Down-regulationofMycIsEssentialforTerminalErythroid ... · tor (MSCV) construct that co-expresses...
15
Down-regulation of Myc Is Essential for Terminal Erythroid Maturation * □ S Received for publication, September 1, 2010 Published, JBC Papers in Press, October 12, 2010, DOI 10.1074/jbc.M110.181073 Senthil Raja Jayapal ‡§1 , Kian Leong Lee ¶ , Peng Ji , Philipp Kaldis**, Bing Lim ‡§‡‡2,3 , and Harvey F. Lodish ‡4 From the ‡ Computation and Systems Biology, Singapore-Massachusetts Institute of Technology Alliance, 4 Engineering Drive 3, Singapore 117576, the § Genome Institute of Singapore, 60 Biopolis Street, Genome, Singapore 138672, the ¶ Cancer Science Institute of Singapore, National University of Singapore, Centre for Life Sciences, 28 Medical Drive, Singapore 117456, the Whitehead Institute for Biomedical Research and Department of Biology, Massachusetts Institute of Technology, Cambridge, Massachusetts 02142 , the **Institute of Molecular and Cell Biology, 61 Biopolis Drive, Proteos, Singapore 138673, and the ‡‡ Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts 02215 Terminal differentiation of mammalian erythroid progenitors involves 4 –5 cell divisions and induction of many erythroid im- portant genes followed by chromatin and nuclear condensation and enucleation. The protein levels of c-Myc (Myc) are reduced dramatically during late stage erythroid maturation, coinciding with cell cycle arrest in G 1 phase and enucleation, suggesting pos- sible roles for c-Myc in either or both of these processes. Here we demonstrate that ectopic Myc expression affects terminal eryth- roid maturation in a dose-dependent manner. Expression of Myc at physiological levels did not affect erythroid differentiation or cell cycle shutdown but specifically blocked erythroid nuclear condensation and enucleation. Continued Myc expression pre- vented deacetylation of several lysine residues in histones H3 and H4 that are normally deacetylated during erythroid maturation. The histone acetyltransferase Gcn5 was up-regulated by Myc, and ectopic Gcn5 expression partially blocked enucleation and inhib- ited the late stage erythroid nuclear condensation and histone deacetylation. When overexpressed at levels higher than the physiological range, Myc blocked erythroid differentiation, and the cells continued to proliferate in cytokine-free, serum-contain- ing culture medium with an early erythroblast morphology. Gene expression analysis demonstrated the dysregulation of erythro- poietin signaling pathway and the up-regulation of several posi- tive regulators of G 1 -S cell cycle checkpoint by supraphysiological levels of Myc. These results reveal an important dose-dependent function of Myc in regulating terminal maturation in mammalian erythroid cells. Mammalian terminal erythroid development is a precisely regulated process involving rapid proliferation and serial mor- phological changes of committed erythroid progenitors (col- ony forming units-erythroid, or CFU-E) 5 to form mature erythrocytes. The initial stages of terminal erythroid matura- tion are highly dependent on erythropoietin (1), whose roles in activation of erythroid specific genes, terminal prolifera- tion, and protection against apoptosis are well understood (2). This is followed by an erythropoietin-independent, fibronec- tin-dependent phase where survival and proliferation of the erythroblasts require signaling by 41 integrins (3). Upon erythropoietin stimulation, CFU-E progenitors undergo 4 –5 cell divisions accompanied by a decrease in cell size, increase in hemoglobin content, and nuclear condensation followed by withdrawal from the cell cycle (4). Late stage mammalian erythroblasts undergo nuclear condensation and enucleate by extrusion of the pycnotic nucleus surrounded by a thin layer of cytoplasm and cell membrane (5–7). The molecular mech- anisms that regulate this hallmark process remain to be fully elucidated. We previously reported that Rac GTPases and their downstream effector mDia2 play important roles in the cytoskeletal reorganization leading to the extrusion of the pycnotic nucleus from late stage erythroblasts (8). Neverthe- less, the mechanisms regulating condensation of the erythroid nucleus preceding its extrusion remain unclear. The proto-oncogene c-Myc (Myc) has been widely studied since its identification as a cellular homolog of the retroviral v-myc oncogene (9), and its functions in the genesis and maintenance of tumors are well established (10, 11). The Myc transcription factor plays important roles in normal, non- transformed cells in regulating proliferation, differentiation, cell growth and apoptosis (12–14) and is critical for several aspects of hematopoietic development and function (15). Studies of epiblast-specific Myc deletion in mice suggest that both primitive and definitive hematopoiesis are impaired in the absence of Myc (16). Forced expression of Myc promoted differentiation of hematopoietic stem cells at the expense of their self-renewal ability (17), indicating that Myc functions in the hematopoietic lineages are not limited to stimulating pro- liferation. Myc expression is rapidly induced upon erythropoi- etin stimulation of committed erythroid progenitor cells (18), * This work was supported, in whole or in part, by National Institutes of Health Grants DK047636 and AI54973 (to B. L.) and P01 HL32262 (to H. F. L.). This study was also supported by Singapore-MIT alliance Grant C-382-641-001-091 (to B. L. and H. F. L.). □ S The on-line version of this article (available at http://www.jbc.org) con- tains supplemental Tables S1 and S2. 1 Supported by a Singapore-MIT Alliance graduate fellowship. 2 Supported by A*STAR funds in Genome Institute of Singapore. 3 To whom correspondence may be addressed: Genome Institute of Singa- pore, 60 Biopolis St., Genome, Singapore 138672. Tel.: 65-64788186; Fax: 65-64789005; E-mail: [email protected]. 4 To whom correspondence may be addressed: Whitehead Institute for Bio- medical Research, 9 Cambridge Center, Suite 601, Cambridge, MA 02142. Tel.: 617-258-5216; Fax: 617-258-6768; E-mail: [email protected]. 5 The abbreviations used are: CFU-E, colony forming units-erythroid; MSCV, murine stem cell retroviral vector; qRT, quantitative real-time; HDAC, his- tone deacetylase; HAT, histone acetyltransferase; MIG, MSCV-IRES-GFP retroviral vector. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 51, pp. 40252–40265, December 17, 2010 © 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 40252 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010 by guest on September 9, 2020 http://www.jbc.org/ Downloaded from
Transcript of Down-regulationofMycIsEssentialforTerminalErythroid ... · tor (MSCV) construct that co-expresses...

Down-regulation of Myc Is Essential for Terminal ErythroidMaturation*□S
Received for publication, September 1, 2010 Published, JBC Papers in Press, October 12, 2010, DOI 10.1074/jbc.M110.181073
Senthil Raja Jayapal‡§1, Kian Leong Lee¶, Peng Ji�, Philipp Kaldis**, Bing Lim‡§‡‡2,3, and Harvey F. Lodish‡�4
From the ‡Computation and Systems Biology, Singapore-Massachusetts Institute of Technology Alliance, 4 Engineering Drive 3,Singapore 117576, the §Genome Institute of Singapore, 60 Biopolis Street, Genome, Singapore 138672, the ¶Cancer ScienceInstitute of Singapore, National University of Singapore, Centre for Life Sciences, 28 Medical Drive, Singapore 117456, the�Whitehead Institute for Biomedical Research and Department of Biology, Massachusetts Institute of Technology, Cambridge,Massachusetts 02142 , the **Institute of Molecular and Cell Biology, 61 Biopolis Drive, Proteos, Singapore 138673, and the‡‡Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts 02215
Terminal differentiation ofmammalian erythroid progenitorsinvolves 4–5 cell divisions and induction ofmany erythroid im-portant genes followed by chromatin and nuclear condensationand enucleation. The protein levels of c-Myc (Myc) are reduceddramatically during late stage erythroidmaturation, coincidingwith cell cycle arrest in G1 phase and enucleation, suggesting pos-sible roles for c-Myc in either or both of these processes. Here wedemonstrate that ectopicMyc expression affects terminal eryth-roidmaturation in a dose-dependentmanner. Expression ofMycat physiological levels did not affect erythroid differentiation orcell cycle shutdown but specifically blocked erythroid nuclearcondensation and enucleation. ContinuedMyc expression pre-vented deacetylation of several lysine residues in histones H3 andH4 that are normally deacetylated during erythroidmaturation.The histone acetyltransferase Gcn5 was up-regulated byMyc, andectopic Gcn5 expression partially blocked enucleation and inhib-ited the late stage erythroid nuclear condensation and histonedeacetylation.When overexpressed at levels higher than thephysiological range,Myc blocked erythroid differentiation, andthe cells continued to proliferate in cytokine-free, serum-contain-ing culturemediumwith an early erythroblast morphology. Geneexpression analysis demonstrated the dysregulation of erythro-poietin signaling pathway and the up-regulation of several posi-tive regulators of G1-S cell cycle checkpoint by supraphysiologicallevels ofMyc. These results reveal an important dose-dependentfunction ofMyc in regulating terminal maturation inmammalianerythroid cells.
Mammalian terminal erythroid development is a preciselyregulated process involving rapid proliferation and serial mor-
phological changes of committed erythroid progenitors (col-ony forming units-erythroid, or CFU-E)5 to form matureerythrocytes. The initial stages of terminal erythroid matura-tion are highly dependent on erythropoietin (1), whose rolesin activation of erythroid specific genes, terminal prolifera-tion, and protection against apoptosis are well understood (2).This is followed by an erythropoietin-independent, fibronec-tin-dependent phase where survival and proliferation of theerythroblasts require signaling by �4�1 integrins (3). Uponerythropoietin stimulation, CFU-E progenitors undergo 4–5cell divisions accompanied by a decrease in cell size, increasein hemoglobin content, and nuclear condensation followed bywithdrawal from the cell cycle (4). Late stage mammalianerythroblasts undergo nuclear condensation and enucleate byextrusion of the pycnotic nucleus surrounded by a thin layerof cytoplasm and cell membrane (5–7). The molecular mech-anisms that regulate this hallmark process remain to be fullyelucidated. We previously reported that Rac GTPases andtheir downstream effector mDia2 play important roles in thecytoskeletal reorganization leading to the extrusion of thepycnotic nucleus from late stage erythroblasts (8). Neverthe-less, the mechanisms regulating condensation of the erythroidnucleus preceding its extrusion remain unclear.The proto-oncogene c-Myc (Myc) has been widely studied
since its identification as a cellular homolog of the retroviralv-myc oncogene (9), and its functions in the genesis andmaintenance of tumors are well established (10, 11). The Myctranscription factor plays important roles in normal, non-transformed cells in regulating proliferation, differentiation,cell growth and apoptosis (12–14) and is critical for severalaspects of hematopoietic development and function (15).Studies of epiblast-specific Myc deletion in mice suggest thatboth primitive and definitive hematopoiesis are impaired inthe absence of Myc (16). Forced expression of Myc promoteddifferentiation of hematopoietic stem cells at the expense oftheir self-renewal ability (17), indicating that Myc functions inthe hematopoietic lineages are not limited to stimulating pro-liferation. Myc expression is rapidly induced upon erythropoi-etin stimulation of committed erythroid progenitor cells (18),
* This work was supported, in whole or in part, by National Institutes ofHealth Grants DK047636 and AI54973 (to B. L.) and P01 HL32262 (toH. F. L.). This study was also supported by Singapore-MIT alliance GrantC-382-641-001-091 (to B. L. and H. F. L.).
□S The on-line version of this article (available at http://www.jbc.org) con-tains supplemental Tables S1 and S2.
1 Supported by a Singapore-MIT Alliance graduate fellowship.2 Supported by A*STAR funds in Genome Institute of Singapore.3 To whom correspondence may be addressed: Genome Institute of Singa-
pore, 60 Biopolis St., Genome, Singapore 138672. Tel.: 65-64788186; Fax:65-64789005; E-mail: [email protected].
4 To whom correspondence may be addressed: Whitehead Institute for Bio-medical Research, 9 Cambridge Center, Suite 601, Cambridge, MA 02142.Tel.: 617-258-5216; Fax: 617-258-6768; E-mail: [email protected].
5 The abbreviations used are: CFU-E, colony forming units-erythroid; MSCV,murine stem cell retroviral vector; qRT, quantitative real-time; HDAC, his-tone deacetylase; HAT, histone acetyltransferase; MIG, MSCV-IRES-GFPretroviral vector.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 51, pp. 40252–40265, December 17, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
40252 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

and its levels are substantially reduced during the final stagesof erythroid maturation (19). The physiological relevance ofthese noticeable changes in Myc expression during terminalerythroid maturation remains unclear due to conflicting re-ports in literature. Studies in G1E erythroid cell lines reportthat forced Myc expression prevented cell cycle arrest but hadminimal effects on erythroid maturation (20). In contrast,Myc expression blocked erythroid differentiation in humanleukemia K562 cells without preventing the cell cycle exit(21). These discrepancies are possibly a result of the differentcell lines used as models of terminal erythroid maturation.Retroviral overexpression of Myc in unfractionated murinefetal liver cells resulted in continuously growing erythroblastcell lines, suggesting that Myc overexpression enhanced theself- renewal ability of early erythroblasts at the expense ofdifferentiation (22). Here we use a recently developed in vitroculture system in which the proliferation and differentiationof purified TER119-negative mouse fetal liver erythroblastscan be monitored quantitatively in a step-by-step manner (23)to investigate the role of Myc in terminal erythroidmaturation.We demonstrate that ectopic Myc expression has a dose-
dependent effect on terminal erythroid differentiation of puri-fied mouse fetal liver erythroblasts cultured in vitro. EctopicMyc expression at physiological levels specifically blocks enu-cleation without affecting terminal erythroid maturation. Incontrast, overexpression of Myc at levels above the physiolog-ical range promotes proliferation of early erythroblasts at theexpense of differentiation and also evokes an apoptotic re-sponse. Our study indicates that Myc levels play a critical rolein regulating the balance between proliferation and differenti-ation in the erythroid lineage. Our data uncover an importantrole for Myc in regulating erythroid nuclear condensation andenucleation.
EXPERIMENTAL PROCEDURES
Purification and Culture of Murine Fetal Liver ErythroidProgenitors—All mouse work was carried out with approvedInstitutional Animal Care and Use Committee protocols atthe Biological Research Centre mouse facility at Biopolis, Sin-gapore. Fetal livers were isolated from E13.5 C57BL/6 miceembryos and mechanically dissociated by pipetting in PBScontaining 20% fetal bovine serum (FBS). The dissociatedcells were passed through 70- and 30-�m strainers to obtainsingle-cell suspensions. For negative selection, cells were la-beled with magnetic microbead-conjugated antibodies forTER119 and CD11b (to remove macrophages) and passedthrough LD columns according to the manufacturer’s instruc-tions (Miltenyi Biotec). The purified TER119-negative eryth-roblasts were seeded in fibronectin-coated plates and culturedin Iscove’s modified Dulbecco’s medium (Invitrogen) contain-ing 15% FBS (StemCell Technologies), 1% detoxified bovineserum albumin (StemCell Technologies), 200 �g/ml holo-transferrin (Sigma),10 �g/ml recombinant human insulin(Sigma), 2 mM L-glutamine (Invitrogen), 10�4 M �-mercapto-ethanol (Invitrogen), and 2 units/ml erythropoietin (R&D sys-tems). For p27 knock-out studies, E13.5 embryos harvestedfrom intercrosses of p27�/� mice (24) were genotyped as de-
scribed previously (25) followed by purification of fetal livererythroid progenitors.Retroviral Constructs—The murine stem cell retroviral vec-
tor (MSCV) construct that co-expresses Myc and GFP(MSCV-Myc-IRES-GFP) was a gift from Dr. Michael H. To-masson (26). The MSCV-Gcn5-IRES-GFP construct to over-express Gcn5 was made by cloning the full-length Gcn5 openreading frame (ORF) into the EcoRI-NotI sites of the MSCV-IRES-GFP vector. The full-length GCN5 ORF was PCR-am-plified from cDNA clones kindly provided by Dr. Sharon Y.R.Dent (27). The retroviral construct expressing shRNA againstc-Myc was purchased from Open Biosystems. The target se-quence for the c-Myc shRNA is 5�-AGCCTTGAAATGTA-AATAACTT-3�, and the target sequence of the non-silencingnegative control (incomplete luciferase shRNA) is 5�-GTGCGTTGCTAGTACCAACTTCAAGAGA-3�.Retrovirus Generation and Infection—Replication-incompe-
tent retroviruses were produced by co-transfection of 293Tcells with the MSCV constructs and the ecotropic packagingvector pCL-Eco using Lipofectamine 2000 (Invitrogen). Ret-roviral supernatants were harvested 48 h after transfection,passed through 0.45-�m filters, and stored in aliquots at�80 °C. The virus titers for the MSCV retroviral supernatantswere estimated by transduction of NIH3T3 cells and quantify-ing the percentage of GFP-positive cells by FACS analysis. Forinfection of purified TER119-negative erythroid progenitorcells, 5 � 105 cells were resuspended in 1 ml of retroviral su-pernatant containing 8 �g/ml Polybrene (Sigma) and centri-fuged at 2000 rpm for 1 h at 37 °C. The virus supernatant wasthen replaced by erythropoietin-containing medium.Immunostaining and Flow Cytometry—Cells were washed
and resuspended in 200 �l of PBS containing 4% FBS for im-munostaining. For analysis of erythroid differentiation, cellswere incubated with 1:200 dilution of phycoerythrin-conju-gated anti-CD71 (BD Biosciences) and allophycocyanin-con-jugated anti-Ter-119 antibody (BD Biosciences) for 15 min atroom temperature. For enucleation analysis, cells were addi-tionally stained with 10 �g/ml Hoechst 33342 (Sigma) atroom temperature for 15 min. Propidium iodide (BD Bio-sciences) at a final concentration of 0.2 �g/ml was added toexclude dead cells from the analysis. Apoptosis was evaluatedby co-staining with allophycocyanin-conjugated annexin V(BD Biosciences) and propidium iodide. To perform cell cycleanalysis, the cells were washed and resuspended in 50 �l ofPBS, fixed in 1 ml of cold 90% ethanol, and stored at 4 °Covernight. The fixed cells were washed and incubated in PBScontaining propidium iodide (20 �g/ml) and RNase-A (200�g/ml) for 1 h at room temperature. Flow cytometry was per-formed using BD LSRII (BD Biosciences), and data analysiswas carried out using BD FACSDiva (BD Biosciences).Benzidine-Giemsa Staining and Hemoglobin Assay—5 �
104 cells were centrifuged onto glass slides using Cytospin 4(Shandon). The cells were air-dried completely and fixed in�20 °C methanol for 2 min. Benzidine solution was preparedby dissolving one benzidine tablet (Sigma, D5905) in 10 ml ofPBS and filtered through a 0.22-�m filter. 10 �l of 30% Hy-drogen peroxide solution (Calbiochem) was added to the ben-zidine solution just before staining and placed as drops di-
Function of Myc in Terminal Erythroid Maturation
DECEMBER 17, 2010 • VOLUME 285 • NUMBER 51 JOURNAL OF BIOLOGICAL CHEMISTRY 40253
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

rectly on fixed cells for 2 h at room temperature. The cellswere rinsed briefly with PBS and stained with 1:20 dilution ofGiemsa (Sigma, GS500) in water for 15 min at room tempera-ture. Excess Giemsa stain was removed by washing with PBS.Images were captured on an Olympus BX51 microscopeequipped with an Olympus DP70 digital camera using DP-Controller image capture software (Olympus). For hemoglo-bin assay, 1 � 106 erythroid cells were lysed in 200 �l ofDrabkin’s reagent (Sigma), and hemoglobin content wasquantified by spectrophotometric measurement of absor-bance at 540 nm on a Tecan Sunrise Reader.Measurement of Nuclear Size by High Content Screening—
Erythroid cells were fixed in 3.7% formaldehyde in PBS for 7min at room temperature, then incubated in 10 �g/ml solu-tion of Hoechst 33342 in PBS for 15 min to stain the nucleusfollowed by 2 washes with PBS. The cells were resuspended inPBS, transferred to flat-bottom 96-well cell culture plates, andcentrifuged at 1000 rpm for 3 min to deposit the cells as a sin-gle layer at the bottom of the wells. Images were acquired bythe automated Arrayscan VTI high content screening reader(Cellomics) using a 20� objective lens from 16 different fieldsin each well. The images were analyzed by Cellomics TargetActivation BioApplication software. Cell clumps and debriswere excluded from analysis by setting appropriate cut-offvalues after visual inspection of the images, and data acquiredconsisted of at least 5000 objects (cells) per sample. The nu-clear size (nuclear area per cell) for each sample was deter-mined as the average Hoechst area per object in the sample.Quantitative Real-time (qRT-PCR)—Total RNA was ex-
tracted from cells using RNeasy mini kits (Qiagen). 1 �g oftotal RNA for each sample was reverse-transcribed using theHigh Capacity cDNA Archive kit (Applied Biosystems). Rela-tive transcript levels of different genes were quantified bySYBR Green real time PCR using ABI Prism 7900HT Se-quence Detection System 2.2 (Applied Biosystems). The re-sults were normalized to GAPDH and analyzed using SDS2.2.2 software. The primer sequences are provided in supple-mental Table S2.Gene Expression Microarrays and Data Analysis—Total
RNA was extracted from 3 biological replicates of the cellsunder each condition with RNeasy mini kits, and 500 ng wasreverse-transcribed into cDNA followed by in vitro transcrip-tion to biotin-labeled cRNA using the Illumina TotalPrepRNA Amplification kit (Applied Biosystems). 750 ng of eachcRNA sample was hybridized to MouseRef-8 Version 2.0 Ex-pression BeadChip microarrays (Illumina) and scanned on theBeadArray Reader (Illumina) at scan factor 1. Backgroundsubtraction was applied on raw intensity values, and subse-quent data were subjected to quantile normalization on theBeadstudio Data Analysis platform (Illumina) with a normal-ized expression value cutoff at 100. Differentially expressedgenes were identified based on an at least a 2-fold change in atleast one condition compared with controls (D0 WT). Indi-vidual gene lists were uploaded into Ingenuity Pathway Analy-sis software (Ingenuity System) to identify the significantlyenriched gene ontology categories based on molecular andcellular functions. Microarray data were deposited into GeneExpression Omnibus (accession number GSE18558).
Western Blots—Cells were lysed in radioimmune precipita-tion assay buffer (Pierce) containing protease inhibitors(Roche Applied Science). 20 �g of total protein for each sam-ple was resolved on a 10% SDS-polyacrylamide gel and trans-ferred to methanol-activated polyvinylidene difluoride mem-branes (GE Healthcare). After washing once with TBST (20mM Tris, pH 7.5, 500 mM NaCl, 0.1% Tween 20), the mem-branes were blocked for 1 h at room temperature in 5% bo-vine serum albumin (BSA) in TBST with mild shaking. Theblots were then incubated with primary antibody at appropri-ate dilution in TBST with 2% BSA overnight with gentle shak-ing at 4 °C, washed 3 times for 15 min with TBST, and incu-bated with horseradish peroxidase-conjugated secondaryantibody for 1 h at room temperature. The blots were washedwith TBST 3 times for 5 min and visualized using a chemilu-minescent substrate (Luminol reagent, Santa Cruz Biotech-nologies). Histones were purified by acid extraction for West-ern blot analysis of histones H3 and H4 acetylation. Cells werewashed once with ice-cold PBS, and cell pellets were incu-bated in PBS containing 0.5% Triton X-100 and protease in-hibitors for 10 min on ice. Pellets were resuspended in 0.2 N
HCl at 4 °C overnight for extraction of histones. The acidicpH of histone extracts was neutralized with 1 M Tris, pH 8.Equal loading of histones was ensured by Ponceau stainingbefore proceeding for immunoblotting with acetylation-spe-cific antibodies. Primary antibodies used for Western blottingwere as follows: Myc (SC-764, Santa Cruz); p27 (610241, BDBiosciences); Gcn5 (#3305, Cell Signaling); GAPDH (ab37187,Abcam); total histone H4 (ab10158, Abcam). The antibodiesfor H3K9Ac, H3K18Ac, H3K23Ac, and H3 (total) were pur-chased from Cell Signaling (#9927). Antibodies for H4K5Ac,H4K8Ac, H4K12Ac, and H4K16Ac were purchased fromMil-lipore (#17–211).Chromatin Immunoprecipitation (ChIP) Assay—TER119-
negative mouse fetal erythroid progenitor cells were cross-linked with 1% formaldehyde for 10 min at room temperature,and chromatin immunoprecipitation was performed accord-ing to Agilent ChIP-on-chip protocol (version 9.1, Nov 2006)using anti-c-Myc antibody (SC-764, Santa Cruz). The enrich-ments of binding sites in the immunoprecipitated DNA werequantified using qRT-PCR. The -fold enrichment values werecalculated by determining the apparent immunoprecipitationefficiency (ratios of the amount of immunoprecipitated DNAto that of the input sample) and normalized to the level ob-served at a control region, which was defined as 1.0. The ChIPqRT-PCR primer sequences are available in supplementalTable S2.Statistical Analysis—Student’s t test was used to determine
the significance of differences between treated samples andcontrols. Statistical analysis was performed using MicrosoftOffice Excel 2003.
RESULTS
Ectopic Myc Expression Has a Dose-dependent Effect onTerminal Erythroid Maturation—As a first step in addressingthe role of Myc in terminal erythropoiesis, we monitored thechanges in Myc protein levels during differentiation ofTER119-negative erythroid progenitor cells purified from
Function of Myc in Terminal Erythroid Maturation
40254 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

E13.5 mouse fetal livers and cultured for 2 days on fibronec-tin-coated plates in medium containing serum and erythro-poietin. These culture conditions support proper terminalproliferation and differentiation of CFU-E progenitors in amanner mimicking erythropoiesis in vivo (23). Myc proteinlevels decreased substantially during the 2-day in vitro culture(Fig. 1A, lanes 1–3). We also quantified changes in p27Kip1(p27) protein levels, as in many cells p27 is repressed by Myc(28), and changes in p27 levels have been suggested to play acritical role in the withdrawal from cell cycle during terminalerythroid maturation (29). As expected, p27 protein levelswere strongly induced during the later stages of erythroid dif-ferentiation (Fig. 1A, lanes 1–3).
We then examined differentiation of TER119-negativeerythroid progenitor cells ectopically expressing Myc. A bicis-tronic retroviral vector (MSCV-Myc-IRES-GFP) that also en-codes green fluorescent protein (GFP) was used to expressMyc in purified TER119-negative erythroid progenitor cells.This allowed for GFP intensity measured by flow cytometry toserve as a reporter for the level of ectopic Myc expression(30). At 24 h post-retroviral infection, the cells were sortedbased on GFP intensity into low GFP (Myc-GFPlow) and highGFP populations (Myc-GFPhigh) followed by culture for an-other 24 h to allow for complete terminal maturation. Cellsinfected with retroviruses encoding GFP only (MIG) served ascontrol. Western blot analysis of retrovirus-transduced cellswas performed at 24 and 48 h post-infection to measureMyc and p27 levels (Fig. 1A, lanes 4–9) and were comparedwith those during differentiation of uninfected erythro-blasts (Fig. 1A, lanes 1–3). The Myc-GFPlow cells showedcontinued Myc expression at levels similar to those of nor-mal cells at 1 day of differentiation (Fig. 1A, lanes 5 and 8)and, thus, within a physiological range. In contrast, theMyc-GFPhigh cells showed Myc expression at levels abovethe physiological range of Myc expression in differentiatingerythroblasts (Fig. 1A, lanes 6 and 9). Myc-GFPhigh cellsdemonstrated Myc protein levels greater than early eryth-roid progenitors (freshly purified TER119-negative fetalliver erythroblasts) at both 24 and 48 h in culture, indicat-ing that these cells represented high level Myc-overex-pressing erythroblasts. At 48 h in culture, the induction ofp27 in Myc-GFPlow cells was similar to that of controlcells, but p27 induction was significantly reduced in Myc-GFPhigh cells (Fig. 1A, lanes 7–9).The terminal differentiation states of Myc-GFPlow,
Myc-GFPhigh, and control (MIG) cells were examined byflow cytometry at 48 h in culture. The percentage ofCD71�TER119� populations was similar in Myc-GFPlow andcontrol cells but was significantly reduced in the Myc-GF-Phigh cells, suggesting that overexpression of Myc at highlevels blocked erythroid differentiation (Fig. 1B, top panel).Enucleation was quantified by measuring the reticulocytesformed in culture, which stain positive for the erythroid dif-ferentiation marker TER119 and negative for Hoechst 33342(Hoechst). The TER119� Hoechst� reticulocyte populationseen in the control cells was absent in both Myc-GFPlow andMyc-GFPhigh cells (Fig. 1B,middle panel), indicating thatsustained expression of Myc at a physiological level was suffi-
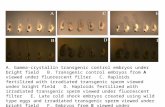
FIGURE 1. Dose-dependent effect of ectopic Myc on terminal erythroidmaturation. A, Western blot analysis of Myc and p27 protein levels isshown. TER119-negative fetal liver erythroblasts purified from E13.5 mouseembryos and cultured in erythropoietin-containing medium in fibronectin-coated plates and cells were harvested at 0 h (D0), 24 h (D1), or 48 h (D2) inculture. Purified erythroblasts were infected with bicistronic retrovirusesencoding GFP only as control or both Myc and GFP at 0 h in culture. At 24 hpost-infection, cells were sorted based on GFP intensity into low (Myc-GF-Plow) and high (Myc-GFPhigh) intensity populations, then cultured for an-other 24 h to allow for differentiation. Cells were harvested at indicatedtime points, and lysates were subjected to Western blotting using the indi-cated antibodies. GAPDH protein levels were determined as a loading con-trol. The Myc bands in the Western blot image were quantified using Im-ageJ, normalized to the GAPDH bands, and are presented as -fold changerelative to D0 levels. Error bars represent S.D. (n � 3). B, flow cytometricanalysis of terminal erythroid maturation is shown. At 48 h post-infection,control, Myc-GFPlow, and Myc-GFPhigh cells were analyzed by flowcytometry. Erythroid differentiation was assayed by quantifying theCD71�TER119� population, which represents the late stage erythroid cells(top). Enucleation was assayed by quantifying the TER119�Hoechst� popu-lations, which correspond to the enucleated reticulocytes (middle). Apopto-sis and cell death were assessed by co-staining with annexin V and pro-pidium iodide (PI, bottom).
Function of Myc in Terminal Erythroid Maturation
DECEMBER 17, 2010 • VOLUME 285 • NUMBER 51 JOURNAL OF BIOLOGICAL CHEMISTRY 40255
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

cient to block erythroid enucleation. Myc-GFPlow cells didnot show any significant change in the levels of apoptosiscompared with control cells, whereas Myc-GFPhigh cellsshowed significantly high levels of apoptosis (Fig. 1B, bottompanel). Taken together, continued expression of Myc at aphysiological level blocked enucleation specifically withoutaffecting the terminal differentiation or survival of primaryerythroblasts, whereas overexpression of Myc at supraphysi-ological levels additionally blocked erythroid differentiationand induced apoptosis.Ectopic Myc Expression at Supraphysiological Levels Inhib-
its Hemoglobin Expression and Exit from the Cell Cycle—Toconfirm the inhibition of terminal erythroid differentiation byoverexpression of Myc at supraphysiological levels, we mea-sured the hemoglobin content of the cultured erythroblasts at48 h post-infection. The hemoglobin content of Myc-GF-Phigh cells was significantly lower than that of control cells,whereas the hemoglobin level of Myc-GFPlow cells was nor-mal (Fig. 2A). We next measured total cell counts at 48 h inculture relative to an equal number of sorted cells at 24 h toevaluate the effect of ectopic Myc on terminal proliferation(Fig. 2B). We normalized the absolute cell numbers to ac-count for enucleation, where one erythroblast forms a nucleusand a reticulocyte, and demonstrated that continued Mycexpression at physiological levels had no effect on terminalproliferation. The absolute cell counts for Myc-GFPhigh cellsat 48 h were significantly lower than that of control (Fig. 2B),and this reduction was explained by the significant inductionof apoptosis by supraphysiological levels of Myc (Fig. 1B). Cell
cycle analysis was performed at 48 h in culture to investigatethe effect of ectopic Myc expression on the withdrawal fromcell cycle observed during the final stages of normal erythroidmaturation (Fig. 2C). Myc-GFPlow cells accumulated in theG1 phase of the cell cycle similar to the control cells. In con-trast, Myc-GFPhigh cells showed a signification population ofcells in the S-phase, indicating that the cells continue to cycle(Fig. 2C). The Myc-GFPhigh cells continued to proliferate inerythropoietin-free culture conditions (DMEM containing10% serum and antibiotics) for at least 8 weeks, prompting usto test whether these cells have been transformed. Mice trans-planted with these cultured Myc-GFPhigh erythroblasts didnot show evidence of erythroleukemia development even after6 months post-transplant, suggesting that ectopic expressionof Myc was not sufficient to fully transform erythroblasts intoerythroleukemias (data not shown). This is consistent withthe conclusions from a previous study (22), where retroviraloverexpression of Myc in murine fetal liver cells gave rise tocontinuously growing erythroblast cell lines but did not fullytransform them. These results indicate that Myc overexpres-sion at supraphysiological levels in early erythroblasts pro-motes proliferation at the expense of differentiation.To test the role of Myc in the rapid terminal prolifera-
tion of erythroid cells, we infected purified TER119-nega-tive erythroblasts with retroviruses harboring shRNAsagainst Myc or with retroviruses encoding non-silencingnegative control shRNA followed by culture for 48 h. Therewas no significant effect of knockdown of Myc on erythroiddifferentiation, enucleation, or apoptosis (Fig. 3A), but pro-
FIGURE 2. Ectopic Myc expression at supraphysiological levels inhibits hemoglobin expression and exit from cell cycle. TER119-negative fetal livererythroblasts were infected with bicistronic retroviruses encoding GFP only as control or both Myc and GFP at 0 h in culture. At 24 h post-infection, cellswere sorted based on GFP intensity into low (Myc-GFPlow) and high (Myc-GFPhigh) intensity populations and then cultured for another 24 h to allow fordifferentiation. A, relative hemoglobin content was quantified at 48 h post-infection by cell lysis in Drabkin’s reagent followed by measurement of absor-bance at 540 nm. B, cell counts were measured at 48 h post-infection for an equal starting cell number (25 � 104) at 24 h after sorting based on GFP inten-sity. The total cell numbers are represented as black bars. The numbers were normalized to account for enucleation using the equation n � total cell num-ber � (1 � percentage enucleation/100) and are shown as white bars. C, cell cycle analysis by flow cytometry at 48 h post-retroviral infection is shown. Allerror bars represent S.D. (n � 3). In all panels, two-tailed t test are indicated by as asterisk, p � 0.01.
Function of Myc in Terminal Erythroid Maturation
40256 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

liferation was significantly reduced in cells expressing MycshRNA (Fig. 3B). Western blot analysis confirmed the re-duction of Myc protein levels in cells expressing MycshRNA (Fig. 3C). The p27 protein was not increased uponknockdown of Myc, presumably because p27 protein ex-pression is already at its maximum in late stage erythro-blasts. Because Myc levels are normally down-regulatedduring terminal erythroid maturation, it was not surprisingthat knockdown of Myc had no adverse effects on terminaldifferentiation or enucleation. Interestingly, accelerateddepletion of Myc significantly reduced the proliferationrates of differentiating erythroid cells, suggesting that Myclevels may regulate the number of cell divisions during ter-minal erythroid differentiation.Terminal Erythroid Differentiation and Cell Cycle Arrest
Are Not Affected by Deficiency of p27—Myc overexpression atsupraphysiological levels in erythroid cells prevented theirwithdrawal from the cell cycle (Fig. 2C) and also diminishedthe induction of p27 during late stage erythroid maturation(Fig. 1A, lanes 7 and 9). p27 has been implicated in the cellcycle arrest during the final stages of erythroid maturation(29). This prompted us to investigate whether p27 protein
levels were the critical mediators of cell cycle arrest duringterminal erythroid differentiation and whether the effect ofMyc on cell cycle in erythroid cells was mediated throughits regulation of p27 protein levels. To this end, we purifiedTER119-negative fetal liver erythroblasts from E13.5 trans-genic embryos that were wild-type (p27�/�), heterozygous(p27�/�) or nullizygous (p27�/�) for the p27 locus andcompared their terminal differentiation in vitro to test ifp27 was necessary for normal terminal erythroid matura-tion and cell cycle arrest. Differentiation, enucleation, andapoptosis were quantified by flow cytometry at 48 h of cul-ture, and no significant difference was observed betweenthe p27�/�, p27�/�, and p27�/� cells (Fig. 4A). There wasalso no significant difference in the extent of G1 arrest (Fig.4B) or terminal proliferation (Fig. 4C) in the absence of oneor both alleles of p27. Western blot analysis was performedto confirm the complete absence of p27 protein in thep27�/� erythroblasts at 48 h in culture (Fig. 4D), and theprotein levels were intermediate in p27�/� cells consistentwith previous studies (24, 31, 32). Adult p27 knock-outmice are larger in body size than their wild-type counter-parts, but there are no apparent defects in the proportions
FIGURE 3. Depletion of Myc by shRNA inhibits terminal proliferation of erythroblasts without affecting terminal differentiation. A, purified TER119-negative erythroblasts were infected with retroviruses encoding shRNA against Myc or control retroviruses encoding non-silencing negative control shRNAfollowed by culture in erythropoietin containing medium. The cells were analyzed by flow cytometry after 48 h in culture. Erythroid differentiation was as-sayed by quantifying the CD71�TER119� population, which represents the late stage erythroid cells (left). Enucleation was assayed by quantifying theTER119�Hoechst� populations, which corresponds to the enucleated reticulocytes (middle). Apoptosis and cell death were assessed by co-staining withannexin V and propidium iodide (PI, right). B, cell counts were measured at 48 h in culture for the same cells as in panel A, for an equal starting cell number(50 � 104). The total cell numbers are represented as black bars. The numbers were normalized to account for enucleation using the equation n � total cellnumber � (1 � percentage enucleation/100) and are shown as white bars. Error bars represent S.D. (n � 3). Two-tailed t test are indicated by double aster-isks, p � 0.01. C, shown is Western blot analysis of the same cells as in panel A using antibodies against Myc and p27. GAPDH protein levels were determinedas a loading control.
Function of Myc in Terminal Erythroid Maturation
DECEMBER 17, 2010 • VOLUME 285 • NUMBER 51 JOURNAL OF BIOLOGICAL CHEMISTRY 40257
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

of various cell types in peripheral blood (32). These resultsdemonstrate that terminal erythroid differentiation andcell cycle arrest occurred normally even in the absence of
p27 and suggest that the effect of Myc overexpression oncell cycle progression in erythroid cells may not be medi-ated by p27 alone.
FIGURE 4. Terminal erythroid differentiation is not affected by deficiency of p27. TER119-negative fetal liver erythroblasts were purified from p27�/�,p27�/�, and p27�/� E13.5 mouse embryos and cultured in erythropoietin-containing medium in fibronectin-coated plates for 48 h. A, differentiation (top),enucleation (middle), and apoptosis (bottom) were quantified by flow cytometry at 48 h in culture, as described in Fig. 1B. PI, propidium iodide. B, shown iscell cycle analysis of cells harvested at 48 h in culture by flow cytometry. C, cell counts were measured at 48 h in culture for an equal starting cell number(1 � 105). The total cell numbers are represented as black bars. The numbers were normalized to account for enucleation as in Fig. 2C and are shown aswhite bars. All error bars represent S.D. (n � 3). D, Western blot analysis to determine p27 protein levels is shown. GAPDH protein levels were determined asa loading control.
Function of Myc in Terminal Erythroid Maturation
40258 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Gene Expression Analysis Demonstrates That Myc-GFPhighCells Are Similar to Early Erythroblasts, and Myc-GFPlowCells Resemble Late Stage Erythroid Cells—Because Myc is apleiotropic transcription factor regulating several diverse cel-lular functions (12–14), we used microarrays to investigatethe genes and pathways perturbed by ectopic Myc expressionat physiological and supraphysiological levels in erythroidcells. Total RNA was extracted fromMyc-GFPlow and Myc-GFPhigh cells at 48 h in culture post retroviral infection andfrom uninfected erythroblasts in culture at 0 (D0 WT), 24 (D1WT), and 48 h (D2 WT) as controls. RNA was then processedfor hybridization to Illumina microarrays. A hierarchical clus-tering analysis of the 2795 differentially expressed genes,identified based on an at least 2-fold change in at least one
condition compared with D0WT, was performed, and theresults are shown in Fig. 5A. Each sample contained 3 biologi-cal replicates (termed 1, 2, and 3) that correlated well witheach other. Cluster analysis showed that the mRNA expres-sion pattern of Myc-GFPhigh cells at 48 h in culture was mostsimilar to that of uncultured TER119-negative erythroblasts(D0 WT). The expression pattern of wild-type cells changedmarkedly during differentiation, comparing erythroblasts cul-tured for 48 h (D2 WT) against uncultured TER119-negativeerythroblasts (D0 WT). Importantly, the expression pattern ofMyc-GFPlow cells at 48 h in culture was most similar to thatof differentiated erythroblasts (D2 WT).The genes that are up-regulated, unchanged, or down-regu-
lated significantly in differentiated control erythroblasts (D2
FIGURE 5. Microarray analysis demonstrates that global gene expression of Myc-GFPhigh cells is similar to early erythroblasts and Myc-GFPlowcells resemble differentiated late stage erythroid cells. A, purified TER119-negative erythroblasts were cultured in erythropoietin-containing medium,and cells were harvested at 0 h (D0 WT), 24 h (D1 (WT), or 48 h (D2 WT) culture or were infected with bicistronic retroviruses encoding Myc and GFP at 0 h inculture and sorted based on GFP fluorescence into low (Myc-GFPlow) and high (Myc-GFPhigh) expression populations at 24 h post-infection followed byculturing for another 24 h. Total RNA was extracted from three biological replicates under each condition and processed for hybridization on Illumina mi-croarrays. Hierarchical clustering of all treatments and replicates was performed on Beadstudio software using Pearson correlation with nesting by averagelinkage method. The numerical scale shows the 1-r distance metric, where r is the Pearson correlation coefficient, and shorter branches denote greater simi-larity. The heatmap shows 2795 differentially expressed genes that are up-regulated (red) or down-regulated (green) by at least 2-fold in at least 1 conditioncompared with the average of the D0 WT replicate controls. The color scale bar indicates relative -fold change in gene expression compared with the aver-age of D0 WT replicate controls on a Log2 scale. B, qRT-PCR analysis of the expression of key erythroid differentiation genes (top), genes downstream oferythropoietin signaling (middle), and cell cycle genes (bottom) in Myc-GFPlow and Myc high cells at 48 h in culture relative to control cells infected withretroviruses encoding GFP only (D2 Control). All error bars represent S.D. (n � 3). The two-tailed t test results are indicated by an asterisks, p � 0.01.
Function of Myc in Terminal Erythroid Maturation
DECEMBER 17, 2010 • VOLUME 285 • NUMBER 51 JOURNAL OF BIOLOGICAL CHEMISTRY 40259
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

WT) relative to early erythroblasts (D0 WT) and are signifi-cantly perturbed upon ectopic Myc expression are denoted asgene subsets i, ii, and iii, respectively (Fig. 5A). The signifi-cantly enriched gene ontology categories for each of thesegene subsets were identified using Ingenuity Pathway Analysissoftware (supplemental Table S1). Gene subset i includesgenes that are normally up-regulated during erythroid differ-entiation. The up-regulation of these erythroid importantgenes is blocked in Myc-GFPhigh cells, consistent with theobserved differentiation block in these cells. A significantnumber of these genes were also perturbed in Myc-GFPlowcells, suggesting that differentiation is not completely normalin these cells, although hemoglobin levels and TER119 ex-pression were normal (Figs. 1 and 2). The genes that are notsignificantly induced or repressed during terminal erythroidmaturation but are significantly perturbed upon ectopic Mycexpression (Gene subset ii) were enriched for gene ontologycategories such as cell-to-cell signaling, cellular movement,general cellular maintenance, and metabolism (supplementalTable S1). The genes down-regulated during normal eryth-roid maturation (Gene subset iii) control cellular processesthat are shut down during late stage erythroid maturation;that is, gene expression, RNA post-transcriptional modifica-tion, cell death, and cell cycle and DNA replication and repair(supplemental Table S1). The expression pattern of thesegenes in Myc-GFPhigh cells was similar to that of early eryth-roblasts (D0 WT), and that of Myc-GFPlow cells was closer todifferentiated late stage erythroblasts (D2 WT).The expression of several key erythroid transcripts such as
the hemoglobin isoforms (Hbb-b1, Hbb-b2, and Hba-a1), theerythroid-specific membrane protein Spectrin �-chain(Spna1), and the erythroid-specific cell surface protein glyco-phorin A (GypA) were repressed in Myc-GFPhigh cells butwere normal in Myc-GFPlow cells, as confirmed by qRT-PCR(Fig. 5B, top panel). The expression of the master regulator ofterminal erythroid maturation GATA1 was not significantlychanged in either Myc-GFPlow or Myc-GFPhigh cells at 48 hin culture, suggesting that the differentiation block observedin Myc-GFPhigh cells is not due to repression of GATA1 ex-pression by Myc. However, the expression of several genesdownstream of the erythropoietin signaling pathway that havebeen implicated to be critical for normal terminal erythroiddifferentiation, such as protein kinase C� (Prkcd) (33), Januskinase 2 (Jak2) (34), protein kinase B� (Akt3) (35, 36), theanti-apoptotic protein Bcl-XL (37, 38), the small GTPase Rac2(8), and the inhibitory cyclin G2 (ccng2) (39), were signifi-cantly repressed in Myc-GFPhigh cells but were only mod-estly repressed or remained relatively normal in Myc-GFPlowcells (Fig. 5B,middle panel). This suggests that the dysregula-tion of the erythropoietin signaling pathway in Myc-GFPhighcells leads to the observed block in differentiation in Myc-GFPhigh cells. The expression of these genes changes mod-estly in Myc-GFPlow cells, suggesting that Myc-GFPlow cellsmay have other minor defects in differentiation that are notimmediately apparent in our assays. We also analyzed the ex-pression of positive regulators of the G1-S transition cell cyclecheckpoint in Myc-GFPlow and Myc-GFPhigh cells. The ex-pression of Cyclin E (25, 40) and cdc25a (41) was up-regulated
significantly in Myc-GFPhigh cells but not in Myc-GFPlowcells, whereas the expression of H-ras (23) and Ebp1 (42) in-creased in a dose-dependent manner upon increase in ectopicMyc expression (Fig. 5B, bottom panel). The expression ofp27, a negative regulator of G1-S transition, was significantlyrepressed in Myc-GFPhigh cells but only mildly reduced inMyc-GFPlow cells (Fig. 5B, bottom panel). Because we dem-onstrated that p27 is dispensable for normal terminal eryth-roid differentiation and cell cycle exit (Fig. 4), this result sug-gests that the induction of several positive regulators of theG1-S transition by ectopic expression of Myc at supraphysi-ological levels abrogates the normal G1-phase cell cycle arrest,whereas the changes in expression of these genes induced bycontinued Myc expression at physiological levels was insuffi-cient to overcome the normal erythroid cell cycle arrest.In summary, gene expression analysis demonstrated that
Myc-GFPlow cells resembled differentiated erythroid cellsand Myc-GFPhigh cells were similar to undifferentiated earlyerythroblasts. Ectopic Myc expression even at a physiologicallevel had significant effects on the gene expression patternsobserved in late stage erythroid cells, suggesting that althoughoverall terminal differentiation was normal in terms of hemo-globin levels and expression of major red cell proteins, thereare other abnormal changes in these cells besides a block inenucleation. Ectopic Myc expression at supraphysiologicallevels prevented the induction of several key erythroid genesand disrupted genes involved in the erythropoietin signalingpathway, leading to a block in terminal erythroid differentia-tion, and also induced the expression of proliferation-promot-ing genes preventing normal erythroid cell cycle exit.Ectopic Myc Expression Inhibits Nuclear Condensation and
Histone Deacetylation in Late Stage Erythroid Cells—Tofurther characterize the block in the ability of the Myc-express-ing cells to undergo enucleation, we performed benzidine-Giemsa staining on the cultured cells (Fig. 6A). The Myc-GFPhigh cells did not stain strongly with benzidine, showinga lack of hemoglobinization, and were similar in appearanceto early erythroblasts; that is, large cells with large nuclei. Thisfurther confirmed that Myc expression at supraphysiologicallevels arrested terminal differentiation at an early erythroblaststage. There was no apparent difference in benzidine stainingbetween Myc-GFPlow cells and control cells at 48 h in cul-ture, confirming that ectopic Myc expression at physiologicallevels had no effect on erythroid maturation and hemoglo-binization (Fig. 6A). Importantly, the nuclei in the late stageMyc-GFPlow erythroblasts were larger and more centralizedcompared with the control cells at 48 h in culture but weresmaller than the nuclei of erythroblasts in earlier stages ofdevelopment harvested at 0 and 24 h in culture (Fig. 6A). Toconfirm this observation, we used Cellomics high contentscreening to measure and compare the erythroid nuclear sizein a quantitative manner (Fig. 6B). The nuclear area per cellfor Myc-GFPhigh cells at 48 h was similar to that of controlerythroblasts at 24 h in culture. The nuclear area per cell wassignificantly higher for Myc-GFPlow cells compared withcontrol cells at 48 h in culture, and both values were signifi-cantly lower than those of erythroblasts in earlier stages ofdevelopment harvested at 0 and 24 h in culture (Fig. 6B).
Function of Myc in Terminal Erythroid Maturation
40260 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

These results demonstrate that ectopic Myc expression inhib-its nuclear condensation in late stage erythroid cells and sug-gests that the observed block in enucleation due to continuedMyc expression at physiological levels was a result of inhibi-tion of nuclear condensation.We, thus, investigated the molecular mechanism underly-
ing the inhibition of nuclear condensation and enucleation byMyc. Pharmacological inhibition of histone deacetylases(HDACs) has been reported to block chromatin condensationand enucleation in erythroid cells, demonstrating that globalhistone deacetylation observed during late stage erythroidmaturation is necessary for erythroid nuclear condensationand enucleation (43, 44). Recent studies have shown that Mycregulates global chromatin structure by influencing wide-spread histone modifications (45–47). We, thus, sought todetermine whether Myc expression induces changes in globalhistone acetylation in erythroid cells, possibly leading to theobserved block in nuclear condensation. Western blot analy-sis of histone acetylation was performed using antibodies de-tecting histones H3 and H4 acetylated at specific lysine resi-dues (Fig. 6C). Global levels of acetylation of histone H3 atLys-9 was maximal at 24 h in culture and decreased slightly at48 h in culture. Ectopic Myc expression reversed the observeddecrease in acetylation at H3K9 during late stage erythroid
maturation. There was no change in levels of H3K18 andH3K23 acetylation during the course of erythroid maturationwith or without ectopic Myc expression. Global levels of his-tone H4 acetylation at Lys-5, -8, -12, and -16 decreased at48 h in culture concomitant with the nuclear condensationobserved during late stage erythroid maturation, and this de-crease did not occur after ectopic Myc expression at eitherphysiological or supraphysiological levels. Hence, continuedMyc expression even at a physiological level inhibited the glo-bal decrease in histone acetylation observed during late stageerythroid maturation, suggesting that histone deacetylationmediated by down-regulation of Myc during terminal eryth-roid differentiation regulates nuclear condensation andenucleation.Because changes in global histone acetylation accompanied
the block in erythroid nuclear condensation by Myc, we in-vestigated the changes in expression of the major histoneacetyltranferases (HATs) and HDACs during terminal eryth-roid maturation and upon ectopic Myc expression (Fig. 7).The expression of the HATs Elp3, Gcn5, Hat1, and Tip60 de-creased during the course of erythroid maturation (Fig. 7A),suggesting that this decrease in expression of HATs may con-tribute to the decrease in global histone acetylation observedduring late stages of erythroid maturation. The expression of
FIGURE 6. Ectopic Myc expression inhibits nuclear condensation and histone deacetylation in late stage erythroid cells. Purified TER119-negativemouse fetal erythroblasts were infected with retroviruses encoding GFP only (MIG) or with retroviruses encoding both Myc and GFP at 0 h in culture andsorted based on GFP intensity into low (Myc-GFPlow) and high (Myc-GFPhigh) intensity populations at 24 h post infection followed by culture for another24 h to allow for differentiation in vitro. Uninfected erythroblasts were harvested at 0 and 24 h in culture as earlier time point controls for comparison.A, cytospin preparations of the harvested cells were stained with benzidine and Giemsa. The arrowhead indicates an enucleated reticulocyte. Arrows indi-cate erythroblasts with condensed nuclei that are at an early stage of enucleation. B, shown is measurement of nuclear area to quantify the degree of nu-clear condensation. The harvested cells were fixed with formaldehyde and incubated with Hoechst 33342 to stain the nucleus. Nuclear area per cell wasdetermined for each sample using high content screening, as described under “Experimental Procedures.” Error bars represent S.D. across three biologicalreplicates. The two-tailed t test are indicated by asterisks, p � 0.01. C, Western blot analysis of acetylation states of histones H3 and H4 was performed usingthe indicated acetylation-specific antibodies. Equal loading of samples was confirmed by stripping and reprobing the membranes with total histone H3 orH4 antibodies.
Function of Myc in Terminal Erythroid Maturation
DECEMBER 17, 2010 • VOLUME 285 • NUMBER 51 JOURNAL OF BIOLOGICAL CHEMISTRY 40261
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

the various HDACs was down-regulated in general during thecourse of erythroid maturation (Fig. 7B), except for HDAC5,which was up-regulated during late stages of erythroid matu-ration, as has been reported previously (43). Because ectopicMyc expression increased global histone acetylation, we ana-lyzed the expression changes of the HATs and HDACs uponectopic Myc expression to identify and select HATs that weresignificantly (at least 2-fold change relative to D2 MIG con-trol) up-regulated or HDACs that were significantly down-regulated for further functional studies. Ectopic Myc expres-sion led to a significant and dose-dependent increase in Gcn5expression at 48 h relative to control (D2 MIG), whereas theincrease in expression of the other HATs upon ectopic Mycexpression was not significant (Fig. 7A), and none of theHDACs tested was significantly down-regulated upon ectopicMyc expression (Fig. 7B). Gcn5 has been reported to functiondownstream of Myc in the regulation of global chromatinstructure (46). This prompted us to investigate the role ofGcn5 in terminal erythroid maturation and enucleation.Ectopic Gcn5 Expression Inhibits Late Stage Erythroid Nu-
clear Condensation and Enucleation—To directly investigatethe role of Gcn5 in terminal erythroid maturation and enucle-ation, we infected TER119-negative mouse fetal erythroblastswith retroviruses encoding Gcn5. The levels of Gcn5, Myc,and p27 proteins in cells infected with retroviruses encodingGcn5 were compared with the expression levels during nor-mal erythroid maturation and also with Myc-expressing cells
using Western blot analysis (Fig. 8A). Gcn5 protein levels nor-mally decreased substantially during late stage erythroid mat-uration, coinciding with the onset of nuclear condensation,similar to the decrease in Myc protein levels at the same stage(Fig. 8A, lanes 1–3). Cells infected with retroviruses encodingMyc showed overexpression of Myc protein at levels withinthe physiological range (Fig. 8A, lanes 1–4) and also displayedsignificantly increased Gcn5 protein expression (Fig. 8A, lanes3 and 4), consistent with the induction of Gcn5 transcripts byMyc (Fig. 7A). Cells infected with retroviruses encoding Gcn5displayed elevated levels of Gcn5 protein expression relativeto controls, but the levels of Myc and p27 were not affected(Fig. 8A, lanes 3 and 5). Ectopic Gcn5 expression inhibitederythroid enucleation significantly without affecting the dif-ferentiation, apoptotic status, or cell cycle arrest of the cul-tured erythroid cells (Fig. 8B). The hemoglobin content ofGcn5 expressing cells was similar to those of the controls (Fig.8C), further confirming that the inhibition of enucleation wasnot due to a block in erythroid differentiation. Ectopic Gcn5expression also did not affect the terminal proliferation ratesof the differentiating erythroblasts (Fig. 8D). Benzidine-Gi-emsa staining of Gcn5-expressing cells at 48 h in culture indi-cated late stage erythroblasts with larger nuclei relative tocontrols, with no apparent difference in hemoglobin content(Fig. 8E). High content screening-based measurement of nu-clear area confirmed that the average nuclear size of Gcn5-expressing erythroblasts was significantly higher than those of
FIGURE 7. Effect of ectopic Myc on the expression of HATs and HDACs. Cells were treated and harvested as described in Fig. 6, qRT-PCR analysis of theexpression of the indicated HATs (A) and HDACs (B) was performed, and the relative expression levels normalized to D2 MIG control cells are shown. All er-ror bars represent S.D. (n � 3). The two-tailed t test results are indicated by asterisks, p � 0.01.
Function of Myc in Terminal Erythroid Maturation
40262 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

control cells at 48 h in culture (Fig. 8F). Furthermore, West-ern blot analysis of global acetylation states of histones H3and H4 demonstrated that Gcn5 expression significantly re-versed the histone deacetylation at H3K9, H4K8, and H4K12normally observed during late stage erythroid maturation,similar to the effects of ectopic Myc expression (Fig. 8G).Gcn5, unlike Myc, did not affect the acetylation at H4K5 andH4K16 significantly. The acetylation states of H3K18 andH3K23, which did not change during the course of normalterminal erythroid maturation (Fig. 6C), remained unchangedupon Gcn5 expression (Fig. 8G). Because Gcn5 expression
was induced by ectopic Myc expression in erythroid cells, andbecause both were down-regulated substantially and simulta-neously in normal late stage erythroblasts (Fig. 8A), we rea-soned that Gcn5 could be a direct transcriptional target ofMyc in erythroid cells. Gcn5 has been reported to be a directMyc target gene in human fibroblasts (46). Thus, ChIP assayswere performed on purified TER119-negative mouse fetalerythroblasts using an antibody against Myc. The immuno-precipitated DNA was significantly enriched for Gcn5 pro-moter sequences relative to negative control regions chosen1500 bp away from the Gcn5 transcriptional start site (Fig.
FIGURE 8. Ectopic Gcn5 expression inhibits late stage erythroid nuclear condensation and enucleation. A, TER119-negative mouse fetal erythroblastswere cultured for 0 h (D0) and 24 h (D1) or were cultured for 48 h after infection with bicistronic retroviruses encoding GFP and Myc (D2 Myc), GFP, andGcn5 (D2 Gcn5) or GFP alone (D2 control). Total protein lysates from the cells were subjected to Western blot analysis using the indicated antibodies.B, TER119-negative mouse fetal erythroblasts were infected with bicistronic retroviruses encoding Gcn5 and GFP (Gcn5) or GFP alone (Control). The cells wereanalyzed by flow cytometry after 48 h in culture to quantify enucleation (i), differentiation (ii), and apoptosis (iii). The cells were fixed in ethanol, and cellcycle analysis was performed by flow cytometry (iv). The percentage of cells in G1, S, and G2 phase of cell cycle are shown. C, relative hemoglobin contentwas quantified by lysing cells in Drabkin’s reagent followed by measurement of absorbance at 540 nm. D, cell counts were measured at 48 h in culture forthe same cells as in panel B, with an equal starting cell number (1 � 105) at 0 h. The total cell numbers are represented as black bars. The number was nor-malized to account for enucleation and is shown as white bars. E, benzidine-Giemsa staining images of cells harvested at 48 h in culture are shown. Scalebars represent 20 �m. F, cells were harvested at 48 h in culture, fixed, and stained with Hoechst 33342, and nuclear area per cell was measured by high con-tent screening. G, TER119-negative fetal erythroblasts were cultured for 48 h after infection with retroviral vectors encoding GFP and Myc, GFP and Gcn5, orGFP alone (Control), and Western blot analysis of acetylation states of histones H3 and H4 was performed using the indicated antibodies. H, a ChIP assayshows Myc binding to the Gcn5 promoter. ChIP was performed on purified TER119-negative mouse fetal liver erythroblasts using an antibody against c-Myc. PCR primers were designed for prospective Myc binding regions in the Gcn5 promoter at �3 to �96 and �44 to �142 bp upstream of the Gcn5 tran-scriptional start site. The enrichment for Gcn5 promoter sequences in the immunoprecipitated DNA was quantified using qRT-PCR, relative to a negativecontrol region at �1596 to �1690 bp upstream of the Gcn5 transcriptional start site. All error bars represent S.D. (n � 3).
Function of Myc in Terminal Erythroid Maturation
DECEMBER 17, 2010 • VOLUME 285 • NUMBER 51 JOURNAL OF BIOLOGICAL CHEMISTRY 40263
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

8H), indicating that Gcn5 is transcriptionally regulated byMyc in erythroid cells. Taken together, these results suggestthat Gcn5 functions downstream of Myc in regulating histonedeacetylation, nuclear condensation, and enucleation inerythroid cells.
DISCUSSION
The present study uncovers the significance of changes inMyc expression in controlling mammalian terminal erythroiddifferentiation. Myc is normally down-regulated beginning atthe CFU-E stage. We show that overexpression of Myc at lev-els above that of these early erythroblasts promotes prolifera-tion at the expense of differentiation and is accompanied byinduction of apoptosis. In contrast, continued Myc expressionat physiological levels blocks enucleation specifically and in-hibits histone deacetylation and nuclear condensation in latestage erythroid cells. Our results support a model wherebydown-regulation of Myc from the high levels seen in earlyerythroblasts is essential for the progression of normal termi-nal erythroid differentiation.Upon erythropoietin induction of terminal differentiation, the
mouse erythroblasts undergo about 4–5 terminal cell divisionsin 48 h followed by a G1-phase cell cycle arrest. Themechanismof regulation of these rapid terminal cell divisions followed by aG1 arrest is not well understood. Up-regulation of p27 duringlate stage erythroid differentiation has been implicated in regu-lating the withdrawal of erythroid cells from cell cycle (29). How-ever, we demonstrated using p27 knock-out erythroblasts thatterminal erythroid differentiation is unaffected by the absence ofp27, suggesting that p27 alone is not sufficient to regulate the latestage G1 arrest in erythroid cells.We provide evidence suggest-ing thatMyc plays a critical role in controlling the number ofterminal cell divisions and the withdrawal from the cell cycle.shRNA-mediated depletion ofMyc in cultured erythroblastssignificantly inhibited terminal cell proliferation without affect-ing terminal differentiation, indicating that a precisely regulateddecrease inMyc levels during late stage erythroid maturationmay control the exact number of terminal cell divisions beforethe cell cycle arrest. Myc levels were normally dramatically re-duced during late stages of erythroid maturation, coinciding withthe arrest in G1-phase of the cell cycle. EctopicMyc expression atlevels above that of early erythroblasts promotes their prolifera-tion in culture for much longer times in an undifferentiated state.This suggests that down-regulation ofMyc levels below a criticalthreshold might be necessary for commitment toward both ter-minal erythroid maturation and the withdrawal from the cellcycle.Because of its well established roles in driving cell prolifera-
tion, repression of Myc during late stage erythroid maturationhas been suggested to be involved mainly in the withdrawalfrom cell cycle (20). Our data demonstrates that normaldown-regulation of Myc in late stage erythroid cells is essen-tial for nuclear condensation and enucleation, revealing anovel role for Myc in mammalian erythropoiesis. Myc isdown-regulated during the late stages of erythroid matura-tion, concomitant with the global histone deacetylation that iscritical for erythroid nuclear condensation and enucleation(43, 44). Ectopic Myc expression at physiological levels that
did not induce cell cycle re-entry was sufficient to block the glo-bal histone deacetylation and inhibit nuclear condensation andenucleation. These findings strongly support the emerging no-tion thatMyc regulates global chromatin structure by influenc-ing genome-wide histonemodifications (45–47).We showedthat the HATGcn5 was down-regulated during normal terminalerythroid maturation and was induced upon ectopicMyc expres-sion and demonstrated its important role in the regulation of thebalance between global histone acetylation and deacetylation andnuclear condensation in late stage erythroblasts. This function ofGcn5 is consistent with previous studies in yeast showing thatGcn5 was required either directly or indirectly for the acetylationof several sites in histones H3 andH4 (48). The inhibition of enu-cleation by Gcn5 expression was not as complete as that ofMyc,suggesting that there are additional mechanisms other than in-duction of Gcn5 by whichMyc can regulate erythroid chromatincondensation and enucleation.We previously reported that RacGTPases regulate the cytoskeletal reorganization leading to theextrusion of the condensed nucleus from late stage erythroblasts(8). EctopicMyc expression led to a modest, dose-dependentdecrease in the expression of Rac2, suggesting that this may addi-tionally contribute to the block in enucleation byMyc.Epigenetic regulation of nuclear condensation can explain
the selective transcription of certain critical genes such asBcl-XL (49) during late stage erythroid maturation when DNAreplication and RNA transcription in the condensed nucleusare largely shut down. It also allows for a rapid condensationand general inactivation of the erythroid nucleus in a mannersimilar to apoptotic nuclear condensation while also leavingthe general nuclear architecture and chromosomal DNA in-tact (50). Although our results are strongly consistent withepigenetic regulation of erythroid chromatin condensation(43), they do not exclude the possibility that other genes acti-vated by repression of Myc during late stage erythroid matu-ration may also act as important regulators of erythroid nu-clear condensation and/or enucleation.In many cell types ectopic Myc expression promotes prolif-
eration and blocks terminal differentiation, which are essen-tial outcomes leading to tumor formation (51). Myc also in-duces apoptosis through the Arf-Mdm2-p53 tumorsuppressor pathway (52, 53), and additional mutations thatovercome the induction of apoptosis by Myc lead to tumori-genesis. We demonstrate here that the dosage of Myc is criti-cal in determining the phenotypic outcome of ectopic Mycexpression on differentiation, proliferation, and survival oferythroid cells. Continued Myc expression at physiologicallevels did not elicit an apoptotic response in cultured eryth-roid cells and broadly allowed for terminal maturation andexit from the cell cycle, with mainly nuclear condensation andenucleation being affected. High levels of ectopic Myc in-duced several proliferation-promoting genes and repressedthe expression of several genes in the erythropoietin signalingpathway, leading to continued proliferation at the expense ofdifferentiation. Supraphysiological levels ofMyc expression alsoinduced significant levels of apoptosis. This supports the theorythat the apoptotic response is a safeguard against the abnormalproliferationmediated by oncogene activation as in the case ofhigh level ectopicMyc overexpression and that low levels of ec-
Function of Myc in Terminal Erythroid Maturation
40264 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 51 • DECEMBER 17, 2010
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

topicMyc that do not induce cell cycle re-entry do not evoke theapoptotic response either. An elegant study using an in vivomodel ofMyc-induced tumorigenesis previously showed thatactivation of the apoptotic pathways required high level Mycoverexpression, and low level deregulatedMyc was potentiallyoncogenic in several but not all tissue types by avoiding the en-gagement of tumor suppression pathways (54). In our study us-ing primary erythroid cells cultured in vitro, low-doseMyc ex-pression did not induce cell cycle re-entry or a complete block indifferentiation. Hence, expression levels ofMyc protein togetherwith other factors such as cell type (26) and developmental stageand context (55) control the phenotypic outcome of ectopic orderegulatedMyc expression. A detailed understanding of thefunctioning of all these factors in combination would lead to bet-ter therapeutic intervention in cancers that involve deregulatedMyc expression.
Acknowledgments—We thank all our colleagues in Biopolis and theWhitehead Institute for valuable discussions and the staff at BiopolisShared Facilities for providing the flow cytometry and High ContentScreening services and advice. We are thankful to Dr. Motomi Osatoand Dr. Bindya Jacob for help with mice bone marrow transplants.
REFERENCES1. Koury, M. J., and Bondurant, M. C. (1988) J. Cell. Physiol. 137, 65–742. Richmond, T. D., Chohan, M., and Barber, D. L. (2005) Trends Cell Biol.
15, 146–1553. Eshghi, S., Vogelezang, M. G., Hynes, R. O., Griffith, L. G., and Lodish,
H. F. (2007) J. Cell Biol. 177, 871–8804. Koury, S. T., Koury, M. J., and Bondurant, M. C. (1988) Exp. Hematol.
16, 758–7635. Skutelsky, E., and Danon, D. (1967) J. Cell Biol. 33, 625–6356. Simpson, C. F., and Kling, J. M. (1967) J. Cell Biol. 35, 237–2457. Repasky, E. A., and Eckert, B. S. (1981) Prog. Clin. Biol. Res. 55, 679–6928. Ji, P., Jayapal, S. R., and Lodish, H. F. (2008) Nat. Cell Biol. 10, 314–3219. Vennstrom, B., Sheiness, D., Zabielski, J., and Bishop, J. M. (1982) J. Vi-
rol. 42, 773–77910. Oster, S. K., Ho, C. S., Soucie, E. L., and Penn, L. Z. (2002) Adv. Cancer
Res. 84, 81–15411. Meyer, N., and Penn, L. Z. (2008) Nat. Rev. Cancer 8, 976–99012. Dang, C. V., O’Donnell, K. A., Zeller, K. I., Nguyen, T., Osthus, R. C.,
and Li, F. (2006) Semin. Cancer Biol. 16, 253–26413. Hoffman, B., and Liebermann, D. A. (2008) Oncogene 27, 6462–647214. Iritani, B. M., and Eisenman, R. N. (1999) Proc. Natl. Acad. Sci. U.S.A.
96, 13180–1318515. Hoffman, B., Amanullah, A., Shafarenko, M., and Liebermann, D. A.
(2002) Oncogene 21, 3414–342116. Dubois, N. C., Adolphe, C., Ehninger, A., Wang, R. A., Robertson, E. J.,
and Trumpp, A. (2008) Development 135, 2455–246517. Wilson, A., Murphy, M. J., Oskarsson, T., Kaloulis, K., Bettess, M. D.,
Oser, G. M., Pasche, A. C., Knabenhans, C., Macdonald, H. R., andTrumpp, A. (2004) Genes Dev. 18, 2747–2763
18. Spangler, R., and Sytkowski, A. J. (1992) Blood 79, 52–5719. Spotts, G. D., and Hann, S. R. (1990)Mol. Cell. Biol. 10, 3952–396420. Rylski, M.,Welch, J. J., Chen, Y. Y., Letting, D. L., Diehl, J. A., Chodosh,
L. A., Blobel, G. A., andWeiss, M. J. (2003)Mol. Cell. Biol. 23, 5031–504221. Acosta, J. C., Ferrandiz, N., Bretones, G., Torrano, V., Blanco, R., Rich-
ard, C., O’Connell, B., Sedivy, J., Delgado, M. D., and Leon, J. (2008)Mol.Cell. Biol. 28, 7286–7295
22. Cory, S., Maekawa, T., McNeall, J., and Metcalf, D. (1991) Cell GrowthDiffer. 2, 165–172
23. Zhang, J., Socolovsky, M., Gross, A. W., and Lodish, H. F. (2003) Blood102, 3938–3946
24. Kiyokawa, H., Kineman, R. D., Manova-Todorova, K. O., Soares, V. C.,Hoffman, E. S., Ono, M., Khanam, D., Hayday, A. C., Frohman, L. A.,and Koff, A. (1996) Cell 85, 721–732
25. Aleem, E., Kiyokawa, H., and Kaldis, P. (2005) Nat. Cell Biol. 7, 831–83626. Luo, H., Li, Q., O’Neal, J., Kreisel, F., Le Beau, M. M., and Tomasson,
M. H. (2005) Blood 106, 2452–246127. Xu, W., Edmondson, D. G., and Roth, S. Y. (1998)Mol. Cell. Biol. 18,
5659–566928. Yang, W., Shen, J., Wu, M., Arsura, M., FitzGerald, M., Suldan, Z., Kim,
D. W., Hofmann, C. S., Pianetti, S., Romieu-Mourez, R., Freedman, L. P.,and Sonenshein, G. E. (2001) Oncogene 20, 1688–1702
29. Hsieh, F. F., Barnett, L. A., Green, W. F., Freedman, K., Matushansky, I.,Skoultchi, A. I., and Kelley, L. L. (2000) Blood 96, 2746–2754
30. Liu, X., Constantinescu, S. N., Sun, Y., Bogan, J. S., Hirsch, D., Weinberg,R. A., and Lodish, H. F. (2000) Anal. Biochem. 280, 20–28
31. Nakayama, K., Ishida, N., Shirane, M., Inomata, A., Inoue, T., Shishido,N., Horii, I., Loh, D. Y., and Nakayama, K. (1996) Cell 85, 707–720
32. Fero, M. L., Rivkin, M., Tasch, M., Porter, P., Carow, C. E., Firpo, E.,Polyak, K., Tsai, L. H., Broudy, V., Perlmutter, R. M., Kaushansky, K.,and Roberts, J. M. (1996) Cell 85, 733–744
33. Leng, L., Yu, F., Dong, L., Busquets, X., Osada, S., Richon, V. M., Marks,P. A., and Rifkind, R. A. (1993) Cancer Res. 53, 5554–5558
34. Neubauer, H., Cumano, A., Muller, M., Wu, H., Huffstadt, U., and Pfef-fer, K. (1998) Cell 93, 397–409
35. Ghaffari, S., Kitidis, C., Zhao, W., Marinkovic, D., Fleming, M. D., Luo,B., Marszalek, J., and Lodish, H. F. (2006) Blood 107, 1888–1891
36. Zhao, W., Kitidis, C., Fleming, M. D., Lodish, H. F., and Ghaffari, S.(2006) Blood 107, 907–915
37. Socolovsky, M., Fallon, A. E., Wang, S., Brugnara, C., and Lodish, H. F.(1999) Cell 98, 181–191
38. Dolznig, H., Habermann, B., Stangl, K., Deiner, E. M., Moriggl, R., Beug,H., and Mullner, E. W. (2002) Curr. Biol. 12, 1076–1085
39. Fang, J., Menon, M., Kapelle, W., Bogacheva, O., Bogachev, O., Houde,E., Browne, S., Sathyanarayana, P., and Wojchowski, D. M. (2007) Blood110, 2361–2370
40. Koff, A., Giordano, A., Desai, D., Yamashita, K., Harper, J. W., Elledge,S., Nishimoto, T., Morgan, D. O., Franza, B. R., and Roberts, J. M. (1992)Science 257, 1689–1694
41. Ray, D., and Kiyokawa, H. (2007) Cell Cycle 6, 3039–304242. Zhang, Y., Lu, Y., Zhou, H., Lee, M., Liu, Z., Hassel, B. A., and Ham-
burger, A. W. (2008) BMC Cell Biol. 9, 6943. Popova, E. Y., Krauss, S. W., Short, S. A., Lee, G., Villalobos, J., Etzell, J.,
Koury, M. J., Ney, P. A., Chasis, J. A., and Grigoryev, S. A. (2009) Chro-mosome Res 17, 47–64
44. Ji, P., Yeh, V., Ramirez, T., Murata-Hori, M., and Lodish, H. F. Haemato-logica (2010) doi: 10.3324/haematol.2010.029827
45. Knoepfler, P. S. (2007) Cancer Res. 67, 5061–506346. Knoepfler, P. S., Zhang, X. Y., Cheng, P. F., Gafken, P. R., McMahon,
S. B., and Eisenman, R. N. (2006) EMBO J. 25, 2723–273447. Guccione, E., Martinato, F., Finocchiaro, G., Luzi, L., Tizzoni, L., Dall’
Olio, V., Zardo, G., Nervi, C., Bernard, L., and Amati, B. (2006) Nat. CellBiol. 8, 764–770
48. Zhang, W., Bone, J. R., Edmondson, D. G., Turner, B. M., and Roth, S. Y.(1998) EMBO J. 17, 3155–3167
49. Gregoli, P. A., and Bondurant, M. C. (1997) Blood 90, 630–64050. Krauss, S. W., Lo, A. J., Short, S. A., Koury, M. J., Mohandas, N., and
Chasis, J. A. (2005) Blood 106, 2200–220551. Janz, S. (2005) Oncogene 24, 3541–354352. Hermeking, H., and Eick, D. (1994) Science 265, 2091–209353. Eischen, C. M., Weber, J. D., Roussel, M. F., Sherr, C. J., and Cleveland,
J. L. (1999) Genes Dev. 13, 2658–266954. Murphy, D. J., Junttila, M. R., Pouyet, L., Karnezis, A., Shchors, K., Bui,
D. A., Brown-Swigart, L., Johnson, L., and Evan, G. I. (2008) Cancer Cell14, 447–457
55. Beer, S., Zetterberg, A., Ihrie, R. A., McTaggart, R. A., Yang, Q., Bradon,N., Arvanitis, C., Attardi, L. D., Feng, S., Ruebner, B., Cardiff, R. D., andFelsher, D. W. (2004) PLoS Biol. 2, e332
Function of Myc in Terminal Erythroid Maturation
DECEMBER 17, 2010 • VOLUME 285 • NUMBER 51 JOURNAL OF BIOLOGICAL CHEMISTRY 40265
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from

LodishSenthil Raja Jayapal, Kian Leong Lee, Peng Ji, Philipp Kaldis, Bing Lim and Harvey F.
Down-regulation of Myc Is Essential for Terminal Erythroid Maturation
doi: 10.1074/jbc.M110.181073 originally published online October 12, 20102010, 285:40252-40265.J. Biol. Chem.
10.1074/jbc.M110.181073Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2010/10/12/M110.181073.DC1
http://www.jbc.org/content/285/51/40252.full.html#ref-list-1
This article cites 55 references, 29 of which can be accessed free at
by guest on September 9, 2020
http://ww
w.jbc.org/
Dow
nloaded from