. E. BEDRAK (1974): Intrascrotal
temperature, testicular histology and fertility of heat-acclimatized rats.
J. Reprod. Fertil. 37, 263-268.
STUART, B., J. DRUMM, D. E. FITZGERALD u. N. M. DUIGNAN (1980): Fetal blood
velocity waveforms in normal pregnancy.
Br. J. Obstet. Gynaecol. 87, 780-785.
TAYLOR, K. J. W. u. S. HOLLAND (1990): Doppler US. Part I. Basic principles,
instrumentation and pitfalls.
Radiol. 174, 297-307.

Literaturverzeichnis
112
TROIANO, L., M. F. FUSTINI, E. LOVATO, A. FRASOLDATI, W. MOLORNI, M.
CAPRI, E. GRASSILLI, P. MARRAMA u. C. FRANCESCHI (1994): Apoptosis and
spermatogenesis: evidence from an in vivo model of testosterone withdrawal in the
adult rat.
Biochem. Biosphys. Res. Commun. 202, 1315-1321.
VERONESI, M. C., E. RICCARDI, A. ROTA u. V. CRIECO (2009): Characteristics of
cryptic/ectopic and contralateral scrotal testes in dogs between 1 and 2 years of age.
Theriogenology 72, 969-977.
VOGLER, C. J., J. H. BAME, J. M. DEJARNETTE, M. L. MCGILLIARD u. R. G.
SAACKE (1993): Effects of the elevated testicular temperature on morphology
characteristics of ejaculated spermatozoa in the bovine
Theriogenology 40, 1207-1219.
WEISS, E. u. I. KÄUFER-WEISS (2007): Männliche Geschlechtsorgane.
in: DAHME, E. u. E. WEISS (Hrsg.): Grundriss der speziellen pathologischen
Anatomie der Haustiere. 6. Aufl.
Enke Verlag Stuttgart, 2007, S. 280-292.
WROBEL, K.-H., R. KUJAT u. R. LUTZ (1993): Expression of the proliferation-
associated Ki-67 antigen in bovine testicular tubular cells during the seminiferous
epithelial cycle, demonstrated with the MIB-1 antibody.
Andrologia 25, 301-305.
WU, C. T. (2005): Durchflusszytometrische Verfahren zur Beurteilung der
Spermaqualität nach einer experimentell induzierten Hyperthermie am Bullenhoden.
München, Ludwig-Maximilian Universität, Diss.

Literaturverzeichnis
113
YIN Y., HWAKINS, K. L., DEWOLF, W. C. u. MORGENTALER, A. (1997): Heat stress
causes testicular germ cell apoptosis in adult mice.
J Androl. 18, 159-165

Anhang
114
9 Anhang
9.1 Histologische und immunhistochemische Verfahren –
Arbeitsprotokolle
4% gepuffertes Formalin: Ansatz für 2 x 2,5 Liter:
4250 ml Aqua dest.
25,95 g Na2HPO4 2 H2O
20,00 g NaH2PO4 H2O
50 ml 1 M NaOH
550 ml 37% Formalin
ins Aqua dest. beide Natriumphosphate und die Natronlauge geben, die Phosphate
müssen durch Rühren aufgelöst werden, dann das Formalin zugeben, mischen und
den pH-Wert überprüfen (in der Regel stimmt 7).
Entwässerung im Gewebeeinbettungsautomaten (Hypercenter II, Shandon, Frankfurt) Nachfixierung für 1 Stunde in 10%igem Formalin, spülen mit Leitungswasser,
dehydrieren in aufsteigender Alkoholreihe (70%, 85%, 96%), entfernen des Alkohols
mit Isopropanol und Essigsäure-n-Butylester
Hämalaun-Eosin (HE)-Färbung Das Entparaffinisieren der Schnitte erfolgt in einer absteigenden Alkoholreihe,
beginnend mit
Xylol (3x je) 5 Min., Isopropanol 5 Min., dann 96%iges, 70%iges und 50%iges
Ethanol je 2-3 Min., dann Aqua dest., anschließend erfolgt die Färbung
15 Min. in Hämalaun nach P. Mayer,
10 Min. bläuen in Leitungswasser und in Aqua dest. spülen,
1 Min. in 1%igem Eosin nach Zugabe von 1 Tropfen Eisessig pro Küvette und wieder
in Aqua dest. spülen.

Anhang
115
Darauf folgt ein Entwässern in einer aufsteigenden Alkoholreihe (70% - 96%ige 2-3
min), Isopropanol 5 Min., Xylol (3x je) 5 Min.
Abschließend erfolgt das Eindecken in Roti-Histokitt (Eindeckmittel Rotihisto-Kitt, Fa.
Roth C. GmbH).
Lösungen für die HE-Färbung Hämalaun nach P. Mayer:
1g Hämatoxylin in 100 ml Aqua dest. lösen,
0,2 g Natriumjodat,
50 g chemisch reines Kalialaun (Kaliumaluminiumsulfat-12-sulfat) dazugeben und
erwärmen;
nach dem Erkalten: 50 g Chloralhydrat und 1 g kristalline Zitronensäure dazugeben,
kalt lösen; filtrieren.
Eosin:
0,1 % in Aqua dest. lösen;
Nach dem Erkalten filtrieren;
Zum Färben auf 100 ml 1% iges Eosin 2 Tropfen Eisessig geben.
Durchführung der TUNEL-Reaktion Der Nachweis TUNEL-positiver Zellen erfolgte mittels ApopTag®Plus (Firma
Appligene Oncur, Heidelberg)
1. Entparaffinisieren der Schnitte in einer absteigenden Alkoholreihe von Roticlear
(2x je 10min, bei Raumtemperatur) über Isopropanol, 96%iges, 70%iges,
50%iges Ethanol, je 5 Minuten bei Raumtemperatur
2. Einmaliges Spülen der Schnitte in PBS1 für je 5 Minuten bei Raumtemperatur
unter ständigem Rühren,
Ansetzen der Endverdünnung von Proteinase K (20µg/ml)2

Anhang
116
3. Objekträger abtrocknen und Schnitte mit Fettstift umranden; bei vielen
Schnitten diese mit PBS abdecken oder in feuchter Kammer lagern, bis alle
Schnitte umrandet sind
4. Unter Umständen zuerst PBS von den Schnitten abschütteln, andauen mit
Proteinase K-Lösung (20µg/ml) für 10 Minuten bei Raumtemperatur in einer
feuchten Kammer
Enzym-Substrat-Lösung (Working-Strength-TdT-Solution)3 ansetzen und bis
zum Gebrauch auf Eis lagern, Stop-Wash-Puffer4 in einer Küvette ansetzen
5. Herausnehmen der Schnitte aus der feuchten Kammer und Umsetzen in eine
Küvette. Viermaliges Spülen in Aqua dest. für je 2 min bei Raumtemperatur
unter ständigem Rühren
6. Hemmung der endogenen Peroxidasen mit 2% H2O2 in PBS für 5 Minuten bei
Raumtemperatur unter ständigem Rühren,
7. zweimaliges Spülen der Schnitte in PBS für je 5 Minuten bei Raumtemperatur
unter ständigem Rühren.
8. Abtrocknen der Objektträger und Verbringen der Schnitte in eine feuchte
Kammer. Auftragen von 75µl Equilibritation Puffer/Schnitt für 15 Sekunden-30
Minuten (10 Minuten) bei Raumtemperatur
9. Überschuss an Equilibration Puffer abschütteln und Objekträger vorsichtig
abtrocknen, umbetten in eine vorgewärmte feuchte Kammer
54µl Working-Strength-TdT-Solution/Schnitt auftragen und mit CoverSlip
abdecken, Inkubation für 60 Minuten bei 37°C im Brutschrank
Küvette mit Stop-Wash-Puffer ins Wasserbad setzen und vorwärmen
10. Entfernen der CoverSlips (Lagerung in A.dest.)
Schnitte in die vorgewärmte Küvette mit Stop-Wash-Puffer stellen, im
Wasserbad für 30 Minuten bei 37°C unter ständiger Bewegung inkubieren
11. Dreimaliges spülen der Schnitte in PBS für je 5min bei Raumtemperatur unter
ständigem Rühren; Reinigung der CoverSlips für Schritt 12
12. Abtrocknen der Objektträger und Verbringen der Schnitte in eine feuchte
Kammer. Auftragen von 55µl Anti-Digoxigenin-Peroxidase/Schnitt für 30
Minuten bei Raumtemperatur unter Abdeckung mit CoverSlips

Anhang
117
13. Herausnehmen der Schnitte aus der feuchten Kammer und Umsetzen in eine
Küvette; viermaliges Spülen der Schnitte in PBS für je 2 Minuten bei
Raumtemperatur unter ständigem Rühren
14. Inkubation mit der DAB-Gebrauchslösung5 in einer Küvette für 5 Minuten bei
Raumtemperatur unter ständigem Rühren
15. Dreimaliges Spülen der Schnitte in PBS für je 5 Minuten bei Raumtemperatur
unter ständigem Rühren
16. Kurzes Eintauchen der Schnitte in Aqua dest.
17. Gegenfärben in Hämalaun nach Meyer für 5 Minuten bis 15 Sekunden bei
Raumtemperatur
18. Bläuen der Schnitte in Leitungswasser für 5 Minuten bei Raumtemperatur
19. Entwässern der Schnitte in einer aufsteigenden Alkoholreihe von Ethanol
50%ig, 70%ig und 96%ig bis zum Isopropanol (je 1 Minute bei
Raumtemperatur), anschließend 2x in Xylol für je 5 Minuten bei
Raumtemperatur
20. Eindecken der Schnitte
1PBS: 40g NaCl und 8,8g NaH2PO4 auf 5 ml Aqua dest.; pH auf 7,0-7,1 einstellen 2Proteinase K-Lösung: 4µl Stammlösung und 1996 µl Verdaupuffer = 2ml Proteinase
K-Lösung, für ca. 20 Schnitte
Proteinase K-Stammlösung (10µg/µl bzw. 10mg/ml): 100mg Proteinase K mit 2ml
Tris-HCl (1M, pH 8,0) sowie 20µl CaCl2 (0,1 M) ad 10ml DEPC-H2O steril ansetzen
und bei -20°C lagern
Verdaupuffer: 5ml Tris-HCl (1M, pH 8,0), EDTA (0,5M, pH 8,0), Tween 20 ad 100ml
DEPC-H2O, autoklavieren und bei 4°C lagern 3Enzym-Subtrat-Lösung (Working-Strength-TdT-Solution): 38µl Reaktions-Puffer und
16µl TdT-Enzyme = 54µl/Schnitt
760µl Reaktionspuffer und 320µl TdT-Enzyme = 1080µl/20Schnitte 4Stop-Wash-Puffer: 1ml Stop-Wash-Konzentrat und 34ml Aqua dest. = 35ml Stop-
Wash-Puffer
5ml Stop-Wash-Konzentrat und 170ml Aqua dest. = 175ml Stop-Wash-Puffer

Anhang
118
5DAB: Diaminobenzidin-Tetrahydrochloruidihydrat
Stammlösung: 100mg 3,3´DAB (Fluka, Schweiz) in 50ml PBS-Puffer unter
Lichtausschluss lösen; Lösung 20 Minuten rühren und bei -2°C lagern
Gebrauchslösung: 50ml DAB-Stammlösung in 150ml PBS (pH 7,2) lösen und durch
zwei Filterpapiere filtrieren, anschließend Zugabe von 2ml 3% H2O2-Lösung
Endkonzentration 0,05% DAB
Durchführung der Caspase-Methode Nachweis von Caspase-positiven Zellen erfolgt mittels der ABC-Methode
1. 2μm dicke Schnitte auf Superfrost® Plus Objektträger (Menzel-Gläser GmbH &
Co KG) aufziehen, anschließend Deparaffinisierung und Rehydration mittels
Xylol 3x 5 Minuten; Isopropanol 5 Minuten; Ethanol 96% 3 Minuten
2. Hemmung der endogenen Peroxidase für 30 Minuten in Methanol mit 0,5%
H2O2 3ml Perhydrol und 197ml 85% Methanol
3. 3x je 5 Minuten spülen in PBS1, dabei langsam rühren
4. Demarkierung mittels Erwärmung über 20 Minuten in Citratpuffer2 (in der
Mikrowelle)
5. Wiederholtes dreimaliges Spülen in PBS, dabei langsam rühren
6. Verbringen der Schnitte in Coverplates (Thermo Electron GmbH), überschichtet
mit inaktivem Ziegen Normalserum in PBS als Blockserum. Inkubation 20
Minuten bei Raumtemperatur
7. 1. Antikörper: in Coverplates Caspase 3 1:200 in PBS mit 1% BSA; jeweils eine
negativ Kontrolle mitlaufen lassen. Inkubation über Nacht bei 4°C im
Kühlschrank
8. 3x 5 Minuten spülen in PBS
9. 2.Antikörper in Coverplates: überschichten mit GAR-b (Goat-anti-Rabbit) 1:200
in PBS, Vector BA-1000; Inkubation 30 Minuten bei Raumtemperatur
10. 3x 5 Minuten spülen in PBS
11. ABC: in Coverplates. ABC-Reagenz (Vector Laboratories Inc.) einfüllen;
Ansatz: 1ml PBS vorlegen, 15μl Reagenz A zufügen und gut schütteln, dann

Anhang
119
15μl Reagenz B dazu pipettieren und schütteln; 30 Minuten vor Gebrauch
ansetzen; Inkubation 20 Minuten bei Raumtemperatur
12. 3x 5 Minuten spülen in PBS
13. DAB3: Enzymhistochemische Reaktion: 0,1g DAB auf 200ml PBS und 2ml 3%
H2O2
14. Spülen in PBS und Verbringen in Leitungswasser, insgesamt 15 Minuten
15. Schwache Gegenfärbung, etwa 20 Sekunden bis 5 Minuten in Hämalaun nach
Mayer; anschließend 10-15 Minuten bläuen unter fließendem Leitungswasser
16. Aufsteigende Alkoholreihe bis Essigsäure-N-Butylester. 50%-96% Ethanol je 2-
3 Minuten
Isopropanol 5 Minuten
Xylol 1-2 Minuten
Anschließend eindecken
1PBS: 40g NaCl und 8,8g NaH2PO4 auf 5ml Aqua dest.; pH auf 7,0-7,1 einstellen 2Citratpuffer: 2,1g Citronensäuremonohydrat ad 1000ml Aqua dest., pH-Wert mit 1N
NaOH auf 6,0 einstellen 3DAB: Diaminobenzidin-Tetrahydrochlordihydrat
Stammlösung: 100mg 3,3´DAB (Fluka, Schweiz) in 50ml PBS-Puffer unter
Lichtausschluss lösen; Lösung 20 Minuten rühren und bei -2°C lagern
Gebrauchslösung: 50ml DAB-Stammlösung in 150ml PBS (pH 7,2) lösen und
durch zwei Filterpapiere filtrieren, anschließend Zugabe von 2ml 3% H2O2-
Lösung
Endkonzentration 0,05% DAB

Anhang
120
Durchführung der Ki-67-Reaktion Nachweis von Ki-67 positiven Zellen erfolgt ebenfalls mittels der ABC-Methode
1. 2μm dicke Schnitte auf Superfrost® Plus Objektträger (Menzel-Gläser GmbH &
Co KG) aufziehen, anschließend Deparaffinisierung und Rehydration mittels
Xylol 3x 5 Minuten; Isopropanol 5 Minuten; Ethanol 96% 3 Minuten
2. Hemmung der endogenen Peroxidase für 30 Minuten in Methanol mit 0,5%
H2O2 3ml Perhydrol und 197ml 85% Methanol
3. 3x je 5 Minuten spülen in PBS1, dabei langsam rühren
4. Demarkierung mittels Erwärmung über 20min in Citratpuffer2 (in der Mikrowelle)
5. Wiederholtes dreimaliges Spülen in PBS, dabei langsam rühren
6. Verbringen der Schnitte in Coverplates (Thermo Electron GmbH), überschichtet
mit inaktivem Ziegen Normalserum in PBS als Blockserum. Inkubation 20
Minuten bei Raumtemperatur
7. 1. Antikörper: in Coverplates Mouse-Anti-Human-Ki 67-Antigen, Klon MIB-1 (
Dako Diagnostika, Hamburg) 1:100 in PBS mit 1% BSA; jeweils eine negativ
Kontrolle mitlaufen lassen. Inkubation über Nacht bei 4°C im Kühlschrank
8. 3x 5 Minuten spülen in PBS
9. 2.Antikörper in Coverplates: überschichten mit GAM-b (Goat-anti-Mouse) 1:200
in PBS, Vector BA-9000; Inkubation 30 Minuten bei Raumtemperatur
10. 3x 5 Minuten spülen in PBS
11. ABC: in Coverplates. ABC-Reagenz (Vector Laboratories Inc.) einfüllen;
Ansatz: 1ml PBS vorlegen, 15μl Reagenz A zufügen und gut schütteln, dann
15μl Reagenz B dazu pipettieren und schütteln; 30 Minuten vor Gebrauch
ansetzen; Inkubation 20 Minuten bei Raumtemperatur
12. 3x 5 Minuten spülen in PBS
13. DAB3: Enzymhistochemische Reaktion: 0,1g DAB auf 200ml PBS und 2ml 3%
H2O2
14. Spülen in PBS und verbringen in Leitungswasser, insgesamt 15 Minuten
15. Schwache Gegenfärbung, etwa 20 Sekunden bis 5 Minuten in Hämalaun nach
Mayer; anschließend 10-15min bläuen unter fließendem Leitungswasser
16. Aufsteigende Alkoholreihe bis Essigsäure N Butylester.

Anhang
121
50%-96% Ethanol je 2-3 Minuten
Isopropanol 5 Minuten
Xylol 1-2 Minuten
Anschließend eindecken
1PBS: 40g NaCl und 8,8g NaH2PO4 auf 5ml Aqua dest.; pH auf 7,0-7,1 einstellen 2Citratpuffer: 2,1g Citronensäuremonohydrat ad 1000ml Aqua dest., pH-Wert mit 1N
NaOH auf 6,0 einstellen 3DAB: Diaminobenzidin-Tetrahydrochlordihydrat
Stammlösung: 100mg 3,3´DAB (Fluka, Schweiz) in 50ml PBS-Puffer unter
Lichtausschluss lösen; Lösung 20 Minuten rühren und bei -2°C lagern
Gebrauchslösung: 50ml DAB-Stammlösung in 150ml PBS (pH 7,2) lösen und durch
zwei Filterpapiere filtrieren, anschließend Zugabe von 2ml 3% H2O2-Lösung
Endkonzentration 0,05% DAB

Anhang
122
9.2 Skrotale Wärmeapplikation und Aufbereitung der Hoden:
Materialien, Chemikalien und Geräte
Bezugsquelle Material
Dr. Fritz Buschhaus ETM (Engineering Technologie Marketing GmBH Schönbrunn 180, 07929 Saalburg-Ebersdors
Thermoplastischer Kautschuk, Verschnitt aus
EPDM (Ethylen-Propylen-Dien-Kautschuk)
und PP Polyprolylen
KiK Textilien und Non-Food GmbH Siemensstraße 21, 59199 Bönen
Babysöckchen: 60% Baumwolle, 20% Nylon,
19% Polyester, 1% Elastane-Lycra
Tierärztebedarf J. Lenecke GmbH & Co.KG Mentestraße 12a, 26419 Schortens
Halskragen Buster, clic collar 25cm
Vaseline Favodent
Handschuhe Nobaglove®-Latex
Ultraschallgel
Rolta Polsterwatte
Yatego GmbH Technologiezentrum Leopoldstr. 1 D-78112 St. Georgen
TFA Klima Logger
Thermo-Hygro-Station
Kat.-Nr. 30.3015
Wirtschaftgenossenschaft deutscher Tierärzte eG Siemensstraße 14, 30827 Garbsen
Brustgeschirr

Anhang
123
Bezugsquelle Material A. Krüss Optronic GmbH, Hamburg
Binocular Mikroskop, MBL 2000
Bauknecht Hausgeräte Mikrowelle, MWS 2924
Carl Roth GmbH Karlsruhe
Citronensäure-Monohydrat, 3958.1
Chloralhydrat
Essigsäure-n-butylester, 4600.4
Hämalaunlösung nach Mayer, T865.1
Hämatoxylin, 3816.2
Natriumjodat p.a.
Roti®-Histokit, 6638.1
Roticlear®, A538.3
Cell Signaling Danvers, USA
Caspase 3-Antikörper, polyclonal Kaninchen-
anti-Caspase 3, 9661
Chemicon® International Ltd. Hampshire, UK
ApopTag®Plus Peroxidase In Situ Apoptosis
Detection Kit, S7101
Engelbrecht Edermünde
Chromalaun-Gelantine beschichtete
Objekträger
Fluka Chemika Buchs, Schweiz
Diaminobenzidin-Tetrahydrochloriddihydrat,
DAB, 32750
Gesellschaft für Labortechnik Burgwedel
Warmwasserbad für paraffinierte
Gewebeschnitte
Heraeus GmbH Hanau
Wärmeschrank 5042
IKA-Werke GmbH & Co.KG, Staufen
Magnetrührer, Typ REO
Leica Microsystems Nussloch GmbH Nussloch
Färbegerät Leica, ST 4040

Anhang
124
Medite Medizintechnik Burgdorf
Objekträger-Eindeckautomat Promounter,
RCM 2000
Menzel-Gläser, Glasaufbereitungswerke GmbH & Co KG Braunschweig
Superfrost®Plus Objekträger, 041300
Merck KGaA Darmstadt
Wasserstoffperoxid 30% H2O2 (Perhydrol®),
107209
Microm International GmbH Walldorf
Rotationsmikrotom
Quartett Immundiagnostica und Biotechnologie GmbH Berlin
Citratpuffer-Konzentrat, pH 6,0
Sigma-Aldrich Chemie GmbH Taufkirchen
Bovines Serumalbumin (BSA), A-3059
3,3´-Diaminobenzidin.tetrahydrochlorid Dihydrat
purum (DAB), p.a., 32750
Kaninchenserum, PK 4505
Thermo Electron GmbH Dreieich
Shandon CoverplatesTM, 72110013
Shandon Sequnenza®-Slide racks, 7331017
Paraplast Plus®
Vector Laboratories Inc. Burlingame, CA, USA Über Biologo Kronshagen
ABC-Reagenz, PK 6100
W. Knittel Glasbearbeitungs GmbH Braunschweig
Deckgläser 24x50mm

Anhang
125
9.3 Verläufe der Skrotaltemperatur
1. Hyperthermie n=7
Stunde 4 5 6 7 8 9 10 1 35,0 35,6 35,5 36,1 35,1 35,9 35,7 2 35,0 35,0 37,2 35,7 35,0 35,2 35,1 3 35,7 34,8 36,6 35,0 35,6 35,2 35,1 4 36,8 35,1 36,1 35,3 35,7 35,4 36,1 5 35,6 35,5 35,8 35,7 35,0 36,0 35,6 6 35,1 35,5 35,6 35,6 34,9 36,6 35,1 7 35,6 35,3 35,6 35,7 35,2 36,1 35,2 8 35,7 35,7 35,6 35,0 35,6 35,4 35,7 9 34,9 35,7 35,0 34,9 36,6 35,3 35,2 10 34,9 35,0 36,2 35,3 36,8 35,4 35,8 11 35,4 35,7 36,1 35,9 36,3 35,4 36,5 12 35,5 36,0 35,5 35,5 36,3 35,2 36,0 13 35,4 35,0 35,7 35,3 36,7 35,3 35,9 14 35,8 34,9 36,1 35,2 36,3 35,1 35,1 15 36,0 34,8 35,9 34,9 36,6 35,1 35,3 16 35,7 35,4 35,4 35,6 36,6 35,3 35,1 17 35,4 34,9 35,8 35,6 36,6 35,8 35,0 18 34,8 36,0 36,0 35,2 35,7 35,6 34,9 19 35,6 35,5 35,6 35,4 35,3 35,0 35,2 20 35,8 34,9 35,6 35,5 35,1 36,2 35,0 21 35,7 35,4 35,7 35,6 35,3 36,7 35,1 22 35,1 35,6 34,8 35,8 35,8 36,1 35,6 23 35,6 35,2 35,3 36,4 36,6 35,9 35,8 24 35,0 35,1 34,9 36,0 36,0 35,4 35,6 25 35,4 35,3 35,4 35,6 35,4 35,7 35,2 26 34,8 35,6 35,6 35,3 35,5 36,6 35,2 27 35,3 35,7 35,7 35,0 36,7 35,5 35,6 28 35,1 36,0 35,1 35,7 37,0 35,6 35,3 29 35,0 35,5 35,3 36,0 37,3 35,3 34,9 30 35,7 35,5 35,4 36,1 37,2 36,1 36,5 31 35,2 35,9 35,7 36,4 37,2 36,2 35,7 32 35,8 35,5 34,9 35,5 36,7 36,7 35,1 33 34,8 35,6 34,9 36,1 36,9 36,5 35,1 34 35,0 35,8 35,8 36,0 37,4 36,1 36,2 35 34,9 35,7 36,0 35,6 37,3 35,4 35,2 36 35,5 35,9 36,1 35,5 35,4 35,5 35,1 37 36,0 36,6 36,4 36,2 36,0 35,2 35,6 38 34,8 37,9 35,7 35,9 35,5 34,9 35,1 39 35,4 39,4 35,8 35,6 35,1 35,4 35,1 40 35,4 39,3 35,3 35,2 35,2 35,3 35,0 41 35,3 39,0 35,3 35,0 35,2 35,3 35,9 42 36,4 39,6 34,9 35,8 35,4 36,0 35,0 43 37,3 40,1 34,8 36,1 35,3 35,1 35,4

Anhang
126
44 37,2 38,5 36,5 35,8 35,0 35,7 37,8 45 37,1 39,9 36,3 35,6 35,6 35,7 35,6 46 37,5 39,0 36,2 36,2 36,7 36,1 36,5 47 37,8 39,5 36,2 35,9 36,8 35,6 35,2 48 37,9 41,6 36,4 36,0 37,0 35,1 35,7
MW 35,8 Min 34,8 Max 41,6 SD 0,9
2. Hyperthermie n= 5
Stunde 6 7 8 9 10 1 35,3 35,9 35,1 35,0 34,0 2 37,0 35,8 36,0 36,1 36,7 3 36,6 35,4 35,6 35,7 36,2 4 36,2 35,3 35,7 35,7 35,8 5 35,8 35,7 35,9 35,4 36,2 6 35,6 35,6 35,5 35,3 37,0 7 35,6 35,7 35,2 35,5 36,3 8 35,6 35,9 36,0 35,4 35,9 9 35,8 35,4 36,6 35,6 36,0
10 36,2 35,3 36,8 35,4 35,7 11 36,1 35,9 36,3 35,3 35,4 12 35,5 35,9 36,3 35,3 35,5 13 35,7 35,3 36,7 35,0 36,2 14 36,1 35,2 36,3 35,4 36,3 15 35,9 34,9 36,6 35,0 36,0 16 35,4 35,6 37,0 35,2 36,1 17 35,8 35,6 36,6 35,7 36,0 18 36,0 36,0 35,7 35,1 36,5 19 35,6 35,7 35,3 35,3 35,9 20 35,6 35,5 35,1 35,6 36,1 21 35,7 35,6 35,3 35,7 36,0 22 35,0 35,8 35,8 35,8 35,9 23 35,3 36,4 36,6 36,0 35,6 24 35,0 36,0 36,3 36,2 35,9 25 35,9 36,1 35,4 36,8 35,8 26 35,6 35,3 35,5 36,4 35,6 27 35,7 35,5 37,0 37,1 35,4 28 35,1 35,7 38,8 37,0 36,5 29 35,3 36,0 37,3 36,1 35,4 30 35,4 36,1 37,2 35,9 35,3 31 35,7 36,4 37,2 35,0 35,1

Anhang
127
32 35,4 35,5 36,7 35,3 36,7 33 35,2 36,1 36,9 35,5 36,3 34 35,8 36,0 37,4 35,6 36,2 35 36,0 35,6 38,0 35,8 36,4 36 36,1 35,5 35,4 35,7 36,1 37 36,4 36,2 36,0 35,7 37,0 38 35,7 35,9 35,5 35,4 36,9 39 35,8 35,6 35,2 35,8 36,3 40 35,3 35,6 35,2 35,5 36,1 41 35,3 35,7 35,2 35,6 35,9 42 34,9 36,3 35,4 35,4 35,7 43 35,8 36,1 35,3 35,3 35,8 44 36,5 35,8 35,8 35,5 35,9 45 36,3 35,6 35,6 35,6 35,8 46 36,2 36,2 36,7 35,7 35,6 47 36,2 36,7 36,8 36,0 35,8 48 36,4 36,0 37,0 36,1 35,8
MW 35,9 Min 34,0 Max 38,8 SD 0,6
9.4 Alter und Gewicht der Rüden
Hund Alter zu Beginn der Studie Durchschnittsgewicht über den Studienzeitraum 1 11 Monate 15,4 kg 2 11 Monate 16,0 kg 3 11 Monate 15,8 kg 4 21 Monate 14,7 kg 5 38 Monate 14,0 kg 6 40 Monate 16,0 kg 7 40 Monate 14,5 kg 8 20 Monate 13,6 kg 9 20 Monate 14,7 kg 10 21 Monate 13,9 kg

Anhang
128
Abbildungen Abbildung 1: Suspensorium zur Erhöhung der Skrotaltemperatur bei Rüden .......... 38
Abbildung 2: Mittlerer (±SD) Skrotalumfang (cm), vor und nach der 1. und 2.
skrotalen Hyperthermie; n = Anzahl der Rüden, T = Tag, SD = Standardabweichung
................................................................................................................................. 47
Abbildung 3: Mittlere (±SD) Länge und Breite des a) linken und b) rechten Hodens,
gemessen mittels Schieblehre, vor und nach der 1. und 2. skrotalen Hyperthermie, n
= Anzahl der Rüden, T = Tag, SD = Standardabweichung....................................... 48
Abbildung 4: Mittlere (±SD) Länge (L) und Breite (B) des linken (li) und rechten (re)
Hodens (H), sowie der Länge (L) und Höhe (H) der Prostata (P) in der
Kontrollgruppe, vor und nach der 1. und 2. skrotalen Hyperthermie; n = Anzahl der
Rüden, SD = Standardabweichung .......................................................................... 49
Abbildung 5: Mittels Ultraschall gemessene mittlere (±SD) Länge und Breite des
linken (a) und rechten (b) Hodens vor und nach der 1. und 2. skrotalen Hyperthermie;
n = Anzahl der Rüden, SD = Standartabweichung ................................................... 50
Abbildung 6: Mittels Ultraschall gemessene mittlere (±SD) Länge und Höhe der
Prostata vor und nach der 1. und 2. skrotalen Hyperthermie; n = Anzahl der Rüden,
SD = Standardabweichung....................................................................................... 51
Abbildung 7: Mittlere (±SD) winkelkorrigierte systolische Maximalgeschwindigkeit
(SPV) der A. marginalis am linken (a) und rechten (b) Hoden vor und nach der 1. und
2. sktoralen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung ....... 52
Abbildung 8: Mittlere (±SD) winkelkorrigierte systolische Maximalgeschwindig-keit
(SPV) der A. marginalis des rechten und linken Hodens vor und nach der 1. und 2.
sktoralen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung ........... 53
Abbildung 9: Mittlerer (±SD) Pulsatilitätsindex (PI) und Widerstandsindex (RI) am a)
linken Hoden und b) rechten Hoden vor und nach der 1. und 2. sktoralen
Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung........................... 54
Abbildung 10: Mittlerer (±SD) Pulsatilitätsindex (PI) und Widerstandsindex (RI) am
linken und rechten Hoden in der Kontrollgruppe, im Vorlauf sowie unmittelbar nach
der 1. und 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 55

Anhang
129
Abbildung 11: a) Vakuolig veränderte Zellen des Keimepithels eines Hundes 40 Tage
nach der 2. skrotalen Hyperthermie (x200); b) unveränderte Hodentubuli eines
Hundes der Kontrollgruppe (x100)............................................................................ 57
Abbildung 12: Zelldebris im Lumen eines Hodentubulus eines Hundes neun Wochen
nach einmaliger skrotaler Hyperthermie (x400) ........................................................ 58
Abbildung 13: Mittlere Anzahl (±SD) der von verschiedenen Degenerations-
erscheinungen betroffenen Hodentubuli in der Kontrollgruppe (n = 3), neun Wochen
nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 (n = 2) Tage
nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 59
Abbildung 14: a) Hodentubuli eines Hundes der Kontrollgruppe, eine einzelne
TUNEL - positive Zelle (↑) in einem Tubulus (x100); b) vier TUNEL - positive Zellen
(↑) in einem Hodentubulus eines Hundes zehn Tage nach der 2. skrotalen
Hyperthermie (x400)................................................................................................. 60
Abbildung 15: Anzahl TUNEL - positiver Zellen pro 100 Sertolizellen in der
Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40
Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 61
Abbildung 16: Prozentualer Anteil aller Tubuli mit TUNEL - positiven Zellen pro 100
Sertolizellen aufgeteilt nach den Stadien des Keimepithelzyklus über alle Gruppen
zusammengefasst (Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen
Hyperthermie (n = 2) sowie zehn (n = 3) und 40 (n = 2) Tage nach der 2. skrotalen
Hyperthermie)........................................................................................................... 62
Abbildung 17: Anzahl TUNEL-positiver Zellen pro 50 Hodentubuli in der
Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40
Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 63
Abbildung 18: a) Hodentubuli eines Kontrolltieres, keine Caspase - 3 - positive Zelle
(x100); b) Hodentubuli eines Rüden neun Wochen nach der 1. skrotalen
Hyperthermie, drei Caspase - 3 - positive Zellen (↑) in einem Hodentubulus (x100) 64

Anhang
130
Abbildung 19: Caspase-3-positive Zelle (↑) im Hodentubulus eines Hundes zehn
Tage nach der 2. skrotalen Hyperthermie (x400) ..................................................... 65
Abbildung 20: Anzahl Caspase-3-positiver Zellen pro 100 Sertolizellen in der
Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40
Tage nach der 2. skrotalen Hyperthermie ................................................................ 66
Abbildung 21: Prozentualer Anteil aller Tubuli mit Caspase-3-positiven Zellen pro 100
Sertolizellen aufgeteilt nach den Stadien des Keimepithelzyklus über alle Gruppen
zusammengefasst (Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen
Hyperthermie (n = 2) sowie zehn (n = 3) und 40 (n = 2) Tage nach der 2. skrotalen
Hyperthermie)........................................................................................................... 67
Abbildung 22: Anzahl Caspase -3 - positiver Zellen pro 50 Hodentubuli in der
Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40
Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 68
Abbildung 23: a) Ki - 67 - positive Spermatogonien (↑) in den Hodentubuli eines
Hundes zehn Tage nach der 2. skrotalen Hyperthermie (x100); b) einzelne Ki - 67 -
positive Spermatogonien in dem Hodentubuli eines Hundes der Kontrollgruppe
(x100); positiv erscheinende Spermatozyten wurden nicht mitgezählt ..................... 70
Abbildung 24: Anzahl Ki-67-positiver Zellen pro 100 Sertolizellen in der
Kontrollgruppe, neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40
Tage nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 71
Abbildung 25: Anzahl Ki-67-positiver Zellen pro 50 Hodentubuli in der Kontrollgruppe,
neun Wochen nach der 1. skrotalen Hyperthermie sowie zehn und 40 Tage nach der
2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung ....... 72
Abbildung 26: a) Vakuolig aufgelockerte Epithelzellen in einem Nebenhodentubulus
eines Hundes zehn Tage nach der 2. skrotalen Hyperthermie (x100); b)
Nebenhodentubulusanschnitt ohne vakuolig verändertes Epithel eines Hundes der
Kontrollgruppe (x100) ............................................................................................... 74
Abbildung 27: Nebenhodentubulus ohne Spermien (↑) eines Hundes neun Wochen
nach einmaliger skrotaler Hyperthermie (x200) ........................................................ 75

Anhang
131
Abbildung 28: Mittlere Anzahl (±SD) der von verschiedenen
Degenerationserscheinungen betroffenen Nebenhodentubulusanschnitte in der
Kontrollgruppe (n = 3), neun Wochen nach der 1. skrotalen Hyperthermie (n = 2)
sowie zehn (n = 3) und 40 (n = 2) Tage nach der 2. skrotalen Hyperthermie; n =
Anzahl der Rüden, SD = Standardabweichung ........................................................ 76
Abbildung 29: Anzahl TUNEL - positiver Zellen pro 25 Nebenhodentubulus-
anschnitte im Stroma, Epithel und Lumen in der Kontrollgruppe (n = 3), neun Wochen
nach der 1. skrotalen Hyperthermie (n = 2), sowie zehn (n = 3) und 40 Tage (n = 2)
nach der 2. skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 77
Abbildung 30: Anzahl Caspase-3-positiver Zellen pro 25 Nebenhoden-
tubulusanschnitte im Stroma, Epithel und Lumen in der Kontrollgruppe (n = 3), neun
Wochen nach der 1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 Tage
(n = 2) nach der 2. zweiten skrotalen Hyperthermie; n = Anzahl der Rüden, SD =
Standardabweichung................................................................................................ 78
Abbildung 31: Ki-67 - positive Zellen (↑) im Nebenhodentubulusepithel eines Hundes
neun Wochen nach einmaliger skrotaler Hyperthermie (x400) ................................. 79
Abbildung 32: Anzahl Ki-67 - positiver Zellen pro 25 Nebenhoden-tubulusanschnitte
im Stroma, Epithel und Lumen in der Kontrollgruppe (n = 3), neun Wochen nach der
1. skrotalen Hyperthermie (n = 2) sowie zehn (n = 3) und 40 Tage (n = 2) nach der
zweiten skrotalen Hyperthermie; n = Anzahl der Rüden, SD = Standardabweichung
................................................................................................................................. 80
Tabellen Tabelle 1: Versuchsablauf 34

Anhang
132
Danksagung Frau Prof. Dr. A.-R. Günzel-Apel danke ich für die Überlassung dieses interessanten
Themas, sowie für die Möglichkeit die Routinesprechstunde im Bereich
Rreproduktionsmedizin kennen zu lernen. Ebenfalls vielen Dank an Frau Prof.
Pfarrer die mir ganz zum Schluss mit Rat und Tat zur Seite stand und trotz der
knappen Zeit an mich geglaubt hat.
Vielen Dank auch an Christine für die Zusammenarbeit und den Spaß den wir trotz
48 Stunden Kälte zusammen hatten. Ebenso vielen Dank an Caro, dass sie die
Arbeit mit mir durchgeboxt hat. Danke auch an den Rest des Reproteams für die Zeit
mit Keksen, Süßigkeiten und dem ein oder anderen Späßchen zwischen den
Terminen.
Herrn Prof. Dr. A. Beinecke danke ich für die Unterstützung und Beratung in allen
histologischen Fragen. Außerdem vielen Dank an B. Buck und P. Grünig für die tolle
Unterstützung und Hilfe bei der Bearbeitung der Proben, bis sie dann endlich alle
geschnitten und gefärbt waren.
Einen außerordentlichen Dank auch an Daniel für die Nerven, dass 2 Jahre
durchzustehen und mir trotzdem noch zu helfen und mich so zu nehmen wie ich bin.
Weiteren Dank verdienen alle meine Hannover-Mädels, denn ohne diesen Rückhalt
wäre schon das Studium eine Qual gewesen und die Doktorarbeit erst Recht! Special
thanks an die Recherche-Königin Nina!!!!
Auch einen Dank verdient hat meine fröhliche durch geknallte Frieda, die doch in
Zeiten der Krise mehr als einmal meine Laune aufgehellt hat.
Ebenfalls einen Dank an meine Chefin Ulrike für die Unterstützung und die häufige
Nachsicht, wenn ich mal wieder gestresst und mit den Nerven am Ende war, sowie
für das großzügige, auch kurzfristige, Freistellen zu mehr als einer Gelegenheit.

Anhang
133
Und last but not least vielen Dank an meine gesamte Familie, besonders meine
Eltern, die mir erst das Studium und die Arbeit ermöglicht haben und dass sowohl mit
finanzieller Unterstützung, aber auch mit der noch um einiges wichtigeren
moralischen und seelischen Unterstützung!!!!!!!!!


















