
e Z j gy - csl.bas-net.bycsl.bas-net.by/xfile/v_med/2010/3/p046p2.pdf · 3. PROCEEDINGS. OF THE...
128
1 СЕРЫЯ МЕДЫЦЫНСКІХ НАВУК 2010 № 3 СЕРИЯ МЕДИЦИНСКИХ НАУК 2010 № 3 ЗАСНАВАЛЬНIК – НАЦЫЯНАЛЬНАЯ АКАДЭМIЯ НАВУК БЕЛАРУСI Часопіс выдаецца са студзеня 2004 г. Выходзіць чатыры разы ў год ЗМЕСТ КЛІНІЧНАЯ І ЭКСПЕРЫМЕНТАЛЬНАЯ МЕДЫЦЫНА Конопля Е. Ф., Сушко С. Н., Маленченко А. Ф., Савин А. О., Кадукова Е. М. Оценка спонтанного и химически индуцированного мутагенеза и опухолеобразования у мышей линии Af в уcловиях повышенного радиационного фона .............................................................................. 5 Титов Л. П., Столярова Е. А., Столярова Т. А. Компьютерная иммунология: сравнительный анализ за- мен нуклеотидных последовательностей CDR и FR фрагментов VH генов иммуноглобулинов больных гепатитом С, криоглобулинемией и лимфомами................................................................... 10 Белецкий А. В., Сошникова Е. В., Тесаков Д. К., Ильясевич И. А. Электрофизиологический контроль состояния функций спинного мозга в процессе хирургической коррекции сколиотической деформации позво- ночника. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19 Зафранская М. М., Федулов А. С., Нижегородова Д. Б., Мотузова Я. М., Колобова М. Ю., Багатка С. С., Миланович Н. Ф., Иванчик Г. И. Влияние мезенхимальных стволовых клеток на пролиферацию Т-клеток па- мяти у пациентов с рассеянным склерозом. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 Эйсмонт О. Л., Пашкевич Л. А., Малюк Б. В., Голутвина Н. О., Борисов А. В., Букач Д. В. Влияние перфорации остеохондральной пластинки на регенерацию поврежденного суставного хряща в эксперименте. 32 Балашевич Т. В., Никандров В. Н. Особенности состояния клеток глиомы С6 в культуре при совместном воздействии плазминогена и глицина................................................................ 38 Середа В. В., Одинцов С. Г., Власов А. П., Фима Д. В., Шубенок Д. В., Кравчук З. И., Кожух Г. В., Шманай В. В., Деев С. М., Марцев С. П. Получение модифицированного полиэтиленгликолем иммуноток- сина scFv4D5-барназа, специфичного к онкогену с-ErbB2, и сравнительное исследование его структурно- функциональных параметров ...................................................................... 42 Новак Н. В. Состояние границы «зуб – пломба» при использовании различных реставрационных мате- риалов........................................................................................... 48 Национальная академия наук Беларуси
Transcript of e Z j gy - csl.bas-net.bycsl.bas-net.by/xfile/v_med/2010/3/p046p2.pdf · 3. PROCEEDINGS. OF THE...

1
СЕРЫЯ МЕДЫЦЫНСКІХ НАВУК 2010 № 3
СЕРИЯ МЕДИЦИНСКИХ НАУК 2010 № 3
ЗАСНАВАЛЬНIК – НАЦЫЯНАЛЬНАЯ АКАДЭМIЯ НАВУК БЕЛАРУСI
Часопіс выдаецца са студзеня 2004 г.
Выходзіць чатыры разы ў год
ЗМЕСТ
КЛІНІЧНАЯ І ЭКСПЕРЫМЕНТАЛЬНАЯ МЕДЫЦЫНА
Конопля Е. Ф., Сушко С. Н., Маленченко А. Ф., Савин А. О., Кадукова Е. М. Оценка спонтанного и химически индуцированного мутагенеза и опухолеобразования у мышей линии Af в уcловиях повышенного радиационного фона . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Титов Л. П., Столярова Е. А., Столярова Т. А. Компьютерная иммунология: сравнительный анализ за-мен нуклеотидных последовательностей CDR и FR фрагментов VH генов иммуноглобулинов больных гепатитом С, криоглобулинемией и лимфомами.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Белецкий А. В., Сошникова Е. В., Тесаков Д. К., Ильясевич И. А. Электрофизиологический контроль состояния функций спинного мозга в процессе хирургической коррекции сколиотической деформации позво-ночника. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
Зафранская М. М., Федулов А. С., Нижегородова Д. Б., Мотузова Я. М., Колобова М. Ю., Багатка С. С., Миланович Н. Ф., Иванчик Г. И. Влияние мезенхимальных стволовых клеток на пролиферацию Т-клеток па-мяти у пациентов с рассеянным склерозом. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
Эйсмонт О. Л., Пашкевич Л. А., Малюк Б. В., Голутвина Н. О., Борисов А. В., Букач Д. В. Влияние перфорации остеохондральной пластинки на регенерацию поврежденного суставного хряща в эксперименте. 32
Балашевич Т. В., Никандров В. Н. Особенности состояния клеток глиомы С6 в культуре при совместном воздействии плазминогена и глицина. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
Середа В. В., Одинцов С. Г., Власов А. П., Фима Д. В., Шубенок Д. В., Кравчук З. И., Кожух Г. В., Шманай В. В., Деев С. М., Марцев С. П. Получение модифицированного полиэтиленгликолем иммуноток-сина scFv4D5-барназа, специфичного к онкогену с-ErbB2, и сравнительное исследование его структурно-функциональных параметров . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
Новак Н. В. Состояние границы «зуб – пломба» при использовании различных реставрационных мате-риалов.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
Национальная
академия наук
Беларуси

Новаковская С. А., Козловская Е. В., Рыжковская Е. Л., Манеева О. А., Арчакова Л. И. Ультраструк-турные изменения в миокарде при дилатационной кардиомиопатии . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
Чернов А. Н. Морфофункциональное состояние клеток глиомы С6 в культуре при действии рекомби-нантного фактора роста нервов. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
Митюкова Т. А., Леонова Т. А., Тузова А. А., Платонова Т. Ю., Лущик М. Л., Кулаковская Л. Г., Дрозд В. М. Биохимические показатели крови у пациентов с карциномой щитовидной железы после тоталь-ной тиреоидэктомии . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
Новикова И. А., Ярец Ю. И. Взаимосвязь степени окисленности плазмы и эритроцитов в условиях акти-вации свободнорадикального окисления. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
Девина Е. А., Принькова Т. Ю., Таганович А. Д. Особенности изменения функциональной активности альвеолярных макрофагов при контакте с сигаретным дымом . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75
Бочарова В. Н., Савчина Е. Н. Гистохимический анализ активности NADPH-диафоразы, ацетилхолин-эстеразы и ферментов энергетического обмена в структурах продолговатого мозга крысы при лихорадке. . . . . 81
Тропникова Г. К. Сравнительный анализ протекторных эффектов интервальной гипоксии и мелатонина на серотонинергические структуры передних отделов мозга крысы . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86
Терпинская Т. И. Влияние трансплантации клеток костного мозга и селезенки на прививаемость и ско-рость роста асцитной карциномы Эрлиха у мышей. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
Козлова Н. М., Касько Л. П., Кутько А. Г., Петрович В. А., Сержан Т. А., Слобожанина Е. И. Актив-ность ферментов антиоксидантной защиты и окисление мембранных белков эритроцитов при некоторых патологиях беременности . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 97
АГЛЯДЫ
Сидоренко Г. И., Герцен М. А. Актуальные аспекты изучения прегипертонии (риск, распространенность, лечебно-профилактические вмешательства). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103
Евстигнеев В. В., Головко В. А., Мастыкин А. С., Анапель Е. Н., Кистень О. В., Лаврентьева С. В. Нейросетевое моделирование и теория хаоса: возможности построения прогнозно-диагностических медицин-ских систем . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 109
ВУЧОНЫЯ БЕЛАРУСІ
Георгий Иванович Сидоренко (К 85-летию со дня рождения). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119Франтишек Иванович Висмонт (К 60-летию со дня рождения) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
ИЗ ВЕСТ ИЯ НАЦ ИО НАЛЬ НОЙ АКА ДЕМИИ НАУК БЕ ЛА РУСИ 2010 № 3
Сер ия медицинских наук
на русском, бе ло русс ком и английском язы ках
Рэ дак тар В. Г. К а л а с о ў с к а я Камп’ю тар ная вёрст ка Ю. В. Д з я н і ш ч ы к
Зда дзе на ў на бор 19.07.2010. Пад пі са на ў друк 18.08.2010. Выхад у свет 25.08.2010. Фар мат 60×841/8. Па пе ра аф сет ная. Ум. друк. арк. 14,88. Ул.-выд. арк. 16,4. Ты раж 65 экз. За каз 366.
Кошт нумару: індывідуальная падпіска – 17 780 руб., ведамасная падпіска – 44 110 руб.
Рэс пуб лі канс кае ўні тар нае прадп рыемст ва «Вы да вецкі дом «Бе ла рус кая на ву ка». ЛИ № 02330/0494405 ад 27.03.2009. Вул. Ф. Ска-рыны, 40, 220141, г. Мінск. Пасведчанне аб рэгістрацыі № 393 ад 18.05.2009.
Надрукавана ў РУП «Выдавецкі дом «Беларуская навука».
© Вы да вецкі дом «Бе ла рус кая на ву ка» Вес ці НАН Бе ла ру сі, се рыя медыцынскіх на вук, 2010
Национальная
академия наук
Беларуси

3
PROCEEDINGSOF THE NATIONAL ACADEMY
OF SCIENCES OF BELARUSMEDICINE SERIES 2010 N 3
FOUNDER IS THE NATIONAL ACADEMY OF SCIENCES OF BELARUS
The Journal has been published since January 2004
Issued four times a year
CONTENTS
CLINICAL AND EXPERIMENTAL MEDICINE
Konoplya E. F., Sushko S. N., Malenchenko A. F., Savin A. O., Kadukova E. M. Estimation of the spontaneous and chemically induced mutagen and tumor process of the mice of the line Af in the conditions of the increased radia-tion background. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Titov L. P., Stolyarova E. A., Stolyarova T. A. Computational immunology: comparative analysis of the nucle-otides substitution in CDR and FR fragments of the VH immunoglobulin’s genes from patients with hepatitis C, cryo-globulinemia and lymphomas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Beletsky A. V., Soshnikova E. V., Tesakov D. K., Ilyasevich I. A. Electrophysiological control of the functions spinal cord during surgery to correct scoliosis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
Zafranskaya М. М., Fedulov A. S., Nizheharodava D. B., Motuzova J. M., Kolobova M. J., Bagatka S. S., Milanovich N. F., Ivanchik G. I. Influence of mesenchymal stem cells on the memory t-cells proliferation in patients with multiple sclerosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
Eismont O. L., Pashkevich L. A., Maluk B. V., Golutvina N. O., Borisov A. V., Bukach D. V. Influence of the osteochondral plate perforation on the osteochondral regeneration of experimental damage of joint cartilage . . . . . . . . 32
Balashevich T. V., Nikandrov V. N. Peculiarities of C6 glioma cells state in culture under the joint effect of plas-minogen and glycine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
Sereda V. V., Оdintsov S. G., Vlasov А. P., Fima D. V., Shubenok D. V., Kravchuk Z. I., Kozhukh G. V., Shmanai V. V., Deyev S. М., Маrtsev S. P. Pegylation of recombinant immunotoxin scFv4D5-barnase directed against с-ErbB2 oncogene and its comparative structural and functional study. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
Novak N. V. State of the border «filling – tooth» in using different filling materials. . . . . . . . . . . . . . . . . . . . . . . . . 48Novakovskaya S. A., Kozlovskaya E. V., Ryzhkovskaya E. L., Maneeva O. A., Archakova L. I. Ultrastruc-
tural changes of the myocardium during dilatational cardiomyopathy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52Chernov A. N. Morphofunctional state of C6 glioma cells culture under the action of the recombinant nerve
growth factor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59Mityukova T. A., Leonova T. A., Tuzova A. A., Platonova T. Yu., Lushchyk M. L., Kulakovskaya L. G.,
Drozd V. M. Biochemical blood indices of patients with thyroid carcinoma after total thyreoidectomy. . . . . . . . . . . . . 63Novikova I. A., Yarets Y. I. Correlation between the oxidation degree of plasma and erythrocytes in the condi-
tions of free radical oxidation activation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70Devina E. A., Prinkova T. Y., Tahanovich A. D. Peculiarities of change in the functional activity of alveolar
macrophages exposed to cigarette smoke. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75Bocharova V. N., Savchina E. N. Histochemical analysis of NADPH-diaphorase, acetylcholinesterase and en-
ergy metabolism enzymes activity in structures of medulla oblongata in rats during fever . . . . . . . . . . . . . . . . . . . . . . . 81Tropnikova G. K. Comparative analysis of protective effects of intermittent hypoxia and melatonin on the sero-
toninergic structures in the rat forebrain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86
Национальная
академия наук
Беларуси

Terpinskaja T. I. Influence of transplantation of bone marrow and spleen cells on the inoculation and growth rate of ascitic Ehrlich carcinoma in rats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
Kozlova N. M., Kasko L. P., Kutko A. G., Petrovich V. A., Serzhan T. A., Slobozhanina E. I. Activity of antioxidant defence enzymes and membrane protein oxidation of erythrocytes of women with some pathologies of pregnancy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 97
SURVEYS
Sidorenko G. I., Gertszen M. A. Current aspects of prehypertension studies (risk, incidence, medical and pre-ventive manipulations) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103
Evstigneev V. V., Golovko V. A., Mastykin A. S., Anapel E. N., Kisten О. V., Lavrentieva S. V. Neuronet modelling and theory of chaos: possibilities for constructing prognosis-diagnosis medical systems . . . . . . . . . . . . . . . . 109
SCIENTISTS OF BELARUS
Georgii Ivanovich Sidorenko (To the 85th Anniversary). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119Frantishek Ivanovich Wismont (To the 60th Anniversary). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
Национальная
академия наук
Беларуси

5
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
Клінічная і эКсперЫментальная медЫцЫна
УДК 612.014.482
Е. Ф. Конопля , С. н. СУшКо, А. Ф. МАлЕнчЕнКо, А. о. САвин, Е. М. КАДУКовА
ОЦЕНКА СПОНТАННОГО И ХИМИЧЕСКИ ИНДУЦИРОВАННОГО МУТАГЕНЕЗА И ОПУХОЛЕОБРАЗОВАНИЯ У МЫШЕй ЛИНИИ AF
В УcЛОВИЯХ ПОВЫШЕННОГО РАДИАЦИОННОГО ФОНА
институт радиобиологии нАн Беларуси, Гомель
(поступила в редакцию 22.02.2010)
Введение. Определение генотоксического потенциала среды обитания в районе 30-киломе-тровой зоны аварии на Чернобыльской АЭС (территория Полесского государственного радиационно-экологического заповедника – ПГРЭЗ) является важной характеристикой ее эколо-гического состояния. Это необходимо для прогноза отдаленных последствий действия мутаге-нов различной природы на живые объекты и для решения комплекса радиобиологических про-блем: эффекты малых доз излучения, их модифицирующее действие, нестабильность генома, индукция скрытых радиационных повреждений. В настоящее время проблеме нестабильности генома уделяется особое внимание, так как она непосредственно связана с пониманием механиз-мов реализации таких важных радиобиологических явлений, как радиационный мутагенез, кан-церогенез, старение, которые являются основными отдаленными последствиями действия иони-зирующих излучений [1, 2].
Проблема оценки биологических эффектов при сочетанном действии ионизирующего излуче-ния и факторов нерадиационной природы является одной из наиболее сложных и дискуссионных в радиационной медицине и радиобиологии. Ее научная и практическая значимость определяется тем, что в реальных экологических условиях на организм действует комплекс загрязнителей раз-личной природы, а ответная реакция представляет собой интеграцию различных механизмов, суммарное проявление которых не всегда адекватно полученной дозе. Накопленный к настоящему времени экспериментальный материал свидетельствует, что при сочетанном действии ионизирую-щего излучения и при низких концентрациях химических агентов регистрируется достоверно вы-сокий вклад сверхаддитивных и антагонистических эффектов. Многофакторные воздействия аген-тов разной природы способны индуцировать в клетке сложные разнонаправленные процессы, в частности индукцию систем репарации или подавление их работы [3]. Поэтому ответную реак-цию биологической системы на сочетанное воздействие сложно прогнозировать, исходя только из информации об эффектах раздельного действия каждого из факторов [4].
Установление закономерностей индуцирования повреждения генома клетки при воздействии облучения и ее реализации при действии нерадиационных факторов позволит корректно оценить генетические последствия в условиях реального сочетанного действия факторов окружающей среды, обосновать комплекс гигиенических регламентов, обеспечивающих безопасное прожива-ние населения в постчернобыльский период, и профилактических мероприятий по снижению негативного действия существующих экологических факторов на здоровье настоящего и буду-щих поколений.
Цель работы – оценка последствий длительного облучения лабораторных мышей линии Af при их экспозиции в зоне отчуждения ЧАЭС и его влияния на спонтанный и химически индуци-рованный мутагенез и онкогенез.
Национальная
академия наук
Беларуси

6
Материалы и методы исследования. Исследования выполнены на 326 мышах линии Af с начальной массой 19–20 г, которые в возрасте 11–12 недель были разделены на две группы. Первая группа была размещена на экспериментальной базе ПГРЭЗ (12-километровая зона отчуждения ЧАЭС – д. Масаны), мощность экспозиционной дозы в местах размещения жи-вотных на поверхности почвы составляла 3,29 ± 0,10 мкГр/ч. Вторая группа мышей анало-гичного возраста содержалась на стандартной диете в виварии г. Минска в качестве контро-ля. По мере выполнения эксперимента (в течение 5 мес.) формировали группы из вивария и зоны отчуждения ЧАЭС, количество животных обоего пола в которых было одинаково.
Схема опыта: через 1, 2 и 4 мес. нахождения в зоне ПГРЭЗ группу мышей вывозили из зоны для снятия фоновых показателей и оценки реакции животных на стандартизированное действие химического мутагена. Для этой цели после транспортировки в виварий животным вводили 10%-ный раствор уретана (l мг/г массы тела). Аналогичные действия в соответствии со сроками вывоза лабораторных мышей из зоны отчуждения ЧАЭС проводили с животными, находивши-мися в виварии г. Минска. Интактным животным вводили одинаковое количество физиологиче-ского раствора.
Другую группу мышей после экспозиции в течение 1 и 4 мес. в зоне ПГРЭЗ перевозили в виварий г. Минска, отсаживали на 7–10 сут (3♀ + 2♂) с целью получения потомства. Для оцен-ки со стояния генома половых клеток родителей потомству Fl в 12-недельном возрасте также вво-дили уретан (1 мг/г). В качестве контроля использовали животных поколения Fl, полученного в условиях вивария г. Минска, которым вводили одинаковое количество физиологического рас-твора.
Генетические нарушения в соматических клетках мышей и их потомства оценивали по коли-честву полихроматофильных эритроцитов (ПХЭ) с микроядрами (МЯ) в клетках костного мозга методом [5]. Процесс образования опухолей анализировали по среднему числу аденом на мышь через 20 недель после введения уретана по методу [6].
Для изучения действия хронического облучения на противоопухолевую резистентность животных была использована экспериментальная модель асцитной карциномы Эрлиха (АКЭ) [7]. Мышам линии Af, которые находились в зоне отчуждения ЧАЭС 5 мес., после вывоза в виварий г. Минска перевивали подкожно клетки АКЭ в дозе 6 млн на мышь согласно мето-дике [8]. Половине животных через неделю после перевивки опухоли однократно внутри-брюшинно вводили противоопухолевый цитостатический препарат циклофосфан (ЦФ) (100 мг/кг), остальные мыши получали эквиобъемные количества физиологического раствора. Таким образом, были сформированы 4 группы животных: 1) зона отчуждения ЧАЭС + АКЭ; 2) зона отчуждения ЧАЭС + АКЭ + ЦФ; 3) контроль (виварий) + АКЭ; 4) контроль (виварий) + АКЭ + ЦФ.
О скорости роста опухоли судили по изменению ее объема. Объем опухоли определяли по формуле: V = (a⋅b⋅c⋅π)/6, где a, b, c – три взаимно перпендикулярных диаметра опухоли на 7, 10, 17, 19, 24 и 31-е сутки с момента перевивки; 1/6π = 0,52 – постоянная величина; V – объем опухо-ли, см3. Также велись наблюдения за выживаемостью мышей-опухоленосителей.
Статистическую обработку экспериментального материала проводили с использованием критерия Стьюдента (t-тест) при уровне значимости P<0,05, U-критерия Манна–Уитни системы Statistica 6.0. Данные представлены как среднее арифметическое ± стандартное отклонение (М±m). Коэффициент взаимодействия (Кв) определяли соотношением полученного сочетанного радиационно-токсического эффекта к вычисленному аддитивному [9].
Результаты и их обсуждение. В табл. 1 приведены данные спонтанной и химически инду-цированной (уретан) частоты ПХЭ с МЯ у лабораторных животных, экспонированных в зоне отчуждения ЧАЭС в течение 1–4 мес., а также у их потомства Fl при тех же сроках экспозиции.
Из представленных данных следует, что после перемещения мышей в зону отчуждения ЧАЭС частота ПХЭ с МЯ у мышей, экспонированных в зоне 1 мес., повысилась в 2,1 раза, при 4 мес. экспозиции – в 2,5 раза по сравнению с аналогичными показателями в группе интактного контроля вивария.
Национальная
академия наук
Беларуси

7
Т а б л и ц а 1. Частота ПХЭ с МЯ костного мозга у мышей и их потомства при экспозиции родителей в зоне отчуждения ЧАЭС и введении уретана
Группа животных
Время экспозиции 1 мес. Время экспозиции 4 мес.
Кол-во живот-
ных
Общее число проанализиро-ванных ПХЭ
Средняя частота ПХЭ
с МЯ, %
Кол-во живот-
ных
Общее число проанализиро-ванных ПХЭ
Средняя частота ПХЭ
с МЯ, %
Контроль (виварий) 12 28 602 0,14 ± 0,04 9 19 225 0,19 ± 0,03Уретан (виварий) 14 31 124 0,56 ± 0,12 8 17 037 0,57 ± 0,09Зона отчуждения ЧАЭС 12 25 830 0,29 ± 0,0* 10 21 404 0,48 ± 0,1*
Зона отчуждения ЧАЭС + уретан 10 24 280 0,72 ± 0,17** 8 17 270 0,76 ± 0,23**
Контроль к поколениям (виварий) 8 18 108 0,16 ± 0,05 10 26 400 0,18 ± 0,05Уретановый контроль к поколению F1
(виварий г. Минска) 10 23 320 0,54 ± 0,10 12 31 500 0,58 ± 0,06F1 от родителей, экспонированных в зоне ЧАЭС 2 27 562 0,22 ± 0,09 12 34 000 0,24 ± 0,05F1 + уретан от родителей, экспонированных
в зоне ЧАЭС 13 27 649 0,63 ± 0,21 11 30 955 0,64 ± 0,11
П р и м е ч а н и е. Достоверность показателей: * – P < 0,05 при сравнении с контролем; ** – Р < 0,05 при сравне-нии с группой уретанового контроля.
Введение уретана мышам, экспонированным в зоне отчуждения ЧАЭС в течение 1 мес., привело к увеличению частоты клеток ПХЭ с МЯ, что на 28,6% превысило данный показатель в группе уре-танового контроля вивария г. Минска. При пролонгировании срока экспозиции до 4 мес. введение уретана вызвало увеличение числа клеток с МЯ по сравнению с уретановым контролем на 33,3%.
Частота МЯ при экспозиции мышей в зоне отчуждения в течение 4 мес. выросла в 1,65 раза по сравнению с таковой при 1-месячной экспозиции, в то же время данный показатель в контрольной группе с учетом возрастного фактора за указанный период увеличился в 1,35 раза. Количество кле-ток с МЯ, индуцированных уретаном, при экспозиции животных 4 мес. на 5,56% превысил данный показатель у мышей, которым вводили уретан после экспозиции в зоне отчуждения ЧАЭС 1 мес.
Анализ фонового и индуцированного уретаном мутагенеза в соматических клетках мышей потомства F1, родители которых в течение 1 и 4 мес. находились в зоне отчуждения ЧАЭС, по-казал, что спонтанная частота клеток с МЯ у потомства F1, полученного от родителей, которые находились в зоне отчуждения ЧАЭС 1 мес., была увеличена по сравнению с контролем на 37,5%, а при экспозиции родителей 4 мес. – на 33,3%. Введение уретана потомству F1 от родителей, экс-понированных в зоне отчуждение ЧАЭС 1 мес., привело к увеличению частоты ПХЭ с МЯ на 16,67% по сравнению с уретановым контролем, а у потомства F1, полученного от родителей, ко-торые находились в зоне отчуждения ЧАЭС 4 мес., – на 10,34%.
Таким образом, вопрос о трансзиготной передаче генетических повреждений от облученных в малых дозах ионизирующего излучения родителей последующим поколениям требует более углубленных исследований в силу неоднозначности количественного проявления эффекта в раз-личные периоды года.
Результаты исследований индуцированного опухолевого процесса в легких у мышей (адено-мы), перемещенных в зону отчуждения ЧАЭС, а также у животных с внутрибрюшинным введе-нием уретана, представлены в табл. 2.
Т а б л и ц а 2 . Частота индуцированных аденом у животных, экспонированных в зоне отчуждения ЧАЭС в течение 4 мес.
Группа животных Кол-во животных
Кол-во аденом на мышь Кв
Контроль-1 (фоновый уровень) 39 0,13 ± 0,06 –Уретан-1 26 7,53 ± 1,51* –Зона отчуждения ЧАЭС (1 мес. экспозиции) 24 0,17 ± 0,07 –Зона отчуждения ЧАЭС (1 мес. экспозиции) + уретан 12 7,67 ± 2,18* 1,01Зона отчуждения ЧАЭС (2 мес. экспозиции) 16 0,25 ± 0,16 –
Национальная
академия наук
Беларуси

8
Группа животных Кол-во животных
Кол-во аденом на мышь Кв
Зона отчуждения ЧАЭС (2 мес. экспозиции) + уретан 15 8,13 ± 2,18* 1,06Зона отчуждения ЧАЭС (4 мес. экспозиции) 28 2,44 ± 0,58* –Зона отчуждения ЧАЭС (4 мес. экспозиции) + уретан 36 7,27 ± 1,71* 1,22Контроль-2 (4 мес. эксперимента) 29 0,41 ± 0,08 –Уретан-2 (4 мес. эксперимента) 25 12,20 ± 1,61* –
П р и м е ч а н и е. * – P ≤ 0,05 – достоверность показателей при сравнении с контролем.
Уретановая интоксикация мышей (группы уретан-1 и уретан-2) способствовала повышению количества индуцированных опухолей в легких по сравнению с соответствующими показателя-ми в разных возрастных группах (контроль-1 – в 58, контроль-2 – в 30 раз), что свидетельствует о более высокой чувствительности к уретану молодых животных (вдвое выше, чем у животных зрелого возраста).
Экспозиция мышей в зоне отчуждения ЧАЭС в течение 1–2 мес. практически не повлияла на опухолевый процесс в легких. При 4-месячной экспозиции в зоне действия ЧАЭС число опухо-лей значительно возросло, превысив соответствующий возрастной контроль в 6 раз и более чем в 14 раз – уровень опухолеобразования в легких у мышей после 1 мес. экспозиции в зоне.
Введение уретана животным при всех сроках экспозиции их в зоне отчуждения ЧАЭС стати-стически достоверно интенсифицировало онкогенез в легких. После 4 мес. экспозиции живот-ных в зоне ЧАЭС и введении уретана количество индуцированных аденом на мышь в 2,25 раза превысило аналогичный показатель опухолевого процесса в легких у мышей, получавших уре-тан, после их 1-месячной экспозиции в зоне действия ЧАЭС.
Коэффициент взаимодействия для мышей, экспонированных в зоне отчуждения ЧАЭС от 1 до 4 мес., составил 1,01–1,22, что свидетельствует о синергическом характере взаимодействия факто-ров, возрастающем с увеличением длительности экспозиции животных в зоне отчуждения ЧАЭС.
Данные по динамике выживаемости и объема сформированной АКЭ после экспозиции экс-периментальных животных в зоне отчуждения ЧАЭС в течение 5 мес. и последующего введения ЦФ представлены в табл. 3 и 4 соответственно.
Т а б л и ц а 3. Выживаемость мышей с перевитой аденокарциномой Эрлиха после экспозиции в зоне отчуждения ЧАЭС и введения ЦФ, %
Группа животных Сроки после перевивки АКЭ
17-е сутки 19-е сутки 24-е сутки 31-сутки
Виварий+АКЭ (n = 7) 28,6 28,6 28,6 14Зона отчуждения ЧАЭС+АКЭ (n = 9) 28,5 14,3 0 0Виварий+АКЭ+ЦФ (n = 9) 89 77 77 22Зона отчуждения ЧАЭС+АКЭ+ЦФ (n = 9) 50 25 25 17
Т а б л и ц а 4. Объем подкожно перевитой аденокарциномы Эрлиха у мышей после экспозиции в зоне отчуждения ЧАЭС и введения ЦФ
Группа животныхОбъем опухоли, см3
7-е сутки 10-е сутки 17-е сутки 19-е сутки 24-е сутки 31-е сутки
Виварий + АКЭ (n = 7) 0,92±0,13 2,29±0,65 1,07±0,51 2,44±1,24 3,67±0,31 8,95Зона отчуждения ЧАЭС + АКЭ
(n = 9) 1,1±0,14 0,65±0,21 1,16±0,19 0,65 – –Виварий + АКЭ + ЦФ (n = 9) – 2,19±0,24 2,27±0,27 3,45±0,36 4,31±0,45 8,17±1,17Зона отчуждения ЧАЭС + АКЭ + ЦФ
(n = 9) – 2,53±0,25 2,95±0,31 3,09±0,48 2,89±0,43 5,96±1,76
Выживаемость мышей с перевитой АКЭ после экспозиции в зоне отчуждения ЧАЭС была снижена на 14,3% по сравнению с таковой у мышей вивария на 19-е сутки после перевивки, а на 24-е сутки все мыши-опухоленосители из зоны ЧАЭС (группа «зона отчуждения ЧАЭС+АКЭ») погибли, тогда как в аналогичной группе вивария выжило 28,6% животных.
продолжение табл. 2Национальная
академия наук
Беларуси

Выживаемость мышей в виварии на фоне противоопухолевой терапии была выше (от 5 до 52% в различные сроки наблюдения) выживаемости животных, предварительно экспонированных в зоне отчуждения ЧАЭС. Терапия ЦФ повлияла и на скорость роста АКЭ у животных сравни-ваемых групп. Так, на 10-е и 17-е сутки после перевивки АКЭ и введения ЦФ у мышей, которые ранее находились в зоне отчуждения ЧАЭС, скорость роста опухоли была выше, чем у живот-ных в виварии. В последующие сроки объем опухоли у мышей вивария превысил аналогичный показатель у животных, которые находились в зоне отчуждения ЧАЭС, в связи с более ранней гибелью последних. Полученные нами данные совпадают с литературными. В частности, в ра-боте [10] отмечен колебательный характер изменения противоопухолевой резистентности крыс Вистар в зависимости от сроков облучения в зоне отчуждения ЧАЭС.
Заключение. Таким образом, полученные в ходе исследований данные показывают, что экс-позиция лабораторных животных в зоне отчуждения ЧАЭС в течение 4–5 мес. активизирует процессы мутагенеза и канцерогенеза, снижает противоопухолевую резистентность и изменяет реакцию организма на стандартизированное действие химического канцерогена. Исходя из по-лученных результатов, можно заключить, что изменение сроков экспозиции экспериментальных животных в зоне отчуждения ЧАЭС оказывает влияние на интенсивность течения метаболиче-ских процессов в клетке и способствует повышению чувствительности животных к действию нерадиационных факторов.
Работа поддержана грантом БРФФИ № Б09БРУ-003.
Литература
1. М а з у р и к В. К., М и х а й л о в В. Ф. // Радиационная биология. Радиоэкология. 2001. Т. 41, № 3. С. 272–289.2. П е л е в и н а И. И., А л е щ е н к о А. В., А н т о щ и н а М. М. и др. // Радиационная биология. Радиоэко-
логия. 2003. Т. 43, № 2. С. 161–166. 3. Г е р а с ь к и н С. А., Д и к а р е в В. Г., У д а л о в а А. А., Д и к а р е в а Н. С. // Генетика. 1996. Т. 32, № 2.
С. 279–288.4. М а л е н ч е н к о А. Ф., С у ш к о С. Н., С а л т а н о в а Н. В. // Экологическая антропология. 2001. С. 203–209.5. Руководство по краткосрочным тестам для выявления мутагенных и канцерогенных химических веществ.
Женева, 19896. Т у р у с о в В. С., П а р ф е н о в Ю. Д. Методы выявления и регламентирования химических канцерогенов.
М., 1986. 7. И в а н о в А. А., Н а с о н о в а Т. А., Д а р е н с к а я Н. Г. и др. // Радиационная биология. Радиоэкология.
2009. Т. 49, № 4. С. 449–455. 8. С о ф ь и н а З. П., С ы р к и н А. Б., Г о л д и н А., К л я й н А. Экспериментальная оценка противоопухо-
левых препаратов в СССР и США. М., 1980. 9. К у з и н A. M. // Изв. АН СССР. Сер. биол. 1983. № 4. С. 485–502.10. С а в ц о в а З. Д., Ф е д о с о в а Н. И., В о е й к о в а И. М. и др. // Experim. Oncol. 2001. Vol. 23. P. 183–188.
E. F. Konoplya , S. n. SuShKo, a. F. MalEnchEnKo, a. o. Savin, E. M. KaduKova
ESTIMATION OF THE SPONTANEOUS AND CHEMICALLY INDUCED MUTAGEN AND TUMOR PROCESS OF THE MICE OF THE LINE AF IN THE CONDITIONS OF THE INCREASED RADIATION BACKGROUND
institute of Radiobiology of naS of Belarus, Gomel
Summary
The main purpose of the present research is to estimate the influence of the radiation factor of the zone of the Chernobyl disaster on the spontaneous and chemically induced mutagen and tumor process of the laboratory animals exhibited in the zone of the Chernobyl disaster within 4–5 months. The research results have shown that the exposition of laboratory animals in the zone of the Chernobyl disaster within 4 months raises the mutagen and tumor induced process and changes the reaction of an organism to the standardized action of chemical carcinogen. The research of changes of the antineoplastic resistance of the mice of the line Af when exhibited in the zone of the Chernobyl disaster within 5 months have shown that the survival rate of mice with the tumours which have arisen at hypodermic introduction of inoculated Ehrlich’s adenocarcinoma has lowered in comparison with a similar indicator of the control group. The tumour growth in the early terms after the inoculation occurred more intensively at the mice preliminarily exhibited in the zone of alienation of the Chernobyl Nuclear Power Plant. In the subsequent terms the tumour volume in the control group has exceeded similar values of the mice that were in the zone of the Chernobyl disaster, in connection with the earlier destruction of the last.
Национальная
академия наук
Беларуси

10
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.36-002+616.153.96+616-006.441
л. п. ТиТов, Е. А. СТоляРовА, Т. А. СТоляРовА
КОМПЬЮТЕРНАЯ ИММУНОЛОГИЯ: СРАВНИТЕЛЬНЫй АНАЛИЗ ЗАМЕН НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕй cDR И FR
ФРАГМЕНТОВ VH ГЕНОВ ИММУНОГЛОБУЛИНОВ БОЛЬНЫХ ГЕПАТИТОМ С, КРИОГЛОБУЛИНЕМИЕй И ЛИМФОМАМИ
Республиканский научно-практический центр эпидемиологии и микробиологии, Минск, Беларусь
(поступила в редакцию 19.04.2010)
Введение. Компьютерные технологии все интенсивнее проникают во все сферы медицин-ской науки и практики. Геном человека насчитывает более 26 000 генов. Функция более трети из них связана с неспецифическими или специфическими иммунологическими механизмами защи-ты [3]. Иммунный ответ организма является универсальной формой защиты, сформированной в ходе биологической эволюции, от инфекций вирусной, бактериальной и иной природы [4]. Ин-дивидуальная композиция генома человека, включая суммарное количество и полиморфизм ге-нов иммунной системы, детерминирует разную степень восприимчивости к инфекционным агентам, разную степень напряженности и длительность развиваемого иммунитета, а также клиническое течение и исход инфекционных заболеваний [5].
Гепатит С – одно из распространенных инфекционных заболеваний вирусной природы, протекающее с поражением РНК-вирусом (Hepatitis C virus) гепатоцитов, клеток костного мозга и периферической крови, включая лимфоциты и их предшественники [1, 4]. Репродукция вируса гепатита С (ВГС) в цитоплазме пре-В-клеток, зрелых В-лимфоцитов и плазматических клеток, вероятно, оказывает эпигенетический эффект на экспрессию генов иммуноглобулинов и разви-тие гуморального противовирусного иммунитета [2]. Связывание поверхностного Е2 гликопро-теина ВГС с молекулой CD81 – рецептором мембраны В-клеток активирует синтез фермента цитидин-ДНК-деаминазы и запускает пролиферацию пораженных вирусом В-лимфоцитов. Ан-титела против Е-2 белка к эпитопам Е2-8 (ак. 481–500) и Е2-1 (ак. 551–579) обладают протектив-ным противовирусным эффектом [12, 26, 28].
Oсновными этапами гуморального иммунного ответа являются: 1) захват вирионов дендрит-сновными этапами гуморального иммунного ответа являются: 1) захват вирионов дендрит-ными клетками, процессирование вирусных антигенов с образованием множества пептидов (8–13 аминокислот) и их комплексов с молекулами главного комплекса гистосовместимости (ГКГ); 2) распознавание комплексов (молекулы ГКГ + пептиды иммунодоминантных эпитопов) рецепторами клонов В- и Т-лимфоцитов; 3) активация и пролиферация антигенспецифических В- и t-лимфоцитов; 4) образование и созревание антителопродуцирующих (плазматических) клеток; 5) продукция антител и накопление пула клеток памяти на протяжении циркуляции возбу-дителя и после его элиминации. Эффективность гуморального иммунитета определяется резерв-ными возможностями и специфичностью антител, интенсивностью их биосинтеза и катаболиз-ма, нейтрализующими и регуляторными свойствами, формирующимися в результате созревания афинности [3, 5, 24]. Вариабельные фрагменты тяжелых (VH) и легких (V�) цепей антител раз-V�) цепей антител раз-) цепей антител раз-личаются по степени мутабельности. P. Casali и соавт. [9], E. D. Charles и соавт. [10] выделили четыре FR области (1–4) с меньшей вариабельностью и три области (1–3) со значительно большей вариабельностью (гипервариабельные области), которые получили название комплементарно-детерминирующих (CDR1, CDR2, CDR3) (см. рисунок). Протяженность каждого из этих участков составляет около 10 аминокислот и кодируется примерно 30 нуклеотидами [13, 14].
Национальная
академия наук
Беларуси

11
Комплементарно-детерминирующие области легких и тяжелых цепей иммуноглобулинов составляют активный центр антител, или паратоп, взаимодействующий с эпитопами антигенов [3, 5]. Конформационное комплексирование гипервариабельных доменов одной тяжелой и одной легкой цепи ведет к образованию петлеобразной трехмерной структуры (кармана, полости), рас-полагающейся на поверхности молекулы антитела и способной связываться с эпитопом. Специ-фичность активного центра, формируемая в результате созревания афинности антител, обуслов-лена заменами нуклеотидов в кодонах и фиксацией в определенных положениях полипептидных цепей тех аминокислот, которые обеспечивают наиболее сильные и стабильные взаимодействия с иммунодоминантным эпитопом [5, 15, 19].
Фрагменты генов, кодирующие вариабельные домены, состоят из 288–400 нуклеотидных осно-ваний. Перестройки генов иммуноглобулинов и изменения в строении ДНК, включая синони-мичные и несинонимичные замены нуклеотидов в кодонах, имеют место уже на ранних этапах эмбрионального развития, в ходе антигеннезависимой дифференцировки В-лимфоцитов, в про-цессе развития иммунного ответа на антигены, т. е. осуществляются на протяжении всей жизни. Рекомбинация генов иммуноглобулинов происходит с участием двух ферментов – рекомбиназ, продуктов соответствующих генов – RAG1 и RAG2 [7, 9]. Рекомбинационный процесс является ключевым в генерации разнообразия специфических антител в ходе иммунного ответа. В свою очередь экспрессия генов рекомбиназ контролируется цитокинами (ИЛ-7 и др.). Механизмы экс-прессии, позитивной и негативной регуляции генов иммуноглобулинов в ходе нормального им-мунного ответа, при иммунопатологических и инфекционных состояниях, а также при разно-образных нозологических формах соматических заболеваниях изучены недостаточно [23, 24, 27].
Цель настоящей работы – сравнительный анализ частоты использования семейств генов им-муноглобулинов и замещений нуклеотидных оснований в CDR и FR фрагментах, кодирующих вируснейтрализующие анти-Е2 антитела у больных гепатитом С и ВГС-ассоциированными лимфо-пролиферативными заболеваниями (криоглобулинемией II типа и лимфомами), на базе совре-II типа и лимфомами), на базе совре- типа и лимфомами), на базе совре-менных биоинформационных технологий.
Материалы и методы исследования. Для анализа семейств иммуноглобулинов и нуклео-тидных последовательностей фрагментов генов, кодирующих вариабельные области иммуно-глобулинов, использовали данные базы NCBI GeneBank. Идентификацию семейств генов, коди-рующих вариабельные области, а также определение гипервариабельных (CDR) и каркасных (FR) регионов вариабельных участков молекул иммуноглобулинов проводили с помощью програм-мы IMGT-V-QEST (http:// imgt. cines. fr/IMGT_vquest/share/textes/).
Для сравнения нуклеотидных последовательностей вариабельных участков генов исполь-зовался метод Identity. Частоты нуклеотидных замен между генами рассчитывали с помощью программ VVK Germline 1.0 и VVK 3.4 (www. barkovsky. hotmail. ru). В качестве первичного материала для исследования были выбраны нуклеотидные последовательности 149 фрагмен-тов длиной от 284 до 396 оснований следующих генов:
Схема строения вариабельных участков тяжелых цепей молекул иммуноглобулинов
Национальная
академия наук
Беларуси

12
12 фрагментов (длина 372–396 н. о.) VН генов тяжелых γ-цепей больных гепатитом С, обу-словленным вирусом генотипа 1а, которые кодируют анти-Е2 антитела;
111 фрагментов (длина 317–389 н. о.) VН генов тяжелых µ-цепей 7 больных с ассоцииро-ванной с ВГС-смешанной криоглобулинемией II типа (пациенты: 389, 960, 1073, 1308, 1432, 1403, 773);
26 фрагментов (длина 284–382 н. о.) VH генов тяжелых γ-цепей от больных гепатитом С с ассоциированными лимфомами.
В ходе исследования нуклеотидных последовательностей генов определяли следующие гено-мические показатели:
1. GC-насыщенность – содержание гуанина и цитозина в FR и CDR регионах генов зароды-GC-насыщенность – содержание гуанина и цитозина в FR и CDR регионах генов зароды--насыщенность – содержание гуанина и цитозина в FR и CDR регионах генов зароды-FR и CDR регионах генов зароды- и CDR регионах генов зароды-шевых линий. Отражает интенсивность мутационных процессов под воздействием мутационно-го давления в результате активизации дезаминирования нуклеотидов, дефектов функции ДНК-полимераз, ошибок репликации и репарации за счет встраивания в ДНК 8-оксо-ГТФ.
2. 3GC-насыщенность – содержание гуанина и цитозина в третьих положениях кодонов. От-GC-насыщенность – содержание гуанина и цитозина в третьих положениях кодонов. От--насыщенность – содержание гуанина и цитозина в третьих положениях кодонов. От-ражает интенсивность наиболее часто возникающих синонимичных замен, не приводящих к за-мене аминокислот в кодируемом белке.
3. Транзиции GC/AT и AT/GC – частота мутаций с заменой одного пуринового основания на другое или одного пиримидинового основания на другое в нуклеотидной последовательности. Транзиции GC/AT отражают активность цитидин ДНК-дезаминазы при активации В-лимфоцитов. Частота замен АТ/GC – частота мутаций, включающих частоту транзиций и трансверсий АТ/GC, осуществляемых ферментом репарации В-лимфоцитов – полимеразой η. При определе-нии ее активности учитывали частоту замен аденина и тимина на гуанин и цитозин.
4. Частота трансверсии (GC/CG) – частота мутаций, при которых в нуклеотидной последова-тельности имеет место замена одного пуринового основания на пиримидиновое или замена одного пиримидинового основания на пуриновое. Отражает уровень активность фермента си-стемы репарации В-лимфоцитов – Rev-1.
Достоверность различий полученных данных определяли с помощью методов вариационной статистики.
Результаты исследования. Характеристика насыщенности гуанином и цитозином FR и cdR регионов генов тяжелых цепей молекул иммуноглобулинов v3 и v4 семейств герминальных ли-ний. В ходе выполнения исследования установлено, что гены тяжелых цепей иммуноглобулинов герминальных линий неравномерно насыщены гуанином и цитозином в FR и CDR регионах. Со-держание GC в гипервариабельных CDR1 и CDR2 регионах составило: для V3 семейства генов – 0,482 ± 0,02 и 355 ± 0,041; для V4 семейства генов – 0,499 ± 0,043 и 0,442 ± 0,044 соответственно, что существенно ниже уровня GC-насыщенности FR областей (для V3 семейства генов значение GC-насыщенности составило: FR1 – 0,649 ± 0,01, FR2 – 0,648 ± 0,018, FR3 – 0,5 ± 0,012; для генов V4 семейства – 0,625 ± 0,009; 0,655 ± 0,022, 0,531 ± 0,01 соответственно) (табл. 1). Достоверность различий по уровню GC-насыщенности для FR и CDR1 регионов составила Р < 0,05, для FR и CDR2 регионов эти различия были более достоверны – Р < 0,001.
Кроме того, установлено, что последовательности нуклеотидов FR регионов в третьих поло-жениях кодонов содержат больше гуанина и цитозина (для V3 семейства FR1 – 0,615 ± 0,043, FR2 – 0,753 ± 0,042, FR3 – 0,693 ± 0,03; для V4 семейства генов – 0,74 ± 0,036; 0,773 ± 0,021, 0,572 ± 0,03 соответственно), чем гипервариабельные CDR1 и CDR2 области (для V3 семейства – 0,623 ± 0,075 и 0,232 ± 0,09; для V4 семейства генов – 0,582 ± 0,068 и 0,519 ± 0,162 соответственно) (табл. 1). До-стоверность различий по уровню 3GC-насыщенности для FR и CDR1 регионов составила Р < 0,05, для FR и CDR2 регионов – Р < 0,001. Полученные результаты свидетельствуют о боль-шей вариабельности CDR регионов по сравнению с FR участками, что обусловлено низким со-CDR регионов по сравнению с FR участками, что обусловлено низким со- регионов по сравнению с FR участками, что обусловлено низким со-держанием гуанина и цитозина в CDR регионах и активностью фермента цитидин-ДНК-дезаминазы, осуществляющего дезаминирование цитозина и гуанина.
Более низкая 3GC-насыщенность гипервариабельных CDR регионов позволяет им сохранять относительную устойчивость к заменам 3GC на 3АТ и не сопровождается заменой аминокислот в кодируемых ими полипептидах. Особенностью генов герминальных линий вариабельных
Национальная
академия наук
Беларуси

13
Т а
б л
и ц
а 1
. Н
асы
щен
ност
ь FR
и C
DR
фра
гмен
тов
гено
в ге
рмин
альн
ых
лини
й V
H3
и V
H4
сем
ейст
в гу
анин
ом и
цит
озин
ом в
тре
тьем
пол
ожен
ии к
одон
а
Пок
азат
ель
Пос
ледо
вате
льно
сти
фра
гмен
тов
гено
в V
H с
емей
ств
FR1
(1)
CD
R1
(2)
FR2
(3)
CD
R2
(4)
FR
3 (5
)
VН
3V
Н4
VН
3V
Н4
VН
3V
Н4
VН
3V
Н4
VН
3V
Н4
GC
-нас
ыщ
енно
сть
(n =
22)
0,64
9 ±
0,01
0,62
5 ±
0,00
90,
482
± 0,
020,
499
± 0,
043
0,64
8 ±
0,01
80,
655
± 0,
022
0,35
5 ±
0,04
10,
442
± 0,
044
0,5
± 0,
012
0,53
1 ±
0,01
3GC
-нас
ыщ
енно
сть
(n =
22)
0,61
5 ±
0,04
3 0,
74 ±
0,0
360,
623
± 0,
075
0,58
2 ±
0,06
80,
753
± 0,
042
0,77
3 ±
0,02
10,
232
± 0,
090,
519
± 0,
162
0,69
3 ±
0,03
0,57
2 ±
0,03
Дост
овер
ност
ь ра
злич
ий G
C-
и 3G
C-н
асы
щен
ност
и FR
и
CD
R р
егио
нов
VH
3 и
VH
4 се
мейс
тв г
енов
Р (1
–2) <
0,0
5 Р
(1–4
) < 0
,001
Р (2
–3) <
0,0
5 Р
(3–4
) < 0
,001
Р (2
–5) <
0,0
5 Р
(4–5
) < 0
,001
Национальная
академия наук
Беларуси

14
участков молекул иммуноглобулинов V4 семейств является более высокий уровень GC-насы-щенности гипервариабельных CDR регионов по сравнению с соответствующими участками V3 семейства генов (для V3 семейства –0,482 ± 0,02 и 0,355 ± 0,041; для V4 семейства генов – 0,499 ± 0,043 и 0,442 ± 0,044 для CDR1 и CDR2 участков соответственно при Р < 0,05) (табл. 1).
частота использования семейств генов иммуноглобулинов у больных гепатитом С и у паци-ентов с ассоциированными лимфопролиферативными заболеваниями. Анализ фрагментов, име-ющихся в базе данных, показал, что в синтез вариабельных участков генов, кодирующих анти-E2 антитела у больных ВГС, включаются преимущественно гены V1 и V3 семейств (44,4 и 44,4% соответственно), в 11,2% случаев для синтеза молекул иммуноглобулинов используется V4 семей-ство генов. В синтез вариабельных участков молекул иммуноглобулинов у пациентов с ВГС-ас - социированной криоглобулинемией включаются преимущественно гены V1 и V3 семейств (53,15 и 36,95% соответственно). Гены V4, V5 и V7 семейств используются в 5,4; 3,6 и 0,9% случаев со-V4, V5 и V7 семейств используются в 5,4; 3,6 и 0,9% случаев со-4, V5 и V7 семейств используются в 5,4; 3,6 и 0,9% случаев со-V5 и V7 семейств используются в 5,4; 3,6 и 0,9% случаев со-5 и V7 семейств используются в 5,4; 3,6 и 0,9% случаев со-ответственно (табл. 2).
Т а б л и ц а 2. Частота использования семейств VH генов у пациентов с ВГС (1), у больных ВСГ с криоглобу-линемией (2) и у пациентов с ВГС и лимфомой (3), %
ГруппаСемейства генов иммуноглобулинов
VH1 VH2 VH3 VH4 VH5 VH7
Больные ВГС (n = 12) 44,4 – 44,4 11,2 – –Больные ВГС с криоглобулинемией (n = 111) 53,15 – 36,95 5,4 3,6 0,9Больные ВГС с лимфомами (n = 26) 26,92 3,85 38,46 30,77 – –Р (1–2) > 0,1 – > 0,05 > 0,05 – –Р (2–3) < 0,05 – > 0,1 < 0,05 – –Р (1–3) > 0,05 – > 0,1 < 0,05 – –
Последовательности нуклеотидов вариабельной области генов молекул иммуноглобулинов, вы-деленных из клеток лимфом, синтезированы на основе V1 семейства генов в 26,92% случаев; 38,46% последовательностей синтезировано на основе V3 семейства генов. Гены V4 и V2 семейств герми-V4 и V2 семейств герми-4 и V2 семейств герми-V2 семейств герми-2 семейств герми-нальных линий вовлекаются в синтез 30,77 и 3,85% последовательностей соответственно (табл. 2).
Характер нуклеотидных замен в кодонах фрагментов генов иммуноглобулинов у больных гепатитом С и у пациентов с ассоциированными лимфопролиферативными состояниями. Сред-нее значение частоты нуклеотидных замен (транзиций и трансверсий) аденина и тимина на гуа-нин и цитозин в нуклеотидных последовательностях фрагментов вариабельных участков моле-кул иммуноглобулинов ВГС-инфицированных пациентов незначительно (в 1,02 раза) превалирует над транзициями GC/AT (разность не достоверна, Р > 0,05), что свидетельствует об активности полимеразы η в В-лимфоцитах ВГС-инфицированных пациентов. Среднее значение частоты транзиций гуанина и цитозина на аденин и тимин незначительно (в 1,1 раза) превышает среднее значение частоты трансверсий гуанина и цитозина на цитозин и гуанин (табл. 3).
Т а б л и ц а 3. Сравнительная характеристика частот нуклеотидных замен во фрагментах генов вариабельных участков иммуноглобулинов у пациентов с ВГС, у больных ВСГ с криоглобулинемией и у пациентов с ВГС
и лимфомой
Частота нуклеотидных замен
Больные ВГС (n = 12)
Больные ВГС с криоглобулинемией
(n= 111)
Больные ВГС с лимфомами
(n = 23)
GC/AT (1) 0,0285 ± 0,009 0,01 ± 0,0038 0,03 ± 0,01AT/GC* (2) 0,0292 ± 0,009 0,0089 ± 0,0038 0,02 ± 0,0055GC/CG (3) 0,0257 ± 0,0078 0,005 ± 0,0018 0,013 ± 0,0036GC/TA 0,008 ± 0,0048 0,002 ± 0,0009 0,0065 ± 0,0033TA/AT 0,018 ± 0,0075 0,0024 ± 0,0009 0,008 ± 0,0029Р (1–2) > 0,05 > 0,05 < 0,05Р (2–3) > 0,05 < 0,01 < 0,05Р (1–3) > 0,05 < 0,001 < 0,001
Национальная
академия наук
Беларуси

15
Гены фрагментов вариабельных участков иммуноглобулинов пациентов с ВГС-ассоцииро-ванной криоглобулинемией характеризуются преобладанием транзиций GC/AT. При этом часто-та нуклеотидных замен (транзиций и трансверсий) аденина и тимина на гуанин и цитозин мень-ше частоты транзиций гуанина и цитозина на аденин и тимин. Соотношение частот замен GC/AT и AT/GC составляет 1,1 (разность между частотой замен GC/AT и AT/GC не достоверна, p > 0,05). Частота транзиций GC/AT в 2 раза превышает частоту трансверсий GC/CG. Различия между за-менами GC/AT и GC/CG достоверны (Р < 0,05), что свидетельствует о снижении активности ме-ханизмов репарации, осуществляемых полимеразой Rev-1 во время репликации, в сравнении с генами ВГС-инфицированных пациентов (табл. 3).
При анализе частоты замен в нуклеотидных последовательностях молекул иммуноглобули-нов, выделенных из клеток ВГС-ассоциированных лимфом, установлено, что частота транзиций GC/AT достоверно превышает частоту замен AT/GC (в 1,5 раза, Р < 0,05) и частоту трансверсии GC/CG (в 2,3 раза, Р < 0,001) (табл. 3).
Обсуждение. Инфицируя клетку, вирусы гепатита С используют потенциал ее биохимиче-ских систем с целью формирования инфекционного потомства. В результате внутриклеточной репродукции модификации подвергаются многие клеточные системы и механизмы, которые на первый взгляд не связаны с репликацией вирусного генома, трансляцией вирусных белков или образованием вирусных частиц [1, 5, 26]. К таковым процессам можно отнести процесс созрева-ния клетки, включая дифференцировку В- и Т-лимфоцитов костного мозга, лимфатических узлов и селезенки, а также жизненно важные цитоплазматические и ядерные процессы (биосин-тез нуклеиновых кислот и белков) [8, 11]. Персистенция ВГС в организме сопровождается гипер-стимуляцией и пролиферацией клонов В-лимфоцитов в результате связывания поверхностного белка вируса Е2 с клеточным рецептором – молекулой CD81. Длительная стимуляция пролифера-ции В-лимфоцитов на относительно высоком уровне приводит к развитию у некоторых пациен-тов иммунопатологических и лимфопролиферативных процессов [11, 14, 16, 26]. Вирусопосредо-ванные механизмы повреждения на этапах онкогенеза В-лимфоцитов могут быть реализованы посредством модуляции белками сердцевины вируса таких процессов, как замещение нуклеоти-дов и использование кодонов и избирательности включения аминокислот в процессе трансляции полипептидов В- и Т-клеточных рецепторов, апоптоза и трансформации клеток [17, 18, 20, 26]. Ранее установлена ассоциация криоглобулинемии II типа с гепатитом С. При этом показано на-личие клональной пролиферации В-лимфоцитов, интенсивность которой снижалась под воздей-ствием химиотерапии [10, 26]. Кроме того, у больных с криоглобулинемией II типа часто разви-ваются лимфомы, генез которых не ясен. Являются ли эти патологические состояния следствием гепатита С, либо они возникают у той части инфицированных вирусом лиц, которые имеют яв-ные или скрытые генетические дефекты дифференциации и созревания В-клеток, предшество-вавшие инфицированию, пока не установлено.
Как свидетельствуют полученные нами данные, в синтез рассматриваемых генов, кодирую-щих вариабельные участки молекул иммуноглобулинов анти-E2 у больных гепатитом С, вклю-E2 у больных гепатитом С, вклю-2 у больных гепатитом С, вклю-чаются гены преимущественно V1 и V3 семейств иммуноглобулинов. Данные семейства генов кодируют синтез большинства фрагментов вариабельных областей иммуноглобулинов, выде-ленных от пациентов с ВГС-ассоциированной криоглобулинемией и лимфомами.
Герминальные линии (зародышевые гены иммуноглобулинов) – первично сформированные кассеты генов иммуноглобулинов и локализованные в хромосомах интактных лимфоцитов – ха-рактерны для эмбриональных стадий развития лимфоцитов или соматических клеток. При ис-следовании генов герминальных линий, кодирующих вариабельные участки молекул иммуно-глобулинов, анализировалась насыщенность гипервариабельных CDR регионов и FR участков гуанином и цитозином.
Мутационное давление факторов внешней среды на геном проявляется повышением частоты замещения нуклеотидов аденина и тимина на гуанин и цитозин (GC-давление) или, наоборот, в кодирующих участках ДНК соответствующих генов [6, 14, 17, 18]. Вирусная инфекция клетки и организма может рассматриваться в качестве мощного фактора мутационного давления. Мате-матический анализ показывает, что при G + C = 0,4–0,6 мутационное давление характеризуется
Национальная
академия наук
Беларуси

16
преимущественно синонимичными заменами [6]. При этом в первую очередь фиксируются за-мены нуклеотидов в третьих положениях кодонов, которые не приводят к заменам аминокислот во фрагментах вариабельных фрагментов иммуноглобулинов. Более того, их фиксация не под-вергается воздействию жесткого естественного отбора. Однако когда насыщение гуанином и ци-тозином третьих положений достигает максимума, возрастает вероятность возникновения и фиксации замен в первых и вторых положениях кодонов, которые обычно являются несинони-мичными, т. е. приводят к замене аминокислот [14, 27]. При этом показатель GC-насыщенности достигает 0,6 и более. При усилении процессов, определяющих AT-давление, GC-насыщенность ДНК, становится менее 0,4.
Полученные нами данные указывают, что гены иммуноглобулинов герминальных линий не-равномерно насыщены гуанином и цитозином в FR и CDR регионах: содержание GC в гиперва-риабельных CDR регионах ниже уровня GC-насыщенности FR областей. FR регионы содержат больше гуанина и цитозина в третьих положениях кодона, чем CDR области. Данное свойство генов герминальных линий позволяет им сохранять относительную устойчивость к заменам 3GC на 3АТ, не ведущим к замене аминокислот, даже в присутствии фермента цитидин-ДНК-дезаминазы, индуцируемого активацией В-лимфоцитов в результате их стимуляции антигенами вируса гепатита С [17, 22, 23, 26].
Особенностью генов герминальных линий вариабельных участков молекул иммуноглобули-нов V4 семейства, чаще использующихся для синтеза молекул иммуноглобулинов у пациентов с ВГС-ассоциированными лимфомами, является более высокий уровень GC-насыщенности ги-GC-насыщенности ги--насыщенности ги-первариабельных CDR регионов, чем соответствующих участков V3 семейства. Избиратель-CDR регионов, чем соответствующих участков V3 семейства. Избиратель- регионов, чем соответствующих участков V3 семейства. Избиратель-V3 семейства. Избиратель-3 семейства. Избиратель-ность (предпочтительность) использования определенных семейств генов иммуноглобулинов в ходе иммунного ответа отмечена при ВИЧ-инфекции, Нib-инфекции, системной красной вол-ib-инфекции, системной красной вол--инфекции, системной красной вол-чанке и атопиях [13, 15, 19, 25].
Процесс возникновения соматических мутаций – один из основных механизмов, обеспечи-вающих созревание аффинности антител. Ферментом, инициирующим процесс мутагенеза, является AICDA – цитидин-ДНК-дезаминаза, индуцируемая при активации В-лимфоцитов. Она осуществляет дезаминирование цитозина с образованием урацила, который распознается и вырезается урацил-ДНК-гликозилазой с образованием АР-сайта (апиримидинового сайта) [7, 10, 12, 18]. Затем полимераза Rev-1 встраивает дезоксицитозин трифосфат (dCtP) напротив АР-сайта. При репликации нерепарированного урацила происходит замена цитозина и гуани-на на аденин и тимин (CG/AT). Особый интерес представляют мутации, возникающие в ре-зультате деятельности фермента системы репарации – полимеразы η, которая характеризуется низкой корректирующей способностью и может допускать генетические ошибки. В случае встраивания полимеразой η дезоксигуанин трифосфата и дезоксицитозин трифосфата напро-тив А:Т-нуклеотидных пар происходит репарация неправильно спаренных нуклеотидов путем активации аденин-ДНК-гликозилазы (Mut�) и тимин-ДНК-гликозилазы (TDG), осуществляю-Mut�) и тимин-ДНК-гликозилазы (TDG), осуществляю-) и тимин-ДНК-гликозилазы (TDG), осуществляю-TDG), осуществляю-), осуществляю-щих удаление аденина и тимина из материнской цепи ДНК [22]. Таким образом, при встраива-нии дезоксигуанина трифосфата и дезоксицитозина трифосфата напротив аденина и тимина ферменты системы репарации не только не ликвидируют последствия неправильного спарива-ния нуклеотидов, но и способствуют заменам нуклеотидов на обеих цепях ДНК. В случае, если А и Т встраиваются напротив гуанина и цитозина материнской цепи, Mut� и TDG обе-Mut� и TDG обе- и TDG обе-TDG обе- обе-спечивают вырезание встроенных аденина и тимина. При этом под влиянием полимеразы η может происходить неправильное спаривание нуклеотидов как по АТ, так и по GC нукле-отидным парам, однако избежать действия системы репарации могут только замены по А:Т-нуклеотидным парам, вследствие чего для определения активности полимеразы η в В-лим фоцитах учитывались замены (как транзиции, так и трансверсии) аденина и тимина на гуанин и цитозин.
Исследование замен нуклеотидов в ДНК-фрагментов генов иммуноглобулинов у пациентов рассматриваемых групп показало, что показатель соотношения среднего значения транзиций GC/AT к трансверсиям GC/CG наименьший в последовательностях молекул иммуноглобулинов, выделенных от ВГС-инфицированных пациентов (разность средних значений частот данных за-
Национальная
академия наук
Беларуси

17
мен не достоверна, P > 0,05). Для фрагментов генов иммуноглобулинов пациентов с ВГС-ас-социированной криоглобулинемией и ВГС-ассоциированными лимфомами характерно повыше-ние показателя данного соотношения (разность средних значений частот замен GC/AT и GC/CG достоверна, p < 0,001), что указывает на снижение активности фермента Rev-1, участвующего в репарации урацила во время репликации ДНК.
Соотношение среднего значения частоты транзиций GC/AT к частоте замен АТ/GC увеличи-вается во фрагментах генов иммуноглобулинов в ряду пациенты ВГС-инфицированные (0,97) – пациенты с ВГС-ассоциированной криоглобулинемией (1,1) – пациенты с ВГС-ассоциированной лимфомой (1,5), что свидетельствует о снижении в данном ряду активности полимеразы η и на-рушении механизмов репарации.
Заключение. Среди направлений развития теоретической и прикладной иммунологии все большее значение приобретают молекулярные методы исследований и технологии мате-матического анализа полученных данных, позволяющие выявить закономерности генетиче-ского кодирования информации у индивидуума и ее воспроизведения в процессе физиоло-гического и патофизиологического вариантов иммунного ответа в условиях воздействия факторов внешней среды. Проведенный сравнительный анализ частоты использования се-мейств генов иммуноглобулинов и замещений нуклеотидных последовательностей CDR и FR фрагментов, кодирующих вируснейтрализующие анти-Е2 антитела у больных гепати-том С и у пациентов с ВГС-ассоциирован ными лимфопролиферативными заболеваниями (криоглобулинемией II типа и лимфомами), показал наличие определенных закономерно-II типа и лимфомами), показал наличие определенных закономерно- типа и лимфомами), показал наличие определенных закономерно-стей микроэволюции генов тяжелых цепей иммуноглобулинов, участвующих в формирова-нии специфического иммунного ответа против вируса гепатита С в зависимости от феноти-па процесса (инфекция) и присоединяющихся патологических состояний (криоглобулинемия и лимфомы), что может быть использовано как для поиска критериев прогнозирования раз-вития лимфопролиферативных иммунопатологических процессов, так и для эффективно-сти химио- и иммунотерапии.
Литература
1. Д у н а е в Н. В., Э с а у л е н к о Е. В. // Вопр. вирусол. 2006. Т. 51, № 2. С. 10–14.2. Л е п к о в С. В., С т о р о ж а к о в Г. И., К о с ю р а С. Д. и др. // Совр. онкол. 2006. Т. 8, № 2. С. 57–62.3. Т и т о в Л. П. Иммунология: терминолог. словарь. М., 2008.4. Т и т о в Л. П. Вирусология: терминолог. словарь. Минск, 2009.5. Т и т о в Л. П. // Мед. журн. 2007. № 1. С. 4–20.6. Х р у с т а л ё в В. В., Б а р к о в с к и й Е. В. // Мед. журн. 2007. № 1. С. 92–96. 7. A g r a w a l A., S c h a t z D. G. // Cell. 1997. N 89. P. 45–53.8. B a n d o �., S h i m i z u A., R a C. // Immun. �ett. 2004. Vol. 94. Iss. 1–2. P. 99–106.9. C a s a l i P., S i l b e r s t a i n �. E. S. // Ann. N. �. Acad. Sci. 1995. P. 764.10. C h a r l e s E. D., G r e e n R. M., M a r u k i a n S. et al. // J. Blood. 2008. Vol. 111, N 3. P. 1344–1356.11. С h a u d h u r i J., A l t F. W. // Nat. Rew. Immunol. 2004. N 4. P. 541–552.12. C u r r y M. P., G o l d e n - M a s o n �., D o h er t y D. G. et al. // J. Hepatol. 2003. Vol. 38. P. 642–650.13. D a v i d D., D e m a i s o n C h., B a n i �. et al. // Eur. J. of Immunol. 2005. Vol. 25. Iss. 6. P. 1524–1528.14. E d w a r d J. // J. Mol. Immunol. 2009. Vol. 46. Iss. 3. P. 305–320.15. F o r e m a n A. �., � e m e r c i e r B., � i m A. et al. // Seminary Autoimmunity. 2008. Vol. 41, N 1. P. 80–86.16. G o r e v i c P. D. // Samters’s Immunologic Diseases. 6th ed. 2001. Vol. 2. P. 1002–1022.17. H e r s h b e r g U., S c h l o m c h i k M. J. // PNAS. 2006. Vol. 103. P. 15963–15968.18. H u a n g � u. K., � i e b e r M. R. // J. Biol. Chem. 2004. N 279. P. 6496–6500.19. H u r r i s R. S., S a l e J. E., P e t e r s e n - M a h r t S. K. // Curr. Biol. 2002. N 12. P. 435–438.20. M a s u d a K., O u c h i d a R., � i �. et al. // J. Immunol. 2009. Vol. 183, N 3. P. 1846–1850.21. N e u b e r g e r M. S., R a d a C. // J. Exp. Med. 2007. Vol. 204, N 1. Р. 7–10. 22. D i N o i a J., N e u b e r g e r M. S. // Nature. 2002. N 419. P. 43–48.23. P a p a v a s i l l o u F. N., S c h a t z D. G. // Cell. 2002. N 109. P. 35–44.24. S n o w R. E., D j u k a n o v i c t R., S t e v e n s o n F. K. // Immunology. Vol. 98. Iss. 4. P. 646–651.25. T o n e g a w a S., B r a c k C., H o z u m i N. // Cold Spring Harbor Symp. Quant. Biol. 1978. N 42. P. 921–931.26. D e R e V., D e V i t a S., M a r z o t t o A. et al. // J. Blood. 2000. Vol. 96, N 10. Р. 3578–3584.27. Z e n g N. Z., W i l s o n K., J a r e d M. // JEM. 2005. Vol. 201, N 9. P. 1467–1478.28. Z i b e r t A., K r a s s W., R o s s R. S. et al. // J. of Hepatol. 1999. Vol. 30. P. 177–184.
Национальная
академия наук
Беларуси

l. p. TiTov, E. a. STolyaRova, T. a. STolyaRova
COMPUTATIONAL IMMUNOLOGY: COMPARATIVE ANALYSIS OF THE NUCLEOTIDES SUBSTITUTION IN CDR AND FR FRAGMENTS OF THE VH IMMUNOGLOBULIN’S GENES FROM PATIENTS
WITH HEPATITIS C, CRYOGLOBULINEMIA AND LYMPHOMAS
Republican Scientific-practical center for Epidemiology and Microbiology, Minsk, Belarus
Summary
Dogma of genomics is that DNA leads to RNA, which leads to proteins that provide function. The genome contains a pool of gene segments for each type of the polypeptides chain. Random assortment of these segments makes the largest contribution to the receptor diversity. In the case of B-cells, the diversity is generated in two steps. First, a variety of B-cell receptors are encoded in the germ line as gene segments that undergo a process of random rearrangement to form an assembled receptor gene. Second, when a pathogen triggers an immune response, B-cells undergo a process of proliferation, death, and mutation of their IgV regions. The mutation rate is high, 10–3 per bp-generation –1, leading to a remarkable diversity of receptors among antigen-stimulated B-cells during the acute phase of immune response. For the heavy (H) chain of BCR (antibodies) there are 51 hypervariable (VH) segments. Immunoglobulin class switching and point mutations may occur while this is going on. B-cells contain an enzyme – activation-induced deaminase (AID) – that converts C in CG pairs to U. These are concentrated in the hypervariable region of the rearranged gene. In attempting to repair the resulting mismatch, mutation may result. Some of these may generate a binding site with the increased affinity for antigen’s epitope. The result is the affinity maturation, the production of antibodies of the ever-increasing affinity for the antigen. Nevertheless, it is also clear that B-cell receptors, as whole, could not have evolved just to recognize specific ligands because the immune system responds to a changing and unpredictable array of antigens.
Национальная
академия наук
Беларуси

19
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.832:612.014.42:[616.711-007.55-089
А. в. БЕлЕЦКиЙ, Е. в. СошниКовА, Д. К. ТЕСАКов, и. А. илЬяСЕвич
ЭЛЕКТРОФИЗИОЛОГИЧЕСКИй КОНТРОЛЬ СОСТОЯНИЯ ФУНКЦИй СПИННОГО МОЗГА В ПРОЦЕССЕ ХИРУРГИЧЕСКОй КОРРЕКЦИИ
СКОЛИОТИЧЕСКОй ДЕФОРМАЦИИ ПОЗВОНОЧНИКА
Республиканский научно-практический центр травматологии и ортопедии, Минск, Беларусь
(поступила в редакцию 06.04.2010)
Введение. В последние годы отмечается тенденция к внедрению в клиническую практику мало-инвазивных методов диагностики и лечения. Одним из наиболее распространенных способов оцен-ки функции нервной системы является метод соматосенсорных вызванных потенциалов (ССВП), позволяющий оценивать проводимость глубоко расположенных нервных структур, не оказывая при этом травмирующего воздействия на ткани. Данный метод основан на изучении параметров вызванных потенциалов, возникающих в мозге при электрической стимуляции периферических нервных стволов. ССВП отражают проведение афферентной волны возбуждения по восходящим путям общей чувствительности спинного мозга (СМ), затем через стволовые ядра по спино-таламическому тракту и далее в первичную соматосенсорную кору головного мозга [5, 6, 18].
По данным зарубежной литературы, метод ССВП широко применяется для контроля невро-логического статуса СМ и его корешков во время хирургического вмешательства на позвоночни-ке. Его применение основано на высокой чувствительности параметров ССВП к изменению воз-будимости нервных структур мозга и его микрогемоциркуляции [5, 13, 14].
Идиопатический (диспластический) сколиоз является одним из наиболее распространенных заболеваний опорно-двигательной системы детского и подросткового возраста, которое характе-ризуется развивающейся в процессе роста специфической трехплоскостной деформацией по-звоночника по типу скручивания. Сколиотическое искривление позвоночника сопровождается выраженными структурными изменениями тел позвонков, суставов и межпозвонковых дисков, что приводит к нарушению топографической анатомии позвоночного канала, его деформации и транспозиции СМ [1, 2, 7, 8, 11]. С нарастанием деформации, выходом ее тяжести на IV степень, когда искривление уже полностью поглотило физиологический анатомо-биомеханический ресурс позвоночника, происходит более выраженное смещение дурального мешка со всем его содержимым в вогнутую часть позвоночника, где он находится в состоянии натяжения за счет перерастянутых и напряженных спинномозговых корешков. СМ оказывается сплющенным, зна-чительно уменьшается его переднезадний диаметр, сдавливаются поверхностные оболочечные и некоторые корешковые артерии, что приводит к расстройствам микроциркуляции, нарушению ликвородинамики и изменению функционального состояния сегментарного аппарата. В прямой зависимости от величины угла деформации находится уменьшение объемного капиллярного кровотока оболочек СМ, наиболее выраженное на вершине искривления позвоночника [4, 13, 14, 19, 20, 22, 23]. Указанные анатомо-биомеханические нарушения в состоянии СМ приводят к сни-жению физиологических ресурсов его микроциркуляторного русла при тяжелых сколиотиче-ских деформациях позвоночника.
Одним из ведущих методов лечения сколиоза IV степени тяжести является хирургическая коррекция и стабилизация зоны искривления с применением имплантируемых металлокон-струкций. Оперативные вмешательства при сколиозе относят к категории высокотравматичных
Национальная
академия наук
Беларуси

20
и продолжительных, имеющих риск провокации неврологических осложнений, связанных с реа-гированием СМ на изменяемые анатомо-топографические и биомеханические параметры дефор-мированного позвоночника [5, 10].
Нейрофизиологический интраоперационный мониторинг осуществляется по динамике пара-метров ССВП, среди которых выделяют: преходящие нарушения (до 30 мин), постоянные (более 30 мин), истинные (74%), ложноположительные (3,3% − значимые изменения ССВП, которые не сопровождаются неврологическими расстройствами после операции). Однако разработка досто-верных критериев ССВП и их диагностическая значимость на различных этапах хирургической коррекции сколиотической деформации позвоночника до сих пор остаются малоосвещенными в литературе и до конца не изученными.
Цель работы – электрофизиологическая оценка выраженности неврологического дефицита в системе восходящих путей СМ у больных с идиопатическим сколизом IV степени в динамике оперативного лечения по данным ССВП.
Материалы и методы исследования. Электрофизиологические наблюдения методом ССВП выполнены у 82 пациентов с идиопатическим (диспластическим) сколиозом в возрасте от 14 до 18 лет с основной дугой искривления позвоночника от 40 до 150° (при измерении по J. A. Cobb) в грудном отделе справа [16]. С учетом значимости углового интервала больные были распреде-лены на четыре клинические группы: с дугой искривления 41−60° (n = 36), 61−90° (n = 27), 91−120° (n = 8), 121−150° (n = 11). По данным клинического и лучевого обследования с примене-нием магнитно-резонансной (МРТ) и рентгеновской компьютерной томографии (РКТ), призна-ков компрессии нервных структур СМ у этих больных не обнаружено.
Всем пациентам была выполнена одноэтапная дорсальная коррекция и стабилизация дефор-мации позвоночника по методологии Котреля−Дюбуссе [17] с применением имплантируемой ти-тановой конструкции BelCD фирмы «Медбиотех» (Республика Беларусь).
Электрофизиологический контроль состояния СМ проводили в режиме динамического на-блюдения по данным ССВП (до операции, затем через 3 недели, 3, 6 и 12 мес. после операции). У 10 пациентов с дугой искривления позвоночника свыше 90° выполнен интраоперационный мониторинг (ИОМ) неврологического статуса СМ во время хирургического вмешательства на позвоночнике. Группу контроля составили 20 здоровых лиц того же возраста.
До и после операции регистрацию ССВП осуществляли в ответ на электрическое раздраже-нии n. tibialis на уровне медиальной лодыжки. Использовали стандартную схему отведения ССВП на трех уровнях: периферическом (pf ) – в подколенной ямке; спинальном (n22) – в проек-ции поясничного утолщения; корковом (P38−n45) – в проекции коркового представительства нижней конечности [5, 6, 9]. Анализировали следующие параметры ССВП: амплитуду (мкВ), ла-тентный период (мс), длительность (мс) и время центрального афферентного проведения (мс), которое рассчитывали как разность между латентными периодами коркового (P38) и спинально-го (n22) потенциалов. Оборудование: электрофизиологическая цифровая установка Nicolet Viking IV (Nicolet Biomedical, США).
При проведении ИОМ использовали модификацию стандартной методики ССВП (n. tibialis), при которой регистрацию ССВП осуществляли по двум отведениям: pf и P38−n45. Для ИОМ ис-пользовали установку Keypoint (Medtronic A/S, Дания).
Во время операции, несмотря на проводимый электрофизиологический мониторинг, у всех больных выполнялся анестезиологический тест на пробуждение (wake-up тест) с целью допол-wake-up тест) с целью допол--up тест) с целью допол-up тест) с целью допол- тест) с целью допол-нительного контроля функций СМ [21].
Статистическая обработка цифровых данных выполнена с применением компьютерных про-грамм Microsoft Excel и GraphPad Prism 5.0.
Результаты и их обсуждение. При оценке диагностической значимости выявленных нару-шений ССВП руководствовались общепринятым мнением, согласно которому достоверными яв-ляются изменения латентного периода потенциалов на 25%, амплитуды – на 50% [5].
Анализ данных ССВП, зарегистрированных до операции у пациентов с углом искривления до 90° (I и II группы) показал, что изучаемые параметры находились в пределах, близких к кон-I и II группы) показал, что изучаемые параметры находились в пределах, близких к кон- и II группы) показал, что изучаемые параметры находились в пределах, близких к кон-II группы) показал, что изучаемые параметры находились в пределах, близких к кон- группы) показал, что изучаемые параметры находились в пределах, близких к кон-трольным значениям, или отклонялись от них незначительно − на 5−10% (P > 0,05).
Национальная
академия наук
Беларуси

21
У пациентов III и IV групп, имев-III и IV групп, имев- и IV групп, имев-IV групп, имев- групп, имев-ших дуги деформации более 90°, изме-нения параметров ССВП были более выраженными (P<0,05) по сравнению с контролем. Они сопровождались снижением амплитуды, увеличением латентного периода и времени цен-трального афферентного проведения на 20–25%. Электрофизиологическая картина, зарегистрированная у боль-ных с дугами искривления позвоноч-ника свыше 90°, свидетельствовала об изменении функций СМ, которое не определялось клинически и не выяв-лялось по данным МРТ. На диаграм-мах (рис. 1) графически представлено изменение времени центрального афферентного проведения импульса (CCT – central conduction time) по восходящим путям СМ. Характер билатеральной асимметрии CCT свидетельствует, что наиболее выраженное изменение функций СМ определяется на выпуклой стороне деформации по-звоночника (по сравнению с вогнутой стороной) и зависит от величины дуги его искривления. На рис. 1 хорошо видно, что наиболее значимое изменение ССТ определяется у пациентов IV группы.
Результаты проведенной диагностики учитывали при планировании тактики и объема хирур-гического вмешательства. Пациентов с признаками нарушения ССВП относили к группе риска, у которых изначально имеется снижение сенсорной проводимости СМ. С учетом полученных дан-ных рекомендовали щадящий режим интраоперационной дистракции позвоночника и проведение после операции консервативной терапии, направленной на улучшение микроциркуляции СМ. При отсутствии признаков нарушения проводимости СМ хирургическое вмешательство планировали в объеме максимально возможной коррекции де-формации позвоночника.
Во время операции ИОМ проводили на про-тяжении всех этапов хирургического вмеша-тельства. Интраоперационные ССВП оценивали в соответствии с классификацией Американско-го электроэнцефалографического общества [15]. Критерием возникновения нежелательных по-следствий считалось снижение амплитуды по-тенциала более чем на 50% и/или увеличение латентного периода до 10% с учетом того, что в условиях длительного наркоза латентный пе-риод может изменяться независимо от тяжести хирургического вмешательства.
В наших исследованиях по сравнению с фоно-выми данными ССВП (до операции) после пода-чи наркоза наблюдали незначительно изменение амплитуды и латентного периода ССВП, отра-жающее общий кумулятивный эффект анесте-зии. В ходе проведения операции, включая все корригирующие и стабилизирующие манипуля-ции, параметры периферического и корковых ССВП характеризовались стабильными значени-ями на протяжении всего периода наблюдения – 6−8 ч (рис. 2).
Рис. 1. Изменение времени центрального афферентного проведения импульса (мс) у пациентов cо сколиотической деформацией позво-cо сколиотической деформацией позво-о сколиотической деформацией позво-ночника IV степени тяжести в зависимости от величины основной
дуги искривления позвоночника
Рис. 2. Интраоперационный мониторинг по данным со-матосенсорных вызванных потенциалов пациентки Д.
со сколиозом IV степени тяжести.Этапы: 1 – до начала операции; 2 – скелетирование позвоночника; 3, 4 –установка винтов; 5 – коррекция деформации позвоночника; 6 – завершение коррекции
деформации позвоночника
Национальная
академия наук
Беларуси

22
В сроки через 3 недели после операции электрофизиологический контроль по данным ССВП (n = 25) свидетельствовал о нормализации функций СМ. Положительная динамика параметров ССВП характеризовалась увеличением амплитуды, снижением латентного времени и степени билатеральной асимметрии показателей, что отражало процессы восстановления проводимости СМ. Электрофизиологические и клинические наблюдения в более поздние сроки (3, 6 и 12 мес.) указывали на стабилизацию параметров ССВП, возможность достижения контрольных значе-ний и в более отдаленные сроки после операции, а также на общее улучшение функционального состояния и самочувствия пациентов (рис. 3).
Заключение. Таким образом, проведенные исследования позволили количественно оценить состояние функций СМ в динамике оперативного лечения сколиоза. Полученные результаты показали, что, несмотря на тяжелую деформацию позвоночника, у больных сколиозом с дугой искривления до 90° сохраняются нормальные показатели сенсорной проводимости СМ, что от-ражает высокую резистентность сегментарных и проводниковых нервных структур мозга к хро-нической компрессии. Увеличение тяжести сколиотической деформации свыше 90° сопровожда-ется появлением электрофизиологических признаков субклинического поражения СМ, основой которых могут являться ишемические нарушения [14, 22]. Установлено, что у пациентов указан-ной категории риск неврологических осложнений при интраоперационной дистракции позво-ночника повышен.
В отличие от хронической компрессии СМ, которая имеет место при дегенеративно-дистро-фических заболеваниях, деформациях или нестабильности позвоночника, острые травматические феномены, в том числе гипоксия или тракция, как правило, приводят к немедленным, необрати-мым и масштабным нейродеструктивным поражениям СМ, имеющим не только очаговый, но и дистантный характер [3, 12]. Применение интраоперационного мониторинга позволяет контро-лировать состояние функций спинного мозга на всех этапах оперативного лечения сколиоза.
Литература
А б а л ь м а с о в а Е. А., К о г а н А. В. // Ортопедия, травматология и протезирование. 1995. № 7. С. 881. −97.Б а л д о в а С. Н. Клинико-нейрофизиологическая характеристика идиопатического сколиоза у детей: 2.
Автореф. дис. … канд. мед. наук. Нижний Новгород, 2009. Б о р щ е н к о И. А., Б а с к о в А. В., К о р ш у н о в А. Г., С а т а н о в а Ф. С. // Вопр. нейрохирургии им. 3.
Н. Н. Бурденко. 2000. № 2. С. 28−31.В е т р и л э С. Т., М о р о з о в А. К., К и с е л ь А. А. и др. // Вестн. травматол. и ортопед. им. Н. Н. Приоро-4.
ва. 2003. № 1. С. 11−20.В и ш н е в с к и й А. А., П о с о х и н а О. В., Р ы ж о в а О. Е. и др. // Хирургия позвоночника. 2005. № 3. 5.
С. 101−110.Г н е з д и ц к и й В. В. Вызванные потенциалы мозга в клинической практике. М., 2003.6. Д у д и н М. Г., П и н ч у к Д. Ю. // Хирургия позвоночника. 2005. № 1. С. 457. −55.Д у д и н М. Г., П и н ч у к Д. Ю., Б е к ш а е в С. С. и др. // Хирургия позвоночника. 2006. № 4. С. 188. −25.
Рис. 3. Изменения соматосенсорных вызванных потенциалов у больной Б., 18 лет, со сколиотической деформацией позвоночника IV степени тяжести до (А) и после (Б) хирургической коррекции: 1 – регистрация ССВП справа;
2 – регистрация ССВП слева (n22 – спинальный пик, P38−n45 – корковый комплекс)
А БНациональная
академия наук
Беларуси

Н и к о л а е в С. Г. Практикум по клинической электромиографии. Иваново, 2003. 9. Т е с а к о в Д. К. // Развитие вертебрологии на современном этапе: материалы науч.-практ. конф. Минск, 10.
2009. С. 204−208.Ц и в ь я н Я. Л. Сколиотическая болезнь и ее лечение. Ташкент, 1972.11. Ш е в е л е в И. Н., Б а с к о в А. В., Я р и к о в Д. Е., Б о р щ е н к о И. А. // Вопр. нейрохирургии им. 12.
Н. Н. Бурденко. 2000. № 3. С. 35−39.Ш е и н А. П., К р и в о р у ч к о Г. А., Щ у р о в а Е. Н. и др. // Хирургия позвоночника. 2007. № 1. 13.
С. 35−43.Щ у р о в а Е. Н., Х у д я е в А. Т., К о в а л е н к о П. И. // Хирургия позвоночника. 2006. № 4. С. 26–32.14. American Electroencepghalograpic Society: Guideline nine: Guidelines on Evoked Potentials // J. Clin. Neurophysi-15.
ol. 1994. Vol. 11. P. 40−73.C o b b J. A. / The Am. Acad. of Orthopaedic Surg. 1948. N 5. P. 26116. −275.C o t r e l �., D u b o u s s e t J. Instrumentation en Chirurgie Rachidienne. Principes, Technique, Erreurs et Pieges. 17.
Sauramps medical, 1992.H a u s m a n n O., M i n K., B o n i T. et al. // Eur. Spine J. 2003. N 12. P. 11718. −123. I n o u e M., M i n a m i S., N a k a t a �. et al. // Spine. 2004. Vol. 30, N 3. P. 10819. −114. M a c h i d a M., S a i t o M., D u b o u s s e t J. et al. // Eur. Spine J. 2005. N 14. P. 84320. −848.P a d b e r g A. M., W i l s o n - H o l d e n T. J., � e n k e �. G., B r i d w e l l K. H. // Spine. 1998. Vol. 23, 21.
N 12. P. 1392−1400. R o b e r t R., B o g e l l i D. M., F a u r e A. et al. Vascularization of the spinal cord. GICD, 1994. P. 3422. −40.V e l d h u i z e n A. G., W e v e r D. J., W e b b P. J. // Eur. Spine J. 2000. N 9. P. 17823. −184.
А. v. BElETSKy, E. v. SoShniKova, d. K. TESaKov, i. a. ilyaSEvich
ELECTROPHYSIOLOGICAL CONTROL OF THE FUNCTIONS SPINAL CORD DURING SURGERY TO CORRECT SCOLIOSIS
Republican Scientific practical center of Traumatology and orthopaedy, Minsk, Belarus
Summary
The results of the electrophysiological study (by somatosensory evoked potentials) have been presented in 82 patients (14−18 years) with scoliosis of grade IV and 20 healthy subjects.
The character of the changes revealed allows determining electrophysiological criteria of the function of conduction of the spinal cord. They could be used for prediction of neurological complications during surgery to correct scoliosis.
Национальная
академия наук
Беларуси

24
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.832-004.2+616-056.22]:612.017.1:602.9
М. М. ЗАФРАнСКАя1, А. С. ФЕДУлов2, Д. Б. ниЖЕГоРоДовА1, я. М. МоТУЗовА2, М. Ю. КолоБовА1, С. С. БАГАТКА1, н. Ф. МилАнович 3, Г. и. ивАнчиК1
ВЛИЯНИЕ МЕЗЕНХИМАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК НА ПРОЛИФЕРАЦИЮ Т-КЛЕТОК ПАМЯТИ У ПАЦИЕНТОВ С РАССЕЯННЫМ СКЛЕРОЗОМ
1Белорусская медицинская академия последипломного образования, Минск, 2Белорусский государственный медицинский университет, Минск,
39-я городская клиническая больница, Минск, Беларусь
(поступила в редакцию 10.03.2010)
Введение. Рассеянный склероз (РС) представляет собой хроническое прогрессирующее воспа-лительно-дегенеративное аутоиммунное заболевание центральной нервной системы (ЦНС), харак-теризующееся нарушением баланса между регуляторными и потенциально миелин-реактивны-ми клонами Т-лимфоцитов. В результате развиваются специфические эффекторные реакции Т-клеточного иммунитета, направленные на повреждение компонентов миелиновой оболочки аксо-нов, одним из которых является миелин-олигодендроцитарный гликопротеин (МОГ) [1, 2]. В на-стоящее время Т-лимфоциты при РС рассматриваются как мишень для патогенетического лечения, в основе которого лежит иммуносупрессия или индукция толерантности. При этом ведущая роль в развитии РС наряду с миелин-реактивными СD4+ Th1-лимфоцитами, которые вовлекаются в про-цессы демиелинизации оболочек аксонов, отводится цитотоксическим СD8+ T-лимфоцитам [3].
Несмотря на определенные успехи в лечении РС, что связано с использованием препаратов, модулирующих течение заболевания (интерферонов, глатирамера ацетата, пептидных лигандов), а также с проведением ДНК- и Т-клеточной вакцинации, существующие методы селективной антиген-специфической иммунотерапии РС не всегда эффективны [4]. Развитие одного из направ-лений патогенетической терапии РС связано с применением клеточных технологий, в частности с трансплантацией мезенхимальных стволовых клеток (МСК), и основывается на использова-нии иммуносупрессивных свойств МСК и их способности стимулировать процессы репарации в ЦНС [5, 6].
Рядом авторов показано, что аутологичные МСК мышей могут применяться для лечения экс-периментального аллергического энцефаломиелита – Т-медиированной модели аутоиммунного заболевания ЦНС, эффективность лечения которого обусловлена супрессией Т-клеточного отве-та. При этом стромальные клетки, полученные из костного мозга, ингибируют ответ наивных и антиген-специфических Т-клеток памяти на родственный антиген, супрессируют функцию зрелых Т-лимфоцитов, активированных неспецифическими митогенами, и восстанавливают ми-елин при экспериментальном аллергическом энцефаломиелите у мышей [7–9].
В настоящее время точный механизм иммуносупрессивного действия МСК изучен недоста-точно. Показано, что, не являясь клетками иммунной системы, они могут взаимодействовать со всеми популяциями иммунокомпетентных клеток, оказывая регуляторное влияние как клеточно-опосредованным путем, так и за счет синтеза растворимых факторов [10, 11]. Способность МСК модулировать иммунный ответ, низкий уровень риска развития побочных эффектов при транс-плантации этих клеток по сравнению с кортикостероидами, цитостатиками и иммунодепрессан-тами, а также имеющиеся данные о МСК-опосредованном снижении активации иммунокомпе-тентных клеток in vitro и in vivo с сохранением их жизнеспособности могут служить основанием для использования МСК в патогенетической терапии РС [12, 13].
Национальная
академия наук
Беларуси

25
Цель исследования – изучение способности МСК ингибировать in vitro митоген- и миелин-индуцированную пролиферацию эффекторных CD4+ и CD8+ Т-клеток памяти у пациентов с рассеянным склерозом.
Материалы и методы исследования. Материалом для исследования послужили МСК кост-ного мозга 2-го пассажа и мононуклеары периферической крови (МПК), полученные от больных с рецидивно-ремиттирующей формой течения РС в фазе обострения, находившихся на лечении в неврологических отделениях 9-й городской клинической больницы г. Минска, и от здоровых доноров. Характеристика больных РС и здоровых доноров представлена в табл. 1. Верификацию диагноза РС проводили на основании критериев W. McDonald и cоавт. [15] в соответствии с уни-W. McDonald и cоавт. [15] в соответствии с уни-. McDonald и cоавт. [15] в соответствии с уни-McDonald и cоавт. [15] в соответствии с уни- и cоавт. [15] в соответствии с уни-cоавт. [15] в соответствии с уни-оавт. [15] в соответствии с уни-фицированными стандартными методами клинического, инструментального и клинико-лабораторного обследования пациентов. До поступления в стационар и забора биологического материала пациенты не получали иммуносупрессивного или иммуномодулирующего лечения в течение не менее чем 6 мес.
Т а б л и ц а 1. Клинико-демографическая характеристика обследованных групп
Группа Возраст, лет Пол, м/ж Продолжительность заболевания, лет EDSS, балл Биологический материал
Пациенты с РС (n = 13) 31 (23–33) 7/6 4,5 (2,5–7,0) 3,0 (2,0–5,0) ПК (n = 13), КМ (n = 7)Здоровые доноры (n = 14) 30 (23–45) 7/7 – – ПК (n = 14), КМ (n = 7)
П р и м е ч а н и е. Возраст, продолжительность заболевания и EDSS представлены в виде медианы (25–75-й про-EDSS представлены в виде медианы (25–75-й про- представлены в виде медианы (25–75-й про-центили). EDSS (Expanded Disability Status Scale) – оценка неврологического статуса пациентов с РС согласно шкале инвалидизации по Куртцке (cредний балл) [14], ПК – периферическая кровь, КМ – костный мозг.
выделение мононуклеаров из периферической венозной крови и пунктата костного мозга че-ловека. Все этапы забора биологического материала (периферической венозной крови, пунктата костного мозга) и выделения мононуклеарных клеток выполняли в стерильных условиях после получения информированного согласия пациентов о предоставлении материала для исследова-ний. Протокол обследования был утвержден локальным этическим комитетом 9-й городской клинической больницы. Образцы костного мозга получали при трепанобиопсии подвздошной кости. Мононуклеары выделяли путем центрифугирования периферической крови и образцов костного мозга на градиенте плотности Histopaque-1077 (Sigma, Германия). Полученные кольца мононуклеаров дважды отмывали в фосфатном буфере PBS (Gibco, Германия), содержавшем 5% эмбриональной телячьей сыворотки (ЭТС), полученной в НИИЭиМ, Беларусь. Мононуклеары костного мозга использовали для получения пассажей МСК.
Криоконсервация МпК и МСК человека. Длительное хранение МПК и МСК осуществляли с помощью их криоконсервации в питательных средах – DMEM-�G или RPMI-1640 (Gibco) соот-DMEM-�G или RPMI-1640 (Gibco) соот--�G или RPMI-1640 (Gibco) соот-�G или RPMI-1640 (Gibco) соот- или RPMI-1640 (Gibco) соот-RPMI-1640 (Gibco) соот--1640 (Gibco) соот-Gibco) соот-) соот-ветственно, содержавших 20% ЭТС и 10% DMSO (Sigma), используя автоматический криозамо-DMSO (Sigma), используя автоматический криозамо- (Sigma), используя автоматический криозамо-Sigma), используя автоматический криозамо-), используя автоматический криозамо-раживатель и криохранилище на –180 ºС (Entrydell, США).
Культивирование МСК костного мозга человека. Мононуклеары костного мозга ресуспендиро-вали в среде DMEM-�G, содержавшей 10% ЭТС, 1% антибиотика и 1% �-глютамина (Sigma), засе-DMEM-�G, содержавшей 10% ЭТС, 1% антибиотика и 1% �-глютамина (Sigma), засе--�G, содержавшей 10% ЭТС, 1% антибиотика и 1% �-глютамина (Sigma), засе-�G, содержавшей 10% ЭТС, 1% антибиотика и 1% �-глютамина (Sigma), засе-, содержавшей 10% ЭТС, 1% антибиотика и 1% �-глютамина (Sigma), засе-�-глютамина (Sigma), засе--глютамина (Sigma), засе-Sigma), засе-), засе-вали в чашки Петри с покрытием для адгезивных культур (Greiner, Германия) и культивировали в увлажненной атмосфере с 5% СО2 при 37 °С. Через 24 ч и далее каждые 3–4 сут производили замену культуральной среды. По достижении культурами 70–80% конфлюэнтности клетки снима-ли с поверхности пластика трипсином/ЭДТА (Gibco), двукратно отмывали в PBS, содержавшем 10% ЭТС, и засевали в чашки Петри для дальнейшего пассирования. Для микроскопии и монито-ринга культур использовали микроскоп CarlZeiss Axiovert 200 и высокочувствительную камеру CarlZeiss AxioCam MRm с выдержкой 100 мс (CarlZeiss, Германия). Полученные культуры МСК 2-го пассажа имели CD90+/CD105+/CD44+/CD119+/CD31–/CD34–/CD45– фенотип. Полипотентность культур МСК была подтверждена направленной адипо- и остеогенной дифференцировкой.
Совместное культивирование МпК c МСК. МПК больных РС и здоровых доноров, предвари-тельно окрашенные 7 мкМ карбоксифлуоресцеином (CFSE, Sigma), культивировали в концентра-Sigma), культивировали в концентра-), культивировали в концентра-ции 2·105 клеток/лунку 96-луночного планшета в полной культуральной среде RPMI-1640, содер-RPMI-1640, содер--1640, содер-
Национальная
академия наук
Беларуси

26
жавшей 2,5 мкг/мл фитогемагглютинина (ФГА, Sigma) или 10 мкг/мл рекомбинантного миелин-олигодендроцитарного гликопротеина (рМОГ1–125), в присутствии МСК 2-го пассажа или в их отсутствие (МПК:МСК – 10:1) в течение 5 дней (при стимуляции ФГА) и 10 дней (при стимуля-ции рМОГ1–125) в увлажненной атмосфере с 5% СО2 при 37 °С. На 6-й день культивирования осуществляли замену половины культуральной среды с добавлением интерлейкина-2 (Sigma) в конечной концентрации 10 Ед/мл.
проточная цитофлуориметрия. Для цитофлуориметрической оценки фенотипа МПК исполь-зовали моноклональные антитела CD3-PC7, CD4-PC5, CD8-FITC/PC5, CCR7-PE, CD45RO-ECD, для определения пролиферации МПК – CD3-PC7, CD4-PE, CD8-PC5, CD45RO-ECD (Beckman Coulter, США; R&DSystem, Канада). МСК и МПК инкубировали с моноклональными антителами или изотипическими контролями в течение 15 мин в темноте при комнатной температуре. Измере-ния осуществляли на 5-канальном проточном цитофлуориметре FC500 (Beckman Coulter).
Для оценки пролиферативного ответа в соответствии с распределением флуоресценции были установлены границы популяции CD3+ Т-клеток среди живых лимфоцитов, в пределах которой выделяли субпопуляции СD3+CD4+, СD3+CD8+, СD3+CD45RO+, СD4+CD45RO+, СD8+CD45RO+
Т-клеток. Пролиферацию Т-лимфоцитов и их субпопуляций оценивали, рассчитывая процент-ное содержание неподелившихся (CFSEhigh) и поделившихся (CFSElow) Т-клеток после 5 и 10 дней культивирования. Результат регистрировали на 50 000 событий в пробе.
Статистический анализ. Статистическую обработку данных проводили с помощью про-граммы Statistica 6.0. Достоверность различий определяли с использованием непараметрических критериев Вилкоксона (в случае зависимых переменных) и Манна–Уитни (в случае независимых переменных). Корреляционный анализ выполняли путем определения коэффициентов корреляции по Спирмену. Результаты считали достоверными при P < 0,05; P < 0,01; P < 0,001. Полученные данные представлены как медиана (25–75-й процентили).
Результаты и их обсуждение. В зависимости от экспрессии хемокинового рецептора CCR7 различают центральные Т-клетки памяти c фенотипом СD3+CD45RO+CCR7+ и эффекторные Т-клетки памяти c фенотипом СD3+CD45RO+CCR7–, причем последние обладают выраженным ци-тотоксическим потенциалом, способны продуцировать перфорины, γ-интерферон и участвовать в повреждении миелиновой оболочки аксонов [16]. Нами было установлено достоверное увеличе-ние количества эффекторных t-клеток памяти в периферической крови больных РС по сравнению с их уровнем в контрольной группе (17,0% (14,5–20,5%) и 13,7% (8,9–15,4%) соответственно, P<0,05), главным образом за счет увеличения популяции CD8+CD45RO+CСR7– Т-лимфоцитов (12,6% (8,4–19,5%) – у больных РС; 5,4% (2,7–6,7%) – у здоровых доноров, P < 0,001).
Известно, что эфферентная фаза иммунной реакции развивается в результате рестимуляции центральных T-клеток памяти резидентными антиген-презентирующими клетками и превращения их в терминальные эффекторные Т-клетки памяти, теряющие СCR7 и СD27 маркеры [17]. Показано, что на Т-лимфоцитах, выделенных из поврежденных очагов больных РС, отсутствует экспрессия CCR7, что указывает на локальную дифференцировку центральных T-клеток памяти после стиму-ляции антигеном в ЦНС, при этом дифференцировка Т-лимфоцитов определяется интенсивностью антигенной стимуляции [18, 19]. Таким образом, повышение количества эффекторных Т-клеток па-мяти в периферической крови больных РС по сравнению с лицами контрольной группы, возможно, обусловлено хронической антигенной стимуляцией при развитии аутоиммунных реакций.
Наряду с этим у больных РС по сравнению с лицами контрольной группы наблюдалось до-стоверное увеличение общего количества CD3+СD45RO+ Т-лимфоцитов, причем как CD4+CD45RO+, так и CD8+CD45RO+ Т-клеток (рис. 1, А, Б). Это согласуется с данными ряда авторов о роли в разви-тии эффекторных специфических реакций при РС не только CD4+ Th1-лимфоцитов, но и цитоток-сических Т-клеток, экспрессирующих CD8+ маркер [20, 21]. Кроме того, нами было установлено, что количество эффекторных Т-клеток памяти достоверно коррелирует с общим процентным со-держанием Т-клеток памяти в периферической крови как у здоровых доноров (R = 0,64; P<0,02), так и у пациентов с РС (R = 0,76; P < 0,01). Полученные данные позволили в дальнейшем исполь-зовать фенотип CD45RO+ Т-лимфоцитов для характеристики функциональных свойств эффек-торных CD4+ и CD8+ Т-клеток памяти in vitro.
Национальная
академия наук
Беларуси

27
Для оценки влияния МСК на пролиферацию Т-клеток памяти у 7 больных РС и у 7 здоро-вых доноров нами были изучены ФГА- и МОГ-индуцированные пролиферативные ответы CD3+CD45RO+ Т-лимфоцитов и их субпопуляций в присутствии аутологичных и аллогенных МСК 2-го пассажа и в их отсутствие.
Оценку пролиферации проводили методом количественного анализа клеточного деления in vitro, основанного на использовании внутриклеточного красителя CFSE, флуоресценция которо-го снижается пропорционально увеличению количества клеточных делений. На рис. 2 представ-
Рис. 1. Количество CD4+CD45RO+ (a) и CD8+CD45RO+ (Б) Т-клеток памяти в периферической крови у пациентов с РС (n = 13) и у здоровых доноров (n = 14). Достоверность различий: * – Р < 0,05; ** – Р < 0,01
Рис. 2. Пролиферативный ответ CD3+CD45RO+ Т-лимфоцитов в ответ на действие ФГА в присутствии аутологичных МСК 2-го пассажа и в их отсутствие у здорового донора. В верхней панели точечных диаграмм по оси Х отображено распределение CFSE+ Т-лимфоцитов, по оси y – CD3+CD45RO+ Т-лимфоцитов. Нижняя панель гистограмм является проекцией точечных диаграмм верхней панели и отображает распределение CFSElow- и CFSEhigh-клеток; на гисто-граммах указано процентное содержание поделившихся CFSElow Т-клеток памяти. А – спонтанная пролиферация Т-клеток памяти; Б – ФГА-индуцированная пролиферация Т-клеток памяти; в – ФГА-индуцированная пролиферация
Т-клеток памяти в присутствии аутологичных МСК 2-го пассажа
Национальная
академия наук
Беларуси

28
лены оригинальные точечные диаграммы и их проекции в виде гистограмм, полученных в результате проточной цитофлуориметрии, которые отражают изменение пролиферации СD3+СD45RO+ Т-лимфоцитов здорового донора в ответ на поликлональный митоген ФГА в при-сутствии аутологичных МСК или в их отсутствие. На рис. 2, А отображена спонтанная пролифе-рация СD3+СD45RO+ Т-лимфоцитов в 5-дневной культуре (количество поделившихся CFSElow Т-клеток памяти – 2,0%). Так, добавление ФГА (рис. 2, Б) значительно стимулировало пролифе-рацию Т-клеток памяти, и доля поделившихся клеток составила 71,1%. В то же время в присут-ствии аутологичных МСК (рис. 2, в) количество пролиферирующих СD3+СD45RO+ Т-лимфоцитов в ответ на действие митогена снизилось до 40,6%, что составило, соответственно, 43% супрессии ФГА-индуцированной пролиферации Т-клеток памяти.
Обобщенные результаты пролиферации Т-клеток памяти и их субпопуляций, культивируе-мых в присутствии МСК или в их отсутствие в условиях митогенной стимуляции, у больных РС и у здоровых доноров представлены в табл. 2. У больных РС отмечено увеличение спонтанной пролиферации Т-лимфоцитов по сравнению с таковой в группе здоровых доноров, при этом до-стоверные изменения выявлены только в популяции СD8+CD45RO+ Т-лимфоцитов (P<0,05, до-стоверность в табл. 2 не указана). При неспецифической стимуляции ФГА в Т-клетках памяти больных РС, так же как и у здоровых доноров, отмечалось одинаковое процентное содержание поделившихся клеток. Добавление МСК в культуру ФГА-стимулированных МПК приводило к достоверному снижению пролиферации СD3+CD45RO+ Т-клеточных субпопуляций и у боль-ных РС, и у здоровых доноров, причем наиболее выраженным было снижение ответа СD4+CD45RO+
Т-лимфоцитов. Наряду с этим отмечалась тенденция к супрессии антиген-неспецифической про-лиферации СD8+CD45RO+ Т-лимфоцитов (табл. 2).
Т а б л и ц а 2. Количество поделившихся Т-клеток памяти (%), культивируемых в присутствии МСК и в их отсутствие, в ответ на действие митогена у пациентов с РС и у здоровых доноров
Условия культивирования
Здоровые доноры (n = 7) Больные РС (n = 7)
CD3+45RO+ CD4+45RO+ CD8+45RO+ CD3+45RO+ CD4+45RO+ CD8+45RO+
Культуральная среда 3,0 (1,6–6,6)
3,5 (2,6–6,6)
9,4 (6,2–12,4)
6,4 (3,5–10,0)
7,9 (3,2–14,2)
24,2 (17,9–38,4)
ФГА 66,5 (57,0–70,0)
58,0 (56,7–79,5)
85,3 (71,5–89,3)
66,1 (52,1–83,7)
66,5 (54,3–80,8)
83,6 (70,0–90,7)
ФГА+МСК 39,7* (39,0–57,1)
39,6* (28,0–39,8)
73,8P = 0,07
(61,6–75,4)42,3*
(29,0–51,8)44,7*
(25,2–61,2)70,4 P = 0,06
(58,8–78,9)
П р и м е ч а н и е. Достоверность различий (* – P < 0,01) указана по отношению к ФГА. ФГА – фитогемагглюти-нин, МСК – мезенхимальные стволовые клетки костного мозга.
Для характеристики степени выраженности ингибирующего влияния МСК на клеточное де-ление Т-лимфоцитов в ответ на действие митогена были рассчитаны индексы супрессии (ИС). Установлено, что ИС МСК в отношении митоген-индуцированной пролиферации СD3+ Т-клеток памяти больных РС и здоровых доноров достоверно не различались, однако в обеих субпопуля-циях исследуемых групп наблюдалась наиболее выраженная супрессия пролиферации СD4+45RO+
Т-лимфоцитов. Так, у больных РС ИС СD3+45RO+ Т-клеток составил 42,3% (40,3–44,3%), СD4+45RO+
Т-клеток – 49,9% (30,2–51,7%) и СD8+45RO+ Т-клеток – 21,7% (15,6–27,8%), а у здоровых – 41,9% (34,6–52,6%), 47,4% (33,2–72,6%) и 15,5% (6,4–25,5%) соответственно. При этом нами не было вы-явлено достоверных отличий в супрессивной активности аллогенных и аутологичных МСК как у больных РС, так и у здоровых доноров, а степень выраженности супрессии МСК у больных РС была аналогична таковой у здоровых доноров (рис. 3, А).
Для оценки влияния МСК на миелин-специфическую пролиферацию Т-клеток памяти боль-ных РС и здоровых доноров был использован один из основных иммунодоминантных миелино-вых белков – рМОГ1–125, который, в отличие от других миелиновых антигенов, обнаруживается не только в компактных миелиновых волокнах оболочки аксонов, но и на внешней поверхности мембраны олигодендроцитов, представляя собой, таким образом, наиболее вероятную мишень для аутореактивных Т-лимфоцитов [2]. Показано, что Т-клетки памяти как пациентов с РС,
Национальная
академия наук
Беларуси
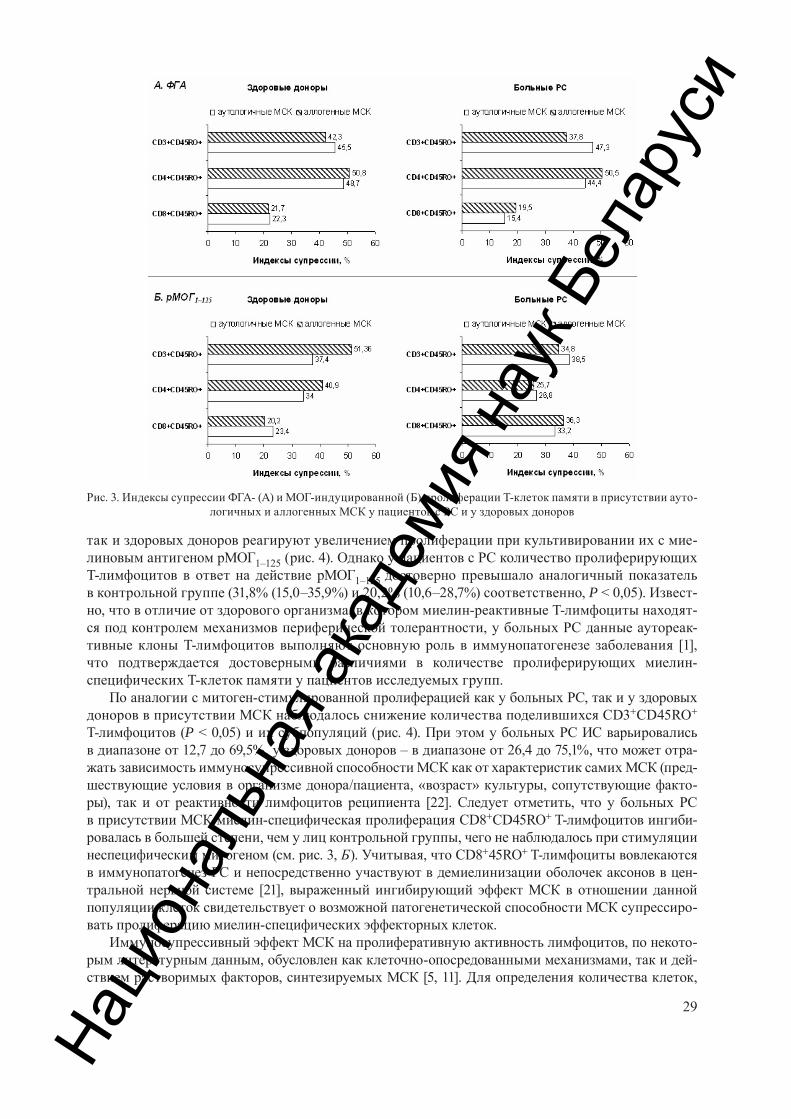
29
так и здоровых доноров реагируют увеличением пролиферации при культивировании их с мие-линовым антигеном рМОГ1–125 (рис. 4). Однако у пациентов с РС количество пролиферирующих Т-лимфоцитов в ответ на действие рМОГ1–125 достоверно превышало аналогичный показатель в контрольной группе (31,8% (15,0–35,9%) и 20,2% (10,6–28,7%) соответственно, P < 0,05). Извест-но, что в отличие от здорового организма, в котором миелин-реактивные Т-лимфоциты находят-ся под контролем механизмов периферической толерантности, у больных РС данные аутореак-тивные клоны Т-лимфоцитов выполняют основную роль в иммунопатогенезе заболевания [1], что подтверждается достоверными различиями в количестве пролиферирующих миелин-специфических Т-клеток памяти у пациентов исследуемых групп.
По аналогии с митоген-стимулированной пролиферацией как у больных РС, так и у здоровых доноров в присутствии МСК наблюдалось снижение количества поделившихся CD3+CD45RO+
Т-лимфоцитов (P < 0,05) и их субпопуляций (рис. 4). При этом у больных РС ИС варьировались в диапазоне от 12,7 до 69,5%, у здоровых доноров – в диапазоне от 26,4 до 75,1%, что может отра-жать зависимость иммуносупрессивной способности МСК как от характеристик самих МСК (пред-шествующие условия в организме донора/пациента, «возраст» культуры, сопутствующие факто-ры), так и от реактивности лимфоцитов реципиента [22]. Следует отметить, что у больных РС в присутствии МСК миелин-специфическая пролиферация CD8+CD45RO+ Т-лимфоцитов ингиби-ровалась в большей степени, чем у лиц контрольной группы, чего не наблюдалось при стимуляции неспецифическим митогеном (см. рис. 3, Б). Учитывая, что CD8+45RO+ Т-лимфоциты вовлекаются в иммунопатогенез РС и непосредственно участвуют в демиелинизации оболочек аксонов в цен-тральной нервной системе [21], выраженный ингибирующий эффект МСК в отношении данной популяции клеток свидетельствует о возможной патогенетической способности МСК супрессиро-вать пролиферацию миелин-специфических эффекторных клеток.
Иммуносупрессивный эффект МСК на пролиферативную активность лимфоцитов, по некото-рым литературным данным, обусловлен как клеточно-опосредованными механизмами, так и дей-ствием растворимых факторов, синтезируемых МСК [5, 11]. Для определения количества клеток,
Рис. 3. Индексы супрессии ФГА- (А) и МОГ-индуцированной (Б) пролиферации Т-клеток памяти в присутствии ауто-логичных и аллогенных МСК у пациентов с РС и у здоровых доноров
Национальная
академия наук
Беларуси

30
поделившихся в результате митогенной/антигенной стимуляции от 1 до 7 раз, были проведены до-полнительные расчеты, основанные на вычислении антилогарифма среднего геометрического пика флуоресценции неподелившихся (не подвергшихся митозу) клеток. На рис. 5 представлено количество клеточных генераций, стимулированных ФГА (рис. 5, А) и рМОГ1–125 (рис. 5, Б), всту-пивших в митоз от 1 до 7 раз (М1–М7), как в присутствии культур МСК, так и в их отсутствие. В присутствии МСК наблюдалось достоверное увеличение (в 1–2 раза) количества клеток, всту-пивших в митоз, и в то же время резкое снижение количества клеток, поделившихся 7 и более раз в результате митогенной и антигенной стимуляции. Таким образом, можно предположить, что МСК проявляют супрессивные свойства в отношении митоген/миелин-индуцированной пролифе-рации лимфоцитов путем остановки клеточного деления ранних (М1–М2) генераций.
R. Newman и соавт. [22] показали, что в зависимости от состояния клетки МСК защищают покоящиеся Т-лимфоциты от клеточной гибели, останавливают Т-клетки в фазе G0/G1 клеточного
Рис. 4. Количество поделившихся CD4+CD45RO+ (a) и CD8+CD45RO+ (Б) Т-клеток памяти, культивируемых в при-сутствии МСК и в их отсутствие, в ответ на действие миелинового антигена у пациентов с РС и у здоровых доноров. Культ. среда – культуральная среда; рМОГ1–125 – рекомбинантный человеческий миелин-олигодендроцитарный гли-копротеин с аминокислотной последовательностью 1–125; МСК – мезенхимальные стволовые клетки костного мозга.
Достоверность различий: * – Р < 0,05; ** – Р < 0,01
Рис. 5. Количество поделившихся CD3+CD45RO+ Т-лимфоцитов в клеточных генерациях (М1–М7), культивируемых в присутствии МСК и в их отсутствие, в ответ на действие митогена (А) и миелинового антигена (Б) у пациентов
с РС и у здоровых доноров. Достоверность различий: * – Р < 0,05; ** – Р < 0,01
Национальная
академия наук
Беларуси

цикла и способствуют развитию апоптоза активированных лимфоцитов. Полученные нами дан-ные согласуются с результатами S. Glennie и соавт. [8], которые установили, что внутриклеточ-ные механизмы МСК-индуцируемой супрессии могут быть обусловлены снижением экспрессии циклина D2, cdk2 и увеличением экспрессии ингибитора клеточного цикла – p27Kip1, приводя-щих к остановке клеточного цикла как в контрольной точке G1/S, так и в точке G2/M.
Заключение. Показано, что в периферической крови пациентов с РС достоверно увеличено количество эффекторных СD4+ и СD8+ Т-клеток памяти. Пролиферация последних при миелин-индуцированной стимуляции in vitro достоверно превышала аналогичный показатель в кон-трольной группе, что отражает вовлечение данных субпопуляций лимфоцитов в патогенетиче-ские механизмы развития РС. Установлено, что аллогенные и аутологичные культуры МСК ингибируют антиген-неспецифическую и миелин-индуцированную пролиферацию Т-клеток па-мяти больных РС. При этом механизм МСК-индуцируемой иммуносупрессии может быть связан с остановкой клеточного деления на уровне ранних (М1–М2) генераций. Полученные результаты могут быть использованы для индивидуального подхода к оценке иммуномодулирующих свойств МСК и дальнейшего применения клеточных культур в патогенетической терапии РС.
Литература
1. S o s p e d r a M., M a r t i n R. // Ann. Rev. Immunol. 2005. N 23. P. 683–747.2. I g l e s i a s A. et al. // Glia. 2001. N 36. P. 220–234.3. H a f l e r D. // J. of Clin. Invest. 2004. N 6. P. 788–794.4. S t e i n m a n �. // Curr. Opin. Immunol. 2001. N 5. P. 597–600.5. U c c e l l i A. et al. // Sum. Exp. Opin. on Biol. Ther. 2006. N 6. P. 17–22.6. F r e e d m a n M. et al. // Multiple Sclerosis. 2010. N 1. P. 1–8.7. K r a m p e r a M. et al. // Stem Cells. 2005. N 24. P. 386–398.8. G l e n n i e S. et al. // Blood. 2005. N 105. P. 2821–2827.9. Z a p i a E. et al. // Blood. 2005. N 106. P. 1755–1761.10. J i a n g �. et al. // Nature. 2002. N 418. P. 41–49.11. A g g a r w a i S., P i t t e n g e r M. // Blood. 2004. N 105. P. 1815–1822.12. G u a n q w u X. et al. // Cell Res. 2007. N 17. P. 240–248.13. R a s m u s s o n I. et al. // Exp. Cell. Res. 2005. N 305. P. 33–41.14. K u r t z k e J. // Neurology. 1983. N 11. P. 1444–1452.15. M c D o n a l d W. еt al. // Ann. Neurol. 2001. N 50. P. 121–127.16. С a m p b e l l J. еt al. // Immunology. 2001. N 166. P. 877–884.17. S a l l u s t o F. еt al. // Annu. Rev. Immunol. 2004. N 22. P. 745–763.18. K i v i s a P. еt al. // Ann. Neurol. 2004. N 55. P. 627–638. 19. W i l l i n g e r T. еt al. // Immunology. 2005. N 175. P. 5895–5903.20. F r i e s e M. еt al. // Brain. 2005. N 128. P. 1747–1763.21. Z a f r a n s k a y a M. еt al. // Immunology. 2007. N 1. P. 29–39.22. N e w m a n R. еt al. // Inflammation and allergy. 2009. N 8. P. 110–123.
М. М. ZaFRanSKaya1, a. S. FEdulov2, d. B. niZhEhaRodava1, J. M. MoTuZova 2, M. J. KoloBova1, S. S. BaGaTKa1, n. F. Milanovich3, G. i. ivanchiK1
INFLUENCE OF MESENCHYMAL STEM CELLS ON THE MEMORY T-CELLS PROLIFERATION IN PATIENTS WITH MULTIPLE SCLEROSIS
1Belarusian Medical academy of postgraduate Education, Minsk, 2Belarusian State Medical university, Minsk,
39th Municipal hospital, Minsk, Belarus
Summary
The perspective tendency in pathogenetic therapy of central nervous system’s inflammatory-degenerative autoimmune diseases is based on mesenchymal stem cells (MSC) immunoregulatory properties. Using the intracellular fluorescent dye CFSE-method to monitor cell division we have shown that allogenic and autologous MSC inhibit in vitro mitogen- and my-elin-induced proliferation of pathogenetically important effector memory СD4 + and СD8 + Т-cells in multiple sclerosis patients. The mechanism of MSC-mediated immunosuppression can be caused by cell division arrest of early (М1–М2) gen-erations. The obtained results may be used for the individual approach for estimating MSC immunomodulatory properties and their further application of cell cultures in pathogenetic therapy of multiple sclerosis.
Национальная
академия наук
Беларуси

32
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.72-018.3-001-003.93:617-089.855
о. л. ЭЙСМонТ, л. А. пАшКЕвич, Б. в. МАлЮК, н. о. ГолУТвинА, А. в. БоРиСов, Д. в. БУКАч
ВЛИЯНИЕ ПЕРФОРАЦИИ ОСТЕОХОНДРАЛЬНОй ПЛАСТИНКИ НА РЕГЕНЕРАЦИЮ ПОВРЕЖДЕННОГО СУСТАВНОГО ХРЯЩА В ЭКСПЕРИМЕНТЕ
Республиканский научно-практический центр травматологиии ортопедии, Минск, Беларусь
(поступила в редакцию 25.03.2010)
Одной из актуальных проблем современной артрологии является проблема лечения локаль-ных повреждений и заболеваний суставного хряща. Известно, что нелеченое повреждение хря-ща, независимо от генеза, не обладает способностью спонтанного излечения и может приводить в дальнейшем к распространенному дегенеративному поражению сустава, прогрессивной поте-ре хрящевой ткани и возникновению необратимых вторичных артрозных изменений [1]. Однако хирургическое вмешательство на изолированном дефекте хряща может приостановить или даже предотвратить развитие генерализованного остеоартроза [2, 3].
К одной из групп хирургических вмешательств при лечении повреждений суставного хряща относят операции, направленные на создание условий для стимуляции хондрогенеза путем перфорации субхондральной костной пластинки [2, 4]. Первоначально они имели полуэмпири-ческое обоснование, суть которого заключалась в воспроизведении классического процесса заживления тканевой раны путем ее реваскуляризации [5, 6]. В последующем появилось много теоретических и экспериментальных данных, указывающих на положительные эффекты форми-рования на перфорированной костной пластинке фибринного сгустка, содержащего мезенхи-мальные стволовые клетки костного мозга и факторы роста [7, 8]. Под влиянием биологических и механических факторов синовиальной среды полипотентные стволовые клетки способны трансформироваться в хондроидные и продуцировать волокнисто-хрящевую ткань, восстанав-ливая поврежденную суставную поверхность [4]. В отличие от гиалинового хряща эта ткань со-держит преимущественно коллаген I типа и в меньшей степени II, VI и IX типов, а также имеет пониженную концентрацию протеогликанов [9]. Наряду с этим в ряде работ была продемонстри-рована возможность формирования в ранних стадиях (19 недель) участков ткани, гистологически подобных строению гиалинового хряща, которые в последующем подвергались фибротизации [4].
Другой предпосылкой для более активного применения остеоперфоративных методик лечения повреждений хряща в клинической практике последнего времени стало внедрение эндоскопиче-ской хирургии, которая сделала хирургическое вмешательство малоинвазивным по сравнению с техникой открытой операции [2, 3]. Таким образом, классическими показаниями к применению остеоперфорации в очаге поражения считается наличие глубоких полнослойных дефектов сустав-ного хряща IV степени по R. E. Outerbridge [10].
Среди данных хирургических вмешательств наибольшую популярность получили туннели-зация и микрофрактурирование [4–6, 10, 11].
Субхондральная туннелизация, впервые предложенная K. H. Pridie в 1959 г. [6], заключается в просверливании хряща и склерозированной субхондральной кости до появления хорошо выражен-ного кровотечения из спонгиозной кости. В результате образуется волокнистый хрящ, восстанав-ливающий в месте дефекта гладкую суставную поверхность. Такие хирургические вмешательства выполняются не только в случаях глубоких дефектов покровного хряща, но и при рассекающем
Национальная
академия наук
Беларуси

33
остеохондрите и остеонекрозе [12, 13], когда происходит некроз подлежащей субхондральной и спонгиозной костной ткани.
Более современной хирургической технологией, которая широко апробируется в клиниче-ской практике в последние десятилетия, является микрофрактурирование субхондральной кост-ной пластинки. По сути данная техника остеоперфорации сводится к образованию микропере-ломов последней с сохранением ее каркасной целостности [10].
Несмотря на то что различные остеоперфоративные вмешательства при повреждениях су-ставного хряща относят к группе операций, стимулирующих полипотентные стволовые клетки костного мозга, оценка их результативности неоднозначна [14, 15]. На этом основании ведущими учеными мира поддерживается мнение о перспективности и приоритетности исследований в об-ласти изучения результативности остеоперфоративных методик лечения повреждения хряща, уточнения показаний к их применению, разработки новых способов стимуляции хондрогенеза и нормализации физиологических параметров суставной среды [1, 11, 15].
Целью настоящего исследования стало изучение процессов хондро- и остеорегенерации при различных видах перфорации субхондральной кости в эксперименте на животных.
Материалы и методы исследования. Исследования проводили на экспериментальной моде-ли полнослойного повреждения суставного хряща коленных суставов. Использовали 6 беспород-ных собак мужского пола массой 13,6 ± 2,4 кг. Моделирование хронического полнослойного по-вреждения суставного гиалинового хряща у животных выполняли путем полного механического удаления хрящевой ткани площадью 0,25 см² по методике Д. А. Маланина [7] (рис. 1). Поврежде-ние суставного хряща производили на внутреннем мыщелке бедра коленного сустава в зоне наи-большей осевой нагрузки, так как данная локализация поражения наиболее характерна при раз-витии посттравматического остеоартроза и рассекающего остеохондрита [16].
После премедикации нейролептиком (0,25%-ный раствор дроперидола из расчета 0,5 мг/кг) в качестве наркотического вещества применяли 5%-ный раствор нембутала из расчета 0,5 мл/кг внутрибрюшинно.
Все экспериментальные животные были разделены на две группы. В первой группе, вклю-чавшей трех животных (6 суставов), через 21 сут после формирования повреждения хряща была выполнена туннелизация по Pridie очага деструкции суставной поверхности медиального мы-Pridie очага деструкции суставной поверхности медиального мы- очага деструкции суставной поверхности медиального мы-щелка бедра коленных суставов. После выполнения миниартротомии с помощью спицы Кирш-нера производили глубокое (не менее 1 см) просверливание в субхондральной пластинке на дне дефекта хряща туннелизационных отверстий диаметром 1,5 мм на расстоянии 1,5–2,0 мм друг от друга [11]. Далее рану послойно ушивали (наглухо, без дренирования).
Во второй группе использовано трое животных (6 суставов), которым после формирования повреждения хряща осуществляли микрофрактурирование очага деструкции. Микрофрактуры выполнялись шилом-перфоратором на заданную глубину (около 2–3 мм) с тем же интервалом, что и в первой группе. В результате этого на субхондральной костной пластинке образовывалась шероховатая поверхность, на которой появлялись капли «росы» из крови и жира [10].
Рис. 1. Формирование повреждения внутреннего мыщелка бедра коленного сустава в эксперименте на собаках: хрящевая рана в момент ее формирования (А) и через 21 день после операции (Б)
А Б
Национальная
академия наук
Беларуси

34
С целью формирования в дефекте суставной поверхности прочной репаративной ткани в по-слеоперационном периоде нагрузка на оперированную конечность была частично ограничена в течение 3–4 недель.
Животных обеих групп выводили из эксперимента одновременно – через 6 мес. после опера-тивного вмешательства. Для исследования забирались мыщелки бедра коленных суставов, их обработку осуществляли по общепринятой методике. Материал фиксировали в 10%-ном раство-ре нейтрального формалина, декальцинацию производили в 10%-ном растворе муравьиной кис-лоты, обезвоживали в спиртах восходящей концентрации и заливали в парафиновые блоки. Па-рафиновые срезы толщиной 3–6 мкм окрашивали гематоксилином и эозином и исследовали под световым микроскопом �eica (Германия) с увеличением от 50 до 400.
Проведение экспериментов и умерщвление наркотизированных животных регламентирова-лись требованиями соответствующих приказов и методических рекомендаций [17].
Результаты и их обсуждение. При микроскопическом исследовании были выявлены следу-ющие изменения.
У животных первой группы, которым была проведена туннелизация мыщелка, зона туннели-зации распространяется за субхондральные отделы кости на значительную глубину. В области дефекта наблюдаются процессы дифференцировки стволовых клеток в хрящевые клеточные эле-менты, продуцирующие хрящевое межклеточное вещество, и в фибробласты, продуцирующие волокнистые структуры. Это приводит к закрытию дефекта грубоволокнистой соединительной и хондроидной тканью, напоминающей по строению волокнистый хрящ с немногочисленными кровеносными сосудами, количество которых увеличивается в периферических и глубоких от-делах дефекта, т. е. вблизи от неповрежденнных структур мыщелка (рис. 2). В толще этой ткани в одном наблюдении определялся очаговый хондрогенез с формированием мелких фокусов зре-лого гиалинового хряща (рис. 3). Во всех наблюдениях дефекты закрыты полностью и целост-ность наружной суставной поверхности восстановлена.
От окружающей ткани зона дефекта отграничена нечетко выраженным слоем местами рых-лой волокнистой, местами более плотной соединительной ткани грануляционного типа, в кото-рой обнаруживается активная пролиферация клеточных элементов и значительное количество расширенных и полнокровных сосудов. Кроме того, на фоне остеокластического рассасывания предсуществующей костной ткани происходило формирование молодого остеоидного вещества с образованием костных балочек, что говорит об активной регенерации костной ткани (рис. 4). В межбалочных пространствах видна рыхлая фибро-ретикулярная ткань с диффузно располо-женными гистиоцитарными и лимфоидными элементами. По линии дефекта новообразованная хондроидная и грубоволокнистая соединительная ткань непосредственно срастается с костны-
Рис. 2. Микрофото суставной поверхности при туннелиза-ции (×50). Окраска гематоксилином и эозином. Определя-ется дефект суставной поверхности, заполненный грубо-волокнистой соединительной тканью. Стрелкой указана
зона хондрогенеза
Рис. 3. Микрофото той же суставной поверхности при тун-нелизации (×200). Окраска гематоксилином и эозином. Среди грубоволокнистой ткани определяется участок зре-лого гиалинового хряща с хорошо прокрашенными хон-дроцитами и намечающейся субхондральной пластинкой
Национальная
академия наук
Беларуси

35
ми и хрящевыми структурами мыщелка, однако можно четко определить границу перехода предсуществующего суставного гиалинового хряща в зону грубоволокнистой соединительной ткани, заполняющей дефект после туннелизации (рис. 5).
У животных второй группы глубина прокола небольшая, затрагивает только субхондраль-ные отделы. Зона дефекта в 4 случаях заполнена незрелой хондроидной тканью (рис. 6, а), в ко-торой четко определяются хондроциты. В глубине дефекта они имеют капсулы и выстраиваются в колонноподобные структуры, а по мере приближения к наружной суставной поверхности приобретают более уплощенную форму (рис. 6, б). По периферии данной ткани определяются немногочисленные кровеносные сосуды. В двух наблюдениях, так же как и в первой группе, де-фект закрыт грубоволокнистой соединительной и хондроидной тканью (рис. 7) с немногочис-ленными кровеносными сосудами и очаговым хондрогенезом и формирующимися участками зрелого гиалинового хряща (рис. 8).
По периферии зоны дефекта в костных структурах мыщелка определяются незначительная пролиферация тонкостенных сосудов и клеточных элементов, слабая остеокластическая резорб-ция; по мере удаления от места микрофрактурирования склероз костной ткани, межбалочное пространство местами заполнено рыхлой волокнистой соединительной тканью с единичными кровеносными сосудами и скудной диффузной кругло-клеточной инфильтрацией, местами – жи-ровым костным мозгом (см. рис. 6, а).
Рис. 4. Микрофото субхондральных отделов кости при туннелизации (×200). Окраска гематоксилином и эози-ном. Определяется выраженное остеокластическое рас-сасывание костных балок и формирование новых, полно-
кровие сосудов
Рис. 5. Микрофото суставной поверхности при тунне-лизации (×200). Окраска гематоксилином и эозином. Четко видна граница между гиалиновым хрящом, по-крывающим сустав, и грубоволокнистой соедини-
тельной тканью, заполняющей дефект
Рис. 6. Микрофото суставной поверхности при микрофрактурировании: а – зона микрофрактурирования, располо-жена в субхондральной области, выполнена волокнистым хрящом, закрыта со стороны суставной поверхности гиа-линовым хрящом (×100); б – участок, напоминающий по строению гиалиновый хрящ (×200). Окраска гематоксили-
ном и эозином
а б
Национальная
академия наук
Беларуси

36
Заключение. При глубокой туннелизации по Pridie костной ткани в зоне дефекта суставно-Pridie костной ткани в зоне дефекта суставно- костной ткани в зоне дефекта суставно-го хряща происходят процессы дифференцировки стволовых клеток в хрящевые клеточные элементы, которые продуцируют хрящевое межклеточное вещество, и в фибробласты, проду-цирующие волокнистые структуры. Это приводит к полному закрытию дефекта грубоволок-нистой соединительной и хондроидной тканью с формированием очагов хондрогенеза внутри нее и восстановлению целостности наружной суставной поверхности. Кроме того, по перифе-рии зоны туннелизации наблюдается активная пролиферация клеточных элементов и крове-носных сосудов, что способствует улучшению тканевого питания и процессам репаративной регенерации костной ткани.
При микрофрактурировании субхондральной костной пластинки очаг деструкции заполняется в большинстве случаев незрелой хондроидной тканью и процессы реваскуляризации и репаратив-ной регенерации в окружающей костной ткани выражены слабее, чем при туннелизации по Pridie.
Таким образом, при рассекающем остеохондрите мыщелков бедра коленного сустава показано применение глубокой туннелизации пораженной костной ткани по Pridie. В отличие от микро-Pridie. В отличие от микро-. В отличие от микро-фрактурирования она позволяет не только закрыть дефект суставной поверхности грубоволокни-стой соединительной тканью с участками гиалинового хряща, но и вызвать в очаге деструкции костной ткани процессы остеорегенерации.
Литература
1. B u c k w a l t e r J. A., M a n k i n H. J. // J. Bone Joint Surg. 1997. Vol. 79A. P. 612–632. 2. B u c k w a l t e r J. A., � o h m a n d e r S. // J. Bone Joint Surg. (Am.) 1994. Vol. 76. Р. 1405–1418.3. � e v y A., � o h n e s J., S c u l l y S. et al. // Am. J. Sports Med. 1996. Vol. 24. P. 634–639. 4. J o h n s o n �. �. // Arthroscopy. 1986. Vol. 2. P. 54–69. 5. F i c a t R. P., F i c a t C., G e d e o n P., T o u s s a i n t J. B. // Clin. Orthop. 1979. Vol. 144. P. 74–83. 6. P r i d i e K. H. // J. Bone Joint Surg. (Br.) 1959. Vol. 41. P. 618–619. 7. М а л а н и н Д. А. // Пластика полнослойных дефектов гиалинового хряща в коленном суставе: эксперимен-
тальные и клинические аспекты репаративного хондрогенеза: автореф. дис. ... д-ра мед. наук: 14.00.15, 14.00.22. Вол--ра мед. наук: 14.00.15, 14.00.22. Вол-ра мед. наук: 14.00.15, 14.00.22. Вол- мед. наук: 14.00.15, 14.00.22. Вол-мед. наук: 14.00.15, 14.00.22. Вол-. наук: 14.00.15, 14.00.22. Вол-наук: 14.00.15, 14.00.22. Вол-: 14.00.15, 14.00.22. Вол-Вол-гоград, 2002.
8. S h a p i r o F., K o i d e S., G l i m c h e r M. // J. Bone Joint Surg. (Am.) 1993. Vol. 75. P. 532–553. 9. B u c k w a l t e r J. A., R o s e n b e r g �. C., H u n z i k e r E. B. // Articular cartilage: composition, structure,
response to injury, and methods of facilitation repair / Eds. J. W. Ewing. N. �., 1990. P. 19–56.10. S t e a d m a n J. R., R o d k e y W. G., B r i g g s K. K. // Sports Med. Arthrosc. Rev. 2003. Vol. 11. P. 236–244.11. I n s a l l J. N. Surgery of the Knee. N. �.; Edinburg; �ondon; Melbourne, 1984. 12. C a i n E. �., C l a n c y W. G. // Clinics in Sports Medicine. 2001. Vol. 20, N 2. P. 236–248. 13. � o u i s i a S., B e a u f i l s P., K a t a b i M., R o b e r t H. // Knee Surg. Sports Traumatol. Arthroscopy. 2003.
Vol. 11. P. 33–39.
Рис. 7. Микрофото суставной поверхности при микро-фрактурировании (×200). Окраска гематоксилином и эози-ном. Зона дефекта выполнена волокнистым хрящом, по-
крыта гиалиновым хрящом обычного строения
Рис. 8. Микрофото суставной поверхности при ми-крофрактурировании (×100). Окраска гематоксили-ном и эозином. Зона дефекта выполнена волокнистым хрящом, в котором определяется участок образова-ния молодого гиалинового хряща. Дефект покрыт
бесклеточным участком гиалинового хряща
Национальная
академия наук
Беларуси

14. A n d e r s S., S c h a u m b u r g e r J., G r i f k a J. // Orthopäde. 2001. B. 30. S. 866–880. 15. J o h n s o n �. �. // Clin. Orthop. 2001. Vol. 391S. P. 306–317. 16. М и р о н о в С. П., О м е л ь я н е н к о Н. П., Т р о ц е н к о В. В. и др. // Вестн. травматол. и ортопед.
им. Н. Н. Приорова. 2009. № 2. С. 3–12.17. Эвтаназия экспериментальных животных: метод. рекомендации. М., 1985.
o. l. EiSMonT, l. a. paShKEvich, B. V. MaluK, n. o. GoluTvina, a. v. BoRiSov, d. v. BuKach
INFLUENCE OF THE OSTEOCHONDRAL PLATE PERFORATION ON THE OSTEOCHONDRAL REGENERATION OF EXPERIMENTAL DАMAGE OF JOINT CARTILAGE
Republican Scientific-practical centre of Traumatology and orthopedic Surgery of Belarus, Minsk
Summary
The research results of hondro- and osteoregeneration processes in treatment of experimental full-layer damage of articular cartilage of canine knee joints by perforation of underlying bone are presented. It was established that after deep tunneling by Pridie of bone tissue the defect zone is characterized by differentiation processes of trunk cells to cartilage cellular elements and fibroblasts, producing fibrous structures. At the periphery of the tunneling zone the active proliferation of cellular elements and blood vessels is observed. This promotes the improvement of tissue nutrition and processes of reparative regeneration of bone tissue.
Национальная
академия наук
Беларуси

38
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 612.014.3:616-006.48:57.085.23+577.152.34:577.112.382.2
Т. в. БАлАшЕвич, в. н. ниКАнДРов
ОСОБЕННОСТИ СОСТОЯНИЯ КЛЕТОК ГЛИОМЫ С6 В КУЛЬТУРЕ ПРИ СОВМЕСТНОМ ВОЗДЕйСТВИИ ПЛАЗМИНОГЕНА И ГЛИЦИНА
институт физиологии нАн Беларуси, Минск
(поступила в редакцию 17.05.2010)
Введение. Исследования последних 20 лет доказали, что плазминоген (Pg) является нейротро-Pg) является нейротро-) является нейротро-фическим фактором, регулирующим ряд витальных функций клеток нервной ткани в центральной и периферической нервной системе [3–5, 9]. Установлено также, что этот белок синтезируется не-посредственно в структурах нервной системы микроглией [9] и отдельными субпопуляциями нейронов [12]. Однако особенности регуляторного действия Pg раскрыты далеко не полностью, учитывая гетерогенность клеток нервной ткани не только в морфологическом, функциональном, но и в метаболическом плане.
Ранее нами было показано, что добавление Pg к культуре глиомы С6 на бессывороточной среде через 24 ч обеспечивает сохранение жизнеспособности клеток, их пролиферацию, способствует уве-личению содержания РНК и белка в клетках, а через 72 ч – и повышению уровня ДНК [3–4, 6–7].
При изменении состояния клеток эффекты Pg имели существенные отличия. Так, в исходных энтерохромаффинных клетках феохромоцитомы PC12 после добавления зимогена в концентра-PC12 после добавления зимогена в концентра-12 после добавления зимогена в концентра-ции 10–9 М активность лактат- и NADH-дегидрогеназ практически не изменялась, тогда как при таком же воздействии на эти клетки, подвергнутые влиянию фактора роста нервов (при этом они трансформировались в нейроноподобные и приобретали ацетилхолинэстеразную активность), активность этих дегидрогеназ увеличивалась на 37 и 33% соответственно [5].
Судя по источникам мировой литературы, более обстоятельных исследований подобного плана ранее не проводилось.
Цель настоящей работы – раскрыть особенности совместного эффекта плазминогена и гли-цина на морфофункциональное состояние клеток глиомы С6 и содержание в них биополимеров.
Известно, что функция глицина в нервной ткани двояка. Эта аминокислота используется в качестве интермедиата для синтеза практически всех белков (включая нейроспецифические), нуклеозидов, ряда пептидов и других соединений. Кроме того, глицин выполняет функцию тор-мозного нейромедиатора.
Материалы и методы исследования. Культура крысиной глиомы С6 получена из Российской коллекции клеточных культур позвоночных (Институт цитологии РАН, Санкт-Петербург, Россия). Культивирование клеток глиомы проводили по отработанной методике [1] в синтетической пита-тельной среде, содержавшей 10% эмбриональной телячьей сыворотки и 25 мкг/мл гентамицина. Для исследования использовали питательную среду, содержавшую 0,5% сыворотки, чтобы исклю-чить влияние ростовых факторов и питательных веществ, содержащихся в сыворотке. Через 3 сут в среду вносили Pg в концентрации 10–7 М и глицин (Applichem, Германия) в концентрациях 0,01; 0,1; 1,0; 10,0; 25,0; 50,0 мМ. Очищенные образцы Pg получены из фракции β-глобулинов белков плазмы крови методом аффинной хроматографии на лизин-сефарозе (Sigma, США); подробно суть метода, характеристика чистоты и активности образцов приведены в работе [11].
По истечении 72 ч клетки глиомы С6 отделяли от кондиционированной среды путем центри-фугирования в течение 5 мин при 2000 об/мин. Содержание РНК, ДНК и белка оценивали с помо-щью двухволновой спектрофотометрии по О. Warburg (1942 г.), описанной Ю. Б. Филипповичем [8].
Национальная
академия наук
Беларуси

39
Определение содержания нуклеиновых кислот проводили в два этапа: путем щелочного и кислот-ного гидролиза [7]. Для обсчета результатов использовали приложение RNA/DNA к программно-RNA/DNA к программно-/DNA к программно-DNA к программно- к программно-му обеспечению Cary WinUV (2003 г.). Количество общего белка измеряли по абсорбции при 280 нм (с поправкой на 260 и 320 нм) на 0,1 М фосфатном буфере, pH 7,5 (Sigma). Содержание нуклеи-pH 7,5 (Sigma). Содержание нуклеи- 7,5 (Sigma). Содержание нуклеи-Sigma). Содержание нуклеи-). Содержание нуклеи-новых кислот и белка выражали в мкг/мл. Статистическую значимость полученных результатов оценивали при помощи теста непараметрического анализа (критерий Манна–Уитни) в программе Statistica 6.0. Результаты представлены как среднее арифметическое ± стандартное отклонение.
Прижизненную микроскопию клеток проводили в фазовом контрасте с помощью микроско-па Opton, Германия (×16).
Результаты и их обсуждение. Прежде всего следует отметить, что в наших экспериментах культура глиомы С6 заметно отличалась от использованной ранее [3–7]. Клетки культивировали на дефицитной по белкам сыворотки крови питательной среде не 24, а 72 ч. В этих условиях их пролиферативная активность снижалась. Заметно изменялась морфология: вместо ранее исполь-зуемых в культурах веретенообразных, с небольшой сомой и 2–3 вытянутыми отростками кле-ток образовывались полигональные клетки с плоскими псевдоподиями и распластанной сомой крупных размеров. Принято считать, что отдельные типы клеток таких культур продуцируют GFAP (глиальный кислый фибриллярный белок) – главный маркер астроцитов [10], а культиви- (глиальный кислый фибриллярный белок) – главный маркер астроцитов [10], а культиви-рование клеток культуры С6 в дефицитной по белкам сыворотки крови среде способствует их дифференцировке преимущественно в астроцитоподобные [13]. В используемых в настоящей ра-боте культурах глиомы С6 доля таких клеток составляла 50%.
При добавлении Pg в концентрации 10–7 М к таким клеткам доля погибших клеток снижалась лишь на 8% в сравнении с контролем. Это принципиально отличалось от действия зимогена на «не-зрелые» клетки глиомы С6 – доля погибших клеток снижалась практически в 6 раз [5]. Воздействие глицина в концентрации 0,01–50,0 мМ также не приводило к существенному снижению доли погиб-ших клеток. При воздействии каждого из факторов в культуре наблюдались доминирование гли-альных клеток округлой формы, потеря адгезивной способности, что выражалось в формировании конгломератов (рис. 1). Это позволяет предположить, что метаболические резервы клеток направле-
Рис. 1. Фазово-контрастная микроскопия клеток культуры глиомы С6 через 72 ч после добавления плазминогена и глицина: а – контроль (0,5% сыворотки в питательной среде); б – 10 мкг/мл плазминогена; в – 0,01 мМ глицина; г – 0,01 мМ
глицина + 10 мкг/мл плазминогена; д – 0,1 мМ глицина; е – 0,1 мМ глицина + 10 мкг/мл плазминогена (×16)
Национальная
академия наук
Беларуси

40
ны скорее на обеспечение выживаемости в неблагоприятных условиях (общее время роста на дефи-цитной по белкам сыворотки крови среде – 6 сут!), чем на пролиферацию и дифференцировку.
Если же к клеткам культуры, выращенной на питательной среде, одновременно добавляли Pg и глицин (0,01 или 0,1 мМ), то доля погибших клеток не превышала 4% при четком проявлении пролиферации. Из этого следует, что Pg способен усиливать действие глицина в метаболических процессах клеток глиомы С6.
В использованных нами культурах глиомы С6 («дифференцирующихся») добавление зимо-гена не вызывало существенных изменений уровней общего белка и нуклеиновых кислот в клет-ках (рис. 2), в то же время в «незрелых» клетках их уровень возрастал в 2,3–4 раза [5].
Воздействие глицина в диапазоне концентраций от 0,01 до 25,0 мМ обусловило повышение уровня общего белка на 37–69% (рис. 2). Принято считать, что для реализации процесса биосин-теза белка необходимо присутствие одновременно всех 20 аминокислот [2]. Дополнительное вве-дение в питательную среду лишь одной из них, видимо, не должно вызывать интенсификации биосинтеза. Можно предположить два пути реализации наблюдаемых изменений: либо добавле-ние глицина вызывает усиление действия других аминокислот, присутствующих в базовой пи-тательной среде, либо оно сопровождается подавлением реакций внутриклеточного протеолиза. Действительно, ранее было показано, что добавление этой аминокислоты к данной культуре со-провождается угнетением на 17–41% 2-кальпаиновой активности [1]. Отмечаемое же нами отсут-ствие существенных изменений уровней РНК или ДНК позволяет предположить, что наиболее вероятным является именно второй вариант.
Совместное действие зимогена и аминокислоты (в концентрациях 0,01; 0,1; 10,0 и 50,0 мМ) сопровождалось снижением уровня внутриклеточного белка на 14–19% по сравнению с кон-тролем, тогда как после добавления Pg совместно с глицином (в концентрациях 0,01; 0,1; 1,0 и 25,0 мМ) наблюдалось повышение уровня РНК на 38–78%. Комбинированное влияние Pg и глицина (в концентрациях 0,01 и 0,1 мМ) вызывало также повышение уровня ДНК в клет-ках – на 38–46%.
Рис. 2. Изменение содержания общего белка (а), РНК (б) и ДНК (в) в клетках глиомы С6 через 72 ч после добавления в культуру плазминогена и глицина: контроль (К); добавление глицина (А), плазминогена (Б) и плазминогена + гли-цина (В). Концентрация глицина, мМ: 1 – 0,01; 2 – 0,1; 3 – 1,0; 4 – 10,0; 5 – 25,0; 6 – 50,0. ∗ – различия достоверны по
отношению к контролю при P ≤ 0,05 (n = 5)
Национальная
академия наук
Беларуси

Заключение. Полученные нами данные свидетельствуют о том, что так называемые «диф-ференцирующиеся» культуры глиомы С6 в сравнении с культурами «незрелых» клеток отлича-ются довольно слабым изменением уровней белка и нуклеиновых кислот в ответ на воздействие Pg. Это еще раз подтверждает факт зависимости эффекта плазминогена (как и других субстан-. Это еще раз подтверждает факт зависимости эффекта плазминогена (как и других субстан-ций) от состояния клеток нервной ткани.
Кроме того, представленные результаты позволили заключить, что эффект Pg существенно изменяется в присутствии глицина, принципиально отличаясь от эффекта каждого из исследуе-мых соединений. Причем в зависимости от концентрации изменения носили сложный характер: ни в одном случае сдвиги не имели противоположной направленности, но проявлялись при дей-ствии не всех концентраций аминокислоты. Это диктует необходимость проведения дальней-ших исследований в данном направлении в целях выяснения механизмов выявленных сдвигов, а также для расширения арсенала метаболических эффекторов нейротрофического действия плазминогена.
Литература
1. Б а л а ш е в и ч Т. В., Н и к а н д р о в В. Н., Г р о н с к а я Р. И., Т у м и л о в и ч М. К. // Журн. Гродн. гос. мед. ун-та. 2009. № 2. С. 61–63.
2. Б е р е з о в Т. Т., К о р о в к и н Б. Ф. Биологическая химия. М., 2002.3. Н и к а н д р о в В. Н., Ж у к О. Н., Г р о н с к а я Р. И. и др. // Materials, methods and technology. Scientific
articles. 2007. P. 48–66.4. Н и к а н д р о в В. Н., Ж у к О. Н., Г р о н с к а я Р. И. и др. // Биомед. химия. 2008. Т. 54, № 2. С. 192–200.5. Н и к а н д р о в В. Н., Ж у к О. Н., П ы ж о в а Н. С. и др. // Весці НАН Беларусі. Сер. мед. навук. 2008.
№ 1. С. 85–97.6. Р о м а н о в с к а я A. A., Ж у к O. Н., Н и к а н д р о в В. Н. // Наука и инновации. 2007. № 3. C. 24–27. 7. Р о м а н о в с к а я А. А. // Весці НАН Беларусі. Сер. мед. навук. 2005. № 5. C. 38–41.8. Ф и л и п п о в и ч Ю. Б., Е г о р о в а Т. А., С е в а с т ь я н о в а Г. А. Практикум по общей биохимии. М.,
1975. С. 182–184.9. K o h s a k a S., N a k a j i m a K., H a m a n o u e M. et al. // Neuropathol. Appl. Neurobiol. 1994. Vol. 20, N 2. P. 190. 10. N a g a m a t s u S., N a k a m i c h i �., I n o u e M. et al. // Biochem. J. 1996. N 319. P. 477–482.11. N i k a n d r o v V. N., M u r a s h k o O. N., V o r o b y o v a G. V. et al. // �ett. Pept. Sci. 1997. Vol. 4. P. 497–502.12. T s i r k a S. E., R o g o v e A. D., B u g g e T. H. et al. // J. Neurosci. 1997. Vol. 17, N 2. P. 543–552. 13. W e i w e i H u, O n u m a T., B i r u k a w a N. et al. // Cell. Mol. Neurobiol. 2008. Vol. 28. P. 519–528.
T. v. BalaShEvich, v. n. niKandRov
PECULIARITIES OF C6 GLIOMA CELLS STATE IN CULTURE UNDER THE JOINT EFFECT OF PLASMINOGEN AND GLYCINE
institute of physiology of naS of Belarus, Minsk
Summary
The plasminogen influence on cellular DNA, RNA, whole protein level and morpho-functional state of the so-called “dif-ferentiated” C6 glioma culture cells (6 days exposition in the medium with the deficiency of serum proteins) against underground of glycine additions has been investigated. The “differentiated” C6 glioma cells are characterized by quite weak changes of the protein and nucleic acids level in response to plasminogen addition as against “immature” cells. In the presence of glycine, the zymogene effect was modified essentially – observed picture fundamentally differed from the effects of each of the investigated compounds. The joint effect of plasminogen (10–7 М) and glycine (0.01 or 0.1 mМ) during 72 h led to nucleic acids and whole protein level increasing and 96%-tage vital activity of C6 glioma cells in the medium with serum deprivation.
Национальная
академия наук
Беларуси

42
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 615.227.3:616-092.9
в. в. СЕРЕДА1, С. Г. оДинЦов1, А. п. влАСов1, Д. в. ФиМА1, Д. в. шУБЕноК1, З. и. КРАвчУК1, Г. в. КоЖУХ1, в. в. шМАнАЙ2, С. М. ДЕЕв3, С. п. МАРЦЕв1
ПОЛУЧЕНИЕ МОДИФИЦИРОВАННОГО ПОЛИЭТИЛЕНГЛИКОЛЕМ ИММУНОТОКСИНА scFv4D5-БАРНАЗА, СПЕЦИФИЧНОГО К ОНКОГЕНУ с-ErbB2, И
СРАВНИТЕЛЬНОЕ ИССЛЕДОВАНИЕ ЕГО СТРУКТУРНО-ФУНКЦИОНАЛЬНЫХ ПАРАМЕТРОВ
1Республиканский научно-практический центр гематологии и трансфузиологии, Минск, Беларусь, 2оДо «праймтех», Минск, Беларусь,
2институт биоорганической химии им. М. М. шемякина и Ю. А. овчинникова РАн, Москва
(поступила в редакцию 02.06.2010)
Введение. В современной клинической практике для направленной терапии онкологических заболеваний активно применяются моноклональные антитела и их производные – гуманизиро-ванные рекомбинантные антитела, антипролиферативное действие которых связано с активаци-ей клеточной и/или комплемент-зависимой цитотоксичности. Прогнозируется, что следующий инновационный этап развития иммунотерапии будет связан с химерными иммунотоксинами. Данные белки обладают собственной цитотоксичностью за счет присутствия в их структуре рекомбинантных фрагментов антител и цитотоксических белков. Среди иммунотоксинов, кон-струируемых для элиминации клеток рака молочной железы, наиболее перспективными явля-ются конструкции, содержащие фрагменты терапевтического гуманизированного антитела 4D5 (герцептина), специфичного к онкогену-маркеру с-ErbB-2 (HER-2/neu, p185HER2). Этот маркер гиперэкспрессирован на клеточной мембране значительной группы опухолей (рак молочной же-лезы, карцинома яичника, легких и др.), что коррелирует с агрессивностью течения заболевания, резистентностью опухоли к химиотерапии и плохим прогнозом [1, 2].
Клинические исследования показали, что в лечении HER-2-позитивного рака молочной железы I–III стадии сочетанная терапия (химиотерапия + герцептин) намного эффективнее монохимио-терапии. Повышение уровня терапевтического ответа при проведении сочетанной терапии сни-жает риск рецидива рака и, по данным предварительных исследований, улучшает показатель выживаемости у таких пациенток [3].
Однако, несмотря на явные успехи в применении антител и их производных, выявлен ряд не-гативных моментов при проведении иммунотерапии. К ним относится неспецифический (гидро-фобный либо связанный с клеточными Fc-рецепторами) захват иммунотерапевтической кон-Fc-рецепторами) захват иммунотерапевтической кон--рецепторами) захват иммунотерапевтической кон-струкции нетрансформированными клетками, а также ограниченный специфический захват, что обусловлено фоновым присутствием большинства онкомаркеров на поверхности нормальных клеток. Оба этих механизма ведут к неспецифической токсичности иммунотерапевтического агента. Еще одним ограничением, присущим в основном укороченным фрагментам антител и иммунотоксинам на их основе, является быстрый клиренс молекул из кровотока за счет клу-бочковой фильтрации и протеолиза в крови, что снижает эффективность иммунотерапии и при-водит к увеличению инъекционной дозы.
Перечисленные ограничения могут быть минимизированы с помощью развиваемой в послед-ние годы технологии, в основе которой лежит химическая модификация структуры иммунотокси-нов и других терапевтических белков полиэтиленгликолем (ПЭГ) с молекулярной массой 3–40 кДа, имеющим линейную или разветвленную структуру. ПЭГилирование лекарственных препаратов
Национальная
академия наук
Беларуси

43
белковой природы дает ряд весомых преимуществ, которые недоступны при использовании на-тивных аналогов: удлинение периода «эффективной» полужизни, замедление скорости выведе-ния, отсутствие пиков плазменной/тканевой концентрации, понижение токсичности и иммуно-генности, за счет чего усиливается их биологическая активность [4, 5].
Данная работа продолжает цикл исследований химерного иммунотоксина 4D5scFv-барназа, включающего фрагмент терапевтического антитела 4D5 и бактериальную рибонуклеазу (барна-зу). Ранее нами были определены основные параметры конформации и стабильности, связыва-ние с культурой опухолевых клеток с различным уровнем экспрессии HER-2 и цитотоксическое действие на эти клетки [6], подтверждающие, что данный иммунотоксин можно рассматривать как потенциально перспективный терапевтический агент.
Цель настоящего исследования – получение ПЭГилированной формы иммунотоксина scFv4D5-барназа и первичный анализ его конформации, стабильности и активности в сравнении с исходным иммунотоксином.
Материалы и методы исследования. пэГилирование рекомбинантного иммунотоксина scFv4D5-барназа. Рекомбинантный иммунотоксин получали в системе гетерологической экс-прессии с использованием клеток Escherichia coli, штамм SB 536, и плазмиды pET23d(scFv4D5-барназа), несущей последовательность, кодирующую scFv4D5-барназу и барстар (природный ци-топлазматический ингибитор барназы) [7, 8]. Очистку белка до гомогенного состояния проводи-ли методом аффинной хроматографии на колонке, заполненной Ni-NTA-сефарозой, по ранее разработанной схеме [6].
ПЭГилирование иммунотоксина осуществляли в 50 мМ натрий-боратном буфере, рН 8,5, N-оксисукцинимидным эфиром ПЭГ-5000, синтезированным ОДО «Праймтех» (Минск, Бела--оксисукцинимидным эфиром ПЭГ-5000, синтезированным ОДО «Праймтех» (Минск, Бела-русь). Модификатор растворяли в 100%-ном диметилсульфоксиде при комнатной температуре и добавляли к раствору белка порциями (каждые 30 мин) в течение не менее 6 ч, постоянно пере-мешивая реакционную смесь. Молярное соотношение иммунотоксин:ПЭГ при добавлении каж-дой порции составляло 1:2–1:4. Непрореагировавший модификатор отделяли путем диализа. Ко-личество молекул ПЭГ, присоединенных к молекуле белка, оценивали методом электрофореза в полиакриламидном геле (ПААГ), определяя молекулярную массу продуктов реакции.
Флуоресцентная спектроскопия. Спектры собственной флуоресценции регистрировали при 25 °С в термостатируемой кювете с длиной оптического пути 1 см при помощи спектрофлуориме-тра SF�-1211 («Солар», Беларусь), оборудованного персональным компьютером. Диапазон длин волн регистрации составлял 290–390 и 305–405 нм (при возбуждении соответственно на 280 и 295 нм). Спектры флуоресценции буферных растворов вычитались из спектров флуоресценции образцов.
Флуоресценцию гидрофобного зонда 1-анилин-8-нафталинсульфоната (АНС) возбуждали при длине волны 360 нм и регистрировали в диапазоне от 400 до 600 нм. Для получения спек-тров АНС, связанного с белком, из кривых вычитали спектры свободного АНС в соответствую-щих буферах. В экспериментах использовали 30-кратный молярный избыток АНС над белком (из расчета 10 молекул АНС на 1 структурный домен белка).
дифференциальная сканирующая калориметрия (дсК). Измерения были выполнены с ис-пользованием дифференциального адиабатного сканирующего микрокалориметра ДАСМ-4 («Биоприбор», Пущино, Россия) в температурном диапазоне от 10 до 100 °С при скорости скани-рования 1 К/мин. Для каждого образца выполняли 3–8 сканирований, при этом погрешность измерения ∆h составляла 5–6%. Энтальпию тепловой денатурации и теплоемкость рассчитыва-ли с использованием программм TERMCA�C и SCA� («Биоприбор», Россия) и DSCfit (Институт молекулярной биофизики, США) [9].
анализ функциональной активности рекомбинантного иммунотоксина scFv4D5-барназа. Для оценки связывания иммунотоксина с клеточными линиями был использован ранее разрабо-танный иммуноферментный анализ с иммобилизованными в лунках полистирольных планше-тов клетками с высоким (BT-474) либо низким (MCF-7) уровнем экспрессии маркера с-ErbB2. В качестве проявляющего реагента использован тандем поликлональных антител кролика к бар-назе и моноклональные антитела овцы к антителам кролика, конъюгированные с пероксидазой хрена [6].
Национальная
академия наук
Беларуси

44
другие методы. Чистоту белков оценивали методом электрофореза в ПААГ в присутствии додецилсульфата натрия, используя 5%-ный формирующий и 12%-ный разделительные гели с бу-ферной системой по способу, описанному в работе [10]. Все образцы были восстановлены 2%-ным β-меркаптоэтанолом и прогреты в течение 5 мин на кипящей водяной бане. Гели окрашивали бриллиантовым голубым R-250 (по Кумасси).
Концентрацию иммунотоксина определяли спектрофотометрически, используя удельный коэффициент экстинкции %1,0
280E = 1,86 л/(г·см). Коэффициент рассчитывали на основе аминокис-лотной последовательности по алгоритму, приведенному в работе [11]. Молекулярная масса бел-ка, согласно аминокислотной последовательности, составляла 42 120 Да.
Результаты и их обсуждение. разработка методики пэГилирования иммунотоксина scFv4D5-барназа. Плазмида pET23d(scFv4D5-барназа) для биосинтеза рекомбинантного иммуно-ET23d(scFv4D5-барназа) для биосинтеза рекомбинантного иммуно-23d(scFv4D5-барназа) для биосинтеза рекомбинантного иммуно-d(scFv4D5-барназа) для биосинтеза рекомбинантного иммуно-(scFv4D5-барназа) для биосинтеза рекомбинантного иммуно-токсина была сконструирована в лаборатории молекулярной иммунологии Института биооргани-ческой химии им. М. М. Шемякина и Ю. А. Овчинникова РАН [12]. В состав плазмиды входила последовательность, кодирующая scFv фрагмент родительского антитела 4D5, шарнирный уча-сток (H), барназу, спейсер (S) и пентагистидиновый тэг (рис. 1, А). Очистка иммунотоксина, про-веденная по ранее разработанному протоколу, позволила получить белок с чистотой не менее 95% по данным электрофореза в ПААГ (рис. 1, Б). Выход целевого белка составлял 4–6 мг/л культу-ры.
Для ПЭГилирования биомолекул ранее показано применение различных активированных производных ПЭГ: малеимидных, сукцинимидных, винилсульфоновых, альдегидных и др. Все они отличаются по механизмам, эффективности присоединения к молекуле белка, условиям ре-акции и устойчивости получаемого продукта. Кроме того, не менее важным является количество молекул ПЭГ, присоединенных к одной молекуле белка. Известно, что модификация большин-ства аминогрупп может приводить к потере антителом или его фрагментами своих функций. Поэтому основной задачей на данном этапе был выбор оптимального подхода для эффективного ПЭГилирования рекомбинантного иммунотоксина scFv4D5-барназа.
В качестве модификатора был выбран N-оксисукцинимидный эфир ПЭГ-5000, способный вступать в реакцию со свободными аминогруппами белка. Реакция взаимодействия сукцини-мидного эфира ПЭГ с аминогруппами происходит намного быстрее, чем других эфиров (к при-меру, малеимидного), однако стабильность сукцинимидного эфира в водном растворе значительно меньше (время полужизни 10–15 мин).
При разработке методики ПЭГилирования был проведен подбор реакционной буферной си-стемы, а также определено оптимальное молярное соотношение модификатор:белок и режим
Рис. 1. Структура плазмиды pET23d(scFv4D5-барназа) (А): lac – lac-промотор; scFv4D5, барназа, барстар – последователь-ности ДНК, кодирующие scFv фрагмент антитела 4D5, барназу и барстар соответственно; Н – шарнирный участок; S – спейсер; (His)5 – пентагистидиновый тэг; AmpR – ген устойчивости к ампициллину; Р – конститутивный промотор; элек-трофоретический анализ иммунотоксина в процессе ПЭГилирования (Б): 1–5 – реакционная смесь через 1–5 ч реакции ПЭГилирования; 6 – белковые маркеры молекулярных масс (116, 66, 45, 35, 25 кДа); 7 – белок перед ПЭГилированием
Национальная
академия наук
Беларуси

45
(кратность) добавления модификатора. Серия скрининговых экспериментов с определением методом гель-электрофореза количества остатков ПЭГ, присоединенных к молекуле белка, по-казала, что наиболее эффективным является соотношение иммунотоксин:эфир ПЭГ, равное 1:2–1:4 при однократном добавлении. Для достижения максимального выхода реакции целесообразно добавлять модификатор многократно в течение нескольких часов, что связано с относительно низкой частотой продуктивных столкновений полимерного модификатора с аминогруппами белка при ограниченном времени жизни модификатора в водном растворе. При однократном до-бавлении большого молярного избытка ПЭГ-сукцинимида (соотношение белок:ПЭГ–1:50–1:100) реакция протекала очень быстро, что приводило к получению молекул иммунотоксина, связан-ных с пятью и более молекулами ПЭГ.
Применение оптимальной на данный момент методики ПЭГилирования, описанной выше, позволило увеличить общий выход иммуноконъюгата scFv-барназа–ПЭГ-5000 до 90% от количе-ства белка, взятого в реакцию. Около 70–80% продуктов реакции при этом составляли молеку-лы иммунотоксина, конъюгированные с 1–2 молекулами ПЭГ (рис. 1, Б).
сравнительный структурный анализ исходного и пэГилированного иммунотоксина scFv4D5-барназа. Рекомбинантные иммунотоксины представляют собой группу молекул, дан-ные о взаимосвязи структуры, конформации и стабильности которых крайне ограничены, что сдерживает конструирование новых иммунотоксинов и адаптацию структуры к условиям кли-нического применения. В данной работе методами спектроскопии и сканирующей калориметрии выполнен первичный конформационный анализ химерного белка scFv4D5-барназа, специфично-го к онкогену с-ErbB2, а также проведено сравнение исходного белка и модифицированного по-лиэтиленгликолем аналога.
Флуоресцентная спектроскопия. Сравнение спектров собственной флуоресценции исходно-го и ПЭГилированного иммунотоксина scFv4D5-барназа свидетельствует об их практически полной идентичности. В обоих случаях спектры имеют максимум эмиссии при длине волны 335–337 нм, что характерно для белков, обладающих сформированной третичной структурой, при которой основные флуорофоры – остатки тирозина и триптофана – вовлечены в гидрофоб-ное ядро и защищены от тушения растворителем (рис. 2, А). Аналогичная картина наблюдается и при сравнении спектров, полученных при возбуждении остатков триптофана полипептидной цепи (длина волны возбуждения 295 нм): спектры белка до и после ПЭГилирования имеют ми-нимальные отличия и сниженную интенсивность флуоресценции.
Спектры собственной флуоресценции как нативного, так и ПЭГилированного иммунотокси-на при денатурации в 8 М мочевине демонстрируют трехкратное снижение интенсивности флуо-ресценции и смещение максимума эмиссии до 350 нм вследствие экспонирования флуорофоров растворителю (рис. 2, А). Следовательно, исходная конформация обоих белков до добавления денатуранта обеспечивает гидрофобное окружение ароматических аминокислотных остатков, что характерно для сформированной третичной структуры белков [13].
Рис. 2. Сравнительный структурно-функциональный анализ ПЭГилированного иммунотоксина scFv4D5-барназа: А – спектры собственной флуоресценции; Б – кривые термоденатурации иммунотоксина по данным ДСК (сплошная линия – исходный иммунотоксин, штрихованная линия – ПЭГилированный белок, пунктир – денатурированное со-стояние); в – кривые связывания scFv-барназы с клетками рака молочной железы линий BT-474 и MCF-7 (темные
и светлые кружки – белок до и после модификации соответственно; на вставке – схема теста)
Национальная
академия наук
Беларуси

46
Дополнительное исследование третичной структуры рекомбинантного иммунотоксина было проведено с использованием зонда АНС. За счет своей гидрофобной природы АНС способен свя-зываться с белками, находящимися в так называемых промежуточных структурных состояниях, при которых часть гидрофобных кластеров экспонируется и становится доступна зонду. При этом АНС демонстрирует хорошо регистрируемый «голубой сдвиг» в спектре эмиссии (с 520 нм у свободного АНС до 470 нм) за счет «погружения» в гидрофобные участки флуктуирующей белковой глобулы и экранирования от водного окружения. В противоположность промежуточ-ным (частично развернутым) конформациям белка, полностью сформированные третичные структуры не позволяют пространственно объемной молекуле АНС проникать в гидрофобное ядро. Полностью развернутые белки даже в случае связывания зонда с экспонированными ги-дрофобными участками не способны обеспечить регистрируемый коротковолновой сдвиг флуо-ресцентной эмиссии. Эта совокупность свойств позволяет использовать АНС как специфический маркер частично развернутых конформаций.
По результатам эксперимента, рекомбинантный иммунотоксин scFv4D5-барназа не связыва-ет АНС ни до, ни после ПЭГилирования (данные не приведены), что наряду с данными по соб-ственной флуоресценции подтверждает сохранение компактной третичной структуры белка.
Дифференциальная сканирующая калориметрия. Сравнительный анализ кривых тепловой дена-турации иммунотоксина до и после ПЭГилирования показал наличие в обоих случаях выраженных пиков теплопоглощения, что свидетельствует о кооперативности тепловых переходов вследствие присутствия в компактной третичной структуре стабилизирующих специфических взаимодействий. Температурный ход кривой теплоемкости исходной и ПЭГилированной scFv4D5-барназы (рис. 2, Б) имеет бимодальный характер с выраженными максимумами теплопоглощения при 51–53 и 67–70 °С, что описывает, соответственно, термоденатурацию барназы (цитотоксический модуль химерной конструкции) и scFv фрагмента антитела 4D5 (узнающий фрагмент). Повторные прогревы образцов не давали пиков теплопоглощения. Это указывает на то, что тепловые переходы являются необрати-мыми вследствие агрегации белка после температурного разворачивания; последнее показано при визуальном контроле белка, извлеченного из калориметрической ячейки после первого нагрева.
Интегральные калориметрические энтальпии (∆h) термоденатурации исходного и модифициро-ванного белка отличались незначительно и были близки к величине 4,7 кал/г, а удельная теплоем-кость (cp) составляла 63 кДж/(К·моль) при 25 °С. Энтальпии низко- и высокотемпературного перехо-дов составляли соответственно 9,5 и 2,4 кал/г, что отражает известную высокую структурную энер-гетику изолированной барназы (∆h ∼ 10 кал/г). В то же время энтальпия термоденатурации scFv фрагмента заметно уступает величинам 4,0–4,5 кал/г, полученным для изолированных доменов ан-тител [14], или 5,5–7,5 кал/г, полученным для природных антител в условиях взаимодействующих доменов [13], что заставляет предполагать дестабилизирующие взаимодействия V� и VH доменов в составе scFv фрагмента 4D5. Полученные нами энтальпии тепловых переходов для барназного и узнающего модуля иммунотоксина принадлежат диапазону, известному для глобулярных белков с полностью компактной третичной структурой, однако лежат в пределах крайних его значений [15].
Неизменная структурная энергетика ПЭГилированного иммунотоксина доказывает, что ПЭГилирование не вносит существенных нарушений в пространственное строение белка. При этом присоединенные полиэтиленгликольные цепочки располагаются поверхностно и не интер-калируют в структуру белка, поскольку интеркаляция привела бы к изменению степени доступ-ности гидрофобных участков белка до и после теплового разворачивания молекулы, что значи-тельно изменяет величины тепловых эффектов калориметрии (cp, ∆cp, ∆h). Отсутствие конфор-мационных дефектов ПЭГилированного иммунотоксина свидетельствует о возможности его дальнейшего исследования как потенциального терапевтического агента.
анализ антигенсвязывающей активности иммунотоксина. В качестве критерия оценки функциональных свойств белка нами использован ранее разработанный метод иммуноферментно-го определения связывания иммунотоксина с с-ErbB2-позитивными клетками на основе анти-тел кролика к барназе. Данный протокол позволяет получить высокий уровень сигнала для клеток с гиперэкспрессией с-ErbB2 (BT-474) и хорошую воспроизводимость анализа, при этом уровень сигнала для с-ErbB2-негативной линии клеток (MCF-7) сопоставим со значением фона (рис. 2, в).
Национальная
академия наук
Беларуси

С использованием разработанной методики была проанализирована эффективность связыва-ния препарата исходного и ПЭГилированного иммунотоксина с линиями клеток рака молочной железы с низким (MCF-7) и высоким (BT-474) уровнями экспрессии рецептора с-ErbB2. Разница в значениях сигнала для с-ErbB2-позитивных и негативный линий, составляющая в среднем 0,5–0,6 отн. ед. (рис. 2, в), свидетельствует о специфическом характере связывания сравнивае-мых образцов иммунотоксина, что является одним из необходимых условий для возможности использования белка в дальнейших экспериментах in vivo. Константа связывания (Ка) исходного и ПЭГилированного иммунотоксина с с-ErbB2 рецептором клеток BT-474 имеет порядок 108 М–1, что близко к значению Ка «родительского» моноклонального антитела 4D5 (герцептина) [16].
Заключение. В результате проведенного исследования разработана и оптимизирована методика получения ПЭГилированного аналога рекомбинантного иммунотоксина scFv4D5-барназа, специфич-ного к онкогену с-ErbB2, и получен конъюгат scFv-барназа–ПЭГ-5000 с выходом реакции около 90%.
Методами флуоресцентной спектроскопии и сканирующей калориметрии показано, что ПЭГилированному иммунотоксину свойственна сформированная компактная третичная струк-тура. По своим функциональным параметрам, в частности по специфичности связывания с клет-ками рака молочной железы in vitro, ПЭГилированный химерный белок scFv4D5-барназа удо-влетворяет требованиям, предъявляемым к иммунотоксинам. В свою очередь это свидетельству-ет о перспективности его дальнейшего изучения как нового потенциального агента для иммунотерапии рака. Следующие этапы работы будут включать проверку способности ПЭГи-лированного иммунотоксина элиминировать c-ErbB2-позитивные клетки рака молочной железы in vitro, а также определение его фармакологических и токсикологических параметров in vivo.
Исследование выполнено в рамках договора с Министерством здравоохранения Республики Беларусь № 8/09-ФИ от 31.03.2009 г.
Литература
1. N a h t a R. et al. // Curr. Pharm. Person. Med. 2009. Vol. 7, N 4. P. 263–274.2. M e t r o G., M o t t o l e s e M., F a b i A. // Expert Opin. Pharmacother. 2008. Vol. 9, N 15. P. 2583–2601.3. М о и с е е н к о В. М. // Практ. онкол. 2002. Т. 3, N 4. С. 253–261.4. M u g g i a F. // From Research to Practiсe. 2001. Vol. 3, N 1. P. 1–3.5. B r u c e A. // From Research to Practice. 2001. Vol. 3, N 1. P. 3–9.6. В л а с о в А. П. и др. // Весцi НАН Беларусi. Сер. мед. навук. 2009. № 1. С. 5–13.7. H a r t l e y R. W. // Meth. Enzymol. 2001. Vol. 341. P. 599–611.8. D e y e v S. M. et al. // Mol. Gen. Genet. 1998. Vol. 259, N 4. P. 379–382.9. G r e k S. B., D a v i s J. K., B l a b e r M. // Prot. and Pept. �ett. 2001. Vol. 6. P. 429–436.10. � a e m m l i U. K. // Nature. 1970. Vol. 227. P. 680–685.11. G i l l S. C., v o n H i p p e l P. H. // Analyt. Biochem. 1989. Vol. 182. P. 319–326.12. D e y e v S. M. et al. // Nature Biotechnol. 2003. Vol. 21. P. 1486–1492.13. M a r t s e v S. P. et al. // Biochemistry. 2000. Vol. 39. P. 8047–8057.14. M a r t s e v S. P. et al. // Biochemistry. 2002. Vol. 41. P. 3389–3395.15. P e r e z J. M. et al. // Biochemistry. 2001. Vol. 40. P. 74–83.16. B a s e l g a J. et al. // Cancer Res. 1998. Vol. 58. P. 2825–2831.
v. v. SEREda1, S. G. оdinTSov1, А. p. vlaSov1, d. v. FiMa1, d. v. ShuBEnoK1, Z. i. KRavchuK1, G. v. KoZhuKh1, v. v. ShManai2, S. М. dEyEv3, S. p. МАRTSEv1
PEGYLATION OF RECOMBINANT IMMUNOTOXIN scFv4D5-BARNASE DIRECTED AGAINST С-ERBB2 ONCOGENE AND ITS COMPARATIVE STRUCTURAL AND FUNCTIONAL STUDY
1Research and production center for hematology and Transfusiology, Minsk, Belarus,2primetech llc, Minsk, Belarus,
3Shemyakin-ovchinnikov institute of Bioorganic chemistry of RaS, Moscow
Summary
This work provides description for modification of the с-ErbB2-targeted recombinant immunotoxin scFv4D5-barnase with PEG-5000 N-hydroxysuccinimide ether. Comparative structural and functional studies of the native and PEGylated im-munotoxin species demonstrated that PEGylation did not result in any significant changes of tertiary structure of the protein, nor did it change the antigen-binding capacity.
Национальная
академия наук
Беларуси

48
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.314-74
н. в. новАК
СОСТОЯНИЕ ГРАНИЦЫ «ЗУБ – ПЛОМБА» ПРИ ИСПОЛЬЗОВАНИИ РАЗЛИЧНЫХ РЕСТАВРАЦИОННЫХ МАТЕРИАЛОВ
Белорусская медицинская академия последипломного образования, Минск
(поступила в редакцию 04.11.2009)
Введение. При восстановлении зубов с дефектами различной этиологии применяют компо-зиционные материалы, которые характеризуются высокой прочностью и обладают эстетически-ми свойствами. Это позволяет использовать их для выполнения реставраций как на жеватель-ных, так и на фронтальных зубах [1].
Эффективность пломбирования зубов во многом зависит от качества краевого прилегания пломбировочного материала к эмали и дентину и от состояния границы «зуб – пломба» [2, 3].
Целью настоящего исследования явилось изучение состояния границы зуба и композицион-ного материала белорусского производства на свежеудаленных зубах.
Материалы и методы исследования. На базе 8-й стоматологической поликлиники г. Мин-ска были проведены исследования четырех образцов нового композиционного материала бело-русского производства на 42 удаленных по различным показаниям интактных зубах. Сразу по-сле экстракции зубы обрабатывали щеткой под проточной водой с целью удаления оставшихся мягких тканей и зубного налета. При необходимости использовали пневматический скелер. За-тем образцы помещали в 0,1%-ный раствор тимола. В ближайшие часы после удаления на ок-клюзионной поверхности жевательных зубов и вестибулярной поверхности резцов формировали полости овальной формы размером 3×4 мм в пределах средних слоев дентина. Обработку поло-стей проводили в соответствии с классическими принципами [1, 2]. Для препарирования исполь-зовали турбинный наконечник и алмазные боры шаровидной формы средней степени зернисто-сти. Края полостей финировали с помощью мелкозернистых алмазных боров конической формы. На всех этапах работы использовали охлаждение водой. Излишки влаги устраняли воздействи-ем слабой струи воздуха в течение 4–5 с, затем осуществляли кислотное травление. После при-менения универсальной адгезивной системы (Single Bond 2 3M ESPE) осуществляли пломбиро-M ESPE) осуществляли пломбиро- ESPE) осуществляли пломбиро-ESPE) осуществляли пломбиро-) осуществляли пломбиро-вание полостей исследуемыми образцами фотополимеров. Поверхность пломб обрабатывали и полировали до естественного блеска. В зависимости от использованного материала образцы были разделены на 5 групп (образцы № 1–4 из композиционного материала белорусского произ-водства и образец № 5 из комозита Filtek Z250 (3М), США).
Состояние поверхности пломб и их краевое прилегание оценивали полуколичественным ме-тодом по критериям, сформулированным на основании данных литературы и собственного кли-нического опыта [4, 5]. Для этого проводили визуальную оценку поверхности пломбы и границы «зуб – пломба», их зондирование и окрашивание, определение глубины проникновения красите-ля в область границы «зуб – пломба». О состоянии поверхности пломбировочного материала судили по наличию или отсутствию объемных дефектов разного размера (крупных раковин и мелких пор, сколов, трещин).
С целью унификации данных каждый из показателей оценивали по шкале от 1 до 3 баллов (1 балл являлся «плохим» показателем, 2 балла – «средним» и 3 балла – «хорошим»).
Визуальную оценку образцов осуществляли невооруженным глазом в условиях стоматоло-гического кабинета при искусственном освещении.
Национальная
академия наук
Беларуси

49
Для более детального осмотра использовали лупу с 4-кратным увеличением.
Параметры визуальной оценки
Балл Граница «зуб – пломба» Поверхность пломбы
3 Нет зазора Поверхность ровная, без пор и раковин2 Зазор в области менее 1/3 длины границы Отдельные мелкие дефекты1 Зазор в области 1/3 – 2/3 длины границы Отдельные крупные объемные дефекты и мелкие поры
Зондирование границы «зуб – пломба» и поверхности пломбы проводили с помощью стома-тологического зонда с острым концом.
Параметры зондирования
Балл Граница «зуб – пломба» Поверхность пломбы
3 Шероховатость отсутствует Поверхность пломбы ровная2 Шероховатость в области до 1/3 длины границы Отдельные мелкие объемные дефекты1 Шероховатость в области 1/3 – 2/3 длины границы Отдельные крупные объемные дефекты и мелкие поры
После высушивания образцов струей обезжиренного воздуха на пломбу и прилегающие к ней участки эмали с помощью ватного шарика наносили 2%-ный водный раствор метиленового си-него на 30 с. Затем краситель смывали водно-воздушной струей в течение приблизительно 15 с, после чего образцы полировали гибкими дисками и силиконовыми инструментами фирмы Shofu (Япония).
Параметры окрашивания
Балл Граница «зуб – пломба» Поверхность пломбы
3 Нет окрашивания Нет окрашивания2 Окрашивание до 1/3 длины границы Интенсивное окрашивание отдельных мелких участков1 Окрашивание от 1/3 до 2/3 длины границы Интенсивное окрашивание отдельных мелких и крупных участков
Для исследования под световым микроскопом соединения «пломба –зуб» изготавливали шлифы зубов.
Приготовление шлифов включало следующие операции:заливку образцов в эпоксидную смолу;предварительную шлифовку блока на абразивной бумаге зернистостью 180, 240, 320 мкм до
получения нужного сечения;окончательную шлифовку на абразивной бумаге зернистостью 400, 600 мкм;грубую полировку на велюре с добавлением порошка оксида алюминия зернистостью 12 мкм;тонкую полировку на велюре с добавлением порошка оксида алюминия зернистостью 0,1 мкм.Поверхность шлифов образцов исследовали с помощью светового микроскопа «MeF-З» (ув.
200) фирмы Buehler (Чехия) и оценивали структуру пломбы и состояние границы «зуб – плом-ба» в области эмали. Определяли однородность, качество и количество крупных и мелких ча-стиц, их форму и расположение в пломбировочном материале, наличие или отсутствие объем-ных дефектов как в пломбе, так и в области границы, их количество, размер и расположение. Ширину границы «зуб – пломба» в области эмали измеряли с помощью окуляр-микрометра [6].
Результаты и их обсуждение. Анализ полученных данных показал, что исследуемые образ-цы фотополимеризующихся композитов обладают свойствами, необходимыми для такого рода материалов: хорошо прилегают к твердым тканям зуба, уплотняются, моделируются стоматоло-гическими инструментами и полируются до сухого блеска.
При визуальном исследовании зубов, реставрацию которых осуществляли пломбировочным материалом № 1, во всех случаях поверхность пломб была ровной и гладкой, без трещин, пор и раковин. Дефекты на границе «зуб – пломба» не обнаружены. При зондировании границы «зуб –
Национальная
академия наук
Беларуси

50
пломба» определялся плавный переход от эмали к поверхности пломбы. Окрашивание границы и поверхности пломб лишь в отдельных случаях имело очень слабую интенсивность. Полимери-зованный материал № 1 обладал невыраженным эффектом «хамелеона» и по цвету практически не отличался от твердых тканей зуба.
При осмотре и зондировании поверхности пломб, изготовленных из композита № 2, отсут-ствовал зазор при зондировании границы, однако имелись небольшие дефекты в виде пор. После воздействия красителя и полирования границы «зуб – пломба» интенсивность окрашивания по-верхности пломбы была слабой и очень слабой. Кроме того, образец № 2 отличался красно-коричневым оттенком, который не совпадал по цвету ни с одним из интактных удаленных зубов. Однако при пломбировании измененных в цвете депульпированных и резорциненных зубов воз-можно применение этого композиционного материала.
Образцы № 3 и № 4 также характеризовались хорошим краевым прилеганием. Объемных дефектов и шероховатостей не выявлено. Во всех случаях после нанесения красителя и полиро-вания поверхности образцов не наблюдалось окрашивания поверхности пломбы и ее границы с эмалью. Оттенки цвета совпадали с тканями зуба.
Изучение образцов, изготовленных из фотополимера Filtek Z250 (3М), показало, что рестав-Filtek Z250 (3М), показало, что рестав- Z250 (3М), показало, что рестав-Z250 (3М), показало, что рестав-250 (3М), показало, что рестав-рации из этого композита имели гладкую и ровную поверхность, без дефектов, граница «пломба – зуб» была плавной. Окрашивания реставрированного участка не наблюдалось. Фотополимер обладал выраженным эффектом «хамелеона».
Микроскопическое исследование шлифов образцов показало, что структура пломбировочно-го материала № 1 представлена гетерогенным конгломератом, состоящим из мелких включе-ний округлой формы, равномерно распределенных в композите.
Граница «материал № 1 – эмаль» представляла собой равномерную четкую волнистую ли-нию шириной 4,0±1,93 на всем протяжении (см. таблицу). В области соединения «материал № 1 – дентин» ширина границы варьировалась от 3,7 до 7,6 мкм.
Результаты исследования размеров границы «зуб – пломба» на шлифах исследованных образцов представлены в таблице.
Ширина границы «зуб – пломба» на шлифах образцов № 1–4
Образец материала
Количество образцов
Ширина границы, мкм
«эмаль – пломба» «дентин – пломба»
№ 1 8 4,0±1,93 5,6±2,23№ 2 8 8,0±2,03 9,0±2,23№ 3 10 2,0±0,96 3,5±1,23№ 4 10 0,5±0,23 1,0±0,33Filtek Z250 (3М) 6 0,6±0,25 0,94±0,31
Пломбировочный материал как в области эмали, так и в области дентина плотно прилегал к тканям зуба. Объемных дефектов в области пломбы и ее границы не обнаружено.
Поверхность шлифа пломбировочного материала № 2 имела вид неоднородного конгломе-рата с частицами наполнителя разной формы с отдельными мелкими объемными дефектами в виде пор размером до 2 мкм.
Граница «материал № 2 – эмаль» была равномерной и, как и в первом образце, имела вид чет-кой волнистой линии шириной 8,0±2,03 мкм, соединение «материал № 2 – дентин» также имела вид слегка волнистой линии шириной 9,0±2,23 мкм.
Исследование образцов композиционного материала № 3 выявило, что его структура сопо-ставима со структурой образца материала № 1. Она представляла собой гетерогенный конгломе-рат, состоявший из включений округлой формы, равномерно распределенных в фотополимере. Однако граница «материал № 3 – эмаль» была неравномерной (шириной 2–3 мкм), нечеткой, в отдельных местах расслаивалась. В области до 1/3 длины границы обнаружены единичные объемные дефекты в виде пор размером от 2 до 9 мкм. Ширина границы «материал № 3 – ден-тин» находилась в пределах от 2,2 до 4,1 мкм.
Национальная
академия наук
Беларуси

Микроскопическое исследование шлифов изученных образцов показало, что структура плом-бировочного материала № 4 идентична таковой материала № 3. Однако граница «пломба – зуб» образца № 4 ровная и четкая, ширина ее минимальная по сравнению с остальными образцами (в среднем 0,5±0,23 мкм для эмали и 1,0±0,33 мкм для дентина).
Ширина границы «пломба – зуб» композита Filtek Z250 (3М) составила в среднем 0,6±0,25 мкм для эмали и 0,94±0,31 мкм для дентина.
Таким образом, наименьшая ширина границы «пломба – эмаль» была в зубах, запломбиро-ванных композиционными материалами № 3, № 4 и Filtek Z250 (3М). Ширина границы «материал № 2 – эмаль» превысила аналогичный показатель фотополимеров № 3 в 4 раза и образца № 4 – в 16 раз.
Заключение. Все исследованные образцы композитов обладают необходимыми для такого рода пломбировочных материалов рабочими свойствами: хорошо прилегают к твердым тканям зуба, уплотняются и моделируются стоматологическими инструментами, полируются до сухого блеска. Непосредственно после пломбирования и при зондировании щель между зубом и плом-бой визуально не определяется, после нанесения красителя и полирования поверхности образ-цов поверхности пломбы и ее границы с эмалью не окрашиваются. Наилучшими рабочими свой-ствами при визуальном обследовании границы «пломба – зуб» обладают образцы материалов № 1, № 3, № 4 и Filtek Z250 (3М).
Микроскопическое исследование шлифов образцов показало, что структура пломбировоч-ных материалов № 1, № 3, № 4 и Filtek Z250 (3М) представляет собой гетерогенный конгломерат, состоящий из мелких включений округлой формы, равномерно распределенных в композите. Поверхность шлифа фотополимера № 2 имеет вид неоднородного конгломерата с частицами наполнителя разной формы. Пломбировочные материалы № 2 и № 3 имеют отдельные мелкие объемные дефекты в виде пор, при этом величина пор наибольшая у образца № 3. Образец № 2 отличается самой неравномерной и широкой границей «материал – эмаль».
Таким образом, самыми перспективными для разработки являются пломбировочные мате-риалы № 1 и № 4.
Выполненная работа является частью научной тематики кафедры терапевтической стомато-логии БелМАПО и проведена в рамках научно-исследовательских работ совместно с кафедрой вяжущих материалов Белорусского технологического института (тема НИР «Разработать состав и процесс получения светоотверждаемого импортзамещающего цемента и освоить его произ-водство», государственная регистрация № 20083532 от 30.12.2008 г.)
Литература
1. Л у ц к а я И. К., Н о в а к Н. В. // Совр. стоматол. 2009. № 2. С. 18–22.2. Ч у х р а й И. Г., М а р ч е н к о Е. И., Н о в а к Н. В. // Совр. стоматол. 2009. № 2. С. 14–17.3. Л у ц к а я И. К., Ч у х р а й И. Г., М а р ч е н к о Е. И. // Совр. стоматол. 2003. № 4. С. 44–46.4. С е н а Л. А. Единицы физических величин и их размерности. М., 1988. С. 170–176.5. Т р е з у б о в И. К., Ш т е й н г а р т М. З., М и ш н е в Л. М. Ортопедическая стоматология. Прикладное
материаловедение. СПб., 1999. С. 52.6. P h i l l i p s R. W. Skanner’s science of dental materials. 9th ed. Philadelphia, 1991. P. 69–71.
n. v. novaK
STATE OF THE BORDER «FILLING – TOOTH» IN USING DIFFERENT FILLING MATERIALS
Belarusian Medical academy of pоstgraduate Education, Minsk
Summary
The data of preclinical research of four samples of Belarusian composite in comparison with import filling material are presented. Microscope research of the border “filling – tooth” was made.
Национальная
академия наук
Беларуси

52
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.127 616-076.4
С. А. новАКовСКАя, Е. в. КоЗловСКАя, Е. л. РыЖКовСКАя, о. А. МАнЕЕвА, л. и. АРчАКовА
УЛЬТРАСТРУКТУРНЫЕ ИЗМЕНЕНИЯ В МИОКАРДЕ ПРИ ДИЛАТАЦИОННОй КАРДИОМИОПАТИИ
институт физиологии нАн Беларуси, Минск
(поступила в редакцию 27.10.2009)
Введение. Дилатационная кардиомиопатия (ДКМП) относится к некоронарогенным заболе-ваниям миокарда. В основе ее формирования лежит повреждение и гибель кардиомиоцитов. Данная патология сердца сопровождается прогрессирующим нарушением гемодинамики и харак-теризуется синдромом малого сердечного выброса, расстройствами сердечного ритма и проводи-мости, нарастанием дилатации полостей сердца, миокардиодистрофией. В результате формиру-ется клинический синдром сердечной недостаточности (СН), который выражается в нарушении сократительной способности органа и застое крови в малом и большом круге кровообращения [1]. Морфологической основой СН является генетическая предрасположенность сердца к пере-стройке на клеточном уровне, что сопровождается гипертрофией его полостей, ведущей к ремо-делированию миокарда, недостаточной васкуляризацией, фиброзом, уменьшением количества функциональных кардиомиоцитов [2–6].
В последние годы основная роль в развитии ДКМП отводится процессам апоптоза, который рассматривается как мембранозависимый процесс программируемой клеточной гибели, связан-ный с экспрессией специфических рецепторов Fas/APO-1 и вызывающий структурную пере-стройку сердца в результате необратимых изменений кардиомиоцитов [7–10].
В индукции апоптоза кардиомиоцитов и эндотелиальных клеток центральную роль отводят пептиду ангиотензину II, принимающему непосредственное участие в развитии кардиоваску-лярного фиброза. Повышенный уровень данного пептида связан с активностью некротического и трансформирующих ростовых факторов, а также специфических апоптозных белков и каспаз. Кроме того, развитию апоптоза кардиомиоцитов способствует накопление в организме оксида азота, который в высоких дозах становится медиатором воспалительных и иммунных реакций. В результате усиливается синтез белка – супрессора опухолей р53, активируются протеазы из семейства каспаз, участвующих в процессах программированной клеточной гибели [11].
Широкое распространение получило предположение о вирусном происхождении ДКМП. У большинства больных выявляют повышение активности Т-киллеров и уменьшение активности Т-супрессоров, увеличение выработки органоспецифических аутоантител, в том числе к β-адрено-рецепторам, митохондриям, миозину легких цепей [12].
Большую роль в развитии и прогрессировании СН играют провоспалительные цитокины – фактор некроза опухоли (ФНО-α), интерлейкин-1α (ИЛ-1α), интерлейкин-1β (ИЛ-1β), интерлей-кин-6 (ИЛ-6), интерлейкин-8 (ИЛ-8), которые содержатся в плазме крови больных с ДКМП и оказывают разрушающее действие на внеклеточный коллагеновый матрикс миокарда [13, 14]. Индуктором синтеза данных цитокинов являются активированные клетки иммунной системы – макрофаги и моноциты, содержащиеся в плазме крови и межтканевой жидкости. Доказано, что у таких больных провоспалительные цитокины способствуют уменьшению контрактильных свойств миокарда [15], развитию дилатации желудочков и гипертрофии кардиомиоцитов [16]. Они играют важную роль в регуляции интенсивности процессов ремоделирования миокарда и сосудов, определяющих степень апоптотических изменений и способствующих прогрессирова-
Национальная
академия наук
Беларуси

53
нию СН, которая сопровождается системной гипоксией. Показано, что уровень провоспалитель-ных цитокинов в крови больных с СН зависит от степени тяжести патологического процесса. Таким образом, активация системы цитокинов у больных с СН, в том числе при ДКМП, явля-ется маркером прогрессирования заболевания. Чем выше функциональный класс СН, тем более выраженной является реакция иммунной системы и более высоким уровень цитокинов. Умень-шение степени декомпенсации сопровождается снижением уровня иммунного ответа [17].
ДКМП нередко протекает с симптоматикой других, значительно чаще встречающихся забо-леваний – ишемической болезни сердца, ревматических пороков, гипертонической болезни, легочной гипертензии. Вследствие этого нередко возникают затруднения в диагностике, что тре-бует проведения дифференциального диагноза. Ввиду диагностических ошибок применение этиопатогенетического лечения может быть несвоевременным, поэтому прогнозировать раз-витие заболевания довольно сложно [17]. В настоящее время для объективизации ДКМП в кли-нически неясных случаях используется метод прижизненной эндомиокардиальной биопсии. Цитологический анализ биоптатов, полученных методом эндомиокардиальной биопсии, в кли-нической кардиологии становится важным диагностическим методом [3, 18, 19].
Ввиду отсутствия до настоящего времени четкой согласованности по ультраструктурным критериям ДКМП нами проведено электронно-микроскопическое исследование реорганизации миоцитов, микрососудов и интерстиция сердечной мышцы у пациентов с клиническим диагно-зом ДКМП. Для этого изучены особенности ультраструктурной организации миокарда и его си-стемы микроциркуляции на биоптатах, взятых при трансвенозной эндомиокардиальной биопсии миокарда у больных с данной патологией сердца. В дальнейшем полученные данные будут трансформированы в расширенный диагностический комплекс с позиций системного подхода.
Материалы и методы исследования. Объектом исследования являлся миокард правого же-лудочка сердца 8 больных ДКМП (4 женщины и 4 мужчины) в возрасте от 20 до 42 лет (средний возраст 31 год). Диагноз ДКМП был поставлен на основании клинико-инструментальных иссле-дований. Больные проходили консервативное лечение в течение 2,5–3 лет. Работа выполнена на биоптатах миокарда, полученных путем трансфеморального доступа с использованием направ-ляющего катетера, из трех зон: верхушки правого желудочка, межжелудочковой перегородки и зоны выходного тракта правого желудочка. В настоящей работе представлены данные, полу-ченные на миокарде выходного тракта правого желудочка.
В работе использован электронно-микроскопический метод исследования. Кусочки ткани фиксировали в растворе, состоявшем из 3%-ного глутарового альдегида и 1%-ного параформа, после чего материал измельчали и обрабатывали 2%-ным раствором четырехокиси осмия. После промывания 0,1 М фосфатным буфером материал обезвоживали в спиртах возрастающей крепо-сти и заключали в аралдит по общепринятой методике [20]. Срезы готовили на ультратоме �KB и просматривали в электронном микроскопе JEM-100 CX.
Результаты и их обсуждение. Ультраструктура кардиомиоцитов в норме и при патологии представлена в руководствах и монографиях ряда авторов [21–24]. Однако работы, посвященные электронно-микроскопическому изучению патологии миокарда при ДКМП, в литературе немно-гочисленны [25–30].
Проведенные нами исследования дают полную картину субмикроскопических изменений в кардиомиоцитах и других компонентах миокарда правого желудочка при ДКМП.
Электронно-микроскопические исследования показали мозаичность инволюционных изме-нений в миокарде при ДКМП. Среди кардиомиоцитов с интактной структурой (рис. 1, а, б) вы-являются клетки с литическими изменениями сократительного аппарата, которые носят диф-фузный характер и проявляются истончением миофибрилл с сохранением их ультраструктурной организации либо полным лизисом (рис. 1, в, г). Происходит дезориентация и изменение формы миофибрилл (приобретение контуров «бамбукового ствола») (рис. 1, в), наблюдается разобщение миофиламентов. Во многих саркомерах образуются пустоты, а в ряде случаев отмечаются то-тальный лизис миофиламентов в пределах саркомера (внутриклеточный миоцитолизис), расши-рение вставочных дисков и, как следствие, разобщение соседних миокардиальных клеток. В ряде случаев определяются диффузно расположенные гипертрофированные кардиомиоциты с разоб-
Национальная
академия наук
Беларуси

54
щенными пучками миофибрилл и большим количеством беспорядочно расположенных мелких митохондрий (рис. 1, г).
Отмечаются и другие характерные изменения ультраструктуры сердечных миоцитов: де-струкция и лизис митохондрий, формирование групп лизосом, содержащих материал высокой электронной плотности, что свидетельствует об очаговой дегенерации цитоплазмы и ее орга-нелл (рис. 2, а, б); запустевание цитоплазмы в околоядерных и межфибриллярных простран-
Рис. 1. Ультраструктурная организация кардиомиоцитов биоптатов миокарда при ДКМП: а – кардиомиоциты с со-хранившейся организацией ядра; б – элементы саркоплазмы – миофибриллы и митохондрии; в – истончение и лизис миофибрилл при дистрофии кардиомиоцита; г – фрагменты миофибрилл и беспорядочно рассеянные митохондрии в саркоплазме дистрофически измененного кардиомиоцита. КМЦ – кардиомиоцит, Я – ядро, Мф – миофибриллы,
М – митохондрия, Л – лизосома
Рис. 2. Ультраструктурные изменения кардиомиоцитов при ДКМП: а – деструкция и лизис митохондрий и миофила-ментов в пределах саркомера; б –накопление лизосом в саркоплазме среди истонченных миофибрилл и митохондрий с гомогенизированным матриксом; в – вакуолизация саркоплазмы и лизис миофибрилл кардиомиоцита; г – разрыв сар-колеммы и выход органелл в интерстициальное пространство. ДМ – дегенерирующие митохондрии, Мф – миофибриллы,
М – митохондрия, Л – лизосома, КМЦ – кардиомиоцит, В – вакуоль, ИП – интерстициальное пространство
Национальная
академия наук
Беларуси

55
ствах на фоне общего снижения плотности миофибрилл; скопление многочисленных пузырьков и крупных электронно-прозрачных вакуолей на фоне полного отсутствия миофибрилл в ряде кардиомиоцитов (рис. 2, в); разрыв сарколеммы кардиомиоцитов с выходом органелл за пределы клеток (рис. 2, г).
Выражен полиморфизм кардиомиоцитов и со стороны клеточного ядра. В ряде клеток отме-чается резкая деформация и перетяжка ядер, классифицируемые как апоптозные. В измененных ядрах наблюдается выраженная складчатость нуклеолеммы, а также маргинальная конденсация хроматина (рис. 3, а, б), который зачастую образует в ядре серповидные «шапки» и осмиофиль-ные полоски с повышенной электронной плотностью.
Одновременно с начальными признаками апоптоза выявляются клетки в развернутой фазе апоптотического процесса, при котором наблюдаются уплотнение и деформация кардиомиоцитов, инвагинации кариолеммы с отшнуровыванием ядерных фрагментов, а в ряде случаев – полная фрагментация кариоплазмы с образованием разобщенных участков ядра, не связанных между собой (рис. 3, в). На конечную фазу апоптоза кардиомиоцитов указывает наличие конгломератов фрагментов клеточных тел в межклеточном пространстве – остаточных телец, имеющих сфери-ческую или веретенообразную форму и состоящих из фрагментов ядра и цитоплазмы кардио-миоцита (рис. 3, г). Указанные особенности ультраструктурных изменений кардиомиоцитов при отсутствии в миокарде воспалительной реакции свидетельствуют о том, что элиминация части клеток при ДКМП осуществляется путем апоптоза. Диффузная локализация апоптотически из-мененных кардиомиоцитов в различных областях миокарда позволяет предположить, что вовле-чение миоцитов в патологический процесс происходит асинхронно.
При ДКМП отмечается ультраструктурная реорганизация сосудов микроциркуляторного русла, которая, по мнению большинства исследователей, носит вторичный характер. Полагают, что наблюдаемые у пациентов с ДКМП признаки ишемии миокарда связаны с нарушениями микроциркуляции вследствие ремоделирования желудочков. Гипертрофия ряда миокардиаль-ных клеток приводит к увеличению массы миокарда при отставании роста капиллярной сети.
При исследовании биоптатов миокарда выявляются капилляры с различной структурой. Одни из них сохраняют структурную организацию (эндотелий не изменен, отсутствует пери-васкулярный отек), имеют широкий просвет (рис. 4, а). В других капиллярах могут опреде-
Рис. 3. Ультраструктурная организация кардиомиоцитов при апоптозе: а – деформация ядер; б – маргинальная конден-сация хроматина; в – ультраструктура деформированного и уплотненного кардиомиоцита с фрагментами ядра и фор-мирующимися ядросодержащими апоптозными телами; г – ядросодержащие фрагменты кардиомиоцита – остаточные
(апоптозные) тельца. Я – ядро, КМЦ – кардиомиоцит, ЯДР – ядрышко, Мф – миофибриллы, ОТ – остаточное тельце
Национальная
академия наук
Беларуси

56
ляться эндотелиальные разрастания, которые заполняют просвет капилляра и создают пре-пятствия для кровоснабжения, нарушая микроциркуляцию. В некоторых капиллярах наблю-дается выраженная вакуолизация эндотелиоцитов. Различные по величине вакуоли округлой формы диффузно рассеяны в цитоплазме эндотелиальных клеток. В просветах капилляров выявляются эритроциты, а также фрагменты эндотелиоцитов, образовавшиеся вследствие ми-кроклазматоза (рис. 4, б). Часто наблюдается тотальная обтурация просвета капилляров фор-менными элементами крови и крупными мембранными образованиями, заполненными электронно-прозрачным или зернистым материалом, в результате чего уменьшается количе-ство функционирующих сосудов. Часть микрососудов миокарда находится в состоянии значи-тельного растяжения и напряжения, их просвет заполнен «пакетами» эритроцитов, что свиде-тельствует о застойных явлениях в микроциркуляторном русле при ДКМП. Другая часть микрососудов находится в спавшемся состоянии и имеет вытянутый просвет неправильной формы вследствие выключения микрососудов из кровотока.
Ультрамикроскопические исследования подтверждают существующую точку зрения [4, 27], что нарушение микроциркуляции при ДКМП наступает вследствие разрастания соединитель-ной ткани и развития диффузного заместительного фиброза, обусловленного гибелью апоптоз-ных кардиомиоцитов (рис. 4, в, г). Сдавление капилляров миокарда коллагеновыми волокнами приводит в дальнейшем к разобщению кардиомиоцитов разросшейся соединительной тканью и отдалению их от базальной мембраны функционирующих кровеносных капилляров, что вле-чет за собой разрушение и гибель новых кардиомиоцитов и возникновение ишемической кар-диомиопатии.
Заключение. Электронно-микроскопические исследования эндомиокардиальных биопта-тов больных с ДКМП указывают на различный характер деструктивных процессов, развиваю-щихся в миокарде при ДКМП. Результаты проведенных исследований показали, что во многих кардиомиоцитах имеются выраженные дистрофические изменения – крупные плотные лизосо-мы, резкая вакуолизация саркоплазмы, деформация ядер с маргинальной агрегацией хромати-на, уплотнение и последующая вакуолизация митохондрий с разрывами наружной мембраны.
Рис. 4. Ультраструктура микрососудов и интерстиция миокарда при ДКМП: а – ультраструктура капилляра, приле-жащего к сарколемме кардиомиоцита; б – вакуолизация и утолщение стенок капилляра, заполненного эритроцитами и фрагментом эндотелиоцита; в – разрастание соединительной ткани в интерстициальном пространстве между тела-ми кардиомиоцитов; г – разрастание волокнистой соединительной ткани в интерстициальном пространстве миокар-да (заместительный фиброз). ПК – просвет капилляра, КМЦ – кардиомиоцит, Э – эритроцит, ЭНД – эндотелий,
ИП – интерстициальное пространство
Национальная
академия наук
Беларуси

57
К неспецифическим, но весьма характерным изменениям следует отнести резкое изменение формы ядер многих кардиомиоцитов и дезориентацию миофибрилл. В дистрофически изменен-ных кардиомиоцитах, как показали наши исследования, наблюдаются разрывы и лизис миофи-брилл, миофибриллы с выпавшими саркомерами, что согласуется с данными других авторов [28]. Подобные изменения завершаются некрозом кардиомиоцитов, при котором отмечаются необратимая деструкция и лизис цитоплазматических органелл и саркоплазмы, нарушение целостности сарколеммы, лизис ядра. Процесс сопровождается врастанием коллагеновых во-локон в распадающиеся кардиомиоциты.
Таким образом, нами установлено, что наряду с некротическими повреждениями клетки мио - карда могут подвергаться и запрограммированной гибели – апоптозу, при котором наблюдаются весьма характерные изменения ультраструктуры кардиомиоцита. В начальной фазе развития апоптоза отмечается резкая деформация ядра и инвагинация кариолеммы, которая усугубляется и завершается фрагментацией ядра и формированием апоптозных тел. Апоптоз в миокарде про-текает асинхронно, апоптозные клетки рассеяны диффузно и могут наблюдаться наряду с некро-зом [7]. При отсутствии воспалительной реакции апоптозные тела разрушаются внеклеточно либо захватываются соседними клетками.
Таким образом, при ДКМП отмечается мозаичность поражения кардиомиоцитов: наряду с клетками без изменения ультраструктуры в миокарде локализуются клетки с выраженными дистрофическими, некротическими и апоптозными изменениями. Ультраструктурная реоргани-зация сосудов микроциркуляторного русла миокарда при ДКМП сопровождается разрастанием соединительной ткани вследствие гибели кардиомиоцитов и развития заместительного фиброза. Вследствие сдавления капиллярной сети миокарда коллагеновыми фибриллами репаративный фиброз неизбежно приводит к нарушениям микроциркуляции, в результате чего прогрессирует нарушение трофической функции микроциркуляторного русла сердечной мышцы и развивается ишемическая кардиомиопатия.
Литература
О к о р о к о в А. Н. Диагностика болезней внутренних органов. М., 2004. Т. 8. Диагностика болезней серд-1. ца и сосудов. Болезни миокарда. Сердечная недостаточность.
U e n o H., K a n e l l a k i s P., A g r o t i s A., B o b i k A. // Hypertension. 2000. Vol. 36. P. 89.2. А м о с о в а Е. Н., К о л ь ч е н о в В. В., В е р и ч Н. М. // Клин. медицина. 1990. № 1. С. 42–47.3. Ш у м а к о в В. И., Х у б у т и я М. Ш., И л ь и н с к и й И. М. Дилатационная кардиомиопатия. Тверь, 4.
2003. Н е п о м н я щ и х Л. М., Л у ш н и к о в а Е. Л., С е м е н о в Д. Е. // Бюл. эксперим. биол. 2000. Т. 130, 5.
№ 10. С. 463–468.Р о з е н б е р г В. Д., Н е п о м н я щ и х Л. М. Дилатационная кардиомиопатия. Общая патология и пато-6.
физиология. М., 2004.С и м о н е н к о В. Б., Б о й ц о в С. А., Г л у х о в А. А. // Клин. медицина. 2000. № 8. С. 12–16.7. B e l t r a m i C. A., F i n a t o N., R o c c o M. et al. // Circulation. 1994. Vol. 89. P. 151–163.8. N a r u l a J., M a i d e r N., V i r m a n i R. et al. // N. Engl. J. Med. 1996. Vol. 335, N 16. P. 1182–1189.9. N a r u l a J., A r b u s t i n i E., C h a n d r a s h e k h a r �. // Cardiol. Clin. 2001. Vol. 19, N 1. P. 113–126.10. E n a r i M., S a k a h i r a H., � o k o y a m a H. et al. // Nature. 1998. Vol. 391. P. 43–50.11. C a f o r i o A. �., M a r t i n e t t i M., S c h w a r z G. et al. // 12. Tissue Antigens. 1992. Vol. 39, N 5. P. 236–240.M a t s u m o r i A., � a m a d a T., S u z u k i H. et al. // Brit. Heart J. 1994. Vol. 72. P. 561–566.13. Н а с о н о в Е. Л., С а м с о н о в М. Ю., Н а у м о в В. Г. // Сердечная недостаточность. 2000. Т. 1, № 4. 14.
С. 15–26.H e g e w i s h S., W e h H. J., H o s s f e l d D. K. // �ancet. 1990. Vol. 335. P. 294–295.15. K u n i s a d a K., H i r o t a H., F u j i o �. et al. // Circulation. 1996. Vol. 94. P. 2626–2632.16. Г у р е в и ч М. А. // 17. Трудный пациент. 2006. № 8. C. 55–60.В и х е р т А. М., Ц ы п л е н к о в а В. Г. // Тер. архив. 1991. Т. 3, № 9. С. 128–133.18. Т е р е щ е н к о В. П., А м о с о в а Е. Н., П а н и ч к и н Ю. В. // Врачеб. дело. 1988. № 8. С. 46–48.19. Б о г о л е п о в Н. Н. Методы электронно-микроскопического исследования мозга. М., 1976. 20. П о л и к а р А., Б о Ш. А. Субмикроскопические структуры клеток и тканей в норме и при патологии. 21.
Л., 1962. Л е в и А., С и к е в и ц Ф. Структура и функция клетки. М., 1971. 22. С е р о в В. В., П а у к о в В. С. Ультраструктурная патология. М., 1975. 23. А в ц ы н А. П., Ш а х л а м о в В. А. Ультраструктурные основы патологии клетки. М., 1979. 24.
Национальная
академия наук
Беларуси

М у х а р л я м о в Н. М. // Тер. архив. 1985. № 4. С. 8–22.25. К у ш а к о в с к и й М. С. Xроническая застойная сердечная недостаточность. Идиопатические миокардио-Xроническая застойная сердечная недостаточность. Идиопатические миокардио-роническая застойная сердечная недостаточность. Идиопатические миокардио-26.
патии. СПб., 1998. С е в е р и н В. В., М о ж е й к о Н. П., А к о п о в а И. С. // Трансплантол. и искусств. органы. 1998. № 4. 27.
С. 11–16.А к о п о в а И. С., У д а л ь ц о в С. Н., П о д л у б н а я З. А. // Вестн. трансплантол. и искусств. органов. 28.
2000. № 1. С. 42–47.M o s s e r i M., S c h a p e r J., A d m o n D. et al. // Circulation. 1991. Vol. 84, N 1. P. 203–210.29. C h a n g C. J., C h o u �. �., � e e �. S. // Clin. Med. J. (Engl.). 1998. Vol. 111, N 10. P. 929–933.30.
S. a. novaKovSKaya, E. v. KoZlovSKaya, E. l. RyZhKovSKaya, o. a. ManEEva, l. i. aRchaKova
ULTRASTRUCTURAL CHANGES OF THE MYOCARDIUM DURING DILATATIONAL CARDIOMYOPATHY
institute of physiology of naS of Belarus, Minsk
Summary
Electron microscope studies of endomyocardial biopsy material taken from patients suffering from dilatational cardiomyopathy evidence for the development of the apoptotic process in cardiac myocytes. A clear sign of ultrastructural myocardium reorganization are the lytic changes of myofibrils, the partial elimination of cardiomyocytes by diffuse apoptosis and the formation the substitutional fibrosis. This process is accompanied by the disorder of the trophic function of the microcirculatory bed of the heart due to the pathology of endothelium and to the squeezing of the capillary network by collagen fibrils. The ultrastructural changes we revealed are indicative of a progressing microcirculation disorder in the cardiac muscle and of the development of the ishemic cardiomyopathy.
Национальная
академия наук
Беларуси

59
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616-006.484.03:577.17
А. н. чЕРнов
МОРФОФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ КЛЕТОК ГЛИОМЫ С6 В КУЛЬТУРЕ ПРИ ДЕйСТВИИ РЕКОМБИНАНТНОГО
ФАКТОРА РОСТА НЕРВОВ
институт физиологии нАн Беларуси, Минск
(поступила в редакцию 18.05.2009)
Введение. К настоящему времени собрано огромное количество фактических данных, пока-зывающих, что заболеваемость злокачественными опухолями в мире неуклонно растет, увели-чиваясь ежегодно на 0,5–7,0% [3]. В большинстве стран смертность от неоплазии занимает вто-рое место после смертности от сердечно-сосудистой патологии. По материалам ВОЗ, в 2007 г. в мире было зарегистрировано 12 млн больных онкологическими заболеваниями и 7,6 млн умер-ших от них. В Республике Беларусь за последние 10 лет (1998–2007) малигнизированные опухо-ли выявлены у 355 376 человек [3]. Частота встречаемости неоплазий ЦНС составляет 10,9–12,8 случая на 100 000 населения [7], причем 55–60% неоплазий имеют глиальную природу, 80–90% являются злокачественными [1], имеющими неблагоприятный исход. В связи с этим по-прежнему актуальным является поиск новых подходов к терапии онкопатологии. Потенциальными канди-датами в борьбе с онкозаболеваниями могут служить ростовые факторы и, в частности, фактор роста нервов (ФРН). Он представляет 140 кДа протеин (7S-форма), состоящий из трех субъеди-S-форма), состоящий из трех субъеди--форма), состоящий из трех субъеди-ниц – α, β и γ, из которых только β несет биологическое начало. Нейротрофический белок вызывает дифференциацию симпатических и чувствительных нейронов, являясь в то же время эндогенным началом, синтезирующимся и секретирующимся во многих клетках, тканях и органах млекопита-ющего [5, 10]. ФРН оказывает как стимулирующее, так и ингибирующее действие на митогенную активность и дифференциацию глиальных опухолевых клеток, что определяется активацией ме-ханизмов сигнальной трансдукции, вовлекаемых в данные процессы, а также типом клеточной линии [12]. Однако результаты исследований носят противоречивый характер [15].
Цель работы – исследовать роль рекомбинантного ФРН на морфофункциональный статус культивируемых в течение 3 и 7 сут клеток глиомы С6.
Материалы и методы исследования. Клеточная линия. Перевиваемые клетки глиомы кры-сы С6 были получены из Российской коллекции клеточных культур позвоночных (Институт ци-тологии РАН, г. Санкт-Петербург).
Культура клеток. Опухолевые элементы в количестве 250 тыс. на чашку культивировали на 35-миллиметровых чашках Петри в среде Игла МЕМ с добавлением 10%-ной телячьей сыворот-ки и 10–4 г/мл раствора сульфата гентамицина. Посевы инкубировали при температуре 37 °С в СО2-инкубаторе с 5%-ным содержанием углекислоты [6].
используемые реактивы: минимальная среда Игла; телячья сыворотка (Sigma-Aldrich, США); растворы трипсина в версене (Sigma-Aldrich), сульфата гентамицина (Sigma-Aldrich), β-форма ре-комбинантного ФРН (Sigma-Aldrich), версен («Диалек», Беларусь), трипановый синий (Германия).
протокол добавления реагентов. Спустя сутки после посева клеточную среду меняли на среду, содержавшую 0,5% телячьей сыворотки, 10–4 г/мл раствора сульфата гентамицина и 100 или 200 нг/мл тестируемого агента. Наблюдения осуществляли на 3-и и 7-е сутки после введе-ния ростового фактора.
Национальная
академия наук
Беларуси

60
визуализация культуры. В указанные сроки наблюдения осуществляли прижизненную ми-кроскопическую цифровую съемку глиомы (×312,5). С применением программного обеспечения IM1000 (Image J, версия 1.38r) оценивали такие морфологические параметры, как размеры кле-ток, длину и степень арборизации отростков. Степень арборизации определяли визуально, а так-же количественным методом, используя программу Image J.
индекс пролиферации. Из чашек Петри с культурой удаляли питательную среду, клетки дважды промывали версеном и затем добавляли 0,5 мл раствора трипсина с версеном в соотно-шении 1:3. Чашки с трипсином помещали в инкубатор на 2–3 мин, после чего действие фермента нейтрализовали добавлением порядка 1 мл телячьей сыворотки. Клетки пипетировали и 20 мкл их переносили в гемацитометр. Количество глиоцитов во всей камере Горяева подсчитывали без разведения [2].
оценка жизнеспособности. Жизнеспособность клеток оценивали по окрашиванию погибших клеток 0,2%-ным раствором трипаного синего в фосфатно-солевом буфере (рН 7,2). Для этого сус-пензию клеток смешивали с раствором красителя в соотношении 1:10 и переносили в камеру Горяева, где и производили подсчет (в %) погибших (окрашенных) и живых (светлых) элементов.
Статистическая обработка данных. Каждая тестируемая группа была представлена пятью независимыми посевами. Достоверность различий между средними величинами устанавливали по критерию Манна–Уитни для непараметрических выборок с помощью компьютерной про-граммы StatPlus 2005. Различия считали значимыми при P < 0,05, P < 0,01.
Результаты и их обсуждение. Результаты воздействия β-формы рекомбинантного ФРН на морфофизиологические параметры 3-суточной культуры глиомы С6 представлены в табл. 1. Соглас-но данным табл. 1, введение β-ФРН в среду в дозах 100 и 200 нг/мл вызывало увеличение размеров глиоцитов от 20,6 ± 0,78 до 24,5 ± 0,93 (P < 0,05) и 28,1 ± 0,59 мкм (P < 0,01) соответственно. При этом под действием нейротрофина в обеих концентрациях арборизация выростов уменьшалась с 13,8 ± 1,66 до 6,2 ± 1,58 и 8,9±2,39% (P < 0,01) сответственно, тогда как их длина при введении 100 нг протеина сокращалась, а при введении его в дозе 200 нг увеличивалась (рис. 1, а–в).
Т а б л и ц а 1. Действие β-рекомбинантного ФРН на морфофункциональные характеристики культуры клеток глиомы С6 на 3-и сутки культивирования
Серия Размеры клеток, мкм Арборизация, % Длина отростков, мкм Индекс пролиферации Погибшие клетки, %
Контроль 20,60 ± 0,78 13,80 ± 1,66 70,70 ± 3,78 3,10 ± 0,20 4,20±0,94β-ФРН, 100 нг/мл 24,50 ± 0,93* 6,20 ± 1,58** 63,90 ± 2,91 0,50 ± 0,16** 4,80 ± 0,7β-ФРН, 200 нг/мл 28,10 ± 0,59**¤ 8,90 ± 2,39 101,50 ± 2,99**¤¤ 0,90 ± 0,36** 15 ± 7,55¤
П р и м е ч а н и е. Достоверность различий экспериментальных показателей по сравнению с контролем на 3-и сутки (* – P < 0,05; ** – P < 0,01), на 7-е сутки (° – P < 0,05; °° – P < 0,01), между собой в одинаковые сроки (¤ – P < 0,05; ¤¤ – P < 0,01) и в разные сроки († – P < 0,05; †† – P < 0,01).
В дозах 100 и 200 нг/мл β-ФРН оказывал сильное цитостатическое действие на культуру, что выражалось в резком падении индекса пролиферации с 3,1 ± 0,20 до 0,5 ± 0,16 (Р < 0,01) и 0,9 ± 0,36 (Р < 0,01) соответственно, и усиливал ее гибель в максимальной концентрации до 15 ± 7,55%.
Рис. 1. Глиома С6 на 3-и сутки в контроле (а) и при воздействии рекомбинантного β-ФРН в дозах 100 нг/мл (б) и 200 нг/мл (в). Ув. 312,5
а б в
Национальная
академия наук
Беларуси

61
Описанные морфологические изменения также могут быть объяснены активным отмиранием клеточных элементов, идущим по некротическому или смешанному апоптоз-некротическому типу, когда имеет место нарушение тургорных свойств клеток и происходит их набухание [4].
Результаты привнесения указанных доз нейротрофического белка на 7-е сутки в среду куль-тивирования глиоцитов С6 представлены в табл. 2.
Как следует из табл. 2, характер изменений исследуемых параметров сохранял тенденцию, свойственную 3-суточной культуре, хотя имелись и некоторые отличия. Если на 3-и сутки раз-меры сом под действием разных доз β-ФРН увеличивались (см. табл. 1), то к 7-м суткам они не менялись или немного уменьшались (в пределах ошибки среднего), причем более значительным было уменьшение арборизации выростов – с 16,9 ± 5,09% в контроле до 2,7 ± 0,60 (Р < 0,01) и 4,2 ± 0,83% (Р < 0,05) в опыте (рис. 2, а–в).
Т а б л и ц а 2. Действие β-рекомбинантного ФРН на морфофункциональные характеристики культуры клеток глиомы С6 на 7-е сутки культивирования
Серия Размеры клеток, мкм Арборизация, % Длина отростков, мкм Индекс пролиферации Погибшие клетки, %
Контроль 32,10±4,00** 16,90±5,09 133,20±11,25** 22,00±2,63** 22,90±3,91**β-ФРН, 100 нг/мл 29,80±3,54* 2,70±0,60**°° 72,80±3,66°°† 1,80±0,34°† 15,10±8,02**†β-ФРН, 200 нг/мл 29,50±0,97** 4,20±0,83**° 88,40±3,12*°°¤† 3,80±0,42**°† 20,20±7,90**
Изменения коснулись и протяженности отростков. Если на 3-и сутки наблюдения под действи-ем β-ФРН (100 нг/мл) она немного сокращалась по отношению к контролю, а в дозе 200 нг/мл – увеличивалась, то к недельному сроку она значительно снижалась (P < 0,01) при любой его кон-центрации (табл. 2). Особенно резко менялся индекс пролиферации, который к 7-м суткам в кон-троле существенно возрастал – от 3,1 ± 0,20 до 22,0 ± 2,63 (P < 0,01) и не менее резко падал: при внесении 100 нг/мл β-ФРН – до 1,8 ± 0,34 (P < 0,05), при внесении 200 нг/мл – до 3,8 ± 0,42 (P < 0,05). При дозе 100 нг/мл показатель гибели глиоцитов не отличался от контрольных значений или из-менения были незначительными (в диапазоне ошибки среднего) (табл. 2), тогда как при внесении 200 нг/мл β-ФРН на 3-и сутки он имел тенденцию к возрастанию (см. табл. 1).
Полученные нами результаты частично согласуются с данными работы [14], согласно кото-рым экзогенный ФРН (100 нг/мл) к 4-м суткам подавлял клеточную численность С6 на 53,4 ± 5,15% от контрольной и индуцировал признаки морфологической дифференциации. Последнее расхо-ждение может быть объяснено различной природой и формой использованного белка. Авторы работы [14] применяли 7S-форму мышиного белка, тогда как нами была использована β-форма рекомбинантного ФРН. Морфологическое «созревание» культуры авторы [14] сочли следствием потенцирования препаратом уровня как самого эндогенного белка, так и его рецепторов. Как по-казали результаты других исследователей [12], ФРН является антимитогенным протеином лишь в том случае, когда глиома С6 стабильно экспрессирует рецептор TrkA. Взаимодействие нейро-TrkA. Взаимодействие нейро-. Взаимодействие нейро-трофической субстанции с указанной сенсорной молекулой запускает дифференциацию глиомы С6 в олигодендроциты, поскольку эта клеточная линия представлена двумя основными компо-нентами – астроцитарным и олигодендроцитарным [15]. Более детально вопрос о распределении
Рис. 2. Глиома С6 на 7-е сутки в контроле (а) и при воздействии рекомбинантного β-ФРН в дозах 100 нг/мл (б) и 200 нг/мл (в). Ув. 312,5
а б в
Национальная
академия наук
Беларуси

рецепторов между цитологическими типами в опухоли освещен в работе [8], где показаны раз-личия в экспрессии мРНК р75, TrkA и TrkВ между первичными астроцитами и олидендроцита-TrkA и TrkВ между первичными астроцитами и олидендроцита- и TrkВ между первичными астроцитами и олидендроцита-TrkВ между первичными астроцитами и олидендроцита-В между первичными астроцитами и олидендроцита-ми. В работе [13] установлено, что для глиомы С6 имеет место и субклоновая рецепторная гете-рогенность: С6–2В лишен TrkA, в противоположность С6–2BD12 клеткам, используемым в работе [8], тогда как оба клона содержат р75 рецепторы. Имеется и другое мнение [12], согласно которому ФРН обладает рост-стимулирующим действием на клеточную линию глиобластомы человека, реализуя свой эффект через связывание с TrkA рецептором. На примере клеточных линий глиомы U373, U251 и U87 показано, что на 3-и сутки он дозозависимо увеличивал числен-U373, U251 и U87 показано, что на 3-и сутки он дозозависимо увеличивал числен-373, U251 и U87 показано, что на 3-и сутки он дозозависимо увеличивал числен-U251 и U87 показано, что на 3-и сутки он дозозависимо увеличивал числен-251 и U87 показано, что на 3-и сутки он дозозависимо увеличивал числен-U87 показано, что на 3-и сутки он дозозависимо увеличивал числен-87 показано, что на 3-и сутки он дозозависимо увеличивал числен-ность клеток (соответственно до 133,9 и 16,0% по отношению к контролю) с выходом на плато при воздействии 100 нг/мл.
Выводы1. β-ФРН способен подавлять пролиферацию и стимулировать клеточное отмирание при
максимальной его концентрации на 3-и сутки.2. Ростовой протеин оказывает супрессирующее влияние на процесс морфологической диффе-
ренциации, о чем свидетельствует сокращение элонгации (на 7-е сутки) и арборизации выростов.Работа выполнена при финансовой поддержке Белорусского фонда фундаментальных иссле-
дований, грант № МС08–007.
Литература
1. А н а н ь е в а И. И., М а л к а р о в М. С., К о р с а к о в а Н. А. и др. // Арх. патол. 2007. № 1. С. 54–60.2. Б о ж к о в а В. П., Б р е ж е с т о в с к и й П. Д., Б у р а в л е в В. П. и др. Руководство по культивированию
нервной ткани. М., 1988. 3. З а л у ц к и й И. В., М а ш е в с к и й А. А., А в е р к и н Ю. И. // Медицина. 2009. № 1. С. 8–11. 4. К а л и н и ч е н к о С. Г., М а т в е е в а Н. Ю. // Морфология. 2007. Т. 131, № 2. С. 16–26. 5. К а л ю н о в В. Н. Фактор роста нервной ткани. Минск, 1984.6. К у л а г о в а Т. А., С е м е н к о в а Г. Н., К в а ч е в а З. Б. и др. // Докл. НАН Беларуси. 2007. Т. 51, № 1.
С. 75–78.7. Р о з у м е н к о В. Д., Х о р о ш у н А. П. // Укр. нейрохирург. журн. 2004. № 2. С. 34–41.8. H u t t o n �. A., d e V e l l i s J., P e r e z - P o l o J. R. // J. Neurosci. Res. 1992. Vol. 32. P. 375–383.9. K i m u r a S h., � o s h i n o A., K a t a y a m a �. еt al. // J. Neuro-Oncol. 2002. Vol. 59. P. 199–205.10. � e v i - M o n t a l c i n i R. // Science. 1987. Vol. 237. P. 1154–1162.11. R a b i n S. J., T o r n a t o r e C., B e l i n d a-C a i r n s B. et al. // Mol. Brain Res. 1998. Vol. 56, N 1–2. P. 273–276.12. S i n g e r H., H a n s e n B., M a r t i n i e D., K a r p C. // J. Neuro-Oncol. 1999. Vol. 45. P. 1–8. 13. T o r n a t o r e C., R a b i n S., B a k e r - C a i r n s B. et al. // Cell Transplant. 1997. Vol. 6. P. 317–326.14. W a t a n a b e T., K a t a y a m a �., K i m u r a S. et al. // J. Neuro-Oncol. 1999. Vol. 41. P. 121–128. 15. W e i s C., W i e s e n h o f e r B., H u m p e l C. // J. Neuro-Oncol. 2002. Vol. 56. P. 59–67.
a. n. chERnov
MORPHOFUNCTIONAL STATE OF C6 GLIOMA CELLS CULTURE UNDER THE ACTION OF THE RECOMBINANT NERVE GROWTH FACTOR
institute of physiology of naS of Belarus, Minsk
Summary
The influences of the β-form of the recombinant nerve growth factor (β-NGF, 100 and 200 ng/ml) on the range of morphological (sizes, length and degree of arborization of processes) and physiological (index of proliferation and downfall) characteristics of C6 rat glioma cells by 3 and 7 days of cultivation were investigated in the article. Results are stated that β-NGF inhibited of index of proliferation of gliocytes in the both quantities on 3 (p < 0.01) and 7 (p < 0.05) days of cultivation. Agent caused decrease of arborisation (p < 0.05) and elongation of processes (p < 0.01) glioma cells in both doses to 7 day. Therefore the factor has antimitogenic activity on C6 culture cells.
Национальная
академия наук
Беларуси

63
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.441-006.6-089.87:612.015
Т. А. МиТЮКовА1, Т. А. лЕоновА1, А. А. ТУЗовА1, Т. Ю. плАТоновА1, М. л. лУЩиК1, л. Г. КУлАКовСКАя2, в. М. ДРоЗД1
БИОХИМИЧЕСКИЕ ПОКАЗАТЕЛИ КРОВИ У ПАЦИЕНТОВ С КАРЦИНОМОй ЩИТОВИДНОй ЖЕЛЕЗЫ ПОСЛЕ ТОТАЛЬНОй ТИРЕОИДЭКТОМИИ
1 Белорусская медицинская академия последипломного образования, Минск, 2 Республиканский центр медицинской реабилитации и бальнеолечения, Минск, Беларусь
(поступила в редакцию 10.03.2010)
Введение. Пациенты, прооперированные по поводу карциномы щитовидной железы (ЩЖ), требуют постоянного наблюдения с целью выявления возможных рецидивов, а также для оцен-ки общего состояния, контроля и коррекции назначенного им медикаментозного лечения [1].
На сегодняшний день тотальная тиреоидэктомия (ТТЭ) и латеральная шейная лимфодиссек-ция признаны оптимальным хирургическим вмешательством при дифференцированном раке щи-товидной железы (ДРЩЖ) [1, 2]. Оценивая хирургическое лечение 675 больных с ДРЩЖ в период с 1993 по 2002 г. в Беларуси, авторы работы [3] показали, что в большинстве случаев проводилось радикальное удаление тиреоидной ткани, а при возобновлении опухолевого роста на фоне первич-ной органосохраняющей операции – повторная операция с удалением остатков ткани ЩЖ.
Таким образом, по данным литературы, среди пациентов, прооперированных по поводу ра-диоиндуцированного рака ЩЖ, за последние 15 лет преобладают случаи радикального удале-ния ткани ЩЖ с диссекцией клетчатки шеи. Последнее, как предполагается, может привести к повреждению паращитовидных желез и нарушению их функции. Наши данные показывают, что среди молодых пациенток с ДРЩЖ, прооперированных более 10 лет тому назад, в 32% слу-чаев наблюдается некомпенсированный гипопаратиреоз [4].
Важнейшей задачей мониторинга больных с ДРЩЖ является лечение супрессивными доза-ми �-тироксина (�-t4), что необходимо для подавления эндогенной продукции тиреотропного гормона (ТТГ) [1]. В процессе наблюдения не менее 2 раз в год проводится определение уровней ТТГ, тиреоидных гормонов, тиреоглобулина, а также комплекса биохимических показателей в сыворотке крови [1].
В литературе, посвященной изучению метаболических особенностей у онкологических боль-ных, широко представлены сведения по антиоксидантному статусу и перекисному окислению липидов [5–7], но практически отсутствуют данные о влиянии продолжительного комплексного лечения на рутинные биохимические показатели, хотя эта информация является крайне важной для оценки клинического состояния пациентов, а также для коррекции проводимого лечения. При ранней диагностике заболевания и адекватном хирургическом лечении ДРЩЖ имеет благо-приятный прогноз, поэтому первостепенной задачей является сохранение удовлетворительного качества жизни пациентов, своевременное выявление сопутствующей патологии, а также сниже-ние риска побочных эффектов от комплексного лечения основного заболевания, что невозможно без анализа рутинных биохимических исследований.
В последнее время предлагается пересмотреть уровень супрессии ТТГ, необходимый для ве-дения пациентов с различными степенями риска по возобновлению рецидивов, что активно дис-кутируется [8, 9]. В группах низкого риска представители Европейской тиреоидной ассоциации B. Biondi и др. [8] рекомендуют поддерживать ТТГ на уровне 0,5–2,5 мМЕ/л. Исходя из рекомен-
Национальная
академия наук
Беларуси

64
даций Американской тироидной ассоциации (ATA) [9], для этой категории пациентов необходи-мо выдерживать интервал супрессии ТТГ в пределах от 0,1 до 0,5 мМЕ/л, а для больных с высо-ким риском – менее 0,1 мМЕ/л. В своей работе мы ориентировались преимущественно на реко-мендации АТА, выделяя пациентов с разными уровнями супрессии ТТГ – менее 0,1 мМЕ/л (суперсупрессия) и менее 0,5 мМЕ/л (супрессия).
Цель работы – изучить особенности биохимического статуса пациентов с тотальной тиреои-дэктомией по поводу ДРЩЖ в зависимости от супрессивной терапии �-t4, проявлений после-операционного гипопаратиреоза и продолжительности лечения.
Материалы и методы исследования. Группа пациентов с ТТЭ состояла из 244 чел. (178 жен-щин и 66 мужчин) в возрасте от 19 до 40 лет (средний возраст 26,69±4,55 года). В качестве сравнения представлена группа больных с гемитиреоидэктомией (ГТЭ) – 24 чел. аналогичного возраста. Из общего количества пациентов были исключены лица, имевшие сопутствующую эндокринную па-тологию и другие заболевания, а также отклонения от нормы по индексу массы тела. Пациенты с ТТЭ были подразделены на две подгруппы: 1) без явных проявлений гипопаратиреоза (нормаль-ный уровень кальция (Са) в сыворотке крови, возможны незначительные транзиторные колебания, субъективные жалобы отсутствуют) – 154 чел.; 2) с проявлениями гипопаратиреоза (при первичном обращении уровень Са ниже нормы, назначаются препараты Са и витаминов группы D, наличие субъективных жалоб) – 78 чел. Группа контроля состояла из 54 практически здоровых молодых лю-дей (42 женщины и 12 мужчин) в возрасте от 18 до 38 лет (средний возраст 26,40±4,32 года).
Уровень тиреоидных гормонов в сыворотке крови – тироксин свободный, трийодтиронин свободный, а также ТТГ определяли ИФА-методом с использованием наборов фирмы «Диалаб» (Австрия), все определения выполняли в дублях. Дополнительно был проведен референтный анализ ТТГ с использованием ИФА-наборов фирмы «Хоффман Ля Рош» (Швейцария) на закры-той автоматической системе «Модуляр 170».
Биохимические исследования выполняли с использованием наборов «Кормэй-Диана» (Поль-ша) на автоматическом анализаторе HITACHI фирмы Boehringer Mannheim (Германия), а также на анализаторе электролитов ROCHE (Швейцария). Определяли уровни Са, фосфора (Р), общего холестерола, холестерола липопротеидов низкой (ХЛНП) и высокой плотности (ХЛВП), три-глицеридов, билирубина, мочевины, креатинина, железа (Fe), активность щелочной фосфата-Fe), активность щелочной фосфата-), активность щелочной фосфата-зы, АЛТ, АСТ, а также уровни электролитов: калия (K), натрия (Na) и ионизированного кальция (Са++). Интервал нормы для общего Са – 2,1–2,7 ммоль/л, для Са++ – 0,9–1,3 ммоль/л.
Статистический анализ полученных данных проводили в таблицах Microsoft Excel с использова-Microsoft Excel с использова- Excel с использова-Excel с использова- с использова-нием критерия Стьюдента для сравнения двух групп и поправки Бонферрони для нескольких групп, результаты представлены в виде средних величин с их стандартными отклонениями (X±Sd) [10].
Результаты и их обсуждение. На момент аварии на ЧАЭС и на момент операции обследованные лица не имели достоверных отличий по возрасту, примерно одинаковым был также средний возраст на период обследования, не было достоверных отличий и по длительности лечения (рис. 1).
Рис. 1. Возрастные характеристики пациентов обследованных групп (контроль, ТТЭ, ГТЭ)
Национальная
академия наук
Беларуси

65
В табл. 1 представлены средние уровни биохимических показателей в сыворотке крови у па-циентов с ТТЭ, ГТЭ и у лиц из группы контроля.
Т а б л и ц а 1. Биохимические показатели сыворотки крови у пациентов с ТТЭ, у больных с ГТЭ и в группе контроля (Х ± Sd)
Биохимический показатель ТТЭ (n = 244) ГТЭ (n = 24) Контроль (n = 54)
Мочевина, ммоль/л 4,06±1,10 5,53±1,18 4,96±1,85Креатинин, ммоль/л 82,15±8,01 81,32±8,47 86,82±8,99Щелочная фосфатаза, ед/л 54,36±23,26 60,88±41,85 64,69±11,42Холестерол, ммоль/л 4,47±0,86 4,53±0,95 4,53±0,74ХЛВП, ммоль/л 1,68±0,40 1,64±0,39 1,51±0,35ХЛНП, ммоль/л 2,66±0,72 2,70±0,95 2,68±0,87Билирубин, ммоль/л 12,72±6,99 11,67±6,91 12,85±5,90Глюкоза, ммоль/л 5,13±0,56 5,27±0,81 5,13±0,41Na, ммоль/л 140,46±2,62 139,44±3,20 140,34±2,07K, ммоль/л 4,56±0,40 4,62±0,39 4,52±0,42Са, ммоль/л 2,29±0,27* 2,48±0,15 2,47±0,11Ca++, ммоль/л 1,17±0,14* 1,26±0,06 1,25±0,06Р, ммоль/л 1,39±0,70* 1,15±0,22 1,24±0,19Fe, мкмоль/л 15,36±5,51* 18,55±10,29 23,09±8,84АСТ, ед/л 20,32±7,05 17,73±3,57 19,32±5,63АЛТ, ед/л 19,99±9,45 16,76±5,37 17,24±6,52
П р и м е ч а н и е. * – достоверные отличия (Р < 0,05) от группы контроля.
У больных с ТТЭ имелся ряд достоверных отличий от контроля, а именно – снижение сред-него содержания Са, Са++ и Fe, а также повышение содержания P в сыворотке крови. Пациенты с ГТЭ не имели достоверных отличий от контроля.
Далее был проведен анализ биохимических показателей у пациентов с ТТЭ в зависимости от эффективности супрессивной терапии, оцениваемой по уровню ТТГ. Средняя доза тироксина со-ставляла 2,8 + 0,6 мкг/кг массы тела, однако только примерно у 70% обследованных больных имелся необходимый уровень снижения ТТГ (ТТГ < 0,5 мМЕ/л). Пациенты были разделены на четыре группы: 1-я – суперсупрессия ТТГ (ТТГ < 0,1 мМЕ/л), 2-я – супрессия ТТГ (ТТГ < 0,5 мМЕ/л), 3-я – ТТГ в интервале нормы (0,5−4,2 мМЕ/л) и 4-я – некомпенсированный гипотиреоз (ТТГ > 4,2 мМЕ/л).
Т а б л и ц а 2. Биохимические показатели сыворотки крови пациентов с ТТЭ в зависимости от уровня ТТГ (Х ± Sd)
Биохимический показатель Группа 1 (ТТГ < 0,1 мМЕ/л)
Группа 2 (ТТГ < 0,5 мМЕ/л)
Группа 3 (ТТГ = 0,5–4,2
мМЕ/л)
Группа 4 (ТТГ > 4,2 мМЕ/л)
Мочевина, ммоль/л 5,95±1,32 4,02±1,29 4,17±1,49 4,18±0,53Креатинин, ммоль/л 82,18±6,16 81,69±7,19 81,44±8,39 91,00±12,25Щелочная фосфатаза, ед/л 59,68±15,47 54,70±25,20 50,29±15,76 65,96±19,93#Холестерол, ммоль/л 4,47±1,03 4,40±0,82 4,54±0,82 5,07±1,18^ХЛВП, ммоль/л 1,58±0,28 1,68±0,38 1,57±0,29 1,93±0,60ХЛНП, ммоль/л 1,59±0,23 2,67±0,73 2,48±0,56 2,96±0,85Билирубин, ммоль/л 12,66±5,78 13,20±7,45 11,66±5,69 10,93±5,19Глюкоза, ммоль/л 5,19±0,49 5,17±0,53 5,04±0,62 4,97±0,64Na, ммоль/л 141,26±2,74 140,57±2,57 140,06±2,52 140,73±3,77K, ммоль/л 4,56±0,41 4,5±0,41 4,56±0,38 4,71±0,28
Национальная
академия наук
Беларуси

66
Биохимический показатель Группа 1 (ТТГ < 0,1 мМЕ/л)
Группа 2 (ТТГ < 0,5 мМЕ/л)
Группа 3 (ТТГ = 0,5–4,2 мМЕ/л)
Группа 4 (ТТГ > 4,2 мМЕ/л)
Ca, ммоль/л 2,25±0,32 2,31±0,26 2,23±0,29 2,31±0,28Ca++, ммоль/л 1,11±0,16 1,17±0,14 1,17±0,15 1,18±0,15P, ммоль/л 1,42±0,33 1,35±0,32# 1,48±0,42 1,49±0,51Fe, мкмоль/л 13,31±3,64 15,22±5,64 16,23±5,92 14,07±2,08АСТ, ед/л 18,71±4,72 19,91±6,50 20,38±6,96 24,80±11,41*АЛТ, ед/л 18,81±6,76 20,22±9,25 18,98±9,37 20,87±12,28
П р и м е ч а н и е. Достоверность отличий (P < 0,05): * – по сравнению с 1-й группой; ^ – по срав-нению со 2-й группой; # – по сравнению с 3-й группой.
Как видно из табл. 2, у лиц с разными уровнями ТТГ обнаруживались некоторые отличия в биохимических показателях, например: повышение активности щелочной фосфатазы при ги-потиреозе по сравнению с ее активностью у больных, у которых ТТГ был в пределах нормы; по-вышение уровня холестерола при гипотиреозе по сравнению с его содержанием у пациентов из группы супрессии (ТТГ < 0,5 мМЕ/л); повышение активности АСТ при гипотиреозе по сравне-нию с данным показателем у больных с суперсупрессией ТТГ (ТТГ < 0,1 мМЕ/л); достоверные отличия по уровню P между пациентами с супрессией и больными с нормальным уровнем ТТГ. Обращает на себя внимание накопление неблагоприятных изменений у пациентов с некомпенси-рованным послеоперационным гипотиреозом. Поскольку целью супрессивной терапии является компенсация гипотиреоза, то выявленные биохимические сдвиги можно рассматривать как вре-менные, подлежащие коррекции.
Рис. 2. Показатели метаболизма липидов и уровень глюкозы в сыворотке крови пациентов с ТТЭ в зависимости от состояния тиреоидного статуса и длительности лечения (А – более 10 лет, Б – менее 10 лет)
продолжение табл. 2Национальная
академия наук
Беларуси

67
Данные по изучению показателей метаболизма в зависимости от уровня ТТГ и продолжи-тельности лечения выборочно представлены на рис. 2.
Как видно из рис. 2, статистически значимое повышение уровня холестерола при неком-пенсированном гипотиреозе отмечалось только у лиц с продолжительным, более 10 лет, сро-ком лечения. При этом наблюдалась тенденция к повышению уровня как ХЛВП, так и ХЛНП, благодаря чему коэффициент атерогенности оставался на относительно стабильном допусти-мом уровне. На фоне менее продолжительного лечения отличия в тиреоидном статусе не при-водили к достоверным сдвигам липидного метаболизма и уровня глюкозы в сыворотке крови.
На фоне длительного лечения выявлены достоверные изменения в фосфорно-кальциевом об-мене (табл. 3).
Т а б л и ц а 3. Средние уровни показателей фосфорно-кальциевого обмена у пациентов с ТТЭ в зависимости от продолжительности лечения (Х±Sd)
Длительность лечения, летБиохимический показатель
Са, ммоль/л Ca++, ммоль/л Р, ммоль/л
10,14±5,15 (общая гр.) 2,29±0,27 1,17±0,14 1,39±0,7014,0±2,61 (более 10 лет) 2,24±0,03 1,13±0,16 1,46±0,385,37±3,01 (менее 10 лет) 2,35±0,20* 1,21±0,11* 1,31±0,32*Контроль 2,47±0,11 1,25±0,06 1,24±0,19
П р и м е ч а н и е. * – достоверные отличия (Р < 0,05) между группами с разной продолжительностью лечения (более 10 лет, менее 10 лет).
У пациентов с продолжительным сроком лечения (в среднем около 14 лет) наблюдались более низкие значения общего Са и Са++ и повышенные – Р (табл. 3). Среди лиц, которые ле-чились более 10 лет, случаи снижения общего Са (менее 2,1 ммоль/л) и Са++ (менее 1,12 ммоль/л) отмечались у 27,6 и 28,6% больных соответственно, а среди лиц, которые лечились менее 10 лет, эти показатели снизились у 9,3 и 10,2% пациентов соответственно. Выявленные отличия у больных с разной продолжительностью лечения были достоверными. Очевидно, это связано с тем, что за последние 5–7 лет тактика оперативного удаления ЩЖ существенно улучшилась, что способствовало резкому снижению осложнений в виде гипопаратиреоза.
Как упоминалось выше, при первичном обращении признаки гипопаратиреоза были зареги-стрированы примерно у одной трети пациентов с ТТЭ. По результатам последних исследований (2009 г.) даже на фоне назначенного лечения у пациентов с гипопаратиреозом не всегда достига-лась нормализация уровня Са в крови (рис. 3, 4). Последнее может быть связано с нерегулярным приемом назначенных препаратов.
Рис. 3. Уровень кальция в сыворотке крови пациентов с ТТЭ в зависимости от сроков лечения и проявлений гипопа-ратиреоза. Достоверность отличий (P < 0,05): # – между группами больных в зависимости от продолжительности
лечения; * – между пациентами с проявлениями и без проявлений гипопаратиреоза
Национальная
академия наук
Беларуси

68
Рис. 4. Процент отклонений уровней общего и ионизированного Са в сыворотке крови пациентов с ТТЭ в зависи-мости от сроков лечения и проявлений гипопаратиреоза. Достоверность отличий (P < 0,05): # ― между группами больных в зависимости от продолжительности лечения; * ― между пациентами с проявлениями и без проявлений
гипопаратиреоза
Как показано на рис. 3, при менее продолжительном сроке лечения отмечается более высо-кий средний уровень Са++ в группе пациентов без проявлений гипопаратиреоза и более высо-кий средний уровень общего Са в группе больных с проявлениями гипопаратиреоза по сравне-нию с соответствующими показателями у пациентов при длительных сроках лечения. Очевидно, что у лиц с проявлениями гипопаратиреоза наблюдаются более низкие средние уровни изучае-мых показателей во всех подгруппах.
У лиц без проявлений гипопаратиреоза при коротких сроках лечения отмечалось достоверно меньше сниженных значений Са++, чем при длительном лечении (рис. 4). У лиц с проявлениями гипопаратиреоза при сроках лечения менее 10 лет наблюдалось достоверно меньше пониженных значений как общего Са, так и Са++, чем при сроках лечения более 10 лет (рис. 4). Кроме того, у лиц с проявлениями гипопаратиреоза отмечалось достоверно большее количество сниженных зна-чений общего Са и Са++ во всех подгруппах, чем у лиц без проявлений гипопаратиреоза (рис. 4).
Заключение. Таким образом, обследованная нами группа пациентов с ТТЭ имела ряд досто-верных отличий от группы контроля, а именно – снижение среднего содержания общего и иони-зированного Са и Fe, а также повышение содержания P в сыворотке крови.
У пациентов с ТТЭ на фоне разных уровней ТТГ обнаруживались следующие отличия:повышение активности щелочной фосфатазы при гипотиреозе по сравнению с ее активно-
стью у пациентов, у которых ТТГ был в пределах нормы;повышение уровня холестерола при гипотиреозе по сравнению с его содержанием у больных
с супрессией ТТГ, наиболее выраженное при длительном лечении (более 10 лет);повышение активности АСТ при гипотиреозе по сравнению с данным показателем у пациен-
тов с суперсупрессией ТТГ (ТТГ < 0,1 мМЕ/л);достоверные отличия по уровню фосфора между больными с супрессией и нормальным
уровнем ТТГ. У больных, получавших более продолжительное лечение (в среднем 14 лет), отмечались бо-
лее низкие средние уровни общего и ионизированного Са по сравнению с аналогичными показа-телями у пациентов, лечившихся менее 10 лет (в среднем 5 лет), что связано с тактикой хирурги-ческого лечения.
У большинства пациентов с проявлениями гипопаратиреоза наблюдалось снижение общего и ионизированного Са (около половины пациентов с длительными сроками лечения), несмотря на назначенное им лечение.
Таким образом, одним из важных вопросов при наблюдении пациентов, прооперированных по поводу ДРЩЖ, является компенсация нарушенного метаболизма кальция. Необходим регу-
Национальная
академия наук
Беларуси

лярный контроль за приемом назначенных препаратов, а также проведение индивидуального подбора доз и препаратов кальция.
Работа выполнена в рамках проекта «Создание международного научно-практического цен-тра заболеваний щитовидной железы».
Литература
1. Л е о н о в а Т. А., Д р о з д В. М., М и т ю к о в а Т. А. и др. Современные аспекты комплексного лечения пациентов с дифференцированным раком щитовидной железы: инструкция по применению метода. Рег. № 153–1108. Минск, 2009.
2. D e m i d c h i k �u. E., D e m i d c h i k E. P., D r o z d V. M. et al. // Диагностика, лечение, профилактика пато-// Диагностика, лечение, профилактика пато-логии щитовидной железы и сопутствующих заболеваний в современных экологических условиях Беларуси: мате-риалы науч.-практ. бел.-нем. конф. Минск, 2007. С. 52–68.
3. Ш е п е т ь к о М. Н., Д е м и д ч и к Ю. Е. // Мед. новости. 2005. № 11. С. 93–95.4. Л е о н о в а Т. А., Д р о з д В. М., М и т ю к о в а Т. А. и др. // Ранняя диагностика, лечение и реабилитация
пациентов с раком щитовидной железы: сб. науч. тр. Минск, 2009. С. 43–52.5. М а ш е в с к а я Е. А., П р о х о р о в а В. И. // Мед. новости. 2007. № 3. С. 80–84.6. М о р о з к и н а Т. С., С у к о л и н с к и й В. Н., М о р о з к и н а Н. В. // О роли биоантиоксидантов в лечении
и профилактике различных патологических состояний: сб. науч. тр. Гомель, 1999. С. 1–7.7. П р о х о р о в а В. И., Д е р ж а в е ц Л. А., Л а п п о С. В., Ю р е в и ч Н. В. // Весці НАН Беларусі. Сер. мед.
навук. 2008. № 3. С. 11–13.8. B i o n d i B., F i l e t t i S., S c h l u m b e r g e r M. // Nat. Clin. Pract. Endocrinol. Metab. 2005. N 1. P. 32–40.9. C o o p e r D. S., D o h r t y G. M., H a u g e n B. R. et al. // Thyroid. 2006. N 16. P. 109–142.10. С т е н т о н Г. Медико-биологическая статистика / Пер. с англ.; под ред. Н. Е. Бузикашвили, Д. В. Са-. Медико-биологическая статистика / Пер. с англ.; под ред. Н. Е. Бузикашвили, Д. В. Са- Бузикашвили, Д. В. Са-Бузикашвили, Д. В. Са- Са-Са-
мойлова. М., 1999.
T. a. MiTyuKova1, T. a. lEonova1, a. a. TuZova1, T. yu. plaTonova1, M. l. luShchyK1, l. G. KulaKovSKaya2, v. M. dRoZd1
BIOCHEMICAL BLOOD INDICES OF PATIENTS WITH THYROID CARCINOMA AFTER TOTAL THYREOIDECTOMY
1Belarusian Medical academy of postgraduate Education, Minsk, 2Republican center for Medical Rehabilitation and Water-Therapy, Minsk, Belarus
Summary
Peculiarities of the metabolic status in patients thyroidectomised due to differentiated papillary thyroid carcinoma were studied in this article. Some unfavorable shifts in biochemical indices were revealed in the general group of operated patients and especially in subjects with non-compensated postoperative hypoparathyroidism. It was shown that the most important practical question in patients with manifestations of hypoparathyroidism is the necessity of impaired calcium metabolism correction. It makes regular serum total and ionized calcium levels check-ups essential, as well as the prescription of adequate medication to such patients.
Национальная
академия наук
Беларуси

70
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.018.51+616.018.54]:541.515
и. А. новиКовА, Ю. и. яРЕЦ
ВЗАИМОСВЯЗЬ СТЕПЕНИ ОКИСЛЕННОСТИ ПЛАЗМЫ И ЭРИТРОЦИТОВ В УСЛОВИЯХ АКТИВАЦИИ СВОБОДНОРАДИКАЛЬНОГО ОКИСЛЕНИЯ
Гомельский государственный медицинский университет, Беларусь(поступила в редакцию 04.09.2009)
Введение. Перекисное окисление липидов (ПОЛ) является неспецифическим универсальным регулятором метаболических процессов в организме [13, 14]. Показано, что интермедиаты ПОЛ играют роль вторичных мессенджеров, обеспечивающих передачу сигналов через мембраны кле-ток, и участвуют в реализации компенсаторно-адаптационных реакций организма [6, 10, 17]. С другой стороны, чрезмерная активация ПОЛ может приводить к повреждению клеточных мем-бран и служить одной из причин возникновения и прогрессирования патологических состояний. Определение содержания продуктов липопероксидации в биологических материалах широко ис-пользуется при исследовании патогенетических механизмов развития заболеваний [12, 16].
В клинической практике для оценки интенсивности ПОЛ обычно определяют один или несколько продуктов окислительных превращений гидроперекисей липидов: первичные – дие-новые коньюгаты (ДК), вторичные – кетодиены, сопряженные триены (СТ) и малоновый диаль-дегид, конечные продукты липопероксидации – основания Шиффа (ОШ). В качестве биологиче-ского материала, как правило, используют плазму крови и/или эритроциты. Считается, что накопление продуктов ПОЛ в плазме и эритроцитах опосредовано выходом из зоны патологиче-ского процесса в различных тканях [2, 12, 15]. Однако имеются сообщения, что показатели ПОЛ мембран эритроцитов в большей степени отражают структурно-функциональное состояние са-мих эритроцитов, чем накопление продуктов липопероксидации в очаге поражения [4, 11, 18]. Взаимосвязь между содержанием гидроперекисей липидов в плазме и эритроцитах при клини-ческих состояниях, сопровождающихся активацией ПОЛ, не исследовалась.
Цель работы – изучить особенности распределения продуктов ПОЛ в плазме и эритроцитах в норме и в условиях активации свободнорадикального окисления.
В качестве модели активации свободнорадикального окисления выбрана локальная гранули-рующая рана, возникшая случайным образом (в быту или на производстве) у практически здоро-вых людей среднего и молодого возраста, что позволяет исключить влияние других факторов, способных изменять содержание ПОЛ.
Материалы и методы исследования. Обследовано 80 больных (58 мужчин, 22 женщины в возрасте от 19 до 53 лет) с локальными гранулирующими ранами различных сроков давности (от 3 до 85 сут), госпитализированных в Гомельский областной центр термической травмы для оперативного лечения. Среди обследованных 53 человека имели локальные ожоги тела и конеч-ностей II-III А-Б – IV степени, составлявшие от 0,3 до 8% от общей поверхности тела с пло - щадью глубокого ожога от 0,07 до 4%. У 27 человек раны носили посттравматический или пост-некротический характер. В зависимости от наличия клинических признаков готовности раны к оперативному восстановлению кожного покрова (отсутствие признаков воспаления, отсутствие выраженной экссудации, высокая адгезивность ран, наличие краевой эпителизации) вышеука-занным больным была проведена операция одномоментной аутодермопластики (АДП).
В качестве контрольной группы обследовано 40 здоровых доноров Гомельской областной станции переливания крови (28 мужчин и 12 женщин), сопоставимых по возрасту.
Национальная
академия наук
Беларуси

71
Материалом для исследования служила гепаринизированная венозная кровь. Содержание пер-вичных (ДК), вторичных (СТ) и конечных (ОШ) продуктов ПОЛ в плазме и эритроцитах крови оценивали спектрофотометрически, отдельно регистрируя уровень липопероксидов в гептановой и изопропанольной фазах (отражает пероксидацию нейтральных жиров и фосфолипидов соответ-ственно). Концентрацию эритроцитов в суспензиях контролировали фотометрически. Результаты выражали в единицах индексов окисления – Е232/220 (содержание ДК), Е278/220 (уровень СТ) и Е400/220 (содержание ОШ) [5, 7]. Для удобства сопоставления уровня ПОЛ в плазме и эритроци-тах рассчитывали индекс распределения (ИР) как отношение содержания ДК, СТ и ОШ в плазме к аналогичному показателю в эритроцитах (ДКп/ДКэ, СТп/СТэ, ОШп/ОШэ соответственно).
Статистический анализ проводили с использованием пакета программ SPSS 13.0. С учетом результатов проверки на нормальность распределения применяли непараметрические методы статистики – критерий Манна–Уитни (для несвязанных выборок), критерий Вилкоксона (в свя-занных выборках). Для сравнения более двух выборок использовали модель однофакторного дисперсионного анализа. Характеристики распределения представляли в виде 25%–М–75%, где 25% – нижний квартиль; М – медиана; 75% – верхний квартиль [9].
Результаты и их обсуждение. Показатели липопероксидации плазмы и эритроцитов здоро-вых лиц и обследованных больных представлены в таблице.
Содержание продуктов ПОЛ в плазме и эритроцитах и значения ИР у больных с ранами и у здоровых лиц (25%–М–75%)
Определяемый параметр
Значения показателей (25%–М–75%)
Здоровые лица (n = 40) ИР Больные (n = 80) ИР
показатели пероксидации нейтральных жиров (гептановая фаза)ДКп 0,535–0,649–0,695
0,98–0,98–10,672–0,730–0,786*
0,94–0,98–1,05ДКэ 0,565–0,644–0,698 0,678–0,718–0,765*СТп 0,097–0,100–0,110
0,93–0,95–0,970,160–0,210–0,236*(**)
0,71–0,84–0,951СТэ 0,100–0,106–0,110 0,180–0,237–0,280*
ОШп 0,015–0,021–0,0230,94–0,96–1
0,016–0,020–0,0230,92–0,94–0,95
ОШэ 0,016–0,021–0,024 0,017–0,021–0,025показатели пероксидации фосфолипидов (изопропанольная фаза)
ДКп 0,402–0,428–0,4450,98–0,98–0,99
0,621–0,678–0,729*(**)0,89–0,94–0,981
ДКэ 0,408–0,428–0,459 0,670–0,702–0,783*СТп 0,210–0,215–0,220
0,96–0,98–10,313–0,372–0,419*(**)
0,9–1,35–1,581СТэ 0,210–0,218–0,223 0,262–0,291–0,325*
ОШп 0,018–0,021–0,0300,75–0,8–1
0,039–0,055–0,078*(**)0,52–0,7–0,821
ОШэ 0,019–0,024–0,035 0,055–0,068–0,111*
П р и м е ч а н и е. Достоверность различий: * – в содержании продуктов ПОЛ между больными и здоровыми лицами (Р < 0,05); ** – между соответствующими показателями плазмы и эритроцитов; 1 – в значениях ИР между больными и здоровыми лицами (Р < 0,05).
Как видно из таблицы, у здоровых лиц интенсивность липопероксидации в плазме и эритроци-тах практически одинакова (ИР по всем показателям, кроме ОШ в изопропанольной фазе, колебался от 0,94 до 1,0). У больных с локальными ранами по сравнению со здоровыми лицами интенсив-ность липопероксидации в целом увеличивалась, что отражалось в повышении содержания про-дуктов ПОЛ как в плазме, так и в эритроцитах (Р < 0,001, Р < 0,01). Исключение составило лишь содержание ОШ в гептановой фазе плазмы и эритроцитов, которое не отличалось от контрольных значений. Одновременно у больных наблюдался выраженный дисбаланс относительно содержания продуктов пероксидации фосфолипидов в плазме и эритроцитах (Р < 0,001), тогда как различия в распределении нейтральных липопероксидов отмечены только для СТ (Р = 0,006). Снижение ИР ДКп/ДКэ и ОШп/ОШэ у больных по сравнению со здоровыми лицами происходило за счет увели-чения окисленности мембран эритроцитов (соответственно 0,89–0,94–0,98 и 0,52–0,7–0,82 у боль-ных и 0,98–0,98–0,99 и 0,75–0,8–1 у здоровых лиц, Р < 0,01). Индекс распределения СТп/СТэ, на-
Национальная
академия наук
Беларуси

72
против, увеличивался за счет значительного превалирования СТ в плазме по сравнению с эритро-цитами (0,9–1,35–1,58 у больных по сравнению с 0,98–0,98–0,99 у доноров, Р < 0,001).
Учитывая, что наиболее выраженные диспропорции между параметрами липопероксидации плазма/эритроциты были выявлены для СТ и ОШ в изопропанольной фазе, нами проанализиро-ваны особенности этих показателей в зависимости от срока давности раны, а также от характера течения послеоперационного периода у больных (наличие или отсутствие лизиса аутодермо-трансплантата).
По срокам давности ран пациенты были разделены на несколько групп на основании критери-ев, приведенных Ю. К. Абаевым и Б. А. Парамоновым [1, 8]. Первая группа (17 человек) – больные со свежими ранами, которым проводилась «ранняя» АДП (до 7 сут от момента получения травмы), вторая группа (56 человек) – пациенты с острыми ранами (от 7 сут до 2 мес. от момента получе- ния травмы) и третья группа (7 человек) – с хроническими ранами (от 2 мес. до 85 сут). Анализ с использованием однофакторного дисперсионного метода показал, что по всем изученным интер-медиатам ПОЛ индексы соотношения плазма/эритроциты значимо не различались между сравни-ваемыми группами. Так, в первой, второй и третьей группах ИР для СТ составил 1,2–1,35–1,5; 0,9–1,4–1,6 и 0,84–1,2–1,44; для ОШ – 0,41–0,72–0,82; 0,56–0,72–0,82 и 0,56–0,58–0,6 соответственно.
Как указывалось выше, у всех больных, вошедших в данное исследование, отмечалась клиниче-ская готовность раны к операции (отсутствие признаков воспаления, отсутствие выраженной экссу-дации, высокая адгезивность ран, а также наличие краевой эпителизации для острых и хронических ран). Поэтому им была проведена операция аутодермопластики, которая у 54 больных завершилась приживлением трансплантата на 7–9-е сутки (первая группа), а у 26 больных в эти же сроки произо-шел частичный или полный лизис лоскута (вторая группа). Сравниваемые группы не различались по половозрастному признаку, давности получения раны и ее этиологии. Сравнительный анализ пара-метров липопероксидации показал, что у пациентов первой группы уровень СТ в плазме в доопера-ционном периоде значительно превалировал над аналогичным показателем у больных второй груп-пы (ИР 1,26–1,47–1,6 и 0,83–0,87–1,4 соответственно, Р = 0,0003). Индексы распределения ОШп/ОШэ практически не различались (0,52–0,76–0,89 для первой группы, 0,52–0,66–0,76 для второй группы, Р = 0,09), и вне зависимости от исхода операции интенсивность накопления конечных продуктов ПОЛ в эритроцитах была всегда выше, чем в плазме.
Таким образом, дооперационные значения индекса распределения вторичных продуктов ПОЛ (сопряженные триены) между плазмой и эритроцитами могут иметь прогностическое зна-чение при оценке риска отторжения кожного аутотрансплантата у больных с локальными рана-ми. Можно предположить, что превалирование уровня СТ в плазме над аналогичным показателем в мембранах эритроцитов при интенсификации процессов липопероксидации в организме явля-ется отражением степени устойчивости мембраны эритроцитов к окислительному стрессу. Срыв механизмов такой устойчивости (в результате несостоятельности антиоксидантной защиты либо по причине нарушений функциональной резистентности эритроцитов) свидетельствует о недо-статке компенсаторно-адаптационных возможностей организма и может рассматриваться как признак неблагоприятного исхода патологического процесса.
Динамика показателей ПОЛ в течение послеоперационного периода у больных сравнивае-мых групп также различалась (рис. 1). У больных первой группы на 3–4-е сутки происходило равномерное снижение содержания СТ в изопропанольной фазе плазмы и эритроцитов, за счет чего ИР СТп/СТэ сохранялся на уровне дооперационных значений (1,3–1,4–1,53). На 7–9-е сутки наблюдалось снижение ИР по сравнению с его дооперационным уровнем (Р < 0,0001), значения которого приближались к показателям здоровых лиц. Соотношение ОШп/ОШэ практически нормализовалось (достигло значений, характерных для здоровых лиц) уже на 3–4-е сутки.
У больных с лизисом кожного аутотрансплантата (вторая группа) исходно сниженный ин-декс распределения СТп/СТэ продолжал снижаться (Р < 0,01) по сравнению с дооперационными значениями за счет преимущественного увеличения содержания СТ в эритроцитах. Кроме того, на фоне лизиса лоскута происходило дальнейшее увеличение концентрации ОШ в изопропа-нольной фазе как экстракта плазмы, так и эритроцитов, однако в плазме в несколько большей степени. В результате ИР ОШп/ОШэ увеличился по сравнению с дооперационными значе-ниями (Р < 0,001) (рис. 2).
Национальная
академия наук
Беларуси

73
Таким образом, полученные результаты показывают, что наиболее выраженные и кли-нически информативные изменения соотноше-ния показателей окисленности плазма/эритро-циты у обследованных больных наблюдались по содержанию СТ в изопропанольной фазе, которые, как известно, являются вторичными продуктами окисления фосфолипидов [3, 5].
Интересная закономерность выявлена нами при анализе взаимосвязей между по-казателями окисленности плазмы и мембран эритроцитов (по содержанию СТ и ОШ). В контрольной группе взаимосвязей между уровнем СТ фосфолипидов плазмы и СТ фосфолипидов эритроцитов не выявлено, но отмечалась сильная прямая связь между со-держанием ОШ в плазме и эритроцитах (r = 0,8; Р = 0,0003). На фоне активации свободнорадикального окисления у обследованных нами больных наличие взаимосвязи между содержанием СТ в фосфолипидах плазмы и эритроцитов зависело от значений ИР СТп/СТэ. При величине индекса СТп/Стэ > 1 (содержание сопряжен-ных триенов в плазме выше, чем в эритроцитах) корреляций не выявлено. При величине ин-декса СТп/Стэ < 1 (содержание сопряженных триенов в плазме ниже, чем в эритроцитах) по-являлась сильная прямая взаимосвязь между содержанием СТ в изопропанольной фазе плаз-мы и эритроцитов (r = 0,77; Р = 0,00008). Между содержанием ОШ в плазме и эритроцитах у обследованных больных выявлены корреляции, аналогичные таковым в контрольной группе (r = 0,7; Р = 0,0007).
Таким образом, в условиях активации процессов ПОЛ (в рассмотренном случае – при локальной гранулирующей ране) на-чинают проявляться различия в накопле-нии продуктов ПОЛ между плазмой и эри-троцитами, тогда как у здоровых лиц степень окисленности плазмы и эритроци-тов в целом одинаковая. В наибольшей сте-пени различия проявляются по показателям содержания вторичных и конечных продук-тов окисления фосфолипидов – СТ и ОШ. Эти данные указывают, что выбор биомате-риала и исследуемых параметров при изуче-нии процессов липопероксидации в организ-ме не должен быть случайным, а также объ-ясняют значительную противоречивость резу - льтатов, полученных разными авторами.
Выводы
У здоровых лиц в условиях отсутствия активации свободнорадикального окисления со-1. держание продуктов липопероксидации в плазме соответствует таковому в эритроцитах.
На фоне активации процессов липопероксидации (пациенты с локальными гранулирую-2. щими ранами) содержание вторичных продуктов окисления фосфолипидов (сопряженные трие-ны) превалирует в плазме крови, а конечных продуктов – в мембранах эритроцитов.
Дооперационные значения индекса распределения вторичных продуктов ПОЛ (сопряжен-3. ные триены) между плазмой и эритроцитами (сопряженные триены плазмы/ сопряженные триены
Рис. 1. Изменение ИР у пациентов первой группы в дина-. 1. Изменение ИР у пациентов первой группы в дина-Изменение ИР у пациентов первой группы в дина-мике послеоперационного периода. На рисунке приведены значения медианы. * – значимые различия в значениях ИР в послеоперационном периоде по сравнению с доопераци-
онным состоянием
Рис. 2. Изменение ИР у пациентов второй группы в динамике послеоперационного периода. На рисунке приведены значения медианы. * – значимые различия в значениях ИР в послеопера-ционном периоде по сравнению с дооперационным состоянием
Национальная
академия наук
Беларуси

эритроцитов) могут служить фактором прогноза риска отторжения аутодермотрансплантата у боль-ных с локальными ранами. У пациентов с хорошим исходом оперативного вмешательства значе-ния индекса соотношения сопряженных триенов в плазме и эритроцитах до операции достовер-но превышали аналогичные значения у больных с отторжением кожного лоскута.
В динамике приживления трансплантата показатели степени окисленности плазмы и эри-4. троцитов выравниваются, тогда как отторжение аутодермотрансплантата сопровождается преиму-щественным накоплением продуктов липопероксидации (сопряженные триены) в эритроцитах по сравнению с плазмой крови.
У здоровых лиц не выявлено взаимосвязи между уровнем сопряженных триенов в фосфо-5. липидах плазмы и аналогичным показателем в эритроцитах. На фоне активации свободно-радикального окисления наличие указанной связи зависит от значений индекса распределения сопряженных триенов между плазмой и эритроцитами: сильная прямая корреляция появляется при величине индекса менее 1.
Литература
А б а е в Ю. К. // Мед. новости. 2006. № 6. С. 34–40. 1. А н о ш и н а М. Ю. // Укр. мед. журн. 2006. Т. 56, № 6. С. 78–82.2. Б и л е н к о М. В. Ишемические и реперфузионные повреждения органов (молекулярные механизмы, пути 3.
предупреждения и лечения). М., 1989.В а с и л ь е в а Е. М. // Биомед. химия. 2005. Т. 51. Вып. 2. С. 118–126.4. В о л ч е г о р с к и й И. А. // Вопр. мед. химии. 1989. Т. 35, № 1. С. 127–135.5. Д у б и н и н а Е. Е. // Вопр. мед. химии. 2001. № 6. С. 561–581.6. Л ь в о в с к а я Е. И. // Вопр. мед. химии. 1991. Т. 37, № 4. С. 93.7. П а р а м о н о в Б. А. Ожоги: руководство для врачей. СПб., 2000.8. П л а т о н о в А. Е. Статистический анализ в медицине и биологии: задачи, терминология, логика, компью-9.
терные методы. М., 2000.Р я б и н и н В. Е., Л и ф ш и ц Р. И. // Вопр. мед. химии. 1990. № 1. С. 7–13.10. С т о р о ж у к П. Г. // Вестн. интенсив. терапии. 1998. № 4. С. 17–21.11. Ш а н и н Ю. И., Ш а н и н В. Ю., З и н о в ь е в Е. В. Антиоксидантная терапия в клинической практике 12.
(теоретическое обоснование и стратегия проведения). СПб., 2003.A r u o m a O. I. // J. Amer. Oil. Chem. Soc. 1998. Vol. 75. P. 199–212.13. C r o s s M. D. // Ann. Intern. Med. 1987. Vol. 107. Р. 526–545.14. H i r o t o �., S h i m a o S., S h i m i z u �. // Burns. 1988. Vol. 14, N 4. Р. 313–319.15. N i s h i g a k i J., H a g i h a r a M., H i r m a t s u M. et al. // Biochem. Med. 1980. Vol. 24. P. 185–189.16. V i v e k a n a n l a J., � i n A., C o a l s o n J. J., K i n g R. J. // J. Biol. Chem. 1994. Vol. 269, N 40. Р. 25057–17.
25061.Z a u g g R. H., W a l d e r J. A., K l o t z I. M. // The J. of Biol. Chem. 1977. Vol. 252, N 23. P. 8542–8548. 18.
i. a. noviKova, y. i. yaRETS
CORRELATION BETWEEN THE OXIDATION DEGREE OF PLASMA AND ERYTHROCYTES IN THE CONDITIONS OF FREE RADICAL OXIDATION ACTIVATION
Gomel State Medical university, Belarus
Summary
We studied the peculiarities of lipid peroxidation products distribution between plasma and erythrocytes in the normal conditions and in the conditions of free radical oxidation activation by the local granulated wound example. Significant differences in the lipoperoxide accumulation between plasma and erythrocytes were found statistically when lipid peroxidation was intensified while in the healthy individuals it was identical. Most of the significant differences were established for the content of the intermediate and end products of phospholipids peroxidation-trien conjugates and Schiff bases. We analyzed the correlations between the indices of plasma and erythrocytes oxidation and its course of the patients with different post-procedural period outcome.
Национальная
академия наук
Беларуси

75
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.24-018-02:613.84:547.914.3/.5
Е. А. ДЕвинА, Т. Ю. пРинЬКовА, А. Д. ТАГАнович
ОСОБЕННОСТИ ИЗМЕНЕНИЯ ФУНКЦИОНАЛЬНОй АКТИВНОСТИ АЛЬВЕОЛЯРНЫХ МАКРОФАГОВ ПРИ КОНТАКТЕ С СИГАРЕТНЫМ ДЫМОМ
Белорусский государственный медицинский университет, Минск(поступила в редакцию 01.10.2009)
В Беларуси от болезней, связанных с курением, ежегодно умирают более 15 тыс. человек. От воздействия табачного дыма страдает в первую очередь бронхо-легочная система, поэтому куре-ние является одной из наиболее важных причин развития таких хронических неспецифических болезней легких, как хронический бронхит, эмфизема легких, бронхиальная астма, хроническая обструктивная болезнь легких [1].
Среди активных факторов неспецифической защиты бронхо-легочной системы важное место занимают высокодифференцированные клетки легких – альвеолярные макрофаги (АМ). Способ-ность АМ поглощать чужеродный материал тесно связана с их жизнеспособностью и является одним из показателей их функционального состояния.
Находясь на поверхности воздухопроводящих путей, АМ продуцируют оксид азота (NO). Он образуется под действием фермента – NO-синтазы (NOS) в результате окисления �-аргинина. В АМ показано существование двух типов NOS, при этом базальный уровень синтеза NO обе-NOS, при этом базальный уровень синтеза NO обе-, при этом базальный уровень синтеза NO обе-NO обе- обе-спечивается эндотелиальной изоформой – eNOS, а увеличение продукции оксида азота после экспозиции к воспалительным стимулам обеспечивает индуцибельная изоформа (iNOS). NO об-iNOS). NO об-). NO об-NO об- об-ладает микробицидным действием не только на внеклеточные, но и на внутриклеточные пато-генные агенты, действуя посредством образования высокореакционных форм, например перок-синитрита (ONOO–) или радикалов диоксида азота (NO2
•) [2]. Кроме того, он является уникаль-ным медиатором межклеточных взаимодействий [3]. Исходя из этого, предполагается, что изменения метаболизма и функциональной активности АМ в условиях воздействия сигаретного дыма лежат в основе патогенеза хронического воспаления легочной ткани [4].
В литературе широко обсуждается влияние сигаретного дыма на функции клеток легких, в том числе на функции АМ [5–7]. Однако до настоящего времени не определена динамика взаи-модействия сигаретного дыма с макрофагами легких, отсутствует оценка жизнеспособности этих клеток в целом в данных условиях. Нет данных и об изменении метаболитов NO, продуци-NO, продуци-, продуци-руемого АМ.
Целью данного исследования явилось изучение влияния экстракта сигаретного дыма (ЭСД) на жизнеспособность альвеолярных макрофагов, их фагоцитарную активность и способность к генерации оксида азота.
Материалы и методы исследования. Эксперимент был проведен на 20 беспородных крысах-самцах массой 180–200 г, находившихся на стандартном рационе вивария БГМУ. АМ получали из бронхоальвеолярной лаважной жидкости. Клеточную суспензию центрифугировали (900 об/мин, 10 мин, +4 °С). Полученный осадок ресуспендировали в модифицированной Дульбекко культу-ральной среде Игла (DME) (Sigma, США), содержавшей 10% эмбриональной телячьей сыворот-DME) (Sigma, США), содержавшей 10% эмбриональной телячьей сыворот-) (Sigma, США), содержавшей 10% эмбриональной телячьей сыворот-Sigma, США), содержавшей 10% эмбриональной телячьей сыворот-, США), содержавшей 10% эмбриональной телячьей сыворот-ки, 2 мМ глутамина (Sigma). Общее количество клеток подсчитывали в камере Горяева. Клеточ-Sigma). Общее количество клеток подсчитывали в камере Горяева. Клеточ-). Общее количество клеток подсчитывали в камере Горяева. Клеточ-ную суспензию высевали на пластиковые чашки Петри диаметром 3,5 см (концентрация 2⋅106 макрофагов на чашку) и помещали на 60 мин в СО2-инкубатор при температуре 37 °C, увлаж-, увлаж-ненной атмосфере и 5% CO2.
Национальная
академия наук
Беларуси

76
ЭСД (2,1 г/л) был приготовлен посредством пропускания дыма от трех сигарет («Корона», РБ; содержание смол в 1 сигарете – 14 мг) со скоростью 1 сигарета/мин через 20 мл культураль-ной среды DМЕ с использованием вакуумного насоса. Концентрации ЭСД 1,4 и 0,7 г/л получали путем разведения исходного экстракта. ЭСД-среду стерилизовали с помощью бактериального фильтра (Sigma) с диаметром пор 0,22 мкм, рН 7,4. ЭСД стандартизировали, измеряя оптическую плотность (ОD 0,74±0,05) при λ= 320 нм, рН 7,4.
АМ, адгезированные к пластиковой поверхности, инкубировали в течение 1 и 24 ч с ЭСД, со-державшим 0,7; 1,4 и 2,1 г/л смол. По истечении данного времени инкубационную среду аккурат-но удаляли, а к клеткам добавляли изотонический раствор NaCl. АМ соскребали с поверхности чашек Петри скрепером (Costar, CША), после чего клеточную суспензию вручную разрушали методом гомогенизации.
По истечении срока инкубации определяли способность макрофагов поглощать Staphylococcus aureus. Определяли фагоцитарное число (ФЧ) – среднее число микроорганизмов, поглощенных одним активным АМ, и фагоцитарный показатель (ФП) – процентное содержание фагоцитирую-щих клеток от общего количества макрофагов.
Жизнеспособность АМ определяли по окраске клеток трипановым синим (Sigma, США). Подсчет клеток проводили с помощью инвертированного микроскопа («ЛОМО», Россия). По числу АМ, не воспринявших окраску из 100 макрофагов, рассчитывали процентное содержание жизнеспособных клеток.
Активность лактатдегидрогеназы (ЛДГ) (КФ 1.1.1.27) в АМ и среде культивирования опреде-ляли кинетическим методом по скорости уменьшения концентрации НАДН⋅Н+ в реакции, ката-лизируемой ЛДГ, что регистрируется спектрофотометрически.
Генерацию NO оценивали путем определения концентрации нитритов в среде инкубации и в АМ, с помощью реактива Грисса – спектрофотометрически [8].
Статистическую обработку проводили с использованием пакета программ Statistica 6.0. Все результаты представлены в виде медиан и интерквартильных размахов. Статистическая значи-мость полученных результатов была оценена при помощи U-теста Манна–Уитни для непараме-U-теста Манна–Уитни для непараме--теста Манна–Уитни для непараме-трических выборок. Различия считали значимыми при P < 0,05.
Результаты и их обсуждение. Через 1 ч инкубации доля жизнеспособных АМ в контроль-ных пробах составила 98%. Спустя такой же период времени, когда клетки инкубировали в при-сутствии ЭСД, их доля уменьшилась (рис. 1): при концентрации смол 0,7 г/л она составила 92%, при 1,4 г/л – 86 и при 2,1 г/л – 83% (P < 0,05).
Через 24 ч инкубации степень снижения жизнеспособности АМ под влиянием смол сигарет-ного дыма была интенсивнее: доля жизнеспособных АМ при концентрации 0,7 г/л составила 75%, при 1,4 г/л – 71, при 2,1 г/л – 65% (в контрольных пробах – в среднем 93%).
Рис. 1. Влияние ЭСД на жизнеспособность АМ и высвобождение ЛДГ в среду инкубации (%). Продолжительность инкубации – 1 ч. Достоверность различий (Р < 0,05): * – по сравнению с контролем; ** – по сравнению с ЭСД-средой
(0,7 г/л) (n = 6)
Национальная
академия наук
Беларуси

77
В присутствии смол сигаретного дыма в среде инкубации АМ активность ЛДГ увеличилась, при этом наблюдалось снижение ее активности в клетках, в среднем на 20%, по сравнению с контролем (рис. 1, 2). При повышении концентрации смол от 0,7 до 1,4 и 2,1 г/л уровень высво-бождения ЛДГ из АМ через 1 ч возрос до 8,0; 14,1 и 17,4% соответственно. При экспозиции АМ в ЭСД-среде в течение 24 ч активность ЛДГ в среде инкубации еще больше увеличилась. В чаш-ках, в которых присутствовал ЭСД с концентрацией смол 2,1 г/л, она увеличилась в наибольшей степени – в 2,5 раза по сравнению с контрольным уровнем. В клетках активность ЛДГ снизилась на 42,6; 48,0 и 55,0% по сравнению с контролем, соответственно концентрации смол (0,7; 1,4 и 2,1 г/л) в ЭСД-среде.
Выявлена корреляционная зависимость между долей жизнеспособных клеток и активностью ЛДГ в культуральной среде (коэффициент корреляции r, подсчитанный по Спирмену, составил –0,9; P < 0,005).
ЭСД подавлял фагоцитарную активность АМ уже при минимальной концентрации смол си-гаретного дыма в культуральной среде. Увеличение концентрации смол в ЭСД до 1,4 г/л, а затем до 2,1 г/л приводило к достоверному снижению ФП – на 22,3 и 29,7% соответственно (см. таблицу). При инкубации АМ в ЭСД-среде в течение 24 ч ФП по сравнению с контролем уменьшился на 7,4; 29,7 и 40,8% соответственно.
Влияние экстракта сигаретного дыма (ЭСД) на фагоцитарную активность альвеолярных макрофагов и концентрацию нитрит-ионов
Показатель КонтрольЭСД, г/л
0,7 1,4 2,1
продолжительность инкубации 1 чФагоцитарный показатель 31,0 (30,0–32,0) 27,0 * (25,0–29,0) 21,0** (19,0–23,0) 19** (18,0–21,0)Фагоцитарное число 3,7 (3,3–4,2) 3,5 (3,3–3,7) 3,1* (2,7–3,5) 3,0* (2,6–3,3)Внутриклеточный уровень нитрит-ионов, нМ/106 кл. 2,1 (1,7–2,3) 1,9 (1,6–1,9) 1,6 (1,6–1,7) 1,1* (1,0–1,2)Концентрация нитрит-ионов в среде инкубации, мкМ/л – 7,8* (6,9–8,5) 12,0* (11,0–13,2) 18,1* (16,8–22,0)Концентрация нитрит-ионов в среде инкубации (с клетками), мкМ/л 1,6 (1,1–2,4) 5,3∆ (4,0–6,4) 7,4∆∆ (5,6–9,1) 8,6*∆ (6,5–10,7)
продолжительность инкубации 24 ч
Фагоцитарный показатель 31,0 (30,0–32,0) 25* (21,0–28,0) 19,0** (18,0–21,0) 16** (14,0–19,0)Фагоцитарное число 3,7 (3,5–4,0) 3,1* (2,8–3,3) 3,1* (2,8–3,5) 3,1* (2,8–3,4)
Рис. 2. Влияние ЭСД на жизнеспособность АМ и высвобождение ЛДГ в среду инкубации (%). Продолжительность инкубации – 24 ч. Достоверность различий (Р < 0,05): * – по сравнению с контролем; ** – по сравнению с ЭСД-
средой (0,7 г/л) (n = 6)
Национальная
академия наук
Беларуси

78
Показатель КонтрольЭСД, г/л
0,7 1,4 2,1
Внутриклеточный уровень нитрит-ионов, нМ/106 кл. 4,4 (4,3–4,5) 1,8* (1,6–2,0) 2,7* (2,6–2,8) 2,9* (2,7–3,1)Концентрация нитрит-ионов в среде инкубации, мкМ/л – 17,3 (16,8–18,2) 18,6 (17,7–22,0) 21,6 (19,8–23,1)Концентрация нитрит-ионов в среде инкубации (с клетками), мкМ/л 5,1 (4,5–5,3) 8,7∆ (6,8–10,5) 6,8∆∆ (5,2–8,3) 7,4*∆ (5,6–9,1)
П р и м е ч а н и е. Данные представлены как медиана и 50%-ный интерквартильный размах между 25-м и 75-м процентилями. Достоверность различий (Р < 0,05): * – по сравнению с контролем; ** – по сравнению с ЭСД-средой (0,7 г/л); ∆, ∆∆, *∆ – по сравнению с ЭСД-средой без клеток (дозы 0,7; 1,4 и 2,1 г/л соответственно) (n = 6).
Результаты экспериментов показали, что при всех изучаемых концентрациях ЭСД и незави-симо от времени экспозиции ФЧ также уменьшается – в среднем на 16,3% в сравнении с контро-лем (см. таблицу).
В культуральной среде, содержавшей ЭСД, в которой не инкубировались клетки, концентра-ция нитрит-ионов увеличивалась (см. таблицу). Причем с ростом концентрации смол сигаретно-го дыма возрастал и уровень нитрит-ионов. К примеру, при концентрации смол 0,7 г/л медиана его составила 7,8 мкМ/л, а при концентрации смол 2,1 г/л она была в 2,5 раза больше – 18,1 мкМ/л. Через 24 ч инкубации культуральной среды (без клеток) с ЭСД образование нитрит-ионов еще больше увеличилось – до 21,6 мкМ/л, если в среде присутствовал ЭСД с концентрацией смол 2,1 г/л (в контрольных пробах (без добавления ЭСД) нитрит-ионы отсутствовали).
Через 1 ч инкубации АМ с ЭСД наблюдалось снижение концентрации нитрит-ионов в куль-туральной среде по сравнению с бесклеточным уровнем: на 32% – при концентрации смол 0,7 г/л, на 38% – при 1,4 г/л и на 52%, если концентрация смол в ЭСД составляла 2,1 г/л.
Содержание нитрит-ионов через 1 ч инкубации в самих АМ в контрольных пробах было наи-большим (2,1 нМ/106 клеток). Присутствие в среде инкубации ЭСД приводило к снижению этого показателя, при концентрации смол 2,1 г/л он был наименьшим (1,1 нМ/106 клеток) и статистиче-ски достоверно отличался от контрольного уровня.
Через 24 ч инкубации АМ с ЭСД концентрация нитрит-ионов в клетках также была суще-ственно ниже, чем в контроле, причем наименьшей она была при инкубации клеток с ЭСД с кон-центрацией смол 0,7 г/л. С увеличением концентрации смол отмечалась тенденция к снижению уровня нитрит-ионов (см. таблицу).
Показано, что основным компонентом табачного дыма, определяющим его токсичность, яв-ляется смола, или твердая фаза [9]. Поэтому выбор концентрации смол в табачном дыме в каче-стве критерия потенциальной токсичности представлялся вполне обоснованным. Использование в экспериментах ЭСД с различным содержанием смол и разной длительностью их воздействия на АМ позволило определить не только направленность внутриклеточных изменений показате-лей функциональной активности и метаболизма, но и ее динамику.
Установленное нами снижение уровня жизнеспособных клеток под влиянием смол сигаретно-го дыма подтверждается данными литературы. В экспериментах in vitro и in vivo ряд исследовате-лей отмечали индуцированный воздействием сигаретного дыма апоптоз АМ крыс и моноцитов крови человека [10]. J. Wickenden и сотр. [11] описали вызванный конденсатом сигаретного дыма некроз в альвеолярных эпителиальных клетках и эндотелиальных клетках пупочной вены.
Результаты проведенного нами исследования продемонстрировали, что снижение жизнеспособ-ности АМ и активности ЛДГ в АМ и сопряженный с этим рост активности ЛДГ в среде инкубации напрямую зависят от концентрации смол и длительности их контакта с клетками. ЛДГ определяется главным образом в цитозоле клеток, поэтому увеличение ее активности в среде инкубации свиде-тельствует о прогрессирующих нарушениях целостности мембранных структур и о гибели клеток.
Очевидно, что уменьшение жизнеспособности АМ под влиянием смол сигаретного дыма не может не сказаться на их функциональной активности. Полученные в ходе исследования
продолжение таблицыНациональная
академия наук
Беларуси

79
данные показывают, что доля фагоцитирующих макрофагов уменьшается по мере нарастания концентрации смол в среде инкубации АМ. Причем наиболее выраженное изменение, пример-но в 2 раза, имеет место при самой высокой из изучаемых концентраций смолы (2,1 г/л) и при наиболее продолжительном (24 ч) контакте с ней АМ. Несколько по-иному изменяется погло-тительная эффективность АМ, о чем свидетельствует ФЧ. Спустя 24 ч инкубации клеток с ЭСД количество поглощенных St. аureus уменьшается, при этом степень уменьшения не за-висит от концентрации смолы.
Снижение интенсивности фагоцитоза под влиянием смол сигаретного дыма отмечалось ра-нее и другими исследователями [12].
Важнейшим регулятором фагоцитарной активности АМ является NO•. В литературе имеют-ся противоречивые данные об уровне его продукции клетками легких при экспозиции их сига-ретным дымом [6, 8]. При этом не оценивалась динамика изменения метаболизма NO.
Полученные данные убедительно показывают наличие в смоле сигаретного дыма значитель-ного количества NO, которое существенно превышает его эндогенный уровень в АМ. В этих условиях контакт ЭСД с клетками приводит к снижению концентрации нитрит-ионов в среде инкубации. Об этом свидетельствуют результаты сравнения уровня этого показателя в среде инкубации без клеток и с клетками, хотя в среде, в которой АМ инкубировались при большей концентрации смолы, уровень нитрит-ионов выше, чем в среде, в которой они инкубировались при меньшей концентрации смолы. Характерно, что в результате контакта со смолами сигарет-ного дыма снижение концентрации нитрит-ионов в самих АМ напрямую зависело от концентра-ции смолы в среде инкубации.
Однако такая закономерность имела место только при относительно коротком периоде инку-бации АМ с сигаретным дымом. При продолжительной экспозиции клеток с ЭСД значительное снижение концентрации нитрит-ионов также имело место, но в основном при минимальной кон-центрации смолы. Возможно, последнее обстоятельство было обусловлено проникновением NO или его конечного метаболита – нитрит-иона обратно из среды инкубации в клетки вслед- или его конечного метаболита – нитрит-иона обратно из среды инкубации в клетки вслед-ствие нарушения проницаемости последних. Аргументом в пользу такого предположения явля-ется описанная выше зависимость жизнеспособности АМ от контакта с ЭСД.
Есть основание полагать, что причиной снижения уровня нитрит-ионов в АМ под влиянием ЭСД является угнетение активности NO-синтазы. Показано, что инкубация эндотелиальных клеток легочной артерии с ЭСД в течение 24 ч снижает активность этого фермента, причем угне-тение носит дозозависимый характер [13]. Авторы предположили, что снижение активности фермента обусловлено прямым ингибирующим действием высокой концентрации NO в сигарет-NO в сигарет- в сигарет-ном дыме на активный центр NO-синтазы, содержащий гемовое железо [14]. Однако это не объ-NO-синтазы, содержащий гемовое железо [14]. Однако это не объ--синтазы, содержащий гемовое железо [14]. Однако это не объ-ясняет, почему в присутствии АМ концентрация нитрит-ионов, поступающих со смолами сига-ретного дыма, эффективно уменьшается по сравнению с первоначальным уровнем (бесклеточная ЭСД-среда), что демонстрируют полученные нами результаты.
Заключение. Таким образом, результаты исследования позволили установить некоторые за-кономерности изменения жизнеспособности, фагоцитарной активности АМ и генерации NO• в этих клетках под влиянием смол сигаретного дыма, которые можно сформулировать следую-щим образом:
1. При инкубации с ЭСД снижается жизнеспособность изолированных АМ и нарушается структурная целостность их мембран.
2. Контакт с ЭСД снижает поглотительную способность АМ как за счет количества клеток, осуществляющих фагоцитоз, так и за счет эффективности поглощения чужеродного материала фагоцитирующими клетками.
3. Под влиянием сигаретного дыма изменяется метаболизм оксида азота в альвеолярных ма-крофагах, снижается внутриклеточный уровень нитрит-ионов.
4. Обнаруженные изменения становятся более выраженными при увеличении концентрации смолы в ЭСД и длительности контакта с ней АМ.
Работа выполнена при финансовой поддержке Фонда фундаментальных исследований НАН Беларуси.
Национальная
академия наук
Беларуси

Литература
1. D e l c l a u x C. // Eur. Respir. J. 2003. Vol. 22. P. 10–14.2. Д м и т р е н к о Н. П. // Укр. биохим. журн. 2005. Т. 77, № 5. С. 5–22.3. С е п и а ш в и л и Р. И., Ш у б и ч М. Г. // Астма. 2001. Т. 2, № 2. С. 5–14.4. E i s n e r M. D., B a l m e s J. // Environ. Health Perspect. J. 2005. Vol. 4. Р. 715–730.5. G r e e n G., C a r o l i n D. // Engl. J. Med. 1987. Vol. 276. P. 421–427.6. A o s h i b a К. // Am. J. Physiol. �ung Cell Mol. Physiol. 2001. Vol. 281. Р. 1392–1401. 7. W i l h i t e C. // J. Carcinogenesis. 2001. Vol. 22, N 8. P. 1173–1178.8. Г о л и к о в П. П., Н и к о л а е в а Н. Ю. // Клин. лаб. диагностика. 2004. № 1. С. 13–15.9. Z h a n g W., V e n a r d o s K. // Hypertension. 2006. Vol. 48. Р. 278–285.10. R a n g a s a m y T., M i s r a V. // Am. J. Physiol. �ung Cell Mol. Physiol. 2009. Vol. 296 (6). Р. 888–900.11. W i c k e n d e n J. // Am. J. of Resp. Cell and Mol. Biol. 2003. Vol. 29. Р. 562–570.12. I n d e n M., W i e s l a n d e r E. // Eur. Respir. J. 1988. Vol. 1 P. 645–650.13. � u n c h a o S u, W e i h o n g H a n // Am. J. Respir. Cell Mol. Biol. 1998. Vol. 19, N 5. Р. 819–825.14. C h a m b e r s D. // Thorax. 1998. Vol. 53. P. 677–679.
E. a. dEvina, T. y. pRinKova, a. d. Tahanovich
PECULIARITIES OF CHANGE IN THE FUNCTIONAL ACTIVITY OF ALVEOLAR MACROPHAGES EXPOSED TO CIGARETTE SMOKE
Belarusian State Medical university, Minsk
Summary
We studied the effect of exposure to cigarette smoke extract (CSE) on the viabity, phagocytosis of St. aureus and NO-generation by alveolar macrophages (AM). AM were isolated from bronchoalveolar lavage fluid of rats. After the incubation with CSE for 1 and 24 h, AM showed a decreased viability and capacity to phagocytosis in dose-dependent manner compared to AM which were incubated without CSE. The inhibition of the phagocytic index of the AM and also the quantity of St. au-reus absorbed by these cells by phagocytosis was observed. CSE influenced the NO-production of AM. We found a significant decrease of the nitrite concentration in AM which were subjected to CSE and in the medium where the cells were incubated.
Национальная
академия наук
Беларуси

81
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 612.57:612.015.1:612.828
в. н. БочАРовА , Е. н. САвчинА
ГИСТОХИМИЧЕСКИй АНАЛИЗ АКТИВНОСТИ NADPH-ДИАФОРАЗЫ, АЦЕТИЛХОЛИНЭСТЕРАЗЫ И ФЕРМЕНТОВ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
В СТРУКТУРАХ ПРОДОЛГОВАТОГО МОЗГА КРЫСЫ ПРИ ЛИХОРАДКЕ
институт физиологии нАн Беларуси, Минск
(поступила в редакцию 08.02.2010)
Введение. Известно, что лихорадка является реакцией организма на воздействие различных патогенов. Эта чрезвычайная реакция характеризуется не только нарушением процессов тепло-обмена и значительным повышением температуры тела, но и структурно-функциональными из-менениями деятельности нервных центров и функциональных систем [1]. К настоящему времени существуют представления о сопряженном функционировании системы терморегуляции и им-мунной системы [1–3]. Вместе с тем пути и механизмы передачи информации от клеток иммун-ной системы в ЦНС по-прежнему изучены недостаточно. Установлено, что эндогенные пироге-ны, образующиеся при введении липополисахарида (ЛПС), обладают нейротропным действием [1, 3], что, как предполагается, может быть обусловлено действием цитокинов, высвобождаемых на периферии, на чувствительные окончания нервов, которые иннервируют брюшную полость [2]. Одним из источников поступления висцеральных импульсов от рецепторов внутренних ор-ганов в ЦНС являются афферентные волокна блуждающих нервов, которые оканчиваются в ядре солитарного тракта (ЯСТ). Это ядро рассматривается как афферентное в системе многочислен-ных висцеральных рефлексов, а также как первое передаточное звено на пути висцеральных им-пульсов к вышележащим структурам мозга. Ранее показано, что ЛПС E. сoli при внутрибрю-шинном введении в дозе 5 мкг/кг вызывает усиление афферентной импульсации в блуждающих нервах [4], а также усиление афферентной и эфферентной активности в шейных отделах блуж-дающих нервов и мультинейронной активности в ЯСТ и дорсальном моторном ядре этих нервов [5]. Параллельно с этим повышаются активность ферментов энергетического обмена и интенсив-ность синтеза монооксида азота (NO) и гидролиза ацетилхолина в нейронах ЯСТ крысы [4]. Предполагается, что передача сигналов от энтероцитов к нервным окончаниям при системном действии ЛПС является нехолинергической и неадренергической, а ее медиатором может быть NO [6]. Именно эта сигнальная молекула наряду с цитокинами может быть связующим звеном между интегральными системами организма. Данные литературы также указывают на особую роль холинергических механизмов в поддержании температурного гомеостазиса [7]. Однако меха-низмы, которые вовлекаются в развитие лихорадочной реакции, до сих пор детально не изучены.
Возможная роль NO и ацетилхолина в дорсальном моторном ядре блуждающего нерва (ДМЯБН) при действии в организме пирогенных веществ изучена недостаточно.
Цель настоящего исследования – определить, какую роль играют NO- и холинергические структуры ДМЯБН (по гистохимическим показателям) в поддержании температурного гомео-стазиса при эндотоксиновой лихорадке.
Материалы и методы исследования. Исследование выполнено на наркотизированных эфиром 25 крысах-самцах массой 250–330 г. Животных содержали в стационарных условиях вивария при температуре 22,0 ± 1,0 °С, они имели свободный доступ к воде и пище. Экспериментальную лихорадку вызывали путем внутрибрюшинного введения ЛПС E. coli в дозе 5 мкг/кг, который
Национальная
академия наук
Беларуси

82
растворяли в 1 мл апирогенного физиологического раствора (АФР) непосредственно перед инъек-цией. Животным контрольной группы вводили АФР в таком же объеме. Ректальную температуру измеряли с помощью трехканального электронного термометра (точность которого составляла ± 0,05 °С) с миниатюрными терморезисторами СТЗ-14, сигнал с которых подавался на цифровой вольтметр B7–38. Датчик вводили в прямую кишку на глубину 5 см. Объектом исследования служило ДМЯБН в продолговатом мозге. Для цитохимических исследований нейронов ДМЯБН головной мозг полностью извлекали и выделяли область в продолговатом мозге, где расположе-но данное ядро [8]. Для идентификации NO-синтезирующих нейронов в ДМЯБН использовали методику определения активности фермента NADPH-диафоразы (NADPH-d) [9]. По данным ряда авторов [10, 11], гистохимический метод определения NADPH-d является маркером конститу-NADPH-d является маркером конститу--d является маркером конститу-d является маркером конститу- является маркером конститу-тивной (нейрональной и эндотелиональной) NO-синтазы. Для этой цели кусочки продолгова-NO-синтазы. Для этой цели кусочки продолгова--синтазы. Для этой цели кусочки продолгова-того мозга фиксировали в 4%-ном растворе параформальдегида, приготовленном на 0,1 М фос-фатном буфере (рН 7,2) в течение 12 ч при температуре 4 °С. Установлено, что при фиксации с использованием параформальдегида инактивируются все NADPH-зависимые ферменты-окислители, за исключением NO-синтазы [12, 13]. Затем материал несколько раз промывали в 15%-ном растворе сахарозы в течение 20 ч при той же температуре, готовили криостатные сре-зы толщиной 15 мкм и монтировали на предметные стекла. Препараты инкубировали 1 ч в сре-де, содержавшей 0,5 мМ β-NADPH (Sigma, США), 0,5 мМ нитросинего тетразолиевого и 0,3% тритона Х-100 в 0,15 М Трис-НCl буфере, рН 8,0, в термостате при температуре 37 °С. Затем их промывали в дистиллированной воде, заключали в глицерин и накрывали покровными стекла-ми. Контрольные образцы, инкубированные в среде без добавления β-NADPH, не давали поло-NADPH, не давали поло-, не давали поло-жительной реакции на NADPH-d. Ингибитор синтеза NO метиловый эфир N-нитро-�-аргинин (�-NAME) (Sigma) растворяли в АФР в дозе 25 мг/кг и вводили внутрибрюшинно за 30 мин до инъекции ЛПС (5 мкг/кг). Для определения активности фермента ацетилхолинэстеразы (АХЭ) при гидролизе ацетилхолина использовали методику El-Вadawi, Shenk (1967). Активность фер-El-Вadawi, Shenk (1967). Активность фер--Вadawi, Shenk (1967). Активность фер-adawi, Shenk (1967). Активность фер-, Shenk (1967). Активность фер-ментов энергетического обмена сукцинат- и лактатдегидрогеназы (СДГ, ЛДГ) определяли на се-рийных срезах тетразолиевым методом по Пирсу (1962). Об активности ферментов судили по плотности образующихся осадков: формазана – для NADPH-d, СДГ и ЛДГ, ферроцианида ме- ди – для АХЭ. Оптическую плотность цитоплазмы нейронов, содержавшей в себе эти осадки, измеряли с помощью микроскопа МРV-2 (фирмы �eitz, Германия), снабженного цифровой каме-V-2 (фирмы �eitz, Германия), снабженного цифровой каме--2 (фирмы �eitz, Германия), снабженного цифровой каме-�eitz, Германия), снабженного цифровой каме-, Германия), снабженного цифровой каме-рой, а также с использованием специальных светофильтров (С0 и С4) и предназначенной для дан-ных целей компьютерной программы Sсion for Windows. Результаты опытов обрабатывали стати-Sсion for Windows. Результаты опытов обрабатывали стати-сion for Windows. Результаты опытов обрабатывали стати-ion for Windows. Результаты опытов обрабатывали стати- for Windows. Результаты опытов обрабатывали стати-for Windows. Результаты опытов обрабатывали стати- Windows. Результаты опытов обрабатывали стати-Windows. Результаты опытов обрабатывали стати-. Результаты опытов обрабатывали стати-стически с применением критерия Стьюдента и однофакторного дисперсионного анализа ANOVA. Морфологические изменения в нейронах изучали методом Ниссля в модификации Шабадаша.
Результаты и их обсуждение. Цитохимический анализ срезов ДМЯБН продолговатого моз-га животных контрольной группы показал, что в данном ядре активность NADPH-d выявляется в нейронах и эндотелиоцитах кровеносных сосудов. Количество NO-позитивных нейронов со-NO-позитивных нейронов со--позитивных нейронов со-ставляло 25,4±1,8%. Продукт гистохимической реакции, определявшийся в цитоплазме таких нейронов, имел цвет от бледно-голубого до темно-фиолетового. В большинстве нейронов ядра оставались неокрашенными. Считается, что плотность образующегося осадка диформазана пря-мо пропорциональна молекулярному содержанию NO-синтазы [9], что дает возможность судить об активности энзима. Реакция на NADPH-d позволяет идентифицировать строение нейронов за счет выявления их тел и отростков. Форма и размер NADPH-d-позитивных клеток в ДМЯБН были весьма разнообразными. Они имели веретеновидные, треугольные, звездчатые и округ-лые сомы с радиально разветвленными отростками. В зависимости от количества отходящих отростков нейроны могут быть классифицированы как уни-, би- и мультиполярные (рис. 1, А). В данном ядре эти клетки располагались диффузно, а их темноокрашенные нейриты разветвля-лись и вступали в сложные взаимоотношения с другими нейронами и кровеносными сосудами как внутри ядра, так и на достаточно большом расстоянии от него. Активность NADPH-d была определена и в сосудистом эндотелии. Однако интенсивность окраски в нем была ниже, чем в нейроцитах. Полагают, что NO, синтезируемый эндотелиальными клетками кровеносных со-NO, синтезируемый эндотелиальными клетками кровеносных со-, синтезируемый эндотелиальными клетками кровеносных со-судов, является ключевым фактором релаксации сосудов [11].
Национальная
академия наук
Беларуси

83
Результаты изучения активности АХЭ показали, что в составе ДМЯБН имеются нейроны с различной степе-нью активности данного фермента (высокой, средней или низкой). Морфологическим критерием ферментной активности являлось наличие плотного темно-корич-невого осадка гистохимической реакции сульфида меди, заполнявшего всю цитоплазму нейронов. Счи-тается, что такого типа нейроны являются холинерги-ческими, поскольку в них с помощью иммуногисто-химических и радиохимических методов исследова-ния была выявлена высокая активность фермента ацетилтрансферазы [14, 15].
Известно, что нейронная активность связана с кле-точным метаболизмом, в котором участвуют окис-лительно-восстановительные ферменты. В контроле нейроны ДМЯБН различались по уровню исследованных ферментов. Средние значения показателей активности ЛДГ были выше, чем СДГ. Это может свидетельство-вать о том, что в ДМЯБН преобладает гликолитиче-ский путь превращения углеводов.
Цитофотометрический анализ срезов показал, что через 120 мин после внутрибрюшинного введения ЛПС E. сoli в дозе 5 мкг/кг и подъема ректальной температу-ры на 0,9–1,2 ºС выше исходной активность NADPH-d в нейронах ДМЯБН повышается на 51,24 ± 5,4% (Р < 0,01) по сравнению с показателями у интактных животных (рис. 2, А). Перикарионы более многочисленных нейро-нов с высокой активностью данного фермента были за-полнены сплошным темно-синим осадком. В некото-рых подобных нейроцитах контур ядра не определялся или определялся недостаточно отчетливо (рис. 1, Б). Интенсивность цитохимической окраски, соответствующей уровню активности NADPH-d, усиливалась также в терминалях отростков нервных клеток (рис. 1, Б). Реакция ана-логичного характера отмечалась и в сосудистом эндотелии. Активность АХЭ в нейронах ДМЯБН
Рис. 1. Активность NADPH-d в нейронах дор-NADPH-d в нейронах дор--d в нейронах дор-d в нейронах дор- в нейронах дор-сального моторного ядра блуждающего нерва в контроле и после экспериментального воздей-ствия: А – контроль; Б – после введения ЛПС
E. coli (5 мкг/кг, 120 мин). Ув. 600
Рис. 2. Диаграмма распределения активности NADPH-d (А), СДГ (Б), ЛДГ (в) и АХЭ (Г) в нейронах моторного дви-гательного ядра блуждающего нерва в разных условиях эксперимента: 1 – контроль; 2 – после введения ЛПС E. сoli (5 мкг/кг, 120 мин); 3 – после введения ингибитора �-NAME (25 мг/кг) интактным животным; 4 – после введения ЛПС E. сoli (5 мкг/кг, 120 мин) на фоне действия ингибитора �-NAME (25 мг/кг). Достоверность различий в сравне-�-NAME (25 мг/кг). Достоверность различий в сравне--NAME (25 мг/кг). Достоверность различий в сравне-NAME (25 мг/кг). Достоверность различий в сравне- (25 мг/кг). Достоверность различий в сравне-
нии с контролем, принятым за 100%: * – Р < 0,05; ** – Р < 0,01
Национальная
академия наук
Беларуси

84
после введения эндотоксина увеличивалась на 64,4 ± 6,5% (Р < 0,01) (рис. 2, Г), а активность СДГ и ЛДГ – на 52,3 ± 4,8 и 31,2 ± 3,9% соответственно (Р < 0,05, Р < 0,01) (рис. 2, Б, в).
Повышение активности NADPH-d, АХЭ и фер-NADPH-d, АХЭ и фер--d, АХЭ и фер-d, АХЭ и фер-, АХЭ и фер-ментов энергетического обмена может свидетель-ствовать об увеличении функциональной активно-сти этих структур. В работах ряда авторов, иссле-довавших как ЦНС, так и периферическую нервную систему, показано, что высокая активность фермен-тов энергетического обмена связана с повышенной функциональной активностью нейронов, регистри-руемой с применением электрофизиологических методов [16, 17]. Результаты исследований других авторов, полученные на модели, аналогичной на-шей, показали, что в условиях эндотоксининдуци-рованной лихорадки усиливается афферентная и эфферентная импульсация в блуждающих нервах и мультинейронная активность в ЯСТ крысы [4, 5]. Параллельно с этим в ЯСТ повышаются активность ферментов энергетического обмена и интенсивность синтеза NO и гидролиза ацетилхолина [4].
При светооптическом исследовании препаратов продолговатого мозга, окрашенных по методу Нис-сля, в ДМЯБН интактных и лихорадящих животных определялись нейроны в различном морфофункцио-нальном состоянии. После введения ЛПС у неко-торых нейронов были отмечены участки с раство-ренной хроматофильной субстанцией. Выявлялись нейроны, тела которых выглядели набухшими, ядро
было сдвинуто к одному из полюсов, а базофильная субстанция уменьшена. Данное состояние клеток свидетельствует об их разном функциональном напряжении (рис. 3, А, Б).
Известно, что относительно небольшое количество NO в организме необходимо для защиты клетки от повреждения. Однако его избыточная продукция во время лихорадки воспалительно-го происхождения, ишемии/реперфузии, теплового шока и др. может приводить к повреждению клетки, нарушению ее кровоснабжения и оказывать цитотоксическое действие на окружающие ткани [18, 19]. Частичная блокада образования NO ингибиторами NO-синтазы нивелирует по-NO ингибиторами NO-синтазы нивелирует по- ингибиторами NO-синтазы нивелирует по-NO-синтазы нивелирует по--синтазы нивелирует по-вреждение [20]. В частности, лечение септического шока предполагает фармакологические вме-шательства, направленные на ингибирование гиперпродукции NO [19].
У интактных животных, взятых на исследование, через 120 мин после инъекции ингибитора NO-синтазы �-NAME в дозе 25 мг/кг активность NADPH-d в нейронах ДМЯБН была на 17,0 ± 1,8% (Р < 0,05) ниже, чем в контроле (см. рис. 2, А). Активность ферментов энергетического обмена и АХЭ достоверно не изменялась (рис. 2, Б, в, Г). Предварительное (за 30 мин до введения ЛПС) вве-дение �-NAME не вызывало характерной для этой дозы и времени лихорадочной реакции и обу-�-NAME не вызывало характерной для этой дозы и времени лихорадочной реакции и обу--NAME не вызывало характерной для этой дозы и времени лихорадочной реакции и обу-NAME не вызывало характерной для этой дозы и времени лихорадочной реакции и обу- не вызывало характерной для этой дозы и времени лихорадочной реакции и обу-словливало снижение активности NADPH-d в нейронах ДМЯБН на 27,9 ± 3,1% (Р < 0,01) (рис. 2, А). Активность СДГ уменьшалась на 16,0 ± 2,1%, а ЛДГ – на 21,8 ± 1,7% (Р < 0,01) (рис. 2, Б, в). Интен-сивность цитоплазматической окраски, отражающей активность АХЭ, не изменялась (рис. 2, Г).
Заключение. Таким образом, на основании анализа полученных данных (активности NA-NA-DPH-d, АХЭ и ферментов энергетического обмена) можно предположить, что NO- и холинергиче--d, АХЭ и ферментов энергетического обмена) можно предположить, что NO- и холинергиче-d, АХЭ и ферментов энергетического обмена) можно предположить, что NO- и холинергиче-, АХЭ и ферментов энергетического обмена) можно предположить, что NO- и холинергиче-NO- и холинергиче-- и холинергиче-ские структуры ДМЯБН продолговатого мозга в условиях эндотоксиновой лихорадки, вызванной действием ЛПС E. сoli, могут принимать участие в системных реакциях организма по поддержа-нию температурного гомеостазиса.
Рис. 3. Распределение глыбок тигроидного веще-ства в нейронах дорсального моторного ядра блуж-дающего нерва у интактных животных (А) и при
эндотоксиновой лихорадке (Б). Ув. 600
Национальная
академия наук
Беларуси

Литература
Г у р и н В. Н. Механизмы лихорадки. Минск, 1993.1. Н о з д р а ч е в А. Д., К о л о с о в а Л. И., М о и с е е в а А. Б., Р я б ч и к о в а О. В. // Физиол. журн. им. 2.
И. М. Сеченова. 2000. Т. 86, № 6. С. 728–742.W a t k i n s �. R., M a i e r S. F., G o e h l e r �. E. // �ife Sci. 1995. Vol. 37. Р. 1011–1026.3. Л а п ш а В. И., Б о ч а р о в а В. Н., Л у к а ш е н к о Т. М. и др. // Нейрофизиология. 2006. Т. 38, № 4. 4.
С. 298–302.Л а п ш а В. И., А з е в О. А., Л у к а ш е н к о Т. М., Ш е л а е в а Е. А. // Нейрофизиология. 2000. № 2. 5.
С. 112–119.H o f e r D., A s a n E., D r e u c h h a h n D. // NIPS. 1989. Vol. 14. P. 18–23. 6. Г у р и н В. Н., Г у р и н А. В. Терморегуляция и биологически активные вещества крови. Минск, 2004.7. P a x i n o s G., W a t s o n C. The Rat Brain in Stereotaxic Coordinates. San Diego, 1998.8. V i n c e n t S. R., K i m u r a H. // Neuroscience. 1992. Vol. 46, N 4. Р. 755–784.9.
К о р ж е в с к и й Д. Э., О т е л л и н В. А., Г р и г о р ь е в И. П. и др. // Морфология. 2007. Т. 132, № 4. С. 77–80.10. О х о т и н В. Е., Ш у к л и н А. В. // Морфология. 2006. Т. 129, № 1. С. 7–15.11. D a w s o n T. M., S n y d e r S. H. // J. Neurosci. 1994. Vol. 14, N 9. Р. 5147–5159.12. M a t s u m o t o T., N a k a n e M., P o l l o c k J. S. et al. // Neurosci. �ett. 1993. Vol. 155. P. 61–64.13. М о т а в к и н П. А., О х о т и н В. Е. // Бюл. эксперим. биол. и мед. 1982. Т. 4, № 12. С. 89–90.14. B r u c e J. // Brain Res. 1981. Vol. 219, N 1. P. 190–195.15. M o w e G. M., G e r s h o n M. D. // J. Comp. Neurol. 1986. Vol. 249, N 3. P. 381–391.16. N e l s o n J. S., W a k e f i e l d �. // J. Histochem. and Cytochem. 1973. Vol. 21, N 2. P. 184–188.17. A r s t a l l M. A., S a w v e r D. B., F u k a z a w a R., K e l l u R. A. // Circ. Res. 1999. Vol. 85. P. 829–840.18. С т о к л е Ж. К., М ю л е Б., А н д р и а н ц и т о х а й н а Р. // Биохимия. 1998. Т. 63, № 7. С. 976–983.19. М а н у х и н а Е. Б., П о к и д ы ш е в Д. А., Г о л у б е в а Л. Ю. и др. // Изв. РАН. Сер. Биол. 1996. № 5. 20.
С. 583–588.
v. n. BochaRova , E. n. Savchina
HISTOCHEMICAL ANALYSIS OF NADPH-DIAPHORASE, ACETYLCHOLINESTERASE AND ENERGY METABOLISM ENZYMES ACTIVITY IN STRUCTURES OF MEDULLA OBLONGATA
IN RATS DURING FEVER
institute of physiology of naS of Belarus, Minsk
Summary
Intraperitonal injection of Escherichia coli lipopolysaccharide (5 mcg/kg) results in the increase of NADPH-diaphorase, acetylcholinesterase and energy metabolism enzymes in the neurons and endotheliocytes of blood vessels within dorsal nu-cleus of the vagus nerve of medulla oblongata.
Национальная
академия наук
Беларуси

86
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 612.8.015
Г. К. ТРопниКовА
СРАВНИТЕЛЬНЫй АНАЛИЗ ПРОТЕКТОРНЫХ ЭФФЕКТОВ ИНТЕРВАЛЬНОй ГИПОКСИИ И МЕЛАТОНИНА НА СЕРОТОНИНЕРГИЧЕСКИЕ
СТРУКТУРЫ ПЕРЕДНИХ ОТДЕЛОВ МОЗГА КРЫСЫ
институт физиологии нАн Беларуси, Минск
(постуила в редакцию 01.03.2010)
Введение. Известно, что длительная экспозиция к гипобарической гипоксии индуцирует ка-скад патологических процессов в мозге, что может привести к его структурно-функциональным повреждениям и развитию состояний, сопровождаемых нарушением двигательной активности, памяти, когнитивных процессов. В связи с этим разработка новых эффективных способов повы-шения устойчивости мозга к повреждающим воздействиям гипоксии/ишемии является актуаль-ной задачей современной биологии и медицины.
Известно, что ключевую роль в нейрональной активности мозга играет серотонин (5-ОТ). 5-ОТ-нейроны эволюционировали как генеральная регуляторная система, отвечающая на внеш-ние стимулы путем модифицирования своей активности. Эти нейроны интенсивно взаимодей-ствуют не только с другими нейронами (через синаптические контакты), но и с эндотелиальны-ми, эпендимальными, эндокринными, астроцитарными и олигодендроглиальными клетками, с которыми они имеют специализированные контакты. Таким образом, 5-ОТ-нейроны имеют структурную организацию, совместимую с функциями 5-ОТ как нейромедиатора и трофического фактора. Как нейротрансмиттер 5-ОТ оказывает быстрые постсинаптические эффекты на нейро-ны, а как трофический фактор он влияет на регенерацию поврежденных клеток-мишеней прямо или путем активации выделения глиального белка S-100 [1] или нейротрофического фактора BDNF (Brain derived neurotrophic factor) [2]. Поэтому изменение уровня 5-ОТ в мозге является сигналом к запуску эволюционно и генетически детерминированных приспособительных меха-низмов, помогая тем самым динамически интегрировать и стабилизировать структуру и функ-ции ЦНС.
Подтверждением этому служат экспериментальные данные, свидетельствующие о том, что крысы, перенесшие пренатальную гипоксию, отстают в физиологическом развитии и у них на-рушается формирование памяти [3]. У таких животных обнаружено снижение количества 5-ОТ-нейронов в мозге [4]. У взрослых крыс в условиях высокогорья также отмечалось нарушение обучения, что сопровождалось снижением функции 5-ОТ-системы мозга [5].
Предшественником мелатонина в организме является 5-ОТ, и в норме между ними поддер-живается определенный баланс. Имеется множество свидетельств о протекторной роли мелато-нина против индуцируемого гипоксией окислительного стресса [6]. В частности, установлено, что мелатонин сохраняет митохондриальный гомеостаз, редуцируя генерацию свободных ради-калов путем повышения уровня митохондриального глутатиона, сохранения протонного потен-циала и синтеза АТФ благодаря стимуляции активности дыхательных комплексов I и IV [7].
Для профилактики и терапии постишемических структурно-функциональных нарушений мозга в настоящее время наряду с фармакологическими подходами большое внимание уделяется немедикаментозным методам лечения, направленным на мобилизацию эндогенных защитных механизмов. Одним из наиболее эффективных способов защиты мозга является прекондициони-рование краткими курсами малоинтенсивной гипобарической гипоксии. Еще в 1990 г. в экспери-
Национальная
академия наук
Беларуси

87
ментах на песчанках было продемонстрировано, что кратковременное воздействие сублетальны-ми дозами ишемии предотвращает гибель чувствительных пирамидных нейронов гиппокампа СА 1 в ответ на последующую глобальную ишемию [8]. Впоследствии этот феномен был воспро-изведен и на других отделах мозга (неокортексе, стриатуме) [9, 10].
Имеются также экспериментальные доказательства, свидетельствующие о возможности ис-пользования гипоксического прекондиционирования для профилактики нейродегенеративных процессов в мозге крыс с экспериментальной болезнью Альцгеймера [11], поведенческих нару-шений при экспериментальной болезни Паркинсона [12] и вызванных стрессами депрессивных состояний [13].
В предыдущих работах нами было установлено, что предварительное воздействие краткими курсами интервальной гипоксии значительно увеличивает физическую выносливость и актив-ность животных в условиях действия непрерывной гипоксии [14]. Этот эффект сопровождался стабилизацией уровня 5-ОТ и общего белка в миокарде [15], значимой активацией его метабо-лизма в вентролатеральной области продолговатого мозга и, напротив, снижением его уровня в дорсомедиальной области [16]. Однако было обнаружено, что введение экзогенного мелатони-на перед каждым сеансом интервальной гипоксии в значительной степени снижает защитный эффект интервальной гипоксии, что выражалось в угнетении исследовательских реакций и дви-гательной активности у животных в условиях непрерывной гипоксии [14].
Настоящее исследование посвящено сравнительному анализу влияния прекондиционирова-ния краткими курсами малоинтенсивной гипобарической интервальной гипоксии и мелатонина на вызываемые непрерывной гипоксией особенности изменения метаболизма 5-ОТ в церебраль-ной коре и гиппокампе крыс с целью дальнейшего изучения роли 5-ОТ-ергических структур мозга на разных уровнях ЦНС в механизме протекторных эффектов мелатонина и гипоксическо-го прекондиционирования.
Материалы и методы исследования. Опыты проводили на беспородных белых крысах-самцах (6 групп по 6 особей в каждой) массой 180–200 г. Животных содержали в помещении ви-вария на стандартном рационе питания при естественном световом режиме, постоянной темпе-ратуре воздуха 21 ˚С и свободном доступе к воде. I (контрольную) группу составили интактные животные. Во II группу вошли крысы, подвергавшиеся 10-минутной гипобарической непрерывной гипоксии (2300 м над уровнем моря). Гипоксию создавали путем разрежения воздуха в сосуде, со-единенным с вакуометром. Для предотвращения накопления в емкости углекислоты использовали натронную известь. III группу составили животные с интервальной гипоксией (ИГ), которых в течение 7 дней 1 раз в день подвергали воздействию гипоксии тех же параметров в следующем режиме: три эпизода 1,5-минутной гипоксии, чередуемые такими же эпизодами дыхания атмо-сферным воздухом, что достигалось путем выравнивания давления в сосуде с окружающей сре-дой. Животных IV группы, так же как и животных III группы, подвергали воздействию ИГ, но за 30 мин до сеанса ИГ им внутрибрюшинно вводили мелатонин (Sigma Aldrich Chemicals, США) в дозе 0,5 мг/кг в объеме 0,2 мл. Животные V и VI групп соответственно в течение 7 дней либо однократно получали мелатонин в той же дозе, но тренировке ИГ не подвергались.
Через 7 дней животных III–VI групп, так же как и животных II группы, подвергали воздей-III–VI групп, так же как и животных II группы, подвергали воздей-–VI групп, так же как и животных II группы, подвергали воздей-VI групп, так же как и животных II группы, подвергали воздей- групп, так же как и животных II группы, подвергали воздей-II группы, подвергали воздей- группы, подвергали воздей-ствию непрерывной 10-минутной гипоксии с целью сравнительного анализа роли предваритель-ного воздействия краткими курсами ИГ и мелатонина на характер ответных реакций 5-ОТ-ергических структур мозга на непрерывное действие гипоксии. Все эксперименты проводили в первой половине дня в весеннее время (апрель–май).
После декапитации животных быстро извлекали головной мозг, выделяли кору больших по-лушарий и гиппокамп, в которых по методу [17] определяли содержание 5-ОТ и его основного метаболита – 5-оксииндолуксусной кислоты (5-ОИУК). Полученные данные обработаны стати-стически с применением метода ANOVA для малых выборок. Данные представлены в виде сред-ANOVA для малых выборок. Данные представлены в виде сред- для малых выборок. Данные представлены в виде сред-ней арифметической ± ошибка среднего (M±m). Достоверность оценивали по t-критерию Стью-дента. Различия считали достоверными при Р < 0,05.
Результаты и их обсуждение. Кора больших полушарий. В коре головного мозга в ответ на непрерывную 10-минутную гипоксию (II группа) достоверных изменений в уровне 5-ОТ и 5-ОИУК
Национальная
академия наук
Беларуси

88
относительно контроля не наблюдалось (табл. 1). Однократное введение мелатонина (VI группа) также не повлияло на ответные реакции 5-ОТ-ергических структур коры на непрерывную ги-поксию, хотя у крыс, получавших мелатонин в течение 7 дней (V группа), отмечалась некоторая активация метаболизма 5-ОТ, о чем свидетельствовало достоверное увеличение уровня 5-ОИУК при сохранении базального уровня 5-ОТ. У животных, предварительно в течение 7 дней подвер-гавшихся действию ИГ (III группа), тест-воздействие, так же как у животных V группы, вызыва-III группа), тест-воздействие, так же как у животных V группы, вызыва- группа), тест-воздействие, так же как у животных V группы, вызыва-V группы, вызыва- группы, вызыва-ло активацию метаболизма 5-ОТ в коре головного мозга (отмечалось достоверное увеличение индекса 5-ОИУК/5-ОТ, хотя уровень 5-ОТ не отличался от контроля). Однако предварительное сочетанное действие ИГ и мелатонина (IV группа) индуцировало в условиях непрерывной ги-IV группа) индуцировало в условиях непрерывной ги- группа) индуцировало в условиях непрерывной ги-поксии достоверное накопление 5-ОТ в коре головного мозга (на 39,9%) на фоне снижения уров-ня его катаболизма (отмечалось достоверное уменьшение индекса 5-ОИУК/5-ОТ на 34,2%).
Т а б л и ц а 1. Влияние интервальной гипоксии (ИГ) и мелатонина на вызываемые 10-минутной непрерывной гипоксией (НГ) изменения содержания 5-ОТ и 5-ОИУК (мкг/г ткани) в коре больших полушарий мозга крыс
(M±m)
Группа 5-ОТ 5-ОИУК 5-ОИУК/5-ОТ
I. Контроль 0,363 ± 0,033 0,362 ± 0,027 1,011 ± 0,067II. НГ 0,438 ± 0,039 0,415 ± 0,038 0,898 ± 0,055III. ИГ (7 дней) + НГ 0,347 ± 0,027 0,436 ± 0,027 1,281 ± 0,074*IV. Мелатонин (7 дней) + ИГ (7 дней) + НГ 0,508 ± 0,044* 0,328 ± 0,011 0,666 ± 0,051*V. Мелатонин (7 дней) + НГ 0,419 ± 0,029 0,464 ± 0,021* 1,200 ± 0,095VI. Мелатонин (однократно) + НГ 0,415 ± 0,025* 0,363 ± 0,031 0,868 ± 0,057
*Изменения достоверны (P < 0,05) по отношению к контролю.
Таким образом, прекондиционирование ИГ способствовало активации корковых 5-ОТ-ергических структур в условиях действия непрерывной гипоксии. Данные литературы свиде-тельствуют, что 5-ОТ модулирует активность пирамидальных нейронов префронтальной коры и облегчает двигательную и другие исполнительные функции ЦНС [18]. Следовательно, актива-ция 5-ОТ-ергических структур церебральной коры участвует в механизме отмечаемого нами ранее [14] увеличения физической активности животных, подвергавшихся тренировке ИГ. До-полнительное введение экзогенного мелатонина сопровождалось угнетением процессов окисли-тельного дезаминирования 5-ОТ и избыточному накоплению его в коре головного мозга, чем, возможно, и объясняется седативный эффект мелатонина. Это подтверждалось подавлением в аналогичных условиях опыта исследовательских реакций и двигательной активности у крыс [14]. В работе [13] также отмечалось, что у золотых рыбок введение мелатонина перед сеансом гипоксии приводит к снижению локомоторной активности и уровня кортизола в плазме.
В работе [3] указывалось также на более слабую выраженность противосудорожных и ан-гиопротекторных эффектов мелатонина при острой гипоксии по сравнению с защитными эф-фектами интервальной гипоксии. Кроме того, некоторые авторы [15] отмечали, что в условиях хронической гипоксии мелатонин утрачивал релаксирующее влияние на сосуды. Поэтому, сравнивая результаты собственных исследований с данными литературы, можно предполо-жить, что под влиянием прекондиционирования в ответ на острую гипоксию происходит выброс эндогенного мелатонина, а при дополнительном введении экзогенного мелатонина не исключена возможность либо десенситизации мелатониновых рецепторов, либо избыточного удаления реактивных кислородных радикалов, которые в определенном количестве активиру-ют защитные силы организма.
Гиппокамп. В ответ на 10-минутную гипоксию достоверных изменений уровня 5-ОТ и 5-ОИУК в гиппокампе, как и в коре головного мозга, у животных II группы не наблюдалось (табл. 2). Однако в отличие от коры головного мозга предварительное в течение 7 дней введение мелатонина не изменило этот паттерн реакции 5-ОТ-ергических структур гиппокампа (V груп-V груп- груп-па) на непрерывную гипоксию, тогда как однократное введение мелатонина перед сеансом сплошной гипоксии приводило к достоверному накоплению 5-ОТ на фоне снижения уровня
Национальная
академия наук
Беларуси

89
5-ОИУК и индекса 5-ОИУК/5-ОТ. У крыс, предварительно подвергавшихся ИГ (III группа), в от-III группа), в от- группа), в от-вет на 10-минутную непрерывную гипоксию, так же как и после однократного применения мела-тонина, отмечалось примерно одинаковое увеличение содержания 5-ОТ – на 23,9%. Однако при этом наблюдалось повышение уровня 5-ОИУК, поэтому индекс 5-ОИУК/5-ОТ не отличался от контрольных значений. Интересно отметить, что паттерн реакции гиппокампальных 5-ОТ-ергических структур на непрерывную гипоксию у крыс, предварительно подвергавшихся воз-действию ИГ в сочетании с мелатонином (IV группа), не отличался от такового при однократном введении мелатонина. У всех животных, получавших мелатонин, наблюдалось ингибирование окислительного дезаминирования 5-ОТ в гиппокампе. Кроме того, у этих животных, в отличие от крыс, подвергавшихся одной ИГ, отмечалась меньшая резистентность к действию непрерыв-ной гипоксии, что выражалось в пассивном поведении, снижении двигательной активности и ориентировочно-исследовательской деятельности [14]. Вероятно, применяемые параметры ИГ более оптимально способствовали активации метаболизма 5-ОТ в гиппокампе, что проявлялось в лучшем сохранении когнитивных способностей и двигательной активности животных в условиях непрерывной гипоксии. Возможно, в основе наблюдаемого явления лежало облегче-ние опосредуемой 5-ОТ-1А рецепторами нейропередачи в гиппокампе, активация которых селективными агонистами, как установлено ранее [22], способствовала восстановлению иссле-довательского рефлекса, нарушенного стрессом. Установлено также, что хроническое лечение селективными блокаторами обратного захвата 5-ОТ ослабляет постишемическое повреждение мозга [23] и уменьшает индуцированный ишемией дефицит пространственной памяти благодаря гиппокампальному неонейрогенезу [24]. Кроме того, в активации гиппокампальных нейронов важную роль играет тесно связанный с 5-ОТ-системой белок BDNF, пробуждающий функцио-нально не активные, так называемые silent, синапсы [25].
Т а б л и ц а 2. Влияние интервальной гипоксии (ИГ) и мелатонина на вызываемые 10-минутной непрерывной гипоксией (НГ) изменения содержания 5-ОТ и 5-ОИУК (мкг/г ткани) в гиппокампе крыс (M±m)
Группа 5-ОТ 5-ОИУК 5-ОИУК/5-ОТ
I. Контроль 0,485 ± 0,026 0,368 ± 0,022 0,768 ± 0,058II. НГ 0,491 ± 0,050 0,343 ± 0,020 0,727 ± 0,051III. ИГ (7 дней) + НГ 0,594 ± 0,018* 0,512 ± 0,036* 0,866 ± 0,071IV. Мелатонин (7 дней) + ИГ (7 дней) + НГ 0,587 ± 0,035* 0,312 ± 0,028 0,531 ± 0,034V. Мелатонин (7 дней) + НГ 0,486 ± 0,053 0,324 ± 0,021 0,729 ± 0,071VI. Мелатонин (однократно) + НГ 0,601 ± 0,023* 0,282 ± 0,017* 0,471 ± 0,034*
*Изменения достоверны (P < 0,05) по отношению к контролю.
Заключение. Прекондиционирование краткими курсами малоинтенсивной гипобарической гипоксии способствовало активации гиппокампальных и корковых 5-ОТ-ергических структур в ответ на действие непрерывной гипоксии. Дополнительное введение экзогенного мелатонина перед каждым сеансом интервальной гипоксии индуцировало избыточное накопление 5-ОТ в коре головного мозга, снижение содержания 5-ОИУК в коре больших полушарий мозга и гип-покампе и сопровождалось угнетением активного поведения у животных. Полученные данные указывают на то, что к применению мелатонина в сочетании с интервальной гипоксией в каче-стве средства повышения выносливости животных в условиях непрерывной гипоксии следует относиться с осторожностью.
Литература
1. A z m i t i a E. Ch. // Int. Rev. Neurobiol. 2007. Vol. 73. P. 31–56.2. M a r t i n o w i c h K., � u B. // Neuropsichopharmacology. 2008. Vol. 33, N 1. P. 73–83.3. Ж у р а в и н И. А., Д у б р о в с к а я Н. М., К о ч к и н а Е. Г. и др. // Технология живых существ. 2007. Т. 4,
№ 5–6. С. 109–123.4. S t r a c k x E., v a n d e n H o v e D. �., S t e i n b u s c h H. P. et al. // Dev. Neurosci. 2008. Vol. 30, N 5.
P. 358–366.
Национальная
академия наук
Беларуси

5. С е м е н о в а Т. П., З а к и р о в Д. З., М е д в и н с к а я Н. И. и др. // Hyp. Med. J. 1996. Vol. 4, N 4. P. 3–6.6. R e i t e r R. J., T a n D. X., J o u M. J. et al. // Neuroendocrinol. �ett. 2008. Vol. 29, N 4. P. 391–398.7. A c u n a - C a s t r o v i e j o D., E s c a m e s G., � e o n J. et al. // Adv. Exp. Med. Biol. 2003. Vol. 527. P. 549–557.8. K i t a g a w a K., M a t s u m o t o M., T a g a y a M. et al. // Brain Res. 1990. Vol. 528, N 1. P. 21–24.9. С а м о й л о в М. О., С е м е н о в Д. Г., Т ю л ь к о в а Е. Н. и др. // Проблемы гипоксии: молекулярные,
физиологические и медицинские. М., 2004. С. 96–111.10. H a s s e n G. W., T i a n D., D i n g D., B e r g o l d P. J. // Brain Res. Brain Res. Protoc. 2004. Vol. 13, N 3.
P. 135–143.11. М а н у х и н а Е. В., Г о р я ч е в а А. В., Б а р с к о в И. В. и др. // Рос. физиол. журн. им. И. М. Сеченова.
2009. Т. 95, № 5. С. 706–715.12. Г у л я е в а Н. В., С т е п а н ы ч е в М. Ю., М и т р о х и н а О. С. // Hyp. Med. 1996. N 4. С. 7–11.13. Р ы б н и к о в а Е. А., С а м о й л о в Е. О., М и р о н о в а В. И. // Журн. неврол. и психиатр. им. Корсакова.
2009. № 7. С. 11–17.14. Т р о п н и к о в а Г. К. // Закономерности развития патологических состояний и их коррекция. Минск, 2009.
С. 263–265.15. Т р о п н и к о в а Г. К., Г о р б у н о в а Н. Б. // Закономерности развития патологических состояний и их
коррекция. Минск, 2009. С. 266–269. 16. Т р о п н и к о в а Г. К. // Новости мед.-биол. наук. 2010. № 2. C. 182–188.17. C u r z o n G., G r e e n A. R. // Br. J. Pharmacol. 1970. Vol. 39, N 3. P. 653–655.18. Z h a n g Z., A r s e n a u l t D. // J. Physiol. 2005. Vol. 566, N 2. P. 379–394.19. A z p e l e t a C., M a r t i n e z - A l v a r e z R. M., D e l g a d o M. J. // Horm. Behav. 2010. Vol. 57, N 3. P. 323–329.20. К у з е н к о в В. С., М о р е н к о в Е. Д., К о ш е л е в В. Б. // Hyp. Med. J. 2000. Vol. 8, N 1–2. P. 4–7.21. D a s R., B a l o n a n �., B a l l a r d H. J., H o S. // Int. J. Cardiol. 2008. Vol. 126, N 3. P. 340–345.22. G r a e f f F. G., G u i m a r a e s F. S., D e A n r a d e T. G., D e a k i n J. F. // Pharmacol. Biochem. Behav. 1996.
Vol. 54, N 1. P. 129–141.23. S h i n T. K., K a n g M. S., � e e H. G., S e o M. S. et al. // Korean J. Physiol. Pharmacol. 2009. Vol. 13, N 3.
P. 257–263.24. � i W. �., C a i H. H., W a n g B. et al. // J. Neurosci. Res. 2009. Vol. 87, N 1. P. 112–122.25. V a n D o n k e l a a r E. �., v a n d e n H o v e D. �., B l o k l a n d A. et al. // Eur. Neuropsychopharmacol. 2009.
Vol. 19, N 11. P. 812–821.
G. K. TRopniKova
COMPARATIVE ANALYSIS OF PROTECTIVE EFFECTS OF INTERMITTENT HYPOXIA AND MELATONIN ON THE SEROTONINERGIC STRUCTURES IN THE RAT FOREBRAIN
institute of physiology of naS of Belarus, Minsk
Summary
Preconditioning the intermittent hypobaric hypoxia (1.5 min × 3 with 1.5 min intervals/day/7days) promoted the activation of hippocampal and cortical serotoninergic structures of the brain under the action of continuous hypoxia. Additional introduction of exogenous melatonin before each session of intermittent hypoxia induced under the action of continuous hypoxia the superfluous accumulation of serotonin in the cortex of the brain, a decrease of 5-hydroxyindole acetic acid in the cortex and hippocampus and was accompanied by the inhibition of the active behavior of animals.
Национальная
академия наук
Беларуси

91
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616-092.9:616-006.6
Т. и. ТЕРпинСКАя
ВЛИЯНИЕ ТРАНСПЛАНТАЦИИ КЛЕТОК КОСТНОГО МОЗГА И СЕЛЕЗЕНКИ НА ПРИВИВАЕМОСТЬ И СКОРОСТЬ РОСТА АСЦИТНОй КАРЦИНОМЫ ЭРЛИХА
У МЫШЕй
институт физиологии нАн Беларуси, Минск
(поступила в редакцию 01.07.2009)
Введение. Возникновение и развитие опухоли зависит как от влияния разнообразных внеш-них факторов канцерогенеза, так и от состояния внутренних систем организма, в частности от иммунного статуса, определяемого балансом клеточных популяций, вовлеченных в ассоцииро-ванный с опухолью иммунный ответ. Патологическая иммунореактивность при раке проявляет-ся в снижении выраженности противоопухолевой защиты и появлении популяций и клонов им-мунных клеток, оказывающих проопухолевый эффект [1, 2]. Вероятно, такие изменения функ-ций иммунитета развиваются постепенно.
Цель исследования – изучение динамики функциональных свойств костного мозга и селезен-ки по мере развития опухоли, а именно их способности подавлять либо стимулировать рост зло-качественных клеток.
Материалы и методы исследования. Работа выполнена на 256 самках мышей линии Af 2–3-ме-сячного возраста массой 20–22 г. Животные находились в стационарных условиях вивария при температуре 21,0 ± 1,0 °С, имели свободный доступ к воде и пище. Мышам подкожно вводили по 6 млн клеток асцитной карциномы Эрлиха (АКЭ) на особь. На 2, 5, 7, 9, 13, 20 и 30-е сутки после прививки животных забивали методом цервикальной дислокации, вырезали опухоли и, взвешивая каждую из них, определяли среднюю массу опухолей на каждом этапе эксперимента; аналогичным образом оценивали массу селезенок. Получали суспензии клеток селезенки и костного мозга опухоленосителей (в каждой серии опытов использовали по 6–7 жи-вотных). Для этого селезенки всех мышей-доноров серии измельчали ножницами, а затем про-давливали через капроновое сито стеклянным пестиком, смывая клетки в пробирку физиологи-ческим раствором Эрла. Клетки костного мозга получали из костей бедра и голени: срезали эпи-физы и промывали из шприца с тонкой иглой физиологическим раствором, вымывая костный мозг; пропускали через капроновое сито, смывая клетки физиологическим раствором. Затем су-спензии центрифугировали 10 мин при 1500 об/мин, жидкость сливали, клетки ресуспензирова-ли в физиологическом растворе. Количество ядросодержащих клеток в суспензии определяли, используя 3%-ный раствор уксусной кислоты [3]. Полученные в разные сроки опухолевого роста суспензии инъецировали внутрибрюшинно в дозе 100 млн ядросодержащих клеток селезенки или 10 млн ядросодержащих клеток костного мозга в 0,3 мл суспензии на мышь через 1 ч после подкожного введения животным 100 тыс. клеток АКЭ. По наличию подкожного роста опухоли оценивали прививаемость последней в описанных условиях; регистрировали продолжитель-ность жизни мышей с привившейся опухолью. В двух сериях вместо суспензий спленоцитов и миелоцитов опухоленосителей трансплантировали аналогичные суспензии, полученные от ин-тактных мышей. Контролем служили животные, которым прививали АКЭ, не подвергая их ни-каким дополнительным воздействиям. Всего поставлено 17 серий опытов, в каждую из которых было включено от 8 до 20 особей. При статистической обработке данных процент прививаемо-
Национальная
академия наук
Беларуси

92
сти опухолей и ошибку среднего в сериях, где АКЭ привили всем животным, рассчитывали по методу Ван-дер-Вардена [4].
Результаты и их обсуждение. При росте опухоли у мышей – доноров спленоцитов и миело-цитов регистрировалось увеличение массы селезенки (рис. 1). На 2-е сутки после прививки опу-холи масса селезенки носителей незначительно отличалась от этого показателя у интактных жи-вотных (увеличение на 7,3%, Р > 0,05). К 5-м суткам превышение ее над массой селезенки ин-тактных мышей составило 47,9% (Р < 0,05), к 9-м суткам масса селезенки опухоленосителей достигала максимума (превышение над интактным контролем – 52,7%, Р < 0,05), затем несколь-ко снижалась, оставаясь, однако, на 13-е сутки роста АКЭ выше этого показателя у интактных особей на 33,4% (Р < 0,05), на 20-е – на 7,7% (Р > 0,05) и на 30-е – на 19,4% (Р > 0,05).
Рис. 1. Масса опухолей и селезенок у мышей – доноров спленоцитов и миелоцитов. * – достоверность различий (Р < 0,05) по сравнению с массой селезенок интактных мышей
При анализе противоопухолевых свойств клеток селезенки и костного мозга установлено, что в контрольной серии (прививка опухоли без дополнительных воздействий) прививаемость АКЭ составила 60,0 ± 11,0%. Внутрибрюшинное введение спленоцитов, полученных у интакт-ных животных, привело к увеличению прививаемости: последняя составила 84,6 ± 10,0%, что в 1,41 раза выше, чем в контроле (0,1 < P <0,05) (рис. 2). Клетки селезенки, полученные на 2-е сутки роста АКЭ у доноров, не влияли значительно на прививаемость гомологичной опухоли, которая развилась у 66,7% мышей (в 1,11 раза больше, чем в контроле, и в 1,27 раза меньше, чем при введении спленоцитов интактных мышей). Введение спленоцитов, взятых на 5-е сутки по-сле прививки АКЭ, способствовало тому, что опухоль развилась у всех животных этой серии (при расчете по методу Ван-дер-Вардена – 92,9 ± 6,6%). Прививаемость увеличилась в 1,54 раза по сравнению с контрольной прививкой (P < 0,05) и в 1,10 раза по сравнению с серией, где парал-лельно с опухолью вводили спленоциты интактных мышей (P > 0,1). При трансплантации клеток селезенки, выделенных на 7-е сутки роста АКЭ, прививаемость составила 58,3 ± 14,2%, прак-тически не изменившись по сравнению с контролем и снизившись в 1,45 раза по сравнению с прививаемостью при введении спленоцитов интактных животных (в обоих случаях P > 0,1). Полученные на 9-е сутки роста АКЭ клетки селезенки способствовали прививаемости, как и 5-суточные: рост опухоли имел место у всех мышей (при расчете по методу Ван-дер-Вардена – 92,0 ± 7,1%), прививаемость увеличивалась по сравнению с контролем и серией, где донорами спленоцитов были интактные животные, в 1,53 (Р < 0,05) и в 1,09 раза (P > 0,1) соответственно. Прививаемость при трансплантации клеток селезенки, полученных на 13-е сутки роста АКЭ, не изменилась по сравнению с этим показателем в контроле и была в 1,48 раза меньше, чем при вве-дении спленоцитов интактных мышей (P > 0,1 в обоих случаях). При внутрибрюшинном введе-
Национальная
академия наук
Беларуси

93
нии параллельно с прививкой АКЭ спленоцитов, выделенных у мышей на последующих этапах опухолевого развития (20-е и 30-е сутки), доля животных с развившимися опухолями значитель-но не отличалась от таковой в контроле, составляя 63,6 ± 14,5 и 61,5 ± 13,5% соответственно, и была несколько меньше (в 1,33 и 1,38 раза соответственно), чем при введении спленоцитов ин-тактных мышей (во всех случаях Р > 0,1).
Рис. 2. Прививаемость АКЭ у мышей при параллельной трансплантации им клеток селезенки и костного мозга синген-ных доноров – носителей гомологичной опухоли. * – достоверность различий (Р < 0,05) по сравнению с контролем
Как показано на рис. 2, клетки костного мозга интактных мышей существенно не повлияли на прививаемость АКЭ, которая в этом случае составила 70,0 ± 14,5% (повышение в 1,17 раза, Р > 0,1). Миелоциты, взятые на 2-е сутки роста АКЭ, также не оказали заметного эффекта на прививае-мость гомологичной опухоли, развившейся у 54,5 ± 15,0% животных (снижение в 1,10 и 1,28 раза по сравнению с контролем и с введением спленоцитов интактных животных соответственно, P > 0,1 в обоих случаях). Миелоциты животных, выделенные на 5-е сутки роста АКЭ, снижали количество привившихся опухолей до 40,0 ± 15,5% (в 1,75 раза по сравнению с миелоцитами ин-тактных животных и в 1,50 раза по сравнению с прививкой опухоли без воздействия), однако это снижение не было статистически значимым (Р > 0,1). Клетки костного мозга, взятые у доноров-опухоленосителей на 7-е сутки роста АКЭ, также не оказали статистически достоверного эффек-та на прививаемость гомологичной опухоли по сравнению с миелоцитами интактных мышей (снижение в 1,40 раза, Р > 0,1) и с прививаемостью в контроле (снижение в 1,20 раза, Р > 0,1); прививаемость на этом этапе составила 50,0 ± 17,7%. При трансплантации клеток костного моз-га, полученных на 9-е сутки роста АКЭ, опухоли привились у всех мышей серии (при расчете по методу Ван-дер-Вардена – 90,0 ± 9,5%), аналогично клеткам селезенки, взятым на том же этапе, что в 1,5 и 1,29 раза больше по сравнению с этим показателем в контроле (Р < 0,05) и при транс-плантации миелоцитов интактных мышей (0,1 < P <0,05). Клетки костного мозга, взятые на 13-е сутки роста АКЭ, снизили прививаемость АКЭ в 1,4 раза по сравнению с миелоцитами интакт-ных мышей и в 1,2 раза по сравнению с контрольной прививаемостью, аналогично 7-суточным (Р > 0,1 в обоих случаях); прививаемость составила 50 ± 15,8%. В то же время миелоциты, выде-ленные у мышей на 20-е сутки опухолевого роста, способствовали усилению прививаемости – опухоли развились у 90,0 ± 9,5% мышей, что в 1,50 раза больше, чем в контроле (Р < 0,05) и в 1,29 раза больше, чем при трансплантации миелоцитов интактных животных (P > 0,1). Незначитель-ное увеличение прививаемости АКЭ обнаружено также при трансплантации клеток костного мозга, полученных у мышей на 30-й день роста опухоли: в 1,36 и 1,17 раза больше по сравнению с прививаемостью в контроле и при введении интактных миелоцитов соответственно (Р > 0,1 в обоих случаях).
Сутки роста опухоли у мышей – доноров клеток селезенки и костного мозга
Национальная
академия наук
Беларуси

94
Продолжительность жизни мышей с привившейся опухолью в контроле составила 101,6 ± 12,7 сут (рис. 3). При трансплантации спленоцитов, взятых у интактных животных или у носителей АКЭ на каждом из этапов эксперимента, средние значения продолжительности жизни в сериях коле-бались от 90,7± 23,5 сут (20-й день роста опухоли) до 140,8 ± 27,6 сут (13-й день роста опухоли), статистически значимых отличий между какими-либо сериями не выявлено (во всех случаях Р > 0,1). При введении клеток костного мозга интактных мышей или миелоцитов, полученных на 13-е сутки роста опухоли, наблюдалась тенденция к сокращению продолжительности жизни опухоленосителей, которая составила 68,7 ± 11,3 сут (в 1,48 раза меньше, чем в контроле) и 62,6 ± 3,9 сут (в 1,48 раза меньше, чем в контроле) соответственно (в обоих случаях 0,1 > P > 0,05). Трансплан-тация клеток костного мозга опухоленосителей во всех других сериях, за исключением исполь-зования донорских клеток, полученных на 7-е сутки роста опухоли, также вела к некоторому сокращению продолжительности жизни носителей АКЭ: мыши в среднем жили от 74,3 ± 9,9 сут (миелоциты забирали у доноров на 5-й день роста опухоли) до 99,8 ± 13,9 сут (миелоциты забира-ли у доноров на 2-й день роста опухоли). При трансплантации миелоцитов, полученных на 7-е сутки после прививки АКЭ, продолжительность жизни опухоленосителей составила 139,5 ± 59,2 сут, что соответственно в 1, 37 и в 2,06 раза дольше, чем продолжительность жизни животных, которые не получали донорских миелоцитов или получали миелоциты интактных мышей; ста-тистически значимых отличий между сериями не выявлено.
Таким образом, при внутрибрюшинном введении мышам параллельно с подкожной прививкой АКЭ спленоцитов интактных животных наблюдалась тенденция к увеличению прививаемости опухоли. При внутрибрюшинном введении спленоцитов носителей АКЭ, полученных у доноров на 5-е или 9-е сутки после прививки им АКЭ, регистрировалось статистически достоверное уси-ление прививаемости этой опухоли у реципиентов по сравнению с животными, не получавшими донорских спленоцитов; увеличение прививаемости также было статистически достоверным при сравнении с сериями, где мышам трансплантировали клетки селезенки, взятые на 2, 5, 9, 11 и 13-е сутки после прививки АКЭ.
Введение миелоцитов интактных животных, взятых на 9-е сутки роста АКЭ, также способ-ствовало увеличению прививаемости по сравнению с мышами, не получавшими миелоциты, а также получавшими миелоциты, взятые на 2, 5, 7 и 13-е сутки роста опухоли у доноров; тен-денция к увеличению прививаемости выявлялась также при сравнении с серией, где трансплан-
Рис. 3. Продолжительность жизни носителей АКЭ при параллельной трансплантации им клеток селезенки и костного мозга сингенных доноров – носителей гомологичной опухоли
Национальная
академия наук
Беларуси

95
тировались миелоциты интактных мышей. Сходную закономерность наблюдали при использо-вании для трансплантации клеток костного мозга, взятых у мышей на 20-е сутки роста АКЭ: увеличение числа животных с развившимися опухолями было статистически достоверным при сравнении с контролем и с сериями, где реципиенты получали миелоциты, взятые на 5-е и 13-е сутки после прививки АКЭ донорам; тенденция к повышению прививаемости выявлялась при сравнении с сериями, где использовали 2- и 7-суточные миелоциты. Наблюдали тенденцию к снижению продолжительности жизни мышей с привившейся опухолью при трансплантации ми-елоцитов интактных мышей.
Наибольшая прививаемость выявлялась при введении спленоцитов, полученных от доноров на тех этапах, когда масса селезенки была максимальной. Вероятно, это свидетельствует о нали-чии в это время в селезенке опухоленосителей клеточных популяций, поддерживающих толе-рантность организма к злокачественному росту. К настоящему времени многочисленными ис-следованиями установлен проопухолевый эффект ряда клеточных популяций, выявляющихся в повышенном количестве у опухоленосителей. К ним относят натуральные CD4+CD25+foxP3+ [5, 6] и индуцированные CD4+CD25+/– [5] Т-регуляторные лимфоциты, подавляющие проти-воопухолевую реакцию антиген-специфическим и антиген-неспецифическим образом соот-ветственно; макрофаги с фенотипом М2, способствующие ангиогенезу опухоли [7]; незрелые натуральные киллеры с регуляторным эффектом, секретирующие цитокины II типа, такие как трансформирующий фактор роста β и ИЛ-10 [8, 9]; супрессорные клетки миелоидного проис-хождения с фенотипом CD11b+TCR-, ингибирующие клональную пролиферацию и функции CD8+ Т-лимфоцитов [10, 11]; CD4+ НКТ-клетки, подавляющие противоопухолевые реакции и индуцирующие супрессоры миелоидного происхождения [2]; незрелые дендритные клетки CD11c+ с нарушенной функцией передачи костимуляторного сигнала, которые не способны ини-циировать цитотоксический ответ CD8+ Т-клеток, а также супрессируют Т-клеточную пролифе-рацию, ускоряют апоптоз Т-клеток и индуцируют активность Т-регуляторных клеток [2, 12]; мезенхимальные стволовые клетки костного мозга (CD105+, CD73+, CD90+, CD166+, CD44+, CD29+, CD14–, CD34–, CD45–, CD31–), супрессирующие функции CD8+, НК и дендритных кле-ток и стимулирующие Т-регу ляторные клетки [13, 14]; В-лимфоциты, вносящие вклад в усиле-ние ангиогенеза, инвазивности и метастазирования опухоли [15–18].
Эффекты трансплантации спленоцитов и миелоцитов на опухолевый рост, а также волнообраз-ный характер изменения свойств этих клеток при злокачественном росте определяются, вероятно, кинетикой ассоциированных с опухолью иммунных реакций, в частности временем индукции и длительностью проявления толерантности – индуцированного антигеном специфического пода-вления иммунного ответа [19]. Можно предположить, что первая волна клеток с проопухолевыми функциями обусловливается индукцией толерантности Т-клеток (через несколько часов после воз-действия опухолевого толерогена) с последующей (4–5-е сутки) пролиферацией стимулированных клонов и, таким образом, выявляется в селезенке на 5-е сутки опухолевого роста. Вторую волну (9-е сутки опухолевого роста), возможно, представляют собой участвующие в развитии толерант-ности к опухоли В-клетки селезенки и костного мозга. Увеличение массы селезенки на 5-е и 9-е сутки роста АКЭ может быть связано с задержкой в этом лимфоидном органе выходящих из цир-куляции специфичных к опухолевым антигенам Т- и В-клеток соответственно и с их клональной пролиферацией. Часть этих лимфоцитов, возможно, вовлечена в процесс формирования онкотоле-рантности и ответственна за усиление прививаемости опухоли. Изменения свойств спленоцитов и миелоцитов могут быть связаны не только с вариацией количества толерогенных и способствую-щих опухолевому росту клеток в селезенке и костном мозге носителей на этих этапах, но и с раз-витием клеточных популяций с противоопухолевыми функциями (NK и НКТ-клетки и CD8+ Т-лимфоциты [1, 9, 12, 20]). Кроме того, существенное влияние могут оказывать местные механиз-мы избегания опухолью действия иммунного ответа, в частности слабая иммуногенность малиг-низированных клеток и выделение ими иммунодепрессивных факторов (например, трансформи-рующего фактора роста β, ИЛ-10, простагландина Е2 и др. [1]).
Заключение. При внутрибрюшинной трансплантации сингенных спленоцитов интактных доноров параллельно с подкожной прививкой АКЭ наблюдается тенденция к усилению приви-
Национальная
академия наук
Беларуси

ваемости солидной формы этой опухоли у реципиентов; при трансплантации клеток селезенки, полученных на 5-е и 9-е сутки после прививки АКЭ, отмечается статистически достоверное уси-ление прививаемости по сравнению с таковой у мышей, не получавших донорские спленоциты. Трансплантация клеток костного мозга, полученных у мышей на 9-е и 20-е сутки роста АКЭ, также увеличивает прививаемость гомологичной опухоли по сравнению с этим показателем у животных, не получавших донорские миелоциты; при трансплантации клеток костного мозга интактных мышей статистически значимого повышения прививаемости не зарегистрировано. Выявляется тенденция к сокращению продолжительности жизни опухоленосителей, получавших миелоциты интактных доноров.
Литература
K i m R., E m i M., T a n a b e K. // Immunology. 2007. Vol. 121. P. 1–14. 1. P a n P.-�., O z a o J., Z h o u Z., C h e n S.-H. // Adv. Drug Deliv. Rev. 2008. Vol. 60, N 2. Р. 91–105.2. Лабораторные методы исследования в клинике: справочник / Под ред. В. В. Меньшикова. М., 1987. 3. Р о к и ц к и й П. Ф. Биологическая статистика. Минск, 1973. 4. C u r e l T. J. // The J. of Clin. Investigation. 2007. Vol. 117, N 5. Р. 1167–1174.5. Q i n F. X. - F. // Cell. & Mol. Immunol. 2009. Vol. 6, N 1. Р. 3–13.6. B i s w a s S. K., S i c a A., � e w i s C. E. // The J. of Immunol. 2008. Vol. 180. Р. 2011–2017.7. Z h a n g C., Z h a n g J., T i a n Z. // Cell & Mol. Immunol. 2006. Vol. 3, N 4. Р. 241–254.8. Z a m a i �., P o n t i C., M i r a n d o l a P. et al. // J. of Immunol. 2007. Vol. 178. Р. 4011–4016.9. F r e y A. B. // The J. of Clin. Investigation. 2006. Vol. 116, N 10. Р. 2587–2590. 10. O c h o a A. 11. C., Z e a A. H., H e r n a n d e z C., R o d r i g u e z P. C. // Clin. Cancer Res. 2007. Vol. 15, N 13.
Р. 721–726.D h o d a p k a r M. 12. V., D h o d a p k a r K. M., P a l u c k a A. K. // Cell Death Differ. 2008. Vol. 15, N 1.
Р. 39–50. N a u t a A. J., F і b b e W. E. // Blood. 2007. Vol. 110, N 10. Р. 3499–3506.13. D u g a s t A. - S., V a n h o v e B. // Clin. and Exper. Immunol. 2009. Vol. 156. P. 25–34. 14. W i j e s u r i y a R., M a r i o S., Z o u J. P. et al. // Int. Immunol. 1998. Vol. 10, N 8. P. 1057–1065.15. N y h u s J. 16. K., W o l f o r d C. C., F r i e c e C. R. et al. // Cancer Immunol. Immunother. 2001. Vol. 50, N 7.
P. 361–372. B a r b e r a - G u i l l e m 17. E., M a y K. F. Jr., N y h u s J. K., N e l s o n M. B. // Neoplasia. 1999. Vol. 1, N 5.
P. 453–460.B a r b e r a - G u i l l e m 18. E., N y h u s J. K., W o l f o r d C. C. et al. // Cancer Res. 2002. Vol. 62, N 23.
P. 7042–7049.Р о й т А., Б р о с т о ф ф Дж., М е й л Д. Иммунология. М., 2000.19. V u k - P a v l o v i c S. // Blood Cells Mol. Dis. 2008. Vol. 40, N 1. Р. 94–100.20.
T. i. TERpinSKaJa
INFLUENCE OF TRANSPLANTATION OF BONE MARROW AND SPLEEN CELLS ON THE INOCULATION AND GROWTH RATE OF ASCITIC EHRLICH CARCINOMA IN RATS
institute of physiology of naS of Belarus, Minsk
Summary
Mice spleen and bone morrow cells at intact animals or at different terms of the subcutaneous Ehrlich carcinoma growth were obtained and intraperitoneally transplanted in a dose of 100⋅106 and 10⋅106 nuclear cells correspondingly to syngeneic animals in one hour after 100⋅105 homologous tumor cells subcutaneous inoculation. At transplantation of the intact mice splenocytes the tendency to increasing in the number of the mice with the full-grown solid Ehrlich carcinoma was revealed; transplantation of the splenocytes, received in 5 and 9 days after tumor inoculation, resulted in the statistically reliable increasing in the number of the mice with full-grown tumor. Bone marrow cells, taken on 9th and 20th tumor growth days, increased in the number of the mice with full-grown subcutaneous carcinoma as compared with animals that didn’t receive the donor mielocytes. Intact myelocyte transplantation didn’t result in reliable increasing in the number of the mice with tumor, the tendency to reduction of the tumor bearing life time was revealed.
Национальная
академия наук
Беларуси

97
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 612.015.1:618.3
н. М. КоЗловА1, л. п. КАСЬКо2, А. Г. КУТЬКо1, в. А. пЕТРович1, Т. А. СЕРЖАн3, Е. и. СлоБоЖАнинА1
АКТИВНОСТЬ ФЕРМЕНТОВ АНТИОКСИДАНТНОй ЗАЩИТЫ И ОКИСЛЕНИЕ МЕМБРАННЫХ БЕЛКОВ ЭРИТРОЦИТОВ
ПРИ НЕКОТОРЫХ ПАТОЛОГИЯХ БЕРЕМЕННОСТИ
1институт биофизики и клеточной инженерии нАн Беларуси, Минск, Беларусь, 2Белорусская медицинская академия последипломного образования, Минск,
3Родильный дом Минской области, Беларусь
(поступила в редакцию 04.01.2010)
Введение. Известно, что окислительный стресс включается в патогенез многих заболева-ний. В литературе имеются данные, что в крови беременных женщин с нормально протекаю-щей беременностью концентрация активных форм кислорода (АФК) и продуктов перекисного окисления липидов (ПОЛ) в плазме и клетках крови повышается по сравнению с их уровнем у небеременных [1], что компенсируется усилением активности основных антиоксидантных ферментов [2]. Концентрация липидных перекисей в крови беременных со сроком гестации 40 недель в 1,6 раза выше, чем у небеременных [3]. В течение беременности наблюдается по-степенное увеличение концентрации всех продуктов ПОЛ в сыворотке крови, наибольших значений эти показатели достигают к моменту родов [4]. Недавно показано [5], что при нор-мально протекающей беременности значительно понижается общая антиоксидантная актив-ность плазмы крови. Это говорит об изменении функционирования антиоксидантной системы, которое может происходить из-за снижения уровня отдельных антиоксидантов. Авторы рабо-ты [5] предполагают, что общая антиоксидантная способность организма – не просто суммар-ная активность различных антиоксидантов, а их совместное действие, которое обеспечивает наилучшую защиту от действия свободных радикалов. Существует динамическое равновесие между различными антиоксидантами. Хотя уровень отдельных антиоксидантов повышается во время беременности, суммарный результат может давать понижение антиоксидантной ак-тивности. Это относится и к патологии беременности.
Повышение интенсивности ПОЛ и снижение антиоксидантной активности может играть важную роль в развитии осложнений беременности, о чем свидетельствуют исследования по-следних лет. Так, у женщин с гестозом напряженность окислительного стресса, регистрируе-мая по динамике уровня общей оксидантной активности плазмы крови и концентрации про-дуктов ПОЛ, прогрессивно нарастает к концу 3-го триместра беременности [6, 7]. Аналогич-ные изменения наблюдаются и при беременности на фоне железодефицитной анемии: повышается уровень малонового диальдегида (МДА) в плазме крови [8], снижается уровень восстановленного глутатиона в эритроцитах [9], существенно возрастает уровень метаболитов [10]. В итоге суммарный потенциал антиоксидантной системы снижается за счет уменьшения уровня восстановленного глутатиона и активности глутатионпероксидазы (ГП) в сыворотке крови [6]. Согласно литературным данным, причиной преждевременных родов при незрелости плода и внутриутробной смертности является окислительный стресс, сопровождающийся уве-личением образования продуктов ПОЛ в плазме крови и уменьшением уровня общей антиок-сидантной активности [11]. Показано, что у беременных с внутриутробной задержкой развития
Национальная
академия наук
Беларуси

98
плода (ВЗРП) уровень МДА в плазме крови также значительно выше, чем у женщин с нор-мально протекающей беременностью [12]. Наряду с этим отмечается высокое содержание дие-новых конъюгатов и липидных гидроперекисей в мембранах эритроцитов [13]. Обнаружено увеличение протеолитической активности в эритроцитах беременных с ВЗРП, что авторы ра-боты [12] считают индикатором чрезмерного окисления белков. Все эти данные указывают на возникновение состояния окислительного стресса в клетках крови при патологии беременно-сти. Имеются сведения о том, что окислительный стресс может вносить значительный вклад и в патогенез преэклампсии. У беременных женщин с этим заболеванием наблюдается повыше-ние продукции АФК, особенно NO. и О2
.-, и повышение уровня ПОЛ в плазме и клетках крови [2, 14]. Многие исследователи рассматривают увеличение продуктов ПОЛ и уменьшение анти-оксидантной активности в плазме крови как маркеры окислительного стресса [11, 15, 16]. Од-ним из маркеров для оценки окислительного стресса in vivo могут служить параметры, харак-теризующие окисление мембранных белков [17, 18].
Цель данной работы – выяснить, могут ли активность ферментов антиоксидантной защиты и концентрация сульфгидрильных групп в мембранных белках эритроцитов являться маркера-ми окислительного стресса у женщин при патологии беременности.
Материалы и методы исследования. Клиническое обследование и наблюдение беремен-ных женщин проводилось в УЗ «Родильный дом Минской области». Обследовано три группы беременных: I – женщины с нормально протекающей беременностью при сроке гестации 38−40 недель (n = 15), II – женщины с угрозой прерывания беременности (n = 21), III – женщи-III – женщи- – женщи-ны, беременность которых осложнилась внутриутробной задержкой развития плода (согласно МКБ-ВОЗ Р05 – замедленный рост и недостаточность питания плода) при сроке гестации 30−40 недель (n = 8). В работе использованы эритроциты периферической крови и изолирован-ные из них мембраны. Эритроцитарные мембраны получали по методу Доджа и сотр. [19]. Концентрацию белка в тенях эритроцитов определяли по модифицированному методу Лоури [20], активность ГП – по методу Моина [21], супероксиддисмутазы (СОД) – непрямым спектро-фотометрическим методом, основанным на использовании реакции супероксид-зависимого окисления кверцетина [22]. Количество мембранных SH-групп оценивали двумя независимы-ми методами: спектрофотометрическим [23] и флуориметрическим (с помощью флуоресциру-ющего SH-реагента N-(1-пирен)малеимида) [24, 25]. Об изменении микровязкости липидов в мембранах эритроцитов судили по параметрам флуоресценции липофильных зондов 1-(4-триметиламмоний)-6-фенил-1,3,5-гексатриена (ТМА-ДФГ) и лаурдана, включенных в ли-пидный бислой изолированных эритроцитарных мембран. Флуориметрические измерения ТМА-ДФГ проводили аналогично методу, описанному в работе [26] (λвозб = 365 нм, λрег = 428 нм), а для лаурдана определяли величину генерализованной поляризации (GP) (λвозб = 340 нм, λрег = 440 нм и λрег = 490 нм) [27]. Флуоресцентные измерения производили на люминесцент-ном спектрофотометре �SF222 («СОЛАР», Беларусь), а спектрофотометрические – на спектро-фотометре Specord M-40 (Германия).
Результаты экспериментов анализировали методом вариационной статистики с использова-нием параметрического критерия Стьюдента.
В работе использованы следующие реагенты: ТМА-ДФГ, лаурдан (Мolecular Probes, США), трет-бутилгидроперекись (t-BHP), 5,5-дитио-бис-2-нитробензойная кислота (ДТНБ), N-(1-пирен)малеимид (ПМ) фирмы Sigma (США), NaCl, NaH2PO4, Na2HPO4 (марки ОСЧ или ХЧ, «Реахим», Россия).
Результаты и их обсуждение. Изучена активность ферментов антиоксидантной защиты ГП и СОД в эритроцитах женщин при патологии беременности. Как видно из данных, представлен-ных в табл. 1, активность СОД в эритроцитах периферической крови беременных снижалась в среднем на 15% как у женщин с угрозой прерывания беременности, так и у женщин, бере-менность которых осложнилась ВЗРП, по сравнению данными показателями крови у женщин с нормально протекающей беременностью. Активность же ГП в эритроцитах женщин с угро-зой прерывания беременности при этом повышалась на 25−35% и не отклонялась от значений, характерных для нормально протекающей беременности, в группе женщин, беременность кото-
Национальная
академия наук
Беларуси
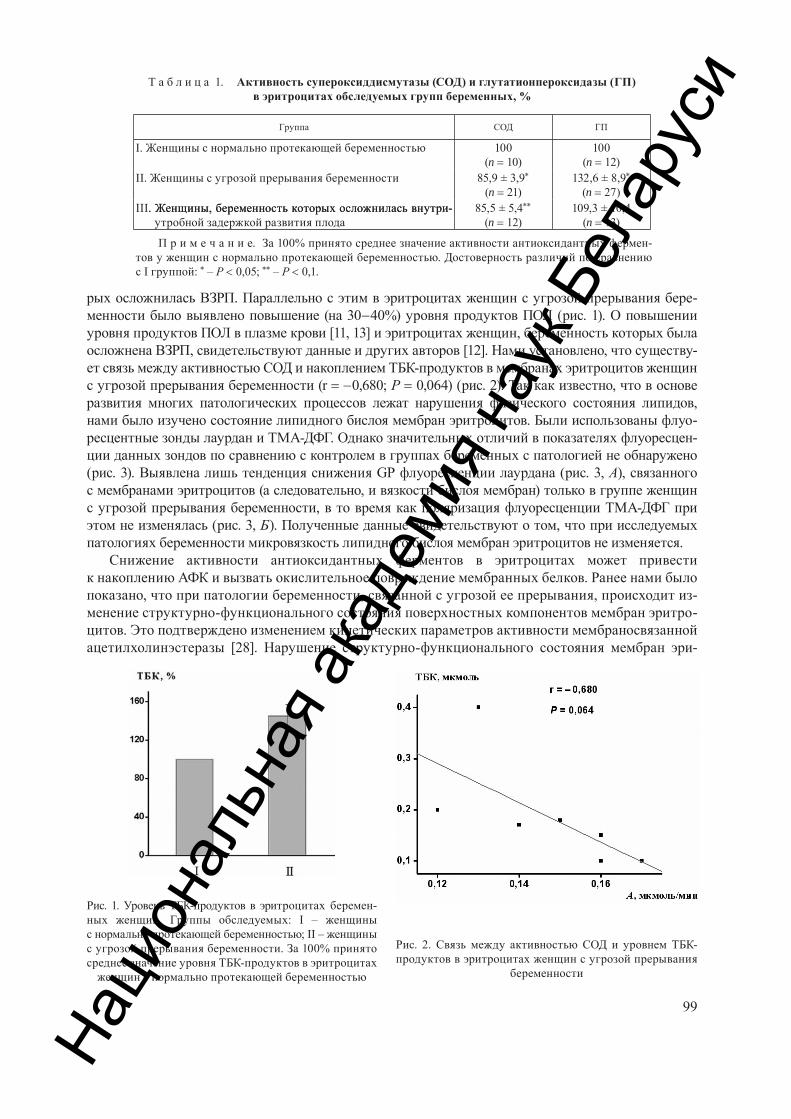
99
Т а б л и ц а 1. Активность супероксиддисмутазы (СОД) и глутатионпероксидазы (ГП) в эритроцитах обследуемых групп беременных, %
Группа СОД ГП
I. Женщины с нормально протекающей беременностью 100 (n = 10)
100 (n = 12)
II. Женщины с угрозой прерывания беременности 85,9 ± 3,9∗
(n = 21)132,6 ± 8,9∗
(n = 27)III. Женщины, беременность которых осложнилась внутри-. Женщины, беременность которых осложнилась внутри-
утробной задержкой развития плода 85,5 ± 5,4∗∗
(n = 12)109,3 ± 10,4
(n = 13)
П р и м е ч а н и е. За 100% принято среднее значение активности антиоксидантных фермен-тов у женщин с нормально протекающей беременностью. Достоверность различий по сравнению с I группой: ∗ – Р < 0,05; ∗∗ – Р < 0,1.
рых осложнилась ВЗРП. Параллельно с этим в эритроцитах женщин с угрозой прерывания бере-менности было выявлено повышение (на 30−40%) уровня продуктов ПОЛ (рис. 1). О повышении уровня продуктов ПОЛ в плазме крови [11, 13] и эритроцитах женщин, беременность которых была осложнена ВЗРП, свидетельствуют данные и других авторов [12]. Нами установлено, что существу-ет связь между активностью СОД и накоплением ТБК-продуктов в мембранах эритроцитов женщин с угрозой прерывания беременности (r = −0,680; Р = 0,064) (рис. 2). Так как известно, что в основе развития многих патологических процессов лежат нарушения физического состояния липидов, нами было изучено состояние липидного бислоя мембран эритроцитов. Были использованы флуо-ресцентные зонды лаурдан и ТМА-ДФГ. Однако значительных отличий в показателях флуоресцен-ции данных зондов по сравнению с контролем в группах беременных с патологией не обнаружено (рис. 3). Выявлена лишь тенденция снижения GP флуоресценции лаурдана (рис. 3, А), связанного с мембранами эритроцитов (а следовательно, и вязкости бислоя мембран) только в группе женщин с угрозой прерывания беременности, в то время как поляризация флуоресценции ТМА-ДФГ при этом не изменялась (рис. 3, Б). Полученные данные свидетельствуют о том, что при исследуемых патологиях беременности микровязкость липидного бислоя мембран эритроцитов не изменяется.
Снижение активности антиоксидантных ферментов в эритроцитах может привести к накоплению АФК и вызвать окислительное повреждение мембранных белков. Ранее нами было показано, что при патологии беременности, связанной с угрозой ее прерывания, происходит из-менение структурно-функционального состояния поверхностных компонентов мембран эритро-цитов. Это подтверждено изменением кинетических параметров активности мембраносвязанной ацетилхолинэстеразы [28]. Нарушение стру ктурно-функционального состояния мембран эри-
Рис. 1. Уровень ТБК-продуктов в эритроцитах беремен-ных женщин. Группы обследуемых: I – женщины с нормально протекающей беременностью; II – женщины с угрозой прерывания беременности. За 100% принято среднее значение уровня ТБК-продуктов в эритроцитах
женщин с нормально протекающей беременностью
Рис. 2. Связь между активностью СОД и уровнем ТБК-продуктов в эритроцитах женщин с угрозой прерывания
беременности
Национальная
академия наук
Беларуси

100
троцитов при патологии беременности может быть связано с окислительной деструкцией как липидов, так и белков. Учитывая разнообразную роль белков в мембранах, можно предполо-жить, что их окислительная модификация носит, в отличие от пероксидации липидов, более из-бирательный и специфический характер. Ранее было обнаружено повышение уровня карбониль-ных групп в плазме крови женщин, беременность которых была осложнена ВЗРП, что свиде-тельствовало об окислительном разрушении белков плазмы крови при данном заболевании [12]. В данной работе c помощью флуоресцентного и спектрофотометрического методов (использова-ние N-(1-пирен)малеимида и реактива Эллмана) установлено достоверное снижение уровня SH-групп белков в мембранах эритроцитов в группе женщин с угрозой прерывания беременности (табл. 2). Это свидетельствует об окислительном повреждении белков в мембранах эритроцитов женщин с угрозой прерывания беременности. Аналогичное изменение уровня SH-групп мем-SH-групп мем--групп мем-бранных белков выявлено и в эритроцитах беременных с внутриутробной задержкой развития плода (табл. 2). Как показано выше, нами обнаружено снижение активности СОД на 15−20% в эритроцитах беременных обследуемых групп с аналогичными показателями в группе женщин с нормально протекающей беременностью. Уменьшение активности СОД в эритроцитах можно рассматривать и как накопление АФК, что может приводить к окислительному повреждению мембранных белков.
Т а б л и ц а 2. Изменение параметров, характеризующих уровень SH- групп в мембранных белках эритроцитов беременных женщин
Группа iфл ПМ, % (λвозб = 342 нм, λрег = 396 нм) [SH], %
I. Женщины с нормально протекающей беременностью (n = 10) 100 100
II. Женщины с угрозой прерывания беременности (n = 21) 81,5 ± 5,2∗ 81,4 ± 3,7∗
III. Женщины, беременность которых осложнилась внутри-утробной задержкой развития плода (n = 8) 88,9 ± 2,0∗∗ 84,6 ± 0,4∗
П р и м е ч а н и е. За 100% принято среднее значение интенсивности флуоресценции N-(1-пирен)малеимида (ПМ) и концентрации SH-групп [SH] мембранных белков эритроцитов женщин с нормально протекающей беременностью. Достоверность различий по сравнению с I группой: ∗ – Р < 0,05; ∗∗ – Р < 0,1.
Рис. 3. Параметры флуоресценции липофильных зондов, включенных в мембраны эритроцитов. А – генерализован-ная поляризация (GP) флуоресценции лаурдана, встроенного в мембраны эритроцитов беременных женщин (λвозб = 340 нм, λрег = 440 нм и λрег = 490 нм); Б – поляризации флуоресценции ТМА-ДФГ, встроенного в мембраны эритроцитов беременных женщин (λвозб = 365 нм, λрег = 428 нм). Группы: I – женщины с нормально протекающей беременностью; II – женщины, беременность которых осложнена внутриутробной задержкой развития плода; III – женщины с угро- – женщины, беременность которых осложнена внутриутробной задержкой развития плода; III – женщины с угро-III – женщины с угро- – женщины с угро-
зой прерывания беременности. ∗ – Р < 0,05 по сравнению с контролем
Национальная
академия наук
Беларуси

101
Заключение. Таким образом, полученные результаты свидетельствуют, что в эритроци-тах женщин при угрозе прерывания беременности и ВЗРП наблюдаются окисление мембран-ных белков и снижение активности супероксиддисмутазы. Это указывает на возможность использования этих параметров в качестве маркеров для диагностики угрозы прерывания беременности и беременности, осложненной ВЗРП, с целью назначения антиокси дантной терапии.
Работа поддержана грантом БРФФИ № Б07–319.
Литература
1. M o r r i s J. M., G o p a u l N. K., E n d r e s e n M. J. R. et al. // Br. J. Obstet. Gynaecol. 1998. Vol. 105. P. 1195–1199.
2. S h a r m a R. K., A g a r w a l A. // Reproduct. Med. and Biol. 2004. N 3. P. 177–199.3. � o s h i k i T a k e h a r a, T a m o t s u � o s h i o k a // Acta Medica Okayama. 1990. Vol. 44, N 2. P. 103–
111.4. М и к а е л я н Н. П., М а к с и н а А. Г., К н я з е в Ю. А., М и к а е л я н А. В. // Бюл. эксперим. биол. и мед.
1997. Т. 123, № 1. С. 26–28.5. A d i g a U. S., A d i g a M. N. S. // Online J. of Health and Allied Sci. (On-line/Unpaginated). 2009. Vol. 8,
N 2.6. А к к е р Л. В., В а р ш а в с к и й Б. Я., Е л ь ч а н и н о в а С. А. и др. // Акушерство и гинекология. 2000.
№ 4. С. 17–20.7. К о л о м и й ц е в а А. Г., Ч е р н е н к о Т. С. // Акушерство и гинекология. 1986. № 4. С. 22–26.8. Б а л а б и н а Н. М. // Сиб. мед. журн. 2009. № 3. С. 41–43.9. N e e l i m a H e m k a r, S h a r m a D. C., M a n m i n d e r R i y a t // J. of Clin. Med. and Res. 2010. Vol. 2, N 2.
P. 012–014.10. Б е г о в а С. В., О с м а н о в а З. М., О м а р о в Н. С.- М. // Вопр. геникологии, акушерства и перинатоло-// Вопр. геникологии, акушерства и перинатоло-Вопр. геникологии, акушерства и перинатоло-. геникологии, акушерства и перинатоло-геникологии, акушерства и перинатоло-, акушерства и перинатоло-акушерства и перинатоло- и перинатоло-и перинатоло- перинатоло-перинатоло-
гии. 2007. Т. 6, № 3. С. 23–27.11. K a r o w i c z - B i l i n s k a A., K d z i o r a - K o r n a t o w s k a K., B a r t o s z G. // Free Radic. Res. 2007.
Vol. 41, N 8. P. 870–873.12. U l l a s K a m a t h, G u r u p r a s a d R a o, S h o b h a U. K a m a t h, � a v a n y a R a i // Ind. J. of Clin. Biochem.
2006. Vol. 21, N 1. P. 111–115.13. K a r o w i c z - B i l i n s k a A. // Ginecol. Pol. 2004. Vol. 75. P. 6–9.14. S h a a m a s h A. H., E l s n o s y a E. D., M a k h l o u f a A. M. et al. // Inter. J. of Gynecol. & Obstetrics. 2000.
Vol. 68, N 3. P. 207–214.15. � a g o d �., P a s z k o w s k i T., S i k o r s k i R., R o l a R. // Ginekol. Pol. 2001. Vol. 72, N 12. P. 1073–
1078.16. K k e n G., K i r S a h i n F., C o s a r E. et al. // Acta Obstetricia et Gynecologica Scandinavica. 2007. Vol. 86,
N 11. P. 1317–1321.17. K a r o w i c z - B i l i n s k a A., M a r s z a l e k M., K o w a l s k a - K o p r e k U. et al. // Ginekol. Pol. 2004.
Vol. 75, N 1. P. 15–20.18. К а с ь к о Л. П., К о з л о в а Н. М., К у т ь к о А. Г. и др. // Медико-социальная экология личности:
состояние и перспективы: материалы конф. Минск, 4–5 апреля 2008 г. Минск, 2008. Ч. 1. С. 138–141.19. D o d g e G. T., M i t c h e l l C., H a n a h a n D. J. // Arch. Biochem. Biophys. 1963. Vol. 100. P. 119–130.20. M a r k w e l l M. A. K., H a a s S. M., T o l b e r t N. E. // Analyt. Biochem. 1978. Vol. 87, N 2. P. 206–210. 21. М о и н В. М. // Лаб. дело. 1986. № 12. С. 724–727.22. K o s t y u k V. A., P o t a p o v i t c h A. I. // Biochem. Int. 1989. Vol. 19. P. 1117–1124.23. K i t a j i m a H., � a m a g u c h i T., K i m o t o E. // J. Biochem. 1990. Vol. 108. P. 1057−1062.24. W u C. W., � a r b r o u g h �. R., W u F. �. H. // Biochemistry. 1976. Vol. 15, N 13. P. 2863−2868.25. S h a r m a M., J o s h i P. G., J o s h i N. B. // Radiat. Phys. Chem. 1997. Vol. 49, N 1. P. 141−143. 26. S o k a l A., P u l a s k i �., R y c h l i k B. et al. // Biochem. Mol. Biol. Int. 1998. Vol. 44, N 1. P. 97–105.27. P a r a s a s s i T., D e S t a s i o G. et al. // Biophys. J. 1990. Vol. 57. P. 1179–1186.28. К а с ь к о Л. П., К о з л о в а Н. М., С л о б о ж а н и н а Е. И. и др. // Докл. НАН Беларуси. 1994. Т. 38,
№ 6. С. 81–83.
Национальная
академия наук
Беларуси

n. M. KoZlova1, l. p. KaSKo2, a. G. KuTKo1, v. a. pETRovich1, T. a. SERZhan3, E. i. SloBoZhanina1
ACTIVITY OF ANTIOXIDANT DEFENCE ENZYMES AND MEMBRANE PROTEIN OXIDATION OF ERYTHROCYTES OF WOMEN WITH SOME PATHOLOGIES OF PREGNANCY
1institute of Biophysics and cell Engineering of naS of Belarus, Minsk, 2Belarusian Medical academy of postgraduate Education, Minsk,
3Maternity hospital of Minsk region, Belarus
Summary
The decrease of superoxide dismutase in erythrocytes of pregnant women with intrauterine growth restriction and with threatened abortion was found. By means of fluorescent and spectrophotometric methods it was shown that the level of SH-groups of proteins in erythrocyte membranes in pregnant women with threatened abortion and intrauterine growth restriction reliably decreases. The results obtained show the possibility of use of these parameters in the capacity of markers for diagnos-tics of intrauterine growth restriction and threatened abortion for the purpose of prescription of antioxidant therapy.
Национальная
академия наук
Беларуси

103
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
аГлядЫ
УДК 616.12-008.331.1:614.8.026.1
Г. и. СиДоРЕнКо, М. А. ГЕРЦЕн
АКТУАЛЬНЫЕ АСПЕКТЫ ИЗУЧЕНИЯ ПРЕГИПЕРТОНИИ (РИСК, РАСПРОСТРАНЕННОСТЬ, ЛЕЧЕБНО-ПРОФИЛАКТИЧЕСКИЕ
ВМЕШАТЕЛЬСТВА)
Республиканский научно-практический центр «Кардиология», Минск, Беларусь
(поступила в редакцию 05.04.2010)
Еще в конце XIX в. выдающийся отечественный клиницист С. П. Боткин предвидел возмож-XIX в. выдающийся отечественный клиницист С. П. Боткин предвидел возмож- в. выдающийся отечественный клиницист С. П. Боткин предвидел возмож-ность повышения артериального давления (АД). Однако в то время проверить гениальную до-гадку было невозможно, ибо отсутствовали методы его измерения.
Своеобразным методическим прорывом явилось открытие Н. С. Коротковым метода прижиз-ненной диагностики АД. Его доклад, основанный на фронтовых наблюдениях, состоялся в 1905 г. Как указал в своем выступлении профессор М. В. Яновский, данным методом можно измерять не только систолическое АД (САД), но и диастолическое (ДАД).
В 1911 г. Е. Франк предложил считать «эссенциальную гипертонию» как самостоятельный диагноз.
Начиная с 1922 г. Г. Ф. Ланг и его школа разрабатывали учение о гипертонической болезни как о неврозе высших нервных отделов нейрогуморального регулирования АД [1].
Следующим важным этапом явилось изучение гипертонической болезни академиком А. Л. Мяс-никовым и его школой, в результате чего была предложена соответствующая классификация [2]. Последующие десятилетия патогенез гипертонической болезни исследовался академиком Е. И. Чазовым и его сотрудниками [3]. В дальнейшем в патогенезе гипертонической болезни изучалась роль дисфункции эндотелия [4]. Все эти этапы сопровождались коррекцией в класси-фикации гипертонии.
В течение последних десятилетий интенсивно велись исследования с целью установить со-стояния, предшествующие развитию гипертонии.
В 1939 г. S. C. Robinson и M. Brucer определили АД в диапазоне 120−139/80−89 мм рт. ст. как прегипертензивное [5]. Ими отмечено, что гипертония берет начало от прегипертонии и что у лиц с прегипертонией смертность примерно в 2 раза выше, чем у тех, чье АД ниже 120/80 мм рт. ст.
В отечественной литературе вопросы пограничной артериальной гипертонии детально раз-рабатывались академиком В. А. Алмазовым и сотр. [6]. Ими изучены патогенетические факторы, ведущие к развитию пограничной гипертонии, при этом особое внимание уделено роли бароре-цепторов. В частности, было устаовлено, какую роль играет повышение порога чувствительности аортальных и синокаротидных барорецепторов и уменьшение порога чувствительности кардио-пульмональных барорецепторов. Кроме того, была выяснена роль чувствительности адренерги-ческих и α- и β-адренергических рецепторов. Спустя несколько лет термин «прегипертония» был использован в докладе JNC 7 для обозначения АД как новой категории [7]. Потребность в таком понятии, по мнению директора Американского Национального Института (National Heart, �ung, and Blood Institute) C. �enfant, генерирована аккумуляцией критической массы ин-
Национальная
академия наук
Беларуси

104
формации. Данные эпидемиологических исследований указывают на то, что уровни давления, соответствующие этой категории, не ассоциируются с оптимальным здоровьем [8−13].
Прегипертония, вероятность развития гипертонии и неблагоприятных сердечно-сосу-дистых исходов. Согласно результатам Фремингемского кардиологического исследования, в возрасте 55 лет у людей с нормальным АД риск развития артериальной гипертонии в последу-ющие годы жизни составляет 90% [14]. Скорость трансформирования прегипертонии в гиперто-нию может быть относительно высокой. Так, у лиц с АД от 120/80 до 129/80 мм рт. ст. в возрасте 30−64 лет частота развития гипертонии в течение 4 лет составила 17,6%, в возрасте от 65 лет и старше – 25,5%, при АД 130−139/85–89 мм рт. ст. − 37,3 и 49,5% соответственно [15].
Согласно результатам исследования по профилактике артериальной гипертонии (TROPH�), включавшего 809 пациентов (средний возраст 49 лет), у 40% индивидуумов с прегипертонией в течение 2 лет развилась артериальная гипертония [16]. Среди подростков (Американская На-циональная база данных) скорость прогрессирования прегипертонии и переход ее в стадию ги-пертонии за аналогичный временной период составила 7% в год [17].
Мета-анализ 61 проспективного наблюдения миллиона человек в возрасте от 40 до 89 лет по-казал, что смертность от сердечно-сосудистых заболеваний прогрессивно повышается начиная с уровня АД 115/75 мм рт. ст. и с каждым приростом на 20/10 мм рт. ст. этот показатель удваива-ется [18].
Длившиеся в течение 18 лет исследования (NHANES и NHEFS) указывают на то, что риск сердечно-сосудистых событий (инфаркт миокарда, инсульт или сердечная недостаточность) у лиц с прегипертонией, определяемой в соответствии с JNC 7, на 31% выше, чем у имеющих нормальное давление [19]. Риск возрастает до 37% при наличии кроме прегипертонии одного и более сердечно-сосудистого фактора риска. Дальнейший анализ показал, что при высокой гра-дации прегипертонии (САД 130−139 мм рт. ст., ДАД 85−89 мм рт. ст.) вероятность сердечно-сосудистых событий еще выше − 42%, при низкой прегипертонии (САД 120–129 мм рт. ст., ДАД 80–84 мм рт. ст.) увеличение риска на 24% было статистически незначимым.
Исследование the Strong Heart Study выявило независимую связь прегипертонии при всех уровнях АД с почти двукратным сердечно-сосудистым риском [20].
Наблюдение в течение 10 лет 11 116 взрослых американцев показало, что прегипертония ас- 116 взрослых американцев показало, что прегипертония ас-116 взрослых американцев показало, что прегипертония ас-социируется с повышенным риском развития ишемической болезни сердца, инфаркта миокарда, но не инсульта [21].
У лиц среднего возраста (40−60 лет) была выявлена обратная линейная зависимость между САД (в том числе ниже 140 мм рт. ст.) и когнитивной функцией. Это позволило предположить, что цере-бральное повреждение начинается с прегипертензивных уровней АД [22]. Установлено, что преги-пертония укорачивает среднюю ожидаемую продолжительность жизни на срок до 5 лет [23−25].
Сопутствующие кардиоваскулярные факторы риска у лиц с прегипертонией. Прегипер-тония, по-видимому, ассоциируется с другими традиционными и нетрадиционными сердечно-сосудистыми факторами риска: дислипидемией, диабетом, инсулинорезистентностью, ожирени-ем, метаболическим синдромом, наличием С-реактивного белка и др. [9, 11, 26, 27]. В сравнении с нормотензивными субъектами (АД<120/80 мм рт. ст.) у лиц со 2-й стадией прегипертонии (130−139/85−89 мм рт. ст.) чаще определяются маркеры воспаления, связанные с атеросклерозом, дисфункция эндотелия, диастолическая дисфункция левого желудочка, сниженный резерв коро-нарного кровотока [28−31].
По данным the Strong Heart Study (3088 участников, средний возраст 60±8 лет), Bogalusa Heart Study (1379 участников, средний возраст 36 лет), при сравнении с нормотензивными паци-ентами лица с прегипертонией имели субклинические признаки кардиоваскулярной патологии – более высокие уровни показателей индекса массы левого желудочка, внутреннего диаметра ле-вого желудочка, толщины комплекса интима-медиа [32, 33]. По другим данным, несмотря на не-благоприятный кардиоваскулярный фенотип и наследственность по артериальной гипертонии, у молодых лиц с прегипертонией (n = 942, средний возраст 17,6 ± 3,3 года) в сравнении с нормо-тензивными субъектами статистически значимые различия структурных параметров левого же-лудочка отсутствовали [34].
Национальная
академия наук
Беларуси

105
Влияние прегипертонии в возрастном диапазоне от 18 до 35 лет ассоциировалось с более ве-роятной частотой обнаружения в последующие 20 лет жизни маркера атеросклероза – кальция в коронарных артериях [35]. Значимость этой связи сохранялась независимо от других сердечно-сосудистых факторов риска и иных характеристик (возраста, пола, дохода, образования) участ-ников исследования.
Распространенность прегипертонии. Распространенность прегипертонии, по данным иссле-дования National Health and Nutrition Examination Survey 1999−2000 гг., составила среди мужчин 40%, среди женщин − 23% [10]. Среди контингента лиц, состоящих на действительной военной службе, прегипертония, по-видимому, еще более распространена, чем в общей популяции. Так, среди многонационального контингента американских вооруженных сил (15 391 чел.), прегипер-тония была выявлена у 62% обследованных [36], среди молодых служащих сил обороны Израи-ля (36 424 чел.) − соответственно у 50,6% мужчин и 35,9% женщин [37].
В случайной выборке студентов БГУ (120 чел., средний возраст 20,4±0,5 года) прегипертен-зивный уровень АД имели 34% обследованных [38].
По данным анализа медицинской регистрации пациентов, в учреждении первичного звена здравоохранения − Центре семейной медицины при университете в американском штате Север-ная Каролина у подавляющего большинства взрослых пациентов АД не соответствовало норме [39]. Так, в группе пациентов от 35 лет и старше без документированного диагноза гипертонии или назначения антигипертензивного лечения в 40% случаев показатели АД находились в пре-делах значений, характерных для прегипертонии.
Лечебно-профилактические вмешательства для лиц с прегипертонией. Вопросы контро-ля прегипертонии остаются предметом дискуссии. В докладе JNC 7 таким лицам рекомендован выбор здорового образа жизни до достижения целевых уровней давления. Исключение состав-ляют прегипертензивные пациенты с диабетом или хроническим заболеванием почек. Им реко-мендовано медикаментозное лечение.
В отдельных исследованиях продемонстрирована эффективность диетических подходов для снижения АД у пациентов с прегипертонией. Так, в результате применения диеты, богатой фруктами и овощами, отмечалось достоверное снижение САД на 2,8 мм рт. ст., а при дополне-нии ее продуктами с низким содержанием общих и насыщенных жиров − снижение САД и ДАД на 5,5 и 3,0 мм рт. ст. соответственно [40−42]. Сокращение потребления натрия с пище-выми продуктами ассоциировалось с достоверным снижением САД с высокого до промежу-точного уровня – на 1,3−2,1 мм рт. ст. в зависимости от типа диеты, с промежуточного до низ-кого уровня – на 1,7−4,6 мм рт. ст. соответственно. В исследовании PREMIER изучалось совместное действие диеты, снижения массы тела, повышения физической активности, сниже-ния потребления натрия и алкоголя в группах пациентов с прегипертонией и гипертонией [43]. В результате многокомпонентной модификации образа жизни спустя 18 мес. у 40−48% пациен-тов с прегипертонией был достигнут уровень давления менее 120/80 мм рт. ст. Вместе с тем авторы проекта не смогли оценить влияние вмешательства на сердечно-сосудистые события из-за ограниченности исследования по времени и небольшой численности обследованного контингента.
Кроме того, недавние результаты исследования дают основание полагать, что даже при регулярном следовании рекомендациям JNC 7, касающимся образа жизни, примерно полови-JNC 7, касающимся образа жизни, примерно полови- 7, касающимся образа жизни, примерно полови-на индивидуумов со 2-й стадией прегипертонии в ближайшие 4 года станут гипертониками [44].
Изучение роли антигипертензивного медикаментозного лечения у субъектов с прегиперто-нией, в частности оценка влияния блокатора рецепторов ангиотензина – кандесартана на предот-вращение развития гипертонии у субъектов с САД 130−139 и/или ДАД 85−89 мм рт. ст., показало, что в сравнении с контрольной (плацебо) группой у лиц, принимавших в течение 2 лет кондесартан, частота развития гипертонии была ниже на две трети [16]. Спустя 2 года после отмены этого препа-рата гипертония развилась у 53% лиц, ранее принимавших кандесартан, аналогичный показатель в группе плацебо составил 63%. Таким образом, польза лекарственной терапии по предотвраще-нию гипертонии была минимальной, иными словами, 2-летний курс профилактического лечения
Национальная
академия наук
Беларуси

106
не оправдал ожиданий исследователей полностью блокировать прогрессирование прегипертонии и переход ее в стадию гипертонии.
Медицинская помощь и наблюдение лиц с прегипертонией, возможности для иннова-ций. Накопление доказательств связи пограничного АД с будущими сердечно-сосудистыми проблемами ставит вопрос в практическую плоскость. Каково клиническое применение всех этих данных [45]?
Верхние и нижние значения АД для таких пациентов, отличающиеся на 10 мм рт. ст. (САД) и 5 мм рт. ст. (ДАД), ассоциируются с различными исходами. Возможно ли устано-вить эти небольшие различия при измерении АД и насколько это удобно для применения в клинической практике, или категория прегипертонии должна применяться для определе-ния лиц, находящихся в зоне сердечно-сосудистого риска в целом? Постоянно ли различима эта категория лиц во времени, другими словами, на каком этапе у каждого конкретного па-циента определять наличие прегипертонии − на 1-й стадии гипертензии или на этапе нор-мального давления?
Какие виды врачебного вмешательства имеют значение для изменения образа жизни и ослаб-ления прегипертонии?
Утверждается, что таким пациентам необходима модификация образа жизни. Она включает снижение веса (при избыточном весе), повышение физической активности, потребления фрук-тов, овощей, умеренность в употреблении алкоголя. Такие рекомендации следует соблюдать не-зависимо от величины АД. Можно предположить, что пациенты с прегипертонией будут полу-чать больше рекомендаций. Соответственно, такие консультации потребуют дополнительного времени у занятого медицинского персонала, а также развития и совершенствования необходи-мых для этого навыков [46]. При этом последствия вмешательства точно не известны. И еще, как это сочетать с другими видами помощи, оказываемой пациенту, и каковы побочные эффекты ведения прегипертонии?
В этом отношении последние руководства передвинули фокус с вмешательства на отдельных факторах риска в направлении высокого общего риска. Это изменение предполагает, что преги-пертония будет оцениваться в сочетании с другими факторами риска или проявлениями сосуди-стого заболевания и что антигипертензивное лечение должно основываться на оценке общего кардиоваскулярного риска, а не только на определении уровней АД.
Не ясно, подобны ли профили риска оптимального давления, достигнутого с помощью фар-макологических средств и естественным образом, а также точно не известны риск и стоимость антигипертензивной терапии для достижения оптимальных уровней АД [47]. В конечном счете важно выяснить, можно ли предотвратить поражение органов-мишеней у пациентов с прегипер-тонией с помощью медикаментозного лечения. Все это служит основанием для проведения соот-ветствующих клинических исследований по данной проблеме.
Итак, введение такого понятия, как «прегипертония», имело целью сфокусировать внимание на лицах с сердечно-сосудистым риском, превышающим нормальный уровень, способствовать преодолению клинической инерции соответствующих действий в отношении пациентов с АД ниже целевых уровней [48]. Однако вызывает беспокойство такой момент, что большая доля здо-ровых лиц популяции может быть отнесена к категории нездоровых. Необходимы дальнейшие исследования для подтверждения ожидаемых выгод от идентификации лиц с прегипертонией, которым показаны лечебно-профилактические вмешательства [21].
Если потребность в «обращении» лиц с прегипертонией адресуется лечебным учреждениям, то врачи, вероятно, нуждаются в инновационных способах консультаций таких пациентов, воз-можно посредством объединения усилий врачей разных медицинских специальностей. Важным также является возможность оценки эффективности лечебно-профилактических мероприятий [49]. Таким образом, проведение дальнейших исследований с целью определения физиологиче-ских механизмов, ассоциирующихся с начальной стадией повышения АД, позволит понять, что реально «работает» на прегипертонию.
Национальная
академия наук
Беларуси

107
Литература
1. Л а н г Г. Ф. Гипертоническая болезнь. Л., 1950. 2. М я с н и к о в А. Л. Гипертоническая болезнь. М., 1954.3. Ч а з о в Е. И. // Руководство по артериальной гипертонии / Под ред. Е. И. Чазова, И. Е. Чазовой. М.,
2005.4. Г о г и н Е. Е. // Руководство по артериальной гипертонии / Под ред. Е. И. Чазова. М., 2005. С. 21−41.5. А л м а з о в В. А., Ш л я х т о Е. В., С о к о л о в а Л. А. Пограничная артериальная гипертензия. СПб.,
1992. 6. R o b i n s o n S. C. and B r u c e r M. // Arch. Intern. Med. 1939. Vol. 64. P. 409−444. 7. C h o b a n i a n A. V., B a k r i s G. �., B l a c k H. R. et al. // JAMA. 2003. Vol. 289. P. 2560−2572. 8. O s t c h e g a �., � o o n S. S., H u g h e s J., � o u i s T. Hypertension awareness, treatment, and control − Con-
tinued disparities in adults. United States, 2005−2006. NCHS Data Brief, N 3, January 2008. 9. G r e e n l u n d K. J., C r o f t J. B., M e n s a c h G. A. // Arch. Intern. Med. 2004. Vol. 164. P. 2113−2118.10. W a n g �., W a n g Q. J. // Arch. Intern. Med. 2004. Vol. 164. P. 2126−2123. 11. K a z u m i T., K a w a g u c h i A., S a k a i et al. // Diabetes Care. 2002. Vol. 25. P. 971−976.12. N e a t o n J. D., K u l l e r �., S t a m l e r J., W e n t w o r t h D. N. Hypertension, Pathophysiology, Diagnosis,
and Management. 2nd ed. / Eds. by J. H. �aragh, B. M. Brenner. N. �., 1995. 13. J u l i u s S., J a m e r s o n K., M e j i a A. et al. // JAMA. 1990. Vol. 264. P. 354−358. 14. V a s a n R. S., B e i s e r A., S e s h a r d i S. et al. // JAMA. 2002. Vol. 287. P. 1003−1010.15. V a s a n R. S., � a r s o n M. G., � e i p E. P. et al. // �ancet. 2001. Vol. 358. P. 1682−1686. 16. J u l i u s S., N e s b i t t S. D., E g a n B. M. et al. // N. Engl. J. Med. 2006. Vol. 354. P. 1685−1697.17. F a l k n e r B., G i d d i n g S., P o r t m a n R., R o s n e r B. // Pediatrics. 2008. Vol. 122. P. 238−242. 18. � e w i n g t o n S., C l a r k e R., Q I z i l b a s h N. et al. // �ancet. 2002. Vol. 360. P. 1903−1913.19. � i s z k a H. A., M a i n o u s A. G., K i n g D. E. et al. // Ann. Fam. Med. 2005. Vol. 3. P. 294−299.20. Z h a n g �., H o w a r d B., R o m a n M. et al. // J. Am. Coll. Cardiol. 2004. Vol. 43 (5 Suppl. A). P. 513A.21. Q u r e s h i A., S u r i M., K i r m a n i J. et al. // Stroke. 2005. Vol. 36. 22. K n e c h t S., W e r s c h i n g H., � o h m a n n H. et al. // Hypertension. 2008. Vol. 51. P. 663−668.23. V a s a n R. S., � a r s o n M. G., � e i p E. P. et al. // N. Engl. J. Med. 2001. Vol. 345. P. 1291−1297. 24. F r a n c o O. H., P e e t e r s A., B o n n e u x �., de � a e t C. // Hypertension. 2005. Vol. 46. P. 280−286. 25. R u s s e l l �. B., V a l i y e v a E., C a r s o n J. �. // Arch. Intern. Med. 2004. Vol. 164. P. 2119−2124. 26. C o r d e r o A., � a c l a u s t r a M., � e o n M. et al. // Am. J. Hypertension. 2006. Vol. 19. P. 189−196.27. K i n g D. E., E g a n B. M., M a i n o u s A. G., G r e e s e y M. E. // J. Clin. Hypertension. 2004. Vol. 6.
P. 562−569. 28. C h r y s o h o o u C., P i t s a v o s C., P a n a g i o t a k o s et al. // Am. J. Hypertension. 2004. Vol. 17.
P. 568−573. 29. M i l l g a r d J., H a g g A., S a r a b i M., � i n d �. // Blood Pressure. 2002. Vol. 11. P. 279−284. 30. M a r t i n i n G., R a b b i a F., G a s t a l d i �. et al. // Clin. Exp. Hypertension. 2001. Vol. 23. P. 77−87. 31. P a l o m b o C., K o z a k o v a M., M a g a g n a A. et al. // J. Hypertension. 2000. Vol. 18. P. 453−459. 32. T o p r a k A., W a n g H., C h e n W. et al. // J. Hypertension. 2009. Vol. 27. P. 243−250.33. Z h a n g �., H o w a r d B., R o m a n M. et al. // J. Am. Coll. Cardiol. 2004. Vol. 43 (5 Suppl. A). P. 513A.34. Z h u H., � a n W., g e D et al. // Am. J. Hypertension. 2007. Vol. 20 (10). P. 1051−1057. 35. P l e t c h e r M., B i b b i n s-D o m i n g o K., � e w i s C. et al. // Ann. Intern. Med. 2008. Vol. 149 (2).
P. 91−99.36. S m o l e y B. A., S m i t h N. �., R u n k l e G. P. // JABFM. 2008. Vol. 21. P. 504−511.37. G r o t t o I., G r o s s m a n E., H u e r t a M., S h a r a b i �. // Hypertension. 2006. Vol. 48. P. 254−259.38. С и д о р е н к о Г. И., А р и н ч и н а Н. Г., Г е р ц е н М. А. Артериальная гипертензия в аспекте решения
проблемы демографической безопасности: материалы V междунар. науч.-практ. конф. Витебск, 2009. С. 70−73. 39. V i e r a A. J. // JABFM. 2007. Vol. 20. P. 45−51.40. A p p e l �. J., M o o r e T. J., O b a r z a n e k E. et al. // N. Engl. J. Med. 1997. Vol. 336. P. 1117–1124. 41. S a c k s F. M., S v e t k e y �. P., V o l l m e r W. M. et al. // N. Engl. J. Med. 2001. Vol. 344. P. 3–10.42. A p p e l �. J., S a c k s F. M., C a r e y V. J. et al. // JAMA. 2005. Vol. 294. P. 2455–2464. 43. Writing Group of the PREMIER Collaborative Research Group. Effects of comprehensive lifestyle modification
on blood pressure control // JAMA. 2003. Vol. 289 (16). P. 2083–2093.44. J u l i u s S., K a c i r o t i N., N e s b i t t S. et al. // J. Am. Soc. Hypertension. 2008. Vol. 2. P. 39−43. 45. S c h u n k e r t H. // N. Engl. J. Med. 2006. Vol. 354. P. 1742−1744.46. O l i v i e r i a S., � a p u e r t a P., Mc C a r t h y B. et al. // Arch. Intern. Med. 2002. Vol. 162. P. 413−420.47. F r a n c o V., O p a r i l S., C a r r e t e r o O. // Circulation. 2004. Vol. 109. P. 2953−2959.48. P h i l i p s �. S., B r a n c h W. T., C o o k C. B. et al. // Ann. Intern. Med. 2001. Vol. 135. P. 825−834.49. С и д о р е н к о Г. И. // Кардиология. 2009. № 2. C. 73−75.
Национальная
академия наук
Беларуси

G. i. SidoREnKo, M. a. GERTSZEn
CURRENT ASPECTS OF PREHYPERTENSION STUDIES (RISK, INCIDENCE, MEDICAL AND PREVENTIVE MANIPULATIONS)
Republican Scientific-practical center “cardiology”, Minsk, Belarus
Summary
In the present article the main problems of prehypertension diagnosing are discussed; the data on the age-related prehypertension incidence are presented. The risk of age-related prehypertension complications is stated. The indications to medical and preventive manipulations and further perspectives of prehypertension studies are given.
Национальная
академия наук
Беларуси

109
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
УДК 616.8-005-072:517.938
в. в. ЕвСТиГнЕЕв1, в. А. ГоловКо2, А. С. МАСТыКин1, Е. н. АпАнЕлЬ3, о. в. КиСТЕнЬ1, С. в. лАвРЕнТЬЕвА2
НЕйРОСЕТЕВОЕ МОДЕЛИРОВАНИЕ И ТЕОРИЯ ХАОСА: ВОЗМОЖНОСТИ ПОСТРОЕНИЯ ПРОГНОЗНО-ДИАГНОСТИЧЕСКИХ МЕДИЦИНСКИХ СИСТЕМ
1Белорусская медицинская академия последипломного образования, Минск, 2Брестский государственный технический университет, Беларусь,
3научно-практический центр неврологии и нейрохирургии, Минск, Беларусь
(поступила в редакцию 17.03.2010)
В последнее время все чаще стали появляться публикации, которые посвящены разработ-кам, в рамках концепции детерминированного хаоса, моделей искусственного интеллекта (ИИ), реализованных с помощью нейросетевых технологий, а также созданию на их базе асси-стирующих медицинских прогнозно-диагностических систем. Сутью нейросетевой методики является возможность оперировать нечеткими данными, знаниями, незавершенными и проти-воречивыми выводами предыдущих исследований. Весь этот нейросетевой методологический комплекс в своем разнообразии прикладного применения объединен единой технологией – мо-делированием когнитивной деятельности мозга человека.
Понятие модели в медико-биологических исследованиях. Моделирование – это опосредо-ванное практическое или теоретическое исследование объекта, при котором непосредственно изучается не сам объект, а некоторая вспомогательная искусственная или естественная система [24]. В качестве такой системы выступают данные клинического обследования и параклиниче-ских исследований пациента, но не сам пациент как физическое лицо.
В медико-биологических исследованиях применяются в основном три вида моделирования: биологическое, физико-химическое и математическое.
Биологическое моделирование – это воспроизведение на лабораторных животных опреде-ленных патологических состояний или заболеваний, встречающихся у человека или животных.
При физико-химическом моделировании биологические структуры, функции или процессы обычно воспроизводятся физическими или химическими средствами, что, разумеется, не позволяет смоделировать то или иное биологическое явление. Так было смоделировано распространение вол-ны возбуждения по нерву с воспроизведением периода рефрактерности и закона «всё или ничего». Такие модели демонстрируют возможность воспроизведения некоторых свойств и проявлений жи-вого посредством физико-химических явлений, основанных на внешнем качественном сходстве.
Более сложные модели строятся по электротехническим и электронным схемам. Так, созда-ны механические устройства с электронным управлением, моделирующие сложные акты пове-дения («черепахи», «собаки»). В настоящее время с использованием этих схем моделирования идет интенсивное развитие робототехники.
Что касается моделирования когнитивной деятельности мозга человека, в частности про-гнозно-диагностического мышления врача, то здесь возникают уже принципиально новые труд-ности, что обусловлено сложностью биологического и физико-химического моделирования [22]. Большинство методов, которыми располагает современная экспериментальная физиология нервной системы, принципиально неприемлемо при исследовании мозга человека, а тем более при иссле-довании его когнитивной функции [9, 10]. В этом контексте надежды возлагаются на математи-ческие (точнее, формально-логические) модели, которые строятся на основе данных эксперимен-
Национальная
академия наук
Беларуси

110
та или умозрительно, на основе формализации принимаемой гипотезы или теории, требующих дальнейшей проверки, конкретизации и корректировки [1, 2, 4–8, 11–19, 22–26, 29–30, 39, 42, 45, 49–51].
В настоящее время интенсивно развивающимся направлением медико-биологического моде-лирования, основанным на нечеткой формальной логике и теории нечетких множеств, являются нечеткие стохастические модели (fuzzy stochastic models). С их помощью предпринимаются шаги по созданию модели когнитивной деятельности мозга человека. В отличие от традиционных ма-тематических вычислений с помощью чисел в нечеткой логике вводится понятие лингвистиче-ской переменной, т. е. вместо традиционного обозначения цифрами градаций качественно варьи-рующей переменной (признака) вводятся слова: «слабо выраженный», «умеренно выраженный», «сильно выраженный» и т. п. Эти понятия входят в конструкцию логически законченной фразы нечеткой логики. Например, «если имеется умеренно выраженная головная боль, то ее причиной может быть повышение внутричерепного давления». Разумеется, возможны и другие причины. Принадлежность признака к диагнозу (его релевантность) характеризуется степенью выражен-ности этой принадлежности, которая, как и вероятность, варьируется в пределах от 0 до 1,0. Тем не менее, отождествлять эти понятия математически не корректно.
Историческая справка. В процесс создания модели когнитивной деятельности мозга, полу-чивший особенно интенсивное развитие с 40-х годов прошлого века, постепенно включались различные научные направления и технические устройства: нейрокибернетика, нейроинформа-тика, нейроматематика, нейрокомпьютеры, нейрочипы, нейроускорители. Активизировались та-кие науки, как нейрофизиология, нейрореабилиталогия и ангионеврология.
Начиная с работ конца первой половины ХХ в. стали более четко обозначаться единые прин-ципы понимания и моделирования биологической когнитивной способности мозга, которые, на-конец, слились в единое обобщающее понятие – нейрокомпьютинг, формально-логический аспект которого базируется на современных представлениях об ИИ, сложившихся на основе тео-ретических исследований, представленных теоремами Колмогорова – Арнольда – Хехт-Нильсона и Б. Коско [40, 49]. Этому способствовали также выводы экспериментальных работ по невроло-гии Н. П. Бехтеревой [9]. Как дальнейшее обоснование и обобщение логических основ нейроком-пьютинга особое место занимают разработки Лотфи Заде по нечеткой формальной логике [52]. Возникло новое научное направление – математическая биология, курс лекций по которой впер-вые был прочитан в МГУ им. М. В. Ломоносова А. А. Ляпуновым.
В 1959 г. Алексей Андреевич Ляпунов предложил создать при Президиуме АН СССР Науч-ный совет по комплексной проблеме «Кибернетика», когда она все еще считалась «буржуазной лженаукой». Этот Совет возглавил А. И. Берг, А. А. Ляпунов стал его заместителем.
В нейрофизиологии и медицине данное направление связано также с именами выдающихся советских ученых – академиками Н. П. Бехтеревой, Н. М. Амосовым, Г. И. Сидоренко, профессо-рами Н. И. Моисеевой, И. М. Тонконогим и многими другими.
Считается, что создание систем искусственного интеллекта, в современном их понимании, началось непосредственно с работ Ф. Розенблатта, М. Минского, и прежде всего с введенного по-нятия фрейма [27, 33].
Фрейм (англ. frame – кадр, рамка) – это своеобразная когнитивная ячейка, содержащая какую-либо стереотипированную ситуацию. Различают фреймы-образцы, фреймы-экземпляры, фреймы-структуры, фреймы-роли, фреймы-сценарии, фреймы-ситуации. Система связанных фреймов может образовывать семантическую сеть, аналогично тому, как отдельные, разрозненные по со-держанию фотокадры объединяются единым смысловым повествованием в одну киноленту. Фреймы составляют основу экспертных интеллектуальных медицинских систем.
Можно с полной уверенностью утверждать, что интенсивная работа в области создания мо-делей искусственного интеллекта в наши дни это вторая (если не третья) волна всплеска интере-са к математическим моделям когнитивной деятельности мозга человека с целью использования их для распознания нозологических единиц в медицинской диагностике.
С конца 50-х годов прошлого века в Советском Союзе стал возрастать интерес к кибернети-ке. В те годы в области кибернетики наиболее известными были имена академиков АН СССР
Национальная
академия наук
Беларуси

111
А. И. Берга, А. А. Ляпунова, В. М. Глушкова, Н. Н. Моисеева, а также профессоров Н. И. Моисее-вой, И. М. Тонконогого и др.
В 1960–1970-е годы на кафедре нервных болезней МГМИ разрабатывались методики созда-ния таких моделей применительно к диагностике и лечению заболеваний мозга, в том числе эпи-лепсии. В дополнение к доминирующей рефлекторной теории высшей нервной деятельности предлагалась, в частности, концепция спонтанности. В результате сопоставления гипотетиче-ских алгоритмов работы мозга с работой кибернетической машины была разработана концепция двойной детерминации нервно-психической деятельности, которая еще раз подтвердила давно установленную истину, что человек не только пассивно приспосабливается к окружающим усло-виям, но и преобразует их в соответствии со своими желаниями. А это возможно лишь тогда, когда деятельность головного мозга основана не только на принципе реагирования, но и на прин-ципе инициативы. И вот этот инициативный приспособительный (адаптивный) аспект, согласно классической рефлекторной теории высшей нервной деятельности, игнорируется ее рамками [28]. В этой же публикации вопросы работы мозга затрагиваются и в энергетическом аспекте, что позволяет рассматривать его работу как диссипативную энергетическую машину, которая спонтанно детерминирует хаотизированное использование энергетических ресурсов в зависи-мости от поступающих в мозг сигналов (раздражений).
В те же годы были предприняты первые попытки использовать принципы ИИ для разработ-ки интеллектуализированного диагностического (дифференциально-диагностического) инстру-ментария, в той или иной степени моделирующего когнитивную работу мозга [26]. В настоящее время интерес к таким разработкам не ослабевает.
Тем не менее, некоторые авторы термин «искусственный интеллект» считают не совсем удач-ным для моделирования работы мозга. По их мнению, современная наука до сих пор не имеет достаточных оснований к тому, чтобы дать исчерпывающее определение понятию «естествен-ный интеллект».
Свое мнение, отличающееся от общепринятых в науке, устоявшихся на постсоветском про-странстве антропологических представлений о работе мозга, высказала академик Н. П. Бехтере-ва: «Всю свою жизнь я посвятила изучению самого совершенного органа – человеческого мозга и пришла к выводу, что возникновение такого чуда невозможно без Творца. Эволюция мозга, как ее рисовали антропологи, практически нереальна. Недаром они сейчас отказываются от многих своих данных». Более четко и полно свой взгляд на достижения в изучении мозга человека она изложила в своей книге «Магия мозга и лабиринты жизни» [10].
Проблема формализации медицинских знаний. В самом общем представлении формализа-ция, в отличие от интуитивных и/или содержательных рассуждений, есть отображение результа-тов мышления в точных понятиях или утверждениях. Формализация тесно связана с абстрактным мышлением и представляет собой его практическую реализацию. Процесс распознавания образа осуществляется по логической цепочке: органолептическое восприятие (созерцание, восприятие на слух, обоняние, осязание) – абстрактное осмысление – формализованная практическая реализа-ция. В логико-математической интерпретации формализация отображает содержательное (пред-метное) знание в виде специальных условных знаков и/или на формализованном алгоритмическом языке. В медицине особый интерес представляет формализация врачебного (клинического) ди-агноза в гармоничном соответствии с формализацией профессиональных знаний врача [22].
Не претендуя на полноту формализации медицинского диагноза вообще, обратим основное вни-мание на формализацию неврологического диагноза. Можно с полной уверенностью утверждать: пока традиционно сложившаяся не стандартизированная произвольность в определении этого поня-тия не будет помещена в строгие рамки формальной логики, проблема так и останется нерешенной.
Конфликт между определенностью и неопределенностью в математической (формальной) логике применительно к нейропатофизиологии. Согласно общей тенденции, неврологические (нейропатофизиологические, ангионеврологические) знания совершенствуются в едином русле об-щего развития научного анализа. Но, с другой стороны, для достижения общей цели необходимым условием является независимость (отделение) информации-знания от алгоритмов ее обработки. К сожалению, на сегодняшний день подобных условий для знаний не существует [48].
Национальная
академия наук
Беларуси

112
В противоположность представлениям Д. Гильберта (конец XIX в.) о завершенности матема-XIX в.) о завершенности матема- в.) о завершенности матема-тических доказательств однозначно установлено, что в любой области знаний невозможно по-лучить исчерпывающие доказательства, позволяющие судить о свойствах вещей (материи) и особенностях процессов. «Обаяние логически законченных теорий без внутреннего противоре-чия, в сущности, давно миновало» [41] вместе с «чистой» математикой Д. Гильберта, не допу-скающей противоречий, неточностей, незавершенности суждений, аксиом, теорем, постулатов. Настоящее время – это время пересмотра прошлых догматических взглядов на Природу, время обобщений накопленного фактического материала.
Всю противоречивость и неоднозначность пользы теоретических основ математики и ее роли в решении прикладных задач в свое время четко определил Ф. Энгельс, считавший, что математика – родная сестра формальной логики: «Столь безупречная некогда матема-тика, введя у себя переменные величины и распространив свойство переменности на об-ласть бесконечно-большого и бесконечно-малого, совершила грехопадение; она вкусила яблоко познания, что открыло перед ней поприще гигантских успехов, но также и заблуж-дений. В вечность кануло девственное состояние абсолютной правильности, неопровержи-мой верности всего математического; открылась эра разногласий, и мы дошли до того, что большинство людей дифференцирует и интегрирует не потому, чтобы понимали, что они делают, а руководясь чистой верой, потому что результат до сих пор всегда получался вер-ный» [44].
Разумеется, все вышеизложенное интересует нас прежде всего в прикладном аспекте – для разработки ассистирующих распознавательно-диагностических систем.
В стремлении создать всеобщую формализованную нейропатофизиологическую парадиг-му знаний, игнорируя основные положения формальной логики, мы находимся в заколдован-ном замкнутом круге. Можно, конечно, не замечать ее присутствия, но установка на непризна-ние целесообразности и полезности прикладного использования разработок по созданию ИИ – это отказ от совершенствования анализа диагностических данных и, как следствие, доб-ровольный отказ от совершенствования методов адресной профилактики и лечения в условиях все возрастающей лавины новых нейровизуализационных и других высокотехнологичных данных, охватить которые и оперативно ими пользоваться только благодаря усилиям есте-ственного интеллекта врача невозможно – нужен соответствующий высокотехнологичный ин-теллектуальный помощник.
История возникновения научной доктрины «теория хаоса». Проблема хаоса актуальна как в биологии, так и в медицине. Еще А. Пуанкаре разрабатывал теорию ИИ параллельно с раз-работкой теории хаоса. Классические теоретические исследования по разработке теории хаоса лауреата Нобелевской премии по химии И. Р. Пригожина ее особенно обострили. Здесь можно провести четкую границу между до- и послепригожинским периодами в понимании и толкова-нии понятия «хаос», в том числе применительно к донозологической диагностике транзиторных ишемических атак (ТИА) и поиску эпилептогенных очагов в структурах мозга на субклиниче-ской стадии [2–5, 7, 8, 16–19, 23, 25, 26, 29, 30, 42, 50].
Основные работы И. Р. Пригожина посвящены неравновесной термодинамике. Он обо-сновал понятие неравновесных термодинамических систем, которые «общаются» с окружа-ющей средой путем энергомассообмена, чем и обеспечивают свое существование и устойчи-вое развитие. Такие системы получили название диссипативных структур (фр. dissipation – трата, растрачивание, в данном контексте – энергии). Процесс такого диссипатизированного энергомассообмена сложной системы с окружающей ее средой не поддается описанию мето-дами классической статистики. В какие-то моменты (пункты, точки) развитие системы при-останавливается и она «раздумывает», каким путем развиваться дальше, причем то или иное решение принимается с вероятностью 50/50. И только после этих колебаний («нерешитель-ности») принимается окончательное решение. Эти «остановки раздумий» получили назва-ние точек бифуркации [32]. Данный термодинамический феномен был использован нами для объяснения явления прекондиции гипоксии-ишемии мозга при эпизодах ТИА. С этих пози-ций бифуркационных «остановок раздумий» нами была предпринята попытка объяснить
Национальная
академия наук
Беларуси

113
феномен прекондиции ишемизации мозга как возможное «предлечение». В какой-то момент мозг «решает», идти ли ему на самоуничтожение (например, по аналогии с апоптозом), или же, включив «второе дыхание» пока еще полностью не изученных триггерных механизмов, продолжать борьбу за свое существование. Это связано, по нашему мнению, с феноменом энтропии [26].
Немецкий физик Рудольф Клаузиус (1822–1888) для формулировки второго закона термо-динамики использовал понятие энтропии. Когда энтропия системы возрастает, то, соответ-ственно, усиливается беспорядок в системе. В таком случае второй закон термодинамики по-стулирует: энтропия замкнутой системы, т. е. системы, которая не обменивается с окруже-нием ни энергией, ни веществом, постоянно возрастает [20]. Это означает, что такие системы эволюционируют в сторону увеличения в них беспорядка, хаоса и дезорганизации, пока не будут достигнуты точки термодинамического равновесия, при котором всякий вид деятельности становится невозможным. Энтропия замкнутой системы, т. е. системы, которая не обменива-ется с окружением ни энергией, ни веществом, постоянно возрастает, а сама система самораз-рушается и гибнет. Идея энтропийного эволюционирования сложных диссипативных систем и связанных с ним условий выживания живого организма нашла свое отражение в современ-ных разработках эволюционных адаптивных нейросетевых и генетических алгоритмов в ан-гионеврологии [5, 17–19, 26, 51].
Проблема хаоса и неопределенности в медицинских исследованиях. Особенно интен-сивно проблема хаоса (хаотичности) дебатируется в кардиологических и психиатрических ис-следованиях. В этом аспекте ЭКГ в кардиологии – это неисчерпаемый поставщик поводов для сомнений и дискуссий. В неврологии этой теме уделяется скромное внимание, хотя в контек-сте хаотичности электроэнцефалограмма заслуживает не меньшего внимания [7, 8, 23, 50].
Вместе с тем, по мере совершенствования знаний и накопления опыта в области разработок по созданию моделей ИИ за ними неотступно следует неопределенность, которая все больше за-являет о себе. Причем влияние неопределенности на успехи и достижения в области разработок ИИ (и не только) достигло таких размеров, что вынудило ученых разных областей знаний объеди-ниться в специальную международную ассоциацию по ее преодолению – Ассоциацию анализа неопределенности в искусственном интеллекте (Association for Uncertainty in Artificial Intelligence, AUAI).
Подробное описание математического аппарата доктрины детерминированного хаоса, полу-чившей наибольшее распространение в прикладных исследованиях, приведено в монографии Г. Шустера [43].
Последовательность представлений о месте теории хаоса в кардиологии варьируется от изна-чальных сомнений в целесообразности внедрения нового сложного математического аппарата в арсенал кардиологических исследований до намерения анализировать аритмии и фибрилляции сокращений сердечной мышцы с позиций нелинейных систем и детерминированного хаоса [31, 36]. Имеются и скептические возражения. Так, по мнению H. Janse [47], «продвижение в изу-H. Janse [47], «продвижение в изу-. Janse [47], «продвижение в изу-Janse [47], «продвижение в изу- [47], «продвижение в изу-чении аритмий может быть достигнуто, если будет установлена связь между «новой» теорией хаоса и «старомодной» электрофизиологией».
Замещение упорядоченной самоорганизации сердечных сокращений детерминированной хаотизацией рассматривается уже в аспекте адаптации, поиска выхода («наугад») из сложивших-ся патологических состояний, вызывающих сбой в нормальной сократительной деятельности сердца [31, 35–38].
Такое предначертание Природы существования и развития сложных биологических систем (живых организмов, человека) в условиях нестабильности, нелинейности развития, детермини-рованного хаоса имеет конкретную практическую реализацию в лечебно-диагностической рабо-те, вынуждая подражать Ей и подстраиваться под Ее директивы в кинезотерапии при анализе физических нагрузок на сердечно-сосудистую систему. Доказана целесообразность перехода от традиционного тестирования реакции организма на физические нагрузки к стохастическому (детерминированно хаотизированному) режиму [35–38]. Сопоставление характеристик нагрузок постоянного и стохастического типа приведено в таблице.
Национальная
академия наук
Беларуси

114
Сравнительная характеристика нагрузок постоянного и стохастического типа (Г. И. Сидоренко, 2002)
Стохастический способ тестирования Традиционный способ тестирования
1. Величина нагрузки меняется по непредсказуемому за-кону. Мощность ступеней меняется случайным образом
2. Частотная характеристика нагру зочного воздействия соответствует характеристике сердечно-сосудистой си-стемы
3. Нагрузка легче переносится больными. Чередование ступеней разной мощности является динамическим от-дыхом
4. Характер нагрузки соответствует большинству быто-вых и трудовых нагрузок человека
1. Нагрузка изменяется закономерно. Динамика нагрузки предсказуема
2. Частотная характеристика нагрузочного воздействия не соответствует характеристике сердечно-сосудистой системы. Согласование отсутствует
3. Больные хуже переносят нагрузку при идентичной ве-личине суммарной мощности
4. Равномерность и предсказуемость нагрузок редко встречается в производстве и в быту
Использование физических нагрузок, меняющихся по случайной хаотичной закономерности, «впервые открывает возможность определять информационную «стоимость» нагрузок, ибо по-является перспектива оценивать физи ческие нагрузки в битах на единицу времени. Вероятно, этот критерий позволит количественно оценить нейромышечную координацию, которая всегда участвует в выполнении нагрузочных тестов» [36].
При стохастическом моделировании* предполагается случайная неточность и нечеткость с не - сколькими возможными результатами решения поставленной задачи, что, однако, не следует трактовать как неизбежную обязательную ошибку, мешающую поиску единственно нужного результата. Так, в проведении функциональных проб при сердечно-сосудистых заболеваниях стохастически меняющиеся нагрузки оказываются более физиологичными и адекватными ре-альным условиям жизнедеятельности и, следовательно, дают более точный результат проведен-ной функциональной пробы [35–38].
Если в кардиологии стохастическое моделирование хаотизированных процессов достаточно распространено, то в неврологических исследованиях интерес к ним более чем скромный.
Проведенные нами ангионеврологические исследования не выявили четкой прямой зависимо-сти степени выраженности какого-либо подтипа ТИА (атеротромботического, кардиоэмболиче-ского, гипертензивного) от уровня атеротромботических поражений стенок сосудов, эмболизации системного кровообращения, артериальной гипертензии, а кроме того, четкой гендерно-возрастной зависимости. Возможны самые различные сочетания вегетативных и кардиальных, психоэмо-циональных и других симптомов вне зависимости от пола и возраста. Этот хаос в симптомати-ческих проявлениях ТИА имитирует другие заболевания, что ведет к постановке ошибочного диагноза. Здесь самое обширное поле для ошибочной констатации кардиологических нозологий, артериальной гипертензии, диэнцефальной патологии, остеохондроза грудного и шейного отде-лов позвоночника с синдромом вертебробазилярной недостаточности, астеноневротического и климактерического синдромов и т. п. Вызывает затруднения и дифференциальная диагностика эпизодов ТИА и мигренозных состояний [3, 4, 16, 21, 28, 43]. Все это делает «чистый» детерми-нистский подход к прогнозной донозологической диагностике неактуальным, ориентируя на создание нечетких ассистирующих адаптивных систем на базе разработок по созданию искус-ственного интеллекта [1–8, 11–19, 22, 23, 25, 26, 29–31, 34, 39, 42, 45, 49–51]. Согласно концепции детерминированного хаоса, хаотичное инстинктивное неформализованное стремление пациен-та, перенесшего инсульт, восстановить нормальные движения в мелких суставах рук может быть
* Под термином «стохастическое моделирование» мы понимаем объединение нескольких разделов математики, изучающих случайные явления: детерминистскую теорию хаоса, теорию вероятностей, математическую статистику, теорию игр, теорию случайных процессов, теорию катастроф. Внимание акцентируется на нестабильных, неустойчивых, хаотизированных системах (или состояниях, в том числе диагнозах состояний), которые в любой момент могут быть подвержены качественным и/или структурным изменениям и перестройкам. В живом организме такой системой является кардиоцереброваскулярный комплекс, очень чувствительный к абнормальным влияниям на него. Прогнозирование в этом аспекте осуществляется с целью предотвращения возможных патологических качественных и/или структурных изменений и перестроек.
Национальная
академия наук
Беларуси

115
замещено более продуктивной детерминизирующей лечебно-физкультурной методикой, осно-ванной на числовой последовательности Фибоначчи [2, 3].
Все изложенное выше указывает на актуальность разработок программных и аппаратных средств для работы с неполными, неопределенными и противоречивыми данными с учетом кон-цепции детерминированного хаоса [1–8, 11–19, 22, 23, 25, 26, 29–31, 34, 39, 42, 49–51].
Прогнозная диагностика методами искусственного интеллекта (нейросетевая техноло-гия) в клинической нейропатофизиологии. В настоящее время в клинической нейропатофи-зиологии, в частности в ангионеврологии и неврологии, четко определились три основных прогнозно-диагностических направления: прогнозная диагностика первичной эссенциальной гипертензии, дифференциальная диагностика транзиторных ишемических атак, превентивная диагностика эпилептических припадков.
С целью решения этих клинических задач проявляется значительный интерес к разработкам ассистирующих прогнозно-диагностических систем (ПДС), в том числе и автоматических.
Основной вектор этих научных направлений – это разработка искусственно-интеллек-туального инструментария для прогнозирования возможности развития патологических состояний, обусловленных различными функционально-структурными нарушениями. Результат прогноза может быть желаемым или нежелательным. В большинстве случаев в клинической диагностике превалирует нежелательный прогноз, на котором акцентируется внимание врача.
Если предпочтение отдается желаемому прогнозированию саногенного события, то фактор времени (минимизация времени ожидания) актуален – скорее бы это событие наступило.
При прогнозировании нежелательного события временнóй фактор утрачивает всякую акту-альность – лишь бы оно вообще никогда не наступило (неважно, через час, день, месяц, год и т. д. до бесконечности). А в подавляющем большинстве клинико-диагностических исследований, особенно профилактических, по прогнозированию возможности возникновения патологическо-го состояния как раз превалирует категория нежелательного события, и, таким образом, фактор времени лишен практического смысла. Здесь актуален прогноз как упреждающее корректирова-ние угрожающего состояния безотносительно времени его наступления.
Таким нежелательным предсказуемым событием является возникновение эпизода ТИА, ко-торый необходимо предупредить и предотвратить независимо от фактора времени, т. е. ни от времени наступления, ни от длительности самого эпизода [2, 4, 5, 16–19, 25, 26, 42, 51]. Анало-гично – при прогнозной диагностике приступов эпилепсии [7, 8, 23, 50].
Сложилась парадоксальная ситуация. Несмотря на то что проблеме ТИА посвящено все боль-ше публикаций, разрабатываются методы лечения и профилактики, конкретизируются показа-ния к их применению, само определение этой нозологии все еще порождает дискуссии и споры.
В июне 2009 г. Американская ассоциация сердца (American Heart Association, AHA) и Амери-канская ассоциация инсультов (American Stroke Association, ASA) опубликовали в журнале Stroke научное соглашение, в котором был существенно обновлен подход к пониманию ТИА. Документ акцентирует внимание на трех аспектах: определении ТИА, риске инсульта после ТИА и опти-мальной диагностике ТИА. Данное научное соглашение рекомендовано Американской академи-ей неврологии (American Academy of Neurology) для использования неврологами США. По реко-мендации AHA/ASA под ТИА следует понимать преходящие эпизоды неврологической дис-функции, обусловленные региональной ишемией тканей головного мозга, спинного мозга или сетчатки, но не приводящие к развитию инфаркта ишемизированного участка [46].
В этом определении внимание привлекает отсутствие временного параметра, что, по нашему мнению, очень удобно для разработки прогнозно-диагностического подхода к превентивному рас-познанию этой нозологии и подавлению ее дальнейшего развития независимо от длительности эпи-зода. Лечебно-профилактические мероприятия должны проводиться неотложно (здесь и сейчас), т. е. во время обследования, без ожидания результатов высокотехнологичных параклинических обследо-ваний, которые, бесспорно, необходимы, но уже для последующих уточнений [2, 4, 5, 16–19, 42, 51].
Ряд наших публикаций посвящен прогнозной диагностике возникновения эпизодов ТИА с учетом этиопатогенетического подтипа. Причем наличие одного подтипа ТИА не исключает присутствия других. Внимание акцентируется на наиболее вероятном лидирующем подтипе
Национальная
академия наук
Беларуси

116
ТИА, однако этиопатогенетическое присутствие других подтипов также не должно упускаться из поля зрения при проведении лечебно-профилактических мероприятий. Первые исследования проводились с применением разведочных методов многомерного статистического анализа – фак-торного и дискриминантного. Были получены удовлетворительные результаты по распознава-нию подтипов ТИА на конкретном материале, но когда встал вопрос о создании ПДС для внедрения в практическую работу, то пришлось столкнуться с явлением «переобученности» (overfitting) обучающей выборки, что снижает дифференциально-диагностическую эффективность и сводит ее на нет. Наши исследования по распознаванию подтипов ТИА окончательно убедили нас в том, что нейросетевые методики лишены этого недостатка «переобученности» [4, 5, 16–19, 25, 26, 29].
Осваивая нейросетевую технологию, мы остановились на адаптивном нейросетевом класси-фикаторе (АНК) с агрегированием нескольких АНК в единый распознавательный дифферен-циально-диагностический «консилиум». Этот набор из нескольких АНК уже лишен феномена «переобученности». Для усиления распознавательной способности такого «консилиума» ему предпосылается предварительная подготовка по отбору наиболее информативных признаков-предикторов (независимых переменных) из общего исходного набора в матрице данных. Такая целенаправленная предподготовка может осуществляться с помощью эволюционного генетиче-ского алгоритма (ГА). Распознавательная (диагностическая, дифференциально-диагностическая) работа, таким образом, осуществляется путем сочетания ГА и АНК [30]. Пример успешного при-менения такого ассемблирования в ангионеврологии приводит авторский коллектив из Бахрейна и Индии [51]. Разработанная авторами классификационная диагностическая система предназна-чена для дифференциальной диагностики заболеваний, входящих в нозологию «инсультная бо-лезнь, the Stroke Disease».
В настоящее время разработаны различные методы анализа биоэлектрической активности мозга, используемые для диагностики эпилепсии. Электроэнцефалография (ЭЭГ) является од-ним из основных методов оценки функциональной активности головного мозга. Фурье-анализ, позволяющий получить усредненные показатели, имеет ряд существенных недостатков. Резуль-таты применения Вейвлет-анализа во многих случаях не дают возможности дифференцировать патологическую активность от артефактов. Нами предпринята попытка оценить сигналы ЭЭГ с помощь такой величины, которая характеризовала бы изменения функциональной активности и фиксировала бы появление аномалий за счет изменения своего значения. Для оценки подоб-ных изменений возможно применение теории хаоса и нелинейной динамики. В качестве харак-теристических величин применимы корреляционная размерность восстановления аттрактора и старший показатель Ляпунова (lmax). Нейросетевой подход позволяет вычислить значение lmax для выборок малого объема за счет обобщающей способности искусственных нейронных сетей. Для прогнозирования временных процессов нами использован многослойный персептрон (Mul-Mul-ti�ayer Perceptron, M�P). Доказано, что данная нейронная сеть способна аппроксимировать лю- Perceptron, M�P). Доказано, что данная нейронная сеть способна аппроксимировать лю-Perceptron, M�P). Доказано, что данная нейронная сеть способна аппроксимировать лю-, M�P). Доказано, что данная нейронная сеть способна аппроксимировать лю-M�P). Доказано, что данная нейронная сеть способна аппроксимировать лю-). Доказано, что данная нейронная сеть способна аппроксимировать лю-бую непрерывную функцию со сколь угодно высокой точностью. Для обучения многослойного персептрона использован алгоритм обратного распространения ошибки. Хаотичные аномаль-ные очаги в записях ЭЭГ расцениваются как маркеры возможного эпилептиформного приступа, зарождающегося в соответствующем участке мозга. Одним из условий хаотичности ЭЭГ как ди-намической системы является чувствительность к исходным значениям начальных условий. Предсказание развития траектории экспоненты осуществляется посредством нейросетевой тех-нологии. Такой подход к прогнозной диагностике приступов эпилепсии дает возможность выяв-лять аномальные участки в записях ЭЭГ, что позволяет предотвратить эти приступы до того, как они успеют развиться [7, 8, 23, 50].
На сегодняшний день можно с полной уверенностью утверждать, что в повседневной про-гнозной прогностической медицине создание прогнозно-диагностических систем на базе разра-боток искусственных интеллектуальных моделей – перспективное и актуальное направление, позволяющее совершенствовать прогнозно-диагностические мероприятия.
Своевременное выявление предпатологических и прегипертонических состояний в клиниче-ской медицине, в ангионеврологии диктует необходимость разработки соответствующих уско-ренных методов распознания в условиях цейтнота, ориентируясь на анамнестические данные
Национальная
академия наук
Беларуси

117
и данные экспресс-тестов, с последующим соответствующим быстрым лечебно-профилакти-ческим реагированием. Не вызывает сомнений актуальность разработок прогнозно-диагно-стических моделей для превентивного выявления возможных приступов эпилепсии по поиску эпилептогенных маркеров (зон, участков) на записях ЭЭГ до того, как разовьется развернутый эпилептический припадок или его эквивалент.
В этих условиях профилактические мероприятия следует проводить с акцентом на диагноз-состояние [36], который необходимо как можно быстрее установить на доклиническом донозоло-гическом этапе, а нейросетевые модели, реализованные алгоритмами и программами ассисти-рующих интеллектуальных медицинских систем, призваны как раз этому способствовать.
Литература
А в е р к и н А. Н. Нечеткие системы поддержания принятия решений. Калинин, 1989. С. 67–75.1. А п а н е л ь Е. Н., М а с т ы к и н А. С. Основы восстановления нарушенных функций после инсульта. 2.
Минск, 2005. А п а н е л ь Е. Н. // Бел. мед. журн. 2005. № 3. С. 113–115.3. А п а н е л ь Е. Н. // Мед. журн. 2008. № 2. С. 117–120.4. А п а н е л ь Е. Н. // Воен. медицина. 2009. № 2. С. 117–124.5. А с а и К., В а т а д а Д., И в а и С. и др. Прикладные нечеткие системы / Пер. с япон.; под ред. Т. Тэрано, 6.
К. Асаи, М. Сугено. М., 1993.Б е з о б р а з о в а С. В., Г о л о в к о В. А., Е в с т и г н е е в В. В. // Вестн. БрГТУ. 2008. № 5. С. 70–76.7. Б е з о б р а з о в а С. В., Г о л о в к о В. А., Е в с т и г н е е в В. В. // Современные информационные и теле-8.
медицинские технологии для здравоохранения: вторая междунар. конф. Минск, 1–3 октября, Минск, 2008. С. 133–137.Б е х т е р е в а Н. П., Б о н д а р ч у к А. В., С м и р н о в В. М., Т р о х а ч е в А. И. Физиология и патофи-9.
зиология глубоких структур мозга человека (очерки) / Под ред. Н. П. Бехтеревой. М., 1967.Б е х т е р е в а Н. П. Магия мозга и лабиринты жизни. М.; СПб., 2008.10. В и л к о в В. Г., О г а н о в Р. Г., Ш а л ь н о в а С. А. // Физиология человека. 2006. № 6. С. 33–37.11. Г о л о в к о В. А. Нейронные сети: обучение, организация и применение: учеб. пособие для вузов: в 4 кн. / 12.
Под ред. А. И. Галушкина. М., 2001.Г о л о в к о В. А. Нейросетевые методы обучения и обработки информации в системах управления и про-13.
гнозирования: автореф.... дис. д-ра техн. наук: 05.13.01. Минск, 2002. Г о л о в к о В. А., Ч у м е р и н Н. Ю., С а в и ц к и й Ю. В. // Вестн. Брест. гос. техн. ун-та. 2002. № 4. 14.
С. 66–70.Г о р б а н ь А. Н., Д у н и н - Б а р к о в с к и й В. Л., К и р д и н А. Н. и др. Нейроинформатика. Новоси-15.
бирск, 1998.Д р и в о т и н о в Б. В., А п а н е л ь Е. Н., Н о в о с е л о в а Н. А., М а с т ы к и н А. С. // Мед. журн. 2009. 16.
№ 1. С. 143–148.Е в с т и г н е е в В. В., А п а н е л ь Е. Н., Н о в о с е л о в а Н. А., М а с т ы к и н А. С. // ARS MEDICA. 17.
2009. № 3. С. 60–72. Е в с т и г н е е в В. В., А п а н е л ь Е. Н., М а с т ы к и н А. С. // Мед. журн. 2009. № 3. С. 11–119. 18. Е в с т и г н е е в В. В., А п а н е л ь Е. Н., М а с т ы к и н А. С. // Актуальные проблемы медицинской 19.
экспертизы и реабилитации больных и инвалидов: материалы междунар. конф. Минск, 20–21 мая. Минск, 2010. С. 349–355.
К л а у з и у с P. // Второе начало термодинамики. М.; Л., 1934. С. 71–158.20. К о з ю к Г. В. // Медицина. 2004. № 4. С. 63–68. 21. К о т о в Ю. Б. Методы формализации профессионального знания врача в задачах медицинской диагности-22.
ки: автореф. … дис. д-ра физ.-мат. наук: 05.13.18. М., 2002. Л а в р е н т ь е в а С. В., К и с т е н ь О. В., Г о л о в к о В. А., Е в с т и г н е е в В. В. // Новости мед.-биол. 23.
наук. 2010. № 1. С. 114–123. Л я п у н о в А. А. // Математическое моделирование жизненных процессов. М., 1968. С. 65–107.24. М а с т ы к и н А. С., Д р и в о т и н о в Б. В., А п а н е л ь Е. Н. // Бел. мед. журн. 2005. № 1. С. 18–21.25. М а с т ы к и н А. С., Д р и в о т и н о в Б. В., А п а н е л ь Е. Н. // Весцi НАН Беларусi. Сер. мед. навук. 26.
2006. № 1. С. 76–82.М и н с к и й М., П е й п е р т С. Персептроны. М., 1971.27. М и с ю к Н. С. Модели механизмов мозга человека. Минск, 1973.28. Н о в о с е л о в а Н. А. Алгоритмы построения гибридного нечеткого классификатора для анализа меди-29.
цинских данных: автореф. … дис. канд. техн. наук: 05.13.18. Минск, 2008. Н о в о с е л о в а Н. А., М а с т ы к и н А. С., Т о м И. Э. // Искусств. интеллект. 2008. № 3. С. 105–112.30. П а р х о м е н к о А. Н. // Тер. архив. 1996. № 4. С. 43–44.31. П р и г о ж и н И., С т е н г е р с И. Порядок из хаоса. М., 1986. 32. Р о з е н б л а т т Ф. Принципы нейродинамики. М., 1965.33.
Национальная
академия наук
Беларуси

Р е б р о в а О. Ю., М а к с и м о в а М. Ю., П и р а д о в М. А. // Журн. неврол. и психиатр. им. С. С. Кор-34. сакова. Инсульт (приложение). 2004. Вып. 12. С. 23–28.
С и д о р е н к о Г. И., Ш и р о к о в А. М., В о п н я р с к и й В. И. // Докл. Акад. наук БССР. 1982. Т. 26, 35. № 7. С. 665–668.
С и д о р е н к о Г. И. Творчество и медицина: поиск неочевидных решений. Минск, 2002. 36. С и д о р е н к о Г. И., Ф р о л о в А. В., К о т о в а О. В., С т а н к е в и ч В. И. // Кардиология. 2004. № 1. 37.
C. 14–19.С и д о р е н к о Г. И. // Кардиология. 2004. № 7. С. 4–9.38. С т е п а н о в а М. Д., С а м о д у м к и н С. А. Прикладные интеллектуальные системы в области медици-39.
ны: учеб.-метод. пособие. Минск, 2000.Х е х т - Н и л ь с е н Р. // Открытые системы. 1998. № 4. С. 32–37.40. У х т о м с к и й А. А. Доминанта. СПб., 2006. С. 396.41. Ш а л ь к е в и ч В. Б., М а с т ы к и н А. С., А п а н е л ь Е. Н., К о т о в а С. Г. Диагностика преходящих 42.
нарушений мозгового кровообращения. Минск, 1998.Ш у с т е р Г. Детерминированный хаос. М., 1980. 43. Э н г е л ь с Ф. Анти-Дюринг. М., 1988. С. 84–85.44. B a x t W. G. // Cancer �ett. 1994. Vol. 77, N 2–3. Р. 85–93.45. E a s t o n J. D., S a v e r J. �., A l b e r s G. W. et al. // Stroke. 2009. Vol. 40, N 6. P. 2276–2293.46. J a n s e M. // Br. Heart. J. 1992. Vol. 67. P. 3–4.47. K r a s n o p r o s h i n V., O b r a z t s o v V., V i s s i a H. // Proceed. of Intern. Conf. on Modeling and Simulation 48.
in Technical and Social Sciences (MS’2002). Girona, Spain, 25–27 June 2002. P. 267–277.K o s k o B. Fuzzy Thinking: The New Science of Fuzzy �ogic. Hyperion, 1993.49. � a u r e n t s y e v a S., G o l o v k o V., E v s t i g n e e v V. // Proceed. of the Tenth Intern. Conf. of Pattern 50.
Recognition and Information Processing (PRIP’2009). Minsk, 19–21 May. Minsk, 2009. P. 327–331.S h a n t h i D., S a h o o G., S a r a v a n a n N. // IJCSNS. 2008. Vol. 8, N 12. Р. 99–107.51. Z a d e h �. A. // Communications of the ACM. 1994. Vol. 37, N 3. P. 77–84.52.
v. v. EvSTiGnEEv1, v. a. GolovKo2, a. S. MaSTyKin1, E. n. apanEl3, o. v. KiSTEn1, S. v. lavREnTiEva2
NEURONET MODELLING AND THEORY OF CHAOS: POSSIBILITIES FOR CONSTRUCTING PROGNOSIS-DIAGNOSIS MEDICAL SYSTEMS
1Belarusian Medical academy of postgraduate Education, Minsk 2Brest State Technical university, Belarus,
3Scientific and clinical center o f neurology and neurosurgery, Minsk, Belarus
Summary
Results of research on application of neural network qualifiers within the limits of the concepts of the theory of chaos and stochastic modeling by examples of recognition of transient ischemic attacks (TIA) subtypes and revealing of epileptogenic zones in structures of the brain on EEG records are presented. Neural network classification can be put into the basis of creation of assisting intellectual prognostic/diagnostic systems on prevention of episodes ТIА at a subclinical stage of their development and to revealing the epileptogenic centers with the purpose of acceptance of prophylactic measures on preven-tion of development of full completed epileptic seizures.
Национальная
академия наук
Беларуси

119
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
ВучонЫя беларусі
ГЕОРГИй ИВАНОВИЧ СИДОРЕНКО(К 85-летию со дня рождения)
21 августа 2010 г. исполняется 85 лет известному ученому, видному клиницисту-кардиологу, лауреату Государственной премии, академику НАН Беларуси, заслуженному деятелю нау-ки, заслуженному изобретателю Республики Беларусь, доктору медицинских наук, профессору Георгию Ивановичу Сидоренко.
Георгий Иванович Сидоренко родился в Киеве в семье слу-жащего. В 1932 г. семья переехала в Минск, и с этого времени Белоруссия стала для него второй родиной. До Великой Отече-ственной войны Георгий Иванович успел окончить 9 классов. В 19 лет он добровольцем ушел на фронт, в боях в Восточной Пруссии в 1944 г. был тяжело ранен, за боевые заслуги награж-ден орденом Отечественной войны I степени и медалью «За от-I степени и медалью «За от- степени и медалью «За от-вагу». После войны окончил с золотой медалью среднюю школу.
В 1950 г. окончил с отличием Минский медицинский инсти-тут, в 1956 г. под руководством члена-корреспондента Акаде-мии наук БССР Б. И. Трусевича успешно защитил кандидатскую диссертацию. С 1952 по 1961 г. работал ассистентом, затем доцентом, с 1961 г. – заведующим кафедрой факультетской терапии Минского медицинского института. В 1968 г. Г. И. Сидо ренко была успешно защищена докторская диссертация, научным консультантом которой был выдающийся ученый-физиолог, основатель космической медицины академик В. В. Парин.
В 1968 г. по инициативе и при непосредственном участии Г. И. Сидоренко при кафедре фа-культетской терапии была создана проблемная научно-исследовательская лаборатория киберне-тических методов диагностики и биоуправления (ПНИЛ). В ПНИЛ впервые в СССР были про-ведены исследования по биоуправляемому лечению больных ишемической болезнью сердца и артериальной гипертензией, изучению психофизиологических механизмов регуляции крово-обращения, появились первые работы по разработке автоматизированных методов лечения и диагностики в кардиологии.
Признанием важности и актуальности избранного направления, новизны и оригинальности идей Г. И. Сидоренко явилось решение Государственного комитета Совета Министров СССР по науке и технике и постановление Совета Министров БССР от 10 августа 1977 г. об открытии на базе ПНИЛ одного из первых в СССР научно-исследовательских институтов кардиологии. Пер-вым директором НИИ кардиологии стал Г. И. Сидоренко.
В 1969 г. Г. И. Сидоренко было присвоено звание профессора, в 1974 г. он избран членом-корреспондентом АН БССР, в 1996 г. – академиком НАН Беларуси.
Мировая новизна и приоритетность исследований, проводимых Г. И. Сидоренко и его учени-ками, были подтверждены 162 авторскими свидетельствами и патентами на изобретение. Под руководством Г. И. Сидоренко было подготовлено и успешно защищено 76 докторских и канди-датских диссертаций.
Г. И. Сидоренко опубликовано 10 монографий и около 900 научных публикаций в ведущих рецензируемых периодических изданиях Беларуси, России, Украины, а также в европейских
Национальная
академия наук
Беларуси

и американских научных журналах. Многочисленные работы Г. И. Сидоренко были посвящены биоуправляемой фармакотерапии, изучению различных фармакологических средств, в том чис-ле в ряде кооперативных рандомизированных исследований.
Под руководством и при участии профессора Г. И. Сидоренко более чем в 250 учреждений практического здравоохранения Беларуси и государств СНГ была разработана и внедрена серия информационных диагностических систем, предназначенных для контроля и исследования на-рушений сердечного ритма и гемодинамики в различных зонах организма. Коллектив разработ-чиков (Г. И. Сидоренко, А. В. Фролов, Л. З. Полонецкий, В. В. Мирончик, Я. Г. Никитин, А. П. Воробьёв) был удостоен Государственной премии Республики Беларусь по науке и технике за 1996 г.
Совместно с НИИ физико-химических проблем при БГУ под руководством Г. И. Сидоренко был создан новый препарат – Нитрамил, на который получены европейский патент и разрешение к клиническому применению (массовое производство начато на Борисовском заводе медпре-паратов).
В 1964 г. академик Г. И. Сидоренко организовал Белорусское научное общество кардиологов. Научные связи Белорусское научное общество кардиологов поддерживает с Европейским научным обществом кардиологов (1996), Всемирной федерацией сердца (2001), Ассоциацией кардиологов СНГ. После создания Ассоциации с 1994 по 1996 г. Г. И. Сидоренко являлся ее президентом.
В течение 16 лет Г. И. Сидоренко работал экспертом в системе ВАК СССР.Последние 10 лет, после реорганизации БелНИИ кардиологии в Республиканский научно-
практический центр «Кардиология», академик Г. И. Сидоренко продолжает успешно трудиться в качестве главного научного сотрудника-консультанта, с октября 2004 г. – заведующего лабора-торией клинической патофизиологии кровообращения.
Академик Г. И. Сидоренко, один из наиболее опытных и всесторонне эрудированных клини-цистов, стоял у истоков создания кардиологической службы Беларуси, под его руководством и при непосредственном участии одними из первых в СССР были организованы кардиологиче-ские бригады и специализированная кардиологическая реанимация, начата санаторная реаби-литация кардиологических больных, получило развитие отечественное медицинское приборо-строение. Г. И. Сидоренко – единственный из врачей Беларуси, кто удостоен золотой медали Е. И. Чазова за выдающиеся достижения в области кардиологии (2000). Кроме того, Г. И. Сидо-ренко удостоен званий «Почетный кардиолог России» за заслуги в развитии кардиологической науки и организацию кардиологической службы (2001), заслуженный деятель науки Белорусской ССР (1985), заслуженный изобретатель Беларуси (1999). Академик Г. И. Сидоренко награжден орденами Трудового Красного Знамени, Отечественной войны I степени, Отечества 3-й сте-I степени, Отечества 3-й сте- степени, Отечества 3-й сте-пени, медалью «За отвагу», 14 медалями, почетными грамотами Верховного Совета БССР, пре-зидиума Верховного Совета Республики Беларусь, золотыми и серебряными медалями ВДНХ и другими знаками отличия.
Сердечно поздравляем Георгия Ивановича с 85-летием, желаем ему крепкого здоровья, сча-стья, благополучия, продолжения плодотворной научной деятельности.
отделение медицинских наук, редколлегия
Национальная
академия наук
Беларуси

121
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
ФРАНТИШЕК ИВАНОВИЧ ВИСМОНТ(К 60-летию со дня рождения)
Исполнилось 60 лет со дня рождения известного ученого в области физиологии и патологии, заведующего кафедрой Бе-лорусского государственного медицинского университета, члена-корреспондента НАН Беларуси, доктора медицинских наук, профессора Франтишка Ивановича Висмонта.
Франтишек Иванович родился 12 июня 1950 г. в г. Лида Гродненской области. После окончания школы в 1967 г. посту-пил в Минский государственный медицинский институт, кото-рый окончил с отличием в 1973 г. По окончании института Ф. И. Висмонт работал врачом-хирургом в Лидской централь-ной районной больнице, а с 1974 по 1976 г. проходил службу в рядах Вооруженных Сил СССР младшим врачом полкового медицинского пункта Белорусского военного округа.
С 1976 г. дальнейшая трудовая, научная и педагогическая деятельность Франтишка Ивановича связана с Белорусским го-сударственным медицинским университетом, где он прошел путь от младшего научного сотрудника Центральной научно-исследовательской лаборатории МГМИ до заведующего кафедрой патологической физиологии.
Основные направления научных исследований, проводимых под руководством Ф. И. Висмонта, связаны с изучением патогенетических механизмов и защитно-приспособительских реакций организма, механизмов нарушения и поддержания системных функций организма, темпера-турного гомеостаза, терморезистентности, роли эндотоксинемии в терморегуляции, в возник-новении предпатологических и патологических состояний, в патогенезе гипертермии и лихо-радочных состояний. С его участием доказана регуляторная и патогенетическая роль системы ограниченного протеолиза и эндогенных ингибиторов протеиназ в центральных механизмах ре-гуляции температуры тела, что послужило основой для формирования нового научного направле ния в термофизиологии. Ф. И. Вис монтом впервые экспериментально установлено зна-чение эндоген ных ингибиторов протеиназ в механизмах развития лихорадочных состояний. Кроме того, им доказано, что увеличение содержания ai-антитрипсина в крови является важным патоге нетическим фактором эндотоксиновой лихорадки, показана тесная взаимосвязь между функциональной активностью терморегуляторных структур мозга и уровнем в крови эн догенных ингибиторов протеиназ, синтезируемых гепатоцитами. Получены принципиаль но новые знания о регуляторной роли гепатоцитов и клеток Купфера, их детоксикационной и эндотоксинобезвре-живающей функции в механизмах формирования тиреоидного статуса организма и регуляции температуры тела. Установлено также, что депрессия клеток Купфера – важнейший фактор фор-мирования гипертиреоидного состояния организма, что имеет важное значение для понимания патогенеза целого ряда состояний, сопровож дающихся повышением температуры тела, для по-нимания механизмов саногенеза и по вышения устойчивости организма к действию эндотокси-нов и температурного фактора, для выяснения системных механизмов адаптации и компенсации при эндотоксинемии.
Франтишек Иванович Висмонт умело сочетает труд ученого с научно-организационной и пе-дагогической деятельностью, успешно продолжая традиции научной школы академика В. Н. Гурина.
Национальная
академия наук
Беларуси

Его научные достижения изложены более чем в 320 научных работах, в том числе монографиях, учебниках и учебных пособиях. Под его руководством подготовлено 8 кандидатов и 1 доктор наук.
Будучи заведующим кафедрой, Ф. И. Висмонт ведет интенсивную научно-педагогическую деятельность, много внимания уделяет подготовке врачебных кадров и совершенствованию методов преподавания в высшей школе. Им разработан и внедрен в учебный процесс метод обуче-ния, в основе создания которого – учебно-экспериментальный практикум с широким использо-ванием учебных видиофильмов и новых учебных компьютерных пособий, заменяющих демон-страции опытов на животных. Это способствует гуманизации и оптимизации учебного процесса на медико-биологических кафедрах медицинских ВУЗов с целью подготовки высококвалифици-рованных специалистов в соответствии с международными требованиями, морально-этическими принципами формирования личности будущего врача.
Франтишек Иванович ведет активную научно-организационную деятельность, являясь пред-седателем специализированного совета по защите диссертаций при БГМУ, проблемно-экспертной комиссии «Общие закономерности заболевания и выздоровления, разработка путей профилактики и патогенетической терапии», заместителем Национального комитета по биоэтике Республики Беларусь и Белорусского общества физиологов, членом Международного общества по патофи-зиологии. Им регулярно проводится работа в рамках российско-белорусского сотрудничества, при поддержке бюро ЮНЕСКО в Москве организуются обучающие семинары членов комите-тов по биоэтике стран СНГ. Ф. И. Висмонт является членом ряда ученых советов и редколле-гий журналов.
За успехи и достижения в научно-исследовательской и педагогической деятельности Ф. И. Вис-монт отмечен званием члена-корреспондента НАН Беларуси, награжден медалью Франциска Скорины, почетными грамотами Высшей аттестационной комиссии, Министерства здравоохра-нения и Министерства образования, является лауреатом стипендии Президента Республики Бела-русь в области здравоохранения и образования.
Франтишек Иванович Висмонт встречает свой юбилей с новыми творческими планами. Же-лаем ему дальнейших успехов в его многогранной научно-исследовательской и организационной деятельности, свершения всех его замыслов, крепкого здоровья на долгие годы.
отделение медицинских наук, редколлегия
Национальная
академия наук
Беларуси

123
ВЕСЦI НА Ц ЫЯ НАЛЬ НАЙ АКА ДЭМII НА ВУК БЕ ЛА РУСI № 3 2010СЕ Р ЫЯ МЕДЫЦЫНСКІХ НА ВУК
реФератЫ
УДК 612.014.482
К о н о п л я Е. Ф. , С у ш к о С. Н., М а л е н ч е н к о А. Ф., С а в и н А. О., К а д у к о в а Е. М. Оценка спонтанного и химически индуцированного мутагенеза и опухолеобразования у мышей линии Af в усло-Af в усло-в усло-
виях повышенного радиационного фона // Весцi НАН Беларусi. Сер. мед. навук. 2010. № 3. С. 5–9.
Дана оценка влияния радиационного фактора зоны отчуждения ЧАЭС на спонтанный и химически инду-цированный мутагенез и опухолеобразование лабораторных животных, экспонированных в зоне отчуждения ЧАЭС в течение 4–5 мес. Результаты исследований показали, что экспозиция лабораторных животных в зоне отчуждения ЧАЭС в течение 4 мес. способствует повышению процессов мутагенеза и канцерогенеза и изме-нению реакции организма на стандартизированное действие химического канцерогена. Исследования измене-ния противоопухолевой резистентности мышей линии Af при экспонировании в зоне отчуждения ЧАЭС в течение 5 мес. показали, что выживаемость мышей с опухолями, возникшими при подкожном введении ас-цитной аденокарциномы Эрлиха, была снижена по сравнению с аналогичным показателем у животных в усло-виях вивария. Рост опухоли в ранние сроки после прививки происходил более интенсивно у мышей, предва-рительно экспонированных в зоне отчуждения ЧАЭС.
Табл. 4. Библиогр.– 10 назв.
УДК 616.36-002+616.153.96+616-006.441
Т и т о в Л. П., С т о л я р о в а Е. А., С т о л я р о в а Т. А. Компьютерная иммунология: сравнительный анализ замен нуклеотидных последовательностей CDR и FR фрагментов VH генов иммуноглобулинов боль-CDR и FR фрагментов VH генов иммуноглобулинов боль- и FR фрагментов VH генов иммуноглобулинов боль-FR фрагментов VH генов иммуноглобулинов боль- фрагментов VH генов иммуноглобулинов боль-VH генов иммуноглобулинов боль- генов иммуноглобулинов боль-ных гепатитом С, криоглобулинемией и лимфомами // Весці НАН Беларусі. Сер. мед. навук. 2010. № 3. С. 10–18.
Центральная догма молекулярной биологии состоит в том, что перенос информации происходит по направлению ДНК – РНК – белок. Геном человека состоит из совокупности генных сегментов для каждо-го типа полипептидной цепи. Стохастическая комбинация этих сегментов обеспечивает огромное разно-образие рецепторов клеток. В случае В-лимфоцитов это разнообразие обеспечивается двумя этапами. Во-первых, многообразные В-клеточные рецепторы кодируются в зародышевой линии как генные сег-менты, которые претерпевают процесс стохастической реаранжировки, формируя конкретную конфигу-рацию гена рецептора. Во-вторых, когда триггерные факторы микроорганизмов запускают иммунный ответ, В-клетки претерпевают процессы пролиферации, клеточной гибели или мутации их иммуноглобу-линовых регионов. Уровень мутации генов иммуноглобулинов высок, что приводит к многообразию ре-цепторов на стимулированных антигеном В-клетках в раннюю фазу иммунного ответа. Тяжелая цепь В-клеточного рецептора имеет 51 гипервариабельный сегмент. В-клетки содержат фермент – дезаминазу, индуцируемую активацией (AID), которая конвертирует основание С в СG парах в U. Они концентриру-AID), которая конвертирует основание С в СG парах в U. Они концентриру-), которая конвертирует основание С в СG парах в U. Они концентриру-ются в гипервариабельном регионе перестраивающихся генов. В процессе репарации возникших повреж-дений также может произойти мутация. Некоторые из них могут генерировать сайт связывания с повы-шенной афинностью для эпитопов антигена. В результате созревания афинности образуются антитела, способные более специфично распознавать комплементарные им лиганды. Репродукция вируса гепатита С в В-лимфоцитах оказывает влияние на перестройку генов иммуноглобулинов и интенсивность возни-кающих мутаций, что может лежать в основе формирования иммунопатологических процессов.
Табл. 3. Ил. 1. Библиогр.– 28 назв.
Национальная
академия наук
Беларуси

124
УДК 616.832:612.014.41:[616.711-007.55-089
Б е л е ц к и й А. В., С о ш н и к о в а Е. В., Т е с а к о в Д. К., И л ь я с е в и ч И. А. Электрофизиологиче-ский контроль состояния функций спинного мозга в процессе хирургической коррекции сколиотиче-
ской деформации позвоночника // Весцi НАН Беларусi. Сер. мед. навук. 2010. № 3. С. 19–23.
Представлены результаты электрофизиологического исследования проводимости спинного мозга (СМ) у 82 пациентов (возраст 14–18 лет) со сколиотической деформацией позвоночника IV степени тяжести в про-IV степени тяжести в про- степени тяжести в про-цессе хирургической коррекции. Контрольную группу составили 20 здоровых лиц.
На основании регистрации и анализа соматосенсорных вызванных потенциалов (ССВП) в ответ на элек-трическую стимуляцию периферических нервных стволов установлены показатели, позволяющие объектив-но оценить функцию проводимости СМ. Электрофизиологический мониторинг по данным ССВП обеспечива-ет контроль неврологического статуса пациентов на всех этапах оперативного лечения сколиоза.
Ил. 3. Библиогр.– 23 назв.
УДК 616.832-004.2+616-056.22]:612.017.1:602.9
З а ф р а н с к а я М. М., Ф е д у л о в А. С., Н и ж е г о р о д о в а Д. Б., М о т у з о в а Я. М., К о л о б о в а М. Ю., Б а г а т к а С. С., М и л а н о в и ч Н. Ф., И в а н ч и к Г. И. Влияние мезенхимальных стволовых клеток на пролиферацию Т-клеток памяти у пациентов с рассеянным склерозом // Весцi НАН Беларусi.
Сер. мед. навук. 2010. № 3. C. 24–31.Изучена способность мезенхимальных стволовых клеток (МСК) ингибировать in vitro митоген- и миелин-
индуцированную пролиферацию эффекторных CD4+ и CD8+ Т-клеток памяти у пациентов с рассеянным склеро-зом (РС). Показано, что в периферической крови больных РС достоверно увеличено количество эффекторных СD4+ и СD8+ Т-клеток памяти. Пролиферация последних при миелин-индуцированной стимуляции in vitro достоверно превышала аналогичный показатель в контрольной группе. С помощью цитофлуориметрического метода количе-ственного анализа клеточного деления установлено, что аллогенные и аутологичные культуры МСК ингибируют антиген-неспецифическую и миелин-индуцированную пролиферацию Т-клеток памяти больных РС. Механизм МСК-индуцируемой иммуносупрессии может быть связан с остановкой клеточного деления на уровне ранних (М1–М2) генераций. Полученные результаты могут быть использованы для индивидуального подхода к оценке им-муномодулирующих свойств МСК и дальнейшего применения клеточных культур в патогенетической терапии РС.
Табл. 2. Ил. 5. Библиогр.– 22 назв.
УДК 616.72-018.3 001-003.93:617-089.855
Э й с м о н т О. Л., П а ш к е в и ч Л. А., М а л ю к Б. В., Г о л у т в и н а Н. О., Б о р и с о в А. В., Б у к а ч Д. В. Влияние перфорации остеохондральной пластинки на регенерацию поврежденного суставного хряща
в эксперименте // Весці НАН Беларусі. Сер. мед. навук. 2010. № 3. С. 32–37.
Представлены результаты изучения процессов хондро- и остеорегенерации при лечении эксперименталь-ного полнослойного повреждения суставного хряща коленных суставов у собак методами перфорации подле-жащей субхондральной кости. Установлено, что при глубокой туннелизации по Pridie костной ткани в зоне дефекта происходят процессы дифференцировки стволовых клеток в хрящевые клеточные элементы и в фи-бробласты, продуцирующие волокнистые структуры. По периферии зоны туннелизации наблюдается актив-ная пролиферация клеточных элементов и кровеносных сосудов, что способствует улучшению тканевого питания и процессам репаративной регенерации костной ткани.
Ил. 8. Библиогр.– 17 назв.
УДК 612.014.3:616-006.48:57.085.23+577.152.34:577.112.382.2
Б а л а ш е в и ч Т. В., Н и к а н д р о в В. Н. Особенности состояния клеток глиомы С6 в культуре при совместном воздействии плазминогена и глицина // Весці НАН Беларусі. Сер. мед. навук. 2010. № 3. С. 38–41.
Исследовано влияние плазминогена на содержание ДНК, РНК, общего белка и морфофункциональное со-стояние так называемых «дифференцирующихся» клеток (экспозиция 6 сут на дефицитной по белкам крови пи-тательной среде) культуры глиомы С6 на фоне глицина. «Дифференцирующиеся» культуры глиомы С6 характе-ризуются довольно слабым изменением уровней белка и нуклеиновых кислот в ответ на воздействие зимогена в сравнении с культурами «незрелых» клеток. Эффект зимогена существенно изменяется в присутствии глици-на, принципиально отличаясь от эффекта каждого из исследуемых соединений. Совместное действие плазмино-гена (10–7 М) с глицином (0,01 или 0,1 мМ) в течение 72 ч вело к увеличению содержания нуклеиновых кислот и общего белка в клетках, 96%-ной выживаемости клеток глиомы С6 в условиях депривации сыворотки.
Ил. 2. Библиогр.– 13 назв.
Национальная
академия наук
Беларуси

125
УДК 615.227.3:616-092.9
С е р е д а В. В., О д и н ц о в С. Г., В л а с о в А. П., Ф и м а Д. В., Ш у б е н о к Д. В., К р а в ч у к З. И., К о ж у х Г. В., Ш м а н а й В. В., Д е е в С. М., М а р ц е в С. П. Получение модифицированного полиэтилен-гликолем иммунотоксина scFv4D5-барназа, специфичного к онкогену с-ErbB2, и сравнительное исследова-
ние его структурно-функциональных параметров // Весцi НАН Беларусi. Сер. мед. навук. 2010. № 3. С. 42–47.
С использованием N-оксисукцинимидного эфира ПЭГ-5000 синтезирован ПЭГилированный рекомби-нантный иммунотоксин scFv-барназа, специфичный к онкогену-маркеру с-ErbB2, при этом 80% продуктов реакции составляют моно- и диПЭГилированные производные.
Методами флуоресцентной спектроскопии и сканирующей калориметрии показано, что как исходный, так и ПЭГилированный иммунотоксины обладают компактной третичной структурой с двумя пиками тепло-поглощения на термограмме, отражающими кооперативное «плавление» антигенсвязывающего scFv фраг-мента и бактериальной РНКазы (барназы) без видимых изменений третичной структуры в результате модифи-кации. ПЭГилированный иммунотоксин способен эффективно связываться с с-ErbB2-позитивными клетками, причем константа связывания сопоставима с константой связывания «родительского» полноразмерного анти-тела – герцептина. Результаты доказывают возможность ПЭГилирования иммунотоксина для улучшения его фармакокинетики как одного из свойств, важных в клиническом применении.
Ил. 2. Библиогр.– 16 назв.
УДК 616.314-74
Н о в а к Н. В. Состояние границы «зуб – пломба» при использовании различных реставрационных ма-териалов // Весцi НАН Беларусi. Сер. мед. навук. 2010. № 3. С. 48–51.
В статье приведены данные предклинических испытаний четырех образцов нового пломбировочного ма-териала белорусского производства в сравнении с материалом импортного производства. Проведено микро-скопическое исследование границы «зуб – пломба».
Табл. 1. Библиогр.– 6 назв.
УДК 616.127:616-076.4
Н о в а к о в с к а я С. А., К о з л о в с к а я Е. В., Р ы ж к о в с к а я Е. Л., М а н е е в а О. А., А р ч а к о в а Л. И. Ультраструктурные изменения в миокарде при дилатационной кардиомиопатии // Весцi НАН Беларусi.
Сер. мед. навук. 2010. № 3. С. 52–58.
Электронно-микроскопические исследования эндомиокардиальных биоптатов пациентов с дилатацион-ной кардиомиопатией показали, что в сердечных миоцитах таких больных развивается апоптотический про-цесс. Характерными признаками ультраструктурной реорганизации миокарда являются литические измене-ния миофибрилл, элиминация части кардиомиоцитов путем диффузного апоптоза и формирование замести-тельного фиброза. Данный процесс сопровождается нарушением трофической функции микроциркуляторного русла сердца, что обусловлено патологией эндотелия и сдавлением капиллярной сети коллагеновыми фибрил-лами. Выявленные ультраструктурные изменения свидетельствуют о прогрессирующем нарушении микро-циркуляции в сердечной мышце и развитии ишемической кардиомиопатии.
Ил. 4. Библиогр.– 30 назв.
УДК 616-006.484.03:577.17
Ч е р н о в А. Н. Морфофункциональное состояние клеток глиомы С6 в культуре при действии рекомби-нантного фактора роста нервов // Весці НАН Беларусі. Сер. мед. навук. 2010. № 3. С. 59–62.
Изучено действие β-формы рекомбинантного ФРН (100 и 200 нг/мл) на ряд морфологических (размеры, длина и арборизация отростков) и физиологических (индекс пролиферации и гибель) параметров глиомы С6, культивируемой in vitro в течение недельного срока. Показано, что β-ФРН в указанных дозах подавлял индекс пролиферации глиоцитов на 3-и (P < 0,01) и 7-е (P < 0,05) сутки культивирования, а кроме того, к 7-м суткам вызывал уменьшение арборизации (P < 0,05) и элонгации (P < 0,01) отростков клеток глиомы. Таким образом, ФРН в указанных дозах обладает антимитогенной активностью на культуры клеток С6.
Табл. 2. Ил. 2. Библиогр.– 15 назв.
Национальная
академия наук
Беларуси

126
УДК 616.441-006.6-089.87:612.015
М и т ю к о в а Т. А., Л е о н о в а Т. А., Т у з о в а А. А., П л а т о н о в а Т. Ю., Л у щ и к М. Л., К у л а - к о в с к а я Л. Г., Д р о з д В. М. Биохимические показатели крови у пациентов с карциномой щитовид-ной железы после тотальной тиреоидэктомии // Весцi НАН Беларусi. Сер. мед. навук. 2010. № 3. С. 63–69.
Изучены особенности метаболического статуса пациентов с тотальной тиреоидэктомией (диагноз «дифференцированный рак щитовидной железы»). Выявлены некоторые неблагоприятные сдвиги биохи-мических показателей у пациентов общей группы, особенно у лиц с некомпенсированным послеоперацион-ным гипотиреозом. Показано, что наиболее важным практическим вопросом для больных с проявлениями гипопаратиреоза является компенсация нарушенного метаболизма кальция, что делает необходимым регу-лярный контроль уровня общего и ионизированного Са в крови таких пациентов и назначение им адекват-ных препаратов.
Табл. 3. Ил. 4. Библиогр.– 10 назв.
УДК 616.018.51+616.018.54]:541.515
Н о в и к о в а И. А., Я р е ц Ю. И. Взаимосвязь степени окисленности плазмы и эритроцитов в условиях активации свободнорадикального окисления // Весці НАН Беларусі. Сер. мед. навук. 2010. № 3. С. 70–74.
Изучены особенности распределения продуктов липопероксидации в плазме и эритроцитах в норме и в условиях активации свободнорадикального окисления на примере локальной гранулирующей раны. При интенсификации перекисного окисления липидов выявлены различия в накоплении липопероксидов между плазмой и эритроцитами, тогда как у здоровых лиц степень окисленности плазмы и эритроцитов одинако-вая. Наибольшие различия установлены в содержании вторичных и конечных продуктов окисления фосфо-липидов – сопряженных триенов и оснований Шиффа. Проанализирована взаимосвязь между показателями окисленности плазмы и мембран эритроцитов и ее динамика у больных с различным исходом оперативного вмешательства.
Табл. 1. Ил. 2. Библиогр.– 18 назв.
УДК 616.24-018-02:613.84:547.914.3/.5
Д е в и н а Е. А., П р и н ь к о в а Т. Ю., Т а г а н о в и ч А. Д. Особенности изменения функциональной активности альвеолярных макрофагов при контакте с сигаретным дымом // Весцi НАН Беларусi. Сер.
мед. навук. 2010. № 3. С. 75–80.
Изучено влияние экстракта сигаретного дыма (ЭСД) с различной концентрацией смол на жизнеспособ-ность альвеолярных макрофагов (АМ), фагоцитарную активность и генерацию оксида азота этими клетками. Установлено, что снижение жизнеспособности и фагоцитарного показателя АМ напрямую зависит от концен-трации смол в ЭСД, контактирующих с клетками в течение 1 и 24 ч. При этом среднее количество микробных тел, поглощаемых одной фагоцитирующей клеткой, снижалось вне зависимости от концентрации ЭСД и вре-мени экспозиции. Отмечено значительное снижение концентрации нитрит-ионов в АМ и среде их инкуба-ции под влиянием ЭСД.
Табл. 1. Ил. 2. Библиогр.– 14 назв.
УДК 612.57:612.015.1:612.828
Б о ч а р о в а В. Н. , С а в ч и н а Е. Н. Гистохимический анализ активности NADPH-диафоразы, аце-тилхолинэстеразы и ферментов энергетического обмена в структурах продолговатого мозга крысы при
лихорадке // Весцi НАН Беларусi. Сер. мед. навук. 2010. № 3. С. 81–85.
В острых опытах на наркотизированных крысах показано, что после внутрибрюшинного введения липо-полисахарида E. coli (5 мкг/кг) в нейронах дорсального моторного ядра блуждающего нерва возрастает актив-ность NADPH-диафоразы, лактатдегидрогеназы, сукцинатдегидрогеназы и ацетилхолинэстеразы (определе-NADPH-диафоразы, лактатдегидрогеназы, сукцинатдегидрогеназы и ацетилхолинэстеразы (определе--диафоразы, лактатдегидрогеназы, сукцинатдегидрогеназы и ацетилхолинэстеразы (определе-ние с помощью цитофотометрических способов). Установлено, что введение ингибитора синтеза NO-синтазы �-NAME (25 мг/кг, внутрибрюшинно) предотвращает развитие лихорадочной реакции, снижает активность
Национальная
академия наук
Беларуси

127
ферментов энергетического обмена в нейронах и интенсивность синтеза NO. Полученные данные позволяют предположить, что NO- и холинергические структуры этого ядра участвуют в системных реакциях организма по поддержанию температурного гомеостазиса.
Ил. 3. Библиогр.– 20 назв.
УДК 612.8.015
Т р о п н и к о в а Г. К. Сравнительный анализ протекторных эффектов интервальной гипоксии и мела-тонина на серотонинергические структуры передних отделов мозга крысы // Весцi НАН Беларусі. Сер.
мед. навук. 2010. № 3. С. 86–90.
Прекондиционирование интервальной гипобарической гипоксией (2300 м над уровнем моря, 3 cеанса по 1,5 мин c 1,5-минутными интервалами в день, 7 дней) способствовало активации гиппокампальных и корко-c 1,5-минутными интервалами в день, 7 дней) способствовало активации гиппокампальных и корко- 1,5-минутными интервалами в день, 7 дней) способствовало активации гиппокампальных и корко-вых серотонинергических структур мозга в условиях действия сплошной 10-минутной гипоксии. Дополни-тельное введение экзогенного мелатонина перед каждым сеансом интервальной гипоксии индуцировало в условиях непрерывной гипоксии избыточное накопление серотонина в коре головного мозга, снижение со-держания 5-оксииндолуксусной кислоты в гиппокампе и коре больших полушарий мозга и сопровождалось угнетением активного поведения у животных.
Табл. 2. Библиогр.– 25 назв.
УДК 616.006.02.009.097
Т е р п и н с к а я Т. И. Влияние трансплантации клеток костного мозга и селезенки на прививаемость и скорость роста асцитной карциномы Эрлиха у мышей // Весці НАН Беларусі. Сер. мед. навук. 2010.
№ 3. С. 91–96.
У интактных мышей или мышей с развивающейся подкожно карциномой Эрлиха на различных этапах опухолевого роста получали клетки селезенки и костного мозга и трансплантировали их сингенным живот-ным внутрибрюшинно в дозе 100 млн и 10 млн ядросодержащих клеток на мышь соответственно через 1 ч после подкожной прививки им 100 тыс. клеток гомологичной опухоли. При трансплантации спленоцитов ин-тактных доноров наблюдалась тенденция к усилению прививаемости карциномы Эрлиха; трансплантация спленоцитов, полученных на 5-е и 9-е сутки роста карциномы Эрлиха у доноров, вела к статистически достоверно-му усилению прививаемости этой опухоли у реципиентов. Транплантация клеток костного мозга, полученных у мышей на 9-е и 20-е сутки опухолевого роста, также увеличивала прививаемость подкожной опухоли по сравнению с этим показателем у животных, не получивших донорских миелоцитов; при трансплантации кле-ток костного мозга интактных мышей статистически значимого повышения прививаемости не зарегистриро-вано. Выявлена тенденция к сокращению продолжительности жизни опухоленосителей, получивших миело-циты интактных доноров.
Ил. 3. Библиогр.– 20 назв.
УДК 612.015.1:618.3
К о з л о в а Н. М., К а с ь к о Л. П., К у т ь к о А. Г., П е т р о в и ч В. А., С е р ж а н Т. А., С л о б о ж а - н и н а Е. И. Активность ферментов антиоксидантной защиты и окисление мембранных белков эри-троцитов при некоторых патологиях беременности // Весцi НАН Беларусi. Сер. мед. навук. 2010.
№ 3. С. 97–102.
Изучены активность ферментов антиоксидантной защиты (глутатионпероксидазы и супероксиддисмутазы), физическое состояние липидов в мембранах эритроцитов и концентрация сульфгидрильных групп в мембран-ных белках эритроцитов женщин с патологией беременности. Обнаружено снижение активности супероксид-дисмутазы в эритроцитах женщин при угрозе прерывания беременности и у беременных с внутриутробной задержкой развития плода (ВЗРП). Показано, что микровязкость мембранных липидов в эритроцитах обследо-ванных не изменяется. С помощью флуоресцентного и спектрофотометрического методов установлено досто-верное снижение уровня SH-групп белков в мембранах эритроцитов женщин с данными патологиями.
Полученные результаты указывают на возможность использования этих параметров в качестве маркеров для диагностики угрозы прерывания беременности и беременности, осложненной ВЗРП, с целью назначения антиоксидантной терапии.
Табл. 2. Ил. 3. Библиогр.– 28 назв.
Национальная
академия наук
Беларуси

УДК 616.12-008.331.1:614.8.026.1
С и д о р е н к о Г. И., Г е р ц е н М. А. Актуальные аспекты изучения прегипертонии (риск, распростра-ненность, лечебно-профилактические вмешательства) // Весці НАН Беларусі. Сер. мед. навук. 2010.
№ 3. С. 103–108.
В статье излагаются вопросы становления диагноза прегипертонии; приведены данные о распространен-ности риска осложнений прегипертонии в возрастном аспекте. Определены показания к лечебно-профи-лактическому вмешательству и дальнейшие перспективы изучения прегипертонии.
Библиогр.– 49 назв.
УДК 616.8-005-072:517.938
Е в с т и г н е е в В. В., Г о л о в к о В. А., М а с т ы к и н А. С., А п а н е л ь Е. Н., К и с т е н ь О. В., Л а в- р е н т ь е в а С. В. Нейросетевое моделирование и теория хаоса: возможности построения прогнозно-
диагностических медицинских систем // Весцi НАН Беларусi. Сер. мед. навук. 2010. № 3. С. 109–118.
Приведены результаты исследования по применению нейросетевых классификаторов в рамках концеп-ций теории хаоса и стохастического моделирования на примерах распознавания подтипов транзиторных ише-мических атак (ТИА) и выявления эпилептогенных зон в структурах мозга по записям ЭЭГ. Нейросетевая классификация может быть положена в основу создания ассистирующих интеллектуальных прогнозно-диагностических систем по предотвращению эпизодов ТИА на субклинической стадии их развития и выявле-нию эпилептогенных очагов с целью принятия превентивных мер по предотвращению развития развернутого эпилептического приступа.
Табл. 1. Библиогр.– 52 назв.
Национальная
академия наук
Беларуси


















