
Dr Gihan E-H Gawish, MSc, PhD Molecular Genetics & Clinical Biochemistry KSU Cell Cycle Control,...
21
Dr Gihan E-H Gawish, MSc, PhD Molecular Genetics & Clinical Biochemistry KSU Cell Cycle Control, Defects and Apoptosis 1 st Lecture
-
Upload
reynaldo-streit -
Category
Documents
-
view
221 -
download
1
Transcript of Dr Gihan E-H Gawish, MSc, PhD Molecular Genetics & Clinical Biochemistry KSU Cell Cycle Control,...

Dr Gihan E-H Gawish, MSc, PhDMolecular Genetics & Clinical Biochemistry
KSU
Cell Cycle Control, Defects and Apoptosis
1st Lecture

2
In vertebrates and diploid yeasts,
cells in G1 have a diploid number of
chromosomes (2n), one inherited
from each parent.
In haploid yeasts, cells in G1 have
one of each chromosome (1n).
Rapidly replicating human cells
progress through the full cell cycle
in about 24 hours: mitosis takes ≈30
minutes; G1, 9 hours; the S phase, 10
hours; and G2, 4.5 hours. In contrast,
the full cycle takes only ≈90 minutes
in rapidly growing yeast cells.
Cell-Cycle Control in Mammalian Cells
The Cell Cycle Is an Ordered Series of Events Leading to Replication of Cells

3
Dr Gihan Gawish
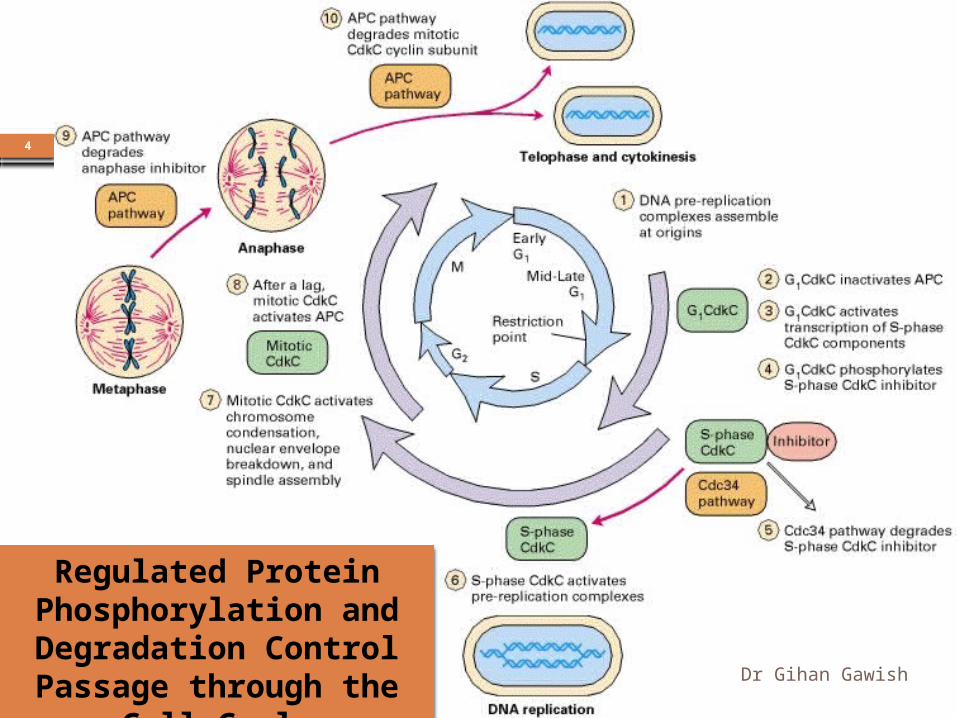
the complex macromolecular events of the eukaryotic cell cycle are regulated by a small number of heterodimeric protein kinases.
Passage through three critical cell-cycle transitions, is irreversible because these transitions are triggered by the regulated degradation of proteins, an irreversible process. As a consequence, cells are forced to traverse the cell cycle in one direction only.
Regulated Protein Phosphorylation and Degradation Control Passage through the Cell Cycle
4
Dr Gihan Gawish
Regulated Protein Phosphorylation and
Degradation Control Passage through the Cell Cycle

5 Amphibian and invertebrate eggs and early embryos
from synchronously fertilized eggs provide sources of
extracts for biochemical studies of cell-cycle events.
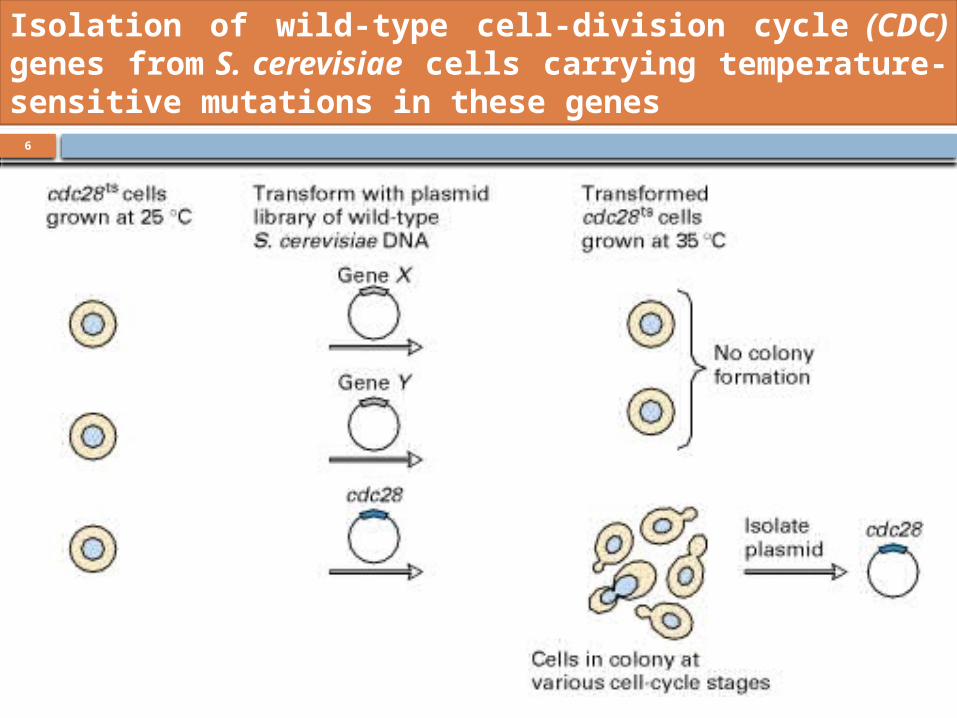
The isolation of yeast cell-division cycle (cdc) mutants
led to the identification of genes that regulate the cell
cycle
Diverse Experimental Systems Have Been Used to Identify and Isolate Cell-Cycle Control Proteins
6
Dr Gihan Gawish
Isolation of wild-type cell-division cycle (CDC) genes from S. cerevisiae cells carrying temperature-sensitive mutations in these genes

7
In multicellular organisms, cell replication is controlled by a complex
network of signaling pathways that integrate signals from the extracellular
environment with intracellular cues about cell size and developmental
program.
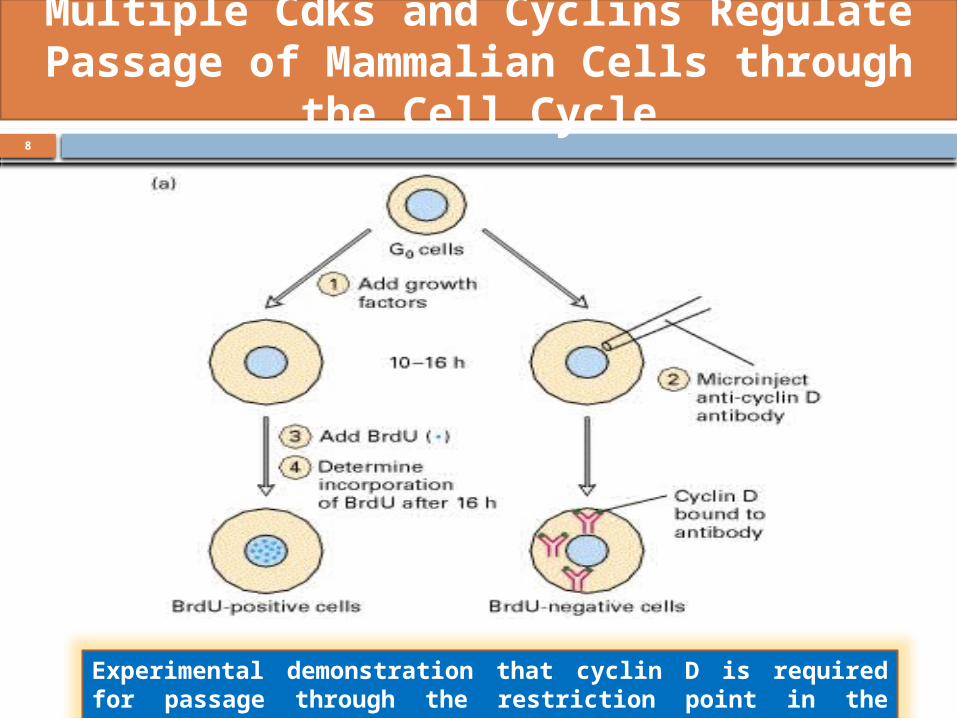
Polypeptide growth factors called mitogens stimulate cultured
mammalian cells to cycle. Once cycling cells pass the restriction point,
they can enter the S phase and complete S, G2, and mitosis in the
absence of growth factors.
Mammalian Restriction Point is Analogous to start in Yeast Cells
8
Multiple Cdks and Cyclins Regulate Passage of Mammalian Cells through
the Cell Cycle
Experimental demonstration that cyclin D is required for passage through the restriction point in the mammalian cell cycle
9
Multiple Cdks and Cyclins Regulate Passage of Mammalian Cells through
the Cell Cycle
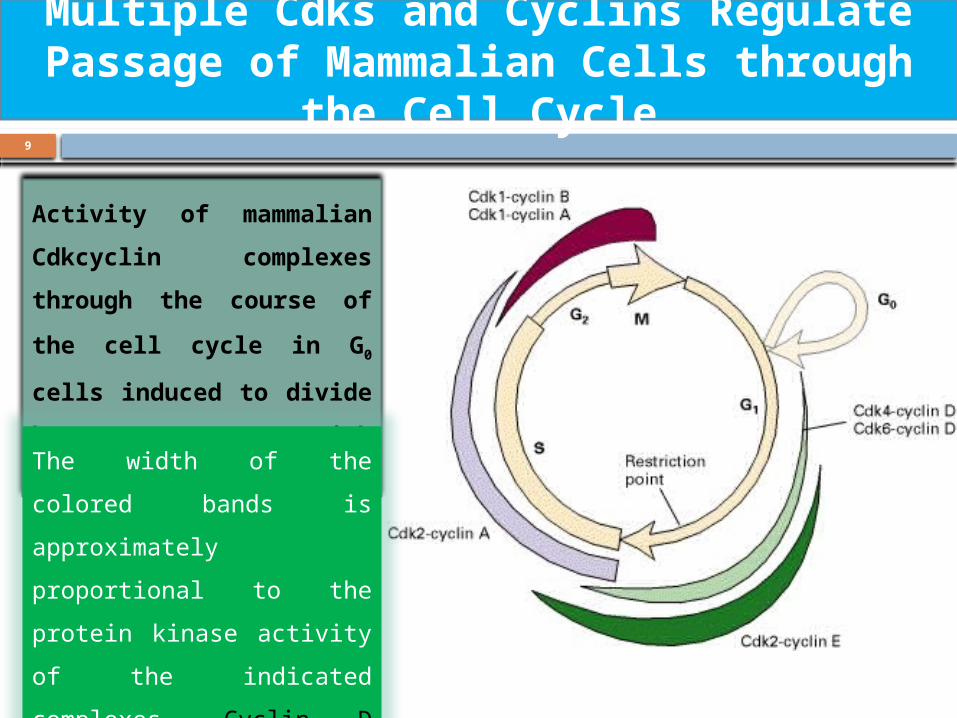
Activity of mammalian
Cdkcyclin complexes
through the course of
the cell cycle in G0 cells
induced to divide by
treatment with growth
factors The width of the colored
bands is approximately
proportional to the protein
kinase activity of the
indicated complexes. Cyclin
D refers to all three D-type
cyclins.
10
Dr Gihan Gawish
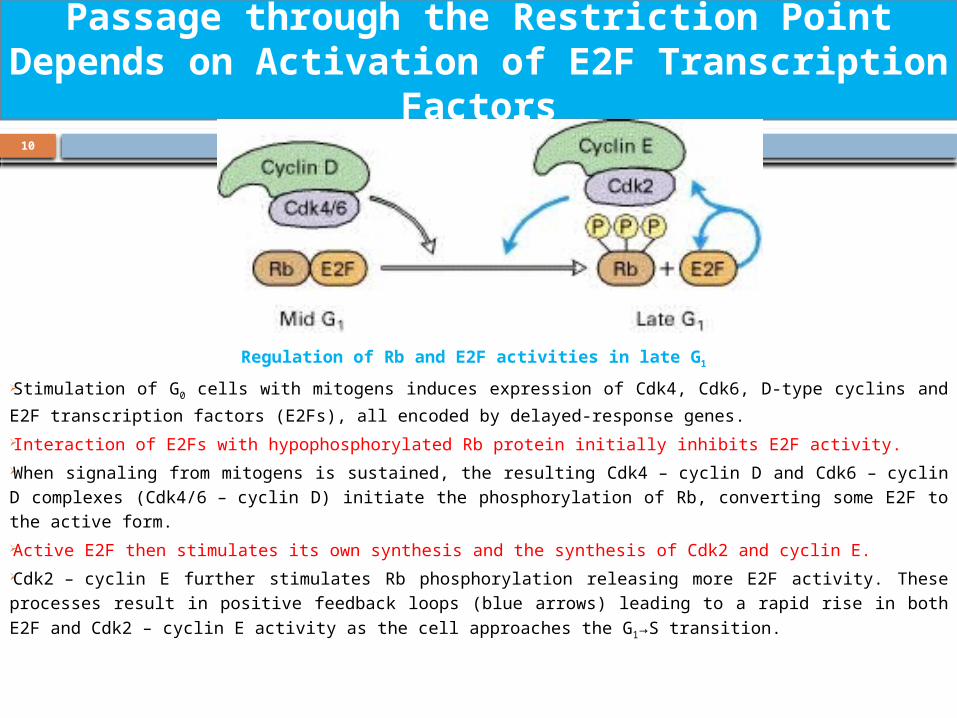
Regulation of Rb and E2F activities in late G1
Stimulation of G0 cells with mitogens induces expression of Cdk4, Cdk6, D-type
cyclins and E2F transcription factors (E2Fs), all encoded by delayed-response genes. Interaction of E2Fs with hypophosphorylated Rb protein initially inhibits E2F activity. When signaling from mitogens is sustained, the resulting Cdk4 – cyclin D and Cdk6 – cyclin D complexes (Cdk4/6 – cyclin D) initiate the phosphorylation of Rb, converting some E2F to the active form. Active E2F then stimulates its own synthesis and the synthesis of Cdk2 and cyclin E. Cdk2 – cyclin E further stimulates Rb phosphorylation releasing more E2F activity. These processes result in positive feedback loops (blue arrows) leading to a rapid rise
in both E2F and Cdk2 – cyclin E activity as the cell approaches the G1→S transition.
Passage through the Restriction Point Depends on Activation of E2F Transcription
Factors

11
Dr Gihan Gawish
Mammalian cells are thought to express a cyclin-kinase inhibitor (CKI)
that functions like S. cerevisiae Sic1.
Mammalian cells are known to express several CKIs that contribute to
cell-cycle control.
These are grouped into two classes:
CIP (Cdk inhibitory protein) family bind and inhibit all Cdk1-, Cdk2-,
Cdk4-, and Cdk6-cyclin complexes
INK4 (inhibitors of kinase 4) family bind and inhibit only Cdk4 –
cyclin D and Cdk6 – cyclin D complexes.
Mammalian Cyclin-Kinase Inhibitors Contribute to Cell-Cycle Control
12
Dr Gihan Gawish
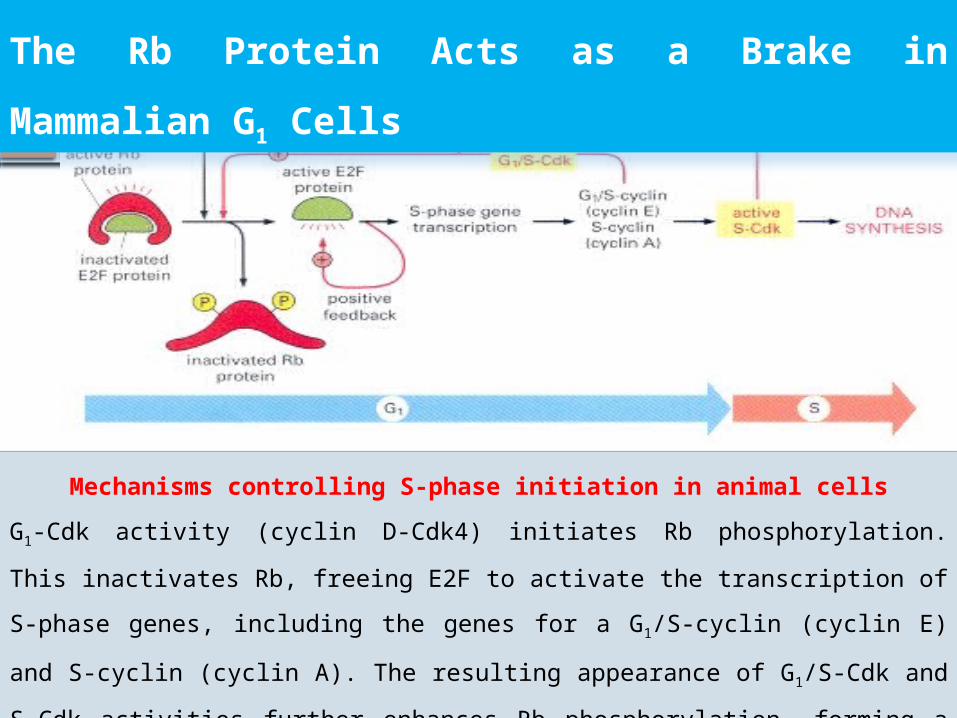
The Rb Protein Acts as a Brake in
Mammalian G1 Cells
Mechanisms controlling S-phase initiation in animal cells
G1-Cdk activity (cyclin D-Cdk4) initiates Rb phosphorylation. This inactivates
Rb, freeing E2F to activate the transcription of S-phase genes, including the
genes for a G1/S-cyclin (cyclin E) and S-cyclin (cyclin A). The resulting
appearance of G1/S-Cdk and S-Cdk activities further enhances Rb
phosphorylation, forming a positive feedback loop. E2F acts back to
stimulate the transcription of its own gene, forming another positive
feedback loop

13
Dr Gihan Gawish
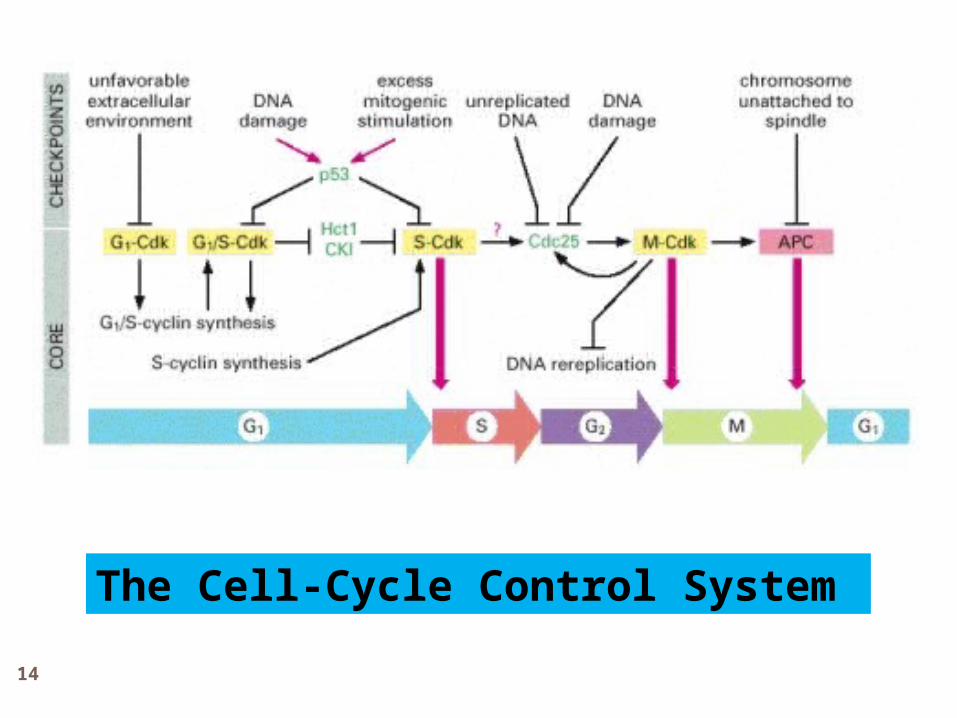
Cell-Cycle Progression is Blocked by DNA Damage and p53:
DNA Damage Checkpoints
When DNA is damaged, protein kinases that phosphorylate p53 are activated.
Mdm2 normally binds to p53 and promotes destruction.
Phosphorylation of p53 blocks its binding to Mdm2; as a result, p53 accumulates to high levels and stimulates transcription of the gene that encodes the CKI protein p21.
The p21 binds and inactivates G1/S-Cdk and S-Cdk complexes, arresting the cell in G1. In some cases, DNA damage also induces either the phosphorylation of Mdm2 or a decrease in Mdm2 production, which causes an increase in p53
14
The Cell-Cycle Control System

15
GENERAL NAME FUNCTIONS AND COMMENTS
Protein kinases and protein
phosphatases that modify Cdks
Cdk-activating kinase (CAK)
phosphorylates an activating site in Cdks
Wee1 kinase phosphorylates inhibitory sites in Cdks; primarily involved in controlling entry into mitosis
Cdc25 phosphataseremoves inhibitory phosphates from Cdks; three family members (Cdc25A, B, C) in mammals; Cdc25C is the activator of Cdk1 at the onset of mitosis
Cdk inhibitory proteins (CKIs)
Sic1 (budding yeast) suppresses Cdk activity in G1; phosphorylation by Cdk1 triggers its destruction
p27 (mammals)suppresses G1/S-Cdk and S-Cdk activities in G1; helps cells to withdraw from cell cycle when they terminally differentiate; phosphorylation by Cdk2 triggers its ubiquitylation by SCF
p21 (mammals) suppresses G1/S-Cdk and S-Cdk activities following DNA damage in G1; transcriptionally activated by p53
p16 (mammals) suppresses G1-Cdk activity in G1; frequently inactivated in cancer
Ubiquitin ligases and their activators
SCFcatalyzes ubiquitylation of regulatory proteins involved in G1 control, including CKIs (Sic1 in budding yeast, p27 in mammals); phosphorylation of target protein usually required for this activity
APCcatalyzes ubiquitylation of regulatory proteins involved primarily in exit from mitosis, including Securin and M-cyclins; regulated by association with activating subunits
Cdc20APC-activating subunit in all cells; triggers initial activation of APC at metaphase-to- anaphase transition; stimulated by M-Cdk activity
Hct1 maintains APC activity after anaphase and throughout G1; inhibited by Cdk activity
Gene regulatory proteins
E2Fpromotes transcription of genes required for G1/S progression, including genes encoding G1/S cyclins, S-cyclins, and proteins required for DNA synthesis; stimulated when G1-Cdk phosphorylates Rb in response to extracellular mitogens
p53promotes transcription of genes that induce cell cycle arrest (especially p21) or apoptosis in response to DNA damage or other cell stress; regulated by association with Mdm2, which promotes p53 degradation
The Major Cell-cycle Regulatory Proteins

16
The number of cells in this community is tightly regulated—not simply by controlling
the rate of cell division, but also by controlling the rate of cell death process known as
programmed cell death, or apoptosis.
Apoptosis is mediated by proteolytic enzymes called caspases, which trigger cell death
by cleaving specific proteins in the cytoplasm and nucleus.
Caspases exist in all cells as inactive precursors, or procaspases, which are usually
activated by cleavage by other caspases, producing a proteolytic caspase cascade.
The activation process is initiated by either extracellular or intracellular death signals,
which cause intracellular adaptor molecules to aggregate and activate procaspases.
Caspase activation is regulated by members of the Bcl-2 and IAP protein families.
Programmed Cell Death (Apoptosis)
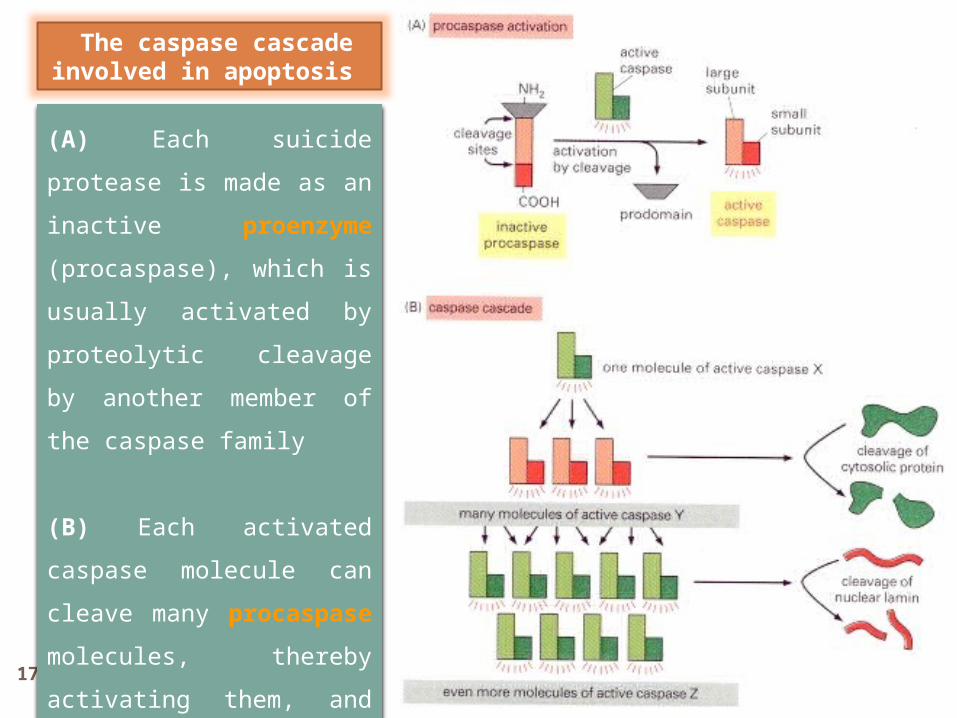
17 Dr Gihan Gawish
(A) Each suicide protease
is made as an inactive
proenzyme (procaspase),
which is usually activated
by proteolytic cleavage by
another member of the
caspase family
(B) Each activated
caspase molecule can
cleave many procaspase
molecules, thereby
activating them, and these
can then activate even
more procaspase
molecules.
The caspase cascade involved in apoptosis
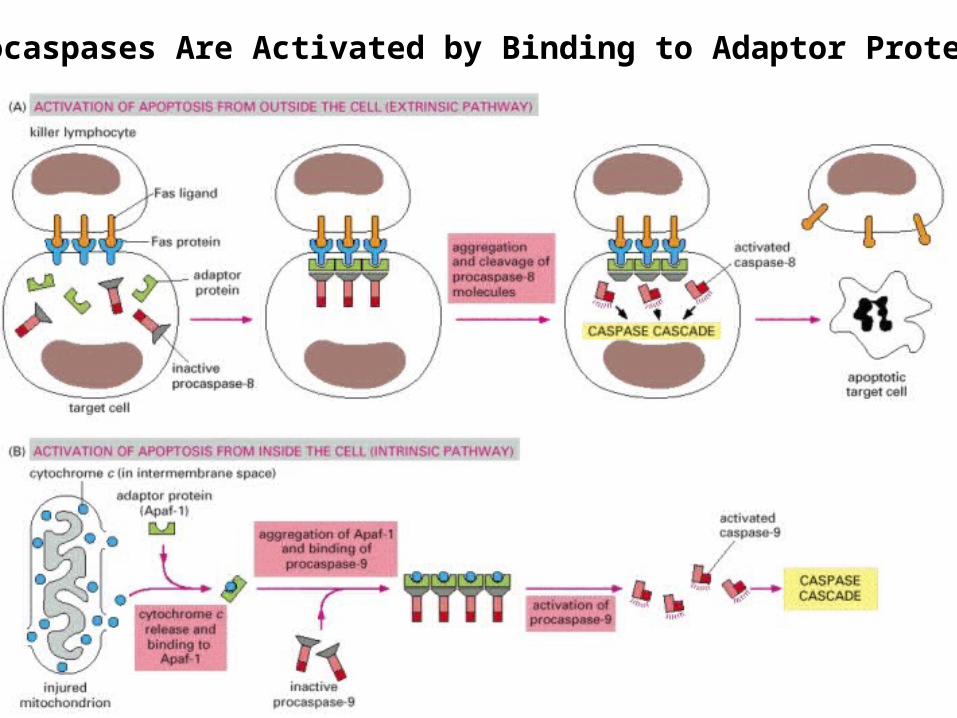
18 Dr Gihan Gawish
Procaspases Are Activated by Binding to Adaptor Proteins

19
Dr Gihan Gawish
Bcl-2 Family Proteins Are the Main Intracellular Regulators of the Cell Death Program
The Bcl-2 family of intracellular proteins helps regulate the activation of procaspases. Some
members of this family, like Bcl-2 itself or Bcl-XL, inhibit apoptosis, at least partly by blocking the
release of cytochrome c from mitochondria.
Other members of the Bcl-2 family are not death inhibitors, but instead promote procaspase
activation and cell death.
Some of these apoptosis promoters, such as Bad, function by binding to and inactivating the death-inhibiting members of the family, whereas others,
like Bax and Bak, stimulate the release of cytochrome c from mitochondria.

20
Dr Gihan Gawish
IAP Proteins Are the Main Intracellular Regulators of the Cell Death Program
they bind to some procaspases to prevent their activation, and they bind to caspases to inhibit
their activity
IAP proteins were originally discovered as proteins produced by certain insect viruses, which use them
to prevent the infected cell from killing itself before the virus has had time to replicate.
When mitochondria release cytochrome c to activate Apaf-1, they also release a protein that
blocks IAPs, thereby greatly increasing the efficiency of the death activation process

21
The factors that promote organ or organism growth can be operationally divided into three major classes:
Mitogens, which stimulate cell division, primarily by relieving intracellular
negative controls that otherwise block progress through the cell cycle.
Growth factors, which stimulate cell growth (an increase in cell mass) by
promoting the synthesis of proteins and other macromolecules and by
inhibiting their degradation.
Survival factors, which promote cell survival by suppressing apoptosis.
Extracellular Control of Cell Division, Cell Growth, and Apoptosis
The extracellular signal molecules that regulate cell size and cell number are generally either soluble secreted proteins, proteins bound to the surface of cells, or components of the extracellular
matrix.


















