
Does abiotic stress cause functional B vitamin deficiency? · ... D.R.M) and Food Science and Human...
46
1 Short title: B Vitamin Deficiency and Abiotic Stress 1 2 Corresponding author: Andrew D. Hanson, [email protected] Phone (352) 273-4856 3 4 Research Area: Topical Reviews (Associate Editor Julia Bailey-Serres) 5 6 Does abiotic stress cause functional B vitamin deficiency in plants? 7 8 Andrew D. Hanson, Guillaume A. Beaudoin, Donald R. McCarty, and Jesse F. Gregory III 9 10 Horticultural Sciences Department (A.D.H., G.A.B., D.R.M) and Food Science and Human Nutrition 11 Department (J.F.G.), University of Florida, Gainesville, FL 32611-0690, USA 12 13 14 One-sentence Summary: 15 Unlike animals, plants produce their own B vitamins but may nonetheless suffer vitamin deficiencies 16 under stress conditions, with wide-ranging metabolic consequences. 17 18 19 Author contributions: 20 A.D.H. conceived the article and wrote the manuscript along with G.A.B., D.R.M., and J.F.G. 21 22 Funding information: 23 This work was supported by National Science Foundation award IOS-1444202. 24 25 Corresponding author: Andrew D. Hanson [email protected] 26 Plant Physiology Preview. Published on November 2, 2016, as DOI:10.1104/pp.16.01371 Copyright 2016 by the American Society of Plant Biologists www.plantphysiol.org on August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
Transcript of Does abiotic stress cause functional B vitamin deficiency? · ... D.R.M) and Food Science and Human...

1
Short title: B Vitamin Deficiency and Abiotic Stress 1
2
Corresponding author: Andrew D. Hanson, [email protected] Phone (352) 273-4856 3
4
Research Area: Topical Reviews (Associate Editor Julia Bailey-Serres) 5
6
Does abiotic stress cause functional B vitamin deficiency in plants? 7 8
Andrew D. Hanson, Guillaume A. Beaudoin, Donald R. McCarty, and Jesse F. Gregory III 9
10
Horticultural Sciences Department (A.D.H., G.A.B., D.R.M) and Food Science and Human Nutrition 11
Department (J.F.G.), University of Florida, Gainesville, FL 32611-0690, USA 12
13
14
One-sentence Summary: 15
Unlike animals, plants produce their own B vitamins but may nonetheless suffer vitamin deficiencies 16
under stress conditions, with wide-ranging metabolic consequences. 17
18
19
Author contributions: 20
A.D.H. conceived the article and wrote the manuscript along with G.A.B., D.R.M., and J.F.G. 21
22
Funding information: 23
This work was supported by National Science Foundation award IOS-1444202. 24
25
Corresponding author: Andrew D. Hanson [email protected] 26
Plant Physiology Preview. Published on November 2, 2016, as DOI:10.1104/pp.16.01371
Copyright 2016 by the American Society of Plant Biologists
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

2
Abstract 27
B vitamins are the precursors of essential metabolic cofactors, but are prone to destruction under 28
stress conditions. It is therefore a priori reasonable that stressed plants suffer B vitamin deficiencies, 29
and that certain stress symptoms are metabolic knock-on effects of these deficiencies. Given the 30
logic of these arguments, and the existence of data to support them, it is a shock to realize that the 31
roles of B vitamins in plant abiotic stress have had minimal attention in the literature (100-fold less 32
than hormones) and continue to be overlooked. In this article, we therefore aim to explain the 33
connections among B vitamins, enzyme cofactors, and stress conditions in plants. We first outline 34
the chemistry and biochemistry of B vitamins, and explore the concept of vitamin deficiency with the 35
help of information from mammals. We then summarize classical and recent evidence for stress-36
induced vitamin deficiencies and for plant responses that counter these deficiencies. Lastly, we 37
consider potential implications for agriculture. 38 39 Key Words: Biotin, folate, niacin, pantothenate, pyridoxine, riboflavin, thiamin 40
41
INTRODUCTION 42
The great pioneers of modern plant physiology pointed out that B vitamins have much in common 43
with hormones, and even classified some vitamins as plant hormones (i.e. plant growth regulators in 44
today’s terms) (Went et al., 1938; Bonner and Bonner, 1948; Thimann, 1963). In plants, as in anim-45
als, B vitamins and hormones are biologically active in minute amounts, are transported, and lead to 46
similarly profound consequences when deficient (Bonner and Bonner, 1948). B vitamins and 47
hormones might consequently be expected to have received comparable research attention. This 48
has broadly been the case in biomedical research since vitamins and hormones were first 49
discovered (Kohler, 1975). It has almost never been the case in any area of plant research, including 50
abiotic stress. Thus, in the Arabidopsis abiotic stress literature, articles involving hormones 51
outnumber those involving B vitamins by a factor of 100 (Supplemental Table S1). 52 53 This stunning disparity suggests two things about past and present thinking in the abiotic stress field. 54
First, it indicates a prevalent default assumption that the whole of B vitamin metabolism always and 55
everywhere continues to work well in stressed plants – and so can be safely ignored. Second, and 56
relatedly, it signals a lack of attention to the possibility that stress-induced defects in B vitamin meta-57
bolism can be intermediate causes of system-wide plant stress responses. Neither the default ass-58
umption nor the inattention is reasonable a priori and they are not justified by the available evidence. 59 60 This review sets out to show that B vitamin deficiency is a simple, probable, but overlooked scenario 61
in abiotic stress responses. We begin by providing key background information on the chemical and 62
metabolic lability of B vitamins and the cofactors derived from them, and on the concept of vitamin 63
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

3
deficiency derived from work in animals. We then revisit classical studies that point clearly to roles 64
for B vitamins in abiotic stress, but are no longer in the modern canon, and assess recent evidence 65
for such roles. Next, we outline how plants combat stress-induced B vitamin deficiencies. Lastly, we 66
consider how understanding stress-induced vitamin deficiency could inform crop breeding and 67
management. 68 69 Two points should be made at the outset. First, there has been much research on the B vitamin con-70
tents of food plants and on B vitamin synthesis and metabolism in plants. However, the main – if not 71
sole – driver for this valuable work has been plants as vitamin sources for humans rather than plants 72
as vitamin sources for themselves (Fitzpatrick et al., 2012; Gerdes et al., 2012). This article takes 73
the complementary position that ‘plants need their vitamins too’ (Smith et al., 2007). Second, re-74
search on vitamin C (ascorbate) shows that plant stress biology does not invariably sideline vitamins 75
(Foyer and Noctor, 2011). This article argues that B vitamins deserve as much attention as vitamin 76
C. 77
78
CHEMICAL AND METABOLIC LABILITY – WHY BAD THINGS HAPPEN TO GOOD VITAMINS 79
B vitamins are indispensable because they are the metabolic precursors of essential cofactors. Fig-80
ure 1 shows the seven B vitamins found in plants, along with the cofactors to which they are 81
converted. In brief: Thiamin (vitamin B1) is converted to thiamin diphosphate; riboflavin (vitamin B2) 82
to flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD); niacin (vitamin B3) to 83
NAD(P)+; pantothenate (vitamin B5) to coenzyme A and the prosthetic group of acyl carrier protein; 84
pyridoxine (vitamin B6) to pyridoxal 5'-phosphate; biotin (vitamin B8) to the biotinyl side-chain of 85
enzymes; and tetrahydrofolate (historically termed vitamin B9) to various one-carbon substituted 86
folates and their polyglutamylated derivatives. 87 88 Cofactors function by participating in biochemical reactions, e.g. thiamin diphosphate forms covalent 89
complexes with carbonyl substrates, FAD and NAD(P)+ become reduced as substrates are oxidized, 90
and coenzyme A forms thioesters with acyl groups. Cofactors and their vitamin precursors are thus 91
chemically reactive by nature. Their chemically reactive groups are therefore prone to undergo spon-92
taneous side-reactions such as oxidation, hydrolysis, racemization, or addition that damage or des-93
troy the molecule – and stress conditions promote such side-reactions (Piedrafita et al., 2015). For 94
example, most abiotic stresses cause accumulation of reactive oxygen species (You and Chan, 95
2015) that can inflict oxidative damage on every compound in Figure 1. Similarly, temperature extr-96
emes, high light levels, pH excursions, and stress-driven accumulations of metabolites with which 97
cofactors react all directly accelerate diverse types of spontaneous cofactor damage (e.g. Treadwell 98
and Metzler, 1972; Baggott, 2000; Mills et al., 2006; Marbaix et al., 2011). Abiotic stresses can also 99
promote cofactor damage indirectly by altering compartmentation (e.g. Akhtar et al., 2010; Moham-100
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

4
madi et al., 2012), by inducing enzymes that break down cofactors (e.g. Rapala-Kozik et al., 2008; 101
Higa et al., 2012), and by creating harsh cellular conditions in which enzymes that normally act on 102
other substrates become more promiscuous (Piedrafita et al., 2015) and mistakenly attack cofactors. 103 104 Figure 1 uses color-coded arrows to show the site and nature of damage reactions that vitamins and 105
cofactors can undergo in physiological conditions, and Table I catalogs the reactions corresponding 106
to the arrows. Note that Figure 1 is bristling with arrows – which helps explain why most of the B 107
vitamins/cofactors made it into a ‘Top 30’ list of damage-prone metabolites (Lerma-Ortiz et al., 2016) 108
and why all organisms have dedicated systems to deal with vitamin/cofactor damage (Linster et al, 109
2013). “Bad things happen” to B vitamins and cofactors because these compounds are basically 110
chemical and metabolic “accidents waiting to happen”. Also, unlike many other reactive metabolites, 111
cofactors are end-products, not short-lived intermediates that are quickly converted to something 112
else. The longer an end-product lives, the higher the probability of its getting damaged at some point. 113 114 Thiamin diphosphate deserves special mention for metabolic lability because, unlike other cofactors, 115
it can be damaged during the catalytic cycle (‘catalysis-induced inactivation’) (McCourt et al., 2006). 116
Oxygen-dependent side-reactions in enzyme active sites convert the thiamin diphosphate cofactor to 117
an inactive thiazolone derivative (Sümegi and Alkonyi, 1983; Bunik et al., 2007). Such use-depend-118
ent damage explains why thiamin is the only B vitamin whose dietary requirement in animals is pro-119
portional to non-fat energy intake (McCourt et al., 2006). More generally, the damage reactions un-120
dergone by all B vitamins account for their dietary requirements; if vitamins were not continuously 121
damaged they would last forever and not need to be constantly resupplied. In this context it is note-122
worthy that the whole-body half-life of thiamin (and its phosphates) in humans is one to two orders of 123
magnitude shorter than those of other B vitamins for which estimates are available, as follows: thia-124
min, 9.5–18.5 days (Ariaey-Nejad et al., 1970); folates, 60–150 days (Gregory and Quinlivan, 2002); 125
vitamin B6, ~500 days (Coburn, 1990); vitamin B12 (absent from plants), 480–1284 days (Hall, 1964). 126
127
WHAT ACTUALLY CONSTITUTES B VITAMIN DEFICIENCY? 128
The concept of vitamin deficiency comes from human and animal nutrition; it refers to the consequ-129
ences of a shortfall in the dietary supply of a specific vitamin. Because plants make their own B 130
vitamins, and indeed are the ultimate source of most of the B vitamins consumed by animals, the 131
concept of ‘B vitamin deficiency’ might seem inapplicable to plants, and hence irrelevant. But, as we 132
will show, this concept is very relevant to plants. 133 134 Key questions about B vitamin nutrition in animals are: ‘What dietary intake of each vitamin is 135
needed for optimal growth and health?’ and ‘What happens when the supply of a vitamin falls below 136
the optimal level?’ Both questions are often answered by measuring weight gain in young animals 137
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

5
given various vitamin doses in the diet, and by tracking levels of the vitamin and its corresponding 138
cofactor in tissues and organs. Figure 2 (left half) summarizes data from such studies, in which 139
various levels of thiamin, pyridoxine, or folate were supplied to rats whose growth and liver cofactor 140
levels were monitored. Two points that emerge from these data are: (i) There is a continuum rather 141
than a sharp divide between vitamin sufficiency and deficiency; and (ii) the cofactor level in liver 142
does not have to fall much before growth is substantially impacted (as are development, metabolic 143
functions, and behavior, not shown), and declines of 30–60% from optimal are devastating. Animals 144
therefore operate their vitamin and cofactor systems with rather narrow margins of safety. 145 146 The same appears to be true of plants. Nutritional trials analogous to those above have not yet been 147
conducted using totally vitamin-deficient mutants. However, mutant plants partially deficient in thi-148
amin or vitamin B6 have been studied (Woodward et al., 2010; Rueschhoff et al., 2013), as have cult-149
ured cells depleted in folate using a reasonably specific antifolate drug (Loizeau et al., 2008) (Fig. 2, 150
right half). Although these experiments produced a single deficient state, not a range as in rats, the 151
data clearly suggest that a 25–40% loss of cofactor leads to severe consequences. To summarize: 152
The red zone on the B vitamin fuel gauge seems to be in roughly the same place in plants and 153
animals, and is quite close to the full mark. 154 155 A subsidiary concept within vitamin deficiency is ‘functional vitamin deficiency’, wherein vitamin and 156
cofactor measurements need not show marked depletion but strong metabolic disturbances never-157
theless ensue. A prime example is vitamin B12 (absent from plants), whose deficiency in humans is 158
better diagnosed by its metabolic consequences (elevated levels of methylmalonic acid and homo-159
cysteine) than by measuring B12 itself (Stabler et al., 1996). Another example is folate, for which – in 160
contrast to B12 – deficiency causes elevation only of homocysteine and not of methylmalonic acid 161
(Green, 2008). Functional deficiencies of other B vitamins in humans generally affect enzymes hav-162
ing the weakest affinity for their cofactor relative to the cofactor’s intracellular concentration, as is the 163
case for vitamin B6 (Ueland et al., 2015). However, the hierarchy of biochemical reactions most sus-164
ceptible to deficiency of B vitamins has not been fully clarified. 165 166 Functional vitamin deficiencies also can arise when the deficiency occurs in a particular (inaccess-167
ible) tissue or compartment but does not affect the whole organism – or at least the part convenient 168
for assessing vitamin or cofactor status. Such a situation could develop in roots for thiamin and 169
niacin because roots of certain species import these vitamins from shoots (Bonner and Bonner, 170
1948). Intracellular deficiencies are also possible given that B vitamins and cofactors are synthes-171
ized and used in different compartments. For example, thiamin diphosphate is made in the cytosol 172
but used mainly in plastids and mitochondria (Rapala-Kozik et al., 2012), and vitamin B6 is made in 173
the cytosol but used throughout the cell (Fitzpatrick et al., 2011). In this connection, note that the 174
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

6
plastids of the vitamin B6 deficient Arabidopsis mutant in Figure 2 were far more severely deficient in 175
B6 than the leaf as a whole (Rueschhoff et al., 2013), and that a thiamin-requiring tobacco mutant 176
appeared to suffer primarily from thiamin deficiency in chloroplasts (McHale et al., 1988). 177
178
EVIDENCE FOR STRESS-INDUCED B VITAMIN DEFICIENCY 179
The previous sections explained that B vitamins and cofactors are chemically and metabolically un-180
stable, that stresses potentially make them even more unstable, and that modest falls in cofactor 181
levels slow growth. We might therefore predict that (i) abiotic stresses cause vitamin and cofactor 182
deficiencies, (ii) the deficiencies degrade plant performance, and (iii) supplementing stressed plants 183
with the deficient vitamin(s) improves performance. Evidence from both classical and modern work 184
indicates that all these things happen. 185
186
Classical Evidence 187
The classical evidence was considered in a short 1957 article with the evocative title ‘The chemical 188
cure of climatic lesions’ (Bonner, 1957). The idea captured in the title was that negative effects of 189
‘climatic lesions’ (i.e. physicochemical environmental stresses) could be ‘cured’ by applying vitamins 190
or other essential metabolites, singly or in mixtures. ‘Chemical cures’ by B vitamins were reported for 191
various plants and various abiotic stresses during the 1940s–1960s; typical data are shown in Figure 192
3. In each case, applying a vitamin or vitamin mixture promoted growth in unfavorable conditions but 193
not in favorable ones. These results are de facto confirmation that stress can lead to functional B 194
vitamin deficiency. Importantly, the Arabidopsis work in Figure 3 gave stress-induced vitamin defic-195
iency a genetic basis by defining ecotypic differences attributable to one or a few genes (Langridge 196
and Griffing, 1959). This is the pattern expected if the enzymes that use a particular cofactor bind to 197
that cofactor with different strengths. When the vitamin starts to run out and cofactor levels fall, the 198
weakest binder loses activity first, and small allelic differences in the affinity of this enzyme for the 199
cofactor become critical determinants of performance of the organism as a whole (Guenther et al., 200
1999). 201
202
Modern Evidence 203
Although the exogenous application approach (‘spray and pray’) has fallen out of fashion, it can be 204
perfectly valid, and beneficial effects of applying B vitamins – particularly thiamin – to stressed crop 205
species continue to be reported, e.g. for salinized wheat, sunflower, and maize (Al-Hakimi and 206
Hamada, 2001; Sayed and Gadallah, 2002; Tuna et al., 2013; Kaya et al., 2015). Also, a rigorous 207
Arabidopsis study showed that thiamin enhances tolerance to oxidative stress imposed by paraquat 208
(Tunc-Ozdemir et al., 2009). Such protective effects of thiamin on stressed plants are generally 209
interpreted in terms of the antioxidant properties of thiamin and its diphosphate (Lukienko et al., 210
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

7
2000), but the in-vivo relevance of these properties has been questioned (Lesgards et al. 2015) and 211
it is not clear that the observed protection is due to direct antioxidant effects (Asensi-Fabado and 212
Munné-Bosch, 2010). The experimental data are equally compatible with the protection being due to 213
relief of a functional deficiency of thiamin diphosphate and the consequent restoration of normal 214
metabolic fluxes. Although bias from prior positive reports may be at work, it is interesting that thia-215
min features so prominently in the literature on vitamin application, given that thiamin and thiamin di-216
phosphate are particularly labile in vivo (see above). It is also interesting in the light of the extremely 217
high energetic cost and inefficiency of thiamin biosynthesis (Box 1), to which we will return later. 218 219 Further modern support for stress-induced deficiency of four different B vitamins has come from mol-220
ecular genetics, as follows. (i) Pyridoxine: The Arabidopsis SOS4 (SALT OVERLY SENSITIVE 4) 221
gene for pyridoxal kinase was cloned via the salt-sensitive phenotype of sos4 mutants, which was 222
reverted by pyridoxine (Shi et al., 2002) – a textbook ‘cure’ of a stress lesion. Arabidopsis mutants in 223
the PDX1.3 gene for pyridoxal 5'-phosphate synthase were found to be hypersensitive to salt, 224
osmotic, and oxidative stress (Chen and Xiong, 2005; Titiz et al., 2006). (ii) Pantothenate/coenzyme 225
A: The Arabidopsis HAL3A (HALOTOLERANCE DETERMINANT 3A) gene, first identified for its relat-226
ion to stress tolerance (Espinosa-Ruiz et al., 1999), was shown to code for a key enzyme in panto-227
thenate conversion to coenzyme A (Kupke et al., 2001), and HAL3A overexpression increased salt 228
tolerance (Espinosa-Ruiz et al., 1999; Yonamine et al., 2001). (iii) Folate: Ablation of the Arabidopsis 229
gene encoding a cytosolic folate synthesis enzyme (HPPK/DHPS) specifically impacted salt and 230
oxidative stress-resistance at germination (Storozhenko et al., 2007; Navarrete et al., 2012). (iv) 231
Riboflavin: The Arabidopsis phs1 (PHOTOSENSITIVE 1) mutant, first identified by its sensitivity to 232
high-light stress (Ouyang et al., 2010), was found to lack a functional domain of the riboflavin 233
biosynthesis enzyme PyrR (Hasnain et al., 2013; Frelin et al., 2015). Note the pattern here: Either a 234
gene identified via a stress-sensitive phenotype turned out to be a B vitamin or cofactor synthesis 235
enzyme, or ablating a known B vitamin/cofactor enzyme caused stress sensitivity. 236
237
COUNTERING STRESS-INDUCED B VITAMIN DEFICIENCY 238
Conceptually, resource deficiencies can be countered by (1) maintaining and then mobilizing re-239
serves, (2) obtaining more of the resource, (3) repairing or recycling it, or (4) using less of it. Plants 240
certainly deploy strategies 2 and 3 to confront B vitamin deficiency, and potentially deploy the others. 241 242 Strategy 1: Storing and Mobilizing B Vitamins and Precursors 243
Storage forms of B vitamins have had little attention in the plant literature, in sharp contrast to stor-244
age forms of hormones (Ludwig-Müller, 2011; Piotrowska and Bajguz, 2011). Storage forms of vit-245
amins and vitamin precursors – mainly glucosides and glucose esters – nevertheless exist and some 246
can reach higher concentrations than the corresponding free forms (Gregory, 1998). The only B vita-247
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

8
min for which no storage forms have been reported is thiamin. Storage forms have not been analyz-248
ed in abiotic stress studies, perhaps because they are not commercially available as standards. Un-249
fortunately, this means that the contribution of storage forms to B vitamin homeostasis during stress 250
remains unresolved; this contribution could, however, be large. The main storage forms are shown in 251
Figure 4 and described below, starting with the two vitamins (B6 and B3) whose storage forms are 252
reported to be more abundant than the free forms in certain species and tissues. 253 254 Pyridoxine (B6). The 5'-β-D-glucoside of pyridoxine, and other glycosides linked via the 4- or 5-hydr-255
oxymethyl groups, can contribute up to 75% of the total vitamin B6 in plant tissues (Gregory, 1998), 256
i.e. they can be the largest item in the B6 budget. A glucosyltransferase responsible for pyridoxine-5'-257
β-D-glucoside synthesis has been detected in pea (Tadera et al., 1982); enzymatic hydrolysis of the 258
glucoside has been shown in vitro (Yasumoto et al., 1976) so it can presumably be mobilized in vivo. 259 260 Niacin (B3). Nicotinic acid-N-β-D-glucoside and N-methylnicotinate (trigonelline) are widespread in 261
plants and can be major metabolites of nicotinamide and nicotinate (Matsui et al., 2007; Ashihara et 262
al., 2010); more complex glycosides also occur (Gregory, 1998). Enzymes that would allow the N-263
glucoside and trigonelline to act as mobilizable storage forms have been characterized (Upmeier et 264
al., 1988; Shimizu and Mazzafera, 2000; Mizuno et al., 2014). Brassicaceae contain the β-D-glucosyl 265
ester of nicotinic acid, which almost certainly acts as a mobilizable storage form (Li et al., 2015). 266 267 Pantothenate (B5). Pantothenate 4'-O-β-D-glucoside has been isolated from tomato fruit (Amachi et 268
al., 1971) and is most likely present in a wide range of species and tissues (Yoshizumi and Amachi, 269
1969). Glycosides of the lactone form of pantoate, the immediate precursor of pantothenate (pant-270
oyllactone-β-D-glucopyranoside and pantoyllactone primeveroside) occur in rice seedlings, the lev-271
els in coleoptile tissue being in the millimolar range (Menegus et al., 2002). These pantoate derivat-272
ives seem not to have been sought in any plant besides rice; they could conceivably be widespread. 273 274 Tetrahydrofolate Precursors. The precursor p-aminobenzoate was converted to its β-D-glucosyl 275
ester by all tissues and species tested and was found to be the major endogenous form of p-amino-276
benzoate (Quinlivan et al., 2003). The esterification reaction is reversible and the ester is stored in 277
vacuoles (Eudes et al., 2008). Glycosides (probably β-D-glucosides) of the tetrahydrofolate precurs-278
ors neopterin and monapterin were found in tomato fruit engineered to overproduce pterins (Díaz de 279
la Garza et al., 2004); it is not known whether such glycosides occur naturally, or whether they can 280
be mobilized. 281 282 Riboflavin (B2). The α and β forms of riboflavin-5′-D-glucoside have been found in germinating barl-283
ey seedlings supplied with riboflavin (Suzuki and Uchida, 1983). As with the pterin glycosides above, 284
it is it is not known whether riboflavin glucosides occur naturally, or whether they can be mobilized. 285 286
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

9
Biotin (B7). While no small-molecule conjugates of biotin have been reported, a biotinyl protein that 287
represents >90% of the total protein-bound biotin has been characterized and cloned from pea 288
seeds (Dehaye et al., 1997), and is conserved among higher plants (Gerdes et al., 2012). 289 290 Strategy 2: Increasing B Vitamin Supply 291
Many analyses of gene and protein expression have indicated that abiotic stresses upregulate flux 292
through some but not all B vitamin biosynthesis pathways and studies of the effects of stress on the 293
levels of vitamins and cofactors tend to support this inference. As is usual in stress research, 294
differences in experimental design (the species and tissue used, the timing, duration, and severity of 295
stress, the analysis methods applied – blots/microarrays/RNAseq, etc) preclude meta-analysis of all 296
the data. Below, we therefore focus first on a high-throughput dataset – the AtGenExpress global 297
stress expression dataset (available at http://jsp.weigelworld.org/expviz/expviz.jsp) – supplemented 298
with broadly comparable data from several independent Arabidopsis stress studies (Supplemental 299
Table S2). We then highlight illustrative papers on pyridoxine and thiamin. 300 301 Note that even when a vitamin biosynthetic pathway is clearly upregulated in response to stress, 302
stress-induced vitamin or cofactor deficiency may still occur for several reasons. Firstly, activating 303
biosynthetic gene expression may fail to increase vitamin production due to downstream constraints, 304
e.g. the energy cost of making THI4 protein for thiamin biosynthesis (see below and Box 1). Sec-305
ondly, as noted above, a high vitamin or cofactor status – as measured in whole organs, tissues, or 306
cells – can obscure functional deficiencies within sub-compartments. Lastly, conversion of vitamins 307
to cofactors may become limiting, so that increases in vitamin levels do not always result in higher 308
cofactor levels. Cofactor levels can consequently fall even when free vitamin levels rise – and cofact-309
or levels – not vitamin levels – determine metabolic outcomes. 310 311 Arabidopsis Gene Expression. The stresses used for the AtGenExpress global stress expression 312
dataset (heat, cold, drought, salt, high osmolarity, UV-B light, and wounding) were essentially single-313
regime shock treatments, lasting 24 hours or less, given to Arabidopsis seedlings (Kilian et al., 314
2007); the protocols in the other studies listed in Supplemental Table S2 were quite similar. The 315
transcriptional responses observed in these experiments therefore necessarily only included part of 316
the plant’s full repertoire. These responses nonetheless follow the coherent pattern seen in Figure 5, 317
a bird’s-eye schematic of B vitamin synthesis and salvage pathways in which enzymes are 318
represented by rectangles. The color density of each enzyme is proportional to the number of differ-319
ent stresses that induce the corresponding gene by two-fold or more. The pathways are arranged in 320
order of prevalence of gene induction. At one extreme (biotin, pantothenate/CoA, and tetrahydro-321
folate pathways) just one or two biosynthetic genes are induced by a single stress, whereas at the 322
other extreme (vitamin B6 and thiamin pathways) all the known biosynthetic genes are induced by at 323
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

10
least three stresses. Except for B6, the prevalence of induction of salvage genes basically tracks that 324
of biosynthetic genes. This overview of gene-level data thus indicates that certain B vitamin biosyn-325
thesis pathways – particularly those for B6 and thiamin – are upregulated by several abiotic stresses 326
and that others – such as the tetrahydrofolate pathway – are largely not. Vitamin/cofactor analysis 327
data and protein-level data for vitamin B6 and thiamin are consistent with this picture, as discussed 328
next. 329 330 Vitamin B6. Pyridoxal 5′-phosphate is made by the pyridoxal synthase complex, which consists of 331
two proteins, PDX1 and PDX2 (the two red-colored boxes in the pyridoxal 5′-phosphate section of 332
Figure 5). Arabidopsis has three PDX1 homologs, of which PDX1.1 and PDX1.3 are active enzymes 333
and PDX1.2 is a positive regulator (Titiz et al., 2006; Moccand et al., 2014). Paralleling the stress 334
induction of Arabidopsis PDX genes (Fig. 5; Supplemental Table S2), PDX1.3 protein accumulated 335
and the total B6 level increased by 60% in response to UV-B exposure (Ristilä et al., 2011); total B6 336
also increased by 60% in response to heat stress (Moccand et al., 2014). Levels of pyridoxal 5'-337
phosphate and free pyridoxal or pyridoxine likewise increased by about two-fold in tobacco exposed 338
to chilling, UV radiation, osmotic stress, oxidative stress, or drought (Huang et al., 2013). 339 340 Thiamin. Thiamin is synthesized by a pathway that is very energetically expensive because the thi-341
azole moiety is made by a suicide enzyme (THI4), and the pyrimidine moiety is made by a radical 342
SAM (S-adenosylmethionine) enzyme (THIC) that probably lasts for only a few catalytic cycles (Box 343
1). Paralleling the stress induction of Arabidopsis THI4, THIC, and other thiamin synthesis genes 344
(Fig. 5; Supplemental Table S2), the level of total thiamin and of its components (thiamin mono- and 345
diphosphates and free thiamin) increased by up to two-fold in Arabidopsis seedlings subjected to 346
cold, osmotic, salinity, or oxidative stress (Tunc-Ozdemir et al., 2009). Similarly, total thiamin 347
increased by up to 60% in Arabidopsis seedlings exposed to salt, osmotic, or oxidative stress, with 348
free thiamin contributing as much or more than thiamin diphosphate to the increase (Rapala-Kozik et 349
al., 2012). Also similarly, in maize seedlings subjected to water, salt, or oxidative stress, total thiamin 350
increased by 70-150%, due to increases in free thiamin and thiamin monophosphate (Rapala-Kozik 351
et al., 2008). However, in this case levels of the cofactor thiamin diphosphate stayed constant or fell 352
by up to 30%, which is quite enough to cause severe deficiency symptoms (Fig. 2) and illustrates the 353
point made above about cofactor levels falling even though free vitamin levels rise. 354 355 Given the suicide nature of the THI4 enzyme, it is striking that THI4 was the most highly or second 356
most highly induced gene in cotton seedlings exposed to cold, drought, salt, or high pH stress; the 357
induction of THI4 relative to unstressed controls ranged from 14-fold to 44-fold (Zhu et al., 2013). It 358
is equally striking that the THI4 protein showed, among all proteins analyzed, the second largest 359
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

11
increase in expression (3.2-fold) in heat-stressed poplar leaves (Ferreira et al., 2006) and the third 360
largest increase in expression (3.7-fold) in heat-stressed wild rice leaves (Scafaro et al., 2010). 361 362 Strategy 3: Repairing or Recycling Damaged B Vitamins and Cofactors 363
Like other organisms, plants have enzyme systems that deal with some of the chemical and enzym-364
atic damage reactions shown in Figure 1 (Linster et al., 2013). These enzyme systems may repair 365
damaged vitamins and cofactors, i.e. directly restore them to the original state, or salvage parts of 366
damaged molecules for re-use in biosynthesis. As vitamin and cofactor repair (Hanson et al., 2016) 367
and recycling (Gerdes et al., 2012) were recently reviewed, we focus here on three cases where the 368
repair or recycling-related activity appears to be upregulated by stress and not simply constitutive. 369 370 Thiamin Salvage Hydrolase TenA_E. TenA_E mediates two successive steps in the recovery of 371
the pyrimidine moiety from thiamin damaged in the thiazole moiety (Zallot et al., 2014). The Ara-372
bidopsis gene encoding TenA_E is upregulated by several abiotic stresses, in common with thiamin 373
synthesis genes (Fig. 4; Supplemental Table S2). The Arabidopsis gene specifying the thiazole 374
salvage enzyme ThiM (Yazdani et al., 2013) is not upregulated (Fig. 4), which fits with the thiazole 375
moiety of thiamin being more prone to irreversible damage than the pyrimidine moiety, and hence 376
not as recyclable (Zallot et al., 2014). 377 378 Thiamin Diphosphate-related Nudix Hydrolases. These enzymes (NUDT20 and NUDT24 in Ara-379
bidopsis) are diphosphatases whose preferred substrates are damaged – and toxic – forms of thiam-380
in diphosphate, including the previously mentioned thiazolone form generated by oxygen-dependent 381
side-reactions (Goyer et al., 2013). The diphosphatase reaction is both a detoxification step and the 382
first step in salvage of the pyrimidine moiety. The Arabidopsis gene(s) (which are not distinguished 383
by microarrays) are induced between two-fold and five-fold by salt, drought, and osmotic stress, and 384
probably also oxidative stress (Kilian et al., 2007; Goyer et al., 2013). 385 386 NAD(P)+ Salvage Module. Two enzymes mediating consecutive steps in NAD(P)+ salvage (nicotin-387
amidase and nicotinate phosphoribosyltransferase) are each induced by two stresses in Arabidopsis 388
(Fig. 5), as is a transporter (NiaP) for nicotinate, the intermediate that these enzymes share (Kilian et 389
al., 2007). Arabidopsis NiaP also transports trigonelline (Fig. 4) (Jeanguenin et al., 2012). NiaP is 390
probably located in the plasma membrane (Jeanguenin et al., 2012), suggesting that nicotinate salv-391
aged in one location could be re-used for NAD(P)+ synthesis in another. This would be consistent 392
with the classical evidence for inter-organ exchange of nicotinate (Bonner and Bonner, 1948). 393
394
Strategy 4: Decreasing Demand for B Vitamins 395
Metabolism can sometimes be re-routed to bypass certain cofactor-dependent steps, as in the phos-396
phate starvation response, in which alternative bypass pathways of cytosolic glycolysis are upregul-397
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

12
ated to spare scarce adenylates and phosphate (Plaxton and Tran, 2011). There are as yet no cases 398
of full-blown re-routing in response to vitamin deficiency, although two studies provide indirect evi-399
dence for redirection of fluxes that may be, at least in part, active responses. In the first study, 400
tobacco leaves overexpressing transketolase, a thiamin diphosphate-dependent enzyme, became 401
moderately thiamin diphosphate-deficient, and showed reduced isoprenoid synthesis via the thiamin 402
diphosphate-dependent enzyme 1-deoxy-D-xylulose 5-phosphate synthase (DXS); this reduction 403
was associated with lower DXS transcript levels (Khozaei et al., 2015). In the second study, a 404
drastic, rapid-onset tetrahydrofolate deficiency in Arabidopsis cells treated with antifolate drugs led 405
to initial depletion of methionine and S-adenosylmethionine pools, reflecting reduction in the tetra-406
hydrofolate-dependent flux of one-carbon units to methionine as one-carbon fluxes were adaptively 407
reprioritized in favor of nucleotides (Loizeau et al., 2007). Surprisingly, the methionine and S-aden-408
osylmethionine pools – and by inference the associated fluxes – subsequently returned to normal, in 409
part via a conserved adaptive mechanism, triggered by tetrahydrofolate deficiency, that involves 410
proteolytic removal of the N-terminal regulatory region of the methionine synthesis enzyme 411
cystathionine γ-synthase (Loizeau et al., 2007). 412
413
AGRICULTURAL IMPLICATIONS 414
As Abraham Blum has emphasized (Blum, 2014), humility and caution are needed when trying to 415
predict agricultural benefits by extrapolating from the responses of seedlings of Arabidopsis or other 416
species to short, sharp stresses in growth chambers (which describes most of the experiments cov-417
ered above). With this warning in mind, what might stress-induced B vitamin and cofactor deficiency 418
imply for crop breeding and management? The primary implication is that breeding for enhanced vit-419
amin content (biofortification) – by transgenic or other approaches – could pay off in terms of stress-420
resistance as well as human nutrition (Fitzpatrick et al., 2012). If stress indeed predisposes to defic-421
iency, a clear prescription for stress-resistance breeding would be: ‘Keep your cofactors safe’. 422 423 The two B vitamins most connected with stress responses, vitamin B6 and thiamin (Fig. 5), are 424
targets of transgenic biofortification efforts in Arabidopsis (Raschke et al., 2011; Pourcel et al., 2013; 425
Vanderschuren et al., 2013; Dong et al., 2015), and stress tests have been run on the engineered 426
plants, albeit mainly with plantlets cultured on agar medium containing sucrose (i.e. not photo-427
synthesizing normally). The total B6 content of leaf tissue was boosted up to four-fold, and included 428
large increases in the cofactor pyridoxal 5'-phosphate and pyridoxine-5'-β-D-glucoside (Raschke et 429
al., 2011). The total thiamin content of leaf tissue was boosted up to 3.4-fold, contributed by up to 430
two-fold increases in the cofactor thiamin diphosphate and up to six-fold increases in thiamin (Dong 431
et al., 2015). The B6-biofortified plants were larger than controls and more resistant to paraquat-432
imposed oxidative stress; no other stress data were reported (Raschke et al., 2011). The thiamin-433
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

13
fortified plants showed no change in resistance to salt, cold, osmotic, drought, or oxidative stress 434
(Dong et al., 2015). To summarize: These first biofortification results for B6 and thiamin are incon-435
clusive and merit follow-up. 436 437 Stress-induced B vitamin deficiency also has implications for crop management (Plaut et al., 2013). 438
Reported benefits of vitamin applications are cited above; such applications are most likely to be 439
useful in practice if they can be given as seed-priming treatments (Al-Hakimi and Hamada, 2001), 440
i.e. applied to seed before sowing. Such seed treatments are cost-efficient (Tanou et al., 2012). 441
442
FUTURE PERSPECTIVES 443
The preceding overview of work relating B vitamins to abiotic stress raises five issues that are listed 444
in the Outstanding Questions Box. Each issue is developed briefly below 445 446 Itemizing Stress-relevant Enzyme Reactions. Although various enzymes and genes for repair or 447
recycling of damaged vitamins and cofactors are known, this is still not the case for about half of the 448
30 damage reactions in Figure 1. While the products of some of these reactions may have no 449
possible fate besides complete degradation, comparative biochemistry suggests that others may 450
well be reclaimed in plants; examples include biotin sulfoxide and chain-shortened biotin (Linster et 451
al., 2013), pyridoxine and nicotinamide N-oxides (Sakuragi and Kummerow, 1960; Shibata et al., 452
1991), and riboflavin-4',5'-phosphate (cyclic FMN) (Fraiz et al., 1998). Nor do we know most of the 453
enzymes or genes for the synthesis and mobilization of the vitamin conjugates in Figure 4, and it is 454
not even clear that we have fully inventoried the conjugates that plants can make. Notably, conjug-455
ated forms of thiamin and its pyrimidine and thiazole precursors have not been reported from plants, 456
but they seem not to have been sought. 457 458 Graduating to Crops and Realistic Stress. As noted previously, most of the modern data linking 459
abiotic stress with B vitamins and cofactors comes from young Arabidopsis plants and unrealistic 460
stress protocols. This is a generic weakness of molecular-level stress research, and the resulting 461
disconnect from agriculture goes a long way toward explaining why molecular work has had a very 462
limited impact on breeding for stress environments (Blum, 2014). It is consequently important to 463
extend stress/vitamin work from Arabidopsis to good genetic model crops such as tomato, canola, 464
maize, and rice, and to adopt stress protocols that mimic field stresses to plants beyond the seedling 465
stage. 466 467 Diagnosing Functional Deficiencies. Vitamin deficiencies, particularly functional ones affecting on-468
ly certain cells or compartments, may be better diagnosed from characteristic changes in metabolite 469
profiles than by directly measuring vitamin levels, as discussed above for vitamin B12 deficiency in 470
humans (Stabler et al., 1996). Given the possibility that direct vitamin measurements can fail to 471
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

14
detect subtle but physiologically critical functional deficiencies, and the high cost and low-throughput 472
nature of many vitamin assays, indirect assessment of plant vitamin and cofactor status via metabol-473
omics is appealing. Confirming or disconfirming the validity of this approach in plants, as in humans 474
(Stabler et al., 2013), will require prior metabolic profiling studies of vitamin-deficient mutants. An 475
alternative indirect, high-throughput way to detect functional vitamin deficiencies may be via charact-476
eristic transcriptome changes, analogous to the induction of specific genes by mineral nutrient defic-477
iencies (e.g. Nikiforova et al., 2003; Zheng et al., 2009). Furthermore, transcriptome analysis could 478
uncover mechanisms that re-route metabolism to mitigate the effects of vitamin deficiency (see 479
above). Again, evaluation of these possibilities will require studies of vitamin-deficient mutants. 480
481
Genotypic Variation. Although there is natural genotypic variation in vitamin content (Hanson and 482
Gregory, 2011; Fitzpatrick et al., 2012), it would be impractical to use vitamin content as a selection 483
criterion to develop experimental populations due to the issues of assay cost and efficacy just outlin-484
ed. On the other hand, were high-throughput indirect methods to assess deficiency available (see 485
above), it would be feasible to assess vitamin/cofactor status in populations subjected to realistic 486
stresses, and then to work back via genome-wide association studies to genes governing 487
vitamin/cofactor status. Once identified, such genes, which would presumably include known and 488
novel genes with repair/recycling, synthesis, storage, or bypass functions, could be used to explore 489
a ‘keep your cofactors safe’ breeding strategy for abiotic stress resistance. 490 491 Is Thiamin an Achilles’ Heel in Stress Metabolism? A thread running through the whole of this re-492
view is that thiamin and thiamin diphosphate are not like the other B vitamins and cofactors. Thiamin 493
diphosphate may turn over exceptionally fast, at least in part because of its unusual feature of 494
catalytic inactivation. Thiamin is energetically costly to make due to the suicidal nature of one of its 495
biosynthetic enzymes and the suicidal tendency of another. Because thiamin synthesis depends 496
absolutely on concurrent protein synthesis it is vulnerable to stress in a way that other B vitamins are 497
not. Unlike most other B vitamins, thiamin and its precursors lack known storage forms. Thiamin 498
synthesis and salvage genes are particularly highly stress-regulated in Arabidopsis seedlings. And 499
lastly, thiamin is more prominent than other B vitamins in the ‘spray and pray’ literature. Taken 500
together, these features suggest that thiamin could be an Achilles’ heel – a crucial weak point – in 501
plant stress metabolism. 502
503
Supplemental Data 504
The following supplemental materials are available. 505
• Supplemental Table S1. Analysis of Arabidopsis stress literature based on PubMed search 506
terms. 507
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

15
• Supplemental Table S2. Stress-induced B vitamin/cofactor synthesis and salvage genes in 508
Arabidopsis. 509
510
ACKNOWLEDGMENTS 511
We apologize to the authors of relevant studies that could not be cited here due to page limitations, 512
and thank Dr. Claudia Lerma Ortiz for a helpful critique of a draft of the review. 513
514
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

16
515
Box 1. Thiamin Biosynthesis – A High-cost, High-risk Operation 516
Enzymes all have finite lifespans, but some have much shorter lifespans than others. These include 517
so-called ‘suicide enzymes’ in which a residue is irreversibly altered after just one turnover, as well 518
as enzymes employing reactive radicals as intermediates, e.g. radical S-adenosylmethionine (SAM) 519
enzymes, which typically catalyze fewer than a dozen turnovers in vitro (Broderick et al., 2014). 520
Enzymes with short lifespans are energetically costly because they have to be replaced so often. 521 522 Thiamin is much more energetically costly to produce than other vitamins because its first biosynth-523
etic enzymes, THI4 and THIC, have extremely short lifespans. THI4 (also called THI1) synthesizes 524
the thiazole moiety of thiamin (Figure). The sulfur atom in the thiazole ring comes from a conserved 525
cysteine in THI4 whose loss permanently inactivates the protein, making THI4 a suicide enzyme 526
(Chatterjee et al., 2013). THIC synthesizes the pyrimidine moiety of thiamin. THIC is a radical SAM 527
enzyme that may achieve no more than five catalytic turnovers in vitro (Palmer and Downs, 2013). 528
Thus THI4 and THIC mediate orders of magnitude fewer catalytic events during their lives than most 529
other enzymes. 530 531 This catalytic inefficiency is manifested in vivo. In proteomic analyses of barley leaves, the THI4 and 532
THIC proteins had, respectively, the first and fifth highest turnover rates of more than 500 proteins 533
studied (Nelson et al., 2014). The half-lives of THI4 and THIC were just a few hours compared to 534
days for most proteins. 535 536 Another consequence may be high vulnerability of the thiamin biosynthetic pathway to stress. Global 537
responses to abiotic stress include shutting down protein synthesis in general (Dhindsa and Cleland, 538
1975; Good and Zaplachinski, 1994) and specifically inducing proteins involved in protein folding and 539
processing (Liu and Howell, 2010). Such reprioritization could adversely impact the replacement of 540
THI4 and THIC, without which thiamin synthesis cannot proceed. 541
542
Box 1 Figure. Catalytic longevity and protein turnover in thiamin synthesis. The number of catalytic 543
turnovers that each pathway enzyme mediates during its lifetime (represented by the width of the 544
blue reaction arrows) varies from thousands to <10 for THIC and one for THI4. The consequent 545
rapid turnover of the THIC and THI4 proteins is schematized by red arrows. Abbreviations: AIR, 546
aminoimidazole ribotide; HET, hydroxyethylthiazole, HMP, hydroxymethylpyrimidine, P, phosphate, 547
PP, diphosphate. 548
549
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

17
550
Advances Box 551
● Recent research and the classical literature on ‘the chemical cure of climatic lesions’ converge to 552
indicate that stressed plants can develop functional B vitamin deficiencies. 553
● Fully appreciating the lability and metabolic centrality of B vitamins and cofactors makes clear that 554
they are crucial stress-sensitive nodes in pathway networks that cells must seek to protect. 555
● Protection strategies include repairing or recycling damaged vitamins and cofactors, increasing 556
biosynthesis by upregulating biosynthetic gene expression, and probably mobilizing reserves. 557
● Thiamin is the most metabolically labile B vitamin and the only one whose synthesis depends on 558
suicidal enzymes, implying that thiamin pools are doubly difficult to maintain in stress conditions. 559
560
561
Outstanding Questions Box 562
● Have we documented all the relevant metabolic reactions of B vitamins and cofactors? 563
● Do the results from model stresses in Arabidopsis apply to realistic stresses in crops? 564
● Can functional B vitamin deficiency be diagnosed from metabolome/transcriptome changes? 565
● How far can genotypic variation in vitamins or cofactors explain variation in stress resistance? 566
● Is thiamin really the most stress-sensitive B vitamin? 567
568
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

18
FIGURE LEGENDS 569
570
Figure 1. Structures and damage reactions of the seven B vitamins found in plants and of repres-571
entative cofactors derived from them. The vitamin moieties are highlighted in blue. Note that folates 572
generally have a short γ-linked polyglutamate chain attached to the glutamyl moiety. Color-coded 573
arrows show the site and nature of spontaneous chemical or enzymatic damage reactions that each 574
vitamin/cofactor can undergo in vivo. The damage reactions involved are documented in Table I. 575
576
Figure 2. Comparing growth responses to deficiencies of thiamin, B6, or folate in rats versus plants. 577
The decline from vitamin sufficiency to deficiency is approximated by a color gradient. For rats, 578
deficiencies were obtained by varying dietary vitamin content; liver was used to assess vitamin 579
status. For plants, the experiments involved vitamin deficient mutants (blk-1 for thiamin in maize, 580
pdx1.3 for B6 in Arabidopsis) or cell cultures treated with an antifolate drug for folate in Arabidopsis. 581
Whole plants, leaves, or cells were used to assess vitamin status. Data sources: rat thiamin, Rains 582
et al. (1997); rat B6, Mackey et al. (2003) and Scheer et al. (2005); rat folate, Clifford et al. (1993); 583
maize thiamin, Woodward et al. (2010); Arabidopsis B6, Rueschhoff et al. (2013); Arabidopsis folate, 584
Loizeau et al. (2008). 585
586
Figure 3. Classical data on the chemical cure of high- or low-temperature growth lesions. Day/night 587
temperatures are in ºC. +, Vitamin(s) added; – , no vitamin; differences significant at * P <0.05, ** P 588
<0.01. The data are for species of the ornamental plant Cosmos (Bonner, 1943), for Arabidopsis 589
(Langridge and Griffing, 1959), and for other species (Ketellapper, 1963). 590
591
Figure 4. Structures of conjugated forms of B vitamins and their precursors that occur in plants. The 592
molecules to which the vitamins and precursors are conjugated are colored blue. 593
594
Figure 5. Stress induction of B vitamin and cofactor synthesis and salvage genes in Arabidopsis. 595
Synthesis and salvage pathways for each vitamin are schematized as in Gerdes et al. (2012) except 596
for the omission of compartmentation and the addition of two thiamin salvage reactions (Yazdani et 597
al., 2013; Zallot et al., 2014). Circles are metabolites, arrows are reactions, and rectangles are 598
enzymes. The full pathway schemes, which include enzyme and metabolite names, are available at 599
http://pubseed.theseed.org/seedviewer.cgi?page=PlantGateway. Enzymes whose genes are induc-600
ed at least two-fold by 0, 1, 2, or ≥3 different abiotic stresses (in shoots or roots) are color coded 601
white and three shades of red, respectively. Gene expression data were taken from AtGenExpress 602
and literature summarized in Supplemental Table S2. 603
604
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

19
Table I. Chemical and enzymatic damage reactions of B vitamins and the corresponding cofactors
Vitamin/cofactor Reactions C/Ea Referencesb
Thiamin (vitamin B1) Thiamin diphosphate
Pyrimidine ring deamination Thiazole ring oxidation Thiazole ring breakdown Oxidation to thiamin acetate Hydrolysis to thiazole + pyrimidine Dephosphorylation Thiamin triphosphate formation
C C,E C E E E E
Windheuser and Higuchi, 1962 Bunik et al., 2007; Dwivedi and Arnold, 1973 Jenkins et al., 2007 Dalvi et al., 1974 Jurgenson et al., 2009 Rapala-Kozik et al., 2009 Linster et al., 2013
Riboflavin (vitamin B2) FMN, FAD
(Photo)oxidative flavin ring loss (Di)phosphate bond hydrolysis 7α- and 8α-hydroxylation Cyclic FMN formation from FAD
C E E
C,E
Choe et al., 2005 Ogawa et al., 2008; Rawat et al., 2011 Ohkawa et al., 1983 Pinto et al., 1999; Sánchez-Moreno et al. 2009
Niacin (vitamin B3) NAD(P)(H)
Hydration of nicotinamide ring Epimerization of β- to α-NAD(P)H Hydrolytic loss of nicotinamide ring Diphosphate bond hydrolysis Nicotinamide ring addition reactions
C,E C E E C
Marbaix et al., 2011 Oppenheimer and Kaplan, 1975 Everse et al., 1975 Ogawa et al., 2008 Everse et al., 1971
Pantothenate (vitamin B5) Coenzyme A, ACPc
Oxidations of the thiol group Diphosphate bond hydrolysis Hydrolysis of the 3' phosphate group
C E E
Huang et al., 2016 Ogawa et al., 2008 Paizs et al., 2008
Pyridoxine (vitamin B6) Pyridoxal 5'-phosphate
Aldehyde group oxidation Aldehyde group condensations Dephosphorylation 6-Hydroxylation
C/E C E C
Gerdes et al., 2012 Dalling et al., 1976 Gerdes et al., 2012 Tadera et al., 1986
Biotin (vitamin B8) Biotinylated enzymes
Oxidation to biotin sulfoxide Side chain β-oxidation
C E
Melville, 1954 Izumi et al., 1973
Tetrahydrofolate C1-Substituted folates
Oxidative cleavage of C9-N10 bond Pteridine ring oxidation 5-Formyltetrahydrofolate formation γ-Glutamyl bond hydrolysis
C C/E C/E E
Gregory, 1989 Noiriel et al., 2007 Baggott, 2000; Goyer et al., 2005 Orsomando et al., 2005; Bozzo et al., 2008
605 aC, chemical (i.e. non-enzymatic, spontaneous) reaction; E, enzymatic reaction or side-reaction. 606 bCertain reactions have so far been reported only from animals or microbes. cACP, acyl carrier protein, which has a bound 4'-phosphopantetheinyl prosthetic group derived from coenzyme A.
607
608
609
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

20
610 LITERATURE CITED 611
Akhtar TA, Orsomando G, Mehrshahi P, Lara-Núñez A, Bennett MJ, Gregory JF 3rd, Hanson AD 612
(2010) A central role for gamma-glutamyl hydrolases in plant folate homeostasis. Plant J 64: 613
256-266 614
Al-Hakimi A, Hamada AM (2001) Counteraction of salinity stress on wheat plants by grain soaking in 615
ascorbic acid, thiamin or sodium salicylate. Biol Plantarum 44: 253-261 616
Amachi T, Imamoto S, Yoshizumi H (1971) A growth factor for malo-lactic fermentation bacteria. Part 617
II. Structure and synthesis of a novel pantothenic acid derivative isolated from tomato juice. Agric 618
Biol Chem 35: 1222-1230 619
Ariaey-Nejad MR, Balaghi M, Baker EM, Sauberlich HE (1970) Thiamin metabolism in man. Am J 620
Clin Nutr 23: 764-778 621
Asensi-Fabado MA, Munné-Bosch S (2010) Vitamins in plants: occurrence, biosynthesis and 622
antioxidant function. Trends Plant Sci 15: 582-592 623
Ashihara H, Yin Y, Deng WW, Watanabe S (2010) Pyridine salvage and nicotinic acid conjugate 624
synthesis in leaves of mangrove species. Phytochemistry 71: 47-53 625
Baggott JE (2000) Hydrolysis of 5,10-methenyltetrahydrofolate to 5-formyltetrahydrofolate at pH 2.5 626
to 4.5. Biochemistry 39: 14647-14653 627
Blum A (2014) Genomics for drought resistance – getting down to earth. Funct Plant Biol 41: 1191-628
1198 629
Bonner J (1943) Effects of application of thiamin to Cosmos. Botan Gaz 104: 475-479 630
Bonner J (1957) The chemical cure of climatic lesions. Eng Sci 20: 28-30 631
Bonner J, Bonner H (1948) The B vitamins as plant hormones. Vitam Horm 6: 225-275 632
Bozzo GG, Basset GJ, Naponelli V, Noiriel A, Gregory JF 3rd, Hanson AD (2008) Characterization 633
of the folate salvage enzyme p-aminobenzoylglutamate hydrolase in plants. Phytochemistry 69: 634
29-37 635
Broderick JB, Duffus BR, Duschene KS, Shepard EM (2014) Radical S-adenosylmethionine 636
enzymes. Chem Rev 114: 4229-4317 637
Bunik VI, Schloss JV, Pinto JT, Gibson GE, Cooper AJ (2007) Enzyme-catalyzed side reactions with 638
molecular oxygen may contribute to cell signaling and neurodegenerative diseases. Neurochem 639
Res 32: 871-891 640
Chatterjee A, Abeydeera ND, Bale S, Pai PJ, Dorrestein PC, Russell DH, Ealick SE, Begley TP 641
(2011) Saccharomyces cerevisiae THI4p is a suicide thiamine thiazole synthase. Nature 478: 642
542-546 643
Chen H, Xiong L (2005) Pyridoxine is required for post-embryonic root development and tolerance to 644
osmotic and oxidative stresses. Plant J 44: 396-408 645
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

21
Choe E, Huang R, Min DB (2005) Chemical reactions and stability of riboflavin in foods. J Food Sci 646
70: R28-R36 647
Clifford AJ, Bills ND, Peerson JM, Müller HG, Burk GE, Rich KD (1993) A depletion-repletion folate 648
bioassay based on growth and tissue folate concentrations of rats. J Nutr 123: 926-932 649
Coburn SP (1990) Location and turnover of vitamin B6 pools and vitamin B6 requirements of 650
humans. Ann N Y Acad Sci 585: 76-85 651
Dalling DK, Grant DM, Horton WJ, Sagers RD (1976) Carbon 13 NMR study of nonenzymatic 652
reactions of pyridoxal 5'-phosphate with selected amino acids and of related reactions. J Biol 653
Chem 251: 7661-7668 654
Dalvi RR, Sauberlich HE, Neal RA (1974) An examination of the metabolism of thiamin by rat liver 655
alcohol dehydrogenase. J Nutr 104: 1476-1483 656
Dehaye L, Duval M, Viguier D, Yaxley J, Job D (1997) Cloning and expression of the pea gene 657
encoding SBP65, a seed-specific biotinylated protein. Plant Mol Biol 35: 605-621 658
Dhindsa RS, Cleland RE (1975) Water stress and protein synthesis 1. Differential inhibition of 659
protein synthesis. Plant Physiol 55: 778-781 660
Díaz de la Garza R, Quinlivan EP, Klaus SM, Basset GJ, Gregory JF 3rd, Hanson AD (2004) Folate 661
biofortification in tomatoes by engineering the pteridine branch of folate synthesis. Proc Natl 662
Acad Sci USA 101: 13720-13725 663
Dong W, Stockwell VO, Goyer A (2015) Enhancement of thiamin content in Arabidopsis thaliana by 664
metabolic engineering. Plant Cell Physiol 56: 2285-2296 665
Dwivedi BK, Arnold RG (1973) Chemistry of thiamine degradation in food products and model 666
systems: a review. J Agric Food Chem 21: 54-60 667
Espinosa-Ruiz A, Belles JM, Serrano R, Culianez-MacIa FA (1999) Arabidopsis thaliana AtHAL 3: a 668
flavoprotein related to salt and osmotic tolerance and plant growth. Plant J 20: 529-539 669
Eudes A, Bozzo GG, Waller JC, Naponelli V, Lim EK, Bowles DJ, Gregory JF 3rd, Hanson AD 670
(2008) Metabolism of the folate precursor p-aminobenzoate in plants: glucose ester formation 671
and vacuolar storage. J Biol Chem 283: 15451-15459 672
Everse J, Cooper Zoll E, Kahan L, Kaplan NO (1971) Addition products of diphosphopyridine nucl-673
eotides with substrates of pyridine nucleotide-linked dehydrogenases. Bioorg Chem 1: 207-233 674
Everse J, Everse KE, Kaplan NO (1975) The pyridine nucleosidases from Bacillus subtilis and 675
Neurospora crassa. Isolation and structural properties. Arch Biochem Biophys 169: 702-713 676
Ferreira S, Hjernø K, Larsen M, Wingsle G, Larsen P, Fey S, Roepstorff P, Salomé Pais M (2006) 677
Proteome profiling of Populus euphratica Oliv. upon heat stress. Ann Bot 98: 361-377 678
Fitzpatrick TB (2011) Vitamin B6 in plants: more than meets the eye. Adv Bot Res 59: 1-38 679
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

22
Fitzpatrick TB, Basset GJ, Borel P, Carrari F, DellaPenna D, Fraser PD, Hellmann H, Osorio S, 680
Rothan C, Valpuesta V, Caris-Veyrat C, Fernie AR (2012) Vitamin deficiencies in humans: can 681
plant science help? Plant Cell 24: 395-414 682
Foyer CH, Noctor G (2011) Ascorbate and glutathione: the heart of the redox hub. Plant Physiol 155: 683
2-18 684
Fraiz FJ, Pinto RM, Costas MJ, Aavalos M, Canales J, Cabezas A, Cameselle JC (1998) Enzymic 685
formation of riboflavin 4',5'-cyclic phosphate from FAD: evidence for a specific low-Km FMN 686
cyclase in rat liver. Biochem J 330: 881-888 687
Frelin O, Huang L, Hasnain G, Jeffryes JG, Ziemak MJ, Rocca JR, Wang B, Rice J, Roje S, Yurgel 688
SN, Gregory JF 3rd, Edison AS, Henry CS, de Crécy-Lagard V, Hanson AD (2015) A directed-689
overflow and damage-control N-glycosidase in riboflavin biosynthesis. Biochem J 466: 137-145 690
Gerdes S, Lerma-Ortiz C, Frelin O, Seaver SM, Henry CS, de Crécy-Lagard V, Hanson AD (2012) 691
Plant B vitamin pathways and their compartmentation: a guide for the perplexed. J Exp Bot 63: 692
5379-5395 693
Good AG, Zaplachinski ST (1994) The effects of drought stress on free amino acid accumulation 694
and protein synthesis in Brassica napus. Physiol Plant 90: 9-14 695
Goyer A, Collakova E, Díaz de la Garza R, Quinlivan EP, Williamson J, Gregory JF 3rd, Shachar-Hill 696
Y, Hanson AD (2005) 5-Formyltetrahydrofolate is an inhibitory but well tolerated metabolite in 697
Arabidopsis leaves. J Biol Chem 280: 26137-26142 698
Goyer A, Hasnain G, Frelin O, Ralat MA, Gregory JF 3rd, Hanson AD (2013) A cross-kingdom Nudix 699
enzyme that pre-empts damage in thiamin metabolism. Biochem J 454: 533-542 700
Green R (2008) Indicators for assessing folate and vitamin B12 status and for monitoring the efficacy 701
of intervention strategies. Food Nutr Bull 29: S52-S63 702
Gregory JF 3rd (1989) Chemical and nutritional aspects of folate research: analytical procedures, 703
methods of folate synthesis, stability, and bioavailability of dietary folates. Adv Food Nutr Res 33: 704
1-101 705
Gregory JF 3rd (1998) Nutritional properties and significance of vitamin glycosides. Annu Rev Nutr 706
18: 277-296 707
Gregory JF 3rd, Quinlivan EP (2002) In vivo kinetics of folate metabolism. Annu Rev Nutr 22: 199-708
220 709
Guenther BD, Sheppard CA, Tran P, Rozen R, Matthews RG, Ludwig ML (1999) The structure and 710
properties of methylenetetrahydrofolate reductase from Escherichia coli suggest how folate 711
ameliorates human hyperhomocysteinemia. Nat Struct Biol 6: 359-365 712
Hall CA (1964) Long term excretion of Co57-vitamin B12 and turnover within plasma. Am J Clin 713
Nutr 14: 156-162 714
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

23
Hanson AD, Gregory JF 3rd (2011) Folate biosynthesis, turnover, and transport in plants. Annu Rev 715
Plant Biol 62: 105-125 716
Hanson AD, Henry CS, Fiehn O, de Crécy-Lagard V (2016) Metabolite damage and metabolite 717
damage control in plants. Annu Rev Plant Biol 67: 131-152 718
Hasnain G, Frelin O, Roje S, Ellens KW, Ali K, Guan JC, Garrett TJ, de Crecy-Lagard V, Gregory JF 719
3rd, McCarty DR, Hanson AD (2013) Identification and characterization of the missing pyrimidine 720
reductase in the plant riboflavin biosynthesis pathway. Plant Physiol 161: 48-56 721
Higa A, Khandakar J, Mori Y, Kitamura Y (2012) Increased de novo riboflavin synthesis and hydro-722
lysis of FMN are involved in riboflavin secretion from Hyoscyamus albus hairy roots under iron 723
deficiency. Plant Physiol Biochem 58: 166-173 724
Huang L, Khusnutdinova A, Nocek B, Brown G, Xu X, Cui H, Petit P, Flick R, Zallot R, Balmant K, 725
Ziemak MJ, Shanklin J, de Crécy-Lagard V, Fiehn O, Gregory JF 3rd, Joachimiak A, Savchenko 726
A, Yakunin AF, Hanson AD (2016) A family of metal-dependent phosphatases implicated in 727
metabolite damage-control. Nat Chem Biol 12: 621-627 728
Huang S, Zhang J, Wang L, Huang L (2013) Effect of abiotic stress on the abundance of different 729
vitamin B6 vitamers in tobacco plants. Plant Physiol Biochem 66: 63-67 730
Izumi Y, Tani Y, Ogata K (1973) The conversion of bisnorbiotin and bisnordethiobiotin to biotin and 731
dethiobiotin, respectively, by microorganisms. Biochim Biophys Acta 326: 485-488 732
Jeanguenin L, Lara-Núñez A, Rodionov DA, Osterman AL, Komarova NY, Rentsch D, Gregory JF 733
3rd, Hanson AD (2012) Comparative genomics and functional analysis of the NiaP family uncover 734
nicotinate transporters from bacteria, plants, and mammals. Funct Integr Genomics 12: 25-34 735
Jenkins AH, Schyns G, Potot S, Sun G, Begley TP (2007) A new thiamin salvage pathway. Nat 736
Chem Biol 3: 492-497 737
Jurgenson CT, Begley TP, Ealick SE (2009) The structural and biochemical foundations of thiamin 738
biosynthesis. Annu Rev Biochem 78: 569-603 739
Kaya C, Ashraf M, Sonmez O, Tuna AL, Polat T, Aydemir S (2015) Exogenous application of thi-740
amin promotes growth and antioxidative defense system at initial phases of development in salt-741
stressed plants of two maize cultivars differing in salinity tolerance. Acta Physiol Plant 37: 1741 742
Ketellapper HJ (1963) Temperature-induced chemical defects in higher plants. Plant Physiol 38: 743
175-179 744
Khozaei M, Fisk S, Lawson T, Gibon Y, Sulpice R, Stitt M, Lefebvre SC, Raines CA (2015) 745
Overexpression of plastid transketolase in tobacco results in a thiamine auxotrophic phenotype. 746
Plant Cell 27: 432-447 747
Kilian J, Whitehead D, Horak J, Wanke D, Weinl S, Batistic O, D'Angelo C, Bornberg-Bauer E, Kudla 748
J, Harter K (2007) The AtGenExpress global stress expression data set: protocols, evaluation 749
and model data analysis of UV-B light, drought and cold stress responses. Plant J 50: 347-363 750
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

24
Kohler RE (1975) The history of biochemistry: A survey. J Hist Biol 8: 275-318 751
Kupke T, Hernandez-Acosta P, Steinbacher S, Culianez-Macia FA (2001) Arabidopsis thaliana 752
flavoprotein AtHAL3a catalyzes the decarboxylation of 4'-phosphopantothenoylcysteine to 4'-753
phosphopantetheine, a key step in coenzyme A biosynthesis. J Biol Chem 276: 19190-19196 754
Langridge J, Griffing B (1959) A study of high temperature lesions in Arabidopsis thaliana. Aust J 755
Biol Sci 12: 117-135 756
Lerma-Ortiz C, Jeffryes JG, Cooper AJ, Niehaus TD, Thamm AM, Frelin O, Aunins T, Fiehn O, de 757
Crécy-Lagard V, Henry CS, Hanson AD (2016) 'Nothing of chemistry disappears in biology': The 758
Top 30 damage-prone endogenous metabolites. Biochem Soc Trans 44: 961-971 759
Lesgards J-F, Lehucher-Michel M-P, Vidal N, Stocker P (2015) Assessment of antioxidative activity 760
of lipid- and water-soluble vitamins in human whole blood. comparative analysis between a 761
biological test and chemical methods. Int J Vitam Nutr Res 75: 11-18 762
Li W, Zhang F, Chang Y, Zhao T, Schranz ME, Wang G (2015) Nicotinate O-glucosylation is an evo-763
lutionarily metabolic trait important for seed germination under stress conditions in Arabidopsis 764
thaliana. Plant Cell 27: 1907-1924 765
Linster CL, Van Schaftingen E, Hanson AD (2013) Metabolite damage and its repair or pre-emption. 766
Nat Chem Biol 9: 72-80 767
Liu JX, Howell SH (2010) Endoplasmic reticulum protein quality control and its relationship to 768
environmental stress responses in plants. Plant Cell 22: 2930-2942 769
Loizeau K, De Brouwer V, Gambonnet B, Yu A, Renou JP, Van Der Straeten D, Lambert WE, 770
Rébeillé F, Ravanel S (2008) A genome-wide and metabolic analysis determined the adaptive 771
response of Arabidopsis cells to folate depletion induced by methotrexate. Plant Physiol 148: 772
2083-2095 773
Ludwig-Müller J (2011) Auxin conjugates: their role for plant development and in the evolution of 774
land plants. J Exp Bot 62: 1757-1773 775
Lukienko PI, Mel'nichenko NG, Zverinskii IV, Zabrodskaya SV (2000) Antioxidant properties of 776
thiamin. Bull Exp Biol Med 130: 874-876 777
Mackey AD, Lieu SO, Carman C, Gregory JF 3rd (2003) Hydrolytic activity toward pyridoxine-5'-beta-778
D-glucoside in rat intestinal mucosa is not increased by vitamin B-6 deficiency: effect of basal 779
diet composition and pyridoxine intake. J Nutr 133: 1362-1367 780
Marbaix AY, Noël G, Detroux AM, Vertommen D, Van Schaftingen E, Linster CL (2011) Extremely 781
conserved ATP- or ADP-dependent enzymatic system for nicotinamide nucleotide repair. J Biol 782
Chem 286: 41246-41252 783
Matsui A, Yin Y, Yamanaka K, Iwasaki M, Ashihara H (2007) Metabolic fate of nicotinamide in higher 784
plants. Physiol Plant 131: 191-200 785
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

25
McCourt JA, Nixon PF, Duggleby RG (2006) Thiamin nutrition and catalysis-induced instability of 786
thiamin diphosphate. Br J Nutr 96: 636-638 787
McHale NA, Hanson KR, Zelitch I (1988) A nuclear mutation in Nicotiana sylvestris causing a 788
thiamine-reversible defect in synthesis of chloroplast pigments. Plant Physiol 88: 930-935 789
Melville DB (1954) Biotin sulfoxide. J Biol Chem 208: 495-501 790
Menegus F, Lilliu I, Scaglioni L (2002) A unique pantoyllactone glycoside system is activated in rice 791
seedlings developing aerobically in the dark. Physiol Plant 116: 299-307 792
Mills PB, Struys E, Jakobs C, Plecko B, Baxter P, Baumgartner M, Willemsen MA, Omran H, Tacke 793
U, Uhlenberg B, Weschke B, Clayton PT (2006) Mutations in antiquitin in individuals with 794
pyridoxine-dependent seizures. Nat Med 12: 307-309 795
Mizuno K, Matsuzaki M, Kanazawa S, Tokiwano T, Yoshizawa Y, Kato M (2014) Conversion of 796
nicotinic acid to trigonelline is catalyzed by N-methyltransferase belonged to motif B' 797
methyltransferase family in Coffea arabica. Biochem Biophys Res Commun 452: 1060-1066 798
Moccand C, Boycheva S, Surriabre P, Tambasco-Studart M, Raschke M, Kaufmann M, Fitzpatrick 799
TB (2014) The pseudoenzyme PDX1.2 boosts vitamin B6 biosynthesis under heat and oxidative 800
stress in Arabidopsis. J Biol Chem 289: 8203-8216 801
Mohammadi PP, Moieni A, Hiraga S, Komatsu S (2012) Organ-specific proteomic analysis of 802
drought-stressed soybean seedlings. J Proteomics 75: 1906-1923 803
Navarrete O, Van Daele J, Stove C, Lambert W, Van Der Straeten D, Storozhenko S (2012) A folate 804
independent role for cytosolic HPPK/DHPS upon stress in Arabidopsis thaliana. Phytochemistry 805
73: 23-33 806
Nelson CJ, Alexova R, Jacoby RP, Millar AH (2014) Protein with high turnover rate in barley leaves 807
estimated by proteome analysis combined with in planta isotope labeling. Plant Physiol 166: 91-808
108 809
Nikiforova V, Freitag J, Kempa S, Adamik M, Hesse H, Hoefgen R (2003) Transcriptome analysis of 810
sulfur depletion in Arabidopsis thaliana: interlacing of biosynthetic pathways provides response 811
specificity. Plant J 33: 633-650 812
Noiriel A, Naponelli V, Gregory JF 3rd, Hanson AD (2007) Pterin and folate salvage. Plants and 813
Escherichia coli lack capacity to reduce oxidized pterins. Plant Physiol 143: 1101-1109 814
Ogawa T, Yoshimura K, Miyake H, Ishikawa K, Ito D, Tanabe N, Shigeoka S (2008) Molecular char-815
acterization of organelle-type Nudix hydrolases in Arabidopsis. Plant Physiol 148: 1412-1424 816
Ohkawa H, Ohishi N, Yagi K (1983) New metabolites of riboflavin appear in human urine. J Biol 817
Chem 258: 5623-5628 818
Oppenheimer NJ, Kaplan NO (1975) The alpha beta epimerization of reduced nicotinamide adenine 819
dinucleotide. Arch Biochem Biophys 166: 526-535 820
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

26
Orsomando G, de la Garza RD, Green BJ, Peng M, Rea PA, Ryan TJ, Gregory JF 3rd, Hanson AD 821
(2005) Plant gamma-glutamyl hydrolases and folate polyglutamates: characterization, compart-822
mentation, and co-occurrence in vacuoles. J Biol Chem 280: 28877-28884 823
Ouyang M, Ma J, Zou M, Guo J, Wang L, Lu C, Zhang L (2010) The photosensitive phs1 mutant is 824
impaired in the riboflavin biogenesis pathway. J Plant Physiol 167: 1466-1476 825
Paizs C, Diemer T, Rétey J (2008) The putative coenzyme B12-dependent methylmalonyl-CoA 826
mutase from potatoes is a phosphatase. Bioorg Chem 36: 261-264 827
Palmer LD, Downs DM (2013) The thiamine biosynthetic enzyme ThiC catalyzes multiple turnovers 828
and is inhibited by S-adenosylmethionine (AdoMet) metabolites. J Biol Chem 288: 30693-30699 829
Piedrafita G, Keller MA, Ralser M (2015) The impact of non-enzymatic reactions and enzyme pro-830
miscuity on cellular metabolism during (oxidative) stress conditions. Biomolecules 5: 2101-2122 831
Piotrowska A, Bajguz A (2011) Conjugates of abscisic acid, brassinosteroids, ethylene, gibberellins, 832
and jasmonates. Phytochemistry 72: 2097-2112 833
Pinto RM, Fraiz FJ, Cabezas A, Avalos M, Canales J, Costas MJ, Cameselle JC (1999) Preparation 834
of riboflavin 4',5'-cyclic phosphate by incubation of flavin-adenine dinucleotide with Mn2+ in the 835
absence of riboflavin 5'-phosphate cyclase. Anal Biochem 268: 409-411 836
Plaut Z, Edelstein M, Ben-Hur M (2013) Overcoming salinity barriers to crop production using 837
traditional methods. Crit Rev Plant Sci 32: 250-291 838
Plaxton WC, Tran HT (2011) Metabolic adaptations of phosphate-starved plants. Plant Physiol 156: 839
1006-1015 840
Pourcel L, Moulin M, Fitzpatrick TB (2013) Examining strategies to facilitate vitamin B1 biofortification 841
of plants by genetic engineering. Front Plant Sci 4: 160 842
Quinlivan EP, Roje S, Basset G, Shachar-Hill Y, Gregory JF 3rd, Hanson AD (2003) The folate 843
precursor p-aminobenzoate is reversibly converted to its glucose ester in the plant cytosol. J Biol 844
Chem 278: 20731-20737 845
Rains TM, Emmert JL, Baker DH, Shay NF (1997) Minimum thiamin requirement of weanling 846
Sprague-Dawley outbred rats. J Nutr 127: 167-170 847
Rapala-Kozik M, Gołda A, Kujda M (2009) Enzymes that control the thiamine diphosphate pool in 848
plant tissues. Properties of thiamine pyrophosphokinase and thiamine-(di)phosphate phosphat-849
ase purified from Zea mays seedlings. Plant Physiol Biochem 47: 237-242 850
Rapala-Kozik M, Kowalska E, Ostrowska K (2008) Modulation of thiamine metabolism in Zea mays 851
seedlings under conditions of abiotic stress. J Exp Bot 59: 4133-4143 852
Rapala-Kozik M, Wolak N, Kujda M, Banas AK (2012) The upregulation of thiamine (vitamin B1) 853
biosynthesis in Arabidopsis thaliana seedlings under salt and osmotic stress conditions is 854
mediated by abscisic acid at the early stages of this stress response. BMC Plant Biol 12: 2 855
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

27
Raschke M, Boycheva S, Crèvecoeur M, Nunes-Nesi A, Witt S, Fernie AR, Amrhein N, Fitzpatrick 856
TB (2011) Enhanced levels of vitamin B6 increase aerial organ size and positively affect stress 857
tolerance in Arabidopsis. Plant J 66: 414-432 858
Rawat R, Sandoval FJ, Wei Z, Winkler R, Roje S (2011) An FMN hydrolase of the haloacid dehalo-859
genase superfamily is active in plant chloroplasts. J Biol Chem 286: 42091-42098 860
Ristilä M, Strid H, Eriksson LA, Strid A, Sävenstrand H (2011) The role of the pyridoxine (vitamin B6) 861
biosynthesis enzyme PDX1 in ultraviolet-B radiation responses in plants. . Plant Physiol Biochem 862
49: 284-292 863
Rueschhoff EE, Gillikin JW, Sederoff HW, Daub ME (2013) The SOS4 pyridoxal kinase is required 864
for maintenance of vitamin B6-mediated processes in chloroplasts. Plant Physiol Biochem 63: 865
281-291 866
Sakuragi T, Kummerow FA (1960) Utilization of pyridoxine N-oxide by rats. Proc Soc Exp Biol Med. 867
103: 185-188 868
Sánchez-Moreno I, Iturrate L, Martín-Hoyos R, Jimeno ML, Mena M, Bastida A, García-Junceda E 869
(2009) From kinase to cyclase: an unusual example of catalytic promiscuity modulated by metal 870
switching. Chembiochem 10: 225-229 871
Sayed SA, Gadallah MA (2002) Effects of shoot and root application of thiamin on salt-stressed 872
sunflower plants. Plant Growth Regul 36: 71-80 873
Scafaro AP, Haynes PA, Atwell BJ (2010) Physiological and molecular changes in Oryza 874
meridionalis Ng., a heat-tolerant species of wild rice. J Exp Bot 61: 191-202 875
Scheer JB, Mackey AD, Gregory JF 3rd (2005) Activities of hepatic cytosolic and mitochondrial forms 876
of serine hydroxymethyltransferase and hepatic glycine concentration are affected by vitamin B-6 877
intake in rats. J Nutr 135: 233-238 878
Shi H, Xiong L, Stevenson B, Lu T, Zhu JK (2002) The Arabidopsis salt overly sensitive 4 mutants 879
uncover a critical role for vitamin B6 in plant salt tolerance. Plant Cell 14: 575-588 880
Shibata S, Mori Y, Onodera M (1991) Nutritional efficiency of nicotinamide N-oxide and N’-methyl-881
nicotinamide as niacin in rats. Agric Biol Chem 55: 2591-2597 882
Shimizu MM, Mazzafera P (2000) A role for trigonelline during imbibition and germination of coffee 883
seeds. Plant Biol 2: 605-611 884
Smith AG, Croft MT, Moulin M, Webb ME (2007) Plants need their vitamins too. Curr Opin Plant Biol. 885
10: 266-275 886
Stabler SP, Korson M, Jethva R, Allen RH, Kraus JP, Spector EB, Wagner C, Mudd SH (2013) 887
Metabolic profiling of total homocysteine and related compounds in hyperhomocysteinemia: 888
utility and limitations in diagnosing the cause of puzzling thrombophilia in a family. JIMD Rep 11: 889
149-163 890
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

28
Stabler SP, Lindenbaum J, Allen RH (1996) The use of homocysteine and other metabolites in the 891
specific diagnosis of vitamin B-12 deficiency. J Nutr 126): 1266S-1272S 892
Storozhenko S, Navarrete O, Ravanel S, De Brouwer V, Chaerle P, Zhang GF, Bastien O, Lambert 893
W, Rebeille F, Van Der Straeten D (2007) Cytosolic hydroxymethyldihydropterin pyrophospho-894
kinase/dihydropteroate synthase from Arabidopsis thaliana: a specific role in early development 895
and stress response. J Biol Chem 282: 10749-10761 896
Sümegi B, Alkonyi I (1983) Paracatalytic inactivation of pig heart pyruvate dehydrogenase complex. 897
Arch Biochem Biophys 223: 417-424 898
Suzuki Y, Uchida K (1983) Formation of riboflavin glucosides from riboflavin in germinating barley 899
seeds. Agric Biol Chem 47: 1145-1147 900
Tadera K, Arima M, Yoshino S, Yagi F, Kobayashi A (1986) Conversion of pyridoxine into 6-hydroxy-901
pyridoxine by food components, especially ascorbic acid. J Nutr Sci Vitaminol (Tokyo) 32: 267-902
277 903
Tadera K, Yagi F, Kobayashi A (1982) Specificity of a particulate glucosyltransferase in seedlings of 904
Pisum sativum L. which catalyzes the formation of 5'-O-(beta-D-glucopyranosyl)pyridoxine. J 905
Nutr Sci Vitaminol (Tokyo) 28: 359-366 906
Tanou G, Fotopoulos V, Molassiotis A (2012) Priming against environmental challenges and 907
proteomics in plants: Update and agricultural perspectives. Front Plant Sci 3: 216 908
Thimann KV (1963) Plant growth substances; past, present and future. Annu Rev Plant Physiol 14: 909
1-19 910
Titiz O, Tambasco-Studart M, Warzych E, Apel K, Amrhein N, Laloi C, Fitzpatrick TB (2006) PDX1 is 911
essential for vitamin B6 biosynthesis, development and stress tolerance in Arabidopsis. Plant J 912
48: 933-946 913
Treadwell GE, Metzler DE (1972) Photoconversion of riboflavin to lumichrome in plant tissues. Plant 914
Physiol 49: 991-993 915
Tuna AL, Kaya C; Altunlu H, Ashraf M (2013) Mitigation effects of non-enzymatic antioxidants in 916
maize (Zea mays L.) plants under salinity stress. Aust J Crop Sci 7: 1181-1188. 917
Tunc-Ozdemir M, Miller G, Song L, Kim J, Sodek A, Koussevitzky S, Misra AN, Mittler R, Shintani D 918
(2009) Thiamin confers enhanced tolerance to oxidative stress in Arabidopsis. Plant Physiol 151: 919
421-432 920
Ueland PM, Ulvik A, Rios-Avila L, Midttun Ø, Gregory JF (2015) Direct and functional biomarkers of 921
vitamin B6 status. Annu Rev Nutr 35: 33-70 922
Upmeier B, Thomzik JE, Wolfgang Barz W (1988) Enzymatic studies on the reversible synthesis of 923
nicotinic acid-N-glucoside in heterotrophic parsley cell suspension cultures. Z Naturforsch C 43: 924
835-842 925
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

29
Vanderschuren H, Boycheva S, Li KT, Szydlowski N, Gruissem W, Fitzpatrick TB (2013) Strategies 926
for vitamin B6 biofortification of plants: a dual role as a micronutrient and a stress protectant. 927
Front Plant Sci 4: 143 928
Went FW, Bonner J, Warner GC (1938) Aneurin and the rooting of cuttings. Science 87: 170-171 929
Windheuser JJ, Higuchi T (1962) Kinetics of thiamine hydrolysis. J Pharm Sci 51: 354-364 930
Woodward JB, Abeydeera ND, Paul D, Phillips K, Rapala-Kozik M, Freeling M, Begley TP, Ealick 931
SE, McSteen P, Scanlon MJ (2010) A maize thiamine auxotroph is defective in shoot meristem 932
maintenance. Plant Cell 22: 3305-3317 933
Yasumoto K, Iwami K, Tsuji H, Okada J, Mitsuda H (1976) Bound forms of vitamin B6 in cereals and 934
seeds. Vitamins 50: 327-333 935
Yazdani M, Zallot R, Tunc-Ozdemir M, de Crécy-Lagard V, Shintani DK, Hanson AD (2013) Identific-936
ation of the thiamin salvage enzyme thiazole kinase in Arabidopsis and maize. Phytochemistry 937
94: 68-73 938
Yonamine I, Yoshida K, Kido K, Nakagawa A, Nakayama H, Shinmyo A (2004) Overexpression of 939
NtHAL3 genes confers increased levels of proline biosynthesis and the enhancement of salt 940
tolerance in cultured tobacco cells. J Exp Bot 55: 387-395 941
Yoshizumi H, Amachi T (1969) Studies on the bacteria isolated from wine. Part IV. Distribution of the 942
growth factor for a bacterium inducing malo-lactic fermentation. Agric Biol Chem 33: 18-24 943
You J, Chan Z (2015) ROS Regulation during abiotic stress responses in crop plants. Front Plant Sci 944
6: 1092 945
Zallot R, Yazdani M, Goyer A, Ziemak MJ, Guan JC, McCarty DR, de Crécy-Lagard V, Gerdes S, 946
Garrett TJ, Benach J, Hunt JF, Shintani DK, Hanson AD (2014) Salvage of the thiamin pyrim-947
idine moiety by plant TenA proteins lacking an active-site cysteine. Biochem J 463: 145-155 948
Zheng L, Huang F, Narsai R, Wu J, Giraud E, He F, Cheng L, Wang F, Wu P, Whelan J, Shou H 949
(2009) Physiological and transcriptome analysis of iron and phosphorus interaction in rice 950
seedlings. Plant Physiol 151: 262-274 951
Zhu YN, Shi DQ, Ruan MB, Zhang LL, Meng ZH, Liu J, Yang WC (2013) Transcriptome analysis 952
reveals crosstalk of responsive genes to multiple abiotic stresses in cotton (Gossypium hirsutum 953
L.). PLoS One 8: e80218 954
955
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
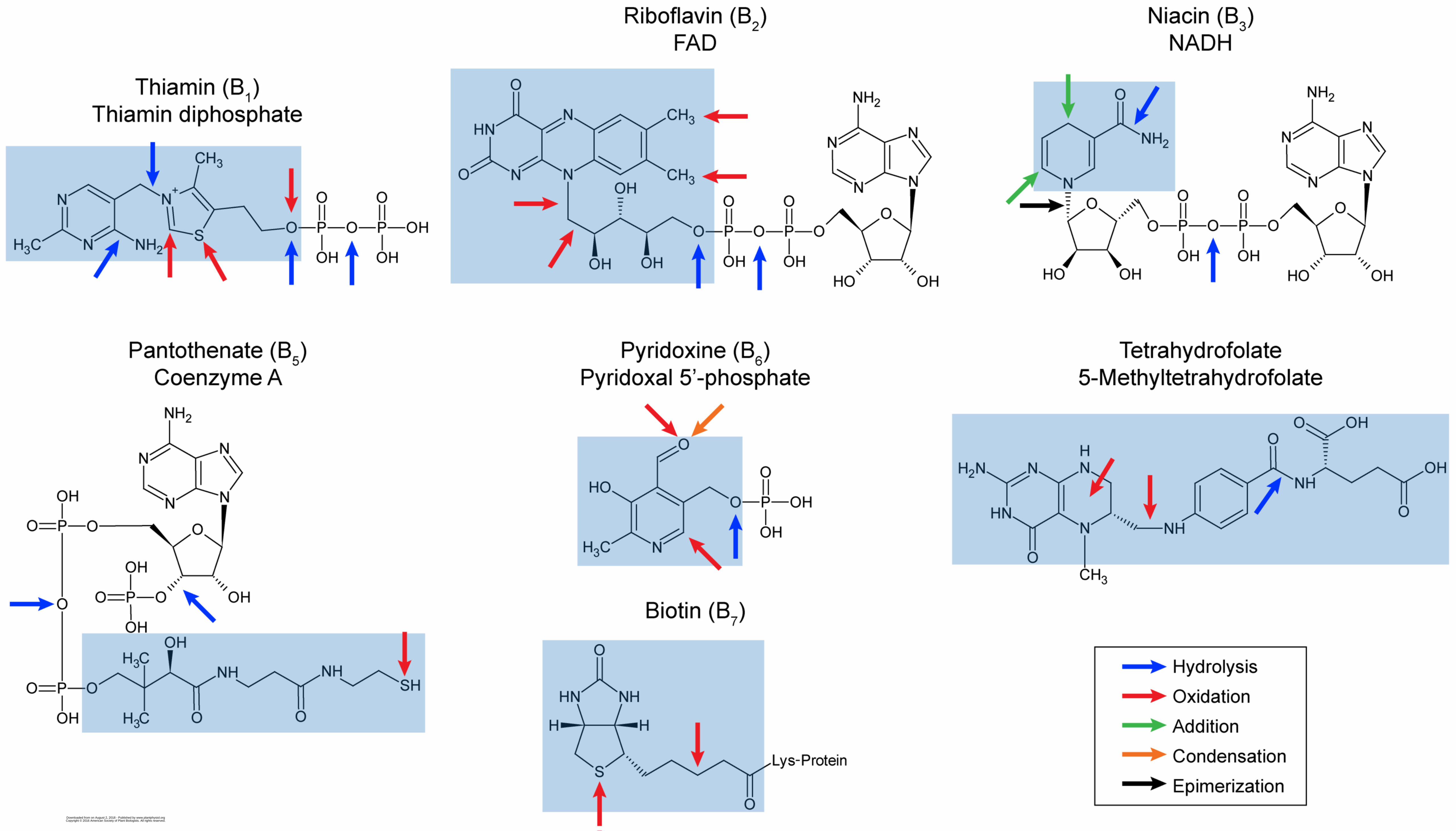
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
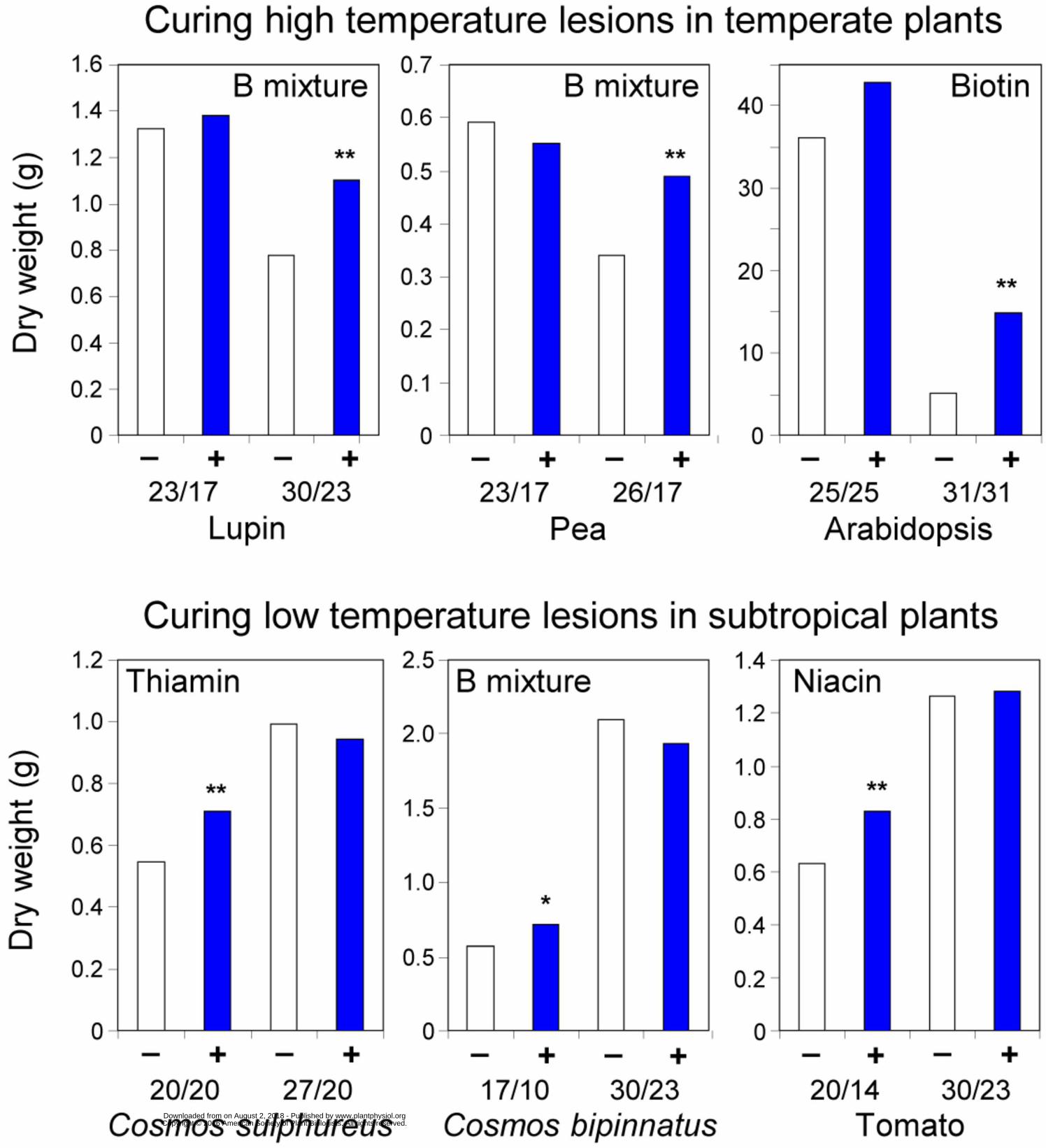
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
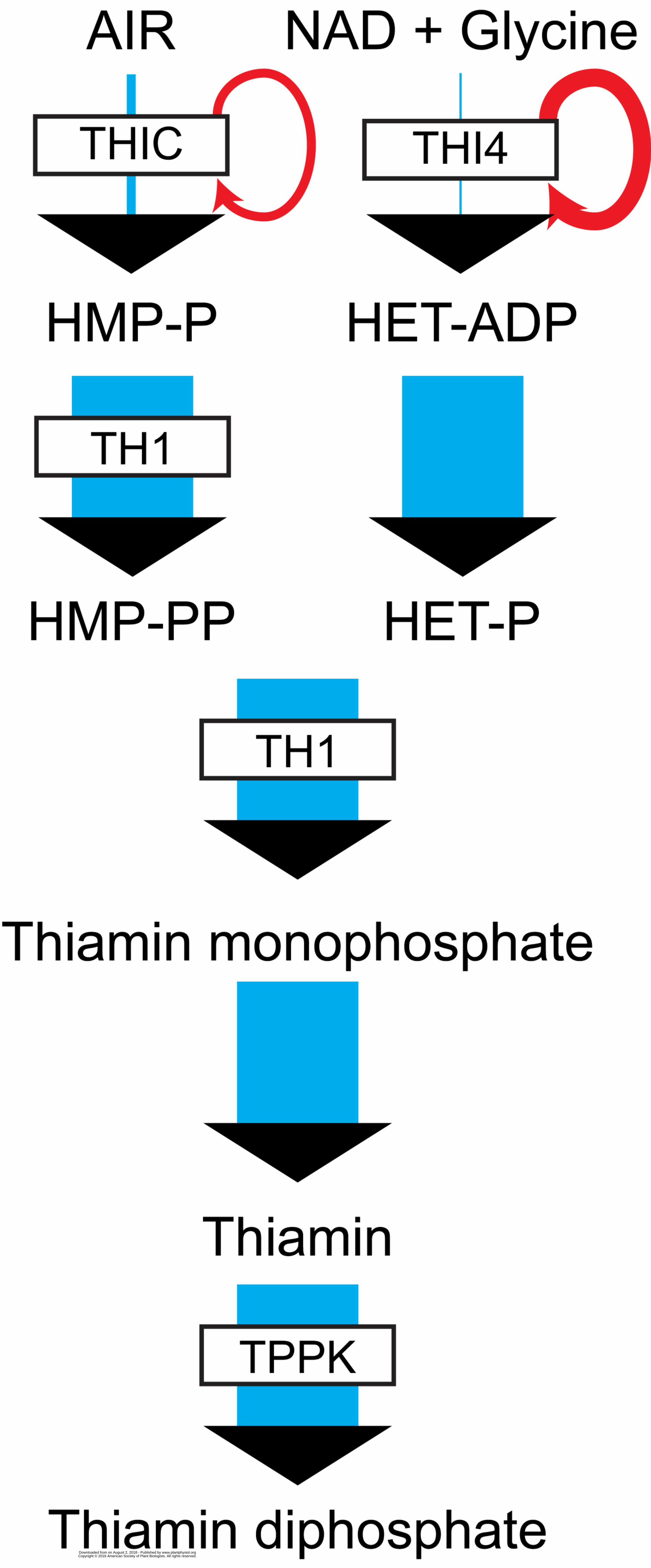
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Parsed CitationsAkhtar TA, Orsomando G, Mehrshahi P, Lara-Núñez A, Bennett MJ, Gregory JF 3rd, Hanson AD (2010) A central role for gamma-glutamyl hydrolases in plant folate homeostasis. Plant J 64: 256-266
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Al-Hakimi A, Hamada AM (2001) Counteraction of salinity stress on wheat plants by grain soaking in ascorbic acid, thiamin orsodium salicylate. Biol Plantarum 44: 253-261
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Amachi T, Imamoto S, Yoshizumi H (1971) A growth factor for malo-lactic fermentation bacteria. Part II. Structure and synthesis of anovel pantothenic acid derivative isolated from tomato juice. Agric Biol Chem 35: 1222-1230
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ariaey-Nejad MR, Balaghi M, Baker EM, Sauberlich HE (1970) Thiamin metabolism in man. Am J Clin Nutr 23: 764-778Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Asensi-Fabado MA, Munné-Bosch S (2010) Vitamins in plants: occurrence, biosynthesis and antioxidant function. Trends Plant Sci15: 582-592
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ashihara H, Yin Y, Deng WW, Watanabe S (2010) Pyridine salvage and nicotinic acid conjugate synthesis in leaves of mangrovespecies. Phytochemistry 71: 47-53
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Baggott JE (2000) Hydrolysis of 5,10-methenyltetrahydrofolate to 5-formyltetrahydrofolate at pH 2.5 to 4.5. Biochemistry 39: 14647-14653
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Blum A (2014) Genomics for drought resistance - getting down to earth. Funct Plant Biol 41: 1191-1198Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bonner J (1943) Effects of application of thiamin to Cosmos. Botan Gaz 104: 475-479Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bonner J (1957) The chemical cure of climatic lesions. Eng Sci 20: 28-30Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bonner J, Bonner H (1948) The B vitamins as plant hormones. Vitam Horm 6: 225-275Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bozzo GG, Basset GJ, Naponelli V, Noiriel A, Gregory JF 3rd, Hanson AD (2008) Characterization of the folate salvage enzyme p-aminobenzoylglutamate hydrolase in plants. Phytochemistry 69: 29-37
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Broderick JB, Duffus BR, Duschene KS, Shepard EM (2014) Radical S-adenosylmethionine enzymes. Chem Rev 114: 4229-4317Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bunik VI, Schloss JV, Pinto JT, Gibson GE, Cooper AJ (2007) Enzyme-catalyzed side reactions with molecular oxygen maycontribute to cell signaling and neurodegenerative diseases. Neurochem Res 32: 871-891
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Chatterjee A, Abeydeera ND, Bale S, Pai PJ, Dorrestein PC, Russell DH, Ealick SE, Begley TP
(2011) Saccharomyces cerevisiae THI4p is a suicide thiamine thiazole synthase. Nature 478: 542-546
Chen H, Xiong L (2005) Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidativestresses. Plant J 44: 396-408
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choe E, Huang R, Min DB (2005) Chemical reactions and stability of riboflavin in foods. J Food Sci 70: R28-R36Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Clifford AJ, Bills ND, Peerson JM, Müller HG, Burk GE, Rich KD (1993) A depletion-repletion folate bioassay based on growth andtissue folate concentrations of rats. J Nutr 123: 926-932
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Coburn SP (1990) Location and turnover of vitamin B6 pools and vitamin B6 requirements of humans. Ann N Y Acad Sci 585: 76-85Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dalling DK, Grant DM, Horton WJ, Sagers RD (1976) Carbon 13 NMR study of nonenzymatic reactions of pyridoxal 5'-phosphatewith selected amino acids and of related reactions. J Biol Chem 251: 7661-7668
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dalvi RR, Sauberlich HE, Neal RA (1974) An examination of the metabolism of thiamin by rat liver alcohol dehydrogenase. J Nutr104: 1476-1483
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dehaye L, Duval M, Viguier D, Yaxley J, Job D (1997) Cloning and expression of the pea gene encoding SBP65, a seed-specificbiotinylated protein. Plant Mol Biol 35: 605-621
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dhindsa RS, Cleland RE (1975) Water stress and protein synthesis 1. Differential inhibition of protein synthesis. Plant Physiol 55:778-781
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Díaz de la Garza R, Quinlivan EP, Klaus SM, Basset GJ, Gregory JF 3rd, Hanson AD (2004) Folate biofortification in tomatoes byengineering the pteridine branch of folate synthesis. Proc Natl Acad Sci USA 101: 13720-13725
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dong W, Stockwell VO, Goyer A (2015) Enhancement of thiamin content in Arabidopsis thaliana by metabolic engineering. PlantCell Physiol 56: 2285-2296
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dwivedi BK, Arnold RG (1973) Chemistry of thiamine degradation in food products and model systems: a review. J Agric FoodChem 21: 54-60
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Espinosa-Ruiz A, Belles JM, Serrano R, Culianez-MacIa FA (1999) Arabidopsis thaliana AtHAL 3: a flavoprotein related to salt andosmotic tolerance and plant growth. Plant J 20: 529-539
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Eudes A, Bozzo GG, Waller JC, Naponelli V, Lim EK, Bowles DJ, Gregory JF 3rd, Hanson AD (2008) Metabolism of the folateprecursor p-aminobenzoate in plants: glucose ester formation and vacuolar storage. J Biol Chem 283: 15451-15459
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Everse J, Cooper Zoll E, Kahan L, Kaplan NO (1971) Addition products of diphosphopyridine nucleotides with substrates of www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

pyridine nucleotide-linked dehydrogenases. Bioorg Chem 1: 207-233Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Everse J, Everse KE, Kaplan NO (1975) The pyridine nucleosidases from Bacillus subtilis and Neurospora crassa. Isolation andstructural properties. Arch Biochem Biophys 169: 702-713
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ferreira S, Hjernø K, Larsen M, Wingsle G, Larsen P, Fey S, Roepstorff P, Salomé Pais M (2006) Proteome profiling of Populuseuphratica Oliv. upon heat stress. Ann Bot 98: 361-377
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fitzpatrick TB (2011) Vitamin B6 in plants: more than meets the eye. Adv Bot Res 59: 1-38Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fitzpatrick TB, Basset GJ, Borel P, Carrari F, DellaPenna D, Fraser PD, Hellmann H, Osorio S, Rothan C, Valpuesta V, Caris-VeyratC, Fernie AR (2012) Vitamin deficiencies in humans: can plant science help? Plant Cell 24: 395-414
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Foyer CH, Noctor G (2011) Ascorbate and glutathione: the heart of the redox hub. Plant Physiol 155: 2-18Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fraiz FJ, Pinto RM, Costas MJ, Aavalos M, Canales J, Cabezas A, Cameselle JC (1998) Enzymic formation of riboflavin 4',5'-cyclicphosphate from FAD: evidence for a specific low-Km FMN cyclase in rat liver. Biochem J 330: 881-888
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Frelin O, Huang L, Hasnain G, Jeffryes JG, Ziemak MJ, Rocca JR, Wang B, Rice J, Roje S, Yurgel SN, Gregory JF 3rd, Edison AS,Henry CS, de Crécy-Lagard V, Hanson AD (2015) A directed-overflow and damage-control N-glycosidase in riboflavin biosynthesis.Biochem J 466: 137-145
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gerdes S, Lerma-Ortiz C, Frelin O, Seaver SM, Henry CS, de Crécy-Lagard V, Hanson AD (2012) Plant B vitamin pathways and theircompartmentation: a guide for the perplexed. J Exp Bot 63: 5379-5395
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Good AG, Zaplachinski ST (1994) The effects of drought stress on free amino acid accumulation and protein synthesis in Brassicanapus. Physiol Plant 90: 9-14
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Goyer A, Collakova E, Díaz de la Garza R, Quinlivan EP, Williamson J, Gregory JF 3rd, Shachar-Hill Y, Hanson AD (2005) 5-Formyltetrahydrofolate is an inhibitory but well tolerated metabolite in Arabidopsis leaves. J Biol Chem 280: 26137-26142
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Goyer A, Hasnain G, Frelin O, Ralat MA, Gregory JF 3rd, Hanson AD (2013) A cross-kingdom Nudix enzyme that pre-empts damagein thiamin metabolism. Biochem J 454: 533-542
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Green R (2008) Indicators for assessing folate and vitamin B12 status and for monitoring the efficacy of intervention strategies.Food Nutr Bull 29: S52-S63
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gregory JF 3rd (1989) Chemical and nutritional aspects of folate research: analytical procedures, methods of folate synthesis,stability, and bioavailability of dietary folates. Adv Food Nutr Res 33: 1-101
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Gregory JF 3rd (1998) Nutritional properties and significance of vitamin glycosides. Annu Rev Nutr 18: 277-296Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gregory JF 3rd, Quinlivan EP (2002) In vivo kinetics of folate metabolism. Annu Rev Nutr 22: 199-220Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Guenther BD, Sheppard CA, Tran P, Rozen R, Matthews RG, Ludwig ML (1999) The structure and properties ofmethylenetetrahydrofolate reductase from Escherichia coli suggest how folate ameliorates human hyperhomocysteinemia. NatStruct Biol 6: 359-365
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hall CA (1964) Long term excretion of Co57-vitamin B12 and turnover within plasma. Am J Clin Nutr 14: 156-162Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hanson AD, Gregory JF 3rd (2011) Folate biosynthesis, turnover, and transport in plants. Annu Rev Plant Biol 62: 105-125Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hanson AD, Henry CS, Fiehn O, de Crécy-Lagard V (2016) Metabolite damage and metabolite damage control in plants. Annu RevPlant Biol 67: 131-152
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hasnain G, Frelin O, Roje S, Ellens KW, Ali K, Guan JC, Garrett TJ, de Crecy-Lagard V, Gregory JF 3rd, McCarty DR, Hanson AD(2013) Identification and characterization of the missing pyrimidine reductase in the plant riboflavin biosynthesis pathway. PlantPhysiol 161: 48-56
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Higa A, Khandakar J, Mori Y, Kitamura Y (2012) Increased de novo riboflavin synthesis and hydrolysis of FMN are involved inriboflavin secretion from Hyoscyamus albus hairy roots under iron deficiency. Plant Physiol Biochem 58: 166-173
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Huang L, Khusnutdinova A, Nocek B, Brown G, Xu X, Cui H, Petit P, Flick R, Zallot R, Balmant K, Ziemak MJ, Shanklin J, de Crécy-Lagard V, Fiehn O, Gregory JF 3rd, Joachimiak A, Savchenko A, Yakunin AF, Hanson AD (2016) A family of metal-dependentphosphatases implicated in metabolite damage-control. Nat Chem Biol 12: 621-627
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Huang S, Zhang J, Wang L, Huang L (2013) Effect of abiotic stress on the abundance of different vitamin B6 vitamers in tobaccoplants. Plant Physiol Biochem 66: 63-67
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Izumi Y, Tani Y, Ogata K (1973) The conversion of bisnorbiotin and bisnordethiobiotin to biotin and dethiobiotin, respectively, bymicroorganisms. Biochim Biophys Acta 326: 485-488
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jeanguenin L, Lara-Núñez A, Rodionov DA, Osterman AL, Komarova NY, Rentsch D, Gregory JF 3rd, Hanson AD (2012)Comparative genomics and functional analysis of the NiaP family uncover nicotinate transporters from bacteria, plants, andmammals. Funct Integr Genomics 12: 25-34
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jenkins AH, Schyns G, Potot S, Sun G, Begley TP (2007) A new thiamin salvage pathway. Nat Chem Biol 3: 492-497Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jurgenson CT, Begley TP, Ealick SE (2009) The structural and biochemical foundations of thiamin biosynthesis. Annu RevBiochem 78: 569-603
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kaya C, Ashraf M, Sonmez O, Tuna AL, Polat T, Aydemir S (2015) Exogenous application of thiamin promotes growth andantioxidative defense system at initial phases of development in salt-stressed plants of two maize cultivars differing in salinitytolerance. Acta Physiol Plant 37: 1741
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ketellapper HJ (1963) Temperature-induced chemical defects in higher plants. Plant Physiol 38: 175-179Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Khozaei M, Fisk S, Lawson T, Gibon Y, Sulpice R, Stitt M, Lefebvre SC, Raines CA (2015)
Overexpression of plastid transketolase in tobacco results in a thiamine auxotrophic phenotype. Plant Cell 27: 432-447
Kilian J, Whitehead D, Horak J, Wanke D, Weinl S, Batistic O, D'Angelo C, Bornberg-Bauer E, Kudla J, Harter K (2007) TheAtGenExpress global stress expression data set: protocols, evaluation and model data analysis of UV-B light, drought and coldstress responses. Plant J 50: 347-363
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kohler RE (1975) The history of biochemistry: A survey. J Hist Biol 8: 275-318Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kupke T, Hernandez-Acosta P, Steinbacher S, Culianez-Macia FA (2001) Arabidopsis thaliana flavoprotein AtHAL3a catalyzes thedecarboxylation of 4'-phosphopantothenoylcysteine to 4'-phosphopantetheine, a key step in coenzyme A biosynthesis. J BiolChem 276: 19190-19196
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Langridge J, Griffing B (1959) A study of high temperature lesions in Arabidopsis thaliana. Aust J Biol Sci 12: 117-135Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lerma-Ortiz C, Jeffryes JG, Cooper AJ, Niehaus TD, Thamm AM, Frelin O, Aunins T, Fiehn O, de Crécy-Lagard V, Henry CS,Hanson AD (2016) 'Nothing of chemistry disappears in biology': The Top 30 damage-prone endogenous metabolites. Biochem SocTrans 44: 961-971
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lesgards J-F, Lehucher-Michel M-P, Vidal N, Stocker P (2015) Assessment of antioxidative activity of lipid- and water-solublevitamins in human whole blood. comparative analysis between a biological test and chemical methods. Int J Vitam Nutr Res 75: 11-18
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li W, Zhang F, Chang Y, Zhao T, Schranz ME, Wang G (2015) Nicotinate O-glucosylation is an evolutionarily metabolic traitimportant for seed germination under stress conditions in Arabidopsis thaliana. Plant Cell 27: 1907-1924
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Linster CL, Van Schaftingen E, Hanson AD (2013) Metabolite damage and its repair or pre-emption. Nat Chem Biol 9: 72-80Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu JX, Howell SH (2010) Endoplasmic reticulum protein quality control and its relationship to environmental stress responses inplants. Plant Cell 22: 2930-2942
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Loizeau K, De Brouwer V, Gambonnet B, Yu A, Renou JP, Van Der Straeten D, Lambert WE, Rébeillé F, Ravanel S (2008) Agenome-wide and metabolic analysis determined the adaptive response of Arabidopsis cells to folate depletion induced bymethotrexate. Plant Physiol 148: 2083-2095
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Ludwig-Müller J (2011) Auxin conjugates: their role for plant development and in the evolution of land plants. J Exp Bot 62: 1757-1773
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lukienko PI, Mel'nichenko NG, Zverinskii IV, Zabrodskaya SV (2000) Antioxidant properties of thiamin. Bull Exp Biol Med 130: 874-876
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mackey AD, Lieu SO, Carman C, Gregory JF 3rd (2003) Hydrolytic activity toward pyridoxine-5'-beta-D-glucoside in rat intestinalmucosa is not increased by vitamin B-6 deficiency: effect of basal
diet composition and pyridoxine intake. J Nutr 133: 1362-1367
Marbaix AY, Noël G, Detroux AM, Vertommen D, Van Schaftingen E, Linster CL (2011) Extremely conserved ATP- or ADP-dependent enzymatic system for nicotinamide nucleotide repair. J Biol Chem 286: 41246-41252
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Matsui A, Yin Y, Yamanaka K, Iwasaki M, Ashihara H (2007) Metabolic fate of nicotinamide in higher plants. Physiol Plant 131: 191-200
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
McCourt JA, Nixon PF, Duggleby RG (2006) Thiamin nutrition and catalysis-induced instability of thiamin diphosphate. Br J Nutr 96:636-638
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
McHale NA, Hanson KR, Zelitch I (1988) A nuclear mutation in Nicotiana sylvestris causing a thiamine-reversible defect in synthesisof chloroplast pigments. Plant Physiol 88: 930-935
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Melville DB (1954) Biotin sulfoxide. J Biol Chem 208: 495-501Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Menegus F, Lilliu I, Scaglioni L (2002) A unique pantoyllactone glycoside system is activated in rice seedlings developingaerobically in the dark. Physiol Plant 116: 299-307
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mills PB, Struys E, Jakobs C, Plecko B, Baxter P, Baumgartner M, Willemsen MA, Omran H, Tacke U, Uhlenberg B, Weschke B,Clayton PT (2006) Mutations in antiquitin in individuals with pyridoxine-dependent seizures. Nat Med 12: 307-309
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mizuno K, Matsuzaki M, Kanazawa S, Tokiwano T, Yoshizawa Y, Kato M (2014) Conversion of nicotinic acid to trigonelline iscatalyzed by N-methyltransferase belonged to motif B' methyltransferase family in Coffea arabica. Biochem Biophys Res Commun452: 1060-1066
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Moccand C, Boycheva S, Surriabre P, Tambasco-Studart M, Raschke M, Kaufmann M, Fitzpatrick TB (2014) The pseudoenzymePDX1.2 boosts vitamin B6 biosynthesis under heat and oxidative stress in Arabidopsis. J Biol Chem 289: 8203-8216
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mohammadi PP, Moieni A, Hiraga S, Komatsu S (2012) Organ-specific proteomic analysis of drought-stressed soybean seedlings. JProteomics 75: 1906-1923
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Navarrete O, Van Daele J, Stove C, Lambert W, Van Der Straeten D, Storozhenko S (2012) A folate independent role for cytosolicHPPK/DHPS upon stress in Arabidopsis thaliana. Phytochemistry 73: 23-33
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nelson CJ, Alexova R, Jacoby RP, Millar AH (2014) Protein with high turnover rate in barley leaves estimated by proteome analysiscombined with in planta isotope labeling. Plant Physiol 166: 91-108
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nikiforova V, Freitag J, Kempa S, Adamik M, Hesse H, Hoefgen R (2003) Transcriptome analysis of sulfur depletion in Arabidopsisthaliana: interlacing of biosynthetic pathways provides response specificity. Plant J 33: 633-650
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Noiriel A, Naponelli V, Gregory JF 3rd, Hanson AD (2007) Pterin and folate salvage. Plants andPubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Escherichia coli lack capacity to reduce oxidized pterins. Plant Physiol 143: 1101-1109
Ogawa T, Yoshimura K, Miyake H, Ishikawa K, Ito D, Tanabe N, Shigeoka S (2008) Molecular characterization of organelle-typeNudix hydrolases in Arabidopsis. Plant Physiol 148: 1412-1424
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ohkawa H, Ohishi N, Yagi K (1983) New metabolites of riboflavin appear in human urine. J Biol Chem 258: 5623-5628Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Oppenheimer NJ, Kaplan NO (1975) The alpha beta epimerization of reduced nicotinamide adenine dinucleotide. Arch BiochemBiophys 166: 526-535
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Orsomando G, de la Garza RD, Green BJ, Peng M, Rea PA, Ryan TJ, Gregory JF 3rd, Hanson AD (2005) Plant gamma-glutamylhydrolases and folate polyglutamates: characterization, compartmentation, and co-occurrence in vacuoles. J Biol Chem 280: 28877-28884
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ouyang M, Ma J, Zou M, Guo J, Wang L, Lu C, Zhang L (2010) The photosensitive phs1 mutant is impaired in the riboflavinbiogenesis pathway. J Plant Physiol 167: 1466-1476
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Paizs C, Diemer T, Rétey J (2008) The putative coenzyme B12-dependent methylmalonyl-CoA mutase from potatoes is aphosphatase. Bioorg Chem 36: 261-264
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Palmer LD, Downs DM (2013) The thiamine biosynthetic enzyme ThiC catalyzes multiple turnovers and is inhibited by S-adenosylmethionine (AdoMet) metabolites. J Biol Chem 288: 30693-30699
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Piedrafita G, Keller MA, Ralser M (2015) The impact of non-enzymatic reactions and enzyme promiscuity on cellular metabolismduring (oxidative) stress conditions. Biomolecules 5: 2101-2122
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Piotrowska A, Bajguz A (2011) Conjugates of abscisic acid, brassinosteroids, ethylene, gibberellins, and jasmonates.Phytochemistry 72: 2097-2112
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pinto RM, Fraiz FJ, Cabezas A, Avalos M, Canales J, Costas MJ, Cameselle JC (1999) Preparation of riboflavin 4',5'-cyclicphosphate by incubation of flavin-adenine dinucleotide with Mn2+ in the absence of riboflavin 5'-phosphate cyclase. Anal Biochem268: 409-411
Pubmed: Author and Title www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

CrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Plaut Z, Edelstein M, Ben-Hur M (2013) Overcoming salinity barriers to crop production using traditional methods. Crit Rev PlantSci 32: 250-291
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Plaxton WC, Tran HT (2011) Metabolic adaptations of phosphate-starved plants. Plant Physiol 156: 1006-1015Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pourcel L, Moulin M, Fitzpatrick TB (2013) Examining strategies to facilitate vitamin B1 biofortification of plants by geneticengineering. Front Plant Sci 4: 160
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Quinlivan EP, Roje S, Basset G, Shachar-Hill Y, Gregory JF 3rd, Hanson AD (2003) The folate precursor p-aminobenzoate isreversibly converted to its glucose ester in the plant cytosol. J Biol
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chem 278: 20731-20737
Rains TM, Emmert JL, Baker DH, Shay NF (1997) Minimum thiamin requirement of weanling Sprague-Dawley outbred rats. J Nutr127: 167-170
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rapala-Kozik M, Golda A, Kujda M (2009) Enzymes that control the thiamine diphosphate pool in plant tissues. Properties ofthiamine pyrophosphokinase and thiamine-(di)phosphate phosphatase purified from Zea mays seedlings. Plant Physiol Biochem47: 237-242
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rapala-Kozik M, Kowalska E, Ostrowska K (2008) Modulation of thiamine metabolism in Zea mays seedlings under conditions ofabiotic stress. J Exp Bot 59: 4133-4143
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rapala-Kozik M, Wolak N, Kujda M, Banas AK (2012) The upregulation of thiamine (vitamin B1) biosynthesis in Arabidopsis thalianaseedlings under salt and osmotic stress conditions is mediated by abscisic acid at the early stages of this stress response. BMCPlant Biol 12: 2
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Raschke M, Boycheva S, Crèvecoeur M, Nunes-Nesi A, Witt S, Fernie AR, Amrhein N, Fitzpatrick TB (2011) Enhanced levels ofvitamin B6 increase aerial organ size and positively affect stress tolerance in Arabidopsis. Plant J 66: 414-432
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rawat R, Sandoval FJ, Wei Z, Winkler R, Roje S (2011) An FMN hydrolase of the haloacid dehalogenase superfamily is active inplant chloroplasts. J Biol Chem 286: 42091-42098
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ristilä M, Strid H, Eriksson LA, Strid A, Sävenstrand H (2011) The role of the pyridoxine (vitamin B6) biosynthesis enzyme PDX1 inultraviolet-B radiation responses in plants. . Plant Physiol Biochem 49: 284-292
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rueschhoff EE, Gillikin JW, Sederoff HW, Daub ME (2013) The SOS4 pyridoxal kinase is required for maintenance of vitamin B6-mediated processes in chloroplasts. Plant Physiol Biochem 63: 281-291
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sakuragi T, Kummerow FA (1960) Utilization of pyridoxine N-oxide by rats. Proc Soc Exp Biol Med. 103: 185-188Pubmed: Author and TitleCrossRef: Author and Title www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Sánchez-Moreno I, Iturrate L, Martín-Hoyos R, Jimeno ML, Mena M, Bastida A, García-Junceda E (2009) From kinase to cyclase: anunusual example of catalytic promiscuity modulated by metal switching. Chembiochem 10: 225-229
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sayed SA, Gadallah MA (2002) Effects of shoot and root application of thiamin on salt-stressed sunflower plants. Plant GrowthRegul 36: 71-80
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Scafaro AP, Haynes PA, Atwell BJ (2010) Physiological and molecular changes in Oryza meridionalis Ng., a heat-tolerant species ofwild rice. J Exp Bot 61: 191-202
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Scheer JB, Mackey AD, Gregory JF 3rd (2005) Activities of hepatic cytosolic and mitochondrial forms of serinehydroxymethyltransferase and hepatic glycine concentration are affected by vitamin B-6 intake in rats. J Nutr 135: 233-238
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shi H, Xiong L, Stevenson B, Lu T, Zhu JK (2002) The Arabidopsis salt overly sensitive 4 mutants uncover a critical role for vitaminB6 in plant salt tolerance. Plant Cell 14: 575-588
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shibata S, Mori Y, Onodera M (1991) Nutritional efficiency of nicotinamide N-oxide and N'-methylnicotinamide as niacin in rats.Agric Biol Chem 55: 2591-2597
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shimizu MM, Mazzafera P (2000) A role for trigonelline during imbibition and germination of coffee seeds. Plant Biol 2: 605-611Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Smith AG, Croft MT, Moulin M, Webb ME (2007) Plants need their vitamins too. Curr Opin Plant Biol. 10: 266-275Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stabler SP, Korson M, Jethva R, Allen RH, Kraus JP, Spector EB, Wagner C, Mudd SH (2013) Metabolic profiling of totalhomocysteine and related compounds in hyperhomocysteinemia: utility and limitations in diagnosing the cause of puzzlingthrombophilia in a family. JIMD Rep 11: 149-163
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stabler SP, Lindenbaum J, Allen RH (1996) The use of homocysteine and other metabolites in the specific diagnosis of vitamin B-12 deficiency. J Nutr 126): 1266S-1272S
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Storozhenko S, Navarrete O, Ravanel S, De Brouwer V, Chaerle P, Zhang GF, Bastien O, Lambert W, Rebeille F, Van Der StraetenD (2007) Cytosolic hydroxymethyldihydropterin pyrophosphokinase/dihydropteroate synthase from Arabidopsis thaliana: a specificrole in early development and stress response. J Biol Chem 282: 10749-10761
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sümegi B, Alkonyi I (1983) Paracatalytic inactivation of pig heart pyruvate dehydrogenase complex. Arch Biochem Biophys 223:417-424
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Suzuki Y, Uchida K (1983) Formation of riboflavin glucosides from riboflavin in germinating barley seeds. Agric Biol Chem 47: 1145-1147
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tadera K, Arima M, Yoshino S, Yagi F, Kobayashi A (1986) Conversion of pyridoxine into 6-hydroxypyridoxine by food components, www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

especially ascorbic acid. J Nutr Sci Vitaminol (Tokyo) 32: 267-277Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tadera K, Yagi F, Kobayashi A (1982) Specificity of a particulate glucosyltransferase in seedlings of Pisum sativum L. whichcatalyzes the formation of 5'-O-(beta-D-glucopyranosyl)pyridoxine. J Nutr Sci Vitaminol (Tokyo) 28: 359-366
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tanou G, Fotopoulos V, Molassiotis A (2012) Priming against environmental challenges and proteomics in plants: Update andagricultural perspectives. Front Plant Sci 3: 216
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Thimann KV (1963) Plant growth substances; past, present and future. Annu Rev Plant Physiol 14: 1-19Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Titiz O, Tambasco-Studart M, Warzych E, Apel K, Amrhein N, Laloi C, Fitzpatrick TB (2006) PDX1 is essential for vitamin B6biosynthesis, development and stress tolerance in Arabidopsis. Plant J 48: 933-946
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Treadwell GE, Metzler DE (1972) Photoconversion of riboflavin to lumichrome in plant tissues. Plant Physiol 49: 991-993Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tuna AL, Kaya C; Altunlu H, Ashraf M (2013) Mitigation effects of non-enzymatic antioxidants in maize (Zea mays L.) plants undersalinity stress. Aust J Crop Sci 7: 1181-1188.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tunc-Ozdemir M, Miller G, Song L, Kim J, Sodek A, Koussevitzky S, Misra AN, Mittler R, Shintani D
(2009) Thiamin confers enhanced tolerance to oxidative stress in Arabidopsis. Plant Physiol 151: 421-432
Ueland PM, Ulvik A, Rios-Avila L, Midttun Ø, Gregory JF (2015) Direct and functional biomarkers of vitamin B6 status. Annu RevNutr 35: 33-70
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Upmeier B, Thomzik JE, Wolfgang Barz W (1988) Enzymatic studies on the reversible synthesis of nicotinic acid-N-glucoside inheterotrophic parsley cell suspension cultures. Z Naturforsch C 43: 835-842
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vanderschuren H, Boycheva S, Li KT, Szydlowski N, Gruissem W, Fitzpatrick TB (2013) Strategies for vitamin B6 biofortification ofplants: a dual role as a micronutrient and a stress protectant. Front Plant Sci 4: 143
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Went FW, Bonner J, Warner GC (1938) Aneurin and the rooting of cuttings. Science 87: 170-171Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Windheuser JJ, Higuchi T (1962) Kinetics of thiamine hydrolysis. J Pharm Sci 51: 354-364Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Woodward JB, Abeydeera ND, Paul D, Phillips K, Rapala-Kozik M, Freeling M, Begley TP, Ealick SE, McSteen P, Scanlon MJ (2010)A maize thiamine auxotroph is defective in shoot meristem maintenance. Plant Cell 22: 3305-3317
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yasumoto K, Iwami K, Tsuji H, Okada J, Mitsuda H (1976) Bound forms of vitamin B6 in cereals and seeds. Vitamins 50: 327-333Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Yazdani M, Zallot R, Tunc-Ozdemir M, de Crécy-Lagard V, Shintani DK, Hanson AD (2013) Identification of the thiamin salvageenzyme thiazole kinase in Arabidopsis and maize. Phytochemistry 94: 68-73
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yonamine I, Yoshida K, Kido K, Nakagawa A, Nakayama H, Shinmyo A (2004) Overexpression of NtHAL3 genes confers increasedlevels of proline biosynthesis and the enhancement of salt tolerance in cultured tobacco cells. J Exp Bot 55: 387-395
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoshizumi H, Amachi T (1969) Studies on the bacteria isolated from wine. Part IV. Distribution of the growth factor for a bacteriuminducing malo-lactic fermentation. Agric Biol Chem 33: 18-24
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
You J, Chan Z (2015) ROS Regulation during abiotic stress responses in crop plants. Front Plant Sci 6: 1092Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zallot R, Yazdani M, Goyer A, Ziemak MJ, Guan JC, McCarty DR, de Crécy-Lagard V, Gerdes S, Garrett TJ, Benach J, Hunt JF,Shintani DK, Hanson AD (2014) Salvage of the thiamin pyrimidine moiety by plant TenA proteins lacking an active-site cysteine.Biochem J 463: 145-155
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zheng L, Huang F, Narsai R, Wu J, Giraud E, He F, Cheng L, Wang F, Wu P, Whelan J, Shou H (2009) Physiological andtranscriptome analysis of iron and phosphorus interaction in rice seedlings. Plant Physiol 151: 262-274
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhu YN, Shi DQ, Ruan MB, Zhang LL, Meng ZH, Liu J, Yang WC (2013) Transcriptome analysis reveals crosstalk of responsivegenes to multiple abiotic stresses in cotton (Gossypium hirsutum L.). PLoS One 8: e80218
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon August 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.


















