
Dissertação apresentada para cumprimento dos requisitos ... (versão final).pdf · do grau de...
119
Dissertação apresentada para cumprimento dos requisitos necessários à obtenção do grau de Mestrado em Arqueologia, realizada sob a orientação científica da Professora Doutora Catarina Tente e coorientaçãodo do técnico superior do Laboratório de Arqueociências da DGPC Carlos Pimenta. 1 1 Versão corrigida e melhora após defesa pública.
Transcript of Dissertação apresentada para cumprimento dos requisitos ... (versão final).pdf · do grau de...

Dissertação apresentada para cumprimento dos requisitos necessários à obtenção
do grau de Mestrado em Arqueologia, realizada sob a orientação científica da
Professora Doutora Catarina Tente e coorientaçãodo do técnico superior do Laboratório
de Arqueociências da DGPC Carlos Pimenta.1
1 Versão corrigida e melhora após defesa pública.

Aos meus pais que sempre me deram liberdade de escolha.

AGRADECIMENTOS
Para a realização desta dissertação de mestrado muitas pessoas contribuíram, às
quais não posso deixar de dizer o meu muito obrigada.
A Zooarqueologia foi-me apresentada pela primeira vez do ponto de vista
prático durante as escavações arqueológicas realizadas em São Gens (Celorico da Beira,
Guarda). Isto tenho de agradecer à Sílvia Casimiro e à Prof. Dra. Catarina Tente, que
vendo o meu interesse por esta área, me propôs realizar um estágio curricular no LARC,
e que desde então e até à realização desta dissertação, como orientadora científica, se
mostrou sempre prestável, portadora de bons conselhos, pronta a preparar e a construir
bons investigadores.
Na impossibilidade de trabalhar com macrofaunas, tive o meu primeiro contacto
com as microfaunas durante o estágio curricular realizado no LARC, no último ano da
Licenciatura.
Neste contexto tenho de agradecer ao Carlos Pimenta, responsável pelo meu
estágio e posteriormente meu coorientador, que desde este primeiro momento me
recebeu sempre com grande simpatia, entusiasmo e paciência por me abrir as portas
para o estudo das microfaunas, partilhando comigo algum do seu conhecimento, sem a
sua orientação a realização desta dissertação não teria sido possível. Tenho ainda de
agradecer ao LARC por me ter recebido em ambas as ocasiões com grande à vontade,
permitindo a utilização dos equipamentos que tornaram possível o estudo do material
aqui apresentado, bem como a todos os seus investigadores, que sempre me trataram de
forma cordial ao longo do moroso período de estudo das microfaunas, e, de uma forma
geral à DGPC.
Neste seguimento tenho de agradecer ao Prof. Dr. António Faustino de
Carvalho, pela disponibilidade dos materiais ostelógicos correspondentes às
microfaunas provenientes da Pena d´Água aqui estudados e pelo facto de se ter
mostrado sempre disponível para me ajudar e esclarecer no que fosse necessário do
ponto de vista arqueológico, como arqueólogo responsável pela escavação, e logístico
durante a preparação das amostras para estudo.
À Dra. Liliana Póvoas, investigadora do Museu Nacional de História Natural e
da Ciência, a quem foi inicialmente pedido para estudar este material, por me ter cedido,
sem reservas, as amostras ainda por estudar, por me ter instruído nas práticas

laboratoriais necessárias para o tratamento deste tipo de amostras e por se ter mostrado
disponível para me auxiliar sempre que necessário, do ponto de vista científico. Neste
sentido tenho ainda de agradecer de um modo geral, ao Museu Nacional de História
Natural e da Ciência por me ter recebido, permitindo aí realizar uma parte significativa
do meu trabalho que tornou possível a realização desta dissertação e, em particular, a
todos os técnicos do Departamento de Mineralogia, Geologia e Paleontologia do Museu
(Sr. João Paulo Lopes, Drs. César Lopes e Bruno Ribeiro) pelo bom acolhimento e por
todo o apoio que me prestaram durante a preparação das amostras de sedimento, tendo-
se mostrado sempre disponíveis a ajudar no que fosse necessário e, claro, ao Wilamy
Rita, também técnico do Museu, que me ajudou a realizar as primeiras lavagens de
sedimento.
Tenho ainda de agradecer ao CSIC por me ter recebido nas suas instalações em
Madrid e permitir a utilização dos equipamentos que tornaram possível a realização das
fotografías dos restos odontológicos apresentadas na Sistemática desta dissertação, e
neste contexto à Dra. Marta Moreno García e ao Carlos Pimenta pela ajuda e boleia.
Não esquecendo todo o trabalho que foi realizado antes de mim, tenho de
agradecer a todos os investigadores que se dedicam a esta arqueociência e sobretudo
àqueles cujo conhecimento contribuiu de algum modo para a realiazação desta
dissertação, e ao LARC por todo o esforço e trabalho que tiveram ao criar aquela que é
hoje uma coleção osteológoca de referência, que permite a muitos investigadores
esclarecer as suas dúvidas.
Um obrigado ainda ao Pedro, por ter aturado com a sua paciência de santo os
meus momentos de maior stress quando a escrita não saía a feição, por reler os meus
textos e por me ouvir lê-los em voz alta vezes e vezes sem conta.
Por último, mas não menos importante – bem pelo contrário – tenho de
agradecer aos meus pais, principais responsáveis pela realização deste trabalho, na
medida que puserem à minha disposição desde sempre todos os meios financeiros
necessários à minha educação e me deixaram sempre seguir com os meus sonhos e
vontades, apoiando-me incondicionalmente nas minhas escolhas.
A vocês devo aquilo que sou!

AS MICROFAUNAS DO ABRIGO DA PENA D´ÁGUA (TORRES NOVAS):
PROPOSTA DE METODOLOGIA PARA O ESTUDO DE MICROFAUNAS EM
CONTEXTOS ARQUEOLÓGICOS
MARTINA SOFIA GONÇALVES MONTEIRO
RESUMO
PALAVRAS-CHAVE: Arqueologia; Neolítico Antigo; Ação Antrópica;
Micromamíferos; Espécies comensais (Mus).
O Abrigo da Pena d'Água localiza-se na freguesia da Chancelaria, concelho de
Torres Novas, distrito de Santarém. Trata-se de um abrigo sob rocha situado na base da
escarpa conhecida na região por Arrife da Serra d'Aire.
As escavações realizadas entre 1992 e 1997 permitiram constatar a existência de
ocupação humana no local durante todo o Neolítico e ocupações esporádicas desde o
Epipaleolítico até à Época Moderna (Carvalho, 1998).
Na presente dissertação são alvo de estudo os restos osteológicos
correspondentes aos micromamíferos recolhidos nos níveis de ocupação humana
datadas do Epipaleolítico e Neolítico Antigo. Pretende-se, através da sua análise e a
partir da metodologia de estudo aqui proposta, caracterizar a distribuição dos
micromamíferos identificados na área envolvente do Abrigo da Pena d'Água, tendo em
conta as alterações ambientais e ecológicas aqui ocorridas após a introdução da
agricultura no início do Neolítico Antigo e compreender o seu significado
paleoclimático. Foi possível constatar que a introdução da agricultura na área da Pena
d´Água teve como consequência a introdução de novas espécies de roedores,
nomeadamente Mus, e, a redistribuição de espécies de roedores já existentes,
nomeadamente Microtus cabrerae, devido as alterações causadas por esta prática no
meio natural.
ABSTRACT
KEYWORDS: Archaeology; Early Neolithic, Anthropic action; Small mammals;
Commensal species (Mus).
The Pena d’Água rock-shelter is located in the village of Chancelaria,
municipality of Torres Novas, district of Santarém. This archaeological site is situated
on an escarpment known as Arrife da Serra d'Aire.
Between 1992 and 1997 were carried out archaeological excavation that showed
evidence of occupation during the Neolithic and some sporadic occupations in the
Epipaleolithic to the Early Modern Period (Carvalho, 1998).

This master dissertation is about the study of osteological remains of small
mammals (micro-fauna) of the archaeological record of Epipaleolítico and Neolithic
occupations.
The principal aim is to characterize the micro-fauna distribution in this levels
and try to characterize and understand the ecological and environmental changes
resulting from the introduction of agriculture in the early Neolithic.

ÍNDICE
LISTA DE ABREVIATURAS
0. NOTAS INTRODUTÓRIAS ……………………………………….… Pág. 1
0.1. OBJETIVOS ………………..…………………………..…...……… Pág. 5
0.2. ESTADO DA ARTE ………………………..………...…..………… Pág. 7
CAPÍTULO I: O ABRIGO DA PENA D´ÁGUA E O SEU CONTEXTO
1. O sítio arqueológico …………………………………………………... Pág. 10
1.1. Contextualização geográfica ……………………………………...… Pag. 12
1.1.1. Implatação e exploração dos recursos animais …………………… Pág. 12
1.1.2. O coberto vegetal e as práticas agrícolas ……………………….… Pág. 15
1.2. Contextualização arqueológica …………………………..…………. Pág. 19
1.2.1. Crono-estratigrafia …………………………………..……………. Pág. 19
1.3. Contextualização cultural: a Neolitização ………………………….. Pág. 23
CAPÍTULO II: AS AMOSTRAS E AS MICROFAUNAS
2. Amostras estudadas ……………………………………………......….. Pág. 25
3. Metodologia de estudo …………………………………….………….. Pág. 27
4. Taxonomia ……………………………………………...…………….. Pág. 36
4.1. Sistemática ……………………………………………………..…… Pág. 38
4.1.1. Micromamíferos …………………….…………………………….. Pág. 38
4.1.1.1. Ordem Soricomorpha …………………………….…………...… Pág. 38

4.1.1.2. Ordem Rodentia ………………………...…………………….… Pág. 42
CAPÍTULO III: DISCUSSÃO DOS RESULTADOS
5. Processos deposicionais – Agentes tafonómicos ………….…........….. Pág. 62
6. Processos pós-deposicionais ……………………………………...…... Pág. 64
6.1. Estado das amostras ……………...…………………………………. Pág. 68
7. Distribuição espacial dos restos analisados ………………………..…. Pág. 70
8. Significado paleoecológico, paleoambiental e paleoclimático das espécies
identificadas durante o Epipaleolítico e o Neolítico Antigo ………………..…... Pág. 73
CONCLUSÃO ……………………………………………...…………… Pág. 78
REFERÊNCIAS BIBLIOGRÁFICAS ……………....……………….…. Pág. 83
ANEXOS
ANEXO 1: TABELAS

LISTA DE ABREVIATURAS
LARC: Laboratório de Arqueociências
CSIC: Consejo Superior de Investigaciones Cientificas
RI: Restos Identificáveis
NI: Não Identificáveis
NMI: Número Mínimo de Indivíduos
NRD: Número de Restos Determinados
NRDa: Número de Restos Determinados anatomicamente
NRDt: Número de Restos Determinados taxonomicamente
ND: Não Determinados
NTR: Número Total de Restos
M1: Primeiro molar superior
M2: Segundo molar superior
M3: Terceiro molar superior
Pm4: Quarto pré-molar superior
M1: Primeiro molar inferior
M2: Segundo molar inferior
M3: Terceiro molar inferior
Pm4: Quarto pré-molar inferior

1
0. NOTAS INTRODUTÓRIAS
O Abrigo da Pena d'Água (Torres Novas) é um sítio arqueológico de grande
importância para o conhecimento do período Neolítico da Estremadura e do atual
território português, na medida em que é o único contexto que preserva uma
estratigrafia que apresenta uma sequência crono-cultural extensa e uma boa conservação
dos restos materiais e orgânicos exumados (Carvalho, 1998). Constitui, assim, o único
sítio arqueológico conhecido até ao momento em Portugal detentor de uma sequência
estratigráfica contínua desde o Neolítico Antigo Cardial até ao Neolítico Final e, o único
identificado na vertente Atlântica Ocidental que permitiu reconhecer indicadores
indiretos da ocorrência de práticas agrícolas e pastoris durante o Neolítico Antigo
(Carvalho 2003b e 2008, Póvoas, 1998; Figueiral, 1998; Valente, 1998). O sítio revelou
ainda uma sequência sedimentar Holocénica com aspetos interessantes para o estudo da
evolução das associações microfaunísticas, e sobretudo de micromamíferos, durante o
período de transição entre o final do Epipaleolítico e o início do Neolítico Antigo
(Póvoas, 1998).
É do ponto de vista zooarqueológico que esta dissertação se insere no estudo do
sítio arqueológico do Abrigo da Pena d'Água, tendo como objeto os restos osteológicos
microfaunísticos recuperados nos níveis estratigráficos correspondentes às ocupações do
Epipaleolítico e do Neolítico Antigo.
A Zooarqueologia, surge como disciplina auxiliar da Arqueologia durante os
anos 40 do século XX como consequência da aplicação de conceitos como
“funcionalismo” e “contexto” à Arqueologia e beneficia do novo entendimento do
comportamento humano que surge neste período, não apenas meramente artefatualista,
mas sobretudo de compreensão da adaptação do Homem ao meio ambiente e animal,
pondo em destaque o papel que os animais desempenharam nesta adaptação (Reitz &
Wing, 1999). Mas é durante o surgimento da Nova Arqueologia que esta disciplina se
desenvolve, assumindo um papel determinante na compreensão da formação de
contextos faunísticos em sítios arqueológicos, adquirindo uma perspetiva sobretudo
tafonómica (Reitz & Wing, 1999). Neste contexto surgem dois conceitos com objetivos
distintos no estudo de faunas provenientes de contextos arqueológicos: a
Zooarqueologia, que se dedica ao estudo de restos osteológicos animais de proveniência

2
arqueológica no sentido de reter informação de tipo antropológico (comportamento do
Homem) e a Arqueozoologia, que dá prioridade à componente biológica dos restos
osteológicos com a finalidade de estudar a evolução das espécies representadas através
dos elementos osteológicos fósseis (Reitz & Wing, 1999). Neste sentido ao longo desta
dissertação optamos pelo termo Zooarqueologia.
A Zooarqueologia como área de investigação que se dedica ao estudo dos restos
osteológicos de origem animal provenientes de contextos arqueológicos (Moreno-
García et al, 2003), aborda diferentes tipos e dimensões de restos faunísticos, para o
trabalho em questão, tratando-se de um estudo microfaunístico, são apenas abordados os
restos faunísticos de menores dimensões, denominados de microfaunas.
Embora esta dissertação se dedique à criação de uma metodologia de estudo de
restos osteológicos de microfaunas provenientes de contextos arqueológicos, importa
clarificar que apenas foram alvo de estudo os restos odontológicos, mandíbulas e
maxilares com dentes inclusos, por serem aqueles que se encontram em maior
abundância nas amostras estudadas e por constituírem restos diagnósticos a nível
taxonómico, de espécies de micromamíferos e dentro deste grupo as correspondentes às
Ordens Rodentia (roedores) e Soricomorpha (insectívoros), por serem as únicas que se
encontravam representadas nas amostras analisadas.
Todavia, estas duas Ordens de micromamíferos integram um conjunto de
espécies, que pelas suas características adaptativas a diferentes biótopos e habitats, nos
permitem reconhecer eventuais alterações ambientais registadas ao longo da sequência
cronológica aqui estudada, tivessem sido elas de origem natural ou provocadas pela
ação antrópica devido ao desenvolvimento da agricultura e da pastorícia na área em
redor do abrigo.
Após esta nota há a necessidade de esclarecer algumas noções básicas dos
grupos abordados, nomeadamente microvertebrados e micromamíferos.
O grupo dos microvertebrados engloba todos os animais vertebrados de menores
dimensões. Estes distribuem-se pelas Classes: Amphibia (anfíbios), que compreende as
Ordens Anura (sapos e rãs) e Caudata (salamandras e tritões) (Ferrand de Almeida,
2001); Reptilia (répteis), que, neste contexto, incluí as espécies de menor dimensão
distribuídas pela Ordem Squamata – Famílias Emydidade e Bataguridae (cágados),
Amphisbaenidae (cobra-cega), Gekkonidade (osgas), Lacertidae (lagartos e lagartixas),

3
Anguidae (licranço ou cobra-de-vidro), Chamaleonidae (camaleão), Scincidadae
(cobras-de-pernas) e Colubridae (serpentes) (Barbadillho, 1999); Aves, considerando as
aves pertencentes às espécies de menores dimensões que, no essencial estão
concentradas na Ordem Passeriformes (excluindo algumas espécies de corvídeos); e
Mammalia, representada pelas espécies que integram os Mamíferos de menores
dimensões denominados de micromamíferos, que incluem as Ordens Soricomorpha
(insectívoros) (Gállego & López, 1982), Chiroptera (morcegos) (Sevilha, 1986),
Rodentia (roedores) (Mitchell-Jones et al 1999; Gállego & Alamany, 1985) e algumas
espécies pertencentes à Ordem Carnívora, tais como pequenos Mustelídeos (doninha e
arminho).
Embora o termo "micromamíferos" não possua valor taxonómico, este é
amplamente utilizado na literatura científica deste tipo para mencionar os Mamíferos
pretendentes às Ordens, Famílias e Géneros em que a maioria das espécies possui uma
dimensão reduzida. Esta denominação geralmente inclui insectívoros, morcegos e
roedores, embora algumas espécies de roedores de maior dimensão como o castor, a
marmota e o ouriço-cacheiro não estejam incluídos neste grupo (López-García, 2008,
pp. 119). O mesmo sucede com os Lagomorfos (coelhos e lebres).
O estudo dos restos osteológicos de micromamíferos provenientes de contextos
arqueológicos é imprescindível para a caracterização ambiental, climática e ecológica
de assentamentos arqueológicos, sobretudo de períodos mais recuados. Constituem-se
como óptimos indicadores paleoecológicos, paleoambientais e paleoclimáticos, por
serem organismos bastante sensíveis à alteração de fatores como a temperatura, a
humidade e do coberto vegetal (Sése, 2011). Devido ao seu reduzido tamanho, altas
taxas de reprodução e baixa mobilidade são altamente sensíveis a mudanças climáticas,
ambientais e ecológicas, permitindo compreender estas variações ao longo do tempo
num determinado sítio arqueológico (Chaline, 1972). Por último, em contextos
sedimentares antigos, estes são ainda indicadores bioestratigráficos muito precisos,
sobretudo no caso roedores, permitindo estabelecer uma cronologia relativa para os
depósitos onde se encontram (Chaline, 1972).
A partir do seu estudo é ainda possível retirar conclusões relacionadas com a
ação antrópica. A existência e/ou ausência ou a diminuição drástica da representação de
determinados microvertebrados num contexto arqueológico, poderá estar diretamente
relacionada com a ação antrópica ocorrida nesse local.

4
A intervenção humana no meio natural, sobretudo a partir do desenvolvimento
da agricultura e da pastorícia, teve profundas implicações na paisagem, originando
variações na distribuição espacial das comunidades de micromamíferos (Mateus et al,
2002). A introdução da agricultura, tal como o desenvolvimento das rotas comerciais,
traduziram-se numa sucessão de introduções involuntárias de microvertebrados que, no
caso dos roedores, teve consequências profundas, quer ao nível da competição
estabelecida com as espécies autóctones, quer da relação oportunista/comensal das
atividades humanas. O ratinho-doméstico (Mus domesticus), o rato-preto (Rattus rattus)
e a ratazana-castanha (Rattus norvegicus), são exemplos disso (Mateus et al, 2002).
Neste sentido a Zooarqueologia assume um papel incontornável para o
conhecimento do passado zoogeográfico dos animais.
Por último, tendo em conta que o estudo microfaunístico é um processo bastante
complexo, quer pela diversidade de Taxones existentes, quer devido às alterações
morfológicas sofridas pelos mesmos ao longo do tempo, a Zooarqueologia, necessita
recorrer a análises cada vez mais aprofundadas nos diferentes ramos das Ciências
Naturais (Biologia, Ecologia, Genética, etc.). É no conhecimento da Biologia e dos
requisitos ecológicos das espécies atuais que residem bases imprescindíveis para
interpretar os vestígios que nos chegam do passado (Cabral, 2005; Palomo & Gisbert,
2007).

5
0.1. OBJETIVOS
A seguinte dissertação de mestrado tem como objetivo principal propor uma
metodologia para o estudo de restos osteológicos de microfaunas provenientes de
contextos arqueológicos.
Através desta proposta metodológica e a partir da identificação das espécies
existentes nas amostras sedimentares analisadas, pretende-se caracterizar as associações
de micromamíferos (roedores e insectívoros) provenientes dos níveis do Epipaleolítico e
Neolítico Antigo do Abrigo da Pena d'Água. Desta forma aspira-se igualmente perceber
se os taxones identificados correspondem a um tipo de zona biótica específica e, deste
modo chegar à compreensão do seu significado paleoecológico, paleoclimático e
paleoambiental, e caracterizar a sua distribuição na área envolvente ao Abrigo da Pena
d'Água, verificando se o aparecimento/desaparecimento de determinadas espécies de
micromamíferos se prende com alterações climáticas e/ou ambientais, com a existência
de pressão antrópica ou devido a relações de competição com outros micromamíferos.
Os micromamíferos sendo animais muito sensíveis a condições meteorológicas
precisas, a ocorrência de alterações climáticas força-os a migrar para outras áreas.
Devido às suas necessidades energéticas e fisiológicas concertas, à grande diversidade
de espécies existentes, que se traduzem em relações complexas de competição, possuem
faixas geográficas de distribuição muito limitadas, habitando biótopos específicos. Estas
características fazem com que este grupo de animais seja uma ferramenta muito útil no
momento de aferir características climáticas e de vegetação dos depósitos arqueológicos
em que aparecem (López-García, 2008, pp. 119).
Por último, pretende-se identificar e compreender os processos deposicionais –
agentes tafonómicos – e pós-deposicionais – agentes diagénicos – que originaram a
acumulação destes restos osteológicos no Abrigo d'Água e condicionaram a sua
preservação desde a sua deposição até à sua recuperação.
A Tafonomia é a ciência que reconstitui os fenómenos que afetaram os restos
esqueléticos animais desde a sua morte, quando este ainda se encontra na biosfera, até à
recuperação do elemento osteológico durante o processo de escavação, ou seja, depois
da sua incorporação na litosfera (Lyman, 2001; Valente, 2000; Moreno García et al,
2003)

6
São vários os fatores tafonómicos que condicionam a conservação dos restos
osteológicos em contextos arqueológicos. Existem fatores de origem natural, animal e
antrópica (Lyman, 2001). Concretamente no que diz respeito às microfaunas os mais
relevantes são os de origem natural, nomeadamente, a acidez dos solos (PH), as
alterações de temperatura e a presença ou ausência de humidade.
A caracterização dos agentes tafonómicos que estiveram na origem da
acumulação destes restos osteológicos será possível através da identificação de marcas
registadas sobretudo nos restos odontológicos, relacionadas com processos de desgaste
(desgaste do esmalte dos dentes devido ao longo período de uso) ou de digestão
(causadas pelos líquidos gástricos existentes no estômago que permitem a digestão dos
alimentos ingeridos). Desta forma, esta acumulação em traços gerais pode ser resultado
da desagregação de dejetos/ regurgitações de predadores que frequentaram o local como
abrigo (Pecharromán, 1993) e/ ou originada pela morte de forma natural de animais que
utilizaram este espaço para o mesmo fim.
Existem diferentes tipos de predadores que incluem micromamíferos na sua
dieta, nomeadamente aves de rapina noturnas e diurnas e pequenos e médios carnívoros,
e que em função do tipo de ingestão e digestão das suas presas, podem ser responsáveis
pela formação de acumulações fósseis de restos destes animais, nomeadamente em
contextos de gruta e de abrigo sob rocha (Andrews, 1990). As aves de rapina noturnas
(sobretudo corujas) são as principais responsáveis por este tipo de acumulações. Devido
ao seu processo de ingestão e digestão (ingestão das suas presas inteiras e posterior
expulsão das partes não digestíveis do seu organismo através de regurgitações) levam à
acumulação de esqueletos na maioria dos casos completos, pelo contrário os pequenos
carnívoros, por possuírem um processo de ingestão e digestão muito destrutivos em
relação aos restantes predadores de micromamíferos, são responsáveis por uma fraca
acumulação de restos osteológicos, que na maioria dos casos possuem um elevado grau
de digestão (López-García, 2008, pp. 119).
As acumulações de restos osteológicos em contextos arqueológicos podem
ainda, embora seja menos frequentemente, ser resultado de armadilhas naturais (onde os
animais morrem de forma natural) (Pimenta et al, 2015) e em raríssimos casos da ação
antrópica (Andrews, 1990).

7
0.2. ESTADO DA ARTE
O Abrigo da Pena d'Água, foi alvo de escavações arqueológicas entre os anos de
1992 e 2000, sob a coordenação do arqueólogo António Faustino de Carvalho,
atualmente docente na Universidade do Algarve.
Os resultados das escavações decorridas entre 1992 e 1997 foram publicados
pelo arqueólogo responsável em 1998 na Revista Portuguesa de Arqueologia (Carvalho,
1998). Neste número foram também publicados outros artigos que abordaram os
resultados de diversos estudos realizados a artefactos e ecofactos recolhidos neste sítio,
nomeadamente cerâmicas2, restos vegetais3, restos macrofaunísticos4 e microfaunístico5.
Após a publicação dos resultados das escavações de 1992-1997, foi publicado um artigo
referente ao estudo das cerâmicas estampilhadas da Idade do Ferro descobertas nos
níveis arqueológicosdo abrigo.6 Por último em 2012, foi publicado o estudo
geoarqueológico do abrigo.7
Apesar do significativo desenvolvimento que a investigação na área da
Zooarqueologia de um modo geral tem conhecido na última década em Portugal,
sobretudo desde a criação do LARC e das suas coleções osteológicas de referência
(Moreno-García et al, 2003), no que diz respeito concretamente ao estudo de restos
microfaunísticos provenientes de contextos arqueológicos, apesar desta arqueociência
2 FERREIRA, Maria Mulize Neves (1998) – “As cerâmicas medievais/modernas do Abrigo da Pena
d'Água (Torres Novas).” Revista Portuguesa de Arqueologia, vol.1, nº2, pp. 97-106.
3 FIGUEIRAL, Isabel (1998) – “Abrigo da Pena d'Água (Torres Novas): a contribuição da antracologia.”
Revista Portuguesa de Arqueologia, vol.1, nº2, pp. 73-80.
4 VALENTE, Maria João (1998) – “Análise preliminar da fauna mamalógica do Abrigo da Pena d'Água
(Torres Novas): campanhas de 1992-1994.” Revista Portuguesa de Arqueologia, vol.1, nº2, pp. 85-96.
5 PÓVOAS, Liliana (1998) – “Faunas de Micromamíferos do Abrigo da Pena d'Água (Torres Novas) e o
seu significado paleoecológico: considerações preliminares.” Revista Portuguesa de Arqueologia, vol.1,
nº2, pp. 81-84.
6 CRAVALHO, António Faustino de (1998) – “Cerâmica estampilhada do Abrigo da Pena d'Água
(Torres Novas): contexto, cronologia e breve enquadramento regional.” In BERNARDES, J. P, org – Sic
memorat. Estudos em homenagem a Teresa Júdice Gamito. Pp. 53-68.
7 SIMÕES, Carlos (2012) – “Estudio geoarqueológico del Abrigo da Pena d'Água (Torres Novas,
Portugal) en el contexto de la Neolitizaciôn de la vertiente atlântica meridional peninsular.” Revista
Estudios de Cuarternario, nº2, pp. 49-69.

8
ser já recorrente em vários países, nomeadamente em Espanha (para uma síntese geral
ver López-García, 2008) e em França (para uma síntese geral ver Chaline, 1972), em
Portugal encontra-se pouco desenvolvida. O fraco incremento de estudos
microfaunísticos em Portugal poderá relacionar-se sobretudo com razões de recolha, por
não se encontrar ainda implementada na maioria das escavações arqueológicas a recolha
de amostras de sedimento para crivagem com água, único meio de recolha de restos
osteológicos de dimensões tão reduzidas.
Até ao presente foram alvo de estudo microfaunístico apenas um número
muitíssimo reduzido de sítios arqueológicos em Portugal. Estes têm sido realizados
sobretudo em contextos arqueológicos de épocas mais recuadas, essencialmente com
ocupações Pré-históricas localizadas em contextos de grutas.
Estes estudos revelaram-se essenciais para a compreensão e caracterização
destes sítios arqueológicos, uma vez que permitiram retirar conclusões relacionadas
com o tipo de ecologia e coberto vegetal existentes e, mais importante, permitiram
através de dados indiretos concluir o tipo de economia aí desenvolvida.
Tendo em conta o facto deste trabalho se dedicar apenas ao estudo de restos de
micromamíferos, abaixo é apresentada uma síntese dos sítios arqueológicos portugueses
nos quais foram analisados restos faunísticos de associações de micromamíferos:
Castro do Zambujal – Torres Vedras – (Storch & Uerpmann, 1976);
Abrigo do Algueirão de Goldra – Loulé – (Antunes et al, 1986; Póvoas et al,
1993; Póvoas, 2001);
Gruta do Caldeirão – Tomar – (Antunes, 1989; Póvoas et al, 1992; Póvoas,
2001);
Castro de Leceia – Oeiras – (Cardoso et al, 1996);
Cidadela Medieval de Mértola – Mértola – (Morales, 1997);
Pena d'Água – Torres Novas – (Póvoas, 1998);
Gruta da Figueira Brava – Setúbal – (Jeannet, 2000);
Abrigo do Lagar Velho – Lapedo – (Moreno-García et al, 2002);
Gruta da Furninha – Peniche – (Brugalet al, 2012);
Bom Santo – Alenquer – (Pimenta, 2013).
Procuro, assim, com este trabalho contribuir de algum modo para o
desenvolvimento do estudo de microfaunas provenientes de contextos arqueológicos, e,

9
acrescentar informação à que foi dada a conhecer por Liliana Póvoas (Póvoas, 1998),
para o caso concreto do Abrigo da Pena d'Água.

10
CAPÍTULO I: O ABRIGO DA PENA D'ÁGUA
1. O sítio arqueológico
O Abrigo da Pena d'Água
localiza-se na zona centro-oeste de
Portugal, na localidade da Rexaldia,
pertencente à freguesia de
Chancelaria, concelho de Torres
Novas e distrito de Santarém.
Trata-se de um abrigo sob
rocha, situado a 120 metros acima do
nível médio das águas do mar, na base
da escarpa de falha, com cerca 30
metros de altura, que separa as serras
do Maciço Calcário Estremenho da
Bacia Terciária do Tejo. Este ponto da
escarpa, conhecida na região por
Arrife da Serra d'Aire, é designado
por Pena d'Água por existirem neste
local pequenas exsurgências
(Carvalho, 1998, pp. 41).
O sítio arqueológico foi descoberto em 1992, aquando da deslocação ao local de
membros da Sociedade Torrejana de Espeleologia e Arqueologia (S.T.E.A) com o
intuito de levar a cabo trabalhos de desobstrução de algumas exsurgências visíveis na
parede do Arrife, e, desse modo verificar a possibilidade de exploração espeleológica
dessas cavidades. O surgimento de fragmentos de cerâmica pré-histórica e de pedra
lascada na terra removida pela retroescavadora utilizada na desobstrução das fissuras,
revelaram a presença de um depósito arqueológico, posteriormente reconhecido pelo
arqueólogo Dr. João Zilhão (Carvalho, 1998, pp. 42).
A observação dos cortes efetuados pela máquina permitiu verificar a ocorrência
de vestígios arqueológicos, resultantes da ocupação humana no local, até uma
profundidade de cerca de 5 metros (Figura 1). Tendo em conta que a área destruída pela
Mapa 1: localização do concelho de Torres Novas
no mapa de Portugal e da freguesia de Chancelaria e
da localidade de Rexaldia no concelho de Torres
Novas (Fonte: pt.wikipedia.org)

11
máquina retroescavadora se localiza na extremidade oriental do abrigo, pode considerar-
se que apenas uma pequena fração do sítio arqueológico foi afetada por esta (Carvalho,
1998, pp. 42).
Figura 1: topografia do Abrigo da Pena d'Água – levantamento realizado pela S.T.E.A em 1992
(Carvalho, 1998,pp. 42).

12
1.1. Contextualização Geográfica
1.1.1. Implantação e exploração dos recursos animais
O ponto da escarpa do Arrife da Serra d'Aire (Figura 2) onde se localiza o
Abrigo da Pena d'Água, contorna a Serra d'Aire ao longo da sua vertente virada ao Tejo,
estendendo-se desde Ourém, a Norte, até Rio Maior, a sul. Possui uma topografia muito
irregular, apresentando como ponto mais elevado o próprio cume da Serra d'Aire,
erguido a 679 metros de altura (Araújo & Zilhão, 1991).
Vis ta
Figura 2: vista do Arrife a partir da planície existente abaixo do Abrigo da Pena d´Água (Fotografia da
autoria de A. F. Carvalho).
No que concerne à implantação na paisagem, o Abrigo da Pena d'Água encontra-
se numa posição de charneira, entre o maciço calcário e o rebordo ocidental da Bacia
Terciária do Tejo. Devido à sua localização entre estas duas realidades geologicamente
distintas, o abrigo integra-se, tal como todo o Arrife, num contexto de ecótono
(Carvalho, 2003a, pp. 137). Se por um lado, o abrigo é sobranceiro à planície do Tejo e
reúne um conjunto de condições favoráveis ao desenvolvimento da agricultura, como a
existência de solos irrigados, devido à proximidade a um dos principais afluentes do Rio

13
Almóada, a Ribeira do Alvorão, por outro lado, é detentor de um bom domínio visual
para Leste e Sul e localiza-se junto a bons acessos naturais à Serra d'Aire, que
facilitavam a existência de pastoreio (Figura 3).
Figura 3: vista para o Vale do Alvorão (Fotografia da autoria de A. F. Carvalho).
Ainda que o abrigo ocupado pelo Homem durante o Neolítico seja atualmente
quase irreconhecível (Figura 4), devido ao derrube da sua pala original, como é possível
constatar nas suas imediações, onde são ainda visíveis grandes blocos de calcário
resultantes da sua queda, a escolha deste local como abrigo prende-se sobretudo com a
existência deste contexto de ecótono, que tornava possível a exploração simultânea dos
recursos espontâneos pelas populações neolíticas e o desenvolvimento da agricultura e
da pastorícia. Localizado junto de solos aráveis e densamente irrigados que permitiam o
desenvolvimento de uma agricultura primitiva nas terras baixas, e próximo da serra que
possibilitava a ocorrência da atividade pastoril nas áreas montanhosas do maciço
(Carvalho, 1998, pp. 43).

14
Figura 4: à esquerda vista da área em escavação desde 1992; à direita vista atual (2014) da área escavada
(Fotografia da autoria de A. F. Carvalho).
A existência de uma economia assente na exploração simultânea de recursos
domésticos e recursos selvagens é comprovada pelo estudo da fauna mamalógica. Ainda
que as principais espécies identificadas para os níveis do Neolítico tenham sido a cabra
e a ovelha domésticas, foram também identificados, embora em menor número, restos
osteológicos pertencentes a animais selvagens, nomeadamente javali e veado (Valente,
1998, pp. 85).
É de salientar, por um lado, o facto de terem sido identificados restos
osteológicos de ovelha e cabra doméstica em todos os níveis do Neolítico, o que
demonstra que a primeira exploração pastoril na Serra d'Aire ocorreu desde o início do
Neolítico Antigo, e por outro lado, o facto de restos osteológicos de veado se
encontrarem também associados a todos os níveis do Neolítico, revelando que a caça
detinha um papel importante na economia neolítica, funcionando como complemento à
pastorícia, embora esta predominasse sobre a caça (Quadro 1) (Valente, 1998, pp. 85;
90).

15
Quadro 1: número de Restos por Taxa identificados em cada uma das camadas arqueológicas escavadas
no Abrigo da Pena d´Água (Valente, 1998, pp. 95).
1.1.2. O coberto vegetal e as práticas agrícolas
A zona onde se insere o Abrigo da Pena d'Água integra-se na zona ecológica
Atlanto-mediterrânea/ Sub-mediterrânea, tendo como principais espécies arbóreas
espontâneas o zambujeiro, o pinheiro bravo, o pinheiro manso, o carvalho e o sobreiro
(Araújo & Zilhão, 1991).
No que concerne ao coberto vegetal espontâneo atualmente existente na área em
torno do abrigo, este é composto sobretudo por espécies rasteiras e arbustivas,
registando-se escassos casos de árvores de grande porte. No que diz respeito à
arboricultura local, esta consiste sobretudo na exploração de laranjeiras e oliveiras,
culturas típicas de zonas de clima mediterrâneo.
A partir do estudo antracológico realizado aos carvões recolhidos durante os
trabalhos arqueológicos desenvolvidos no Abrigo da Pena d'Água, foi possível constatar
que a paisagem envolvente seria, já no Neolítico Antigo, de tipo mediterrâneo, sendo o
coberto vegetal composto sobretudo pela variedade brava da oliveira, o zambujeiro. De
facto foi possível constatar que já durante o Epipaleolítico ou Paleolítico Superior Final,
o coberto vegetal seria de tipo mediterrâneo, uma vez que os resultados verificados para
16
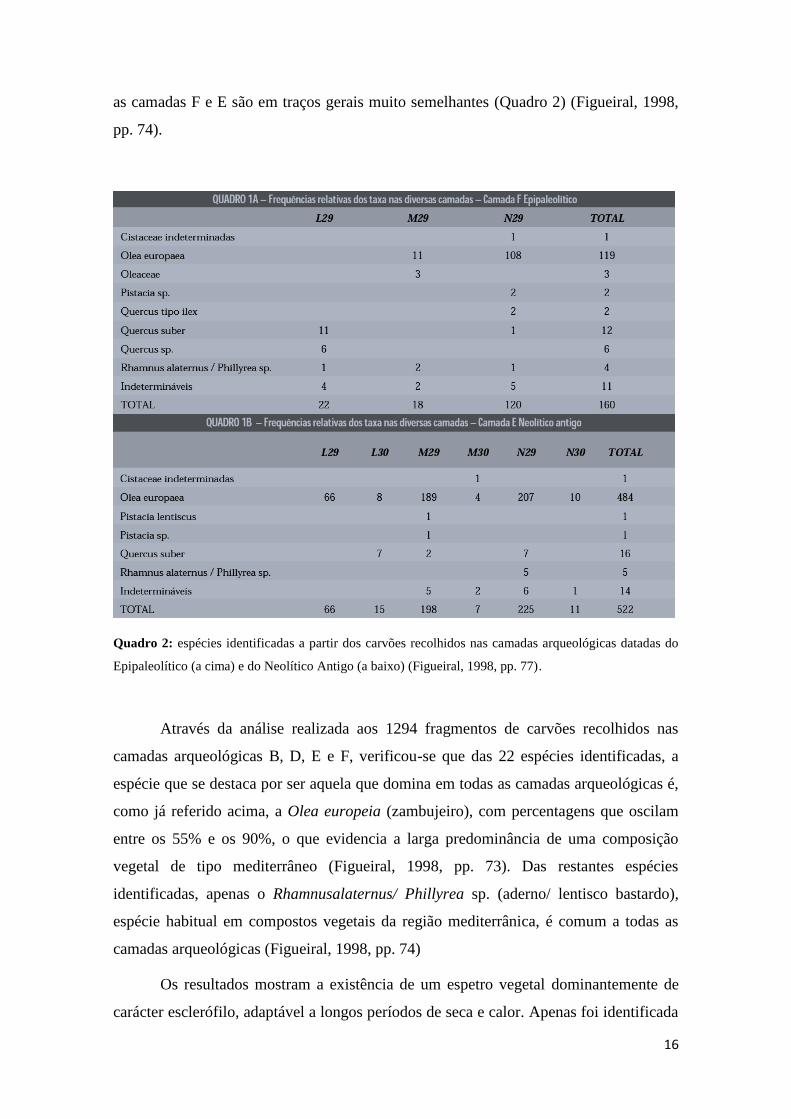
as camadas F e E são em traços gerais muito semelhantes (Quadro 2) (Figueiral, 1998,
pp. 74).
Quadro 2: espécies identificadas a partir dos carvões recolhidos nas camadas arqueológicas datadas do
Epipaleolítico (a cima) e do Neolítico Antigo (a baixo) (Figueiral, 1998, pp. 77).
Através da análise realizada aos 1294 fragmentos de carvões recolhidos nas
camadas arqueológicas B, D, E e F, verificou-se que das 22 espécies identificadas, a
espécie que se destaca por ser aquela que domina em todas as camadas arqueológicas é,
como já referido acima, a Olea europeia (zambujeiro), com percentagens que oscilam
entre os 55% e os 90%, o que evidencia a larga predominância de uma composição
vegetal de tipo mediterrâneo (Figueiral, 1998, pp. 73). Das restantes espécies
identificadas, apenas o Rhamnusalaternus/ Phillyrea sp. (aderno/ lentisco bastardo),
espécie habitual em compostos vegetais da região mediterrânica, é comum a todas as
camadas arqueológicas (Figueiral, 1998, pp. 74)
Os resultados mostram a existência de um espetro vegetal dominantemente de
carácter esclerófilo, adaptável a longos períodos de seca e calor. Apenas foi identificada

17
uma espécie arbórea associada a cobertos vegetais típicos de zonas húmidas, o Fraxinus
excelsior (freixo) (Figueiral, 1998, pp. 75).
É ainda de salientar a identificação de espécies arbustivas, como o medronheiro,
as urzes, as cistáceas e algumas leguminosas (giestas) que testemunham a existência de
áreas de matagal, como as ainda hoje registadas em torno do Abrigo da Pena d'Água, e a
total ausência de carvões pertencentes a pinheiros em todas as camadas arqueológicas, o
que poderá estar relacionado com o declínio dos pinhais durante o Holocénico Médio
devido à expansão da vegetação de carácter esclerófilo, e, muito provavelmente com a
intensificação das atividades humanas a partir do Neolítico, que terão levado à
desflorestação dos pinhais da Estremadura e à expansão do zambujeiro (Figueiral, 1998,
pp. 75 e 76).
A existência de um coberto vegetal de tipo mediterrâneo na área envolvente do
Abrigo da Pena d'Água é também comprovada pelos resultados obtidos do estudo
preliminar realizado a uma fração de restos microfaunísticos provenientes deste sítio
arqueológico (Póvoas, 1998).
Segundo o estudo realizado, os micromamíferos identificados nas amostras
sedimentares recolhidas no Abrigo da Pena d'Água estão representados por cinco
espécies de roedores, dos quais os géneros Apodemus e Mus são os mais frequentes. A
predominância dos murídeos (Apodemus sylvaticus e Mus) face aos arvicolídeos
(Microtus), é característica das regiões de clima e coberto vegetal de tipo mediterrâneo
(Póvoas, 1998, pp. 81).
A predominância dos murídeos permite-nos retirar duas conclusões
determinantes na caracterização da área envolvente ao abrigo: se por um lado o domínio
de Apodemus silvaticus em todas as camadas arqueológicas, a par da percentagem
constante de Eliomys quercinus, são observações que demonstram a existência de
manchas de bosque onde estas espécies encontram os biótopos favoráveis ao seu
desenvolvimento, por outro lado, a presença de Mus, nomeadamente de Mus spretus e
Mus musculus, espécies associadas à expansão da agricultura, a partir da camada Ea,
demonstra a consolidação da neolitização nesta área a partir do Neolítico Antigo
Evoluído (Póvoas, 1998, pp. 83). A presença de ambas as espécies a partir da camada
Ea indica igualmente que a instalação da paisagem agrícola que sustenta a sua
existência terá sido um processo com início numa etapa anterior, o que por seu lado
permite deduzir a existência de práticas agrícolas pelo menos a partir das ocupações

18
humanas registadas na camada Eb-topo, subjacente àquela. Note-se, porém, que embora
a campanha de 1995 tenha sido integralmente consignada à recolha de elementos
carpológicos (através da escavação de um setor neolítico antigo e sua crivagem
sistemática a água), não foi possível até ao momento identificar indicadores diretos de
agricultura neolítica (isto é, sementes de leguminosas ou cereais domésticos).
Por último, a identificação de insectívoros, representados sobretudo pelo género
Crocidura, evidencia a existência de áreas detentoras de biótopos secos de cobertura
vegetal arbustiva, de tipo matagal, habitat onde este Género se desenvolve, e, a baixa
percentagem de Microtus indica uma reduzida extensão das áreas de solos húmidos que
estes arvicolídeos escavam (Póvoas, 1998. pp. 83).
A ocorrência maioritariamente de áreas de bosques é também confirmada pelo
estudo realizado aos restos de mesofauna e de macrofauna recolhidos dos mesmos
níveis arqueológicos da Pena d'Água, o qual, entre outros pontos, revelou uma elevada
ocorrência de javali (Sus scrofa) e veado (Cervus elaphus), no que concerne aos animais
selvagens (Valente, 1998). Ambas as espécies desenvolvem-se preferencialmente em
ambientes de bosque com clareiras, como o registado antracologicamente na área
envolvente do Abrigo da Pena d'Água.
Em suma, as conclusões retiradas dos estudos faunísticos vêm ao encontro dos
resultados verificados pelo estudo antracológico, comprovando a existência na área em
torno do abrigo de áreas arborizadas alternadas por cobertos arbustivos e por clareiras
desflorestadas, e em suma a dominância de um espetro vegetal de carácter esclerófilo
em detrimento dos cobertos vegetais mais húmidos (Figueiral, 1998), o que de um modo
geral está de acordo com as características bioclimáticas típicas do Holoceno Médio.

19
1.2. Contextualização arqueológica
1.2.1. Crono-estratigrafia
As oito campanhas arqueológicas levadas a cabo no local do Abrigo da Pena
d'Água, entre os anos de 1992 e 2000, permitiram constatar a existência de uma
sequência sedimentar com 2 m de potência estratigráfica incorporando vestígios de
ocupação humana datáveis desde o Epipaleolítico, nos níveis mais antigos (camada F),
até à Época Moderna, nos níveis mais recentes (camada A) (para uma síntese, ver
Carvalho, 1998).
Ao longo dos trabalhos arqueológicos foram exumados artefactos que apontam
para ocupações humanas contínuas durante todo o período do Neolítico, ocupações
esporádicas durante o Epipaleolítico, a Idade do Ferro e a Época Romana, e para
ocupações muito efémeras durante a Época Medieval e Moderna.
Neste contexto foi possível identificar seis camadas estratigráficas distintas,
onde são visíveis os vários momentos de ocupação do abrigo:
Camada A
Corresponde ao nível de derrube da pala do abrigo. É sobretudo constituída
por grandes blocos de pedra, sendo os artefactos muito raros, registando-se apenas
alguns fragmentos de cerâmica de Época Medieval e Moderna, relacionados com
ocupações humanas muito efémeras, talvez resultantes da passagem de pastores ou
caçadores pelo local após o derrube da pala do abrigo (Ferreira, 1998, pp. 97;106), e
escassos restos osteológicos de microfaunas e de macrofauna recente, com deposição
de origem antrópica, tendo em conta a marcas de corte e sinais de classificação que
apresentam (Carvalho, 1998, pp. 48).
Camada B
Corresponde sobretudo ao nível de ocupação do Neolítico Final, embora
ocorram também escassos fragmentos cerâmicos datados da Idade do Ferro e da
Época Romana (designadamente integrados numa lareira datada do início da Era
cristã), o que poderá significar que o topo desta camada serviu de superfície deste
depósito arqueológico durante pelo menos 3 mil anos, desde finais do Neolítico até ao
colapso da pala do abrigo no início da Época Romana. Este nível é caracterizado pela

20
ocorrência sobretudo de cerâmica lisa carenada com esféricos mamilares de pastas
vermelhas, vasos esféricos e hemisféricos e cerâmica estampilhada, no que concerne
as ocupações datadas da Idade do Ferro (Carvalho, 2008, pp.13-14). No que concerne
aos utensílios líticos, há no Neolítico Final uma preferência pela utilização de rochas
locais – quartzo e quartzito – para a produção de utensílios simples em lascas, e de
sílex apenas na produção de lâminas e pontas de seta. Esta camada é datável dos
finais do IV milénio BC, a julgar pelos paralelos obtidos noutros contextos
estremenhos (Carvalho, 1998, pp. 48; 62).
Camada C
Corresponde ao episódio de transição entre o Neolítico Final e o Neolítico
Médio, tendo em conta a ausência dos líticos registados na camada B e a menor
frequência de vasos carenados. Registam-se ainda escassos fragmentos intrusivos de
vasos da Idade do Ferro (Carvalho, 1998, pp. 49).
Camada D
Corresponde ao nível de ocupação do Neolítico Médio. Esta camada encontra-
se subdividida na camada Da, que corresponde ao Neolítico Médio Final, e Db, que
por sua vez corresponde ao Neolítico Médio Inicial. É de notar o reduzido número de
vasos decorados, apresentando como técnica decorativa predominante a incisão,
sobretudo na camada Db, predominando a cerâmica lisa com formas geométricas
simples não carenadas. No que diz respeito aos utensílios líticos em pedra lascada,
são sobretudo utilizas matérias-primas locais — quartzo e quartzito. O sílex apenas é
utilizado para a debitagem de lamelas. Estão ainda presentes micrólitos geométricos.
Datável da 1ª metade do IV milénio BC. (Carvalho, 1998, pp.49; 64).
Camada E
Corresponde ao nível de ocupação do Neolítico Antigo. Encontra-se também
subdividida em camada Ea e camada Eb-topo, ambas correspondentes ao Neolítico
Antigo Evoluído, e na camada Eb-base, correspondente ao Neolítico Antigo Cardial.
As camadas Ea e Eb-topo são caracterizadas pela ocorrência de cerâmica com
decoração impressa e incisa sob o bordo e por utensílios líticos muito semelhantes, no
que concerne à tipologia e técnicas de talhe, com os registados para as ocupações do

21
Neolítico Médio, enquanto que a base da camada Eb-topo é caracterizada pela
ocorrência sobretudo de cerâmica de tipo cardial e por um elevado número de
utensílios em sílex, o maior registado em toda a sequência estratigráfica. Deste
conjunto de realidades estratigráficas há diversas datações de radiocarbono realizadas
sobre carvões, a mais antiga das quais de 6390 ± 150 BP correspondente ao Neolítico
Antigo Cardial (Carvalho, 1998, pp. 49; 67; 70).
Camada F
Corresponde aos níveis de ocupação do Epipaleolítico. Destaca-se a indústria
lítica sobre lascas, algumas lâminas em sílex e a utensilagem macrolítica. Datada por
datação absoluta de 7370 ± 110 BP (Carvalho, 1998, pp. 49; 71).
Os trabalhos arqueológicos desenvolvidos no Abrigo da Pena d'Água revelaram
ser de grande importância para o conhecimento do Neolítico em Portugal, uma que vez
permitiram a elaboração de um primeiro modelo crono-estratigráfico deste período na
Estremadura portuguesa, devido à grande sequência sedimentar escavada e ao elevado
grau de conservação dos artefactos e ecofactos exumados.
Na sequência deste modelo crono-estratigráfico, e tendo em conta a
interpretação realizada por Carvalho (1998), é possível verificar a existência de cinco
fases de deposição de sedimentos, que correspondem a três tramos distintos:
o tramo superior, que corresponde à queda da pala do abrigo (camada A),
e ao qual não está associada nenhuma ocupação contínua;
o tramo intermédio, que corresponde as ocupações contínuas registadas
no abrigo (da camada B à camada E), mais bem conservadas no registo estratigráfico
devido à abundância de artefactos e à acelerada e crescente deposição de sedimentos
registada durante o Neolítico;
o tramo inferior que corresponde à primeira ocupação humana do abrigo
(camada F) que subjaz aos blocos depositados aquando o primeiro episódio de
abatimento da pala do mesmo.

22
Modelo crono-estratigráfico:
Fase 1
Corresponde à camada F e à ocupação datada do Epipaleolítico.
Fase 2
Marcada pelo surgimento da 1ª descontinuidade estratigráfica: primeira fase
de abatimento da pala do abrigo e deposição dos grandes blocos que marcam a
transição da camada F para a camada E. Caracteriza-se pela baixa deposição
sedimentar, o que poderá estar relacionado com a fixação dos solos causada pela
expansão das florestas durante o Holocénico Inicial.
Fase 3
Marcada pela possível introdução da agricultura. Grande aumento do ritmo de
sedimentação de origem argilosa registada nas camadas E a B, provavelmente
relacionado com a desflorestação de origem antrópica para abertura de campos de
cultivo e pastoreio durante o Neolítico Antigo e com a ocupação contínua do abrigo.
Fase 4
2ª Descontinuidade estratigráfica após a formação da camada B, devido ao
provável abandono do Maciço Calcário pelas comunidades Calcolíticas e consequente
fim da exploração agro-pastoril deste território, que por sua vez levou a um novo
desenvolvimento das florestas e fixação dos solos. Fim das ocupações contínuas do
abrigo. Deposições posteriores de vestígios datados da Idade do Ferro e da Época
Romana, resultantes de ocupações esporádicas durante estes períodos.
Fase 5
Abatimento definitivo da pala do abrigo, selando as ocupações anteriores.
Marcada pela ocorrência de ocupações esporádicas durante a Época Medieval e
Moderna.

23
1.3. Contextualização Cultural: A Neolitização
A Neolitização do extremo ocidental peninsular ocorre a partir de cerca de
5.400 B.C., aquando da chegada das primeiras populações portadores do chamado
“pacote neolítico” e de uma cultura material e rituais funerários próprios, que os
distinguia dos grupos mesolíticos então fixados em Muge, no Sado e no litoral
alentejano (ver Carvalho, 2003a, 2003b, 2008 e referências citadas por este autor).
Estas populações seriam, segundo os estudos realizados até ao momento acerca desta
temática, grupos humanos exógenos, originários de outras regiões do Ocidente
mediterrâneo, que chegaram à fachada atlântica peninsular por via marítima, e que se
instalaram ex novo em determinadas regiões, em territórios marginais aos ocupados
pelas populações mesolíticas, onde predominavam condições ambientais de tipo
mediterrâneo e que ofereciam boas condições para o desenvolvimento de uma
economia produtora, criando os chamados "enclaves neolíticos".
No que diz respeito concretamente à Estremadura portuguesa, tendo por base
datações absolutas atualmente disponíveis para o VI milénio a.C., os testemunhos
mais antigos da presença de populações neolíticas até ao momento identificados no
extremo ocidental peninsular, parecem ocorrer nesta região, nas grutas do Caldeirão e
Almonda, no Abrigo da Pena d'Água e no sítio de ar livre do Cerradinho do Ginete
(Carvalho, 2008, pp. 50).
Os primeiros grupos neolíticos que chegaram à Estremadura portuguesa
instalaram-se em territórios despovoados ou marginalmente explorados pelas
comunidades mesolíticas (Carvalho, 2003b, pp. 80). Após o estudo dos vários
contextos arqueológicos identificados na área envolvente do Arrife da Serra d'Aire,
foi possível concluir que este parece ter funcionado como a espinha dorsal da
neolitização na região estremenha, atraindo as primeiras populações neolíticas que
aqui se instalaram por terem encontrado condições geomorfológicas propícias à
instalação de uma economia produtora (Simões, 2012, pp. 52). Uma vez que, e apesar
da precariedade dos resultados da prospeção na planície, todos os assentamentos
humanos datados do Neolítico Antigo se localizam ao longo do Arrife, de forma a
aproveitar as potencialidades oferecidas por esta região de contexto ecótono, em
contacto com dois ambientes ecológicos e geográficos distintos. Houve, assim, a
ocupação sistemática de locais-chave onde se poderia praticar a exploração conjunta e

24
simultânea da serra e da planície (Carvalho, 2003a, pp. 135;142).
Apesar de o Epipaleolítico se encontrar bem documentado na região, tendo
sido identificadas ocupações humanas datadas deste período ao longo do Arrife,
inclusive na Pena d'Água, é de salientar o facto de se ter verificado que não existe
continuidade Mesolítico-Neolítico (Carvalho, 2003a, pp. 135). Em termos
económicos, é visível na transição do Mesolítico para o Neolítico uma tendência
generalizada para a substituição de uma economia assente em partes iguais em
recursos aquáticos e terrestres para uma economia baseada em exclusivo em recursos
terrestres, através da introdução de uma economia produtora na fachada atlântica da
Península Ibérica a partir de meados do VI milénio a.C., a que se fez referência acima
(Carvalho, 2008, pp. 50). Ainda que a caracterização económica do Neolítico Antigo
do Portugal meridional padeça de várias lacunas, devido sobretudo a limitações de
ordem tafonómica e ao facto de até ao momento ainda não ter sido possível identificar
restos carpológicos que comprovem a existência de práticas agrícolas desde o início
do Neolítico Antigo, foi possível verificar através de análises arqueozoológicas de
mamíferos (Valente, 1998; Póvoas, 1998; Carvalho, 2008, pp. 48), o
desenvolvimento da agricultura e da pastorícia desde o início da Neolitização nesta
região.
Relativamente às características residenciais, os assentamentos humanos
identificados em torno do Arrife da Serra d'Aire seriam, segundo os indicadores
económicos-funcionais disponíveis, geralmente de curta duração, uma vez que apenas
foram exumados vasos cerâmicos de pequena dimensão, como é possível verificar no
caso da Pena d'Água, à exceção dos vasos de armazenamento registados no sítio do
Forno do Terreirinho, que indicariam a existência de um habitat de carácter mais
permanente, ou pelo menos de maior duração, que obrigaria ao armazenamento dos
produtos produzidos (Carvalho, 2003a, pp. 135; 143).
Assim, tendo em conta a sua localização em ambiente ecótono, a prática de
caça associada ao pastoreio, o predomínio de rochas locais e o domínio de utensílios
domésticos, é possível afirmar que estes assentamentos praticariam um sistema de
povoamento assente numa estratégia de mobilidade residencial, onde por vezes estão
bem documentadas atividades económicas específicas, como a caça, a recolha e/ou
processamento de vegetais e a pastorícia (Carvalho, 2003a, pp. 135; 150).

25
CAPÍTULO II: AS AMOSTRAS E A MICROFAUNA
2. Amostras estudadas
Para a realização deste estudo foi apenas utilizada uma fração da totalidade dos
sacos de sedimento recolhidos durante as campanhas arqueológicas realizadas entre
1992 e 2000 no Abrigo da Pena d'Água. Foram estudadas no total oito amostras de
sedimento provenientes dos quadrados N29, N30, L29 e L30 e das camadas
arqueológicas F e E, correspondentes às ocupações humanas do abrigo mais antigas
datadas do Epipaleolítico e do Neolítico Antigo. Estas foram recolhidas no decorrer das
campanhas arqueológicas realizadas em 1993 e 1994.
Na figura 5, abaixo apresentada, é possível visualizar a localização destes
quadrados e respetivas camadas arqueológicas na planta geral da escavação.
Figura 5: planta geral da escavação (quadrados escavados à esquerda, camadas arqueológicas em baixo e
linha denteada correspondente ao limite do depósito após ação da retroescavadora (Carvalho, 1998, pp.
45).

26
No quadro seguinte (Quadro 3) é possível verificar os quadrados e respetivas
camadas arqueológicas alvo de estudo neste trabalho.
Cada amostra possuía cerca de 10 Litros de sedimento (entre 7 a 10 Kg de
sedimento), que correspondiam a cada um dos níveis artificiais escavados com cerca de
5 cm de altura por cada camada arqueológica.
As amostras encontravam-se armazenadas em sacos de plástico acompanhados
por etiquetas com a respetiva informação arqueológica (sigla do sítio arqueológico,
quadrado, camada e data da recolha).
Camada Quadrado Data da recolha
L30 F2 16/09/1993
N29 E10 07/10/1993
N30 E1 23/09/1993
N30 E3 07/09/1993
N30 E4 07/10/1993
L29 E1 23/09/1993
L29 E2 24/09/1993
L29 E5 06/09/1994
Quadro 3: quadrados e respetivas camadas arqueológicas das amostras de sedimento arqueológico
analisado.

27
3. Metodologia de estudo
A fim de realizar este estudo foi necessário estabelecer uma metodologia de
estudo, dividida em três fases principais, cada uma delas realizada em locais diferentes:
a primeira fase teve lugar no Museu Nacional de História Natural e da Ciência (Lisboa),
a segunda no Laboratório de Arqueociências (LARC) (Lisboa) e a terceira no Consejo
Superior de Investigaciones Cientificas (CSIC) (Madrid).
A primeira fase do trabalho consistiu no tratamento e na crivagem a água das
amostras de sedimento a fim de recolher os restos osteológicos das microfaunas.
Antes de iniciar a crivagem do sedimento das amostras, este foi pesado e
procedeu-se ao registo manual do quadrado, da camada e da data a que cada um destes
correspondia, bem como do peso e da data do dia em que foi realizado o processamento
da amostra. De seguida o sedimento foi retirado do saco de plástico em que se
encontrava e deitado em dois recipientes plásticos (alguidares) em igual quantidade
(para facilitar a desagregação da argila), aos quais foi anexada uma etiqueta autocolante
com a respetiva informação arqueológica, tendo sido sujeito a um tratamento de
desagregação dos elementos argilosos através de uma solução preparada com água
oxigenada a 120 volumes diluída em água a 20% (1/5 de água oxigenada por cada litro
de solução, num total de 3 litros de solução por cada amostra de sedimento processada).
Durante o processo de desagregação dos elementos argilosos (que durou pelo
menos 48 horas) o sedimento permaneceu no interior de uma câmara de exaustão de
vapores (Figura 6).
Figura 6: à esquerda balança utilizada para pesar os sacos de sedimento; ao centro garrafa da água
oxigenada a 120 volumes utilizada para desagregação dos elementos argilosos; e à direita os alguidares
com sedimento no interior da câmara de exaustão de vapores (Local: Museu Nacional de História Natural
e da Ciência).

28
A metodologia utilizada na crivagem foi a de crivagem com recurso a água
corrente e a duas malhas de crivo. Na primeira fase da crivagem a malha de crivo
utilizada foi de 4 mm para retenção dos elementos de maiores dimensões (fração grossa)
e na segunda fase foi utilizada uma malha de crivo de 0,6 mm para retenção dos
elementos de menores dimensões (fração fina) (Figura 7), que tornou possível a recolha
dos elementos osteológicos de dimensões muito reduzidas (como dentes soltos).
Figura 7: à esquerda malha de crivo de 4 mm utilizada na recolha da fração grossa; à direita malha de
crivo de 0,6 mm utilizada na recolha da fração fina (Local: Museu Nacional de História Natural e da
Ciência).
Após a crivagem do sedimento correspondente a cada amostra os materiais
arqueológicos daí resultantes foram armazenados em tabuleiros metálicos, aos quais foi
anexada uma etiqueta autocolante com a respetiva informação arqueológica, e
colocados numa estufa (marca Memmert) (Figura 8) para secagem dos mesmos a uma
temperatura nunca superior a 40º C, durante cerca de 24 horas.
Após a secagem, os materiais foram armazenados em dois sacos de plástico tipo
zip, um contendo a fração grossa e outro a fração fina, acompanhados por uma etiqueta
autocolante com a respetiva informação arqueológica.

29
Figura 8: tabuleiros metálicos utilizados na armazenagem dos materiais arqueológicos, no interior da
estufa Memmert utilizada para a secagem dos materiais arqueológicos após a sua lavagem (Local: Museu
Nacional de História Natural e da Ciência).
Concluída a crivagem da totalidade do sedimento das amostras deu-se início à
segunda fase do trabalho que consistiu na triagem, análise e estudo dos restos
osteológicos resultantes da crivagem.
Foi realizada uma primeira triagem com o objetivo de separar os restos
osteológicos e os restantes materiais arqueológicos que poderão ocorrer (sementes,
carvões, fragmentos de cerâmica, esquirolas…) dos restos pétreos que passaram na
malha do crivo. Foi analisada uma fração grossa, correspondente a restos faunísticos de
macro e microfauna, a carvões de maiores dimensões e a fragmentos líticos e cerâmicos
(Figura 9), e, uma fração fina, correspondente a microfauna e a carvões de menores
dimensões (Figura 10). No caso da fração fina houve recurso à lupa binocular (Olympus
SZ 60) (Figura 11).

30
Figura 9: elementos arqueológicos retidos na fração grossa: em cima restos faunísticos e carvões de
maiores dimensões; em baixo fragmentos de cerâmica e de líticos (Local: CSIC).
Figura 10: elementos arqueológicos retidos na fração fina: restos faunísticos e carvões de menores
dimensões (Local: CSIC).
Tendo em conta a elevada quantidade do conteúdo das amostras correspondentes
à fração fina e o tempo disponível para realização do estudo, foi necessário adotar uma
metodologia de triagem em que foram apenas triados 250 grs de cada saco com cerca de
1 kg, em vez da totalidade da amostra como aconteceu com as amostras da fração
grossa. O restante conteúdo das amostras foi armazenado em novos sacos de plástico
tipo zip, acompanhados por uma etiqueta com a respetiva informação arqueológica, a
fim de ser sujeito a estudo no futuro.

31
Figura 11: lupa binocular Olympus SZ 60, existente no LARC, utilizada na triagem e análise dos restos
microfaunísticos (Local: LARC).
Os restos osteológicos correspondentes à microfauna separados foram alvo de
uma nova triagem com recurso à lupa binocular, com o intuito de separar os restos
identificáveis (RI) de cada Ordem/ Família, dos restos osteológicos não identificáveis
(NI). Foram apenas separados os restos odontológicos e algumas mandíbulas por serem
aqueles que pelas suas características morfológicas permitem realizar identificações
taxonómicas específicas. Estes são denominados de restos diagnósticos.
Os restos diagnósticos considerados para cada uma das Ordens, Famílias e
Géneros existentes nas amostras foram:
Ordem Soricomorpha – foram separados:
Do Género Crocidura, todas as mandíbulas e dentes isolados exceto
incisivos.
Ordem Rodentia – foram separados:
Da Família Arvicolinae – Géneros Terricola, Microtus e Arvicola todos
os dentes molares superiores e inferiores (M1; M2; M3; M1; M2; M3) isolados e
mandíbulas com dentes inclusos, embora apenas tenham sido considerados os M1 para
realizar identificações ao nível do Género.

32
Da Família Muridae – Géneros Apodemus e Mus, todos os dentes
molares superiores e inferiores (M1; M2; M3; M1; M2; M3) isolados, embora apenas
tenham sido considerados os M1 para realizar identificações ao nível do Género.
Da Família Gliridae – Género Eliomys, todos os dentes pré-molares e
molares superiores e inferiores (Pm4; M1; M2; M3; Pm4; M1; M2; M3).
O critério utilizado é aquele que é habitualmente considerado na bibliografia de
referência (López-García, 2008).
Os restos odontológicos diagonóstico foram então alvo de uma análise
individual à lupa binocular tendo sido divididos por Ordem (Rodentia e Soricomorpha),
Família (Muridae, Arvicolidae e Gliridae), e por elemento anatómico (Pm4; M1; M2;
M3; Pm4; M1; M2; M3), e colocados em pasta adesiva (bostik) organizados em fiadas
paralelas (Figura 12). Esta tipo de organização (em fiadas paralelas no bostik) permitiu
por um lado a referenciação individual de cada dente (ver Tabela 1 em Anexo) e
permitirá se necessário uma fácil revisão do material no futuro, e por outro lado,
mantem os restos odontológicos intactos dado a sua fragilidade. Ao longo deste
procedimento foi realizada a identificação taxonómica, verificada a lateralidade e a
ocorrência de alterações relacionadas com os processos tafonómicos e pós-
deposicionais de cada dente.
Figura 12: restos odontológicos correspondentes à Família Muridae (à esquerda) e Arvicolidae (à
esquerda) organizados no bostik (Local: CSIC).
Durante este processo houve o registo sistemático manual dos resultados obtidos
em três tabelas distintas (ver Tabela 1, Tabela 2, Tabela 3 em Anexo). Uma primeira
onde foram registados todos os M1 correspondentes à Família Muridae existentes em

33
cada amostra (Tabela 1 em anexo), uma segunda onde foram registados por amostra
todos os restos odontológicos e mandíbulas correspondentes a cada uma das Ordens/
Famílias identificadas, tendo sido separados por Taxon e por elemento anatómico
(Tabela 2 em anexo), e, uma última, uma tabela geral onde foram incluídos todos os
resultados da análise e foram tidos em conta os seguintes parâmetros: informação
arqueológica original (quadrado, camada arqueológica, data e fração), Classe, Ordem,
Família, Género, Taxon, identificação anatómica, lateralidade, quantidade e processo
tafonómico quando existente (Tabela 3 em anexo). Estes dados foram posteriormente
introduzidos numa base de dados informatizada (Programa SPSS), para tratamento
estatístico.
A identificação anatómica e taxonómica dos restos osteológicos foi realizada
com recurso a comparações com os esqueletos disponíveis nas coleções de referência
existentes no Laboratório de Arqueociências (Moreno-Garcia et al, 2003; Pimenta &
Moreno-Garcia, 2004), e em alguns casos através da consulta de fontes bibliográficas
específicas mencionadas no texto.
Para atingir algumas determinações específicas, caso do Género Microtus,
tornou-se necessário efetuar medições de parâmetros odontométricos (Figura 13), apesar
de estes se terem mostrado pouco conclusivos dado o estado de conservação dos restos
odontológicos.
Figura 13: medições realizadas num M1 de Microtus (Terricola) duodecimcostatus/ lusitanicus: à
esquerda comprimento total e à direita largura do complexo T4 e T5 (Quadrado L30/ Camada F2) (Local:
CSIC).

34
Para o efeito foi utilizada a Lupa binocular Leica M 165 C com câmara
fotográfica DFC 490 (Figura 14), disponibilizada pelo Centro de Ciencias Humanas y
Sociales (CCHS) do Instituto de História do CSIC, em Madrid, Espanha.
Figura 14: lupa binocular Leica M 165 C, existente no CSIC em Madrid, utilizada na realização das
medições de parâmetros odontométricos e das fotografias ao material odontológico (Local: CSIC).
Após a análise dos restos odontológicos procedeu-se à quantificação do material
faunístico separado em cada uma das amostras: contagem dos restos determinados
(NRD), dos restos não determinados (ND) e da totalidade de restos (NTR). A totalidade
dos restos apenas identificados a nível anatómico (NRDa) foi incluída na totalidade dos
restos não determinados (ND). Não se procedeu à contagem da totalidade de restos
osteológicos existentes em cada amostra devido à elevada quantidade e elevadíssimo
grau de fragmentação dos mesmos, tendo-se optado apenas pela contagem dos restos
odontológicos separados em cada uma das amostras analisadas.
Foi também realizado o cálculo do Número Mínimo de Indivíduos (NMI), com o
intuito de estabelecer a abundância relativa dos Taxones presentes na totalidade das
amostras analisadas, utilizando o elemento odontológico diagnóstico mais frequente e
tendo em consideração a sua lateralidade (Andrews, 1990). O cálculo do NMI permite
conhecer a proporção de indivíduos existentes em cada uma das espécies identificadas,

35
e, é apropriado para casos em que os processos tafonómicos tendem a concentrar os
restos osteológicos, como os causados por armadilhas naturais e pela ação direta de
predadores. (López-García, 2008, pp. 129)
A quantificação faunística além de permitir obter uma noção da quantidade de
restos osteológicos estudados, permite através do NMI compreender a abundância
relativa das espécies identificadas (Valente, 1997, pp. 83).
A última etapa do trabalho consistiu no registo fotográfico dos restos
odontológicos considerados mais relevantes para a ilustração de cada uma das espécies
identificadas, na Lupa binocular Leica M 165 C, com câmara fotográfica DFC 490,
disponibilizada pelo Centro de Ciencias Humanas y Sociales (CCHS) do Instituto de
História do CSIC, de Madrid, em Espanha.
No final da análise os restos odontológicos alvo de identificação anatómica e
taxonómica permaneceram armazenados em caixas de plástico individuais para cada
uma das amostras, acompanhadas por uma etiqueta autocolante com a respetiva
informação arqueológica original (Figura 15).
Figura 15: restos odontológicos de uma das amostras estudadas armazenados nas caixas individuais
(Local: CSIC).
36
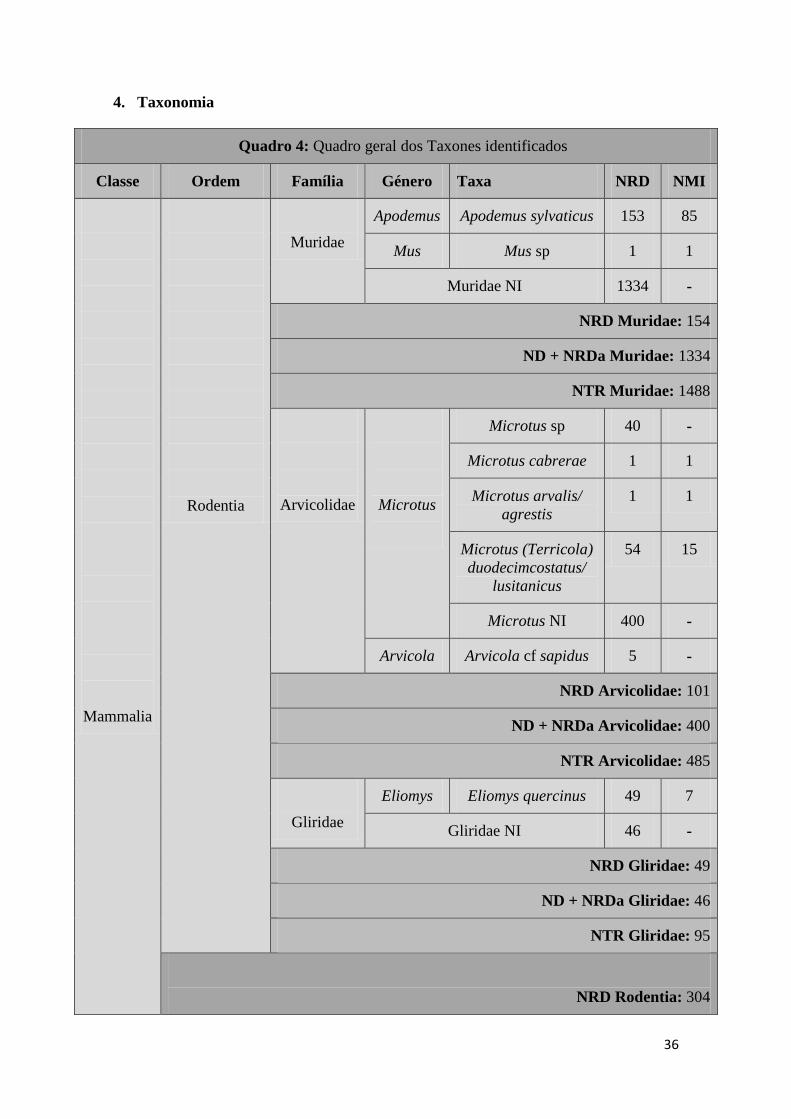
4. Taxonomia
Quadro 4: Quadro geral dos Taxones identificados
Classe Ordem Família Género Taxa NRD NMI
Mammalia
Rodentia
Muridae
Apodemus Apodemus sylvaticus 153 85
Mus Mus sp 1 1
Muridae NI 1334 -
NRD Muridae: 154
ND + NRDa Muridae: 1334
NTR Muridae: 1488
Arvicolidae
Microtus
Microtus sp 40 -
Microtus cabrerae 1 1
Microtus arvalis/
agrestis
1 1
Microtus (Terricola)
duodecimcostatus/
lusitanicus
54 15
Microtus NI 400 -
Arvicola Arvicola cf sapidus 5 -
NRD Arvicolidae: 101
ND + NRDa Arvicolidae: 400
NTR Arvicolidae: 485
Gliridae
Eliomys Eliomys quercinus 49 7
Gliridae NI 46 -
NRD Gliridae: 49
ND + NRDa Gliridae: 46
NTR Gliridae: 95
NRD Rodentia: 304
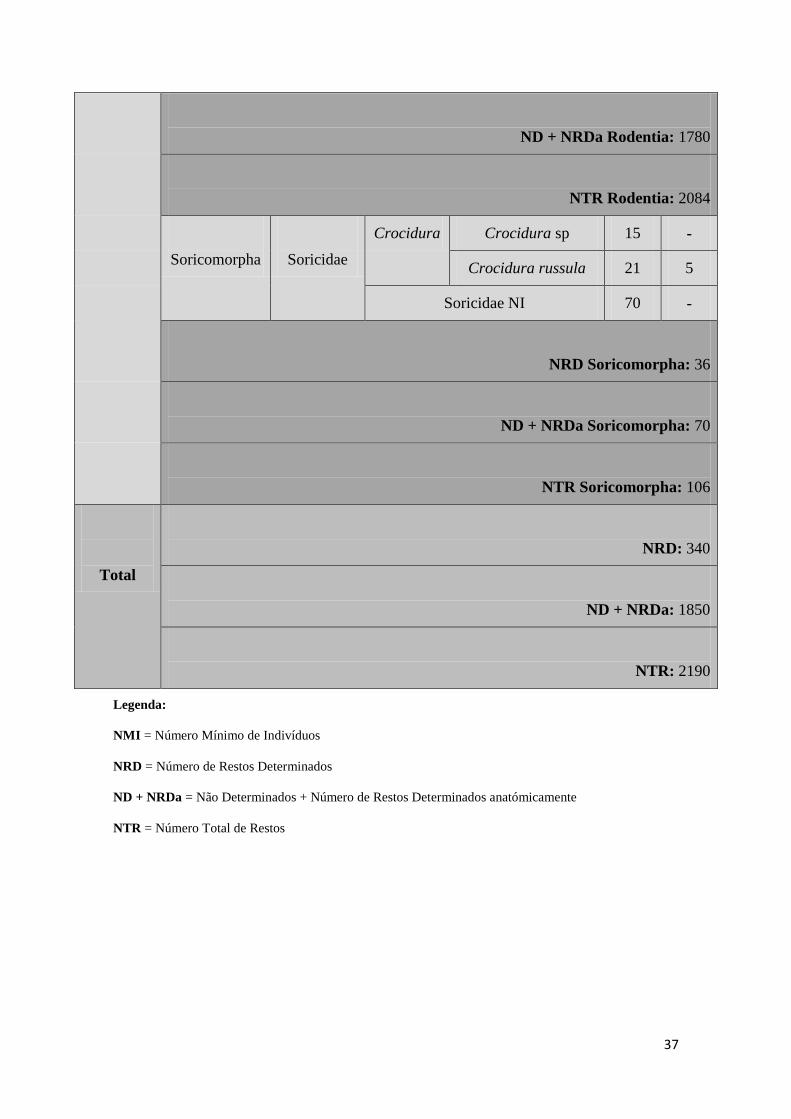
37
ND + NRDa Rodentia: 1780
NTR Rodentia: 2084
Soricomorpha
Soricidae
Crocidura Crocidura sp 15 -
Crocidura russula 21 5
Soricidae NI 70 -
NRD Soricomorpha: 36
ND + NRDa Soricomorpha: 70
NTR Soricomorpha: 106
Total
NRD: 340
ND + NRDa: 1850
NTR: 2190
Legenda:
NMI = Número Mínimo de Indivíduos
NRD = Número de Restos Determinados
ND + NRDa = Não Determinados + Número de Restos Determinados anatómicamente
NTR = Número Total de Restos

38
4.1. Sistemática
O critério utilizado na Sistemática tem por base o manual de Zooarqueologia de
J. Reitz e S. Wing (J. Reitz & S.Wing, 1999).
4.1.1. Micromamíferos
4.1.1.1. Ordem Soricomorpha
A Ordem Soricomorpha possui nas amostras analisadas uma amostragem
muitíssimo reduzida face à Ordem Rodentia, representando apenas 6% do Número Total
de Restos (Gráfico 4).
O material odontológico e osteológico (fragmentos de mandíbula e de maxilar)
além de se encontrar de modo geral em número reduzido, encontrava-se com fraco grau
de conservação, na maioria dos casos muito fragmentado. Deste modo, foram apenas
identificados 106 restos odontológicos e osteológicos correspondentes a esta Ordem
(Quadro 4), entre os quais mandíbulas sem dentes inclusos e incompletas, sendo que
nenhuma delas possuía o côndilo articular conservado, através do qual é possível
realizar identificações a nível taxonómico, fragmentos de maxilares, um deles com P4
incluso e dentes soltos, que por si só à expecção de um Pm4, também não permitiram
identificações ao nível da espécie, apenas ao nível do Género.
Após a análise da totalidade dos restos osteológicos e odontológicos
pertencentes a esta Ordem foi apenas identificada uma Família – Soricidae – e uma
espécie – Crocidura russula.
Sistemática
Ordem: Soricomorpha (Gregory, 1910)
Família: Soricidae (Ficher, 1817)
Subfamília: Crocidurinae (Wagler, 1832)
Género: Crocidura (Wagler, 1832)
Espécie: Crocidura russula (Wagler, 1832)

39
Nome comum: Musaranho-de-dentes-brancos
NMI: 5
Material diagnóstico: Pm4
Figura 16: fragmento de maxilar esquerdo com Pm4 incluso de Crocidura russula em vista labial e
carnial (Quadrado L29/ Camada E1) (Local: CSIC).
Observações morfológicas: o Género Crocidura caracteriza-se por possuir
dentes totalmente brancos, a parte oclusal do incisivo inferior lisa e as duas faces do
côndilo articular completamente fundidas.
Na Península Ibérica existem duas espécies de Crocidura: Crocidura russula e
Crocidura suaveolens. A diferenciação das duas espécies a partir da morfologia dos
dentes é por vezes difícil, incidindo sobretudo na observação de um conjunto de
caracteres odontométricos e das mandíbulas, que no caso de restos provenientes de
contextos arqueológicos, como é o caso dos restos aqui apresentados, nem sempre são
possíveis de observar tendo em conta o estado de conservação dos mesmos.
Neste caso, apesar de não ter sido identificada nenhuma mandíbula completa,
que possuísse o côndilo articular conservado, que permitiria fazer a diferenciação das
duas espécies (López-García, 2008), foi possível verificar em duas das amostras
analisadas que os restos odontológicos e osteológicos identificados de Crocidura
correspondem a Crocidura russula, uma vez que se verificou que as mandíbulas
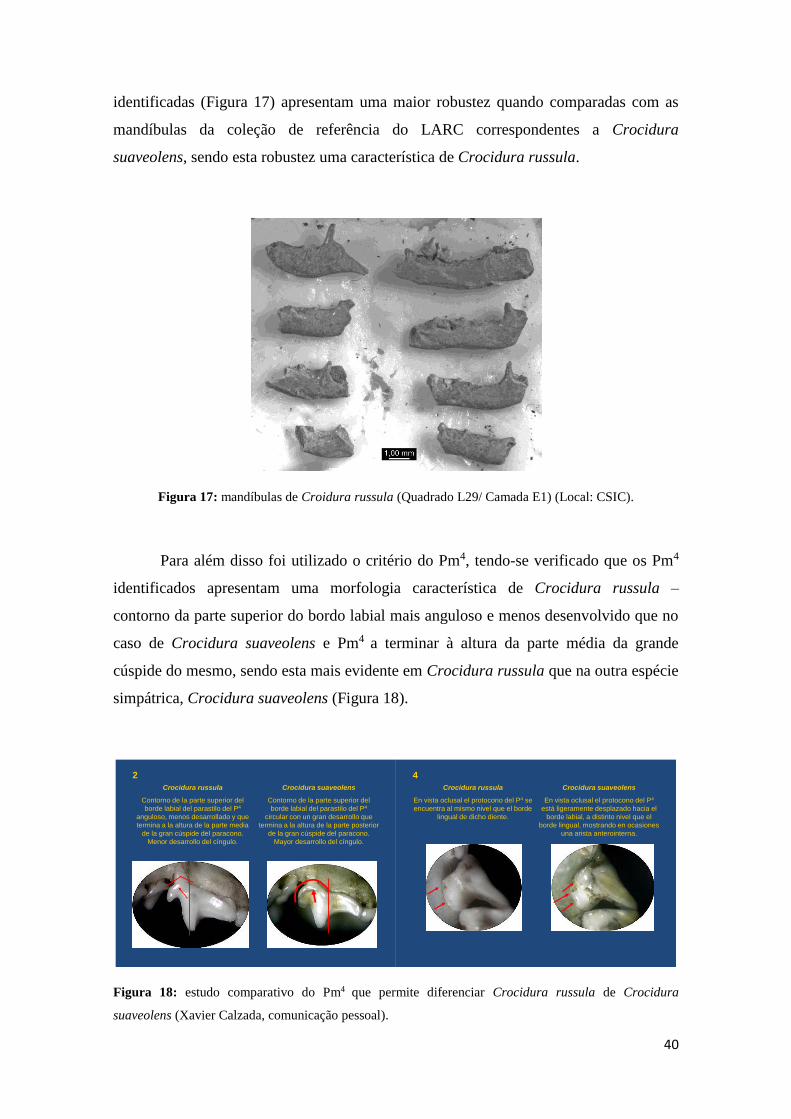
40
identificadas (Figura 17) apresentam uma maior robustez quando comparadas com as
mandíbulas da coleção de referência do LARC correspondentes a Crocidura
suaveolens, sendo esta robustez uma característica de Crocidura russula.
Figura 17: mandíbulas de Croidura russula (Quadrado L29/ Camada E1) (Local: CSIC).
Para além disso foi utilizado o critério do Pm4, tendo-se verificado que os Pm4
identificados apresentam uma morfologia característica de Crocidura russula –
contorno da parte superior do bordo labial mais anguloso e menos desenvolvido que no
caso de Crocidura suaveolens e Pm4 a terminar à altura da parte média da grande
cúspide do mesmo, sendo esta mais evidente em Crocidura russula que na outra espécie
simpátrica, Crocidura suaveolens (Figura 18).
2
Crocidura russula Crocidura suaveolens
Contorno de la parte superior del
borde labial del parastilo del P4
anguloso, menos desarrollado y que
termina a la altura de la parte media
de la gran cúspide del paracono.
Menor desarrollo del cíngulo.
Contorno de la parte superior del
borde labial del parastilo del P4
circular con un gran desarrollo que
termina a la altura de la parte posterior
de la gran cúspide del paracono.
Mayor desarrollo del cíngulo.
4
Crocidura russula Crocidura suaveolens
En vista oclusal el protocono del P4 se
encuentra al mismo nivel que el borde
lingual de dicho diente.
En vista oclusal el protocono del P4
está ligeramente desplazado hacia el
borde labial, a distinto nivel que el
borde lingual, mostrando en ocasiones
una arista anterointerna.
Figura 18: estudo comparativo do Pm4 que permite diferenciar Crocidura russula de Crocidura
suaveolens (Xavier Calzada, comunicação pessoal).

41
Habitat e distribuição geográfica: o musaranho-de-dentes-brancos é uma
espécie autóctone da Península Ibérica, atualmente amplamente distribuída por toda a
Europa Ocidental. Na Península Ibérica ocorre em todas as regiões, exceto em zonas de
grande altitude (Mapa 2). É uma espécie generalista, que apresenta requisitos de tipo
mediterrâneo e que ocupa áreas preferencialmente com espaços abertos e margens de
bosques com boa cobertura a nível do solo, até 1200/ 1600 metros de altitude, com uma
precipitação anual inferior a 1000 mm e com temperaturas médias acima de 5° C,
aparecendo ainda em zonas de cultivo (Palomo & Gisbert, 2007, pp. 129).
Mapa 2: distribuição atual de Crocidura russula na Península Ibérica (López-García, 2008, pp. 154).

42
4.1.1.2. Ordem Rodentia
A Ordem Rodentia é aquela que se encontra mais representada na totalidade das
amostras analisadas, representando 94% (Gráfico 4) do Número Total de Restos (NTR),
tendo sido identificados 2084 restos odontológicos (Quadro 4).
Os restos odontológicos correspondentes à Ordem Rodentia encontravam-se de
um modo geral na totalidade das amostras analisadas muito fragmentados e, sobretudo
no que concerne ao Género Apodemus num estado de grande fragilidade, possuindo em
muitos casos apenas o esmalte exterior do dente (Figura 19).
Figura 19: à esquerda restos odontológicos de Muridae onde é visível o elevado grau de fragmentação
dos dentes (Quadrado L29/ Camada E5); à direita M1 direito de Apodemus sylvaticus em que apenas se
conservou o esmalte lateral do dente (Quadrado N30/ Camada E3) (Local: CSIC).
No que concerne aos Géneros Microtus, Terricola e Eliomys estes encontravam-
se em reduzido número face ao Género Apodemus (Gráfico 5) e, na maioria dos casos
muito fragmentados, impossibilitando no caso do Género Microtus realizar
identificações a nível específico num grande número de restos odontológicos (ver ND
do Género Microtus apresentados no Quadro 4). Para além do grau de fragmentação, no
caso do Género Microtus os restos odontológicos possuíam em alguns casos concreções
que dificultavam a análise dos triângulos e a sua medição, e como tal impossibilitaram a
identificação da espécie (Figura 20).

43
Figura 20: à esquerda restos odontológicos de Arvicolidae onde é visível o elevado grau de fragmentação
dos dentes (Quadrado L29/ Camada E5); à direita M1 esquerdo de Microtus (Terricola)
duodecimcostatus/ lusitanicus onde é visível a existência de uma concreção em torno do dente (Quadrado
L29/ Camada E2) (Local: CSIC).
Ainda assim, foi possível identificar sete espécies de roedores, agrupadas em três
Famílias – Muridae, Arvicolidae e Gliridae e em cinco Géneros – Apodemus, Mus,
Microtus, Arvicola e Eliomys. A abundância de cada espécie foi calculada através do
Número Mínimo de Indivíduos (NMI), tendo por base o elemento osteológico mais
abundante e a sua lateralidade.
Sistemática
Murídeos
Ordem: Rodentia (Bowdich, 1821)
Família: Muridae (Illiger, 1811)
Género: Apodemus (Kaup, 1829)
Espécie: Apodemus sylvaticus (Linnaeus, 1758)
Nome comum: Rato-dos-Bosques
NMI: 85
Material diagnóstico: M1 esquerdo

44
Figura 21: M1 esquerdo de Apodemus sylvaticus (Quadrado L29/ Camada E1) (Local: CSIC).
Observações morfológicas: a principal característica morfológica de Apodemus
sylvaticus é o M1 possuir uma superfície oclusal baixa com seis cúspides principais
(López-García, 2008, pp. 223), e o facto dos tubérculos anteriores formarem um
conjunto de cinco lóbulos e possuírem dois tubérculos acessórios no lado externo,
principal característica deste Género (Moreno, 2007) e que o distingue do Género Mus.
Para a identificação desta espécie foram tidos em conta os aspetos morfológicos
nos M1 acima descritos.
Habitat e distribuição geográfica: Apodemus sylvaticus encontra-se
amplamente distribuído por toda a Europa possuindo uma distribuição homogénea em
todos os países mediterrânicos. Na Península Ibérica é um dos micromamíferos mais
abundantes encontrando-se presente em todo o seu território (Mapa 3). Atualmente
existem duas espécies de Apodemus na Península Ibérica: Apodemus sylvaticus e
Apodemus flavicollis. Contudo, o segundo ocorre apenas no norte peninsular (López-
García, 2008, pp. 223). Apodemus sylvaticus ocupa quase todos os tipos de habitats
existentes na Península Ibérica, mas sobretudo zonas de abundante e diversificada
cobertura arbustiva ou arbórea, dando preferência a áreas marginais, com cobertura
arbustiva ou arbórea homogéneas, em detrimento das áreas de bosque de grande escala
(Palomo & Gisbert, 2007, pp. 450). A razão do sucesso da sua distribuição em toda a

45
Península Ibérica reside no fato de se adaptar com grande facilidade aos diferentes
recursos alimentares disponíveis em cada momento (Jubete, 2007).
A região do Abrigo da Pena d'Água insere-se na zona de distribuição de
Apodemus sylvaticus, uma vez que a outra espécie deste Género apenas ocorre a norte.
Apodemus silvaticus é a espécie da Ordem Rodentia mais representativo no total
das amostras analisadas representando 45% do Número de Restos Determinados (NRD)
(Gráfico 5). Esta notável representação de Apodemus silvaticus no total das amostras
analisadas deve-se ao facto de este ser o micromamífero mais amplamente representado
em todo território da Península Ibérica.
Mapa 3: distribuição atual de Apodemus sylvaticus na Península Ibérica (López-García, 2008, pp. 224).

46
Género: Mus (Lataste, 1883)
Nome comum: Rato-doméstico/ Rato-das-hortas
NMI: 1
Material diagnóstico: M1 esquerdo
Figura 22: M1 esquerdo de Mus sp (Quadrado L29/ Camada E1) (Local: CSIC).
Observações morfológicas: a diferenciação morfológica dos restos
odontológicos correspondente às duas espécies deste Género é realizada através da
visualização de parâmetros morfológicos existentes no M1 e de parâmetros
odontométricos realizados a partir deste dente, contudo estes parâmetros nem sempre
são visíveis, não sendo nestes casos possível fazer a separação entre as duas espécies.
Tendo em conta que apenas foi identificado um M1 correspondente ao Género
Mus no total das amostras analisadas, não tendo sido possível identificar com certeza a
qual das duas espécies correspondia, optou-se por uma identificação deste resto apenas
ao nível do Género.

47
Habitat e distribuição geográfica: existem duas espécies simpátricas de Mus
na Península Ibérica: Mus musculus e Mus spretus. Mus spretus ocupa zonas de tipo
mediterrâneo sub-húmidas e semi-áridas, uma vez que devido aos seus escassos
requisitos hídricos pode ocupar territórios mais áridos e inacessíveis a outras espécies
de roedores, ainda assim mostra preferência por espaços abertos, áreas de matagal,
ambientes rochosos com alguma vegetação herbácea e áreas de cultivo de cereais,
enquanto que Mus musculus, comummente designado de rato-caseiro, por ser um
animal comensal por excelência, ocupa sobretudo zonas habitacionais ou com presença
do Homem e zonas em redor de campos de cultivo que detenham vegetação herbácea,
evitando os bosques e as áreas extremamente secas (Palomo & Gisbert, 2007).
Ambas as espécies são alóctones da Península Ibérica, tendo colonizado este
território a partir do Neolítico Antigo, encontrando-se associadas à expansão e
desenvolvimento da Neolitização na Europa e por conseguinte na Península Ibérica. A
sua introdução na Península Ibérica está diretamente relacionada com a introdução da
agricultura neste território, ainda que Mus spretus ao contrário de Mus musculus não
seja uma espécie comensal do Homem, contudo habita como referido acima
preferencialmente zonas de cultivo de cereais e espaços desflorestados, os quais foram
sendo criados através das desmatações levadas a cabo a partir do Neolítico Antigo.
Mus spretus é uma espécie silvestre de “exterior”, não florestal, proveniente do
Norte de África, que migrou para a Península Ibérica durante o Neolítico Antigo graças
ao desenvolvimento da navegação no Mar Mediterrâneo, colonizando toda a Europa
Ocidental mediterrânea e sobretudo a Península Ibérica e o Sul de França. As
desmatações crescentes realizadas pelo Homem a partir do Neolítico Antigo para a
criação de campos de cultivo e de pastorícia, tornaram possível a criação de biótopos
favoráveis ao desenvolvimento desta espécie, incapaz de viver em zonas de bosque e,
possibilitaram a sua colonização (Bonhomme & Thaler, 1988).
Por outro lado, Mus musculus como já mencionado acima é uma espécie
comensal, proveniente provavelmente do Próximo Oriente e que migrou para Ocidente
durante o Neolítico Antigo acompanhando o Homem durante a Neolitização, à medida
que este ia desenvolvendo a agricultura e construindo abrigos para guardar os
excedentes produzidos e, que tal como a sua espécie simpátrica colonizou toda a Europa
Ocidental mediterrânea e sobretudo a Península Ibérica e o Sul de França (Bonhomme
& Thaler, 1988).
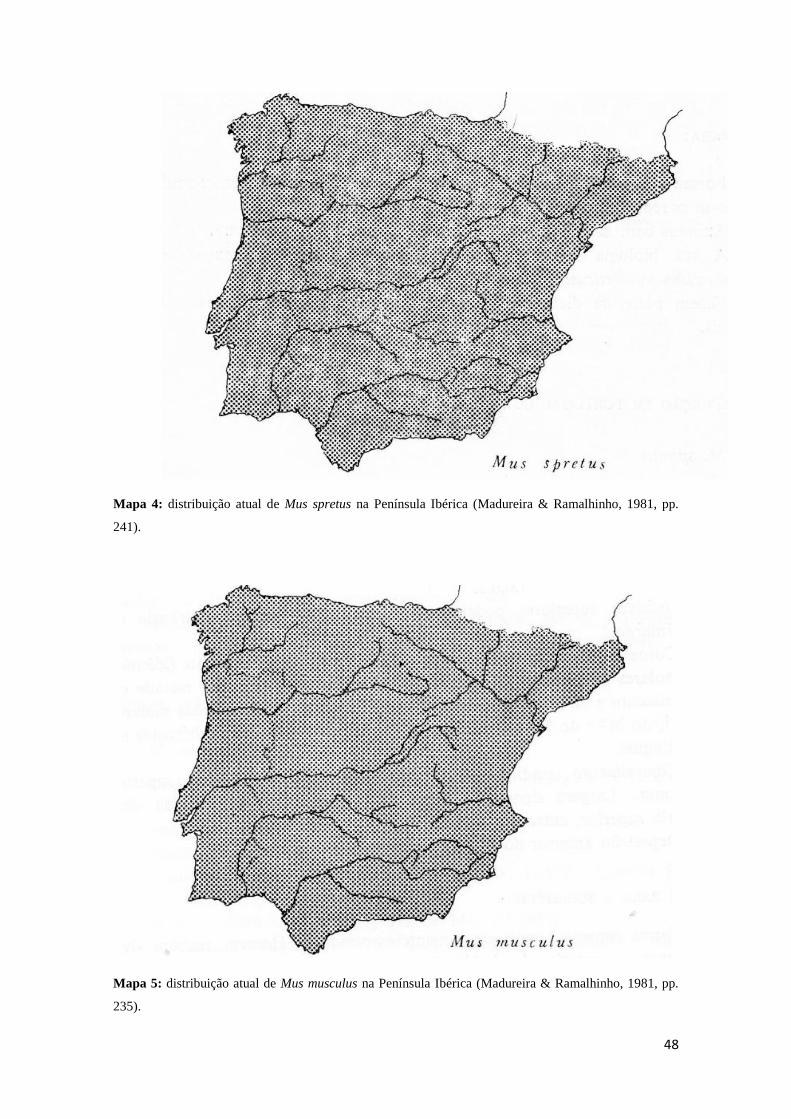
48
Mapa 4: distribuição atual de Mus spretus na Península Ibérica (Madureira & Ramalhinho, 1981, pp.
241).
Mapa 5: distribuição atual de Mus musculus na Península Ibérica (Madureira & Ramalhinho, 1981, pp.
235).

49
Arvicolídeos
Género Microtus
Este Género agrupa um grande número de roedores de pequena dimensão,
vulgarmente conhecidos como ratos-toupeiros. O critério morfológico que os caracteriza
é a presença do complexo T4-T5 do M1 ser fechado. Todas as espécies deste Género,
bem como as do Sub-género Terricola, possuem molares de crescimento contínuo,
como tal não apresentam raízes, e um esmalte distinto, comummente denominado de
esmalte tipo “Microtus”, por ser mais largo na metade mesial que na distal dos
triângulos oclusais dos molares (López-García, 2008, pp. 195).
Família: Arvicolidae (Gray, 1821)
Género: Microtus (Schrank, 1798)
Espécie: Microtus cabrerae (Thomas, 1906)
Nome comum: Rato-de-cabrerae
NMI: 1
Material diagnóstico: mandíbula direita com M1 e M2 inclusos.
Figura 23: mandíbula direita de Microtus cabrerae com M1 e M2 inclusos (Quadrado L29/ Camada E2)
(Local: CSIC).

50
Observações morfológicas: em termos morfológicos Microtus cabrerae
caracteriza-se pelo elevado tamanho e largura do M1 e pelo facto de todos os triângulos
serem fechados.
Habitat e distribuição geográfica: Microtus cabrerae é descendente do fóssil
Microtus brecciensis e constituí o único representante deste Género na Península
Ibérica. É uma espécie endémica da Península Ibérica. A sua distribuição atual
encontra-se fragmentada muito provavelmente devido à pressão antrópica exercida
desde o Neolítico após a introdução da agricultura, que levou à redução das populações
de Microtus cabrerae causada pelo desaparecimento progressivo do seu habitat em
determinadas zonas (López-García, 2008, pp. 194). Atualmente em Portugal encontra-se
apenas a Sul (Mapa 6). Ocupa exclusivamente áreas de clima mediterrâneo detentoras
de cobertura herbácea que se mantenha verde ao longo de todo o ano e com um nível
freático elevado, tais como juncais, gramíneas perenes, margens de ribeiras temporárias,
solos alagados ou margens de cursos de água permanentes (Palomo & Gisbert, 2007,
pp. 430; Cabral, 2005).
Mapa 6: distribuição atual de Microtus cabrerae na Península Ibérica (López-García, 2008, pp. 195).

51
Espécie: Microtus arvalis/ agrestis (Linnaeus, 1761)
Nome comum: Rato-do-campo
NMI: 1
Material diagnóstico: M1 direito
Figura 24: M1 direito de Microtus arvalis/ agrestis (Quadrado N29/ Camada E10) (Local: CSIC).
Observações morfológicas: existem duas espécies simpátricas deste Micortus
na Peninsula Ibérica: Microtus agrestis e Microtus arvalis. Microtus agrestis
caracteriza-se por possuirno M1 seis triângulos fechados (três no complexo posterior e
três no complexo anterior) e por os triângulos T4/ T5 e T6/ T7 possuírem uma elevada
assimetria e alternância, característica que o distingue de Microtus arvalis. Contudo,
devido à grande semelhança existente entre os M1 destas duas espécies, a maioria dos
autores cita-as como Microtus arvalis/ agrestis, não fazendo a distinção entre as duas
espécies (López-García, 2008, pp. 198).
Neste caso optou-se também por manter a designação Microtus arvalis/ agrestis
por não ter sido possível diferenciar com certeza a qual das duas espécies correspondia
o M1 identificado, ainda que tendo em conta a atual distribuição das duas espécies no

52
território da Península Ibérica, é mais provável tratar-se de Microtus agrestis (ver Mapa
7 e 8).
Habitat e distribuição geográfica: Microtus agrestis ocupa atualmente uma
franja da Península Ibérica que se estende desde os Pirenéus até a Galiza e desde esta
zona até ao Sul de Portugal, na zona de Lisboa (Mapa 7). Ocupa preferencialmente
espaços abertos com pouca ou nenhuma pressão de pastoreio, húmidos com rios ou
zonas encharcadas nas proximidades, próximos de bosques e detentores de erva em
abundância (Palomo & Gisbert, 2007, pp. 435). Por seu lado, Microtus arvalis embora
possua uma distribuição geográfica semelhante a Microtus agrestis (Mapa 8), é uma
espécie mais generalista, com uma distribuição que se estende mais para Sul e que
ocupa múltiplos habitats desde pastos e prados, a boques, desde que não muito cerrados,
dando preferência a espaços abertos e pouco húmidos, evitando as zonas encharcadas e
com vegetação alta (Palomo & Gisbert, 2007, pp. 437).
Mapa 7: distribuição atual de Microtus agrestis na Península Ibérica (López-García, 2008, pp. 203).

53
Mapa 8: distribuição atual de Microtus arvalis na Península Ibérica (López-García, 2008, pp. 203).
Sub-género Terricola
As características morfológicas que diferenciam o Género Microtus do seu Sub-
género Terricola são a presença no M1 de um triângulo aberto em T4 e o triângulo T4 e
T5 formarem o chamado rombo “pitymiano” (López-García, 2008, pp. 211).
Atualmente existem na Península Ibérica duas espécies simpátricas deste Sub-
género: Microtus (Terricola) duodecimcostatus e Microtus (Terricola) lusitanicus.
Família: Arvicolidae (Gray, 1821)
Género: Microtus (Schrank, 1798)
Sub-género: Terricola (Fatio, 1867)
Espécie: Terricola duodecimcostatus/ lusitanicus (Sellys-
Longchamps, 1839)
Nome comum: Rato-toupeiro/ Rato-toupeiro-mediterrânico

54
NMI: 15
Material diagnóstico: M1 esquerdo
Figura 25: M1 esquerdo de Microtus (Terricola) duodecimcostatus/ lusitanicus (Quadrado N30 / Camada
E4) (Local: CSIC).
Observações morfológicas: a diferenciação morfológica destas duas espécies é
complicada contudo, Microtus (Terricola) duodecimcostatus apresenta no M1 o
triângulo T6 aberto e com uma disposição quase simétrica e em paralelo com o
triângulo T7, formando um segundo rombo “pitymiano”, enquanto que Microtus
(Terricola) lusitanicus apresenta no M1 o segundo rombo pitimiano (com disposição
simétrica e em paralelo do triângulo T6 com T7) relativamente isolado da parte anterior
do dente (López-García, 2008, pp. 211). Além dos parâmetros morfológicos é ainda
possível realizar alguns parâmetros odontométricos no M1 que permitem separar as duas
espécies, nomeadamente: a relação do comprimento máximo do M1 com o comprimento
máximo do rombo “pitymiano”, obtido na diagonal entre o T4 (4ª triângulo) e o T5 (5ª
triângulo) na face interna do esmalte.
Contudo, no caso de restos odontológicos provenientes de contextos
arqueológicos esta separação nem sempre é possivel, tendo em conta o grau de
conservação destes restos, nem sempre possuem os triângulos completos. Neste sentido,

55
neste caso optou-se igualmente por identificar os M1 correspondentes a este Sub-género
mantendo o nome das duas espécies simpátricas, uma vez que foram identificadas
características morfológicas correspondentes às duas e, na impossibilidade de realizar
parâmetros odontométricos conclusivos nos M1, optou-se por não fazer a separação das
duas espécies.
Habitat e distribuição geográfica: Microtus (Terricola) duodecimcostatus é
uma espécie endémica da Península Ibérica, encontrando-se em grande parte do
território do quadrante peninsular sul (Mapa 10). Ocupa zonas de clima mediterrâneo,
com altitudes que vão desde o nível do mar até aos 3000 m de altitude, detentoras de
espaços abertos, tanto naturais como agrícolas, comsolos húmidos, fáceis de escavar e
com abundante cobertura herbácea. Por sua vez, Microtus (Terricola) lusitanicus,
também uma espécie endémica da Península Ibérica, distribui-se pelo quadrante
peninsular norte (Mapa 11), em zonas que vão desde o nível do mar até 200 m de
altitude, sendo que a sua ocorrência é condicionada pela existência de solos macios e
húmidos com uma densa cobertura vegetal (Palomo & Gisbert, 2007, pp. 423/425).
A zona ocupada pelo Abrigo da Pena d'Água localiza-se numa zona de simpatria
das duas espécies do Sub-género Terricola (Mapa 9).
Mapa 9: distribuição de Microtus (Terricola) duodecimcostatus e de Microtus (Terricola) lusitanicus na
Península Ibérica, e a zona de simpatria entre as duas espécies (Adaptado por Santos, 2009, a partir de
Madureira, 1984, Mitchell-Jones et al. 1999, Cotilla & Palomo, 2007, Mira e Mathias, 2007).
56
Mapa 10: distribuição atual de Microtus (Terricola) duodecimcostatus na Península Ibérica (López-
García, 2008, pp. 215).
Mapa 11: distribuição atual de Microtus (Terricola) lusitanicus na Península Ibérica (López-García,
2008, pp. 215).

57
Género Arvicola
A principal característica morfológica que define este Género, além da sua
grande dimensão, é que o facto da parte posterior do M1 ser precedida por três triângulos
fechados. Por outro lado, o Género Arvicola possui um esmalte mais espesso que o
Género Microtus.
Atualmente, na Península Ibérica existem duas espécies de Arvicola: Arvicola
sapidus e Arvicola terrestris (López-García, 2008, pp. 217).
Família: Arvicolidae (Gray, 1821)
Género: Arvicola (Miller, 1908)
Espécie: Arvicola cf sapidus (Miller, 1908)
Nome comum: Rato-de-água
Devido ao elevado grau de fragmentação
dos cinco restos odontológicos identificados
correspondentes a este Género, não foi possível
calcular o Número Mínimo de Indivíduos nem
realizar uma identificação segura ao nível da
espécie, contudo pode considerar-se tratar-se de
Arvicola sapidus, uma vez que a área onde se
encontra o Abrigo da Pena d'Água insere-se na
zona de distribuição deste Arvicola (Mapa 12),
sendo que a outra espécie deste Género, Arvicola
terrestris, embora ocupe grande parte da Europa
não se encontra representado na parte central e sul
da Península Ibérica, fixando-se apenas
exclusivamente no norte de Espanha (Mapa 13)
(López-García, 2008, pp. 218). Para além disto,
nos restos odontológicos analisados não foram
identificadas as características morfológicas de Arvicola terrestris.
Figura 26: fragmento de M1 esquerdo de
Arvicola cf sapidus (Quadrado N30/
Camada E3) (Local: CSIC).

58
Observações morfológicas: morfologicamente Arvicola sapidus diferencia-se
de Arvicola terrestris por possuir no loop anterior do M1 no lado lingual um ângulo
agudo junto à extremidade interior, enquanto que Arvicola terrestris no loop anterior do
M1 possui um contorno arredondado. Além disso, Arvicola sapidus possui o esmalte
mais espesso na parte distal dos triângulos que na parte mesial, ao contrário do que
acontece com Arvicola terrestris, que possui o esmalte mais espesso na parte mesial dos
triângulos que na parte distal (López-García, 2008, pp. 217).
Habitat e distribuição geográfica: Arvicola sapidus é uma espécie endémica
da Península Ibérica, distribuindo-se por todo o seu território (Mapa 12). É um roedor
semi-aquático que vive associado a cursos ou massas de água estáveis, com abundante
vegetação herbácea ou matagal nas margens, dando preferência a solos brandos, como
prados húmidos e charcas, que lhes permitem escavar facilmente tocas (Ventura, 2007).
Mapa 12: distribuição atual de Arvicola sapidus na Península Ibérica (López-García, 2008, pp. 220).

59
Mapa 13: distribuição atual de Arvicola terrestris na Península Ibérica (López-García, 2008, pp. 220).
Glirideos
Os Glirideos caracterizam-se morfologicamente pela existência de raízes
molares braquidontes e por possuírem molares com uma superfície oclusal côncava
atravessada por sulcos transversais, separados por vales (lofodontos), em que é quase
impossível distinguir as cúspides (López-García, 2008, pp. 225).
Família: Gliridae (Thomas, 1897)
Género: Eliomys (Wagner, 1840)
Espécie: Eliomys quercinus (Linnaeus, 1766)
Nome comum: Leirão ou Rato-dos-pomares
NMI: 7
Material diagnóstico: M2 direito

60
Figura 27: M1 direito de Eliomys quercinus (Quadrado L29 / Camada E1) (Local: CSIC).
Observações morfológicas: morfologicamente os molares de Eliomys quercinus
caracterizam-se por possuírem uma superfície oclusal côncava em que as cristas
principais atravessam o dente em sentido lábio-lingual destacando as cúspides
principais (López-García, 2008, pp. 226).
Habitat e distribuição geográfica: Eliomys quercinus encontra-se em quase
toda a Europa. Na Península Ibérica ocupa a generalidade do seu território desde o nível
do mar até níveis superiores a 1500 m de altitude (Mapa 14) (Palomo & Gisbert, 2007,
pp. 392). É uma espécie generalista capaz de habitar numerosos habitats terrestres e
arbóreos. Apresenta uma grande variação morfológica em tamanho, que diminui com o
aumento da altitude, na cor e nas características biológicas (Palomo & Gisbert, 2007,
pp. 393). Encontra-se frequentemente associado a zonas pedregosas, de matagal, a
campos desflorestados, diferentes tipos de bosque, sobretudo bosques caducifólios
pouco densos, e é ainda frequente em zonas rurais.

61
Mapa 14: distribuição atual de Eliomys quercinus na Península Ibérica (Fonte: López-garcìa, 2008, pp.
228).

62
CAPÍTULO III: DISCUSSÃO DOS RESULTADOS
Após realizada a análise dos restos odontológicos e os osteológicos passíveis de
identificação importa agora expor determinados pontos considerados relevantes para a
caracterização do depósito microfaunístico fóssil em questão.
5. Processos deposicionais – agentes tafonómicos
Existem dois principais tipos de agentes tafonómicos responsáveis pelos
processos deposicionais de restos faunísticos em contextos arqueológicos, um de origem
animal e outro de origem antrópica (o Homem) (Andrews, 1990).
Uma vez que não foram identificadas marcas nos restos osteológicos aqui
analisados que pudessem relacionar a sua deposição com a atividade humana e tratando-
se de amostras microfaunísticas, em que apenas em raras exceções a sua deposição se
encontra relacionada com agentes de origem antrópica, podemos afirmar que o agente
tafonómico responsável por esta acumulação é de origem animal.
Assim a nível tafonómico a composição dos Taxones identificados nas amostras
analisadas sugere que a sua presença neste contexto estará relacionada com dois
processos tafonómicos:
Acumulação originada por predadores, através da desagregação de
dejetos e/ ou regurgitações, que frequentaram o local como abrigo. Micromamíferos,
como os roedores e os insectívoros, são as presas que maioritariamente integram o
regime alimentar de predadores como as aves de rapina diurnas e noturnas e os
carnívoros de pequena e média dimensão, que em função do tipo de ingestão e digestão
das suas presas, são potenciais responsáveis pela acumulação de restos osteológicos de
microfaunas em depósitos arqueológicos (Andrews, 1990; Pecharromán, 1993).
Acumulação originada pela morte natural destes animais no interior do
abrigo. Muitos microvertebrados procuram refúgio no interior de grutas e abrigos
naturais acabando por aí morrer de forma natural.
Nomeadamente, Apodemus sylvaticus tem por hábito procurar refúgio no interior
de grutas ou abrigos naturais como a Pena d'Água. Também Eliomys quercinus procura
refúgio nestes contextos para hibernar nos meses de Inverno.
Contudo, para este caso em concreto, podemos considerar que a acumulação
destes micromamíferos embora possa ser resultado destes dois processos tafonómicos,

63
se deve sobretudo à ação de predadores, muito provavelmente aves de rapina noturnas,
sobretudo corujas ebufos, principais responsáveis por acumulações deste tipo em
contextos arqueológicos (Pecharromán, 1993).
Isto porque por um lado, foram sobretudo identificados restos osteológicos
correspondentes a roedores, sendo estes as principais presas deste tipo de aves noturnas.
Sendo que, efetivamente, em alguns casos podemos considerar que os restos
odontológicos de roedores analisados possuem vestígios relacionados com processos de
digestão embora pouco agressivos, típicos do processo de ingestão e digestão pouco
destrutivo destas aves de rapina (ingestão das suas presas inteiras e posterior expulsão
das partes não digestíveis do seu organismo através de regurgitações) (Andrews, 1990;
Pecharromán, 1993). E por outro lado, porque apenas foram registados graus de
desgaste significativos do esmalte num dos restos odontológicos analisados (M1 de
Apodemus sylvaticus – Figura 28), que nos permitui confirmara provável morte de
forma natural deste micromamíferono interior do abrigo por velhice. Embora fosse
visível a existência de exemplares mais jovens e outros adultos, sobretudo no caso dos
restos odontológicos de Apodemus sylvaticus, nos quais em alguns casos são bastante
proeminentes os lóbulos e noutros estes encontram-se já desgastados, de uma forma
geral não foram identificados graus de desgaste significativos, típico de um longo
período de uso.
Figura 28: M1 de Apodemus sylvaticus onde são visíveis maiores e menores níveis de desgaste do esmalte
(Quadrado L29/ Camada E1) (Local: CSIC).

64
6. Processos pós-deposicionais– Diagénese
Após a deposição de um determinado elemento osteológico em ambiente
sedimentar têm lugar uma série de processos que atuam no tecido destes elementos. A
estes processos dá-se o nome de diagénese (Lyman, 2001).
A diagénese pode ser desencadeada por fatores intrínsecos ao osso, tais como a
estrutura molecular e química do osso, a porosidade da sua superfície e o seu tamanho,
mas também por fatores extrínsecos, como o pH do sedimento em que foi depositado, a
presença ou ausência de água/ humidade, a existência de variações de temperatura e a
ação bacteriana (Lyman, 2001).
Assim, além dos agentes tafonómicos de origem animal e antrópica acima
mencionados, que atuam na formação de registos faunísticos fósseis, existem ainda
agentes tafonómicos de origem antrópica, animal e natural que afetam estes registos
após a sua deposição, estes denominam-se de agentes pós-deposicionais (Lyman, 2001;
Valente, 2000).
Os agentes pós-deposicionais de origem antrópica e animal apresentam-se para o
caso das microfaunas sobretudo como agentes perturbadores das deposições originais.
Nos restos osteológicos aqui analisados a perturbação provocada por agentes de
origem antrópica é visível através do estado carbonizado em que alguns elementos
odontológicos se encontram, sobretudo nos Géneros Apodemus, Eliomys e Crocidura
(Figura 29).
Figura 29: restos odontológicos correspondentes aos Géneros Apodemus (à esquerda) (Quadrado N30/
Camada E1), Crocidura (ao centro) (Quadrado N30/ Camada E3) e Eliomys (à direita) (Quadrado L29/
Camada E1) carbonizados (Local: CSIC).

65
O fogo é utilizado pelo Homem para várias finalidades, como iluminação, para
aquecimento, na confeção de alimentos, na defesa contra animais ferozes, entre outras
finalidades ligadas à esfera ritual e simbólica. Se o fogo for efetuado sobre depósitos
faunísticos fósseis pode deixar a sua marca sobre estes.
A outra possibilidade que pode ser apresentada como a causa do estado
carbonizado destes dentes é a sua exposição aos fogos florestais de ordem natural,
recorrentes nas florestas mediterrânicas, ou a partir do Neolitico Antigo causados
intencionalmente pelo Homem com o intuito de desflorestar e criar campos de cultivo.
A coloração que o osso exibe revela a intensidade e duração da exposição deste
ao fogo, correspondendo a diferentes etapas de intensidade térmica. No caso dos
elementos odontológicos da Pena d'Água identificados com marcas de fogo, estes
revelam ter estado em contacto com o fogo durante um período de tempo prolongado,
causado por uma combustão de alguma intensidade térmica, pois possuem uma
coloração negra uniforme, causada pela carbonização do colagénio do osso (Stiner et al,
1995).
Por último, os agentes de origem natural consistem em fenómenos físicos,
químicos e biológicos que atuam sobre a superfície do osso alterando a sua composição
física. Destacam-se os fenómenos erosivos – vento e circulação de água; fenómenos
químicos próprios do sedimento – pH ácido; e biológicos – atividade de invertebrados
(Micróbios) incorporados no sedimento e nas raízes (Jans, 2003).
Um dos principais fenómenos erosivos que se pensa poder ter condicionado a
conservação dos restos osteológicos e odontológicos aqui alvo de estudo foi a ação da
água proveniente da nascente sazonal existente junto à parede do abrigo e, que escorre
sazonalmente pela zona escavada (Póvoas, 1998). Esta terá causado alterações físico-
químicas substanciais nos restos, comprometendo a sua conservação. Como já
mencionado foi identificada uma quantia significativa de restos odontológicos,
sobretudo no caso dos murídeos, em que apenas subsistiu o esmalte do dente, tendo
desaparecido toda a dentina (Figura 30). O esmalte sendo a parte do dente mais
resistente sobreviveu à ação erosiva da água.

66
Figura 30: M1 de Apodemus sylvaticus em que apenas subsistiu o esmalte do dente (Quadrado L29/
Camada E1) (Local: CSIC).
A decomposição química causada pela ação da água no osso vai provocar a
perda do cologénio levando à sua dissolução, sendo que a velocidade de dissolução
aumenta com a existência de temperaturas elevadas, de pH´s extremos e quanto maior
for o tempo de exposição (Lyman, 2001).
No caso da Pena d'Água sabemos que a exposição dos restos osteológicos à
escorrência de água embora sazonal ocorria pelo menos anualmente, cessando apenas
nos períodos do ano de maior seca (Verão).
Quanto às alterações de temperatura, apesar do concelho de Torres Novas se
encontrar inserido numa região de clima mediterrâneo, é visível a existência de um
microclima de tipo continental devido à proximidade à Serra de Aire e Candeeiros, que
se traduz na existência de temperaturas baixas durante o Inverno e no Verão de
temperaturas bastante elevadas (Araújo & Zilhão, 1991).
Aquando da realização do estudo preliminar das microfaunas da Pena d'Água,
Liliana Póvoas fez referência ao mau estado de conservação dos restos osteológicos por
ela analisados, dando também como causa provável a atividade erosiva causada pela
ação da água da nascente, que terá “lavado” o sedimento e causado a dissolução dos
restos osteológicos (Póvoas, 1998). Também Maria João Valente, que se dedicou ao

67
estudo da macro e mesofauna proveniente deste mesmo sítio, dá conta do mau estado de
conservação destes restos, que se encontravam muito fragmentados, à exceção dos
provenientes da camada C, sendo praticamente impossível observar a sua superfície
original, devido à existência de alterações físico-químicas, também para Valente
causadas pelos movimentos que os materiais sofreram causados pela ação hídrica e pela
ação química da água sobre os restos faunísticos (Valente, 1998).
Apesar disto, os restos odontológicos em que se observou este fenómeno foram
na sua maioria passíveis de identificação. Contudo, este acontecimento condicionou a
quantidade original dos restos depositados dado o elevado nível de fragmentação que
estes apresentavam. Liliana Póvoas refere mesmo que o estado e conservação das
amostras comprometeu a identificação taxonómica dos restos odontológicos (Póvoas,
1998).
Além da ação química causada pela água ocorreu ainda outro tipo de ação
química, para a qual não foi possível identificar a causa, que levou a existência de
concreções em torno de alguns dos restos odontológicos analisados, sobretudo no que
diz respeito aos dentes de arvicolídeos, que em alguns casos não tornaram possível
retirar as medições dos seus triângulos para identificação da espécie correspondente
(caso do Sub-género Terricola como já mencionado acima na Sistemática), não tendo
sido possível proceder a limpeza dos mesmos na câmara de ultra-sons disponibilizada
pelo Museu Nacional da História Natural e da Ciência, sendo estes passíveis de uma
revisão futura.
No que diz respeito aos fenómenos pós-deposicionais de origem biológica, no
conjunto das amostras analisadas apenas foi identificado num dos restos osteológicos
analisados, numa mandíbula de Crocidura russula, marcas de carácter tafonómico
relacionadas com a ação radicular (raízes) (Figura 31).
As marcas de raízes na superfície dos ossos são provocadas pela acidez que as
raízes de algumas plantas possuem, provocando um padrão irregular de dissolução
(Andrews, 1997). As raízes podem ainda causar a fragmentação dos ossos ao atravessa-
los.

68
Figura 31: mandíbula com dentes inclusos de Crocidura russula com marcas relacionadas com a
atividade radicular (Quadrado N30/ Camada E1) (Local: CSIC).
6.1. Estado das amostras
À semelhança do já observado na publicação de 1998 (Póvoas, 1998), os restos
que integram as amostras analisadas apresentam um elevado grau de fragmentação e são
essencialmente compostos por material odontológico.
Deste fator resulta o fato do Número de Restos Determinados (NRD) ser muito
inferior ao Número Total de Restos (NTR) (Gráfico 1).
15,50%
84,50%NTR
NRD
0,00% 20,00% 40,00% 60,00% 80,00% 100,00%
Gráfico 1: Percentagem do NTR face ao NRD

69
Os elementos ósseos do esqueleto pós-cranial encontram-se escassamente
representados e num estado de fragmentação que impede a sua identificação anatómica.
Figura 32: vista geral do material osteológico de umas das amostras estudadas onde é visível o seu
elevado grau de fragmentação (Local: CSIC).
Este facto leva-nos a levantar questões relacionadas com a capacidade de
conservação dos restos osteológicos face aos restos odontológicos.
Por um lado, sabemos à partida que nem todas as partes do esqueleto,
especialmente no caso dos M amíferos, resistem da mesma forma aos processos pós-
deposicionais e aos agentes erosivos, situação que implica diferentes graus de
conservação. Os ossos de estrutura esponjosa, como os que constituem o esqueleto
axial, são menos resistentes que os ossos de estrutura densa, que compõem as
extremidades dos membros (Beherensmeyeer, 1978). Por sua vez, os dentes devido ao
facto de se encontrarem revestidos por esmalte são a parte do esqueleto que melhor se
conserva, uma vez que o esmalte pelas suas características físicas constituiu a parte mais
resistente do esqueleto de um Mamífero (Willson, 1990).
Por outro, partindo do princípio que estes dentes se encontravam inseridos nos
crânios ou nas mandíbulas, ou numa parte do mesmos, aquando da sua deposição, tendo
em conta a fraca representação destes elementos osteológicos, salvo algumas
mandíbulas incompletas e um fragmento de crânio, podemos assumir que ocorreu um
processo de decomposição química destes ossos. Este fator, além de estar diretamente
relacionado com os processos erosivos que aqui tiveram lugar, pode ainda dever-se ao
facto de os dentes analisados corresponderem sobretudo a exemplares juvenis ou sub-
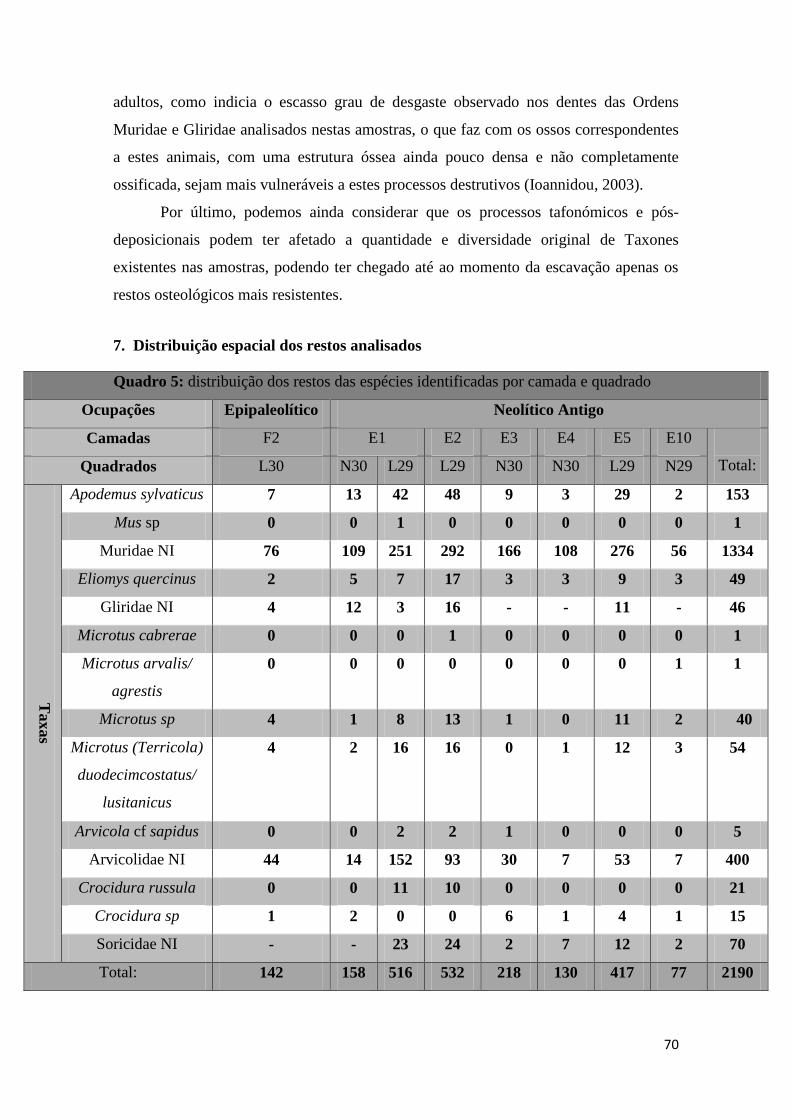
70
adultos, como indicia o escasso grau de desgaste observado nos dentes das Ordens
Muridae e Gliridae analisados nestas amostras, o que faz com os ossos correspondentes
a estes animais, com uma estrutura óssea ainda pouco densa e não completamente
ossificada, sejam mais vulneráveis a estes processos destrutivos (Ioannidou, 2003).
Por último, podemos ainda considerar que os processos tafonómicos e pós-
deposicionais podem ter afetado a quantidade e diversidade original de Taxones
existentes nas amostras, podendo ter chegado até ao momento da escavação apenas os
restos osteológicos mais resistentes.
7. Distribuição espacial dos restos analisados
Quadro 5: distribuição dos restos das espécies identificadas por camada e quadrado
Ocupações Epipaleolítico Neolítico Antigo
Camadas F2 E1 E2 E3 E4 E5 E10
Total: Quadrados L30 N30 L29 L29 N30 N30 L29 N29
Taxas
Apodemus sylvaticus 7 13 42 48 9 3 29 2 153
Mus sp 0 0 1 0 0 0 0 0 1
Muridae NI 76 109 251 292 166 108 276 56 1334
Eliomys quercinus 2 5 7 17 3 3 9 3 49
Gliridae NI 4 12 3 16 - - 11 - 46
Microtus cabrerae 0 0 0 1 0 0 0 0 1
Microtus arvalis/
agrestis
0 0 0 0 0 0 0 1 1
Microtus sp 4 1 8 13 1 0 11 2 40
Microtus (Terricola)
duodecimcostatus/
lusitanicus
4 2 16 16 0 1 12 3 54
Arvicola cf sapidus 0 0 2 2 1 0 0 0 5
Arvicolidae NI 44 14 152 93 30 7 53 7 400
Crocidura russula 0 0 11 10 0 0 0 0 21
Crocidura sp 1 2 0 0 6 1 4 1 15
Soricidae NI - - 23 24 2 7 12 2 70
Total: 142 158 516 532 218 130 417 77 2190
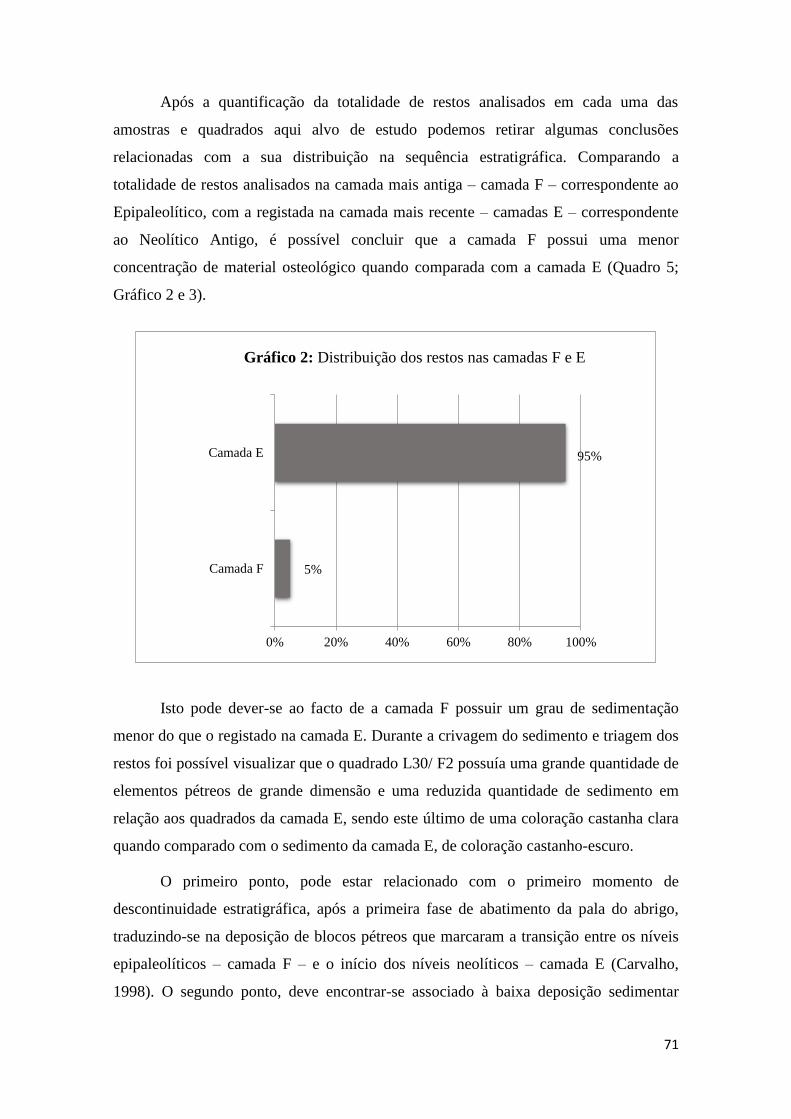
71
Após a quantificação da totalidade de restos analisados em cada uma das
amostras e quadrados aqui alvo de estudo podemos retirar algumas conclusões
relacionadas com a sua distribuição na sequência estratigráfica. Comparando a
totalidade de restos analisados na camada mais antiga – camada F – correspondente ao
Epipaleolítico, com a registada na camada mais recente – camadas E – correspondente
ao Neolítico Antigo, é possível concluir que a camada F possui uma menor
concentração de material osteológico quando comparada com a camada E (Quadro 5;
Gráfico 2 e 3).
Isto pode dever-se ao facto de a camada F possuir um grau de sedimentação
menor do que o registado na camada E. Durante a crivagem do sedimento e triagem dos
restos foi possível visualizar que o quadrado L30/ F2 possuía uma grande quantidade de
elementos pétreos de grande dimensão e uma reduzida quantidade de sedimento em
relação aos quadrados da camada E, sendo este último de uma coloração castanha clara
quando comparado com o sedimento da camada E, de coloração castanho-escuro.
O primeiro ponto, pode estar relacionado com o primeiro momento de
descontinuidade estratigráfica, após a primeira fase de abatimento da pala do abrigo,
traduzindo-se na deposição de blocos pétreos que marcaram a transição entre os níveis
epipaleolíticos – camada F – e o início dos níveis neolíticos – camada E (Carvalho,
1998). O segundo ponto, deve encontrar-se associado à baixa deposição sedimentar
5%
95%
Camada F
Camada E
0% 20% 40% 60% 80% 100%
Gráfico 2: Distribuição dos restos nas camadas F e E

72
provocada pela fixação dos solos em torno do abrigo devido à expansão florestal
ocorrida durante o Holocénico Inicial, sendo que a partir da camada E é observável
precisamente o fenómeno oposto, havendo uma maior deposição sedimentar de tipo
argiloso, relacionada com a introdução da agricultura na área em torno do abrigo e,
consequentemente com a desflorestação levada a cabo pelo Homem para criação de
campos de cultivo e pastorícia a partir do início do Neolítico Antigo (Carvalho, 1998).
Quando analisado o Gráfico 3, olhando para os quadrados da camada E podemos
concluir por um lado, que os quadrados N29 e N30 possuem uma menor concentração
de restos osteológicos quando comparados com o quadrado L29. Este acontecimento
deve-se certamente ao facto dos quadrados N29 e N30 se encontrarem na proximidade
da escorrência de água proveniente da nascente, ocorrendo uma maior “lavagem” do
sedimento e dissolução dos elementos osteológicos, enquanto que os quadrados L29 por
se encontrarem mais afastados da escorrência, os restos osteológicos foram menos
afetados pelo processo erosivo causado pela água. Durante a crivagem do sedimento e
triagem do material foi possível verificar que o sedimento dos quadrados N29 e N30 era
sobretudo constituído por elementos arenosos, ao contrário do que acontecia no
quadrado L29, em que o sedimento era argiloso, o que revela também a constante
“lavagem” a que o sedimento dos quadrados N29 e N30 esteve sujeito.
7%
23%
25%
10%
6%
19%
4%
6%
N30/E1
L29/E1
L29/E2
N30/E3
N30/E4
L29/E5
N29/E10
L30/F2
0% 5% 10% 15% 20% 25% 30%
Gráfico 3: Percentagem de restos por quadrado e camada

73
8. Significado paleoecológico, paleoambiental e paleoclimático das espécies
identificados durante o Epipaleolítico e o Neolítico Antigo
Na totalidade das amostras analisadas a Ordem Rodentia é claramente
dominante face à Ordem Soricomorpha (ver Gráfico 4).
As Ordens Rodentia e Soricomorpha são as duas Ordens de microvertebrados
mais referenciadas no registo arqueológico português quando analisadas amostras
microfaunísticas (Cardoso et al, 1996; Storch & Uerpmann, 1976; Antunes, 1989;
Póvoas et al, 1992; Póvoas, 1998; 2001; Antunes et al, 1986; Póvoas et al, 1993;
Póvoas, 2001; Brugal et al, 2012; Morales, 1997; Jeannet, 2000; Pimenta, 2013).
O facto de terem sido identificadas sete espécies de roedores indicia a presença
na envolvente do Abrigo da Pena d'Águade um mosaico de habitats e a existência de
zonas bióticas específicas de cada uma delas. Sendo que, uma grande percentagem são
endémicas da Península Ibérica, nomeadamente Arvicola cf sapidus, Microtus cabrerae
e Microtus (Terricola) duodecimcostatus/ lusitanicus, ou autóctones da Península
Ibérica, nomeadamente Apodemus sylvaticus e Crocidura russula, destacando-se apenas
a introdução do Género Mus, alóctone da Península Ibérica e proveniente no caso do
Mus spretus do Norte de África e no caso da sua espécie simpátrica, Mus musculus, do
Médio Oriente.
A maioria espécies identificadas possui requisitos típicos de zonas de clima de
influência mediterrânea, nomeadamente Apodemus sylvaticus, Mus sp, Microtus
94%
6%
Ordem Rodentia
Ordem Soricomorpha
0% 20% 40% 60% 80% 100%
Gráfico 4: Distribuição das Ordens Soricomorpha e Rodentia

74
(Terricola) duodecimcostatus/ lusitanicus, Microtus cabrerae, Eliomys quercinus e
Crocidura russula.
Como já mencionado o coberto vegetal existente na área em que se insere o
Abrigo da Pena d'Água é próprio de zonas de clima mediterrâneo (Figueiral, 1998),
sendo que a predominância notória dos murídeos (Apodemus sylvaticus) face aos
arvicolídeos (Microtus, Terricola e Arvicola) (ver Gráfico 5) vem ao encontro desta
realidade, uma vez que é característica das regiões de clima e coberto vegetal de tipo
mediterrâneo existentes no sul da Península Ibérica, como já observado por Liliana
Póvoas no estudo preliminar que realizou (Póvoas, 1998, pp. 81).
As principais espécies identificadas foram Apodemus sylvaticus, com um
número de restos osteológicos determinados muito elevado face aos restantes, seguido
de Microtus duodecimcostatus/ lusitanicus e de Eliomys quercinus (ver Gráfico 5).
Importa referir que no trabalho citado (Póvoas, 1998) a presença do Género Microtus
foi pouco significativa, sendo que os principais Géneros identificados foram Apodemus
e Mus, tendo sido aqueles que sofreram um acentuado aumento nas camadas mais
recentes da sequência estratigráfica.
A predominância de Apodemus sylvaticus reflete a grande abundância desta
espécie autóctone em todos os tipos de habitat da Península Ibérica, constituindo uma
presa muito frequente de diferentes predadores, responsáveis pela sua acumulação em
contextos arqueológicos (Pechorroman, 1993)
Esta predominância de Apodemus sylvaticus em todas as camadas analisadas
(ver Quadro 5), permite-nos retirar algumas conclusões acerca da caracterização da área
envolvente do abrigo, nomeadamente no que diz respeito à existência de manchas de
bosque com boa cobertura arbustiva e arbórea, onde tanto Apodemus sylvaticus como
Eliomys quercinus encontram os biótopos favoráveis ao seu desenvolvimento (Jubete,
2007; Moreno, 2007). Por outro, o facto de Apodemus sylvaticus se encontrar em
dominância face ao Género Mus, sugere que a expansão da agricultura ainda se
encontrava nas primeiras camadas E numa fase muito inicial, sendo ainda pouco
expressiva a existência de campos de cultivo face ao bosque. Embora seja de referir que
Eliomys quercinus além dos bosques e das zonas de matagal com afloramentos
rochosos, como as existentes junto ao abrigo, ocupa ainda campos desflorestados e
zonas rurais, que estariam a emergir na área em torno do Abrigo da Pena d'Água a partir
do Neolítico Antigo.

75
Ainda em relação aos murídeos é de destacar a identificação de um M1 de Mus
sp, uma vez que a ocorrência deste Género encontra-se diretamente relacionada com a
introdução da agricultura na Península Ibérica e com a criação de excedentes por parte
do Homem (Bonhomme & Thaler, 1988). Tanto Mus musculus como Mus spretus são
como referido espécies alóctones da Península Ibérica e a sua introdução e expansão
estão diretamente relacionadas com o desenvolvimento da Neolitização neste território a
partir do Neolítico Antigo (Bonhomme & Thaler, 1988).
Apesar de Mus spretus não ser uma espécie comensal do Homem, como
acontece com Mus musculus, a sua introdução na Península deve-se tal como a da sua
espécie simpátrica à ação do Homem, uma vez que este sendo autóctone do Norte de
África chegou à Península Ibérica devido à expansão da navegação (Bonhomme &
Thaler, 1988). O sucesso da sua distribuição encontra-se indiretamente relacionado com
a introdução da agricultura, uma vez que a desflorestação levada a cabo pelo Homem a
partir do Neolítico Antigo para abertura de campos de cultivo e desenvolvimento da
pastorícia, tornou possível a criação dos biótopos favoráveis ao seu desenvolvimento,
45%
0,30%
0,30%
0,30%
16%
11,70%
1,40%
14,40%
6,20%
4,40%
Apodemus sylvaticus
Mus sp
Microtus cabrerae
Microtus arvalis/agrestis
Microtus duodecimocustatus/…
Microtus sp
Arvicola cf sapidus
Eliomys quercinus
Crocidura russula
Crocidura sp
0% 10% 20% 30% 40% 50%
Gráfico 5: Distribuição do NRD

76
sendo esta uma espécie incapaz de sobreviver em zonas de bosque (Bonhomme &
Thaler, 1988).
Podemos com isto concluir que a introdução do Género Mus na zona geográfica
em que se insere o Abrigo da Pena d'Água foi consequência do desenvolvimento da
paisagem agrícola que terá tido início nos primórdios do Neolítico Antigo, situação que
se encontra refletida desde as camadas mais antigas (E-a e E-b).
No que concerne ao Género Microtus, este encontra-se na totalidade das
amostras analisadas sobretudo representado pelo Sub-género Terricola.
A percentagem de Terricola embora significativa é bastante inferior à registada
para Apodemus, o que indica uma reduzida extensão das áreas com um nível freático
elevado e detentoras de solos brados.
Este Sub-género encontra-se representado pelas espécies Microtus (Terricola)
duodecimcostatus e Microtus (Terricola) lusitanicus. Dada a fragmentação da maior
parte dos dentes hipsodontes destes arvicolídeos, não foi possível efetuar a
diferenciação específica das duas espécies presentes na Península Ibérica, sendo que a
zona do abrigo encontra-se como já referido numa zona de simpatria, podendo ocorrer
ambas. A sua presença, além de refletir a existência de um clima de tipo mediterrâneo,
sugere a existência na envolvente do Abrigo da Pena d'Água de espaços abertos de
origem natural ou antrópica, no caso de espaços agrícolas, com solos brados e húmidos
fáceis de escavar, a que esta espécie se encontra associada (Cotilla & Palomo, 2007;
Mira & Mathias, 2007).
No caso do Género Microtus sobressai o registo de Microtus cabrerae, ainda que
se tenha identificado apenas um resto correspondente a esta espécie, tendo em conta o
facto das suas populações terem vindo a sofrer uma acentuada fragmentação da sua
distribuição na Península Ibérica desde o Neolítico Antigo, devido à pressão antrópica
relacionada coma desflorestação e com a introdução de práticas agro-pastoris, que
levaram em determinadas zonas ao desaparecimento do seu habitat (Cabral, 2005). Esta
espécie ocupa também exclusivamente áreas de clima mediterrâneo, detentoras de
cobertura herbácea que se mantenha verde ao longo de todo o ano, na margem de
ribeiras temporárias (Fernandez-Salvador, 2007), como a existente junto ao Abrigo da
Pena d'Água.

77
Dentro do Género Microtus destaca-se ainda a identificação de um resto
atribuído ao grupo Microtus arvalis/ agrestis. Microtus agrestis, a espécie deste grupo
que ocorre atualmente no nosso território, ocupa espaços abertos, com pouca ou
nenhuma pressão de pastoreio, em zonas húmidas ou encharcadas (Palomo & Gisbert,
2007), pouco expressivas na área em torno do abrigo.
A baixa percentagem do Género Arvicola releva a existência pouco significativa
de solos húmidos e zonas encharcadas a que este roedor semi-aquático vive associado
(Ventura, 2007), embora as margens da nascente de água existente junto ao abrigo fosse
um local propício ao seu desenvolvimento.
Por último, a identificação de insectívoros, nomeadamente do Género
Crocidura, evidencia a existência de áreas abertas e de margens de bosques ou matagais
detentoras de biótopos secos e de cobertura vegetal arbustiva, habitat onde este Género
se desenvolve (López-Fuster, 2007) e corrobora a existência de um clima de tipo
mediterrâneo que lhe é favorável.

78
CONCLUSÃO
A sequência crono-estratigráfica extensa do Abrigo da Pena d'Água revelou a
existência de ocupações humanas contínuas durante todo o Neolítico e ocupações
esporádicas desde o Epipaleolítico até à Época Moderna.
O seu estudo revelou-se de grande importância para a caracterização do processo
de Neolitização na Estremadura portuguesa e atual território português. A partir da sua
sequência sedimentar holocénica foi possível reconhecer indicadores indiretos da
ocorrência de práticas agrícolas e pastoris nesta área geográfica desde o Neolítico
Antigo, tendo por base o estudo das associações de micromamiferos, e, sobretudo a
partir da identificação do Género Mus.
A identificação destes indicadores indiretos apenas foi possível graças à
excelente metodologia de trabalho levada a cabo durante os trabalhos de escavação, que
salvaguardou a totalidade dos materiais arqueológicos e permitiu o estudo de restos de
dimensões reduzidas, como os carvões e os restos osteológicos de microfaunas, a partir
da recolha de sedimento para flutuação e craivagem a água. Sendo de salientar o
excelente trabalho de caracterização do sítio que teve por base a existência de uma
abordagem pluridisciplinar através da conciliação das várias Arqueociências,
nomeadamente da Antracologia (Figueiral, 1998), da Zooarqueologia (Valente, 1998;
Póvoas, 1998) e da Geoarqueologia (Simões, 2012).
Importa também referir que a metodologia de crivagem a água do sedimento
utilizada com recurso às malhas de crivo de 4 mm, para recuperação da fração grossa, e
da malha de crivo de 0,6 mm, para recuperação da fração fina, se mostrou eficaz, uma
vez que permitiu a recuperação da totalidade dos restos osteológicos e odontológicos
presentes nas amostras, permitindo inclusive a recuperação de dentes soltos,
nomeadamente de M3 de Apodemus sylvaticus e Pm4 de Crocidura russula e de Eliomys
quercinus.
O estudo das associações microfaunísticas revelou-se de extrema importância
para a caracterização do Abrigo da Pena d'Água porque, para além de ter permitido
reconhecer indicadores indiretos da ocorrência de práticas agrícolas como referido,
permitiu caracterizar a nível climático, ecológico e ambiental o território em torno do
abrigo, a partir da identificação de habitats e áreas bióticas específicas de cada uma das
espécies identificadas.

79
A maioria das espécies identificadas, nomeadamente Apodemus sylvaticus, Mus
sp, Microtus (Terricola) duodecimcostatus/ lusitanicus, Microtus cabrerae e Crocidura
russula, possui requisitos bióticos de influência mediterrânea, revelando a existência de
um coberto de espetro vegetal dominantemente de carácter esclerófilo. Este é
característico de bosques e matagais de ifluência mediterrânea, com boa cobertura
arbustiva e arbórea, intercaladas por zonas de espaços abertos também com boa
cobertura arbustiva, em detrimento do coberto vegetal característico de zonas húmidas.
A existência deste tipo de coberto vegetal é atestada pela predominância de Apodemus
sylvaticus a par de Eliomys quercinus, de Crocidura russula e de Microtus
duodecimcostatus/ lusitanicus, espécies que se desenvolvem em ambientes de bosque,
no caso dos dois primeiros, ou em espaços abertos com solos macios em prados abertos
nas margens de matagais e de bosques no caso dos dois últimos.
A fraca expressão de Microtus arvalis/ agrestis, é explicável pelo facto de este
se distribuir sobretudo a norte, onde ocorre um clima de influência continental e se
desenvolve um espetro vegetal característico de zonas húmidas. O mesmo ocorre com
Arvicola cf sapidus, que se desenvolve sobretudo em zonas encharcadas, que na área
em torno do Abrigo da Pena d'Água teriam uma expressão muitíssimo reduzida.
As conclusões retiradas a partir da caracterização da distribuição das espécies
identificadas vêm confrimar os resultados dos estudos de Antracologia (Figueiral, 1998)
e de Zooarqueologia (Póvoas, 1998; Valente, 1998), realizados em materiais recolhidos
neste mesmo sítio arqueológico.
Contudo, a principal conclusão retiradada caracterização dos micromamiferos
identificados relaciona-se com a intervenção humana no meio natural em torno do
Abrigo da Pena d'Água durante a Neolitização. Esta teve um impacto significativo na
paisagem natural, como consequência direta da introdução da agricultura e
desenvolvimento da pastorícia, tendo implicações ao nível do coberto vegetal,
alterando-o. As alterações causadas por estas práticas originaram variações na
distribuição de determinadas comunidades de micromamíferos, que em alguns casos se
traduziram na diminuição drástica da distribuição de algumas espécies que habitavam
esta área, como foi o caso de Microtus cabrerae, ou, na introdução involuntária de
novas espécies, como foi o caso de Mus spretus e de Mus musculus.
No caso de Microtus cabrerae a intervenção do Homem no meio natural poderá
ter provocado uma redução das populações deste roedor, uma vez que a desflorestação

80
levada a cabo a partir do Neolítico Antigo para a criação de campos de cultivo e de
pastorícia, traduziu-se em muitos casos na destruição do seu habitat e na existência de
uma distribuição atual muito fragmentada. A desflorestação levada a cabo pelo Homem
na área em torno do Abrigo da Pena d'Água é atestada pelo estudo de Antracologia, no
sentido em que não foram identificados carvões correspondentes a pinheiros em
nenhuma das camadas analisadas, o que revela o declínio dos pinhais em benefício dos
zambujais desde o Holocénico Médio, como causa direta da intensificação da
desflorestação para criação de campos de cultivo e de pastorícia a partir do Neolítico
Antigo (Figueiral, 1998).
A criação de campos de pastorícia pode ainda ter levado à diminuição de
Microtus arvalis/ agrestis, a partir do Neolítico Antigo, no sentido em que Microtus
agrestis dá preferência a habitats em que exista pouca ou nenhuma pressão de pastoreio,
que se começava agora a desenvolver na área em torno do abrigo.
Noutros casos, a desflorestação revelou-se determinante na criação de biótopos
favoráveis ao desenvolvimento de determinadas espécies, nomeadamente Mus spretus.
Este, apesar de não manter relações de comensalidade com o Homem, habita sobretudo
zonas de campos abertos e de campos de cultivo de cereais de sequeiro. Esta prática
poderá ainda ter influenciado o desenvolvimento de Microtus duodecimcostatus/
lusitanicus, tendo em conta a sua preferência por habitats onde predominem os espaços
abertos de origem natural ou antrópica.
Em relação aos agentes tafonómicos que levaram à acumulação destes restos
osteológicos no interior do Abrigo da Pena d'Água podemos concluir que o agente
tafonómico responsável pela sua acumulação é de origem animal. Esta acumulação terá
sido originada pela ação de predadores, provavelmente aves de rapina noturnas,
sobretudo bufos e corujas, que frequentaram este local como refúgio ou abrigo e, que
naturalmente aqui depositaram as suas regurgitações que continham os restos das suas
presas, o que deu origem à acumulação de grande parte do conjunto de micromamíferos
estudada.
Não obstante, alguns dos restos aqui depositados podem ainda ser resultado da
morte natural destes animais no interior do abrigo, que lhes servia de refúgio. Tanto
Apodemus sylvaticus, como Eliomys quercinus, têm por hábito frequentar o interior de
abrigos naturais e grutas para procurar abrigo ou para hibernar nos meses de Inverno,

81
especificamente no caso do Eliomys quercinus, embora este panorama para este
contexto em concreto seja pouco provável.
Quanto aos processos pós-deposicionais foram detetados agentes tafonómicos de
origem antrópica numa percentagem reduzida de restos odontológicos dos Géneros
Apodemus, Eliomys e Crocidura, nomeadamente marcas de fogo. Foram também
detetados processos de origem natural, nomeadamente agentes erosivos, como a ação da
água proveniente da nascente sazonal localizada junto ao abrigo. Esta conciliada com as
temperaturas elevadas dos meses de estio, terá causado alterações físico-químicas
substanciais nos restos osteológicos que poderam ter comprometido a sua conservação.
Há ainda a registar processos pós-deposicionais de origem biológica, relacionados com
a atividade radicular da vegetação que terá coberto a superfície daqueles depósitos.
Concluindo, a existência de agentes tafonómicos pós-deposicionais erosivos
(ação da água), bem como as próprias características do sedimento, foram responsáveis
pelo mau estado de conservação em que a amostra faunística aqui alvo de estudo se
encontra. Os fragmentos de restos osteológicos são de dimensões reduzidas, o que não
permitiu identificações ao nível anatómico nem taxonómico. Os restos odontológicos
encontravam-se também estes com um grau de fragmentação significativo, o que
dificultou nalguns casos (sobretudo nos arvicolídeos) a identificação taxonómica.
Devemos assim considerar que os processos tafonómicos pós-deposicionais podem ter
afetado a quantidade e a diversidade original de espécies existentes nas amostras.
Para finalizar, no que diz respeito à distribuição espacial e estratigráfica dos
restos osteológicos podemos concluir que existe uma maior concentração na
generalidade dos quadrados da camada E, correspondente ao Neolítico Antigo, quando
comparados com o quadrado da camada F, correspondente ao Epipaleolítico. Este facto
relaciona-se provavelmente com o efeito/ causa da menor sedimentação registada na
camada F face à camada E. Há igualmente uma acentuada concentração de material
osteológico nos quadrados L29 face aos restantes, o que poderá derivar da circunstância
de os quadrados N29 e N30 se encontrarem na proximidade da escorrência de água e de
o seu sedimento ter sido alvo de uma constante ação erosiva, que provavelmente terá
originado a dissolução dos restos osteológicos que originalmente se encontrariam nestes
quadrados.
O estudo microfaunístico do Abrigo da Pena d'Água não se esgota nesta
dissertação tendo em conta a existência de amostras de sedimento ainda por estudar e de

82
uma grande quantidade de material osteológico anteriormente analisado pela Dra.
Liliana Póvoas que poderá ser no futuro alvo de uma revisão, tendo por base a mesma
metodologia de estudo no que concerne aos métodos de identificação e quantificação
aqui utilizados.

83
REFERÊNCIAS BIBLIOGRÁFICAS
A
ANDREWS, P. (1990) – "Owls, Caves and Fossils. Predation, Preservation and
Accumulation of Small Mammal Bones in Caves", The University Chicago Press.
ANDREWS, P. (1997) – "What taphonomy can and cannot tell us", Cuadrenos de
Geologia Ibérica, nº 23, pp. 53-72.
ANTUNES, M. T.; MANEPELLA, G.; MEIN, P. & ZBYSZEWSKI, G. (1986) –
"Goldra: permier gisement karstique en Algarve, faune at industries", Ciências da
Terra, nº 8, Lisboa, Universidade Nova de Lisboa, pp. 3-42.
ANTUNES, M. T. (1989) – "Castor fiber na Gruta do Caldeirão. Existência,
distribuição e extinção do castor em Portugal", Ciências da Terra, nº 10. Lisboa,
Universidade Nova de Lisboa, pp. 23-40.
ARAÚJO, A. C.; ZILHÃO, J. (1991) – "Arqueologia do Parque Natural das Serras de
Aire e dos Candeeiros", Colecção Estudos, nº 8. Lisboa, Serviço Nacional de Parques,
Reservas e Conservação da Natureza.
B
BARBADILLO, L. J.; IGNACIO LACOMBA, J.; PÉREZ-MELLADO, V.; SANCHO,
V. & LOPÉZ-JURADO, L.F. (1999) – "Anfibios y Reptiles de La Peninsula Ibérica,
Baleares y Canarias", in Guia Ilustrada para identificar y conocer todas las especies,
Geo Planeta.
BEHERENSMEYEER, A. K. (1978) – "Taphonomic and ecologic information from
bone weathering", Paleobiology, nº 4, pp. 150-162.
BRUGAL, J.; CRISPIM, J. A.; FIGUEIREDO, S.; SERRA, A. M. & PALMQUIST, P.
(2012) – "The complex carnivore – rich assemblages from Furninha (Peniche,
Portugal)", Jornal of Taphonomy, nº 10, pp. 417-438.
BONHOMME, F. ; THALER, L. (1988) – "L’évolution de la souris", La Recherche, pp.
606-616.

84
C
CABRAL, M. J.; ALMEIDA, J.; ALMEIDA, P. R.; FERRAND de ALMEIDA N.;
OLIVEIRA, M. E.; PALMEIRIM, J. M.; QUEIROZ, A. I.; ROGADO, L. & SANTOS -
REIS eds. (2005) – Livro Vermelho dos Vertebrados de Portugal. Lisboa, Instituto de
Conservação da Natureza, pp. 660.
CARDOSO, J. L.; ANTUNES, M. T. & MEIN, P. (1996) –"Pequenos Mamíferos do
povoado de Leceia (Oeiras)", Estudos Arqueológicos de Oeiras, nº 6. Oeiras, Câmara
Municipal de Oeiras, pp. 121-133.
CARVALHO, A. F. (1998) – "O Abrigo da Pena de Água (Rexaldia, Torres Novas):
resultados dos trabalhos de 1992- 1997", Revista Portuguesa de Arqueologia, Vol. 1, nº
2, pp. 39-72.
CARVALHO, A. F. (2008) – "Cerâmica estampilhada do Abrigo da Pena d'Água
(Torres Novas): contexto, cronologia e breve enquadramento regional", BERNARDES,
J. P., org., Sic memorat. Estudos em homenagem a Teresa Júdice Gamito, Faro,
Universidade do Algarve, pp. 53-68.
CARVALHO, A. F. (2003a) – "O Neolítico antigo no Arrife da Serra d'Aire. Um case
study da neolitização da Média e Alta Estremadura", in GONÇALVES, V.S., ed. –
Muita gente, poucas antas? Origens, espaços e contextos do Megalitismo, Actas do II
Colóquio Internacional sobre Megalitismo, Trabalhos de Arqueologia nº 25, Lisboa,
Instituto Português de Arqueologia, pp.135-154.
CARVALHO, A. F. (2003b) – "A emergência do Neolítico no actual território
português: pressupostos teóricos, modelos interpretativos e evidência empírica", O
Arqueólogo Português; IV: 21, pp. 65-150.
CARVALHO, A. F. (2007) – "A Neolitização do Portugal Meridional no contexto
Mediterrâneo Ocidental do IV milénio a.C.", in CARDOSO, J.L., coord. – A
Arqueologia portuguesa e o espaço europeu balanços e perspectivas, Actas do
Colóquio da Sociedade de Geografia de Lisboa, Vol. 15, Lisboa, Centro de Estudos
Arqueológicos de Oeiras, pp. 47-77.
CARVALHO, A. F. (2008) – "A Neolitização do Portugal Meridional. Os exemplos do
Maciço Calcário Estremenho e do Algarve ocidental", Promotoria Monográfica, nº 12,
Faro, Universidade do Algarve.

85
CARVALHO, A. F. (s.d.) – "The Pena d’Água Rock-shelter (Torres Novas, Portugal):
two distinct life ways within a Neolithic sequence", in JUAN-CABANILLES, J., ed. -
Homenaje a Bernart Martí,Valencia, Museo de Prehistòria de Valencia, no prelo.
CHALINE, J. (1972) – "Les Rongeurs du Pleistocéne Moyer et Supérior de France",
Editions du Centre National de la RechercheScientifique. Paris
COTILLA, I. & PALOMO, L. J. (2007) – "Microtus duodecimcostatus (de Sélys –
Longchamps, 1839)", in PALOMO, L.J.; GISBERT, J.; BLANCO, J. C. eds..Atlas y
Libro Rojo de los Mamíferos Terrestres de España., Madrid, pp. 422-425.
F
FERNÁNDEZ-SALVADOR, R. (2007) – "Microtus cabrerae (Thomas, 1906)", in
PALOMO, L.J.; GISBERT, J. & BLANCO, J. C. eds..Atlas y Libro Rojo de los
Mamíferos Terrestres de España. Madrid, pp. 429-433.
FERRAND DE ALMEIDA, N.; FERRAND DE ALMEIDA, P.; GONÇALVES, H.;
SEQUEIRA, F.; TEIXEIRA, J.& FERRAND DE ALMEIDA, F. (2001) – GUIAS
Fapas – Anfíbios e Répteis de Portugal.
FERREIRA, M. M. N. (1998) – As cerâmicas medievais/modernas do Abrigo da Pena
d'Água (Torres Novas)", Revista Portuguesa de Arqueologia, Vol. 1, nº 2, pp. 97-106.
FIGUEIRAL, I. (1998) –"Abrigo da Pena d'Água (Torres Novas): a contribuição da
antracologia", Revista Portuguesa de Arqueologia, Vol. 1, nº2, pp. 73-80.
G
GÁLLEGO, L. C. & LÓPEZ, S. (1982) –Vertebrados Ibéricos: Mamíferos Insectívoros.
Universidad de Sevilla, Vol. 5, pp. 5-60.
GÁLLEGO, L. C. & ALAMANY, A. (1985) – Vertebrados Ibéricos, Roedores y
Lagomorfos. Universidad de Sevilla, Vol. 6, pp. 8-63.
H
HILLSON, S. (1990) – Theeth. Cambridge University Press. Cambridge.
I
IOANNIDOU, E. (2003) – "Tafonomy of animal bones: species, sex, age and breed.
Variability of sheep, cattle and pig bone density", Journal of Archaeological Science, nº
30, pp. 355-365.

86
J
JANS, M. M. E. (2003) – "Characterisation of microbial attack on archaeological bone",
Journal of Archaeological Science, nº 31, pp. 87-95.
JEANNET, M. (2000) – "Gruta da Figueira Brava: les Rongeurs", in ANTUNES, M. T.
ed.. Memória da academia das ciências de Lisboa: Colóquio: Últimos Neandertais em
Portugal: evidência, odontológica e outra,Lisboa, pp. 179-243.
J. REITZ, Elizabeth &S. WING, Elizabeth. (1999) – Zooarcheology. Manuals in
Archaeology. Cambridge.
JUBETE, F. (2007) – "Apodemus sylvaticus (Linnaeus, 1758)", in PALOMO, L.J.;
GISBERT, J. & BLANCO, J. C. eds..Atlas y Libro Rojo de los Mamíferos Terrestres de
España, Madrid, pp. 449-451.
L
LYMAN, R. L. (2001) – Vertebrate Taphonomy. Cambridge University Press.
Cambridge.
LÓPEZ-FUSTER, M. J. (2007) – "Crocidura russula (Herman, 1780) ", in PALOMO,
L.J.; GISBERT, J. & BLANCO, J. C. eds..Atlas y Libro Rojo de los Mamíferos
Terrestres de España, Madrid, pp. 128-130.
LOPEZ-GARCÍA, J. M. (2008) – "Evolución de la diversidad taxonómica de los
micromamíferos en la Península Ibérica y câmbios Paleoambientales durante el
Pleistoceno Superior", Tesis doctural, Universitat Rovira I Virgili, Tarracona.
M
MADUREIRA, M. L. & RAMALHINHO, M. G. (1981) – "Notas sobre a distribuição,
diagnose e ecologia dos Insectívora e Rodentia portugueses", Arquivos do Museu
Bocage, série A, Vol. 1, nº 10. Publicação do Museu e Laboratório Zoológico e
Antropológico. Faculdade de Ciências de Lisboa. Lisboa.
MATEUS, J. E.; QUEIROZ, P. F. & LEEUWAARDEN, W. (2002) – "O Laboratório
de Paleoecologia e Arqueobotânica – uma visita guiada aos seus programas, linhas de
trabalho e perspectivas", in MATEUS, J. E.; MORENO-GARCÍA, M. eds..
Paleoecologia Humana e Arqueociências: Um Programa Multidisciplinar para a

87
Arqueologia sob a Tutela da Cultura. IPA, Trabalhos de Arqueologia, nº 29, Lisboa, pp.
105-188.
MIRA, A. & MATHIAS, M. L. (2007) – "Microtus lusitanicus (Gerbe, 1879) ", in
PALOMO, L.J.; GISBERT, J.; BLANCO, J. C. eds..Atlas y Libro Rojo de los
Mamíferos Terrestres de España ,Madrid, pp. 418-421.
MITCHELL – JONES, A. J.; AMORI, G.; BOGDANOWICZ, W.; KRYŠTUFEK, B.;
REIJNDERS, J. P. H.; SPITZENBERGER, F.; STUBBE, M.; THISSEN, J. B. M.;
VOHRALIK, V. & ZIMA, J. (1999) – The Atlas of European Mammals. London, pp.
484.
MORALES, A. & RODRIGUEZ, J. (1997) – "Black Rats (Rattus rattus) from medieval
Mértola (Baixo Alentejo, Portugal)", The Zoological Society of London, pp. 623-642.
MORENO, S. (2007) – "Eliomys quercinus (Linnaeus, 1766)", in PALOMO, L.J.;
GISBERT, J. & BLANCO, J. C. eds..Atlas y Libro Rojo de los Mamíferos Terrestres de
España, Madrid, pp. 392-394.
MORENO-GARCÍA, M.; PIMENTA, C. M (2002) – "The Paleofaunal contexto", in
ZILHÃO, J.; TRINKAUS, E., eds. – Portrait of the artist as a child. The Gravettian
Human skeleton from the Abrigo do Lagar Velho and its Archaeological Context.
Trabalhos de Arqueologia, nº 22, Instituto Português de Arqueologia, Lisboa, pp.112-
131.
MORENO-GARCÍA, M.; DAVIS, S.; PIMENTA, C. (2003) –"Arqueozoologia: estudo
da fauna do passado", in MATEUS, J. E. & MORENO-GARCÍA, M.
eds..Paleoecologia Humana e Arqueociências: Um Programa Multidisciplinar para a
Arqueologia sob a Tutela da Cultura, Capítulo 5, Trabalhos de Arqueologia, nº 29,
Instituto Português de Arqueologia, Lisboa, pp. 191-234
MORENO-GARCÍA, M.; PIMENTA, C. M.; DAVIS, S. & GABRIEL, S. (2003) – "A
osteoteca: uma ferramenta de trabalho", in MATEUS, J. E. & MORENO-GARCÍA, M.
eds..Paleoecologia Humana e Arqueociências: Um Programa Multidisciplinar para a
Arqueologia sob a Tutela da Cultura, Capítulo 6, Trabalhos de Arqueologia, nº 29,
Instituto Português de Arqueologia, Lisboa, pp. 235-261.

88
P
PALOMO, L. J.; GISTERT, J. & BLANCO, J. C. (2007) – Libro Rojo de los los
mamíferos terrestres de Espanhã. Ministerio do Medio Ambiente. Madrid.
PECHARROMÁN, M. A. C. (1993) – "Las rapaces nocturnas como acumuladores
potenciales de restos faunísticos en yacimientos arqueológicos: los micromamíferos de
Peñalosa", Archaeofauna – International Journal of Archaeozoology, vol. 2, pp. 219-
230.
PIMENTA, C. & MORENO-GARCÍA, M. (2004) – "A Osteoteca do Instituto
Português de Arqueologia. Um património aberto à comunidade científica", Vol. 22.
Lisboa, pp. 18-19.
PIMENTA, C. (2013) – "Microvertebrados do Algar do Bom Santo (Alenquer) ",
Trabalhos do Larc, nº 4. Lisboa.
PIMENTA, C.; PANDO, A.; ALDANA, P.; MONTEIRO, M. (2015) – "Libaciones o
trampeo involuntário? Los micro-vertebrados de las tumbas T68 y T98 de la necrópolis
romana del Prado de San Sebastián (Sevilha, España)",Comunicação realizada no
encontro de Zooarqueologia: O mundo animal na romanização da Península Ibérica,
Trabalhos no Larc, nº 9. Lisboa.
PÓVOAS, L.; ZILHÃO, J.; CHALINE, J. & BRUNET-LECOMTE, P. (1992) – "La
faune de Rongeurs du Pleistocene Superieur de la Grotte de Caldeirão (Tomar,
Portugal) ", Quaternaire – International Jornal of the french Quarternary Association,
vol. 3, nº1, pp. 40-47.
PÓVOAS, L.; BRUNET-LECOMTE, P. & CHALINE, J. (1993) – "Présence de Mus
spretus fossile dans l´Holocénedu Portugal", III Reunião do Quaternário Ibérico,
Coimbra, pp. 485-489.
PÓVOAS, L. (1998) – "Faunas de micromamíferos do Abrigo da Pena d'Água (Torres
Novas) e seu significado paleoecológico: considerações preliminares", Revista
Portuguesa de Arqueologia, Vol. 1, nº 2, pp. 81-84.
PÓVOAS, L. (2001) – "Contribuição do estudo de associações de Roedores para o
conhecimento de variações climáticas durante o Quarternário em Portugal", I Congresso
do Quaternário de Países de línguas ibéricas – V Reunião do Quaternário Ibérico,
Lisboa, pp. 133-136.

89
S
SANTOS, S. M. (2009) – "Factors influencing the distribution of the Lusitanian and the
mediterrnean pine voles (Microtus lusitanicus and Microtus duodecimcostatus) in
Portugal: a multiscale approach". Tese de doutoramento em Biologia apresentada à
Faculdade de Ciências de Lisboa. Tese não publicada.
SESÉ, C. (2011) – "Micromamíferos (Erinaceomorfos y Roedores) del final del
Pleistoceno Superior y la primera parte del Holoceno de Cova Fosca (Alto Maestrazgo,
Castellón): Reconstrucción paleoambiental del entorno del yacimiento", Archaeofauna
–International Journal of Archaeozoology, vol. 20, pp. 119-137.
SEVILHA, P. G. (1986) – "Identificación de los principales quirópteros ibéricos a partir
de sus dientes aislados. Valor sistemático de los caracteres morfológicos y métricos
dentários", Acta Vertebrata, nº 13, pp. 111-130.
SIMÕES, C. D. (2012) – "Estudio geoarqueológico del Abrigo da Pena d'Água (Torres
Novas, Portugal) en el contexto de la Neolitización de la vertiente atlântica meridional
peninsular", Revista Estudios de Cuarternario, nº 2, pp. 49-69.
STINER, M. C. et al. (1995) – "Differential Burning, Recrystallization and
Fragmentation of Archaeological Bone", Journal of Archaeological Science, nº 22, pp.
223-237.
STORCH, G. & UERPMANN, H. P. (1976) – "Die Kleinsaugerknochen vom Castro do
Zambujal. In Studien uber fruhe Tierknochenfunde von der Iberischen Halbinsel",
Deutsches Archaologisches Institut, nº 5, Madrid, pp.129-138.
V
VALENTE, M.J. (1997) – "A quantificação faunística: principais unidades, alguns
parâmetros, regras e problemas", Estudos do Quaternário, nº 1, APEQ, Lisboa, pp. 86-
96.
VALENTE, M. J. (1998) –"Análise preliminar da fauna mamalógica do Abrigo da Pena
d'Água (Torres Novas): campanhas de 1992-1994", Revista Portuguesa de Arqueologia,
vol. 1, nº 2, pp. 85-96.
VALENTE, M. J. (2000) – "Arqueozoologia e Tafonomia em contexto paleolítico. A
Gruta do Pego do Dabo (Loures) ". Dissertação de mestrado apresentada à Faculdade de
Letras da Universidade de Lisboa, Lisboa.

90
VENTURA, J. (2007) – "Arvicola sapidus (Miller, 1908)", in PALOMO, L.J.;
GISBERT, J. & BLANCO, J. C. eds..Atlas y Libro Rojo de los Mamíferos Terrestres de
España, Madrid, pp. 405-409.
VIGNE, J. D. (1995) – "Determination ostéologique des principaux éléments du
squelette appendiculaire d’Arvicola, d’ Eliomys, de Glis et de Rattus", in DESSE, J. &
DESSE – BERSET, N. eds..Fiches d’Osteologie animale pour l’ Archéologie, série B:
Mammiféres, Nº 6, CNRS, Paris, pp. 3-12.

91
ANEXOS

92
ANEXO 1: TABELAS

93
Tabela 1: Registo dos M1 deMuridae
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos M1 de Muridae
Quadrado:L29 Camada:E1
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 IIIIIII IIII
Apodemus sylv. 2 IIII IIIIIII
Apodemus sylv. 3 IIII IIIIIII
Mussp 3 I
Apodemus sylv. 4 IIIIII III II
Total: 21 Total: 22 Total: 2
Total: 45
Quadrado: N30 Camada:E1 (c.Ea)
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 IIIIIII IIIIIII
Total: 7 Total: 6 Total: 0
Total: 13
Quadrado: N30 Camada:E4 (c.Eb)
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 I II
Total: 1 Total: 2 Total: 0
Total: 3
Quadrado: N30 Camada:E3 (c.Eb)
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 IIII IIIII
Total: 4 Total: 5 Total: 0
Total: 9
Quadrado: N29 Camada:E10
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 II
Total: 2 Total: 0 Total: 0
Total: 2
Quadrado:L30 Camada:F2
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 IIIIII I
Total: 7 Total: 0 Total: 1
Total: 8
Quadrado:L29 CamadaE5
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 IIII IIIII I

94
Apodemus sylv. 2 III IIIIIII
Apodemus sylv. 3 III IIIIIII
Total: 10 Total: 20 Total: 1
Total: 31
Quadrado:L29 Camada:E2
Espécie Fila M1 Direito M1Esquerdo NI
Apodemus sylv. 1 I II
Apodemus sylv. 2 IIII IIIIIII
Apodemus sylv. 3 IIIII IIIIII
Apodemus sylv. 4 IIIII IIIIII
Apodemus sylv. 5 II IIIIIIIIII
Total: 17 Total: 31 Total: 0
Total: 48
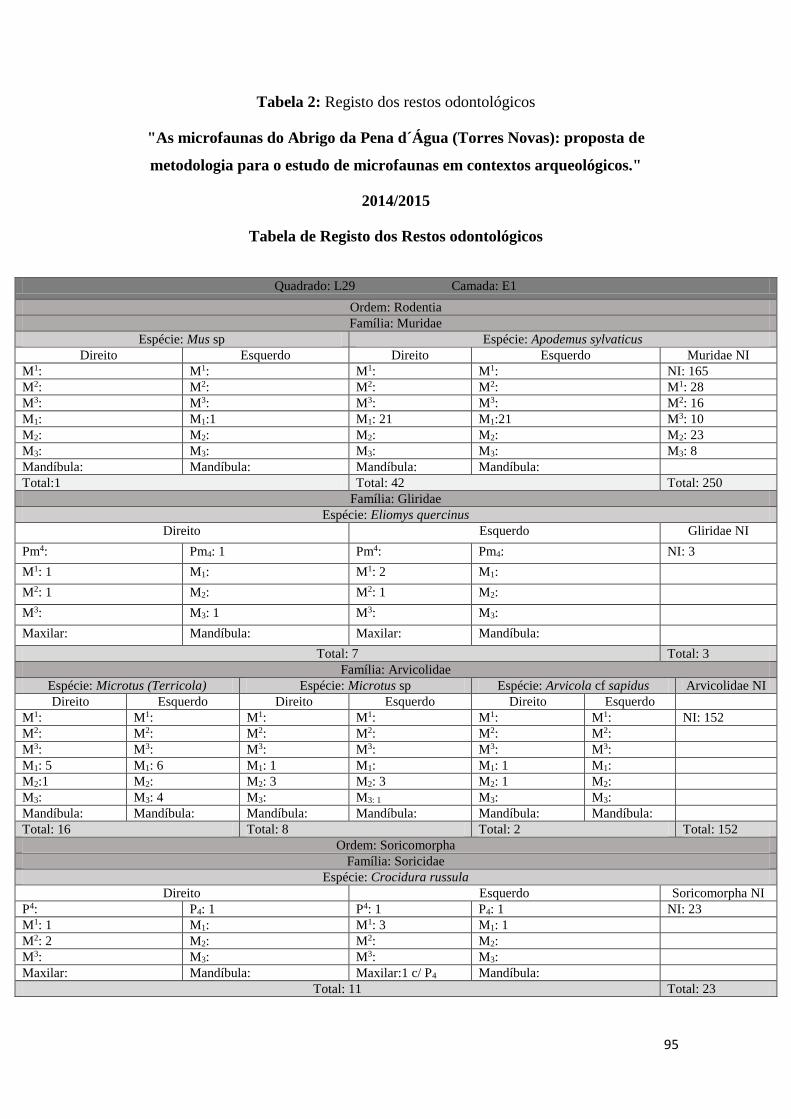
95
Tabela 2: Registo dos restos odontológicos
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológicos
Quadrado: L29 Camada: E1
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: NI: 165
M2: M2: M2: M2: M1: 28
M3: M3: M3: M3: M2: 16
M1: M1:1 M1: 21 M1:21 M3: 10
M2: M2: M2: M2: M2: 23
M3: M3: M3: M3: M3: 8
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total:1 Total: 42 Total: 250
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: Pm4: 1 Pm4: Pm4: NI: 3
M1: 1 M1: M1: 2 M1:
M2: 1 M2: M2: 1 M2:
M3: M3: 1 M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 7 Total: 3
Família: Arvicolidae
Espécie: Microtus (Terricola) Espécie: Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo
M1: M1: M1: M1: M1: M1: NI: 152
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
M1: 5 M1: 6 M1: 1 M1: M1: 1 M1:
M2:1 M2: M2: 3 M2: 3 M2: 1 M2:
M3: M3: 4 M3: M3: 1 M3: M3:
Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 16 Total: 8 Total: 2 Total: 152
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura russula
Direito Esquerdo Soricomorpha NI
P4: P4: 1 P4: 1 P4: 1 NI: 23
M1: 1 M1: M1: 3 M1: 1
M2: 2 M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar:1 c/ P4 Mandíbula:
Total: 11 Total: 23

96
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológicos
Quadrado: N30 Camada:E1 (c.Ea)
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: NI: 67
M2: M2: M2: M2: M1: 8
M3: M3: M3: M3: M2: 8
M1: M1: M1: 7 M1:6 M3: 2
M2: M2: M2: M2: M2: 14
M3: M3: M3: M3: M3:10
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total:0 Total: 13 Total: 109
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: Pm4: Pm4: Pm4: NI: 12
M1: M1: M1: 1 M1:
M2: M2: 1 M2: 1 M2: 1
M3: M3: 1 M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 5 Total: 12
Família: Arvicolidae
Espécie: Microtus (Terricola) Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo NI: 14
M1: M1: M1: M1: M1: M1:
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
M1: M1: 1 M1: M1: M1: M1:
M2: M2: M2: 1 M2: M2: M2:
M3: 1 M3: M3: M3: M3: M3:
Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 2 Total: 1 Total: 0 Total: 14
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura sp
Direito Esquerdo Soricomorpha NI
P4: P4: P4: P4:
M1: M1:1 M1: M1:
M2: M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: 1 c/ P4, M1, M2 Maxilar: Mandíbula:
Total: 2 Total: 0
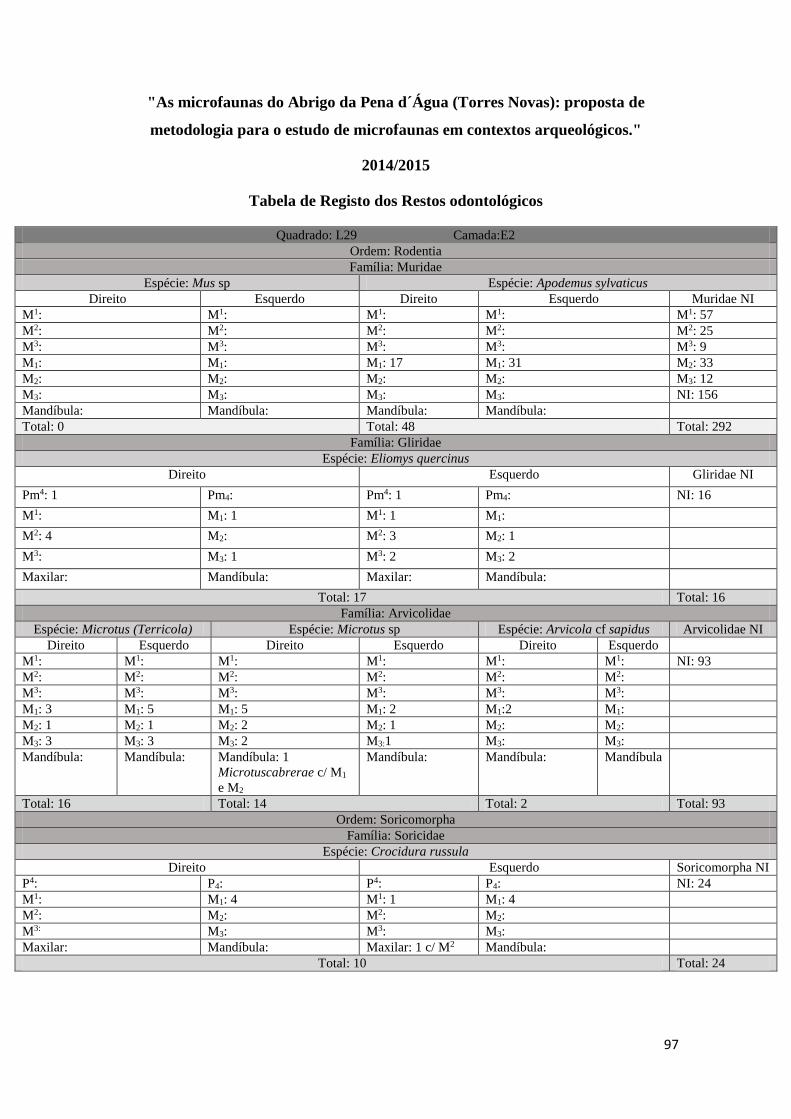
97
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológicos
Quadrado: L29 Camada:E2
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: M1: 57
M2: M2: M2: M2: M2: 25
M3: M3: M3: M3: M3: 9
M1: M1: M1: 17 M1: 31 M2: 33
M2: M2: M2: M2: M3: 12
M3: M3: M3: M3: NI: 156
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 0 Total: 48 Total: 292
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: 1 Pm4: Pm4: 1 Pm4: NI: 16
M1: M1: 1 M1: 1 M1:
M2: 4 M2: M2: 3 M2: 1
M3: M3: 1 M3: 2 M3: 2
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 17 Total: 16
Família: Arvicolidae
Espécie: Microtus (Terricola) Espécie: Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo
M1: M1: M1: M1: M1: M1: NI: 93
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
M1: 3 M1: 5 M1: 5 M1: 2 M1:2 M1:
M2: 1 M2: 1 M2: 2 M2: 1 M2: M2:
M3: 3 M3: 3 M3: 2 M3:1 M3: M3:
Mandíbula: Mandíbula: Mandíbula: 1
Microtuscabrerae c/ M1
e M2
Mandíbula: Mandíbula: Mandíbula
Total: 16 Total: 14 Total: 2 Total: 93
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura russula
Direito Esquerdo Soricomorpha NI
P4: P4: P4: P4: NI: 24
M1: M1: 4 M1: 1 M1: 4
M2: M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar: 1 c/ M2 Mandíbula:
Total: 10 Total: 24

98
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológicos
Quadrado: N30 Camada: E4 (c.Eb)
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: M1: 3
M2: M2: M2: M2: M2: 3
M3: M3: M3: M3: M3: 7
M1: M1: M1: 1 M1: 2 M2: 7
M2: M2: M2: M2: M3: 7
M3: M3: M3: M3: NI: 81
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 0 Total: 3 Total: 108
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: Pm4: Pm4: Pm4:
M1: M1: M1: 1 M1:
M2: M2: M2: 1 M2:
M3: M3: M3: M3: 1
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 3 Total: 0
Família: Arvicolidae
Espécie: Microtus (Terricola) Espécie: Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo NI: 7
M1: M1: M1: M1: M1: M1:
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
M1: M1: 1 M1: M1: M1: M1:
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 1 Total: 0 Total: 0 Total: 7
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura sp
Direito Esquerdo Soricomorpha NI
P4: P4: P4: P4: NI: 7
M1: M1: 1 M1: M1:
M2: M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 1 Total: 7
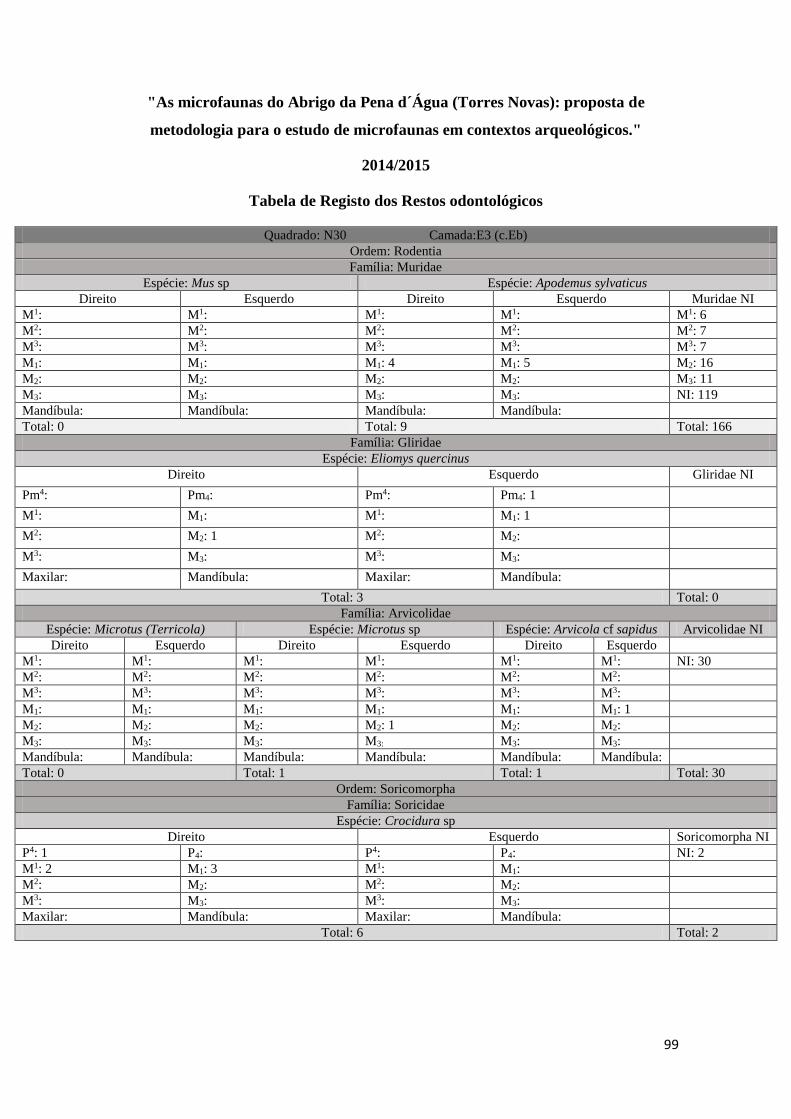
99
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológicos
Quadrado: N30 Camada:E3 (c.Eb)
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: M1: 6
M2: M2: M2: M2: M2: 7
M3: M3: M3: M3: M3: 7
M1: M1: M1: 4 M1: 5 M2: 16
M2: M2: M2: M2: M3: 11
M3: M3: M3: M3: NI: 119
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 0 Total: 9 Total: 166
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: Pm4: Pm4: Pm4: 1
M1: M1: M1: M1: 1
M2: M2: 1 M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 3 Total: 0
Família: Arvicolidae
Espécie: Microtus (Terricola) Espécie: Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo
M1: M1: M1: M1: M1: M1: NI: 30
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
M1: M1: M1: M1: M1: M1: 1
M2: M2: M2: M2: 1 M2: M2:
M3: M3: M3: M3: M3: M3:
Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 0 Total: 1 Total: 1 Total: 30
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura sp
Direito Esquerdo Soricomorpha NI
P4: 1 P4: P4: P4: NI: 2
M1: 2 M1: 3 M1: M1:
M2: M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 6 Total: 2
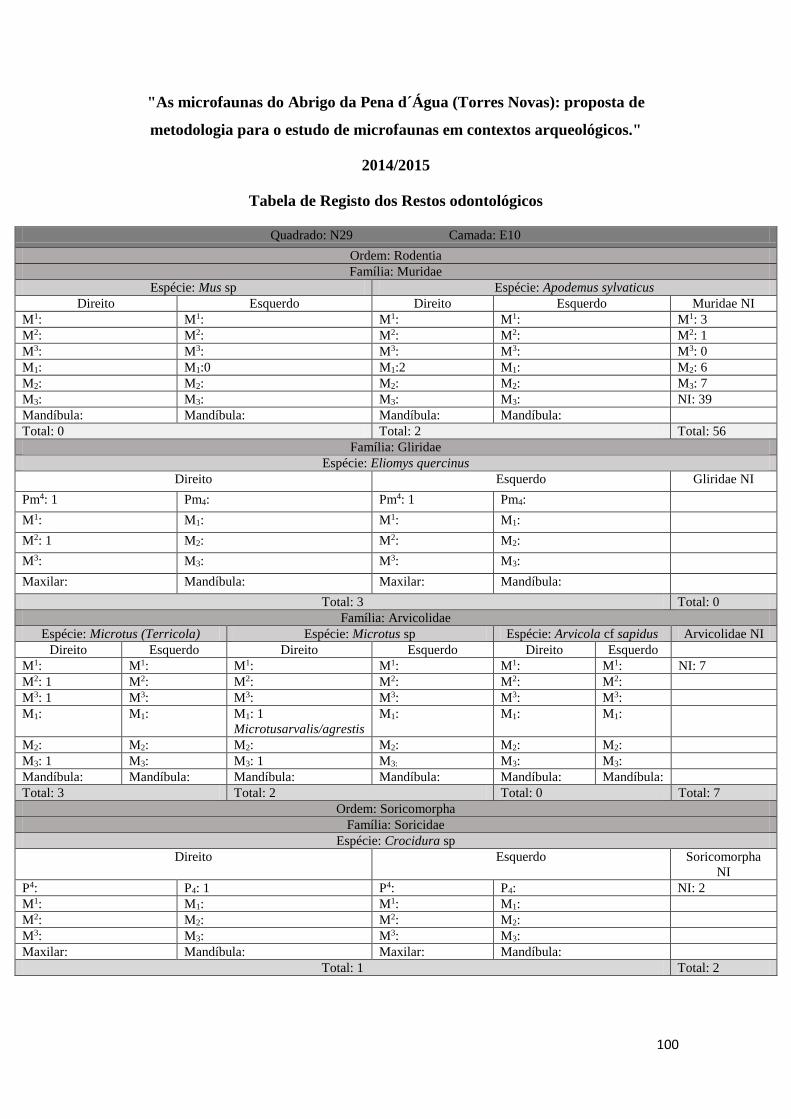
100
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológicos
Quadrado: N29 Camada: E10
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: M1: 3
M2: M2: M2: M2: M2: 1
M3: M3: M3: M3: M3: 0
M1: M1:0 M1:2 M1: M2: 6
M2: M2: M2: M2: M3: 7
M3: M3: M3: M3: NI: 39
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 0 Total: 2 Total: 56
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: 1 Pm4: Pm4: 1 Pm4:
M1: M1: M1: M1:
M2: 1 M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 3 Total: 0
Família: Arvicolidae
Espécie: Microtus (Terricola) Espécie: Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo
M1: M1: M1: M1: M1: M1: NI: 7
M2: 1 M2: M2: M2: M2: M2:
M3: 1 M3: M3: M3: M3: M3:
M1: M1: M1: 1
Microtusarvalis/agrestis
M1: M1: M1:
M2: M2: M2: M2: M2: M2:
M3: 1 M3: M3: 1 M3: M3: M3:
Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 3 Total: 2 Total: 0 Total: 7
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura sp
Direito Esquerdo Soricomorpha
NI
P4: P4: 1 P4: P4: NI: 2
M1: M1: M1: M1:
M2: M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 1 Total: 2

101
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológico
Quadrado: L30 Camada: F2
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: M1: 6
M2: M2: M2: M2: M2: 3
M3: M3: M3: M3: M3: 1
M1: M1: M1:6 M1: 1 M2: 6
M2: M2: M2: M2: M3: 1
M3: M3: M3: M3: NI: 59
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 0 Total: 7 Total: 75
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: Pm4: Pm4: Pm4: NI: 4
M1: M1: M1: M1:
M2: 1 M2: 1 M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 2 Total: 4
Família: Arvicolidae
Espécie: Microtus (Terricola) Espécie: Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo
M1: M1: M1: M1: M1: M1: NI: 44
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
M1: 1 M1: 2 M1: M1: M1: M1:
M2: M2: M2: 2 M2: 2 M2: M2:
M3: M3: 1 M3: M3: M3: M3:
Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 4 Total: 4 Total: 0 Total: 44
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura sp
Direito Esquerdo Soricomorpha NI
P4: P4: P4: P4:
M1: M1: M1: M1:
M2: M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: 1 s/ dentes Maxilar: Mandíbula:
Total: 1 Total: 0

102
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela de Registo dos Restos odontológicos
Quadrado: L29 Camada: E5
Ordem: Rodentia
Família: Muridae
Espécie: Mus sp Espécie: Apodemus sylvaticus
Direito Esquerdo Direito Esquerdo Muridae NI
M1: M1: M1: M1: M1: 29
M2: M2: M2: M2: M2: 17
M3: M3: M3: M3: M3: 4
M1: M1: M1:10 M1: 19 M2: 18
M2: M2: M2: M2: M3: 4
M3: M3: M3: M3: NI: 202
Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total:1 Total: 29 Total: 274
Família: Gliridae
Espécie: Eliomys quercinus
Direito Esquerdo Gliridae NI
Pm4: Pm4: 1 Pm4: Pm4: NI: 11
M1: M1: 2 M1: 1 M1: 1
M2: 2 M2: 1 M2: M2:
M3: M3: M3: M3: 1
Maxilar: Mandíbula: Maxilar: Mandíbula:
Total: 9 Total: 11
Família: Arvicolidae
Espécie: Microtus (Terricola) Espécie: Microtus sp Espécie: Arvicola cf sapidus Arvicolidae NI
Direito Esquerdo Direito Esquerdo Direito Esquerdo
M1: M1: M1: M1: M1: M1: NI: 53
M2: M2: M2: M2: M2: M2:
M3: M3: M3: M3: M3: M3:
M1: 2 M1: 1 M1: 3 M1: 1 M1: M1:
M2: 1 M2: 2 M2: 3 M2: 2 M2: M2:
M3: 4 M3: 2 M3: 1 M3:1 M3: M3:
Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula: Mandíbula:
Total: 12 Total: 11 Total: 0 Total: 53
Ordem: Soricomorpha
Família: Soricidae
Espécie: Crocidura sp
Direito Esquerdo Soricomorpha NI
P4: P4: P4: P4: NI: 12
M1: 1 M1: 1 M1: M1:
M2: M2: M2: M2:
M3: M3: M3: M3:
Maxilar: Mandíbula: 2 s/ dentes Maxilar: Mandíbula:
Total: 4 Total:12

103
Tabela 3: Tabela Geral de registos
"As microfaunas do Abrigo da Pena d´Água (Torres Novas): proposta de
metodologia para o estudo de microfaunas em contextos arqueológicos."
2014/2015
Tabela Geral de Registo
Folha Quad Cam. Data Classe Ordem Fam. Taxon Resto Lado Quant. Observações
1 L29 E1 23/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 21 A maioria
completos
1 L29 E1 23/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 E 21 A maioria
completos
1 L29 E1 23/09/93 Mam. Rod. Muridea Mus sp
M1 E 1 Completo
1 L29 E1 23/09/93 Mam. Rod. Muridea NI NI NI 2 Muito
fragmentados
1 N30 E1
(c.Ea)
23/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 7 A maioria
completos
1 N30 E1
(c.Ea)
23/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 E 6 A maioria
completos
1 N30 E4
(c.Eb)
07/10/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 1 Completo
1 N30 E4
(c.Eb)
07/10/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 E 2 Completos
1 N30 E3
(c.Eb)
07/10/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 4 A maioria
completos
1 N30 E3
(c.Eb)
07/10/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 E 5 A maioria
completos
1 N29 E10 07/10/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 2 Completos
1 L30 F2 16/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 6 A maioria
completos
2 L30 F2 16/09/93 Mam. Rod. Muridea NI NI NI 1 Muito
fragmentados
2 L29 E5 06/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 10 A maioria
completos
2 L29 E5 06/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 E 19 A maioria
completos
2
L29 E5 06/09/93 Mam. Rod. Muridea NI NI NI 1 Completo
2
L29 E2 24/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 D 17 A maioria
completos
2
L29 E2 24/09/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 E 31 A maioria
completos
2
N29 E10 07/10/93 Mam. Rod. Muridea Apodemus
sylvaticus
M1 NI 3
2
N29 E10 07/10/93 Mam. Rod. Muridea NI M2 NI 1
2
N29 E10 07/10/93 Mam. Rod. Muridea NI M2 NI 6
2
N29 E10 07/10/93 Mam. Rod. Muridea NI M3 NI 7
2
N29 E10 07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
Pm4 D 1
2 N29 E10 07/10/93 Mam. Rod. Gliridae Eliomys Pm4 E 1
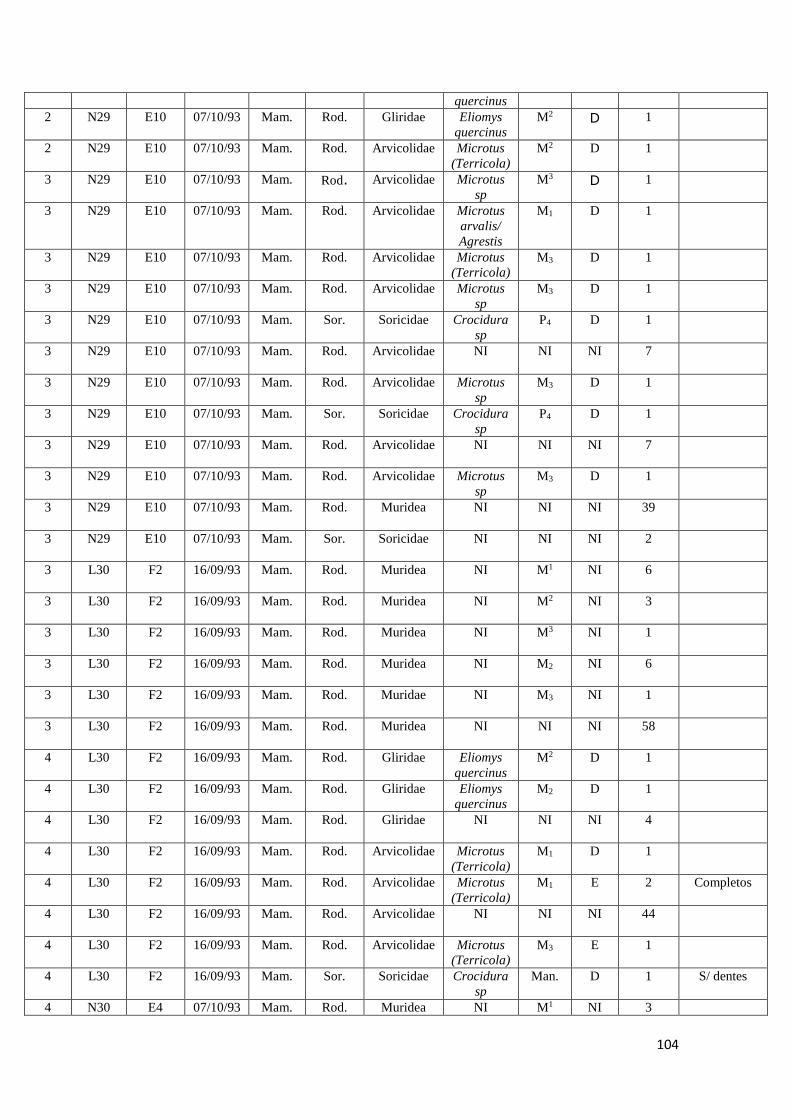
104
quercinus
2
N29 E10 07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 D 1
2
N29 E10 07/10/93 Mam. Rod. Arvicolidae Microtus (Terricola)
M2 D 1
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
Microtus
sp
M3 D 1
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
Microtus
arvalis/
Agrestis
M1 D 1
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
Microtus (Terricola)
M3 D 1
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
Microtus
sp
M3 D 1
3 N29 E10 07/10/93 Mam. Sor. Soricidae Crocidura
sp
P4 D 1
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
NI NI NI 7
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
Microtus
sp
M3 D 1
3 N29 E10 07/10/93 Mam. Sor. Soricidae Crocidura
sp
P4 D 1
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
NI NI NI 7
3 N29 E10 07/10/93 Mam. Rod. Arvicolidae
Microtus
sp
M3 D 1
3 N29 E10 07/10/93 Mam. Rod. Muridea
NI NI NI 39
3 N29 E10 07/10/93 Mam. Sor. Soricidae NI
NI NI 2
3 L30 F2 16/09/93 Mam. Rod. Muridea NI M1 NI 6
3 L30 F2 16/09/93 Mam. Rod. Muridea NI
M2 NI 3
3 L30 F2 16/09/93 Mam. Rod. Muridea NI
M3 NI 1
3 L30 F2 16/09/93 Mam. Rod. Muridea NI
M2 NI 6
3 L30 F2 16/09/93 Mam. Rod. Muridae NI
M3 NI 1
3 L30 F2 16/09/93 Mam. Rod. Muridea NI
NI NI 58
4 L30 F2 16/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 D 1
4 L30 F2 16/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 D 1
4 L30 F2 16/09/93 Mam. Rod. Gliridae NI
NI NI 4
4 L30 F2 16/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 D 1
4 L30 F2 16/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 E 2 Completos
4 L30 F2 16/09/93 Mam. Rod. Arvicolidae NI
NI NI 44
4 L30 F2 16/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M3 E 1
4 L30 F2 16/09/93 Mam. Sor. Soricidae Crocidura
sp
Man. D 1 S/ dentes
4 N30 E4 07/10/93 Mam. Rod. Muridea NI M1 NI 3
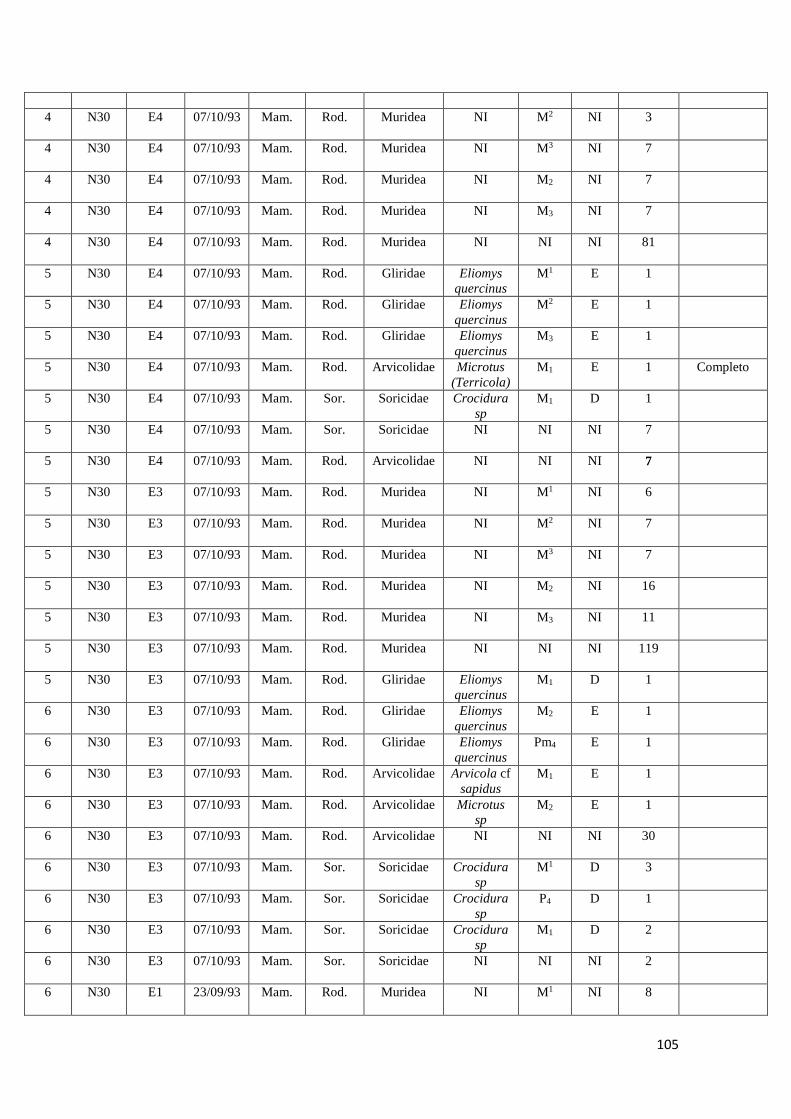
105
4 N30 E4 07/10/93 Mam. Rod. Muridea NI
M2 NI 3
4 N30 E4 07/10/93 Mam. Rod. Muridea NI
M3 NI 7
4 N30 E4 07/10/93 Mam. Rod. Muridea NI
M2 NI 7
4 N30 E4 07/10/93 Mam. Rod. Muridea NI
M3 NI 7
4 N30 E4 07/10/93 Mam. Rod. Muridea NI
NI NI 81
5 N30 E4 07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 E 1
5 N30 E4 07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 E 1
5 N30 E4 07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
M3 E 1
5 N30 E4 07/10/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 E 1 Completo
5 N30 E4 07/10/93 Mam. Sor. Soricidae Crocidura
sp
M1 D 1
5 N30 E4 07/10/93 Mam. Sor. Soricidae NI NI NI
7
5 N30 E4 07/10/93 Mam. Rod. Arvicolidae
NI NI NI 7
5 N30 E3 07/10/93 Mam. Rod. Muridea NI
M1 NI 6
5 N30 E3 07/10/93 Mam. Rod. Muridea NI
M2 NI 7
5 N30 E3 07/10/93 Mam. Rod. Muridea NI
M3 NI 7
5 N30 E3 07/10/93 Mam. Rod. Muridea NI M2 NI 16
5 N30 E3 07/10/93 Mam. Rod. Muridea NI
M3 NI 11
5 N30 E3 07/10/93 Mam. Rod. Muridea NI
NI NI 119
5 N30 E3
07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 D 1
6 N30 E3
07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 E 1
6 N30 E3
07/10/93 Mam. Rod. Gliridae Eliomys
quercinus
Pm4 E 1
6 N30 E3
07/10/93 Mam. Rod. Arvicolidae Arvicola cf
sapidus
M1 E 1
6 N30 E3
07/10/93 Mam. Rod. Arvicolidae Microtus
sp
M2 E 1
6 N30 E3
07/10/93 Mam. Rod. Arvicolidae NI NI NI 30
6 N30 E3
07/10/93 Mam. Sor. Soricidae Crocidura
sp
M1 D 3
6 N30 E3
07/10/93 Mam. Sor. Soricidae Crocidura
sp
P4 D 1
6 N30 E3
07/10/93 Mam. Sor. Soricidae Crocidura
sp
M1 D 2
6 N30 E3
07/10/93 Mam. Sor. Soricidae NI NI NI 2
6 N30 E1
23/09/93 Mam. Rod. Muridea NI M1 NI 8
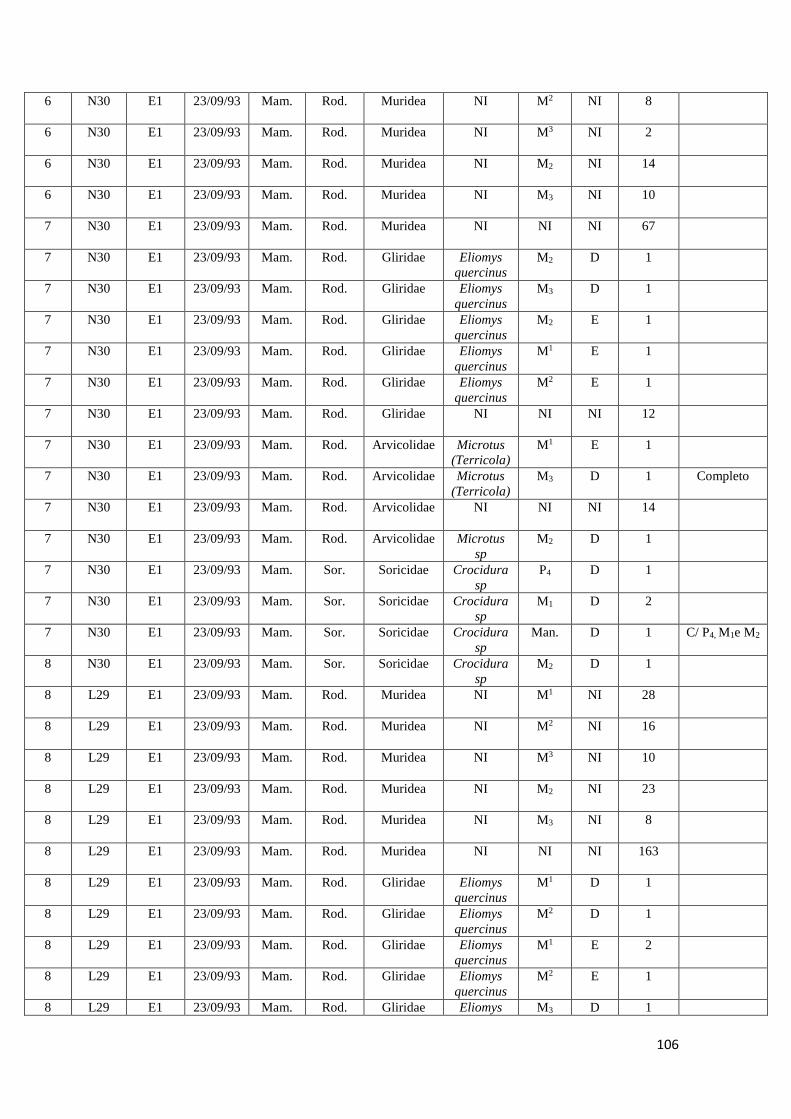
106
6 N30 E1 23/09/93 Mam. Rod. Muridea NI
M2 NI 8
6 N30 E1 23/09/93 Mam. Rod. Muridea NI
M3 NI 2
6 N30 E1 23/09/93 Mam. Rod. Muridea NI
M2 NI 14
6
N30 E1 23/09/93 Mam. Rod. Muridea NI M3 NI 10
7
N30 E1 23/09/93 Mam. Rod. Muridea NI NI NI 67
7
N30 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 D 1
7
N30 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M3 D 1
7
N30 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 E 1
7
N30 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 E 1
7
N30 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 E 1
7
N30 E1 23/09/93 Mam. Rod. Gliridae NI NI NI 12
7
N30 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 E 1
7
N30 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M3 D 1 Completo
7 N30 E1 23/09/93 Mam. Rod. Arvicolidae NI
NI NI 14
7 N30 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
sp
M2 D 1
7 N30 E1 23/09/93 Mam. Sor. Soricidae Crocidura
sp
P4 D 1
7 N30 E1 23/09/93 Mam. Sor. Soricidae Crocidura
sp
M1 D 2
7
N30 E1 23/09/93 Mam. Sor. Soricidae Crocidura
sp
Man. D 1 C/ P4, M1e M2
8 N30 E1 23/09/93 Mam. Sor. Soricidae Crocidura
sp
M2 D 1
8 L29 E1 23/09/93 Mam. Rod. Muridea NI
M1 NI 28
8 L29 E1 23/09/93 Mam. Rod. Muridea NI
M2 NI 16
8 L29 E1 23/09/93 Mam. Rod. Muridea NI
M3 NI 10
8 L29 E1 23/09/93 Mam. Rod. Muridea NI
M2 NI 23
8 L29 E1 23/09/93 Mam. Rod. Muridea NI
M3 NI 8
8 L29 E1 23/09/93 Mam. Rod. Muridea NI
NI NI 163
8 L29 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 D 1
8 L29 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 D 1
8 L29 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 E 2
8 L29 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 E 1
8 L29 E1 23/09/93 Mam. Rod. Gliridae Eliomys M3 D 1

107
quercinus
8 L29 E1 23/09/93 Mam. Rod. Gliridae Eliomys
quercinus
Pm4 D 1
8 L29 E1 23/09/93 Mam. Rod. Gliridae NI
NI NI 3
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 E 6
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M3 E 4
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M2 D 1
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 D 5
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
sp
M2 D 3
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
sp
M3 D 1
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
sp
M1 E 1
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Microtus
sp
M2 E 3
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Arvicola cf
sapidus
M1 D 1
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae Arvicola cf
sapidus
M2 D 1
9 L29 E1 23/09/93 Mam. Rod. Arvicolidae
NI NI NI 93
9
L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
M1 D 1
9 L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
M2 D 2
9 L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
P4 E 1
10 L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
M1 E 3
10 L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
P4 D 1
10 L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
P4 E 1
10 L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
M1 E 1
10 L29 E1 23/09/93 Mam. Sor. Soricidae Crocidura
russula
Max. E 1 C/ P4
10 L29 E1 23/09/93 Mam. Sor. Soricidae NI NI NI
23
10 L29 E2 24/09/93 Mam. Rod. Muridea NI
M1 NI 57
10 L29 E2 24/09/93 Mam. Rod. Muridea NI
M2 NI 25
10 L29 E2 24/09/93 Mam. Rod. Muridea NI
M3 NI 9
10 L29 E2 24/09/93 Mam. Rod. Muridea NI
M2 NI 33
10 L29 E2 24/09/93 Mam. Rod. Muridea NI
M3 NI 12
10 L29 E2 24/09/93 Mam. Rod. Muridea NI
NI NI 156
10 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
Pm4 D 1

108
10 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
Pm4 D 1
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 D 4
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 D 1
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M3 D 1
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
Pm4 E 1
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 E 1
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 E 3
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M3 E 2
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 E 1
11 L29 E2 24/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M3 E 2
11 L29 E2 24/09/93 Mam. Rod. Gliridae NI
NI NI 16
11 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
cabrerae
Man. D 1 C/ M1 e M2
11 L29 E2 24/09/93 Mam. Rod. Arvicolidae Arvicolacf
sapidus
M1 D 2
11
L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 E 5
11 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M2 E 1
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M3 E 3
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 D 3
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M2 D 1
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M3 D 3
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
sp
M1 E 2
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
sp
M2 E 1
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
sp
M3 E 1
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
sp
M1 D 5
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
sp
M2 D 2
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae Microtus
sp
M3 D 2
12 L29 E2 24/09/93 Mam. Rod. Arvicolidae
NI NI NI 93
12 L29 E2 24/09/93 Mam. Sor. Soricidae NI
NI NI 24
12
L29 E2 24/09/93 Mam. Sor. Soricidae Crocidura
russula
M1 E 1
12 L29 E2 24/09/93 Mam. Sor. Soricidae Crocidura
russula
M1 D 4
13 L29 E2 24/09/93 Mam. Sor. Soricidae Crocidura M1 D 4

109
russula
13 L29 E2 24/09/93 Mam. Sor. Soricidae Crocidura
russula
Max. E 1 C/ M2
13 L29 E5 06/09/93 Mam. Rod. Muridea NI
M1 NI 29
13 L29 E5 06/09/93 Mam. Rod. Muridea NI
M2 NI 17
13 L29 E5 06/09/93 Mam. Rod. Muridea NI
M3 NI 4
13 L29 E5 06/09/93 Mam. Rod. Muridea NI M2 NI 18
13 L29 E5 06/09/93 Mam. Rod. Muridea NI
M3 NI 4
13 L29 E5 06/09/93 Mam. Rod. Muridea NI
NI NI 201
13 L29 E5 06/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M3 D 2
13 L29 E5 06/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 D 2
13 L29 E5 06/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M2 D 1
13 L29 E5 06/09/93 Mam. Rod. Gliridae Eliomys
quercinus
Pm4 D 1
13
L29 E5 06/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 D 1
13 L29 E5 06/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M1 D 1
14 L29 E5 06/09/93 Mam. Rod. Gliridae Eliomys
quercinus
M3 D 1
14 L29 E5 06/09/93 Mam. Rod. Gliridae NI
NI NI 11
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
sp
M1 D 3
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
sp
M2 D 3
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
sp
M3 D 1
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
sp
M1 E 1
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
sp
M2 E 2
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
sp
M3 E 1
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola)
M1 D 2
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola) M2 D 1
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola) M3 D 4
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola) M1 E 1
14
L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola) M2
E 2
14 L29 E5 06/09/93 Mam. Rod. Arvicolidae Microtus
(Terricola) M3 E 2
15 L29 E5 06/09/93 Mam. Sor. Soricidae Crocidura
sp
M1 D 1
15 L29 E5 06/09/93 Mam. Sor. Soricidae Crocidura
sp
M1 D 1
15 L29 E5 06/09/93 Mam. Sor. Soricidae Crocidura Man. D 2 S/ dentes

110
sp
15 L29 E5 06/09/93 Mam. Sor. Soricidae NI NI NI
12


















