
Disruption of Kif3a in osteoblasts results in defective...
13
Disruption of Kif3a in osteoblasts results in defective bone formation and osteopenia Ni Qiu 1 , Zhousheng Xiao 1, *, Li Cao 1 , Meagan M. Buechel 2 , Valentin David 1 , Esra Roan 2 and L. Darryl Quarles 1, * 1 Department of Medicine, the University of Tennessee Health Science Center, Memphis, TN, 38165, USA 2 Department of Mechanical Engineering, the University of Memphis, Memphis, TN 38152 USA *Authors for correspondence ([email protected]; [email protected]) Accepted 17 December 2011 Journal of Cell Science 125, 1945–1957 ß 2012. Published by The Company of Biologists Ltd doi: 10.1242/jcs.095893 Summary We investigated whether Kif3a in osteoblasts has a direct role in regulating postnatal bone formation. We conditionally deleted Kif3a in osteoblasts by crossing osteocalcin (Oc; also known as Bglap)–Cre with Kif3a flox/null mice. Conditional Kif3a-null mice (Kif3a Oc-cKO ) had a 75% reduction in Kif3a transcripts in bone and osteoblasts. Conditional deletion of Kif3a resulted in the reduction of primary cilia number by 51% and length by 27% in osteoblasts. Kif3a Oc-cKO mice developed osteopenia by 6 weeks of age unlike Kif3a flox/+ control mice, as evidenced by reductions in femoral bone mineral density (22%), trabecular bone volume (42%) and cortical thickness (17%). By contrast, Oc-Cre;Kif3a flox/+ and Kif3a flox/null heterozygous mice exhibited no skeletal abnormalities. Loss of bone mass in Kif3a Oc-cKO mice was associated with impaired osteoblast function in vivo, as reflected by a 54% reduction in mineral apposition rate and decreased expression of Runx2, osterix (also known as Sp7 transcription factor 7; Sp7), osteocalcin and Dmp1 compared with controls. Immortalized osteoblasts from Kif3a Oc-cKO mice exhibited increased cell proliferation, impaired osteoblastic differentiation, and enhanced adipogenesis in vitro. Osteoblasts derived from Kif3a Oc-cKO mice also had lower basal cytosolic calcium levels and impaired intracellular calcium responses to fluid flow shear stress. Sonic hedgehog-mediated Gli2 expression and Wnt3a-mediated b-catenin and Axin2 expression were also attenuated in Kif3a Oc-cKO bone and osteoblast cultures. These data indicate that selective deletion of Kif3a in osteoblasts disrupts primary cilia formation and/or function and impairs osteoblast-mediated bone formation through multiple pathways including intracellular calcium, hedgehog and Wnt signaling. Key words: Primary cilia, Conditional deletion, Kif3a, Bone formation, Osteoblast functions, Signaling Introduction Primary cilia are present on most mammalian cells, including osteoblasts and osteocytes (Berbari et al., 2009; Davis et al., 2006; Xiao et al., 2006), but their function is not known. Primary cilia are dynamic organelles in which the kinesin-2 motor mediates the anterograde transport of intraflagellar transport (IFT) rafts to the distal tip of the cilium and the dynein motor facilitates the retrograde transport of IFT cargos to the basal body of the cilium (Davis et al., 2006; Gerdes et al., 2009). Kinesin-2 forms a heterotrimeric complex of two motor subunits, kinesin-2 family protein A (Kif3a) and B (Kif3b) and a nonmotor subunit, kinesin-associated protein (Kap3). Primary cilia house many signaling pathways that might be involved in osteoblast development and postnatal osteoblast functions (Christensen et al., 2007; Gerdes et al., 2009; Han et al., 2008; Huangfu and Anderson, 2005; Kovacs et al., 2008; Serra, 2008; Veland et al., 2009; Wong et al., 2009), including polycystin-1 (Pkd1) and 2 (Pkd2), which form a mechanosensing signaling complex that localizes to primary cilia. The C-terminal region of Kif3a has recently been shown to bind to the C-terminus of PC2 (also known as Pkd2), resulting in its localization and function in primary cilia (Li et al., 2006). Other studies have reported that Kif3b, the other motor subunit of kinesin-2, serves as a linker between PC2 and fibrocystin (also known as polyductin), the gene product of PKHD1, the gene responsible for autosomal recessive polycystic kidney disease (ARPKD) (Wu et al., 2006). Polycystins and primary cilia have interdependent functions in renal epithelial cells, and mutations in either of the polycystin genes, as well as in other genes required for ciliogenesis, lead to a common PKD phenotype. We have recently identified an important role of Pkd1 in osteoblastic development and mechanosensing in responses to skeletal loading (Xiao et al., 2011; Xiao, Z. et al., 2010; Xiao et al., 2008; Xiao et al., 2006). This suggests that primary cilia also have a role in regulating osteoblast development and function. There is additional evidence supporting the role of primary cilia in bone development. Global deletion and/or mutations of IFT88 (also known as Tg737 and polaris) or Kif3a, which blocks cilia formation (Goetz and Anderson, 2010; Pedersen et al., 2008; Serra, 2008) are associated with abnormalities of the skeleton. Disruption of gene products involved in cilium assembly and IFT also results in abnormalities of skeletal development (Serra, 2008), including left–right asymmetry (Marszalek et al., 1999), limb patterning (Haycraft and Serra, 2008; Zhang et al., 2003), endochondral bone formation (Haycraft et al., 2007), postnatal growth plate (McGlashan et al., 2007; Song et al., 2007) and craniofacial development (Kolpakova-Hart et al., 2007; Zhang et al., 2003). However, global deletion of Ift88 or Kif3a in mice, which disrupts ciliogenesis, is embryonic lethal, making it difficult to determine the role of primary cilia in postnatal osteoblast function (Marszalek et al., 1999; Murcia et al., 2000). Research Article 1945 Journal of Cell Science
Transcript of Disruption of Kif3a in osteoblasts results in defective...

Disruption of Kif3a in osteoblasts results in defectivebone formation and osteopenia
Ni Qiu1, Zhousheng Xiao1,*, Li Cao1, Meagan M. Buechel2, Valentin David1, Esra Roan2 and L. Darryl Quarles1,*1Department of Medicine, the University of Tennessee Health Science Center, Memphis, TN, 38165, USA2Department of Mechanical Engineering, the University of Memphis, Memphis, TN 38152 USA
*Authors for correspondence ([email protected]; [email protected])
Accepted 17 December 2011Journal of Cell Science 125, 1945–1957� 2012. Published by The Company of Biologists Ltddoi: 10.1242/jcs.095893
SummaryWe investigated whether Kif3a in osteoblasts has a direct role in regulating postnatal bone formation. We conditionally deleted Kif3a in
osteoblasts by crossing osteocalcin (Oc; also known as Bglap)–Cre with Kif3aflox/null mice. Conditional Kif3a-null mice (Kif3aOc-cKO)had a 75% reduction in Kif3a transcripts in bone and osteoblasts. Conditional deletion of Kif3a resulted in the reduction of primary cilianumber by 51% and length by 27% in osteoblasts. Kif3aOc-cKO mice developed osteopenia by 6 weeks of age unlike Kif3aflox/+ control
mice, as evidenced by reductions in femoral bone mineral density (22%), trabecular bone volume (42%) and cortical thickness (17%).By contrast, Oc-Cre;Kif3aflox/+ and Kif3aflox/null heterozygous mice exhibited no skeletal abnormalities. Loss of bone mass inKif3aOc-cKO mice was associated with impaired osteoblast function in vivo, as reflected by a 54% reduction in mineral apposition rateand decreased expression of Runx2, osterix (also known as Sp7 transcription factor 7; Sp7), osteocalcin and Dmp1 compared with
controls. Immortalized osteoblasts from Kif3aOc-cKO mice exhibited increased cell proliferation, impaired osteoblastic differentiation,and enhanced adipogenesis in vitro. Osteoblasts derived from Kif3aOc-cKO mice also had lower basal cytosolic calcium levels andimpaired intracellular calcium responses to fluid flow shear stress. Sonic hedgehog-mediated Gli2 expression and Wnt3a-mediated
b-catenin and Axin2 expression were also attenuated in Kif3aOc-cKO bone and osteoblast cultures. These data indicate that selectivedeletion of Kif3a in osteoblasts disrupts primary cilia formation and/or function and impairs osteoblast-mediated bone formation throughmultiple pathways including intracellular calcium, hedgehog and Wnt signaling.
Key words: Primary cilia, Conditional deletion, Kif3a, Bone formation, Osteoblast functions, Signaling
IntroductionPrimary cilia are present on most mammalian cells, including
osteoblasts and osteocytes (Berbari et al., 2009; Davis et al.,
2006; Xiao et al., 2006), but their function is not known. Primary
cilia are dynamic organelles in which the kinesin-2 motor
mediates the anterograde transport of intraflagellar transport
(IFT) rafts to the distal tip of the cilium and the dynein motor
facilitates the retrograde transport of IFT cargos to the basal body
of the cilium (Davis et al., 2006; Gerdes et al., 2009). Kinesin-2
forms a heterotrimeric complex of two motor subunits, kinesin-2
family protein A (Kif3a) and B (Kif3b) and a nonmotor subunit,
kinesin-associated protein (Kap3).
Primary cilia house many signaling pathways that might be
involved in osteoblast development and postnatal osteoblast
functions (Christensen et al., 2007; Gerdes et al., 2009; Han
et al., 2008; Huangfu and Anderson, 2005; Kovacs et al., 2008;
Serra, 2008; Veland et al., 2009; Wong et al., 2009), including
polycystin-1 (Pkd1) and 2 (Pkd2), which form a mechanosensing
signaling complex that localizes to primary cilia. The C-terminal
region of Kif3a has recently been shown to bind to the C-terminus
of PC2 (also known as Pkd2), resulting in its localization and
function in primary cilia (Li et al., 2006). Other studies have
reported that Kif3b, the other motor subunit of kinesin-2, serves as
a linker between PC2 and fibrocystin (also known as polyductin),
the gene product of PKHD1, the gene responsible for autosomal
recessive polycystic kidney disease (ARPKD) (Wu et al., 2006).
Polycystins and primary cilia have interdependent functions in
renal epithelial cells, and mutations in either of the polycystin
genes, as well as in other genes required for ciliogenesis, lead to
a common PKD phenotype. We have recently identified an
important role of Pkd1 in osteoblastic development and
mechanosensing in responses to skeletal loading (Xiao et al.,
2011; Xiao, Z. et al., 2010; Xiao et al., 2008; Xiao et al., 2006).
This suggests that primary cilia also have a role in regulating
osteoblast development and function.
There is additional evidence supporting the role of primary
cilia in bone development. Global deletion and/or mutations of
IFT88 (also known as Tg737 and polaris) or Kif3a, which blocks
cilia formation (Goetz and Anderson, 2010; Pedersen et al.,
2008; Serra, 2008) are associated with abnormalities of the
skeleton. Disruption of gene products involved in cilium
assembly and IFT also results in abnormalities of skeletal
development (Serra, 2008), including left–right asymmetry
(Marszalek et al., 1999), limb patterning (Haycraft and Serra,
2008; Zhang et al., 2003), endochondral bone formation
(Haycraft et al., 2007), postnatal growth plate (McGlashan
et al., 2007; Song et al., 2007) and craniofacial development
(Kolpakova-Hart et al., 2007; Zhang et al., 2003). However,
global deletion of Ift88 or Kif3a in mice, which disrupts
ciliogenesis, is embryonic lethal, making it difficult to determine
the role of primary cilia in postnatal osteoblast function
(Marszalek et al., 1999; Murcia et al., 2000).
Research Article 1945
Journ
alof
Cell
Scie
nce

There is strong evidence that primary cilia regulate embryonic
endochondral bone formation through hedgehog (Hh) signalingduring both embryonic and postnatal endochondral boneformation. Mice with conditional deletion of Ift88 or Kif3a
resulting from the use of both Prx1–Cre and Dermo1–Cre, thatare expressed in limb mesenchyme (including chondrocytes andperichondrium) have shorter bones in the limbs because ofalterations in embryonic endochondral bone formation, similar to
that seen in mice with germline mutations in indian hedgehog(Ihh) (Haycraft et al., 2007; Kolpakova-Hart et al., 2007; St-Jacques et al., 1999). By contrast, mice with conditional deletion
of Ift88 or Kif3a from the use of Col2a–Cre, which is expressedin the chondrocyte lineage, show no skeletal abnormalities duringthe embryonic stages, but develop postnatal dwarfism because of
a progressive loss of the growth plate between postnatal day (P) 7and P15 (Haycraft et al., 2007; Koyama et al., 2007; Song et al.,2007), similar to mice with conditional deletion of Ihh induced in
postnatal cartilage (Col2a-CreER;Ihhflox/flox) (Maeda et al.,2007). So far, the focus on mice with mutations in IFT- andcilia-related genes has been on embryonic skeletal developmentand chondrocyte function; the role of primary cilia in osteoblast
function and postnatal bone homeostasis remains to be studied.
In the study reported here we used genetic approaches toconditionally delete Kif3a (disruption of ciliogenesis) in
osteoblasts of mice to gain insights into whether IFT andprimary cilia have a direct function in regulating postnatal boneformation. We demonstrated that conditional deletion of Kif3a
from osteoblasts of heterozogyous Kif3anull/flox mutant mice
resulted in defective osteoblast function in vivo and in vitro andosteopenia, a condition in which bone mineral density is lowerthan normal. In addition, Kif3a-deficient osteoblasts exhibited a
gene dose-dependent reduction in response to fluid flow shearstress in vitro and an impairment of many cilia-related pathwaysincluding Hh and Wnt signaling. These results indicate that IFT
and cilia have a direct role in regulating osteoblast function andskeletal homeostasis.
ResultsOsteocalcin (Oc)-Cre-mediated conditional deletion ofKif3a in different tissues
The four genotypes from the breeding strategy crossingosteocalcin (Oc; also known as Bglap)–Cre with Kif3aflox/null
mice (Oc-Cre;Kif3aflox/null or Kif3aOc-cKO, Oc-Cre;Kif3aflox/+,Kif3aflox/null and Kif3aflox/+; see Materials and Methods) wereborn at the expected Mendelian frequency, and the survival of all
Kif3a-deficient mice was indistinguishable from that of controlmice (Kif3aflox/+, equivalent to wild type). The normal survival ofconditional Kif3aOc-cKO null mice (Oc-Cre;Kif3aflox/null) contrastswith the perinatal lethality of homozygous Kif3anull/null mice
(Marszalek et al., 1999). To confirm that the Kif3a-floxed allelewas selectively deleted in bone, we performed PCR analysis byusing a combination of primers that specifically detect floxed
Kif3a alleles (Kif3aflox) and the excised floxed Kif3a alleles(Kif3aDflox), as well as wild-type alleles (Kif3a+) in Oc-Cre;Kif3aflox/+ mice (Fig. 1A). Oc-Cre expression is limited to
mature osteoblasts (bone surface osteoblasts that synthesize newbone and osteocytes embedded in bone that regulate boneremodeling) with onset of expression just before birth (E18.5)
(Zhang et al., 2002). We demonstrated that Oc-Cre-mediatedfloxed recombination occurred exclusively in tissues that containosteoblasts, whereas nonskeletal tissues retained the intact floxed
Kif3a alleles (Kif3aflox; Fig. 1B). Consistent with the lack of Cre
expression in the kidney, Kif3aOc-cKO, Oc-Cre;Kif3aflox/+ andKif3aflox/null mice had no cysts in the kidney (data not shown).
To quantify the effect of combined use of the floxed Kif3aflox
allele with the nonfunctional Kif3anull allele to increase the net
efficiency of Kif3a inactivation by Cre recombinase, weexamined the percentage of Kif3a, conditional deleted(Kif3aDflox) and null (Kif3anull) allele expressions in bone
tissues from 6-week-old mice by real-time RT-PCR. Asexpected, Kif3aflox/null mice expressed 50% of the Kif3anull nullallele, whereas Oc-Cre;Kif3aflox/+ mice exhibited approximately
25% excision of the floxed exon 2 from Kif3a, indicating that Oc-Cre-mediated bone-specific deletion of the floxed Kif3a allelewas incomplete (Fig. 1C). The combined effect of Kif3aDflox andKif3anull alleles in Oc-Cre;Kif3aflox/null mice resulted in a net
reduction of Kif3a expression by ,75% in bone (Fig. 1C).Unlike a time-dependent increase of total Pkd1 transcripts duringosteogenic culture (Xiao et al., 2011), we observed no further
increase of total Kif3a transcripts in control Kif3aflox/+
osteoblasts. Consistent with the lower expression of Kif3a
expression in bone, conditional Kif3aOc-cKO null osteoblasts
showed more than 75% inactivation of Kif3a transcripts by real-time RT-PCR during 18 days of osteogenic culture (Fig. 1D). Inaddition, whereas in both Oc-Cre;Kif3aflox/+ and Kif3aflox/null
heterozygous mice there was no change in the appearance ofprimary cilia, in conditional Kif3aOc-cKO null mice there wassubstantially fewer primary cilia in osteoblast cultures(Fig. 1E,F), in association with a 75% lower expression of
Kif3a transcript in osteoblasts derived from conditionalKif3aOc-cKO null mice (Fig. 1D).
A gene dose-independent role of global and Oc-Cre-mediated conditional deletion of Kif3a in postnatalbone formation
We observed no differences in body weight, lean body mass or
fat mass in any of the three Kif3a-deficient mice compared withcontrols at 6 weeks of age (data not shown). Consistent with ourprevious report (Qiu et al., 2010), we found that single
heterozygous Oc-Cre;Kif3aflox/+ or Kif3aflox/null mice had nodemonstrable bone abnormalities (Fig. 2). Bone mineral density(BMD) and bone structure (Fig. 2A,B), as well as mineralapposition rates (MAR; Fig. 2C) were not different in single
heterozygous Oc-Cre;Kif3aflox/+, Kif3aflox/null and age-matchedcontrol mice (Kif3aflox/+). In addition, bone samples from Oc-Cre;Kif3aflox/+ and Kif3aflox/null mice had no detectable changes
in markers of either osteoblast or osteoclast gene expression(Table 1). However, Kif3aflox/null mice had significantly(P,0.05) lower expression of adipocyte-related marker genes,
including peroxisome proliferator-activated receptor gamma(Pparg), adipocyte-specific fatty acid binding protein (aP2; alsoknown as Fabp4) and lipoprotein lipase (Lpl) in long-bone
samples (Table 1).
We observed a significantly (P,0.05) lower BMD, of11,21%, in both female and male conditional Kif3aOc-cKO
mice at 6 weeks of age compared with age-matched control mice
(Kif3aflox/+; Fig. 2A). MicroCT (mCT) analysis revealed that thelower bone mass in male conditional Kif3aOc-cKO null mice wascaused by reduced trabecular bone volume (BV/TV, 42%) and
cortical bone thickness (Ct.Th, 17%; Fig. 2B). These reductionsin bone volume and cortical thickness were associated with a50% decrease in periosteal MAR in male conditional Kif3aOc-cKO
Journal of Cell Science 125 (8)1946
Journ
alof
Cell
Scie
nce
null mice compared with age-matched control mice (Fig. 2C). To
investigate the effects of the combined Kif3aDflox and Kif3anull
deficiency on gene expression profiles in bone, we examined, by
real-time RT-PCR, the expression levels of a panel of
osteoblast lineage-, osteoclast- and adipocyte-related mRNAs
from the tibias of 6-week-old male control and conditional
Kif3aOc-cKO null mice (Table 1). Bone derived from conditional
Kif3aOc-cKO null mice had significantly lower levels of
osteoblast-lineage gene transcripts, including Runx2, osterix
(also known as Sp7 transcription factor 7; Sp7), osteocalcin,
rank ligand (RankL; also known as tumor necrosis factor ligand
superfamily member 11; Tnfsf11) and Dmp1 mRNA, but no
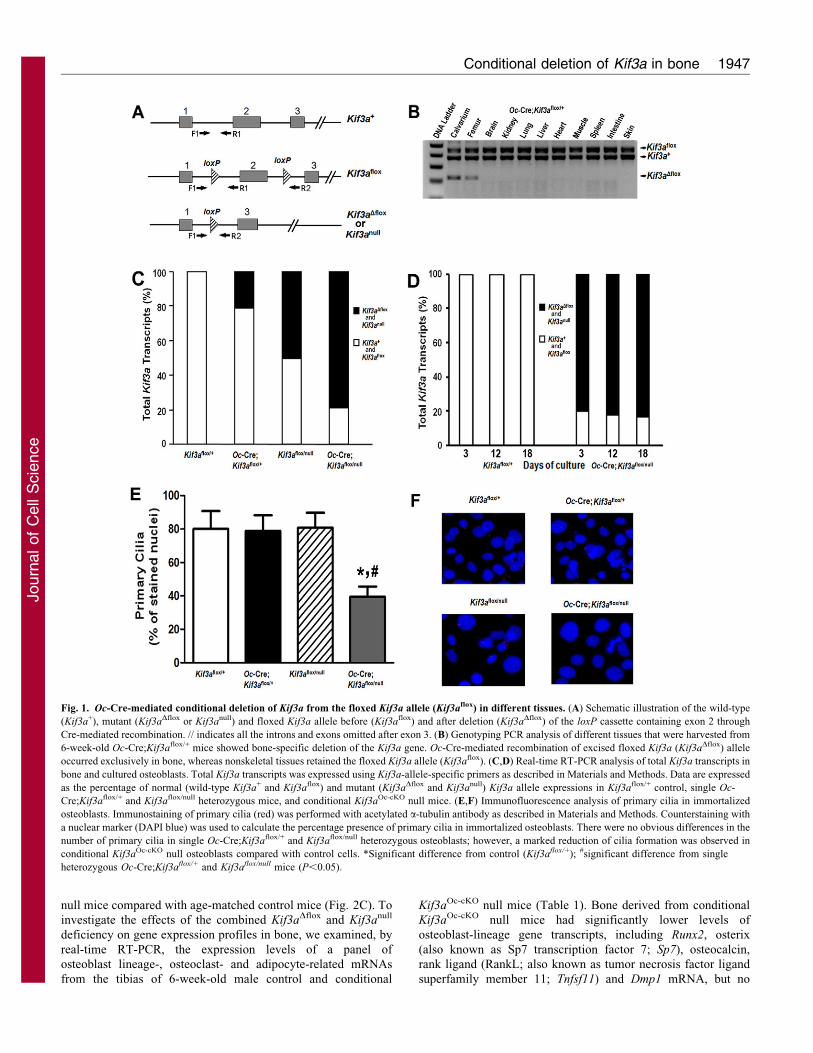
Fig. 1. Oc-Cre-mediated conditional deletion of Kif3a from the floxed Kif3a allele (Kif3aflox) in different tissues. (A) Schematic illustration of the wild-type
(Kif3a+), mutant (Kif3aDflox or Kif3anull) and floxed Kif3a allele before (Kif3aflox) and after deletion (Kif3aDflox) of the loxP cassette containing exon 2 through
Cre-mediated recombination. // indicates all the introns and exons omitted after exon 3. (B) Genotyping PCR analysis of different tissues that were harvested from
6-week-old Oc-Cre;Kif3aflox/+ mice showed bone-specific deletion of the Kif3a gene. Oc-Cre-mediated recombination of excised floxed Kif3a (Kif3aDflox) allele
occurred exclusively in bone, whereas nonskeletal tissues retained the floxed Kif3a allele (Kif3aflox). (C,D) Real-time RT-PCR analysis of total Kif3a transcripts in
bone and cultured osteoblasts. Total Kif3a transcripts was expressed using Kif3a-allele-specific primers as described in Materials and Methods. Data are expressed
as the percentage of normal (wild-type Kif3a+ and Kif3aflox) and mutant (Kif3aDflox and Kif3anull) Kif3a allele expressions in Kif3aflox/+ control, single Oc-
Cre;Kif3aflox/+ and Kif3aflox/null heterozygous mice, and conditional Kif3aOc-cKO null mice. (E,F) Immunofluorescence analysis of primary cilia in immortalized
osteoblasts. Immunostaining of primary cilia (red) was performed with acetylated a-tubulin antibody as described in Materials and Methods. Counterstaining with
a nuclear marker (DAPI blue) was used to calculate the percentage presence of primary cilia in immortalized osteoblasts. There were no obvious differences in the
number of primary cilia in single Oc-Cre;Kif3aflox/+ and Kif3aflox/null heterozygous osteoblasts; however, a marked reduction of cilia formation was observed in
conditional Kif3aOc-cKO null osteoblasts compared with control cells. *Significant difference from control (Kif3aflox/+); #significant difference from single
heterozygous Oc-Cre;Kif3aflox/+ and Kif3aflox/null mice (P,0.05).
Conditional deletion of Kif3a in bone 1947
Journ
alof
Cell
Scie
nce
obvious differences were found in osteoprotegerin (Opg;
Tnfrsf11b) expression compared with that in control mice. In
this regard, the Opg/RankL gene expression ratio was higher in
conditional Kif3aOc-cKO null mice (Table 1). Consistent with a
ratio of Opg/RankL genes that favored the lowerosteoclastogenesis, bone expression of Trap and Mmp9,
markers of bone resorption, were also lower in conditionalKif3aOc-cKO null mice (Table 1). By contrast, the gene encodingPparc, an adipocyte transcription factor, and adipocyte markerssuch as aP2 and Lpl were significantly (P,0.05) higher in tibias
of conditional Kif3aOc-cKO null mice compared with singleheterozygous Kif3aflox/null mice, and the expression of Lpl washigher in conditional Kif3aOc-cKO null mice than that in control
Kif3aflox/+ mice (Table 1).
Changes in gene expression in bone correlated with in serumbiomarkers. In this regard, control Kif3aflox/+ mice had reduced
osteoblastic and osteoclastic markers as a function of age,consistent with an age-dependent decrease in bone formation andresorption. At 6 weeks of age, Kif3aOc-cKO null mice had lowerlevels of certain markers than age-matched control Kif3aflox/+
mice: the osteoblast markers osteocalcin (4268 vs 59610 g/ml)and RankL (79628 vs 122624 pg/ml), as well as the osteoclasticmarker tartrate-resistant acid phosphatase (TRAP; 4.560.43 vs
5.660.29 IU/l). At 24 weeks of age, there was no longer adifference in osteocalcin, but RankL (54616 vs 95622 pg/ml)and TRAP (0.860.11 vs 1.760.32 IU/l) remained lower than in
the age-matched control Kif3aflox/+ mice. Although there was nodifference in the OPG level, the RankL/OPG ratio, which is anindicator of osteoclastogenesis, was lower by approximately 29%
at 6 weeks of age and 47% at 24 weeks of age in Kif3aOc-cK nullmice compared with age-matched control Kif3aflox/+ mice. Thesedata suggest that conditional deletion of Kif3a in osteoblastsresults in diminished osteoblast-mediated bone formation and
osteoclast-mediated bone resorption, resulting in low-turnoverosteopenia. Kif3aOc-cKO null mice had no change in serum ureanitrogen (BUN), calcium or phosphorus levels at either 6 or 24
weeks (data not shown).
Age-dependent effects of global and Oc-Cre-mediatedconditional deletion of Kif3a on bone mass, structure,geometry and mechanical properties
We observed no differences in bone mass and bone structurebetween single heterozygous Oc-Cre; Kif3aflox/+, Kif3aflox/null and
control Kif3aflox/+ mice from 6 to 24 weeks of age. However, weobserved an age-dependent partial recovery of BMD from 21%lower at 6 weeks of age to 7% lower at 24 weeks of age in male
conditional Kif3aOc-cKO null mice compared with age-matchedcontrol mice, indicating age-dependent effects that attenuate theeffects of deleted Kif3a on bone mass (Fig. 3A). mCT analysis
revealed that the increase in bone mass was caused by a recoveryin cortical bone thickness. Indeed, the differences in cortical bonethickness at 6 weeks of age were no longer significant in the fourgenotypes at 24 weeks of age (Fig. 3B). By contrast, there was
less recovery of BV/TV with age in Kif3aOc-cKO mice. In thisregard, BV/TV remained significantly (P,0.05) lower inKif3aOc-cKO mice compared with control mice at 24 weeks of
age (Fig. 3C), a finding that suggests a site-specific interactionbetween Kif3a deficiency and age-dependent changes in bonestructure. Moreover, we found that the mechanism of BMD and
cortical bone thickness recovery resulted from alterations in bonegeometry and led to a compensatory increase in bone mechanicalproperties between 6 and 24 weeks in Kif3aOc-cKO mice
(Table 2). Also, conditional Kif3aOc-cKO null mice showed agreater total bone area, moment of inertia (Ix) and distance fromthe neutral axis to the plane where the load is applied (c) at 6
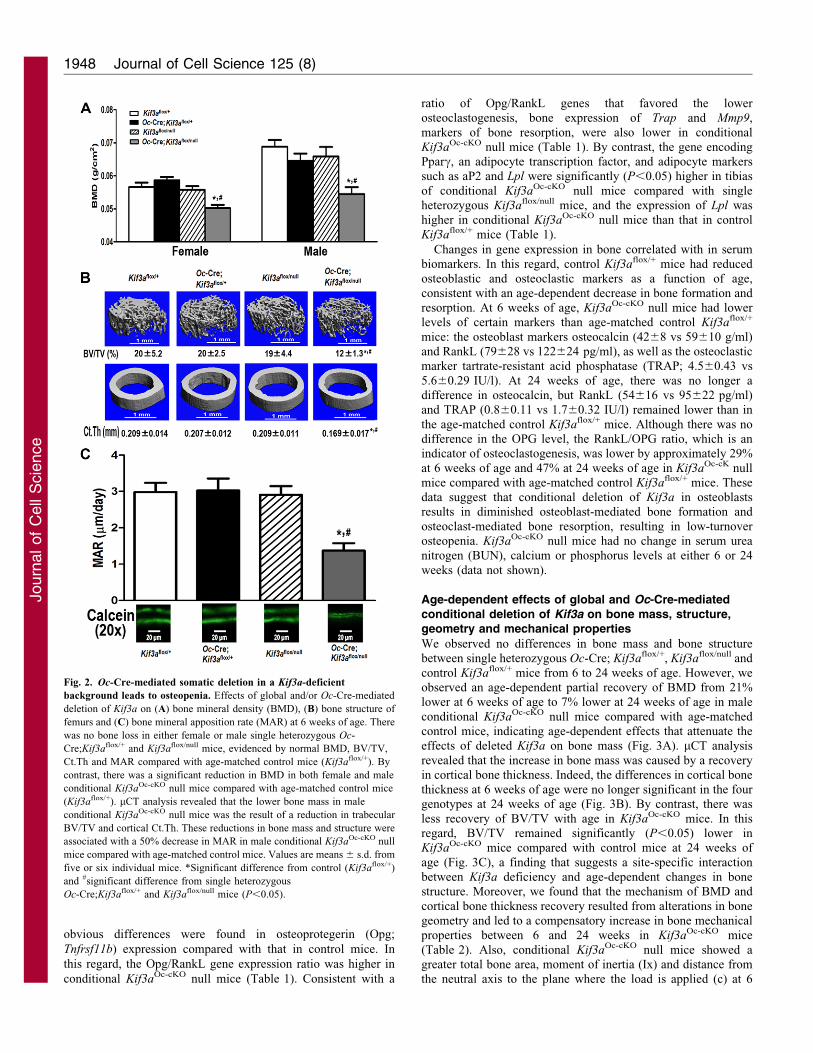
Fig. 2. Oc-Cre-mediated somatic deletion in a Kif3a-deficient
background leads to osteopenia. Effects of global and/or Oc-Cre-mediated
deletion of Kif3a on (A) bone mineral density (BMD), (B) bone structure of
femurs and (C) bone mineral apposition rate (MAR) at 6 weeks of age. There
was no bone loss in either female or male single heterozygous Oc-
Cre;Kif3aflox/+ and Kif3aflox/null mice, evidenced by normal BMD, BV/TV,
Ct.Th and MAR compared with age-matched control mice (Kif3aflox/+). By
contrast, there was a significant reduction in BMD in both female and male
conditional Kif3aOc-cKO null mice compared with age-matched control mice
(Kif3aflox/+). mCT analysis revealed that the lower bone mass in male
conditional Kif3aOc-cKO null mice was the result of a reduction in trabecular
BV/TV and cortical Ct.Th. These reductions in bone mass and structure were
associated with a 50% decrease in MAR in male conditional Kif3aOc-cKO null
mice compared with age-matched control mice. Values are means 6 s.d. from
five or six individual mice. *Significant difference from control (Kif3aflox/+)
and #significant difference from single heterozygous
Oc-Cre;Kif3aflox/+ and Kif3aflox/null mice (P,0.05).
Journal of Cell Science 125 (8)1948
Journ
alof
Cell
Scie
nce

weeks of age. However, there was no difference in cortical bone
area compared with age-matched control mice (Table 2), but the
marrow cavity was larger, resulting in a greater midshaft
diameter compared with that in control mice. However, these
structural differences were no longer evident in 24-week-old
Kif3aOc-cKO mice because of a recovery in femoral midshaft
diameter and the size of the marrow cavity with age (Table 2).
To examine whether changes of femoral bone geometry might
affect bone mechanical properties, we used these femurs to
perform three-point bending experiments (Table 2). At 6 weeks
of age, conditional Kif3aOc-cKO null mice had a lower elastic
modulus in three-point bending experiments but no significant
differences in bending stiffness or maximum force compared
with age-matched control mice (Table 2), indicating that the
changes in bone geometry and bone structure at 6 weeks of age
had an impact on bone strength in conditional Kif3aOc-cKO null
mice. Again, the recovery of bone geometry and structure in 24-
week-old Kif3aOc-cKO normalized these mechanical properties
(Table 2). Both the alterations in geometry and mechanical
properties in Kif3aOc-cKO are similar to observations in Dmp1–
Cre-mediated conditional Pkd1Dmp1-cKO-null mice, consistent
with the known interdependent functions of polycystins and
primary cilia (Xiao et al., 2011).
Effect of conditional deletion of Kif3a on osteoblastic
function ex vivo
To determine the impact of conditional deletion of Kif3a on
osteoblast function ex vivo, we used immortalized osteoblasts
derived from E17.5 control Kif3aflox/+ mice, single heterozygous
Oc-Cre;Kif3aflox/+ mice, single heterozygous Kif3aflox/null mice
and conditional Kif3aOc-cKO null mice. Immortalized osteoblasts
in culture undergo progressive alterations in cell proliferation and
osteoblastic differentiation that recapitulates the osteoblastic
developmental program (Xiao et al., 2004). Consistent with our
previous report (Qiu et al., 2010), we found that heterozygous
Kif3a-deficient mice had no abnormalities in cell proliferation or
osteoblastic differentiation. In this regard, the osteoblasts from
Oc-Cre;Kif3aflox/+ mice and heterozygous Kif3aflox/null mice
exhibited a time-dependent increase of total BrdU incorporation
during 48 hours of osteogenic culture (Fig. 4A) as well as
increased alkaline phosphatase (ALP) activity (a marker of
differentiated osteoblasts) and calcium deposition in long-term
cultures similar to controls (Fig. 4B,C), However, we found that
conditional Kif3aOc-cKO null osteoblasts had a higher BrdU
incorporation than the other three groups, indicating a greater
proliferation rate in Kif3aOc-cKO null osteoblasts (Fig. 4A). In
addition, Kif3aOc-cKO null osteoblasts had impaired osteoblastic
differentiation and maturation, as evidenced by culture duration-
dependent reductions in ALP activity (Fig. 4B), less calcium
deposition in extracellular matrix (Fig. 4C), and lower expression
of osteoblastic differentiation markers, including Runx2-II,
osterix and osteocalcin, compared with control osteoblasts
(Fig. 4D–F). The immortalized primary calvarial osteoblasts
derived from Kif3aOc-cKO mice also exhibited evidence for
divergence from the osteoblastic development program,
analogous to the higher levels of adipogenic markers observed
in vivo. Indeed, under osteogenic culture condition, Kif3aOc-cKO-
derived osteoblasts exhibited a marked increase of adipocyte
markers such as Pparg2 and the aP2 gene (Fig. 4G,H),
suggesting that impairment of osteogenesis was associated with
enhancement of adipogenesis in conditional Kif3aOc-cKO null
osteoblast cultures.
Effects of conditional deletion of Kif3a on intracellularcalcium, Wnt and Hh signaling in immortalized osteoblasts
Loss of Kif3a in a gene-dose-dependent fashion resulted in a
reduction in the length of primary cilia in primary cultured
osteoblasts and altered responses to flow shear stress, consistent
with alterations of cilia functions. Primary cilia were
approximately 27% shorter in Kif3aOc-cKO null osteoblasts
(Fig. 5A). In addition, we found that Kif3aOc-cKO null osteoblasts
had a significantly (P,0.05) lower basal intracellular calcium
([Ca2+]i) concentration compared with control Kif3aflox/+ cells,
whereas [Ca2+]i levels in single heterozygous Oc-Cre;Kif3aflox/+
and Kif3aflox/null osteoblasts were similar to controls (Fig. 5B).
These findings are consistent with previously reported effects of
primary cilia and/or polycystic complex regulation of intracellular
calcium signaling in renal epithelial cells (Nauli et al., 2003; Nauli
et al., 2006; Praetorius et al., 2003; Praetorius and Spring, 2001;
Shiba et al., 2005; Yoder, 2007). The change in intracellular
Table 1. Gene expression profiles in 6-week-old mice
Gene Accession no. Oc-Cre;Kif3aflox/+ Kif3aflox/null Oc-Cre;Kif3aflox/null P-value
Osteoblast lineageRunx2 NM_009820 1.1060.39 0.9660.16 0.6060.13*,# 0.0124Osterix NM_130458 0.9960.24 0.9660.24 0.6260.11*,# 0.0058osteocalcin NM_007541 1.1360.19 1.1260.27 0.7160.15*,# 0.0102Opg MMU94331 1.1860.16 1.0660.16 0.9460.17 0.1310RankL NM_011613 0.9760.22 0.9660.16 0.6660.09*,# 0.0236Dmp1 MMU242625 1.1160.12 1.1260.34 0.6560.08*,# 0.0054
OsteoclastTrap NM_007388 1.1960.18 1.1760.12 0.7360.09*,# 0.0002Mmp9 NM_013599 1.1160.11 1.0460.12 0.8460.11*,# 0.0032
AdipocytePparg NM_009505 0.8160.11 0.626 0.12* 1.0960.23# 0.0031aP2 NM_024406 0.8660.11 0.716 0.10* 1.0860.22# 0.0112Lpl NM_008509 0.7560.13 0.516 0.12* 1.5160.54*,# 0.0001
Data are means 6 s.d. from five to six tibias of 6-week-old individual mice, expressed as the fold differences relative to the housekeeping gene cyclophilin A,subsequently normalized to wild-type mice.
*Significant difference from control Kif3aflox/+ mice; #significant difference from single heterozygous Oc-Cre;Kif3aflox/+Kif3a+/D and Kif3aflox/null mice(P,0.05).
Conditional deletion of Kif3a in bone 1949
Journ
alof
Cell
Scie
nce
calcium was proportional to the magnitude of the fluid flow shear
stress (FFSS, 0.69,9.5 dynes/cm2) in control Kif3aflox/+
osteoblasts (Fig. 5C), with the highest peak-flow-induced
intracellular calcium response of 6.24 dynes/cm2. No further
significant increase in intracellular calcium was observed with 9.5
dynes/cm2, indicating that 6.24 dynes/cm2 is the optimal FFSS to
induce intracellular calcium responses in immortalized osteoblasts
under these experimental conditions. Applying this amount of
FFSS to heterozygous Oc-Cre;Kif3aflox/+, Kif3aflox/null and
Kif3aOc-cKO null osteoblasts demonstrated a gene-dose-dependent
reduction in flow-induced intracellular calcium response
compared with controls (Fig. 5D), indicating that FFSS is a
more sensitive measure of primary cilia abnormalities than the
assessment of cilia length or number. Indeed, an intermediate
calcium response was observed in single heterozygous cells
whereas minimal calcium influx was observed in conditional
Kif3aOc-cKO null osteoblasts in response to FFSS (Fig. 5D).
However, 10 mM caffeine resulted in normal calcium influx in
conditional Kif3aOc-cKO null cells after flow stimulus (data not
shown), indicating the viability of the conditional Kif3aOc-cKO null
cells. Finally, we assessed the effects of FFSS on expression of
‘mechanosensing’ genes by real-time RT-PCR and protein
expression by western blot analysis using RNA and cytoplasmic
proteins isolated from control and Kif3aOc-cKO null cells with or
without FFSS. FFSS increased both the mRNA and protein of
Cox2, a mechanoresponsive gene, in control osteoblasts
(Fig. 5E,F), whereas these changes were less in Kif3aOc-cKO null
cells exposed to identical FFSS (Fig. 5E,F).
To explore additional mechanisms whereby loss of Kif3a leads
to alterations in osteoblast functions, we examined other
signaling molecules associated with primary cilia, including the
patched (Ptch1)–smoothened (Smo)–hedgehog (Hh) and Wnt–b-
catenin pathways. Using total RNA from Kif3aOc-cKO null tibias
and cultured osteoblasts, we found that expression of Gli2, a
downstream gene of Hh signaling, was significantly lower in
Kif3aOc-cKO-derived bone samples compared with Kif3aflox/+
controls (Fig. 6A). Addition of sonic hedgehog (Shh; 1 mg/ml) to
immortalized osteoblast cultures also resulted in significant
(P,0.05) increases in Gli-responsive-promoter luciferase
activity, Gli2 mRNA expression and Gli2 protein levels in
Kif3aflox/+ control cells (Fig. 6B–D). By contrast, Shh failed to
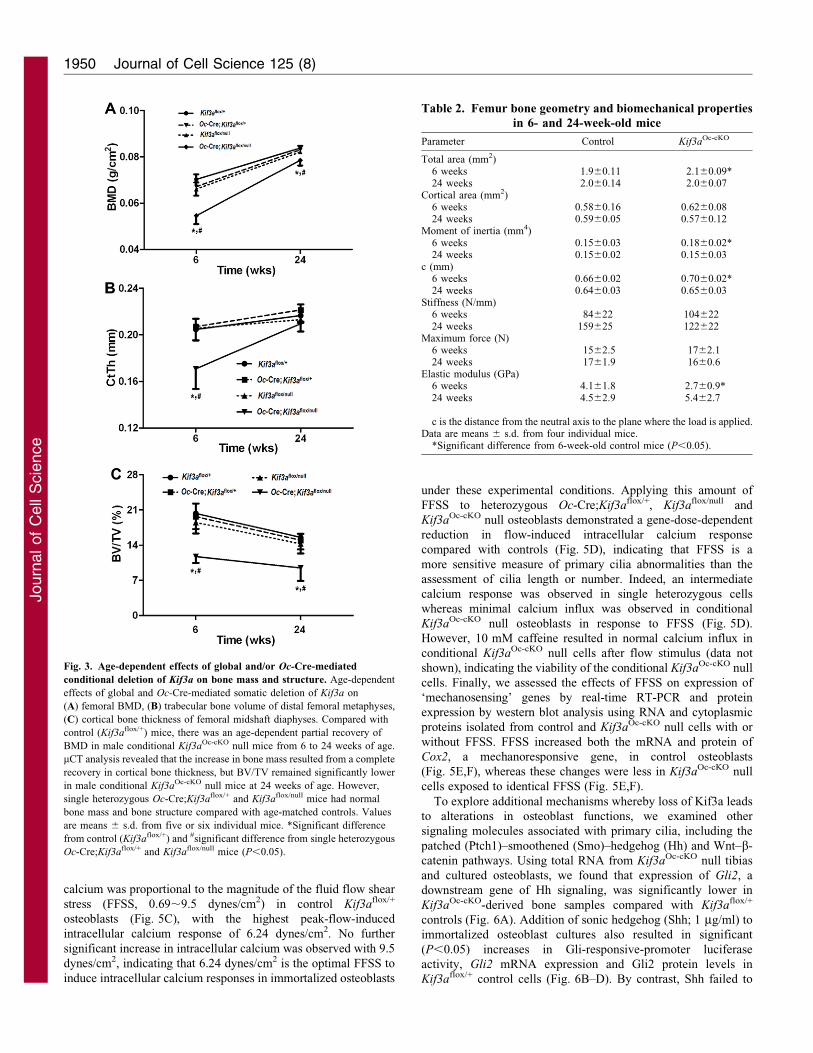
Fig. 3. Age-dependent effects of global and/or Oc-Cre-mediated
conditional deletion of Kif3a on bone mass and structure. Age-dependent
effects of global and Oc-Cre-mediated somatic deletion of Kif3a on
(A) femoral BMD, (B) trabecular bone volume of distal femoral metaphyses,
(C) cortical bone thickness of femoral midshaft diaphyses. Compared with
control (Kif3aflox/+) mice, there was an age-dependent partial recovery of
BMD in male conditional Kif3aOc-cKO null mice from 6 to 24 weeks of age.
mCT analysis revealed that the increase in bone mass resulted from a complete
recovery in cortical bone thickness, but BV/TV remained significantly lower
in male conditional Kif3aOc-cKO null mice at 24 weeks of age. However,
single heterozygous Oc-Cre;Kif3aflox/+ and Kif3aflox/null mice had normal
bone mass and bone structure compared with age-matched controls. Values
are means 6 s.d. from five or six individual mice. *Significant difference
from control (Kif3aflox/+) and #significant difference from single heterozygous
Oc-Cre;Kif3aflox/+ and Kif3aflox/null mice (P,0.05).
Table 2. Femur bone geometry and biomechanical properties
in 6- and 24-week-old mice
Parameter Control Kif3aOc-cKO
Total area (mm2)6 weeks 1.960.11 2.160.09*24 weeks 2.060.14 2.060.07
Cortical area (mm2)6 weeks 0.5860.16 0.6260.0824 weeks 0.5960.05 0.5760.12
Moment of inertia (mm4)6 weeks 0.1560.03 0.1860.02*24 weeks 0.1560.02 0.1560.03
c (mm)6 weeks 0.6660.02 0.7060.02*24 weeks 0.6460.03 0.6560.03
Stiffness (N/mm)6 weeks 84622 10462224 weeks 159625 122622
Maximum force (N)6 weeks 1562.5 1762.124 weeks 1761.9 1660.6
Elastic modulus (GPa)6 weeks 4.161.8 2.760.9*24 weeks 4.562.9 5.462.7
c is the distance from the neutral axis to the plane where the load is applied.Data are means 6 s.d. from four individual mice.
*Significant difference from 6-week-old control mice (P,0.05).
Journal of Cell Science 125 (8)1950
Journ
alof
Cell
Scie
nce

stimulate any of these parameters in the Kif3aOc-cKO null
osteoblasts (Fig. 6B–D), suggesting that either loss of Kif3a
and/or disruption of primary cilium function impairs Hh signaling
in osteoblasts.
To examine the effect of conditional deletion of Kif3a we
examined the expression of Axin2, a direct downstream gene of
the Wnt–b-catenin pathway, in bone and immortalized
osteoblasts. We found that Axin2 was significantly (P,0.05)
lower in Kif3aOc-cKO null bone and osteoblasts compared with
Kif3aflox/+controls (Fig. 6E). Wnt3a–b-catenin transcriptional
activity was also lower, by approximately 35%, in Kif3aOc-cKO
null osteoblasts. In this regard, Wnt3a induced a respective 45-
and 12-fold increase in Super 8xTOPFlash promoter luciferase
activity and Axin2 expression in control osteoblasts (Fig. 6F,G),
but only a 28- and 8-fold increase in these parameters in
Kif3aOc-cKO null osteoblasts (Fig. 6F,G). In agreement with the
changes of Wnt–b-catenin downstream signaling described
above, Wnt3a-induced accumulation of cytoplasmic b-catenin
protein was lower in Kif3aOc-cKO null osteoblasts compared with
controls (Fig. 6H), indicating that conditional deletion of Kif3a
significantly attenuates Wnt–b-catenin signaling in osteoblasts.
DiscussionOur studies show that the selective deletion of Kif3a, a subunit of
kinesin II, an anterograde ciliary motor protein, from osteoblasts
in mice causes defects in osteoblast-mediated bone formation and
abnormalities of multiple cilia-related signaling pathways,
resulting in osteopenia. We found that Kif3aOc-cKO mice had a
Fig. 4. Effects of global and/or Oc-Cre-mediated
conditional deletion of Kif3a on osteoblastic
proliferation and maturation, as well as gene
expression profiles ex vivo. (A) BrdU incorporation.
There was a time-dependent increase of total BrdU
incorporation during 48 hours in osteogenic culture
in osteoblasts from each of the four genotypes.
However, a higher BrdU incorporation was observed
in Kif3aOc-cKO null osteoblasts compared with the
other three groups of osteoblasts at the indicated
times. (B) ALP activity. All osteoblasts from each of
the four genotypes displayed time-dependent
increases in ALP activity during 14 days in culture,
but ALP activity was significantly lower in
Kif3aOc-cKO null osteoblasts compared with the other
three groups of osteoblasts at day 14.
(C) Quantification of mineralization. Alizarin Red-S
was extracted with 10% cetylpyridinium chloride and
quantified as described in Materials and Methods. All
osteoblasts from each of the four genotypes had time-
dependent increases in Alizarin Red-S accumulation
during the 21 days in culture, but the accumulation
was significantly lower in Kif3aOc-cKO null
osteoblasts compared with the other three groups of
osteoblasts at day 21. (D–H) Gene expression
profiles by real-time RT-PCR. Immortalized
Kif3aOc-cKO null osteoblasts in osteogenic medium
showed time-dependent increases in osteogenesis,
evident from increases in osteoblastic markers
(Runx2, osterix and osteocalcin) during 18 days in
culture which were significantly lower than in control
(Kif3aflox/+) osteoblasts at 12 and 18 days (D–F). By
contrast there was a marked increase in adipocyte
markers (Pparg2; PPARc2; and aP2) in the
Kif3aOc-cKO null osteoblasts compared with control
osteoblasts. Values are means 6 s.d. from three
independent experiments. *Significant difference
from control (Kif3aflox/+), and #significant difference
from single heterozygous Oc-Cre;Kif3aflox/+ and
Kif3aflox/null mice (P,0.05).
Conditional deletion of Kif3a in bone 1951
Journ
alof
Cell
Scie
nce

75% less Kif3a transcripts in bone. Loss of Kif3a resulted in
reductions in both the number and length of primary cilia in
osteoblasts, consistent with the known role of Kif3a in primary
cilia formation and function (Lin et al., 2003; Marszalek et al.,
1999). Adult Kif3aOc-cKO null mice exhibited lower bone mineral
density, trabecular bone volume, cortical thickness, bone
formation rate and osteoblast-related gene expression, and had
impaired bone mechanical properties. Because Oc-Cre directs
expression of the recombinase to postnatal osteoblasts, bone
abnormalities in Kif3aOc-cKO mice reflect the function of Kif3a in
differentiated osteoblasts. A direct effect of Kif3a on postnatal
osteoblast function was also demonstrated in ex vivo experiments
in primary osteoblast cultures. These osteoblasts displayed
impairment of intracellular calcium signaling, Shh-induced
upregulation of Hh and Gli2 signaling, and Wnt–b-catenin and
Axin2 activation in osteoblasts.
There are several potential mechanisms whereby disruption of
Kif3a in mature osteoblasts leads to abnormal function in the adult
mouse. Because the C-terminal region of Kif3a binds to the C-
terminus of PC2 and mediates polycystin complex colocalization
Fig. 5. Effects of global and/or Oc-Cre-mediated conditional deletion of Kif3a on baseline and flow-induced intracellular calcium ([Ca2+]i) response, as
well as mechanoresponsive gene expression in osteoblasts. (A) Length of primary cilia (n515,20). A gene dose-dependent reduction of cilium length was
observed in the Kif3a-deficient osteoblasts compared with control Kif3aflox/+ cells. (B) Basal [Ca2+]i levels. Only Kif3aOc-cKO null osteoblasts (n553) showed a
significantly lower basal [Ca2+]i levels compared with the other three group of osteoblasts (n553). (C) Flow-induced [Ca2+]i response curves using different flow
rates. Control Kif3aflox/+ osteoblasts exhibited an FFSS-dependent intracellular Ca2+ response. Of the four flow rates (n54), 6.24 dynes/cm2 was the optimal FFSS
to induce the intracellular Ca2+ response in the immortalized osteoblasts. There was no significant difference between 6.24 dynes/cm2 and 9.50 dynes/cm2 loading.
(D) Flow-induced [Ca2+]i response curves obtained using cells of different genotypes (n54). A gene dose-dependent reduction of flow-induced [Ca2+]i responses
was observed in the Kif3a-deficient osteoblasts compared with control Kif3aflox/+ cells, which is in agreement with the length of primary cilia in Kif3a-deficient
osteoblasts. There was no significant difference between Oc-Cre;Kif3aflox/+ and Kif3aflox/null cells. (E–F) Expression of mechanoresponsive genes. Real-time RT-
PCR and western blot analyses from control Kif3aflox/+ and Kif3aOc-cKO null osteoblasts showed that mRNA and protein levels of Cox-2 were markedly increased
in the loaded control cells but the increase in the loaded Kif3aOc-cKO null cells was much less. Values are means 6 s.d. from three independent experiments.
*Significant difference from control Kif3aflox/+ cells (P,0.05); #significant difference from Oc-Cre;Kif3aflox/+ or Kif3aflox/null cells at P,0.05; @significant
difference from 9.50 dynes/cm2 and 6.24 dynes/cm2 (P,0.05); &significant difference from 1.56 dynes/cm2 at P,0.05. Values sharing the same superscript are
not significantly different, P,0.05.
Journal of Cell Science 125 (8)1952
Journ
alof
Cell
Scie
nce
to primary cilia (Li et al., 2006), the effects of Kif3a on osteoblast
function might be mediated through polycystins. This possibility is
supported by several observations. First, Kif3a and the polycystins,
Pkd1 and Pkd2, are known to be functionally interrelated in other
organs, such as the kidney, where loss of Kif3a or polycystins
cause a common phenotype (i.e. polycystic kidney disease) (Hou
et al., 2002; Lin et al., 2003; Pazour et al., 2002; Ward et al., 2003;
Yoder et al., 2002a; Yoder et al., 2002b). Second, conditional
Kif3aOc-cKO null mice exhibit similar bone phenotypes to
conditional Pkd1Oc-cKO and Pkd1Dmp1-cKO-null mice (Xiao et al.,
2011; Xiao, Z. et al., 2010), consistent with the possibility that
Kif3a regulates osteoblast function through its mediation of the
localization of polycystins to primary cilia in osteoblasts and
osteocytes in bone (Xiao et al., 2008; Xiao et al., 2006). Third, both
immortalized conditional Kif3aOc-cKO- and Pkd1Dmp1-cKO-null
primary osteoblasts exhibit increased cell proliferation and
impaired osteoblast gene expression (Qiu et al., 2010; Xiao
et al., 2011; Xiao, Y. et al., 2010; Xiao, Z. et al., 2010). Fourth,
Kif3aOc-cKO- and Pkd1Dmp1-cKO-null primary osteoblasts exhibit
similar defects in flow-induced changes in intracellular calcium,
consistent with the fact that mutations in Pkd1 and Pkd2 result in
impaired intracellular calcium response to fluid flow in renal
epithelial cells (Nauli et al., 2003; Nauli et al., 2006). Finally, Cox2
expression, Shh-induced Gli2 expression and Wnt3a-stimulated
Axin2 transcription are abnormal in both Kif3aOc-cKO and
Pkd1Dmp1-cKO mice (Qiu et al., 2010; Xiao et al., 2011; Xiao, Z.
et al., 2010).
Our studies do not assess the role of Kif3a in the early stage of
the osteoblast lineage or in skeletal development. Indeed, we found
no evidence for abnormalities in skeletogenesis, because Oc-Cre is
expressed in mature osteoblasts at E18.5, after completion of bone
development. Additional studies in which Kif3a is ablated earlier
in the osteoblast lineage, during embryogenesis, will be needed to
establish a role of Kif3a in osteoblast development and
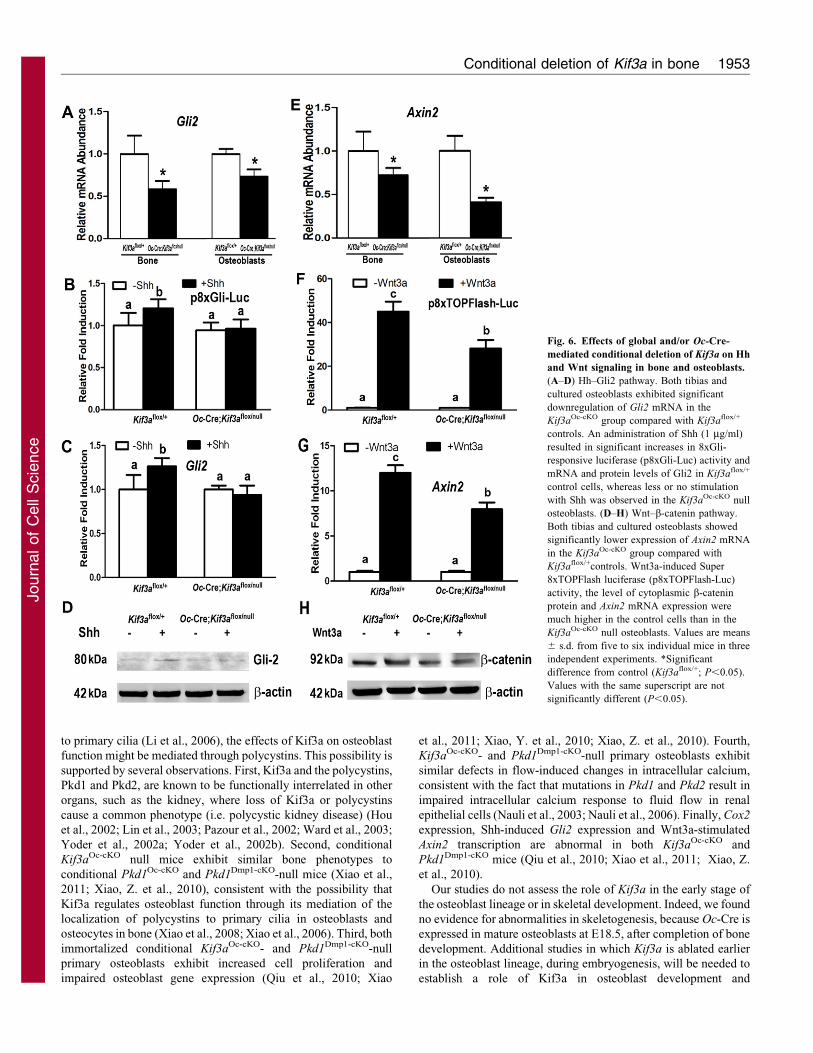
Fig. 6. Effects of global and/or Oc-Cre-
mediated conditional deletion of Kif3a on Hh
and Wnt signaling in bone and osteoblasts.
(A–D) Hh–Gli2 pathway. Both tibias and
cultured osteoblasts exhibited significant
downregulation of Gli2 mRNA in the
Kif3aOc-cKO group compared with Kif3aflox/+
controls. An administration of Shh (1 mg/ml)
resulted in significant increases in 8xGli-
responsive luciferase (p8xGli-Luc) activity and
mRNA and protein levels of Gli2 in Kif3aflox/+
control cells, whereas less or no stimulation
with Shh was observed in the Kif3aOc-cKO null
osteoblasts. (D–H) Wnt–b-catenin pathway.
Both tibias and cultured osteoblasts showed
significantly lower expression of Axin2 mRNA
in the Kif3aOc-cKO group compared with
Kif3aflox/+controls. Wnt3a-induced Super
8xTOPFlash luciferase (p8xTOPFlash-Luc)
activity, the level of cytoplasmic b-catenin
protein and Axin2 mRNA expression were
much higher in the control cells than in the
Kif3aOc-cKO null osteoblasts. Values are means
6 s.d. from five to six individual mice in three
independent experiments. *Significant
difference from control (Kif3aflox/+; P,0.05).
Values with the same superscript are not
significantly different (P,0.05).
Conditional deletion of Kif3a in bone 1953
Journ
alof
Cell
Scie
nce

skeletogenesis. Although global Kif3a-null mice die beforeskeletal development, global deletion of Pkd1 and Pkd2 results
in severe abnormalities of skeletogenesis, probably because oftheir effect on primary cilia and polycytins in the bonedevelopment process. However, in this study, we focused on thefunction of Kif3a in mature osteoblasts, and we cannot exclude the
possibility that disruption of Kif3a through polycystin signalingmight affect the differentiation state of osteoblast as previouslyreported in renal epithelia cells (Rankin et al., 1992). Our current
results show that lower osteoblast-specific gene expressionconcomitant with higher adipocyte-specific gene expression inbones of 6-week-old mice is similar to the data from the osteoblast
cultures in vitro, and probably reflects abnormalities in the boneremodeling process and alterations in marrow fat during theprocess of postnatal bone generation, which recapitulates aspectsof embryonic bone development (Griffith et al., 2009).
Other signaling pathways that are known to regulate osteoblastfunction, including Hh–Gli (Han et al., 2008; Huangfu andAnderson, 2005; Kovacs et al., 2008; Serra, 2008; Veland et al.,
2009; Wong et al., 2009) and Wnt–b-catenin (Christensen et al.,2007; Corbit et al., 2008; Gerdes et al., 2009) are also present inprimary cilia. We found that expression of Gli2, a downstream
gene of Hh signaling, was significantly decreased in bone andosteoblasts from conditional Kif3aOc-cKO null mice comparedwith Kif3aflox/+ controls. We also found that Shh-inducedexpression of Gli2 mRNA and protein were markedly
attenuated in osteoblasts from conditional Kif3aOc-cKO nullmice. With regards to Wnt signaling, we observed that theexpression of Axin2, a direct downstream gene of the Wnt–b-
catenin signaling, was significantly lower in bone and osteoblastsfrom conditional Kif3aOc-cKO null mice compared with Kif3aflox/+
controls. In addition, Wnt3a-induced accumulation of
cytoplasmic b-catenin protein and b-catenin transcriptionalactivity were significantly attenuated in osteoblasts fromconditional Kif3aOc-cKO null mice. These findings suggest that
the skeletal phenotype in Kif3aOc-cKO mice results fromabnormalities in multiple cilia-related pathways including Pkd1,Pkd2, Ca2+, hedgehog and Wnt signaling. Our studies do notseparate specific functions of Kif3a from more global effects of
primary cilium dysfunction caused by loss of Kif3a (Praetoriusand Spring, 2001; Praetorius and Spring, 2003; Yoder, 2007).Comparative analysis of the bone phenotype in additional mouse
genetic models in which ciliogenesis is selectively disrupted bytargeting genes other than Kif3a, such as Ift88, as well asinvestigations of specific downstream signaling pathways, will be
necessary to determine the precise mechanisms whereby Kif3aregulates osteoblast functions.
Because Oc-Cre-mediated deletion of Kif3a in osteoblasts affectsthe terminally differentiated osteocytes, we cannot exclude a
possible role of the osteocyte in the bone phenotype of Kif3aOc-cKO
null mice. There is growing support for a central role of osteocytesin bone mechanosensing (Bonewald and Johnson, 2008; Galli et al.,
2010; Rochefort et al., 2010; Santos et al., 2009; Tatsumi et al.,2007). Using a 6.24 dynes/cm2 FFSS regime, we observed thatglobal and/or conditional deletion of Kif3a produced a gene-dose
effect on flow-induced intracellular calcium response inimmortalized Kif3a-deficient osteoblasts, consistent with reportsby others of cilia-dependent calcium signaling in renal epithelial
cells (Nauli et al., 2003; Nauli et al., 2006). Our findings differ fromthose of Malone et al. who found that disruption of primary cilia inosteoblasts and osteocytes impairs mechanosensing through
mechanisms independent of Ca2+ flux in a high flow shear stress
(12 dynes/cm2) (Malone et al., 2007). The reason for these different
results is not clear. Mechanical loading experiments in vivo, are
needed to establish the mechanosensing function of Kif3a in
osteoblasts and osteocytes.
In conclusion, the disruption of Kif3a in postnatal osteoblasts
either directly, through Kif3a regulation of specific signaling
pathways, or indirectly, through the broader effects of Kif3a
through disruption of primary cilium formation and/or function,
results in osteopenia. Disruption of Kif3a ex vivo affects cilia-
related pathways, such as intracellular calcium, Hedgehog and
Wnt signaling.
Materials and MethodsMice
We obtained the floxed Kif3a mice from Lawrence S. B. Goldstein at theUniversity of California San Diego (Marszalek et al., 1999) and osteocalcin (Oc)-Cre mice from Thomas Clemens at the University of Alabama (Zhang et al., 2002).
The Kif3anull/+ heterozygous mice were from our laboratory stock, as previouslydescribed (Qiu et al., 2010). These mice were bred and maintained on a C57BL/6Jbackground. Because Cre-recombinase-mediated deletion of a single flox/nullallele reduces the risk of mosaicism that could occur because of the less than 100%efficiency of Cre recombinase to excise two floxed alleles (flox/flox) (Kwan,
2002), we created double heterozygous Oc-Cre;Kif3anull/+ mice and homozygousKif3aflox/flox mice. Double heterozygous Oc-Cre;Kif3anull/+ mice were mated withhomozygous Kif3aflox/flox mice to generate excised floxed Kif3a heterozygous (Oc-Cre;Kif3aflox/+) and null mice (Oc-Cre;Kif3aflox/null or Kif3aOc-cKO), as well as
Kif3a heterozygous mice (Kif3anull/flox) and Oc-Cre-negative control mice(Kif3aflox/+, equivalent to wild type). These last four genotypes were used forskeletal phenotype analysis and primary osteoblast cultures. All animal researchwas conducted according to guidelines provided by the National Institutes of
Health and the Institute of Laboratory Animal Resources, National ResearchCouncil. The University of Tennessee Health Science Center’s Animal Care andUse Committee approved all animal studies (Protocol number 1885 and 1889).
Genotyping polymerase chain reaction (PCR) to detect bone-specific
deletions
Genomic DNA was prepared from tail and other tissue specimens using standard
procedures (Xiao et al., 2005). PCR genotyping was performed using the followingprimers (Fig. 1A) (Lin et al., 2003; Qiu et al., 2010): Kif3a wild-type (Kif3a+) andfloxed (Kif3aflox) alleles, F1, 59-AGGGCAGACGGAAGGGTGG-39, R1, 59-TCTGTGAGTTTGTGACCAGCC-39; Kif3a null (Kif3anull) and conditional null
(Kif3aDflox) alleles, F1, 59-AGGGCAGACGGAAGGGTGG-39, R2, 59-TGGCAGGTCAATGGACGCAG-39. The Kif3a floxed (Kif3aflox), wild-type(Kif3a+) and Kif3a null (or conditional null; Kif3anull or Kif3aDflox) alleles wereidentified, using 2% agarose gels as 490 bp, 360 bp and 200 bp bands, respectively
(Fig. 1B) (Lin et al., 2003; Qiu et al., 2010).
Bone densitometry, histomorphometric and microcomputed tomography
analyses
BMD of femurs was assessed at 6 and 24 weeks of age with a LUNARPIXIMUS
bone densitometer (Lunar Corp., Madison, WI). Calcein (Sigma, St. Louis, MO)double labeling of bone and histomorphometric analyses of periosteal MAR in
tibias were performed using the osteomeasure analysis system (OsteoMetrics,Decatur, GA) (Glass et al., 2005; Xiao et al., 2005). The distal femoral metaphyseswere also scanned with a Scanco mCT 40 (Scanco Medical AG, Bruttisellen,Switzerland). Three-dimensional images were analyzed to determine bone volume,
trabecular volume and cortical thickness as previously described (Xiao et al.,2005).
Bone geometry and mechanical property testing
For each femur, mCT data (1520 slices per femur) were imported into the imagingsoftware Amira (Pro Medicus Limited, Richmond, Australia), and saved in aDICOM file format. Measurements of the inner and outer diameters in both the x
and y directions were taken using a DICOM viewer (Santa DICOM Viewer FREE,Santesoft, Athens, Greece). To increase the accuracy of the measurements, athreshold on the luminosity was applied with a lower limit of 1000 and an upperlimit of 1500. For each specimen, three slices were measured and the averageswere used for the calculations. From these data, the moment area of inertia (I) was
calculated using the formula I~p
64(X1Y 3
1 {X2Y 32 ), where X1 and Y1 are the outer
diameters and X2 and Y2 are the inner diameters. After I was calculated, it wascombined with the force–displacement data from the three-point bending tests to
Journal of Cell Science 125 (8)1954
Journ
alof
Cell
Scie
nce

calculate the apparent elastic modulus, Eapp, using the following expression:
Eapp~FL3 FL3
48dI, where F is the applied force, L is the span and d is the deflection.
Eight femurs from 6-week-old male mice and eight femurs from 24-week-oldmale were used for the three-point bend testing. Femurs were stored at 220 Cprior to data acquisition from the three-point bending experiments. On the day oftesting, the bones were thawed on ice and rehydrated with 16 phosphate-bufferedsaline (PBS) at room temperature for 5 minutes before testing. The distancebetween the supports was held constant for all femurs, at 6 mm, and the radius ofsupports was 0.5 mm. The femurs were tested using an Instron 33R (Instron,Norwood, MA) at a rate of 2 mm/second to a 40% decline in maximum load. Loadmagnitude and displacement data were collected using BluehillH Materials TestingSoftware (Instron).
BiochemistrySerum osteocalcin levels were measured a mouse Osteocalcin EIA (BiomedicalTechnologies, Inc., Stoughton, MA). Serum blood urea nitrogen (BUN) wasdetermined using a BUN diagnostic kit from Pointe Scientific, Inc. (Canton, MI).Serum calcium was measured by the colorimetric cresolphthalein binding method,and phosphorus was measured by the phosphomolybdate–ascorbic acid method(Stanbio Laboratory, Boerne, TX). Serum OPG and RankL were measured usingmouse ELISA kits (QuantikineH, R&D Systems, Minneapolis, MN), and serumTRAP was assayed with the ELISA-based SBA Sciences mouse TRAPTM assay(Immunodiagnostic Systems, Fountain Hills, AZ).
Bone RNA isolation and real-time RT-PCRFor quantitative real-time RT-PCR, 1.0 mg total RNA isolated from whole tibias of6-week-old control and Kif3a-deficient mice was reverse transcribed as previouslydescribed (Xiao et al., 2004). PCR reactions contained 20 ng template (cDNA orRNA), 375 nM each forward and reverse primers and 16 SsoFastTM EvaGreenHsupermix (Bio-Rad, Hercules, CA) in a total of 10 ml reaction volume. Thethreshold cycle (Ct) of tested gene product from the indicated genotype wasnormalized to the Ct for cyclophilin A. Expression of total Kif3a transcripts wasperformed using the following Kif3a-allele-specific primers in exon 2: forwardprimer of normal Kif3a+ transcript (Kif3a+ plus Kif3aflox): 59-GCTATAGACA-GGCCGTCAGC-39 and reverse primer: 59-GTCTTTGGAGGTTCGTTGGA-39.The normal Kif3a+ versus cyclophilin A was normalized to the mean ratio of fivecontrol mice, which was set to 1. The percentage of Kif3a null (Kif3anull) and/orconditional deleted (Kif3aDflox) transcripts was calculated from the relative levelsof the normal Kif3a+ transcripts in different Kif3a-deficient mice (Xiao, Z. et al.,2010). All primer information of other genes used in real-time RT-PCR can befound in our previous report (Xiao et al., 2011).
Cell proliferation, osteoblastic differentiation and gene expression profilesin immortalized osteoblast cultures
Calvaria from E17.5 control and Kif3a-deficient embryos were used to isolate primaryosteoblasts by sequential collagenase digestion at 37 C. To engineer immortalosteoblast cell lines, isolated primary osteoblasts were infected using a retroviral vectorcarrying SV40 large and small T antigen as previously described (Borton et al., 2001;Xiao et al., 2004). Briefly, cells were grown in 100-mm plates at 50–60% confluencythe day before infection. On the day of infection, the medium was removed andmedium containing SV40 large and small T antigen-helper-free viral supernatant wasadded in the presence of 4 mg/ml polybrene (Sigma, St. Louis, MO) for 48 hours. Thecells were allowed to recover for 72 hours followed by selection with 1 mg/mlpuromycin (Sigma) for up to 15 days. The immortalized osteoblasts were cultured in a-MEM containing 10% FBS and 1% penicillin and streptomycin (P/S) and characterizedfollowing the protocols below. Cell proliferation was detected by BrdU incorporationassays following the manufacturer’s directions (QIA58, Calbiochem, Gibbstown, NJ).To induce differentiation, the immortalized osteoblasts were plated at a density of26104 cells per well in a 12-well plate and 4604 cells per well in a 6-well plate andgrown for 21 days in a-MEM containing 10% FBS supplemented with 5 mM b-glycerophosphate and 25 mg/ml ascorbic acid. ALP activity and Alizarin Red-Shistochemical staining for mineralization were performed as previously described(Xiao et al., 2006; Xiao et al., 2004). Total DNA content was measured with aPicoGreenH dsDNA quantitation kit (Molecular Probes, Eugene, OR). Proteinconcentrations of the supernatant were determined with a Bio-Rad protein assay kit.For gene expression profiles, 1.0 mg total RNA were isolated from primary osteoblastscultured for 3, 12 and 18 days in differentiation medium. The cDNAs were generatedusing an iScript reverse transcription kit (Bio-Rad). PCR reactions contained 20 ngtemplate (cRNA or cDNA), 375 nM each forward and reverse primer, 16 SsoFastEvaGreenH supermix (Bio-Rad), in a total of 10 ml reaction volume. The Ct of testedgene product from the indicated genotype was normalized to the Ct for cyclophilin A aspreviously described (Xiao et al., 2008; Xiao et al., 2006; Xiao et al., 2004).
ImmunofluorescenceThe immortalized control and Kif3a-deficient osteoblasts were grown on collagen-coated 4-well chambers at 16105 cells per well and kept at confluence for at least 3days. The end of the culture, the cells were washed three times with PBS, fixed with
cold 4% paraformaldehyde, 0.2% Triton X-100 for 10 minutes at room temperature,and washed with PBS three times. The cells were incubated for 30 minutes in 1%
BSA before incubation with primary acetylated a-tubulin antibody (1:4000; T6793,Sigma) for 1 hour at room temperature. After washing three times in PBS, cells were
treated with secondary Texas-Red-labeled anti-mouse IgG (1:400; 715-076-150,Jackson ImmunoResearch, West Grove, PA) in 1% BSA for 1 hour at roomtemperature and washed three times in PBS before mounting with ProLongH Gold
antifade reagent (P36935, Invitrogen). Nuclei were counterstained with DAPI blue.Photographs were taken under a microscope with magnification of 406 and 1006to count the number and measuring the length of primary cilia, respectively, in theseosteoblasts as previously described (Xiao et al., 2006).
Intracellular calcium measurements in vitro
We measured basal intracellular calcium ([Ca2+]i) concentration and flow-induced
intracellular calcium response in immortalized control and Kif3a-deficientosteoblasts as previously described (Xiao et al., 2011). Briefly, the immortalizedcells were cultured on type I rat tail collagen-coated 40-mm diameter glass slides
at 80–90% confluency in a-MEM containing 2% FBS and 1% P/S for 3 days. Thecells were loaded with 3 mM Fura2-AM (Molecular Probes, Eugene, OR), a
fluorescent Ca2+ probe, in Hank’s balanced salt solution (HBSS) that contained 2%FBS and 20 mM HEPES for 30 minutes at 37 C. Loaded cells were incubated for
an additional 45 minutes with HBSS alone to ensure complete de-esterification ofthe fluorescent molecule. A glass slide was then placed in an FCS2 parallel plateflow chamber (Bioptechs, Inc., Butler, PA), 0.25614622 mm. A fresh bolus of
flow medium was added to the chamber and the cells were left undisturbed for30 minutes. The flow medium consisted of phenol-free a-MEM and 2% FBS
equilibrated with 5% CO2, 95% air at 37 C. The chamber was mounted on thestage of an inverted microscope with a CCD camera to allow real-time recordingof fluorescence intensity (F340/F380 ratio) to generate ratiometric video images of
individual static cells or cells exposed to pulsatile laminar fluid flow (IntracellularImaging, Inc., Cincinnati, OH). To obtain the optimal FFSS to induce response of
intracellular calcium in individual osteoblasts, the immortalized control cells wereexposed to various pulsatile laminar fluid flow rates resulting in shear stresses of
0.69, 1.56, 6.24 and 9.5 dynes/cm2. To assess mechanoresponsive gene expression,total RNA was harvested and the cells subjected to FFSS (6.24 dynes/cm2) for30 minutes and then returned to static culture for 30 minutes (post-FFSS), which
was based on previous studies in osteoblasts (Mehrotra et al., 2006).
Transient transfection and western blot analysis
The immortalized control and conditional Kif3a null osteoblasts were cultured ina-MEM containing 10% FBS and 1% P/S. To examine whether conditional
deletion of Kif3a (disruption of ciliogenesis) has an impact on Hh signaling inosteoblasts, 1.56106 cells were transfected with 3.0 mg Gli-responsive luciferase
reporter construct (8xGli-Luc) (Zhao et al., 2009), 3.0 mg pcDNA3.1 empty vectorand 0.6 mg Renilla luciferase-null (RL-null) as an internal control plasmid by
electroporation using a Cell Line Nucleofector Kit R according to themanufacturer’s protocol (Amaxa, Inc., Gaithersburg, MD). The cells werecultured in a-MEM supplemented with 1% FBS and the relative luciferase
activity of cell lysates was measured using a luciferase assay kit (Promega,Madison, WI) 72 hours after transfection in the presence or absence of 1 mg/ml of
recombinant mouse Shh N-terminus (Shh-N) for the last 8 hours (Qiu et al., 2010;Xiao, Z. et al., 2010). Total RNA was also isolated for real-time RT-PCR analysis.
To explore potential abnormalities of the Wnt pathway in conditional Kif3a null
mice, control (Kif3aflox/+) and conditional Kif3a null (Kif3aOc-cko) osteoblasts weretransiently cotransfected with 3.0 mg Super 8xTOPFlash luciferase reporter plasmid
(8xTOPFlash-Luc), 3.0 mg pcDNA3.1 empty vector or 0.6 mg Renilla luciferase-null(RL-null; Promega) as an internal control, by electroporation. Promoter activity was
assessed by measuring luciferase activity 48 hours after transfection in the presenceor absence of 100 ng/ml recombinant Wnt3a for the last 8 hours (Qiu et al., 2010;Xiao, Z. et al., 2010). Total RNA was also isolated for real-time RT-PCR analysis.
To examine the amounts of cytoplasmic Cox-2, Gli-2 and b-catenin, the cellswere prepared using 16 passive lysis buffer for 30 minutes at 4 C (Promega) and
centrifuged at 100,000 g for 45 minutes at 4 C. Protein concentrations of thesupernatant were determined with a Bio-Rad protein assay kit. Equal quantities ofprotein were applied to a NuPAGETM 4–12% Bis-Tris gel (Invitrogen, Carlsbad,
CA) and analyzed with standard western blot protocols (HRP-conjugatedsecondary antibodies from Santa Cruz Biotechnology, CA, and ECL from
Amersham Biosciences, Little Chalfont, Buckinghamshire, UK). Antibody againstCox2 (4842S) was from Cell Signaling Technology (Beverly, MA). Antibodyagainst Gli2 (ab7195, 80 kDa) was purchased from Abcam (San Francisco, CA).
Anti-b-catenin (sc-7199) and anti- b-actin (sc-47778) antibodies were from SantaCruz Biotechnology.
Statistical analysis
We evaluated differences between two groups using unpaired t-tests and multiplegroups using one-way analysis of variance. All values are expressed as means 6
Conditional deletion of Kif3a in bone 1955
Journ
alof
Cell
Scie
nce

s.d. All computations were performed using GraphPad Prism5 (GraphPad SoftwareInc., La Jolla, CA, USA).
FundingThis work was supported by the National Institutes of Health [grantnumbers R21-AR056794 to Z.S.X. and R01-DK083303 to L.D.Q.].Deposited in PMC for release after 12 months.
ReferencesBerbari, N. F., O’Connor, A. K., Haycraft, C. J. and Yoder, B. K. (2009). The
primary cilium as a complex signaling center. Curr. Biol. 19, R526-R535.
Bonewald, L. F. and Johnson, M. L. (2008). Osteocytes, mechanosensing and Wntsignaling. Bone 42, 606-615.
Borton, A. J., Frederick, J. P., Datto, M. B., Wang, X. F. and Weinstein, R. S.
(2001). The loss of Smad3 results in a lower rate of bone formation and osteopeniathrough dysregulation of osteoblast differentiation and apoptosis. J. Bone Miner. Res.
16, 1754-1764.
Christensen, S. T., Pedersen, L. B., Schneider, L. and Satir, P. (2007). Sensory ciliaand integration of signal transduction in human health and disease. Traffic 8, 97-109.
Corbit, K. C., Shyer, A. E., Dowdle, W. E., Gaulden, J., Singla, V., Chen, M. H.,
Chuang, P. T. and Reiter, J. F. (2008). Kif3a constrains beta-catenin-dependent Wntsignalling through dual ciliary and non-ciliary mechanisms. Nat. Cell Biol. 10, 70-76.
Davis, E. E., Brueckner, M. and Katsanis, N. (2006). The emerging complexity of the
vertebrate cilium: new functional roles for an ancient organelle. Dev. Cell 11, 9-19.
Galli, C., Passeri, G. and Macaluso, G. M. (2010). Osteocytes and WNT: themechanical control of bone formation. J. Dent. Res. 89, 331-343.
Gerdes, J. M., Davis, E. E. and Katsanis, N. (2009). The vertebrate primary cilium indevelopment, homeostasis, and disease. Cell 137, 32-45.
Glass, D. A., 2nd, Bialek, P., Ahn, J. D., Starbuck, M., Patel, M. S., Clevers, H.,
Taketo, M. M., Long, F., McMahon, A. P., Lang, R. A. et al. (2005). Canonical
Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev.
Cell 8, 751-764.
Goetz, S. C. and Anderson, K. V. (2010). The primary cilium: a signalling centreduring vertebrate development. Nat. Rev. Genet. 11, 331-344.
Griffith, J. F., Yeung, D. K., Ahuja, A. T., Choy, C. W., Mei, W. Y., Lam, S. S.,
Lam, T. P., Chen, Z. Y. and Leung, P. C. (2009). A study of bone marrow andsubcutaneous fatty acid composition in subjects of varying bone mineral density.Bone 44, 1092-1096.
Han, Y. G., Spassky, N., Romaguera-Ros, M., Garcia-Verdugo, J. M., Aguilar, A.,
Schneider-Maunoury, S. and Alvarez-Buylla, A. (2008). Hedgehog signaling andprimary cilia are required for the formation of adult neural stem cells. Nat. Neurosci.
11, 277-284.
Haycraft, C. J. and Serra, R. (2008). Cilia involvement in patterning and maintenanceof the skeleton. Curr. Top. Dev. Biol. 85, 303-332.
Haycraft, C. J., Zhang, Q., Song, B., Jackson, W. S., Detloff, P. J., Serra, R. and
Yoder, B. K. (2007). Intraflagellar transport is essential for endochondral boneformation. Development 134, 307-316.
Hou, X., Mrug, M., Yoder, B. K., Lefkowitz, E. J., Kremmidiotis, G., D’Eustachio,
P., Beier, D. R. and Guay-Woodford, L. M. (2002). Cystin, a novel cilia-associatedprotein, is disrupted in the cpk mouse model of polycystic kidney disease. J. Clin.
Invest. 109, 533-540.
Huangfu, D. and Anderson, K. V. (2005). Cilia and Hedgehog responsiveness in themouse. Proc. Natl. Acad. Sci. USA 102, 11325-11330.
Kolpakova-Hart, E., Jinnin, M., Hou, B., Fukai, N. and Olsen, B. R. (2007). Kinesin-2 controls development and patterning of the vertebrate skeleton by Hedgehog- andGli3-dependent mechanisms. Dev. Biol. 309, 273-284.
Kovacs, J. J., Whalen, E. J., Liu, R., Xiao, K., Kim, J., Chen, M., Wang, J., Chen,
W. and Lefkowitz, R. J. (2008). Beta-arrestin-mediated localization of smoothenedto the primary cilium. Science 320, 1777-1781.
Koyama, E., Young, B., Nagayama, M., Shibukawa, Y., Enomoto-Iwamoto, M.,
Iwamoto, M., Maeda, Y., Lanske, B., Song, B., Serra, R. et al. (2007). Conditional
Kif3a ablation causes abnormal hedgehog signaling topography, growth platedysfunction, and excessive bone and cartilage formation during mouse skeletogenesis.Development 134, 2159-2169.
Kwan, K. M. (2002). Conditional alleles in mice: practical considerations for tissue-specific knockouts. Genesis 32, 49-62.
Li, Q., Montalbetti, N., Wu, Y., Ramos, A., Raychowdhury, M. K., Chen, X. Z. and
Cantiello, H. F. (2006). Polycystin-2 cation channel function is under the control ofmicrotubular structures in primary cilia of renal epithelial cells. J. Biol. Chem. 281,37566-37575.
Lin, F., Hiesberger, T., Cordes, K., Sinclair, A. M., Goldstein, L. S., Somlo, S. and
Igarashi, P. (2003). Kidney-specific inactivation of the KIF3A subunit of kinesin-IIinhibits renal ciliogenesis and produces polycystic kidney disease. Proc. Natl. Acad.
Sci. USA 100, 5286-5291.
Maeda, Y., Nakamura, E., Nguyen, M. T., Suva, L. J., Swain, F. L., Razzaque, M. S.,
Mackem, S. and Lanske, B. (2007). Indian Hedgehog produced by postnatalchondrocytes is essential for maintaining a growth plate and trabecular bone. Proc.
Natl. Acad. Sci. USA 104, 6382-6387.
Malone, A. M., Anderson, C. T., Tummala, P., Kwon, R. Y., Johnston, T. R.,
Stearns, T. and Jacobs, C. R. (2007). Primary cilia mediate mechanosensing in bone
cells by a calcium-independent mechanism. Proc. Natl. Acad. Sci. USA 104, 13325-13330.
Marszalek, J. R., Ruiz-Lozano, P., Roberts, E., Chien, K. R. and Goldstein, L. S.
(1999). Situs inversus and embryonic ciliary morphogenesis defects in mouse mutantslacking the KIF3A subunit of kinesin-II. Proc. Natl. Acad. Sci. USA 96, 5043-5048.
McGlashan, S. R., Haycraft, C. J., Jensen, C. G., Yoder, B. K. and Poole, C. A.
(2007). Articular cartilage and growth plate defects are associated with chondrocytecytoskeletal abnormalities in Tg737orpk mice lacking the primary cilia proteinpolaris. Matrix Biol. 26, 234-246.
Mehrotra, M., Saegusa, M., Voznesensky, O. and Pilbeam, C. (2006). Role of Cbfa1/Runx2 in the fluid shear stress induction of COX-2 in osteoblasts. Biochem. Biophys.
Res. Commun. 341, 1225-1230.
Murcia, N. S., Richards, W. G., Yoder, B. K., Mucenski, M. L., Dunlap, J. R. andWoychik, R. P. (2000). The Oak Ridge Polycystic Kidney (orpk) disease gene isrequired for left-right axis determination. Development 127, 2347-2355.
Nauli, S. M., Alenghat, F. J., Luo, Y., Williams, E., Vassilev, P., Li, X., Elia, A. E.,Lu, W., Brown, E. M., Quinn, S. J. et al. (2003). Polycystins 1 and 2 mediatemechanosensation in the primary cilium of kidney cells. Nat. Genet. 33, 129-137.
Nauli, S. M., Rossetti, S., Kolb, R. J., Alenghat, F. J., Consugar, M. B., Harris, P. C.,
Ingber, D. E., Loghman-Adham, M. and Zhou, J. (2006). Loss of polycystin-1 in humancyst-lining epithelia leads to ciliary dysfunction. J. Am. Soc. Nephrol. 17, 1015-1025.
Pazour, G. J., San Agustin, J. T., Follit, J. A., Rosenbaum, J. L. and Witman, G. B.(2002). Polycystin-2 localizes to kidney cilia and the ciliary level is elevated in orpkmice with polycystic kidney disease. Curr. Biol. 12, R378-R380.
Pedersen, L. B., Veland, I. R., Schrøder, J. M. and Christensen, S. T. (2008).Assembly of primary cilia. Dev. Dyn. 237, 1993-2006.
Praetorius, H. A. and Spring, K. R. (2001). Bending the MDCK cell primary ciliumincreases intracellular calcium. J. Membr. Biol. 184, 71-79.
Praetorius, H. A. and Spring, K. R. (2003). Removal of the MDCK cell primary ciliumabolishes flow sensing. J. Membr. Biol. 191, 69-76.
Praetorius, H. A., Frokiaer, J., Nielsen, S. and Spring, K. R. (2003). Bending theprimary cilium opens Ca2+-sensitive intermediate-conductance K+ channels in MDCKcells. J. Membr. Biol. 191, 193-200.
Qiu, N., Cao, L., David, V., Quarles, L. D. and Xiao, Z. (2010). Kif3a deficiencyreverses the skeletal abnormalities in Pkd1 deficient mice by restoring the balancebetween osteogenesis and adipogenesis. PLoS ONE 5, e15240.
Rankin, C. A., Grantham, J. J. and Calvet, J. P. (1992). C-fos expression ishypersensitive to serum-stimulation in cultured cystic kidney cells from the C57BL/6J-cpk mouse. J. Cell. Physiol. 152, 578-586.
Rochefort, G. Y., Pallu, S. and Benhamou, C. L. (2010). Osteocyte: the unrecognizedside of bone tissue. Osteoporos. Int. 21, 1457-1469.
Santos, A., Bakker, A. D. and Klein-Nulend, J. (2009). The role of osteocytes in bonemechanotransduction. Osteoporos. Int. 20, 1027-1031.
Serra, R. (2008). Role of intraflagellar transport and primary cilia in skeletaldevelopment. Anat. Rec. (Hoboken) 291, 1049-1061.
Shiba, D., Takamatsu, T. and Yokoyama, T. (2005). Primary cilia of inv/inv mouserenal epithelial cells sense physiological fluid flow: bending of primary cilia and Ca2+
influx. Cell Struct. Funct. 30, 93-100.
Song, B., Haycraft, C. J., Seo, H. S., Yoder, B. K. and Serra, R. (2007). Developmentof the post-natal growth plate requires intraflagellar transport proteins. Dev. Biol. 305,202-216.
St-Jacques, B., Hammerschmidt, M. and McMahon, A. P. (1999). Indian hedgehogsignaling regulates proliferation and differentiation of chondrocytes and is essentialfor bone formation. Genes Dev. 13, 2072-2086.
Tatsumi, S., Ishii, K., Amizuka, N., Li, M. Q., Kobayashi, T., Kohno, K., Ito, M.,
Takeshita, S. and Ikeda, K. (2007). Targeted ablation of osteocytes inducesosteoporosis with defective mechanotransduction. Cell Metab. 5, 464-475.
Veland, I. R., Awan, A., Pedersen, L. B., Yoder, B. K. and Christensen, S. T. (2009).Primary cilia and signaling pathways in mammalian development, health and disease.Nephron Physiol. 111, 39-53.
Ward, C. J., Yuan, D., Masyuk, T. V., Wang, X., Punyashthiti, R., Whelan, S.,
Bacallao, R., Torra, R., LaRusso, N. F., Torres, V. E. et al. (2003). Cellular andsubcellular localization of the ARPKD protein; fibrocystin is expressed on primarycilia. Hum. Mol. Genet. 12, 2703-2710.
Wong, S. Y., Seol, A. D., So, P. L., Ermilov, A. N., Bichakjian, C. K., Epstein, E. H.,Jr, Dlugosz, A. A. and Reiter, J. F. (2009). Primary cilia can both mediate andsuppress Hedgehog pathway-dependent tumorigenesis. Nat. Med. 15, 1055-1061.
Wu, Y., Dai, X. Q., Li, Q., Chen, C. X., Mai, W., Hussain, Z., Long, W., Montalbetti,
N., Li, G., Glynne, R. et al. (2006). Kinesin-2 mediates physical and functionalinteractions between polycystin-2 and fibrocystin. Hum. Mol. Genet. 15, 3280-3292.
Xiao, Y., Lv, X., Cao, G., Bian, G., Duan, J., Ai, J., Sun, H., Li, Q., Yang, Q., Chen, T.
et al. (2010). Overexpression of Trpp5 contributes to cell proliferation and apoptosisprobably through involving calcium homeostasis. Mol. Cell. Biochem. 339, 155-161.
Xiao, Z., Awad, H. A., Liu, S., Mahlios, J., Zhang, S., Guilak, F., Mayo, M. S. andQuarles, L. D. (2005). Selective Runx2-II deficiency leads to low-turnoverosteopenia in adult mice. Dev. Biol. 283, 345-356.
Xiao, Z., Zhang, S., Mahlios, J., Zhou, G., Magenheimer, B. S., Guo, D., Dallas,
S. L., Maser, R., Calvet, J. P., Bonewald, L. et al. (2006). Cilia-like structures andpolycystin-1 in osteoblasts/osteocytes and associated abnormalities in skeletogenesisand Runx2 expression. J. Biol. Chem. 281, 30884-30895.
Xiao, Z., Zhang, S., Magenheimer, B. S., Luo, J. and Quarles, L. D. (2008).Polycystin-1 regulates skeletogenesis through stimulation of the osteoblast-specifictranscription factor RUNX2-II. J. Biol. Chem. 283, 12624-12634.
Journal of Cell Science 125 (8)1956
Journ
alof
Cell
Scie
nce

Xiao, Z., Zhang, S., Cao, L., Qiu, N., David, V. and Quarles, L. D. (2010).
Conditional disruption of Pkd1 in osteoblasts results in osteopenia due to direct
impairment of bone formation. J. Biol. Chem. 285, 1177-1187.
Xiao, Z., Dallas, M., Qiu, N., Nicolella, D., Cao, L., Johnson, M., Bonewald, L. and
Quarles, L. D. (2011). Conditional deletion of Pkd1 in osteocytes disrupts skeletal
mechanosensing in mice. FASEB J. 25, 2418-2432.
Xiao, Z. S., Hjelmeland, A. B. and Quarles, L. D. (2004). Selective deficiency of the
‘‘bone-related’’ Runx2-II unexpectedly preserves osteoblast-mediated skeletogenesis.
J. Biol. Chem. 279, 20307-20313.
Yoder, B. K. (2007). Role of primary cilia in the pathogenesis of polycystic kidney
disease. J. Am. Soc. Nephrol. 18, 1381-1388.
Yoder, B. K., Hou, X. and Guay-Woodford, L. M. (2002a). The polycystic kidney
disease proteins, polycystin-1, polycystin-2, polaris, and cystin, are co-localized in
renal cilia. J. Am. Soc. Nephrol. 13, 2508-2516.
Yoder, B. K., Tousson, A., Millican, L., Wu, J. H., Bugg, C. E., Jr, Schafer, J. A. andBalkovetz, D. F. (2002b). Polaris, a protein disrupted in orpk mutant mice, is requiredfor assembly of renal cilium. Am. J. Physiol. Renal Physiol. 282, F541-F552.
Zhang, M., Xuan, S., Bouxsein, M. L., von Stechow, D., Akeno, N., Faugere, M. C.,
Malluche, H., Zhao, G., Rosen, C. J., Efstratiadis, A. et al. (2002). Osteoblast-specificknockout of the insulin-like growth factor (IGF) receptor gene reveals an essential role ofIGF signaling in bone matrix mineralization. J. Biol. Chem. 277, 44005-44012.
Zhang, Q., Murcia, N. S., Chittenden, L. R., Richards, W. G., Michaud, E. J.,
Woychik, R. P. and Yoder, B. K. (2003). Loss of the Tg737 protein results inskeletal patterning defects. Dev. Dyn. 227, 78-90.
Zhao, M., Ko, S. Y., Liu, J. H., Chen, D., Zhang, J., Wang, B., Harris, S. E.,
Oyajobi, B. O. and Mundy, G. R. (2009). Inhibition of microtubule assembly inosteoblasts stimulates bone morphogenetic protein 2 expression and bone formationthrough transcription factor Gli2. Mol. Cell. Biol. 29, 1291-1305.
Conditional deletion of Kif3a in bone 1957
Journ
alof
Cell
Scie
nce


















