
DOKTORA TEZİ - Hoşgeldinizacikarsiv.ankara.edu.tr/browse/25121/Binder1.pdf · pü ve rektal swab...
126
ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ DOKTORA TEZİ Enterococcus faecium KLİNİK İZOLATLARINDA ÇOKLU İLAÇ DİRENÇLİLİK MEKANİZMALARININ GENETİK DOĞASI Kübra KASAROĞLU BİYOLOJİ ANABİLİM DALI ANKARA 2013 Her Hakkı Saklıdır
Transcript of DOKTORA TEZİ - Hoşgeldinizacikarsiv.ankara.edu.tr/browse/25121/Binder1.pdf · pü ve rektal swab...

ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
DOKTORA TEZİ
Enterococcus faecium KLİNİK İZOLATLARINDA ÇOKLU İLAÇ DİRENÇLİLİK MEKANİZMALARININ GENETİK DOĞASI
Kübra KASAROĞLU
BİYOLOJİ ANABİLİM DALI
ANKARA 2013
Her Hakkı Saklıdır

i
ÖZET
Doktora Tezi
Enterococcus faecium KLİNİK İZOLATLARINDA ÇOKLU İLAÇ DİREÇLİLİK
MEKANİZMALARININ GENETİK DOĞASI
Kübra KASAROĞLU
Ankara Üniversitesi
Fen Bilimleri Enstitüsü
Biyoloji Anabilim Dalı
Danışman: Prof. Dr. Mustafa AKÇELİK
İnsanlarda gastrointestinal sistem florasının elemanı olan enterokoklar, önemli hastane
infeksiyonu etkenleri arasındadır. Çalışmamızda Akdeniz Üniversitesi Tıp Fakültesi
hastanesinde çeşitli klinik servislerde yatmakta olan hastalardan alınan idrar, bos, kan, katater,
pü ve rektal swab örneklerinden elde edilen kültür koleksiyonundan seçilen 64 Enterococcus
faecium suşunda çoklu ilaç dirençliliklerinin belirlenmesi amaçlanmıştır. Suşların
identifikasyonunda Phonex antibiyogram cihazı ve rapid ID 32 Strep APİ kiti kullanılmıştır.
Antibiyogram duyarlılıkları ise Phonex antibiyogram cihazı ve disk difüzyon yöntemleri
kullanılarak gerçekleştirilmiştir. Suşlardaki direnç yüzdeleri; ampsiline % 100, streptomisine %
87.5, kanamisine % 100, gentamisine % 67.18, kloramfenikole % 32.81, tetrasikline % 21.87,
rifampisine % 100, vankomisine % 96.87, eritromisine % 100 ve teikoplanine % 79.68 olarak
belirlenmiştir. Antibiyotik direncinden sorumlu genlerin belirlenmesi amacıyla polimeraz zincir
reaksiyonu (PZR) yöntemi kullanılmıştır. PZR ürünleri dizi analizi ile doğrulanmıştır.
Çalışmamızda Van B tipi direnç içeren suş saptanmış ve Türkiye’den bildirilen ikinci olgu
olduğu görülmüştür. Ayrıca antibiyotik direnç genlerinin konjugasyon yolu ile aktarımı
araştırılmış vanA direnç geni duyarlı suşlara aktarılabilmiştir.
Şubat 2013, sayfa 116
Anahtar Kelimeler: E. faecium, Antibiyotik Direnç, Konjugasyon

ii
ABSTRACT
Ph.D. Thesis
GENETİC NATURE OF MULTİ DRUG RESİSTANCE MECHANİSMS İN THE CLİNİCAL
ISOLATES OF Enterococcus faecium
Kübra KASAROĞLU
Ankara University
Graduate School of Natural and Applied Sciences
Department of Biology
Supervisor: Prof. Dr. Mustafa AKÇELİK
The enterococci which are the elements of gastrointestinal system flora, are some of the
important factors of nosocomial infections. In our study, our intention was to determine muti-
drug resistancy of 64 Enterococcus faecium strains which are chosen from bacterial cultures
obtained from the urine, CSF, blood, catheter, pus and rectal swab specimens taken from
hospitalized patients at various clinical departments of Akdeniz University Faculty of Medicine.
Phonex antibiogram device and rapid ID 32 Strep API kit were used for the identifications of
strains. Antibiogram susceptibilities were carried out by using Phonex antibiogram device and
disc diffusion methods. The percentages of resistance in strains are determined as; ampsiline %
100, streptomycin % 87.5, kanamycin % 100, gentamycin % 67.18, chloramphenicol % 32.81,
tetracycline % 21.87, rifampycin % 100, vancomycin % 96.87, erythromycin % 100 and
teicoplanin % 79.68. Polymerase chain reaction (PCR) method was used in order the determine
the genes responsible for antibiotic resistance. PCR products was confirmed by sequence
analysis. In our study, strains with Van B type resistance are detected and found to be the
second case reported from Turkey. Also transfer of the antibiotic resistance genes by
conjugation was researched, vanA resistance gene were able to transfered to sensitive strains.
February 2013, 116 pages
Key Words: E. faecium, Antibiotic Resistance, Conjugation

iii
TEŞEKKÜR
Lisans eğitimimim başından itibaren tüm çalışmalarımda beni yönlendiren, bilgi, sabır
ve tecrübelerini benden esirgemeyen, yanında çalışmaktan onur duyduğum danışman
hocam Prof. Dr. Mustafa AKÇELİK’e (Ankara Üniversitesi Biyoloji Anabilim Dalı),
çalışmam süresince yardımını esirgemeyen Yrd. Doç. Dr. Nefise AKÇELİK’e (Ankara
Üniversitesi Biyoteknoloji Enstitüsü);
Çalışmalarım sırasında tezime önemli katkılarda bulunan Akdeniz Üniversitesi
Mikrobiyoloji Anabilim Dalı Öğretim Üyelerinden Sayın Prof. Dr. Dilara Öğünç’e
Çalışmalarımın deneysel aşamalarında bana yardımcı olan başta Fatma Neslihan
YÜKSEL’e ve Prokaryot Genetiği Laboratuvarındaki arkadaşlarım Arş. Gör. Başar
KARACA (Ankara Üniversitesi Biyoloji Anabilim Dalı), Arş. Gör. Neslihan
KARATUĞ (Ankara Üniversitesi Biyoloji Anabilim Dalı), Özlem GÜNAY, Maryam
DİANİ, Muhammad Nima BADALİ, Barış YAVAŞ’a ve diğer çalışma arkadaşlarıma,
Benden hiçbir zaman destek ve yardımlarını esirgemeyen Akdeniz Üniversitesi Kültür
Laboratuvarındaki arkadaşlarım, Serpil TOSUN, Özlem KARAGÖZ, Bengü ÇAĞLAR,
Sedat KUZU ve diğer çalışma arkadaşlarıma,
Bu günlere gelmemi benden daha çok arzulayan babam İsmail YILDIRIM, annem
Neyran YILDIRIM, kardeşlerim Aslıhan YILDIRIM, Nihal YILDIRIM’a ve beni her
zaman izlediklerini bildiğim sevgili amcam Ali YILDIRIM ve Mehmet YILDIRIM’a
Manevi desteklerini hep yanımda hissettiğim ikinci ailem A. Nurhan KASAROĞLU,
Serap KASAROĞLU ve Z. Müge KASAROĞLU’na
Her anını beraber yürüdüğüm hayatta, elimi hiç bırakmayan ve başarmamı sağlayan ruh
eşim S. Volkan KASAROĞLU’na
Sonsuz teşekkürlerimi sunarım.
Kübra KASAROĞLU
Ankara, Şubat 2013

iv
İÇİNDEKİLER
ÖZET ................................................................................................................................ i
ABSTRACT ..................................................................................................................... ii
TEŞEKKÜR ................................................................................................................... iii
SİMGELER DİZİNİ ..................................................................................................... vii
ŞEKİLLER DİZİNİ ..................................................................................................... viii
ÇİZELGELER DİZİNİ ................................................................................................. ix
1. GİRİŞ ........................................................................................................................... 1
2. KURAMSAL TEMELLER ........................................................................................ 2
2.1 Enterococcus Cinsinin Genel Özellikleri ................................................................ 2
2.2 Enterokok Enfeksiyonlarının Epidemiyolojisi ....................................................... 7
2.3 Enterokok Enfeksiyonları ........................................................................................ 9
2.3.1 Üriner sistem enfeksiyonları ............................................................................... 10
2.3.2 Endokardit ............................................................................................................ 10
2.3.2 Bakteriyemi ........................................................................................................... 10
2.3.4 İntraabdominal ve pelvik enfeksiyonlar ............................................................ 11
2.3.5 Yara ve yumuşak doku enfeksiyonları ............................................................... 11
2.3.6 Neonatal enfeksiyonlar ........................................................................................ 11
2.3.7 Menenjit ................................................................................................................ 12
2.3.8 Solunum yolu enfeksiyonları ............................................................................... 12
2.4 Enterokoklarda Bulunan Virülans Faktörler ve Patojenite ............................... 12
2.5 Enterokoklarda Antibiyotik Direnci ve Direnç Mekanizmaları ......................... 15
2.5.1 Doğal (kromozomal) direnç ................................................................................. 15
2.5.2 Kazanılmış direnç ................................................................................................ 16
2.5.2.1 Beta laktam direnci ........................................................................................... 17
2.5.2.2 Glikopeptit direnci ............................................................................................ 18
2.5.2.2.1 Van A tipi direnç ............................................................................................ 21
2.5.2.2.2 Van B tipi direnç ............................................................................................ 22
2.5.2.2.3 Van C tipi direnç ............................................................................................ 23
2.5.2.2.4 Van D tipi direnç ............................................................................................ 23
2.5.2.3 Aminoglikozitlere direnç .................................................................................. 24

v
2.5.2.3.1 Permeabiliteye bağlı direnç .......................................................................... 24
2.5.2.3.2 Aminoglikozit modifiye edici enzimlere bağlı direnç .................................. 24
2.5.2.3.3 Ribozomal direnç .......................................................................................... 25
2.5.2.4 Kloramfenikole direnç ..................................................................................... 25
2.5.2.5 Tetrasiklinlere direnç ....................................................................................... 25
2.5.2.6 Kinolonlara direnç ............................................................................................ 26
2.3.2.7 Makrolitlere direnç ........................................................................................... 26
3. MATERYAL VE YÖNTEM ................................................................................... 28
3.1 Materyal ................................................................................................................... 28
3.1.1 Bakteriler .............................................................................................................. 28
3.1.2 Besiyerleri ............................................................................................................. 28
3.1.3 Bakterilerin tür teşhisi ......................................................................................... 29
3.1.4 Antibiyotikler ....................................................................................................... 31
3.1.5 Primerler ............................................................................................................... 32
3.2 Yöntem ..................................................................................................................... 33
3.2.1 İzolatlarının saflık kontrolü ve tür teşhisinin doğrulanması ........................... 33
3.2.2 Antimikrobiyal dirençliliğin belirlenmesi ......................................................... 36
3.2.3 İzolatların plazmid profillerinin belirlenmesi ................................................... 37
3.2.3.1 Plazmid izolasyonu ............................................................................................ 37
3.2.3.2 Elektroforez ....................................................................................................... 39
3.2.3.3 Plazmid büyüklüklerinin saptanması .............................................................. 40
3.2.4 Genomik DNA izolasyonu .................................................................................. 41
3.2.5 Polimeraz zincir reaksiyonu (PZR) ................................................................... 43
3.2.5.1 Antibiyotik dirençliliği kodlayan gen bölgelerinin PZR ile araştırılması .... 44
3.2.6 DNA dizi analizi ................................................................................................... 54
3.2.7 Konjugasyon denemeleri ..................................................................................... 54
4. BULGULAR VE TARTIŞMA ................................................................................. 55
4.1 Enterococcus faecium Suşlarının Antibiyotik Duyarlılık Düzeylerinin
Tanımlanması .......................................................................................................... 55
4.2 Enterococcus faecium Suşlarının Plazmid Profillerinin Tanımlanması ............. 77
4.3 Antibiyotik Direnç Genlerinin PZR ile Araştırılması ......................................... 82
4.3.1 vanA ve vanB gen bölgeleri .................................................................................. 82

vi
4.3.2 tetM ve tetL gen bölgeleri ..................................................................................... 85
4.3.3 ermB gen bölgesi ................................................................................................... 87
4.3.4 rpoB1 ve rpoB2 gen bölgeleri ............................................................................... 89
4.3.5 cat gen bölgesi ....................................................................................................... 92
4.3.6 Yüksek düzey aminoglikozit direncinden sorumlu gen bölgeleri .................... 94
4.3.7 blaZ gen bölgesi .................................................................................................... 99
4.4 Konjugasyon ............................................................................................................ 99
5. SONUÇ ..................................................................................................................... 103
KAYNAKLAR ............................................................................................................ 104
ÖZGEÇMİŞ ................................................................................................................ 115

vii
SİMGELER VE KISALTMALAR DİZİNİ
µg Mikrogram
AME Aminoglikozit Modifiye Edici Enzim
Bç Baz Çifti
BLAST Basic Local Alignment Search Tool
CLSI Klinik Laboratuar Standartları Enstitüsü
dNTP Deoksinükleotidtrifosfat
EARSS Avrupa Antibiyotik Direnci Sürveyans Sistemi
EDTA Etilen Diamin Tetraasetik Asit
HLR Yüksek Seviyede Direnç
Kb Kilobaz
M Molar
MHB Müller Hinton Broth
MİK Minimal İnhibisyon Konsantrasyonu
mL Mililitre
mM Milimolar
PAI Patojenite Adası
PBP Penisilin Bağlayan Protein
PZR Polimeraz Zincir Reaksiyonu
UV Ultra Viyole
VRE Vankomisin Dirençli Enterokok

viii
ŞEKİLLER DİZİNİ
Şekil 2.1 Enterokokların diğer Gram pozitif ve katalaz negatif koklardan ayrımı .......... 4
Şekil 2.2 Enterokok türlerinin fenotipik özellikleri ......................................................... 6
Şekil 2.3 Avrupa’da VRE prevalansı (2010) ................................................................... 8
Şekil 2.4 Avrupa’da VRE prevalansı (2011) ................................................................... 8
Şekil 2.5 Vankomisin antibiyotiğinin ve vankomisine direncli enterokoklarda
VanA direncinin hücre duvar öncülerine etki mekanizması .......................... 20
Şekil 2.6 Vankomisin direncinden sorumlu Tn1546 transpozonundaki van A gen
kümesinin genetik haritası .............................................................................. 21
Şekil 3.1 Enterokok izolatlarının tanısında kullanılan testler ......................................... 34
Şekil 3.2 rapid ID 32 Strep Api testi ile Enterococcus faecium suşlarının
doğrulanması……………………………………………………………….....35
Şekil 4.1 Enterococcus faecium suşlarının disk difüzyon testi ..................................... .55
Şekil 4.2 Enterococcus faecium suşlarının antibiyotik dirençlilik yüzdeleri................. 74
Şekil 4.3 Enterococcus faecium suşlarının plazmid içerikleri ....................................... 78
Şekil 4.4 vanA gen bölgesi ............................................................................................. 83
Şekil 4.5 vanB gen bölgesi ............................................................................................. 84
Şekil 4.6 tetM gen bölgesi ............................................................................................. 86
Şekil 4.7 tetL gen bölgesi ............................................................................................... 86
Şekil 4.8 ermB gen bölgesi ............................................................................................ 87
Şekil 4.9 rpoB1 gen bölgesi ........................................................................................... 90
Şekil 4.10 rpoB2 gen bölgesi .......................................................................................... 91
Şekil 4.11 cat gen bölgesi ............................................................................................... 93
Şekil 4.12 aac6-aph2 gen bölgesi ................................................................................... 94
Şekil 4.13 ant6 gen bölgesi ............................................................................................. 96
Şekil 4.14 aphA3 gen bölgesi ......................................................................................... 97
Şekil 4.15 vanA geninin konjugal aktarımı ................................................................... 101
Şekil 4.16 Transkonjugantta vankomisin direncinin disk difüzyon yöntemi ile
doğrulanması ................................................................................................ 101

ix
ÇİZELGELER DİZİNİ
Çizelge 2.1 Enterococcus cinsinde tanımlanan virülans faktörler ................................. 14
Çizelge 2.2 Enterokoklarda glikopeptid direnç tipleri .................................................... 19
Çizelge 3.1 Çalışmada kullanılan bakteriler ve izolasyon materyalleri ......................... 30
Çizelge 3.2 Çalışmada kullanılan antibiyotikler ve disk konsantrasyonları ................... 31
Çizelge 3.3 Antibiyotik direnç genlerinin tespiti için kullanılan primerler .................... 32
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik
duyarlılıkları ................................................................................................ 56
Çizelge 4.2 Standart E. faecalis ATCC29212 suşunun disk difüzyon ve MİK
testlerine göre antibiyotik duyarlılığı .......................................................... 72
Çizelge 4.3 E. faecium suşlarının antibiyotik gruplarına karşı direnç oranları .............. 73

1
1. GİRİŞ
Enfeksiyon hastalıklarının tedavisinde karşılaşılan en önemli sorun, antibiyotik
kullanımına bağlı olarak bakterilerde yaygınlaşan antibiyotik dirençlilik olarak
tanımlanmaktadır. Önceleri önemsiz olarak görülen pek çok bakteri şu anda ciddi
hastane enfeksiyonlarına neden olmaktadır. Normal florada bulunan ve ilk
tanımlandıkları yıllarda sadece endokardit etkeni olarak bildirilen enterokoklar,
bakterisidal antimikrobiyal terapiyi gerektiren nozokomiyal patojenler olarak son 20
yılda önem kazanmışlardır (Huycke vd. 1998). Bu artışın en önemli nedeni;
enterokokların hastanelerde sıklıkla kullanılan 3. kuşak sefalosporinler gibi birçok
antibiyotiğe doğal olarak dirençlilik içermeleri ve kullanılabilen tüm antibiyotiklere
karşı direnç geliştirebilme özelliğine sahip olmalarıdır. β-laktamlara ve düşük
konsantrasyonlardaki aminoglikozitlere karşı doğal direnç içermeleri nedeniyle, her iki
sınıfa karşı yüksek düzeyde direnç oluşumu gerçekleştiğinde ilaçların sinerjetik etkisi
ortadan kalkmakta ve tedavi güçleşmektedir.
Enterokoklar klasik virülans faktörlerine sahip olmasa da, çoklu antibiyotik
dirençlilikleri onlara antibiyotik tedavisi altında yaşama ve çoğalma olanağı
sağlamaktadır. Direnç determinantlarının enterokoklar arasında ve özellikle diğer Gram-
pozitif bakterilere plazmid ve transpozonlar aracılığıyla aktarımı, direncin hızla
yayılmasına neden olmaktadır. Enterokoklarda doğru tür teşhisi, antibiyotik duyarlılık
testlerinin optimize edilmesi ve direnç kaynaklarının genetik temelinin araştırılması;
direncin önlenmesi ve yeni antibiyotik kullanım politikaları geliştirilmesi açısından
önemlidir.
Enterokok enfeksiyonları içinde E. faecalis ile oluşan enfeksiyonların oranı diğer türlere
göre 10 kat fazladır. Ancak son yıllarda vankomisin dirençli enterokoklar (VRE)’lerin
ortaya çıkması ile bu oran gittikçe düşmüş ve E. faecium izolatları ön plana çıkmaya
başlamıştır. Bu sebeple çalışmamızda hastane ortamında yaygın olarak bulunan ve
enfeksiyona neden olan E. faecium suşlarının çoklu antibiyotik direnç özellikleri
araştırılmış, bu özelliklerin yatay gen transferi yolu ile aktarımı analiz edilmiştir.

2
2. KURAMSAL TEMELLER
2.1 Enterococcus Cinsinin Genel Özellikleri
İlk defa 1899’da Fransa’da yayınlanan bir yazıda intestinal orjinli Gram- pozitif koklar
‘enterocoque’ olarak tanımlanmıştır (Thiercelin 1899). Streptococcus faecalis ismi
1906’da endokarditli bir hastanın kanından izole edilen organizmaya verilmiştir
(Andrews ve Horder 1906). S. faecium’un ilk olarak tanımlanması ise 1919 yılında
Orlo-Jensen tarafından yapılmıştır (Kandler vd. 1968). 1980’ lerde DNA-DNA ve
DNA-ribozomal RNA hibridizasyon çalışmaları bu bakterilerin Streptococcus
genusunun üyesi olmadığını göstermiş ve Enterococcus genusu içinde yeniden
sınıflandırılmışlardır (Schleifer ve Kilpper-Balz 1984). Enterococcus cinsi, DNA-DNA
hibridizasyon çalışmaları, 16S rRNA dizi analizi ve toplam hücre protein profillerinin
(WCP) belirlenmesi çalışmaları sonucunda 32 farklı türe ayrılmıştır (Facklam 2002).
E. faecalis insan enterokokal enfeksiyonlarının % 80-90’ ına yol açmaktadır. E. faecium
ise geriye kalan enfeksiyonlardan sorumludur. E. avium, E. casseliflavus, E. durans, E.
gallinorum, E. hirae, E. raffinosus, E. malodoratus, E. dispar ve E. mundtii diğer sık
rastlanan türlerdir (Murray 1990, Ruoff vd. 1990, Jett vd. 1994). Ayrıca son on yıl içinde E. haemoperoxidus, E. villorum, E. phoeniculicola, E. canis, E. moraviensis, E.
columbae ve E. cecorum gibi yeni türler de enterokok cinsi içinde tanımlanmıştır
(Koneman vd. 2005).
Enterokoklar tekli, ikili, ya da kısa zincirler halinde görülebilen, fakültatif anaerob,
Gram- pozitif koklardır. Mikroskopik olarak streptokok türlerinden ayrılamazlar.
Agarda üreyen kolonilerden yapılan Gram boyamada bazen Gram- pozitif kokobasil
şeklinde görülebilirler. İdeal üreme ısıları 35 οC olmakla birlikte 10-45 οC arasında
değişen bir üreme aralığına sahiplerdir. 60 οC’ye 30 dakika dayanıklıdırlar. Optimal
üreme pH’ ları 7.2 ±0.2 dir. Kanlı agarda 0,5-1,5 mm boyutunda, kabarık, gri - beyaz
renkte koloniler oluştururlar. Bazen zayıf bir alfa hemoliz meydana getirebilirlerse de

3
genellikle hemolitik özellik içermezler (Murray 1990, Koneman vd. 1992, Gültekin
2004).
Enterokokların tür düzeyinde identifikasyonu; tedavi yanında, epidemiyolojik ve
enfeksiyonel kontrol amacı ile de yapılmaktadır. Gram- negatif bakterileri de içeren
karışık örneklerden soyutlanmaları için, selektif besiyeri olarak, azid içeren safra-
eskulin-azid veya Enterococcosel agar, Columbia-Kolistin-Nalidiksik asit agar (CNA)
veya feniletil alkol agar (PEA) kullanılabilir. CNA, PEA’ ya göre bakterinin hemoliz
özelliğini değerlendirmede daha değerlidir. Kontamine alanlardan özellikle E. faecium
izolasyonu için de sefaleksin-aztreonam-arabinoz agar kullanılabilir. Vankomisin
dirençli enterokokların büyük bir kısmı E. faecium suşlarıdır. Vankomisin dirençli
enterokok saptanması için 6 µg/mL vankomisin ilave edilmiş brain-heart infüzyon
(BHI) agar veya Enterococcosel sıvı besiyeri kullanılabilir (Facklam ve Sahm 1995).
Enterokoklar bakteriyosin üretirler. Bunların bazıları diğer Gram- pozitif
mikroorganizmalar üzerinde etkilidir (Moreno vd. 2006).
Enterokoklar; sitokrom enzimleri olmadığından katalaz negatiftirler. Fakat E. faecalis
suşları ilk izolasyon sırasında görülüp seri pasajlarda kaybolan “pseudo-katalaz” üretir
ve katalaz testinde zayıf bir pozitiflik gösterebilir. Ayrıca E. haemoperoxidus türlerinde
de katalaz pozitifliği tanımlanmıştır (Ruoff vd. 1990).
Tüm suşlar % 6,5’luk NaCl içeren buyyonda ve % 40 safra varlığında ürerler. L-
pironil-beta-naftilamit (PYR) ve eskulini hidrolize ederler. Enterokoklar, E. cecorum, E.
columbae ve E. saccharolyticus dışında PYR (+) dir (Facklam and Sahm 1995).
Glukozu gaz yapmadan fermente ederler ve laktik asit oluştururlar. DNA’larının G+C
oranı % 37-45 oranındadır. E. flavescens, E. casseliflavus, ve E. gallinarum gibi bazı
suşlar hareketli olup 1-5 flagella içerirler. Endospor ve kapsül gibi yapıları
sentezleyemezler. Bütün enterokok suşları lösin aminopeptidaz (LAP) üretirler (Murray
1990, Ruoff vd. 1990, Koneman vd. 1992). Enterokokların hücre duvar yapısı diğer
Gram- pozitif koklara benzer peptidoglikan, teikoik asit, lipoproteinler ve yüzey protein
antijenlerinden oluşur. E. faecium Lys-Ala 2-3 tipinde bir peptidoglikana sahiptir
(Murray 1990, Korten 2002). Lancefield’ın grup D antijeni, hücre duvarı ile bağlantılı

4
bir gliserol olan teikoik asitten oluşur. Enterokoklar D grubu antiserumlarla % 80
oranında aglütinasyon verirler. Bu sonucu, ekstraksiyon işlemi ve antiserum kalitesi
etkileyebilir. Grup D antijeni tespiti, enterokoklar dışında, S. bovis, S. equinus, S. suis,
Pediococcus spp. ve Leuconostoc spp. gibi diğer Gram- pozitif bakterilerde de
bulunabildiğinden, özellikle doğal olarak glikopeptit direnci bulunan Leuconostoc spp.
ve Pediococcus spp. den ayırımı için PYR hidrolizinden yararlanılır (Şekil 2.1).
Şekil 2.1 Enterokokların diğer Gram- pozitif ve katalaz negatif koklardan ayrımı (Klein 2003, İşleroğlu vd. 2008)

5
Enterokokların karbon metabolizması, solunum, iyon transportu, pirimidin ve folat
yolakları, stres cevapları ve reaktif oksijen türleri metabolizmaları dikkat çekicidir.
Enterokokların bazal metabolik faaliyetleri için, B1, B6 ve B12 vitaminlerine, nükleik
asit bazlarına ve bir karbon kaynağına ihtiyaçları vardır. E. faecalis’in çoğalması için
histidin, izolösin, metionin ve triptofan gerekli iken, diğer bazı türler arjinin, glutamat,
glisin, lösin ve valin’e ihtiyaç duyarlar (Murray vd. 1993, Facklam ve Teixeria 1998).
Ancak bazı türler de bu aminoasitlere ihtiyaç duyulmaz. Vankomisin bağımlı suşlarda
olduğu gibi yoğun antimikrobiyal baskı, metabolik ihtiyaçları etkileyebilir. Bu da farklı
enterokok türlerinin metabolik ihtiyaçlarının suştan suşa bile farklılık gösterdiği
anlamına gelmektedir (Facklam ve Teixeria 1998).
Enterokokların hızlı tanı sistemleri ile teşhis edilmesini sağlayan otomatize (Phonex,
Vitek, MicroScan, API 20 Strep) sistemler mevcuttur. Bu sistemler içerdikleri Gram-
pozitif teşhis kartları ile tür düzeyinde teşhis yapabilirler. Ayrıca tiplendirmede;
bakteriyosin tiplendirmesi, faj tiplendirmesi, biyokimyasal reaksiyon profilleri,
antimikrobiyal direnç paternleri ve serolojik yöntemler de kullanılabilir (Facklam ve
Sahm 1995). Enterokok türlerini teşhis etmede en kolay yol anahtar fenotipik
karakterlere göre tür seviyesinde gruplar oluşturmaktır. Bu şekilde çeşitli test
ortamlarındaki reaksiyonlarına göre 5 grupta toplanmış, ilave testler ve karakteristik
özelliklerine göre de tür ayırımları yapılmıştır (Şekil 2.2). Biyokimyasal, fizyolojik,
serolojik ve antibiyotik direnç paternine dayanarak ayrım yapılamadığı durumlarda
moleküler tekniklere yönelinmektedir. Moleküler olarak enterokokların tiplendirilmesi
çalışmalarında çok sayıda teknikten yararlanılmaktadır. Bu tekniklere; dalgalı alan jel
elektroforezi (PFGE), multilokus sekans tiplendirme (MLST), rastgele çoğaltılmış
polimorfik DNA (RAPD) analizi, gen sekans analizi, çoğaltılmış fragment uzunluk
polimorfizmi (AFLP) ve gerçek zamanlı polimeraz zincir reaksiyonu (RT-PZR) örnek
olarak verilebilir (Nourse vd. 1998, Mascini vd. 2006, Çelik ve Alhan 2008).

6
Şekil 2.2 Enterokok türlerinin fenotipik özellikleri (Facklam ve Teixeria 1998)

7
2.2 Enterokok Enfeksiyonlarının Epidemiyolojisi
Enterokoklar doğada yaygın olarak (toprakta, suda, bitkilerde, hayvanlarda, kuşlarda ve
böceklerde) bulunurlar. Esas konakları insan ve hayvanların gastrointestinal sistem
florasıdır.
Enterokoklar son yıllarda tüm dünyada giderek artan sıklıkta yol açtıkları hastane
enfeksiyonu salgınları ile dikkatleri üzerlerine çekmişlerdir. İlk yapılan epidemiyolojik
çalışmalar, enterokokların hastadan hastaya ve hatta hastaneler arasında
yayılabilmesinin temelinde, bu bakterilerin normal bağırsak florasında bulunmalarının
yattığı tespit edilmiştir. Yapılan çalışmalarda E. faecalis’in hastane enfeksiyonlarının %
85-95’ inde, E. faecium’un ise % 5-10’unda başlıca etken olduğu saptanmıştır
(Çetinkaya vd. 2000, Zırakzadeh ve Patel 2006). Ancak vankomisin dirençli
enterokokların sebep oldukları enfeksiyonların kliniğine ait ilk bilgilerin açıklanması ve
direnç özelliklerinin tespit edilmesinden sonra enterokoklara bağlı hastane
enfeksiyonları epidemiyolojisinde iki önemli değişiklik yaşanmıştır (Uttley vd. 1988,
Leclercq vd. 1988). Bunlar insidansdaki hızlı artış ve tür dağılımındaki değişimdir.
Özellikle vankomisinin hastanelerde yaygın olarak kullanılmaya başlandığı 1989
yılından itibaren, Avrupa ülkeleri ve ABD’den başta glikopeptit grubu antibiyotikler
olmak üzere beta-laktam ve aminoglikozit grubu antibiyotiklere karşı dirençli çok
sayıda klinik izolat ve vaka bildirimi yapılmış, VRE’lerin özellikle E. faecium’un klonal
seleksiyona uğrayarak hastane ortamına hakim olduğu, ya tek başlarına ya da
stafilokoklarla birlikte hastane enfeksiyonlarındaki insidanslarını hızla artırdıkları tespit
edilmiştir (Çetinkaya vd. 2000).
Avrupa Antibiyotik Direnci Sürveyans Sistemi (EARSS)’nin 2010-2011 verilerine göre
E. faecium izolatları arasında en yüksek vankomisin direnç oranı; İrlanda % 38.7-34.9
(2010-2011), ikinci sırada ise Yunanistan olarak bildirilmiştir (% 22.5-2010 ve % 23.1-
2011) (Şekil 2.3-2.4).

8
Şekil 2.3 Avrupa’da VRE prevalansı (www.earss.rivm.nl- 2010)
Şekil 2.4 Avrupa’da VRE prevalansı (www.earss.rivm.nl- 2011)

9
2000’li yıllarda Uluslararası Hastane Enfeksiyonu Sürveyans Sistemi (NNIS)
sonuçlarına göre Amerika Birleşik Devletleri (ABD)’nde enfeksiyon etkeni olarak izole
edilen enterokoklar içerisinde VRE insidansının % 25’in üzerine çıktığı, hastanede
yatan hastalarda VRE kolonizasyon oranının da % 5-18 arasında değiştiği, ancak
hastane dışındaki popülasyonda intestinal kolonizasyon olmadığı bildirilmiştir.
Ülkemizde ise ilk vankomisin direçli enterokok (VRE) suşu Akdeniz Üniversitesi’nden
bildirilmiştir. Bu suş malign histiyositozis tanısı almış, bronkopulmoner enfeksiyonu
olan bir bebekte plevra sıvısından izole edilmiştir (Vural vd. 1998). Bunu 1999 yılında
İstanbul Üniversitesi, İstanbul Tıp Fakültesi ve Ankara Gülhane Askeri Tıp Akademisi’
nden bildirilen suşlar izlemiştir (Başustaoğlu ve Özyurt 2000). 2003 yılı itibariyle VRE
sorunu ile karşılaşan merkez sayısı onu aşmıştır (Yetkin ve Ateş 2004).
2.3 Enterokok Enfeksiyonları
Hastanede yatan hastalarda, endojen kökenli veya ekzojen kökenli hastane
enfeksiyonları ortaya çıkar. İnfekte hastaların varlığıyla geniş bir alanı enfekte eden
enterokokların, duyarlı hastalarda idrar yolları, kan, bos gibi steril vücut bölgelerine
yerleşerek, üriner sistem enfeksiyonları, endokardit, sepsis veya bakteriyemi, intra-
abdominal enfeksiyonlar, pelvik enfeksiyonlar, menenjit ve cerrahi yara
enfeksiyonlarına yol açabildikleri gösterilmiştir (Yıldırım 2007).
Klinik örneklerden izole edilen suşların % 80-90’ını E. faecalis, % 5-10 kadarını ise E.
faecium oluşturur. Son yıllarda çoklu ilaç direnci gösteren E. faecium suşlarının hastane
enfeksiyonlarında artış gösterdiği belirlenmiştir. Ayrıca nadir olarak E. durans, E.
casseliflavus, E. raffinosus, E. gallinorum, E. mundtii, E. flavescens, E. avium ve E.
hirae gibi diğer enterokok türleri hastane enfeksiyonlarda klinik materyalden izole
edilirken E. malodoratus, E. pseudoavium ve E. sulfureus türleri ile PYR testi negatif
olan ve atipik enterokoklar olarak adlandırılan E. columbae, E. cecorum ve E.
saccharolyticus türleri henüz insanlardan izole edilmemiştir (Sood vd. 2008)

10
2.3.1 Üriner sistem enfeksiyonları
Enterokokların sebep olduğu enfeksiyonların en sık görülen tipi üriner sistem
enfeksiyonlarıdır ve çoğu nozokomiyal kaynaklıdır. ABD’de enterokoklar nozokomiyal
üriner sistem enfeksiyonları arasında ikinci en sık etkendir. Üriner sistemde hastane
kökenli VRE enfeksiyonları; sistit, piyelonefrit, prostatit ve perinefritik apse gibi klinik
tablolarla seyredebilir. Üriner kateterizasyon en sık görülen risk faktörü olmakla
beraber, üriner sistem anomalileri, uzun süreli sonda kullanımı, cerrahi girişim ve
antibiyotik kullanımıda enterokokal risk faktörleri arasında yer alır (Kaye 1982, Krieger
vd. 1983). Toplum kaynaklı üriner sistem enfeksiyonlarında, sağlıklı kadınlarda % 5’
inden azında enterokok izole edilmektedir (Yıldırım 2007). Enterokokal üriner
enfeksiyonlarda bakteriyemi gelişmediği sürece mortalite düşüktür (Linden vd. 1996).
2.3.2 Endokardit
Enterokoklar infektif endokarditlerin S. viridans ve S. aureus’dan sonra en sık
rastlanılan üçüncü önemli etkenidir. En sık görülen tür E. faecalis’ dir. Hastalık cerrahi
yolla veya çeşitli maniplasyonlarla bakterinin gastrointestinal sistemden veya
çoğunlukla genitoüriner sistemden translokasyonu ile endojen kaynaklı subakut
endokardit şeklinde başlar (Anderson vd. 2004). Ayrıca intravenöz ilaç bağımlılarında
da % 5-53 oranında enterokokal endokardit görülebilir. Vakaların çoğunda altta yatan
bir kapak hastalığı veya prostetik kapak bulunmakla beraber, enterokoklar normal
kapaklarda da enfeksiyon oluşturabilir (Yıldırım 2007). Çoğunlukla sol taraf
endokarditine neden olur ve mitral kapak, aort kapağına göre daha sık tutulmaktadır.
2.3.3 Bakteriyemi
Enterokokkal bakteriyemi, endokarditlerden daha sık görülen bir klinik formdur.
Hastane enfeksiyonları içerisinde giderek artan öneme sahip olan VRE bakteriyemileri,
hastane kökenli tüm bakteriyemiler içerisinde Avrupa’da 4. ABD’de ise 3. sıklıkta
görülmektedir. VRE bakteriyemisi için risk faktörleri, hemodiyaliz, organ

11
transplantasyonu, kortikosteroit kullanımı, kemoterapi veya parenteral beslenme, cerrahi
girişimler, ciddi hastalıklar, uzun süreli antibiyotik kullanımı, üriner kateterler,
nötropeni ve mukozittir (Devriese vd. 2006, Yıldırım 2007).
2.3.4 İntraabdominal ve pelvik enfeksiyonlar
İntraabdominal ve pelvik enfeksiyonlar genellikle polimikrobiyaldir. Enterokoklar
bağırsakta normalde bulunan diğer fakültatif ve anaerob bakterilerle birlikte etken
olarak saptanmaktadır ve bu enfeksiyonlarda enterokokların esas rolünün ne olduğu
açıklık kazanmamıştır. Enterokoklar, nefrotik sendrom veya sirozlu hastalarda gelişen
spontan peritonit ve periton diyalizi uygulanan vakalarda gelişen abdominal enfeksiyon
veya intraabdominal abselerde tek mikroorganizma olarak izole edilebilmektir. Ayrıca
salpenjit, endometrit gibi peripartum maternal enfeksiyonlar ve sezeryan sonrası
apselerde de etken olabildikleri gösterilmiştir (Yıldırım 2007).
2.3.5 Yara ve yumuşak doku enfeksiyonları
Enterokoklar nadiren selülit veya derin doku enfeksiyonlarına yol açarlar. Cerrahi yara
enfeksiyonları, dekübitis ülserleri ve diyabetik ayak enfeksiyonlarında çoğunlukla
Gram- negatifler ve anaeroblarla birlikte izole edilirler. Enterokoklar nadiren diyabetik
olan veya olmayan kişilerde kronik osteomiyelite neden olmaktadırlar (Murray 1998).
2.3.6 Neonatal enfeksiyonlar
Enterokoklar yenidoğan için de önemli bir patojendir. Yenidoğanlarda erken ve geç
sepsise neden olur. Yenidoğan sepsisi tablosu menenjit ve/veya bakteriyemi ile birlikte
solunum güçlüğü ve ateşle karakterize bir tablodur. Prematüre veya düşük doğum
ağırlıklı bebeklerde, özellikle de nazogastrik tüple beslenme, intravasküler kateter
bulunması gibi durumlarda E. faecalis veya E. faecium’un etken olduğu sepsis salgınları
bildirilmiştir. Enterokokkal suşların % 82’ si E. faecalis, % 14’ ü E. faecium olarak

12
tespit edilmiştir. Uygun antibiyotik tedavisine iyi cevap vermektedir (Taşova ve İnal
2004).
2.3.7 Menenjit
Enterokokal menenjit nadir olmakla beraber, genellikle santral sinir sisteminde
anatomik bir defekt, geçirilmiş beyin cerrahi girişimi veya kafa travması gibi
predispozan faktörlerin varlığında görülmektedir (Murray 1990, Başustaoğlu 2004).
Menenjit, enterokok endokarditli hastalarda gelişen bakteriyeminin nadir bir
komplikasyonu olarak da oluşabilir. AIDS ve akut lösemi gibi ciddi immün yetmezlikli
hastalarda da VRE menenjiti görülebilir. Etken diğer enterokok enfeksiyonlarında
olduğu gibi sıklıkla E. faecalis olup daha seyrek olmak üzere E. faecium’dur (Zeana vd.
2001, Yıldırım 2007).
2.3.8 Solunum yolu enfeksiyonları
Geniş spektrumlu antimikrobiyal tedavi (özellikle sefalosporinler) ve enteral beslenme
uygulanan hastalarda da nadiren enterokok pnömonisi gelişebilmektedir. Enterokokların
neden olduğu solunum yolu enfeksiyonu sıklığının giderek arttığı bildirilmektedir
(Robert ve Moellering 2005).
2.4 Enterokoklarda Bulunan Virülans Faktörler ve Patojenite
Enterokoklar, düşük virülanslı bakterilerdir. Buna rağmen toplum kaynaklı ve özellikle
hastane kaynaklı enfeksiyonlarda önemli etkenlerdir. Pek çok antibiyotiğe karşı doğal
olarak dirençli olmaları, diğer antibiyotiklere de kolaylıkla direnç geliştirebilmeleri ve
çevreye adaptasyonlarının iyi olması nedeni ile diğer patojenlerden daha avantajlı hale
gelmektedirler.
Enterokoklara özgü virülans faktörlerin büyük bir çoğunluğu plazmidler üzerinde
kodlanır. Gastrointestinal sistem gibi doğal çevredeki suşlar arasında horizontal (yatay)

13
yayılım gösteren bu özellikler, daha az virülanslıktaki endojenik floraya hızla yayılmayı
sağlar (Jett vd. 1994).
Virülans faktörler çoğunlukla multifaktöriyeldir ve Patojenite Adaları (PAI) olarak
adlandırılan ve genom üzerinde farklı bölgelerde kümelenmiş olan virülans genler ile
düzenlenmektedir. PAI’ları virülans genlerin yatay transferinden sorumlu olmaları
nedeniyle patojen bakterilerin evriminde plazmidler, bakteriyofajlar ve transpozonlar
kadar öneme sahiptir (Hacker ve Kaper 2000, Tendolkar vd. 2003).
Enterokokların patojenite adaları ilk kez 1980 yılında nozokomiyal salgına yol açan E.
faecalis’te tanımlanmıştır (Huycke vd. 1991, Shankar vd. 2002). Bu PAI’ın
bulunmasından iki yıl sonra E. faecium suşlarında E. faecalis’dekinden farklı olan bir
PAI varlığı bulunmuştur. E. faecium klinik izolatlarında epidemik yayılma özelliği
kazandıran “esp” geninin görülmesi bu türdeki patojenite adası için bir indikatör olarak
kabul edilmiştir (Leavis vd. 2004). Bunun aynı zamanda nozokomiyal salgınlar ile
ilişkili izolatların ortak özelliği olduğu öngörülmektedir. Farklı VRE suşlarındaki PAI
dizilerinin karşılaştırılması sonucu; yüksek derecede dizi benzerlikleri olduğu ve
spesifik gen bölgelerinin değişen sıklıkta delesyona uğradıkları görülerek, bu
mikroorganizmaların virülans düzenleme yeteneği kazandıkları öne sürülmüştür. Diğer
taraftan bu çalışmalarda PAI’larının en az üçte birinin konjugatif bir plazmidin
kromozoma integrasyonu ile oluştuğu ispatlanmıştır. Ayrıca PAI’larda fonksiyonu izah
edilemeyen 18 ORF bölgesi kodlanmaktadır. Kommensal enterokokların PAI’larında bu
ORF bölgeleri gösterilememiş, bu sebeple de, bunların enterokokların hastane
ortamında yaşamasına veya hastalık geçişine veya patogeneze katkıda bulundukları
düşünülmüştür (Upadhyaya vd. 2009). Enterokoklarda tanımlanan virülans faktörler
çizelge 2.1’de verilmiştir.

14
Çizelge 2.1 Enterococcus cinsinde tanımlanan virülans faktörler (Mundy vd. 2000, Clewell vd. 2000, Eaton ve Gasson 2001, Shankar vd. 2001, Toledo-Arana vd. 2001)
Faktör/Gen Ürünlerin Virülanslıktaki Rolü efaAfm, efaAfs E. faecalis ve E. faecium serumlarıyla ifade olan hücre duvarı ilişkili
adhezinler agg Ökaryotik hücrelere tutunmada gerekli olan agregasyon proteini;
hücresel agregasyon ve konjugasyon
gelE Toksin; ekstraselüler metaloendopeptidaz, jelatin, kollojen ve
hemoglobini hidrolize eder
cylLL, cylLS Sitolizin (hemolizin, bakteriyosin) öncüleri; ökaryotik ve Gram-
pozitif hücrelerin büyük çoğunluğunu lize eden aktif sitolizin
üretiminde gerekli olan cylLL, -LS, -M, -B, ve -A genlerinin ifadesi
ürünleri
cylM Sitolizinin posttranslayonel modifikasyonu
cylB Sitolizinin transportu
cylA Sitolizinin aktivasyonu
esp İmmün cevaptan kaçmada rol oynayan hücre duvarı ilişkili protein;
ptojenite adası üzerinde bulunan cyl genleri ile ilişkili olabilir
cpd, cob, ccf,
cad
Seks feremonları; insan lökositleri için kemotaktiktirler ve
konjugasyonu kolaylaştırırlar
Lipotaykon asidi Üretilmiş insan monositleri yoluyla sitokin üretimin uyarılması
Süperoksit Bakteremi virülanslığı ile ilişkilidir; kültürdeki insan intestinal
epitelyum hücrelerinde hücre-DNA hasarının indüklenmesi
Hiyaluronidaz Bağlayıcı dokularda hücre yüzeyi ile ilişkili enzimlerin yayılmasını
sağlar

15
2.5 Enterokoklarda Antibiyotik Direnci ve Direnç Mekanizmaları
Enterokoklardaki antibiyotik direnci doğal ve kazanılmış olmak üzere iki gruba
ayrılabilir. Doğal direnç türe/cinse özgüdür. Enterokok türlerinin tamamında görülen
kromozomal direnci ifade eder. Doğal direnç kromozomlar üzerindeki genler aracılığı
ile oluşurken, kazanılmış direnç plazmidler veya transpozonlar üzerinde bulunan
genlerle gerçekleştirilmektedir (Sarantinopoulos vd. 2001, İşleroğlu vd. 2008).
Enterokoklarda antibiyotik direnci detaylı olarak araştırılmıştır (Fisher ve Philips 2009).
2.5.1 Doğal (kromozomal) direnç
Enterokok türleri penisilinlere, sefalosporinlere, linkozamidlere, trimetoprim-
sulfametaksazole (TMP-SMX), aminoglikozitlere (düşük düzeyde), polimiksinlere,
vankomisine (E. gallinarum, E. casseliflavus ve E. flavescens gibi bazı türler düşük
seviyede), monobaktamlara ve kinopristin/dalfopristin’e (E. faecalis) karşı kalıtsal
olarak dirençlidir (Poeta vd. 2005).
Enterokoklar β-laktam antibiyotiklere karşı karakteristik olarak tolerans gösterirler yani
tedavi dozunda minimum bakterisidal konsantrasyon/minimum inhibitör konsantrasyon
(MBK/MİK) oranı 1/32’nin üzerindedir. Bu nedenle β-laktam antibiyotikler
enterokoklara karşı bakterisidal değil, bakteriyostatik etkilidir. Enterokoklarda doğal
penisilin direnci, β-laktam antibiyotiklere düşük bağlanma afinitesi gösteren penisilin
bağlayan protein 5 (PBP-5) enziminin varlığına bağlıdır. E. faecalis suşlarında penisilin
MİK değeri streptokoklardan 10-100 kat daha yüksektir. Enterokoklarda yarı sentetik ve
penisilinaza dirençli β-laktam antibiyotiklere karşı da doğal direnç oldukça yüksek
oranda bulunmuştur. Ampisilin direnci E. faecium suşlarının % 85–90’ında, E. faecalis
suşlarının ise % 2-3’ünde görülmektedir (Murray 1998, Çetinkaya vd. 2000, Marothi
vd. 2004).
Enterokoklar düşük düzeyde aminoglikozit direnci de gösterirler. Bu tip dirençte iki
mekanizma söz konusudur. Birinci mekanizma tüm enterokok türlerinde bulunur ve

16
bakteri duvarının bu grupta bulunan antibakteriyel ilaçlara karşı geçirgenliğinin az
olmasından kaynaklanır. İkinci mekanizma sadece E. faecium’da bulunur. E. faecium
aac6’-li geni tarafından kodlanan, 6’ asetiltransferaz (AAC-6’) enzimine sahiptir. Bu
enzim aminoglikozit yapısındaki bir amino grubunun asetil CoA’ya bağımlı olarak
asetilasyonuna yol açar. Böylece sitoplazmaya geçen ilaç inaktive edilir. Enzim
kanamisin, netilmisin, sisomisin, isepamisin ve tobramisini modifiye eder. Ancak
gentamisine etkisi yoktur (Şardan 2004). Aminoglikozit grubu antibakteriyel ilaçlar,
beta-laktam antibiyotik ya da vankomisin gibi hücre duvarı sentezini engelleyen
antibiyotikler ile kombine edilecek olursa, zedelenen hücre duvarından bu gruptaki
antibakteriyeller daha kolay geçeceğinden MİK değerleri önemli ölçüde düşecektir.
Enterokoklara karşı, beta-laktam veya glikopeptit grubu antibakteriyel ilaçlar ile
aminoglikozit grub ilaçların kombinasyonunun sinerjistik mekanizması bu şekilde
açıklanmaktadır (Başustaoğlu 2004).
Enterokoklar linkozomit grubu antibiyotiklere de düşük düzeyde doğal olarak
dirençlidir (Lefort vd. 2000, Şardan 2004).
Enterokokların ekzojen folat kullanma yetenekleri bulunmaktadır. Bu nedenle
trimetoprim-sulfametaksazole (TPM-SMX) de doğal olarak dirençlidirler. Bu
antibiyotik in vitro olarak etkin görünmesine rağmen in vivo etkin değildir. Bu nedenle
antibiyotik duyarlılık deneylerinde TPM-SMX kullanılmamalıdır (Lefort vd. 2000,
Korten 2005).
2.5.2 Kazanılmış direnç
Enterokoklar plazmid ve transpozonlar yoluyla son yıllarda belirgin bir şekilde
kazanılmış direnç geliştirmiştir. Bunlar arasında en önemli olanları yüksek düzey
aminoglikozit direnci, beta laktamaz yapımı veya diğer mekanizmalarla gelişen yüksek
düzey penisilin direncidir. Enterokokların çok önemli bir kısmı bu yol ile eritromisin,
klindamisin ve tetrasiklinlere direnç kazanmış durumdadır (Korten 2002).

17
2.5.2.1 Beta laktam direnci
Enterokokların iki ayrı direnç mekanizması ile beta-laktam antibiyotiklere direnç
kazandığı saptanmıştır. Bunlardan biri E. faecium suşlarında görülen, kromozomal olan
ve penisilin afinitesinin azalması sonucu PBP 5’in miktarının artması ile ortaya çıkan
dirençtir. Bu mekanizmada esas, üremelerini inhibe edecek konsantrasyondaki β-laktam
antibiyotiklerinin varlığında, onlara düşük bağlanma afinitesi gösteren düşük molekül
ağırlıklı PBP5 sentezleyebilmeleridir. Bu sentez PBP değişikliklerini kodlayan DNA
bölgesinde meydana gelen mutasyon sonucu veya plazmid kazanımı ile
gerçekleşmektedir (Derbentli 1998).
İkinci direnç mekanizması ise beta-laktamaz üretimidir. Bu enzimler antibiyotiğin hedef
bölgeye ulaşamadan modifiye edilmesine veya parçalanmasına yol açarlar. Beta
laktamaz oluşturan suş ilk olarak 1981 yılında ABD’de tanımlanmıştır (Derbentli 1998,
Lefort vd. 2000). Ülkemizde yapılan çeşitli çalışmalarda beta-laktamaz üreten
enterokok saptanmamıştır (Moaddab ve Töreci 2000). Enterokoklarda β-laktamaz
enzimini kodlayan gen, transfer edilebilen bir plazmid üzerindedir. Bazı E. faecalis
suşlarında, bu genin kromozomlarda yerleştiği gösterilmiştir (Moellering 1992, Çınar
vd. 1999). Beta laktamaz üreten enterokok suşlarının büyük çoğunluğunda, yüksek
düzeyde gentamisin direnci de tespit edilmiştir (Markowitz vd. 1991).
E. faecium’ un klinik izolatları penisilinlere E. faecalis’den daha dirençlidir (Wax vd.
2008). PBP5 ilişkili düşük seviyeli ampisilin direnci enterokoklarda kromozomal
kökenlidir. Yüksek seviyede (MİK > 100 µg/mL) ampisilin direnci E. faecium’da düşük
afiniteli PBP üretimi, E. faecalis’te ise β-laktamaz üretimi ile oluşmaktadır
(Handwerger vd. 1992, Lavery vd. 1997). Vankomisine dirençli E. faecium penisilin ve
ampisiline daha dirençlidir.
β-laktam antibiyotikler üriner sistem infeksiyonları gibi bazı infeksiyonlarda tek başına
kullanılabilseler de menenjit, endokardit gibi bakterisidal aktivite gerektiren vakalarda
standart kombinasyon tedavisi kullanmak gereklidir. Vankomisin ve teikoplanin gibi
hücre duvarına etkili antibiyotiklerde beta-laktamaz üreten suşların etken olduğu

18
infeksiyonların tedavisinde yer alabilir (Kutlu ve Dokuzoğuz 2004). β-laktam
antibiyotiklerin aminoglikozitlerle kombinasyonu, sinerjistik bakterisidal etki
göstermektedir (penisline duyarlı ve yüksek düzey aminoglikozit direnci yok ise)
(Robert ve Moellering 2000).
2.5.2.2 Glikopeptit direnci
Enterokoklarda glikopeptit direnci, Van A’dan Van G’ye kadar çeşitlilik gösterebilen
farklı direnç fenotipleri ile ifade edilir. Fenotipik sınıflandırmada; bakterinin sadece
vankomisin ya da hem vankomisin hem de teikoplanine dirençli olması, direncin
indüklenebilir veya yapısal olması ve diğer bakterilere geçirilebilir olup olmamasına
göre yapılır. Bu glikopeptit direnç tipleri içerisinde en iyi tanımlanmış olanları Van A,
Van B, Van C ve Van D dirençleridir (Çizelge 2.2).

19
Çizelge 2.2 Enterokoklarda glikopeptit direnç tipleri (Murray 2000, Guardabassi ve Dalsgaard 2004)
Özellik VanA VanB VanC VanD VanE
MİK( µg/mL) Vankomisin Teikoplanin
64 - >1000 16 - > 512
4 - >1000 0.5 - > 32
2 - 32 0.5 - 1
16 - 64 2 - 4
16 0.5
Transfer edilebilme
+ + - - ?
Direnç genlerinin lokasyonu
Plazmidler Kromozom (plazmidler)
Kromozom Kromozom Kromozom
İndüklenme ile ekspresyon Vankomisin Teikoplanin
+ +
+ -
- -
+ -
+ -
Hareketli element
Tn 1546
Tn 1547
-
?
?
Ligaz geni van A van B van C-1 ve
van C-2/van
C-3
van D van E
Direnç tipi Kazanılmış Direnç
Kazanılmış Direnç
Doğal Direnç
Kazanılmış Direnç
Kazanılmış Direnç
Direnç proteinin M.A’lığı(kDa)
39-40 39.5 38 ? ?
Modifiye edilmiş hedef
D-ala-D-lac
D-ala-D-lac
D-ala-D-ser
D-ala-D-lac
D-ala-D-ser
Türler
E. faecalis
E. mundtii
E. faecium
E. raffinossus
E. avium
E. gallinarum
E. durans
E.casseliflavus
E. faecalis
E. faecium
E. gallinarum
E.casseliflavus
E. flavescens
E. faecium
E. faecalis

20
Ortamda vankomisin gibi bir indükleyici bulunduğunda, sensör kinaz enziminin bir
regülatör yanıt proteini ile ilişkiye girmesi sonucu, vankomisin direncinden sorumlu
genlerin transkripsiyonu uyarılır. Transkripsiyona uğrayan genler, translasyona
uğradığında; vankomisinin çok düşük bir ilgi ile bağlanabildiği ve D-ala-D-laktat veya
D-ala-D-serin ile sonlanan hücre duvarı öncülerinin oluşumunu sağlayan ligaz
enzimlerine dönüşürler. Diğer gen ürünleri ise D-ala-D-ala dipeptitlerini keserek hücre
duvarının esas yapısında bulunan glikopeptitlere duyarlı hedefleri ortadan kaldırırlar
(Şekil 2.5). VRE’ lerde peptidoglikan öncülüğündeki bu değişiklikler, glikopeptit
ajanların hedefine 1000 kat daha düşük bir ilgi ile bağlanmasına neden olur ve hücre
duvarı sentezinin inhibisyonu, bu temel mekanizma ile engellenir (Murray 2000,
Guardabassi ve Dalsgaard 2004).
Şekil 2.5 Vankomisin antibiyotiğinin ve vankomisine dirençli enterokoklarda Van A direncinin hücre duvar öncülerine etki mekanizması (V= vankomisin) (Patel 1999)

21
2.5.2.2.1 Van A tipi direnç
Van A tipi direnç en sık karşılaşılan dirençtir. Dirençten 39-40 kDa ağırlığında, Van A
adlı bir membran proteini sorumludur. Van A izolatları hem vankomisine (MIC ≥ 64
µg/mL) hem de teikoplanine (MIC ≥ 16 µg/mL) yüksek düzeyde dirençlidir. Dirence
neden olan membran proteini ancak vankomisin varlığında sentezlenebilir. vanA geni ve
direnç ekspresyonu ile regülasyondan sorumlu vanS, vanR, vanH, vanX ve vanZ gibi
diğer genler, E. faecium’da sıklıkla bir plazmid üzerinde bulunan 10581 bp’lik bir
transpozon (Tn 1546) da yerleşmektedir (Şekil 2.6). Bu genlerin ekspresyonu sonucu,
peptidoglikan terminalinde D-Ala-D-Ala yerine D-Ala-D-Laktat sentezlenmekte ve
vankomisin bu bileşiğe daha düşük ilgi ile bağlanmaktadır (French 1998, Çetinkaya vd.
2000, Reynolds ve Courvalin 2005).
Şekil 2.6 Vankomisin direncinden sorumlu Tn1546 transpozonundaki van A gen
kümesinin genetik haritası (Arthur ve Courvalin 1993) (Açık oklar sekansların kodlanma yönünü; transpozonun başındaki ve sonundaki IRL ve IRR sırasıyla sol ve sağ inverted tekrarlarını; B, BamHI; Bg, BglII; EI, EcoRI; EV, EcoRV; H, HindIII; P, PstI; X, XbaI restriksiyon enzimi kesim bölgelerini göstermektedir.)
Van R ve Van S proteinleri, vanHAX gen kümesinin transkripsiyonunu düzenleyen iki
elemanlı bir regülatör sistem oluşturur. vanS, sensör gendir ve vankomisin varlığını
algılamada veya tanımada rol alan Van S proteinini kodlar. vanR, regulatör gendir ve
diğer direnç genlerinin transkripsiyonel aktivasyonunda görev alan Van R proteini
kodlar. Van S, Van R’ye sinyal verir ve dirençte yer alan Van H, Van A ve Van X gibi

22
diğer bazı proteinlerin sentezi veya aktivasyonu ile sonuçlanan regülatör cevabı
oluşturur. Van A tipi suşlarda hem vankomisin hem de teikoplanin transkripsiyonu
indükleyebilir, ancak kesin sinyaller hala bilinmemektedir. Van A, Van H ve Van X
proteinleri kor proteinleridir
vanA, D-Ala-D-Laktat oluşumunu sağlayan bir ligaz proteini kodlar. vanA tek başına
vankomisin direncini sağlayamaz, çünkü D-Laktat gibi D-hidroksi asitler,
enterokoklarda genellikle sentezlenmez ve enterokokların çevresinde doğal olarak
bulunmazlar. Bu yüzden D-Laktat sentezlemek için enterokoklar, Van A için substrat
üretiminde gerekli olan vanA operonu içindeki genleri kazanmalıdır. vanH, piruvattan
D-Laktat sentezini sağlayan bir dehidrogenaz enzimini kodlar ve D-Laktat havuzunun
oluşumunu sağlar. vanX, bir DD-dipeptidaz enzimi olan Van X proteinini kodlar. D-
Ala- D-Ala havuzunu doğal bir enterokokal ligaz ile azaltarak, yarışma sureti ile normal
pentapeptit sentezini azaltır. D-Ala-D-Laktata karşı aktivitesi yoktur. vanY, DD
karboksipeptidaz olan Van Y proteinin kodlar ve peptidin sonundaki D-Ala’ yı ayırır.
Bu yolla orta düzeyde direnci sağlar. vanZ, ılımlı olarak teikoplanin MİK değerini
arttırır, ancak vankomisinin MİK değeri üzerinde etkili değildir. Bunun mekanizması
henüz aydınlatılmamıştır. vanY ve vanZ genleri dirence katkıda bulunabilir, ancak
direnç fenotipi için zorunlu değildir (Malathum ve Murray 1999, Çetinkaya vd. 2000).
vanA geni ilk olarak E. faecium’da, sonraları ise E. faecalis, E. durans, E. gallinorum,
E. avium, E. mundtii, E. casseliflavus, E. raffinosus gibi diğer enterokok türlerinde de
belirlenmiştir. Van A tipi direnç Avrupa’ da en çok görülen direnç tipidir. ABD’de Van
B suşları oldukça yaygın olmasına rağmen, Van A hala daha baskın direnç tipidir (Clark
vd. 1993).
2.5.2.2.2 Van B tipi direnç
Van B tipi direnç; Tn1547 transpozonunu üzerinde kodlu olan vanB geninin
sentezlediği 39.5 kDa moleküler ağırlıklı Van B membran proteini ile oluşturulur.
Vankomisin direnci orta düzeyde (MİK: 4-1000 µg/mL) olup, teikoplanine duyarlıdır
(MİK: 0.5-1 µg/mL). Modifiye edilmiş hedefi D-ala-D-laktat’dır. E. faecium ve E.

23
faecalis türlerinde görülür. Sadece Van B direncine özgü bir gen dışında diğer altı
genin, vanA kümesindeki genlerle % 77 homoloji gösterdiği saptanmıştır (Arthur ve
Courvalin 1993). Direnç genleri genellikle kromozom üzerinde yerleşmiştir ve
konjugasyon ile aktarılabilir ancak bazı VRE vakalarında plazmidler üzerinde
bulunabileceği de saptanmıştır (Handwerger vd. 1992, Lavery vd. 1997, Leclercq ve
Courvalin 1997, Hanrahan vd. 2000). Van B tipi dirençte vankomisin duyarlılığında
azalma, gentamisin sinerjisinde kaybolma, streptomisin sinerjisinde azalma olmuştur.
Teikoplanin tek başına Van B direnci taşıyan suşlara karşı etkili bulunmuş ama bu
tedavi teikoplanine dirençli suşların seleksiyonuna neden olmuştur. Teikoplanin
gentamisin kombinasyonunun ise dirençli mutant gelişimini engellediği belirlenmiştir.
Buna göre Van B tipi dirençli enterokokkal enfeksiyonlarda teikoplanin aminoglikozit
kombine kullanımı tercih edilmelidir (Çetinkaya vd. 2000, Rice 2001).
2.5.2.2.3 Van C tipi direnç
Van C tipi direnç, E. gallinarum, E. casseliflavus ve E. flavescens türlerinde görülen
doğal bir direnç türüdür. Van C gen kümesi D-ala-D-serin ile sonlanan pentapeptidin
oluşumunu sağlar. Bu direnç türünün üç alt tipi bulunmaktadır: Van C-1, Van C-2, Van
C-3. Bu genlerin türe spesifik olduğu düşünülmektedir (E. gallinorum– vanC-1, E.
casseliflavus– vanC-2 ve E. flavescens– vanC-3). Van A veya Van B ile % 37-39
oranında benzerlik gösterir. Van C tipi dirence sahip olan suşlar teikoplanine duyarlıdır.
Yapısal olarak indüklenemez ve transfer edilemezler (Çetinkaya vd. 2000).
2.5.2.2.4 Van D tipi direnç
Sadece E. faecium’da bildirilmiştir. VanD geni izolatları yapısal olarak hem
vankomisine (MIK 64-128 µg/mL) hem de teikoplanine (MIK 4-64 µg/mL) dirençlidir.
vanD geni kromozomaldir ve konjugasyon yolu ile transfer edilemez (Başustaoğlu
2004, Koneman vd. 2005). Modifiye edilmiş hedef D-ala-D-Laktat’dır. Glikopeptit
direnci gen grubu olan vanD; vanA ve vanB operonları ile benzer organizasyon gösterir
(Perichon vd. 2000).

24
2.5.2.3 Aminoglikozitlere direnç
Enterokoklarda aminoglikozit direnci üç farklı mekanizma ile meydana gelir.
2.5.2.3.1 Permeabiliteye bağlı direnç
Aminoglikozitlere karşı kromozomal mutasyon sonucunda membrandaki
permeabilitenin azalması ile oluşan direnç yüksek düzeyde olmamakla birlikte tüm
aminoglikozitlere karşı çapraz direnç şeklindedir. Bu tip direnç β-laktam antibiyotikler
ile birlikte kullanılarak ortadan kaldırılabilir (Başustaoğlu ve Aydoğan 2002).
2.5.2.3.2 Aminoglikozit modifiye edici enzimlere bağlı direnç
Aminoglikozitlerdeki en sık gözlenen yüksek düzeydeki (MIK ≥ 2000 µg/mL) (HLR)
edinsel direnç, plazmid veya transpozon kaynaklı asetiltransferaz (AAC),
adeniltransferaz (ANT), fosfatransferaz (APH) gibi modifiye edici enzimlerle
antibiyotiğin inaktive edilmesidir (Murray 1998). Yüksek düzey gentamisin direncine
neden olan enzim 6’- asetiltransferaz- 2” fosfo- transferaz enzim kompleksi olup
streptomisin hariç klinik kullanımda olan tüm aminoglikozitlere (gentamisin,
tobramisin, amikasin ve netilmisin) yüksek düzeyli direncin ortaya çıkmasında etkilidir.
Streptomisine, enzimatik yoldan kazanılan yüksek düzey direnç ise adeniltranferaz
(ANT-6’) enzimi ile olmaktadır. Bu enzim varlığında sadece streptomisine karşı yüksek
düzeyde direnç gelişmektedir (Leclercq vd. 1992). E. faecium kökenlerinde yüksek
düzey aminoglikozit direnci bulunmasa da penisilin ile sadece gentamisin ve
streptomisin sinerjik etkili olabilir. Çünkü E. faecium kökenleri doğal olarak tobramisin,
netilmisin, kanamisin ve sisomisini modifiye eden 6’ asetiltransferaz (AAC- 6’)
enzimini oluştururlar. AAC 6’ enzimi aac(6’)- li geni tarafından kodlanır. Bu durumda
yüksek düzey aminoglikozit direnci olmamakla beraber (MIK< 2000 µg/mL) hücre
duvarına etkili antibiyotikler ile sayılan bu 4 aminoglikozit arasındaki sinerji
bozulmaktadır (Simjee ve Gill 1997).

25
2.5.2.3.3 Ribozomal direnç
Bir ribozomal proteinde oluşan tek bir aminoasit değişikliği, o ribozomun antibiyotiğe
karşı düşük ilgi göstermesine neden olur. Yalnız streptomisine karşı gelişen ve transfer
edilemeyen bu tür direnç nadir görülmekte ve diğer aminoglikozitlere karşı çapraz
direnç oluşmamaktadır (Başustaoğlu ve Aydoğan 2002).
2.5.2.4 Kloramfenikole direnç
Yapılan çesitli çalısmalarda enterokokların % 20–42’sinin kloramfenikole dirençli
olduğu ve dirençten en sık sorumlu mekanizmanın kloramfenikol asetil transferaz
üretimi olduğu bildirilmektedir (Murray 1990). Kloramfenikolün aktivitesi,
kloramfenikol asetiltransferaz (CAT) enzimi tarafından inhibe edilir ve direnç gelişir.
Asetillenme sonucu kloramfenikol ribozomlara bağlanamaz ve protein sentezi
engellenemez. Enzimi kodlayan cat geni enterokoklarda konjugatif ve konjugatif
olmayan plazmidler ya da kromozom üzerinde bulunabilir. Streptokokal plazmid
pIP501 üzerinde bulunan kloramfenikol asetiltransferaz direnç geni ile akraba olan gen
dizisi, E. faecalis ve E. faecium izolatlarında bulunmuştur bu da genin yatay gen
transferi ile aktarılabildiğini göstermiştir (Aarestrup vd. 2000). Kloramfenikol
enterokoklara karşı bakteriyostatik etkilidir. Klinikte kloramfenikolün in vitro kullanımı
yaygın değildir çünkü çoklu ilaç direnci gösteren E. faecium’a karşı in vitro aktivitesini
korumaktadır. Ancak bazı VRE suşlarında kloramfenikole karşı da direnç gösterilmiştir
(Robert ve Moellering 2005).
2.5.2.5 Tetrasiklinlere direnç
Tetrasiklin direnci enterokoklarda konjugasyon yolu ile kazanılan direncin en tipik
örneğidir (Murray 1998). Enterokoklarda tetrasiklin grubu antibiyotiklere dirençten
sorumlu olan çok sayıda gen tanımlanmıştır. Bunlardan tetM geni Tn916 transpozonu
üzerinde taşınır ve en yaygın olarak bulunan direnç genidir (Aarestrup vd. 2000, Huys
vd. 2004). tetM, tetO ve tetS genleri tetrasiklinlerin ribozomlar üzerindeki etkisini

26
inhibe eder. tetL geni ise enterokokal bir plazmid üzerinde taşınır. Bu direnç geni
mikroorganizma tetrasiklinle karşılaştığında amplifiye olur. tetL ve tetK geni
tetrasiklinlerin hücre dışına pompalanmasını sağlayan aktif transport sistemini de kodlar
(Murray 1990).
2.5.2.6 Kinolonlara direnç
Direnç gyrA (DNA giraz alt ünitesi) ve parC (topoizomeraz) genlerindeki mutasyonlara
bağlı gelişir. Enterokokal suşların çoğunluğu kinolonlara orta seviyede duyarlılık veya
direnç gösterir (Klare vd. 2003). Kinolonlar üriner sistem infeksiyonlarında tek başına
kullanılabilir. Ancak üriner sistem infeksiyonu dışında başka bir infeksiyon varsa
kullanılmamalıdır. Yeni kinolonlar, özellikle klinafloksasin ve sitafloksasin VRE’lere
karşı, eski kinolonlara göre daha etkilidir. VRE ile oluşturulan deneysel endokarditlerde
klinafloksasin tek başına veya penisilinle birlikte etkili bulunmuştur (Kutlu ve
Dokuzoğuz 2004).
2.3.2.7 Makrolitlere direnç
Makrolidlere direnç genellikle ermB geni ile ilişkilidir. Eritromisin dirençli izolatların
çoğunda belirlenen ermB geni 23 rRNA’yı modifiye eden ve makrolidlere karşı
dirençliliği sağlayan erm metilazı kodlamaktadır (Del Campo vd. 2003). Makrolidler
yanında, linkozamidlere ve streptogramin B’ye karşı da direçlilik sağlayan bu gen söz
konusu antibiyotiklere dirençli enterokok izolatlarında çok yaygın bir şekilde
görülmektedir (Aarestrup vd. 2000, Shepard ve Gilmore 2002).
E. faecalis’te yürütülen analizler sonucu, bu bakteride tanımlanan ermB geninin S.
aureus transpozonu 554 üzerinde taşınan ermB geni ile tamamen aynı olduğu
saptanmıştır (Stovcik vd. 2008). Ayrıca enterokoklarda lakton halkasının hidrolizi veya
efluks mekanizması sonucu antibiyotiğin uzaklaştırılmasıyla da direnç oluşabilir (Klare
vd. 2003).

27
Rifampisin antibiyotikleri DNA-bağımlı RNA polimerazın β-altünitesine
bağlanmaktadır. Pek çok bakteride, rifampisin direnci RNA polimerazın β-altünitesini
kodlayan rpoB genindeki mutasyonlar ile gerçekleşmektedir (Enne vd. 2004). Özellikle
rpoB genleri bakterilerde yüksek düzeyde korunmuş merkezi metabolizmaya ait
genlerdir. Tek başına kullanımında direnç gelişeceğinden klinikte kullanımı önerilmez.

28
3. MATERYAL VE YÖNTEM
3.1 Materyal
3.1.1 Bakteriler
Çalışmada Akdeniz Üniversitesi Tıp Fakültesi Hastanesi’nde çeşitli klinik servislerde
yatmakta olan hastalardan alınan idrar, bos, kan, katater, pü ve rektal swab
örneklerinden elde edilen bakteri kültür koleksiyonundan sağlanan suşlar kullanılmıştır.
Yapılan araştırmada, daha önce Enterococcus faecium olduğu saptanmış 95 izolat
arasından çoklu ilaç dirençliliği içeren 64 tanesi incelenmiştir (Çizelge 3.1).
Stok kültürler -80 °C’de % 15 gliserol ilave edilen Trypton Soy Broth (TSB) sıvı besi
ortamında saklanmıştır. Çalışma süresince denenecek kültürler, TSB besi ortamında 37
°C’de 1 gece geliştirildikten sonra denemeye alınmıştır.
3.1.2 Besiyerleri
Trypton Soy Broth (Oxoid, UK) Kazein hidrolizat 17 g
Soya hidrolizat 3 g
Glukoz 5 g
NaCl 5 g
K2 HPO4 2.5 g
Destile su 1 lt
pH 7.3 ± 0.2 (sterilizasyondan önce)
30 Gram toz besiyeri 1 litre saf su ile eritilerek karıştırıldı ve 121 °C otoklavda 15
dakika sterilize edildi.

29
Müller-Hinton Broth (Oxoid, UK)
Et ekstraktı 30 g
Kazein hidrolizat 17.5 g
Nişasta 1.5 g
pH 7.3 ± 0.1 (sterilizasyondan önce)
38 Gram toz besiyeri 1 litre saf su ile karıştırıldı ve 121 °C otoklavda 15 dakika sterilize
edildi.
Trypticase Soy Agar (Oxoid, UK)
Kazein hidrolizat 15 g
Soya hidrolizat 5 g
NaCl 5 g
Agar 15.0 g
pH 7.3 ± 0.1 (sterilizasyondan önce)
40 Gram toz besiyeri 1 litre distile su ile karıştırıldı ve 121 °C otoklavda 15 dakika
sterilize edildi. 50 °C’ye kadar soğutuldu ve % 5 steril defibriniz koyun kanı ilave
edildi.
3.1.3 Bakterilerin tür teşhisi
Klinik izolatlardan enterokokların teşhisi ve MİK testleri için BD (Becton Drive, USA)
firmasına ait Phonex sistemi, Enterococcus faecium olarak tür teşhisini doğrulamak
amacıyla da Rapid ID 32 Strep Api (Biomerıeux, France) testi kullanılmıştır.

30
Çizelge 3.1 Çalışmada kullanılan bakteriler ve izolasyon materyalleri
Kod Bakteri Türü İzolasyon
materyali
Bakteri Türü Kod İzolasyon
materyali
1
2
3
4
5
6
8
11
12
13
15
17
19
21
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
41
42
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
Rektal
Kan
İdrar
Kan
İdrar
Kan
İdrar
Kan
Kan
Rektal
Rektal
Rektal
Rektal
Kan
İdrar
İdrar
İdrar
Pü
Kan
İdrar
İdrar
Katater
İdrar
İdrar
Bos
İdrar
Rektal
İdrar
Rektal
Rektal
Rektal
Rektal
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faeciums
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
E. faecium
43
46
48
49
50
52
55
56
57
59
60
63
64
65
67
68
69
70
71
74
75
76
77
79
80
81
84
87
89
90
94
95
İdrar
Kan
Pü
İdrar
İdrar
İdrar
Rektal
İdrar
İdrar
Rektal
İdrar
Rektal
Pü
Rektal
İdrar
Rektal
Kan
Rektal
Kan
İdrar
rektal
İdrar
Kan
İdrar
İdrar
Kan
Rektal
Rektal
Rektal
Rektal
İdrar
Rektal

31
3.1.4 Antibiyotikler
Antibiyotik duyarlılık deneylerinde kullanılan antibiyotikler SİGMA (Oxoid, UK)
firmasından satın alınmıştır. Araştırmada kullanılan antibiyotiklerin disk
konsantrasyonları çizelge 3.2’de verilmiştir. Antibiyotik duyarlılık deneylerinde
standart suş Enterococcus faecalis ATCC 29212 kullanılmıştır.
Çizelge 3.2 Çalışmada kullanılan antibiyotikler ve disk konsantrasyonları (Oxoid, UK)
Antimikrobiyal Madde Sembol Grubu Disk konsantrasyonu
(µg)
Ampisilin AMP β -laktam 10
Streptomisin STR Aminoglikozit 300
Kanamisin KAN Aminoglikozit 30
Gentamisin GEN Aminoglikozit 120
Kloramfenikol CHL Fenikol 30
Tetrasiklin TET Tetrasiklin 30
Rifampin RIF Ansamisin 5
Vankomisin VA Glikopeptit 30
Eritromisin
Teicoplanin
ERT
TEC
Makrolid
Glikopeptit
15
30

32
3.1.5 Primerler
Çizelge 3.3 Antibiyotik direnç genlerinin tespiti için kullanılan primerler
Gen
Primer dizisi
Bant
büyüklüğü
(bç)
Referans
van A
Vankomisin
5’ GGGAAAACGACAATTGC 3’
732
Dutka-Malen
vd. 1995 3’ GTACAATGCGGCCGTTA 5’
ant(6) - la
Streptomisin
5’ ACTGGCTTAATCAATTTGGG 3’
577
Kobayashi vd.
2001 3’ GCCTTTCCGCCACCTCACCG 5’
aac(6’)-aph(2’’)
Gentamisin
5’ CCAAGAGCAATAAGGGCATACC 3’
675
Kobayashi vd.
2001 3’ ACCCTCAAAAACTGTTGTTGC 5’
aphA-3
Kanamisin
5’ GCCGATGTGGATTGCGAAAA 3’
292
Braak vd.
1999 3’ GCTTGATCCCCAGTAAGTCA 5’
rpoB1
Rifampisin
5’ GTCCGTTTCGGCTTTAATATA 3’
1000
Enne vd. 2004 3’ AAGAAACGAGCATTCAGCAA 5’
rpoB2
Rifampisin
5’ CGCAAGCGACTCAAGAACAG 3’
1000
Enne vd. 2004 3’ GAGCAAATGTTCCATCTTCA 5’
bla(Z)
Ampisilin
5’ TACAACTGTAATATCGGAGGG 3’
778
Stovcik vd.
2008 3’ CAATAGGTTCAGATTGGCCC 5’
ermB
Eritromisin
5’ CATTTAACGACGAAACTGGC 3’
422
Aarestrup vd.
2000 3’ GGAACATCTGTGGTATGGCG 5’
catpIP
Kloramfenikol
5’ GGATATGAAATTTATCCCTC 3’
486
Aarestrup vd.
2000 3’ CAATCATCTACCCTATGAAT 5’
tet M
Tetrasiklin
5’ GTTAAATAGTGTTCTTGGAG 3’
657
Aarestrup vd.
2000 3’ CTAAGATATGGCTCTAACAA 5’
tet L
Tetrasiklin
5’ CATTTGGTCTTATTGGATCG 3’
475
Aarestrup vd.
2000 3’ ATTACACTTCCGATTTCGG 5’
vanB
vankomisin
5’ ATGGGAAGCCGATAGTC 3’
635
Mcgregor ve
Young 2000 3’ GATTTCGTTCCTCGACC 5’

33
3.2 Yöntem
3.2.1 İzolatlarının saflık kontrolü ve tür teşhisinin doğrulanması
Daha önce E. faecium olarak tanımlanmış ve antibiyotik duyarlılıkları incelenmiş olan
95 izolat arasından çoklu ilaç dirençli oldukları bilinen 64 izolat seçilmiş ve tekrar saflık
kontrolü yapmak için % 5 koyun kanlı TSA besiyerine ekilmiştir. E. faecium izolatları α
hemolitik olabilir yada hemolitik özellik göstermeyebilir. Bu nedenle suşlar öncelikle
besiyeri üzerinde α-hemolitik, β-hemolitik veya hemolitik olmama özelliklerine göre
incelenmiştir. Daha sonrasında spesifik koloniler seçilerek Gram boyamaları
yapılmıştır. Tekli, ikili veya kısa zincirler halinde görünen Gram- pozitif koklar
değerlendirmeye alınmıştır. Katalaz testi; öze ile temiz bir lam üzerine alınan bakteri
kolonisine % 3’lük H2O2 damlatılarak yapılmıştır. Hava kabarcığı oluşturmayan lamlar
katalaz negatif olarak değerlendirilmiştir. % 6.5 NaCl’de üremelerine ve lateks
yöntemine göre serolojik olarak D grubu yüzey antijeni içerip içermediklerine
bakılmıştır (Şekil 3.1). Bu özelliklerine göre yeniden Enterokok cinsine ait oldukları
saptanan 64 bakterinin tür teşhisi yapılmıştır (Facklam vd. 1997).
Teşhis işlemi ve MİK (Minimal İnhibisyon Konsantrasyonu) için Gram- negatif ve
Gram- pozitif bakterilerin otomatik tanı ve antibiyogram duyarlılık testlerinde
kullanılan, Akdeniz Üniversitesi Hastanesi Mikrobiyoloji Laboratuvarına ait Phonex
cihazı (Becton Drive, USA) kullanılmıştır. Phonex panelinin ID kısmı, kurutulmuş
kimyasal substratlı 45 kuyu ve 2 floresans kontrol kuyusu içerir. Bakteri tiplendirmesini
saptamak için bir seri konvansiyonel, kromatojenik ve florejenik biyokimyasal
maddeler kullanır. Üreme tabanlı ve enzimatik maddelerin her ikiside taksonomi
sınırları içinde çeşitli reaktif tiplerine döner. Testler bakteri tarafından kullanım ve
indirgeme olaylarının çeşitli indikatörlerle gösterilmesi presibine dayanır. Phonex ID
paneli ile çalışmada kullanılan tüm suşların E. faecium olduğu kanıtlanmıştır. Ayrıca tür
teşhisini doğrulamak amacıyla rapid ID 32 Strep Api testinden de yararlanılmıştır (Şekil
3.2). Tür teşhisi doğrulanan bakterilerden % 15 gliserol içeren TSB besiyerine stok
kültürler hazırlanmıştır.

34
Şekil 3.1 Enterokok izolatlarının tanısında kullanılan testler (Faclam vd. 1997)
%5 koyun kanlı Tripticase soya agar
Gram Boyama
Gram- pozitif koklar (katalaz(-)
%40 safralı ortamda üreme ve escülini hidrolize etme
D grubu Streptekok
%6.5 NaCl’de üreme
PYR (+)
Enterococcus spp.
Rapid ID 32 Strep Api ile tür teşhisi
Enterococcus faecium

35
Şekil 3.2 rapid ID 32 Strep Api testi ile Enterococcus faecium suşlarının doğrulanması

36
3.2.2 Antimikrobiyal dirençliliğin belirlenmesi
Antibiyotik duyarlılık testinde kullanılan Phonex cihazı AST paneli buyyon tabanlı bir
mikrodilüsyon testidir ve kuru antimikrobiyal ajanları içeren 84 kuyu ile üreme kontrol
kuyusu içerir. Bakteriyel bulanıklık, bakteriyel üremenin belirlenmesinde kullanılır.
Phonex sistemi, organizmanın üremesinin belirlenmesi için redox göstergesini kullanır.
Phonex VRE, yüksek düzey aminoglikozit direnci testleri NCCLS standart broth mikro
seyreltme ve NCCLS taramalı agar testi ile geliştirilerek optimize edilmiştir
(Anonymous 2000).
% 5 koyun kanlı TSA plaklarında saf olarak elde edilen enterokok kolonilerinden ID
broth içerisine steril eküvyonla BBL CrystalSpec nefelometre (Becton Drive, USA) ile
0.5-0.6 Mac Farland ölçülecek şekilde bakteri süspansiyonu hazırlanmıştır. Hazırlanan
süspansiyondan 25 µl indikatör çözeltisi eklenen AST broth içerisine alınmıştır. Bu
şekilde hazırlanan panel cihaza yerleştirilmiştir. Panel üzerindeki kuyucuklarda oluşan
biyokimyasal reaksiyonlara göre her bir izolatın türü teşhis edilmiş ve belirli
antibiyotiklere karşı MİK değerleri saptanmıştır. Ayrıca suşların antibiyotik
dirençlilik/duyarlılık profillerinin araştırılmasında disk difüzyon yöntemi de
kullanılmıştır. Kültürler 5mL MHB ortamına inoküle edilerek 37 °C’de inkübasyona
bırakılmıştır. İnkübasyondan sonra kültür süspansiyonunun bulanıklılığı McFarland 0.5
standardının absorbansı 625 nm’de 0.08 - 0.1 değerinde olacak şekilde ayarlanmıştır.
Absorbansı ayarlanan kültürler % 0.7 oranında agar içeren 5 mL yumuşak agar (MHB)
ortamına 100 µl inoküle edilerek, Müller Hinton (Oxoid, UK) agar plakları üzerine
homojen bir şekilde yayılmıştır. Antimikrobiyel ajanlar emdirilmiş 6 mm’ lik kağıt
diskler, agar yüzeyine yerleştirilmiş ve 37 ºC’de 18 saat inkübe edilmiştir. Vankomisin
antibiyotiği için ise 24 saat inkübasyon uygulanmıştır. Disk difüzyonu için, 10 farklı
antimikrobiyel madde kullanılmıştır. Kullanılan antimikrobiyel diskler ve içerikleri
çizelge 3.2’de verilmiştir. İnkübasyon sonrası, disklerin etrafındaki zon çapları
ölçülmüş ve Klinik Laboratuar Standartları Enstitüsü (CLSI)’ nün belirlediği kırılma
noktaları ile karşılaştırılmıştır. Buna göre sonuçlar duyarlı (S), orta dirençli (I) ve
dirençli (R) olarak belirlenmiştir (Anonymous 2011).

37
3.2.3 İzolatların plazmid profillerinin belirlenmesi
3.2.3.1 Plazmid izolasyonu
TSB broth besiyerinde 37 °C’de 18 saat geliştirilen E. faecium kültürlerinden, 10 mL’
lik TSB ortamlarına 1 mL’lik inokülasyonlar yapılarak, kültürler 37 °C’de 3–3.5 saat
inkübasyona tabi tutulmuştur. Bu süre bitiminde santrifüj tüplerine aktarılan bakteri
kültürleri, 6000 devirde 15 dakika santrifüj edilmiştir. Hücre çökeltisi kurutulduktan
sonra, 380 µL sakkaroz tamponunda çözülmüş ve steril mikrosantrifüj tüplerine
aktarılmıştır. 37 °C’ye kadar ısıtılan bu ortama 96.5 µL lizozim çözeltisi ilave edilerek
37 °C su banyosunda 5 dakika bekletilmiştir. 48.5 µL Tris-EDTA–1 uygulamasından
sonra, mikrosantrifüj tüplerine % 20 sodyum dodesil sülfat (SDS) çözeltisinden 28 µL
aktarılarak karıştırılmıştır. Bu aşamada ortamda viskozitenin artışı, lizizin başladığını
göstermektedir. Lizizin tamamlanması için, mikrosantrifüj tüpleri 37 ºC su banyosunda
10 dakika bekletilmiştir. Bu süre sonunda tüpler mekanik karıştırıcıda ve yüksek
devirde 30 saniye karıştırılarak kromozomal DNA’nın kırılması sağlanmıştır. Ortama,
yeni hazırlanmış 3 N NaOH çözeltisinden 28 µL ilave edilmiş ve tüpler düz bir zemin
üzerinde 10 dakika süre ile yavaş bir şekilde çevrilerek kromozomal DNA’nın alkali
denatürasyon koşulları oluşturulmuştur. Ortam pH’sının uygulama sırasında 12.1 – 12.3
arasında olup olmadığı kontrol edilmiştir. Denatürasyon aşamasının sonunda
mikrosantrifüj tüplerine 50 µL 2 M Tris-HCl çözeltisi aktarılarak, 3 dakika süre ile yine
düz bir zeminde karıştırılmıştır. Ortam pH’ sının 8.5–9.0 arasına düşüşü ile
nötralizasyonun sağlandığı belirlenmiştir. Mikrosantrifüj tüplerine, +4 °C’ de tutulan 5
M NaCl çözeltisinden 72 µL ve % 3 NaCl ile doyurulmuş fenol çözeltisinden 700 µL
ilave edilerek +4 °C’de 12500 devirde 20 dakika santrifüj işlemi uygulanmıştır.
Tüplerde oluşan üst faz, mikropipet ile yeni mikrosantrifüj tüplerine aktarılmış ve
deproteinasyonun sağlanması için kloroform/izoamilalkol (24:1) çözeltisinden 700 µL
ilave edilmiştir. Tüpler +4 ºC’de 12500 devirde 20 dakika santrifüj edilerek üst faz
alınmış ve 500 µL etanol ilave edilmiştir. Ekstraktlar - 20 ºC’de 1 gece bekletilmiş ve
plazmid DNA, 12500 devirde 20 dakika santrifüj edilmek suretiyle çöktürülmüştür. Son
aşamada sıvı faz akıtılarak DNA çökeltisi kurutulmuştur. Çökeltiler, 20 µL Tris-EDTA-

38
2 içerisinde çözülmüş ve elektroforez uygulamasından önce RNaz A stok çözeltisinden
2 µL ilave edilerek 37 ºC’de 45 dakika inkübe edilmiştir (Anderson ve McKay 1983).
Sakkaroz Çözeltisi
Tris 0.655 g
EDTA 0.0372 g
Sakkaroz 6.7 g
Distile su 100 mL
pH 8.0 ± 0.02
Kaba filtreden geçirildi ve 121 °C’de sterilize edildi.
Lizozim Çözeltisi
Tris 0.3 g
Lizozim 0.1 g
Distile su 10 mL
pH 8.0 ± 0.02
Membran filtre ile sterilize edildi.
Tris-EDTA-1
Tris 0.6 g
EDTA 9.31 g
Distile su 100 mL
pH 8.0 ± 0.02
Kaba filtreden geçirildi ve 121 °C’de sterilize edildi.
Tris-HCl
Tris-HCl 31.52 g
Distile su 100 mL
pH 7.0 ± 0.02
Kaba filtreden geçirildi ve 121 °C’de sterilize edildi.

39
SDS Çözeltisi
Tris 0.6 g
EDTA 0.74 g
SDS 20 g
Distile su 100 mL
pH 8.0 ± 0.02
Kaba filtreden geçirildi ve 121 °C’de sterilize edildi.
Tris_EDTA-2
Tris 0.121 g
EDTA 0.037 g
Distile su 100 mL
pH 7.5 ± 0.02
Kaba filtreden geçirildi ve 121 °C’de sterilize edildi.
%3 NaCI ile Doyurulmuş Fenol Çözeltisinin Hazırlanışı: 100 g fenol üzerine 20 mL
destile su ve 3 g NaCl aktarılarak 45 ºC’deki su banyosunda çözülmüştür. Ortama 0.1 g
hidroksiguinolin ilave edilmiş ve karıştırılarak oda sıcaklığında tutulmuştur.
RNaz A Çözeltisinin Hazırlanışı: 5 mL steril destile su içinde hazırlanan 0.05 M
sodyum asetat çözeltisinin pH’sı, asetik asit ile 5’e ayarlanmış ve üzerine 5 mg RNaz A
ilave edilmiştir. Kaynar su içerisinde 5 dakika tutulduktan sonra, ortam –20 ºC’de
saklanmıştır.
3.2.3.2 Elektroforez
DNA örneklerinin elektroforezi, % 0.7 agaroz içeren jellerde yapılmıştır (Meyers vd.
1976). Yatay jel sistemleri için agaroz, 100 mL tris-asetat elektroforez tamponu
içerisinde ve kaynar su banyosunda çözülmüştür. 45 ºC’ye kadar soğutulan ortam
elektroforez plakalarına 30–50 mL olacak şekilde aktarılmış ve jel tarakları
yerleştirilerek 30 dakika bekletilmiştir. Bu süre sonunda tampon çözelti, jeli kapatacak

40
şekilde elektroforez tanklarına dökülmüş ve taraklar dikkatlice çıkartılmıştır. 37 ºC’de
45 dakika inkübe edilen DNA örnekleri su banyosundan alınarak, 2 µL marker boya
çözeltisi ile karıştırılmış ve mikropipet aracılığı ile jel kuyucuklarına aktarılmıştır.
Elektroforez, 80 voltta 2–2.5 saat süreyle yapılmıştır. Marker boyanın jel sistemleri terk
etmesinden sonra, elektrik akımı kesilmiş ve ortamdan alınan jeller, kullanılan
elektroforez tamponunun yeni hazırlanmış 0.2 µg/mL etidyum bromit içeren
çözeltisinde 30 dakika boya işlemine tabi tutulmuştur. Boyama işlemi bitiminde jeller,
366 nm dalga boyunda ultraviyole ışıkta incelenmiş ve Kodak Gel Logic 200 Imaging
System kullanılarak fotoğrafları alınmıştır (Macrina vd. 1982).
TRİS-Asetat Tampon
Tris 4.84 g
Sodyum asetat 4.08 mL
EDTA 0.37 mL
Distile su 1000 mL
pH 8.0 ± 0.02
Marker Boya
Bromfenol blue 0.25 g
Sakkaroz 40 g
Distile su 100 mL
3.2.3.3 Plazmid büyüklüklerinin saptanması
E. faecium suşlarından izole edilen plazmidlerin büyüklüklerinin belirlenmesinde;
moleküler büyüklükleri bilinen (Sigma Chem. Co. D5292) ve 2.067-16.210 baz çifti
arasında değişen 11 süpersarmal parça içerikli ccc DNA marker’larının elektroforetik
hareketlilikleri ile büyüklüklerinin logaritmaları arasında tanımlanan doğrusal ilişkiden
yararlanılmıştır (Macrina vd. 1978, Southern 1979, Schaffer ve Sederof 1981). Marker
ccc DNA moleküllerinin agaroz jel fotoğrafları üzerinde ölçülen göç aralıkları ile

41
bilinen büyüklüklerinin logaritmik değerlerine bağlı olarak eğrileri çıkarılmıştır.
İstatistik analizlerle her farklı jel için korelasyon katsayısı ve eğrinin eğimi belirlenerek,
bakterilerden izole edilen plazmidlerin büyüklükleri saptanmıştır (Campbell 1974, Elder
ve Southern 1983, Elder vd. 1983).
Moleküler Büyüklük (W) = Antilog10 [ I. (α-G) + H ]
X = marker DNA moleküllerinin agaroz jel üzerindeki göç aralığı (mm)
Y = Marker DNA moleküllerinin büyüklüğü (kb)
A = X1 + X2 + X3 + ...........................+ Xn
B = X12 + X2
2 + X32 + ...........................+ Xn
2
C = log10Y1 + log10Y2 + log10Y3 + .....................+ log10Yn
D = (log10Y1 )2+ (log10Y2 )
2+ (log10Y3 )2+ .....................+( log10Yn)
2
E = X1 (log10Y1 )+X2 ( log10Y2 ) + X3 ( log10Y3 )+ .....................+Xn ( log10Yn)
G = Ortalama X = A/N
H = Ortalama Y = C/N
αααα = Moleküler büyüklüğü bilinmeyen plazmidin jel üzerindeki göçü (mm)
Eğrinin Eğimi (I) =
Korelasyon Katsayısı (J) =
3.2.4 Genomik DNA izolasyonu
TSB besiyeri ortamında 37 °C’de 18 saat geliştirilen E. faecium kültürleri, 10 mL’lik
steril santrifüj tüplerine aktarılmış ve 6000 rpm’de 15 dakika santrifüj işlemine tabi
tutulmuştur. Hücre çökeltisi 500 µl TE tampon içerisinde çözülerek 14000 g’de 1
dakika oda sıcaklığında tekrar çöktürülmüştür. Oluşan hücre çökeltisi SET (Sakkaroz-
EDTA-TrisHCl) tampon içerisinde çözüldükten sonra 50 µl lizozim eklenmiş ve 30 dk
37 °C’de inkübe edilmiştir. Bu içerik üzerine 200 µl TE tampon ve 30 µl % 20’ lik SDS
).(
).(
AGB
CGE
−
−
)]._()].[._([
).(
AGBCHD
CGE

42
eklenip, birkaç kez elde çevrilerek karıştırılmıştır. Karışım üzerine 100 µl 5 M NaCl
ilave edilip tekrar karıştırılmıştır. Daha sonra ortama eşit hacimde (~500 µl) fenol
aktarılmış ve emülsiyon oluşana kadar karıştırılmıştır. Ortam 14000 g’de 15 dk +4 °C’
de santrifüj edilerek fazların ayrılması sağlanmıştır. Üst faz mikropipet aracılığı ile yeni
bir mikrosantrifüj tüpüne aktarılmış ve üzerine eşit hacim (~500 µl) kloroform/izoamil
alkol eklenerek karıştırılmıştır. Karışım, 14000 g’de 15 dk oda sıcaklığında santrifüj
işlemine tabi tutulmuş ve oluşan üst faz yeni mikrosanrifüj tüplerine aktarılmıştır. Daha
sonra bu tüpte bulunan üst sıvının iki katı hacminde % 100’lük etanol tüpe ilave
edilmiştir. Bu karışım, 12000 rpm’ de +4 °C’de 15 dk santrifüj edilmiştir. Oluşan
çökelti havada kurutulduktan sonra 100 µl TE tampon ile çözülerek 2 µl RNaz A
eklenmiş ve 37 °C’de 45 dk inkübe edilmiştir. DNA saflığı spektrofotometrede
A260/A280 oranına bakılmak suretiyle ölçülmüştür.
Tris (Tris Hidroksimetil Amino Metan) / EDTA (Etilen Diamin Tetra Asetik Asit)
Tris 1,21g
EDTA 0.37g
Distile su 1000mL
pH 2N HCl kullanılarak 7.4’ e ayarlandı. 121 °C’de sterilize edildi.
SET (Sakkaroz-EDTA-Tris HCl) Tampon
Tris HCl 0,788g
EDTA 1,86g
Sakkaroz 20g
Distile su 100mL
pH 8’ e ayarlanıp 121 °C’de sterilize edildi.
% 20 SDS Çözeltisi
SDS 20g
Steril distile su 100mL

43
Kloroform/İzoamil Alkol Çözeltisi
Kloroform 24mL
İzoamil Alkol 1mL
3.2.5 Polimeraz zincir reaksiyonu (PZR)
PZR amplifikasyonları için Techne TC-512 Thermal Cycler (England) cihazı
kullanılmıştır. Ta (bağlanma sıcaklığı, °C) her bir primer için ayrı ayrı Ta= 2 x (A+T) +
4 x (G+C) formülü kullanılarak hesaplanmıştır. PZR sonrasında, elde edilen ürünlerden
5µl alınarak 2µl 6x yükleme boyası ile karıştırılmış ve % 1 oranında agaroz içeren jelde
100 volt elektrik akımı altında koşturulmuştur. Bant büyüklüklerinin belirlenmesi amacı
ile 1 kb DNA Marker’dan (Fermentas, Finland) yararlanılmıştır. Elektroforez sonrası
jeller, 0.2 µg/mL etidyum bromit (BioShop, Canada) çözeltisinde 20 dk süre ile boyama
işlemine tabi tutularak ve 366 nm dalga boyunda UV ışık altında translüminatör
kullanılarak görüntülenmiş ve fotoğrafları çekilmiştir (Kodak Gel Logic 200 Imaging
System).

44
3.2.5.1 Antibiyotik dirençliliği kodlayan gen bölgelerinin PZR ile araştırılması
rpoB1 ve rpoB2 gen bölgelerinin çoğaltılması (Enne vd. 2004)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 29,75 µl
5X PZR tamponu* 10 µl 1 X
dNTP (2mM)** 1 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 2 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 3 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
95 0C …………………………………… 5 dakika
95 0C …………………………………… 1 dakika
57 0C …………………………………... 1 dakika 35 döngü
72 0C …………………………………... 1 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

45
bla Z gen bölgesinin çoğaltılması (Stovcik vd. 2008)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 31,55 µl
5X PZR tamponu* 10 µl 1 X
dNTP (10mM)** 0.2 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 2 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 2 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
94 0C …………………………………… 5 dakika
94 0C …………………………………… 1 dakika
58 0C …………………………………... 30 saniye 35 döngü
72 0C …………………………………... 2 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

46
tetM ve tetL gen bölgelerinin çoğaltılması (Aarestrup vd. 2000)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 29,75 µl
5X PZR tamponu* 10 µl 1 X
dNTP (2mM)** 1 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 2 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 3 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
94 0C …………………………………… 5 dakika
94 0C …………………………………… 1 dakika
53.5 0C ………………………………... 30 saniye 35 döngü
72 0C …………………………………... 2 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

47
aphA3 gen bölgesinin çoğaltılması (Braak vd. 1999)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 28,55 µl
5X PZR tamponu* 10 µl 1 X
dNTP (10mM)** 0.2 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 6 µl 3 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 3 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
94 0C …………………………………… 5 dakika
94 0C …………………………………… 30 saniye
54.5 0C ………………………………... 45 saniye 32 döngü
72 0C …………………………………... 2 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

48
aac6’-aph2’’ gen bölgesinin çoğaltılması (Kobayashi vd. 2001)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 30,55 µl
5X PZR tamponu* 10 µl 1 X
dNTP (10mM)** 0.2 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 2 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 3 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
94 0C …………………………………… 5 dakika
94 0C …………………………………… 1 dakika
58 0C ……………….………………...... 1 dakika 30 döngü
72 0C …………………………………... 2 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

49
vanA gen bölgelerinin çoğaltılması (Dutka-Malen vd. 2005)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 31,05 µl
5X PZR tamponu* 10 µl 1 X
dNTP (10 mM)** 0.2 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 3 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 2.5 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
94 0C …………………………………… 5 dakika
94 0C …………………………………… 1 dakika
54 0C ……………….………………...... 1 dakika 30 döngü
72 0C …………………………………... 1 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

50
vanB gen bölgelerinin çoğaltılması (Mcgregor ve Young 2000)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 30,55 µl
5X PZR tamponu* 10 µl 1 X
dNTP (10 mM)** 0.2 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 3 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 3µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
94 0C …………………………………… 5 dakika
94 0C …………………………………… 1 dakika
53 0C ……………….………………...... 1 dakika 30 döngü
72 0C …………………………………... 2 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

51
ant6 gen bölgesinin çoğaltılması (Kobayashi vd. 2001)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 30,55 µl
5X PZR tamponu* 10 µl 1 X
dNTP (10 mM)** 0.2 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 2 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 3 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
95 0C …………………………………… 10 dakika
94 0C …………………………………… 30 saniye
54 0C ……………….………………...... 30 saniye 30 döngü
72 0C …………………………………... 30 saniye
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

52
cat gen bölgesinin çoğaltılması (Aarestrup vd. 2000)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 29,75 µl
5X PZR tamponu* 10 µl 1 X
dNTP (2 mM)** 1 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 4 µl 2 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 3 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
94 0C …………………………………… 5 dakika
94 0C …………………………………… 1 dakika
50 0C ……………….………………...... 1 dakika 30 döngü
72 0C …………………………………... 2.5 dakika
72 0C …………………………………… 7 dakika
+ 4 0C ………………………………….. ∞

53
ermB gen bölgesinin çoğaltılması (Aarestrup vd. 2000)
PZR karışım içeriği Reaksiyon hacmi Son hacim
Steril distile su 32,75 µl
5X PZR tamponu* 10 µl 1 X
dNTP (2 mM)** 1 µl 0,04 mM
İleri primer 1 µl 0,2 µM
Geri primer 1µl 0.2 µM
MgCl2 (25mM)* 2 µl 1 mM
Taq DNA Polimeraz (5u/mL)* 0,25 µl 0.025u/mL
DNA 2 µl
Toplam 50µl
* Promega, USA ** Fermentas, Finland
PZR koşulları
93 0C …………………………………… 5 dakika
93 0C …………………………………… 1 dakika
56 0C ……………….………………...... 1 dakika 35 döngü
72 0C …………………………………... 1 dakika
72 0C …………………………………… 10 dakika
+ 4 0C ………………………………….. ∞

54
3.2.6 DNA dizi analizi
Antibiyotik dirençliliğini kodlayan gen bölgelerine özel olarak tasarlanan primerler
kullanılarak PZR gerçekleştirilmiştir. Elde edilen PZR ürünleri saflaştırılmış ve dizi
analizi REFGEN (ODTÜ Teknokent, Ankara) firması tarafından yapılmıştır. Sonuçlar
PSI-BLAST programı kullanılarak değerlendirilmiştir.
3.2.7 Konjugasyon denemeleri
Konjugal verici ve alıcı suşlar antibiyotik ilaveli TSB ortamlarında 37 ºC’de 18 saat
geliştirilmiş ve aynı ortamlarda 10–1 düzeyinde dilüsyonları hazırlanmıştır. Bu
dilüsyonlardan alınan 2 mL verici ve 3 mL alıcı suş kültürleri karıştırıldıktan sonra,
filtrasyon işlemi ile steril membran filtreler (0.45 µm, Sartorius, Germany) üzerinde
toplanmıştır. Alıcı ve verici suş kültür karışımlarını içeren filtreler TSA plaklarına
yerleştirilerek, 37 ºC’de 18 saat inkübe edilmiştir. Bu süre bitiminde TSA plaklarından
alınan filtreler, 1 mL steril fizyolojik su içerisinde çözülmüş ve 10–8 düzeyine kadar seri
dilüsyonları hazırlanmıştır. Her bir dilüsyondan alıcı ya da verici suşun duyarlı olduğu
her iki antibiyotiği içeren agar plaklarına sürme ekim yapılarak 37 ºC’de 18 saat
inkübasyona bırakılmıştır (Lampkowska vd. 2008).

55
4. BULGULAR VE TARTIŞMA
4.1 Enterococcus faecium Suşlarının Antibiyotik Duyarlılık Düzeylerinin Tanımlanması
Araştırmada kullanılan Akdeniz Üniversitesi Hastanesi Mikrobiyoloji Laboratuvarı
kültür koleksiyonundan sağlanan 64 E. faecium izolatının 25’ini idrar, 22’sini rektal,
12’sini kan, 3’ünü pü ve geri kalan 2 tanesini bos ve katater kültürlerinden elde edilen
suşlar oluşturmaktadır. Enterokokların her tür nozokomiyal enfeksiyonlardan, özellikle
üriner sistem enfeksiyonlarından izolasyon sıklığı gittikçe artmaktadır. Ülkemizde
yapılan araştırmaların çoğunda enterokoklar en sık idrardan izole edilmişlerdir (Gür
1997, Esen vd. 2001). Çalışmamızda da diğer çalışmalarla uyumlu olarak E. faecium
izolatlarına en sık idrarda rastlanmıştır.
E. faecium suşlarının antibiyotik duyarlılık profillerinin belirlenmesinde bölüm 3.2.2’de
anlatıldığı gibi Phonex sistemi ve disk difüzyon yöntemi kullanılmıştır. Phonex
antibiyogram sisteminde çalışılan antibiyotik disklerinin MİK konsantrasyonları
sonuçlarına göre direnç özellikleri incelenecek 10 farklı antibiyotik disk difüzyon
yöntemi ile de doğrulanmıştır (Şekil 4.1). Çalışmada kullanın suşların antibiyotik
duyarlılık sonuçları çizelge 4.1’de verilmiştir.
Şekil 4.1 Enterococcus faecium suşlarının disk difüzyon testi

56
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları
Bakteri Adı= E. faecium (1) Bakteri Adı= E. faecium (2)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 20mm Kloramfenikol X 24mm
Gentamicin SYN X 24mm S<=500 Gentamicin SYN X - R>500
Streptomicin SYN X 4mm R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 28mm S<=0.5 Tetrasiklin X 24mm S<=0.5
Teikoplanin X 5mm R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (3) Bakteri Adı= E. faecium (4)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X 10mm R>4 Eritromisin X - R>4
Rifampin X - Rifampin X 13mm
Kanamicin X - Kanamicin X -
Kloramfenikol X 15mm Kloramfenikol X 19mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X - R>8 Tetrasiklin X 27mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X R>16
*Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

57
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (5) Bakteri Adı= E. faecium (6)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X 17mm I 4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 12mm Kloramfenikol X 17mm
Gentamicin SYN X - R>500 Gentamicin SYN X 23mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X 23mm S<1000
Tetrasiklin X 10mm R>8 Tetrasiklin X 30mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (8) Bakteri Adı= E. faecium (11)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X 14mm R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 18mm Kloramfenikol X 24mm
Gentamicin SYN X - R>500 Gentamicin SYN X 26mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 28mm S<=0.5
Teikoplanin X 9mm R>16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

58
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (12) Bakteri Adı= E. faecium (13)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X 12mm Rifampin X 16mm
Kanamicin X - Kanamicin X -
Kloramfenikol X 17mm Kloramfenikol X 15mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 26mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (15) Bakteri Adı= E. faecium (17)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X 16mm I 4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X 10mm Kanamicin X -
Kloramfenikol X 30mm Kloramfenikol X 26mm
Gentamicin SYN X 28mm S<=500 Gentamicin SYN X - R>500
Streptomicin SYN X 26mm S<=1000 StreptomicinSYN X - R>1000
Tetrasiklin X 30mm S<=0.5 Tetrasiklin X 30mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

59
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (19) Bakteri Adı= E. faecium (21)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X 13mm R>16 Vankomisin X 14mm R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X 17mm Rifampin X -
Kanamicin X 13mm Kanamicin X -
Kloramfenikol X 24mm Kloramfenikol X 26mm
Gentamicin SYN X 22mm S<=500 Gentamicin SYN X 24mm S<=500
Streptomicin SYN X 20mm S<=1000 StreptomicinSYN X - R>1000
Tetrasiklin X 24mm S<=0.5 Tetrasiklin X 32mm S<=0.5
Teikoplanin X 9mm R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (24) Bakteri Adı= E. faecium (25)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X 10mm R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X 10mm R>4 Eritromisin X - R>4
Rifampin X - Rifampin X 14mm
Kanamicin X - Kanamicin X -
Kloramfenikol X 17mm Kloramfenikol X 19mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 18mm R>8 Tetrasiklin X 27mm S 2
Teikoplanin X - R>16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

60
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (26) Bakteri Adı= E. faecium (27)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X 23mm S<=1
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X 9mm
Kanamicin X - Kanamicin X -
Kloramfenikol X 17mm Kloramfenikol X 22mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 25mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X 20mm S<=1
Bakteri Adı= E. faecium (28) Bakteri Adı= E. faecium (29)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X 15mm I 8
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X 13mm Kanamicin X -
Kloramfenikol X 19mm Kloramfenikol X 19mm
Gentamicin SYN X 25mm S<=500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X 6mm R>1000
Tetrasiklin X 10mm R>8 Tetrasiklin X 30mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X 18mm S<=1
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

61
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (30) Bakteri Adı= E. faecium (31)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X 10mm R>16 Vankomisin X 12mm R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 25mm Kloramfenikol X 24mm
Gentamicin SYN X - R>500 Gentamicin SYN X 29mm S<=500
Streptomicin SYN X 6mm R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 29mm S<=0.5 Tetrasiklin X 26mm S<=0.5
Teikoplanin X 6mm R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (32) Bakteri Adı= E. faecium (33)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X 22mm S<=1 Vankomisin X 22mm S<=1
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 23mm Kloramfenikol X 20mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 26mm S<=0.5
Teikoplanin X 23mm S<=1 Teikoplanin X 20mm S<=1
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

62
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (34) Bakteri Adı= E. faecium (35)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X 24mm S<=1 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X 11mm Rifampin X 12mm
Kanamicin X - Kanamicin X 12mm
Kloramfenikol X 23mm Kloramfenikol X 19mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 30mm S<=0.5
Teikoplanin X 22mm S<=1 Teikoplanin X - R>16
Bakteri Adı= E. faecium (36) Bakteri Adı= E. faecium (37)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X 11mm R>16 Vankomisin X 23mm S 2
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X 13mm
Kanamicin X - Kanamicin X -
Kloramfenikol X 24mm Kloramfenikol X 27mm
Gentamicin SYN X 25mm S<=500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 26mm S<=0.5 Tetrasiklin X 28mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X 20mm S<=1
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

63
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (38) Bakteri Adı= E. faecium (39)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X 11mm R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 25mm Kloramfenikol X 29mm
Gentamicin SYN X 26mm S<=500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 28mm S<=0.5 Tetrasiklin X 28mm S<=0.5
Teikoplanin X 6mm R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (41) Bakteri Adı= E. faecium (42)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X 14mm Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 24mm Kloramfenikol X 18mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 29mm S<=0.5 Tetrasiklin X 9mm R>4
Teikoplanin X 13mm I 16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

64
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (43) Bakteri Adı= E. faecium (46)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X 28mm S<=1
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin I 20mm I 4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X 14mm
Kloramfenikol X 18mm Kloramfenikol X 25mm
Gentamicin SYN X - R>500 Gentamicin SYN X 23mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 9mm R>4 Tetrasiklin X 24mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X 25mm S<=1
Bakteri Adı= E. faecium (48) Bakteri Adı= E. faecium (49)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X 24mm S<=1 Vankomisin X 14mm R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X 13mm
Kanamicin X - Kanamicin X 12mm
Kloramfenikol X 24mm Kloramfenikol X 28mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X 22mm S<=1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 30mm S<=0.5
Teikoplanin X 21mm S<=1 Teikoplanin X 8mm R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

65
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (50) Bakteri Adı= E. faecium (52)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X 21mm S 2 Vankomisin X 21mm S 2
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 19mm Kloramfenikol X 18mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 30mm S<=0.5 Tetrasiklin X 28mm S<=0.5
Teikoplanin X 22mm S<=1 Teikoplanin X 20mm S<=1
Bakteri Adı= E. faecium (55) Bakteri Adı= E. faecium (56)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X 24mm S<=1
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X 12mm Rifampin X 10mm
Kanamicin X - Kanamicin X -
Kloramfenikol X 26mm Kloramfenikol X 24mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 32mm S<=0.5 Tetrasiklin X 30mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X 25mm S<=1
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

66
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (57) Bakteri Adı= E. faecium (59)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X 9mm R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X 13mm
Kloramfenikol X 15mm Kloramfenikol X 17mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X 25mm S<=1000
Tetrasiklin X 10mm R>8 Tetrasiklin X 30mm S<=0.5
Teikoplanin X 6mm R>16 Teikoplanin X 16mm R>16
Bakteri Adı= E. faecium (60) Bakteri Adı= E. faecium (63)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 16mm Kloramfenikol X 22mm
Gentamicin SYN X - R>500 Gentamicin SYN X 28mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 28mm S<=0.5 Tetrasiklin X 26mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

67
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (64) Bakteri Adı= E. faecium (65)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 16mm Kloramfenikol X 20mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 9mm R>8 Tetrasiklin X 21mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (67) Bakteri Adı= E. faecium (68)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X 15mm I 2
Rifampin X 12mm Rifampin X -
Kanamicin X - Kanamicin X 12mm
Kloramfenikol X 22mm Kloramfenikol X 25mm
Gentamicin SYN X - R>500 Gentamicin SYN X 26mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X 21mm S<=1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 24mm S<=0.5
Teikoplanin X 8mm R>16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

68
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (69) Bakteri Adı= E. faecium (70)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X 22mm S<=1 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X 12mm
Kloramfenikol X 24mm Kloramfenikol X 24mm
Gentamicin SYN X - R>500 Gentamicin SYN X 24mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X 21mm S<=1000
Tetrasiklin X 27mm S<=0.5 Tetrasiklin X 30mm S<=0.5
Teikoplanin X 20mm S<=1 Teikoplanin X - R>16
Bakteri Adı= E. faecium (71) Bakteri Adı= E. faecium (74)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X 22mm S<=1 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X 9mm R>4 Eritromisin X - R>4
Rifampin X 11mm Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 19mm Kloramfenikol X 16mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X - R>4 Tetrasiklin X 10mm R>8
Teikoplanin X 20mm S<=1 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

69
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (75) Bakteri Adı= E. faecium (76)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X 16mm R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X 11mm Kanamicin X -
Kloramfenikol X 16 mm Kloramfenikol X 15mm
Gentamicin SYN X 26 mm S<=500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X - R>8 Tetrasiklin X 36mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (77) Bakteri Adı= E. faecium (79)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 16 mm Kloramfenikol X 15mm
Gentamicin SYN X - R>500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 9mm R>4 Tetrasiklin X 9mm R>8
Teikoplanin X 6mm R>16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

70
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (80) Bakteri Adı= E. faecium (81)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X 16mm
Kanamicin X 12mm Kanamicin X -
Kloramfenikol X 16mm Kloramfenikol X 17mm
Gentamicin SYN X 26 mm S<=500 Gentamicin SYN X - R>500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 26mm S<=0.5 Tetrasiklin X 26mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X 11mm I 16
Bakteri Adı= E. faecium (84) Bakteri Adı= E. faecium (87)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X 13mm R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X 13mm Kanamicin X -
Kloramfenikol X 22mm Kloramfenikol X 16mm
Gentamicin SYN X 24mm S<=500 Gentamicin SYN X 28mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 30mm S<=0.5 Tetrasiklin X 30mm S<=0.5
Teikoplanin X 6mm R>16 Teikoplanin X 8mm R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

71
Çizelge 4.1 E. faecium suşlarının disk difüzyon ve MİK testlerine göre antibiyotik duyarlılıkları (devam)
Bakteri Adı= E. faecium (89) Bakteri Adı= E. faecium (90)
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ R S I
ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X -
Kloramfenikol X 24mm Kloramfenikol X 16mm
Gentamicin SYN X 15mm S<=500 Gentamicin SYN X 13mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 26mm S<=0.5 Tetrasiklin X 30mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Bakteri Adı= E. faecium (94) Bakteri Adı= E. faecium (95)
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
ANTİBİYOTİK DİSKLERİ
R S I ZON ÇAPI
MİK PHONEX
Vankomisin X - R>16 Vankomisin X - R>16
Ampicilin X - R>8 Ampicilin X - R>8
Eritromisin X - R>4 Eritromisin X - R>4
Rifampin X - Rifampin X -
Kanamicin X - Kanamicin X 11mm
Kloramfenikol X 22mm Kloramfenikol X 17mm
Gentamicin SYN X - R>500 Gentamicin SYN X 26mm S<=500
Streptomicin SYN X - R>1000 StreptomicinSYN X - R>1000
Tetrasiklin X 9mm R>4 Tetrasiklin X 30mm S<=0.5
Teikoplanin X - R>16 Teikoplanin X - R>16
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli

72
Çizelge 4.2 Standart E. faecalis ATCC29212 suşunun disk difüzyon ve MİK testlerine göre antibiyotik duyarlılığı
Bakteri Adı= E. faecalis ATCC 29212 (Kontrol)
ANTİBİYOTİK DİSKLERİ R S I ZON ÇAPI MİK PHONEX
Vankomisin X 18mm S 2
Ampicilin X 26mm S<=2
Eritromisin X 17mm I 2
Rifampin X 12mm
Kanamicin X -
Kloramfenikol X 18mm
Gentamicin SYN X 17mm S<=500
Streptomicin SYN X 15mm S<=1000
Tetrasiklin X 17mm I 8
Teikoplanin X 17mm S<=1
Sadece disk difüzyon testi yapılmıştır. MİK=Minimal İnhibisyon Konsantırasyonu R=Dirençli, S=Duyarlı, I=Orta Dirençli
64 adet E. faecium suşunun tamamı ampsiline, kanamisine, eritromisine ve rifampisine;
52 tanesi vankomisine, 56 tanesi yüksek düzey streptomisine, 43 tanesi yüksek düzey
gentamisine, 22 tanesi kloramfenikole, 14 tanesi tetrasikline ve 51 tanesi teikoplanine
dirençli bulunmuştur (Çizelge 4.3).

73
Çizelge 4.3 E. faecium suşlarının antibiyotik gruplarına karşı direnç oranları
ANTİBİYOTİK GRUBU ANTİBİYOTİK ADI
DİSK İÇERİĞİ DUYARLILIK (%)
GLİKOPEPTİDLER S I R
Vankomisin 30 µg 18.75 1.56 79.6
Teicoplanin 30 µg 20.3 3.1 76.5
Grup Ortalaması 19.5 2.33 78.05
MAKROLİDLER
Eritromisin 15 µg - 6.25 93.75
Kloramfenikol(Fenikoler)
30 µg - 34.3 -
Rifampin (Ansamisinler) 5 µg - 1.56 98.4
Grup Ortalaması - 14.03 64.05
TETRASİKLİNLER
Tetrasiklin 30 µg 78.1 - 21.8
Grup Ortalaması 78.1 - 21.8
AMİNOGLİKOZİDLER
Gentamisin 120 µg 32.8 - 67.18
Kanamisin 30 µg - 1.56 98.4
Streptomisin 300 µg 12.5 - 87.5
Grup ortalaması 15.1 1.56 84.36
β - LAKTAMLAR
Ampicilin 10 µg - - 100
Grup ortalaması - - 100
Sadece disk difüzyon testi yapılmıştır. R=Dirençli, S=Duyarlı, I=Orta Dirençli
Suşlarının toplam antibiyotik dirençlilik yüzdeleri; ampsiline % 100, streptomisine %
87.5, kanamisine % 100, gentamisine % 67.18, kloramfenikole % 32.81, tetrasikline %
21.87, rifampisine % 100, vankomisine % 96.87, eritromisine % 100 ve teikoplanine %
79.68 olarak belirlenmiştir (Şekil 4.2).
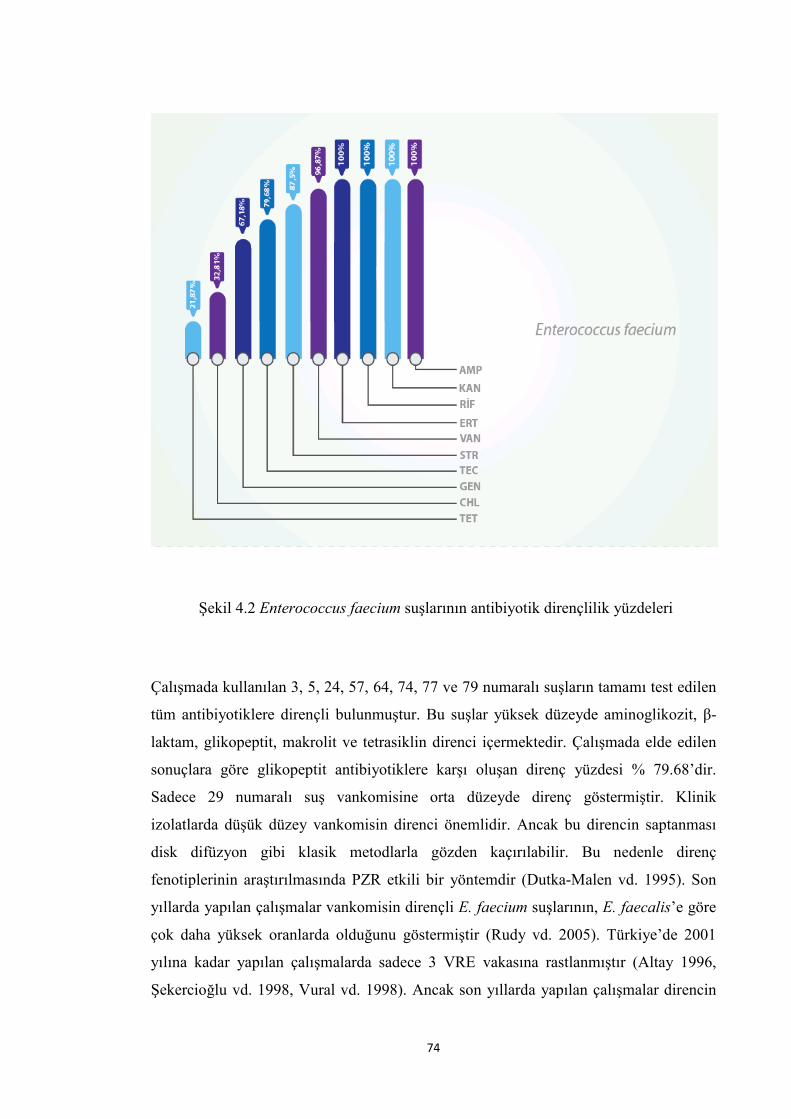
74
Şekil 4.2 Enterococcus faecium suşlarının antibiyotik dirençlilik yüzdeleri
Çalışmada kullanılan 3, 5, 24, 57, 64, 74, 77 ve 79 numaralı suşların tamamı test edilen
tüm antibiyotiklere dirençli bulunmuştur. Bu suşlar yüksek düzeyde aminoglikozit, β-
laktam, glikopeptit, makrolit ve tetrasiklin direnci içermektedir. Çalışmada elde edilen
sonuçlara göre glikopeptit antibiyotiklere karşı oluşan direnç yüzdesi % 79.68’dir.
Sadece 29 numaralı suş vankomisine orta düzeyde direnç göstermiştir. Klinik
izolatlarda düşük düzey vankomisin direnci önemlidir. Ancak bu direncin saptanması
disk difüzyon gibi klasik metodlarla gözden kaçırılabilir. Bu nedenle direnç
fenotiplerinin araştırılmasında PZR etkili bir yöntemdir (Dutka-Malen vd. 1995). Son
yıllarda yapılan çalışmalar vankomisin dirençli E. faecium suşlarının, E. faecalis’e göre
çok daha yüksek oranlarda olduğunu göstermiştir (Rudy vd. 2005). Türkiye’de 2001
yılına kadar yapılan çalışmalarda sadece 3 VRE vakasına rastlanmıştır (Altay 1996,
Şekercioğlu vd. 1998, Vural vd. 1998). Ancak son yıllarda yapılan çalışmalar direncin

75
ne derece tehlikeli boyutlara ulaştığının göstergesidir. Aygün vd. (2008) yapmış
oldukları çalışmalarda 467 hastanın % 1.9’unda VRE kolonizasyonu belirlenmiş ve
bunların tamamının vankomisin ve teikoplanine dirençli E. faecium olduğu
tanımlanmıştır. Ergani- Özcan vd. (2008) pediatri bölümünde yatan hastalarla yaptığı
bir yıllık sürveyans taraması sonucunda, 586 hastadan sadece 10 tanesinde VRE suşu
saptamıştır. Aynı yılda Sayıner (2008) 250 hastada aldığı rektal sürüntü örneğinde VRE
oranını % 15 olarak bildirmiştir. Yiş vd. (2010) 123 perirektal sürüntü örneği ile yaptığı
çalışmada VRE kolonizasyon oranını % 14.6 olarak belirlemiştir. Brezilya’da
D’azevedo ve Furtado (2008) tarafından yapılan bir çalışmada 81 hastanın fekal
sürveyans kültürlerinde, tamamı vankomisin ve teikoplanine dirençli, 37 VRE suşu
izole edilmiş ve bunların 20 tanesi (% 54,05) E. faecium olarak tanımlanmıştır. Bu
oranlarla kıyaslandığında çalışmamızda elde edilen veriler oldukça yüksek olup, elde
ettiğimiz bulgular vankomisin dirençli enterokok sayısının ülkemizde artığını
göstermektedir. Çalışmada kullandığınız 32 numaralı suş, hem vankomisine ve hem de
yüksek düzey gentamisin direncine sahiptir. Vankomisine dirençli suşlar genellikle
diğer antibiyotiklere de dirençlilik içermektedir (French 1998, Gordon vd. 1998).
Yüksek düzey aminoglikozit direncinin giderek yaygınlaşması, enterokokların neden
olduğu, aminoglikozitlerin sinerjik etkisi ile tedavi edilen enfeksiyonlar için önemli bir
sorundur. Çünkü böyle bir direncin varlığında penisilin ve aminoglikozit
kombinasyonunun etkisi ortadan kalkar. Bu nedenle enterokoklar ile oluşan ağır
enfeksiyonlarda yüksek düzey aminoglikozit direncinin belirlenmesi gerekmektedir
(Robert ve Moellering 2000). Çalışmamızda aminoglikozit antibiyotiklerden yüksek
seviyede gentamisin direnci (HLGR) içeren suşlar % 67.18, yüksek seviyede
streptomisin direnci (HLSR) içeren suşlar ise % 87.5 olarak bulunmuştur. Hem HLGR
hem de HLSR gösteren suşların yüzdesi % 62.5 oranındadır. Çalışmamızda kanamisine
yüksek düzey direnç oranı % 98.4 olarak bulunmuştur. Sadece 46 numaralı suş orta
düzeyde kanamisin direnci göstermiştir. Karadenizli vd. (2002) enterokok türlerine göre
yüksek düzey aminoglikozit düzeyleri üzerine yaptığı çalışmada, E. faecium suşları için
direnci; HLGR % 25.6 ve HLSR % 11.6 olarak belirlenmiştir. Mert-Dinç vd. (2009)’
nın yaptığı çalışmada aynı antibiyotiklere direnç oranlarını E. faecium suşunda sırasıyla
% 52 ve % 61.5 bulunmuştur. Aral vd. (2011) klinik kökenli enterokoklarla yaptıkları

76
çalışmada, izole ettikleri 96 E. faecium suşunun % 60’ında HLGR, % 65’inde HLSR
direnci saptamıştır. Kobayashi vd. (2001) yaptıkları çalışmada E. faecium için HLGR’
yi % 43 olarak saptamıştır. Mounir vd. (2011) ise HLGR oranını % 29.5, HLSR oranını
ise % 25 olarak belirlemiştir. Çalışmamızda elde edilen yüksek düzey aminoglikozit
direnç oranları, bu konuda yapılan çalışma bulgularıyla uyumlu bulunmuştur.
Makrolit grubu antibiyotiklere karşı E. faecium için belirlenen direnç oranları oldukça
yüksektir. Aarestrup vd. (2000) yaptıkları çalışmada E. faecium klinik örneklerde
eritromisin direnç oranını % 20, kloramfenikol direnç oranını % 12, tetrasiklin direnç
oranını ise % 12 olarak bulmuşlardır. Trieu-Cuot vd. (1993) klinik E. faecium suşlarının
yaklaşık % 50’sini kloramfenikole dirençli bulmuştur.
Çalışmamızda direnç yüzdeleri eritromisin ve rifampisin için % 100, kloramfenikol için
% 32.8’dir. Çalışmadaki 5 numaralı suş dışında tüm suşların kloramfenikole orta
düzeyde direnç göstermeleri dikkat çekicidir. Araştırmamızda, test edilen E. faecium
suşlarında tetrasiklin antibiyotiklerine direnç oranı % 21.87 bulunmuştur. Klinik
enfeksiyonlardan, gıda maddelerinden ve hayvansal gıdalardan izole edilen E. faecium
suşları arasında söz konusu dirençlilikler yaygın bir şekilde bulunmaktadır (Aarestrup
vd. 2000).
Beta laktamaz üreten enterokoklar ilk olarak 1983 yılında tespit edilmiştir. Bazı
araştırıcılar, penisiline dirençli izolatların hiçbirinin β-laktamaz üreticisi olmadığını
saptamıştır (Murray ve Mederski –Samaroj 1983, Aarestrup vd. 2000). Bu durum
enterokoklarda penisilin dirençliliğin farklı bir mekanizma ile sağlandığını
kanıtlamaktadır. Ülkemizde yapılan çalışmalarda beta- laktam antibiyotiklere dirençli
enterokokların giderek arttığına dikkat çekilerek, bu artışta penisilinlere dirençli, beta-
laktamaz üretmeyen suşların rol aldığı ileri sürülmüştür (Gür 1997). Çalışmamızda grup
temsilcisi olan ampisilin incelenmiş ve tüm suşlar ampisiline dirençli bulunmuştur.

77
4.2 Enterococcus faecium Suşlarının Plazmid Profillerinin Tanımlanması
64 adet E. faecium suşunun ve kontrol olarak kullanılan E. faecalis ATCC 29212
suşunun plazmid içerikleri alkali denatürasyon yöntemi kullanılarak araştırılmıştır
(Anderson ve McKay 1983). Plazmid izolasyonu ve agaroz jel elektroforezi çalışmaları
sonucu incelenen suşlarda; sayıları 1 ile 10 arasında değişen farklı moleküler
büyüklüklere sahip plazmid içerikleri belirlenmiştir. Saptanan plazmidlerin en küçüğü
2.2 kb en büyüğü ise 35.8 kb moleküler büyüklükte bulunmuştur (Şekil 4.3 ).
Enterokok suşları ile yürütülen diğer çalışmalarda; 2-8 adet arasında değişen sayıda
plazmid içerdikleri saptanmıştır. Bu plazmidlerin büyüklükleri ise 4-111 kb arasında
bulunmuştur. Son vd. (1999) vankomisin dirençli 19 E. faecium suşuna ait plazmid
profillerini ve antibiyotik dirençliliklerini incelemiş ve bu suşların 15’inde büyüklükleri
1.5-34 kb arasında değişen 5 ile 8 adet arasında plazmid tanımlamıştır. Rosvoll vd.
(2010), 93 E. faecium izolatının 88’inde 1-7 adet plazmidin olduğunu belirlemiştir.
Çöleri vd. (2004), klinik enterokok izolatlarında büyüklükleri 2.08 ile 56.15 kb arasında
olan ve sayıları 1 ile 11 adet arasında değişen plazmid içeriği saptamıştır. Ayrıca bu
araştırmada enterokoklarda plazmid üzerinde kodlanan bir antibiyotik direnci tespit
edilmiştir.
Bu çalışmada 36 ve 38, 50 ve 52, 77 ve 79, 80, 84 ve 90 numaralı suşlarının aynı
plazmid içeriğine sahip oldukları belirlenmiştir. Diğer tüm suşlarda plazmid içerikleri,
sayı ve büyüklük bakımından değişim göstermiştir. Aynı plazmid içeriğine sahip
suşların farklı hastalardan izole edilerek tanımlanması, söz konusu izolatların klonal
yayılımına işaret etmektedir. Özellikle çoklu antibiyotik direncine sahip suşların klinik
yayılımının araştırılması açısından bu veriler önem taşımaktadır.

78
Şekil 4.3 Enterococcus faecium suşlarının plazmid içerikleri. M; Süper Sarmal ccc DNA Marker,
1 E. faecium (kb) 35.3, 27.2, 16.5, 14.8, 12.3, 10.8, 7.9, 5.5, 3.5, 2.8
2 E. faecium (kb) 31.5, 23.6, 16.8, 2.8
3 E. faecium (kb) 32.9, 23.2, 17.0, 12.5, 3.4
4 E. faecium (kb) 35.8, 25.9, 17.1, 12.4, 7.7, 6.4, 2.2
5 E. faecium (kb) 29.5, 16.8, 11.5, 6.9, 5.4, 3.6, 2.3
6 E. faecium (kb) 35.0, 21.9, 17.2, 12.9, 7.3, 6.1, 4.4, 3.4, 2.2
8 E. faecium (kb) 29.6, 20.1, 15.8, 13.1, 9.7, 3.7, 2.2
11 E. faecium (kb) 31.5, 28.6, 17.7, 13.1, 10.2, 5.9, 3.6, 2.9
12 E. faecium (kb) 33.8, 27.7, 17.9, 13.5, 10.4, 5.2, 4.3, 2.2
13 E. faecium (kb) 29.7, 20.1, 16.5, 12.7, 7.8, 4.4, 3.8, 2.2
15 E. faecium (kb) 21.1, 18.4, 12.7, 8.8, 6.1, 4.8, 4.1, 2.4
17 E. faecium (kb) 26.8, 19.3, 18.0, 13.5, 3.4, 2.3
19 E. faecium (kb) 4.6
21 E. faecium (kb) 25.7, 22.4, 18.0, 16.2, 13.7, 7.1, 6.2, 3.9
24 E. faecium (kb) 23.9, 17.2, 13.5, 3.8
25 E. faecium (kb) 24.6, 17.5, 13.3, 8.3, 5.7, 4.8, 2.4
26 E. faecium (kb) 24.4, 17.7, 13.1, 7.9, 4.7, 2.4
27 E. faecium (kb) 21.2, 18.1
28 E. faecium (kb) 21.2, 19.6, 14.1, 4.2, 2.5
29 E. faecium (kb) 24.6, 17.5, 15.2, 4.1
30 E. faecium (kb) 24.8, 18.9, 14.5, 7.8, 5.6, 4.8, 3.2, 2.5
M Marker (bç) 16210, 14174, 12138, 10102, 8066, 7045, 6030, 5012, 3990, 2972, 2067
Supercolied ccc DNA marker (Sigma Chem. Co., USA, D 5292)
1 2 3 4 5 6 8 11 12 13 15 17 19 21 24 25 26 27 28 29 30 M

79
Şekil 4.3 Enterococcus faecium suşlarının plazmid içerikleri (devam). M; Süper sarmal ccc DNA Marker
31 E. faecium (kb) 20.2, 18.9, 17.1, 10.5, 7.6, 4.8, 3.7, 2.8
32 E. faecium (kb) 34.5, 23.2, 21.2, 15.8, 5.3, 3.4
33 E. faecium (kb) 21.7,19.6, 16.7, 14.5, 9.6, 6.8, 3.4
34 E. faecium (kb) 20.8, 19.1, 16.4, 8.7, 6.4, 4.6
35 E. faecium (kb) 22.7, 20.6, 18.7, 16.3, 11.5, 6.9, 5.4, 3.2, 2.2
36 E. faecium (kb) 22.5, 19.4, 17.9, 16.2, 13.4, 7.0, 6.3, 3.1
37 E. faecium (kb) 21.9, 18.6, 15.8, 5.8, 2.9
38 E. faecium (kb) 22.5, 19.4, 17.9, 16.2, 13.4, 7.0, 6.3, 3.1
39 E. faecium (kb) 18.8, 15.3, 2.9
41 E. faecium (kb) 21.3, 18.9, 14.9, 11.7, 6.1, 4.5, 3.7, 2.8
42 E. faecium (kb) 18.7, 14.8, 7.9, 5.9, 2.7
43 E. faecium (kb) 18.6, 14.7, 10.4, 6.1, 2.7
46 E. faecium (kb) 20.5, 17.4, 14.3, 5.8, 3.6, 2.6
48 E. faecium (kb) 20.6, 18.7, 17.5, 14.3
49 E. faecium (kb) 21.2, 17.9, 8.9, 5.0, 3.2
50 E. faecium (kb) 20.0, 16.8, 14.4, 2.6
52 E. faecium (kb) 20.0, 16.8, 14.4, 2.6
55 E. faecium (kb) 18.6, 17.1, 13.4, 7.7, 5.6, 3.5,2.4
56 E. faecium (kb) 16.3, 14.4, 9.1, 6.2, 2.4
57 E. faecium (kb) 13.7, 8.7, 5.3, 3.0, 2.2
59 E. faecium (kb) 13.8, 8.2, 5.5, 3.4
M Marker (bç) 16210, 14174, 12138, 10102, 8066, 7045, 6030, 5012, 3990, 2972, 2067
Supercolied ccc DNA marker (Sigma Chem. Co., USA, D 5292)
31 32 33 34 35 36 37 38 39 41 42 43 46 48 49 50 52 55 56 57 59 M

80
Şekil 4.3 Enterococcus faecium suşlarının plazmid içerikleri (devam). M; Süper sarmal ccc DNA Marker
M Marker (bç) 16210, 14174, 12138, 10102, 8066, 7045, 6030, 5012, 3990, 2972, 2067
63 E. faecium (kb) 21.4, 17.5, 15.7, 14.2, 11.3, 9.7, 6.5, 5.3
64 E. faecium (kb) 20.7, 18.3, 15.6, 11.4, 6.5, 5.3
65 E. faecium (kb) 21.1, 15.4, 10.0, 6.9, 5.3
67 E. faecium (kb) 21.2, 19.0, 14.8, 10.4, 9.6, 6.3
68 E. faecium (kb) 23.2, 17.6, 15.0, 11.4, 7.2, 6.3, 5.2
69 E. faecium (kb) 17.6, 14.9,12.1
70 E. faecium (kb) 24.5, 21.2, 17.6, 14.9,12.3, 5.1
71 E. faecium (kb) 21.2, 16.8, 14.8, 10.4, 4.9, 3.3
74 E. faecium (kb) 28.0, 21.2, 16.8, 14.8, 11.4, 8.2, 4.9, 3.3
75 E. faecium (kb) 35.8, 28.0, 21.2, 18.5, 14.7, 11.4, 4.2, 3.3,2.3
76 E. faecium (kb) 33.1, 27.1, 21.2, 4.2, 3.3,2.3
77 E. faecium (kb) 28.0, 26.5, 21.2, 16.8, 9.6, 5.5, 2.5
79 E. faecium (kb) 28.0, 26.1, 22.5, 16.6, 9.6, 5.5, 2.5
80 E. faecium (kb) 18.5, 13.4, 12.1, 5.8
81 E. faecium (kb) 28.0, 26.1, 19.0, 17.3, 13.7, 9.7, 6.4, 4.8
84 E. faecium (kb) 18.5, 13.4, 12.1, 5.8
87 E. faecium (kb) 24.5, 15.5, 13.4, 9.7, 7.2, 5.8
89 E. faecium (kb) 24.3,15.9, 13.5, 9.6, 7.8,4.7
90 E. faecium (kb) 18.5, 13.4, 12.1, 5.8
94 E. faecium (kb) 24.2, 15.8, 13.1, 5.7, 4.7
95 E. faecium (kb) 15.6, 13.1, 5.8
Supercolied ccc DNA marker (Sigma Chem. Co., USA, D 5292)
M 63 64 65 67 68 69 70 71 74 75 76 77 79 80 81 84 87 89 90 94 95

81
Şekil 4.3 Enterococcus faecium suşlarının plazmid içerikleri (devam). M; Süper sarmal ccc DNA Marker K; Standart suş E. faecalis ATCC 29212
M Marker (bç) 16210, 14174, 12138, 10102, 8066, 7045, 6030, 5012, 3990, 2972, 2067
K E. faecalis
ATCC29212(kb)
35.8, 30.4, 28.0
60 E. faecium (kb) 33.1, 30.4, 26.1, 12.6, 10.4, 9.6, 6.3
Supercolied ccc DNA marker (Sigma Chem. Co., USA, D 5292)
M K 60

82
4.3 Antibiyotik Direnç Genlerinin PZR ile Araştırılması
64 adet E. faecium suşunda, çoklu antibiyotik dirençlilik genlerinin varlığı, antibiyotik
dirençlilik genine özgü tasarlanan primerlerle gerçekleştirilen PZR çalışmaları ile
araştırılmıştır (Çizelge 3.3). Elde edilen PZR ürünleri ile dizi analizi gerçekleştirilmiş,
nükleotit dizileri NCBI Blast programı kullanılanarak karşılaştırılmıştır
(http://blast.ncbi.nlm.nih.gov/, 2012).
4.3.1 vanA ve vanB gen bölgeleri
Vankomisin direnç primerleri ile gerçekleştirilen PZR sonucunda vanA gen bölgesine
spesifik primerlerle 732 bç’lik gen bölgesi çoğaltılmıştır. Yüksek düzeyde vankomisine
direnci gösteren suşların hepsinin van A tipi direnç içerdiği belirlenmiştir. Fenotiple
uyumlu olarak tüm suşlar teikoplanine duyarlı bulunmuştur. Negatif kontrol olarak E.
faecalis ATCC 29212 kullanılmıştır. PZR ürünlerinin dizi analizleri sonucunda
çoğaltılan bölgelerin vankomisin direnç gen bölgesi olduğu doğrulanmıştır (Şekil 4.4).
Van A direnç fenotipine neden olan vanA geni kromozom veya plazmidler üzerinde
bulunabilir (Fisher ve Phillips 2009). Kazanılmış antibiyotik direnci genellikle
plazmidler üzerinde lokalize olur ve transpozonlar aracılığı ile aktarılır (Hanrahan vd.
2000) Çalışmamızda elde edilen bulgulara göre; 29 numaralı suş dışında tüm suşlarda
vanA genine kromozom üzerinde; 31, 67, 76 numaralı suşlar dışında geriye kalan 48
suşta hem kromozom hem de plazmid üzerinde rastlanmıştır. 29 numaralı suşun
vankomisine orta düzey dirençli olması Van B direncine işaret etmiştir.
Yapılan çalışmalarda glikopeptit dirençli E. faecium izolatlarının süratle arttığı
belirlenmiştir. Özellikle hastane enfeksiyonu olarak oranının artması ve stafilokoklar
gibi diğer türlere konjugasyon yoluyla aktarılabiliyor olması, vankomisin direncinin
gelecekte de önemli bir risk faktörü olduğunun göstergesidir. Vankomisin direncinin
öneminin artmasındaki nedenlerden biri de E. faecium’da yüksek derecede
indüklenebilir glikopeptit direnci gösteren Van A fenotipi içerikli Tn1546
transpozonunun tanımlanmasıdır (Leclercq vd. 1988).

83
Şekil 4.4 vanA gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.4 vanA gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.4 vanA gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M 1 2 3 4 5 6 8 11 12 13 15 17 19 21 24 25 26 28 K N M
732 bç
M 30 31 35 36 38 39 41 42 43 46 49 55 57 59 60 63 64 65 67 K N M
M 68 70 74 75 76 77 79 80 81 84 87 89 90 94 95 K N M
732 bç
732 bç

84
Fenotipik olarak vankomisine orta düzeyde direnç gösterip, teikoplanine duyarlı olan 29
numaralı suş Van B tipi direnç açısından incelenmiştir. Vankomisin direnç primerleri ile
gerçekleştirilen PZR sonucunda vanB gen bölgesine spesifik primerlerle 635 bç’lik
bölge çoğaltılmış ve sonuçlar DNA dizi analizi verileri ile doğrulanmıştır (Şekil 4.5).
Vankomisin dirençlilik determinantları kromozom üzerinde bulunmakla birlikte
konjugasyon yolu ile aktarılabilir özelliktedir (Hanrahan vd. 2000). Tn1547, Tn1549 ve
Tn5382 transpozonları vanB operonunu içerir. Türkiye’den bildirilen ilk Van B tipi
direnç 2012 yılında Ankara Numune Eğitim ve Araştırma Hastanesi’den bildirilmiştir
(Coşkun vd. 2012). Çalışmamızda tanımlanan idrar izolatı 29 numaralı suş, vanB direnç
geni içeren ikinci suş olması açısından önemlidir. Ayrıca Van B tipi direncin
saptanması, ülkemizde de glikopeptit direncinin geliştiğini göstermektedir. Ülkemizde
yaygın olmamakla birlikte Amerika ve diğer gelişmiş ülkelerde Van B tipi direnç
önemli hastane enfeksiyonu nedenleri arasındadır (Mcgregor ve Young 2000).
Şekil 4.5 vanB gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
635 bç
M 29 K N M

85
4.3.2 tetM ve tetL gen bölgeleri
Tetrasiklin direnci enterokoklarda konjugasyon yolu ile kazanılan direncin en tipik
örneğidir (Murray 1998). Enterokoklarda tetrasiklin grubu antibiyotiklere dirençten
sorumlu olan çok sayıda gen tanımlanmıştır. Bunlardan tetM geni Tn916 transpozonu
üzerinde taşınır ve en yaygın olarak bulunan direnç genidir (Aarestrup vd. 2000, Huys
vd. 2004). tetM, tetO, tetK ve tetS genleri tetrasiklinlerin ribozomlar üzerindeki etkisini
inhibe eder. tetL geni ise enterokokal bir plazmid üzerinde taşınır.
Tetrasiklin direnci enterokoklar arasında oldukça yaygındır. En sık bulunan gen tetM
genidir. Aarestrup vd. (2000) tarafından klinik, gıda ve hayvan kökenli örneklerden elde
edilen suşlarla yapılan çalışmada, 81 E. faecium izolatının 77’sinde (% 95) tetM geni,
% 16’sında tetL geni saptanmıştır. tetO ve tetK direnç genlerine ise rastlanmamıştır.
tetM geninin plazmid aracılı yatay gen transferi yolu ile aktarımının tanımlanması, bu
genin klinik önemini arttırmıştır (Poeta vd. 2005).
tetM ve tetL gen bölgelerine özgü primerler kullanılarak yürütülen PZR sonucunda,
tetrasikline dirençli 14 E. faecium suşunun kromozomu üzerinde tetM direnç geni
belirlenmiş ve sonuçlar DNA dizi analizi verileri ile doğrulanmıştır (Şekil 4.6). 74
numaralı suşta ise hem tetM hem de tetL direnç geni belirlenmiştir (Şekil 4.7). Her iki
direnç geninin aynı suş üzerinde bulunması litaratür verileri ile uyum göstermektedir
(Aarestrup vd. 2000).

86
Şekil 4.6 tetM gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.7 tetL gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M 3 5 24 28 42 43 57 64 71 74 75 77 79 94 K N M
475 bç
657 bç
M 74 K N M

87
4.3.3 ermB gen bölgesi
Makrolidlere direnç genellikle 23S rRNA’nın metilasyonundan sorumlu ermB geni ile
ilişkilidir ve metilasyon sonucu eritromisin ribozomlara bağlanamaz. ermB geni Tn917
transpozonu ile çeşitli plazmidler üzerinde taşınabilmektedir. Ancak kromozomal
lokasyonlu olanları da belirlenmiştir (Jensen vd. 1999, Wax vd. 2008).
Çalışmadaki ermB geni tüm suşlarda kromozomal DNA üzerinde pozitif olarak
bulunmuştur. Çoğaltılan 422 bç’lik gen bölgesi dizi analiz sonuçları, PZR ürününün
ermB gen bölgesine ait olduğunu doğrulamıştır (Şekil 4.8).
Şekil 4.8 ermB gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M K 1 2 3 4 5 6 7 8 11 12 13 15 17 19 21 24 25 26 N M
422 bç

88
Şekil 4.8 ermB gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.8 ermB gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
422 bç
M K 28 29 30 31 32 33 34 35 36 37 38 39 41 42 43 46 48 49 N M
M K 50 52 55 56 57 59 60 63 64 65 67 68 69 70 71 74 75 76 N M
422 bç

Şekil 4.8 ermB gen bölgesi29212, N; Negatif
4.3.4 rpoB1 ve rpoB2
rpoB1 ve rpoB2 gen bölgelerine spesifik primerlerle gerçekleştiri
sırasıyla 1000 bç ve 1094
ATCC 29212 suşu pozitif kontrol olarak kullanılmıştır. Dizi analizleri sonuçlarıyla PZR
ürünlerinin rpoB1 ve
kullanılan suşların tamamı
numaralı suşlar dışındaki diğer suşlarda
Rifampisinin E. faecium
olacağından tercih edilmez. Antibiyotik baskısının yarattığı mutasyonlar ile gerçekleşen
rifampisin dirençliliği tüm bakteri gruplarında görülmüştür. Klinik kökeni örneklerde
yüksek oranda rifampisin direnç genotipinin bulunması, kontrolsüz antibiyotik
kullanımına işaret etmektedir. Söz konusu mutasyonların bakterilerde oluşum ve
yayılma sıklığı yaln
taşımaktadır. Bu nedenle bu mutasyonların enterokoklarda taranması, patojen ilişkili
suşların tespiti açısından zorunludur (
M K 77
422 bç
89
gen bölgesi (devam). M; Marker, K; Kontrol suş 29212, N; Negatif kontrol (su)
rpoB2 gen bölgeleri
gen bölgelerine spesifik primerlerle gerçekleştiri
sırasıyla 1000 bç ve 1094 bç’lik gen bölgeleri çoğaltılmıştır (Şekil 4.9
ATCC 29212 suşu pozitif kontrol olarak kullanılmıştır. Dizi analizleri sonuçlarıyla PZR
ve rpoB2 gen bölgelerinden çoğaltıldığı doğrulanmıştır. Çalışmada
kullanılan suşların tamamı rpoB2 tipi direnç genini bulundururken, 43, 49, 50, 55,
numaralı suşlar dışındaki diğer suşlarda rpoB1 genine rastlanmıştır.
E. faecium klinik örneklerin tedavisinde kullanımı, dirençli suşlara neden
olacağından tercih edilmez. Antibiyotik baskısının yarattığı mutasyonlar ile gerçekleşen
sin dirençliliği tüm bakteri gruplarında görülmüştür. Klinik kökeni örneklerde
yüksek oranda rifampisin direnç genotipinin bulunması, kontrolsüz antibiyotik
kullanımına işaret etmektedir. Söz konusu mutasyonların bakterilerde oluşum ve
yayılma sıklığı yalnız klinik açıdan değil, bakteri evrimi açısından da önem
taşımaktadır. Bu nedenle bu mutasyonların enterokoklarda taranması, patojen ilişkili
suşların tespiti açısından zorunludur (Enne vd. 2004).
79 80 81 84 87 89 90 94 95 N M
. M; Marker, K; Kontrol suş E. faecalis ATCC
gen bölgelerine spesifik primerlerle gerçekleştirilen PZR sonucunda
l 4.9- 4.10). E. faecalis
ATCC 29212 suşu pozitif kontrol olarak kullanılmıştır. Dizi analizleri sonuçlarıyla PZR
gen bölgelerinden çoğaltıldığı doğrulanmıştır. Çalışmada
tipi direnç genini bulundururken, 43, 49, 50, 55, 56
genine rastlanmıştır.
klinik örneklerin tedavisinde kullanımı, dirençli suşlara neden
olacağından tercih edilmez. Antibiyotik baskısının yarattığı mutasyonlar ile gerçekleşen
sin dirençliliği tüm bakteri gruplarında görülmüştür. Klinik kökeni örneklerde
yüksek oranda rifampisin direnç genotipinin bulunması, kontrolsüz antibiyotik
kullanımına işaret etmektedir. Söz konusu mutasyonların bakterilerde oluşum ve
ız klinik açıdan değil, bakteri evrimi açısından da önem
taşımaktadır. Bu nedenle bu mutasyonların enterokoklarda taranması, patojen ilişkili
81 84 87 89 90 94 95 N M

90
Şekil 4.9 rpoB1 gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.9 rpoB1 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.9 rpoB1 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M K 1 2 3 4 5 6 8 11 12 13 15 17 19 21 24 25 26 27 N M
M K 28 29 30 31 32 33 34 35 36 37 38 39 41 42 46 48 52 57 N M
M K 59 60 63 64 65 67 68 69 70 71 74 75 76 77 79 80 81 84 N M
1000 bç
1000 bç
1000 bç

91
Şekil 4.9 rpoB1 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.10 rpoB2 gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.10 rpoB2 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M K 87 89 90 94 95 N M
M K 1 2 3 4 5 6 8 11 12 13 15 17 19 21 24 25 26 27 N M
M K 28 29 30 31 32 33 34 35 36 37 38 39 41 42 43 46 48 49 N M
1094 bç
1094 bç
1000 bç

92
Şekil 4.10 rpoB2 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.10 rpoB2 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
4.3.5 cat gen bölgesi
cat gen bölgesine spesifik primerlerle gerçekleştirilen PZR sonucunda 506 bç’lik gen
bölgesi çoğaltılmıştır. Negatif kontrol olarak E. faecalis ATCC 29212 kullanılmıştır
(Şekil 4.11). DNA dizi analizi sonuçları PZR ürünlerinin cat gen bölgesine ait olduğunu
doğrulamıştır.
Çalışmada kullanılan 5 numaralı suş dışında tüm suşlar kloramfenikole orta düzeyde
dirençli bulunmuştur. Klinik enterokok suşlarında kloramfenikol dirençlilik oldukça
yaygın bir özellik olarak belirlenmiştir. Enzimi kodlayan genler enterokoklarda
M K 77 79 80 81 84 87 89 90 94 95 N M
M K 50 52 55 56 57 59 60 63 64 65 67 68 69 70 71 74 75 76 N M
1094 bç
1094 bç

93
konjugatif ve konjugatif olmayan plazmidler ya da kromozom üzerinde bulunabilir.
Streptokokal plazmid pIP501 üzerinde bulunan kloramfenikol asetiltransferaz direnç
geni ile akraba olan gen dizisi, E. faecalis ve E. faecium izolatlarında bulunmuştur bu da
genin yatay gen transferi ile aktarılabildiğini göstermiştir (Aarestrup vd. 2000).
Şekil 4.11 cat gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.11 cat gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M K 3 4 5 6 12 13 24 26 57 59 60 N M
M K 64 74 75 76 77 79 80 81 87 90 95 N M
506 bç
506 bç

94
4.3.6 Yüksek düzey aminoglikozit direncinden sorumlu gen bölgeleri
Yüksek düzeyde aminoglikozit direnci (AME); gentamisin, streptomisin ve kanamisin
antibiyotiklerine yüksek düzeydeki direnci ifade eder. Enterokokların düşük düzeyde
doğal olarak aminoglikozitlere dirençli olması tedavide sinerjistik etki oluşturmak
amacıyla hücre duvarı sentezini inhibe eden bir antibiyotikle aminoglikozit grubu
antibiyotiğin kombine kullanımını gerektirir. Ancak plazmid ve transpozonlar aracılığı
ile kazanılmış direnç gelişimi nedeniyle yüksek düzey aminoglikozit direncinin ortaya
çıkışı tedavide aminoglikozitler ile kombine olarak hücre duvarı sentezini inhibe eden
ajanların kullanılması ile elde edilen sinerjistik etkiyi de ortadan kaldırmaktadır
(Shepard ve Gilmore 2002). Bu nedenle AME direncinin belirlenmesi önemlidir.
Çalışmamızda AME grubuna ait genlerin spesifik primerleriyle PZR yapılmıştır. PZR
ürünlerinin dizi analizleri sonucunda çoğaltılan bölgelerin aminoglikozit direnç gen
bölgeleri olduğu saptanmıştır. Yüksek düzey gentamisin direncinden sorumlu aac(6’)-
aph(2’’) gen bölgesinin spesifik primerlerle gerçekleştirilen PZR sonucunda 675 bç’lik
gen bölgesi çoğaltılmıştır. Negatif kontrol olarak E. faecalis ATCC 29212 suşu
kullanılmıştır (Şekil 4.12).
Şekil 4.12 aac(6’)-aph(2’’) gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M 2 3 4 5 8 12 13 17 24 25 26 27 29 30 32 33 34 35 K N M
675 bç

95
Şekil 4.12 aac(6’)-aph(2’’) gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.12 aac(6’)-aph(2’’) gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
ant6 gen bölgesine spesifik primerlerle gerçekleştirilen PZR sonucunda 596 bç’lik gen
bölgesi çoğaltılmıştır. Negatif kontrol olarak E. faecalis ATCC 29212 kullanılmıştır
(Şekil 4.13). Bu PZR ürünleri ile yürütülen dizi analizi sonucunda çoğaltılan bölgenin
ant6 gen bölgesi olduğu saptanmıştır.
Enterokoklarda streptomisin direnci diğer aminoglikozit grubu antibiyotiklerden farklı
olarak ribozomal mutasyonlar veya Adenil transferaz sentezlenmesi sonucu
oluşmaktadır.
M 37 39 41 42 43 48 49 50 52 55 56 57 59 60 64 65 67 69 K N M
M 71 74 76 77 79 81 94 K N M
675 bç
675 bç

96
Şekil 4.13 ant6 gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.13 ant6 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
Şekil 4.13 ant6 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M 31 32 33 34 35 36 37 38 39 42 43 46 48 50 52 55 56 57 K N M
M 60 63 64 65 67 69 71 74 75 76 77 79 80 81 84 87 89 90 K N M
M 1 2 3 4 5 8 11 12 13 17 21 24 25 26 27 28 29 30 K N M
596 bç
596 bç
596 bç

97
Şekil 4.13 ant6 gen bölgesi (devam). M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
aphA3 gen bölgesine spesifik primerlerle gerçekleştirilen PZR sonucunda 292 bç’lik
gen bölgesi tüm suşlarda çoğaltılmıştır. Çalışmada pozitif kontrol olarak E. faecalis
ATCC 29212 kullanılmıştır (Şekil 4.14). Dizi analizi sonuçları PZR verilerini
doğrulamıştır.
Şekil 4.14 aphA3 gen bölgesi. M; Marker, K; Kontrol suş E. faecalis ATCC 29212, N; Negatif kontrol (su)
M 94 95 K N M
596 bç
M K 1 2 3 4 5 6 8 11 12 13 15 17 19 21 24 25 26 27 N M
292 bç

Şekil 4.14 aphA3 gen bölge29212, N; Negatif kontrol (su)
Şekil 4.14 aphA3 gen bölge29212, N; Negatif kontrol (su)
Şekil 4.14 aphA3 gen bölge29212, N; Negatif kontrol (su)
M K 28 29 30 31 32 33 34 35 36 37 38 39 41 42 43 46 48 49 N M
M K 50 52 55 56 57 59 60 63 64 65 67 68 69 70 71 74 75 76 N M
M K 70 79
98
gen bölgesi (devam). M; Marker, K; Kontrol suş 29212, N; Negatif kontrol (su)
gen bölgesi (devam). M; Marker, K; Kontrol suş 29212, N; Negatif kontrol (su)
gen bölgesi (devam). M; Marker, K; Kontrol suş 29212, N; Negatif kontrol (su)
28 29 30 31 32 33 34 35 36 37 38 39 41 42 43 46 48 49 N M
50 52 55 56 57 59 60 63 64 65 67 68 69 70 71 74 75 76 N M
79 80 81 84 87 89 90 94 95
M; Marker, K; Kontrol suş E. faecalis ATCC
M; Marker, K; Kontrol suş E. faecalis ATCC
M; Marker, K; Kontrol suş E. faecalis ATCC
28 29 30 31 32 33 34 35 36 37 38 39 41 42 43 46 48 49 N M
50 52 55 56 57 59 60 63 64 65 67 68 69 70 71 74 75 76 N M
94 95 N M
292 bç
292 bç
292 bç

99
4.3.7 blaZ gen bölgesi
blaZ gen bölgesinin araştırılması için S. aureus NCTC 8325 suşu pozitif kontrol olarak
kullanılmıştır. Pozitif kontrol ile belirlenen 831 bç’lik bölge, PZR analizi sonucunda
tüm suşlar fenotipik olarak dirençli olmasına karşın direnç geni E. faecium suşlarında
saptanmamıştır. Enterokok türlerini de içeren Gram- pozitif koklarda plazmid kodlu
blaZ geninin genellikle β-laktamaz üretiminden sorumlu olmasına karşın, çalışmamızla
benzer olarak ulusal ve yurt dışı çalışmalarda blaZ geni tespit edilememiştir.
Stovcik vd. (2008), ampisilin dirençli izolatlarda PZR analizleri sonucunda β-laktamaz
genlerinin saptanamadığını ileri sürmüştür. Aynı zamanda Türkiye’de yapılan
çalışmalarda beta-laktam antibiyotiklere dirençli enterokokların giderek arttığına dikkat
çekilerek, bu artışta penisilinlere dirençli, beta-laktamaz üretmeyen suşların rol aldığı
gösterilmiştir (Gür 1997). Bu sebeple β-laktam direnç mekanizmalarının daha detaylı
olarak incelenmesi gerekmektedir.
4.4 Konjugasyon
Yatay gen transferi, mikrobiyal evrimin en önemli unsurlarından biri olarak kabul
edilmektedir. Yatay gen transferinin doğal mekanizmaları; konjugasyon,
transformasyon ve transdüksiyondur. Konjugasyon; bakteriyel genlerin, verici ve alıcı
hücrelerin fiziksel yüzey teması yolu ile oluşan konjugasyon köprülerinden tek yönlü
aktarımı olarak tanımlanmaktadır. Enterococcus türleri arasındaki genetik değişim
transdüksiyonla olsa da, en yaygın mekanizma konjugasyondur. Bunun yanında
enterokoklarda; kromozomlar veya plazmidler üzerinde bulunan, hareketli genetik
determinantlar olan transpozonlar ile yapılan gen atarımı da gösterilmiştir (Simjee ve
Gill 1997). VRE’lerde yüksek seviyede glikopeptit direnç fenotipi gösteren genler
çoğunlukla yüksek kopya sayısına ve yüksek stabiliteye sahip plazmidler ve plazmidlere
ilave olmuş transpozonlar üzerinde bulunurlar. Plazmid veya kromozomlara ilave olmuş
konjugatif transpozonlar aracılığıyla kazanılmış glikopeptit direnci de yine insersiyon
dizileri içeren transpozonun başka bir konukçu DNA’ya ilave olabilmesi ile kolaylıkla
aktarılabilir. Direnç genini içeren plazmidin stabilitesinin ve kopya sayısının yüksek

100
olması da direncin sürdürülebilme ve transfer edilebilme olasılığını sağlamlaştırır
(Clewell 1993, Dunny vd. 1995). Handwerger vd. (1990) nozokomiyal bir E. faecium
suşu ile yaptıkları çalışmada hem vankomisin direnci hem de beta hemoliz ile feromon
cevaptan sorumlu 55 kb’lık bir pHKK100 konjugatif plazmidi izole etmişlerdir.
Bozdoğan ve Leclercq (1999) E. faecium ile yaptıkları araştırmada, 60 kb’dan büyük
bir plazmid aracılı vankomisin ve streptogramin direnci saptanmışlardır.
Çalışmamızda E. faecium suşların konjugatif özelliklerinin belirlenmesi için antibiyotik
duyarlı suşlar alıcı, dirençli suşlar ise verici suş olarak kullanılarak filtre eşleşme
yöntemi ile konjugasyon denemeleri yürütülmüştür. Konjugasyon denemelerinde
tetrasiklin, vankomisin ve kloramfenikol antibiyotik direnç genlerinin aktarım
özelliklerine bakılmıştır. Çalışmamızda direncin aktarımının belirlenmesinde alıcı suşlar
gentamisin duyarlı olarak seçilmiştir. Eritromisin ve kanamisin antibiyotik direnç
genlerinin aktarılabilirliği literatürde bulunmuş ancak çalışma yaptığımız tüm suşların
bu antibiyotiklere dirençli olmasından dolayı konjugasyon denemelerine alınmamıştır
(Clewell 1993, Dunny vd. 1995). Sadece vankomisin direnç geninin aktarımına bakılan,
6 (vanAR/GenS) ve 32 (vanAS/GenR) numaralı suşlarla yapılan konjugasyon
denemesinde transkonjugant elde edilmiştir (Şekil 4.15). vanA genini içeren plazmidinin
konjugasyon sıklığı, verici hücre başına 18.9 x 10-3 olarak belirlenmiştir. 4.6 kb plazmid
alıcı suşta bulunmazken konjugant üzerinde bulunmuştur. Bu plazmidin vankomisin
direncini kodladığını doğrulamak için disk difüzyon testi yapılmış ve 37 oC de 24 saat
inkübasyon sonunda konjugantlar hem vankomisine hem de gentamisine dirençli olarak
bulunmuştur (Şekil 4.16).

101
Şekil 4.15 vanA geninin konjugal aktarımı M; Marker, 32; Alıcı suş, 6; Verici suş, 32/6; Konjugant
M Marker (bç) 16210, 14174, 12138, 10102, 8066, 7045, 6030, 5012, 3990, 2972, 2067
32 Alıcı suş 25.6, 21.9, 15.8, 5.6, 3.4
6 Verici suş 22.6, 16.2, 12.5, 9.9, 5.7, 4.6,3.3
32/6 Konjugant 25.6, 22.0, 15.8, 5.6, 4.6
Supercolied ccc DNA marker (Sigma Chem. Co., USA, D 5292)
Şekil 4.16 Transkonjugantta vankomisin direncinin disk difüzyon yöntemi ile doğrulanması
Vankomisin
Gentamisin
M 32 6 32/6 M
4.6 kb

102
Enterokokların diğer patojenlere nazaran glikopeptit direnç genlerinin konjugal
transferinde başarılı olmasının ve konjugasyon transfer sıklığının daha yüksek
olmasının pek çok sebebi mevcuttur. Konjugatif özellik içeren plazmidlerin
bulundukları konakçılardaki stabiliteleri, yüksek sıklıkta aktarım yetenekleri ile
doğrudan ilişkilidir. Bu nedenle tanımlanan plazmidlerde konjugatif yeteneğin
biyokimyasal ve genetik doğasının belirlenmesi de önem taşımaktadır (Clewell 1993).
Çalışmada elde edilen plazmidin büyüklüğünün litaratür bulgularına göre çok daha
düşük olması dikkat çekicidir. Bu nedenle söz konusu plazmidin DNA dizi analizinin
yapılması, evrimsel kökeni hakkında daha detaylı bilgi sunabilir.

103
5. SONUÇ
Çalışmamızda Akdeniz Üniversitesi Tıp Fakültesi Hastanesi’nde yatmakta olan
hastalardan sağlanan kültür koleksiyonundan elde edilen klinik kökenli 64 Enterococcus
faecium suşunda antibiyotik direnç determinantlarının genetik doğası ve bu özellikleri
aktarım yolları incelenmiştir. Bu suşların, çoklu ilaç fenotipi gelişimini indükleyen
determinantlar bakımından incelenmesi sonucu; kazanılan antibiyotik dirençliğine sebep
olan gen bölgelerini içerdikleri belirlenmiştir.
Elde edilen veriler; gelişen çoklu antibiyotik dirençliliği içeren bu suşların yol açacağı
enfeksiyonların tedavisinde antibiyotik kullanımının girerek ekinliğini yitirdiğine işaret
etmektedir. Özellikle yüksek oranlarda bulunan glikopeptit dirençi tedavinin son
basamağında kullanılan vankomisin gibi önemli antibiyotikleri devre dışı bırakacaktır.
Yüksek oranda bulduğumuz ampisilin ve yüksek düzey aminoglikozit direnci de bu iki
antibiyotik grubu ile oluşturulan kombinasyon tedavi olasılığını ortadan kaldırmaktadır.
1990’lı yıllardan bu yana yapılan çalışmalarda direnç yüzdelerinin giderek artmış
olması, doğru antibiyotik kullanım politikaları oluşturulmaması sonucunda sadece
varolan antibiyotiklere değil, yeni nesil ilaçlara da hızla direncin gelişebileceğinin
göstergesidir. Ayrıca çalışmamızda Van B tipi direncin tespit edilmiş olması sadece
direnç oranlarının artmadığının aynı zamanda şekillendiğinin de işaretidir.
Enterokoklarda direnç genlerinin aktarımı özellikle plazmid aracılığıyla olmaktadır.
Yapılan konjugasyon denemeleri sonucunda sadece vanA geninin aktarımı
gerçekleştirilmiş ve konjugasyon sıklığı çok yüksek bulunmuştur. Ancak özellikle
enterokoklarda konjugasyon yoluyla kazanılan direncin en tipik örneği olarak tetrasiklin
direncinin konjugasyonla aktarılamamış olması, yatay gen transferi mekanizmalarını
etkileyen koşulların doğru incelenmesi ve belirlenmesi gerektiğine işaret etmektedir.

104
KAYNAKLAR
Andrews, F. and Horder, T. 1906. A study of streptococci pathogenic for man. Lancet ii: pp. 708-713.
Aarestrup, F. M., Agerso, Y., Gerner– Smidt, P., Madsen, M., Jensen, L.B. 2000. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagnostic Microbiology and Infectious Disease, vol. 37, pp. 127–137.
Altay, F. M. 1996. Hastanede yatan hastaların dışkı kültürlerinde Enterokok kolonizasyonu ve vanomisin direncinin araştırılması. Ankara Eğitim Hastanesi (Uzmanlık tezi), s. 1-57.
Anderson, D. G. and McKay, L.L. 1983. Simple and rapid method for ısolating large plasmid dna from lactic streptococci. Applıed and Envıronmental Mıcrobıology, vol. 3, pp. 549-552.
Anderson, D. J., Murdoch, D.R., Sexton, D.J. 2004. Risk factors for infective endocarditis in patients with enterococcal bacteremia: a case-control study. Infection, vol. 32, pp. 72-77.
Anonymous, 2000. NCCLS. Performance Standarts for AntimicrobialSuspceptibility Testing; Tenth İnformational Supplement. January, M100-S10.
Anonymous, 2011. Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial susceptibility testing: 30th informational suplement. CLSI, M100-S21 Clinical and Laboratory Standards Institute, PA, USA, 940.
Aral, M., İsmihan, N., Paköz, E., Aral, İ., Doğan, S. 2011. Antibiotic resistance of Enterococcus faecalis and Enterococcus faecium strains isolated from various clinical samples. Türk Hijyen ve Deneysel Biyoloji Dergisi Cilt 68, (2); s. 85-92.
Arthur, M., and Courvalin, P. 1993. Genetics and mechanisms of glycopeptide resistance in Enterococci. Antimicrob Agents Chemotherapy, vol. 37, pp. 1593-1571.
Aygün, H., Memikoğlu, O.K., Tekeli, A., Azap, A., Yörük, F. 2008. Hastanede Yatan Riskli Hasta Gruplarında Vankomisine Dirençli Enterokok Kolonizasyonunun Sürveyansı. Türk Anest Rean Dergisi, (36); s. 168-173.
Başustaoğlu, A. ve Özyurt, M. 2000. Kan kültürlerinden izole edilen glikopeptit dirençli E. faecium. 3118-3120. Flora, 5, s. 142-7.
Başustaoğlu, A. ve Aydogan, H. 2002. Enterokoklar. Infeksiyon Hastalıkları Serisi. 5, s. 45-60.

105
Başustaoğlu, A. 2004. Enterokoklarda antibakteriyel direnç mekanizmaları ve direnç sorunu. Ed: Ulusoy S, Usluer G, Ünal S. Gram Pozitif Bakteri İnfeksiyonları. Bilimsel Tıp Yayınevi Ankara, s. 141-158.
Bozdoğan, B. and Leclercq, R. 1999. Plasmid-mediated coresistance to streptogramins and vancomycin in Enterococcus faecium HM103. Antimicrobial Agents Chemotherapy, 43, s. 2097-2098.
Braak, N., van Belkum, A., Kreft, D., Witt, R., Verbrugh, H. A., Endtz, H. 1999. The prevalence and clonal expansion of high-level gentamicin-resistant enterococci isolated from blood cultures in a Dutch university hospital. Journal of Antimicrobial Chemotherapy, vol. 44, pp. 795-798.
Campbell, R.C. 1974. Statistics for biologists. Cambridge University Pres, Second Edition, pp. 385.
Clewell, D.B. 1993. Bacterial sex pheromone-induced plazmid transfer. Cell, vol. 73; pp. 9-12
Clewell, D.B., An, F.Y., Flannagan, S.E., Antiporta, M. and Dunny, G., M. 2000. Enterococcal sex pheromone precursors are part of signal sequences for surface lipoproteins. Molecular Microbiology, vol. 35(1), pp. 246-247.
Clark, C. N., Cooksey, R.C., Hill, C. B., Swenson, M. J., Tenover, F. C. 1993. Characterization of glycopeptide resistant Enterococci from U.S hospitals. Antimicrobial Agents and Chemotherapy, November, pp. 2311-2317
Coleri, A., Cokmus, C., Ozcan, B., Akcelik, M. and Tukel, C. 2004. Determination of antibiotic resistance and resistance plasmids of clinical Enterococcus species. The Journal of General and Applied Microbiology, vol. 50, pp. 213–219.
Coşkun, A.F., Mumcuoğlu, İ., Aksu, N., Karahan, C.Z. vd. 2012. Phenotypic and Genotypic Traits of Vancomycin-Resistant Enterococci in a Public Hospital: The First vanB-Positive Enterococcus faecium Isolates Mikrobiyol Bul, 46(2); s. 276-282
Çelik, Ü., Alhan, E. 2008. Pediatrik Enfeksiyonlarda Zorlu Patojen: Enterokoklar. Çocuk Enfeksiyonları Dergisi, 2, s. 58-66.
Çetinkaya, Y., Falk, P., Mayhall, C.G. 2000. Vancomycin-Resistant Enterococci. Clinical Microbiology Reviews, vol. 13, pp. 686-707.
Çınar, T., Leblebicioglu, H., Eroglu, C., Sünbül, M. 1999. Enterokoklarda penisilin aminoglikozit sinerjizminin araştırılması. Klimik Dergisi, 12 (1); s. 39-42.
D’azevedo, P.A., Furtado, G.H.C. 2008. Molecular characterization of Vancomycin Resistant Enterococci strains eight years apart from its first isolation in Sao Paulo, Brazil. Revista do Instituto de Medicina tropical de Sao Paulo, vol. 50, pp. 195-198.

106
Del Campo, R., Ruiz-Garbajosa, P., Sa´nchez-Moreno, M., P., Baquero, F., Torres, C., Canto´ n, R., and Coque, T., M. 2003: Antimicrobial resistance in recent fecal enterococci from healthy volunteers and food handlers in Spain: genes and phenotypes. Microbial Drug Resistance, vol. 9, pp. 47–60.
Derbentli, Ş. 1998. Nozokomiyal enterokok infeksiyonları. Galenos, 23, s. 14-17
Devriese, L., Baele M., Butaye P. 2006. The Genus Enterococcus: Taxonomy. Prokaryotes, vol. 4, pp. 163-174.
Dunny, G.M, Leonard, B.A.B. and Heberg PJ. 1995. Pheromoneinducible conjugation in Enterococcus faecalis: interbacterial and host-parasite chemical communication. Journal Bacteriology, vol. 177, pp. 871-876.
Dutka-Malen, S., Evers, S.,Courvalın, P. 1995. Detection of Glycopeptide Resistance Genotypes and Identification to the Species Level of Clinically Relevant Enterococci by PCR: Journal of Clınıcal Mıcrobıology, January, pp. 24–27
Eaton, T., J., and Gassaon, M., J. 2001. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical ısolates. Applıed and Envıronmental Mıcrobıology, vol. 67, pp. 1628–1635.
Elder, J.K. and Southern, E.M. 1983. Measurement of DNA length by gel electrophoresis (II): comparison of methods for planting mobility of fragment length. Analytical Biochemistry, vol. 170, pp. 38–44.
Elder, J.K., Amos, A., Southern, E.M. and Shippey, G.A. 1983. Measurement of DNA length by electrophoresis (I). Analytical Biochemistry, vol. 128, pp. 223–226.
Enne, V. I., Delsol, A. A., Roe, J. M., Bennett, P. M. 2004. Rifampicin resistance and its fitness cost in Enterococcus faecium. Journal of Antimicrobial Chemotherapy, vol. 53, pp. 203–207.
Ergani-Ozcan, A. Naas, T., Ozhak Baysan, B., Ogunc,D., Inan, D., Colak, D and Nordmann, P. 2008. Nosocomial outbreak of vancomycin-resistant Enterococcus faecium in a paediatric unit at a Turkish university hospital Journal of Antimicrobial Chemotherapy, vol. 61, pp. 1033–1039
Esen, Ş., Sünbül, M., Şener, B., Eroğlu, C., Saniç, A., Leblebicioğlu, H. 2001. Glikopeptit, beta-laktam ve aminoglikozit grubu antibiyotiklerin enterokoklara in-vitro etkinliği. ANKEM Dergisi, 15(1); s. 59-63.
Facklam, R.R., Sahm, F.D 1995. Enterococcus. Manual of Clin. Microbiol. 6th edition. Washington D.C. American Society for Microbiology, pp. 308 – 14.
Faclam, R.R., Sahm, D.S and Teixera, L.M. 1997. Enterococci, Chapter 18, Manual of Clinical Microbiology, 7th Edition, ASM Press,Washington DC.

107
Facklam R.R., Teixeria L.M. 1998. Enterococcus. In: Collier L, Bolows A,Sussman (eds). Topley & Wilson’s Microbiology and Microbial infections. Vol 2 (Systematic Bacteriology). Ed: Edvard Arnold, 9th edition. London, pp. 669-682.
Facklam, R.R. 2002. What happened to the streptococci: overview of taxonomic and nomenclature changes. Clinical Microbiology Reviews, vol. 15, pp. 613-630.
Fisher, K. and Phillips, C. 2009. The ecology, epidemiology and virulence of Enterococcus. Microbiology, vol. 155, pp. 1749-1757.
French, G.N. 1998. Enterococci and Vankomycin resistance Clinic İnfection Disseases, vol. 27 (suppl 1); pp. 75-83
Gordon, S., Swenson, J.M., Hill, B.C. 1998. Antimicrobial susceptibilitiy patterns of common and unysyal species of enterococcus caming infections in the US. Journal of Clinic Microbiology, vol. 9, pp. 372-8
Guardabassi, L. and Dalsgaard, A. 2004. Occurence, structure, and mobility of TN1546-
like elements in environmental isolates of vancomycin-resistant enterococci. Applied and Environmental Microbiology, vol. 70, pp. 984-990.
Gültekin, M. 2004. Enterokoklar: Mikrobiyoloji, epidemiyoloji ve patogenez. Önemli
ve Sorunlu Gram Pozitif Bakteri İnfeksiyonları, Bilimsel Tıp Yayınevi, 121 s. 40.
Gür, D. 1997. Flora Betalaktamazlar. Ek, 2; s. 1.
Hacker, J. and Kaper, J. B. 2000. Pathogenicity islands and the evolution of microbes. Annual Review of Microbiology, vol. 54, pp. 641–679.
Handwerger, S., Pucci, M.J. and Kolokathis, A. 1990. Vancomycin resistance is encoded on a pheromone response plasmid in Enterococcus faecium 228. Antimicrob Agents Chemotherapy, vol. 34, pp. 358-360.
Handwerger, S., Perlman, D.C., Altarac, D. and McAuliffe, V. 1992. Concominant high-level vancomycin and penicillin resistance in clinical isolates of Enterococci. Clinical Infection Disseases, vol. 14, pp. 655-661.
Hanrahan, J., Hoyen, C. and Rice, L.B. 2000. Geographic distribution of a large mobile element that transfers ampicillin and vancomycin resistance between Enterococcus faecium strains. Antimicrobial Agents Chemotherapy, vol. 44, pp. 1349-1351.
Huycke, M. M., Spiegel, C. A., Gilmore, M. S. 1991. Bacteremia caused by hemolytic, high-level gentamicin-resistant Enterococcus faecalis. Antimicrobial Agents and Chemotherapy, vol. 35, pp. 1626–1634.

108
Huycke, M. M., Sahm, D., Gilmore, M. 1998. Multiple drug resistant enterococci: The nature of the problem and an agenda for the future. Emerging Infection Disseases, vol. 4, pp. 239-49.
Huys, G., D’Haene, K., Collard, J., M., Swings, J. 2004. Prevalence and molecular characterization of tetracycline resistance in Enterococcus ısolates from food. Applıed and Envıronmental Mıcrobıology, vol. 70, pp. 1555–1562.
İşleroğlu, H., Yıldırım, Z., Demirpençe, Y., Yıldırım, M. 2008. Enterokoklar: biyokimyasal, fizyolojik ve fonksiyonel özellikleri ile patojenitesi. Akademik Gıda, 6(3), s. 16-26.
Jensen, L. B., Frimodt-Möller, N., Aarestrup, F.M. 1999. Presence of erm gene classes in Gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiology Letters, vol. 170, pp. 151-158.
Jett, B. D., Huycke, M. M., Gilmore, M. S. 1994. Virulence of Enterococci. Clinical Microbiology Reviews, vol. 7, pp. 462-478.
Kandler, O., Schleifer, K.H., Dandl, R. 1968. Differantiation of Streptococcus fecalis Andrewes and Horder and Streptococcus faecium Orla-Jensen based on the Amino acid composition of their murein. Journal of Bacteriology, vol. 96, pp. 1935-1939.
Karadenizli, A., Kolaylı, F. 2002. Kocaeli Üniversitesi Tıp Fakültesi Hastanesinde izole edilen enterokok türleri ve antibiyotiklere duyarlılıkları. Türk Mikrobiyol Cem Dergisi, 32(3-4); s. 212-5.
Kaye, D. 1982. Enterococci. Biologic and epidemiologic characteristics and in vitro suspectibility. Archives of Internal Medicine, vol. 142, pp. 2006 – 9.
Klare, I., Konstabel, C., Badstubner, D., Werner, G., Witte, W. 2003. Occurrence and spread of antibiotic resistances in Enterococcus faecium. International Journal of Food Microbiology, vol. 88, pp. 269–290.
Klein, G. 2003. Taxonomye, cologya nd antibiotic resistance of enterococci from food and the gastrointestinal tract. International Journal of Food Microbiology, vol. 88, pp. 123-131.
Kobayashi, N., Alam, M., M., Nıshımoto, Y., Urasawa, S., Uehara, N. and Watanabe, N. 2001. Distribution of aminoglycoside resistance genes in recent clinical isolates of Enterococcus faecalis, Enterococcus faecium and Enterococcus
avium. Epidemiology and Infection, vol. 126, pp. 197-204.
Koneman, E. W., Allen, S. D., Janda, W. M. Screckenberger, P. C., Winn, W. C. 1992. The gram positive cocci part 2: Streptococci and streptococcus-like bacteria. Diag. Microbiol. 4th edition. Philadelphia, J. B. Lippincott Company, pp. 431 – 66

109
Koneman, E.W, Allen, S.D, Janda, W.M, Schreckenberger, P.C, Winn, W.C. Procop, G., Woods, G. 2005. Color Atlas and Textbook of Diagnostik Microbiology, (Sixth edition) Philadelphia Lippincott Co; pp. 700-711
Korten, V. 2002. Enterokoklar. In Willke TA, Söyletir G, Doganay M (eds). Infeksiyon Hastalıkları. İkinci baskı, İstanbul. Nobel Tip Kitabevleri Ltd. Sti, 2, s. 1497-1506.
Korten, V. 2005. Enterokok infeksiyonları. In: İliçin G, Ünal S, Biberoğlu K, Akalın S, Süleymanlar G (eds). Temel İç Hastalıkları. İkinci baskı. Güneş Kitabevi Ltd. Şti, Ankara.
Krieger, J. N., Kaiser, D. L., Wenzel, R. P. 1983. Urinary tract etiology of bloodstream infections in hospitalize patients. Jornal Infection Disseases, vol. 148, pp. 57 –62.
Kutlu, S.S., Dokuzoğuz, B. 2004. Enterokok infeksiyonlarında tedavi seçenekleri. Ed:
Ünal S,Vahapoğlu H. Yeni ve Yeniden Gündeme Gelen İnfeksiyonlar. Bilimsel Tıp Yayınevi. Ankara, s. 23-32.
Lampkowska, J., Feld, L., Monaghan, A., Toomey, N., Schjorring, S., Jacobsen, B., van der Voet, H., Andersan, S., R., Bolton, D., Aarts, H., Krogfelt, K., A., Wilcks, A., Bardowski, J. 2008. A standardized conjugation protocol to ases antibiotic resistance transfer between lactococcal species. International Journal of Food Microbiology, vol. 127, pp. 172-175.
Lavery, A., Rossney, A.S., Morrison, D., Power, A. and Keane, C.T. 1997. Incidence and detection of multi-drug-resistant Enterococci in Dublin hospitals. Journal Medical Microbiology, vol. 46, pp. 150-156.
Leavis, H., Top, J., Shankar, N., Borgen, K., Bonten, M., Embden, J.V., Willems, R.J.L. 2004. A Novel Putative Enterococcal Pathogenicity Island Linked to the esp Virulence Gene of Enterococcus faecium and Associated with Epidemicity. Journal of Bacteriology, vol. 186, pp. 672-682.
Leclercq, R., Derlot, E., Duval, J., Courvalin, P. 1988. Plasmid-mediated resistance to vancomycin and teicoplanin in Enterococcus faecium. New England Journal of Medicine, vol. 319, pp. 157-161.
Leclercq R., Dutka-Malen S., Brisson-Noel A., Molinas C., Derlot E., Arthur M., Duval J., Courvalin P. 1992. Resistance of enterococci to aminoglycosides and glycopeptides. Clinic Infection Disseases, vol. 15, pp. 495-501.
Leclercq, R., and Courvalin, P. 1997. Resistance to hglycopeptides in enterococci. Clinic Infection Disseases, vol. 24, pp. 545-554.
Lefort, A., Mainandi, J.L., Tod, M., Lortholory, O. 2000. Antienterococcal antibiotics. Med Clin North Ame, vol. 6, pp. 1471-1495.

110
Linden, P.K., Pasculle, A.W., Manez, R. 1996. Differences in outcomes for patients with bacteremia due to vancomycin-resistant E. faecium or vancomycin suspectible E. faecium. Clinic Infection Disseases, vol. 22, pp. 663 -70.
Macrina, F.L., Kopecko, D.J., Jones, K.R., Ayers, D.S. and McCoven, S.M. 1978. A multiple plasmid containing Escherichia coli strain: convenient source of size reference plasmid molecules. Plasmid, vol. 1, pp. 417–420.
Macrina, F.L., Tobian, J.A., Jones, K.R., Evans, R.P. and Clewell, D.B. 1982. A cloning
vector able to replicate in Escherichia coli and Streptococcus sangius. Gene, vol. 19, pp. 345–353.
Malathum, K., Murray, B. E. 1999. Vancomycin-resistant enterococci: recent advances
in genetics, epidemiology and therapeutic options drug resistance updates. vol. 2; pp. 224-43.
Markowitz, S. M., Wells, V. D., Williams, D. S., Stuart, C. G. 1991. Antimicrobial susceptibility and moleculer epidemiology of β-lactamase-producing aminoglycoside resistant isolates of Enterococcus faecalis. Antimicrob. Agents Chemother, 35 (6); pp. 1075- 80.
Marothi, Y.A., Agrihotri, H., Dubey, D. 2004. Enterococcal resistance –An overview. Indian Journal of Medical Microbiology, vol. 23, pp. 214-219.
Mascini, E. M., Troelstra, A., Beitsma, M. 2006. Genotyping and preemptive isolation to control an outbreak of vancomycin-resistant Enterococcus faecium. Clinical Infectious Diseases, vol. 42, pp. 739-46.
Mcgregor, K. F. and Young, H. K. 2000. Identification and characterization of vanB2 glycopeptide resistance elements in enterococci ısolated in Scotland. Antımıcrobıal Agents and Chemotherapy, vol. 44, pp. 2341–2348.
Mert-Dinç, B., Aykut-Arca, E., Yağcı, S., Karabiber, N. 2009. Çeşitli Klinik örneklerden izole edilen Enterococcus faecalis ve Enterococcus faecium suşlarında in-vitro antibiyotik duyarlılığı. Türk Hij Den Biyol Dergisi, 66 (3); s. 117-21.
Meyers, J.A., Sanches, D., Elwell, L.P. and Falkow, S. 1976. Simple agarose gel electrophoretic method for the identification and characterization of plasmid deoxyribonucleic acid. Journal of Bacteriology, vol. 127, pp. 1529-1537.
Moaddab, S., Töreci, K,. 2000. Enterokok suşlarında tür tayini, vankomisin ve diğer bazı antibiyotiklere direnç aranması. Türk Mikrobiyoloji Cemiyeti Dergisi, (30); s. 77-84.
Moellering R. C. 1992. Emergance of Enterococcus as a significant pathogen. Clin. Infect. Dissease, vol. 14, pp. 1173-78.
Moreno, M., Sarantinopoulos, P., Tsakalidou, E., De Vuyst, L. 2006. The role and application of enterococci in food and health. International Journal of Food Microbiology, vol. 106, pp. 1 – 24.

111
Mounir, M., Salem-Bekhit, M.I., Moussa, I., Muharram M. M. 2011. Increasing prevalence of high-level gentamicin resistant enterococci: An emerging clinical problem African Journal of Microbiology Research. 23 December, 5(31) ; pp. 5713-5720.
Mundy, L.M., Sahm, D.F. and Gilmore, M. 2000. Relationships between enterococcal virulence and antimicrobial resistance. Clinical Microbiology Reviews, vol. 13, pp. 513– 522.
Murray, B.E., Mederski–Samaroj, B. 1983. Transferable β-lactamase. A new mechanism for in vitro penicillin resistance in Streptococcus faecalis. Journal of Clinical Investigation, vol. 72, pp. 1168–1171.
Murray, B. E. 1990. The life and times of the Enterococcus. Clinical Microbiology Reviews, vol. 3, pp. 46–65.
Murray, B.E., Singh, K.V., Ross, R.P., Don Heath, J., Dunny, G.M., Weinstock, M.G. 1993. Generation of Restriction Map of Enterococcus faecalis OG1 and Investigation of Growth requirements of Growth requirements and regions encoding biosynthetic function. Journal of Bacteriology, vol. 175, pp. 5216-5223.
Murray, B.E. 1998. Enterococci. In: Gorbach, Bartlett, Blacklow (ed). Infectious Diseases Philadelphia. WB Saunders, pp.1723-1730.
Murray, B.E. 2000. Vancomycine-resistant Enterococcal infections. The New England J Medical, vol. 342, pp. 710-721.
Nourse, C., Murphy, H., Byrne, C. 1998. Control of a nosocomial outbreak of vancomycin resistant Enterococcus faecium in a paediatric oncology unit: risk factors for colonisation. European Journal of Pediatrics, vol. 157, pp. 20-7.
Patel, R. 1999. Vancomycine-resistant enterococci in solid organ transplantation. Curr Opin Organ Transplantation, vol. 4, pp. 271-280.
Perichon, B., Casadewall, B., Reynolds, P. and Courvalin, P. 2000. Glycopeptide Resistant Enterococcus faecium BM4416 Is a Van D-Type Strain with an Impaired D-Alanine:D-Alanine Ligase. Antimicrob. Agents Chemotherapy, vol. 44, pp. 1346-1348.
Poeta, P., Costa, D., Sa´enz, Y., Klibi, N., Ruiz-Larrea, F., Rodrigues, J., Torres, C. 2005. Characterization of antibiotic resistance genes and virulence factors in faecal enterococci of wild animals in Portugal. The Journal of Veterinary Medical, vol. 52, pp. 396–402.
Reynolds, P.E., Courvalin, P. 2005. Vancomycin Resistance in Enterococci Due to Synthesis of Precursors Terminating in D-Alanyl-D-Serine. Antımicrobıal Agents And Chemotherapy, vol. 49, pp. 21–25.

112
Rice, L. B. 2001. Emergence of vancomycin-resistant enterococci. Emerging Infectious Diseases. vol. 7, pp. 183-7
Robert, C., Moellering J. R. 2000. Enterococcus species, Streptococcus bovis and
Leuconostoc species. Principles and Practise of Infectious Disease 5th edition. Churchill – Livingstone, pp. 2147 – 53.
Robert C., Moellering R.C. 2005. Enterococcus species, Streptococcus bovis and Leuconostoc spesies. In: Mandell GL, Bennet JE, Dolin R (edc). Principles and Practice of Infectious Disease. Vol 2. Sixth edition. Elsevier Churchıll Livingstone, pp. 2411-2417.
Rosvoll, T., C., Pedersen, T., Sletvold, H., Johnsen, P., J., Sollid, J., E., Simonsen, G., S., Jensen, L., B., Nielsen, K., M., Sundsfjord, A. 2010. PCR-based plasmid typing in Enterococcus faecium strains reveals widely distributed pRE25-, pRUM-, pIP501- and pHTbeta-related replicons associated with glycopeptide resistance and stabilizing toxin-antitoxin systems. FEMS Immunology & Medical Microbiology, vol. 58, pp. 254-268.
Rudy, M., Zientara, M., Bek, T. 2005. Occurrence of antibiotic resistant enterococci in clinical specimens from a pediatric hospital. Pol J Microbiology, vol. 54, pp. 77-80.
Ruoff, K. L., Maza, L., Murtagh, M. S. 1990. Species identities of enterococci isolated from clinical specimens. Journal of Clinical Microbiology, vol. 28, pp. 435-7.
Sarantinopoulos, P., Andrighetto, C., Georgalaki, M. D., Rea, M. C., Lombardi, A., Cogan, T. M., Kalantzopoulos, G., Tsakalidou, E. 2001. Biochemical properties of enterococci relevant to their technological performance. International Dairy Journal, vol. 11, pp. 621-647.
Sayıner, H.S. 2008. Sürveyansla saptanan VRE’ lerin dağılımı, antibiyotik duyarlılıkları ve kolonize hastalarda risk faktörlerinin değerlendirilmesi. Okmeydanı Eğitim ve Araştırma Hastanesi (uzmanlık tezi ), s. 1-66
Schaffer, H.E. and Sederof, R.R. 1981. Improvement estimation of DNA fragment lengths from agarose gels. Analytical Biochemistry, vol. 115, pp. 122–133.
Schleifer, K. H. and R. Kilpper-Balz. 1984. Transfer of Streptococcus faecalis and Streptococcus faecium to the genus Enterococcus nom. rev. As Enterococcus faecalis comb. nov. and Enterococcus faecium comb. nov. nt. J.Syst. Bacteriology, vol. 34, pp. 31–34.
Shankar, N., Lockatell, C. V., Baghdayan, A. S., Drachenberg, C., Gilmore, M. S., Johnson, D. E. 2001. Role of Enterococcus faecalis surface protein Esp in the pathogenesis of ascending urinary tract infection. Infection and Immunity, vol. 69, pp. 4366-4372.

113
Shankar, N., Baghdayan, A. S., Gilmore, M. S. 2002. Modulation of virulence within a pathogenicity island in vancomycin resistant Enterococcus faecalis. Nature, vol. 417, pp. 746– 750.
Shepard, B.D., Gilmore, M.S. 2002. Antibiotic-resistant enterococci: the mechanisms and dynamics of drug introduction and resistance. Microbes and Infection, vol. 4, pp. 215–224.
Simjee, S., Gill, M.J. 1997. Gene transfer, gentamicin resistance and enterococci. Journal of Hospital Infection, vol. 36, pp. 249-259.
Son, R., Nimita, F., Rusul, G., Nasreldin, E., Samuel, L., and Nishibuchi, M. 1999. Isolation and molecular characterization of vancomycin-resistant in Enterococcus faecium Malaysia. Letters in Appl Microbiology, vol. 29, pp. 118-122.
Sood, S., Malhotra, M., Das-Arti Kapil, B.K. 2008. Enterococcal infections - antimicrobial resistance. Indian Journal of Medical Research, vol. 128, pp. 111-112.
Southern, E.M. 1979. Measurement of DNA lengths by gel electrophoresis. Analytical Biochemistry, vol. 100, pp. 319–323.
Stovcik, V., Javorsky, P., Pristas, P. 2008. Antıbıotıc resıstance patterns and resıstance genes in Enterococcı isolated from sheep gastroıntestınal tract in Slovakıa. Bulletin of the Veterinary Institue in Pulavy, vol. 52, pp. 53-57.
Şardan, Y.Ç. 2004. Enterokoklarda direnç sorunu. Ed: Şardan YÇ. Yeni ve Yeniden Gündeme Gelen İnfeksiyonlar. Bilimsel Tıp Yayınevi. Ankara, s. 10-16.
Şekercioğlu, A.O., Vural, T., Öğünç, D.,Çolak,D. ve Örgüt, G. 1998. Klinik örneklerden izole edilen Enterokok türlerinin identifikasyonu, antibiyotik duyarlılıkları, yüksek düzeyde gentamisin direnç özelliklerinin ve Beta laktamaz aktivitelerinin araştırılması. XXVII. Türk Mikrobiyoloji Kongresi, Antalya, Serbest bildiri,12-182.
Taşova, Y., İnal, A.S. 2004. Enterokok infeksiyonlarında klinik. E. faecium Yeni ve Yeniden Gündeme Gelen İnfeksiyonlar. Bilimsel Tıp Yayınevi, s. 17-22.
Tendolkar, P. M., Baghdayan, A. S., Shankar, N. 2003. Pathohenic enterococci: new developments in the 21st century. Cellular and Molecular Life Sciences, vol. 60, pp. 2622-2636.
Thiercelin, M. E. 1899. Sur un diplocoque saprophyte del’intestin susceptible de devenir pathogene. C. R. Seances Soc. Biol Fil. vol. 51, pp. 269-271.
Toledo-Arana, A., Valle, J., Solano, C., Arrizubieta, M. J., Cucarella, C., Lamata, M., Amorena, B., Leiva, J., Penadés, J. R., Lasa, I. 2001. The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Applied and Environmental Microbiology, vol. 67, pp. 4538–4545.

114
Trieu–Cuot, P., de Cespedes, G., Bentorcha, F., Delbos, F., Gaspar, E., & Horaud, T. 1993. Study of heterogeneity of chloramphenicol acetyltransferase (CAT) genes in streptococci and enterococci by polymerase chain reaction (PCR): characterization of a new CAT determinant. Antimicrobial Agents and Chemotherapy, vol. 37, pp. 2593–2598.
Upadhyaya, P.M.G., Ravikumar, K.L., Umapathy, B.L. 2009. Review of virulence factors of enterococcus: An emerging nosocomial pathogen. Indian Journal of Medical Microbiology, vol. 27, pp. 301-305.
Uttley, A.H.C., Collins, C.H., Naidoo, J. and George, R.C. 1988. Vancomycin resistant enterococci. Letter. Lancet; vol. 1, pp. 57-58.
Vural, T, Şekercioğlu, A.O., Öğünç, D., Gültekin, M., Çolak, D., Yeşilipek, A., Kocagöz, S., Ünal, S., Mutlu, G. 1998. Vankomisine dirençli Enterococcus
faecium suşu. ANKEM Dergisi, l2; s. 113.
Wax, R. G., Lewis, K., Salyers, A., Taber, H. 2008. Bacterial Resistance to Antimicrobials. Taylor and Francis Group, LLC.
Yetkin, M.A., Ateş, A.N. 2004. Vankomisine direncli enterokok bakteremisi: Olgu sunumu. Hastane İnfeksiyonları Dergisi, 8 (4); s. 310-4.
Yıldırım, M. 2007. Enterokoklar ve Enterokoklarla Gelisen Infeksiyonlar. Düzce Üniversitesi Tip Fakültesi Dergisi, (2); s. 46-52.
Yiş, R., Aslan, S., Çıtak, Ç., Değirmenci, S. 2010. Evaluation of Vancomycin-Resistant Enterococcus Colonization at Gaziantep Children’s Hospital, Turkey. Bu çalışma XXXIV. Türk Mikrobiyoloji Kongresi poster olarak sunulmuştur.
Zeana, C., Kubin, J.C., Della-Latta, P., Hammer, S.M. 2001. Vancomycin-resistant Enterococcus faecium meningitis successfully managed with linezolid: case report and review of the literature. Clinic İnfection Diseases, vol. 33, pp. 477-482.
Zırakzadeh, A., Patel, R. 2006. Vancomycin-Resistant Enterococci: Colonization, Infection, Detection, and Treatment. Mayo Clinic Proceedings, vol. 81, pp. 529-536.

115
ÖZGEÇMİŞ
Adı Soyadı: Kübra KASAROĞLU
Doğum Yeri: Ankara
Doğum Tarihi: 26.08.1983
Medeni Hali: Evli
Yabancı Dili: İngilizce
Eğitim Durumu (Kurum ve Yıl)
Lise : Ankara Cumhuriyet Lisesi (1998-2001)
Lisans : Ankara Üniversitesi Fen Fakültesi Biyoloji Bölümü (2001-2005)
Yüksek Lisans: Ankara Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı (2005-2007)
Çalıştığı Kurum/Kurumlar ve Yıl:
Akdeniz Üniversitesi Tıp Fakültesi Hastanesi Merkez Laboratuvarı (2007-)
YAYIN LİSTESİ
1. KASAROĞLU, K., AKÇELİK, N., ŞANLIBABA, P., AKÇELİK, M.
2012.Nisin üreticisi Lactococcus lactis subsp. Lactis LL27 suşunun beyaz
peynirde Listeria monocytogenes’in gelişiminin engellenmesi üzerine etkisi.
Selçuk Tarım ve Gıda Bilimleri Dergisi, 26(1); 77-83
2. OZHAK BAYSAN, B., KILINÇKAYA, H., ÖĞÜNÇ, D., ÖNGÜT, G.,
KASAROĞLU, K., GUNSEREN, F., ÇOLAK, D. 2013. Evaluation of
Brilliance VRE agar for detection of vancomycin-resistant enterococci (VRE) in
rectal swab specimens. Journal of Medical Microbiology, January, 25.

116
Sempozyum Tebliğleri
1. ŞİMŞEK, Ö., UYMAZ, B., AKKOÇ, N., YILDIRIM, K., AKÇELİK, M.
2007. L. lactis subsp. lactis LL27 Suşunda Nisin Üretiminin Artırılmasına
Yönelik Bir Genetik Yaklaşım. 15. Biyoteknoloji Kongresi (Ekim 2007,
Antalya) Bildiri Kitabı, 363-366.


















