
Effects of high atmospheric CO2 concentration on root hydraulic conductivity of conifers depend on...
7
Effects of high atmospheric CO 2 concentration on root hydraulic conductivity of conifers depend on species identity and inorganic nitrogen source Tobias Gebauer, Hormoz BassiriRad * University of Illinois at Chicago, Department of Biological Sciences, 840 West Taylor Street, Chicago, IL 60607, United States article info Article history: Received 20 May 2011 Received in revised form 10 August 2011 Accepted 15 August 2011 Keywords: Ammonium Elevated CO 2 Nitrate Root hydraulic conductivity N deposition abstract We examined root hydraulic conductivity (L p ) responses of one-year-old seedlings of four conifers to the combined effects of elevated CO 2 and inorganic nitrogen (N) sources. We found marked interspecific differences in L p responses to high CO 2 ranging from a 37% increase in P. abies to a 27% decrease in P. menziesii, but these effects depended on N source. The results indicate that CO 2 effects on root water transport may be coupled to leaf area responses under nitrate (NO 3 ), but not ammonium (NH 4 þ ) dominated soils. To our knowledge, this is the first study that highlights the role of inorganic N source and species identity as critical factors that determine plant hydraulic responses to rising atmospheric CO 2 levels. The results have important implications for understanding root biology in a changing climate and for models designed to predict feedbacks between rising atmospheric CO 2 , N deposition, and ecohydrology. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction The projected rise in atmospheric CO 2 concentration will have serious consequences for plant and ecosystem processes (Ainsworth and Long, 2005; Bazzaz, 1990; Norby et al., 1999; Wullschleger et al., 2002), but the magnitude and direction of many of these processes are still the focus of considerable debate. One such process is the flow of water within the soileplante atmosphere-continuum (SPAC). While there is general agreement that stomatal conductance (g s ) is often reduced in response to CO 2 enrichment (Drake et al., 1997; Field et al.,1995; Medlyn et al., 2001; Saxe et al.,1998; Wullschleger et al., 2002), the consequences for the SPAC or whole plant water loss is less clear (Domec et al., 2010; Wullschleger et al., 2002). In fact, CO 2 -induced reduction in g s does not always result in similar changes in whole plant water fluxes (Bhattacharya et al., 1994; Leakey et al., 2009; Schäfer et al., 2002; Tricker et al., 2009). This apparent divergence between g s and whole plant responses to high CO 2 can arise if changes in g s are offset by changes in leaf area and/or conductivities in other parts of the SPAC (Domec et al., 2009). There is, however, a paucity of information on how resistances to water flow in other parts of the plant respond to rising atmospheric CO 2 . A substantial proportion of resistance to water flow between the soil and the leaf is in the root system. In fact, it has been argued that root hydraulic conductivity (L p ) may be the largest component of resistance to water flow inside the plant (Nardini and Tyree, 1999; Nobel and Cui, 1992; North and Peterson, 2005; Saliendra and Meinzer, 1989; Tsuda and Tyree,1997; Tyree et al., 1998). Yet, the direct effect of CO 2 enrichment on root hydraulic conductivity has rarely been examined. A handful of studies that have addressed this important physiological response to increased atmospheric CO 2 concentration have produced inconsistent patterns of responses ranging from increase (Atwell et al., 2009) to decrease (Bunce, 1996; Huxman et al., 1999) to no changes (Bunce, 1996; Huxman et al., 1999; Sarker and Hara, 2009). The meager yet diverse pattern of L p responses to high CO 2 reported in the literature may have resulted from varying experimental protocols and/or use of widely differing functional groups of plants. Therefore, we are currently unable to separate species dependent responses of L p to CO 2 from those caused by different experimental protocols. Here, we propose to examine the L p responses of four conifer seedlings to CO 2 enrichment including Picea pungens, P. abies, Pseudotsuga menziesii, and Abies fraseri. These species represent a relatively wide range of genetic backgrounds; hence, any potential differ- ences will signify a species-specific nature of the responses. An important consideration in examining potential responses of plants and ecosystems to rising CO 2 is the confounding effect of other environmental factors. Nitrogen availability is arguably one of the most important factors that can significantly interact with * Corresponding author. E-mail address: [email protected] (H. BassiriRad). Contents lists available at SciVerse ScienceDirect Environmental Pollution journal homepage: www.elsevier.com/locate/envpol 0269-7491/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.envpol.2011.08.021 Environmental Pollution 159 (2011) 3455e3461
-
Upload
tobias-gebauer -
Category
Documents
-
view
213 -
download
1
Transcript of Effects of high atmospheric CO2 concentration on root hydraulic conductivity of conifers depend on...

at SciVerse ScienceDirect
Environmental Pollution 159 (2011) 3455e3461
Contents lists available
Environmental Pollution
journal homepage: www.elsevier .com/locate/envpol
Effects of high atmospheric CO2 concentration on root hydraulic conductivityof conifers depend on species identity and inorganic nitrogen source
Tobias Gebauer, Hormoz BassiriRad*
University of Illinois at Chicago, Department of Biological Sciences, 840 West Taylor Street, Chicago, IL 60607, United States
a r t i c l e i n f o
Article history:Received 20 May 2011Received in revised form10 August 2011Accepted 15 August 2011
Keywords:AmmoniumElevated CO2
NitrateRoot hydraulic conductivityN deposition
* Corresponding author.E-mail address: [email protected] (H. BassiriRad).
0269-7491/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.envpol.2011.08.021
a b s t r a c t
We examined root hydraulic conductivity (Lp) responses of one-year-old seedlings of four conifers to thecombined effects of elevated CO2 and inorganic nitrogen (N) sources. We found marked interspecificdifferences in Lp responses to high CO2 ranging from a 37% increase in P. abies to a 27% decrease inP. menziesii, but these effects depended on N source. The results indicate that CO2 effects on root watertransport may be coupled to leaf area responses under nitrate (NO3
�), but not ammonium (NH4þ)
dominated soils. To our knowledge, this is the first study that highlights the role of inorganic N sourceand species identity as critical factors that determine plant hydraulic responses to rising atmospheric CO2
levels. The results have important implications for understanding root biology in a changing climate andfor models designed to predict feedbacks between rising atmospheric CO2, N deposition, andecohydrology.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
The projected rise in atmospheric CO2 concentration will haveserious consequences for plant and ecosystem processes(Ainsworth and Long, 2005; Bazzaz, 1990; Norby et al., 1999;Wullschleger et al., 2002), but the magnitude and direction ofmany of these processes are still the focus of considerable debate.One such process is the flow of water within the soileplanteatmosphere-continuum (SPAC). While there is general agreementthat stomatal conductance (gs) is often reduced in response to CO2enrichment (Drake et al.,1997; Field et al.,1995;Medlyn et al., 2001;Saxe et al.,1998;Wullschleger et al., 2002), the consequences for theSPAC or whole plant water loss is less clear (Domec et al., 2010;Wullschleger et al., 2002). In fact, CO2-induced reduction in gsdoes not always result in similar changes inwhole plantwaterfluxes(Bhattacharya et al., 1994; Leakey et al., 2009; Schäfer et al., 2002;Tricker et al., 2009). This apparent divergence between gs andwhole plant responses to high CO2 can arise if changes in gs areoffset by changes in leaf area and/or conductivities in other parts ofthe SPAC (Domec et al., 2009). There is, however, a paucity ofinformation on how resistances to water flow in other parts of theplant respond to rising atmospheric CO2.
All rights reserved.
A substantial proportion of resistance towater flow between thesoil and the leaf is in the root system. In fact, it has been argued thatroot hydraulic conductivity (Lp) may be the largest component ofresistance to water flow inside the plant (Nardini and Tyree, 1999;Nobel and Cui, 1992; North and Peterson, 2005; Saliendra andMeinzer, 1989; Tsuda and Tyree, 1997; Tyree et al., 1998). Yet, thedirect effect of CO2 enrichment on root hydraulic conductivity hasrarely been examined. A handful of studies that have addressed thisimportant physiological response to increased atmospheric CO2concentration have produced inconsistent patterns of responsesranging from increase (Atwell et al., 2009) to decrease (Bunce,1996; Huxman et al., 1999) to no changes (Bunce, 1996; Huxmanet al., 1999; Sarker and Hara, 2009). The meager yet diversepattern of Lp responses to high CO2 reported in the literature mayhave resulted from varying experimental protocols and/or use ofwidely differing functional groups of plants. Therefore, we arecurrently unable to separate species dependent responses of Lp toCO2 from those caused by different experimental protocols. Here,we propose to examine the Lp responses of four conifer seedlings toCO2 enrichment including Picea pungens, P. abies, Pseudotsugamenziesii, and Abies fraseri. These species represent a relativelywide range of genetic backgrounds; hence, any potential differ-ences will signify a species-specific nature of the responses.
An important consideration in examining potential responses ofplants and ecosystems to rising CO2 is the confounding effect ofother environmental factors. Nitrogen availability is arguably oneof the most important factors that can significantly interact with

T. Gebauer, H. BassiriRad / Environmental Pollution 159 (2011) 3455e34613456
plant responses to CO2 (Finzi et al., 2006; Luo et al., 2004). Nitrogenavailability can affect CO2 responses of plants through a number ofcommonly accepted mechanisms, e.g., source:sink relations(Barrett and Gifford, 1995; Prior et al., 1997), production of newmeristematic tissue (Pritchard et al., 1999), acclimation or down-regulation of photosynthetic machinery (Barrett and Gifford, 1995;Farage et al., 1998; Pettersson et al., 1994; Rogers and Humphries,2000; Rogers et al., 1998; Wolfe et al., 1998), and stomatalconductance (McDonald et al., 2002). Nitrogen is, however,a resource that can specifically affect root hydraulic properties(Radin, 1990; Radin and Boyer, 1982; Trubat et al., 2006), but in thatcapacity, its interactions with CO2 enrichment have not previouslybeen examined. Emerging literature provides a convincing case thatLp is highly sensitive to N availability and that this sensitivity isspecific to NO3
� but not NH4þ (Adler et al., 1996; Gloser et al., 2007;
Gorska et al., 2008a, 2010; Guo et al., 2007; Schulze-Till et al.,2009). Therefore, another objective of this study is to examine CO2responses of Lp in the target species to different inorganic N sources.
It is worth noting that the focus on inorganic N form goesbeyond academic interest, i.e., there are reasons to believe that inmany native systems relative plant preference and/or availability ofN forms may shift dramatically in a future climate. Native ecosys-tems are increasingly exposed to chronic anthropogenic N depo-sition (Galloway, 1998; Galloway et al., 2008; Holland et al., 2005;Vitousek et al., 1997), and it is well recognized that, dependingupon the prevailing sources of N pollutants, the deposited materialcan disproportionately be one form or another. For example,industrial-based N pollution tends to be almost exclusively NO3
�,whereas farming-based N deposition is highly enriched in NH4
þ
(Holland et al., 2005; Lovett, 1994; Stulen et al., 1998). Additionally,BassiriRad and colleagues (BassiriRad, 2000; BassiriRad et al., 1996,1997, 2001; Zerihun and BassiriRad, 2001) have shown thatelevated CO2 can result in a species-specific shift in preference forNH4
þ vs. NO3�. The choice of conifers in this study is justified because
these species adapted to soils that are characterized with low pHand high levels of NH4
þ compared to NO3� (Rothstein and Cregg,
2005; Stark and Hart, 1997). However, many of these systemsare currently undergoing increasing atmospheric NO3
� inputs(Aber et al., 1989; Schulze, 1989).
Table 1Three-way analysis of variances (ANOVA) of species, CO2 and N form effects andtheir interactive terms on LpDW. Statistical significance (p< 0.05) are shown in boldtypeface.
Sources DF F p
Species 3 25.60 <0.001CO2 2 0.85 0.43N form 2 18.94 <0.001CO2�N form 4 6.74 <0.001CO2� species 6 8.87 <0.001N form� species 6 6.40 <0.001CO2�N form� species 12 2.00 0.03
2. Material and methods
2.1. Plant growth and conditions
One-year-old seedlings of Colorado spruce (Picea pungens), Norway spruce (Piceaabies), Douglas-fir (Pseudotsuga menziesii), and Fraser fir (Abies fraseri) werepurchased from Lawyers Nursery, Inc. (Montana, USA). Seedlings were initiallyplaced into 40 L Rubbermaid containers filled with vigorously aerated half-strengthcomplete Hoagland solution for four months. The solutionwas replaced weekly and,at the end of this period, healthy seedlings with a significant new root and shootgrowth were used for the experiments. Individual seedlings were transplanted into1 L rocket pots (Stuewe & Son, Tangent, Oregon, USA) containing reviver washedsand. To minimize the effect of ontogenic variability, we chose seedlings of even sizeand rooting vigor within each species. The potted seedlings were then transferredinto one of the three walk-in growth chambers set at 400 (hereon referred to asambient), 600, or 800 (�25) mL L�1 CO2 concentrations. Thermal and photo periodswere identical for all treatments (16/8 h light, 26 �C/18 �C day/night, relativehumidity (RH) 50/80�10% at day/night, and photon flux density at the top of eachseedling was roughly 500 mmolm�2 s�1). The variability in RH reported here indi-cates within-chamber differences in humidity resulting from short-term deviationfrom the set points (often less than an hour) immediately following dailywatering orfertilizing. There were no detectable differences in RH and temperature regimesbetween the chambers; hence, vapor pressure deficits were similar between the CO2
treatments. Seedlingswere grown under these conditions for the next sevenmonths.All seedlings were fertilized daily with a nitrogen-free half-strength Hoaglandsolution (pH adjusted to 6.0) containing all essential macro andmicronutrients (Laneand BassiriRad, 2002). There were three N treatments that were added to the N-freebase solution to obtain a final concentration of 5 mMof N provided as either NH4
þeN,NO3
�eN, or NH4NO3. Each pot was irrigated in the morning with roughly 100 mL oftheir respective N treatment solution and rinsed to free drainage with de-ionized
water in the evening to avoid salt accumulation. Pots were rotated inside thechamber weekly so as to minimize differences from one spot to another.
2.2. Root hydraulic conductivity measurements
Root hydraulic conductivities of intact whole root systems were estimated asdescribed by Gorska et al. (2008a, 2008b). Briefly, we measured exudation rates ofdetopped whole root systems at various hydrostatic pressure gradients. Eachseedling was gently removed from the growing medium and rinsed clean of theadhered sand. The shoot was then cut under water at about 2 cm above the root/shoot junction and the detopped stump was connected by tygon tubing to a beakerplaced on an automatically recording digital analytical balance (model CPA225D,Sartorius, Göttingen, Germany). The root system was submerged in well-aerated,degassed, de-ionized water in a pressure-stable PVC tube that was connected tobuilding air capable of generating up to 0.5 MPa. A pressure transducer simulta-neously monitored and recorded the applied pressure and the weight changes dueto sap flow in 10 s intervals. Pressure inside the root chamber was increased step-wise with 35 kPa increments between 70 and 275 kPa. At each increment, wemade sure that a steady flow rate was reached and maintained for 2 min. We usedthe slope of the linear portion of the flow rate J (mg s�1) vs. applied pressure P (MPa)to calculate Lp expressed on a root dry weight basis (LpDW, in mg s�1MPa�1 g�1 DW).Preliminary results had shown that, in all species tested here, the linear portion of Jvs. P was achieved at external pressures� 100 kPa. We also calculated Lp on a leafarea (LA) basis (LpLA, in mg s�1MPa�1m�2 LA) as described by Tyree et al. (1998). Todo this, we removed and determined the one-sided needle area using a LI-3100 leafarea meter (LI-COR, Lincoln, Nebraska, USA) (Becker et al., 1999). Admittedly, one-sided needle area is considerably lower than total LA, but it can provide reliableestimates of relative changes in total LA.
2.3. Data analysis
Statistical analyses were performed using the GLM (general linear models) andLSMEANS (least square sums of the means) procedures of the SAS statistical soft-ware package (SAS 9.13, SAS Institute, Cary, NC, USA). Two- and three-way analysesof variances (ANOVA) were used to determine statistical differences in the main andinteractive terms using type II sum of squares (Lansgrud, 2003) followed by post-hocpair-wise comparisons of the means for CO2 and nitrogen form effects on Lp. Thelevel of correlations between Lp data expressed on root DWand LA bases were testedby regression analysis using SigmaPlot (version 10.0, Systat Software, San Jose, CA,USA). A linear model provided the best fit for these regressions.
3. Results
A three-way analysis of variance (ANOVA) revealed that speciesdiffered significantly in terms of their Lp responses to CO2 enrich-ment and inorganic N form. Using ambient CO2 and NH4NO3treatment for the purpose of relative comparison among species,we found that Lp in P. pungens, P. menziesii, and A. fraseri weresimilar, but that they were higher than those of P. abies by roughlytwofold. Across species, CO2 did not significantly affect Lp (Table 1,p¼ 0.43), but there was a significant CO2 by species interaction(p< 0.001), indicating that the effect of CO2 on Lp is highly species-specific. Within species, CO2 always had a significant effect on Lp(two-way ANOVA in Fig. 1), but the direction of this effect was notconsistent among species. For example, across N form treatments,CO2 enrichment significantly increased Lp by an average of 37% and17% in P. abies and P. pungens, respectively, but significantlydecreased it in P. menziesii and A. fraseri by an average of 27% and22%, respectively (Figs. 1 and 2). Overall, there appears to be
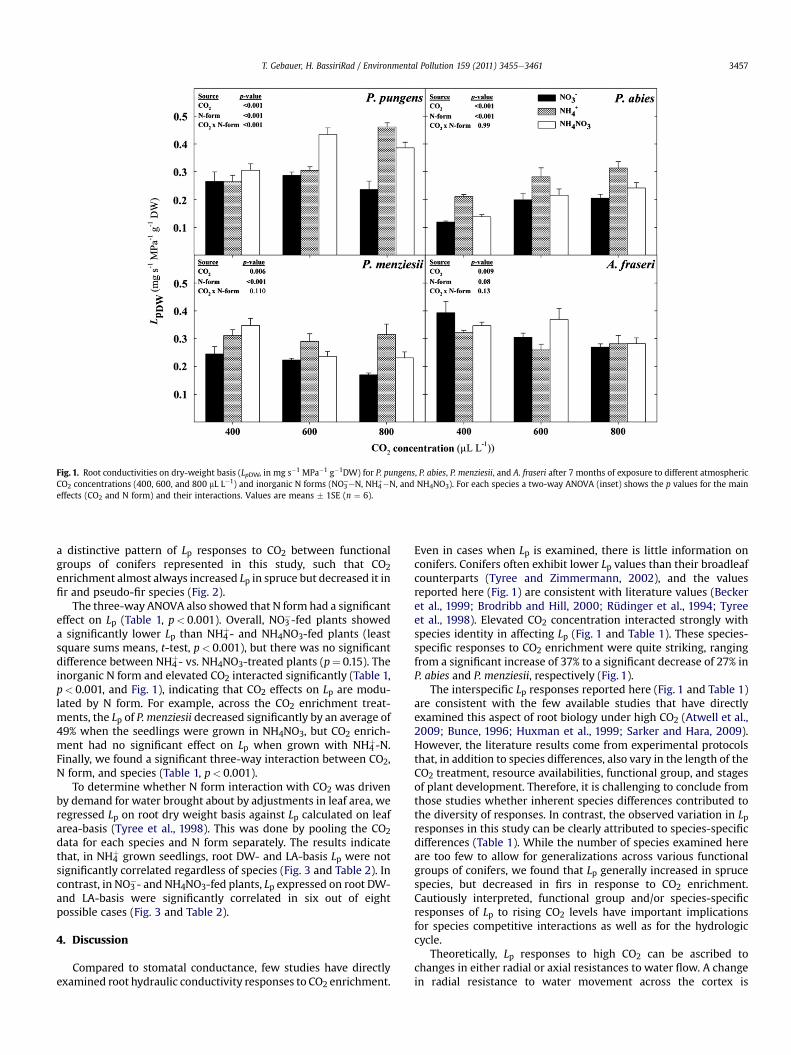
Fig. 1. Root conductivities on dry-weight basis (LpDW, in mg s�1 MPa�1 g�1DW) for P. pungens, P. abies, P. menziesii, and A. fraseri after 7 months of exposure to different atmosphericCO2 concentrations (400, 600, and 800 mL L�1) and inorganic N forms (NO3
eeN, NH4þ�N, and NH4NO3). For each species a two-way ANOVA (inset) shows the p values for the main
effects (CO2 and N form) and their interactions. Values are means � 1SE (n ¼ 6).
T. Gebauer, H. BassiriRad / Environmental Pollution 159 (2011) 3455e3461 3457
a distinctive pattern of Lp responses to CO2 between functionalgroups of conifers represented in this study, such that CO2enrichment almost always increased Lp in spruce but decreased it infir and pseudo-fir species (Fig. 2).
The three-way ANOVA also showed that N form had a significanteffect on Lp (Table 1, p< 0.001). Overall, NO3
�-fed plants showeda significantly lower Lp than NH4
þ- and NH4NO3-fed plants (leastsquare sums means, t-test, p< 0.001), but there was no significantdifference between NH4
þ- vs. NH4NO3-treated plants (p¼ 0.15). Theinorganic N form and elevated CO2 interacted significantly (Table 1,p< 0.001, and Fig. 1), indicating that CO2 effects on Lp are modu-lated by N form. For example, across the CO2 enrichment treat-ments, the Lp of P. menziesii decreased significantly by an average of49% when the seedlings were grown in NH4NO3, but CO2 enrich-ment had no significant effect on Lp when grown with NH4
þ-N.Finally, we found a significant three-way interaction between CO2,N form, and species (Table 1, p< 0.001).
To determine whether N form interaction with CO2 was drivenby demand for water brought about by adjustments in leaf area, weregressed Lp on root dry weight basis against Lp calculated on leafarea-basis (Tyree et al., 1998). This was done by pooling the CO2data for each species and N form separately. The results indicatethat, in NH4
þ grown seedlings, root DW- and LA-basis Lp were notsignificantly correlated regardless of species (Fig. 3 and Table 2). Incontrast, in NO3
�- and NH4NO3-fed plants, Lp expressed on root DW-and LA-basis were significantly correlated in six out of eightpossible cases (Fig. 3 and Table 2).
4. Discussion
Compared to stomatal conductance, few studies have directlyexamined root hydraulic conductivity responses to CO2 enrichment.
Even in cases when Lp is examined, there is little information onconifers. Conifers often exhibit lower Lp values than their broadleafcounterparts (Tyree and Zimmermann, 2002), and the valuesreported here (Fig. 1) are consistent with literature values (Beckeret al., 1999; Brodribb and Hill, 2000; Rüdinger et al., 1994; Tyreeet al., 1998). Elevated CO2 concentration interacted strongly withspecies identity in affecting Lp (Fig. 1 and Table 1). These species-specific responses to CO2 enrichment were quite striking, rangingfrom a significant increase of 37% to a significant decrease of 27% inP. abies and P. menziesii, respectively (Fig. 1).
The interspecific Lp responses reported here (Fig. 1 and Table 1)are consistent with the few available studies that have directlyexamined this aspect of root biology under high CO2 (Atwell et al.,2009; Bunce, 1996; Huxman et al., 1999; Sarker and Hara, 2009).However, the literature results come from experimental protocolsthat, in addition to species differences, also vary in the length of theCO2 treatment, resource availabilities, functional group, and stagesof plant development. Therefore, it is challenging to conclude fromthose studies whether inherent species differences contributed tothe diversity of responses. In contrast, the observed variation in Lpresponses in this study can be clearly attributed to species-specificdifferences (Table 1). While the number of species examined hereare too few to allow for generalizations across various functionalgroups of conifers, we found that Lp generally increased in sprucespecies, but decreased in firs in response to CO2 enrichment.Cautiously interpreted, functional group and/or species-specificresponses of Lp to rising CO2 levels have important implicationsfor species competitive interactions as well as for the hydrologiccycle.
Theoretically, Lp responses to high CO2 can be ascribed tochanges in either radial or axial resistances to water flow. A changein radial resistance to water movement across the cortex is
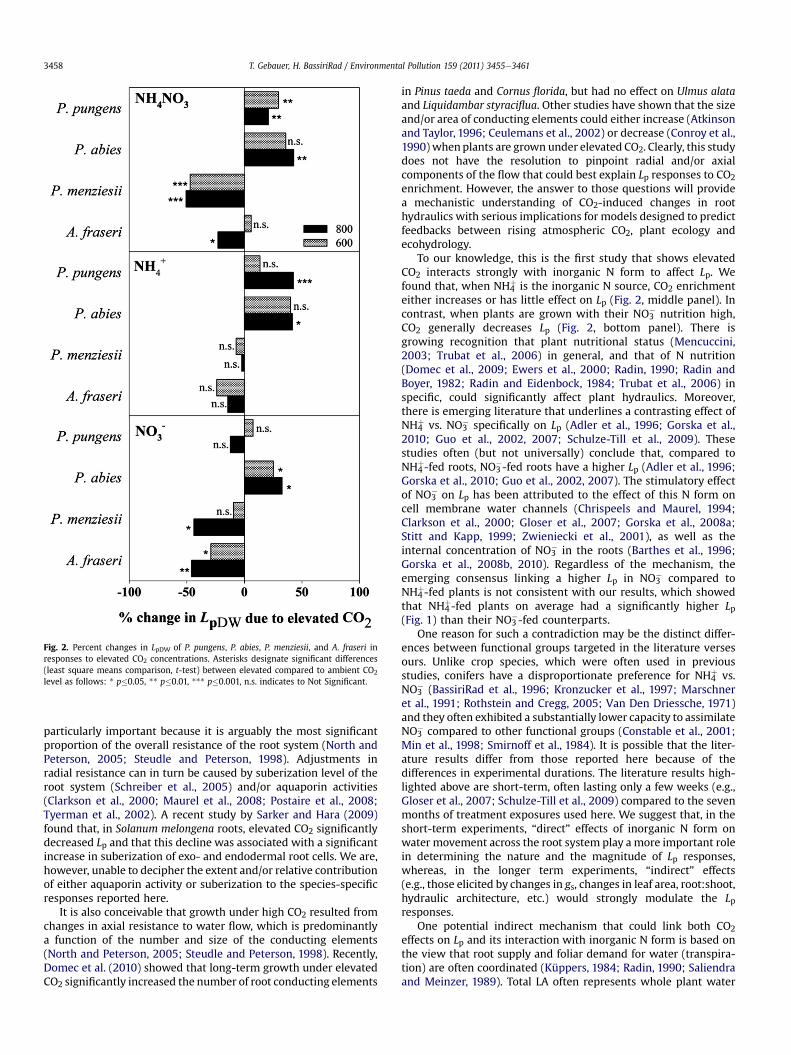
Fig. 2. Percent changes in LpDW of P. pungens, P. abies, P. menziesii, and A. fraseri inresponses to elevated CO2 concentrations. Asterisks designate significant differences(least square means comparison, t-test) between elevated compared to ambient CO2
level as follows: * p�0.05, ** p�0.01, *** p�0.001, n.s. indicates to Not Significant.
T. Gebauer, H. BassiriRad / Environmental Pollution 159 (2011) 3455e34613458
particularly important because it is arguably the most significantproportion of the overall resistance of the root system (North andPeterson, 2005; Steudle and Peterson, 1998). Adjustments inradial resistance can in turn be caused by suberization level of theroot system (Schreiber et al., 2005) and/or aquaporin activities(Clarkson et al., 2000; Maurel et al., 2008; Postaire et al., 2008;Tyerman et al., 2002). A recent study by Sarker and Hara (2009)found that, in Solanum melongena roots, elevated CO2 significantlydecreased Lp and that this decline was associated with a significantincrease in suberization of exo- and endodermal root cells. We are,however, unable to decipher the extent and/or relative contributionof either aquaporin activity or suberization to the species-specificresponses reported here.
It is also conceivable that growth under high CO2 resulted fromchanges in axial resistance to water flow, which is predominantlya function of the number and size of the conducting elements(North and Peterson, 2005; Steudle and Peterson, 1998). Recently,Domec et al. (2010) showed that long-term growth under elevatedCO2 significantly increased the number of root conducting elements
in Pinus taeda and Cornus florida, but had no effect on Ulmus alataand Liquidambar styraciflua. Other studies have shown that the sizeand/or area of conducting elements could either increase (Atkinsonand Taylor, 1996; Ceulemans et al., 2002) or decrease (Conroy et al.,1990) whenplants are grown under elevated CO2. Clearly, this studydoes not have the resolution to pinpoint radial and/or axialcomponents of the flow that could best explain Lp responses to CO2enrichment. However, the answer to those questions will providea mechanistic understanding of CO2-induced changes in roothydraulics with serious implications for models designed to predictfeedbacks between rising atmospheric CO2, plant ecology andecohydrology.
To our knowledge, this is the first study that shows elevatedCO2 interacts strongly with inorganic N form to affect Lp. Wefound that, when NH4
þ is the inorganic N source, CO2 enrichmenteither increases or has little effect on Lp (Fig. 2, middle panel). Incontrast, when plants are grown with their NO3
� nutrition high,CO2 generally decreases Lp (Fig. 2, bottom panel). There isgrowing recognition that plant nutritional status (Mencuccini,2003; Trubat et al., 2006) in general, and that of N nutrition(Domec et al., 2009; Ewers et al., 2000; Radin, 1990; Radin andBoyer, 1982; Radin and Eidenbock, 1984; Trubat et al., 2006) inspecific, could significantly affect plant hydraulics. Moreover,there is emerging literature that underlines a contrasting effect ofNH4
þ vs. NO3� specifically on Lp (Adler et al., 1996; Gorska et al.,
2010; Guo et al., 2002, 2007; Schulze-Till et al., 2009). Thesestudies often (but not universally) conclude that, compared toNH4
þ-fed roots, NO3�-fed roots have a higher Lp (Adler et al., 1996;
Gorska et al., 2010; Guo et al., 2002, 2007). The stimulatory effectof NO3
� on Lp has been attributed to the effect of this N form oncell membrane water channels (Chrispeels and Maurel, 1994;Clarkson et al., 2000; Gloser et al., 2007; Gorska et al., 2008a;Stitt and Kapp, 1999; Zwieniecki et al., 2001), as well as theinternal concentration of NO3
� in the roots (Barthes et al., 1996;Gorska et al., 2008b, 2010). Regardless of the mechanism, theemerging consensus linking a higher Lp in NO3
� compared toNH4
þ-fed plants is not consistent with our results, which showedthat NH4
þ-fed plants on average had a significantly higher Lp(Fig. 1) than their NO3
�-fed counterparts.One reason for such a contradiction may be the distinct differ-
ences between functional groups targeted in the literature versesours. Unlike crop species, which were often used in previousstudies, conifers have a disproportionate preference for NH4
þ vs.NO3
� (BassiriRad et al., 1996; Kronzucker et al., 1997; Marschneret al., 1991; Rothstein and Cregg, 2005; Van Den Driessche, 1971)and they often exhibited a substantially lower capacity to assimilateNO3
� compared to other functional groups (Constable et al., 2001;Min et al., 1998; Smirnoff et al., 1984). It is possible that the liter-ature results differ from those reported here because of thedifferences in experimental durations. The literature results high-lighted above are short-term, often lasting only a few weeks (e.g.,Gloser et al., 2007; Schulze-Till et al., 2009) compared to the sevenmonths of treatment exposures used here. We suggest that, in theshort-term experiments, “direct” effects of inorganic N form onwater movement across the root system play a more important rolein determining the nature and the magnitude of Lp responses,whereas, in the longer term experiments, “indirect” effects(e.g., those elicited by changes in gs, changes in leaf area, root:shoot,hydraulic architecture, etc.) would strongly modulate the Lpresponses.
One potential indirect mechanism that could link both CO2effects on Lp and its interaction with inorganic N form is based onthe view that root supply and foliar demand for water (transpira-tion) are often coordinated (Küppers, 1984; Radin, 1990; Saliendraand Meinzer, 1989). Total LA often represents whole plant water
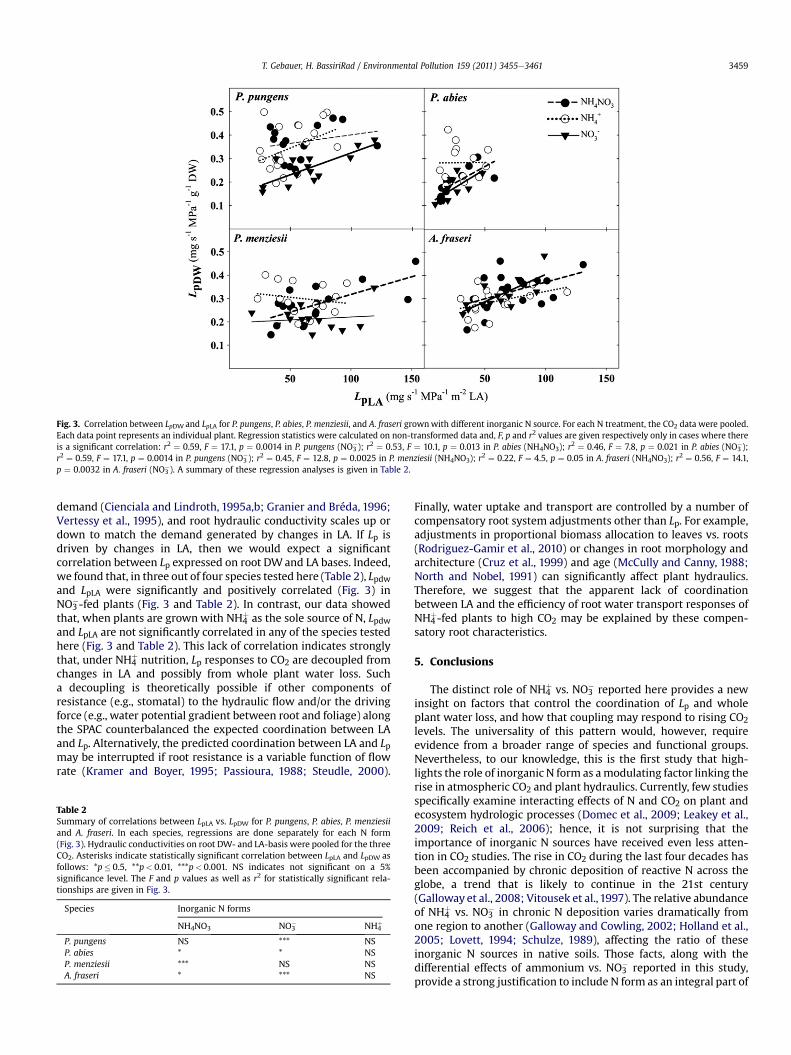
Fig. 3. Correlation between LpDW and LpLA for P. pungens, P. abies, P. menziesii, and A. fraseri grownwith different inorganic N source. For each N treatment, the CO2 data were pooled.Each data point represents an individual plant. Regression statistics were calculated on non-transformed data and, F, p and r2 values are given respectively only in cases where thereis a significant correlation: r2 ¼ 0.59, F ¼ 17.1, p ¼ 0.0014 in P. pungens (NO3
�); r2 ¼ 0.53, F ¼ 10.1, p ¼ 0.013 in P. abies (NH4NO3); r2 ¼ 0.46, F ¼ 7.8, p ¼ 0.021 in P. abies (NO3�);
r2 ¼ 0.59, F ¼ 17.1, p ¼ 0.0014 in P. pungens (NO3�); r2 ¼ 0.45, F ¼ 12.8, p ¼ 0.0025 in P. menziesii (NH4NO3); r2 ¼ 0.22, F ¼ 4.5, p ¼ 0.05 in A. fraseri (NH4NO3); r2 ¼ 0.56, F ¼ 14.1,
p ¼ 0.0032 in A. fraseri (NO3�). A summary of these regression analyses is given in Table 2.
T. Gebauer, H. BassiriRad / Environmental Pollution 159 (2011) 3455e3461 3459
demand (Cienciala and Lindroth, 1995a,b; Granier and Bréda, 1996;Vertessy et al., 1995), and root hydraulic conductivity scales up ordown to match the demand generated by changes in LA. If Lp isdriven by changes in LA, then we would expect a significantcorrelation between Lp expressed on root DWand LA bases. Indeed,we found that, in three out of four species tested here (Table 2), Lpdwand LpLA were significantly and positively correlated (Fig. 3) inNO3
�-fed plants (Fig. 3 and Table 2). In contrast, our data showedthat, when plants are grownwith NH4
þ as the sole source of N, Lpdwand LpLA are not significantly correlated in any of the species testedhere (Fig. 3 and Table 2). This lack of correlation indicates stronglythat, under NH4
þ nutrition, Lp responses to CO2 are decoupled fromchanges in LA and possibly from whole plant water loss. Sucha decoupling is theoretically possible if other components ofresistance (e.g., stomatal) to the hydraulic flow and/or the drivingforce (e.g., water potential gradient between root and foliage) alongthe SPAC counterbalanced the expected coordination between LAand Lp. Alternatively, the predicted coordination between LA and Lpmay be interrupted if root resistance is a variable function of flowrate (Kramer and Boyer, 1995; Passioura, 1988; Steudle, 2000).
Table 2Summary of correlations between LpLA vs. LpDW for P. pungens, P. abies, P. menziesiiand A. fraseri. In each species, regressions are done separately for each N form(Fig. 3). Hydraulic conductivities on root DW- and LA-basis were pooled for the threeCO2. Asterisks indicate statistically significant correlation between LpLA and LpDW asfollows: *p� 0.5, **p< 0.01, ***p< 0.001. NS indicates not significant on a 5%significance level. The F and p values as well as r2 for statistically significant rela-tionships are given in Fig. 3.
Species Inorganic N forms
NH4NO3 NO3� NH4
þ
P. pungens NS *** NSP. abies * * NSP. menziesii *** NS NSA. fraseri * *** NS
Finally, water uptake and transport are controlled by a number ofcompensatory root system adjustments other than Lp. For example,adjustments in proportional biomass allocation to leaves vs. roots(Rodriguez-Gamir et al., 2010) or changes in root morphology andarchitecture (Cruz et al., 1999) and age (McCully and Canny, 1988;North and Nobel, 1991) can significantly affect plant hydraulics.Therefore, we suggest that the apparent lack of coordinationbetween LA and the efficiency of root water transport responses ofNH4
þ-fed plants to high CO2 may be explained by these compen-satory root characteristics.
5. Conclusions
The distinct role of NH4þ vs. NO3
� reported here provides a newinsight on factors that control the coordination of Lp and wholeplant water loss, and how that coupling may respond to rising CO2levels. The universality of this pattern would, however, requireevidence from a broader range of species and functional groups.Nevertheless, to our knowledge, this is the first study that high-lights the role of inorganic N form as amodulating factor linking therise in atmospheric CO2 and plant hydraulics. Currently, few studiesspecifically examine interacting effects of N and CO2 on plant andecosystem hydrologic processes (Domec et al., 2009; Leakey et al.,2009; Reich et al., 2006); hence, it is not surprising that theimportance of inorganic N sources have received even less atten-tion in CO2 studies. The rise in CO2 during the last four decades hasbeen accompanied by chronic deposition of reactive N across theglobe, a trend that is likely to continue in the 21st century(Galloway et al., 2008; Vitousek et al., 1997). The relative abundanceof NH4
þ vs. NO3� in chronic N deposition varies dramatically from
one region to another (Galloway and Cowling, 2002; Holland et al.,2005; Lovett, 1994; Schulze, 1989), affecting the ratio of theseinorganic N sources in native soils. Those facts, along with thedifferential effects of ammonium vs. NO3
� reported in this study,provide a strong justification to include N form as an integral part of

T. Gebauer, H. BassiriRad / Environmental Pollution 159 (2011) 3455e34613460
future studies and models designed to predict plant and ecosystemresponses to high CO2.
Acknowledgments
We thank greenhouse staff, James Scios and Noranne Magee, fortheir help with the experimental setups. We also thank Drs.Zwieniecki and Islam, who helped with setting up and trouble-shooting the root hydraulic conductivity system. Assistance fromLynne Wiora on editing and formatting and maintenance andupkeep of the plants by undergraduate students, Mark Vaughn,Rupak Basu and Joshua Kannankeril, is also appreciated. Commentsand suggestions of an anonymous reviewer were very helpful indrafting the final version of the paper. This study was funded byNSF grant IOS-0823315.
References
Aber, J.D., Nadelhoffer, K.J., Steudler, P., Melillo, J.M., 1989. Nitrogen saturation innorthern forest ecosystems. BioScience 39, 378e386.
Adler, P.R., Wilcox, G.E., Markhart III, A.H., 1996. Ammonium decreases musk-melon root system hydraulic conductivity. Journal of Plant Nutrition 19,1395e1403.
Ainsworth, E.A., Long, S.P., 2005. What have we learned from 15 years of free-airCO2 enrichment (FACE)? A meta-analytic review of the responses of photo-synthesis, canopy properties and plant production to rising CO2. New Phytol-ogist 165, 351e372.
Atkinson, C.J., Taylor, C.M., 1996. Effects of elevated CO2 on stem growth, vessel areaand hydraulic conductivity of oak and cherry seedlings. New Phytologist 133,617e626.
Atwell, B.J., Henery, M.L., Ball, M.C., 2009. Does soil nitrogen influence growth,water transport and survival of snow gum (Eucalyptus pauciflora Sieber exSprengel.) under CO2 enrichment? Plant, Cell and Environment 32, 553e566.
Barrett, D.J., Gifford, R.M., 1995. Photosynthetic acclimation to elevated CO2 inrelation to biomass allocation in cotton. Journal of Biogeography 22, 331e339.
Barthes, L., Deleens, E., Bousser, A., Hoarau, J., Priol, J.L., 1996. Xylem exudation isrelated to nitrate assimilation pathway in detopped maize seedlings: use ofnitrate reductase and glutamine synthetase inhibitors as tools. Journal ofExperimental Botany 47, 485e495.
BassiriRad, H., 2000. Kinetics of nutrient uptake by roots: response to global change.New Phytologist 147, 155e169.
BassiriRad, H., Thomas, R.B., Reynolds, J.F., Strain, B.R., 1996. Differential response ofroot kinetics of NH4
þ and NO3� to enriched atmospheric CO2 concentration in
field-grown loblolly pine. Plant, Cell and Environment 19, 367e371.BassiriRad, H., Griffin, K.L., Reynolds, J.F., Strain, B.R., 1997. Changes in root NH4
þ andNO3
� absorption rates of loblolly and ponderosa pine in response to CO2enrichment. Plant and Soil 190, 1e9.
BassiriRad, H., Gutschick, V.P., Lussenhop, J., 2001. Root system adjustments:regulation of plant nutrient uptake and growth responses to elevated CO2.Oecologia 126, 305e320.
Bazzaz, F.A., 1990. The response of natural ecosystems to the rising global CO2.Annual Reviews in Ecology and Systematics 21, 167e196.
Becker, P., Tyree, M.T., Tsuda, M., 1999. Hydraulic conductances of angiospermsversus conifers: similar transport sufficiency at the whole-plant level. TreePhysiology 19, 445e452.
Bhattacharya, N.C., Radin, J.W., Kimball, B.A., Mauney, J.R., Hendrey, G.R., Nagy, J.,Lewin, K.F., Ponce, D.C., 1994. Leaf water relations of cotton in free-airCO2-enriched environment. Agricultural and Forest Meteorology 70, 171e182.
Brodribb, T.J., Hill, R.S., 2000. Increases in water potential gradient reduce xylemconductivity in whole plants. Evidence from a low-pressure conductivitymethod. Plant Physiology 123, 1021e1028.
Bunce, J.A., 1996. Growth at elevated carbon dioxide concentration reduceshydraulic conductance in alfalfa and soybean. Global Change Biology 2,155e158.
Ceulemans, R., Jach, M.E., Van de Velde, R., Lin, J.X., Stevens, M., 2002. Elevatedatmospheric CO2 alters wood production, wood quality and wood strength ofScots pine (Pinus sylvestris L) after three years of enrichment. Global ChangeBiology 8, 153e162.
Chrispeels, M.J., Maurel, C., 1994. Aquaporins: the molecular basis of facilitatedwater movement through living plant cells? Plant Physiology 105, 9e13.
Cienciala, E., Lindroth, A., 1995a. Gas-exchange and sap flow measurements ofwillow trees in short-rotation forest. I. Transpiration and sap flow. Trees 9,1e12.
Cienciala, E., Lindroth, A., 1995b. Gas-exchange and sap flow measurements of Salixviminalis trees in short rotation forest. II. Diurnal and seasonal variation ofstomatal response and water use efficiency. Trees 9, 295e301.
Clarkson, D.T., Carvajal, M., Henzler, T., Waterhouse, R.N., Smyth, A.J., Cooke, D.T.,Steudle, E., 2000. Root hydraulic conductance: diurnal aquaporin expressionand the effects of nutrient stress. Journal of Experimental Botany 51, 61e70.
Conroy, J.P., Milham, P.J., Mazur, M., Barlow, E.W.R., 1990. Growth, dry weight par-titioning and wood properties of Pinus radiata D. Don after 2 years of CO2enrichment. Plant, Cell and Environment 13, 329e337.
Constable, J.V.H., BassiriRad, H., Lussenhop, J., Zerihun, A., 2001. Influence ofelevated CO2 and mycorrhizae on nitrogen acquisition: contrasting responses inPinus taeda and Liquidambar styraciflua. Tree Physiology 21, 83e91.
Cruz, C., Lips, S.H., Martins-Loucao, M.A., 1999. Changes in the morphology of rootsand leaves of carob seedlings induced nitrogen source and atmospheric carbondioxide. Annals of Botany 80, 817e823.
Domec, J.C., Palmroth, S., Ward, E., Maier, C.A., Therezien, M., Oren, R., 2009.Acclimation of leaf hydraulic conductance and stomatal conductance of Pinustaeda (loblolly pine) to long-term growth in elevated CO2 (free-air CO2enrichment) and N fertilization. Plant, Cell and Environment 32, 1500e1512.
Domec, J.C., Schäfer, K., Oren, R., Kim, H.S., McCarthy, H.R., 2010. Variable conduc-tivity and embolism in roots and branches of four contrasting tree species andtheir impacts on whole-plant hydraulic performance under future atmosphericCO2 concentration. Tree Physiology 30, 1001e1015.
Drake, B.G., Gonzalez-Meler, M.A., Long, S.P., 1997. More efficient plants: a conse-quence of rising atmospheric CO2? Annual Reviews of Plant Physiology andMolecular Biology 48, 609e639.
Ewers, B.E., Oren, R., Sperry, J.S., 2000. Influence of nutrient versus water supply onhydraulic architecture and water balance in Pinus taeda. Plant, Cell and Envi-ronment 23, 1055e1066.
Farage, P.K., McKee, I.F., Long, S.P., 1998. Does a low nitrogen supply necessarily leadto acclimation of photosynthesis to elevated CO2? Plant Physiology 118,573e580.
Field, C.B., Jackson, R.B., Mooney, H.A., 1995. Stomatal responses to increased CO2:implications from the plant to the global scale. Plant, Cell and Environment 18,1214e1225.
Finzi, A.C., Moore, D.J.P., DeLucia, E.H., Lichter, J., Hofmuckel, K.S., Jackson, R.B.,Kim, H.S., Matamala, R., McCarthy, H.R., Oren, R., Pippen, J.S., Schlesinger, W.H.,2006. Progressive nitrogen limitation of ecosystem processes under elevatedCO2 in a warm-temperate forest. Ecology 87, 15e25.
Galloway, J.N., 1998. The global nitrogen cycle: changes and consequences. Envi-ronmental Pollution 102, 15e24.
Galloway, J.N., Cowling, E.N., 2002. Reactive nitrogen and the world: 200 years ofchange. AMBIO: A Journal of the Human Environment 31, 64e71.
Galloway, J.N., Townsend, A.R., Erisman, J.W., Bekunda, M., Cai, Z., Freney, J.R.,Martinelli, L.A., Seitzinger, S.P., Sutton,M.A., 2008. Transformation of the nitrogencycle: recent trends, questions, and potential solutions. Science 320, 889e892.
Gloser, V., Zwieniecki, M.A., Orians, C.M., Holbrook, N.M., 2007. Dynamic changes inroot hydraulic properties in response to nitrate availability. Journal of Experi-mental Botany 58, 2409e2415.
Gorska, A., Ye, Q., Holbrook, N.M., Zwieniecki, M.A., 2008a. Nitrate control of roothydraulic properties in plants: translating local information to whole plantresponse. Plant Physiology 148, 1159e1167.
Gorska, A., Zwieniecka, A., Holbrook, N.M., Zwieniecki, M.A., 2008b. Nitrateinduction of root hydraulic conductivity in maize is not correlated with aqua-porin expression. Planta 228, 989e998.
Gorska, A., Lazor, J.W., Zwieniecka, A., Benway, C., Zwieniecki, M.A., 2010. Thecapacity for nitrate regulation of root hydraulic properties correlates withspecies’ nitrate uptake rate. Plant and Soil 337, 447e455.
Granier, A., Bréda, N., 1996. Modeling canopy conductance and stand transpiration ofan oak forest fromsapflowmeasurements. Annals of Forest Science 53, 537e546.
Guo, S., Brück, H., Sattelmacher, B., 2002. Effects of supplied nitrogen form ongrowth and water uptake of French bean (Phaseolus vulgaris L.) plants. Nitrogenform and water uptake. Plant and Soil 239, 267e275.
Guo, S., Zhou, Y., Shen, Q., Zhang, F., 2007. Effect of ammonium and nitrate nutritionon some physiological processes in higher plants e growth, photosynthesis,photorespiration, and water relations. Plant Biology 9, 21e29.
Holland, E.A., Braswell, B.H., Sulzman, J., Lamarque, J.F., 2005. Nitrogen depositiononto the United States and Western Europe: synthesis of observations andmodels. Ecological Applications 15, 38e57.
Huxman, K.A., Smith, S.D., Neuman, D.S., 1999. Root hydraulic conductivity of Larreatridentata and Helianthus annuus under elevated CO2. Plant, Cell and Environ-ment 22, 325e330.
Kramer, P.J., Boyer, J.S., 1995. Water Relations of Plants and Soils. Academic Press,Orlando.
Kronzucker, H.J., Siddiqi, M.Y., Glass, A.D.M., 1997. Conifer root discriminationagainst soil nitrate and the ecology of forest succession. Nature 385, 59e61.
Küppers, M., 1984. Carbon relations and competition between woody species ina Central European hedgerow. II. Stomatal responses, water use, and hydraulicconductivity in the root/leaf pathway. Oecologia 64, 344e354.
Lane, D.R., BassiriRad, H., 2002. Interspecific variation in the growth and physiologyof tallgrass prairie species in response to nitrate loading. Plant FunctionalEcology 29, 1227e1235.
Lansgrud, O., 2003. ANOVA for unbalanced data: use type II instead of type III sumsof squares. Statistics and Computing 13, 163e167.
Leakey, A.D.B., Ainsworth, E.A., Bernacchi, C.J., Rogers, A., Long, S.P., Ort, D.R., 2009.Elevated CO2 effects on plant carbon, nitrogen, and water relations: siximportant lessons from FACE. Journal of Experimental Botany 60, 2859e2876.
Lovett, G.M., 1994. Atmospheric depositions of nutrients and pollutants in northAmerica: an ecological perspective. Ecological Applications 4, 629e650.
Luo, Y., Su, B., Currie, W., Dukes, J.S., Finzi, A., Hartwig, U., Hungate, B., McMurtie, R.E.,Oren, R., Parton, W.J., Pataki, D.E., Shaw, M.R., Zak, D.R., Field, C.B., 2004.

T. Gebauer, H. BassiriRad / Environmental Pollution 159 (2011) 3455e3461 3461
Progressive nitrogen limitation of ecosystem response to rising atmosphericcarbon dioxide. BioScience 54, 731e739.
Marschner, H., Häussling, M., George, E., 1991. Ammonium and nitrate uptake ratesand rhizosphere pH in non-mycorrhizal roots of Norway spruce [Picea abies (L.)Karst.]. Trees 5, 14e21.
Maurel, C., Verdoucq, L., Luu, D.T., Santoni, V., 2008. Plant aquaporins: membranechannels with multiple integrated functions. Annual Reviews of Plant Biology99, 595e624.
McCully, M.E., Canny, M.J., 1988. Pathways and processes of water and nutrientmovement in roots. Plant and Soil 111, 159e170.
McDonald, E.P., Kruger, E.L., Riemenschneider, D.E., Isebrands, J.G., 2002. Competi-tive status influences tree-growth responses to elevated CO2 and O3 inaggrading aspen stands. Functional Ecology 16, 792e801.
Medlyn, B.E., Barton, C.V.M., Broadmeadow, M.S.J., Ceulemans, R., De Angelis, P.,Forstreuter, M., Freeman, M., Jackson, S.B., Kellomäki, S., Laitat, E., Rey, A.,Roberntz, P., Sigurdsson, B.D., Strassemeyer, J., Wang, K., Curtis, P.S., Jarvis, P.G.,2001. Stomatal conductance of forest species after long-term exposure toelevated CO2 concentration: a synthesis. New Phytologist 149, 247e264.
Mencuccini, M., 2003. The ecological significance of long-distance water transport:short-term regulation, long-term acclimation and the hydraulic costs of statureacross plant life forms. Plant, Cell and Environment 26, 163e182.
Min, X., Siddiqi, M.Y., Guy, R.D., Glass, A.D.M., Kronzucker, H.J., 1998. Induction ofnitrate uptake and nitrate reductase activity in trembling aspen and lodgepolepine. Plant, Cell and Environment 21, 1039e1046.
Nardini, A., Tyree, M.T., 1999. Root and shoot hydraulic conductance of sevenQuercus species. Annals of Forest Science 56, 371e377.
Nobel, P.S., Cui, M., 1992. Hydraulic conductance of the soil, the rootesoil air gap,and the root: changes for desert succulents in drying soil. Journal of Experi-mental Botany 43, 319e326.
Norby, R.J., Wullschleger, S.D., Gunderson, C.A., Johnson, D.W., Ceulemans, R., 1999.Tree responses to rising CO2 in field experiments: implications for the futureforest. Plant, Cell and Environment 22, 683e714.
North, G.B., Nobel, P.S., 1991. Changes in hydraulic conductivity and anatomy causedby drying and rewetting roots of Agave deserti (Agavaceae). American Journal ofBotany 78, 906e915.
North, G.B., Peterson, C.A., 2005. Water flow in roots: structural and regulatoryfeatures. In: Holbrook, N.M., Zwieniecki, M.A. (Eds.), Vascular Transport inPlants. Physiological Ecology Series. Elsevier Academic Press, Burlington, USA,pp. 131e156.
Passioura, J.B., 1988. Water transport in and to roots. Annul Reviews in PlantPhysiology and Plant Molecular Biology 39, 245e256.
Pettersson, R., McDonald, A.J.S., Stadenberg, I., 1994. Response of small birch plants(Betula pendula Roth.) to elevated CO2 and nitrogen supply. Plant, Cell andEnvironment 16, 1115e1121.
Postaire, O., Verdoucq, L., Maurel, C., 2008. Aquaporins in plants: from molecularstructure to integrated functions. Advances in Botanical Research 46, 75e136.
Prior, S.A., Pritchard, S.G., Runion, G.B., Rogers, H.H., Mitchell, R.J., 1997. Influence ofatmospheric CO2 enrichment, soil N, and water stress on needle surface waxformation in Pinus palustris (Pinaceae). American Journal of Botany 84,1070e1077.
Pritchard, S.G., Rogers, H.H., Prior, S.A., Peterson, C.M., 1999. Elevated CO2 and plantstructure: a review. Global Change Biology 5, 807e837.
Radin, J.W., 1990. Responses of transpiration and hydraulic conductance to roottemperature in nitrogen- and phosphorus-deficient cotton seedlings. PlantPhysiology 92, 855e857.
Radin, J.W., Boyer, J.S., 1982. Control of leaf expansion by nitrogen nutrition insunflower plants. Plant Physiology 69, 771e775.
Radin, J.W., Eidenbock, M.P., 1984. Hydraulic conductance as a factor limiting leafexpansion of phosphorus-deficient cotton plants. Plant Physiology 75, 372e377.
Reich, P.B., Hungate, B.A., Luo, Y., 2006. Carbonenitrogen interactions in terrestrialecosystems in response to rising atmospheric carbon dioxide. Annual Reviewsin Ecology, Evolution and Systematics 37, 611e636.
Rodriguez-Gamir, J., Intrigliolo, D.S., Primo-Millo, E., Former-Giner, M.A., 2010.Relationship between xylem anatomy, root hydraulic conductivity, leaf/rootratio and transpiration in citrus trees on different rootstocks. Biologia Planta-rum 139, 159e168.
Rogers, A., Humphries, S.W., 2000. A mechanistic evaluation of photosyntheticacclimation at elevated CO2. Global Change Biology 6, 1005e1011.
Rogers, A., Fischer, B.U., Bryant, J., Frehner, M., Blum, H., Raines, C.A., Long, S.P., 1998.Acclimation of photosynthesis to elevated CO2 under low-nitrogen nutrition isaffected by the capacity for assimilate utilization. Perennial ryegrass under free-air CO2 enrichment. Plant Physiology 118, 683e689.
Rothstein, D.E., Cregg, B.M., 2005. Effects of nitrogen form on nutrient uptake andphysiological performance of Fraser fir (Abies fraseri). Forest Ecology andManagement 219, 69e80.
Rüdinger, M., Hallgren, S.W., Steudle, E., Schulze, E.D., 1994. Hydraulic and osmoticproperties of spruce roots. Journal of Experimental Botany 45, 1413e1425.
Saliendra, N.Z., Meinzer, F.C., 1989. Relationship between root/soil hydraulic prop-erties and stomatal behavior in sugarcane. Australian Journal of Plant Physi-ology 16, 241e250.
Sarker, B.C., Hara, M., 2009. Effects of elevated CO2 and water stress on rootstructure and hydraulic conductance of Solanum melongena L. BangladeshJournal of Botany 38, 55e63.
Saxe, H., Ellsworth, D.S., Heath, J., 1998. Tree and forest functioning in an enrichedCO2 atmosphere. New Phytologist 139, 395e436.
Schäfer, K.V.R., Oren, R., Laj, C.T., Katul, G., 2002. Hydrologica balance in an intacttemperate forest ecosystem under ambient and elevated atmospheric CO2concentration. Global Change Biology 8, 895e911.
Schreiber, L., Franke, R., Hartmann, K.D., Ranathunge, K., Steudle, E., 2005. Thechemical composition of suberin in apoplastic barriers affects radial hydraulicconductivity differently in the roots of rice (Oryza sativa L. cv. IR64) and corn(Zea mays L. cv. Helix). Journal of Experimental Botany 56, 1427e1436.
Schulze, E.D., 1989. Air pollution and forest decline in a spruce (Picea abies) forest.Science 244, 776e783.
Schulze-Till, T., Kaufmann, I., Sattelmacher, B., Jakob, P., Haase, A., Guo, S.,Zimmermann, U., Wegner, L.H., 2009. A 1H NMR study of water flow in Pha-seolus vulgaris L. roots treated with nitrate or ammonium. Plant and Soil 319,307e321.
Smirnoff, N., Todd, P., Stewart, G.R., 1984. The occurrence of nitrate reduction in theleaves of woody plants. Annals of Botany 54, 363e374.
Stark, J.M., Hart, S.C., 1997. High rates of nitrification and nitrate turnover inundisturbed coniferous forests. Nature 385, 61e64.
Steudle, E., 2000. Water uptake by plants: an integration of views. Plant and Soil226, 45e56.
Steudle, E., Peterson, C.A., 1998. How does water get through roots? Journal ofExperimental Botany 49, 775e788.
Stitt, M., Kapp, A., 1999. The interaction between elevated carbon dioxide andnitrogen nutrition: the physiological and molecular background. Plant, Cell andEnvironment 22, 583e621.
Stulen, I., Perez-Soba, M., De Kok, L.J., Van Der Eerden, L., 1998. Impact of gaseousnitrogen deposition on plant functioning. New Phytology 139, 61e70.
Tricker, P.J., Pecchiari, M., Bunn, S.M., Vaccari, F.P., Peressotti, A., Miglietta, F.,Taylor, G., 2009. Water use of a bioenergy plantation increases in a future highCO2-world. Biomass & Bioenergy 33, 200e208.
Trubat, R., Cortina, J., Vilagrosa, A., 2006. Plant morphology and root hydraulics arealtered by nutrient deficiency in Pistacia lentiscus (L.). Trees 20, 334e339.
Tsuda, M., Tyree, M.T., 1997. Whole-plant hydraulic resistance and vulnerabilitysegmentation in Acer saccharinum. Tree Physiology 17, 351e357.
Tyerman, S.D., Niemietz, C.M., Bramley, H., 2002. Plant aquaporins: multifunctionalwater and solute channels with expanding roles. Plant, Cell and Environment26, 173e194.
Tyree, M.T., Velez, V., Dailing, J.W., 1998. Growth dynamics of root and shoothydraulic conductance in seedlings of five neotropical tree species: scaling toshow possible adaptation to differing light regimes. Oecologia 114, 293e298.
Tyree, M.T., Zimmermann, M.H., 2002. Xylem Structure and the Ascent of Sap,second ed. Springer-Verlag, Berlin, p. 283.
Van Den Driessche, R., 1971. Response of conifer seedlings to nitrate and ammo-nium sources of nitrogen. Plant and Soil 34, 421e439.
Vertessy, R.A., Benyon, R.G., O’Sullivan, S.K., Gribben, P.R., 1995. Relationshipbetween stem diameter, sapwood area, leaf area and transpiration in a youngmountain ash forest. Tree Physiology 15, 559e567.
Vitousek, P.M., Aber, J.D., Howarth, R.W., Likens, G.E., Matson, P.A., Schindler, D.W.,Schlesinger, W.H., Tilman, D.G., 1997. Human alteration of the global nitrogencycle: sources and consequences. Ecological Applications 7, 737e750.
Wolfe, D.W., Gifford, R.M., Hilbert, D., Luo, Y., 1998. Integration of photosyntheticacclimation to CO2 at the whole-plant level. Global Change Biology 4, 879e893.
Wullschleger, S.D., Tschapilinski, T.J., Norby, R.J., 2002. Plant water relations atelevated CO2 e implications for water-limited environments. Plant, Cell andEnvironment 25, 319e331.
Zerihun, A., BassiriRad, H., 2001. Interspecies variation in nitrogen uptake kineticresponses of temperate forest species to elevated CO2: potential causes andconsequences. Global Change Biology 7, 211e222.
Zwieniecki, M.A., Melcher, P.J., Holbrook, N.M., 2001. Hydrogel control of xylemhydraulic resistance in plants. Science 291, 1059e1062.


















