Effect of tri-n-butyltin on intracellular Ca2+ concentration of mouse thymocytes under Ca2+-free...
6
environmental toxicology European Journal of Pharmacology and pharmacology ELSEVIER Environmental Toxicology and Pharmacology Section 270 (1994) 137-142 Effect of tri-n-butyltin on intracellular Ca 2 + concentration of mouse thymocytes under Ca 2+-free condition Yasuo Oyama *, Toshiko Ueha ~, Akemi Hayashi 2, Lumi Chikahisa 3 Laboratory of Cell Signalling (Pharmacology), Faculty of lntegrated Arts and Sciences, The University of Tokushima, Minami-Jyozanjima, Tokushima 770, Japan Received 8 September 1993, accepted 28 September 1993 Abstract Effect of tri-n-butyltin at concentrations ranging from 100 nM to 1 /~M on the intracellular Ca 2+ concentration of mouse thymocytes was examined under Ca2+-free conditions in comparison with those of 50 nM A23187, 100 nM thapsigargin and 10 /xM cyclopiazonic acid, using the fluorescent dye for intracellular Ca 2÷, fluo-3. Tri-n-butyitin persistently increased the intensity of fluo-3 fluorescence while A23187, thapsigargin and cyclopiazonic acid produced a transient augmentation of the fluorescence. Pretreatment with A23187 greatly decreased the fluorescence responses induced by 1 /xM tri-n-butyltin. However, the effect of thapsigargin and cyclopiazonic acid on the tri-n-butyltin-induced response was much weaker than that of A23187. In the presence of tri-n-butyltin, the transient response produced by A23187 was greatly prolonged. Results may suggest that tri-n-butyltin increases the membrane Ca 2÷ permeability of the intracellular organelles (cellular calcium stores) and decreases the Ca 2+ pump activity of thymocyte membrane, resulting in a sustained increase in the intracellular Ca 2+ concentration under Ca2+-free concentration. Key words: Tri-n-butyltin; Thymocyte; Ca 2+ pump; Thapsigargin; A23187; Cyclopiazonic acid 1.Introduction The biocidal activity of the triorganotin compounds was first recognized by Van der Kerk and Luijten (1958). Along with the expansion of technical applica- tions such as agricultural fungicide and marine an- tifouling, the concern about possible environmental and health effects of triorganotin compounds increased (Luijten, 1971; Sheldon, 1975; Wilkinson, 1984; for a review, Snoeij et al., 1987). Of triorganotin compounds, we have previously re- ported that tri-n-butyltin increases the intracellular Ca 2+ concentration ([Ca2+]i) of mouse thymocytes and dissociated rat cerebellar neurons, suggesting a type of cytotoxic action (Chikahisa and Oyama, 1992; Oyama et al., 1993). Although the efficacy of tri-n-butyltin in increasing the [Ca 2+]i of both preparations was greatly * Corresponding author. Fax: (81X886)55-2108. Present address: St. Luke's International Hospital, Tokyo 104, Japan. 2 Present address: Ono Pharmaceutical Co., Osaka 619, Japan. 3present address: Taiho Pharmaceutical Co., Tokushima 771-01, Japan. 0926-6917/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0926-6917(93)E0069-D reduced under Ca2+-free condition, tri-n-butyltin still increased the [Ca2+]i, being independent of the pres- ence of external Ca 2+. It was reminiscent of the possi- bility that tri-n-butyltin also affected the intracellular organelles regulating the [Ca2+] i. In this study, in order to characterize the property of tri-n-butyltin in increas- ing the [Ca2+] i under Ca2+-free condition, we have made a comparison of the effect of tri-n-butyltin on the [Ca2+]i of mouse thymocytes with those of a Ca 2+ ionophore, A23187, and the inhibitors for CaZ+-ATPase of intracellular organelles, thapsigargin and cyclopia- zonic acid (Seidler et al., 1989; Thastrup, 1990), using the fluorescent dye for intracellular Ca e+, fluo-3 (Kao et al., 1989). 2. Materials and methods 2.1. Chemicals Pentaacetoxymethyl ester of fluo-3 (Kao et al., 1989), a fluorescent dye for intracellular Ca 2+, was obtained from Dojindo Laboratory (Kumamoto, Japan). Tri-n-
Transcript of Effect of tri-n-butyltin on intracellular Ca2+ concentration of mouse thymocytes under Ca2+-free...

environmental toxicology European Journal of Pharmacology and pharmacology
ELSEVIER Environmental Toxicology and Pharmacology Section 270 (1994) 137-142
Effect of tri-n-butyltin on intracellular Ca 2 + concentration of mouse thymocytes under Ca 2+-free condition
Y a s u o O y a m a *, T o s h i k o U e h a ~, A k e m i H a y a s h i 2, L u m i C h i k a h i s a 3
Laboratory of Cell Signalling (Pharmacology), Faculty of lntegrated Arts and Sciences, The University of Tokushima, Minami-Jyozanjima, Tokushima 770, Japan
Received 8 September 1993, accepted 28 September 1993
Abstract
Effect of tri-n-butyltin at concentrations ranging from 100 nM to 1 /~M on the intracellular Ca 2+ concentration of mouse thymocytes was examined under Ca2+-free conditions in comparison with those of 50 nM A23187, 100 nM thapsigargin and 10 /xM cyclopiazonic acid, using the fluorescent dye for intracellular Ca 2÷, fluo-3. Tri-n-butyitin persistently increased the intensity of fluo-3 fluorescence while A23187, thapsigargin and cyclopiazonic acid produced a transient augmentation of the fluorescence. Pretreatment with A23187 greatly decreased the fluorescence responses induced by 1 /xM tri-n-butyltin. However, the effect of thapsigargin and cyclopiazonic acid on the tri-n-butyltin-induced response was much weaker than that of A23187. In the presence of tri-n-butyltin, the transient response produced by A23187 was greatly prolonged. Results may suggest that tri-n-butyltin increases the membrane Ca 2÷ permeability of the intracellular organelles (cellular calcium stores) and decreases the Ca 2+ pump activity of thymocyte membrane, resulting in a sustained increase in the intracellular Ca 2+ concentration under Ca2+-free concentration.
Key words: Tri-n-butyltin; Thymocyte; Ca 2+ pump; Thapsigargin; A23187; Cyclopiazonic acid
1.Introduction
The biocidal activity of the triorganotin compounds was first recognized by Van der Kerk and Luijten (1958). Along with the expansion of technical applica- tions such as agricultural fungicide and marine an- tifouling, the concern about possible environmental and health effects of triorganotin compounds increased (Luijten, 1971; Sheldon, 1975; Wilkinson, 1984; for a review, Snoeij et al., 1987).
Of triorganotin compounds, we have previously re- ported that tri-n-butyltin increases the intracellular Ca 2+ concentration ([Ca2+]i) of mouse thymocytes and dissociated rat cerebellar neurons, suggesting a type of cytotoxic action (Chikahisa and Oyama, 1992; Oyama et al., 1993). Although the efficacy of tri-n-butyltin in increasing the [Ca 2+]i of both preparations was greatly
* Corresponding author. Fax: (81X886)55-2108. Present address: St. Luke's International Hospital, Tokyo 104, Japan.
2 Present address: Ono Pharmaceutical Co., Osaka 619, Japan. 3present address: Taiho Pharmaceutical Co., Tokushima 771-01, Japan.
0926-6917/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0 9 2 6 - 6 9 1 7 ( 9 3 ) E 0 0 6 9 - D
reduced under Ca2+-free condition, tri-n-butyltin still increased the [Ca2+]i, being independent of the pres- ence of external Ca 2+. It was reminiscent of the possi- bility that tri-n-butyltin also affected the intracellular organelles regulating the [Ca2+] i. In this study, in order to characterize the property of tri-n-butyltin in increas- ing the [Ca2+] i under Ca2+-free condition, we have made a comparison of the effect of tri-n-butyltin on the [Ca2+]i of mouse thymocytes with those of a Ca 2+ ionophore, A23187, and the inhibitors for CaZ+-ATPase of intracellular organelles, thapsigargin and cyclopia- zonic acid (Seidler et al., 1989; Thastrup, 1990), using the fluorescent dye for intracellular Ca e+, fluo-3 (Kao et al., 1989).
2. Materials and methods
2.1. Chemicals
Pentaacetoxymethyl ester of fluo-3 (Kao et al., 1989), a fluorescent dye for intracellular Ca 2+, was obtained from Dojindo Laboratory (Kumamoto, Japan). Tri-n-

138 Y. Oyama et al. /European Journal of Pharmacology - Em.,ironmental Toxicolo,~' and Pharmacology Section 270 (1994) 137-142
butyltin chloride was purchased from Kishida Chemi- cals (Osaka, Japan). A23187, thapsigargin and cyclopia- zonic acid were from Sigma Chemical Company (St. Louis, MO). Other chemical reagents were purchased from Katayama Chemical Industries, Co. (Osaka, Japan). Since tri-n-butyltin at concentrations of 3 /xM or more caused strong depolarization of thymocyte membrane, suggesting a type of non-specific action on the membrane, the concentrations used in this study were 100 nM to 1 /xM which hyperpolarized the mem- brane, due to the activation of Ca2+-dependent K + conductance, during 10 rain after the application (Chikahisa and Oyama, 1992). A23187 at the concen- tration ranging from 1 nM to 30 nM produced a dose-dependent hyperpolarization of thymocyte mem- brane in the presence of external Ca 2+ (Oyama et al., 1992). However, A23187 at the concentrations of 100 nM or more sometimes seemed to deteriorate the thymocytes since the membrane of thymocytes greatly depolarized after a transient hyperpolarization. There- fore, 50 nM of A23187 was used in this study. The concentrations of cyclopiazonic acid and thapsigargin used as a specific inhibitor for CaZ+-ATPase of intra- cellular organelles (Seidler et al., 1989; Thastrup, 1990), were respectively 10 p~M and 100 nM, since further increase in the concentration of respective agent did not produced further augmentation of fluo-3 fluores- cence under external Ca2+-free condition.
2.2. Preparation and solution
The thymocytes were dissociated from thymus glands of 4-week-old ddY mice (from Nissin, Tokushima, Japan) in the CaZ+-free Tyrode's solution at 1-4°C. The Ca2+-free Tyrode's solution used had the follow- ing composition (mM): NaC1 148, KC1 2, MgCI 2 3, E G T A 2, Hepes 10, NaOH 2 and D-glucose 5. The pH of the solution was 7.3. The solution containing thymo- cytes was filtered through a mesh (diameter of 53 p~m) to remove the residues of the thymus. The cell suspen- sion was incubated at 35-36°C for 50-60 rain at least before any fluorescence measurement.
2.3. Fluorescence measurement and analysis
The thymocytes were incubated in the Ca2+-free Tyrode's solution with pentaacetoxymethyl ester of fluo-3 at the concentration of 300 nM for 60 min for loading fluo-3 into the cells as previously described (Oyama et al., 1992). Thereafter , the fluorescence mea- surement from the thymocytes was done on a flow cytometer equipped with argon laser (Cyto-ACE 150, Japan Spectroscopic Co., Tokyo, Japan). Excitation wavelength produced by argon laser was 488 nm and the emission was detected at the wavelength of 530 + 20 nm using a bandpass filter. Fluorescence obtained from
individual thymocytes was analysed on a personal com- puter (PC-8100XR, Nippon Electronics Co., Tokyo, Japan) with software (JASCO System program Ver.3.XX) developed by Japan Spectroscopic Co., Tokyo, Japan. Data aquisition from 4000-5000 cells required about 20 s. Therefore, the aquisition started 10 s before the time described in the text and figure legends. Changes in the fluo-3 fluorescence to higher and lower intensities (arbitrary unit) correspond to the increase and decrease in the [Ca2+]i, respectively. Ten- tative calibration of fluo-3 fluorescence for the [Ca 2+]i was made by the method using ionomycin and Mn 2+ (Kao et al., 1989). Data are presented as means___ S.E.M. from four to six experiments. Statistical signifi- cance was determined by ANOVA procedures. The effects were considered significant if P < 0.05.
3. Results
3.1. Comparison of the action of tri-n-butyltin on the [ Ca2 +]i with those ofA23187, thapsigargin and cyclopi- azonic acid
Although tri-n-butyltin at the concentrations of 100-300 nM augmented the fluo-3 fluorescence of mouse thymocytes under Ca2+-free condition, the de- gree of augmentation in the fluorescence was not sig- nificant. The peak intensity of fluo-3 fluorescence his- togram was greatly increased from the control value of 22.2 _+ 1.3 to 48.5 + 7.5 (mean + S.E.M. of five experi- ments, in arbitrary unit) by 1 /zM tri-n-butyltin in a t ime-dependent manner and the maximum increase was observed within 10 min after applying tri-n-butyltin into the cell suspension (Fig. 1). Mean values of cali- brated [Ca2+]~ in the control and at 10 min after tri-n-butyltin application were 33.8 _+ 5.0 nM and 38.5 +_ 4.8 nM in the case of 100 nM tri-n-butyltin, 34.5 _+ 6.2 nM and 49.9 _+ 10.5 nM for 300 nM, and 31.8 + 6.5 nM and 208.6 _+ 66.4 nM for 1 /xM, respectively (mean _+ S.E.M. of four experiments). The change in calibrated [Ca2+]~ induced by 1 /zM tri-n-butyltin was significant (P < 0.01). A23187, thapsigargin and cyclopiazonic acid also augmented the fluo-3 fluorescence under Ca 2 +-free condition (Fig. 2). However, their augmentation of the fluo-3 fluorescence was transient while 1 /zM tri-n- butyltin produced a sustained increase in the intensity of fluorescence. Although A23187 exerted the most potent action on the fluo-3 fluorescence, the fluores- cence augmented transiently by A23187 decreased to the control level or less within 3 min after its applica- tion while it required 5 min or longer in the case of thapsigargin and cyclopiazonic acid (Fig. 2). These results suggest a difference in the mode of action in increasing the [Ca2+] i between tri-n-butyltin and other agents used in this study.

Y. Oyama et al. /European Journal of Pharmacology - Enuironmental Toxicology and Pharmacology Section 270 (1994) 137-142 139
3.2. Pretreatment with A23187 on tri-n-butyltin-induced increase in the [Ca 2 +]i
To further characterize the effect of tri-n-butyltin on the [Ca2+]i under CaZ+-free condition, the influence of the pretreatment with A23187 on tri-n-butyltin-induced augmentation of the fluo-3 fluorescence was examined. Tri-n-butyltin was added into the cell suspension after the intensity of fluo-3 fluorescence returned to the level similar to control fluorescence in the presence of A23187 (Fig. 2). Pretreatment with 50 nM A23187 greatly decreased the augmentation of fluo-3 fluores- cence by 1 IzM tri-n-butyltin (Fig. 3). Thus, 1 /zM tri-n-butyltin increased the intensity of fluo-3 fluores- cence from 20.5 + 0.8 to 46.2 + 10.5 in the control while it was from 19.2 _+ 1.2 to 25.4 _+ 2.2 in the cells pretreated with 50 nM A23187. The maximum levels of calibrated [Ca2+]i increased by 1 /zM tri-n-butyltin in the control and in pretreatment with A23187 were 310.2_+ 70.4 nM and 118.6_+ 24.9 nM, respectively (mean + S.E.M. of four experiments). The difference
m A. lm in B. 3min C. 5min
Z I I I I I I I I I [ I I 100 101 102 103 100 101 102 103 100 101 102 103
FLUORESCENCE INTENSITY (arbitrary unit)
-tOOnM1 - - - " o 3 0 0 n M I / -
Zzso • j.MMI /
t r > - 0 1 - - --2_ D ~ - J z [ ] 13 ...................................... -- LLLu
1 0 - z t J J i i I I I I I I I I I -3 0 5 10
TIME AFTER DRUG APPLICATION (min) Fig. 1. Effect of tri-n-butyltin on the profile of fluo-3 fluorescence obtained from mouse thymocytes under Ca2+-free conditions. Upper panel: the effect of 1 /.LM tri-n-butyltin on the fluorescence his- togram constructed from 2500 thymocytes. The abscissa corresponds to the intensity of fluo-3 fluorescence in arbitrary units. Increasing the intensity of fluo-3 fluorescence indicates the elevation of intracel- lular Ca 2+ concentration. The ordinate shows the number of cells at the respective intensity. Histograms without and with arrow were obtained from the thymocytes in the control and at 1 rain (A), 3 min (B) and 5 min (C) after the application of 1 /~M tri-n-butyltin. Lower panel: the time course for tri-n-butyltin-induced increase in the mean intensity of fluorescence histogram obtained from 5000 thymocytes at the respective concentrat ions of tri-n-butyltin. The dotted line and the open squares indicate the control level of fluorescence intensity. The arrow indicates the application of tri-n-butyltin at the respective concentrations. The result is a representative one of three experi- ments.
co A. A23187 -4, 128 r
~ 64
z I I I I
B. TPG C. CPA
. . . . . . . . .
I I I I I I I I
z ~
rr>- O~- D~ - J z LLIj J
7 0 -
5 0 - -
[ ]
1 0 - ! I -3
100 101 102 103 100 101 102 103 100 101 102 103 FLUORESCENCE INTENSITY (arbitrary unit)
e A23187 ] 0 TPG [
i I t t I t I t I ~ I i 0 5 10
TIME AFTER DRUG APPLICATION (min)
Fig. 2. Effects of A23187, thapsigargin (TPG) and cyclopiazonic acid (CPA) on the profile of fluo-3 fluorescence in the thymocytes under Ca2+-free conditions. Upper panel: the effects of 50 nM A23187 (A), 100 nM thapsigargin (B) and 10 /xM cyclopiazonic acid (C) on the fluorescence histogram constructed from 3000 thymocytes at 1 rain after their applications. Abscissa and ordinate are the same as those of Fig. 1. The histograms without and with the arrow were con- structed from the control thymocytes and the drug-treated thymo- cytes, respectively. Lower panel: the time course of individual agents for increasing the mean intensity of fluo-3 fluorescence histogram of 5000 thymocytes. The arrow shows the respective drug applications. The dotted line and open square indicate the control level of fluorescence intensity. Result is a representative one of three experi- ments.
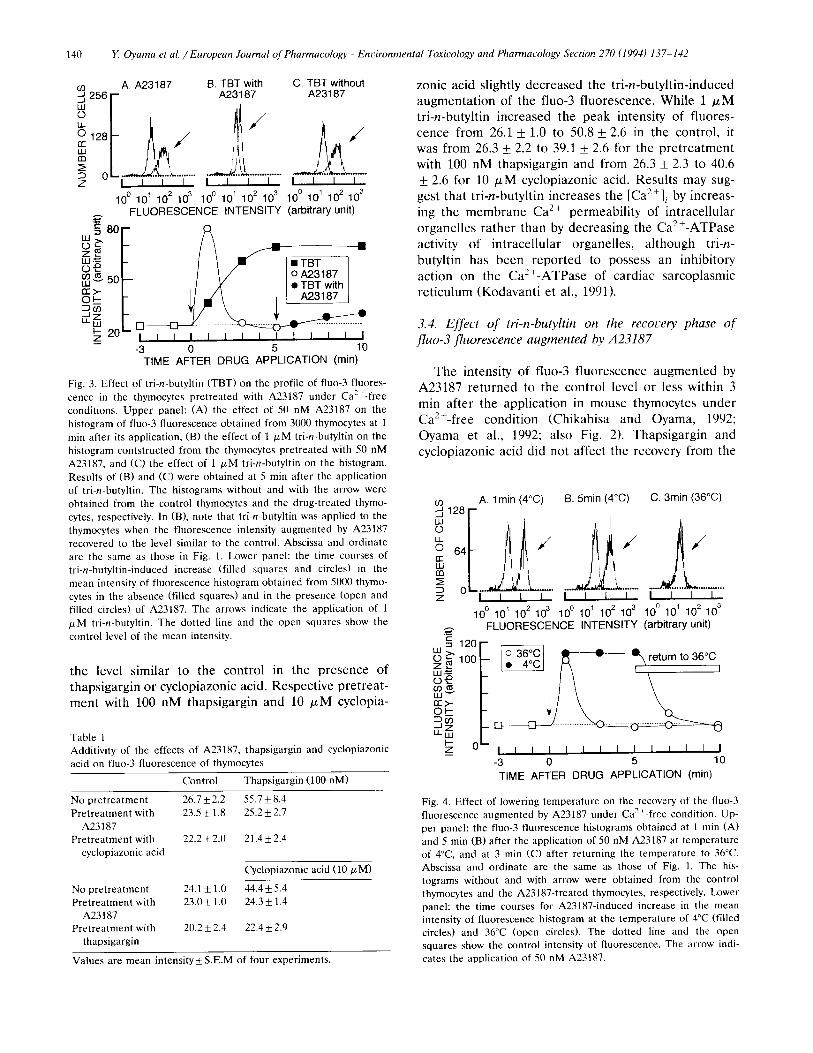
was significant (P < 0.01). On the contrary, the pre- treatment with 1 ~M tri-n-butyltin produced only a slight decrease in the fluorescence augmented by A23187. The peak intensities of fluorescence achieved by 50 nM A23187 were 64.9 _+ 12.4 in the control and 57.8 ___ 9.8 in the pretreatment with 1 ~M tri-n-butyltin. Pretreatment with 50 nM A23187 alomost completely suppressed the augmentations of fluo-3 fluorescence by 100 nM thapsigargin and 10 # M cyclopiazonic acid (Table 1).
3.3. Pretreatment with thapsigargin and cyclopiazonic acid on tri-n-butyltin-induced increase in the [Ca 2 +]i
Pretreatment of thymocytes with 100 nM thapsigar- gin or 10/xM cyclopiazonic acid abolished the fluores- cence response elicited by each other (Table 1). Thus, their effects on the [Ca2+]i were not additive as previ- ously shown in rat thymocytes (Manson et al., 1991a). Therefore, the same site of intracellular organelles in mouse thymocytes seems to be affected by thapsigargin and cyclopiazonic acid.
Tri-n-butyltin was added into the cell suspension after the intensity of fluo-3 fluorescence recovered to
140 Y. Oyama et al. / European Journal of Pharmacology - Em,ironmental Toxicology and Pharmacology Section 270 (1994) 137-142
A. A23187 uJ ~ 256 f 0
~) 1 2 8 ~ ~ /
r n
~ 0 '- . . . . . . . . . z I I I I
B. TBT with C. TBT without A23187 A23187
. . . . . .
I I I I I I I I 10 0 101 10 2 10 3 10 o 101 10 2 10 3 10 o 101 10 2 10 3
==8o ILl>, Z ~
~ 50 rr>- O~- -JZ U-LU
~ 2o
FLUORESCENCE INTENSITY (arbitrary unit)
I Z I • TBT with
I I I I I I I I I I I I I I -3 0 5 10
TIME AFTER DRUG APPLICATION (min)
Fig. 3. Effect of tri-n-butyltin (TBT) on the profile of fluo-3 fluores- cence in the thymocytes pretreated with A23187 under Ca2+-free conditions. Upper panel: (A) the effect of 50 nM A23187 on the histogram of fluo-3 fluorescence obtained from 3000 thymocytes at 1 rain after its application, (B) the effect of 1 /xM tri-n-butyltin on the histogram contstructed from the thymocytes pretreated with 50 nM A23187, and (C) the effect of 1 txM tri-n-butyltin on the histogram. Results of (B) and (C) were obtained at 5 min after the application of tri-n-butyltin. The histograms without and with the arrow were obtained from the control thymocytes and the drug-treated thymo- cytes, respectively. In (B), note that tri-n-butyltin was applied to the thymocytes when the fluorescence intensity augmented by A23187 recovered to the level similar to the control. Abscissa and ordinate are the same as those in Fig. 1. Lower panel: the time courses of tri-n-butyltin-induced increase (filled squares and circles) in the mean intensity of fluorescence histogram obtained from 5000 thymo- cytes in the absence (filled squares) and in the presence (open and filled circles) of A23187. The arrows indicate the application of 1 /xM tri-n-butyltin. The dotted line and the open squares show the control level of the mean intensity.
t h e l eve l s i m i l a r to t h e c o n t r o l in t h e p r e s e n c e o f
t h a p s i g a r g i n o r c y c l o p i a z o n i c ac id . R e s p e c t i v e p r e t r e a t -
m e n t w i t h 100 n M t h a p s i g a r g i n a n d 10 p ,M cyc lop i a -
Table 1 Additivity of the effects of A23187, thapsigargin and cyclopiazonic acid on fluo-3 fluorescence of thymocytes
Control Thapsigargin (100 nM)
No pretreatment 26.7 + 2.2 55.7 + 8.4 Pretreatment with 23.5 _+ 1.8 25.2 _+ 2.7
A23187 Pretreatment with 22.2 _+ 2.0 21.4 + 2.4
cyclopiazonic acid
Cyclopiazonic acid (10/xM)
No pretreatment 24.1 +_ 1.0 44.4_+ 5.4 Pretreatment with 23.0 _+ 1.0 24.3 _+ 1.4
A23187 Pretreatment with 20.2 _+ 2.4 22.4 _+ 2.9
thapsigargin
Values are mean intensity_+ S.E.M of four experiments.
z o n i c ac id s l igh t ly d e c r e a s e d t h e t r i - n - b u t y l t i n - i n d u c e d
a u g m e n t a t i o n o f t h e f luo-3 f l u o r e s c e n c e . W h i l e 1 / xM
t r i - n - b u t y l t i n i n c r e a s e d t h e p e a k i n t e n s i t y o f f l u o r e s -
c e n c e f r o m 26.1 _+ 1.0 to 50.8 + 2.6 in t h e c o n t r o l , it
was f r o m 26.3 + 2.2 to 39.1 _+ 2.6 fo r t h e p r e t r e a t m e n t
w i t h 100 n M t h a p s i g a r g i n a n d f r o m 26.3 +_ 2.3 to 40.6
+_ 2.6 fo r 10 / xM c y c l o p i a z o n i c acid . R e s u l t s m a y sug-
ge s t t h a t t r i - n - b u t y l t i n i n c r e a s e s t h e [Ca2+] i by i n c r e a s -
ing t h e m e m b r a n e C a 2+ p e r m e a b i l i t y o f i n t r a c e l l u l a r
o r g a n e l l e s r a t h e r t h a n by d e c r e a s i n g t h e C a 2 + - A T P a s e
ac t iv i ty o f i n t r a c e l l u l a r o r g a n e l l e s , a l t h o u g h t r i -n -
b u t y l t i n h a s b e e n r e p o r t e d to p o s s e s s a n i n h i b i t o r y
a c t i o n o n t h e C a 2 + - A T P a s e o f c a r d i a c s a r c o p l a s m i c
r e t i c u l u m ( K o d a v a n t i e t al., 1991).
3.4. Effect o f tri-n-butyltin on the recouery phase o f fluo-3 fuorescence augmented by A23187
T h e i n t e n s i t y o f f luo -3 f l u o r e s c e n c e a u g m e n t e d by
A 2 3 1 8 7 r e t u r n e d to t h e c o n t r o l leve l o r less w i t h i n 3
m i n a f t e r t h e a p p l i c a t i o n in m o u s e t h y m o c y t e s u n d e r
C a 2 + - f r e e c o n d i t i o n ( C h i k a h i s a a n d O y a m a , 1992;
O y a m a e t al., 1992; a l so Fig. 2). T h a p s i g a r g i n a n d
c y c l o p i a z o n i c ac id d id n o t a f f ec t t h e r e c o v e r y f r o m t h e
A. lmin (4°C) 1281-
,,=,
. . . . . . . .
z I I I I
rr~. O ~ -Jz U-LU
F- z
B. 5min (4°C) C. 3min (36°C)
2: I L I I I I I I
100 101 102 103 100 101 102 103 10 o 101 102 103 FLUORESCENCE INTENSITY (arbitrary unit)
120 - 100 ~ ~ return to 36°C
[] . . . . . . @
- I I I I I I I I I I I I I I -3 0 5 10 TIME AFTER DRUG APPLICATION (min)
Fig. 4. Effect of lowering temperature on the recovery of the fluo-3 fluorescence augmented by A23187 under Ca2+-free condition. Up- per panel: the fluo-3 fluorescence histograms obtained at 1 min (A) and 5 min (B) after the application of 50 nM A23187 at temperature of 4°C, and at 3 min (C) after returning the temperature to 36°C. Abscissa and ordinate are the same as those of Fig. 1. The his- tograms without and with arrow were obtained from the control thymocytes and the A23187-treated thymocytes, respectively. Lower panel: the time courses for A23187-induced increase in the mean intensity of fluorescence histogram at the temperature of 4°C (filled circles) and 36°C (open circles). The dotted line and the open squares show the control intensity of fluorescence. The arrow indi- cates the application of 50 nM A23187.

Y Oyama et al./European Journal of Pharmacology - Environmental Toxicology and Pharmacology Section 270 (1994) 137-142 141
A. I rain
z
UJ 0 Z > - < F - T ~ O z
I----- 5___ W n-
B. 3min C. 5min
i i ~ I . I I ~ l I I I I 100 101 10 2 10 ~ 100 101 10 2 10 3 100 101 10 2 103
FLUORESCENCE INTENSITY (arbitrary unit)
1.0 " O ~ •Control I [] 100 nM I • 300 a M ]
0
-0.2 m
I I I I I I I I I I 1 5 10
TIME AFTER A23187 APPLICATION (min)
Fig. 5. Effect of tri-n-butyltin on the recovery of fluo-3 fluorescence augmented by A23187 under CaZ+-free conditions. Upper panel: the recovery of the histogram of fluo-3 fluorescence augmented by 50 nM A23187 in the presence of 300 nM tri-n-butyltin. Histogram without the arrow was obtained from 3000 thymocytes treated with 300 nM tri-n-butyltin for 10 min before the application of 50 nM A23187. Histograms indicated with the arrow were obtained at 1 min (A), 3 rain (B) and 5 min (C) after the application of A23187. Abscissa and ordinate are the same as those of Fig. 1. Lower panel: the time course of the recovery of the mean intensity of fluo-3 fluorescence augmented by A23187 in the absence (filled squares) and presence of 100 nM (open squares) and 300 nM (filled circles) tri-n-butyltin. Ordinate shows the normalized augmentation of fluo-3 fluorescence induced by A23187. The zero at the ordinate (with the dotted line) means the intensity before the application of A23187. Result, a representative one of three experiments, was obtained from same cell suspension.
A23187-induced increase in the fluo-3 fluorescence. However, the recovery process was completely inhib- ited by lowering the incubation temperature to 1-4°C (Fig. 4). Result plausibly suggests an involvement of Ca 2+ pump activity of cell membrane, but not Ca 2+ pump activity of intracellular Ca 2+ store sites, in the recovery process of fluo-3 fluorescence.
While the fluorescence augmented by 50 nM A23187 returned to the control level or less within 3 rain after the application in the absence of tri-n-butyltin (Fig. 2), the fluorescence returned to the control level within 5 rain in the presence of 100 nM tri-n-butyltin and it required 10 min or more in the case of 300 nM tri-n- butyltin (Fig. 5). Tri-n-butyltin at 300 nM also delayed the recovery process of fluorescence augmented by 100 nM thapsigargin and 10 /xM cyclopiazonic acid (not shown). Effect of 1 /xM tri-n-butyltin on the recovery process of A23187-induced augmentation of fluo-3 flu- orescence was not examined since the fluorescence intensity before application of A23187 was too high to examine the recovery process in the presence of 1 ~M
tri-n-butyltin. However, the result suggests the possibil- ity that tri-n-butyltin at concentrations of 100 nM or more decreases the activity of membrane Ca 2+ pump of mouse thymocytes.
4. Discussion
We have previously reported that tri-n-butyltin greatly increases the [Ca2+]i of mouse thymocytes and rat cerebellar neurons in the presence of external Ca 2+ and the removal of external Ca 2+ greatly reduces the effect of tri-n-butyltin in increasing the [Ca2+]i, sug- gesting the modification of Ca 2+ permeability of cell membrane by tri-n-butyltin (Chikahisa and Oyama, 1992; Oyama et al., 1993). The present study has re- vealed another mechanism of tri-n-butyltin for increas- ing the [CaZ+],, being independent of the external Ca 2+. Thus, it is likely that 1/xM tri-n-butyltin releases Ca 2+ from the intracellular organelles (calcium stores) and simultaneously decreases the activity of Ca 2+ pump of cell membrane, resulting in a persistent increase in the [Ca;+]i of mouse thymocytes under external Ca 2+- free condition, as discussed below.
The pretreatment of thymocytes with A23187 greatly reduced the tri-n-butyltin-induced increase in the [CaZ+]i under external Ca2+-free condition (Fig. 3). It has been reported that A23187 increases the Ca 2+ permeability of intracellular store sites more selectively than the cell membrane (Itoh et al., 1985) and the increased [CaZ+] i is lowered by the membrane Ca 2+ pump even in the presence of a Ca 2+ ionophore (Fur- ukawa et al., 1989). This seems to be also the case for mouse thymocytes since A23187 produced only a tran- sient increase in the [Ca2+]i under Ca2+-free condi- tions (Fig. 2). On the contrary, pretreatment with thap- sigargin and cyclopiazonic acid slightly decreased the tri-n-butyltin-induced increase in the [Ca2+]~. There- fore, it is suggested that tri-n-butyltin may increase the [Ca~'+]~ of thymocytes by increasing the Ca 2+ perme- ability of the intracellular organelles rather than by inhibiting the Ca2+-ATPase of intracellular organells.
The recovery of the [Ca2+]i increased by A23187 under Ca2+-free conditions occurred in the presence of thapsigargin and cyclopiazonic acid. Therefore, the Ca 2+ pump of intracellular organelles was unlikely to be involved in the recovery process. Tri-n-butyltin at concentrations of 100 nM and 300 nM delayed the recovery process of the [CaZ+]~ increased by A23187 (Fig. 5). These results may suggest a decreased Ca 2+ pump activity of thymocyte membrane in the presence of tri-n-butyltin, resulting in a persistent increase in the [ C a 2 + ] i o f mouse thymocytes.
From the results of this study, it is likely that tri-n- butyltin releases Ca z+ from the intracellular stores, probably resulting in a decreased calcium content of

142 Y. Oyama et al. / European Journal of Pharmacology - Em~ironmental Toxicology and Pharmacology Section 270 (1994) 137-142
i n t r a c e l l u l a r s tores . It has b e e n p r o p o s e d tha t t he
c a l c i u m c o n t e n t o f i n t r a c e l l u l a r o r g a n e l l e s ( in t r ace l lu -
lar c a l c i u m s to res ) is an i m p o r t a n t d e t e r m i n a n t for t h e
Ca 2+ p e r m e a b i l i t y in t he m e m b r a n e o f u n s t i m u l a t e d
t h y m o c y t e s ( M a n s o n et al., 1991a,b). T h e r e f o r e , it is
p l aus ib l e to sugges t t ha t t r i -n -bu ty l t in ind i r ec t ly in-
c r e a s e s t h e Ca 2+ p e r m e a b i l i t y o f t h y m o c y t e m e m b r a n e
by r e d u c i n g the c a l c i u m c o n t e n t o f i n t r a c e l l u l a r s tores .
A n e l e v a t i o n o f t h e [Ca2+] i is o n e o f t h e ea r l i e s t
e v e n t s fo l l owing m i t o g e n i c s t i m u l a t i o n o f l y m p h o c y t e s
( T s i e n et al., 1982). T h e ini t ial r ise in t he [Ca2+]i is
u n a f f e c t e d by r e m o v a l o f ex t e rna l Ca 2+, wh i l e t he
s e c o n d p h a s e o f t he r e s p o n s e is a b o l i s h e d by the re-
mova l o f e x t e r n a l Ca 2+. T h e m o d u l a t i o n o f [Ca2+]i o f
l y m p h o c y t e s fo l l owing m i t o g e n i c s t i m u l a t i o n m a y be
d i s t u r b e d in t he p r e s e n c e o f t r i -n -bu ty l t in , r e s u l t i n g in
t he m a l f u n c t i o n o f l ymphocy t e s . F u r t h e r m o r e , it has
b e e n d e m o n s t r a t e d tha t a p o p t o s i s is d e p e n d e n t on a
s u s t a i n e d i n c r e a s e in t h e [Ca2+]~ ( M c C o n k e y et al.,
1989). T h e r e f o r e , a s u s t a i n e d e l e v a t i o n o f t he [Ca2+]i
c a u s e d by t r i -n -bu ty l t in may ac t iva te a p o p t o s i s o f thy-
mocy tes . T h i s poss ib i l i ty is s u p p o r t e d by t h e fact t ha t
b i s ( t r i -n -bu ty l t i n )ox ide at c o n c e n t r a t i o n s o f 0 . 1 - 1 p ,M
i n d u c e d the a p o p t o s i s o f ra t t h y m o c y t e s ( R a f f r a y and
C o h e n , 1991).
5. Acknowledgements
W e t h a n k the r e f e r e e s fo r t he i r v a l u a b l e sugges t i ons
on this m a n u s c r i p t . T h i s s tudy was s u p p o r t e d by the
r e s e a r c h g r a n t a w a r d e d to Y. O y a m a f r o m t h e N i p p o n
Li fe I n s u r a n c e F o u n d a t i o n (Osaka , J apan ) .
6. References
Chikahisa, L. and Y. Oyama, 1992, Tri-n-butyltin increases intracel- lular Ca 2+ in mouse thymocytes, Pharmacol. Toxicol. 71, 190.
Furukawa, K, Y. Tawada-Iwata and Shigekawa, M., 1989, Modula- tion of plasma membrane Ca 2+ pump by membrane potential in cultured vascular smooth muscle cells, J. Biochem. 106, 1068.
ltoh, T., Y. Kanmura and H. Kuriyama, 1985, A23187 increases
calcium permeability of store sites more than of surface mem- branes in the rabbit mesenteric artery, J. Physiol. 358, 467.
Kao, J.P.Y., A.T. Haroontunian and R.Y. Tsein, 1989, Photochemi- cally generated cytosolic calcium pulses and their detection by fluo-3, J. Biol. Chem. 264, 8179.
Kodavanti, P.R.S., J.A. Cameron, P.R. Yallapragada, P.J.S. Vig and D. Desaiah, 1991, Inhibition of Ca2+-transport associated with cAMP-dependent protein phosphorylation in rat cardiac sar- coplasmic reticulum by triorganotins, Arch. Toxicol. 65, 311.
Luijten, J.G., 1971, Applications and biological effects of organotin compounds, in: Organotin Compounds, ed. A.K. Sawyer (Decker, New York) p. 931.
Manson, M.J., M.P. Mahaut-Smith and S. Grinstein, 1991a, The role of intracellular Ca 2+ in the regulation of the plasma membrane Ca :+ permeability of unstimulated rat lymphocytes, J. Biol. Chem. 266, 10872.
Manson, M.J., C. Garcia-Rodriguez and S. Grinstein, 1991b, Cou- pling between intracellular Ca 2+ stores and the Ca 2+ permeabil- ity of the plasma membrane, J. Biol. Chem. 266, 20856.
McConky, D.J., P. Nicotera, P. Hartzell, G. Bellomo, A.H. Wyllie and S. Orrenius, Gluocorticoids activate a suicide process in thymocytes through an increase in cytosolic Ca2 + concentration, Arch. Biochem. Biophys. 269, 365.
Oyama, Y., L. Chikahisa, K. Noda, F. Tomiyoshi and H. Hayashi, 1992, Characterization of the triphenyltin-induced increase in intracellular Ca 2÷ of mouse thymocytes: Comparison with the action of A23187, Jpn. J. Pharmacol. 60, 159.
Oyama, Y., T. Ueha and A. Hayashi, 1993, Effect of tri-n-butyltin on in t racel lu lar Ca 2+ concentration of rat cerebellar neurons, Eur. J. Pharmacol. 258, 89.
Raffray, M. and G.M. Cohen, 1991, Bis(tri-n-butyltin)oxide induces programmed cell death (apoptosis) in immature rat thymocytes, Arch. Toxicol. 65, 135.
Seidler, N.W., I. Jova, M. Vegh and A. Martonosi, 1989, Cyclopia- zonic acid is a specific inhibitor of the Ca 2 ~-ATPase of sarcoplas- mic reticulum, J. Biol. Chem. 264, 17816.
Sheldon, A.W., 1975, Effects of organotin anti-fouling coatings on man and his environment, J. Paint Technol. 47, 54.
Snoeij, N.J., A.H. Penninks and W.Seinen, 1987, Biological activity of organotin compounds - an overview, Environ. Res. 44, 335.
Thastrup, O., 1990, Role of Ca2+-ATPases in regulation of cellular Ca 2~ signaling, as studied with the selective microsomal Ca 2+-
ATPase inhibitor, thapsigargin, Agents Actions 29, 8. Tsien, R,Y, T. Pozzan and T.J. Rink, 1982, T-cell mitogens cause
early changes in cytoplasmic free Ca 2+ and membrane potential in lymphocytes, Nature 295, 68.
Van der Kerk, G.J.M. and J.G.A. Luiiten, 1958, Investigations on organotin compounds. III. The biocidal properties of organotin compounds, J. Appl. Chem. 4, 314.
Wilkinson, R.R., 1984, Technoeconomic and environmental assess- ment of industrial organotin compounds, Neurotoxicology 5, 141.







![Evidence of Ca2+-Dependent Carbohydrate Association ... · Ca2+I2+ and [2Lex + Ca2+]2+. The CID experiments of the [2Lex-LacCer + Ca2+I2+ dimers resulted in a neutral loss covalently](https://static.fdocuments.net/doc/165x107/5f8af1f17b5f935beb015692/evidence-of-ca2-dependent-carbohydrate-association-ca2i2-and-2lex-ca22.jpg)

![Thymoglobulin (globuline anti-thymocytes [Lapin])](https://static.fdocuments.net/doc/165x107/586dfe2c1a28ab114f8babb9/thymoglobulin-globuline-anti-thymocytes-lapin.jpg)