
East Gondwana
17
The oldest bivalved arthropods from the early Cambrian of East Gondwana: Systematics, biostratigraphy and biogeography Timothy P. Topper a, ⁎, Christian B. Skovsted b,c , Glenn A. Brock a , John R. Paterson d a Department of Biological Sciences, Macquarie University, NSW 2109, Australia b Department of Earth Sciences, Uppsala University, Villavägen 16, SE-752 36 Uppsala, Sweden c Department of Palaeozoology, Swedish Museum of Natural History, Box 50007, SE-104 05 Stockholm, Sweden d Division of Earth Sciences, School of Environmental & Rural Science, University of New England, Armidale NSW 2351, Australia abstract article info Article history: Received 15 February 2010 Received in revised form 30 April 2010 Accepted 23 May 2010 Available online 4 June 2010 Keywords: Cambrian Arthropoda Bradoriida Phosphatocopida South Australia The oldest bradoriid fauna from Australia, occurring in the lower Cambrian Ajax and Wirrapowie limestones of the Flinders Ranges, South Australia consists of eleven taxa, including one new genus and species, Quadricona madonnae gen. et sp. nov. and two new species, Liangshanella circumbolina sp. nov. and Zepaera jagoi sp. nov. In the Ajax Limestone, Liangshanella circumbolina sp. nov. occurs c. 20 m below the FAD of the zonal trilobite Abadiella huoi. This pre-trilobitic occurrence represents the oldest bivalved arthropod hitherto known from East Gondwana and suggests a lower Cambrian (Series 2, Stage 3) age for the assemblage. The recognition of distinct bradoriid assemblages associated with the Abadiella huoi (Atdabanian), Pararaia tatei, P. bunyerooensis and P. janeae (all Botoman) trilobite biozones in South Australia indicates great potential for future regional biostratigraphic correlation. Quantitative biogeographic analysis including new taxonomic data from the lower Cambrian of South Australia, highlights the strong endemism displayed by early Cambrian bradoriid communities and strengthens the close faunal affinities with South China and Antarctica. © 2010 International Association for Gondwana Research. Published by Elsevier B.V. All rights reserved. 1. Introduction Bradoriids are small, bivalved, marine arthropods that formed an important component of Cambrian faunal assemblages before their extinction in the middle Ordovician (e.g. Melnikova et al., 1997; Siveter and Williams, 1997; Hou et al., 2002; Vannier et al., 2005; Zhang, 2007; Jones and Kruse, 2009). The group had a worldwide distribution and are present in all of the major Cambrian Lagerstätten, including the Buen Formation (Siveter et al., 1996), Qiongzhusi Formation (Hou and Bergström, 1991; Hou et al., 2002), Burgess Shale Formation (Conway Morris, 1986; Siveter and Williams, 1997) and the Alum Shale Formation (e.g. Müller, 1964, 1979, 1982; Maas et al., 2003). Historically, bradoriids were suggested to be the ancestors of ostracods (Sylvester Bradley, 1961; Hinz-Schallreuter, 1993a, b, 1999; Zhang and Pratt, 1993; McKenzie et al., 1999; Gozalo and Hinz-Schallreuter, 2002), but investigation of appendage morphology in exceptionally preserved specimens of Kunmingella Huo, 1956 and Kunyangella Huo, 1956 from China (Hou et al., 1996, 2002; Shu et al., 1999; Hou et al., 2010) suggests that bradoriids share few synapomorphies with ostracods and are also not directly related to the other widespread group of Cambrian bivalved arthropods, the Phosphatocopida (see also Hinz-Schallreuter and Schallreuter, 2009). Current evidence suggests phosphatocopids are a sister group to the Eucrustacea, a group that includes all crustacean taxa with extant derivatives (Maas et al., 2003; Jones and Laurie, 2006; Williams et al., 2007, 2008). The possession of five head appendages in Kunmingella, accompanied by endopods that consist of fewer than seven podomeres indicate that bradoriids most probably represent stem group crustaceans (Hou et al., 1996, 2010; Shu et al., 1999; Williams et al., 2007, 2008). A total of 41 bradoriid and 12 phosphatocopid genera have been described from latest early and middle Cambrian [Series 2 (Stage 4) and Series 3 (Stage 5 to Guzhangian)] equivalent successions across central and northern Australia (Öpik, 1961, 1963, 1967, 1968; Fleming, 1973; Jones and McKenzie, 1980; Hinz, 1991, 1992a,b, 1993; Hinz and Jones, 1992; Hinz-Schallreuter, 1993a, 1999; Hinz-Schallreuter and Jones, 1994; Jones and Laurie, 2006; Jones and Kruse, 2009). In stark contrast, the diversity and abundance of bivalved arthropods from the thick lower Cambrian successions of South Australia have, until recently, been seriously neglected. The authors have recently documented 13 bradoriid and a single phosphatocopid species from the lower Cambrian (Series 2, Stage 4) Mernmerna Formation in the Arrowie Basin (Skovsted Gondwana Research 19 (2011) 310–326 ⁎ Corresponding author. Tel.: +61 2 9850 7719; fax: +61 2 9850 6053. E-mail address: [email protected] (T.P. Topper). 1342-937X/$ – see front matter © 2010 International Association for Gondwana Research. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.gr.2010.05.012 Contents lists available at ScienceDirect Gondwana Research journal homepage: www.elsevier.com/locate/gr
-
Upload
maximillian-heartwood -
Category
Documents
-
view
41 -
download
1
description
East Gondwana
Transcript of East Gondwana

Gondwana Research 19 (2011) 310–326
Contents lists available at ScienceDirect
Gondwana Research
j ourna l homepage: www.e lsev ie r.com/ locate /gr
The oldest bivalved arthropods from the early Cambrian of East Gondwana:Systematics, biostratigraphy and biogeography
Timothy P. Topper a,⁎, Christian B. Skovsted b,c, Glenn A. Brock a, John R. Paterson d
a Department of Biological Sciences, Macquarie University, NSW 2109, Australiab Department of Earth Sciences, Uppsala University, Villavägen 16, SE-752 36 Uppsala, Swedenc Department of Palaeozoology, Swedish Museum of Natural History, Box 50007, SE-104 05 Stockholm, Swedend Division of Earth Sciences, School of Environmental & Rural Science, University of New England, Armidale NSW 2351, Australia
⁎ Corresponding author. Tel.: +61 2 9850 7719; fax:E-mail address: [email protected] (T.P. Top
1342-937X/$ – see front matter © 2010 International Adoi:10.1016/j.gr.2010.05.012
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 February 2010Received in revised form 30 April 2010Accepted 23 May 2010Available online 4 June 2010
Keywords:CambrianArthropodaBradoriidaPhosphatocopidaSouth Australia
The oldest bradoriid fauna from Australia, occurring in the lower Cambrian Ajax and Wirrapowielimestones of the Flinders Ranges, South Australia consists of eleven taxa, including one new genus andspecies, Quadricona madonnae gen. et sp. nov. and two new species, Liangshanella circumbolina sp. nov.and Zepaera jagoi sp. nov. In the Ajax Limestone, Liangshanella circumbolina sp. nov. occurs c. 20 m belowthe FAD of the zonal trilobite Abadiella huoi. This pre-trilobitic occurrence represents the oldest bivalvedarthropod hitherto known from East Gondwana and suggests a lower Cambrian (Series 2, Stage 3) age forthe assemblage. The recognition of distinct bradoriid assemblages associated with the Abadiella huoi(Atdabanian), Pararaia tatei, P. bunyerooensis and P. janeae (all Botoman) trilobite biozones in SouthAustralia indicates great potential for future regional biostratigraphic correlation. Quantitativebiogeographic analysis including new taxonomic data from the lower Cambrian of South Australia,highlights the strong endemism displayed by early Cambrian bradoriid communities and strengthens theclose faunal affinities with South China and Antarctica.
© 2010 International Association for Gondwana Research. Published by Elsevier B.V. All rights reserved.
1. Introduction
Bradoriids are small, bivalved, marine arthropods that formedan important component of Cambrian faunal assemblages beforetheir extinction in the middle Ordovician (e.g. Melnikova et al.,1997; Siveter and Williams, 1997; Hou et al., 2002; Vannier et al.,2005; Zhang, 2007; Jones and Kruse, 2009). The group had aworldwide distribution and are present in all of the majorCambrian Lagerstätten, including the Buen Formation (Siveter etal., 1996), Qiongzhusi Formation (Hou and Bergström, 1991; Hou etal., 2002), Burgess Shale Formation (Conway Morris, 1986; Siveterand Williams, 1997) and the Alum Shale Formation (e.g. Müller,1964, 1979, 1982; Maas et al., 2003). Historically, bradoriids weresuggested to be the ancestors of ostracods (Sylvester Bradley,1961; Hinz-Schallreuter, 1993a, b, 1999; Zhang and Pratt, 1993;McKenzie et al., 1999; Gozalo and Hinz-Schallreuter, 2002), butinvestigation of appendage morphology in exceptionally preservedspecimens of Kunmingella Huo, 1956 and Kunyangella Huo, 1956from China (Hou et al., 1996, 2002; Shu et al., 1999; Hou et al.,2010) suggests that bradoriids share few synapomorphies with
+61 2 9850 6053.per).
ssociation for Gondwana Research.
ostracods and are also not directly related to the other widespreadgroup of Cambrian bivalved arthropods, the Phosphatocopida (seealso Hinz-Schallreuter and Schallreuter, 2009). Current evidencesuggests phosphatocopids are a sister group to the Eucrustacea, agroup that includes all crustacean taxa with extant derivatives(Maas et al., 2003; Jones and Laurie, 2006; Williams et al., 2007,2008). The possession of five head appendages in Kunmingella,accompanied by endopods that consist of fewer than sevenpodomeres indicate that bradoriids most probably represent stemgroup crustaceans (Hou et al., 1996, 2010; Shu et al., 1999;Williams et al., 2007, 2008).
A total of 41 bradoriid and 12 phosphatocopid genera have beendescribed from latest early and middle Cambrian [Series 2 (Stage 4)and Series 3 (Stage 5 to Guzhangian)] equivalent successionsacross central and northern Australia (Öpik, 1961, 1963, 1967,1968; Fleming, 1973; Jones and McKenzie, 1980; Hinz, 1991,1992a,b, 1993; Hinz and Jones, 1992; Hinz-Schallreuter, 1993a,1999; Hinz-Schallreuter and Jones, 1994; Jones and Laurie, 2006;Jones and Kruse, 2009). In stark contrast, the diversity andabundance of bivalved arthropods from the thick lower Cambriansuccessions of South Australia have, until recently, been seriouslyneglected. The authors have recently documented 13 bradoriid anda single phosphatocopid species from the lower Cambrian (Series 2,Stage 4) Mernmerna Formation in the Arrowie Basin (Skovsted
Published by Elsevier B.V. All rights reserved.

311T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
et al., 2006; Topper et al., 2007) adding substantially to thediversity of bivalved arthropods previously documented fromthe Parara Limestone in the Stansbury Basin (Chapman, 1918;Bengtson et al., 1990).
Bivalved arthropods co-occur with the first trilobites on severalcontinents (Williams et al., 2007), but appear slightly earlier thanthe first recorded trilobite (Abadiella) in southern China (Hou et al.,2002; Williams et al., 2007; Zhang, 2007; Zhang, et al., 2008). Thefaunal abundance and early stratigraphic occurrence of bivalvedarthropods in South China has prompted some authors (Hou et al.,2002; Williams et al., 2007) to suggest that bradoriids mayhave originated in the South China region. The oldest bradoriidspreviously documented from South Australia were possiblesvealutids from the lower Cambrian (Atdabanian equivalent)Parara Limestone at Curramulka Quarry on Yorke Peninsula(Chapman, 1918). Studies of small shelly fossil (SSF) assemblagesfrom the Parara Limestone on Yorke Peninsula have also estab-lished the presence of species belonging to the Hipponicharionidae(Albrunnicola bengtsoni Hinz-Schallreuter, 1993a) and Monaster-iidae (Epactridion portax Bengtson in Bengtson et al., 1990). Thesebradoriids occur within the Abadiella huoi trilobite Biozone, whichbroadly correlates with the Abadiella trilobite Zone in South China(Jell in Bengtson et al., 1990; Steiner et al., 2001; Zang et al., 2001;Paterson and Brock, 2007) and the Atdabanian Stage of Siberia(Zhuravlev and Gravestock, 1994; Zhuravlev, 1995). The oldestphosphatocopid in Australia (Indianidae gen. et sp. indet. A) hasalso been documented from the Abadiella huoi Biozone in the AjaxLimestone, South Australia (Bengtson et al., 1990).
Detailed collections along measured stratigraphic sectionsthrough the Ajax Limestone reveal that the oldest bradoriid occursc. 20 m below the first appearance datum (FAD) of the zonaltrilobite Abadiella huoi. This pre-trilobitic occurrence suggests anearly Series 2, Stage 3 age and represents the oldest bivalvedarthropod hitherto known from the lower Cambrian of EastGondwana. In a further contribution to the understanding ofearly Cambrian bivalved arthropod biodiversity from East Gond-wana, three new bradoriid species are described herein from thelower Cambrian Ajax Limestone (Series 2, Stage 3) in the MountScott Range and the upper part of the Wirrapowie Limestone(Series 2, Stages 3-4) in the Elder Range. The new data, whencombined with the recent documentation of a number of SouthAustralian bivalved arthropod assemblages (e.g. Skovsted et al.,2006; 2009; Topper et al., 2007) provides the opportunity to shednew light on the biogeography, biodiversity and evolution of earlyCambrian South Australian assemblages. A quantitative analysis ofAustralian and East Gondwanan bradoriid biogeography is provid-ed, using a pair-group cluster analysis for presence-absence data(Raup-Crick similarity index) utilising the statistical package ‘PAST’(Hammer et al., 2001).
2. Localities, biostratigraphy and age
2.1. Localities
The bivalved arthropods described herein were collected fromtwo lower Cambrian (Series 2, Stages 3-4) stratigraphic sections inthe Arrowie Basin (Figs. 1, 2). The first stratigraphic section (AJX-M) was measured through the Ajax Limestone outcropping in theMount Scott Range, northern Flinders Ranges, South Australia(Figs. 1B, 2A). The base of the AJX-M section is located atcoordinates 30°35'49" S, 138°19'59.3" E [WGS84] and is equivalentto section M of Gravestock (1984, fig. 2). The stratigraphy andsedimentology of the Ajax Limestone at section AJX-M has beensummarised by Brock et al. (2006) and Skovsted et al. (2009); thelatter paper documents specimens of the early Cambrian stemgroup brachiopod Mickwitzia from the upper part of the section.
The carbonate-dominated Ajax Limestone is approximately 280 mthick at AJX-M and conformably overlies the siliciclastic ParachilnaFormation (Fig. 2A). The lower 120 m of AJX-M section isdominated by massive stromatolitic boundstones and dolomitisedbioclastic and cryptalgal, laminated limestones. Shelly fossils aregenerally absent from this part of the section. The upper 160 m ofsection consists of richly fossiliferous (frequently silicified) bedsincluding grey to buff coloured, thin nodular limestone beds andred, massive, bioclastic limestone with minor nodular interbeds(Skovsted et al., 2009).
The second section, ER-9, was measured through the Wirrapo-wie Limestone and conformably overlying Mernmerna Formationoutcropping on the eastern limb of a north plunging syncline in thenorthern Elder Range (Figs. 1C, 2B). The base of the ER-9 section isat coordinates 31°40'2.6"S; 138°26'11.5"E [WGS84]. The upper47 m of massive, algal dominated, mottled and ribboned carbo-nates of the Wirrapowie Limestone was sampled at section ER-9,with sampling continuing into the Mernmerna Formation. Theconformably overlying Mernmerna Formation reaches a maximumthickness of 191 m in ER-9 and is dominanted by argillitic tonodular limestones interbedded with calcareous mudstone in thelower two-thirds of the exposure, grading into a calcareousmudstone with minor limestone nodules in the upper 60 m ofoutcrop. The majority of bivalved arthropod specimens are derivedfrom the upper Wirrapowie Limestone; only a few specimens arepresent in the overlying Mernmerna Formation (Fig. 2B).
2.2. Biostratigraphy and age
The International Submission on Cambrian Stratigraphy (ISCS)has recently adopted a four-Series, ten-Stage framework for theCambrian chronostratigraphic timescale (Babcock et al., 2005;Babcock and Peng, 2007; Peng and Babcock, 2008; Babcock et al.,2009). Whilst the majority of stages are undefined and keyhorizons are yet to be resolved (Babcock and Peng, 2007), twoSeries (Terreneuvian and Furongian) and four stages (Fortunian,Drumian, Guzhangian and Paibian) have been ratified (Babcock etal., 2009). The traditional lower Cambrian has been replaced bytwo Series and four Stages, with only the base of the Cambriansystem, currently defined (Brasier et al., 1994; Landing, 1994;Landing et al., 2007).
The fossiliferous part of the Ajax Limestone as represented insection AJX-M broadly coincides with the currently unnamedCambrian Series 2, Stages 3-4. These stages are approximatelyequivalent to the Atdabanian and Botoman of the Siberian stagenomenclature. The FAD of Abadiella huoi, the eponym of the oldestAustralian trilobite Biozone, at 137 m above the base of the sectionmarks the first occurrence of a trilobite taxon in the AjaxLimestone, and most likely falls within Cambrian Series 2, Stage 3(Babcock et al., 2005; Babcock and Peng, 2007; Babcock et al.,2009). Jell (in Bengtson et al., 1990) has described fragmentaryspecimens of “Redlichiid indet. 3” – possibly representing a speciesof Redlichia (cf. Paterson and Brock, 2007) – from a stratigraphiclevel that coincides with the Pararaia tatei Biozone in the AjaxLimestone. Thus, the upper Ajax Limestone that equates to theP. tatei Biozone may be early Stage 4 in age, given that one of theproposed candidate horizons to mark the base of this stage is theFAD of Redlichia (Babcock et al., 2005; Babcock and Peng, 2007).Consequently, the entire Ajax Limestone at section AJX-M maypotentially span from the late Terreneuvian Series, Stage 2 throughto the unnamed Cambrian Series 2, Stage 4. Ongoing investigationsto determine the FAD and LAD of SSFs, brachiopods and molluscs(see Topper et al., 2009), in conjunction with chemostratigraphicanalyses, will help to constrain the position of Series and Stageboundaries in the lower Cambrian succession of South Australia.

Fig. 1. A, locality map showing study areas, the Mount Scott and Elder ranges, Flinders Ranges, South Australia. B, simplified geological map of the Mount Scott Range showing thepositions of section AJX-M through the Ajax Limestone. C, geological map of the Elder Range showing position of the ER9 section through the Hawker Group.
312 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
The AJX-M section contains an abundance of silicified, phos-phatic or phosphatised macro- and microfossils, including speciesof trilobites, lingulate and rhynchonelliform brachiopods,molluscs, tommotiids, problematic small shelly fossils, spongio-morphs, plus eight species of bivalved arthropods documentedherein. Archaeocyaths from the AJX-M section were described byGravestock (1984; his section M) and Bengtson et al. (1990, fig. 6)documented a variety of taxa including trilobites, molluscs,sponge spicules and SSFs from spot localities along this creeksection. Recently, Skovsted et al. (2009) described the first EastGondwanan occurrence of the stem group brachiopod Mickwitziafrom this locality. The stratigraphic ranges of trilobites and
bivalved arthropods documented through the entire AJX-Mstratigraphic section are plotted against the lithostratigraphiccolumn in Fig. 2A. Two distinct trilobite biozones can bedifferentiated in the AJX-M section (see Jago et al., 2006 for themost recent review of the biostratigraphy of the lower Cambrian ofSouth Australia). The first appearance datum (FAD) of the indextrilobite taxon Abadiella huoi at 137 m true thickness above thebase of the section defines the base of the eponymous biozone. TheLAD of A. huoi occurs at 197.67 m above the base of the sectionindicating that the entire A. huoi Biozone is 60.67 m thick in thesection. The base of the succeeding Pararaia tatei trilobite Biozone,defined by the FAD of the eponymous species, is located at

Fig. 2. A, stratigraphic ranges of trilobite and bivalved arthropod taxa through the Ajax Limestone in the AJX-M section, Mount Scott Range. B, stratigraphic ranges of stem groupbrachiopod, trilobite, and bivalved arthropod taxa through the upper Wirrapowie Limestone and Mernmerna Formation in the ER-9 section in the Elder Range.
313T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
199.63 m (true thickness) above the base of the section and 1.96 mabove the last appearance datum (LAD) of Abadiella huoi. Thisleaves a small stratigraphic gap of 1.96 m, where it is difficult toascertain the exact boundaries of the two trilobite biozones. ThePararaia tatei Biozone represents at least a 39.7 m stratigraphicinterval in the upper half of the section, with the LAD of theeponymous species occurring at 239.33 m above the base of thesection, which equates with a distinctive erosional surfaceundoubtedly reflecting a short hiatus in sedimentation. No trilobitespecimens were recovered from the red and grey dolomotised,
stromatolitic limestones representing the top 40 metres of sectionAJX-M, and the presence of the normally much older tommotiidsMicrina etheridgei and Dailyatia ajax in these beds strongly suggestssubstantial reworking (or fault repetition) in this interval.
The FAD of the oldest bradoriid taxon, Liangshanella circumbolinasp. nov., occurs at 117 m above the base of the section, c. 20 mbelow the FAD of Abadiella huoi. Despite detailed samplingthroughout the AJX-M section, we acknowledge that pervasivedolomitisation has destroyed original fabrics, and so uncertaintiesremain about the presence (or not) of trilobites (and other fossils)

314 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
in the lower dolomitised part of the Ajax Limestone succession. Ifthe pre-trilobitic occurrence of Liangshanella circumbolina sp. nov. isaccepted, then this suggests an early Series 2, Stage 3 age for thistaxon, representing the oldest bivalved arthropod hitherto describedfrom the lower Cambrian succession of South Australia. The rest ofthe bivalved arthropod assemblage (7 species in total – see Fig. 2A)from AJX-M, correlates with the A. huoi Biozone and is stillsignificantly older that the bivalved arthropods recently documen-ted from the Mernmerna Formation (P. bunyerooensis and P. janeaebiozones) in the central Flinders Ranges by Skovsted et al. (2006)and Topper et al. (2007). Only two species, Mongolitubulus squamiferMissarzhevsky, 1977 and Zepaera sp. range into the succeedingPararaia tatei Biozone (Fig. 2A).
Two species that occur in the Ajax Limestone have beenpreviously documented from South Australia: Dabashanellahemicyclica Huo, Shu and Fu in Huo et al., 1983, from the Pararaiabunyerooensis Biozone of the Mernmerna Formation (Skovstedet al., 2006) and Mongolitubulus squamifer from the P. janeaeBiozone of the Mernmerna Formation (Topper et al., 2007).Unfortunately, since both are long-ranging taxa they have limitedbiostratigraphic value. Other long-ranging taxa with low biostrati-graphic utility in the AJX-M and ER-9 sections include genericforms Euzepaera, Zepaera. and Mongolitubulus. Specimens of eachgenus from AJX-M and ER-9 are either too poorly preserved (e.g.Euzepaera sp. and Zepaera sp.) or display substantial morphologicalvariation and fragmentation (e.g. Mongolitubulus sp.) that pre-cludes assessment at species level.
The bradoriid assemblage at section ER-9 is mainly restricted tothe Wirrapowie Limestone, which equates to the basal 47 m (truethickness) of the section. The dearth of trilobite specimensthroughout the ER-9 stratigraphic section does not allow preciseplacement of the zonal trilobite boundaries. The first trilobiterecovered in the section, Alanisia guillermoi Richter and Richter,1940 occurs 106 m above the base of the section and is generallytaken to represent the Pararaia tatei Biozone (Jell in Bengtson et al.,1990). Although no trilobites indicative of the Abadiella huoiBiozone were recovered from the basal 47 m of the ER-9 section,these beds do contain typical Atdabanian representativesincluding the tannuoliniid tommotiid Micrina etheridgei Tate,1892 and the paterinid brachiopod Askepasma cf. toddense, whichwidely co-occur with Abadiella huoi elsewhere in the Arrowie Basinand provide strong evidence in support of an A. huoi Biozoneequivalent age for the bradoriid assemblage in the WirrapowieLimestone at section ER-9. The occurrence of Quadricona madonnaegen. et sp. nov. provides additional correlation with section AJX-Mand supports a A. huoi equivalent biostratigraphic age for theassemblage.
Two species, Alutella sp. and Spinospitella coronata Skovsted,Brock and Paterson, 2006 are also present in the overlyingMernmerna Formation at ER-9. Spinospitella coronata co-occurswith the P. tatei Biozone trilobite A. guillermoi and continues torange 25 m (true thickness) above the occurrence of A. guillermoi.Alutella sp. is confined to the uppermost part of the section whichcan be confidently assigned a Pararaia janeae Biozone biostrati-graphic age, based on the presence of the eponym along withSerrodiscus gravestocki Jell in Bengtson et al., 1990, Hebediscinayuqingensis Zhang in Zhang et al., 1980, Atops rupertensis Jell, Jagoand Gehling, 1992, and a possible edelsteinaspidid.
3. Biogeographic implications
The distribution of bradoriids, and to a much lesser extentphosphatocopids, has often been utilised in Cambrian palaeobio-geographical analyses (e.g. Melnikova et al., 1997; Siveter andWilliams, 1997; Williams and Siveter, 1998; Hou et al., 2002). Shu(1981), in an early biogeographical review of bradoriids, identified
a ‘western’ faunal realm, characterized by the genera Indiana andBradoria and an ‘eastern’ faunal realm, characterized by the socalled ‘alutids’ (e.g. Zepaera, Liangshanella and Kunmingella). Theterm ‘alutids’ was used to describe bradoriids with apparent outerborder structures on their shields (Shu and Chen, 1994). However,recent taxonomic revisions have demonstrated that the nominatetaxa Aluta should be considered a nomen dubium and the term‘alutids’ be abandoned (Siveter and Williams, 1997; Williams et al.,2007).
Palaeobiogeographic studies have tended to concentrate oncomparisons of Cambrian bradoriid biogeographical patterns withthat of trilobites (e.g. Melnikova et al., 1997; Hou et al., 2002;Williams et al., 2007; Zhang, 2007). The ‘western’ faunal realmidentified by Shu (1981) is generally associated with cool to coldwater systems broadly mirroring the Olenellid trilobite Provincewhilst the ‘eastern’ realm reflects warm water, largely equatorialcommunities equivalent to the Redlichiid trilobite province(Melnikova et al., 1997; Hou et al., 2002; Williams et al., 2007).Bradoriid taxa characteristic of the Redlichiid trilobite provinceinclude Shangsiella, Comptaluta and Zepaera, whereas others, suchas Indiana, Walcottella and Dielymella, epitomize the Olenellidtrilobite province. The trilobite-based Acadobaltic Province sensuSdzuy (1972), a region that includes Baltica, Avalonia, Armoricaand Morocco also displays strong provinciality. This is supported bythe presence of genera such as Beyrichona Matthew, 1886, Indianaand Wimanicharion Hinz-Schallreuter, 1993a, which are almostexclusively known from these areas in the late early and middleCambrian (Streng et al., 2008). However, uncertainties have arisenregarding the degree of differentiation between the olenellid andredlichiid provinces (Zhang, 2003; Palmer, 2005). Zhang (2003)and Palmer (2005) have noted that some early Cambrian trilobitesfrom both provinces display similarities that suggest that faunalprovinciality was not as well developed in the early Cambrian.
The use of the term ‘western’ and ‘eastern’ was abandoned byShu and Chen (1994) who established the “European” realm andthe “4A” realm, based predominantly on temperature-latitudegradients. The “European” faunal realm consisted of Baltica,Avalonia, northern France and North Africa and represented acool to cold water fauna and the “4A” faunal realm consisted ofLaurentia, Asia (North and South China), Siberia, Tarim,Kazakhstan, Mongolia, Australia and Antarctica, representing anequatorial, warm water fauna (Shu and Chen, 1994). Shu and Chen(1994) suggested a number of possible dispersal routes forbradoriids, including migration from South China to Australiaduring the late early Cambrian, based on the presence of Zepaeraon both palaeocontinents. The late early Cambrian timing of themigration was based solely on the lack of bradoriid taxa from theearly Cambrian of Australia. Zepaera has subsequently beendiscovered in the early Cambrian of Australia (Skovsted et al.,2006; Topper et al., 2007; herein) in conjunction with a diversesuite of bradoriid genera, suggesting that if a migration event didoccur between South China and Australia, it must have transpiredmuch earlier than previously recognised. Williams et al. (2007)broadly supported the idea of ‘western’ and ‘eastern’ faunal realms;however, stressed that the patterns of bradoriid biogeography wasmuch more complicated than previous works indicated; theyidentified a number of distribution patterns and assemblages,reflecting tropical and temperate climates as well as those groupsthat had a cosmopolitan distribution.
Williams et al. (2007) provided a detailed account of theaffinities and biogeography of bradoriids for the duration of theCambrian until their eventual extinction in the mid-Ordovician.Williams et al. (2007) reviewed the bradoriid faunas from anumber of palaeocontinental regions including Laurentia, Avalonia,Siberia, South China, central Asia and Gondwana commenting onthe palaeogeographical positions of these regions pertaining to

Fig. 3. A, reproduced cluster analysis of early Cambrian bradoriid genera conducted by Williams et al. (2007, fig. 10A, page 224). B, pair-group cluster analysis for presence-absencedata (Raup-Crick similarity index, correlation coefficient 0.6587) for 60 bradoriid genera from the early Cambrian. C, pair-group cluster analysis for presence-absence data (Raup-Crick similarity index, correlation coefficient 0.6684) with Australia and Antarctica as amalgamated provinces of the palaeocontinental region of East Gondwana.
Fig. 4. A, pair-group cluster analysis for presence-absence data (Raup-Crick similarityindex, correlation coefficient 0.6361) for 18 bradoriid genera from the early Cambrian,omitting bradoriid genera that have been documented from only a single palaeoncon-tinental region. B, pair-group cluster analysis for presence-absence data (Raup-Cricksimilarity index, correlation coefficient 0.6876) for 18 bradoriid genera from the earlyCambrian, omitting bradoriid genera that have been documented from only a singlepalaeoncontinental region andwith Australia and Antarctica as amalgamated provincesof the palaeocontinental region of East Gondwana.
315T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
bradoriid distribution. Sedimentary successions from West Gond-wana have yielded cambriids from the lower Cambrian of France(Vannier et al., 2005), hipponicharionids from Spain (Gozalo andHinz-Schallreuter, 2002; Gozalo et al., 2004) and a number of taxafrom the middle Cambrian of Morocco (Hinz-Schallreuter, 1993a).East Gondwana yields taxa from the lower Cambrian of Antarctica(Rode et al., 2003; Wrona, 2009) along with prolific faunas fromthe lower Cambrian through to the Furongian of Australia (e.g.Öpik, 1968; Jones and McKenzie, 1980). According to Williams etal. (2007, see table 2), at the generic level, Australia produces oneof the most diverse bradoriid assemblages (23 genera documentedin total) during the Cambrian to Early Ordovician interval.
The large majority of East Gondwanan bradoriid taxa have beendescribed from the Ordian and succeeding Stages across centraland northern Australia (e.g. Öpik, 1968; Fleming, 1973; Hinz-Schallreuter, 1999). Previously, the Australian Ordian Stage wasconsidered to be early Middle Cambrian (e.g. Shergold, 1996 andreferences therein), however evidence continues to amass suggest-ing that this stage should be regarded as latest Early Cambrian (seeLaurie, 2006 and Paterson and Brock, 2007 for discussion).Consequently, all bradoriid genera described from the OrdianStage of Australia are treated as early Cambrian in age andincorporated into the following analyses. The taxonomic diversityand biogeographic analysis of bivalved arthropods reported byWilliams et al. (2007) only included two lower Cambrian(Cambrian Stage 2, Series 3-4) taxa from South Australia (Albrun-nicola bengtsoni Hinz-Schallreuter, 1993a and Epactridion portaxBengtson in Bengtson et al., 1990). Recent work by Skovsted et al.(2006), Topper et al. (2007), Wrona (2009) and herein documentanother thirteen bivalved arthropod genera from the lowerCambrian of South Australia and Antarctica, greatly increasing thedatabase of early Cambrian bivalved arthropod biodiversity fromEast Gondwana.
As part of their original evaluation of bradoriid biogeography,Williams et al. (2007, fig. 10) performed a pair-group clusteranalysis using the statistical package ‘PAST’, based on presence-absence data (Raup-Crick similarity index) of 32 bradoriid generafrom the early Cambrian and 33 genera from the middle Cambrian.The cluster analysis presented by Williams et al. (2007, fig. 10,reproduced here in Fig. 3A) indicated a wide distribution ofbradoriid genera during the early Cambrian, with close faunal
associations between Avalonia, Laurentia, Baltica and West Gond-wana (Morocco and Amorica). Bradoriid assemblages from SouthChina clustered with those of Kazakhstan as part of an easternhemisphere, warm water fauna, whilst Australian bradoriid data(based on two taxa) for the early Cambrian display only weakstatistical similarity to South China, Kazakhstan and Siberia(Fig. 3A). We have performed a number of presence-absencecluster analyses (Fig. 3B, C) using ‘PAST’ (Raup-Crick similarityindex), employing the biogeographical distribution data providedby Williams et al. (2007, table 2 and appendix A), but alsoincorporating new taxonomic data published in recent years that

316 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
was not used in the Williams et al. (2007) analysis (e.g. Skovsted,2006; Skovsted et al., 2006; Topper et al., 2007; Zhang, 2007; DiesΑlvarez et al., 2008; Hinz-Schallreuter et al., 2008; Wrona, 2009),in addition to data provided herein. Any bradoriid taxon that wasquestionably assigned at generic level have been omitted from thefollowing analysis. Given the scope of our data, the biogeographiccluster analyses were limited to the early Cambrian; resultsdiscussed below.
3.1. Bradoriid assemblages from East Gondwana
At the specific level the bradoriid assemblages from the earlyCambrian of East Gondwana are largely endemic. Strong faunalprovinciality among bradoriid species is not unusual at this timeand has been recognised by numerous authors (e.g. Siveter andWilliams, 1997; Williams et al., 2007), in particular faunas fromSouth China frequently display high levels of endemism (Hou et al.,2002; Zhang, 2007). However, several genera from East Gondwanaexhibit a wider distribution in the early Cambrian, includingLiangshanella Huo, 1956 that has a near cosmopolitan distribution.As a result of particular palaeocontinental regions displayinghigh levels of endemism during the early Cambrian (e.g. EastGondwana and South China), two cluster analyses were undertak-en (Figs. 3B-C, 4). The first analysis (Fig. 3B-C) incorporates thepresence-absence of 60 bradoriid genera from the early Cambrian.The second analysis (Fig. 4) only incorporates bradoriid generathat have been documented from two or more early Cambrianpalaeocontinental regions, thereby removing all endemic taxa.We have also run the analysis with Australia and Antarctica asseparate entities during the early Cambrian (Figs. 3B, 4A) and asamalgamated provinces of the palaeocontinental region of EastGondwana (Figs. 3C, 4B). We note that differences in taxonomicdiversity between the analysed faunas do impose some limita-tions on the cluster analysis. Some regions remain inadequatelyexplored in terms of bradoriid diversity (see Appendix), forexample, Antarctica only has four described genera for the earlyCambrian (e.g. Rode et al., 2003; Stigall, 2008; Wrona, 2009).Bradoriid diversity in other regions is much higher, such asAustralia where 21 bradoriid genera have been documented fromthe early Cambrian (e.g. Skovsted et al., 2006; Topper et al., 2007;herein). The South China terrain hosts the most diverse bradoriidfaunal assemblage during the early Cambrian with 32 generadocumented.
The first pair-group cluster analysis (Fig. 3B) displays manysimilarities with the original cluster analysis (Fig. 3A) produced byWilliams et al. (2007). Williams et al. (2007) recognised a widedistribution of congeners in the western hemisphere, betweenAvalonia, Baltica, Laurentia and West Gondwana, a view supportedby data presented here. Both analyses also illustrate an associationbetween South China and Kazakhstan. Close faunal connectionsexist between West Avalonia and West Gondwana (Morocco andArmorica) based on the presence of taxa including HipponicharionMatthew, 1886 and Matthoria Siveter and Williams, 1997. Thebradoriid fauna of Baltica displays close similarities with the faunaof West Avalonia, but also with Armorica and Morocco, supportingthe previously recognised (Gozalo et al., 2004; Dies Αlvarez et al.,2008) Acadobaltic Province sensu Sdzuy (1972). The main discrep-ancy between the analyses is the close faunal association betweenSiberia, Laurentia and East Avalonia (Fig. 3C) based on the presenceof taxa including Liangshanella Huo, 1956 and Matthoria, a groupingnot recognised in the analysis presented by Williams et al. (2007).Australia, similar to data presented Williams et al. (2007), displaysonly weak statistical similarity to Siberia, Laurentia, East Avaloniaand Antarctica.
Incorporating the early Cambrian faunal assemblages ofAustralia and Antarctica to form an East Gondwana assemblage
(Fig. 3C) produces a dendrogram that is very similar to the resultsobtained by Williams et al. (2007). The only noticeable difference isthe close association of Siberia with Laurentia and East Avalonia,discussed above and West Avalonia displaying a stronger faunalassociation with Armorica and Morocco (Fig. 3C). Recent publica-tions (e.g. Skovsted et al., 2006; Topper et al., 2007; Zhang, 2007;Hinz-Schallreuter et al., 2008; Dies Αlvarez et al., 2008; Jones andKruse, 2009; Wrona, 2009 and herein) have greatly increased thedatabase of early Cambrian bivalved arthropod biodiversity,especially from East Gondwana, where nine new genera havebeen established. Despite this increase in faunal diversity, theresulting cluster analysis (Fig. 3C) produced only minor changes tothe results of the original analysis conducted by Williams et al.(2007).
A possible explanation for the limited variation in analyses isthe high levels of endemism displayed in recently describedbradoriid assemblages (Skovsted et al., 2006; Topper et al., 2007;Zhang, 2007; Wrona, 2009). Recent documentation of earlyCambrian bradoriid assemblages from South Australia contain ahigh portion of endemic genera, including Spinospitella Skovsted,Brock and Paterson, 2006, Amphikeropsis Topper, Skovsted, Brockand Paterson, 2007, Onagrocharion Topper, Skovsted, Brock andPaterson, 2007 and Quadricona gen. nov. A diverse early Cambrianfauna from the Yu'anshan and Shuijingtuo formations of SouthChina documented by Zhang (2007) included three new endemicgenera, Retaluta, Spinaluta and Yucola. To highlight the strongendemism displayed by South Chinese faunal assemblages duringthe early Cambrian, of the total 32 genera documented, 19 areentirely restricted to the South China province. The addition ofendemic taxa into a quantitative cluster analysis would notsignificantly change the resulting biogeographic groupings of thepalaeocontinental regions; merely change the similarity indexbetween them. To gain a better understanding of the associationof faunal assemblages during the early Cambrian we decided toconduct the cluster analysis again by omitting bradoriid generathat have only been documented from a single lower Cambrianpalaeocontinental region, effectively removing all endemic taxa.
The resulting cluster analysis (Fig. 4A) displays four distinctbiogeographic groupings for the early Cambrian. Similar toprevious results, close faunal associations exist between thepalaeocontinental regions of the Acadobaltic Province (sensuSdzuy, 1972), including West Avalonia, West Gondwana (Moroccoand Armorica) and Baltica. Siberia has been biogeographicallylinked with Kazakhstan based solely on the occurrence of AlutellaKobayshi and Kato, 1951 on both palaeocontinents. Laurentianfaunal assemblages have curiously been closely associated withAntarctica, based on the presence of Albrunnicola Martinsson, 1979and Liangshanella Huo, 1956. However, this appears superficial,considering the strong evidence supporting the close earlyCambrian faunal associations of Australia and Antarctica, basedon trilobites (Palmer and Rowell, 1995; Brock et al., 2000; Paterson,2005; Paterson and Jago, 2006), brachiopods (Holmer et al., 1996;Brock et al., 2000), archaeocyaths (Zhuravlev and Gravestock,1994), molluscs (Wrona, 2003) and small shelly fossils (Evans andRowell, 1990; Wrona, 2004). The apparent close relationshipbetween Laurentia and Antarctica highlights one of the challengeswith utilising this quantitative technique as differences in taxo-nomic diversity between faunal assemblages can distort theoutcomes. Only two non-endemic bradoriid genera have beenconfirmed from Antarctica (Albrunnicola and Liangshanella), sixgenera have been documented from Laurentia, whereas twelvegenera have been documented from the early Cambrian ofAustralia (see Appendix). Despite Albrunnicola and Liangshanellaoccurring on all three continental terranes, statistically, Antarcticawill display a higher similarity to Laurentia because of the highertaxonomic diversity of Australian faunal assemblages.

Fig. 5. Liangshanella circumbolina sp. nov. All specimens from the Ajax Limestone, Mount Scott Range. A-D, holotype, SAMP44789, sample AJX-M/209.2; A, lateral view of right valve,stereo pair, scale bar 500 μm, B, interior view of right valve, scale bar 500 μm, C, external surface ornament, scale bar 10 μm, D, detail of external surface ornament, scale bar 5 μm;E, detail of external surface ornament, scale bar 5 μm, SAMP44790, sample AJX-M/209.2; F-I, SAMP44791, sample AJX-M/209.2; F, lateral view of left valve, scale bar 500 μm,G, interiorview of left valve, scale bar 500 μm, H, detail of interior of left valve, scale bar 10 μm, I, detail of sealed, tubular structures, protruding from the interior of the valve, scale bar 10 μm.
317T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
Australia and South China are shown to have a very strong faunalassociation, with a similarity index of 0.94 (Fig. 4A), a markedcontrast with the information presented by Williams et al. (2007)and by our previous analysis (Fig. 3A-B). Incorporating the earlyCambrian faunal assemblages of Australia and Antarctica to form anEast Gondwana assemblage (Fig. 4B) barely changes the originaldendrogram. East Gondwana and South China again display a verydefinitive faunal association, with a slightly stronger similarity indexthan previously of 0.96 (Fig. 4B). This result is not surprising, asbradoriid taxa such as Zepaera Fleming, 1973, Haoia Shu, 1990,Hipponicharion, Liangshanella, Parahoulongdongella Shu, 1990,Albrunnicola (Albrunnicola bengtsoni=Beyrichona chinensis Zhang,2007, p. 145) and Mongolitubulus (Mongolitubulus unispinoa=Spinella unialata) documented from the Mernmerna Formation(Skovsted et al., 2006; Topper et al., 2007) and the Ajax andWirrapowie limestones herein (Figs. 5-7), have all been documentedfrom the lower Cambrian (Eoredlichia-Wutingaspis Biozone) of China(e.g. Shu, 1990; Hou et al., 2002; Zhang, 2007). Based on similarshield morphologies, we consider Spinella unialata Zhang, 2007(pl. 17, figs. 1-8) to be conspecific with Mongolitubulus unispinosaTopper, Skovsted, Brock and Paterson, 2007 (figs. 6A-J, 7A-C) andBeyrichona chinensis Shu, 1990 documented by Zhang (2007, p. 145,pl. 17, figs. 9-16) to be conspecific with Albrunnicola bengtsoni Hinz-Schallreuter, 1993a.
The biogeographic ties between the two palaeoncontinentalregions of South China and East Gondwana has been previouslyrecognised based on trilobites (see Paterson and Brock, 2007).However, the results of the original cluster analysis (Fig. 3C) showa low similarity index between South China and East Gondwana(0.3). This evidently appears to be a direct result of differences intaxonomic diversity between the analysed faunas, due to highlevels of endemism. Ten out of the total 12 non-endemic bradoriidgenera documented from the early Cambrian of South Australia are
present in the early Cambrian of South China, further strengtheningthe biogeographic ties between the two palaeocontinental regions.The biogeographic groupings shown in Fig. 4B strongly reflect thedistribution of trilobites during the early Cambrian. South Chinaand East Gondwana are characterised by trilobites that distinguishthe Redlichiid trilobite Realm, whilst Laurentia, Avalonia andBaltica are characterised by trilobites distinguishing the Olenellidtrilobite Realm. Intervening regions, such as Siberia, Morocco,Armorica and potentially Kazakhstan are characterised by trilobitesof the transitional Bigotinid trilobite Realm (McKerrow et al., 1992;Pillola, 1993).
Early Cambrian trilobites and other non-mineralized arthropodshave recently been subjected to a number of quantitativebiogeographic analyses (e.g. Lieberman, 1997, 2003a, b; Meertand Lieberman, 2004; 2008; Hendricks and Lieberman, 2007;Z. Zhang et al., 2008). In particular, the results of Lieberman(2003a, text-fig. 2; 2003b, fig. 2) and Meert and Lieberman (2004,fig. 4) show similar major biogeographic groupings to our results,especially when compared with the cluster analysis that omittedendemic bradoriid genera and treated Australia and Antarctica as asingle region (Fig. 4B). However, it is important to note that somediscrepancies may be the result of the type of analysis conducted(phylogeographic vs. cluster, or a cladistic vs. phenetic approach,respectively) and the fact that the trilobite analyses were stronglybiased towards the inclusion of olenelline taxa, which are absent inEast Gondwana and South China. The analysis involving non-mineralized arthropods (Hendricks and Lieberman, 2007) pro-duced results that show significant differences in area relationshipsto those produced by bradoriid and trilobite data, although thegrouping of Baltica and Africa and the close relationship of Siberiaand Laurentia (Hendricks and Lieberman, 2007, fig. 2) is supportedby some of our results (Figs. 3B-C, 4A). Hendricks and Lieberman(2007) provided a number of explanations for these differences,

Fig. 6. A-F, Zepaera jagoi sp. nov. All from the Ajax Limestone, Mount Scott Range, all scale bars 200 μm, unless otherwise stated. A-C, holotype, SAMP44792, sample AJX-M/266;A, lateral view of right valve, stereo pair, B, detail of external surface ornament, scale bar 20 μm, C, detail of spinose margin, scale bar 20 μm; D, lateral view of left valve, SAMP44793,sample AJX-M/256; E-F, SAMP44794, sample AJX-M/266; E, internal view of left valve, F, internal view showing duplicature, scale bar 20 μm; G-M, Quadricona madonnae gen. et sp.nov. G-J, holotype from theMernmerna Formation, Elder Range, SAMP44795, sample ER-9/67.3; G, lateral view of left valve, H, oblique lateral view of left valve, I, oblique lateral viewfrom posterior, J, detail of external surface ornament, scale bar 10 μm; K, lateral view of right valve, scale bar 500 μm, SAMP44796, sample AJX-M/268.7; L-M, SAMP44797, sampleAJX-M/271.3; L, dorsal view of shield, scale bar 500 μm, M, oblique view of shield, scale bar 500 μm.
318 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
but it is also worth noting that their analysis included taxa rangingfrom the early Cambrian to latest Ordovician, potentially introduc-ing noise to the biogeographic signal.
Recent palaeogeographic reconstructions of the early Cambrianindicate that Australia and the South China block occupied thetropical carbonate development zone (30°±5° north and southlatitudes) (Brock et al., 2000; Meert and Lieberman, 2004; Williamset al., 2007). During the early Cambrian in South China, thedominant groups of bivalved arthropods are the kunmingellids andthe comptalutids. Kunmingellids frequently cover bedding surfaces(e.g. in the Heilinpu Formation), with the type species, Kunmingella
douvillei (Mansuy, 1912) accounting for over 80% of recoveredindividuals in the Chengjiang biota (Hou and Bergstrom, 1991; Houet al., 2002). In contrast, East Gondwanan bradoriid assemblagescontain an equal proportion of many different groups, includingthe svealutids (Liangshanella), hipponicharionids (Hipponicharionand Albrunnicola), beyrichonids (Parahoulongdongella), comptalu-tids (Zepaera, Alutella), monasteriids (Epactridion) and many othergenera that are yet to be confidently assigned at family level(e.g. Onagrocharion, Amphikeropsis and Spinospitella).
Williams et al. (2007) identified a number of faunal assem-blages that reflected tropical and temperate climates during the

Fig. 7. A-G, Parahoulongdongella bashanensis Shu, 1990. From theMernmerna Formation, Elder Range and the Ajax Limestone,Mount Scott Range, all scale bars 200 μm, unless otherwisestated. A, lateral view of right valve, SAMP44798, sample ER-9/16.5; B-D, SAMP44799, sample ER-9/16.5; B, lateral view of right valve, C, oblique lateral view of right valve, D, detail ofexternal surface ornament, scale bar 10 μm, E-G, SAMP44800, sample ER-9/53.5; E, lateral view of right valve, F, oblique lateral view of right valve, G, detail of external surface ornament,scale bar 50 μm; H, Zepaera sp., lateral view of right valve, SAMP44801, sample AJX-M/410; I,Mongolitubulus sp., lateral view of spine, SAMP44802, sample ER-9/10.8; J-K,MongolitubulussquamiferMissarzhevsky, 1977. J-K, SAMP44803, sample AJX-M/415; J, lateral view of spine, scale bar 60 μm, K, detail of external surface ornament, scale bar 20 μm; L, Comptalutid gen et.sp. indet., lateral view of right valve, scale bar 100 μm, SAMP44804, sample AJX-M/266; M, Euzepaera sp., lateral view of right valve, scale bar 500 μm, SAMP44805, sample ER-9/53.5;N, Spinospitella coronata Skovsted, Brock and Paterson, 2006, lateral viewof valve fragment, SAMP44806, sample ER-9/0;O,Dabashanella hemicyclicaHuoand Shu inHuoet al., 1983, lateralview of left valve, SAMP44807, sample AJX-M/274.4; P, Indet. phosphatocopid, lateral view of left valve, scale bar 100 μm, SAMP44808, sample AJX-M/256.5.
319T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
Cambrian. The East Gondwana bradoriid assemblage in the earlyCambrian broadly conforms to the climatic distribution patternsidentified by Williams et al. (2007). Comptalutids, prevalent inSouth Chinese and Australian assemblages are interpreted as warmwater tropical and sub-tropical bradoriids that cover a latitudinalrange from about 30° North to 15° South (Williams et al., 2007).Species of Liangshanella have been documented from earlyCambrian faunal assemblages in East Gondwana, South China,Avalonia, Laurentia and Baltica and thus seems capable of survivingin both low and mid-latitudes (Williams et al., 2007).
The beyrichonids and hipponicharionids have been recognised asmid- to high latitude bradoriids (further north and south than 30°),based on their preference for Avalonia and Baltica during theCambrian (Williams et al., 2007). Regardless, some hipponicharionids,like Neokunmingella Zhang, 1974,Meishucunella Jiang, 1982 and occur
amongst the early Cambrian faunal assemblages of South China (Houet al., 2002). Hipponicharion, a genus previously thought to be re-stricted to the Acadobaltic Province (sensu Sdzuy, 1972; Gozalo andHinz-Schallreuter, 2002; Gozalo et al., 2004) has since beendocumented from South China and South Australia (Topper et al.,2007; Zhang, 2007), extending the biogeographic range of the genusinto low latitude, tropical regions. The beyrichonids, BeyrichonaMatthew, 1886 and Parahoulongdongella have also been describedfrom the low latitude faunas of southern Kazakhstan (Melnikova et al.,1997), South Australia and South China (Topper et al., 2007; Zhang,2007). While both groups may be more abundant in Avalonian andBaltic faunas, holistically, representatives of the Hipponicharionidaeand Beyrichonidae, were evidently able to disperse between thewarmclimates of South Australia and South China and the cooler climates ofAvalonia and Baltica.

320 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
4. Systematic palaeontology
All specimens described and illustrated herein are housed in thepalaeontological collection of the South Australian Museum (acro-nym: SAMP). Stratigraphic ranges for all taxa are provided in terms oftrue thickness above the base of the AJX-M and ER-9 sections. Theterminology used to describe the bivalved arthropods largely followsthat employed to describe Ordovician and later ostracods (as adoptedin Siveter and Williams, 1997; Williams and Siveter, 1998; Hou et al.,2002). We follow Siveter et al. (2003, p. 13) in using the more neutralterm “head-shield”, or “shield” instead of the ostracod term“carapace” (see also Maas et al., 2003). The majority of additionalbivalved arthropod taxa in the Ajax andWirrapowie assemblages (seeFigs. 2, 7) have been described previously from the MernmernaFormation; refer to Skovsted et al. (2006) and Topper et al. (2007) fordescriptions and discussions.
Phylum Arthropoda Siebold and Stannius, 1845Order Bradoriida Raymond, 1935Family Svealutidae Öpik, 1968Liangshanella Huo, 1956Type species. Liangshanella liangshanensis Huo, 1956Liangshanella circumbolina sp. nov. (Fig. 5)Type material. Holotype: SAMP44789 (Fig. 5A-D) from sample
number AJX-M/209.2, 117 m true thickness above the base of thesection, Ajax Limestone, Mount Scott Range. Paratypes: SAMP44790(Fig. 5E) and SAMP44791 (Fig. 5F-I) from sample number AJX-M/209.2, 117 m true thickness above the base of the section, AjaxLimestone, Mount Scott Range.
Etymology. Dervied from the Latin prefix circum, circle and bolus,lump. In reference to the distinctive circlet of raised papillate struc-tures on the outer valve surface.
Diagnosis. Moderately to strongly inflated postplete valves witha straight hinge line. Latero-admarginal ridge present, entirebetween cardinal corners, anterodorsal or posterodorsal lobationor sulci absent. Outer surface with numerous, shallow depressionseach hosting a submicron sized, centrally located circular perfora-tion girt by a circlet of raised, closely packed papillate structures;individual depressions are sealed on the internal valve surface byraised, cylindrical pillars.
Description. Moderately to strongly inflated valves, with a straighthinge line. Postplete valves exhibiting a distinct lateroadmarginalridge, entire between cardinal corners and separated from the lateralsurface by a continuous furrow. Maximum dimensions of shield are1.48 mm in length and 1.27 mm in height. Lateral surface lacks dis-tinct anterodorsal or posterodorsal nodes or sulci. Valve surfaceconsists of numerous, small depressions, varying from 7 – 12 µm indiameter that are evenly distributed over the entire shield (Fig. 5C).Each individual depression hosts a submicron sized, centrally locatedcircular perforation (Fig. 5D-E) surrounded by a circlet of raised,closely packed papillate structures; raised circlet structure has anapproximate diameter of 4 µm (Fig. 5D-E). Internally, each depressionis sealed by raised cylindrical pillars that protrude from thevalve some 10 - 13 µm (Fig. 5H-I); neighbouring pillars can be fusedor isolated (Fig. 5H, I), depending on space between individualdepressions.
Remarks. Postplete bradoriid valves from the Cambrian thatdisplay little or no lobation are often referred to the genusLiangshanella Huo, 1956. Originally described for svealutidsthat exhibit no lobation (Huo, 1956), the generic diagnosis hassince been modified slightly to incorporate an anterior lobe thatmay range from well developed to virtually lacking (Siveter andWilliams, 1997; Hou et al., 2002). The simplicity of shieldmorphology makes it difficult to distinguish Liangshanella fromother bradoriid genera that also possess a non-lobate shield,such as Indiana Matthew, 1902, Hanchungella Huo, 1956, OvalutaZhang, 1987 and Indota Öpik, 1968. Such difficulties are exempli-
fied by Hou et al. (2002) who synonymised both Hanchungella andOvaluta with Liangshanella citing similar shield morphologies. Houet al. (2002) considered that the alleged differences in anteriorcardinal angle – traditionally used to discriminate these genera –
was the result of deformational processes, and thus of notaxonomic significance. Zhang (2007) disputed this synonymyand suggested that a lack of ontogenetic information made suchclaims premature. Zhang (2007) considered all generic namesvalid, and noted that the presence of an anterior lobe can be usedto distinguish Hanchungella from a lobeless Liangshanella. Zhang(2007) was also unconvinced that Ovaluta represented a juvenileinstar of Liangshanella, instead noting the strong similarities inshield size and features of Ovaluta with Bullaluta Copeland, 1986,an upper Cambrian (Furongian) bradoriid of western Newfound-land, and suggested that the former may be a junior synonym.
Liangshanella circumbolina sp. nov. displays the postplete shield,lateroadmarginal rim and lack of lobation typical of the genus andis similar in general shield outline to L. liangshanensis Huo, 1956,L. rotundata Huo, 1956 and L. yunnanensis Zhang, 1974 describedfrom South China and L. burgessensis Siveter and Williams, 1997from the Burgess Shale in Canada. However, all four of thesespecies have a weakly to moderately developed anterodorsal node,a feature absent in L. circumbolina sp. nov. Liangshanella baensisdescribed by Zhang (2007, pl. 15, figs. 1-5) from the ShuijingtuoFormation of southern China differs by possessing a broad, shallowsulcus underneath the anterodorsal margin and a narrow marginalrim. Liangshanella sayutinae (Melnikova, 1988) from the Trans-Baikal area in the Russian Far-East and Greenland (Melnikova et al.,1997; Skovsted, 2006) lack an anterodorsal node and has a valveprofile similar to L. circumbolina sp. nov., but differ by the surfaceornament of fine cancelate ridges. Liangshanella circumbolina sp.nov. possesses a very distinctive exterior valve ornament consis-ting of evenly distributed depressions each containing a raisedcirclet of closely joined papillae surrounding a single, sub-micron sized central perforation (Fig. 5C-E). This highly distinctiveexterior valve ornament has not been previously documented fromany Liangshanella species, nor indeed from any other bradoriidspecies. However, it should be noted that the majority ofLiangshanella species have been described based on “crack-out”macrofossil specimens from fine-grained siliciclastic rocks (e.g.Siveter and Williams, 1997; Hou et al., 2002), a mode ofpreservation not ideal for preserving fine, microstructural detail.Minute surface perforations have been previously interpreted aspotential openings for sensory setae (Skovsted et al., 2006; Topperet al., 2007), analogous to those found in modern ostracods. Thefunction of the raised circlet of papillae surrounding the centralperforation is unknown, but may have provided some protection tothe base of sensory hairs that may have developed inside thesealed cylindrical pillars on the inner surface of the valve. Similarstructures are found in the recent ostracod, Elofsonia baltica(Hirschmann, 1909), which possesses sieve-pores that have acentral sensory hair, surrounded by a ring of mound-like papillae(see Whittaker, 1973, pl. 1:37:200, fig. 2). However, the exactfunctional significance of the mound-like papillae in this taxon alsoremains mysterious.
The FAD of Liangshanella circimbolina sp. nov. occurs some 20 mbelow the FAD of the zonal trilobite Abadiella huoi in the AjaxLimestone. The pre-trilobitic occurrence of this species means that,potentially, Liangshanella circumbolina sp. nov. represents theoldest bradoriid taxon hitherto documented from the earlyCambrian of East Gondwana. Previously, the oldest bradoriidsdocumented from East Gondwana were possible svealutids fromthe lower Cambrian (Atdabanian equivalent) Parara Limestone atCurramulka Quarry on Yorke Peninsula described by Chapman(1918) as ostracods. Jell in Bengtson et al. (1990, fig. 7) indicatedthat the lower levels of the Parara Limestone at Curramulka

321T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
contained A. huoi, and thus can be correlated with the A. huoiBiozone. Jones and Laurie (2006) suggest that the drawings byChapman (1918, pl. 9, figs 1, 2, 5) bear some resemblance to theSvealutidae, however without a more accurate representation ofthe specimens, comparison with L. circumbolina sp. nov. is notpossible.
Distribution. Ajax Limestone, lower Cambrian (Series 2, Stage 3),Flinders Ranges, Arrowie Basin, South Australia.
Stratigraphic range. All specimens recovered from a singlestratigraphic level, 117 m above the base of the AJX-M section.
Family Comptalutidae Öpik, 1968Zepaera Jones and McKenzie, 1980Type species. Zepaera rete Jones and McKenzie, 1980Zepaera jagoi sp. nov (Fig. 6A-F)Type material. Holotype: SAMP44792 (Fig. 6A-C) from sample
number AJX-M/266, 148.75 m true thickness above the base ofthe section, Ajax Limestone, Mount Scott Range. Paratypes:SAMP44793 (Fig. 6D) from sample number AJX-M/255, 143 mtrue thickness above the base of the section, Ajax Limestone,Mount Scott Range and SAMP44794 (Fig. 6E-F), from samplenumber AJX-M/266, 148.75 m true thickness above the base of thesection, Ajax Limestone, Mount Scott Range.
Etymology. Named for Dr. Jim Jago, in recognition for his con-tributions to South Australian Cambrian geology, palaeontology andbiostratigraphy.
Diagnosis. Valves postplete, with a marked retral swing and awell developed, straight hinge line. Lateroadmarginal ridge present,entire between cardinal corners with valves showing weak anteriorsulcation. Valve exterior with widely spaced, circular punctae;surface ornament consists of irregularly concentric wrinkledtexture. Valve margin well defined, ornamented by a series ofregularly spaced, short, tapering spines.
Description. Valves small (maximum height 810 µm, maximumlength 960 µm), postplete, with a well developed, straight hingeline. Valves have a thin (approximately 7.5 µm in width) later-oadmarginal ridge, entire between cardinal corners. Lateroadmar-ginal ridge separated from the lateral surface by a continuous, butweakly defined shallow depression (Fig. 6C). Valve margin welldefined, ornamented by a series of regularly spaced, short, taperingspines (Fig. 6C). Spines are approximately 10 µm in length and varybetween 13-19 µm in width and are continuous along the valvemargin, between cardinal corners (Fig. 6A). Lateral surfacemoderately to strongly inflated, without distinct anterodorsal orposterodorsal nodes. A weak, V-shaped centrodorsal depression ispresent in some specimens (Fig. 6D). Surface is infrequentlypunctate (Fig. 6C) ornamented with an irregularly concentricwrinkled texture (Fig. 6B). Some specimens exhibit low, flatcircular welt-like structures, varying in diameter from 26 µm to63 µm (Fig. 6B). Interior displays a well defined duplicature, entirebetween the cardinal corners (Fig. 6F).
Remarks. Comptalutids, like svealutids, frequently exhibit mor-phologically simple shields that result in taxonomic confusionbetween some early Cambrian bradoriids such as HoulongdongellaLee, 1975, Flemingopsis Jones and McKenzie, 1980, Zepaera Jonesand McKenzie, 1980 and Quetopsis Hinz-Schallreuter, 1999. Theexact taxonomic status and relationship of these taxa has beenthe focus of frequent discussion, but remains controversial (Shu,1990; Hinz-Schallreuter, 1999; Skovsted et al., 2006; Zhang, 2007).Due to the simplicity of the comptalutid shield, soft-part morpho-logical information will undoubtedly play an important role inthe taxonomic resolution of many comptalutid taxa. New soft-part preservation has assisted in the taxonomic classification ofthe comptaluid Kunyangella Huo, 1965 documented from theChengjiang Lagerstätte of China (Hou et al., 2010). A specimen ofKunyangella cheni Hou, Williams, Siveter, Siveter, Aldridge andSansom, 2010 interpreted as a penultimate instar displays
remnants of three head and two trunk appendages. Based on theontogentic patterns of segmentation in Crustacea, Hou et al. (2010)reconstructed the adult as having four head appendages. However,to date only two bradoriid species (Kumingella douvelli andKunyangella cheni) have been described with their soft-partanatomy preserved (Hou et al., 1996, 2010; Shu et al., 1999).
In the absence of soft-part preservation, Zhang (2007)suggested detailed ontogenetic assessment may provide a solutionfor elucidating the taxonomic status of comptalutids. Whilst thevarious growth stages of some bradoriids, such as Zepaera reteJones and McKenzie, 1980, Flemingopsis dua (Jones and McKenzie,1980) and Houlongdongella xichuanensis Zhang, 1987, have beenwell illustrated, the ontogeny of the vast majority of bradoriidsremain obscure. This is primarily due to a lack of available material,since an accurate ontogenetic framework requires a considerablecatalogue of specimens. Zhang (1987), for example, investigated131 specimens of Houlongdongella xichuanensis and Hinz-Schall-reuter (1999) examined 240 specimens of Zepaera rete and 221specimens of Flemingopsis dua for ontogenetic analysis. Thedifficulties associated with charting bradoriid ontogeny usingexternal features of the shield was recently highlighted by Houet al. (2010) who described pre-adult and adult specimens ofKumingella douvelli and Kunyangella cheni that displayed changes insoft-part anatomy between growth stages, but little or nomorphologic change was manifested in shield morphology. Thegenus Zepaera was originally reserved for comptatulid bradoriidswith distinctive lobes forming a characteristic omega-shaped ridgeon the anterior part of the valve. However, the omega-shaped ridgewas found to be only distinct in juvenile stages, graduallybecoming effaced, until it fades completely in adult stages (Jonesand McKenzie, 1980; Hinz-Schallreuter, 1999). This ontogeneticvariation has also been observed in other comptalutids (e.g.Flemingopsis dua Hinz-Schallreuter, 1999). Zepaera jagoi sp. nov.lacks these characteristic ridges and with the large majority ofspecimens approaching a maximum length of approximately900 µm, all specimens are herein considered to represent fullymature adults. Zepaera jagoi sp. nov. displays the typical postpleteshield outline and marked retral swing characteristic of adultZepaera species. Zepaera jagoi sp. nov. displays a weak V-shapedanterodorsal sulcus but lacks any obvious anterior lobation thatcharacterises the type species, Z. rete. The irregularly concentric,wrinkled ornament of Z. jagoi sp. nov. also distinguished it from thewell developed reticulate ornament in the type species. Thereare a number of Cambrian bradoriid genera with short spine-like protuberances from their shield; however the regularlyspaced, short spines that adorn the valve margin of Z. jagoi sp.nov. (Fig. 6A, C) have not been previously described in any knownbradoriid taxon.
Distribution. Ajax Limestone, lower Cambrian (Series 2, Stage 3),Abadiella huoi Biozone, Flinders Ranges, Arrowie Basin, South Australia.
Stratigraphic range. A total of 5.6 m between 143.15-148.75 mabove the base of the section AJX-M.
Family Hipponicharionidae Sylvester Bradley, 1961Remarks. A subtriangular shield in conjunction with distinct,
anterodorsal and posterodorsal lobes is indicative of two familieswithin the Bradoriida, the Hipponicharionidae and the Beyrichoni-dae. The beyrichonids can be discriminated from the hipponichar-ionids by having a less inflated shield, with subdued anterior andposterior lobes that are frequently less ventrally extended (Hinz-Schallreuter, 1993a, c; Siveter and Williams, 1997). The closemorphological similarity between the two families was recognisedby Öpik (1968), who regarded the Hipponicharionidae as asubjective synonym of the Beyrichonidae based on the comparablemorphology of Hipponicharion Matthew, 1886 and BeyrichonaMatthew, 1886, the type genera of their respective families. Thisview has not been widely accepted. Hinz-Schallreuter (1993a),

322 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
Siveter and Williams (1997) and Williams et al. (2007) allemphasized the different lobation between the families, inparticular the anterior and posterior lobes that ventrally extendfurther in the Hipponicharionidae than in the Beyrichonidae. Inaddition, Zhang (2007) noted that the posterior node or linear lobeis distinct in the Hipponicharionidae, whereas it often forms assulcation in the Beyrichonidae, a morphological feature thatcharacterises the type species, Beyrichona papilio Matthew, 1886(see Siveter and Williams, 1997, pl.4, figs. 9-10). Based on thedifferences in shield inflation and lobation, both families arerecognised as valid, separate taxonomic groupings.
Quadricona madonnae gen. et. sp. nov. displays the amplete shield,subtriangular shield outline and distinct lateroadmarginal ridgereminiscent of the Hipponicharionidae and Beyrichonidae, howeverthe lobation is strikingly different. Quadricona madonnae gen. et. sp.nov. displays short, pointed anterior and posterior nodes (Fig. 6E-F, L),unlike the rounded nodes, or ridge-like lobes frequently encounteredin the Beyrichonidae and the Hipponicharionidae. The anterior andposterior nodes are prominent in all specimens with no examples ofposterior sulcation observed, a feature frequently displayed by thebeyrichonids. The moderately inflated shield and prominent lobationdisplayed by Quadricona madonnae gen. et. sp. nov. supports des-ignation to the Hipponicharionidae.
Quadricona gen. nov.Etymology. Derived from the Latin prefix quattuor, four and conus,
cone. In reference to the four, short nodes on the complete shield of thebradoriid.
Diagnosis. Valves small, subamplete, with a subtriangular shieldoutline and well developed, straight hinge line. Narrow lateroad-marginal ridge, entire between cardinal corners. Anterodorsal andposterodorsal nodes are discrete, short, nipple-like protuberances.Surface with many small, closely spaced circular pores, each in awell defined u-shaped depression; some pores with raised, circularmargin. Surface sculpture partially wrinkled, more prominenttowards the valve margin.
Remarks. Discrimination of genera within the Hipponicharionidaeis predominantly related to shield shape, size and ventral extent ofthe anterior and posterior lobes. For example, Wimanicharion Hinz-Schallreuter, 1993a, Andresia Hinz-Schallreuter, 1993a and Neokun-mingella Zhang, 1974, typically exhibit confluent anterior and pos-terior lobes. Albrunnicola Martinsson, 1979, one of the most commonbradoriids in early Cambrian assemblages from South Australia, ischaracterised by a weakly developed posterior lobe, with both lobesrestricted to the dorsal half of the valve.Quadricona gen. nov. is similarin shield outline to Albrunnicola and also has a very similar surfacesculpture (see Topper et al., 2007, fig. 10F-G). However, the degreeand development in shield lobation between the two genera isstrikingly different. The discrete short, nipple-like projections dis-played by Quadricona gen. nov. is a unique morphological character-istic that easily distinguishes it from other genera within theHipponicharionidae.
Quadricona madonnae gen. et sp. nov. (Fig. 6G-M)Type material. Holotype: SAMP44795 (Fig. 6G-J) from sample
number ER-9/67.3, 38.6 m true thickness above the base of thesection, Wirrapowie Limestone, Elder Range. Paratypes: SAMP44796(Fig. 6K) from sample number AJX-M/268.7, 150.2 m true thicknessabove the base of the section and SAMP 44797 (Fig. 6 L-M) fromsample number AJX-M/271.3, 151.7 m true thickness above the baseof the section, Ajax Limestone, Mount Scott Range.
Etymology. Named for the American entertainer Madonna; inreference to the nodes on each valve resembling her conical brassieremade famous during the 1980s and 1990s, particularly her “BlondAmbition” tour in 1990.
Diagnosis. As for genus, by monotypy.Description. Valves subamplete with a subtriangular outline and
a well developed, straight hinge line. The valves are small in size,
measuring on average 1.19 cm in length and 0.922 cm in height,with a maximum length of 1.3 cm and height of 1.05 cm. Valves aremoderately convex with generally rounded posterior and anteriormargins. The anterior and posterior borders extend slightlybeyond the hinge line (Fig. 6G). A distinct, narrow, lateroadmar-ginal ridge is entire between cardinal corners. Anterodorsal andposterodorsal nodes are discrete, short, sharp, nipple-like pro-tuberances (Fig. 6E-F, L). Posterodorsal node is positioned slightlymore dorsally than the anterodorsal node (Fig. 6G). A slight central,dorsal swelling close to the dorsal valve margin, slightly anterior ofthe midline forms a weak and indistinct central lobe (Fig. 6G).Exterior surface with numerous small u-to v-shaped indentations,hosting a single, minute perforation with a distinctive, but simple,raised rim approximately 3 µm in diameter (Fig. 6J). Surfacesculpture partially wrinkled, especially towards the margins ofthe valves (Fig. 6I). Interior surface smooth.
Remarks. Quadricona madonnae gen. et sp. nov. is based on eightspecimens recovered from the Ajax Limestone in the Mount ScottRange and the upper Wirrapowie Limestone in the Elder Range.Quadricona madonnae gen. et sp. nov. is the only taxon unequiv-ocally common to both localities and the short stratigraphicranges in both sections (Fig. 2) may have potential for futureregional correlation. The shield outline and surface ornament ofQ. madonnae is closely comparable to a number of Cambrianbradoriids such as the hipponicharionid Albrunnicola bengtsoniHinz-Schallreuter, 1993a and the beyrichonid Parahoulongdongellabashanensis Shu, 1990 (see Figs. 6J, 7D). This suggests that similartypes of surface ornamentation may appear in many differentbradoriid lineages, restricting the value of this feature in higherlevel bradoriid taxonomy.
Distribution. Ajax and Wirrapowie Limestones, Early Cambrian(Series 2, Stage 3), Abadiella huoi Biozone, Flinders Ranges, ArrowieBasin, South Australia.
Stratigraphic range. A total interval range of 1.45 m from 150.26-151.71 m above the base in section AJX-M (Ajax Limestone) and atotal of 4.71 m from 38.6-43.31 m above the base in section ER-9(Wirrapowie Limestone).
Acknowledgements
Financial support towards fieldwork and laboratory costs comefrom a Macquarie University Research Development Grant to GABand CBS. Funding support for CBS was also provided by apostdoctoral grant from the Swedish Research Council (VR). Weextend warm thanks to Mr Graham Ragless, owner of BeltanaStation and Mr Glen Gabe, Manager at Mt. Little Station for accessto the field areas. We are indebted to B. Jonak, P. Cockle, L.E.Holmer, J.B. Jago, R. Smart, M. Smith, T. Bradley, R. Callow and M.Fuller for their assistance during the field seasons when thismaterial was collected. We are indebted to Dean Oliver (DeanOliver Graphics) for drafting Figs. 1-2 with characteristic skill andspeed. We also acknowledge Elsevier for permission to reproducethe cluster analysis presented by Williams et al. (2007, fig. 10,reproduced here in Fig. 3A). The manuscript benefited from theconstructive reviews of two anonymous reviewers.
Appendix A
Global distribution of bradoriid genera of the major palaeoconti-nents during the early Cambrian. Presence of bradoriid generaindicated by shaded box. Taxa that have only been documented froma single palaeocontinental region and subsequently removed fromthe second cluster analysis (Fig. 4) are indicated with an asterix.Principal sources of revised taxonomy and biogeographic informa-tion from Morocco (Hinz-Schallreuter, 1993a), Australia (Opik,1968; Fleming, 1973; Jones and McKenzie, 1980; Hinz-Schallreuter,
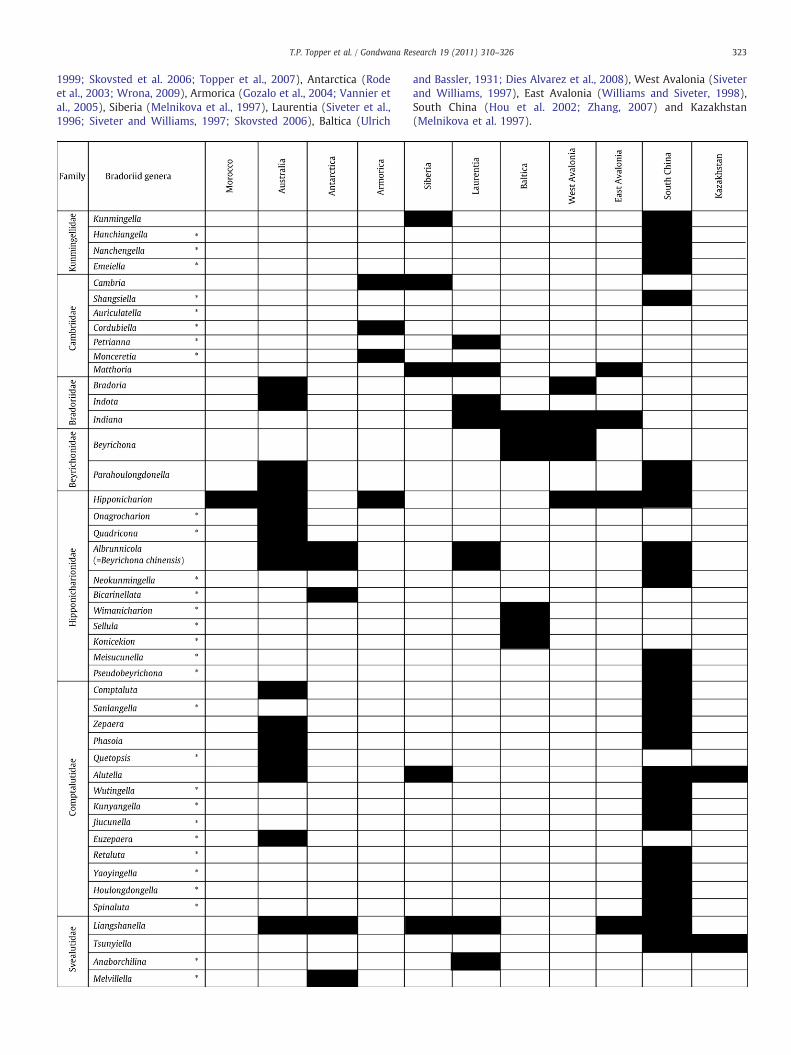
323T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
1999; Skovsted et al. 2006; Topper et al., 2007), Antarctica (Rodeet al., 2003; Wrona, 2009), Armorica (Gozalo et al., 2004; Vannier etal., 2005), Siberia (Melnikova et al., 1997), Laurentia (Siveter et al.,1996; Siveter and Williams, 1997; Skovsted 2006), Baltica (Ulrich
and Bassler, 1931; Dies Αlvarez et al., 2008), West Avalonia (Siveterand Williams, 1997), East Avalonia (Williams and Siveter, 1998),South China (Hou et al. 2002; Zhang, 2007) and Kazakhstan(Melnikova et al. 1997).

324 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
References
Babcock, L.E., Peng, S., Geyer, G., Shergold, J.H., 2005. Changing perspectives onCambrian chronostratigraphy and progress toward subdivision of the CambrianSystem. Geosciences Journal 9, 101–106.
Babcock, L.E., Peng, S., 2007. Cambrian chronostratigraphy: Current state and futureplans. Palaeogeography, Palaeoclimatology, Palaeoecology 254, 62–66.
Babcock, L.E., Peng, S., Sanchi Ellwood, B.B., 2009. Progress towards completion of aCambrian chronostratigraphic scale. 14th International Field Conference of theCambrian Stage Subcommission Working Group. Maly Karatau Range, Kazakhstan,pp. 7–8 (Abstracts and Short papers).
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., Runnegar, B.N., 1990. EarlyCambrian fossils from South Australia. Memoirs of the Association of AustralasianPalaeontologists 9, 1–364.
Brasier, M.D., Cowie, J., Taylor, M., 1994. Decision on the Precambrian-Cambrianboundary startotype. Episodes 17, 3–8.
Brock, G.A., Alexander, E.M., Paterson, J.R., Jago, J.B., Gatehouse, C.G., 2006. Mt ScottRange and Ajax Mine. In: Jago, J.B., Zhang, Wenlong (Eds.), South Australia 2006. XIInternational Conference of the Cambrian Stage Subdivision Working Group.Geological Society of Australia. South Australian Division, Adelaide.
Brock, G.A., Engelbretsen, M.J., Jago, J.B., Kruse, P.D., Laurie, J.R., Shergold, J.H., Shi, G.R.,Sorauf, J.E., 2000. Palaeobiogeographic affinities of Australian Cambrian faunas.Memoirs of the Association of Australasian Palaeontologists 23, 1–61.
Chapman, F., 1918. Ostracoda from the Upper Cambrian Limestone of South Australia.Proceedings of the Royal Society of Victoria 31, 108–112 (new series).
Conway Morris, S., 1986. The community structure of the middle Cambrian PhyllopodBed (Burgess Shale). Palaeontology 29 423-267.
Copeland, M.J., 1986. Bullaluta kindlei n. gen., n. sp. (Ostracoda, Archaeocopida)from Zone 5 (Late Cambrian, Cedaria-Crepicephalus) of the Cow Head Group,western Newfoundland. Current Research, Part B. Geological Survey of Canada(Paper 86-1B).
Dies Αlvarez, M.E., Gozalo, R., Cederstrom, P., Ahlberg, P., 2008. Bradoriid arthropodsfrom the lower-middle Cambrian of Scania, Sweden. Acta Palaeontologica Polonica53 (4), 647–656.
Evans, K.R., Rowell, A.J., 1990. Small Shelly Fossils from Antarctica: An Early CambrianFaunal Connection with Australia. Journal of Paleontology 64, 692–700.
Fleming, P.J., 1973. Bradoriids from the Xystridura Zone of the Georgina Basin,Queensland. Publications of the Queensland Geological Survey 356: ProfessionalPapers, 31, pp. 1–9.
Gozalo, R., Hinz-Schallreuter, I., 2002. Biostratigraphy and palaeobiogeography ofthe Cambrian genus Hipponicharion (Ostracoda). Paläontologishe Zeitscrift 76,65–74.
Gozalo, R., Dies, M.A., Chirivella, J.B., 2004. New occurrence of the family Hipponichar-ionidae (Bradoriida, Arthropoda), in the lower andmiddle Cambrian of the CadenasIbéricas, Spain. Geobios 37, 191–197.
Gravestock, D., 1984. Archaeocyatha from the lower parts of the Cambrian carbonatesequence in South Australia. Memoirs of the Association of AustralasianPalaeontologists 2, 1–139.
Hammer, Ø., Harper, D.A.T., Ryan, P.D., 2001. PAST: palaeontological statistics softwarepackage for education and data analysis. Palaeontologia Electronica 4, 1–9.
Hendricks, J.R., Lieberman, B.S., 2007. Biogeography and the Cambrian radiation ofarachnomorph arthropods. Memoirs of the Association of Australasian Palaeontol-ogists 34, 461–471.
Hinz, I., 1991. On Capricambria cornucopiae gen. et sp. nov. Stereo-Atlas of OstracodShells 18 (16), 65–68.
Hinz, I., 1992a. On Pejonesia sestina (Fleming). Stereo-Atlas of Ostracod Shells 19 (2), 5–8.Hinz, I., 1992b. OnMonasterium oepiki Fleming. Stereo-Atlas of Ostracod Shells 19 (29),
123–130.Hinz, I., 1993. Evolutionary trends in archaeocopid ostracods. In: McKenzie, K.G.,
Jones, P.J. (Eds.), Ostracoda in the Earth and life Sciences. A.A. Balkema,Rotterdam, pp. 3–12.
Hinz, I., Jones, P.J., 1992. On Tubupestis tuberHinz and jones gen. et sp. nov. Stereo Atlastof Ostracod Shells 19, 9–12.
Hinz-Schallreuter, I., 1993a. Cambrian ostracods mainly from Baltoscandia andMorocco. Archiv für Geschiebekunde 1 (369, 370), 385–448.
Hinz-Schallreuter, I., 1993b. Ostracodes from the Middle Cambrian of Australia. NeuesJahrbuch für Geologie und Paläontologie Abhandlungen 188, 305–326.
Hinz-Schallreuter, I., 1993c. Ein Mittelkambrischer hesslandonider Ostracod sowie zurMorphologie und systematischen Stellung der Archaeocopa. Archiv für Geschie-bekunde 1 (359-350).
Hinz-Schallreuter, I., 1999. Systematics and phylogeny of the Oepikalutidae (CambrianOstracoda). Greifswalder Geowissenschaftliche Beiträge 6, 23–53.
Hinz-Schallreuter, I., Jones, P.J., 1994. Gladioscutum lauriei n. gen. et n. sp. (Archae-ocopida) from the Middle Cambrian of the Georgina Basin, central Australia.Paleontologicheskii Zhurnal 68, 361–375.
Hinz-Schallreuter, I., Gozalo, R., Linan, E., 2008. New bradorid arthropods from theLower Cambrian of Spain. Micropalaeontology 53 (6), 497–510.
Hinz-Schallreuter, I., Schallreuter, R., 2009. Phylogeny of Phosphatocopa. Memoirs ofthe Association of Australasian Palaeontologists 37, 151–164.
Hirschmann, N., 1909. Beitrag zur Kenntnis der Ostracodenfauna des FinnischenMeerbusens. Mediterranean Society Fauna flora Fennica 35, 282–296.
Holmer, L.E., Popov, L.E., Wrona, R., 1996. Early Cambrian lingulate brachiopods fromglacial erratics of King George Island (South Shetland Islands), Antarctica. In:Gaździck, A.i (Ed.), Palaeontological Results of the Polish Antarctic Expedition,Part II: Palaeontologia Polonica, 55, pp. 37–50.

325T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
Hou, Xianguang, Bergstrom, J., 1991. The arthropods of the Lower CambrianChengjiang fauna, with relationships and evolutionary significance. In: Simo-netta, A.M., Conway Morris, S. (Eds.), The early evolution of Metazoa andthe significance of problematic taxa. Cambridge University Press, Cambridge,pp. 179–187.
Hou, Xianguang, Siveter, D.J., Williams, M., Xianghong, Feng, 2002. A monograph of thebradoriid arthropods from the Lower Cambrian of SW China. Transactions of theRoyal Society of Edinburgh, Earth Science 92, 347–409.
Hou, Xianguang, Siveter, D.J., Williams, M., Waloszek, D., Bergström, J., 1996.Appendages of the arthropod Kunmingella from the early Cambrian of China: itsbearing on the systematic position of the bradoriids and the fossil record of theOstracoda. Philosophical Transactions of the Royal Society of London B 351,1131–1145.
Hou, Xianguang, Williams, M., Siveter, D.J., Siveter, D.J., Aldridge, R.A., Sansom, R., 2010.Soft-part anatomy of the Early Cambrian bivalve arthropods Kunyangella andKunmingella: significance for the phylogenetic relationships of Bradoriida.Proceedings of the Royal Society, Series B. doi:10.1098/rspb.2009.2194.
Huo, Shicheng, 1956. Brief notes on Lower Cambrian Archaeostraca from Shensi andYunnan. Acta Palaeontologica Sinica 4, 425–445 (In Chinese with English abstract).
Huo, Shi-cheng, 1965. Additional notes on Lower Cambrian Archaeostraca from Shensiand Yunnan. Acta Palaeontologica Sinica 13, 291–307.
Huo, Shi-cheng, Shu, De-gan, Zhang, Xi-guang, Cui, Zhi-lin, Tong, Hao-wen, 1983. Noteson Cambrian bradoriids from Shaanxi, Yunnan, Sichuan, Guizhou, Hubei andGuangdong. Journal of Northwest University 39, 56–75.
Jago, J.B., Zang, Wen-long, Sun, X., Brock, G.A., Paterson, J.R., Skovsted, C.B., 2006. Areview of the Cambrian biostratigraphy of South Australia. Palaeoworld 15,406–423.
Jell, P.A., Jago, J.B., Gehling, J.G., 1992. A new conocoryphid trilobite from the LowerCambrian of the Flinders Ranges, South Australia. Alcheringa 16, 189–200.
Jiang, Z.W., 1982. Archaeostracoda. In: Luo, H.L., Jiang, Z.W., Wu, X.C., Song, X.L.,Ouyang, L. (Eds.), The Sinian-Cambrian boundary in eastern Yunnan, China, YunnanPeoples Publishing House, Yunnan, 46-47, 211-215.
Jones, P.J., Kruse, P.D., 2009. New Middle Cambrian bradoriids (Arthropoda) from theGeorgina Basin, central Australia. Memoirs of the Association of AustralasianPalaeontologists 37, 55–86.
Jones, P.J., Laurie, J.R., 2006. Bradoriida and Phosphatocopida (Arthropoda) from theArthur Creek Formation (Middle Cambrian), Georgina Basin, central Australia.Memoirs of the Association of Australasian Palaeontologists 32, 205–223.
Jones, P.J., McKenzie, K.G., 1980. Queensland Middle Cambrian Bradoriida(Crustacea): new taxa, palaeobiogeography and biological affinities. Alcheringa 4,203–225.
Kobayshi, T., Kato, F., 1951. On the ontogeny and the ventral morphology of Redlichiachinensiswith description of Alutella nakamurain new gen. and sp. Japanese Journalof Geology and Geography 11, 161–169.
Landing, E., 1994. Precambrian-Cambrian boundary global stratotype ratified and a newperspective of Cambrian time. Geology 22, 179–182.
Landing, E., Peng, S.C., Babcock, L.E., Geyer, G., Moczydlowska-Vidal, M., 2007. Globalstandard names for the lowermost Cambrian series and stage. Episodes 30,287–289.
Laurie, J., 2006. Early Middle Cambrian trilobites from Pacific Oil & Gas Baldwin 1 well,southern Georgina Basin, Northern Territory. Memoirs of the Association ofAustralasian Palaeontologists 32, 127–204.
Lee, Yu-wen, 1975. On the Cambrian Ostracoda with new material from Sichuan,Yunnan and southern Shaanxi, China. Professional papers on Stratigraphy andPalaeontology 2, 37–72.
Lieberman, B.S., 1997. Early Cambrian paleogeography and tectonic history: Abiogeographic approach. Geology 25, 1039–1042.
Lieberman, B.S., 2003a. Biogeography of the Trilobita during the Cambrian radiation:deducing geological processes from trilobite evolution. Special Papers in Palaeon-tology 70, 59–72.
Lieberman, B.S., 2003b. Taking the pulse of the Cambrian radiation. Integrative andComparative Biology 43, 229–237.
Maas, A., Waloszek, D., Müller, K.J., 2003. Morphology, ontogeny and phylogeny of thePhosphatocopina (Crustacea) from the Upper Cambrian “Orsten” of Sweden. Fossilsand Strata 49, 1–238.
Mansuy, H., 1912. Etude Geologique de Yun-Nan Oriental. Pt. 2, Palaeontologie.Memoire service geologie 1'Indochine 1, 1–23.
Martinsson, A., 1979. Albrunnicola, a new name for the Cambrian ostracode genusLongispina Andres, 1969. Lethaia 12, 27.
Matthew, G.F., 1886. On the Cambrian faunas of Cape Breton and Newfoundland.Transactions of the Royal Society of Canada 4, 147–157.
Matthew, G.F., 1902. Ostracoda of the basal Cambrian rocks in Cape Breton. CanadianRecord of Science 8, 437–470.
McKenzie, K.G., Aangel, M.V., Becker, G., Hinz-Schallreuter, I., Kontrovitz, M., Parker,A.R., Schallreuter, R.E.L., Swanson, K.M., 1999. Ostracodes. In: Savazzi, E. (Ed.),Functional Morphology of the Invertebrate Skeleton. John Wiley & Sons Ltd.,Chichester, pp. 459–507.
McKerrow, W.S., Scotese, C.R., Brasier, M.D., 1992. Early Cambrian continentalreconstructions. Geological Society of London Journal 149, 599–606.
Meert, J.G., Lieberman, B.S., 2004. A palaeomagnetic and palaeobiogeographicalperspective on latest Neoproterozoic and Early Cambrian tectonic events. Journalof the Geological Society of London 161, 1–11.
Melnikova, L.M., 1988. Nekotoryye bradoriidy(Crustacea) iz botomskogo yarusavostochnogo Zabaykal’ya. Paleontologicheskiy Zhurnal 114–117.
Meert, J.G., Lieberman, B.S., 2008. The Neoproterozoic assembly of Gondwana and itsrelationship to the Ediacaran-Cambrian radiation. Gondwana Research 14, 5–21.
Melnikova, L.M., Siveter, D.J., Williams, M., 1997. Cambrian Bradoriida and Phospha-tocopida (Arthropoda) of the former Soviet Union. Journal of Micropalaeontology16, 179–191.
Missarzhevsky, V.V., 1977. Konodonty (?) i fosfatnye problematiki kembriya Mongolii iSibiri. Bespozvonochnye Paleozoya Mongolii,. Trudy Sovmestaya Sovetsko-Mon-golskaya Paleontologicheskaya Ekspeditsiya, pp. 10–19.
Müller, K.J., 1964. Ostracoda (Bradoriina) mit phosphatischen Gehäusen aus demOberkmabrium von Schweden. Neues Jahrbuch für Geologie und PaläontologieAbhandlungen 121, 1–46.
Müller, K.J., 1979. Phosphatocopine ostracodes with preserved appendages from theUpper Cambrian of Sweden. Lethaia 12, 1–27.
Müller, K.J., 1982. Hesslandona unisulcata sp. nov. with phosphatized appendages fromUpper Cambrian Orsten of Sweden. In: Bate, R.H., Robinson, E., Sheppard, L.M.(Eds.), Fossil and Recent Ostracoda. Ellis Horwood, Chichester, pp. 276–304.
Öpik, A.A., 1961. The geology and palaeontology of the headwaters of the Burke River,Queensland. Bulletin of the Bureau of Mineral Resources. Geology and Geophysics53, 1–249.
Öpik, A.A., 1963. Early Upper Cambrian fossils from Queensland. Bureau of MineralResources, Geology and Geophysics 64, 1–133.
Öpik, A.A., 1967. The Mindyallan fauna of northwestern Queensland. Bureau of MineralResources, Australian Bulletin, 74 (1), 1-404, 74 (2), 1-167.
Öpik, A.A., 1968. Ordian (Cambrian) Crustacea Bradoriida of Australia. Bulletin. Bureauof Mineral Resources. Geology and Geophysics 103, 1–44.
Palmer, A.R., 2005. East-Gondwana/Laurentia trilobite connections — what do they tellus? Geosciences Journal 9 (2), 75–79.
Palmer, A.R., Rowell, A.J., 1995. Early Cambrian trilobites from the ShackletonLimestone of the Central Transantarctic Mountains. Paleontological Society.Memoir 45, 1–28.
Paterson, J.R., 2005. Revision of Discomesites and Estaingia (Trilobita) from the LowerCambrian Cymbric Vale Formation, western New South Wales: Taxonomic,biostratigraphic and biogeographic implications. Proceedings of the Royal Societyof New South Wales 126, 81–93.
Paterson, J.R., Brock, G.A., 2007. Early Cambrian trilobites from Angorichina, FlindersRanges, South Australia, with a new assemblage from the Pararaia bunyerooensisZone. Journal of Paleontology 81, 116–142.
Paterson, J.R., Jago, J.B., 2006. New trilobites from the Lower Cambrian Emu Bay ShaleLagerstätte at Big Gully, Kangaroo Island, South Australia. Memoirs of theAssociation of Australasian Palaeontologists 32, 43–57.
Peng, S.C., Babcock, L.E., 2008. Cambrian Period. In: Ogg, J.G., Ogg, G., Gradstein, F.M.(Eds.), A Concise Geologic Time scale. Cambridge University Press, pp. 37–46.
Pillola, G.L., 1993. The Lower Cambrian trilobite Bigotina and allied genera.Palaeontology 36, 855–881.
Raymond, P.E., 1935. Leanchoilia and other mid-Cambrian Arthropoda. Bulletin of theMuseum of Comparative Zoology 76, 205–230.
Richter, R., Richter, E., 1940. Die Saukianda – Stufe von Andalusien, eine fremde Faunaim europaeischen, Ober-Kambrium (Studien im Palaeozoikum der Mittelmeer-Laender 5). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft.(88 p).
Rode, A.L., Lieberman, B.S., Rowell, A.J., 2003. A new Early Cambrian bradoriid(Arthropoda) from East Antarctica. Journal of Paleontology 77, 691–697.
Sdzuy, K., 1972. Das Kambrium der acadobaltischen Faunen-provinz. Zentralblatt fürGeologie und Paläontologie II (1-2), 1–91.
Shergold, J.H., 1996. Cambrian (Chart 1). In: Young, G.C., Laurie, J.R. (Eds.), An AustralianPhanerozoic Timescale. Oxford University Press, Melbourne, pp. 63–76.
Shu, D., 1981. Bradoriids from Guizhou with discussion of several of the principalrelated problems. Masters Thesis. Northwest University, 123–129.
Shu, D., 1990. Cambrian and Lower Ordovician Bradoriida from Zhenjiang. Hunnan andShaanxi Provinces. Northwest University Press, Xian. (90 p).
Shu, D., Chen, L., 1994. Cambrian palaeobiogeography of Bradoriida. Journal ofSoutheast Asian Earth Sciences 9 (3), 289–299.
Shu, D., Vannier, J., Huilin, Luo, Ling, Chen, Xingliang, Zhang, Shixue, Hu., 1999. Anatomyand lifestyle of Kunmingella (Arthropoda, Bradoriida) from the Chengjiang fossilLagerstätte (lower Cambrian; Southwest China). Lethaia 32, 279–298.
Siebold, C.T.E., Stannius, H., 1845. Lehrbuch der VergleichendenAnatomie. ConVeit, Berlin.Siveter, D.J., Williams, M., Peel, J.S., Siveter, D.J., 1996. Bradoriida (Arthropoda) from the
early Cambrian of North Greenland. Transactions of the Royal Society of Edinburgh,Earth Science 86, 113–121.
Siveter, D.J., Williams, M., 1997. Cambrian bradoriid and phosphatocopid arthropods ofNorth America. Special Papers in Palaeontology 57, 1–69.
Siveter, D.J., Waloszek, D., Williams, M., 2003. An Early Cambrian phosphatocopidcrustacean with three-dimensionally preserved soft parts from Shropshire,England. Special Papers in Palaeontology 70, 9–30.
Skovsted, C.B., 2006. Small Shelly Fauna from the upper Lower Cambrian Bastion and EllaIsland Formations, North-East Greenland. Journal of Paleontology 80, 1087–1112.
Skovsted, C.B., Brock, G.A., Paterson, J.R., 2006. Bivalved arthropods from the LowerCambrian Mernmerna Formation of South Australia and their implications for theidentification of Cambrian ‘small shelly fossils’. Memoirs of the Association ofAustralasian Palaeontologists 32, 7–41.
Skovsted, C.B., Brock, G.A., Holmer, L.E., Paterson, J.R., 2009. First report of the earlyCambrian stem group brachiopod Mickwitzia from East Gondwana. GondwanaResearch 16, 145–150.
Steiner, M., Zhu, M., Weber, B., Geyer, G., 2001. The Lower Cambrian of Eastern Yunnan:Trilobite-based biostratigraphy and related faunas. Acta Palaeontologica Sinica 40,63–79 (Supp).
Stigall, A.L., 2008. Bicarinellata a new name for Bicarinella Rode, Lieberman and Rowell,2003 (Arthropoda, Bradoriida) preoccupied by Bicarinella Waterhouse, 1966

326 T.P. Topper et al. / Gondwana Research 19 (2011) 310–326
(Mollusca, Gastropoda) and Bicarinella Akopyan, 1976 (mollusca, Gastropoda).Journal of Paleontology 82 (6), 1220.
Streng, M., Ebbestad, J.O.R., Moczydlowska, M., 2008. A Walcottella-like bradoriid(Arthropoda) from the lower Cambrian of Sweden. GFF 130, 11–19.
Sylvester Bradley, P.C., 1961. Archaeocopida. Q100–10.5. In: Moore, R.C. (Ed.), Treatiseon Invertebrate Paleontology, Pt. Q, Arthropoda 3. Geological Society of America.Boulder and University of Kansas Press, Lawrence.
Tate, R., 1892. The Cambrian fossils of South Australia. Transactions of the Royal Societyof South Australia 15, 183–189.
Topper, T.P., Brock, G.A., Skovsted, C.B., Paterson, J.R., 2009. Shelly fossils from the lowerCambrian Pararaia bunyerooensis Zone, Flinders Ranges, South Australia. Memoirsof the Association of Australasian Palaeontologists 37, 199–246.
Topper, T.P., Skovsted, C.B., Brock, G.A., Paterson, J.R., 2007. New bradoriids from thelower Cambrian Mernmerna Formation, South Australia: systematics, biostratigraphyand biogeography. Memoirs of the Association of Australasian Palaeontologists 33,67–100.
Ulrich, E.O., Bassler, R.S., 1931. Cambrian bivalved Crustacea of the Order Conchostraca.Proceedings of the US National Museum 78 (4), 1–130.
Vannier, J., Williams, M., Álvaro, J.J., Vizcaïno, D., Monceret, S., Monceret, E., 2005. New EarlyCambrianbivalvedarthropods fromsouthernFrance.GeologicalMagazine142, 751–763.
Williams, M., Siveter, D.J., 1998. British Cambrian and Tremadoc bradoriid andphosphatocopid arthropods. Monograph of the Palaeontolographical SocietyLondon 1–49 (Publ. No. 607, part of Vol. 153 for 1998).
Williams, M., Siveter, D.J., Popov, L.E., Vannier, J.M.C., 2007. Biogeography and affinitiesof the bradoriid arthropods: Cosmopolitan microbenthos of the Cambrian seas.Palaeogeography, Palaeoclimatology, Palaeoecology 248, 202–232.
Williams, M., Siveter, D.J., Jose Salas, M., Vannier, J.M.C., Popov, L.E., Ghobadi Pour, M.,2008. The earliest ostracods: the geological evidence. Senckenbergiana Lethaea 88(1), 11–21.
Whittaker, J.E., 1973. On Elofsonia baltica. In: Sylvester-Bradley, P.C., Siveter, D.J. (Eds.),A Stereo-Atlas of Ostracod Shells, Volume 1. Department of Geology in theUniversity of Leicester, England (Part 3, August 31st).
Wrona, R., 2003. Early Cambrian mollusks from glacial erratics of King George Island,West Antarctica. Polish Polar Research 24, 181–216.
Wrona, R., 2004. Cambrian microfossils from glacial erratics of King George Island,Antarctica. Acta Palaeontologica Polonica 49, 13–56.
Wrona, R., 2009. Early Cambrian bradoriidae and phophatocopidae arthropods fromKing George Island, West Antarctica; Biogeographic implications. Polish PolarResearch 30 (4), 347–377.
Zang,W.-L., Jago, J.B., Lin, T.-R., 2001. EarlyCambrianacritarchs, trilobitesandarchaeocyathidsfrom Yalkalpo 2, eastern Arrowie Basin, South Australia. Report Book, 2001/00002.Department of Primary Industry and Resources, South Australia, p. 41.
Zhang W.-T., 1974. Bradoriida. In: Nanjing Institute of Geology and Palaeontology,Academica Sinica (Ed.), Handbook of Stratigraphy and Palaeontology of SouthwestChina. Science Press, Beijing, 107-111
Zhang, W.-T., 2003. Cambrian correlation between North America and China based ontrilobite and conodont faunas. Acta Palaeontologica Sinica 42, 305–316.
Zhang, W.-T., Lu, Y.-H., Zhu, Z.-L., Qian, Y.-Y., Lin, H.-L., Zhou, Z.-Y., Zhang, S.-G., Yuan, J.-L.,1980. Cambrian trilobite faunas of southwestern China. Palaeontologia Sinica 159,1–497 (new series B, no. 16).
Zhang, X.-G., 1987. Moult stages and dimorphism of Early Cambrian bradoriids fromXichuan, Henan, China. Alcheringa 11, 1–19.
Zhang, X.G., 2007. Phosphatised Bradoriids (Arthropoda) from the Cambrian of China.Palaaeontographica Abteilung A: Palaeozoologie-Stratigraphie 281, 93–173.
Zhang, X.G., Pratt, B.R., 1993. Early Cambrian ostracode larvae with a univalvedcarapace. Science 262, 93–94.
Zhang, Z., Robson, S.P., Emig, C., Shu, D., 2008. Early Cambrian radiation of brachiopods:A perspective from South China. Gondwana Research 14, 241–254.
Zhuravlev, A. Yu., 1995. Preliminary suggestions on the global Early Cambrian zonation.In: Geye, G., Landing, E. (Eds.), MOROCCO ’95—The Lower-Middle Cambrianstandard of western Gondwana, Beringeria Special Issue 2. Freunde derWürzburger Geowissenschaften e. V., Wrzberg, pp. 147–160.
Zhuravlev, A. Yu., Gravestock, D.I., 1994. Archaeocyaths from Yorke Peninsula, SouthAustralia and archaeocyathan Early Cambrian zonation. Alcheringa 18, 1–54.


















