
Effect of potassium, sodium and its mixture ... · Effect of potassium, sodium and its mixture...
8
~ 270 ~ Journal of Entomology and Zoology Studies 2014; 2 (6): 270-277 ISSN 2320-7078 JEZS 2014; 2 (6): 270-277 © 2014 JEZS Received: 27-10-2014 Accepted: 02-12-2014 Essawy M. M. Economic entomology Department, Faculty of Agricultural, Kafr El- Sheikh University, Egypt. Saad I. A. I Department of Sericulture, Plant Protection Institute, Agricultural Research Center, Giza, Egypt. Correspondence: Essawy M. M. Economic entomology Department, Faculty of Agricultural, Kafr El- Sheikh University, Egypt. Effect of potassium, sodium and its mixture supplementation on: II- differential haemocyte counts and absolute differential haemocyte counts of Mulberry Silkworm Bombyx mori (L.). Essawy M. M., Saad I. A. I. Abstract Two bio salt minerals, potassium sulphate, sodium sulphate and a mixture of both bio salts at the effective concentration of 1% which were added to the mulberry leaves of silkworm B. mori L., to evaluate their effect on the differential haemocyte counts (DHC) and absolute differential haemocyte counts (ADHC). Daily changes of five haemocyte types were investigated as, prohaemocytes, plasmatocytes, coagulocytes, spherules and oenocytoides. The data revealed that Larvae treated with potassium sulphate positively affected the DHC and ADHC of prohaemocytes during the 8 th day of mature stage. Larvae treated with potassium and sodium sulphate mixture presented the maximum increase of DHC and ADHC of plasmatocytes during the end of larval stage. The DHC of coagulocytes was fluctuated during the last larval instar in both treated and untreated larvae but ADHC of treated larvae with sodium sulphate was higher than control during the most developmental periods of the last larval instar. When larvae treated with sodium sulphate an increase in spherule cell ADHC was observed during the larval stage , the maximum peak was (61x103 cell/larva) at 5 th day in comparison to (28 x103 cell/larva) of control. Larvae treated with potassium sulphate showed also a maximum peak (81x103cell/larva) comparing to (26x103 cell/larva) during the 10 th day for control. Larvae treated with potassium sulphate presented an increases of DHC of oenocytoids about (3.1%) in comparison to (1 and 2%) in untreated larvae during the 4 th and 7 th days, and the ADHC trend was positively enhanced during the last larval instar of treated larvae. Keywords: Bombyx mori (L.) - Hemocyte types - Differential haemocyte counts (DHC) - Absolute differential haemocyte counts (ADHC). 1. Introduction Mulberry silkworm is the most important sources of natural silk in the world. Much research has been done on the diet supplementation of mulberry leaves fed to silkworms. Although most of the insect species have similar nutritional needs due to the similarities between their metabolisms [1] , the silkworm’s nutritive requirements are very different and they are almost exclusively covered by the use of mulberry leaves, due to the presence of morin [2] . Mulberry leafs contain minerals up to 10 - 28% of the larval structure of the silkworm, in different ages, is represented by the absorbed minerals [3] . Minerals are major constituents of the silkworm’s diets, playing a crucial role in osmotic pressure regulation of the intra and extracellular liquids and participating as co-factors in different enzyme systems. The production parameters of silkworms depend on the larval nutrition and health status. In order to improve these production parameters, both in quantity and quality, a large number of minerals have been used, many studies focusing on the effect of these minerals on silkworms [4, 5, 6, 7, 8, 9-10, 11, 12, 13, 14-15] . Haemocytes perform various physiological functions in the body of insect. They direct nutrients to various tissues and store them. They perform phagocytosis, encapsulation of foreign bodies in the insect body cavity, coagulation to prevent loss of blood, nodule formation, transport of food materials, hormones and detoxification of metabolites and biological active materials [16] . Five haemocyte types were identified in the haemolymph of Lepidopterous larvae: prohemocytes, plasmatocytes, granulocytes (Coagulocytes), spherulocytes and oenocytoides [17, 18, 19, 20-21] . As potassium, sodium and its mixture supplementation enhanced blood volume, total and absolute haemocyte counts in mulberry silkworm Bombyx mori (L.) at low concentration [22] .
Transcript of Effect of potassium, sodium and its mixture ... · Effect of potassium, sodium and its mixture...

~ 270 ~
Journal of Entomology and Zoology Studies 2014; 2 (6): 270-277 ISSN 2320-7078 JEZS 2014; 2 (6): 270-277 © 2014 JEZS Received: 27-10-2014 Accepted: 02-12-2014 Essawy M. M. Economic entomology Department, Faculty of Agricultural, Kafr El-Sheikh University, Egypt. Saad I. A. I Department of Sericulture, Plant Protection Institute, Agricultural Research Center, Giza, Egypt. Correspondence: Essawy M. M. Economic entomology Department, Faculty of Agricultural, Kafr El-Sheikh University, Egypt.
Effect of potassium, sodium and its mixture
supplementation on: II- differential haemocyte counts and absolute differential haemocyte counts
of Mulberry Silkworm Bombyx mori (L.). Essawy M. M., Saad I. A. I. Abstract Two bio salt minerals, potassium sulphate, sodium sulphate and a mixture of both bio salts at the effective concentration of 1% which were added to the mulberry leaves of silkworm B. mori L., to evaluate their effect on the differential haemocyte counts (DHC) and absolute differential haemocyte counts (ADHC). Daily changes of five haemocyte types were investigated as, prohaemocytes, plasmatocytes, coagulocytes, spherules and oenocytoides. The data revealed that Larvae treated with potassium sulphate positively affected the DHC and ADHC of prohaemocytes during the 8th day of mature stage. Larvae treated with potassium and sodium sulphate mixture presented the maximum increase of DHC and ADHC of plasmatocytes during the end of larval stage. The DHC of coagulocytes was fluctuated during the last larval instar in both treated and untreated larvae but ADHC of treated larvae with sodium sulphate was higher than control during the most developmental periods of the last larval instar. When larvae treated with sodium sulphate an increase in spherule cell ADHC was observed during the larval stage , the maximum peak was (61x103 cell/larva) at 5th day in comparison to (28 x103 cell/larva) of control. Larvae treated with potassium sulphate showed also a maximum peak (81x103cell/larva) comparing to (26x103 cell/larva) during the 10th day for control. Larvae treated with potassium sulphate presented an increases of DHC of oenocytoids about (3.1%) in comparison to (1 and 2%) in untreated larvae during the 4th and 7th days, and the ADHC trend was positively enhanced during the last larval instar of treated larvae.
Keywords: Bombyx mori (L.) - Hemocyte types - Differential haemocyte counts (DHC) - Absolute differential haemocyte counts (ADHC). 1. Introduction Mulberry silkworm is the most important sources of natural silk in the world. Much research has been done on the diet supplementation of mulberry leaves fed to silkworms. Although most of the insect species have similar nutritional needs due to the similarities between their metabolisms [1], the silkworm’s nutritive requirements are very different and they are almost exclusively covered by the use of mulberry leaves, due to the presence of morin [2]. Mulberry leafs contain minerals up to 10 - 28% of the larval structure of the silkworm, in different ages, is represented by the absorbed minerals [3]. Minerals are major constituents of the silkworm’s diets, playing a crucial role in osmotic pressure regulation of the intra and extracellular liquids and participating as co-factors in different enzyme systems. The production parameters of silkworms depend on the larval nutrition and health status. In order to improve these production parameters, both in quantity and quality, a large number of minerals have been used, many studies focusing on the effect of these minerals on silkworms [4, 5, 6, 7, 8, 9-10, 11, 12, 13,
14-15]. Haemocytes perform various physiological functions in the body of insect. They direct nutrients to various tissues and store them. They perform phagocytosis, encapsulation of foreign bodies in the insect body cavity, coagulation to prevent loss of blood, nodule formation, transport of food materials, hormones and detoxification of metabolites and biological active materials [16]. Five haemocyte types were identified in the haemolymph of Lepidopterous larvae: prohemocytes, plasmatocytes, granulocytes (Coagulocytes), spherulocytes and oenocytoides [17, 18, 19, 20-21]. As potassium, sodium and its mixture supplementation enhanced blood volume, total and absolute haemocyte counts in mulberry silkworm Bombyx mori (L.) at low concentration [22].

~ 271 ~
Journal of Entomology and Zoology Studies
The present study will complete the previous results to deal with the effect of biosalt (potassium, sodium sulphate and its mixture) at the effective concentration [22] on differential and absolute differential haemocyte counts. 2. Materials and methods 2.1. Insects Chinese hybrid (9F7X) of mulberry silkworm Bombyx mori L., imported from China was used during the present study. Larvae were reared at Kafr El-Sheikh Faculty of Agriculture under the laboratory hygrothermal conditions of 25±2 0C and 65-70 R.H., according to the conventional method in trays and provided with suitable amounts of fresh mulberry leaves [23]. All larvae which molted to the last instar at the same time were grouped and used in the experiments. The determination of the last larval instar was determined by (zero hour) from the last larval ecdysis [24]. 2.2. Biosalt minerals compounds as food additives The mineral salt compounds potassium sulphate anhydrous (k2SO4), sodium sulphate anhydrous (Na2SO4) (99%), provided from El Nasr Pharmaceutical Chemicals Co. (ADWIC), Egypt. A mixture of both sodium and potassium sulphate (1:1) were also used as supplementary nutrient. The bio salt crystals were dissolved in distilled water and diluted to 1% concentrations to determine the haemocyte counts. Fresh mulberry leaves were soaked for 15 minutes at the concentration then left to dry. Treated mulberry leaves were offered to silkworm larvae at the first day of last larval instar for one day only at three times. While during the next days until the end of last instar fed on untreated leaves. Control larvae were fed on mulberry leaves soaked in distilled water only for one day also. 2. 3 Differential haemocyte counts (DHC) The haemolymph was allowed to fall on five clean glass slide every day, and a smear was made [25-26]. The smears were allowed to dry and were stained by using Giemsa and Wright's blood stain according to [25, 27-28]. The smear was examined under oil immersion and a maximum of 100 haemocytes\slide were differentiated according to the classification of [17, 18 - 24]. 2.4 Absolute differential haemocyte counts (ADHC) The ADHC's were calculated from the DHC and the AHC [22] which is the first part of the present work for each treated and untreated larva. 3. Results and discussion Based on previous classification [19-24], five types of hemocytes were identified in the 5th instar larvae of Bombyx mori. They are the prohemocytes (PRs), plasmatocytes (PLs), coagulocytes (COs) which were found as coagulocytes with and without granules, spherule cells (SPs), oenocytoids (OEs). Two more shapes, the vermiform (VEs) and podocytes (POs) have also been observed and they were grouped as a forms of plasmatocytes. 3.1 Differential and absolute differential haemocyte counts (DHC and ADHC) Differential haemocyte counts DHC and absolute differential haemocyte counts ADHC were counted in both untreated and
treated larvae at the low concentration 1% which enhanced the biological parameters in our previous work [22]. 3.1.1. Prohaemocytes Prohaemocytes appears mostly rounded but some are oval, and they are the smallest type of the haemocytes, with characteristically large and filling nucleus that reduce the cytoplasm to a narrow peripheral band. No cytoplasmic projection found in prohaemocytes and usually found in divission. Their cytoplasm is strongly basophilic (Figure). Treated larvae with potasium sulphate showed a non significant peak of prohaemocyte counts during the last larval instar at the 8th day before the maturation of larvae. At the10th day DHC decreased to (4±1%) but it reached a maximum (8±2%) in control.
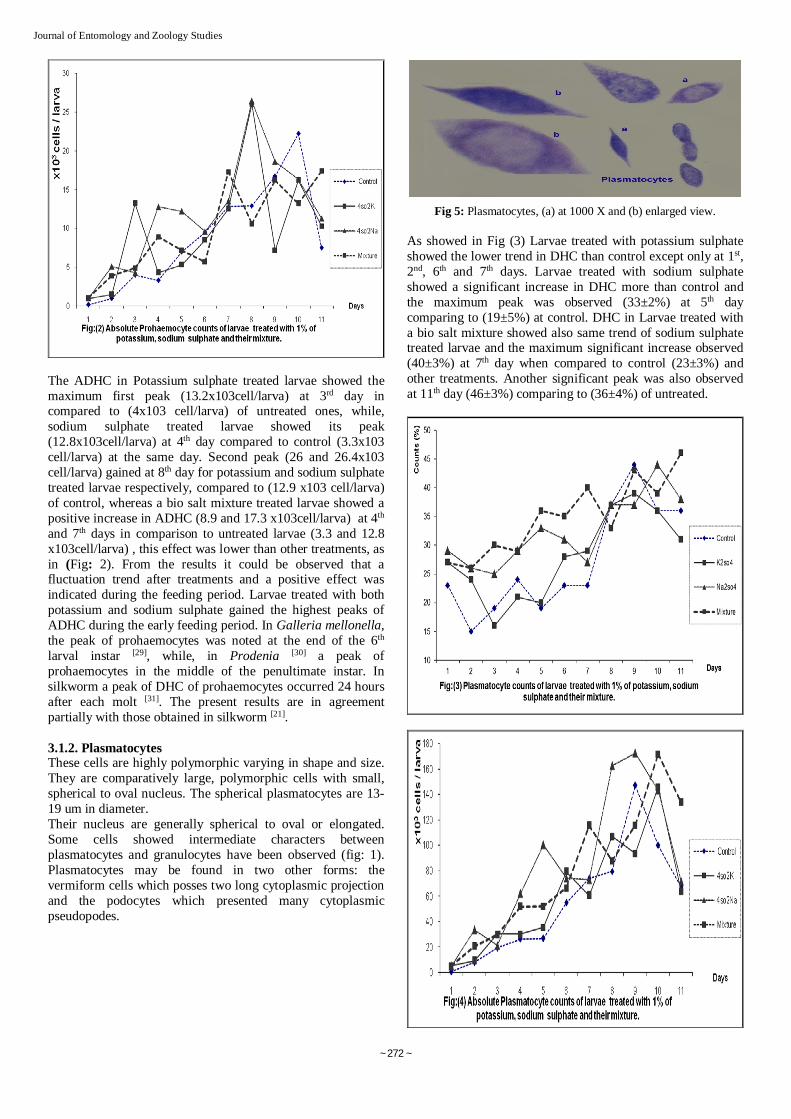
Fig 1: Prohaemocytes (a) at 1000 X and (b) enlarged view. A similar trend was observed in larvae treated with sodium sulphate. Prohaemocyte percentage showed a significant increase at 4rd and 11th day. The larvae treated with a bio salt mixture of potassium and sodium sulphate recorded the same percentage of DHC (6±2%) at 1st, 7th, 9th and 11th days as in Fig (1).
~ 272 ~
Journal of Entomology and Zoology Studies
The ADHC in Potassium sulphate treated larvae showed the maximum first peak (13.2x103cell/larva) at 3rd day in compared to (4x103 cell/larva) of untreated ones, while, sodium sulphate treated larvae showed its peak (12.8x103cell/larva) at 4th day compared to control (3.3x103 cell/larva) at the same day. Second peak (26 and 26.4x103 cell/larva) gained at 8th day for potassium and sodium sulphate treated larvae respectively, compared to (12.9 x103 cell/larva) of control, whereas a bio salt mixture treated larvae showed a positive increase in ADHC (8.9 and 17.3 x103cell/larva) at 4th and 7th days in comparison to untreated larvae (3.3 and 12.8 x103cell/larva) , this effect was lower than other treatments, as in (Fig: 2). From the results it could be observed that a fluctuation trend after treatments and a positive effect was indicated during the feeding period. Larvae treated with both potassium and sodium sulphate gained the highest peaks of ADHC during the early feeding period. In Galleria mellonella, the peak of prohaemocytes was noted at the end of the 6th larval instar [29], while, in Prodenia [30] a peak of prohaemocytes in the middle of the penultimate instar. In silkworm a peak of DHC of prohaemocytes occurred 24 hours after each molt [31]. The present results are in agreement partially with those obtained in silkworm [21]. 3.1.2. Plasmatocytes These cells are highly polymorphic varying in shape and size. They are comparatively large, polymorphic cells with small, spherical to oval nucleus. The spherical plasmatocytes are 13-19 um in diameter. Their nucleus are generally spherical to oval or elongated. Some cells showed intermediate characters between plasmatocytes and granulocytes have been observed (fig: 1). Plasmatocytes may be found in two other forms: the vermiform cells which posses two long cytoplasmic projection and the podocytes which presented many cytoplasmic pseudopodes.
Fig 5: Plasmatocytes, (a) at 1000 X and (b) enlarged view. As showed in Fig (3) Larvae treated with potassium sulphate showed the lower trend in DHC than control except only at 1st, 2nd, 6th and 7th days. Larvae treated with sodium sulphate showed a significant increase in DHC more than control and the maximum peak was observed (33±2%) at 5th day comparing to (19±5%) at control. DHC in Larvae treated with a bio salt mixture showed also same trend of sodium sulphate treated larvae and the maximum significant increase observed (40±3%) at 7th day when compared to control (23±3%) and other treatments. Another significant peak was also observed at 11th day (46±3%) comparing to (36±4%) of untreated.

~ 273 ~
Journal of Entomology and Zoology Studies
ADHC in larvae treated with potassium sulphate showed a gradual increase, the first peak (100.5 x103 cell/larva) gained at 5th day of feeding period in comparison to (26.8 x103cell/larva) of control. The second peak was (172.4 x103cell/larva) gained at 9th day in comparison to (147.1x103 cell/larva) of untreated. Larvae treated with a bio salt mixture showed the first ADHC's maximum peak during 7th day of feeding period (115.4 x103cell/larva) compared to (73.8 x103cell/larva) of control and the second peak was (171.8 x103cell/larva) in compared to (99.9 x103cell/larva) of control at 10th day as in (Fig: 4). In Philosamia ricini, DHC of plasmatocytes in untreated and treated larvae with IGR's presented its maximum peak at prepupal and pupal stage. While, in treated insects the ADHC was higher than control from the beginning of last instar until feeding period, the maximum value was occurred at the 9th day in both treated and untreated larvae. In Eri-silkworm [32, 33]; and in B. mori [30, 31]. ADHC's of plasmatocytes were greater in treated larvae than untreated in most developmental periods which suggests that the immune response of treated larvae is more than untreated [34]. 3.1.3 Coagulocytes Coagulocytes in present study are recognized as spherical or oval cells which vary considerably in size. Their nucleus are relatively small, and the cytoplasm contains numerous cytoplasmic inclusions (granules). In many coagulocytes the cytoplasm were filled with granules (coaguloctes with granules) and in some others little number of inclusions were found (coaguloctes without granules). Under the light microscope granules were found in different sizes and shapes (Fig: 2).
Fig 2: Coagulocytes (a) at 1000 X and (b) enlarged view 3.1.31. Coagulocytes with granules As showed in Fig (5) larvae treated with potassium sulphate presented the highly increase peaks gained during the feeding period at 5th day (59±8%) was higher than those of control (42±4%). Larvae treated with sodium sulphate showed only one significant increase over than control were (45±5%) comparing to (28±9 %) of untreated ones during 1st day. Larvae treated with a biosalt mixture showed a positive increase in DHC at first and the last five days of last larval instar, the highly peak was (44±1%) higher than those (30±5%) of untreated ones during 8th days. ADHC of Coagulocytes with granules in larvae treated with potassium sulphate showed an increases during the feeding period, and reached the maximum (104.5 x103cell/larva) at 5th day in comparing to (97.5 x103cell/larva) of control. Larvae treated with sodium sulphate presented the higher increase in ADHC during the 2nd, 5th and 8th days of feeding period, the
maximum peak (153.8 x103cell/larva) was gained at 8th day, positively higher than those (64.4 x103cell/larva) of untreated. Larvae treated with a bio salt mixture showed two peaks of ADHC. The first peak observed during the feeding period was (116.8 x103cell/larva) gained at 8th day in comparing to (64.4 x103cell/larva) of untreated ones. The higher second peak was obtained at 10thday (163x103cell/larva) comparing to (77.7 103cell/larva) of control as in (Fig 6).
3.1.3.2. Coagulocytes without granules Data in Fig (7) indicated that treated and untreated larvae exhibited a fluctuation trend in DHC during the last larval instar of Chinese silkworm. Larvae treated with potassium sulphate showed a significant reduction in DHC during the first 5 days of feeding period. While, from the 8th day a significant increase of DHC was noted until reach the maximum (42±3%) at 11th day which was positively higher than those (26±4%) of untreated. Larvae treated with sodium sulphate showed the same trend during the feeding period. The only maximum increase of DHC was (32±3%) at 9th day in comparison to (24±4%) of control. larvae treated with a bio salt mixture presented the lowest trend of DHC in comparison to control and other treatments.
~ 274 ~
Journal of Entomology and Zoology Studies
The ADHC of coagulocyte without granules in larvae treated with potassium sulphate positively increased at 8th, 10th and 11th days and the maximum peak was (137.7 x103cell/larva) more than of untreated one (66.6 x103cell/larva) at 10th day. Larvae treated with sodium sulphate showed a gradual increase during the feeding period until reached the first peak (91.3 x103cell/larva) at 5th day comparing to (42.2 x103cell/larva) of untreated. While, during the 9th day the maximum peak of ADHC (149.1 x103 cell/larva) was observed higher than (80.2 x103 cell/larva) of control and other treatments. As in Fig (8) Larvae treated with a bio salt mixture showed the lowest trend of ADHC in comparison to treated and untreated larvae. 3.1.3.3. Total Coagulocytes From data shown in Fig (9) no significant change was observed of total coagulocytes DHC in larvae treated with potassium sulphate. Larvae treated with sodium sulphate presented the same trend. While, larvae treated with a bio salt mixture showed a decrease from the second day to the7 th day.
As shown in Fig (10) potassium sulphate treated larvae showed a slight changes in ADHC of total coagulocytes, the maximum peak was observed at 10th day (230.9 x103 cell/larva) in comparison to (144.3 x103cell/larva) of control. Larvae treated with sodium sulphate presented three significant increases in ADHC during early feeding period of last larval instar. The maximum peak gained at 9th day (260.9 x103cell/larva) comparing to (160.5 103cell/larva) of control. Larvae treated with a bio salt mixture showed the maximum high peak (242.3 x103cell/larva) which gained at 10th day in comparison to (144.3 x103cell/larva) of control. In Ph. Ricini 5th instar larvae treated with IGR's, IKI-7899 at 0.5 ppm, DHC of total coagulocytes showed the maximum peak during the 12th day higher than control, this increase seems to be true as the BV decrease at this time, while ADHC showed its maximum peak at the 9th day, the ADHC of treated larvae was higher than untreated by about 29.1% [32 33]. The DHC trend of total coagulocytes showed in the present study similar to the trend of coagulocytes with granules. Such results were also obtained in H. armigera [26]. However, when silkworm larvae fed on leaves treated with a biosalt minerals, the DHC of total coagulocytes and coagulocytes with granules modified in comparison to untreated during the last larval

~ 275 ~
Journal of Entomology and Zoology Studies
instar. From these results it could attribute this modification to the effect of minerals on the blood volume which directly affect the ADHC of coagulocytes. Present results are similar to the finding in Eri- silkworm [33]; and in mulberry silkworm [22]. 3.1.4. Spherule cells Spherule cells are oval and contain many characteristic spherules in their cytoplasm which stained pale blue in light microscope, the nucleus is usually small and contains a nucleolus, the cytoplasm contains many small spherules, which appear dark in phase-contrast microscopy (Fig:3). The size and number of the spherules seem to vary. The cells appear lobulated due to the marginal bulges possibly caused by accumulation of spherules.
Fig 3: Spherule cells and Oenocytoids (a) at 1000 X and (b) enlarged view.
Data shown in Fig (11) exhibited that DHC of spherule cells showed a non significant fluctuation in both treated and untreated larvae during the last larval instar. As in Fig(12) Larvae treated with potassium sulphate positively increased the ADHC of Spherules, maximum peak (8.1x103 cell/larva) observed during the 10th days comparing to (2.6 x103 cell/larva) of control. Larvae treated with sodium sulphate showed two increase peak in ADHC higher than control at 5th and 7th days were (6.1 and 5.4 x103 cell/larva) in comparison to (2.8 and 3.2 x103cell/larva) of untreated larvae, respectively. Larvae treated with a bio salt mixture showed its high peak (3.6 x103cell /larva) of ADHC at 4th day which was higher than control (2.2 x103 cell/larva).
The larval blood cells of 301 silkworm strains [31], in 26 strains, spherule cells were not present, and the author concluded that those strains produced less silk than those strains containing spherule cells. It was noted a high mitotic index and a rapid turnover of spherule cells, possibly as a mechanism of releasing products of their metabolism into the haemolymph [35]. In addition, spherule cells were noted to play a role in the secretion of some haemolymph protein [36]. Moreover, spherule cells attributed to silk production [31, 37]. The maximum value of ADHC of spherule cells in treated larvae with minerals corresponds to the maximum value of the blood volume. In present study the maximum peak of ADHC of spherule cells was observed at the 10th day. Also, it was reported that spherule cells played a significant role in silk production [33]. 3.1.5. Oenocytoides In the present study, oenocytoides have been to be the largest cellular types observed, being round or oval with small eccentric nucleus, homogenous cytoplasmic and plasmic membrane without projection. Oenocytoides characterized sometimes by one or two cytoplasmic pale staining inclusions. (Fig: 3 and Fig: 13). Oenocytoides were not found in division.

~ 276 ~
Journal of Entomology and Zoology Studies
Treated larvae with potassium sulphate showed insignificant changes in DHC of oenocytoides. Also, no significant observations were noted in larvae treated with sodium sulphate. Larvae treated with a bio salt mixture showed the same trend as in Fig (13). ADHC of oenocytoides treated larvae with potassium sulphate were increased during the 1st, 7th days of feeding period, the maximum increase was (6.3 x103cell/larva) at 7th day comparing to (3.2 x103cell/larva) of control which present about 97% increase. A reduction was appeared at day nine. While, during the10th day an increase of ADHC (8.1x103cell/larva) comparing to (2.6. x103cell/larva) of control. Larvae treated with sodium sulphate showed the highest increase value in ADHC were observed at 9th, 10th and 11th days and the maximum was (9.3 x103cell/larva) gained at 9th day comparing to (6.7 x103cell/larva) of untreated. Larvae treated with a bio salt mixture showed an increase in ADHC of oenocytoides was observed at 10th day (8.8 x103cell/larva) in comparing to (2.6x103cell/larva) of untreated ones Fig (14). In Galleria mellonela proportion of oenocytoides increased as the larvae increased in weight. Oenocytoides usually decreased after pupation in B. mori [31]. The overall Figure of the haemocytes of larvae treated with bio salt were increased in number with both, potassium sulphate, sodium sulphate and their mixture. It is well established that changes in DHC during insect development are common [38-26] and appear to be under endocrine control [39, 40, 26-37]. Changes in the number of circulating haemocytes might be associated with changes in blood volume [41]. From the current results it was observed that DHC and ADHC of prohaemocytes, plasmatocytes and spherule cells was greater in treated larvae than untreated one during the spinning period, while coagulocytes and oenocytoides were less than untreated. It has been investigated the role of spherule cells in secretion of some haemolymph protein [36] and in silk production [31-37]. According to Yamashita and Iwabuchi [42] they showed that over 60% of the non-dividing prohemocytes differentiated to plasmatocytes or granulocytes. Some of the granulocytes subsequently transformed to spherulocytes, 59.2% of the dividing prohemocytes, the daughter cells differentiated into other types of haemocytes such as plasmatocytes, granulocytes and spherulocytes, and the remainder divided into new prohaemocytes. In conclusion the use of minerals as food
supplements for silkworm not only lead to an improvement of the final body weight of the larvae, pupae, silk gland, cocoon shell and cocoon cortex [15] but also enhance the blood volume and total haemocyte counts specially at low concentration [22].The present work showed also the effect of minerals as food supplements on DHC and ADHC of the different types of haemocytes in last larval instar of B. mori. 4. References 1. Etebari K, Fazilati M. Effect of feeding on mulberry’s
supplementary leaves with N, P and K in some biological and biochemical characteristics of silkworm. J Sci Technology Agriculture and Natural Resources 2003; 7:233-244.
2. Vlaic B, Mǎrghitaş LAl, Dezmirean D, Benţea M. Research concerning the influence of biostimulating apicultural products administered in silkworm feeding on body weight gain during V-th age, Buletin UASVM-Animal Science and Biotechnologie, 60/2004, ISSN 2004; 1454-2382: 274-277.
3. Ito T. Silkworm Nutrition; in the Silkworm an Important Laboratory Tool. Tazima, Y (ed). Kodansha Ltd., Tokyo, Japan, 1978, 121-157.
4. Dasmahapatra AK, Chakrabarti MK, Medda AK. Effect of potassium iodide, cobalt chloride, calcium chloride and potassium nitrate on protein, RNA and DNA content of silkgland of silkworm nistari race. Sericologia 1989; 29:355-359.
5. Quader MA, Sarker A, Rab MA, Barman AC. Effects of potassium iodide and iodised salt on the larval growth and cocoon characters of the silkworm, Bombyx mori L. Race BSR-83. Sericologia 1993; 33:595-598.
6. Sarker A, Haque M, Rab M, Absar N. Effects of feeding mulberry (Morus sp.) supplemented with different nutrients to silkworm. Bombyx mori L Curr Sci 1995; 69: 185-188.
7. Zaman K, Ashfaq M, Akram şi W. Effect of feeding Mg and N treated mulberry leaves on larval development of silkworm Bombyx mori L. and silk yield. Pak Entomol 1995; 18:78-79.
8. Nirwani RB, Kaliwal BB. Increase of silk production and quantitative changes of carbohydrate and protein in the fat body and haemolymph after feeding potassium sulphate to bivoltine Bombyx mori L. Sericologia 1996; 36:523-530.
9. Hugar II, Nirwani RB, Kaliwal BB. Effect of zinc chloride on the bio-chemical changes in the fat body and haemolymph of the bivoltine silkworm. Bombyx mori L. Sericologia 1996; 38:299-303.
10. Rajashekhargouda R, Gopalan M, Jayaraj S, Natarajan N. Influence of thyroxine sodium tablets on mulberry silkworm Bombyx mori L. Sericologia 1998; 38:651-655.
11. Etebari K, Ebadi R, Matindoost L. Effect of feeding mulberrys enriched leaves with ascorbic acid some biological, biochemical and economical characteristics of silkworm Bombyx mori L. J Indust Entomol 2004; 7:81-87.
12. Bhattacharya A, Kaliwal BB. Influence on mineral potassium permanganate on the economic parameters of the silkworm, Bombyx mori L. Proceeding of National Conference on Tropical Sericulture for Global Competitiveness, 2003, 91.
13. Islam R, Abdul A, Dipak P, Shaheen S, Nilufa B, Islam R. Effect of salt, nickel cloride supplimentation on the growth of silkworm Bombyx mori L. J Biological Science

~ 277 ~
Journal of Entomology and Zoology Studies
2004; 4:170-172. 14. Khan MA, Akram W, Ashfaq M, Khan HAA, Kim YK,
Lee JJ. Effects of optimum doses of nitrogen, phosphorus, potassium and calcium on silkworm. Bombyx mori L., growth and yield. Entomological Research 2010; 40:285-289.
15. Saad IAI, Essawy MM, Souad MM, El- Kho Dary AS, Tama-Nadia M, El-Dakhakhny. Effect of potassium, sodium and its mixture supplementation on different biological aspects of mulberry silkworm Bombyx mori(L.). Egy J Plant Pro Res 2013; 1(1):100-114.
16. Patton RL. Introductory insect physiology. W.B. Saunders Co. Philadelphia. 1983; 47:65.
17. Jones JC. Current concepts concerning insect haemocytes Amer. Zool 1962; (2):209-246.
18. Akai H, Sato S. Ultrastructure of the larval haemocytes of the silkworm, Bombyx mori L. (Lepidoptera: Bombycidae). J of Insect Morphol & Embryol 1973; 2(3):207-231.
19. Essawy M, Maleville A, Brehelin M. The hemocytes of the Heliothis armigera: Ultrastructure, functions and evolution in the course of larval development. J morphol 1985; 186:255-264.
20. Essawy MM. Comparative light and electron microscopic accounts of immunocytes and other haemocytes of Philosamia ricini (Boisd.), and the evolution of haemogram after changes of rearing temperature. Adv Agric Res 1999; 41:661-684.
21. Saad IAI. Biological and Physiological Studies on the Silkworm. Ph.D. Thesis. Fac Agric Tanta Univ, 2005, 1-121.
22. Essawy MM, Saad IAI. Effect of potassium, sodium and its mixture supplementation on I-Blood volume, total and absolute haemocyte counts in mulberry silkworm Bombyx mori (L.). Journal of entomology and zoology studies 2013; 1(6):92-96.
23. Gomaa AA. Effect of temperature on the silk production of the Eri-silkworm Attacus ricini in Egypt. Zeit Ang Entomol 1973; (74):271-274.
24. Essawy MM. Hemogram changes of last larval, prepupal and newly pupal stage of the silkworm Bombyx mori (L.) after treatment with cholesterol. J Agric Sci Mansoura Univ 1997; 21(4):1169 – 1185.
25. Shapiro M. Changes in the haemocyte population of the wax moth, Galleria mellonella, during wound healing. J Insect Physiol 1968; 14:1725-1733.
26. Essawy MM. Relations cytophysiologues entre la glande prothoracique et le tissue sanguin Durant le dernier stade larvaire. d'. Etat, U. S. T. L. Montpellier, France, 1985.
27. Arnold JW, Hinks CF. Haemopoiesis in Lepidoptera. I. The multiplication of circulating haemocytes. Can J Zool 1976; 54:1003-1012.
28. Essawy MM. Changes in the different haemocyte counts of the last larval instar of Spodoptera littoralis (Boisd.) during wound healing. Alex Sci Exch 1990; 11(4):151-176.
29. Shapiro M. Pathologic changes in the blood of the greater wax moth, Galleria mellonella (L.), during the course of starvation and nucleopolyhedrosis. Ph. D. Thesis,
California Univ., Berkeley, California, 1966. 30. Yeager JE. The blood picture of the southern armyworm,
Prodenia eridenia. J Agric Res 1945; 71:1-40. 31. Nittono Y. Studies on the blood cells in the silkworm,
Bombyx mori L. Bull Seric. Exp Stn (Tokyo) 1960; 16: 171-266.
32. Saad IA. Studies on certain beneficial insects. M.sc. Thesis, Fac Agric Tanta Univ, 1996.
33. Essawy MM, Tama-Nadia El-Dakhakhini, Saad IA. The effect of chlorfluazuron (IKI-7899) as chitin synthesis inhibitor on some biological and physiological parameters of the last larval instar of Philosamia ricini (Boisd.). J Agric Res Tanta Univ 1999; 25(2):269-291.
34. Sanjayan ΚΡ, Ravikumar Τ, Albert S. Changes in the haemocyte profile of Spilostethus hospes (Fab) (Heteroptera: Lygaeidae) in relation to eclosion, sex and mating. J Biosci 1996; 21(6):781-788.
35. Arnold JW, Hinks CF. Haemopoiesis in Lepidoptera. III. note on the spherule cells and granular haemocytes. Can J Zool 1983; (61):275-277.
36. Akai H, Sato S. Surface and internal ultrastructure of haemocytes of some insects. In: Insect haemocytes, Development, Forms, Functions, and Techniques. (Ed. Gupta. A.P.) Cambridge Univ. Press London, New York, 1997.
37. Essawy MM, Gadelhak G, El-Karaksy I, Idriss M. Does juvenile hormone regulate haemogram in Bombyx mori L.? Proc. 1st, Int. Conf. of Silk” ICSAI”, 1997, 101-117.
38. Arnold JW. The Haemocytes of insects. In: The physiology of Insecta. (Ed. Rockstein, M.) Vol. 5, 2 nd Ed. Academic Press, New York, 1974.
39. Judy KJ, Marks EP. Effect of ecdysteron in vitro on hindgut and haemocytes of Manduca sexta (Lepidoptera). J Gen Comp Physiol 1971; 17:351-359.
40. Takeda N. Brain hormone carrier haemocytes in the moth Monema flavescens. J Insect Physiol 1977; 23:1245-1254.
41. Shapiro M. Changes in haemocytes population in: insect haemocytes, development, forms, function and techniques. (Gupta. A.P. Ed) Cambridge University Press, London, New York, 1979.
42. Yamashita M, Iwabuchi K. Bombyx mori prohemocyte division and differentiation in individual microcultures. J Insect Physiol 2001; 47(4-5):325-331.


















