
Effects of systemic or oral ad libitum monosodium ...web.mit.edu/dick/www/pdf/897.pdf · Effects of...
4
ELSEVIER 1 BRAIN RESEARCH Brain Research 660 (\994) 337-340 Short communication Effects of systemic or oral ad libitum monosodium glutamate administration on striatal glutamate release, as measured using microdialysis in freely moving rats Mikhail B. Bogdanov, Richard J. Wurtman * MassachllseltS Instiwte of Technology. Deparrmem of Brain and Cogniti!'e Sciences. £25-604. 0/5 Carleton Street. Cambridge. AlA 011-11. L'SA Accepted 26 July 1994 Abstract We examined effects of high doses of monosodium glutamate (MSG) on extracellular glutamate levels in rat striata. using. in vivo microdialysis. Parenteral doses (0.5. 1.0 and 2.0. but not 0.25. g/kg, i.pJ caused dose- and time-dependent increases. peaking after 40 min (at 174:t 47%. 485 :t 99% and 1021:t 301% of basal levels. respectively). In contrast. dietary MSG (I.49 :t 0.10 g/kg/h) was ineffective. Keywords: Monosodium glutamate; Glutamate release; Microdialysis; Striatum Glutamate is the principal excitatory neurotrans- mitter in the brain. It interacts with two main subtypes of membrane receptors, ionotropic and metabotropic. coupled to ion channels and G proteins, respectively. The ionotropic receptors are further subdivided. based on selective agonists. into N-methyl-D-aspartate (NM- DA), kainate. and a-amino-3-hydroxy-5-methyl-4-iso- xazolepropionate (AMP A) subtypes [18]. Interactions of glutamate with its ionotropic, mainly NMDA, recep- tors can lead to neurotoxic changes in some experi- mental situations by allowing excessive amounts of calcium to enter the neuron [5]. That exogenous glutamate could be neurotoxic was first proposed by Lucas and Newhouse [10] who de- scribed neuronal degeneration in the inner layer of the retina following subcutaneous administration of MSG to the neonatal mouse. Subsequently, Olney [11] ob- served necrosis in the hypothalamic (arcuate nucleus) neurons of neonatal mice given MSG systemically. Other investigators have also found neurodegenerative changes after MSG administration in various rodent species, usually when the compound was administered subcutaneously or by forced gavage. It appears, however, that the neurotoxic potential of * Corresponding author. Fax: (1) (617) 253-6882. 0006-8993/94/$07.00 @ 1994 Elsevier Science B.V. All rights reserved SSDI 0006.8993(94)00902-3 ---- exogenous glutamate in vivo is critically dcpendcnt upon its route of administration. Neuronal lesions have never been observed after ad libitum consumption of very high MSG doses. Thus, Heywood et al. [8] failed to observe hypothalamic lesions in weanling mice. probably the most sensitive species. following ad lihi- tum MSG administration in the dict or drinking watcr at doses as high as 45.5 g/kg or 20.9 g/kg. respectively. (These doses are 10-20 times higher than those rc- quired to induce neurodegenerative changes following parenteral or forced oral administration). This differ- ence is probably related to differences in the pharma- cokinetics of MSG depending on its route of adminis- tration which, in turn. determine its effects on extracel- lular brain glutamate concentrations. When glutamate is consumed orally, its effects on the brain are buffered by metabolism in the gastrointestinal tract. extrusion from the brain by active blood-brain transport systems, and local mechanisms mediating its uptake and metabolism in brain; these cause brain extracellular glutamate concentrations to remain relatively stable. (The mechanism of MSG's neurotoxic effects has been attributed to a prolonged increase in extracellular glu- tamate concentrations [12]). No data are available at present on possible effects of MSG on extracellular glutamate concentrations within the brain. Hence we have examined this rela-
Transcript of Effects of systemic or oral ad libitum monosodium ...web.mit.edu/dick/www/pdf/897.pdf · Effects of...

ELSEVIER
1BRAINRESEARCH
Brain Research 660 (\994) 337-340
Short communication
Effects of systemic or oral ad libitum monosodium glutamateadministration on striatal glutamate release, as measured
using microdialysis in freely moving rats
Mikhail B. Bogdanov,Richard J. Wurtman *
MassachllseltS Instiwte of Technology. Deparrmem of Brain and Cogniti!'e Sciences. £25-604. 0/5 Carleton Street. Cambridge. AlA 011-11. L'SA
Accepted 26 July 1994
Abstract
We examined effects of high doses of monosodium glutamate (MSG) on extracellular glutamate levels in rat striata. using. invivo microdialysis. Parenteral doses (0.5. 1.0 and 2.0. but not 0.25. g/kg, i.pJ caused dose- and time-dependent increases.peaking after 40 min (at 174:t 47%. 485 :t 99% and 1021:t 301% of basal levels. respectively). In contrast. dietary MSG(I.49 :t 0.10 g/kg/h) was ineffective.
Keywords: Monosodium glutamate; Glutamate release; Microdialysis; Striatum
Glutamate is the principal excitatory neurotrans-mitter in the brain. It interacts with two main subtypesof membrane receptors, ionotropic and metabotropic.coupled to ion channels and G proteins, respectively.The ionotropic receptors are further subdivided. basedon selective agonists. into N-methyl-D-aspartate (NM-DA), kainate. and a-amino-3-hydroxy-5-methyl-4-iso-xazolepropionate (AMP A) subtypes [18]. Interactionsof glutamate with its ionotropic, mainly NMDA, recep-tors can lead to neurotoxic changes in some experi-mental situations by allowing excessive amounts ofcalcium to enter the neuron [5].
That exogenous glutamate could be neurotoxic wasfirst proposed by Lucas and Newhouse [10] who de-scribed neuronal degeneration in the inner layer of theretina following subcutaneous administration of MSGto the neonatal mouse. Subsequently, Olney [11] ob-served necrosis in the hypothalamic (arcuate nucleus)neurons of neonatal mice given MSG systemically.Other investigators have also found neurodegenerativechanges after MSG administration in various rodentspecies, usually when the compound was administeredsubcutaneously or by forced gavage.
It appears, however, that the neurotoxic potential of
* Corresponding author. Fax: (1) (617) 253-6882.
0006-8993/94/$07.00 @ 1994 Elsevier Science B.V. All rights reservedSSDI 0006.8993(94)00902-3
----
exogenous glutamate in vivo is critically dcpendcntupon its route of administration. Neuronal lesions havenever been observed after ad libitum consumption ofvery high MSG doses. Thus, Heywood et al. [8] failedto observe hypothalamic lesions in weanling mice.probably the most sensitive species. following ad lihi-tum MSG administration in the dict or drinking watcrat doses as high as 45.5 g/kg or 20.9 g/kg. respectively.(These doses are 10-20 times higher than those rc-quired to induce neurodegenerative changes followingparenteral or forced oral administration). This differ-ence is probably related to differences in the pharma-cokinetics of MSG depending on its route of adminis-tration which, in turn. determine its effects on extracel-lular brain glutamate concentrations. When glutamateis consumed orally, its effects on the brain are bufferedby metabolism in the gastrointestinal tract. extrusionfrom the brain by active blood-brain transport systems,and local mechanisms mediating its uptake andmetabolism in brain; these cause brain extracellularglutamate concentrations to remain relatively stable.(The mechanism of MSG's neurotoxic effects has beenattributed to a prolonged increase in extracellular glu-tamate concentrations [12]).
No data are available at present on possible effectsof MSG on extracellular glutamate concentrationswithin the brain. Hence we have examined this rela-

338 M.B. Bogdanol". R.I. Wurtman / Brain Research 660 (199.J! 337-3.J0
tionship, using brain microdialysis, in freely movingrats, receiving MSG systemically or via the diet adlibitum.
Male Sprague-Dawley rats were housed for 1 weekbefore each experiment. 2 per cage, under a 12 hdarkj 12 h light cycle, with food (Prolab Animal Diet3000, Agway Inc.. Syracuse, NY, 22~ protein) andwater available ad libitum.
The effects of MSG on striatal glutamate releasewere studied following systemic intraperitoneal MSGadministration or elective consumption of MSG-con-taining diet.
In the first study the rats <250-300 g) were im-planted with microdialysis probes 18-24 h before thestart of perfusion. Dialysate samples were collecteduntil stable glutamate levels were obtained. MSG wasthen injected intraperitoneally (4 ml/kg, dissolved indistilled water) and the measurements were continuedfor an additional 3 h. Control animals received aninjection of saline.
In the second study the animals were trained, inadvance of the microdialysis experiments, to consumetheir total daily food intake within 1 h. One week afterarrival the rats (200-250 g at the start of the trainingperiod) were housed I per cage with water available adlibitum. The above rat diet. was provided ad libitumover 1 h, from noon to 1:00 p.m. The training periodcontinued for 2-3 weeks. After the last training ses-sion, the rats were implanted with microdialysis probes,and perfusion experiments were started the next morn-ing. After collection of basal dialysate samples, MSG-containing or control diet was provided to the animalsbetween noon and 1:00 p.m. Measurements were con-tinued during the feeding period and for 3 h after-wards. The MSG-based diet contained: 25.2SCprotein,.+.40Cfat. 4.40C crude fiber. 70'c ash, 50.20'c carbo-hydrate and 6.6% MSG. In the control diet an equiva-lent amount of protein was substituted for the MSG(i.e. protein content of the control diet was 31.89c).
Microdialysis probes of concentric design (mem-brane length 4 mm: o.d. 250 .urn. in vitro recovery forglutamate at 1 .ul/min 20-25%) were used. The ani-mals were anaesthetized with ketamine jxylazine om j10 mgjkg i.p.) and placed in a Kopf stereotaxic frame.After the skull was exposed and a burr hole drilled, theprobe was slowly (2-3 min) lowered into the rightstriatum and secured to the skull. Coordinates of theprobe's tip: AP + 0.5; ML 2.7; V -7.2 [16]. Aftercompletion of the experiments the animals were sacri-ficed, and the localization of the probes in the striatumverified.
The probe was perfused with modified Ringer'ssolution (145 mM NaCl, 2.7 mM KCI, 1.2 CaCI2, 1.0mM MgCI2, pH 7.4 with 2 mM phosphate buffer) at 1.uljmin using a microperfusion pump (CMAj100,CMA, Sweden). Dialysate samples were collected every
20 min and analyzed by HPLC with electrochemicaldetection after precolumn derivatization with o-phta-laldehyde (OPA>. as described by Donzanti and Ya-mamoto [6], with minor modifications. The derivatiza-tion stock reagent included 27 mg of OPA. I ml ofmethanol. 5 .ul of 2-mercaptoethanol (2-ME) and Ymlof 0.1 M sodium tetraborate. pH 9.3. The workingsolution was prepared by diluting the stock solutionwith 0.1 M sodium tetraborate (1: 2.+)' Precolumnderivatization of ammo acids was performed by mixing10 .ul of the dialysate sample or standard with 10 .ul ofthe working OPAj2-ME reagent for 2 min beforeinjection onto the column. The HPLC system consistedof a dual piston pump (L-6000. Hitachi); an ESA 465autosampler (ESA. Bedford. MA): a 3-.um ODS SOX4.6 mm column (HR-80, ESA), and an ESA 5200coulometric detector with an ESA 5014 dual-electrodeanalytical cell. The first electrode was set at + 200 mY.and the second at + 400 mY. The mobile phase deliv-ered at 1.2 mljmin was 0.1 M sodium dibasic phos-phate buffer, 25% (vjv) methanol and 5% (vjv) aceto-nitrile, pH 6.4.
Basal levels of glutamate in 20-min striatal dialysatesamples were 1.45:t 0.058 .uM (n = 9). Once theselevels stabilized (usually 3-4 h after the start of perfu-sion), they remained so for the next 3 h of perfusion.Injection of saline did not modify glutamate dialysatelevels.
At a 0.25 gjkg i.p. dose, MSG failed to affectglutamate output during the 3 h after its injection. 0.5gjkg i.p. of MSG did cause an increase in glutamatelevels. This was observed in the initial 20-min sample(up to 156:t 36% of basal values. P < 0.05). and be-came maximal with the second 20-min sample (174:t47q. of basal values, P < 0.05, Fig. n remaining ele-vated for an additional hour. The two highest MSGdoses studied caused massive increases in glutamatelevels, prominent in the first 20-min sample after injec-tion and peaking in the second 20-min sample. The 1and 2 gjkg doses increased glutamate levels maximallyto 485 :t 99% (P < 0.05) and 1021:t 301% (P < 0.05)of basal values, respectively (Fig. 1). Glutamate levelsreturned to basal values within 2 h after MSG adminis-tration.
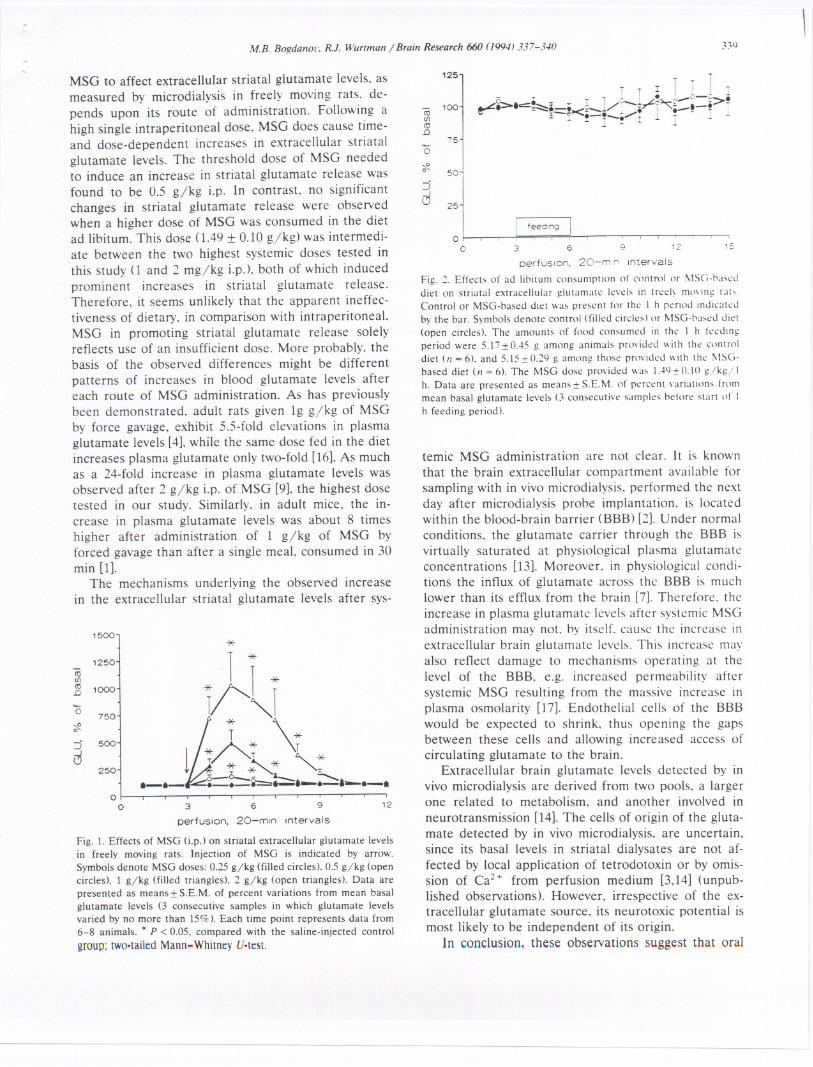
The amount of food consumed by the rats in the 1 hperiod during microdialysis perfusion was the same inboth groups of animals: i.e. 5.17:t 0.45 g in animalsprovided with the control diet (n = 6) and 5.15 :t 0.29 gin animals given the MSG-based diet (n = 6). Aftercorrection for body weight, the oral dose level of MSGwas calculated to have been 1.49:t 0.10 gjkgj 1h. Inboth groups of animals, dialysate glutamate levels dur-ing the 1 h feeding period and the subsequent 3 h, didnot differ from basal levels, ascertained before thestart of feeding (Fig. 2).
These data clearly demonstrate that the ability of
-----
M.B. Bogdallol". R.I. Wurtmall j Braill Research 660 (19941337-340
MSG to affect extracellular striatal glutamate levels. asmeasured by microdialysis in freely moving rats. de-pends upon its route of administration. Following ahigh single intraperitoneal dose. MSG does cause time-and dose-dependent increases in extracellular striatalglutamate levels. The threshold dose of MSG neededto induce an increase in striatal glutamate release wasfound to be 0.5 g/kg i.p. In contrast. no significantchanges in striatal glutamate release were observedwhen a higher dose of MSG was consumed in the dietad libitum. This dose (1.49 J: 0.10 g/kg) was intermedi-ate between the two highest systemic doses tested inthis study (1 and 2 mg/kg i.pJ. both of which inducedprominent increases in striatal glutamate release.Therefore, it seems unlikely that the apparent ineffec-tiveness of dietary. in comparison with intraperitoneal.MSG in promoting striatal glutamate release solelyreflects use of an insufficient dose. More probably. thebasis of the observed differences might be differentpatterns of increases in blood glutamate levels aftereach route of MSG administration. As has previouslybeen demonstrated. adult rats given Ig g/kg of MSGby force gavage, exhibit 5.5-fold elevations in plasmaglutamate levels [4], while the same dose fed in the dietincreases plasma glutamate only two-fold [16]. As muchas a 24-fold increase in plasma glutamate levels wasobserved after 2 g/kg i.p. of MSG [9]. the highest dosetested in our study. Similarly. in adult mice, the in-crease in plasma glutamate levels was about 8 timeshigher after administration of 1 g/kg of MSG byforced gavage than after a single meal, consumed in 30min [1].
The mechanisms underlying the observed increasein the extracellular striatal glutamate levels after sys-
Fig. 1. Effects of MSG (j.p') on striatal extracellular glutamate levelsin freely moving rats. Injection of MSG is indicated by arrow.Symbols denote MSG doses: 0.25 gjkg (filled circles). 0.5 gjkg (opencircles), 1 gjkg (filled triangles). 2 gjkg (open triangles). Data arepresented as means:!: S.E.M. of percent variations from mean basalglutamate levels (3 consecutive samples in which glutamate levelsvaried by no more than 15%). Each time point represents data from6-8 animals. * P < 0.05. compared with the saline-injected control
group; tWo.tailed Mann-Whitney V.test.
----
'25
1
- '00co<f)co.D
75'0
50
25
oo 3 6 9 12 15
perfuSion. 20-m,r' .ncervals
Fig. 2. Effects of ad lioitum consumption of contml or !\ISG-oaseddiet on striatal extracellular glutamate levels in treel~ mmlllg rab.Control or MSG-based diet was present for the I h period indicatedby the bar. Symools denote control (filled circles) or MSG-oased diet(open circles). The amounts of food consumed in the I h feedingperiod were 5.I7 :!:O..t5g among animals pnwided with the controldiet (II = 6), and 5.l5:!: 0.29 g among those pnwided with the !\ISG-based diet (11= 6). The MSG dose provided was l..N:!: O.W gjkg/ Ih. Data are presented as means:!: S.E.M. of percent variations frommean basal glutamate levels (3 consecutive samples odore start of Ih feeding period).
temic MSG administration are not clear. It is knownthat the brain extracellular compartment available forsampling with in vivo microdialysis. performed the nextday after microdialysis probe implantation. is locatedwithin the blood-brain barrier (BBB) [2]. Under normalconditions. the glutamate carrier through the BBB isvirtually saturated at physiological plasma glutamateconcentrations [13]. Moreover. in physiological condi-tions the influx of glutamate across the BBB is muchlower than its efflux from the brain [7]. Therefore. theincrease in plasma glutamate levels after systemic MSGadministration may not. by itself. cause the increase inextracellular brain glutamate levels. This increase mayalso reflect damage to mechanisms operating at thelevel of the BBB. e.g. increased permeability aftersystemic MSG resulting from the massive increase inplasma osmolarity [17]. Endothelial cells of the BBBwould be expected to shrink. thus opening the gapsbetween these cells and allowing increased access ofcirculating glutamate to the brain.
Extracellular brain glutamate levels detected by invivo microdialysis are derived from two pools. a largerone related to metabolism. and another involved inneurotransmission [14]. The cells of origin of the gluta-mate detected by in vivo microdialysis, are uncertain,since its basal levels in striatal dialysates are not af-fected by local application of tetrodotoxin or by omis-sion of Ca2+ from perfusion medium [3,14] (unpub-lished observations). However, irrespective of the ex-tracellular glutamate source, its neurotoxic potential ismost likely to be independent of its origin.
In conclusion, these observations suggest that oral

340 M.B. BogdanoL'. R.I. Wurtman / Brain Research 660 (1994) 337-340
MSG. consumed ad libitum evenfails to affect in vivo glutamatebrain.
at very high doses,release within the
These studies were supported infrom the Center for Brain SciencesCharitable Trust.
part by a grantand Metabolism
[I] Airoldi. L.. Bizzi. A.. Salmona. M.. and Garratini. S.. Attemptsto establish the saqfety margin for neurotoxicity of monosodiumglutamate. In Filer. L.J.. Garratini. S.. Kare. M.R.. Reynolds.W.A.. and Wurtman. R.J. (EdsJ. GllI/amic Acid: Adl"Dllces inBiochemisrry alld Physiology. Raven. New York. 1979. pp. 321-331.
[2) Benveniste. H.. Brain microdialysis. 1. Nellrochem.. 52 (1989)1667-1679.
[3} Butcher. S.P. and Hamberger. A.. In vivo studies of the extracel-lular. and veratrine-releasable. pools of endogenous amino acidsin the rat striatum: effects of corticostriatal deafferentation andkainic acid lesion. 1. Nellrochem.. 48 (1987) 713-721.
[4} Caccia. S.. Garattini. S.. Ghezzi. P. and Zanini. M.G.. Plasmaand brain levels of glutamate and pyroglutamate after oralmonosodium glutamate to rats. Toxico/. Lerr.. 10 (1982) 169-175.
[5} Choi. D.W.. Glutamate neurotoxicityin cortical cell culture iscalcium dependent. Nellrosci. Lerr.. 58 (1985) 293-297.
[6} Donzanti. B.A. and Yamamoto. B.K.. An improved and rapidHPLC.EC metod for the isocratic separation of amino acidneurotransmitters from brain tissue and microdialysis per.fusates. Life Sci.. 43 (1988) 913-922.
[7} Fonnum. F.. Glutamate: a neurotransmitter in mammalian brain.1. Nellrochem.. 42 (1984) I-II.
[8} Heywood. R.. James. R.W. and Worden. A.N.. The ad libitumfeeding of monosodium glutamate to wealning mice. Toxico/.Leu.. I (1977) 151-155.
[9] Liebshutz. J.. Airoldi. L.. Brownstein. M.J.. Chinn. N.G. andWurtman. R.J.. Regional distribution of endogenous and par.enteral glutamate. aspartate and glutamine in rat brain.Biochem. Pharmacal.. 26 (1977) 443-446.
[10] Lucas. D.R. and Newhouse. J.P.. The toxic effect of sodiumL-glutamate on the inner layers of the retina. A.\IA ArchOprhalmol.. 58 (1957) 193-201.
[II} Olney. J.W.. Brain lesions. obesity and other disturbances inmice treated with monosodium glutamate. SC/ellcc. 164 (19091719-721.
[12] Olney. J.W.. Excitotoxic amino acids: Research applications andsafety implications. In Filer. L.J.. Garratini. S.. Kare. M.R..Reynolds. W.A. and Wurtman. R.J. (EdsJ. GllI/amic Acid: Ad.l"Dncesin Biochemisrry and Physiology. Raven. New York. 1979.pp.287-319.
[13) Pard ridge. W.M.. Regulation of amino acids availability to hrain:selective control mechanisms for glutamate. In Filer. L.J.. Gar.ratini. S.. Kare. M.R.. Reynolds. W.A. and Wurtman. R.J.(Eds,). Glmamic Acid: Adl'allces in Biochemisrry and Phniology.Raven. New York. 1979. pp. 125-138.
[14} Paulsen. R.E. and Fonnum. F.. Role of glial cells for the basaland Ca~--dependent K--evoked release of transmitter aminoacids investigated by microdialysis.1. Nellrochem..52 (1989)1823-1829.
[15} Paxinos. G. and Watson. c.. The Rar Brain ill SrereOTaxicCoordi-lIares.AcademicPress. New York. 1986.
[16} Peng. Y.. Gubin. 1.. Harper. A.E.. Yavich.M.G. and Ko:mmer.A.R.. Food intake regulation: Amino acid toxicity and chango:sin rat brain and plasma amino acids. 1. Nmr.. 103 (1973)608-617.
[17] Torii. K.. Takasaki. Y.. Iwata. S. and Wurtman. R.J.. Changes inblood osmolarity. electrolytes. and metabolites among adult ratstreated with a neurotoxic dose of MSG. L~(e Sci.. 28 (l9!H)2855-2864.
[18] Zomurski. C.F. and Thio. L.L.. Properties of vertehrate gluta-mate receptors: Calcium mobilization and desentization. Prog.Neurobiol.. 39 (1992) 295-336.
--- - -----












![Monosodium Glutamate Induced Hepatotoxicity and Oxidative ......modified Ishak's staging system [24]. Modified Ishak staging system “Histological Activity Index (HAI)” staging:](https://static.fdocuments.net/doc/165x107/5e4b7971dc56d95ddf0af434/monosodium-glutamate-induced-hepatotoxicity-and-oxidative-modified-ishaks.jpg)





