
Effects of Repeated Antidepressant Treatment on Type 4A Phosphodiesterase (PDE4A) in Rat Brain
6
Effects of Repeated Antidepressant Treatment on Type 4A Phosphodiesterase (PDE4A) in Rat Brain Ying Ye, Keasha Jackson, and James M. O’Donnell Department of Pharmacology and Therapeutics, Louisiana State University Medical Center, Shreveport, Louisiana, U.S.A. Abstract: In a previous study, an up-regulation of rolip- ram-sensitive, low-K m , cyclic AMP phosphodiesterase (PDE4) subtype PDE4A in rat cerebral cortex following repeated treatment of desipramine was observed. To determine whether this effect is shared by antidepres- sants from different pharmacological classes, PDE4A ex- pression was examined using immunoblot analyses fol- lowing repeated treatment with the norepinephrine re- uptake inhibitor desipramine, the monoamine oxidase inhibitor phenelzine, the atypical antidepressant traz- odone, and the serotonin reuptake inhibitor fluoxetine. Desipramine, phenelzine, and fluoxetine all increased the intensities of the PDE4A bands in hippocampal prepara- tions; trazodone did not. In preparations of cerebral cor- tex, the intensities of the PDE4A bands were increased following desipramine treatment, not changed following phenelzine or fluoxetine treatment, and decreased fol- lowing trazodone treatment. It appears that repeated treatment with antidepressant drugs from different phar- macological classes produces similar effects on the ex- pressions of PDE4A variants in hippocampus. This effect is not correlated with the changes in b-adrenergic recep- tor densities, suggesting these antidepressants may at some point alter intracellular signal transduction path- ways in a similar manner. Key Words: Desipramine— Phenelzine—Fluoxetine—Trazodone—b-Adrenergic re- ceptor—Rolipram-sensitive, low-K m , cyclic AMP phos- phodiesterase. J. Neurochem. 74, 1257–1262 (2000). Rolipram-sensitive, low-K m , cyclic AMP phosphodi- esterase (PDE4) has been suggested to be a target for a new class of antidepressants. PDE4-specific inhibitors, such as rolipram and Ro 20-1724, exhibit antidepressant effects in both animal models and clinical trials (Wachtel and Schneider, 1986; Fleischhacker et al., 1992; O’Donnell, 1993). PDE4 is the primary form of phos- phodiesterase hydrolyzing cyclic AMP associated with the central noradrenergic system (Whalin et al., 1988; Challiss and Nicholson, 1990; Ye and O’Donnell, 1996). This neurotransmitter system has been implicated in the actions of proven antidepressant drugs (Frazer and Con- way, 1984; Charney, 1998). PDE4 is further divided into four subtypes, which are products of four individual genes. Among them, PDE4A, PDE4B, and PDE4D are expressed in brain (Swinnen et al., 1989). Each subtype consists of multiple variants due to alternative splicing and the use of different initi- ation sites. At least some forms of PDE4 are regulated, either through phosphorylation (Sette et al., 1994) or via altered expression (Torphy et al., 1992; O’Donnell, 1993; Ye et al., 1997). PDE4A is differentially down- regulated following a chronic reduction in central norad- renergic activity (Ye et al., 1997). By contrast, chronic enhancement of noradrenergic activity, produced by re- petitive desipramine administration, up-regulates PDE4A (Ye et al., 1997). The aim of the present study is to examine whether PDE4A is up-regulated following repeated treatment of antidepressants from different pharmacological classes. This would begin to show whether regulation of PDE4A, possibly via altered noradrenergic activity, is an effect shared by antidepressant drugs. Rats were repeatedly administered antidepressants, including the norepineph- rine reuptake inhibitor desipramine, the monoamine ox- idase (MAO) inhibitor phenelzine, the atypical antide- pressant trazodone, and the serotonin reuptake inhibitor fluoxetine; the expression of PDE4A subtypes was as- sessed using quantitative immunoblot analysis. MATERIALS AND METHODS Animals Male Sprague–Dawley rats (weighing 225–300 g; Harlan, Indianapolis, IN, U.S.A.) were housed in a temperature (22– 24°C)- and light (on from 6:00 a.m. to 6:00 p.m.)-controlled room and were allowed free access to food pellets and water. The studies reported in this article have been carried out in accordance with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the National Institutes of Health. Received April 14, 1999; revised manuscript received September 10, 1999; accepted October 13, 1999. Address correspondence and reprint requests to Dr. J. M. O’Donnell at Department of Pharmacology and Therapeutics, Louisiana State University Medical School, 1501 Kings Highway, Shreveport, LA 71130-3932, U.S.A. E-mail: [email protected] Abbreviations used: MAO, monoamine oxidase; PAGE, polyacryl- amide gel electrophoresis; PDE4, rolipram-sensitive, low-K m , cyclic AMP phosphodiesterase; SDS, sodium dodecyl sulfate. 1257 Journal of Neurochemistry Lippincott Williams & Wilkins, Inc., Philadelphia © 2000 International Society for Neurochemistry
Transcript of Effects of Repeated Antidepressant Treatment on Type 4A Phosphodiesterase (PDE4A) in Rat Brain

Effects of Repeated Antidepressant Treatment on Type 4APhosphodiesterase (PDE4A) in Rat Brain
Ying Ye, Keasha Jackson, and James M. O’Donnell
Department of Pharmacology and Therapeutics, Louisiana State University Medical Center, Shreveport, Louisiana, U.S.A.
Abstract: In a previous study, an up-regulation of rolip-ram-sensitive, low-Km, cyclic AMP phosphodiesterase(PDE4) subtype PDE4A in rat cerebral cortex followingrepeated treatment of desipramine was observed. Todetermine whether this effect is shared by antidepres-sants from different pharmacological classes, PDE4A ex-pression was examined using immunoblot analyses fol-lowing repeated treatment with the norepinephrine re-uptake inhibitor desipramine, the monoamine oxidaseinhibitor phenelzine, the atypical antidepressant traz-odone, and the serotonin reuptake inhibitor fluoxetine.Desipramine, phenelzine, and fluoxetine all increased theintensities of the PDE4A bands in hippocampal prepara-tions; trazodone did not. In preparations of cerebral cor-tex, the intensities of the PDE4A bands were increasedfollowing desipramine treatment, not changed followingphenelzine or fluoxetine treatment, and decreased fol-lowing trazodone treatment. It appears that repeatedtreatment with antidepressant drugs from different phar-macological classes produces similar effects on the ex-pressions of PDE4A variants in hippocampus. This effectis not correlated with the changes in b-adrenergic recep-tor densities, suggesting these antidepressants may atsome point alter intracellular signal transduction path-ways in a similar manner. Key Words: Desipramine—Phenelzine—Fluoxetine—Trazodone—b-Adrenergic re-ceptor—Rolipram-sensitive, low-Km, cyclic AMP phos-phodiesterase.J. Neurochem. 74, 1257–1262 (2000).
Rolipram-sensitive, low-Km, cyclic AMP phosphodi-esterase (PDE4) has been suggested to be a target for anew class of antidepressants. PDE4-specific inhibitors,such as rolipram and Ro 20-1724, exhibit antidepressanteffects in both animal models and clinical trials (Wachteland Schneider, 1986; Fleischhacker et al., 1992;O’Donnell, 1993). PDE4 is the primary form of phos-phodiesterase hydrolyzing cyclic AMP associated withthe central noradrenergic system (Whalin et al., 1988;Challiss and Nicholson, 1990; Ye and O’Donnell, 1996).This neurotransmitter system has been implicated in theactions of proven antidepressant drugs (Frazer and Con-way, 1984; Charney, 1998).
PDE4 is further divided into four subtypes, which areproducts of four individual genes. Among them, PDE4A,
PDE4B, and PDE4D are expressed in brain (Swinnenet al., 1989). Each subtype consists of multiple variantsdue to alternative splicing and the use of different initi-ation sites. At least some forms of PDE4 are regulated,either through phosphorylation (Sette et al., 1994) or viaaltered expression (Torphy et al., 1992; O’Donnell,1993; Ye et al., 1997). PDE4A is differentially down-regulated following a chronic reduction in central norad-renergic activity (Ye et al., 1997). By contrast, chronicenhancement of noradrenergic activity, produced by re-petitive desipramine administration, up-regulates PDE4A(Ye et al., 1997).
The aim of the present study is to examine whetherPDE4A is up-regulated following repeated treatment ofantidepressants from different pharmacological classes.This would begin to show whether regulation of PDE4A,possibly via altered noradrenergic activity, is an effectshared by antidepressant drugs. Rats were repeatedlyadministered antidepressants, including the norepineph-rine reuptake inhibitor desipramine, the monoamine ox-idase (MAO) inhibitor phenelzine, the atypical antide-pressant trazodone, and the serotonin reuptake inhibitorfluoxetine; the expression of PDE4A subtypes was as-sessed using quantitative immunoblot analysis.
MATERIALS AND METHODS
AnimalsMale Sprague–Dawley rats (weighing 225–300 g; Harlan,
Indianapolis, IN, U.S.A.) were housed in a temperature (22–24°C)- and light (on from 6:00 a.m. to 6:00 p.m.)-controlledroom and were allowed free access to food pellets and water.The studies reported in this article have been carried out inaccordance with theGuide for the Care and Use of LaboratoryAnimalsas adopted and promulgated by the National Institutesof Health.
Received April 14, 1999; revised manuscript received September 10,1999; accepted October 13, 1999.
Address correspondence and reprint requests to Dr. J. M. O’Donnellat Department of Pharmacology and Therapeutics, Louisiana StateUniversity Medical School, 1501 Kings Highway, Shreveport, LA71130-3932, U.S.A. E-mail: [email protected]
Abbreviations used:MAO, monoamine oxidase; PAGE, polyacryl-amide gel electrophoresis; PDE4, rolipram-sensitive, low-Km, cyclicAMP phosphodiesterase; SDS, sodium dodecyl sulfate.
1257
Journal of NeurochemistryLippincott Williams & Wilkins, Inc., Philadelphia© 2000 International Society for Neurochemistry

Drug administrationThirty rats were divided randomly into five groups of six rats
each. These rats received injections intraperitoneally for 2weeks, consisting of 1 ml/kg normal saline twice daily, 10mg/kg desipramine twice daily, 5 mg/kg phenelzine once daily,10 mg/kg trazodone once daily, and 5 mg/kg fluoxetine oncedaily. At 24 h after the last injection, the rats were killed bydecapitation, and their cerebral cortices and hippocampi weredissected and immediately frozen in liquid nitrogen.
Sodium dodecyl sulfate (SDS)–polyacrylamide gelelectrophoresis (PAGE) and immunoblot analysis
Each sample was homogenized in RIPA buffer (150 mMNaCl, 0.1% SDS, 0.5% deoxycholate, 1% NP-40, and 50 mMTris-HCl, pH 8) with a Dounce homogenizer. The proteinlevels of the homogenates were measured using the bicincho-ninic acid protein assay (Smith et al., 1985). Samples from bothcontrol and experimental rats were treated identically.
When the effects of drug treatments were determined, sam-ples from control and experimental groups were examinedwithin single gels. Equal amounts of protein from both controland treated rats, suspended in 23 Laemmli sample buffer [10%2-mercaptoethanol, 4% SDS, 20% glycerol, 120 mM Tris-HCl(pH 6.8), and 0.001% bromophenol blue], were boiled for 3min and subjected to SDS-PAGE (4% stacking gel, 7.5%resolving gel). The electrophoresis was carried out at constantcurrent (40 mA per gel) for 3.5 h with cooling. The separatedproteins were transferred to nitrocellulose membranes at con-stant voltage (20 V) overnight. Following blocking with 5%bovine serum albumin for 2 h, membranes were blotted withthe PDE4A-selective antibody 271 (1:500 dilution in 0.1%bovine serum albumin and 0.5% normal goat serum) for 1 h andsubsequently for another hour with goat anti-rabbit antiserum.Either the ECL (Amersham Life Science, Little Chalfont, U.K.)or alkaline phosphatase (GibcoBRL, Gaithersburg, MD,U.S.A.) method was used for staining.
Antibody C-PDE4A (batch 271) was generated against a12-amino acid peptide (located at amino acid positions 617–628 of PDE4A5) that was contained in the C terminus of bothPDE4A1 and PDE4A5 (McPhee et al., 1995; Ye et al., 1997).
b-Adrenergic receptor densityThe density ofb-adrenergic receptors was measured by
saturation binding of [125I]iodopindolol to membrane prepara-tions of the cerebral cortices and hippocampi (O’Donnell andFrazer, 1985; O’Donnell, 1990). Rat brain tissues were homog-enized in ice-cold homogenization buffer (2 mM HEPES and 1mM EDTA, pH 7.5). The homogenates were centrifuged at40,000g for 20 min at 4°C. The pellets were washed threetimes with homogenization buffer. The final pellets were re-suspended in incubation buffer (140 mM NaCl, 10 mM HEPES,and 10 mM MgCl2, pH 7.5).
The membrane suspensions (30–60mg of protein) wereincubated with [125I]iodopindolol (50–1,000 pM) in the pres-ence and absence of 100mM isoproterenol, at 37°C for 30 min.Samples were immediately filtered through no. 25 glass fiberfilters (Schleicher and Schuell, Keene, NH, U.S.A.). The filterswere washed three times with 6 ml of ice-cold incubationbuffer, and radioactivity was measured at an efficiency of 75%.The density (Bmax) of b-adrenergic receptors and their affinity(KD) for [125I]iodopindolol were determined by nonlinear re-gression analysis (O’Donnell and Frazer, 1985).
Data analysisThe intensities of the bands in immunoblots were analyzed
by densitometry using NIH Image Analysis Software. Differ-ences in intensities between samples from control and treatedrats within individual immunoblots were assessed using Mann–Whitney U tests (Siegel, 1956). ANOVA was applied tob-adrenergic receptor binding data, and the difference betweenthe control group and the treated group was analyzed by Fish-er’s PLSD (Winer, 1971).
MaterialsAntibody C-PDE4A (batch 271) was generously provided
by Prof. Miles D. Houslay (University of Glasgow, Glasgow,Scotland). Goat anti-rabbit IgG antibody was obtained fromPierce (Rockford, IL, U.S.A.). ECL western blotting detectionreagents and Hyperfilm were purchased from Amersham. Al-kaline phosphatase substrates 5-bromo-4-chloro-3-indolylphosphate and nitro blue tetrazolium were purchased fromGibcoBRL. The SeeBlue prestained molecular weight markerset was obtained from Novel Experimental Technology (SanDiego, CA, U.S.A.). [125I]Iodopindolol was purchased fromDuPont-NEN (Boston, MA, U.S.A.). Trazodone was obtainedfrom Research Biochemicals International (Natick, MA,U.S.A.). Fluoxetine was generously provided by Eli Lilly (In-dianapolis). The rest of the chemicals were obtained fromSigma Chemical Co. (St. Louis, MO, U.S.A.) or Fisher Scien-tific (Dallas, TX, U.S.A.).
RESULTS
Effects of repeated antidepressant treatments onPDE4A and b-adrenergic receptors in hippocampus
In rat hippocampal samples, immunoblots revealedtwo major protein bands of molecular masses 109 and 73
FIG. 1. Representative immunoblot images of expression ofPDE4A in hippocampus following repeated treatments with an-tidepressants. Rats received intraperitoneal injections of saline(1 ml/kg twice daily), desipramine (10 mg/kg twice daily; A),phenelzine (5 mg/kg once daily; B), trazodone (10 mg/kg oncedaily; C), or fluoxetine (5 mg/kg once daily; D). After receivinginjections for 14 days, the rats were killed, and brains weredissected for PDE4A immunoblot analysis. Hippocampal ho-mogenates with equal amounts of protein from control andtreated rats were subjected to SDS-PAGE. Separated proteinswere transferred to a nitrocellulose membrane and probed byantibody C-PDE4A.
J. Neurochem., Vol. 74, No. 3, 2000
1258 Y. YE ET AL.

kDa (Fig. 1), as labeled by the PDE4A-selective anti-body. Based on the results of previous studies, these areidentified as PDE4A5 and PDE4A1, respectively(Shakur et al., 1995; Ye et al., 1997).
The immunoblot analyses showed that in hippocam-pus the intensities of the PDE4A bands were signifi-cantly increased following desipramine, phenelzine, andfluoxetine treatment (Figs. 1A, B, and D and 2A, B, andD). By contrast, trazodone treatment failed to alter theexpression of the PDE4A variants (Figs. 1C and 2C).Repeated treatments with desipramine increased the in-tensity of the 109-kDa band by 91% and that of the73-kDa band by 2,125% (Fig. 2A). The phenelzine treat-ment increased the intensities of the 109-kDa band by58% and the 73-kDa band by 384% (Fig. 2B). Thefluoxetine treatment increased the 109-kDa band by112% and the 73-kDa band by 204% (Fig. 2D).
b-Adrenergic receptor binding to [125I]iodopindololusing saturation analysis showed that theBmax was sig-
nificantly decreased following desipramine, phenelzine,and trazodone treatment but not following fluoxetinetreatment. The affinity of the receptors for [125I]iodopin-dolol was not changed by any of the treatments (Table 1).
Effects of repeated antidepressant treatments onPDE4A and b-adrenergic receptorsin cerebral cortex
In rat cerebral cortical samples, immunoblotting re-vealed two major protein bands of molecular masses 109and 73 kDa (Fig. 3), as labeled by antibody C-PDE4A(batch 271). Again, they are known to be PDE4A5 andPDE4A1, respectively (Shakur et al., 1995; Ye et al.,1997). In addition, there was a minor, 102-kDa band thathas been shown previously to be a presently uncharac-terized PDE4A variant termed PDE4AX (Ye et al.,1997).
Following repeated desipramine treatment, the inten-sities of these protein bands in the immunoblots were
FIG. 2. Densitometric analyses of immu-noblots examining the effects of re-peated antidepressant treatments onPDE4A in rat hippocampus: (A) desipra-mine, (B) phenelzine, (C) trazodone, and(D) fluoxetine. Data are mean 6 SEM(bars) values from six rats. *p , 0.05, **p, 0.01, significantly different from thecorresponding control.
TABLE 1. Effects of repeated antidepressant treatments on [125I]iodopindolol binding tob-adrenergic receptors
Control Desipramine Phenelzine Trazodone Fluoxetine
HippocampusBmax (pmol/mg) 35.16 2.0 19.26 1.8a 23.26 3.2a 22.76 2.4a 36.86 1.7KD (nM) 0.186 0.02 0.306 0.12 0.166 0.02 0.166 0.03 0.226 0.01
Cerebral cortexBmax (pmol/mg) 104.06 8.35 59.66 6.9a 102.06 16.9 105.96 15.6 125.56 21.5KD (nM) 0.206 0.02 0.156 0.01 0.216 0.03 0.206 0.01 0.246 0.03
Rats received intraperitoneal injections of normal saline (1 ml/kg, twice daily), desipramine (10 mg/kg, twicedaily), phenelzine (5 mg/kg, once daily), trazodone (10 mg/kg, once daily), or fluoxetine (5 mg/kg, once daily).After injection for 14 days, the rats were killed, and hippocampi and cerebral cortices were dissected for PDE4Aimmunoblot and [125I]iodopindolol binding. Data are mean6 SEM values from three pairs of rats.
a p , 0.05, significantly different from the corresponding control.
J. Neurochem., Vol. 74, No. 3, 2000
1259PDE4 REGULATION BY REPEATED ANTIDEPRESSANT TREATMENTS
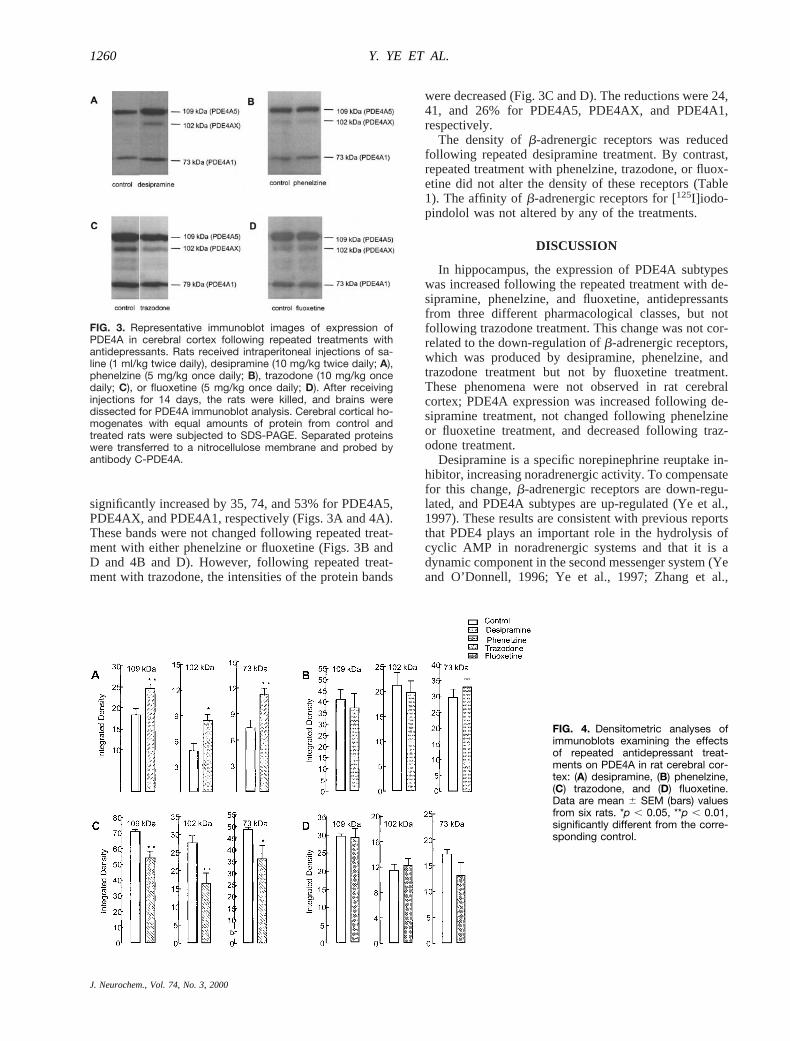
significantly increased by 35, 74, and 53% for PDE4A5,PDE4AX, and PDE4A1, respectively (Figs. 3A and 4A).These bands were not changed following repeated treat-ment with either phenelzine or fluoxetine (Figs. 3B andD and 4B and D). However, following repeated treat-ment with trazodone, the intensities of the protein bands
were decreased (Fig. 3C and D). The reductions were 24,41, and 26% for PDE4A5, PDE4AX, and PDE4A1,respectively.
The density ofb-adrenergic receptors was reducedfollowing repeated desipramine treatment. By contrast,repeated treatment with phenelzine, trazodone, or fluox-etine did not alter the density of these receptors (Table1). The affinity ofb-adrenergic receptors for [125I]iodo-pindolol was not altered by any of the treatments.
DISCUSSION
In hippocampus, the expression of PDE4A subtypeswas increased following the repeated treatment with de-sipramine, phenelzine, and fluoxetine, antidepressantsfrom three different pharmacological classes, but notfollowing trazodone treatment. This change was not cor-related to the down-regulation ofb-adrenergic receptors,which was produced by desipramine, phenelzine, andtrazodone treatment but not by fluoxetine treatment.These phenomena were not observed in rat cerebralcortex; PDE4A expression was increased following de-sipramine treatment, not changed following phenelzineor fluoxetine treatment, and decreased following traz-odone treatment.
Desipramine is a specific norepinephrine reuptake in-hibitor, increasing noradrenergic activity. To compensatefor this change,b-adrenergic receptors are down-regu-lated, and PDE4A subtypes are up-regulated (Ye et al.,1997). These results are consistent with previous reportsthat PDE4 plays an important role in the hydrolysis ofcyclic AMP in noradrenergic systems and that it is adynamic component in the second messenger system (Yeand O’Donnell, 1996; Ye et al., 1997; Zhang et al.,
FIG. 3. Representative immunoblot images of expression ofPDE4A in cerebral cortex following repeated treatments withantidepressants. Rats received intraperitoneal injections of sa-line (1 ml/kg twice daily), desipramine (10 mg/kg twice daily; A),phenelzine (5 mg/kg once daily; B), trazodone (10 mg/kg oncedaily; C), or fluoxetine (5 mg/kg once daily; D). After receivinginjections for 14 days, the rats were killed, and brains weredissected for PDE4A immunoblot analysis. Cerebral cortical ho-mogenates with equal amounts of protein from control andtreated rats were subjected to SDS-PAGE. Separated proteinswere transferred to a nitrocellulose membrane and probed byantibody C-PDE4A.
FIG. 4. Densitometric analyses ofimmunoblots examining the effectsof repeated antidepressant treat-ments on PDE4A in rat cerebral cor-tex: (A) desipramine, (B) phenelzine,(C) trazodone, and (D) fluoxetine.Data are mean 6 SEM (bars) valuesfrom six rats. *p , 0.05, **p , 0.01,significantly different from the corre-sponding control.
J. Neurochem., Vol. 74, No. 3, 2000
1260 Y. YE ET AL.

1999a,b). Similar increases in the expression and activityof PDE4 produced by chronic but not acute antidepres-sant treatment have been described (Suda et al., 1998;Takahashi et al., 1999).
Phenelzine is an irreversible MAO inhibitor, whichalso increases noradrenergic activity. However, follow-ing repeated phenelzine treatment PDE4A is up-regu-lated only in hippocampus but not in cerebral cortex.This coincides with the change ofb-adrenergic density,in that the receptors were down-regulated in hippocam-pus but not changed in cerebral cortex. Similar, region-ally specific effects onb-adrenergic receptors have beenreported (Ordway et al., 1991). An increase in PDE4expression in prefrontal cortex has been reported follow-ing repeated treatment with the MAO inhibitor tranyl-cypromine (Takahashi et al., 1999); this drug seems to bemore efficient at down-regulatingb-adrenergic receptorsin cerebral cortex (McManus et al., 1991). The lack ofchange in PDE4A expression in cerebral cortex follow-ing phenelzine treatment may be due to less extensiveincrease in noradrenergic activity in cerebral cortex thanin hippocampus.
Fluoxetine, a specific serotonin reuptake inhibitor, be-longs to a class of antidepressants distinct from desipra-mine and phenelzine. It is interesting that repeated treat-ment with fluoxetine also up-regulated the expression ofPDE4A in hippocampus, without altering the density ofb-adrenergic receptors. This finding leads to the hypoth-esis that at some point these antidepressants affect com-mon intracellular signal transduction pathways, possiblythrough cross-talk between different neurotransmitter/receptor systems. However, the possibility that PDE4Ahydrolyzes cyclic AMP associated with serotonergic sys-tems and is regulated by changes in serotonergic activitycannot be excluded. This suggestion is supported by thefinding that fluoxetine potentiates the effects of desipra-mine in down-regulatingb-adrenergic receptors, eventhough fluoxetine alone has no effects on these receptors(Goodnough and Baker, 1994). Up-regulation of PDE4 isnot limited to the antidepressant drug treatments; chronicelectroconvulsive seizure also increases PDE4 activityand enhances transcription of PDE4 (Suda et al., 1998;Takahashi et al., 1999). However, this phenomenon maynot be universal to antidepressant treatments becausetrazodone did not increase the expression of PDE4Avariants in the cerebral cortex or hippocampus. It re-mains to be determined whether the different actions oftrazodone on PDE4A subtypes result in a therapeuticdifference.
Affective disorders have been effectively treated byvarious antidepressants. It has always been an intriguingquestion how antidepressants of different pharmacolog-ical classes produce similar clinical effects. Several hy-potheses have been proposed, often involving the down-regulation of postsynaptic receptors, includingb-adren-ergic and serotonergic receptors. There are numerousreports indicating that repeated treatments with norepi-nephrine reuptake inhibitors, MAO inhibitors, and someserotonin reuptake inhibitors decrease the density of
b-adrenergic receptors (Ordway et al., 1991; Paetsch andGreenshaw, 1993; Goodnough and Baker, 1994; Palvi-maki et al., 1994). However, not all antidepressantsdown-regulateb-adrenergic receptors. Some serotoninreuptake inhibitors and atypical antidepressants, such astrazodone, fluoxetine, sertraline, and citalopram, fail todown-regulateb-adrenergic receptors (Ordway et al.,1991; Goodnough and Baker, 1994). As for serotonergicreceptors, repeated treatment of antidepressants from dif-ferent classes also produces different effects. For exam-ple, serotonin 5-HT1A receptors are up-regulated follow-ing repeated treatment with citalopram, imipramine, mi-anserin, and levoprotiline (Klimek et al., 1994), notchanged following fluoxetine treatment (Klimek et al.,1994), and down-regulated following amitriptyline, de-sipramine, doxepin, and trazodone treatment (Akiyoshiet al., 1992). These diverse phenomena are also observedfor serotonin 5-HT2 receptors. Serotonin 5-HT2 receptorsare down-regulated after repeated treatment with citalo-pram, imipramine, and mianserin (Butler et al., 1993;Klimek et al., 1994), not changed following amitriptylinetreatment (Cadogan et al., 1993), and up-regulated fol-lowing levoprotiline, fluoxetine, and electroconvulsiveshock treatment (Butler et al., 1993; Hrdina and Vu,1993; Klimek et al., 1994). Adaptation ofb-adrenergicor serotonergic receptors may not be the ultimate answerto the mechanism of action of antidepressants.
The current finding that antidepressants from differentpharmacological classes produced similar effects onPDE4A expression in hippocampus suggests the possi-bility that intracellular signal transduction pathways maybe a common mechanism mediating antidepressant ac-tivity. This notion is further supported by the finding thatthese antidepressant-induced changes in PDE4A expres-sion are not correlated with down-regulation ofb-adren-ergic receptors. In addition, recent reports show thatelectroconvulsive shock increases expression of PDE4Aand PDE4B in prefrontal cortex (Suda et al., 1998;Takahashi et al., 1999), further supporting this hypo-thesis.
Inhibitors of PDE4 are a novel class of antidepres-sants. They possess antidepressant effects on both clini-cal trials and animal studies (Wachtel and Schneider,1986; Fleischhacker et al., 1992; O’Donnell, 1993).However, their side effects make them unable to beclinically useful. In a previous study, we have shown thatPDE4A as the subtype of PDE4 most susceptible to thechange in noradrenergic activity (Ye et al., 1997; Zhanget al., 1999). The current study revealed that traditionalantidepressants from three different pharmacologicalclasses all up-regulated PDE4A expression in hippocam-pus. These data suggest that the PDE4A subtype mayserve as a selective target for novel antidepressants.
Acknowledgment: This research was supported by grantsand an Independent Scientist Award from the National Instituteof Mental Health. The authors thank Francheska Hamilton andGeneva Evans for excellent technical and secretarial support.
J. Neurochem., Vol. 74, No. 3, 2000
1261PDE4 REGULATION BY REPEATED ANTIDEPRESSANT TREATMENTS

REFERENCES
Akiyoshi J., Tsuchiyama K., Yamada K., Mizobe Y., Nakamura M.,and Nagayama H. (1992) Effect of IAP and chronic antidepressantadministration on the 5HT1A receptor in rat cortical membranes.Prog. Neuropsychopharmacol. Biol. Psychiatry16, 339–349.
Butler M. O., Morinobu S., and Duman R. S. (1993) Chronic electro-convulsive seizures increase the expression of serotonin2 receptormRNA in rat frontal cortex.J. Neurochem.61, 1270–1276.
Cadogan A. K., Marsden C. A., Tulloch I., and Kendall D. A. (1993)Evidence that chronic administration of paroxetine or fluoxetineenhances 5-HT2 receptor function in the brain of the guinea pig.Neuropharmacology32, 249–256.
Challiss R. A. and Nicholson C. D. (1990) Effects of selective phos-phodiesterase inhibition on cyclic AMP hydrolysis in rat cerebralcortical slices.Br. J. Pharmacol.99, 47–52.
Charney D. (1998) Monoamine dysfunction and the pathophysiologyand treatment of depression.J. Clin. Psychiatry59, 11–14.
Fleischhacker W. W., Hinterhuber H., Bauer H., Pflug B., Berner P.,Simhandl C., Wolf R., Gerlach W., Jaklitsch H., Sastre-y-Hernan-dez M., et al. (1992) A multicenter double-blind study of threedifferent doses of the new cAMP-phosphodiesterase inhibitor ro-lipram in patients with major depressive disorder.Neuropsycho-biology 26, 59–64.
Frazer A. and Conway P. (1984) Pharmacologic mechanisms of actionof antidepressants.Psychiatr. Clin. North Am.7, 575–586.
Goodnough D. B. and Baker G. B. (1994) 5-Hydroxytryptamine2 andb-adrenergic receptor regulation in rat brain following chronictreatment with desipramine and fluoxetine alone and in combina-tion. J. Neurochem.62, 2262–2268.
Hrdina P. D. and Vu T. B. (1993) Chronic fluoxetine treatment up-regulates 5-HT uptake sites and 5-HT2 receptors in rat brain: anautoradiographic study.Synapse14, 324–331.
Klimek V., Zak-Knapik J., and Mackowiak M. (1994) Effects ofrepeated treatment with fluoxetine and citalopram, 5-HT uptakeinhibitors, on 5-HT1A and 5-HT2 receptors in the rat brain.J. Psychiatry Neurosci.19, 63–67.
McManus D. J., Mousseau D. D., Paetsch P. R., Wishart T. B., andGreenshaw A. J. (1991) Beta-adrenoceptors and antidepressants:possible 2-phenylethylamine mediation of chronic phenelzine ef-fects.Biol. Psychiatry30, 1122–1130.
McPhee I., Pooley L., Lobban M., Bolger G., and Houslay M. D.(1995) Identification, characterization and regional distribution inbrain of RPDE-6 (RNPDE4A5), a novel splice variant of thePDE4A cyclic AMP phosphodiesterase family.Biochem. J.310,965–974.
O’Donnell J. M. (1990) Behavioral effects of beta adrenergic agonistsand antidepressant drugs after down-regulation of beta-2 adrener-gic receptors by clenbuterol.J. Pharmacol. Exp. Ther.254,147–157.
O’Donnell J. M. (1993) Antidepressant-like effects of rolipram andother inhibitors of cyclic adenosine monophosphate phosphodies-terase on behavior maintained by differential reinforcement of lowresponse rate.J. Pharmacol. Exp. Ther.264,1168–1178.
O’Donnell J. M. and Frazer A. (1985) Effects of clenbuterol andantidepressant drugs on beta adrenergic receptor/N-protein cou-pling in the cerebral cortex of the rat.J. Pharmacol. Exp. Ther.234,30–36.
Ordway G. A., Gambarana C., Tejani-Butt S. M., Areso P., HauptmannM., and Frazer A. (1991) Preferential reduction of binding of125I-iodopindolol to beta-1 adrenoceptors in the amygdala of ratafter antidepressant treatments.J. Pharmacol. Exp. Ther.257,681–690.
Paetsch P. R. and Greenshaw A. J. (1993) Effects of chronic antide-pressant treatment on beta-adrenoceptor subtype binding in the ratcerebral cortex and cerebellum.Mol. Chem. Neuropathol.20,21–31.
Palvimaki E. P., Laakso A., Kuoppamaki M., Syvalahti E., and HietalaJ. (1994) Up-regulation of beta 1-adrenergic receptors in rat brainafter chronic citalopram and fluoxetine treatments.Psychophar-macology (Berl.)115,543–546.
Sette C., Vicini E., and Conti M. (1994) The ratPDE3/IVd phospho-diesterase gene codes for multiple proteins differentially activatedby cAMP-dependent protein kinaseJ. Biol. Chem.269, 18271–18274. [Published erratum appears inJ. Biol. Chem.269, 20806(1994)].
Shakur Y., Wilson M., Pooley L., Lobban M., Griffiths S. L., CampbellA. M., Beattie J., Daly C., and Houslay M. D. (1995) Identificationand characterization of the type-IVA cyclic AMP-specific phos-phodiesterase RD1 as a membrane-bound protein expressed incerebellum.Biochem. J.306,801–809.
Siegel S. (1956) The case of two independent samples, inNonpara-metric Statistics for the Behavioral Sciences(Morgan C. T., ed),pp. 95–158. McGraw-Hill, New York.
Smith P. K., Krohn R. I., Hermanson G. T., Mallia A. K., Gartner F. H.,Provenzano M. D., Fujimoto E. K., Goeke N. M., Olson B. J., andKlenk D. C. (1985) Measurement of protein using bicinchoninicacid. Anal. Biochem.150, 76–85. [Published erratum appears inAnal. Biochem.163,279 (1987)].
Suda S., Nibuya M., Ishiguro T., and Suda H. (1998) Transcriptionaland translational regulation of phosphodiesterase type IVisozymes in rat brain by electroconvulsive seizure and antidepres-sant drug treatment.J. Neurochem.71, 1554–1563.
Swinnen J., Joseph D., and Conti M. (1989) The mRNA encoding ahigh-affinity cAMP phosphodiesterase is regulated by hormonesand cAMP.Proc. Natl. Acad. Sci. USA86, 8197–8201.
Takahashi M., Terwilliger R., Lane C., Mezes P., Conti M., and DumanR. (1999) Chronic antidepressant administration increases theexpression of cAMP-specific phosphodiesterase 4A and 4B iso-forms.J. Neurosci.19, 610–618.
Torphy T. J., Zhou H. L., and Cieslinski L. B. (1992) Stimulation ofbeta adrenoceptors in a human monocyte cell line (U937) up-regulates cyclic AMP-specific phosphodiesterase activity.J. Phar-macol. Exp. Ther.263,1195–1205.
Wachtel H. and Schneider H. H. (1986) Rolipram, a novel antidepres-sant drug, reverses the hypothermia and hypokinesia of monoam-ine-depleted mice by an action beyond postsynaptic monoaminereceptors.Neuropharmacology25, 1119–1126.
Whalin M. E., Garrett R. L., Jr., Thompson W. J., and Strada S. J.(1988) Correlation of cell-free brain cyclic nucleotide phosphodi-esterase activities to cyclic AMP decay in intact brain slices.Second Messengers Phosphoproteins12, 311–325.
Winer B. (1971) Inference with respect to means and variance, inStatistical Principles in Experimental Design(Maytham W., Sha-piro A., and Stern J., eds), pp. 4–54. McGraw-Hill, New York.
Ye Y. and O’Donnell J. M. (1996) Diminished noradrenergic stimula-tion reduces the activity of rolipram-sensitive, high-affinity cyclicAMP phosphodiesterase in rat cerebral cortex.J. Neurochem.66,1894–1902.
Ye Y., Conti M., Houslay M. D., Farooqui S. M., Chen M., andO’Donnell J. M. (1997) Noradrenergic activity differentially reg-ulates the expression of rolipram-sensitive, high-affinity cyclicAMP phosphodiesterase (PDE4) in rat brain.J. Neurochem.69,2397–2404.
Zhang K., Farooqui S. M., and O’Donnell J. M. (1999a) Ontogeny ofrolipram-sensitive, low-Km, cyclic AMP-specific phosphodiester-ase in rat brain.Brain Res. Dev. Brain Res.112,11–19.
Zhang K., Farooqui S. M., Jackson K. T., and O’Donnell J. M. (1999b)Effects of noradrenergic lesions on the development of rolipram-sensitive, low-Km, cyclic AMP specific phosphodiesterase in ratbrain.Brain Res. Dev. Brain Res.116,181–189.
J. Neurochem., Vol. 74, No. 3, 2000
1262 Y. YE ET AL.


















