Efectos de -Sinucleína en neuronas del Sistema Nervioso ...
147
1 Universidad de Concepción Dirección de Postgrado Facultad de Ciencias Biológicas-Programa de Doctorado en Ciencias Biológicas, Área Biología Celular y Molecular Efectos de -Sinucleína en neuronas del Sistema Nervioso Central CARLA ROCÍO PACHECO MURILLO CONCEPCIÓN-CHILE 2013 Profesores Guía: Dr. Luis Aguayo Hernández Dr. Carlos Opazo Martínez Dpto. de Fisiología, Facultad de Ciencias Biológicas Universidad de Concepción
Transcript of Efectos de -Sinucleína en neuronas del Sistema Nervioso ...

1
Universidad de Concepción
Dirección de Postgrado Facultad de Ciencias Biológicas-Programa de Doctorado en Ciencias Biológicas,
Área Biología Celular y Molecular
Efectos de -Sinucleína en neuronas del
Sistema Nervioso Central
CARLA ROCÍO PACHECO MURILLO CONCEPCIÓN-CHILE
2013 Profesores Guía: Dr. Luis Aguayo Hernández
Dr. Carlos Opazo Martínez Dpto. de Fisiología, Facultad de Ciencias Biológicas
Universidad de Concepción

2
Esta tesis ha sido realizada en el Departamento de Fisiología de la Facultad de Ciencias Biológicas, Universidad de Concepción. Profesores integrantes Comisión Evaluadora: _______________________
Dr. Luis Aguayo Hernández Profesor Guía de Tesis Facultad de Ciencias Biológicas _______________________
Dr. Carlos Opazo Martínez Profesor Guía de Tesis Externo
Facultad de Ciencias Biológicas _______________________ Dr. Francisco Nualart Facultad de Ciencias Biológicas _______________________ Dr. Juan Pablo Henríquez
Facultad de Ciencias Biológicas _______________________ Dr. Pablo Caviedes
Profesor Evaluador Externo
Universidad de Chile _______________________ Dr. José Guzmán G.
Director Programa Doctorado en Ciencias Biológicas

3
UNIVERSIDAD DE CONCEPCION
ESCUELA DE GRADUADOS
Efectos de -Sinucleína en neuronas del
Sistema Nervioso Central
CARLA ROCÍO PACHECO MURILLO
Tesis para optar al grado de Doctor en Ciencias Biológicas
Mención Biología Celular y Molecular
Tutores: Dr. Luis G. Aguayo H.
Dr. Carlos Opazo M.
Laboratorio de Neurofisiología, Departamento de Fisiología
Facultad de Ciencias Biológicas, Universidad de Concepción
Concepción – Chile
2013

4
ÍNDICE GENERAL
ÍNDICE GENERAL .................................................................................................. 4
ÍNDICE DE FIGURAS Y TABLAS ........................................................................... 9
LISTA DE ABREVIACIONES ................................................................................ 13
RESUMEN ............................................................................................................ 16
ABSTRACT ........................................................................................................... 18
1.- INTRODUCCION GENERAL ........................................................................... 22
Características patológicas de la enfermedad de Parkinson ................................. 22
Localización de -sinucleína ................................................................................. 26
Características fisiológicas de -sinucleína .......................................................... 30
Características patológicas de -sinucleína .......................................................... 30
Agregados de -sinucleína ................................................................................... 32
Rol del Ca2+ en la enfermedad de Parkinson ........................................................ 36
Formación de estructuras tipo poro/perforado de -sinucleína ............................. 37
HIPÓTESIS ........................................................................................................... 43
OBJETIVO GENERAL .......................................................................................... 44
OBJETIVOS ESPECÍFICOS ................................................................................. 44
Objetivo Específico I. Caracterizar bioquímicamente agregados de -sinucleína y
determinar si forman perforados en membranas neuronales. ............................... 44

5
Objetivo Específico II. Caracterizar efectos funcionales de -sinucleína en
neuronas hipocampales. ....................................................................................... 44
2.- MATERIALES Y MÉTODOS ............................................................................ 45
Cultivo de neuronas hipocampales. ...................................................................... 45
Cultivo de células RCSN-3 .................................................................................... 45
Cultivo de células SHSY-5Y .................................................................................. 46
Agregación de -sinucleína ................................................................................... 46
Obtención de rebanadas de neuronas de sustancia nigra. ................................... 47
Disociación de rebanadas de neuronas de sustancia nigra. ................................. 47
Electrofisiología. .................................................................................................... 49
Western blot. ......................................................................................................... 50
Tinción de plata. .................................................................................................... 50
Tinción con rojo congo .......................................................................................... 50
Viabilidad celular ................................................................................................... 51
Microscopía y análisis de fluorescencia para FM1-43 y Fluo4-AM. ....................... 52
Inmunocitoquímica ................................................................................................ 52
Marcaje de vesículas sinápticas con FM1-43. ....................................................... 54
Análisis de datos.. ................................................................................................. 54
3.- RESULTADOS ................................................................................................. 55

6
Objetivo Específico I. Caracterizar los agregados de -sinucleína y determinar si
forman perforados en membranas neuronales...................................................... 55
Objetivo 1.1. Caracterizar bioquímicamente y estructuralmente agregados de -
sinucleína .............................................................................................................. 56
Resultados ............................................................................................................ 57
Oligómeros de -sinucleína son principalmente especies solubles, de bajo peso
molecular y sensibles a proteínasa K .................................................................... 57
Objetivo 1.2. Determinar si agregados de -sinucleína modifican la conductancia
de membrana en neuronas hipocampales ............................................................ 64
Resultados ............................................................................................................ 65
Oligómeros de -sinucleína incrementan la conductancia de membrana en
neuronas hipocampales ........................................................................................ 65
Oligómeros de -sinucleína no forman “estructuras tipo poro” en la línea celular
RCSN-3 ................................................................................................................. 70
Objetivo 1.3. Caracterizar la asociación de -sinucleína en neuronas y células
RCSN-3 ................................................................................................................. 76
Resultados ............................................................................................................ 77
La asociación de oligómeros de -sinucleína a la membrana plasmática es célula-
dependiente........................................................................................................... 77
La formación de grandes agregados de -sinucleína en células RCSN-3 es
dependiente de la temperatura .............................................................................. 93

7
Oligómeros de -sinucleína inducen la formación de agresomas en células RCSN-
3 ............................................................................................................................ 99
Objetivo Específico II. Caracterizar efectos funcionales de -sinucleína en
neuronas hipocampales ...................................................................................... 103
Objetivo 2.1 Determinar si agregados de -sinucleína alteran los niveles de Ca2+
intracelular y la actividad sináptica neuronal ....................................................... 104
Resultados .......................................................................................................... 104
Oligómeros de -sinucleína aumentan los niveles de Ca2+ intracelular en neuronas
hipocampales y células RCSN-3 ......................................................................... 104
Oligómeros de -sinucleína aumentan la transmisión sináptica de neuronas
hipocampales ...................................................................................................... 108
Objetivo 2.2. Determinar la toxicidad mediada por agregados de -sinucleína .. 119
Resultados .......................................................................................................... 119
En condiciones crónicas de 24 horas oligómeros de -sinucleína no causan
toxicidad en neuronas hipocampales .................................................................. 119
4.- DISCUSIÓN Y CONCLUSIONES .................................................................. 123
La formación de oligómeros de -sinucleína ocurre por una vía alternativa a la
fibrilización convencional ..................................................................................... 123
Oligómeros de -sinucleína se asocian a neuronas y causan daños a nivel de la
membrana plasmática ......................................................................................... 125

8
La asociación de -sinucleína a la membrana plasmática es un proceso
fundamental en la formación de “estructuras tipo poro” ...................................... 128
-Sinucleína actúa como una proteína priónica en células RCSN-3 ................... 130
Células RCSN-3 forman agresomas de -sinucleína .......................................... 130
Oligómeros de -sinucleína causan sinaptotoxicidad en neuronas hipocampales
en condiciones crónicas ...................................................................................... 131
Modelo de acción de oligómeros de -sinucleína en neuronas hipocampales ... 133
Formación e impacto de agregados intracelulares de oligómeros de -sinucleína
............................................................................................................................ 135
5.- AGRADECIMIENTOS .................................................................................... 137
6.- BIBLIOGRAFÍA .............................................................................................. 139

9
ÍNDICE DE FIGURAS Y TABLAS
Figura 1. Progresión de la enfermedad de Parkinson. ..................................... 25
Figura 2. Estructura primaria y secundaria de -sinucleína ........................... 29
Figura 3. Modelo hipótetico de la secreción y de los blancos sinápticos de
agregados de -sinucleína. ............................................................................. 35
Figura 4. Modelo de perforación de -sinucleína ............................................. 42
Figura 5. Esquema de la configuración de Patch Clamp Perforado ............... 48
Figura 6. Agregación de -sinucleína. ............................................................... 58
Figura 7. Oligómeros de -sinucleína son especies principalmente solubles
........................................................................................................................... 60
Figura 8. Tinción de agregados de -sinucleína con tioflavina T y tratamiento
con proteinasa K .............................................................................................. 62
Figura 9. Tinción de agregados de -sinucleína con rojo congo. ................... 63
Figura 10. Efecto de los oligómeros de -sinucleína en la corriente
capacitativa de membrana en neuronas hipocampales ................................ 66
Figura 11. Difusión de glucosa fluorescente a través de estructuras ............ 68
Figura 12. Efecto de fosfolipasa C en la capacidad perforante de oligómeros
de -sinucleína en neuronas hipocampales .................................................. 69
Figura 13. Efecto de los oligómeros de -sinucleína en la corriente
capacitativa de membrana en células RCSN-3 .............................................. 71
Figura 14. Efecto de rotenona sobre las células RCSN-3. ............................... 73

10
Figura 15. Viabilidad de neuronas hipocampales luego del tratamiento con
rotenona. ........................................................................................................... 74
Figura 16. Efecto de los oligómeros de -sinucleína en la corriente
capacitativa de membrana en neuronas disociadas dopaminérgicas de SN
a P20. ................................................................................................................. 75
Figura 17. Asociación de oligómeros de -sinucleína a neuronas
hipocampales. .................................................................................................. 78
Figura 18. Distribución de oligómeros de -sinucleína en células gliales. .... 79
Figura 19. Asociación de oligómeros de -sinucleína a células RCSN-3. ..... 81
Figura 20. Cuantificación de la asociación de oligómeros de -sinucleína en
neuronas hipocampales y células RCSN-3. ................................................... 82
Figura 21. Neuronas hipocampales presentan grandes agregados de -
sinucleína asociados a núcleos apoptóticos. ................................................ 83
Figura 22. Proyección en 3D de oligómeros de -sinucleína en neuronas
hipocampales. .................................................................................................. 84
Figura 23. Proyección en 3D de oligómeros de -sinucleína en células RCSN-
3. ........................................................................................................................ 85
Figura 24. Asociación de oligómeros de -sinucleína a neuronas
hipocampales y células RCSN-3. .................................................................... 86
Figura 25. Efecto de tripsina y -MCD sobre la asociación de oligómeros de
-sinucleína en neuronas hipocampales. ...................................................... 88
Figura 26. Cuantificación de la asociación de oligómeros de -sinucleína en
neuronas hipocampales luego del tratamiento con tripsina y -MCD. ........ 89

11
Figura 27. Efecto de tripsina y -MCD sobre la asociación de oligómeros de
-sinucleína en células RCSN-3. ..................................................................... 91
Figura 28. Cuantificación de la asociación de oligómeros de -sinucleína en
células RCSN-3 luego del tratamiento con tripsina y β-MCD. ...................... 92
Figura 29. Incubación por 1 hora a 4°C inhibe la aparición de agregados de
gran tamaño de -sinucleína en células RCSN-3. ......................................... 93
Figura 30. Cuantificación de la asociación de oligómeros de -sinucleína en
células RCSN-3 a 4°Cy 37ºC. ........................................................................... 94
Figura 31. Proyección en 3D de oligómeros de -sinucleína en células RCSN-
3. ........................................................................................................................ 96
Figura 32. Oligómeros de -sinucleína exógena inducen la agregación de -
sinucleína endógena en células RCSN-3. ...................................................... 97
Figura 33. -Sinucleína exógena y -sinucleína endógena forman agregados
mixtos en células RCSN-3. .............................................................................. 98
Figura 34. Efecto de colchicina y EHNA sobre la formación de agregados de
-sinucleína en células RCSN-3. ................................................................... 100
Figura 35. Determinación de la ubiquitinación de agregados de -sinucleína
en células RCSN-3. A .................................................................................... 102
Figura 36. Efecto de oligómeros de -sinucleína en los niveles intracelulares
de Ca2+ en neuronas hipocampales y células RCSN-3. .............................. 106
Figura 37. Efecto de oligómeros de -sinucleína en los niveles intracelulares
de Ca2+ en neuronas hipocampales en ausencia de Ca2+ externo. ............ 107

12
Figura 38. Asociación tiempo dependiente de los oligómeros de -sinucleína
a cultivo primario neuronal. .......................................................................... 108
Figura 39. Oligómeros de -sinucleína incrementan las transitorias de Ca2+
en neuronas hipocampales. .......................................................................... 110
Figura 40. Oligómeros de -sinucleína incrementan las corrientes sinápticas
en miniatura en neuronas hipocampales. .................................................... 111
Figura 41. Oligómeros de -sinucleína alteran el reciclamiento vesicular de
neuronas hipocampales. ............................................................................... 114
Figura 42. Oligómeros de -sinucleína incrementan la inmunoreactividad de
SNAP25 en neuronas hipocampales. ........................................................... 116
Figura 43. Oligómeros de -sinucleína disminuyen la inmunoreactividad de
SV2 en neuronas hipocampales después de aplicación crónica. .............. 118
Figura 44. Efecto crónico de oligómeros de -sinucleína recombinante
humana sobre neuronas hipocampales. ...................................................... 120
Figura 45. Efecto crónico de medio condicionado con -sinucleína sobre
cultivo neuronal. ............................................................................................. 122
Figura 46. Modelo propuesto para acción de los oligómeros de -sin
extracelular en neuronas hipocampales y células RCSN-3. ....................... 136

13
LISTA DE ABREVIACIONES
-Sin, -sinucleína
A, péptido -Amiloide
AFM, microscopia de fuerza atómica
-MCD, -metilciclodextrina
CL, cuerpo de Lewy
Bapta-AM, ácido 1,2-bis-2-aminofenoxietano-N,N,N´,N´- tetra acético-acetoximetil
éster
Ca+2, ión calcio
Co+2, ión cobalto
DIV, días in vitro
DMEM/F12, medio de Dubelcco modificado por Eagle/Mezcla de nutrientes F12
DPBS, buffer Salino Fosfato de Dubelcco
EP, enfermedad de Parkinson
EA, enfermedad de Alzheimer
FITC, fluoresceína isotiocianato
Fluo-4AM, 2-[2-(2-5-[bis(carboximetil)amino]-2-metilfenoxietoxi)-4-(2,7-difluoro-
6-hidroxi-3-oxo-3H-xanteno-9-yl)fenil](carboximetil)amino acético
FM1-43, N-(-3-trietilamoniopropil)-4-(4-(dibutilamino) estiril) piridinio dibromuro
FLC, fosfolipasa C
GPI, glicosilfosfatidilinositol
hrs, horas

14
HBSS, Hank's Balanced Salt Solution
HEPES, ácido 4-(2-hidroxietil)-1-piperazinaetanosulfónico
HRP, peroxidasa de rábano
Hz, hertz (1/segundo)
K+, ión potasio
kDa, kilodalton (1000g/mol)
MAP2, proteína asociada a microtúbulos 2
MEM, Medio esencial mínimo
min, min
MET, microscopia electrónica de transmisión
Mg, miligramo
mM, milimolar
ms, milisegundo
NIH, Instituto Nacional de Salud de los Estados Unidos
nM, nanomolar
pA, picoamperes
PBS, buffer salino fosfato
PK, proteinasa K
pM, picomolar
Rot, rotenona
rpm, revoluciones por minuto
SN, sustancia nigra
ThT, tioflavina T
TTX, tetrodotoxina

15
U.F.R, unidades de fluorescencia relativa
g, microgramo
m, micrómetro
M, micromolar
L, microlitro
V, voltios
Ω, ohm
RESUMEN

16
RESUMEN
La enfermedad de Parkinson es un desorden neurodegenerativo progresivo
que se caracteriza por la presencia de agregados proteicos denominados cuerpos
de Lewy, que están constituidos principalmente por la proteína -sinucleína, cuyo
rol en esta enfermedad aún no se ha determinado. Sin embargo, estudios
genéticos en humanos y biología molecular en ratones, sugieren que -sinucleína
estaría involucrada en la aparición temprana de la enfermedad de Parkinson. Al
respecto, se ha postulado que la acumulación de -sinucleína en el espacio
extracelular podría alterar membranas neuronales a través de la formación de
“estructuras tipo poro”, lo que llevaría a alteraciones en la homeostasis iónica de la
célula. No obstante, esto nunca ha sido demostrado en membranas neuronales.
En la presente tesis se realizaron una serie de experimentos
electrofisiológicos, bioquímicos y de biología celular que indican que oligómeros
de -sinucleína (500 nM) rápidamente se asocian de manera puntiforme a
neuronas hipocampales, dado principalmente por el componente proteico de la
membrana, lo que resulta en un incremento en la conductancia (5 veces sobre el
control) con el subsecuente influjo de Ca2+ y de un análogo fluorescente de
glucosa. Este incremento en el Ca2+ intracelular (1.7 veces sobre el control) causa
un incremento significativo en diferentes parámetros de la transmisión sináptica
tales como la frecuencia de las transitorias de Ca2+, la frecuencia de las corrientes
sinápticas en miniatura, el decaimiento de la fluorescencia asociada a FM1-43 y el
aumento en el número de puntas para SNAP25, lo que ocurre sin cambios en la

17
viabilidad neuronal. Sin embargo, se observaron algunos cambios morfológicos
apoptóticos asociados a neuronas hipocampales circundadas por “macro”
agregados de -sinucleína.
Al realizar incubaciones prolongadas con oligómeros de -sinucleína (5 M
por 24 horas), todavía en ausencia de cambios en la viabilidad celular, se observó
una depleción vesicular representada por la disminución en el número de puntas
para SV2, sugiriendo que oligómeros de -sinucleína inducirían un falla sináptica
en forma concentración-tiempo dependiente. Esto concuerda con los resultados
obtenidos con otras proteínas amiloides formadoras de poros, como el A, el cual
en condiciones crónicas de tratamiento induce una disminución en la transmisión
sináptica de neuronas hipocampales.
Es importante destacar que el efecto perforante de oligómeros de -
sinucleína es célula dependiente, ya que la línea celular dopaminérgica RCSN-3,
fue resistente a la acción permeabilizante de -sinucleína, incluso en presencia de
un conocido inductor de Parkinsonismo como lo es el pesticida rotenona, lo que se
correlacionó con la formación de cuerpos de inclusión constituidos por la proteína
exógena con características tipo agresoma. Nuestros resultados indican además
que éstos agregados intracelulares indujeron la formación de agregados de -
sinucleína endógena. Sin embargo, a pesar de la resistencia presentada en las
células RCSN-3, -sinucleína indujo un incremento en los niveles de Ca2+
intracelular, efecto bloqueado en presencia de Co2+, bloqueador inespecífico de
canales de Ca2+.

18
En conclusión, las evidencias presentadas permiten proponer que las
acciones extracelulares de oligómeros de -sinucleína podrían estar mediadas por
la formación de perforados de -sinucleína en la membrana plasmática como
también por la formación de cuerpos de inclusión de esta proteína, lo cual
explicaría las etapas iniciales de la sinaptotoxicidad que proponemos conduce a la
neurodegeneración que se observa en las etapas tardías de las enfermedades
neurodegenerativas asociadas con la acumulación de -sinucleína extracelular.
Esta información podría ser de utilidad para implementar nuevas estrategias
terapéuticas para aliviar los problemas motores y cognitivos que presentan los
pacientes con la enfermedad de Parkinson.
ABSTRACT

19
ABSTRACT
Parkinson's disease is a progressive neurodegenerative disorder
characterized by the presence of protein aggregates called Lewy bodies, which are
mainly composed of -synuclein protein, whose role in this disease has not yet
been determined. However, genetic studies in humans and molecular biology
studies in mice strongly indicate that -synuclein is involved in the early onset of
Parkinson's disease. In this regard, it has been postulated that the accumulation of
-synuclein in the extracellular space may alter neuronal membranes through the
formation of "pore-like structures", which would lead to alterations in the ionic
homeostasis of the cell. However, this has never been demonstrated in neuronal
membranes.
This thesis describes a series of electrophysiological, biochemical and cell
biology experiments that indicates that -synuclein oligomers (500 nM) rapidly
associates in a punctate fashion to hippocampal membranes, mainly by the protein
component of the plasma membrane, resulting in an increase in membrane
conductance (5 fold over control) and leading to the influx of both Ca2+ and a
fluorescent glucose analogue. This increase in intracellular Ca2+ (1.7 fold over
control) caused a large increase on several synaptic transmission parameters such
as frequency of the Ca2+ transients, frequency of miniature synaptic currents, the
fluorescence destaining associated to FM1-43 and an increase in the number of
SNAP25 puncta, which occurs without changes in neuronal viability. However,

20
there were some apoptotic morphological changes associated with hippocampal
neurons surrounded by "macro" -synuclein aggregates.
When performing prolonged incubations with -synuclein oligomers (5 M
for 24 hours), even in the absence of changes in cell viability, was observed
vesicular depletion represented by the decrease in the number of SV2 puncta,
suggesting that -synuclein oligomers would induce synaptic failure in a
concentration-time dependent manner. This is consistent with results obtained with
other amyloid pore-forming proteins, such as A, which in chronic conditions
treatment causes synaptic transmission failure of hippocampal neurons.
Importantly, the perforating effect of -synuclein oligomers was cell-type
dependent, since a dopaminergic cell line RCSN-3, was highly resistant to the
permeabilization action of -synuclein, even in the presence of a known inducer of
Parkinsonism such as pesticide rotenone, which correlated with the formation of
inclusion bodies formed by the exogenous protein with aggresome-like
characteristics. Our results also indicate that these aggregates induce the
formation of intracellular aggregates of endogenous -synuclein. However,
although RCSN-3 cells presented resistance, -synuclein induced an increase in
intracellular Ca2+ levels, an effect blocked in the presence of Co2+, a nonspecific
Ca2+ channel blocker.
In conclusion, the evidence presented in this thesis allow us to propose that
the actions of extracellular -synuclein oligomers could be mediated by the
formation of -synuclein perforation in the plasma membrane as well as by the
formation of inclusion bodies of this protein, which would explain the early stages

21
of synaptotoxicity that we propose leads to neurodegeneration observed in late
stages of neurodegenerative diseases associated with accumulation of
extracellular -synuclein. This information could be useful to implement novel
therapeutic strategies to alleviate the motor and cognitive problems observed in
patients that present Parkinson’s disease.
1.- INTRODUCCION GENERAL

22
1.- INTRODUCCION GENERAL
Características patológicas de la enfermedad de Parkinson
La enfermedad de Parkinson (EP) es el desorden del control motor
humano más común, junto con ser el segundo trastorno neurodegenerativo más
frecuente después de la enfermedad de Alzheimer (EA) (Goedert 2001). En
nuestro país, al año 2007, se han diagnosticado 90 pacientes/100 mil habitantes
con EP (MINSAL) y en el mundo se estiman entre 7 a 10 millones de personas
que sufren de EP, afectando principalmente a hombres (OMS). Clínicamente se
caracteriza por rigidez muscular, bradiquinesia (lentitud en los movimientos) y
temblor en estado de reposo (Jankovic 2008). Patológicamente, la EP se
caracteriza por la pérdida de neuronas dopaminérgicas presentes en la sustancia
nigra pars compacta, núcleo perteneciente a los ganglios basales ubicado en la
base del mesencéfalo, cuya función es la correcta coordinación de los
movimientos motores voluntarios mediante la regulación de la actividad de las
neuronas motoras superiores de la corteza premotora y motora primaria, a través
de su proyección hacia el núcleo caudado y putamen, favoreciendo e inhibiendo
las vías motoras directa e indirecta, respectivamente (Willingham et al. 2001).
Por otro lado, patológicamente, también la EP se caracteriza por la presencia de
cuerpos de inclusión en las neuronas que sobreviven en esta región (Bethlem
and Den Hartog Jager 1960), denominados cuerpos de Lewy (CL). Los síntomas
y signos de la EP no se desarrollan hasta la pérdida de un 70-80% de las células
dopaminérgicas (Schapira 1999). Aunque la causa de la EP aún sigue siendo un

23
tema incierto, existe una gran cantidad de evidencia que indica que los
agregados de la proteína -sinucleína (-Sin) estarían participando en la
patogénesis de esta enfermedad. Primero, la presencia de los CL es la principal
característica histológica de la EP (Spillantini et al. 1997, 1998). Segundo,
análisis de asociación genética han permitido identificar 3 mutaciones de -Sin
asociada a las formas heredadas de la EP (Polymeropoulos et al. 1997; Kruger et
al. 1998; Zarranz et al. 2004) donde cada una de las variantes mutadas ha
demostrado acelerar la oligomerización o fibrilización de esta proteína. Tercero,
se ha determinado que la duplicación o triplicación del gen de -Sin resulta en un
incremento en la expresión de esta proteína, lo que se asocia a la EP de tipo
familiar (Singleton et al. 2003; Chartier-Harlin et al. 2004; Ibanez et al. 2004).
Esto sugiere que la acumulación de -Sin sería un factor crítico en la EP.
Disfunción cognitiva en la enfermedad de Parkinson
La EP es principalmente descrita como una enfermedad motora, pero a
medida que la enfermedad progresa distintas zonas asociadas a funciones
autonómicas, límbicas y somatomotoras se encuentran afectadas (Figura 1)
(Braak et al. 2004). Por otra parte, la demencia, un síntoma no motor, es
detectado en una gran proporción de pacientes con EP; sin embargo, el momento
y tasa de declive cognitivo es altamente variable (Aarsland et al. 2007). Resulta
interesante destacar que el 20-40% de los pacientes con EP poseen fallas
cognitivas una vez que la enfermedad es diagnosticada, y que el 80% de los
pacientes con EP eventualmente desarrollará demencia (Aarsland et al. 2007). Los
mecanismos de iniciación y de propagación de la EP asociada a la acumulación

24
de la proteína -Sin se desconocen hasta el momento. Una posible respuesta a
esta interrogante es que los agregados de -Sin puedan afectar distintas zonas
del cerebro, en una forma similar a lo que ocurre en enfermedades priónicas. En el
caso de estas últimas enfermedades, se postula que los agregados del prión
patogénico formados en una célula, podrían “infectar” a células vecinas
favoreciendo la agregación de proteínas priónicas normales (Aguzzi et al. 2006).
En el caso de -Sin ha sido señalado que células de tejido sano injertado en el
cerebro de personas afectadas por EP forman CL en su interior en forma tiempo-
dependiente (Li et al. 2008). Por lo tanto, parece posible la formación de CL en
neuronas sanas circundantes.

25
Figura 1. Progresión de la enfermedad de Parkinson. -Sin puede ensamblarse
en agregados amiloides formando cuerpos de Lewy (CL), los cuales se encuentran
localizados inicialmente en zonas específicas del cerebro (colores oscuros),
progresando a distintas áreas (colores claros) en una secuencia topográficamente
predecible (Modificado de Braak et al. 2004). La flecha a la derecha del esquema
señala algunas de las regiones cerebrales relacionadas con las fases pre-
sintomáticas y sintomáticas de la EP. Los colores oscuros y claros indican los
estadios tempranos y tardíos de la enfermedad, respectivamente.
Neocorteza
Primaria
Secondaria
Neocorteza
de
Asociación
Mesocorteza
Tálamo
Sustancia
Nigra
Amígdala
Núcleo
Dorsal
Núcleo
Motor X
Fase
Presintomática
Fase
Sintomática
Médula
espinal
Tronco
Encefálico
Tálamo
Corteza
Putamen
Estriado
Caudado
Núcleo Subtalámico
Substancia
nigra
Glóbulo
pálido

26
Localización de -sinucleína
La familia de la sinucleína se encuentra compuesta por 3 miembros: -, -
y -Sin, los que presentan una localización variable (Lavedan, 1998). Estudios de
inmunohistoquímica han señalado que - y -Sin se encuentran principalmente a
nivel de los terminales nerviosos en estrecha proximidad con vesículas sinápticas
del sistema nervioso central (Clayton and George, 1999). Por el contrario, -Sin
parece estar presente a lo largo de las células nerviosas en el sistema nervioso
periférico (Clayton and George, 1999). Sobre la localización celular de los
agregados de -Sin estos han sido descritos, principalmente, a nivel intracelular
asociado a los CL (Goedert 2001). Sin embargo, recientemente, se ha
determinado que estos agregados se ubicarían a nivel extracelular (Bininosti et
al. 2012; El-Agnaf et al. 2003; El-Agnaf et al. 2006; Paleologou et al. 2009;
Tokuda et al. 2010). Pacientes que padecen de la EP presentan niveles tres
veces más elevados de oligómeros de -Sin en el líquido cefalorraquídeo en
comparación con los pacientes control (El-Agnaf et al. 2003; El-Agnaf et al.
2006). Además se ha detectado que los niveles de -Sin aumentan al doble en
el plasma de pacientes con EP de tipo familiar que presentan triplicación del gen
de -Sin (Miller et al. 2004). Estos estudios sugieren que tanto la presencia como
los cambios en los niveles extracelulares de -Sin se asociarían con la EP.

27
Estructura de -sinucleína
-Sin presenta 4 isoformas compuesta por 98, 112, 126 y 140
aminoácidos, siendo ésta última la más abundante (Bisaglia et al. 2009). En
virtud de su conformación nativa no estructurada en estado soluble, -Sin forma
parte del conjunto de proteínas intrínsecamente no estructuradas (IUP), las
cuales no poseen una estructura terciaria estable lo que les permite cambiar de
conformación dependiendo de las condiciones del medio (Uversky et al. 2002).
Estudios de resonancia magnética nuclear (RMN) han determinado que en
presencia de micelas aniónicas, cada monómero de -Sin estaría compuesto por
una región N-terminal tipo -hélice interrumpido por un “loop”, además de
presentar un extremo C-terminal no estructurado y móvil (Ulmer et al. 2005). La
región N-terminal de -Sin se encuentra casi completamente compuesta por el
sitio consenso XXKTKEGVXXXX donde se localizan las tres mutaciones
asociadas al desarrollo temprano de la EP: A30P, E46K y A53T. Este dominio se
asemeja bastante al presente en apolipoproteínas caracterizado por presentar -
hélices anfipáticas en su estructura lo que les permite unirse a lípidos de
membrana. En su región central -Sin presenta un dominio altamente
amiloidogénico rico en aminoácidos hidrofóbicos (60-95) responsable del proceso
de agregación y formación de hojas -plegada. Este segmento fue identificado en
los pacientes con enfermedad de Alzheimer (EA) como el componente no
amiloidogénico de las placas seniles (NAC) (Masliah et al. 1996). Cabe
mencionar que la estructura de hojas antes mencionada y que promueve la
agregación de esta proteína, ha sido descrita también para otras proteínas

28
amiloides que sufren agregación y tienen un importante rol en la etiología de
diferentes enfermedades neurodegenerativas denominadas Enfermedades
conformacionales (Bossy-Wetzel et al. 2004). Con respecto a la región C-terminal
de -Sin, ésta se encuentra conformada principalmente por residuos ácidos los
que han mostrado ser responsables de regular la formación de fibras, impidiendo
la formación de agregados (Bisaglia et al. 2009) (Figura 2). Poco se conoce
acerca de modificaciones postraduccionales de -Sin en el cerebro. En células
transfectadas con -Sin, ésta se encuentra constitutivamente fosforilada en los
residuos serina 87 y 129, siendo predominante ésta última modificación (Okochi
et al. 2000).

29
Figura 2. Estructura primaria y secundaria de -sinucleína. A, El presente
esquema muestra los tres dominios principales de -Sin. La región N-terminal
(rosa) es un dominio anfipático en el cual se encuentran tres mutaciones
puntuales ligadas a formas autosómicas dominantes de la EP (barras blancas,
Bisaglia et al. 2009). La región central (verde) es un dominio altamente
hidrofóbico que fue originalmente encontrado en pacientes con EA y Demencia
con Cuerpos de Lewy (DCL), denominado como el precursor del componente no-
amiloidogénico de las placas seniles (NAC, Ueda et al. 1993). El dominio C-
terminal (azul) es de carácter acídico, el cual posee propiedades anti-
amiloidogénicas (Hoyer et al. 2004). B, Diagrama en cinta derivado de la
estructura secundaria del monómero de -Sin obtenido en presencia de micelas
aniónicas a través de RMN [PDB code 1XQ8] (Ulmer et al. 2005).
..................................................................................................................................
..................................................................................................................................
..................................................................................................................................
..................................................................................................................................
..................................................................................................................................
B)
=Proteína soluble
N C61 95 140
Región Central Región C-terminalRegión N-terminal
NAC
A)

30
Características fisiológicas de -sinucleína
Referente al rol fisiológico de -Sin no existe aún una opinión clara,
principalmente, porque ratones knockout para esta proteína no presentan un
cambio sustancial en su fenotipo, indicando de alguna manera que probablemente
su función sea redundante (Chandra et al. 2004). Además, ha sido determinado
que -Sin sería expresada luego del desarrollo sináptico, por lo tanto, esta
proteína tampoco jugaría un papel crítico en la formación de la sinapsis (Murphy et
al. 2000). Sin embargo, actualmente existe evidencia substancial que indica que -
Sin jugaría un rol clave en la regulación de la dinámica de liberación vesicular
(Murphy et al, 2000; Chandra et al, 2004, 2005; Burre et al, 2010). Se ha
demostrado que -Sin actuaría como una chaperona molecular auxiliar
complementando la acción ejercida por la proteína CSP (cysteine-string-protein
), cuya función consiste en el correcto ensamblaje del complejo proteico SNARE
(soluble N-ethylmaleimide-sensitive factor attachment protein receptor) a través de
la proteína SNAP25 que participa en la liberación de vesículas sinápticas
(Chandra et al. 2005).
Características patológicas de -sinucleína
-Sin es una proteína citosólica, por lo tanto, se ha supuesto que los
cambios patogénicos inducidos por esta proteína ocurrirían sólo a nivel
citoplasmático (Lee et al. 2006). Dentro de los mecanismos por los que -Sin
intracelular contribuiría a la patogénesis de la EP se encuentran: I) inhibición del
sistema ubiquitina-proteosoma (Emmanouilidou et al. 2008), II) aumento en los

31
niveles de estrés del retículo endoplasmático (Kim et al. 2008), III) disfunción
mitocondrial (Hsu et al. 2000) y IV) producción de especies reactivas del oxígeno
(Barnham et al. 2004). Todo esto llevaría, finalmente, a la disfunción y muerte
neuronal (Cookson et al. 2008). Sin embargo, poco es lo que se conoce hasta el
momento con respecto a los mecanismos de toxicidad que ejercería -Sin
extracelular. Se proponen principalmente tres hipótesis: I) -Sin extracelular
induciría la activación microglial causando neuroinflamación (Zhang et al. 2005), II)
-Sin extracelular sería endocitada para ejercer su acción tóxica en el citosol
(Desplats et al. 2009) y III) -Sin extracelular alteraría directamente la membrana
neuronal causando la formación de estructuras tipo poro/perforado (Volles et al.
2003; Feng et al. 2010; Schmidt et al. 2012). (Figura 3).
Estos estudios junto con la evidencia previa indican que -Sin tendría un
rol dual: un rol fisiológico involucrado en la regulación de la transmisión sináptica
y un rol patológico, asociado a procesos neurodegenerativos. Una pregunta clave
que surge de ello es: ¿Cómo estos dos roles están
secuencialmente/temporalmente/directamente conectados? Aún no existe una
respuesta concisa a esta pregunta, por lo que resulta de suma importancia
entender cuáles son las diferencias y similitudes entre los mecanismos
patogénicos y fisiológicos de -Sin. Parte de esta respuesta podría encontrarse
en la flexibilidad estructural que presenta -Sin que le permite la transición de
estados solubles a estados agregados. Otra pregunta importante de responder
es: ¿El proceso de agregación es una ganancia o pérdida de función de -Sin?
Se ha sugerido que en condiciones fisiológicas la forma molecular predominante

32
de -Sin sería la forma monomérica, que participaría en el reciclaje de las
vesículas sinápticas. Sin embargo, en condiciones patológicas, los oligómeros de
-Sin serían los predominantes y los causantes de la neurodegeneración. Por lo
tanto, y considerando que los ratones knockout de sinucleína, carecen de un
fenotipo Parkinsoniano, se ha sugerido que en condiciones patogénicas -Sin
adquiriría una función patológica, lo cual sería dependiente de concentraciones
locales elevadas, punto crítico para la formación de oligómeros (Wood et al.
1999).
Agregados de -sinucleína
En relación al proceso de agregación de -Sin, estudios in vitro indican la
presencia de un mecanismo dependiente de nucleación que se caracteriza por
una fase inicial lenta y una subsiguiente fase de crecimiento exponencial que
culmina en un estado estacionario (Wood et al. 1999). Este proceso depende
tanto de la identidad de -Sin (nativa o mutante) (Li et al. 2001), como también
de las condiciones de incubación, por ejemplo, pH, temperatura (Uversky et al.
2001), concentración de iones metálicos (Uversky et al. 2001) y otros agentes
como pesticidas (Uversky et al. 2001). Estructuralmente los agregados in vitro se
asemejan mucho a los agregados provenientes de cerebros de pacientes
afectados por la EP, los cuales exhiben una típica morfología fibrilar amiloidea
(Conway et al. 2000), sin embargo, la agregación in vitro permite generar
además de las fibras maduras, pequeños agregados denominados protofibras
(Goldberg et al. 2000). Se ha reportado también que -Sin nativa formaría
tetrámeros estables con una conformación secundaria rica en -hélices. El

33
tetrámero mostraría poca propensión a la agregación, lo que sugiere que debe
disociarse antes de formar oligómeros tóxicos de -Sin in vivo (Bartels et al.
2011). Referente a las mutaciones asociadas a la EP se ha determinado que
tanto A53T como A30P promueven la generación de especies oligoméricas
prefibrilares mientras que E46K reduce la formación de estos agregados
(Bisaglia et al. 2009). Dentro de las especies potencialmente tóxicas se
encuentran agregados de bajo peso molecular como dímeros, trímeros,
tetrámeros, pentámeros y hexámeros, los que poseen un peso molecular entre
los 20-100 kDa y que se caracterizarían en un principio por poseer una
conformación globular para luego dar lugar a estructuras anulares tipo poro con
un diámetro externo de 9-15 nm e interno de ~2-5 nm (Tsigelny et al. 2007). El
mecanismo de formación a través del cual los monómeros de -Sin se convierten
en oligómeros tóxicos y luego en fibras se encuentra actualmente bajo intensa
investigación (Tsigelny et al. 2007). Se sugiere que este proceso ocurriría a nivel
de la membrana plasmática, preferencialmente, en micro dominios ricos en
colesterol y esfingolípidos llamados balsas lipídicas (lipid rafts) (Bar-On et al.
2008) e involucraría interacciones entre lípidos y residuos hidrofóbicos de las -
hélices anfipáticas presentes en el extremo N-terminal de -Sin (Tsigelny et al.
2007).

34
Secreción de -sinucleína
¿Cuál es el mecanismo de secreción de -Sin?. Aunque el mecanismo
exacto no ha sido caracterizado aún, se ha determinado que una pequeña
porción de -Sin sería secretada al medio extracelular a través de exocitosis del
tipo no convencional, alternativo a la vía clásica de RE/Golgi (Lee et al. 2005). El
mecanismo de exocitosis no convencional aún no es del todo claro, por lo que
más de un vía podría estar implicada en este mecanismo (Níquel y Rabouille
2009). El modelo que se presenta en la Figura 3 describe varias formas de
explicar la secreción de las formas solubles y agregadas de -Sin. La secreción
de -Sin ocurriría en condiciones fisiológicas aunque sería más eficiente en
condiciones patológicas como la disfunción mitocondrial y proteosomal, lo que se
asocia a la EP (Lee et al. 2005).

35
Figura 3. Modelo hipotético de la secreción y de los blancos sinápticos de
agregados de -sinucleína. La figura muestra cuatro mecanismos diferentes
para la secreción no convencional de -Sin (Nickel et al. 2009). El mecanismo 1
representa la translocación no convencional de -Sin mediada por una proteína
transmembrana desconocida localizada en la membrana plasmática. Los
mecanismos 2-4 representan mecanismos vesiculares para la secreción de -Sin
que involucran lisosomas (2), microvesículas (3) o cuerpos multivesiculares (4).
Los mecanismos 3 y 4 secretan -Sin dentro de exosomas.
Una vez en el espacio extracelular (Lee, 2008) oligómeros de -Sin
podrían ser removidos a través de degradación proteolítica (A) o endocitosis (D)
(Desplats et al. 2009). La acumulación de -Sin en el espacio extracelular
activaría la microglia (B) o alteraría la membrana plasmática a través de la
formación de estructuras tipo poro/perforado (C) que podrían causar una
desregulación en la homeostasis del Ca2+ (Danzer et al. 2007). Modificado de
Pacheco et al. 2012.
ESPACIO EXTRACELULAR
NE
UR
ON
A P
RE
-SIN
ÁP
TIC
AN
EU
RO
NA
PO
ST-S
INÁ
PT
ICA
Agregados
de -Sin
Monómeros
de -Sin
Endocitosis
Mecanismo
no vesicular
1
D
Endocitosis
2
Lisosomas secretores
Exosoma
Desprendimiento de microvesículas
3
Cuerpos
multivesiculares
4
Endosomas
Activación
Microglial
B
Degradación
Proteolítica
A
Ca2+Ca2+
poro/perforado de
-Sin
C
Ca2+
poro/perforado
de -Sin
Ca2+

36
Rol del Ca2+ en la enfermedad de Parkinson
Existe numerosa evidencia que indica la participación del ión Ca2+ en la
EP, en relación a esto, algunos estudios indican que -Sin controlaría la función
de canales de Ca2+ voltaje dependientes (Adamczyk and Strosznajder 2006;
Hettiarachchi et al. 2009), mientras que otros estudios indican que oligómeros de
esta proteína incrementarían los niveles de Ca2+ intracelular (Danzer et al. 2007)
producto, probablemente, de alteraciones en la permeabilidad de la membrana
causando la muerte neuronal. Esto concuerda con estudios realizados en células
humanas que sobreexpresan mutantes de -Sin cuya permeabilidad iónica se ve
afectada por quelantes de Ca2+ (Furukawa et al. 2006), descartándose el
aumento en la conductividad de canales de Ca2+ dependientes de voltaje, ya que
al utilizar bloqueadores específicos para este tipo de canales (nifedipina y -
conotoxina-GVIA) no se observa protección frente a la muerte celular (Furukawa
et al. 2006). En este contexto, se ha determinado que agregados de A
(implicado en la EA) forman estructuras tipo poro/perforado en membranas
neuronales (Sepulveda et al. 2010). La formación de estos perforados se
acompaña de un incremento agudo en los niveles de Ca2+ intracelular y una
disminución significativa en proteínas sinápticas de neuronas tratadas
crónicamente con agregados de A (Parodi et al. 2010; Sepulveda et al. 2010).
Considerando el hecho de que otra proteína amiloideogénica como A se inserta
en la membrana y forma perforados (Sepulveda et al. 2010), se podría proponer
un mecanismo similar para explicar la neurodegeneración inducida por -Sin. Es
decir, la formación de estructuras tipo poro/perforado de -Sin en la membrana

37
plasmática conduciría a un aumento de la concentración libre de Ca2+ intracelular
lo que llevaría a la muerte celular. Esta idea se apoya por otro estudio que
muestra que células que sobreexpresan -Sin y luego son tratadas con A
presentan un aumento en la amplitud de corriente junto con un influjo de Ca2+
consistente con la formación de poros a nivel de la membrana plasmática
(Tsigelny et al. 2008).
Formación de estructuras tipo poro/perforado de -sinucleína
Uno de los primeros pasos para la formación estructuras tipo
poro/perforado es la asociación de -Sin a la membrana plasmática, lo que se
discute en las siguientes dos secciones:
a) Asociación de -sinucleína a membranas: Se ha demostrado que -Sin se
asocia con bicapas fosfolípidicas de membranas artificiales, especialmente, con
aquellas membranas que contienen fosfolípidos ácidos (Davidson et al 1998), ya
que la región N-terminal de -Sin se encuentra cargada positivamente a pH
fisiológico (Rhoades et al. 2006). Esta asociación estaría estabilizada mediante la
presencia de -hélices en la estructura secundaria de -Sin (Davidson et al.
1998; Jo et al. 2000; Eliezer et al. 2001; Chandra et al. 2003). Otro factor crítico
para la asociación de -Sin sería el tamaño de la membrana. Se ha demostrado
que -Sin se asocia, preferentemente, a vesículas con diámetros pequeños (20-
25 nm) (Davidson et al. 1998). Por otra parte, estudios recientes demuestran que
la interacción de -Sin con membranas vesiculares es un proceso rápido y
reversible, donde la unión de -Sin a membranas neutras y cargadas

38
negativamente se produce aparentemente por diferentes mecanismos
(Shvadchak et al. 2011).
En membranas de origen natural, -Sin ha demostrado asociarse, como
una proteína periférica de membrana, con las vesículas sinápticas (Maroteaux et
al. 1988), vesículas de transporte axonal (Jensen et al. 1998), micelas (Cole et al.
2002) y vesículas de levadura (Outeiro and Lindquist 2003). Se ha determinado
que la unión de -Sin se produce en la cara externa de la membrana plasmática,
especialmente, en las balsas lipídicas, dominios ricos en colesterol y
esfingolípidos (Fortin et al. 2004; Bar-On et al. 2008). Estos micro dominios se
caracterizan por una difusión lateral lenta, así como también de una resistencia a
los detergentes (Allen et al. 2007). Además, las balsas lipídicas están
involucradas en la interacción entre los lípidos y los residuos hidrófobicos de la -
hélice anfipática presente en el extremo N-terminal de -Sin (Tsigelny et al.
2007). La deleción del extremo N-terminal conduce a una disminución de la
toxicidad de -Sin, lo que sugiere que la toxicidad depende del dominio de unión
a la membrana, (Vamvaca et al. 2009). Esto fue confirmado por mutaciones
puntuales en el extremo N-terminal de -Sin (A30P, E46K, y A53T) asociada con
EP de tipo familiar (Kruger et al. 1998; Polymeropoulos et al. 1997; Zarranz et al.
2004).
b) Modelos de poros de -sinucleína en la membrana: Quizás uno de los
mayores inconvenientes para entender la estructura del perforado de -Sin, es la
carencia de modelos de alta resolución en membranas biológicas que permitan
entender el mecanismo por el cual perforados de -Sin ejercen su función

39
patológica. Sin embargo, existen, principalmente, dos tipos de modelos de poro
que podrían explicar las propiedades de perforación de -Sin, el toroidal y el de
barril (Yang et al. 2001). El modelo toroidal (Figura 4A) consiste en la unión
secuencial de monómeros de -Sin a las membranas lipídicas que resulta en la
formación de poros con una conformación -helicoidal (Zakharov et al. 2007). De
hecho, se cree que el carácter básico de las repeticiones KXKE del extremo N-
terminal actuaría como un sensor de voltaje de membrana (Zakharov et al. 2007).
Además, estudios in silico de modelamiento y dinámica molecular han
demostrado que la unión secuencial de -Sin a la membrana ocurre entre los
extremos N-terminal de cada uno de los monómeros, llevando a la formación de
pentámeros y hexámeros que generan estructuras tipo anillo cuyo tamaño fluctúa
entre los 9-15 nm, con un diámetro interno de 2-5 nm (Tsigelny et al. 2007).
Incluso muestras envejecidas de -Sin, con una esperada conformación de hoja
plegada, se unirían a vesículas en una estado -helicoidal (Smith et al. 2008).
Esto contrasta con el modelo de barril (Figura 4B) que indica que oligómeros de
-Sin (ricos en hojas ) formarían estructuras anulares con un poro central
(Volles et al. 2003; Schmidt et al. 2012). Estos agregados se unirían a la
membrana (Volles et al. 2001) causando su permeabilización, de manera
concentración dependiente, lo que ha sido demostrado en liposomas (Volles et
al. 2001) y bicapas de membrana artificial (Kayed et al. 2004), no demostrándose
el mismo efecto para monómeros ni para fibras de -Sin (Ding et al. 2002). De
acuerdo con esto, se ha observado a través microscopía de fuerza atómica
(AFM) que agregados de -Sin forman poros sobre membranas artificiales, con

40
un diámetro externo de 8-10 nm y un diámetro interno de 1-2 nm (Quist et al.
2005). Análisis de corrientes de canal único en membranas artificiales indican
que agregados de -Sin forman poros con distintas conductancias, lo que
sugiere que diversas especies oligoméricas componen estas estructuras (Quist
et al. 2005). En relación a las mutaciones presentes en -Sin, estudios de
sobreexpresión con mutantes de -Sin, A53T y A30P en células SH-SY5Y, han
indicado que el potencial de depolarización de membrana inducida por estas
mutantes es mayor a la observada por la sobreexpresión de -Sin nativa, lo que
nos indica de alguna manera un aumento en la permeabilidad de membrana
(Furukawa et al. 2006). Esto sugiere que mutaciones asociadas a la EP afectan
directamente la arquitectura del poro o favorecen la formación de especies
permeabilizantes o perforantes. Esta propiedad de formar estructuras tipo poro
estaría, específicamente, asociada a protofibras de -Sin ya que protofibras de -
Sin (también ricas en estructuras hojas ), no son capaces de permeabilizar o de
unirse a las vesículas lipídicas (Park et al. 2003). Esta propiedad permeabilizante
tipo poro, exhibiría selectividad de acuerdo al tamaño de la molécula, ya que
Ca2+ y dopamina son liberados en presencia de protofibras, mientras que
moléculas de mayor tamaño como citocromo c no pueden hacerlo (Volles et al.
2001). Referente a la sensibilidad de estas estructuras perforantes se ha
determinado que serían sensibles a Zn2+ producto, probablemente, de la
interacción entre el Zn2+ y los residuos de cisteína, histidina o arginina de -Sin;
que en el caso de la histidina se ha descrito previamente en las estructuras tipo
poro de A sensibles a Zn2+ (Kawahara et al. 1997). Específicamente, el residuo

41
de histidina 50 del monómero de -Sin se considera como un posible candidato
para la interacción con Zn2+, principalmente, porque se encuentra situado cerca
del sitio putativo del poro (Zakharov et al. 2007). Esto concuerda con estudios
realizados en células humanas que sobreexpresan mutantes de -Sin cuya
permeabilidad iónica se ve afectada por quelantes de cationes divalentes
(Furukawa et al. 2006).

42
Figura 4. Modelo de perforación de -sinucleína. De acuerdo al modelo
operacional de perforación de membrana, una vez que α-Sin interactúa con la
membrana plasmática ésta adquiere una nueva estructura secundaria para
formar estructuras tipo poros/perforados, el cual podría ser explicado por dos
modelos diferentes: El modelo toroidal (A) involucra la unión secuencial de
monómeros de -Sin a la membrana plasmática resultando en la formación de
poros con una conformación -helicoidal (Zakharov et al. 2007), mientras que el
modelo de barril (B) involucra la unión de oligómeros de -Sin (ricos en hoja )
resultando en la formación de estructuras tipo anillo con un poro central (Schmidt
et al. 2012).
Proteína soluble
-hélice
monómeroModelo toroidal
A)
B)
Modelo de
Barril
Oligómeros
monómero
Fibras amiloides
Hoja -plegada
Proteína soluble

43
HIPÓTESIS
Los antecedentes bibliográficos indican que:
1) Niveles elevados de oligómeros de -Sin extracelular parecen asociarse a la
EP.
2) Estudios previos sugieren que -Sin perforaría membranas artificiales, de la
misma forma que oligómeros de A.
3) Oligómeros de -Sin aumentan los niveles de Ca2+ intracelular de manera
similar que oligómeros de A.
4) Hasta el momento se desconoce cuál es el mecanismo de propagación y
toxicidad por el cual -Sin extracelular produce neurodegeneración en distintas
zonas cerebrales.
Lo anteriormente descrito permite proponer como hipótesis de trabajo que:
“Oligómeros de -sinucleína extracelular causan disrupción de la membrana
plasmática neuronal, aumento en los niveles de Ca2+ intracelular y
sinaptoxicidad”.
Este tipo de estudio posee una relevancia significativa en el conocimiento
de la propagación de la EP a distintas zonas del cerebro como, por ejemplo, el
hipocampo, siendo sus resultados de vital importancia para la generación de
fármacos modificantes para el tratamiento de pacientes con EP.

44
OBJETIVO GENERAL
Estudiar los mecanismos celulares y moleculares que median la
sinaptotoxicidad de -sinucleína a través de la formación de perforados en la
membrana plasmática neuronal.
OBJETIVOS ESPECÍFICOS
Objetivo Específico I. Caracterizar bioquímicamente agregados de -
sinucleína y determinar si forman perforados en membranas neuronales.
1.1 Caracterizar bioquímicamente y estructuralmente agregados de -sinucleína.
1.2 Determinar si agregados de -sinucleína modifican la conductancia de
membrana en neuronas hipocampales.
1.3. Caracterizar la asociación de -sinucleína en neuronas hipocampales y
células RCSN-3.
Objetivo Específico II. Caracterizar efectos funcionales de -sinucleína en
neuronas hipocampales.
2.1 Determinar si agregados de -sinucleína alteran los niveles de Ca2+
intracelular y la actividad sináptica neuronal.
2.2 Determinar la toxicidad mediada por agregados de -sinucleína.

45
2.- MATERIALES Y MÉTODOS
Cultivo de neuronas hipocampales. Ratones C57BL/J6 o ratas Sprague Dawley
fueron tratados y manipulados de acuerdo a las guías éticas de manejo y cuidado
de animales de experimentación establecidos por el NIH (NIH, Maryland, EE.UU.)
y el Comité de ética de Universidad de Concepción. Animales preñados de 18-19
días fueron ubicados en una cámara con un papel embebido en éter y sacrificados
por dislocación cervical. Luego se removieron los embriones y rápidamente
decapitados para disecar el hipocampo. Las neuronas fueron sometidas a
disociación mecánica y enzimática para ser plantadas a una densidad de 180.000
cel/ml en cubreobjetos de vidrio de 35 mm recubiertos con poli-L-lisina (0.25%
P/V; PM> 350 kDa, Sigma, EE.UU). Los cultivos fueron mantenidos a 37ºC y 5%
CO2 y el medio fue reemplazado cada 3 días, el medio de cultivo consistió en 90%
minimal essential medium (MEM, GIBCO, Rockvile, EE.UU), 5% V/V de suero de
caballo (Hyclone) inactivado por temperatura, 5% de suero fetal bovino y N3 (una
mezcla de nutrientes). Los experimentos se realizaron en neuronas de 11-13 DIV.
Cultivo de células RCSN-3. La línea celular RCSN-3 (generosamente donada por
el Dr. Pablo Caviedes del laboratorio de Terapia Celular, Instituto de Ciencias
Biomédicas (ICBM), Facultad de Medicina, Universidad de Chile), fue crecida en
monocapa hasta una confluencia del 70%, el medio de crecimiento utilizado fue
DMEM/F12 (1:1) (GIBCO), suero de caballo (HyClone) al 6% y suero bovino fetal
(HyClone) al 6%. Se adicionó bicarbonato (SIGMA-ALDRICH) 1 g/L y HEPES

46
(GIBCO) 15 mM, para luego ajustar pH y alcanzar valor de 7.4. Las células fueron
incubadas a 37ºC, con 100% de humedad en una atmósfera de 5% de CO2.
Cultivo de células SHSY-5Y. La línea celular estable SHSY-5Y es una línea
derivada de neuroblastoma humano que ha sido transfectada para sobreexpresar
wild type -Sin y como control se utilizaron células SHSY-5Y que sobreexpresan
establemente -galactosidasa (-gal), donación del investigador Kostas Vekrellis,
del Departamento de Biología celular y Biofísica de la Facultad de Biología en la
Universidad de Atenas. Se trataron de acuerdo al método descrito por (Vekrellis et
al. 2009). En resumen fueron mantenidas con RPMI 1640 (Hyclone) suplementado
con 10% de suero fetal bovino (FBS 10%) con G418 (Sigma) 250 g/mL, y con
Hygromicina B (Sigma) 50 g/mL. La expresión de -Sin fue silenciada por un
vector “Tet Off”, regulado por Doxiciclina (Vekrellis et al. 2009), lo que permite
controlar los niveles de -Sin secretada en el medio (Emmanouilidou et al. 2010).
Las células fueron sembradas con densidad de aproximadamente 4x104
células/cm2 y se obtuvo el medio condicionado a diferentes días de crecimiento,
hasta llegar a un máximo de 80% de confluencia, el que se mantuvo a -20ºC.
Agregación de -sinucleína. -Sin recombinante humana (1-140) (Anaspec,
USA) fue disuelta en agua estéril y mantenida en alícuotas de una concentración
346 M a -20ºC. Éstas alícuotas fueron luego diluidas en DPBS (Dulbecco’s
Phosphate-Buffered Saline; Gibco, EE.UU.) a pH 4.0, dejando un stock de
proteína soluble a una concentración 59 M el cual fue mantenido a 4ºC. Para la

47
formación de agregados -Sin soluble fue agitada a 800 rpm por 24 horas a 37ºC
en un Thermomixer Compact (Eppendorf, Alemania).
Obtención de rebanadas de neuronas de sustancia nigra. Se prepararon
rebanadas de sustancia nigra de ratón C57BL/6 (P20) de 300 μm en solución de
corte (concentraciones mM: Sucrosa 194, NaCl 30, KCl 4.5, MgCl2 1, NaHCO3 26,
NaH2PO4 1.2, Glucosa 10) usando el vibratomo Leica VT1200S. Las rebanadas
fueron incubadaa en solución de aCSF (concentración mM: NaCl 124, KCl 4.5,
MgCl2 1, NaHCO3 26, NaH2PO4 1.2, Glucosa 10, CaCl2 2) y saturadas con 95% de
O2/ 5% de CO2 por 1 hora a 37ºC.
Disociación de rebanadas de neuronas de sustancia nigra. Las rebanadas
fueron disociadas mecánicamente con una pipeta de vidrio con extremo curvo a
una frecuencia de 5 Hz, disgregando la zona deseada por identificación
histológica. Se esperaron 15 minutos en solución externa normal para que se
adhirieran las células disociadas al pocillo de trabajo. De esta manera se
obtuvieron neuronas disociadas de ratón para posteriores análisis
electrofisiológicos.
Electrofisiología. El medio de cultivo del plato se cambió por una solución
externa normal que contiene 150 mM NaCl, 5.4 mM KCl, 2.0 mM CaCl2, 1.0 mM
MgCl2, 10 mM glucosa y 10 mM HEPES (pH 7.4). La solución interna contiene 120
mM KCl, 2.0 mM MgCl2, 2 mM ATP-Na2, 10 mM BAPTA, 0.5 mM GTP, 10 mM
HEPES (pH 7.4). Para la configuración de patch clamp perforado (Figura 5) el
agente perforante fue agregado en la solución interna (pipeta) y un pulso de 5 mV

48
fue utilizado para monitorear la perforación utilizando un amplificador Axopatch
200B (Molecular Devices, EE.UU.) como previamente ha sido descrito (Sepulveda
et al. 2010). Para las corrientes postsinápticas en miniatura las cuales fueron
registradas en presencia de 25 nM de tetrodotoxina (TTX, Sigma,St Louis M.O,
EE.UU.) se utilizó el sistema de whole-cell patch clamp (Hamill et al. 1981). El
potencial de membrana se ajustó a -60 mV. Los electrodos se construyeron a
partir de capilares de borosilicato (WPI, Sarasota, FL) en un “puller” horizontal
(Sutter Instruments P-89, Novato, CA). La corriente se adquirió a intervalos de 5
ms y se filtró a 2 kHz utilizando un computador conectado al sistema de registro
mediante una tarjeta de adquisición (Digidata 1200, Axon Instruments, Inc.) y el
programa computacional AxoScope 9.0 (Axon Instruments, Inc.).
Figura 5. Esquema de la configuración de Patch Clamp Perforado. A,
Configuración de sello cuya resistencia está a nivel de gigaohms luego de una
pequeña succión sobre la pipeta de patch. B, configuración de perforado, la cual
se adquiere al utilizar compuestos con carácter perforante en la pipeta de patch.
C, Representación esquemática de la corriente capacitativa, Q1 y Q2 representan
la carga de la membrana, Tau es la constante de decaimiento del Peak
capacitativo, Rm y Ra son la resistencia de membrana y de acceso,
respectivamente, Cm es la capacitancia (Cm = Q/V).
Q1 = C x V
DI = AV / (Ra+Rm)
I1 (at V1)
I2 (at V2)
Cm = Q
V
Q2 = DI x Tau
Tau
C
Agentes
Perforantes
Configuración
de Sello
Configuración
Perforado
BA

49
Inmunogold. Control (DPBS pH4) y agregados de -Sin (5 M) fueron adsorbidos
en grillas de níquel cubiertas con una resina de soporte Formvar por 5 minutos. La
unión no específica fue bloqueada con ASB (albúmina de suero bovino) 3% en
TBS (Tris-buffer salino) 1X pH 7.4, por 20 minutos. Luego, las grillas fueron
colocadas sobre una gota del anticuerpo monoclonal 211 producido contra los
aminoácidos 121-125 de -Sin de origen humano (dilución 1:50 en TBS 1X pH 7.4,
con ASB 0.1%; Santa Cruz Biotechnology, CA, EE.UU.) por 30 minutos para,
posteriormente, ser lavadas 6 veces sobre una gota de solución de TBS 1X pH
7.4, por dos minutos. En seguida, una gota de anticuerpo anti-conejo IgG
conjugado a partículas de oro de 10 nM (dilución 1:50 en TBS 1X pH 7.4, con ASB
0.1%; Sigma, EE.UU.) fue agregado por 20 minutos para luego ser lavado
(Naslund et al. 1995). Antes de la examinación final bajo el microscopio electrónico
JEOL 100-B, los especímenes fueron teñidos negativamente con 0.2% de ácido
fosfotúngstico en agua.
Western blot. Cantidades de 10 g de proteínas obtenidas de lisados totales de
neuronas de hipocampo (11 a 13 DIV) tratadas con agregados -Sin (0.5 M),
fueron cargadas en un gel denaturante de Tris-tricina. Luego las proteínas fueron
transferidas a una membrana de nitrocelulosa, la cual posteriormente fue
bloqueada 1 hora en leche al 5%, lavadas 5 veces en TBS-tween e incubadas 1
hora con el anticuerpo anti--Sin 1:1000 (Santa Cruz Biotechnology, CA, EE.UU.)
y anti- actina 1:1000 (Santa Cruz, Biotechnology); lavadas nuevamente 5 veces
en TBS-tween e incubadas 1 hora con el anticuerpo secundario conjugado a HRP

50
(1:5000, SCBT, EE.UU). Posteriormente fueron reveladas utilizando un kit de
quimioluminiscencia (PerkinElmer, Boston, MA, EE.UU.).
Tinción con tioflavina-T. Agregados de -Sin fueron teñidos con tioflavina-T
(ThT, 1 mM, Sigma Aldrich, EE.UU.) por 15 minutos a 37ºC. Para el tratamiento
con proteinasa K (PK, 50 g/mL), los agregados fueron incubados con la enzima
por 30 minutos a 37ºC. La fluorescencia de ThT fue adquirida luego de la
excitación de las muestras a 488 nm.
Tinción de plata. Para tinción de plata se utilizó el kit Silver SNAP Stain II® (Bio-
Rad, Pierce, Rockford, EE.UU.). De acuerdo a las instrucciones del fabricante, una
vez realizada la electroforesis el gel se lava varias veces en agua ultrapura y en
mezcla etanol 30% y ácido acético 10%, posteriormente en solución sensitizer® y
se tiñe por 30 minutos en solución stain®. Se realiza un lavado en solución
developer® hasta que las bandas aparezcan y se detiene la reacción en ácido
acético al 5%.
Tinción con rojo congo. La tinción de los agregados de -Sin, con rojo congo se
realizó según el método modificado de Putchler. (Putchler et al., 1962). Se
preparó una solución alcalina de NaCl saturado en etanol 100% (80% etanol,
0.005% NaOH 1M, saturado NaCl) donde se disolvió rojo congo (SIGMA)
llevándolo a un stock de 0.002 g/mL. De este preparado se tomó una alícuota de
trabajo y se filtró, utilizando filtros estériles de celulosa-acetato VWR, de 0.45 m.
Para teñir los agregados se dejó incubar la preparación de rojo congo filtrado con
los agregados en proporción 1:1 a temperatura ambiente por 30 minutos. Se

51
centrifugó a 14000 rpm por 4 minutos, se eliminó el sobrenadante y se
deshidrataron las muestras en etanol 100% para quitar el exceso de rojo congo, al
menos 3 veces. Luego se resuspendió el pellet en la solución alcalina de NaCl
saturado en etanol y se montaron 20 L de los agregados teñidos en portaobjetos
para llevar a visión con luz polarizada en Microscopio invertido. Las imágenes
fueron captadas con cámara digital 5.0 Mpixeles (Canon A460, Japón) y
calibradas utilizando una cámara de Newbauer y el software ImageJ 1.47h (NIH,
EE.UU.), con un zoom de 60x.
Viabilidad celular. La viabilidad celular fue determinada por el ensayo de Alamar
Blue (Invitrogen), que se basa en la conversión de Resazurina (no fluorescente) a
Resofurina (fluorescente) en el medio reductor de las células vivas (Life
technologies). Los cultivos fueron incubados por 1-4 horas a 37°C con Resazurina
en una razón de 1:10 de reactivo:medio de cultivo. Las muestras fueron excitadas
a 570 nm y la señal fluorescente de Resazurina fue determinada a 585 nm. La
fluorescencia fue medida en equipo NOVOstar (BMG LABTECH).
Incremento de Ca2+ intracelular. Las células fueron incubadas por 30 minutos a
37ºC con Fluo4-AM (5 M, Invitrogen, USA), lavadas por 30 minutos a 37ºC luego
montadas en una cámara de perfusión y llevada al microscopio. Se seleccionaron
regiones de interés (ROIs) correspondientes a regiones somáticas de las neuronas
y se determinó los cambios en los niveles de Ca2+ intracelular en neuronas control
y tratadas a diferentes tiempos con agregados de -Sin (0.5 M). Las imágenes
fueron recolectadas a intervalos de 20 segundos durante un periodo continuo de

52
75 minutos y para las transitorias de Ca2+ (sólo neuronas hipocampales) a
intervalos de 2 segundos durante 5 minutos. Los experimentos se realizaron en
presencia de Ca2+ extracelular y/o en presencia de Co2+ 10 M en todas estas
condiciones las células fueron lavadas por 15 minutos en solución externa
después de cada tratamiento.
Microscopía y análisis de fluorescencia para FM1-43 y Fluo4-AM. Los
registros fueron realizados utilizando un microscopio invertido Nikon (Eclipse TE,
Nikon) equipado con una lámpara de Xenón, objetivos de 100X y 40X, una rueda
de filtros Lambda 10-2 (Sutter Instruments) controlada por el software Axon
Workbench 2.2 (Axon Instruments). El microscopio posee acoplado una cámara
CCD de 12 bits (SensiCam, PCO, Germany) y el binning para todos los
experimentos fue de 2x2 pixeles Fluo4-AM y FM1-43 se excitó a 480 nm y la
emisión fue colectada con un filtro de 510 nm.
Inmunocitoquímica. Neuronas hipocampales y células RCSN-3 se fijaron durante
15 minutos con paraformaldehído a -4°C y se permeabilizaron con Tritón X-100 al
0.1% en PBS 1x. En seguida se lavó 3 veces por 5 minutos para a continuación
bloquear las células por 1 hora con suero de caballo (Hyclone) diluido 1:10. Luego
del bloqueo se incubó durante la noche con los siguientes anticuerpos primarios:
anti -Sin recombinante humana (Santa Cruz) (1:300), anti -Sin endógena de
rata y ratón (Cell Signaling) (1:300), Lámina B (Santa Cruz) (1:100) y anti
Ubiquitina (Dako Cytomation) (1:100), anti MAP-2 (Santa Cruz)(1:400) y anti-
SNAP25 (Synaptic System)(1:50). Posteriormente, se lavó 3 veces con PBS para
incubar por 1 hora y en oscuridad con el anticuerpo secundario correspondiente

53
(Jackson ImmunoReasearch). A continuación, los núcleos se tiñeron con 4',6-
diamino-2-fenilindol (DAPI, 300 nM, Invitrogen, EE.UU.) durante 1-2 minutos y se
lavó con PBS 1X 3 veces durante 5 minutos. Para las células de RCSN-3,
posteriormente, se tiñeron con Faloidina Alexa Fluor 488 (50 nM, Invitrogen,
EE.UU.). Por último, las células fueron montadas en portaobjetos utilizando medio
de montaje (Dako). La adquisición de imágenes se realizó en el Centro de
Microscopía Avanzada (CMA) con el microscopio confocal LSM700 y el
microscopio espectral LSM780 (Zeiss, Alemania); mientras que el procesamiento
de éstas se ejecutó en el software ImageJ 1.47h (NIH, EE.UU.). Se realizaron
pruebas de reactividad para descartar detección cruzada para los anticuerpos anti
-Sin humana y anti -Sin endógena, observándose especificidad de los
anticuerpos por la especie descrita en los respectivos datasheets.
Marcaje de vesículas sinápticas con FM1-43. Neuronas de hipocampo (10-13
DIV) fueron lavadas con solución HBSS (Hank's Balanced Salt Solution, Gibco,
USA) e incubadas en solución externa alto K+ (60 mM) a 37ºC por 5 minutos,
luego de removida esta solución fueron cargadas con FM1-43 10 M (Molecular
Probes, EE.UU.) por 5 minutos a 37ºC y lavadas nuevamente en solución HBSS
por 20 minutos a 37ºC y por 1 hora a 4ºC para ser rápidamente montadas en una
cámara de perfusión acoplada a un microscopio de epifluorescencia (Nikon,
Eclipse 3000). Para evaluar el decaimiento de la fluorescencia asociada a FM1-43
(destaining) para neuronas tratadas y controles se utilizó solución externa alto K+
60 mM (reemplazo equivalente de Na+ por K+) como estímulo despolarizante. Para
evaluar el efecto agudo de agregados α-Sin se seleccionaron las distintos ROIs y

54
se determinó el decaimiento de la fluorescencia asociada a FM1-43 en presencia y
ausencia de agregados -Sin en el medio externo.
Para el estudio de endocitosis post-estímulo, el protocolo utilizado fue
modificado de uno descrito por Ryan en 1996, el cual se basa en estimular y
esperar intervalos de tiempo entre el estímulo despolarizante y la carga con FM1-
43. La cantidad de vesículas cargadas (FM1-43 positivas) disminuye a medida que
se aumenta el intervalo de tiempo entre el estímulo y la carga con FM1-43, ya que
han sido re-endocitadas previamente sin FM1-43.
Análisis de datos. Para el análisis cuantitativo de los datos inmunocitoquímicos,
las neuronas hipocampales fueron escogidas al azar para la adquisición de
imágenes (5-10 células por condición, en 3 o más experimentos independientes)
usando un microscopio espectral LSM780 (objetivo de inmersión en aceite 63X,
Zeiss, Alemania). Las imágenes fueron procesadas utilizando el programa ImageJ
1.47h (NIH, EE.UU.). Para el análisis cuantitativo de los registros en patch clamp
perforado se utilizó el programa ClampFit 10.1 (Axon Instruments) y para las
corrientes en miniatura se utilizó el programa Mini Analysis 6.0.7 (Synaptosoft Inc,
EE.UU.). Todos los valores son presentados como promedios ± error estándar
(ES) para 3 o más experimentos independientes. Los análisis estadísticos fueron
hechos utilizando test de Student o ANOVA seguido de un post-test Bonferroni.
Valores de *p<0.05 fueron considerados estadísticamente significativos. Todos los
análisis estadísticos fueron realizados utilizando GraphPad Prism5.

55
3.- RESULTADOS
Objetivo Específico I. Caracterizar los agregados de -sinucleína y
determinar si forman perforados en membranas neuronales
La EP es principalmente descrita como una enfermedad motora. Sin
embargo, la demencia, un síntoma no motor, es detectada en una gran proporción
de pacientes con EP (Aarsland et al. 2007), la cual puede estar acompañada de
perturbaciones del sueño, constipación, impotencia y pérdida del olfato (Barone et
al. 2009; Ferrer et al. 2011), lo que sugiere la participación de múltiples áreas
cerebrales.
Los mecanismos de iniciación y de propagación de la EP asociada a la
acumulación de la proteína citosólica -Sin no están bien entendidos, siendo aún
una incógnita la función que ejercería a nivel extracelular (El-Agnaf et al. 2006).
Una de las hipótesis plantea que -Sin extracelular se uniría a nivel de la
membrana plasmática neuronal favoreciendo procesos de permeabilización (Quist
et al. 2005; Zakharov et al. 2007; Tsigelny et al. 2007; Kim et al. 2009; Feng et al.
2010; Kostka et al. 2010; Schmidt et al. 2012). Actualmente diversos estudios
sugieren que los agregados de -Sin son capaces de formar estructuras tipo poro
en membranas lipídicas artificiales (Quist et al. 2005; van Rooijen et al. 2010;
Schmidt et al. 2012), sin embargo, aún se desconoce su existencia a nivel de
membranas celulares. Esto resultaría sumamente interesante, ya que explicaría la
progresión de la EP a distintas zonas cerebrales tanto motoras como cognitivas.

56
De este modo, se ha intentado determinar la formación de perforados por
parte de agregados de -Sin a nivel de membrana plasmática tanto de neuronas
hipocampales, modelo neuronal asociado a procesos de aprendizaje y memoria,
como de células RCSN-3, modelo celular dopaminérgico asociado a procesos
motores.
Objetivo 1.1. Caracterizar bioquímicamente y estructuralmente agregados de
-sinucleína
Diversos estudios señalan que son las especies amiloidogénicas de bajo
peso molecular de -Sin las responsables de la sinaptotoxicidad, neurotoxicidad y
progresión de la enfermedad a través del cerebro (Kayed et al. 2003; Tsika et al.
2010; Colla et al. 2012).
De este modo, el desarrollo del presente objetivo involucró la formación de
agregados de -Sin de bajo peso molecular (oligómeros) para los experimentos a
realizar en la tesis.

57
Resultados
Oligómeros de -sinucleína son principalmente especies solubles, de bajo
peso molecular y sensibles a proteínasa K
En una primera etapa examinamos las constantes de cinética de
agregación proteica utilizando el Modelo de Finke-Watsky (Figura 6B; Solomon et
al., 2011). Un estudio reciente indica que para la obtención de estructuras
anulares de -Sin es necesaria la utilización de pHs en el rango de 3.8 a 4.2
(Lasagna-Reeves et al., 2011), por lo tanto, evaluamos la utilización de un DPBS
pH 4 y pH 7 para la agregación de -Sin. De acuerdo a los datos obtenidos (Figura
6C) los resultados indican que K1, constante que cuantifica el proceso de
nucleación, es 11 veces mayor utilizando DPBS pH 4 v/s pH 7 (0,3251 v/s
0,02989). Por otro lado, K2 constante que cuantifica el proceso de elongación
proteica no varía al utilizar ambos buffers (pH 4= 1 * 10 exp -007 y pH 7= 1 * 10 exp -
007) (Figura 6D). Dado los resultados obtenidos, que sugieren que la utilización de
un buffer pH 4 favorece la formación de núcleos de agregación sin promover la
elongación proteica decidimos utilizar agregados de -Sin obtenidos en presencia
de DPBS pH 4 para los posteriores estudios funcionales.

58
Figura 6. Agregación de -sinucleína. A, Esquema de la agregación proteica a
través del tiempo. B, Ecuación de Finke-Watzky. C, Curso de tiempo de la
agregación de α-Sin soluble a 800 rpm por 48 horas a 37ºC monitoreado por la
dispersión óptica a 405 nm (Turbidez). D, Constantes K1 and K2 de la agregación
de -Sin obtenidos a pH 4 y pH 7. Las barras corresponden al promedio ± SEM
obtenido desde 3 experimentos independientes (n=3).
Y= A-((k1/k2)+A)
(1+(k1/(k2*A)exp((k1+k2*A)*X)))+C
Ecuación de Finke-Watzky
A B
A+B 2B
k1
k2
% P
rote
ína
ag
reg
ad
a
Tiempo (hrs)
t inducción
Pendiente k2
C
A B
D
-Sin K1
(h -1 )
K2
( μM-1 h -1 )
pH 4 0.325 1 e-007
pH 7 0.03 1 e-007
0 10 20 30 40 500.0
0.2
0.4
0.6
0.8 -Sin pH 4
-Sin pH 7
Tiempo (hrs)
Tu
rbid
ez (
D.O
.)
(405 n
m)

59
Luego del proceso de agregación por 24 horas, se caracterizaron las
propiedades estructurales y bioquímicas de los agregados de -Sin utilizando MET
y Western blot. La realización de inmunogold, con el anticuerpo especifico 211
contra los aminoácidos 121-125 de -Sin humana, sugirió la presencia de
oligómeros con agrupaciones de 2 (dímeros de -Sin) y 3 (trímeros de -Sin)
nanopartículas de oro (Figura 7B, B2-B3), a diferencia de lo que se obtiene luego
de la agregación de A1-42 donde claramente se observa la presencia de
estructuras fibrilares al mismo tiempo de agregación (Figura 7C). Los datos
estructurales de -Sin concordaron con los resultados obtenidos mediante
Western blot que revelaron la existencia de especies de -Sin SDS-resistentes de
bajo peso molecular presentes, principalmente, en la fracción soluble luego de 15
minutos de centrifugación a 10.000 rpm (Figura 7E).

60
Figura 7. Oligómeros de -sinucleína son especies principalmente solubles.
Las microfotografías electrónicas corresponden a muestras del vehículo de -Sin
(A), a oligómeros de α-Sin (B) y a agregados de A1-42 (C) teñidas negativamente
con ácido fosfotúngstico obtenidas luego de agitación a 800 rpm por 24 horas a
37ºC. La inmunorreactividad del anticuerpo primario correspondiente para -Sin y
A1-42 fue detectada con anticuerpos secundarios unidos a nanopartículas de oro
(10 nm). Los recuadros indican las zonas amplificadas en los paneles B1-B3. D,
Cuantificación del número de nanopartículas de oro por agrupación (cluster) de los
oligómeros de α-Sin. E, Patrón electroforético de oligómeros de α-Sin, analizado
por Western blot con el anticuerpo especifico 211. En la figura se muestra la
inmunorreactividad observada en la fracción total (T) y en las fracciones obtenidas
(fracción soluble (S) y respectivo precipitado (P)) después de centrifugar la
fracción total a 10.000 rpm por 15 minutos. Las barras corresponden al promedio ±
SEM obtenido desde 3 experimentos independientes (n=3, * p< 0.05).
1 2 30
10
20
30
40
50
Partículas de oro
por agrupación
**N
º d
e p
art
ícu
las
de o
ro p
or
m
2
D
E
Vehículo α-Sin 24 hrs
B1
B2
B3 B1
B2
B3
50 nm300 nm 300 nm
300 nm
A 1-42
95
72
26
17
34
170
55
43
130
kDa T S P
M
D
T
A B
C

61
Las especies oligoméricas constituyen intermediarios en el proceso de
agregación: los oligómeros “on-pathway” se dirigen hacia a la formación de fibras
amiloides que se caracterizan por una estructura secundaria rica en hojas -
plegada, que les permite tener una alta afinidad por colorantes como tioflavina T
(ThT) y rojo congo (con el cual emite birrefringencia verde-manzana bajo luz
polarizada), una alta resistencia a la degradación proteolítica y una apariencia
fibrilar bajo MET (Ross and Poirier 2004); mientras que los oligómeros “off-
pathway” no permiten la formación de este tipo de estructuras. Por lo tanto,
investigamos el efecto de la enzima proteínasa K (PK) en los grandes agregados
de -Sin obtenidos luego de 48 horas de agitación para determinar cuál vía de
agregación siguen los oligómeros de -Sin previamente caracterizados. En
ausencia de PK, observamos la presencia de agregados de -Sin ThT-positivos
ricos en estructura hoja -plegada con un rango de tamaño de 2-22 µm2 (n= 100)
que disminuyó significativamente a 1-11 µm2 en la presencia de PK (n= 141,
Figura 8B), resultado que fue corroborado por análisis en electroforesis en geles
tris-tricina con posterior tinción de plata (Figura 8C). Este resultado concordó con
los datos obtenidos por MET y tinción con rojo congo donde se observó la
presencia de estructuras fibrilares difusas (Figura 8D), congofílicas y sin
birrefringencia verde-manzana bajo luz polarizada (Figura 9).
En conjunto, los resultados indican que -Sin recombinante luego de 24
horas de agitación corresponde a una mezcla de agregados -Sin, rica en
oligómeros de bajo peso molecular, con estructuras altamente estables resistentes

62
a SDS y principalmente solubles, siguiendo una vía de agregación alternativa a la
fibrilización convencional.
Figura 8. Tinción de agregados de -sinucleína con tioflavina T y tratamiento
con proteinasa K. A, Imágenes de fluorescencia de agregados de -Sin (5 µM) y
controles teñidos con tioflavina T (ThT, 1 mM; Ex:Em 450:480 nm) obtenidos luego
de agitación a 800 rpm por 48 horas a 37ºC y posterior tratamiento con proteinasa
K (PK, 50 μg/mL) por 30 minutos a 37 ºC. Los recuadros indican las zonas
amplificadas que se muestran en los paneles inferiores. B, Frecuencia de
distribución del tamaño de los agregados de -Sin. C, Tinción de plata de los
agregados de -Sin controles y tratados con PK. D, Microfotografía electrónica de
agregados más amorfos de -Sin obtenida luego de 48 horas de agregación,
inmunomarcados con anticuerpo 211 y anticuerpo secundario unido a
nanopartículas de oro (10 nm). Las imágenes representan al menos 9 imágenes
obtenidas desde 3 experimentos independientes (n=3).
ThT (1 mM)Vehicle
kDa PK - +
9572
26
17
34
170
55
43
130
-Sin
A
B
C
-Sin 48 hrs -Sin 48 hrs + PK
A2
A1
A1
20 m
5 m
A2
20 m
5 m
D
0 5 10 15 20 250
10
20
30
40
50-Sin
-Sin + PK
Area (m2)
Fre
cu
en
cia
de d
istr
ibu
ció
n
(n
º d
e a
gre
gad
os d
e
-Sin
)
-Sin 48 hrs
300 nm
-Sin 48 hrs
M
D
T
T

63
Figura 9. Tinción de agregados de -sinucleína con rojo congo. A, Los
paneles de la izquierda presentan imágenes obtenidas bajo campo claro y los
paneles del medio presentan imágenes obtenidas bajo luz polarizada, de muestras
controles (vehículo) y de los agregados de -Sin (5 M) obtenidos luego agitación
a 800 rpm por 48 horas a 37ºC y posteriormente teñidos con rojo congo (1
mg/mL). Los recuadros A1 y A2 indican las zonas amplificadas que se muestran
en los paneles de la derecha. Las imágenes representan al menos 9 imágenes
obtenidas desde 3 experimentos independientes (n=3).
ALuz polarizadaCampo claro
20 m
20 m
20 m
20 m
-Sin
Vehículo
A2A1
A1
A2
4 m
4 m

64
Objetivo 1.2. Determinar si agregados de -sinucleína modifican la
conductancia de membrana en neuronas hipocampales
¿Cómo puede -Sin extracelular afectar las funciones sinápticas y
celulares?. Un posible mecanismo es que -Sin se asocie a la membrana
induciendo el adelgazamiento de la membrana en un proceso tipo-detergente o
insertarse en la membrana formando “estructuras tipo poro” (Quist et al. 2005;
Zakharov et al. 2007; Tsigelny et al. 2007; Kim et al. 2009; Feng et al. 2010;
Kostka et al. 2010; Schmidt et al. 2012). Al respecto, estudios electrofisiológicos
utilizan antibióticos/antifúngicos en la pipeta de patch para perforar membranas
celulares y registrar corrientes iónicas de la célula completa con la técnica de
patch clamp conocida como configuración de perforado (Ebihara et al. 1995;
Tajima et al. 1996; Andersen et al. 2005).
Por consiguiente, el desarrollo del presente objetivo involucró la utilización
de la técnica de patch clamp perforado para determinar si oligómeros de -Sin
afectan la conductancia de la membrana a través de la formación de poro o algo
de similar función.

65
Resultados
Oligómeros de -sinucleína incrementan la conductancia de membrana en
neuronas hipocampales
Los datos revelaron que utilizando una solución control (vehículo de -Sin)
en la pipeta de patch fueron obtenidos registros estables por 20 minutos o más
(Figura 10). Por ejemplo, la aplicación de un pulso de voltaje de 5 mV produjo una
pequeña carga transferida (dada principalmente por la pipeta de patch), la cual fue
compensada electrónicamente. Esta carga transferida fue significativamente
aumentada 5 veces sobre el control cuando agregados de -Sin (0.5 M) fueron
añadidos en la solución interna presente en la pipeta de patch luego de ~20
minutos de incubación (efecto no observado en presencia de -Sin soluble). Sin
embargo, la carga inducida por oligómeros de -Sin fue menor (30%) que la
producida mediante la ruptura de la membrana con presión positiva (configuración
de célula completa). Adicionalmente, el efecto de -Sin fue bloqueado por la co-
incubación con el anticuerpo 211.

66
Figura 10. Efecto de los oligómeros de -sinucleína en la corriente
capacitativa de membrana en neuronas hipocampales. A, Trazos
representativos de la corriente capacitativa de membrana en condiciones
controles, o luego de la aplicación de oligómeros de -Sin (0.5 M) en ausencia o
presencia del anticuerpo 211 (1:50). Como control de oligomerización se utilizó -
Sin soluble (1 M). Se muestra la corriente de una célula sometida a presión
negativa (célula completa). B, Cuantificación de la carga de membrana transferida
(veces sobre el control). Las barras corresponden al promedio ± SEM obtenido de
al menos 3 experimentos independientes (*** p< 0.001, ** p< 0.01).
Contr
ol M
-Sin
0.5
Cél
ula c
omple
ta
M +
211
-Sin
0.5
M s
oluble
-Sin
1
0
5
10
15
20
25
**
***
n= 10 n= 6 n= 10 n= 4n= 3
Carg
a d
e m
em
bra
na
tran
sfe
rid
a
(veces s
ob
re e
l co
ntr
ol)
A B
-Sin 0.5 M
Control
50403020100
Time (ms) Sw eep:1 Visible:1 of 13
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 10
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 21
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 47
IN 0
(pA
)
-2500
-2000
-1500
-1000
-500
0
500
1000
1500
2000
2500
1 2 43
-Sin 0.5 M + 211
50403020100
Time (ms) Sw eep:1 Visible:1 of 41
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 192
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 15
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
Célula completa
50 p
A
10 ms
0 min 20 min
0 min 20 min
0 min 20 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 1144
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 3 4
50403020100
Time (ms) Sw eep:1000 Visible:1 of 3290
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 3 4
0 min 20 min
-Sin 1 M soluble

67
Para determinar sí los perforados de -Sin permiten la entrada dentro de la
célula de moléculas biológicamente activas, utilizamos un análogo no hidrolizable
de glucosa fluorescente, 6-NBDG, con un diámetro de van der Waals de ~ 1 nm.
Los datos (Figura 11) muestran que la aplicación de oligómeros de -Sin (0.5 M)
causan un incremento sobre las unidades de fluorescencia relativa de 5 veces
sobre el control, lo que se correlacionó con el aumento en la corriente capacitativa
de membrana determinada anteriormente (Figura 10). El efecto de -Sin
incrementando la fluorescencia de la célula fue completamente bloqueado con el
anticuerpo 211. Por lo tanto, estos datos sugieren que la presencia de oligómeros
de -Sin generan grandes rupturas a nivel de la membrana plasmática que
permiten el paso de moléculas con un diámetro cercano a los ~1 nm, más grande
que un átomo de Ca2+ hidratado.
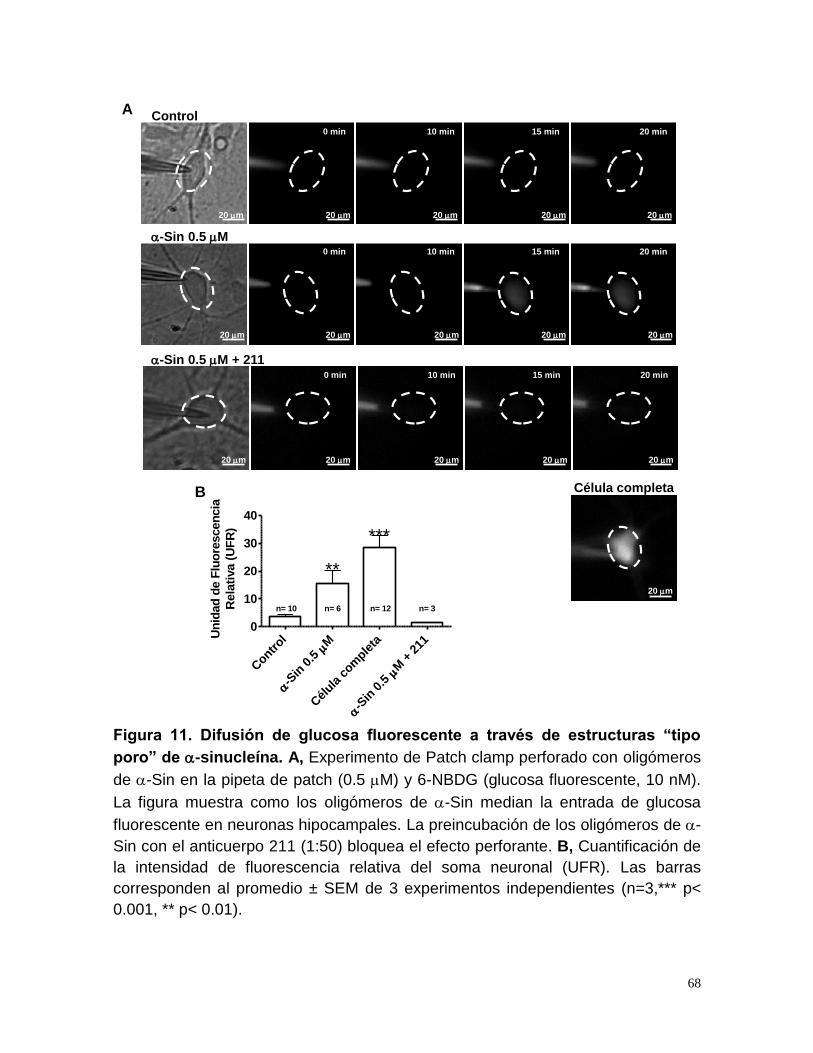
68
Figura 11. Difusión de glucosa fluorescente a través de estructuras “tipo
poro” de -sinucleína. A, Experimento de Patch clamp perforado con oligómeros
de -Sin en la pipeta de patch (0.5 M) y 6-NBDG (glucosa fluorescente, 10 nM).
La figura muestra como los oligómeros de -Sin median la entrada de glucosa
fluorescente en neuronas hipocampales. La preincubación de los oligómeros de -
Sin con el anticuerpo 211 (1:50) bloquea el efecto perforante. B, Cuantificación de
la intensidad de fluorescencia relativa del soma neuronal (UFR). Las barras
corresponden al promedio ± SEM de 3 experimentos independientes (n=3,*** p<
0.001, ** p< 0.01).
Contr
ol M
-Sin
0.5
Cél
ula c
omple
ta
M +
211
-Sin
0.5
0
10
20
30
40
n= 10 n= 6 n= 12 n= 3
**
***
Un
idad
de F
luo
rescen
cia
Rela
tiva (
UF
R)
0 10 20 30 minutes
-Sin 0.5 M
Control
-Sin 0.5 M + 211
Célula completa
A
B
20 m 20 m 20 m 20 m 20 m
20 m 20 m 20 m 20 m 20 m
20 m 20 m 20 m 20 m 20 m
20 m
0 min 10 min 15 min 20 min
0 min 10 min 15 min 20 min
0 min 10 min 15 min 20 min

69
Recientemente se ha descrito que proteínas con tallo GPI como, por
ejemplo, el prión se encuentran asociadas al proceso de unión del péptido A y
por, ende, también implicadas en la formación de estructuras tipo poro (datos no
publicados del laboratorio). Al realizar pretratamientos con fosfolipasa C en
neuronas hipocampales los resultados indicaron que existe una pérdida de la
capacidad perforante de los agregados de -Sin 1 M (Figura 12). En
consecuencia, estos datos sugieren que en el proceso de perforación por parte de
-Sin podría estar implicada alguna proteína de membrana plasmática con tallo
GPI.
Figura 12. Efecto de fosfolipasa C en la capacidad perforante de oligómeros
de -sinucleína en neuronas hipocampales. A, Trazos representativos de la
corriente capacitativa de membrana luego de la aplicación de oligómeros de -Sin
(1 µM). El pretratamiento de los cultivos con FLC (0.2 ug/mL) causa el bloqueo del
efecto perforante de los oligómeros de -Sin. B, Cuantificación de la carga de
membrana transferida (veces sobre el control). Las barras corresponden al
promedio ± SEM de 3 experimentos independientes (*** p< 0.001).
50403020100
Time (ms) Sw eep:1 Visible:1 of 21
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 12
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 22
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
-Sin 1 M
Control FLC
-Sin 1 M + FLC
50 p
A
10 ms
0 min
0 min
0 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 15
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 32
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
50403020100
Time (ms) Sw eep:1 Visible:1 of 12
IN 0
(pA
)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 43
15 min
15min
15 min
Contr
ol FLC M
-Sin
1
M
+ F
LC
-Sin
1
0
2
4
6***
n= 3 n= 6 n= 5Carg
a d
e m
em
bra
na
tran
sfe
rid
a
(veces s
ob
re e
l co
ntr
ol)
A B

70
Oligómeros de -sinucleína no forman “estructuras tipo poro” en la línea
celular RCSN-3
Dado que la EP es una condición neurodegenerativa que afecta
inicialmente a la sustancia nigra, estudiamos si -Sin alteraba la permeabilidad de
membrana de células RCSN-3, línea celular derivada de sustancia nigra de rata
(Paris et al., 2008). Éstas células se caracterizan por crecer en monocapa y no
requerir diferenciación para expresar rasgos catecolaminérgicos, tales como (i)
tirosina hidroxilasa, (ii) dopa decarboxilasa, (iii) dopamina hidroxilasa, (iv)
liberación dopamina, (v) transporte de dopamina, (vi) transporte de norepinefrina,
(vii) expresión de monoaminooxidasa (MAO)-A, (viii) formación de neuromelanina
y (ix) expresión del transportador vesicular de monoaminas 2 (VMAT-2). Además,
esta línea celular expresa transportadores de serotonina, transportadores de
metales divalentes (DMT1), receptores de dopamina D1 pero no D2. Todo lo
anterior apoya la idea que las células RCSN-3 son un buen modelo de estudio in
vitro, ya que bajo condiciones de experimentación expresa todos los marcadores
presentes en neuronas dopaminérgicas del sistema nervioso central.
Como resultado más importante, encontramos que utilizando una
concentración de -Sin (2.5 M) mayor que la empleada en neuronas
hipocampales, no se logró incrementar la corriente capacitativa de membrana
luego de 30 minutos de incubación (Figura 13).

71
Figura 13. Efecto de los oligómeros de -sinucleína en la corriente
capacitativa de membrana en células RCSN-3. A, Trazos representativos de la
corriente capacitativa de membrana de células RCSN-3 luego de la aplicación en
la pipeta de patch de oligómeros de -Sin (2.5 M). B, Cuantificación de los trazos
obtenidos en “A” (veces sobre el control). Las barras corresponden al promedio ±
SEM obtenido de al menos 3 experimentos independientes (*** p< 0.001).
30 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 56
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 3 4
0 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 21
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1 2 3 4
30 min
41704160415041404130
Time (ms)
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
0 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 22
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 3 4
50403020100
Time (ms) Sw eep:1 Visible:1 of 44
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1 2 3 4
-Sin 2.5 M
Control
Célula completa
50 p
A
10 ms
Contr
ol M
-Sin
2.5
Cél
ula c
omple
ta0
5
10
15
20 ***
n= 13 n= 6 n= 11
Carg
a d
e m
em
bra
na
tran
sfe
rid
a
(veces s
ob
re e
l co
ntr
ol)
A B

72
Es conocido que el estrés oxidativo altera la integridad de membranas
lipídicas (Barnham et al. 2004; Axelsen et al. 2011), por lo tanto, se promovió en
las células RCSN-3 la formación de estructuras tipo poro/perforado de -Sin
utilizando rotenona como un sensibilizador de membrana. Rotenona es un
pesticida inhibidor del Complejo I mitocondrial, el que se ha visto promoviendo
estados de estrés oxidativo e induciendo parkinsonismo en mamíferos expuestos
a éste compuesto (Cannon et al. 2009). Para determinar la concentración óptima
de rotenona, se realizaron tratamientos por 24 horas a concentraciones entre 10
nM y 10 M. Con 10 nM se observó inocuidad y una actividad de reductora
(indicador de células vivas) similar a la del control. Hubo una disminución a un 84
± 10 % de la actividad a una concentración de 100 nM. Las siguientes dos
concentraciones evaluadas, 1 y 10 M, provocaron una disminución de la actividad
a un 75 ± 6 % y 65 ± 4 %, respectivamente (Figura 14B). Así, se determinó que 50
nM sería la concentración óptima de rotenona a utilizar en los tratamientos
posteriores. Las incubaciones fueron realizadas 24 horas previas al ensayo
electrofisiológico. El resultado fue similar a lo observado anteriormente, y contrario
a lo esperado, no se observó un incremento significativo en el valor de carga
transferida a través de la membrana celular (Figura 14D).

73
Figura 14. Efecto de rotenona sobre las células RCSN-3. A, Imágenes en
campo claro de células RCSN-3 tratadas con concentraciones crecientes de
rotenona (0.01-10 M) por 24 horas. B, Cuantificación de la viabilidad celular de
los cultivos mostrados en “A” a través de la técnica de Alamar Blue. C, El
pretratamiento de las células RCSN-3 con rotenona 50 nM por 24 horas a 37ºC no
causó un incremento en la corriente capacitiva luego de la aplicación de
oligómeros de -Sin (2.5 M). D, Cuantificación de los trazos obtenidos en “C”
(veces sobre el control). Las barras corresponden al promedio ± SEM obtenido de
3 experimentos independientes (n=3, *** p< 0.001, ** p< 0.01).
A B
30 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 55
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 3 4
0 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 54
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 3 4
0 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 13
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1 2 3 4
30 min
50403020100
Time (ms) Sw eep:1 Visible:1 of 58
-1200
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1200
1 2 3 4
50403020100
Time (ms) Sw eep:1 Visible:1 of 44
-1000
-800
-600
-400
-200
0
200
400
600
800
1000
1 2 3 4
Rotenona 50 nM
Célula completa
-Sin 2.5 M + Rotenona 50 nM
50 p
A
10 ms
Rot 5
0 nM
M +
Rot 5
0 nM
-Sin
2.5
Cél
ula c
omple
ta0
5
10
15
20 ***
n= 13 n= 10 n= 11
Carg
a d
e m
em
bra
na
tran
sfe
rid
a
(veces s
ob
re e
l co
ntr
ol)
C D
Contr
ol
DM
SO
Rot 1
0 nM
Rot 1
00 n
M M
Rot 1
M
Rot 1
0
0
50
100
150
*****
Inte
nsid
ad
de
Flu
ore
scen
cia
(% c
on
tro
l)
Control
Rot 1 M
Rot 10 nM
Rot 10 M
Rot 100 nM
DMSO
30 m
30 m
30 m
30 m
30 m
30 m

74
Cabe destacar que se realizaron de manera paralela, ensayos de viabilidad
en neuronas hipocampales, donde a concentraciones de 100 nM, se observó una
reducción de la viabilidad celular a un 84 ± 2 %, lo que se incrementó a
concentraciones micromolares llegando a un 95 ± 1% en presencia de rotenona 10
M (Figura 15).
Figura 15. Viabilidad de neuronas hipocampales luego del tratamiento con
rotenona. A, Imágenes en campo claro de neuronas hipocampales tratadas con
concentraciones crecientes de rotenona (0.01-10 M) por 24 horas. B,
Cuantificación de la viabilidad celular de los cultivos mostrados en “A” a través de
la técnica de Alamar Blue. Las barras corresponden al promedio ± SEM obtenido
de 3 experimentos independientes (n=3, *** p< 0.001).
Contr
ol
DM
SO
Rot 1
0 nM
Rot 1
00 n
M M
Rot 1
M
Rot 1
0
0
50
100
150
*** ******
Inte
nsid
ad
de
Flu
ore
scen
cia
(%
co
ntr
ol)
A B
Control
Rot 1M
Rot 10 nM
DMSO
Rot 100 nM
Rot 10 M
30 m
30 m
30 m
30 m
30 m
30 m

75
Por consiguiente, para investigar si la ausencia de cambios en la
permeabilidad de membrana dada por los agregados de -Sin en la línea celular
RCSN-3 era producto, probablemente, de su naturaleza dopaminérgica o de su
naturaleza clonal, utilizamos neuronas dopaminérgicas de ratón disociadas de la
sustancia nigra a P20. Los resultados muestran que la aplicación de agregados de
-Sin (0.5 M) sobre este tipo de neuronas de la sustancia nigra incrementó la
corriente capacitativa 2 veces sobre el control luego de ~7 minutos de incubación
(Figura 16). En consecuencia, estos datos sugieren que la formación de
“estructuras tipo poro” por parte de -Sin se encuentran favorecidas en células
primarias con naturaleza neuronal.
Figura 16. Efecto de los oligómeros de -sinucleína en la corriente
capacitativa de membrana en neuronas disociadas dopaminérgicas de SN a
P20. A, Trazos representativos de la corriente capacitativa de membrana luego de
la aplicación de oligómeros de -Sin (0.5 M). B, Cuantificación de la carga de
membrana transferida (veces sobre el control). Las barras corresponden al
promedio ± SEM obtenido de 3 experimentos independientes (* p< 0.05, ** p<
0.01).
B
Contr
ol M
-Sin
0.5
Cél
ula c
omple
ta0
2
4
6
8
10
*
**
n= 6 n= 7 n= 3
Carg
a d
e m
em
bra
na
tran
sfe
rid
a
(veces s
ob
re e
l co
ntr
ol)
A
-Sin 0.5 M
Control
Célula completa
0 min 7 min
0 min 7 min
2 ms
50 p
A
2520151050
Time (ms) Sw eep:1 Visible:1 of 30
IN 0
(pA
)
-150
-100
-50
0
50
100
150
21
252015105
Time (ms) Sw eep:1 Visible:1 of 10
IN 0
(pA
)
-150
-100
-50
0
50
100
21
252015105
Time (ms) Sw eep:1 Visible:1 of 32
IN 0
(pA
)
-150
-100
-50
0
50
100
150
21
2520151050
Time (ms) Sw eep:1 Visible:1 of 15
IN 0
(pA
)
-150
-100
-50
0
50
100
150
21
2520151050
Time (ms) Sw eep:1 Visible:1 of 364
IN 0
(pA
)
-150
-100
-50
0
50
100
150
21

76
Objetivo 1.3. Caracterizar la asociación de -sinucleína en neuronas y
células RCSN-3
Los eventos de unión entre las proteínas amiloidogénicas y la membrana
plasmática resultan en perturbaciones estructurales mutuas que favorecen ó
inhiben el proceso formación de perforados (Figura 4; Butterfield et al. 2010). Por
ende, la sensibilidad o resistencia de las neuronas hipocampales y las células
RCSN-3 a la formación de “estructuras tipo poro” de -Sin, respectivamente,
podría deberse a diferencias en la asociación a la membrana plasmática.
Para responder a ésta interrogante, en el presente objetivo examinamos por
inmunofluorescencia indirecta la asociación de oligómeros de -Sin (0.5 M) a
neuronas hipocampales y células RCSN-3 en experimentos paralelos por 1 hora a
37ºC.

77
Resultados
La asociación de oligómeros de -sinucleína a la membrana plasmática es
célula-dependiente
Como se muestra en la Figura 17, la superposición de secciones ópticas
secuenciales (0.4 m) de las inmunorreactividades asociadas a MAP2 y a -Sin,
nos indican una asociación puntiforme de los oligómeros de -Sin en neuronas
hipocampales, presentes tanto en el soma neuronal (Figura 17D) como en los
procesos primarios (Figura 17E-G). Con respecto a la localización de los
oligómeros de -Sin, 22 secciones ópticas secuenciales (0.4 m) desde la zona
basal hasta la zona apical del soma neuronal (Figura 17D) nos sugieren que en
neuronas hipocampales los oligómeros de -Sin se asocian principalmente a nivel
de la superficie celular (Figura 17H), dado que la inmunorreactividad asociada a -
Sin se encuentra por fuera de los límites de la inmunorreactividad asociada a
MAP2 (secciones ópticas del 7-18), sin observarse una localización significativa a
nivel glial, dado que al realizar tinción inmunoespecífica para la proteína ácida
fibrilar glial (GFAP) y -Sin se observa que sólo un 7.7 ± 1.8 % (n=9) del total de
los agregados de -Sin se encuentra asociado a células GFAP positivas (Figura
18).

78
Figura 17. Asociación de oligómeros de -sinucleína a neuronas
hipocampales. A, Superposición de secciones ópticas secuenciales (0.4 m) de
la inmunorreactividad asociada a MAP2 (Cy3, Rojo). B, Superposición de
secciones ópticas secuenciales (0.4 m) de la inmunorreactividad asociada a
oligómeros de -Sin (0.5 M; 1 hora de incubación, FITC, Verde). C,
Superposición de secciones ópticas secuenciales (0.4 m) de las
inmunorreactividades asociadas a MAP2 y a -Sin, las flechas indican las zonas
amplificadas en D-G. D, Distribución somática neuronal de los oligómeros de -
Sin. E-G, Distribución a nivel de los procesos neuronales de oligómeros de -Sin.
H, Secciones ópticas secuenciales (0.4 m) de las inmunorreactividades
asociadas a MAP2 y a -Sin, desde la región basal hasta la zona apical del soma
neuronal indicado en “D”. Las imágenes confocales representan al menos 9
imágenes obtenidas desde 3 experimentos independientes (n=3).
A1
20 m
AMAP2-Cy3
20 m
-Sin-FITC B
C D
E
F
G
D
E
F
G
10 m
10 m
H
5 m
20 m
5 m
5 m
1 6
7 12
13 18
19 22

79
Figura 18. Distribución de oligómeros de -sinucleína en células gliales. A-E,
Imágenes confocales obtenidas luego del tratamiento con oligómeros de -Sin (0.5
M) por 1 hora a 37ºC. A, Inmunorreactividad asociada a GFAP (Cy3, Rojo). B,
Inmunorreactividad asociada a -Sin (FITC, Verde). C, Superposición de
inmunorreactividades asociadas a GFAP y a -Sin, las flechas indican las zonas
amplificadas en D-E. D, Distribución citoplasmática de los oligómeros de -Sin. E,
Distribución a nivel de procesos gliales de los oligómeros de -Sin. F, Número de
oligómeros de -Sin presentes a nivel de células GFAP positivas. Las imágenes
confocales representan al menos 9 imágenes obtenidas desde 3 experimentos
independientes (n=3).
A
20 m
B
E
D
20 m
20 m
4 m
GFAP-Cy3
-Sin-FITC
C D
E
5 m
Total
GFA
P posi
tivas
0
20
40
60
80
100
120
***
nº
de p
un
tas d
e
-Sin
(% t
ota
l)
F

80
En el caso de las células RCSN-3, la superposición de secciones ópticas
secuenciales (0.4 m) de la fluorescencia asociada a los microfilamentos de actina
y de la inmunorreactividad asociada a -Sin nos indican la presencia de una
población heterogénea de pequeños y grandes agregados de -Sin (Figura 19C-
E). Por otro lado, con respecto a la localización celular, éstos agregados se
ubicarían tanto a nivel intra como extracelular (Figura 19F) dado, principalmente,
por la existencia de inmunorreactividad asociada a -Sin dentro de los límites de la
fluorescencia asociada a los microfilamentos de actina, como también de su
presencia en las 19 secciones ópticas secuenciales (0.4 m) obtenidas desde la
zona basal hasta la zona apical de la célula RCSN-3 indicada (Figura 19D).

81
Figura 19. Asociación de oligómeros de -sinucleína a células RCSN-3. A,
Superposición de secciones ópticas secuenciales (0.4 m) de la tinción
fluorescente de los microfilamentos de actina (Faloidina, Verde). B, Superposición
de secciones ópticas secuenciales (0.4 m) de la inmunorreactividad asociada a
-Sin (0.5 M; 1 hora de incubación, Cy3, Rojo). C, Superposición de secciones
ópticas secuenciales (0.4 m) de la fluorescencia asociada a los microfilamentos
de actina y a la inmunorreactividad asociada a -Sin, las flechas indican las zonas
amplificadas en D-E. D-E, Distribución citoplasmática de los agregados de -Sin.
F, Secciones ópticas secuenciales (0.4 m) de la fluorescencia asociada a los
microfilamentos de actina y a la inmunorreactividad asociada a -Sin desde la
región basal hasta la zona apical de la célula indicada en “D”. Las imágenes
confocales representan al menos 9 imágenes obtenidas desde 3 experimentos
independientes (n=3).
10 µM
2 m
20 m
AFaloidina
20 m
-Sin-Cy3 B
C D
E
D
E
10 m
10 µM
10 µM
20 m
H
10 m
F1 6
7 12
13 18
19

82
Análisis cuantitativo de los rangos de tamaños de los agregados de -Sin
asociados a las células RCSN-3 nos señalan valores entre 0.01 - 2.6 µm2 (similar a
lo obtenido en neuronas hipocampales) y 10 - 140 µm2 (Figura 20). Es importante
destacar que la presencia de grandes agregados de -Sin en células RCSN-3
representa a sólo el 2% del total de los agregados (Figura 20B), sin embargo, se
presenta en casi todas las muestras a 1 hora de incubación.
Figura 20. Cuantificación de la asociación de oligómeros de -sinucleína
recombinante humana en neuronas hipocampales y células RCSN-3. Los
resultados obtenidos corresponden al tratamiento paralelo de neuronas
hipocampales y células RCSN-3 con oligómeros de -Sin (0.5 M) por 1 hora a
37ºC. A, Frecuencia de distribución del tamaño de los agregados de -Sin
presente en neuronas hipocampales (n= 588). B, Frecuencia de distribución del
tamaño de los agregados de -Sin presentes en células RCSN-3 (n=478). El
inserto en el gráfico muestra la frecuencia de distribución entre los tamaños < 5
m2 (n=459). Las barras corresponden a 3 experimentos independientes.
0 20 40 60 80 100 120 1400
2
4
6
8
Tamaño (m2)
Fre
cu
en
cia
de d
istr
ibu
ció
n
(nº
de p
un
tas d
e
-Sin
)
A
B
0.0 0.5 1.0 1.5 2.0 2.5 3.00
10
20
300
400
500
Tamaño (m2)
Fre
cu
en
cia
de d
istr
ibu
ció
n
(nº
de p
un
tas d
e
-Sin
)
0 1 2 3 4 50
20
40
60
200
300
400
Tamaño (m2)
Fre
cu
en
cia
de d
istr
ibu
ció
n
( n
º d
e p
un
tas d
e
-Sin
)

83
Por otro lado, a pesar de que en la mayoría de las neuronas hipocampales
se presentan agregados pequeños de -Sin del tipo puntiforme, inusualmente, se
observan grandes agregados asociados a núcleos apoptóticos, dado que al
realizar tinción fluorescente nuclear con DAPI se observa la fragmentación del
ADN que colocaliza con la inmunorreactividad asociada con -Sin (Figura 21).
Figura 21. Neuronas hipocampales presentan grandes agregados de -
sinucleína asociados a núcleos apoptóticos. A, Superposición de la
inmunorreactividad asociada a MAP2 (Cy3, Rojo), a -Sin (FITC, Verde) y tinción
fluorescente nuclear (DAPI, Azul) en neuronas hipocampales luego del tratamiento
con oligómeros de -Sin (0.5 M) por 1 hora a 37ºC. B, Inmunorreactividad
asociada a MAP2 (Cy3, Rojo). C, Tinción fluorescente nuclear (DAPI, Azul). D,
Inmunorreactividad asociada a -Sin (FITC, Verde). Los recuadros indican las
zonas amplificadas en A1, B1, C1 y D1. Las imágenes confocales representan al
menos 9 imágenes obtenidas desde 3 experimentos independientes (n=3).
DAPI
10 m
C1
-Sin-FITC
10 m
D1
-Sin-FITC
MAP2-Cy3
DAPI
10 m
A11
MAP2-Cy3
10 m
B11
A1
B1
C1
D1
4 m
4 m
4 m
4 m
A B
C D

84
Con el fin de obtener una mejor visualización de la unión diferencial de
oligómeros de -Sin a neuronas hipocampales y células RCSN-3, se realizaron
reconstrucciones y proyecciones en 3D con el programa Imaris (Bitplane, USA) de
las distintas secciones ópticas secuenciales (0.4 m) adquiridas en las imágenes
confocales previamente señaladas (Figura 17, Figura 19). Los resultados
reafirman una asociación puntiforme extracelular para los oligómeros de -Sin en
neuronas hipocampales (Figura 22) y una asociación intra y extracelular de
pequeños y grandes agregados de -Sin en las células RCSN-3 (Figura 23).
Figura 22. Proyección en 3D de oligómeros de -sinucleína en neuronas
hipocampales. Reconstitución y proyección en 3D con el programa Imaris de
secciones ópticas secuenciales (0.4 m) de las inmunorreactividades asociadas a
MAP2 (Rojo) y a -Sin (Verde) de una neurona hipocampal tratada con oligómeros
de -Sin (0.5 M) por 1 hora a 37ºC. La reconstrucción de imagen representa al
menos 9 imágenes obtenidas desde 3 experimentos independientes (n=3).
3 µm
4 µm
5 m
10 m 10 m
10 m
MAP2-Cy3/ -Sin-FITC

85
Figura 23. Proyección en 3D de oligómeros de -sinucleína en células RCSN-
3. Reconstitución y proyección en 3D con el programa Imaris de secciones ópticas
secuenciales (0.4 m) de la fluorescencia asociada a los microfilamentos de actina
(Verde) y a la inmunorreactividad asociada a -Sin (Rojo) de una célula RCSN-3
tratada con oligómeros de -Sin (0.5 M) por 1 hora a 37ºC. La reconstrucción de
imagen representa al menos 9 imágenes obtenidas desde 3 experimentos
independientes (n=3).
Cabe destacar que al realizar inmunocitoquímica indirecta utilizando los
mismos anticuerpos primarios y secundarios para -Sin y MAP2 en ambos tipos
celulares, observamos el mismo efecto mencionado anteriormente (Figura 24).
Interesantemente, las células RCSN-3 también presentaron inmunomarcaje para
MAP2. MAP2 es una proteína estabilizadora de microtúbulos cuya expresión se
presenta sólo en neuronas. Esto nos señala que las células RCSN-3 a pesar de
ser una línea celular clonal mantienen en parte su fenotipo neuronal.
20 m
5 m 10 m
5 m
Faloidina/ -Sin-Cy3

86
Figura 24. Asociación de oligómeros de -sinucleína a neuronas
hipocampales y células RCSN-3. A, Inmunorreactividad asociada a MAP2 (Cy3,
Rojo), a -Sin (0.5 M; 1 hora de incubación, FITC, Verde) y tinción fluorescente
nuclear (DAPI, Azul) en neuronas hipocampales. Los recuadros indican las zonas
amplificadas en A1. B, Inmunorreactividad asociada a MAP2 (Cy3, Rojo) y -Sin
(0.5 M; 1 hora de incubación, FITC, Verde) y tinción fluorescente nuclear (DAPI,
Azul) en células RCSN-3. Los recuadros indican las zonas amplificadas en B1. Las
imágenes confocales representan al menos 9 imágenes obtenidas desde 3
experimentos independientes (n=3).
MAP2-Cy3/ -Sin-FITC
Neuronas hipocampales
MAP2-Cy3/ -Sin-FITCA B
9 m 9 m
20 m
Células RCSN-3
A1
20 m
B1
A1
B1

87
Dadas las diferencias de asociación entre ambos tipos celulares, evaluamos
qué determinantes moleculares estarían involucrados en la asociación de -Sin
(0.5 M) por 1 hora a 37ºC. Con fin de remover el componente proteico y
colesterol se realizaron pretratamientos con tripsina 0.00025% y -MCD 2 mM,
respectivamente, tanto en neuronas hipocampales, como en células RCSN-3.
Neuronas hipocampales tratadas con tripsina (Figura 25) señalaron una
disminución significativa del número total de agregados de -Sin a un 14 ± 8.0 %
en los procesos primarios (20 m) (Figura 26A) y a un 28 ± 5 % en el soma
neuronal con respecto a las neuronas control (sin pretratamiento) (Figura 26B) sin
observarse cambios significativos en el tamaño de los agregados (Figura 26C). Sin
embargo, neuronas hipocampales tratadas con -MCD no generaron cambios
significativos ni en el número total ni en el tamaño de los agregados de -Sin
(Figura 25-26).

88
Figura 25. Efecto de tripsina y -MCD sobre la asociación de oligómeros de
-sinucleína a neuronas hipocampales. A, Inmunorreactividad asociada a
MAP2 (Cy3, Rojo), a -Sin (FITC, Verde) y tinción fluorescente nuclear (DAPI,
Azul) en neuronas hipocampales luego del tratamiento con oligómeros de -Sin
(0.5 M) por 1 hora a 37ºC. Los recuadros indican las zonas amplificadas en A1-
A2. B, Imágenes obtenidas luego del pretratamiento con tripsina (0.00025%) por
30 minutos a 37ºC y posterior tratamiento con oligómeros de -Sin (0.5 M) por 1
hora a 37ºC. Los recuadros indican las zonas amplificadas en B1-B2. C, Imágenes
obtenidas luego del pretratamiento con -MCD (2 mM) por 30 minutos a 37ºC y
posterior tratamiento con oligómeros de -Sin (0.5 M) por 1 hora a 37ºC. Los
recuadros indican las zonas amplificadas en C1-C2. Las imágenes confocales
representan al menos 9 imágenes obtenidas desde 3 experimentos
independientes (n=3).
A1
A2
B1
B2
C1
C2
10 m
3 m
3 m
10 m
3 m
3 m
10 m
3 m
3 m
A1
B1
C1A2
C2
B2
-Sin 0.5 M Tripsina 0.00025%
+ -Sin 0.5 M
-MCD 2 mM
+ -Sin 0.5 M
-Sin-FITC
MAP2-Cy3
DAPI
-Sin-FITC
MAP2-Cy3
DAPI
-Sin-FITC
MAP2-Cy3
DAPI
A B C

89
Figura 26. Cuantificación de la asociación de oligómeros de -sinucleína en
neuronas hipocampales luego del tratamiento con tripsina y -MCD. Los
resultados obtenidos corresponden al pretratamiento de neuronas hipocampales
con tripsina (0.00025%) por 30 minutos a 37ºC, con -MCD (2 mM) por 30 minutos
a 37ºC y posterior tratamiento con oligómeros de -Sin (0.5 M) por 1 hora a
37ºC. A, Número de agregados de -Sin presentes en los procesos primarios de
neuronas hipocampales (20 m). B, Número de agregados de -Sin presentes en
el soma neuronal. C, Tamaño de los agregados de -Sin asociados a neuronas
hipocampales (control= células sin pre-tratamiento). Las barras son el promedio ±
SEM obtenido a partir de 3 experimentos independientes (n=3, ** p< 0.01, *** p<
0.001).
-Sin
-S
in
Tripsi
na +
-Sin
-MCD +
0
30
60
90
120
***
Nº
de p
un
tas d
e
-Sin
/
so
ma (%
co
ntr
ol)
-Sin
-S
in
Tripsi
na +
-Sin
-MCD +
0
50
100
150
**
nº
de p
un
tas d
e
-Sin
/
20
m
(%
co
ntr
ol)
A B
C
-Sin
-S
in
Tripsi
na +
-Sin
-MCD +
0
20
40
60
80
100
120
Tam
añ
o d
e p
un
tas
de
-S
in (
% c
on
tro
l)

90
A diferencia de lo obtenido en neuronas hipocampales, en células RCSN-3
no se observó una diferencia significativa entre los tratamientos y el control (Figura
27). El número de agregados totales disminuyó, aunque de modo no significativo,
sólo en presencia de -MCD a un 84 ± 10% respecto del control (células sin
pretratamiento) (Figura 28A). El tamaño tampoco disminuyó de manera
significativa respecto del control, con valores de un 80 ± 7 % para células tratadas
con tripsina y de un 82 ± 9% para células tratadas con -MCD (Figura 28B). No
obstante, al cuantificar los agregados de gran tamaño, considerados aquellos con
área mayor o igual a 1.0 m2, el número disminuye a un 47 ± 11% con el pre
tratamiento con tripsina y a un 24 ± 10% con -MCD (Figura 28C).
En conjunto, estos resultados sugieren que la membrana plasmática de
neuronas hipocampales y células RCSN-3, es un factor crítico y diferencial en la
asociación de oligómeros de -Sin.

91
Figura 27. Efecto de tripsina y -MCD sobre la asociación de oligómeros de
-sinucleína en células RCSN-3. A, Inmunorreactividad asociada a -Sin (Cy3,
Rojo), tinción fluorescente de los microfilamentos de actina (Faloidina, Verde) y
nuclear (DAPI, azul) en células RCSN-3 luego del tratamiento con oligómeros de
-Sin (0.5 M) por 1 hora a 37ºC. B, Imágenes obtenidas luego del pretratamiento
con tripsina (0.00025%) por 30 minutos a 37ºC y posterior tratamiento con
oligómeros de -Sin (0.5 M) por 1 hora a 37ºC. C, Imágenes obtenidas luego del
pretratamiento con -MCD (2 mM) por 30 minutos a 37ºC y posterior tratamiento
con oligómeros de -Sin (0.5 M) por 1 hora a 37ºC. Los recuadros indican las
zonas amplificadas en A1, B1 y C1. Las imágenes confocales representan al
menos 9 imágenes obtenidas desde 3 experimentos independientes (n=3).
-Sin 0.5 µM Tripsina 0.00025%
+ -Sin 0.5 M
-MCD 2 mM
+ -Sin 0.5 M
Faloidina
-Sin- Cy3
DAPI
A B C
A1 B1 C1
A1
B1
C1
10 m 10 m 10 m
20 m 20 m 20 m

92
Figura 28. Cuantificación de la asociación de oligómeros de -sinucleína en
células RCSN-3 luego del tratamiento con tripsina y -MCD. Los resultados
obtenidos corresponden al pretratamiento de células RCSN-3 con tripsina
(0.00025%) por 30 minutos a 37ºC, con -MCD (2 mM) por 30 minutos a 37ºC y
posterior tratamiento con oligómeros de -Sin (0.5 M) por 1 hora a 37ºC. A,
Número de agregados de -Sin asociados a células RCSN-3. B, Tamaño de los
agregados de -Sin asociados a células RCSN-3. C, Número de agregados de -
Sin cuyo tamaño fue mayor o igual a 1 m2. Las barras son el promedio ± SEM
obtenido a partir de 3 experimentos independientes (n=3, * p< 0.05, ** p< 0.01).
A B
C
-Sin
-S
in
Tripsi
na +
-Sin
-MCD +
0
20
40
60
80
100
120
Tam
añ
o d
e p
un
tas
de
-S
in (
% c
on
tro
l)
-Sin
-S
in
Tripsi
na +
-Sin
-MCD +
0
20
40
60
80
100
120
Nº
de p
un
tas d
e
-Sin
(% c
on
tro
l)
-Sin
-S
in
Tripsi
na +
-Sin
-MCD +
0
20
40
60
80
100
120
140
***
1m2
nº
de p
un
tas d
e
-Sin
(% c
on
tro
l)

93
La formación de grandes agregados de -sinucleína en células RCSN-3 es
dependiente de la temperatura
Considerando que agregados de -Sin forman grandes estructuras a nivel
intracelular en las células RCSN-3, quisimos evaluar si la formación de éstos se
veía alterada por incubación a 4ºC, donde uno de los posibles mecanismos
afectados sea la endocitosis (Pastan et al. 1981). Para ello, se realizaron
incubaciones por 1 hora a esta temperatura, obteniendo en este caso una señal
puntiforme para la inmunorreactividad asociada a -Sin que se distribuyó de
manera homogénea en la célula, con una disminución significativa del número de
agregados a un 39 ± 16% con respecto al control (37ºC) (Figura 29).
Figura 29. Incubación por 1 hora a 4°C inhibe la aparición de agregados de
gran tamaño de -sinucleína en células RCSN-3. A, Inmunorreactividad
asociada a -Sin (Cy3, Rojo), tinción fluorescente de los microfilamentos de actina
(Faloidina, Verde) y nuclear (DAPI, azul) en células RCSN-3 luego del tratamiento
con oligómeros de -Sin (0.5 M) por 1 hora a 4ºC. El recuadro indica la zona
amplificada en A1. B, Número de oligómeros de -Sin asociados a células RCSN-
3 a 37ºC (control) y 4ºC. Las barras corresponden al promedio ± SEM obtenido a
partir de 3 experimentos independientes (n=3, * p< 0.05).
-Sin
37
ºC
-S
in 4
ºC
0
20
40
60
80
100
120
140
*
Nº
de p
un
tas d
e
-S
in (%
co
ntr
ol)
Faloidina
-Sin- Cy3
DAPI
A1
A1
3 m
10 m
A B

94
Análisis de frecuencia de distribución de los agregados por tamaños indican
que tanto agregados pequeños, medianos y grandes se encuentran afectados a
4ºC, especialmente, aquellos cuyo tamaño supera las 10 m2 (Figura 30).
Figura 30. Cuantificación de la asociación de oligómeros de -sinucleína
recombinante humana en células RCSN-3 a 4 y 37°C. Los resultados obtenidos
corresponden al tratamiento de células RCSN-3 con oligómeros de -Sin (0.5 M)
por 1 hora a 37ºC y a 4ºC. A, Frecuencia de distribución del tamaño de los
agregados de -Sin cuya área es menor a 1 m2 en células RCSN-3 (n= 7417). B,
Frecuencia de distribución del tamaño de los agregados de -Sin cuya área se
encuentra entre 1-10 m2 en células RCSN-3 (n= 207). C, Frecuencia de
distribución del tamaño de los agregados de -Sin cuya área es mayor a 10 m2
en células RCSN-3 (n= 9). Los tratamientos se realizaron a 37 y 4ºC. Las barras
fueron obtenidas a partir de 3 experimentos independientes.
0 2 4 6 8 100
20
40
60-Sin 37 ºC
-Sin 4 ºC
Tamaño (m2)
Fre
cu
en
cia
de
dis
trib
ució
n
(nº
pu
nta
s d
e
-Sin
)
0 20 40 60 80 1000
1
2
3
4-Sin 37 ºC
-Sin 4 ºC
Tamaño (m2)
Fre
cu
en
cia
de
dis
trib
ució
n
(nº
pu
nta
s d
e
-Sin
)
0.0 0.2 0.4 0.6 0.8 1.00
500
1000
1500-Sin 37 ºC
-Sin 4 ºC
Tamaño (m2)
Fre
cu
en
cia
de
dis
trib
ució
n
(nº
pu
nta
s d
e
-Sin
)
A
B
C

95
Para una mejor visualización, representaciones en 3D con el programa Zen
(Zeiss, Alemania) de las distintas secciones ópticas secuenciales (0.4 m)
adquiridas en la imagen confocal previamente señalada (Figura 29A), nos
confirman la presencia de pequeños agregados de -Sin en las células RCSN-3
(Figura 31).
En consecuencia, de estos resultados se infiere que la formación de los
agregados de mayor envergadura de -Sin se encuentran inhibidos a 4°C y, por lo
tanto, son dependientes de la temperatura para su generación.
Figura 31. Proyección en 3D de oligómeros de -sinucleína en células RCSN-
3 obtenidas a 4°C. Reconstitución y proyección en 3D con el programa Zen
(Zeiss) de secciones ópticas secuenciales (0.4 m) de la fluorescencia asociada a
los microfilamentos de actina (Verde) y a la inmunorreactividad asociada a -Sin
(Rojo) de una célula RCSN-3 tratada con oligómeros de -Sin (0.5 M) por 1 hora
a 37ºC. La reconstrucción de imagen representa al menos 9 imágenes obtenidas
desde 3 experimentos independientes (n=3).
Faloidina / -Sin-Cy3
37ºC 4ºC
10 m
10 m 10 m
10 m
10 m 10 m

96
Oligómeros de -sinucleína exógena inducen la agregación de -sinucleína
endógena en células RCSN-3
Actualmente, existe la hipótesis que propone que la iniciación y la
propagación de la EP es producto de la progresión (avance) de agregados de -
Sin a distintas zonas cerebrales, en una manera similar al prión (Aguzzi et al.
2006). Por lo que se determinó si oligómeros de -Sin exógena inducían la
agregación de -Sin endógena. Para ello se realizó inmunofluorescencia indirecta
utilizando anticuerpos específicos, donde para oligómeros de -Sin exógena (-
Sin recombinante humana) se utilizó el anticuerpo monoclonal 211 hecho en ratón
contra los aminoácidos 121-125 de -Sin de origen humano (Santa Cruz, USA) y
para -Sin endógena se utilizó el anticuerpo monoclonal D37A6 hecho en conejo
contra el aminoácido Glu105 de -Sin de origen de rata. Las células RCSN-3
expresan -Sin (endógena) (Paris et al. 2008), proteína ampliamente distribuida en
la célula, y que se presenta además en forma de pequeños agregados
perinucleares (Figura 32A). La incubación de células RCSN-3 en presencia de
oligómeros de -Sin exógena, modificó el tamaño de los agregados perinucleares
de -Sin endógena, incrementándolo, y co-localizando a su vez con la señal de -
Sin exógena con valores para el coeficiente de Manders, M1, que corresponde a la
razón de -Sin endógena que colocaliza con -Sin exógena, de 0.25 ± 0.01 y M2,
que corresponde a la razón de -Sin exógena que colocaliza con -Sin endógena,
de 0.9573 ± 0.03 (Figura 32B). Cortes ortogonales en el eje XY e ZY de la imagen
obtenida en la Figura 33A luego del oligómeros de -Sin exógena, nos reafirman

97
la capacidad de -Sin exógena para ingresar a la célula y promover la formación
de agregados de -Sin endógena en el citosol (Figura 33).
Figura 32. Oligómeros de -sinucleína exógena inducen la agregación de -
sinucleína endógena en células RCSN-3. A, Superposición de secciones ópticas
secuenciales (0.4 m) de la inmunorreactividad asociada a -Sin endógena (FITC,
Verde), inmunorreactividad asociada a -Sin exógena (Cy3, Rojo) y tinción nuclear
(DAPI, azul). B, Cuantificación del coeficiente de colocalización Mander entre la
señal inmunofluorescente para -Sin endógena (-Sin EN) y luego del tratamiento
con oligómeros de -Sin exógena (-Sin EX; 0.5 µM) por 1 hora sobre células
RCSN-3. C, Dot blot de lisados de células HEK-293 (de origen humano) y células
RCSN-3 (de origen de rata). Se utilizaron los anticuerpos 211 (Santa Cruz) que
reconoce -Sin humana y el anticuerpo D37A6 que reconoce -Sin endógena. Las
imágenes confocales representan al menos 9 imágenes obtenidas desde 3
experimentos independientes (n=3).
B C Célula
HEK-293
211
D37A6
Célula
RCSN-3
0.0
0.5
1.0
1.5
M1-Sin EN
-Sin EX
M2-Sin EX
-Sin EN
Co
eficie
nte
Man
der'
s
A Control
-Sin 0.5 M
-Sin
endógena-FITC
-Sin
exógena-Cy3
Merge + DAPI
-Sin
endógena-FITC
-Sin
exógena-Cy3
Merge + DAPI
10 m10 m10 m
10 m10 m10 m

98
Figura 33. -Sinucleína exógena y -sinucleína endógena forman agregados
mixtos en células RCSN-3. A, Superposición de secciones ópticas secuenciales
(0.4 m) de la inmunorreactividad asociada a -Sin endógena (FITC, Verde),
inmunorreactividad asociada a -Sin exógena (Cy3, Rojo) y tinción nuclear (DAPI,
azul). En el panel lateral derecho e inferior de la imagen central se muestran
cortes ortogonales de ella en el eje XY e ZY de la zona trazada por las líneas
blancas. La reconstrucción de imagen representa al menos 9 imágenes obtenidas
desde 3 experimentos independientes (n=3).
Y
Z
X
Y
-Sin endógena-FITC
-Sin exógena-Cy3
DAPI
10 m

99
Oligómeros de -sinucleína inducen la formación de agresomas en células
RCSN-3
La evidencia encontrada hasta ahora sugiere que las células RCSN-3
poseen un mecanismo de resistencia que podría estar mediado por la formación
de cuerpos de inclusión y/o agresomas. Cuando la producción de proteínas mal
plegadas excede la capacidad del sistema de replegamiento por chaperonas y la
vía de degradación ubiquitina-proteosoma, las proteínas mal plegadas son
activamente transportadas a una estructura citoplasmática yuxtanuclear llamada,
agresoma (Olzmann et al. 2011). La formación de agresomas es reconocida como
una respuesta citoprotectora que sirve para secuestrar proteínas mal plegadas
potencialmente tóxicas, involucrando el reconocimiento de la proteína, su
acoplamiento al motor de díneina, y su transporte retrógrado a través de los
microtúbulos hacia el centrosoma (Kopito 2000; Garcia-Mata et al. 2002). Para
determinar si los agregados de -Sin forman agresomas en las células RCSN-3 se
realizaron coincubaciones de agregados de -Sin (0.5 M) con colchicina,
inhibidor de la polimerización de microtúbulos, (10 g/L) ó EHNA, inhibidor del
motor de dineína, (50 g/L) por 1 hora a 37ºC. Los resultados obtenidos a través
de inmunofluorescencia indirecta señalaron una leve pero significativa disminución
con respecto al control (células sin co-tratamiento) en el tamaño de los agregados
de -Sin en presencia de colchinina y EHNA con valores de un 66 ± 7 % y de un
66 ± 9 %, respectivamente (Figura 34D). Al analizar el número de los agregados
de -Sin se genera una disminución de un 47 ± 18 % en presencia de colchinina,
sin obtenerse diferencias significativas en presencia de EHNA (Figura 34E).

100
Figura 34. Efecto de colchicina y EHNA sobre la formación de agregados de
-sinucleína en células RCSN-3. A, Imágenes obtenidas luego del tratamiento
con oligómeros de -sinucleína exógena (0.5 M) por 1 hora a 37ºC. B, Imágenes
obtenidas luego del co-tratamiento con oligómeros de -Sin exógena (0.5 M) y
colchicina (10 g/L) por 1 hora a 37ºC. C, Imágenes obtenidas luego del co-
tratamiento con oligómeros de -Sin exógena (0.5 M) y EHNA (50 g/L) por 1
hora a 37ºC. Las imágenes representan la inmunorreactividad asociada a -Sin
exógena (Dyl 649, Azul), a -Sin endógena (Cy3, Rojo) y a Lamina b (FITC,
Verde). D, Tamaño de los agregados de -Sin (por célula) luego del tratamiento
con colchicina y EHNA. E, Número de agregados de -Sin (por célula) luego del
tratamiento con colchicina y EHNA. Las barras corresponden al promedio ± SEM
obtenido de 3 experimentos independientes (n=3,* p< 0.05).
0
10
20
30
40
* *
-Sin -Sin +Colchicina
-Sin +EHNA
Tam
añ
o d
e p
un
tas
de
-S
in/c
élu
la (
m
2)
0
10
20
30
*
-Sin -Sin +Colchicina
-Sin +EHNA
Nº
de p
un
tas d
e
-S
in /célu
la
D E
EHNA + -Sin-Sin Colchicina + -Sin
A B C
A1 B1 C1
10 m10 m 10 m
100 m
B1
A1
C1
100 m100 m
-Sin exógena-Dyl 649/ -Sin endógena-Cy3/ Laminina b-FITC

101
Una característica general de los CL y los agresomas es la presencia de la
proteína ubiquitina. Para determinar si los agregados de -Sin son
inmunopositivos para ubiquitina, se realizaron ensayos de inmunocitoquímica
indirecta de células RCSN-3 tratadas con agregados de -Sin 0.5 M por 1 hora a
37ºC. Análisis de la señal de ubiquitina total señala una disminución en presencia
de los agregados de -Sin a un 76 ± 7 % con respecto al control (Figura 35B).
Interesantemente, el patrón de ubiquitina presenta zonas que colocalizan con -
Sin (color magenta, Figura 35A).
En conclusión, los datos obtenidos hasta el momento sugieren que
oligómeros de -Sin exógena inducen la formación de agresomas en las células
RCSN-3 y la agregación de -Sin endógena.

102
Figura 35. Determinación de la ubiquitinación de agregados de -sinucleína
en células RCSN-3. A, Imágenes obtenidas luego del tratamiento de células
RCSN-3 con oligómeros de -Sin (0.5 M) por 1 hora a 37ºC. Las imágenes
muestran la inmunorreactividad asociada a -Sin exógena (Dyl 649, Azul), a
ubiquitina (Cy3, Rojo) y tinción fluorescente de los microfilamentos de actina
(Faloidina, Verde). B, Cuantificación de la inmunorreactividad total asociada a
ubiquitina. Las imágenes confocales representan al menos 9 imágenes obtenidas
desde 3 experimentos independientes (n=3).
Contr
ol M
-Sin
0.5
0
50
100
150
*
Inte
nsid
ad
To
tal/ U
FR
(ub
iqu
itin
ació
n)
B
Faloidina
Ubiquitina-Cy5
-Sin-Cy3
Control -Sin 0.5 M
Faloidina
Ubiquitina-Cy5
-Sin-Cy3
10 m10 m
A

103
Objetivo Específico II. Caracterizar efectos funcionales de -sinucleína en
neuronas hipocampales
Diversos estudios indican que los efectos neurotóxicos de -Sin estarían
mediados por cambios en el Ca2+ intracelular (Danzer et al. 2007). Recientemente
se ha determinado que agregados de A (implicados en la EA) forman estructuras
tipo poro/perforado en las membranas neuronales (Sepulveda et al. 2010). Esta
acción induce un aumento agudo en la concentración de Ca2+ intracelular y una
disminución significativa de proteínas sinápticas en neuronas tratadas
crónicamente con agregados de A (Parodi et al. 2010).
Considerando que otras proteínas amiloidogénicas como el A se insertan
en la membrana plasmática formando perforaciones, en la presente sección se
describen los experimentos del objetivo específico de “Caracterizar efectos
funcionales de -sinucleína en neuronas hipocampales”. De este modo, se evaluó
si la formación de estructuras tipo poro/perforado de -Sin en neuronas
hipocampales promueve la entrada del Ca2+ el cual está implicado en múltiples
procesos como, por ejemplo, la transmisión sináptica y la neurodegeneración
(Mattson et al. 2007).

104
Objetivo 2.1 Determinar si agregados de -sinucleína alteran los niveles de
Ca2+ intracelular y la actividad sináptica neuronal
Debido a la importancia del Ca2+ a nivel celular y sobre todo en la liberación
de vesículas sinápticas, decidimos evaluar en el presente objetivo si oligómeros de
-Sin alteran la transmisión sináptica de neuronas hipocampales. Para ello,
utilizamos técnicas como: fluorimetría de calcio, patch clamp en la modalidad de
célula completa, fluorimetría con FM1-43 e inmunocitoquímica indirecta.
Resultados
Oligómeros de -sinucleína aumentan los niveles de Ca2+ intracelular en
neuronas hipocampales y células RCSN-3
Considerando los antecedentes anteriormente expuestos, estudiamos el
efecto de oligómeros de -Sin sobre los niveles totales de Ca2+ intracelular de
neuronas hipocampales y células RCSN-3, esta última como control negativo de
perforación. Las determinaciones de Ca2+ intracelular se realizaron utilizando la
sonda fluorescente Fluo4-AM específica para este catión (Kd= 345 nM). Para ello,
se seleccionaron ROIs localizados en el soma y se registró cada 20 segundos
(iluminadas por 200 milisegundos) durante 75 minutos. Luego que el registro se
mantuvo estable durante 10 minutos, con el fin de corroborar que el aumento de
Ca2+ no fuese espontáneo, los cultivos fueron perfundidos con oligómeros de -
Sin (0.5 M) o con la solución control (vehículo de -Sin). La aplicación de
oligómeros de -Sin en neuronas hipocampales indujo un aumento significativo en

105
los niveles totales de Ca2+ intracelular que comenzaron aproximadamente a los 20
minutos post adición de -Sin y que alcanzaron un valor de 1.7 veces con
respecto al valor control (vehículo) a los 75 minutos de registro (Figura 36A, 36C).
Al contrario de lo esperado, se observó un aumento en los niveles de Ca2+
intracelular en células RCSN-3 expuestas a oligómeros de -Sin, a partir de los 10
minutos post adición de -Sin y que alcanzó un valor de 1.3 veces con respecto al
valor control (vehículo) a los 75 minutos de registro (Figura 36B, 36D).
Con el fin de investigar si la entrada de Ca2+ extracelular estaba mediada
por la activación de canales de Ca2+ voltaje-dependientes o por la formación de
“estructuras tipo poro” de -Sin, se realizaron experimentos en presencia de Co2+
10 M, un inhibidor no específico de los canales de Ca2+ voltaje-dependientes
(Relini et al. 2009). Los datos señalan que mientras el incremento en los niveles
de Ca2+ intracelular en las neuronas hipocampales fue mantenido en la presencia
de Co2+, este incremento fue bloqueado en las células RCSN-3 (Figura 36). Por lo
tanto, estos resultados indican que el aumento en los niveles de Ca2+ intracelular
mediado por oligómeros de -Sin estaría mediado por diferentes mecanismos en
estos dos tipos de células. Mientras que en las neuronas hipocampales las
“estructuras tipo poro” de -Sin estarían implicadas en la entrada de Ca2+, en las
células RCSN-3 el incremento de Ca2+ intracelular inducido por oligómeros de -
Sin se explicaría por la apertura de canales de Ca2+ dependientes de voltaje.

106
Figura 36. Efecto de oligómeros de -sinucleína en los niveles intracelulares
de Ca2+ en neuronas hipocampales y células RCSN-3. A través de una técnica
fluorimétrica y utilizando la sonda Fluo-4AM se observó que al adicionar
oligómeros de -Sin (0.5 M) a los 10 minutos después de iniciado el registro, se
produjo un incremento en los niveles de Ca2+ intracelular (rojo), con respecto al
control (vehículo, negro) tanto en neuronas hipocampales (A) como en células
RCSN-3 (B), respectivamente. El uso de un inhibidor no específico de los canales
de Ca2+ como el Co2+ causó un efecto diferencial en los dos tipos de células. C y
D, Cuantificación de los datos obtenidos en “A” y “B”, respectivamente, luego de
75 minutos de comenzado el registro. Los resultados son el promedio ± SEM
obtenido de 3 experimentos diferentes (n=3, **p <0.01, ***p <0.001).
0 10 20 30 40 50 60 700.0
0.10.8
1.0
1.2
1.4
1.6
1.8
2.0
2.2Control + vehículo
-Sin 0.5 M
Co2+ 10 M + vehículo
Co2+ 10 M + -Sin 0.5 M
perfusión
Tiempo (s)
Inte
nsid
ad
de
Flu
ore
scen
cia
No
rmaliz
ad
a (
DF
/F0)
0 10 20 30 40 50 60 700.0
0.10.8
1.0
1.2
1.4
1.6
1.8
2.0
2.2Control + vehículo
-Sin 0.5 M
Co2+ 10 M + vehículo
Co2+ 10 M + -Sin 0.5 M
perfusión
Tiempo (s)
Inte
nsid
ad
de
Flu
ore
scen
cia
No
rmaliz
ad
a (
DF
/F0)
Neuronas hipocampales
Veh
ículo
-Sin
+ ve
hículo
2+
Co
-Sin
+
2+
Co
0.0
0.5
1.0
1.5
2.0
*** ***
n=22 n=34 n=14 n=19
Inte
nsid
ad
de
Flu
ore
scen
cia
No
rmaliz
ad
a (
DF
/F0)
Células RCSN-3
Veh
ículo
-Sin
+ v
ehíc
ulo
2+
Co
-Sin
+
2+
Co
0.0
0.5
1.0
1.5
2.0
**
n=20 n=52 n=40 n=21
Inte
nsid
ad
de
Flu
ore
scen
cia
No
rmaliz
ad
a (
DF
/F0)
A
B
C
D

107
Finalmente, para comprobar que el aumento observado en los niveles de
Ca2+ intracelular en neuronas hipocampales luego del tratamiento con oligómeros
de -Sin es producto sólo de la entrada de Ca2+ extracelular y no de la liberación
de reservas de calcio intracelular, se realizaron experimentos en ausencia de Ca2+
externo. Los resultados indican que en ausencia de Ca2+ externo no se genera un
incremento en los niveles de Ca2+ intracelular (Figura 37).
Figura 37. Efecto de oligómeros de -sinucleína en los niveles intracelulares
de Ca2+ en neuronas hipocampales en ausencia de Ca2+ externo. A, A través
de una técnica fluorimétrica y utilizando la sonda Fluo-4AM se observó que al
adicionar oligómeros de -Sin (0.5 M) a los 10 minutos después de iniciado el
registro, no se produjo un incremento en los niveles de Ca2+ intracelular (verde) en
ausencia de Ca2+ externo, con respecto al control (vehículo, negro). B,
Cuantificación de los datos obtenidos en “A” y “B”, respectivamente, luego de 75
minutos de comenzado el registro. Los resultados son el promedio ± SEM
obtenido de 3 experimentos diferentes (n=3, **p <0.01, ***p <0.001).
0 10 20 30 40 50 60 700.0
0.10.8
1.0
1.2
1.4
1.6
1.8
2.0
2.2Vehículo + SEN s/calcio
-Sin 0.5 M + SEN s/calcio
perfusión
Tiempo (minutos)
Inte
nsid
ad
de
Flu
ore
scen
cia
No
rmaliz
ad
a (
DF
/F0)
Veh
ículo
+ S
EN s
/cal
cio
-Sin
+ S
EN s
/cal
cio
0.0
0.5
1.0
1.5
n=7 n=5
Inte
nsid
ad
de
Flu
ore
scen
cia
No
rmaliz
ad
a (
DF
/F0)
A B

108
Oligómeros de -sinucleína aumentan la transmisión sináptica de neuronas
hipocampales
Dado el incremento observado en los niveles de Ca2+ intracelular en
neuronas hipocampales luego del tratamiento con oligómeros de -Sin, se
evaluaron diferentes parámetros que son indicadores de la actividad sináptica.
Estos análisis se realizaron a partir del rango de tiempo en que se observó la
mayor asociación inicial (temprana) de los oligómeros de -Sin a los cultivos
primarios de hipocampo de rata (Figura 38).
Figura 38. Asociación tiempo dependiente de los oligómeros de -sinucleína
a cultivo primario neuronal. A, El Western blot superior muestra la
inmunorreactividad contra -Sin en lisados de cultivos primarios de hipocampo de
rata tratados con oligómeros de -Sin (0.5 M) por los tiempos indicados en la
figura. El Western blot inferior muestra la inmunorreactividad contra -actina en las
mismas muestras. B, El grafico muestra la cuantificación de los niveles totales de
-Sin mostrados en “A”. Las barras corresponden al promedio ± SEM obtenido
desde 3 experimentos independientes (n=3, ** p< 0.01).
(min) 0 15 30 60 90 kDa
55
17
7295
130
170
43
34
26
-actina
0 15 30 60 900
1
2
3
4
**
-Sin 0.5 M
Tiempo (min)
-s
in/
-acti
na
(un
idad
es r
ela
tivas)
A B

109
Primero se evaluaron las transitorias de Ca2+, que corresponden a la
entrada del Ca2+ al medio intracelular del componente postsináptico mediante la
apertura de canales activados por ligando, específicamente, los receptores de N-
metil-D-aspartato (NMDA) (Markram et al. 1994). Considerando esto último, las
transitorias de Ca2+ son dependientes de la liberación del neurotransmisor al
medio extracelular, por lo tanto, dependientes de la funcionalidad del componente
presináptico. En el desarrollo de estos experimentos se incubaron neuronas
hipocampales con oligómeros de -Sin (0.5 M) por 1.5, 3 y 6 horas,
registrándose las transitorias de Ca2+ a intervalos de 2 segundos (iluminadas por
200 ms) durante 5 minutos. Los resultados indican que oligómeros de -Sin (0.5
M) en condiciones sub agudas generan un incremento significativo en la
frecuencia de las transitorias de Ca2+, con un efecto máximo de 3 veces sobre el
control a las 3 horas de incubación (Figura 39).

110
Figura 39. Oligómeros de -sinucleína incrementan las transitorias de Ca2+
en neuronas hipocampales. A, Trazos representativos de las transitorias de Ca2+
de neuronas hipocampales tratadas con oligómeros de -Sin (0.5 M) por 1.5, 3 y
6 horas. B, Cuantificación de la frecuencia de los trazos señalados en “A”. Las
barras corresponden al promedio ± SEM obtenido a partir 3 experimentos
diferentes (n=3, * p< 0.05, ** p< 0.001).
Las corrientes en miniatura son el reflejo de la actividad postsináptica
producto de la liberación espontánea de vesículas presinápticas. Para evaluar éste
segundo parámetro sináptico utilizamos TTX (25 nM) para inhibir la generación de
potenciales de acción. Como se muestra en la Figura 40 los resultados revelaron
un aumento significativo en la frecuencia de 2.5 veces sobre el control (n=9) sin
verse afectada la amplitud de los registros de neuronas hipocampales luego de 3
horas de incubación con oligómeros de -Sin (0.5 M).
Contr
ol
1.5
hrs
3 hrs
6 h
rs0
100
200
300
400
500**
*
*
-Sin 0.5 M
n= 5 n= 5 n= 5 n= 3
Fre
cu
en
cia
(% c
on
tro
l)
Control 1.5 hrs
3 hrs 6 hrs
50 s
100 U
FR
A B

111
Figura 40. Oligómeros de -sinucleína incrementan las corrientes sinápticas
en miniatura en neuronas hipocampales. A, Trazos representativos de las
corrientes sinápticas en miniatura (TTX, 25 nM) de neuronas hipocampales
tratadas con oligómeros de -Sin (0.5 M) por 1.5, 3 y 6 horas. B, Cuantificación
de la frecuencia y amplitud de los trazos señalados en “A”. Las barras
corresponden al promedio ± SEM obtenido a partir de 3 experimentos diferentes
(n=3, ** p< 0.01).
Contr
ol
1.5
hrs
3 hrs
6 hrs
0
50
100
150
n=12 n=10 n=13 n=19
-Sin 0.5 M
Am
plit
ud
(% c
on
tro
l)
Contr
ol
1.5
hrs
3 hrs
6 hrs
0
100
200
300
400
**
n= 12 n= 10 n= 13 n= 19
-Sin 0.5 M
Fre
cu
en
cia
(% c
on
tro
l)
A B
Control
6050403020100
Time (s)
IN 0
(pA
)
-200
-150
-100
-50
0
50
2 3 41
6050403020100
Time (s)
IN 0
(pA
)
-200
-150
-100
-50
0
50
2 3 41
4003002001000
Time (s)
IN #
0
(pA
)
-300
-200
-100
0
100
200
300
23 41
6050403020100
Time (s)
IN 0
(pA
)
-200
-150
-100
-50
0
50
23 41
10 ms
100 p
A
1.5 hrs
3 hrs
6 hrs

112
El tercer parámetro sináptico que evaluamos fue la liberación y reciclaje de
vesículas sinápticas de neuronas hipocampales, en presencia de oligómeros de -
Sin (0.5 M) utilizando la sonda fluorescente FM1-43 (Molecular probes, USA). Su
carácter anfipático le permite interaccionar mediante su región hidrofóbica con el
componente lipídico de las membranas, donde la intensidad de la señal asociada
a FM1-43 es 10 veces mayor que en solución. Una vez que las neuronas
endocitan parte de su membrana unida a FM1-43 se genera una marca específica
para vesículas sinápticas (Betz et al. 1992). Esta metodología permite estudiar la
liberación y reciclamiento de vesículas sinápticas frente a variados estímulos.
Primero, determinamos si oligómeros de -Sin de aumentan la liberación de
vesículas sinápticas luego de 3 horas de incubación como consecuencia del
aumento en las transitorias de Ca2+ y en la actividad sináptica. Neuronas control y
tratadas con -Sin fueron cargadas con FM1-43 y, subsecuentemente, expuestas
a un pulso de K+ 60 mM, lo que permite la depolarización de la membrana,
causando una disminución en la fluorescencia asociada a la sonda. Los datos
señalan que luego del tratamiento con oligómeros de -Sin, neuronas
hipocampales presentaron un mayor decaimiento de la fluorescencia asociada a
FM1-43 con respecto al control (n=21, **p < 0.01), indicando que la presencia de
oligómeros de -Sin facilitaría la liberación de vesículas sinápticas (Figura 41A).
Los valores para la fracción de depleción (ΔF/F0) de FM1-43 luego de 300 s de
registro fueron de 0.7129 ± 0.02 para el control (vehículo) y 0.6445 ± 0.01 para las
neuronas tratadas con -Sin, existiendo una diferencia significativa entre ambos
tratamientos (Figura 41B).

113
Para continuar el estudio de los efectos de oligómeros de -Sin sobre el
reciclaje de las vesículas sinápticas, caracterizamos una metodología que nos
permitiera medir la endocitosis de FM1-43, como reflejo de la endocitosis de
vesículas sinápticas. Brevemente, el protocolo utilizado fue descrito por Ryan en
1996, el cual se basa en estimular (despolarizar) y esperar un intervalo de tiempo
(∆t) entre el estímulo despolarizante y la carga con FM1-43. La cantidad de
vesículas marcadas disminuye a medida que se aumenta el ∆t entre el estímulo y
la carga con FM1-43. Utilizando este protocolo encontramos que el decaimiento
asociado a FM1-43 disminuye significativamente cuando se da un ∆t 180 s entre el
estímulo despolarizante y la carga con FM1-43 con valores de 0.7186 ± 0.02 para
el control (vehículo) y 0.8174 ± 0.03 para las neuronas hipocampales tratadas con
oligómeros -Sin (Figura 41C).

114
Figura 41. Oligómeros de -sinucleína alteran el reciclamiento vesicular de
neuronas hipocampales. A, Imágenes de epifluorescencia de neuronas
hipocampales control y neuronas tratadas con oligómeros de -Sin (0.5 M),
incubadas con la sonda fluorescente FM1-43 en ausencia o presencia de KCl (K+;
60 mM). B, Cuantificación de la fluorescencia asociada a la sonda FM1-43. C,
Previo al marcaje con la sonda se realizo un ∆t de 180 s con el fin de determinar la
endocitosis vesicular. Cuantificación de la fluorescencia asociada a la sonda FM1-
43. Los gráficos corresponden al promedio ± SEM obtenido a partir de 3
experimentos diferentes (n=3, ** p< 0.01).
0 50 100
150
200
250
300
0.0
0.1
0.7
0.8
0.9
1.0
1.1
K+ 60 mM
**
-Sin 0.5 M
Control
Tiempo (s)
Flu
ore
scen
cia
FM
1-4
3
(DF
/F0)
B
Dt = 0 s Dt = 50 s Dt = 100 s
Dt = 150 s Dt = 200 s Dt = 250 s
3 m
-Sin 0.5 M
Dt = 0 s Dt = 50 s Dt = 100 s
Dt = 150 s Dt = 200 s Dt = 250 s
3 m
ControlA
0 50 100
150
200
250
300
0.00.1
0.7
0.8
0.9
1.0
1.1
Control
-Sin 0.5 M
K+ 60 mM
**
Tiempo (s)
Flu
ore
scen
cia
FM
1-4
3
(DF
/F0)
C

115
Finalmente, el último parámetro sináptico que evaluamos fue determinar si
los cambios a nivel de transmisión sináptica se encuentran acompañados de
cambios sinápticos morfológicos a nivel neuronal. Para esto, los cambios a nivel
del componente presináptico fueron analizados utilizando un anticuerpo que
reconoce SNAP25, una proteína SNARE que permite la correcta fusión entre las
vesículas sinápticas y la membrana plasmática. Para ello, neuronas hipocampales
fueron tratadas con oligómeros de -Sin (0.5 M) por 3 y 24 horas a 37ºC. Análisis
cualitativo y cuantitativo de las imágenes nos revelaron que oligómeros de -Sin
generan un aumento significativo en el número de puntas para SNAP25 a las 3
horas de incubación con un 166 ± 18% respecto al control, observándose una
tendencia a la disminución a las 24 horas con un 87 ± 8% respecto al control
(Figura 42).

116
Figura 42. Oligómeros de -sinucleína incrementan la inmunorreactividad de
SNAP25 en neuronas hipocampales. A,B,C, Inmunorreactividad asociada a
MAP2 (Cy3, Rojo), SNAP25 (FITC, Verde) y tinción fluorescente nuclear (DAPI,
Azul) en neuronas hipocampales controles o tratadas con oligómeros de -Sin (0.5
M) por 3 y 24 horas a 37ºC. D, Cuantificación del número de puntas de SNAP25
obtenidas en “A”. Las imágenes confocales representan al menos 9 imágenes
obtenidas desde 3 experimentos independientes (n=3, * p< 0.05).
Control 3 hrs 24 hrs0
50
100
150
200*
-Sin 0.5 M
Nº
de p
un
tas d
e
SN
AP
25/
20
m
(%
co
ntr
ol)
D
SNAP25-FITC
MAP2- Cy3
DAPI
-Sin 3 hrs -Sin 24 hrsControl
10 m 10 m 10 m
A B C
A1 B1 C1

117
Por otro lado, para evaluar la sinaptotoxicidad de -Sin realizamos
incubaciones con oligómeros de -Sin 5 M por 24 horas a 37ºC y analizamos una
proteína presente en las vesículas sinápticas, llamada SV2 (Synaptic vesicle
protein 2). Los resultados indican una disminución significativa en el número de
puntas inmunoreactivas presentes en los procesos primarios (20 m) en un 52 ±
10% respecto al control (Figura 43).
En conjunto, hemos encontrado que oligómeros de -Sin producen en
condiciones subagudas un incremento en la transmisión sináptica lo que lleva a
finalmente a una depleción vesicular.

118
Figura 43. Oligómeros de -sinucleína disminuyen la inmunorreactividad de
SV2 en neuronas hipocampales después de aplicación crónica. A,
Inmunorreactividad asociada a MAP2 (FITC, Verde) y SV2 (Cy3, Rojo) en
neuronas hipocampales controles o tratadas con oligómeros de -Sin (5 M) por
24 horas a 37ºC. B, Cuantificación del número de puntas de SV2 obtenidas en “A”.
Las imágenes confocales representan al menos 9 imágenes obtenidas desde 3
experimentos independientes (n=3, * p< 0.05).
SV2-Cy3MAP2-FITC Merge
Control
A1
-Sin 24 hrs
A
15 µm
SV2-Cy3MAP2-FITC Merge B1
20 m 20 m 20 m
20 m 20 m 20 m
4 m
4 m
Contr
ol M
-Sin
5
0
50
100
150
*
Nº
de p
un
tas d
e S
V2/
20
m
(%
co
ntr
ol)
C
A1
B1
B

119
Objetivo 2.2. Determinar la toxicidad mediada por agregados de -sinucleína
Actualmente, se sabe que antibióticos/antifúngicos como gramicidina o
anfotericina son extremadamente tóxicos debido a su capacidad de unirse a la
célula y producir un aumento sostenido de la permeabilidad iónica producto de la
formación de poros a nivel de la membrana plasmática (Tajima et al. 1996). Por lo
tanto, el fin de este objetivo es evaluar si la formación de estructuras tipo
poro/perforado por parte de oligómeros de -Sin media fenómenos de toxicidad en
neuronas hipocampales.
Resultados
En condiciones crónicas de 24 horas oligómeros de -sinucleína no causan
toxicidad en neuronas hipocampales
Dado lo anteriormente expuesto y considerando los resultados que indican
que oligómeros de -Sin en condiciones crónicas disminuyen el número de puntas
para SV2, se incubaron neuronas hipocampales con oligómeros de -Sin 5 M por
24 horas y realizamos ensayos de viabilidad con Alamar Blue. Como resultado, se
observó una actividad reductora celular similar a la del control (Figura 44B). Sin
embargo, al analizar las imágenes en campo claro observamos que en la
presencia de oligómeros de -Sin la integridad de la célula si se encuentra
afectada, lo cual no fue detectado por el ensayo de viabilidad utilizado (Figura
44A).

120
Figura 44. Efecto crónico de oligómeros de -sinucleína recombinante
humana sobre neuronas hipocampales. A, Imágenes en campo claro de
neuronas hipocampales tratadas con oligómeros de -Sin (5 M) por 24 horas. B,
Cuantificación de la viabilidad celular utilizando ensayos de Alamar Blue. Las
barras corresponden al promedio ± SEM de 3 experimentos independientes (n=3).
Posteriormente, con el objetivo de determinar la toxicidad de agregados de
-Sin obtenida desde fuentes naturales utilizamos la línea celular SHSY-5Y,
derivada de neuroblastoma humano, la cual fue transfectada establemente con el
gen para sobreexpresar -Sin wild-type humana (Vekrellis et al. 2009). Ésta línea
celular tiene la característica de secretar oligómeros de -Sin al medio extracelular
(Emmanouilidou et al. 2010). Como control se utilizaron células SHSY-5Y,
Contr
ol
Veh
ículo M
-Sin
5
0
20
40
60
80
100
120
Inte
nsid
ad
de
Flu
ore
scen
cia
(%
co
ntr
ol)
A
B
Control Vehículo -Sin 5 M
15 m

121
transfectadas establemente con el plásmido control -galactosidasa (-gal). Para
evaluar la presencia de -Sin en el medio condicionado se realizó un ensayo de
Dot blot a distintos días de cultivo desde que fueron plaqueadas las células. En
estos análisis se obtuvo que la línea celular SHSY-5Y secreta -Sin al medio
extracelular, de forma estable según la señal detectada en el Dot blot, desde el
primer día de crecimiento. Al cuantificar las señales obtenidas, determinamos que
fue en el día 4 de crecimiento donde se presentó la mayor diferencia con respecto
a la señal control (Figura 45A), lo que corresponde a una concentración
aproximada de 7 nM de -Sin (Emmanouilidou et al. 2010). Para los ensayos de
viabilidad se utilizaron neuronas corticales, principalmente, por su fácil
disponibilidad, las cuales fueron incubadas con el medio condicionado control ó el
medio enriquecido en -Sin por 24, 48 y 72 horas. Los resultados obtenidos
utilizando ensayos de Alamar blue indican que a las 24 y 48 horas el medio
condicionado con -Sin no generó una pérdida significativa en viabilidad respecto
a las neuronas control (Figura 45B, 45C). Sin embargo, luego de 72 horas de
incubación si se observó una disminución importante en la viabilidad celular con
respecto a las neuronas control en un 70 ± 1%, sin detectarse una diferencia
significativa en el efecto mediado por ambos medios condicionados (-gal y -Sin)
(Figura 45D).
Finalmente, concluimos en este objetivo que tanto oligómeros de -Sin
recombinante humana como -Sin obtenida desde células que sobreexpresan
esta proteína, no afectan la viabilidad neuronal en las condiciones de estudio
anteriormente mencionadas.

122
Figura 45. Efecto crónico de medio condicionado con -sinucleína sobre
cultivo neuronal. A, Inmunorreactividad (Dot Blot) y cuantificación de la señal
asociada a -Sin obtenida desde medios condicionados de células SHSY-5Y
transfectadas establemente con -Sin humana. Cuantificación de la viabilidad
celular utilizando ensayos de Alamar Blue de neuronas corticales tratadas por 24
(B), 48 (C) y 72 horas (D) con los medios condicionados enriquecidos en -Sin.
Como medio de control se utilizaron células SH-SY5Y establemente transfectadas
con -galactosidasa (-gal). Las barras corresponden el promedio ± SEM obtenido
a partir de 3 experimentos independientes (n=3, ** p< 0.01, *** p< 0.001).
Contr
ol-g
al
-Sin
0
50
100
150
*****
In
ten
sid
ad
de
Flu
ore
scen
cia
(%
co
ntr
ol)
Contr
ol-g
al
-Sin
0
50
100
150
Inte
nsid
ad
de
Flu
ore
scen
cia
(%
co
ntr
ol)
Contr
ol-g
al
-Sin
0
50
100
150
Inte
nsid
ad
de
Flu
ore
scen
cia
(% c
on
tro
l)
A B
Contr
ol
Día
40
2
4
6
Inte
nsid
ad
de s
eñ
al
(U
FR
)
C D

123
4.- DISCUSIÓN Y CONCLUSIONES
La formación de oligómeros de -sinucleína ocurre por una vía alternativa a
la fibrilización convencional
Similar a A, la agregación de -Sin in vitro no es una simple transición
entre dos estados, desde monómeros a fibras; sino que más bien representa un
proceso muy complejo que involucra la formación de intermediarios oligoméricos
de varios tamaños y morfologías (Wood et al. 1999). Si bien las formas
moleculares de los oligómeros de -Sin in vitro se asemejan a las encontradas
en el cerebro (El-Agnaf et al. 2006), el rol de estos oligómeros en la etiología de
la EP y en particular si éstos efectivamente corresponden a las especies
moleculares responsables de la EP, es un aspecto que aún requiere
confirmación empírica (Kayed et al. 2003; Tsika et al. 2010; Colla et al. 2012).
La caracterización estructural y bioquímica de los oligómeros de -Sin
formados a pH 4.0 indicó que estos correspondían a una mezcla de oligómeros
solubles de bajo peso molecular y monómeros, siendo estos últimos los más
abundantes. Además, se encontró que las estructuras oligoméricas fueron
resistentes a beta mercaptoetanol y SDS (Figura 7). En este contexto, algunos
trabajos (Hoyer et al. 2002) indican que la disminución del pH del medio de 7 a 4
inhibe la formación de fibras de -Sin favoreciendo la generación de agregados
amorfos producto de cambios en la estructura secundaria de la proteína,
acompañado por un incremento en la hidrofobicidad y una disminución en la
eficiencia de fibrilización (Hoyer et al. 2002). En este sentido, diversos estudios

124
han señalado que la toxicidad de las proteínas amiloides es dependiente de su
morfología, siendo los oligómeros de bajo peso molecular mucho más tóxicos
que los agregados fibrilares (Bucciantini et al. 2004).
Considerando que la secreción de -Sin por medio del sistema vesicular
requeriría que ésta fuera primero translocada desde el citoplasma a las vesículas
de secreción (Figura 3; Nickel et al. 2009), y que el ambiente vesicular tiene un
pH más ácido que el citosol (Hoyer et al. 2004), se podría esperar que al interior
de las vesículas se favoreciera la formación de oligómeros similares a los
descritos en esta tesis (Figura 7). Por lo tanto, -Sin intravesicular estaría más
propensa a la oligomerización que -Sin citosólica dado, principalmente, a
pequeñas diferencias presentes en el microambiente del lumen vesicular que
podrían alterar las propiedades fisicoquímicas de la proteína y afectar la cinética
de agregación (Lee et al. 2005). De hecho, algunos factores y componentes
luminales de ciertos tipos de vesículas, aceleran la oligomerización de -Sin,
como por ejemplo, el pH ácido y ciertos lípidos (Bar-On et al. 2008).
Por otro lado, en concordancia con los resultados de esta tesis que indican
que al realizar incubaciones de -Sin por 48 horas se forman agregados
amiloides difusos (ThT-positivos,sensibles a la acción proteolítica de PK,
congofílicos no birrefringentes y no fibrilares; Figuras 8 y 9), se ha descrito que
protofibras aisladas desde cerebros post-mortem de pacientes diagnosticados
con EA se obtienen principalmente de placas seniles difusas (Lasagna-Reeves et
al. 2011).

125
Aunque no se ha determinado con certeza cuál es la vía de agregación
por la cual se forman las especies permeabilizantes de -Sin, nosotros
proponemos, en base a los resultados obtenidos en esta tesis, que esto ocurriría
a través de una vía alternativa a la fibrilogénesis convencional dentro de
sistemas vesiculares.
Oligómeros de -sinucleína se asocian a neuronas y causan daños a nivel de
la membrana plasmática
La evidencia experimental acumulada hasta la fecha apoya la idea de que
oligómeros de -Sin alteran la permeabilidad de la membrana a través de la
generación de poros (Quist et al. 2005; Zakharov et al. 2007; Tsigelny et al. 2007;
Kim et al. 2009; Feng et al. 2010; Kostka et al. 2010; Schmidt et al. 2012). Aún no
se han determinado con precisión la importancia de la formación de poros a nivel
celular y menos si estas estructuras se puedan formar en membranas
neuronales. En el presente estudio, utilizando microscopía confocal y técnicas de
patch clamp perforado encontramos que oligómeros de -Sin se asocian a la
membrana plasmática de neuronas hipocampales lo que conlleva a la formación
de "estructuras tipo poro", incrementándose la conductancia de membrana y el
influjo de un análogo fluorescente de la glucosa (Figuras 10 y 11). Estos
resultados son muy interesantes, ya que indican que moléculas con relevancia
biológica podrían difundir dentro o afuera de la neurona a través de estas
grandes disrupciones de la membrana formada por agregados de -Sin.
Consistente con nuestros datos, estudios previos de AFM y dinámica molecular
de -Sin en membranas artificiales apoyan la presencia de distintos poros

126
transmembrana con diferentes tamaños y conductancias (Tsigelny et al. 2012;
Kostka et al. 2008; Quist et al. 2005; Tsigelny et al. 2007) que posiblemente
corresponden a la perforación funcional descrita en este estudio, que poseerían
un diámetro interno mínimo estimado de ~1 nm, de acuerdo a los datos
obtenidos con el análogo no hidrolizable de glucosa 6-NBDG (Figura 11). Con
respecto al modelo de perforación, los resultados obtenidos apoyan la hipótesis
del modelo de barril, que se basa en la unión de oligómeros y no de monómeros
de -Sin (modelo toroidal) a la membrana plasmática, ya que al realizar patch
clamp perforado con -Sin soluble (previa al proceso de agregación) no se
observaron cambios significativos en la conductancia de membrana como
sucedió con oligómeros de -Sin.
Otro resultado importante a considerar es que la disrupción de la
membrana fue altamente dependiente del tipo celular. Por ejemplo, la línea
celular RCSN-3 derivada de sustancia nigra de rata, fue completamente
resistente a la acción permeabilizante de oligómeros de -Sin (Figura 13) y
creemos que esto se relaciona con la formación de cuerpos de inclusión,
detectados por microscopía confocal (Figura 18). Investigaciones previas han
descrito que las neuronas dopaminérgicas son particularmente vulnerables al
estrés oxidativo, posiblemente producto de la propia oxidación de la dopamina,
generándose especies reactivas del oxígeno (Lotharius et al. 2002) que
promueven la agregación de -Sin (Lee et al. 2011) formándose dímeros y
agregados insolubles que probablemente corresponden a CL (Hashimoto et al.

127
1999). Por lo tanto, estas investigaciones concuerdan con los presentes
resultados obtenidos en las células RCSN-3.
Experimentos realizados con la técnica de patch perforado, demostraron
que las células RCSN-3 son más resistentes a la perforación por parte de
oligómeros de -Sin, incluso en presencia del pesticida rotenona, conocido
inductor de Parkinsonismo (Figura 14) (Engel et al. 2001; Ascherio et al. 2006).
Esto podría ser explicado por la naturaleza clonal de las células RCSN-3, que les
entrega resistencia a ciertos agentes como -Sin o rotenona, más que por su
naturaleza dopaminérgica, ya que neuronas disociadas dopaminérgicas de la
sustancia nigra a P20 fueron sensibles a las propiedades perforantes de los
oligómeros de -Sin (Figura 16). Estudios previos han determinado que mientras
más indiferenciada es la célula menor es la expresión del Complejo I
mitocondrial, por lo tanto, esto podría ser uno de los factores que explican la
resistencia de las células RCSN-3 a -Sin (Hoegger et al. 2008).
Otra posible explicación para los resultados obtenidos con células RCSN-3
podría estar relacionado con el hecho de que patológicamente la EP se
caracteriza por la acumulación intracelular de CL en las neuronas sobrevivientes
a la neurodegeneración en la sustancia nigra (Spillantini et al. 1997), en
consecuencia, la formación de cuerpos de inclusión en las células RCSN-3
podría ser un mecanismo de protección frente a las especies más pequeñas
(oligómeros) responsables de la formación de “estructuras tipo poro” de -Sin, al
contrario de lo que se observa en neuronas hipocampales, en las cuales no se
observaron cuerpos de inclusión de -Sin (Figura 17).

128
En conjunto, es sumamente importante considerar que la formación de
estructuras tipo poro/perforado de -Sin no es un fenómeno general aplicable a
todos los modelos celulares, sino más bien un fenómeno particular de las
neuronas estudiadas.
La asociación de -sinucleína a la membrana plasmática es un proceso
fundamental en la formación de “estructuras tipo poro”
La toxicidad de las proteínas amiloides se correlaciona con el tipo de
asociación a la membrana (Butterfield et al. 2010). Por un lado, la superficie de la
membrana, dependiendo de su composición química, podría servir como sitio de
de “andamiaje patológico” para promover la agregación de proteínas amiloides
(Chi et al. 2008). Mientras que por otro lado, las proteínas amiloides podrían
alterar la integridad estructural de la membrana permitiendo el paso de pequeñas
moléculas y iones a través de ella (Ding et al. 2009).
Dadas las diferencias en la asociación de -Sin en neuronas
hipocampales y células RCSN-3, investigamos cuáles eran los determinantes
moleculares de membrana que estaban involucrados en este proceso. En
neuronas hipocampales luego de la remoción del componente proteico de
membrana a través de la utilización de tripsina observamos una disminución
significativa de asociación de -Sin, sin observarse cambios significativos al
remover el colesterol de membrana (Figuras 25 y 26). Es importante mencionar
que al realizar experimentos de patch clamp perforado con neuronas pre tratadas
con Fosfolipasa C (FLC), se bloqueo la formación de perforados de -Sin,
sugiriendo que proteínas con tallo GPI participarían en la unión y formación de

129
“estructuras tipo poro” de -Sin (Figura 12). Apoyando nuestros resultados,
estudios previos realizados en células Hela que sobreexpresan -Sin han
demostrado que -Sin co-fracciona con proteínas con tallo GPI como CD55
(Complement decay-accelerating factor) y Thy-1 (Cluster of Differentiation 90)
presentes en las balsas lipídicas de esas células (Fortin et al. 2004).
En el caso de las células RCSN-3, se encontró que la remoción de
componentes proteicos y la reducción del colesterol de membrana, influyeron en
el número de los grandes agregados de -Sin asociados (Figuras 27 y 28). Estos
resultados concuerdan con estudios previos, que sugieren que -Sin se une a la
membrana principalmente en las zonas de balsas lipídicas, ricas en colesterol y
glicoesfingolípidos (Fortin et al. 2004; Fantini et al. 2011). En relación a lo
anterior, es importante mencionar que mecanismos de endocitosis
independientes de clatrina, asociados a balsas lipídicas y por consecuencia
dependientes de colesterol (Le Roy et al. 2005; Mayor et al. 2007), podrían estar
implicados en la internalización de -Sin extracelular y en la formación de
agregados de mayor tamaño, y que a su vez podría estar relacionado con el
efecto visto en la incubación a 4ºC, ya que a baja temperatura se disminuye la
fluidez de la membrana (Figuras 29, 30 y 31).
En resumen diferentes mecanismos estarían implicados en la unión de -
Sin. En neuronas hipocampales, proteínas transmembrana y ancladas por tallos
de GPI, permitirían asociar pequeños oligómeros de -Sin a nivel de membrana
plasmática, facilitando la formación de “estructuras tipo poro”; mientras que en
células RCSN-3 la asociación de oligómeros de -Sin, dependería de proteínas

130
de membrana y lípidos como el colesterol, que posibilitarían la endocitosis y
posterior formación de agregados intracelulares de -Sin.
-Sinucleína actúa como una proteína priónica en células RCSN-3
-Sin sería una proteína clave en la EP, no sólo porque es el mayor
componente de los CLs, sino porque también está implicada en varios procesos
celulares que están alterados en la EP (Spillantini et al., 1997). Últimamente, se
ha demostrado que -Sin al estar mal plegada se propagaría de una célula a
otra, en una manera similar al prión, favoreciendo la agregación de -Sin en la
célula blanco (Desplats et al. 2009). Al respecto, los resultados obtenidos en esta
tesis, a través de inmunocitoquímica indirecta, utilizando anticuerpos específicos
tanto para -Sin exógena como para la -Sin endógena presente en las células
RCSN-3, sugieren que los oligómeros de -Sin exógena servirían como centros
de nucleación para la agregación de -Sin endógena, cambiando su patrón de
distribución normal de pequeños agregados a grandes agregados
yuxtanucleares, dando cuenta de su capacidad para ingresar a la célula,
asociarse y agregarse junto con -Sin endógena (Figura 32). Esto resulta,
sumamente interesante dado que sugiere que las células RCSN-3 serían un
buen modelo celular para explicar, tanto la formación como la propagación de los
CLs en la EP.
Células RCSN-3 forman agresomas de -sinucleína
Aunque los agresomas no representan, necesariamente, los cuerpos de
inclusión característicos de la EP (Spillantini et al. 1997), nos planteamos

131
determinar a través de inmunocitoquímica indirecta si los agregados de -Sin
presentes en las células RCSN-3 eran agresomas. La utilización de colchicina
disminuyó de manera significativa tanto el tamaño, como el número de
agregados de -Sin, sin embargo, al utilizar EHNA sólo observamos una
disminución significativa en el tamaño y no en el número de agregados de -Sin
(Figura 34). Esto es producto, probablemente, de fallas en el transporte
retrógrado de -Sin, pero no de su inhibición total como ocurrió al inhibir la
depolarización de los microtúbulos con colchinina. De manera interesante, al
analizar un marcador de agresomas como lo es ubiquitina, observamos que los
agregados de -Sin colocalizan con ésta proteína, cambiando su patrón de
distribución desde una amplia a una más restringida disposición, además de
disminuir su señal total (Figura 35).
La presencia de inclusiones citoplasmáticas (agresomas) presentes en las
células RCSN-3, concuerda con investigaciones previas que sugieren que la
sobreexpresión de -Sin y fallas a nivel mitocondrial resultan en la formación de
inclusiones tipo agresomas basados, principalmente, en su inmunorreactividad a
ubiquitina, la subunidad 20S del proteosoma y Hsp70 (Kopito 2000).
Oligómeros de -sinucleína causan sinaptotoxicidad en neuronas
hipocampales en condiciones crónicas
La toxicidad asociada a agregados de -Sin aún no ha sido completamente
dilucidada (Bartels et al. 2011). En este contexto, al evaluar la toxicidad de
agregados de -Sin (5 M) por 24 horas en neuronas hipocampales no se

132
encontró una disminución significativa en la viabilidad respecto al control (Figura
44B). Sin embargo, al analizar las imágenes en campo claro sí se observaron
cambios morfológicos, característicos de degeneración celular, como por ejemplo,
retracción del soma y de procesos, que no lograron ser detectados por la técnica
empleada (Figura 44A). El reactivo Alamar blue funciona como un indicador de
homeostasis celular que utiliza todo el poder reductor de la célula viva, y no sólo
de la funcionalidad de la reductasa mitocondrial como ocurre en los ensayos de
viabilidad por MTT, por lo tanto, para detectar cambios significativos en la
disminución de la viabilidad es necesario que el agente empleado tenga un efecto
citotóxico potente, como sucedió en el caso de rotenona (Figura 15).
Varios estudios previos han examinado la neurotoxicidad de amiloides
sintéticos, pero existe una gran barrera para entender la patogenicidad de las
distintas especies, principalmente, por la dificultad para especificar y controlar el
estado de agregación de las proteínas (Cleary et al. 2005). Por lo tanto,
evaluamos la toxicidad de oligómeros de -Sin secretados fisiológicamente desde
línea celular SHSY-5Y. La toxicidad del medio condicionado con -Sin sobre
cultivos primarios de corteza fue significativa luego de 72 horas de incubación, sin
embargo, al comparar con los datos obtenidos con medio condicionado control
(secreción de -gal) no se observó una diferencia significativa entre ambos, lo que
sugiere que la toxicidad observada pudo ser efecto del medio condicionado, y no,
específicamente, de -Sin (Figura 45D).
Dado que los agregados de -Sin tanto de origen recombinante, como de
origen fisiológico corresponden a una mezcla de especies de bajo peso molecular

133
(Emmanouilidou et al. 2010) sugerimos que, probablemente, para obtener un
efecto citotóxico más potente sería necesario, para un estudio futuro, purificar las
distintas especies de -Sin para evaluar cuál es la especie oligomérica implicada
en la toxicidad de -Sin.
La disfunción sináptica y la axonopatía se han descrito como un posible
marcador de las etapas tempranas y presintomáticas de la EP (Lundblad et al.
2012). Esto es consistente con nuestros resultados donde la acumulación -Sin
extracelular promueve la sinaptotoxicidad de neuronas hipocampales en
condiciones crónicas, representado por la disminución de la inmunorreactividad
para SV2 (Figura 43). Por otra parte, se ha descrito que la sobreexpresión de -
Sin humana reduce la sobrevivencia y el desarrollo dendrítico de neuronas en el
giro dentado (Winner et al. 2012). Por lo tanto, el efecto de los oligómeros de -Sin
en la zona del hipocampo podría explicar el déficit cognitivo observado en la EP.
Modelo de acción de oligómeros de -sinucleína en neuronas hipocampales
Estudios previos han sugerido que la pérdida de homeostasis en el Ca2+
intracelular sería crucial en la patogénesis de diversos desórdenes
neurodegenerativos (Mattson 2007). En la presente tesis determinamos que
oligómeros de -Sin alteran la integridad de la membrana plasmática formando
“estructuras tipo poro” permitiendo el incremento en los niveles de Ca2+ intracelular
(Figura 36) con un subsecuente incremento en la transmisión sináptica
relacionado con un efecto primario en la transmisión presináptica. Por ejemplo, la
incubación de neuronas hipocampales con oligómeros de -Sin causó un

134
incremento en diferentes parámetros tales como la frecuencia de las transitorias
de Ca2+ (Figura 39), la frecuencia de las corrientes sinápticas en miniatura (Figura
40), el decaimiento de la fluorescencia asociada a FM1-43 (Figura 41) y la
inmunotinción para SNAP25 (Figura 42). Estos resultados concuerdan con los
resultados obtenidos con otras proteínas amiloides, como el A que también
puede insertarse en la membrana, formar de perforaciones (Sepulveda et al. 2010)
y, finalmente, alterar la transmisión sináptica (Parodi et al. 2010).
Es pertinente mencionar que el rol fisiológico de -Sin como proteína
SNARE puede, al menos en un contexto de neurodegeneración causado por la
deleción de CSP, prevenir la neurodegeneración (Chandra et al. 2005). Esto
permite plantear la siguiente pregunta: ¿Los efectos neuropatológicos de
agregados de -Sin están causados por una ganancia o una pérdida de función?.
Es importante considerar que diversas evidencias indican que un evento temprano
en los procesos de neurodegeneración es la disfunción presináptica (Gray et al.
2009; Nemani et al. 2010), por lo tanto, una falla en la mantención del complejo
SNARE en la liberación de neurotransmisores podría llevar a la EP (Sharma et al.
2012; Burre et al., 2010; Greten-Harrison et al. 2010; Nemani et al.2010). Según
los datos obtenidos sugerimos que los efectos potenciadores de -Sin en la
transmisión sináptica de neuronas hipocampales se deben a una ganancia de
función que, finalmente, lleva a fenómenos de sinaptotoxicidad (Figura 46).

135
Formación e impacto de agregados intracelulares de oligómeros de -
sinucleína
Fue interesante encontrar que células RCSN-3, una línea clonal derivada de
la sustancia nigra y que dispone de un fenotipo dopaminérgico (Paris et al. 2008),
fuera resistente a oligómeros de -Sin (Figura 13), en comparación con las
neuronas hipocampales (Figura 10) y disociadas de la sustancia nigra a P20
(Figura 16). Del mismo modo, estas células no fueron afectadas por la rotenona
indicando una resistencia notable (Figura 14). Sin embargo, a pesar de que estas
células fueron resistentes a oligómeros de -Sin en términos de permeabilidad de
membrana, la aplicación de estos oligómeros a las células RCSN-3 indujeron un
aumento en el Ca2+ intracelular (Figura 36B). Este aumento pareció estar mediado
por un mecanismo diferente al presente en neuronas hipocampales (Figura 36A) y
que planteamos estaría asociado a la activación de los canales de Ca2+ voltaje-
dependientes sensibles a cobalto, lo que concuerda con lo señalado en literatura,
que indica que -Sin también podría modular la entrada de Ca2+ a la célula a
través de la activación de canales de Ca2+ voltaje dependiente del tipo N
(Adamczyk et al. 2006; Hettiarachchi et al. 2009).

136
Figura 46. Modelo propuesto para la acción diferencial de los oligómeros de
-sinucleína exógena en neuronas hipocampales y células RCSN-3. Sobre la
base del presente estudio postulamos que los mecanismos que conducen a la
perforación, endocitosis y propagación temprana de oligómeros de -Sin son
diversos dependiendo del tipo de celular. En neuronas hipocampales, oligómeros
de -Sin exógena estarían implicados en la sinaptotoxicidad neuronal a través de
la formación de “estructuras tipo poro” en la membrana plasmática que causan un
incremento en la concentración libre de Ca2+ intracelular con posteriores
alteraciones en la transmisión sináptica. En las células RCSN-3, oligómeros de -
Sin exógena promueven la formación de cuerpos de inclusión complejos,
constituidos por -Sin exógena y endógena, con características de agresoma.
Este mecanismo podría ser parte de un sistema citoprotector o ser parte del
mecanismo que permite la agregación y propagación de oligómeros de -Sin, a
diferentes áreas del cerebro de pacientes con la EP (Aguzzi et al. 2009).
poro/perforado
de -Sin
Ca2+
Agresoma
Oligómeros de
-SinNeurona hipocampal Célula RCSN-3
Aumento
transmisión
sináptica
Depleción
Vesicular Sinaptotoxicidad
-Sin endógena
-Sin exógena
Endocitosis

137
5.- AGRADECIMIENTOS
Me gustaría que estas líneas sirvieran para expresar mi más profundo y
sincero agradecimiento a todas aquellas personas que con su ayuda han
colaborado en la realización del presente trabajo, en especial a mis cotutores el
Dr. Carlos Opazo y al Dr. Aguayo, por la orientación, el seguimiento y la
supervisión continúa de la misma, pero sobre todo por la motivación y el apoyo
recibido a lo largo de estos años.
También quiero agradecer a los miembros de mi comisión de tesis, los
Dres. Francisco Nualart, Juan Pablo Henríquez y Pablo Caviedes por haber
contribuido en mi formación doctoral y a mejorar este trabajo de tesis, tanto en su
desarrollo como en su etapa final. De igual modo, al Dr. Francisco J. Muñoz por
haberme recibido en su laboratorio España.
Quisiera hacer extensiva mi gratitud a mis compañeros y profesores del
Departamento de Fisiología, en particular a los miembros del laboratorio de
Neurobiometales y Neurofisiología por su amistad y colaboración. Como también
al personal técnico en especial a Laurie.
A su vez agradezco el apoyo para la asistencia a eventos en el extranjero
de CONICYT y de la Escuela de Graduados de la Universidad de Concepción,
como también el apoyo a estadías en el extranjero de FEBS.
Finalmente, el desarrollo de esta tesis y de mis estudios ha sido posible
gracias a la Beca de Doctorado CONICYT, la Beca de Apoyo a Tesis Doctoral

138
CONICYT, al financiamiento del Anillo-PBCT ACT-04 (Luis Aguayo & Carlos
Opazo) y del FONDECYT Nº 1100502 (Luis Aguayo & Carlos Opazo).
A mi familia, mi querido René y amigos, muchas gracias por la comprensión,
paciencia y el ánimo recibidos.

139
6.- BIBLIOGRAFÍA
Aarsland, D., K. Bronnick, et al. (2007). "Neuropsychiatric symptoms in patients with Parkinson's disease and dementia: frequency, profile and associated care giver stress." J Neurol Neurosurg Psychiatry 78(1): 36-42.
Adamczyk, A., M. Kacprzak, et al. (2007). "Alpha-synuclein decreases arachidonic acid incorporation into rat striatal synaptoneurosomes." Folia Neuropathol 45(4): 230-235.
Adamczyk, A. and J. B. Strosznajder (2006). "Alpha-synuclein potentiates Ca2+ influx through voltage-dependent Ca2+ channels." Neuroreport 17(18): 1883-1886
Aguzzi, A. and M. Heikenwalder (2006). "Pathogenesis of prion diseases: current status and future outlook." Nat Rev Microbiol 4(10): 765-775.
Andersen, O.S., Koeppe, R.E., et al. (2005). Gramicidin channels. IEEE Trans Nanobioscience. 4, 10-20.
Bar-On, P., L. Crews, et al. (2008). "Statins reduce neuronal alpha-synuclein aggregation in in vitro models of Parkinson's disease." J Neurochem 105(5): 1656-1667.
Barnham, K. J., C. L. Masters, et al. (2004). "Neurodegenerative diseases and oxidative stress." Nat Rev Drug Discov 3(3): 205-214.
Bartels, T., J. G. Choi, et al. (2011). "alpha-Synuclein occurs physiologically as a helically folded tetramer that resists aggregation." Nature 477(7362): 107-110.
Betz, W. J., F. Mao, et al. (1992). "Activity-dependent fluorescent staining and destaining of living vertebrate motor nerve terminals." J Neurosci 12(2): 363-375.
Bisaglia, M., S. Mammi, et al. (2009). "Structural insights on physiological functions and pathological effects of alpha-synuclein." FASEB J 23(2): 329-340.
Braak, H., E. Ghebremedhin, et al. (2004). "Stages in the development of Parkinson's disease-related pathology." Cell Tissue Res 318(1): 121-134.

140
Bucciantini, M., G. Calloni, et al. (2004). "Prefibrillar amyloid protein aggregates share common features of cytotoxicity." J Biol Chem 279(30): 31374-31382.
Burre, J., M. Sharma, et al. (2012). "Systematic mutagenesis of alpha-synuclein reveals distinct sequence requirements for physiological and pathological activities." J Neurosci 32(43): 15227-15242.
Butterfield, S. M. and H. A. Lashuel (2010). "Amyloidogenic protein-membrane interactions: mechanistic insight from model systems." Angew Chem Int Ed Engl 49(33): 5628-5654.
Cannon, J. R., V. Tapias, et al. (2009). "A highly reproducible rotenone model of Parkinson's disease." Neurobiol Dis 34(2): 279-290.
Cleary, J. P., D. M. Walsh, et al. (2005). "Natural oligomers of the amyloid-beta protein specifically disrupt cognitive function." Nat Neurosci 8(1): 79-84.
Colla, E., P. H. Jensen, et al. (2012). "Accumulation of toxic alpha-synuclein oligomer within endoplasmic reticulum occurs in alpha-synucleinopathy in vivo." J Neurosci 32(10): 3301-3305.
Conway, K. A., J. D. Harper, et al. (2000). "Fibrils formed in vitro from alpha-synuclein and two mutant forms linked to Parkinson's disease are typical amyloid." Biochemistry 39(10): 2552-2563.
Cookson, M. R. and M. van der Brug (2008). "Cell systems and the toxic mechanism(s) of alpha-synuclein." Exp Neurol 209(1): 5-11.
Chandra, S., F. Fornai, et al. (2004). "Double-knockout mice for alpha- and beta-synucleins: effect on synaptic functions." Proc Natl Acad Sci U S A 101(41): 14966-14971.
Chandra, S., G. Gallardo, et al. (2005). "Alpha-synuclein cooperates with CSPalpha in preventing neurodegeneration." Cell 123(3): 383-396.
Chartier-Harlin, M. C., J. Kachergus, et al. (2004). "Alpha-synuclein locus duplication as a cause of familial Parkinson's disease." Lancet 364(9440): 1167-1169.
Danzer, K. M., D. Haasen, et al. (2007). "Different species of alpha-synuclein oligomers induce calcium influx and seeding." J Neurosci 27(34): 9220-9232.

141
Desplats, P., H. J. Lee, et al. (2009). "Inclusion formation and neuronal cell death through neuron-to-neuron transmission of alpha-synuclein." Proc Natl Acad Sci U S A 106(31): 13010-13015.
Ding, T. T., S. J. Lee, et al. (2002). "Annular alpha-synuclein protofibrils are produced when spherical protofibrils are incubated in solution or bound to brain-derived membranes." Biochemistry 41(32): 10209-10217.
El-Agnaf, O. M., S. A. Salem, et al. (2006). "Detection of oligomeric forms of alpha-synuclein protein in human plasma as a potential biomarker for Parkinson's disease." FASEB J 20(3): 419-425.
Ebihara, S., Shirato, K., et al. (1995).Gramicidin-perforated patch recording:
GABA response in mammalian neurones with intact intracellular chloride. J Physiol. 484,77-86
Emmanouilidou, E., K. Melachroinou, et al. (2010). "Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival." J Neurosci 30(20): 6838-6851.
Fantini, J., D. Carlus, et al. (2011). "The fusogenic tilted peptide (67-78) of alpha-synuclein is a cholesterol binding domain." Biochim Biophys Acta 1808(10): 2343-2351.
Feng, L. R., H. J. Federoff, et al. (2010). "Alpha-synuclein mediates alterations in membrane conductance: a potential role for alpha-synuclein oligomers in cell vulnerability." Eur J Neurosci 32(1): 10-17.
Fortin, D. L., M. D. Troyer, et al. (2004). "Lipid rafts mediate the synaptic localization of alpha-synuclein." J Neurosci 24(30): 6715-6723.
Furukawa, K., M. Matsuzaki-Kobayashi, et al. (2006). "Plasma membrane ion permeability induced by mutant alpha-synuclein contributes to the degeneration of neural cells." J Neurochem 97(4): 1071-1077.
Garcia-Mata, R., Y. S. Gao, et al. (2002). "Hassles with taking out the garbage: aggravating aggresomes." Traffic 3(6): 388-396.
Goedert, M. (2001). "Alpha-synuclein and neurodegenerative diseases." Nat Rev Neurosci 2(7): 492-501.
Goedert, M. (2001). "The significance of tau and alpha-synuclein inclusions in neurodegenerative diseases." Curr Opin Genet Dev 11(3): 343-351.

142
Goldberg, M. S. and P. T. Lansbury, Jr. (2000). "Is there a cause-and-effect relationship between alpha-synuclein fibrillization and Parkinson's disease?" Nat Cell Biol 2(7): E115-119.
Hamill, O. P. and B. Sakmann (1981). "Multiple conductance states of single acetylcholine receptor channels in embryonic muscle cells." Nature 294(5840): 462-464.
Hashimoto, M., L. J. Hsu, et al. (1999). "Oxidative stress induces amyloid-like aggregate formation of NACP/alpha-synuclein in vitro." Neuroreport 10(4): 717-721.
Hettiarachchi, N. T., A. Parker, et al. (2009). "alpha-Synuclein modulation of Ca2+ signaling in human neuroblastoma (SH-SY5Y) cells." J Neurochem 111(5): 1192-1201.
Hoegger, M. J., C. J. Lieven, et al. (2008). "Differential production of superoxide by neuronal mitochondria." BMC Neurosci 9: 4.
Hoyer, W., T. Antony, et al. (2002). "Dependence of alpha-synuclein aggregate morphology on solution conditions." J Mol Biol 322(2): 383-393.
Hoyer, W., D. Cherny, et al. (2004). "Impact of the acidic C-terminal region comprising amino acids 109-140 on alpha-synuclein aggregation in vitro." Biochemistry 43(51): 16233-16242.
Ibanez, P., A. M. Bonnet, et al. (2004). "Causal relation between alpha-synuclein gene duplication and familial Parkinson's disease." Lancet 364(9440): 1169-1171.
Jankovic, J. (2008). "Parkinson's disease: clinical features and diagnosis." J Neurol Neurosurg Psychiatry 79(4): 368-376.
Kawahara, M., N. Arispe, et al. (1997). "Alzheimer's disease amyloid beta-protein forms Zn(2+)-sensitive, cation-selective channels across excised membrane patches from hypothalamic neurons." Biophys J 73(1): 67-75.
Kayed, R., E. Head, et al. (2003). "Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis." Science 300(5618): 486-489.
Kayed, R., Y. Sokolov, et al. (2004). "Permeabilization of lipid bilayers is a common conformation-dependent activity of soluble amyloid oligomers in protein misfolding diseases." J Biol Chem 279(45): 46363-46366.

143
Kopito, R. R. (2000). "Aggresomes, inclusion bodies and protein aggregation." Trends Cell Biol 10(12): 524-530.
Kruger, R., W. Kuhn, et al. (1998). "Ala30Pro mutation in the gene encoding alpha-synuclein in Parkinson's disease." Nat Genet 18(2): 106-108.
Lasagna-Reeves, C. A., C. G. Glabe, et al. (2011). "Amyloid-beta annular protofibrils evade fibrillar fate in Alzheimer disease brain." J Biol Chem 286(25): 22122-22130.
Le Foll, F., H. Castel, et al. (1998). "Gramicidin-perforated patch revealed depolarizing effect of GABA in cultured frog melanotrophs." J Physiol 507 ( Pt 1): 55-69.
Le Roy, C. and J. L. Wrana (2005). "Clathrin- and non-clathrin-mediated endocytic regulation of cell signalling." Nat Rev Mol Cell Biol 6(2): 112-126.
Lee, H. J., S. M. Baek, et al. (2011). "Dopamine promotes formation and secretion of non-fibrillar alpha-synuclein oligomers." Exp Mol Med 43(4): 216-222.
Lee, H. J., S. Patel, et al. (2005). "Intravesicular localization and exocytosis of alpha-synuclein and its aggregates." J Neurosci 25(25): 6016-6024.
Lee, S. J. (2008). "Origins and effects of extracellular alpha-synuclein: implications in Parkinson's disease." J Mol Neurosci 34(1): 17-22.
Li, J., V. N. Uversky, et al. (2001). "Effect of familial Parkinson's disease point mutations A30P and A53T on the structural properties, aggregation, and fibrillation of human alpha-synuclein." Biochemistry 40(38): 11604-11613.
Li, J. Y., E. Englund, et al. (2008). "Lewy bodies in grafted neurons in subjects with Parkinson's disease suggest host-to-graft disease propagation." Nat Med 14(5): 501-503.
Lotharius, J. and P. Brundin (2002). "Pathogenesis of Parkinson's disease: dopamine, vesicles and alpha-synuclein." Nat Rev Neurosci 3(12): 932-942.
Markram, H. and B. Sakmann (1994). "Calcium transients in dendrites of neocortical neurons evoked by single subthreshold excitatory postsynaptic potentials via low-voltage-activated calcium channels." Proc Natl Acad Sci U S A 91(11): 5207-5211.

144
Masliah, E., A. Iwai, et al. (1996). "Altered presynaptic protein NACP is associated with plaque formation and neurodegeneration in Alzheimer's disease." Am J Pathol 148(1): 201-210.
Mattson, M. P. (2007). "Calcium and neurodegeneration." Aging Cell 6(3): 337-350.
Mayor, S. and R. E. Pagano (2007). "Pathways of clathrin-independent endocytosis." Nat Rev Mol Cell Biol 8(8): 603-612.
Miller, D. W., S. M. Hague, et al. (2004). "Alpha-synuclein in blood and brain from familial Parkinson disease with SNCA locus triplication." Neurology 62(10): 1835-1838.
Murphy, D. D., S. M. Rueter, et al. (2000). "Synucleins are developmentally expressed, and alpha-synuclein regulates the size of the presynaptic vesicular pool in primary hippocampal neurons." J Neurosci 20(9): 3214-3220.
Naslund, J., J. Thyberg, et al. (1995). "Characterization of stable complexes involving apolipoprotein E and the amyloid beta peptide in Alzheimer's disease brain." Neuron 15(1): 219-228.
Nickel, W. and C. Rabouille (2009). "Mechanisms of regulated unconventional protein secretion." Nat Rev Mol Cell Biol 10(2): 148-155.
Okochi, M., J. Walter, et al. (2000). "Constitutive phosphorylation of the Parkinson's disease associated alpha-synuclein." J Biol Chem 275(1): 390-397.
Olzmann, J. A. and R. R. Kopito (2011). "Lipid droplet formation is dispensable for endoplasmic reticulum-associated degradation." J Biol Chem 286(32): 27872-27874.
Pacheco C, Aguayo LG, Opazo C (2012). “An extracellular mechanism that can explain the neurotoxic effects of α-synuclein aggregates in the brain". Front Physiol. 26;3:297.
Paris, I., J. Lozano, et al. (2008). "The catecholaminergic RCSN-3 cell line: a model to study dopamine metabolism." Neurotox Res 13(3-4): 221-230.
Park, J. Y. and P. T. Lansbury, Jr. (2003). "Beta-synuclein inhibits formation of alpha-synuclein protofibrils: a possible therapeutic strategy against Parkinson's disease." Biochemistry 42(13): 3696-3700.

145
Parodi, J. and F. Romero (2008). "Synaptic effects of low molecular weight components from Chilean Black Widow spider venom." Neurotoxicology 29(6): 1121-1126.
Parodi, J., F. J. Sepulveda, et al. (2010). "Beta-amyloid causes depletion of synaptic vesicles leading to neurotransmission failure." J Biol Chem 285(4): 2506-2514.
Pastan, I. H. and M. C. Willingham (1981). "Receptor-mediated endocytosis of hormones in cultured cells." Annu Rev Physiol 43: 239-250.
Polymeropoulos, M. H., C. Lavedan, et al. (1997). "Mutation in the alpha-synuclein gene identified in families with Parkinson's disease." Science 276(5321): 2045-2047.
Quist, A., I. Doudevski, et al. (2005). "Amyloid ion channels: a common structural link for protein-misfolding disease." Proc Natl Acad Sci U S A 102(30): 10427-10432.
Relini, A., O. Cavalleri, et al. (2009). "The two-fold aspect of the interplay of amyloidogenic proteins with lipid membranes." Chem Phys Lipids 158(1): 1-9.
Rockenstein, E., L. A. Hansen, et al. (2001). "Altered expression of the synuclein family mRNA in Lewy body and Alzheimer's disease." Brain Res 914(1-2): 48-56.
Ryan, T. A., L. Li, et al. (1996). "Synaptic vesicle recycling in synapsin I knock-out mice." J Cell Biol 134(5): 1219-1227.
Ryan, T. A., S. J. Smith, et al. (1996). "The timing of synaptic vesicle endocytosis." Proc Natl Acad Sci U S A 93(11): 5567-5571.
Schapira, A. H. (1999). "Science, medicine, and the future: Parkinson's disease." BMJ 318(7179): 311-314.
Schmidt, F., J. Levin, et al. (2012). "Single-channel electrophysiology reveals a distinct and uniform pore complex formed by alpha-synuclein oligomers in lipid membranes." PLoS One 7(8): e42545.
Sepulveda, F. J., J. Parodi, et al. (2010). "Synaptotoxicity of Alzheimer beta amyloid can be explained by its membrane perforating property." PLoS One 5(7): e11820.

146
Shankar, G. M., S. Li, et al. (2008). "Amyloid-beta protein dimers isolated directly from Alzheimer's brains impair synaptic plasticity and memory." Nat Med 14(8): 837-842.
Singleton, A. B., M. Farrer, et al. (2003). "alpha-Synuclein locus triplication causes Parkinson's disease." Science 302(5646): 841.
Spillantini, M. G., M. L. Schmidt, et al. (1997). "Alpha-synuclein in Lewy bodies." Nature 388(6645): 839-840.
Tajima, Y., K. Ono, et al. (1996). "Perforated patch-clamp recording in cardiac myocytes using cation-selective ionophore gramicidin." Am J Physiol 271(2 Pt 1): C524-532.
Tsigelny, I. F., P. Bar-On, et al. (2007). "Dynamics of alpha-synuclein aggregation and inhibition of pore-like oligomer development by beta-synuclein." FEBS J 274(7): 1862-1877.
Tsigelny, I. F., L. Crews, et al. (2008). "Mechanisms of hybrid oligomer formation in the pathogenesis of combined Alzheimer's and Parkinson's diseases." PLoS One 3(9): e3135.
Tsika, E., M. Moysidou, et al. (2010). "Distinct region-specific alpha-synuclein oligomers in A53T transgenic mice: implications for neurodegeneration." J Neurosci 30(9): 3409-3418.
Ueda, K., H. Fukushima, et al. (1993). "Molecular cloning of cDNA encoding an unrecognized component of amyloid in Alzheimer disease." Proc Natl Acad Sci U S A 90(23): 11282-11286.
Ulmer, T. S., A. Bax, et al. (2005). "Structure and dynamics of micelle-bound human alpha-synuclein." J Biol Chem 280(10): 9595-9603.
Uversky, V. N. and A. L. Fink (2002). "Amino acid determinants of alpha-synuclein aggregation: putting together pieces of the puzzle." FEBS Lett 522(1-3): 9-13.
Uversky, V. N., H. J. Lee, et al. (2001). "Stabilization of partially folded conformation during alpha-synuclein oligomerization in both purified and cytosolic preparations." J Biol Chem 276(47): 43495-43498.
Uversky, V. N., J. Li, et al. (2001). "Metal-triggered structural transformations, aggregation, and fibrillation of human alpha-synuclein. A possible molecular NK between Parkinson's disease and heavy metal exposure." J Biol Chem 276(47): 44284-44296.

147
Uversky, V. N., J. Li, et al. (2001). "Pesticides directly accelerate the rate of alpha-synuclein fibril formation: a possible factor in Parkinson's disease." FEBS Lett 500(3): 105-108.
van Rooijen, B. D., M. M. Claessens, et al. (2010). "Membrane interactions of oligomeric alpha-synuclein: potential role in Parkinson's disease." Curr Protein Pept Sci 11(5): 334-342.
Vekrellis, K., M. Xilouri, et al. (2009). "Inducible over-expression of wild type alpha-synuclein in human neuronal cells leads to caspase-dependent non-apoptotic death." J Neurochem 109(5): 1348-1362.
Volles, M. J. and P. T. Lansbury, Jr. (2003). "Zeroing in on the pathogenic form of alpha-synuclein and its mechanism of neurotoxicity in Parkinson's disease." Biochemistry 42(26): 7871-7878.
Volles, M. J., S. J. Lee, et al. (2001). "Vesicle permeabilization by protofibrillar alpha-synuclein: implications for the pathogenesis and treatment of Parkinson's disease." Biochemistry 40(26): 7812-7819.
Wood, S. J., J. Wypych, et al. (1999). "alpha-synuclein fibrillogenesis is nucleation-dependent. Implications for the pathogenesis of Parkinson's disease." J Biol Chem 274(28): 19509-19512.
Zakharov, S. D., J. D. Hulleman, et al. (2007). "Helical alpha-synuclein forms highly conductive ion channels." Biochemistry 46(50): 14369-14379.
Zarranz, J. J., J. Alegre, et al. (2004). "The new mutation, E46K, of alpha-synuclein causes Parkinson and Lewy body dementia." Ann Neurol 55(2): 164-173.
Zhang, W., T. Wang, et al. (2005). "Aggregated alpha-synuclein activates microglia: a process leading to disease progression in Parkinson's disease." FASEB J 19(6): 533-542.