
discovery: Ornithischia Dinosaurs of Great Britain - Darren Naish
12
doi:10.1144/0016-76492007-154 2008; v. 165; p. 613-623 Journal of the Geological Society DARREN NAISH and DAVID M. MARTILL discovery: Ornithischia Dinosaurs of Great Britain and the role of the Geological Society of London in their Journal of the Geological Society service Email alerting to receive free email alerts when new articles cite this article click here request Permission to seek permission to re-use all or part of this article click here Subscribe to subscribe to Journal of the Geological Society or the Lyell Collection click here Notes Downloaded by on 9 April 2008 © 2008 Geological Society of London
Transcript of discovery: Ornithischia Dinosaurs of Great Britain - Darren Naish

doi:10.1144/0016-76492007-154 2008; v. 165; p. 613-623 Journal of the Geological Society
DARREN NAISH and DAVID M. MARTILL
discovery: OrnithischiaDinosaurs of Great Britain and the role of the Geological Society of London in their
Journal of the Geological Society
serviceEmail alerting to receive free email alerts when new articles cite this article click here
requestPermission to seek permission to re-use all or part of this article click here
Subscribe to subscribe to Journal of the Geological Society or the Lyell Collection click here
Notes
Downloaded by on 9 April 2008
© 2008 Geological Society of London

Journal of the Geological Society, London, Vol. 165, 2008, pp. 613–623. Printed in Great Britain.
613
Review
Dinosaurs of Great Britain and the role of the Geological Society of London in
their discovery: Ornithischia
DARREN NAISH & DAVID M. MARTILL
Palaeobiology Research Group, School of Earth and Environmental Sciences, University of Portsmouth,
Portsmouth PO1 3QL, UK (e-mail: [email protected])
Abstract: Completing our survey of British non-avian dinosaurs, we here review the ornithischians of Britain.
Heterodontosaurids are present in the Lower Cretaceous Lulworth Formation of Dorset, and a few earlier
possible records imply a long presence in the region of this clade. Britain’s thyreophoran record is rich and
includes the earliest well-represented taxon, Scelidosaurus, as well as Middle Jurassic stegosaurs and
ankylosaurs including a reasonably good Cretaceous record of polacanthids and nodosaurids. Cretaceous
stegosaurs are known only from fragmentary remains, but the proposal that stegosaurs were present as early as
the Rhaetian is rejected. Among British iguanodontian ornithopods, the possible dryosaurid Callovosaurus is the
oldest global record whereas the proposed synonymy of Cumnoria with Camptosaurus requires confirmation.
Iguanodon has become a taxonomic dumping ground for assorted iguanodontians and is in need of revision:
most of the British species referred to this genus are almost certainly not closely allied to the neotype species
I. bernissartensis and require new generic names. Fragmentary remains suggest the early presence of
hadrosaurids in Britain. The only British record of Marginocephalia, the Wessex Formation skull roof named
Yaverlandia bitholus, appears not to belong to this clade but seems to be from a maniraptoran theropod.
Together with their saurischian relatives, ornithischian dinosaurs
dominated terrestrial life during the Jurassic and Cretaceous
periods. Characterized by an opisthopubic pelvis, a predentary
bone and other distinctive characters, they include the familiar
stegosaurs, ankylosaurs, hadrosaurs, pachycephalosaurs and cer-
atopsians. Their feeding mechanics, evolutionary interactions
with plants, and evolution of armour and diverse cranial
structures have proved highly fertile areas of research. The first
ornithischian to be described was the ornithopod Iguanodon,
based on isolated teeth from the Hastings Beds Group of East
Sussex (Mantell 1825), and the second was the ankylosaur
Hylaeosaurus from the same strata (Mantell 1833). The term
‘ornithischian’, referring to a pelvic girdle that superficially
resembles that of birds, was published by Seeley (1888).
Although ornithischians originated in the Triassic (Butler et al.
2007), their fossils from this time are rare and many alleged
Triassic ornithischians have recently been reinterpreted (Irmis et
al. 2006). The British ornithischian record has recently been
extended to the Upper Triassic (Rhaetian), where two fragmen-
tary bones have been referred to Stegosauria (Galton 2005).
Convincing remains of thyreophorans are well documented from
the Hettangian and Sinemurian of Dorset, where partial skeletons
of Scelidosaurus, sometimes with skin preserved, have been
known since the 1860s (Owen 1861). Isolated elements and rare
partial skeletons of ornithischians have been reported from
marine strata in the Bathonian, Callovian, Oxfordian and Kim-
meridgian of central and eastern England. However, the most
substantial remains come from the Lower Cretaceous Hastings
Beds, Weald Clay and Wealden groups, where near-complete and
partial skeletons of the ankylosaurs Hylaeosaurus and Pola-
canthus and the ornithopods Hypsilophodon, Valdosaurus, Man-
tellisaurus and Iguanodon have been recovered. The youngest
non-avian British dinosaurs include ornithischian taxa from the
Albian–Cenomanian Lower Chalk, Folkestone Beds, Gault Clay
and Cambridge Greensand: among them are highly fragmentary
ankylosaurs and hadrosaurids.
In a previous paper (Naish & Martill 2007), we reviewed the
fossil record, taxonomy and phylogenetic relationships of the
saurischian dinosaurs from the British Mesozoic. Here, we
review British ornithischian dinosaurs. As discussed previously
(Naish & Martill 2007, p. 496), many names coined for British
dinosaur taxa are no longer considered adequate in terms of
establishing taxonomic validity (supplementary information is
available online at http://www.geolsoc.org.uk/SUP18267), mean-
ing that approximately 54% of named British dinosaur taxa are
nomina dubia. In view of inconsistencies in the literature, we
have used the original spellings for specific names, although this
results in an apparent inconsistency within our own text. A
number of British ornithischians have been incorrectly allocated
to a genus (e.g. most species referred to Iguanodon) and, in most
cases, it is the type species alone that should be regarded as
being correctly associated with the generic name. All other
designations to generic level should be assumed to be historical
artefacts and, following convention, we use quotation marks.
Heterodontosauridae
Heterodontosaurids were small, gracile ornithischians, best
known for the caniniform teeth and elongate hands present in
most genera. Conventionally regarded as ornithopods (Sereno
1986; Norman et al. 2004), they may be basal ornithischians
outside of Genasauria (Butler et al. 2008) or the sister-taxon to
Marginocephalia (Xu et al. 2006). Most heterodontosaurids are
from the Lower Jurassic of South Africa but Echinodon becklesii

Owen, 1861, probably from the Berriasian Warbarrow Tout
Member of the Lulworth Formation of Durlston Bay, Dorset,
appears to belong to this group (Norman & Barrett 2002) (Fig.
1). Teeth like those of E. becklesii are known from the Bathonian
Forest Marble of Kirtlington and the Tithonian Portland Stone of
Wiltshire (Norman & Barrett 2002). Small, conical osteoderms
from the Purbeck Limestone Group, dubbed ‘granicones’, were
considered referable to E. becklesii when this taxon was regarded
as a possible thyreophoran, but are now known to be osteoderms
from solemydid turtles (Barrett et al. 2002). Heterodontosaurids
have recently been reported from the Lower Cretaceous Camar-
illas Formation of Spain (Sanchez-Hernandez et al. 2007) and
this group may prove to have been more widespread in the Early
Cretaceous in Europe than realized so far.
Thyreophora
Thyreophora is the armoured ornithischian clade: in addition to
the familiar ankylosaurs and stegosaurs it includes several basal
forms from the Lower Jurassic. Conventionally regarded as one
of these basal forms, Scelidosaurus harrisonii Owen, 1861 is one
of Britain’s most historically significant ornithischians. First
described from an incomplete hindlimb and an associated skull
and skeleton from a different individual (Owen 1861), it is from
the Sinemurian Charmouth Mudstone Formation of Charmouth,
Dorset. The hindlimb on which Owen (1861) based the generic
name proved to belong to a tetanuran theropod (Newman 1968)
and, accordingly, Charig & Newman (1992) proposed that the
skull and skeleton be regarded as the lectotype. Owen made little
of the fact that S. harrisonii provided support for his view of
dinosaurs as comparatively heavy-limbed quadrupeds (Norman
2000, 2001).
Owen (1861) also described other elements (dorsal centrum,
phalanges and right hind limb bones) as belonging to a juvenile
S. harrisonii. This identification was doubted by Newman
(1968), who proposed that these were from a ‘hypsilophodontid’.
Another specimen suggested to be a juvenile S. harrisonii (Rixon
1968; Charig 1972), BMNH R6704, was thought by Thulborn
(1974, 1977) to represent a different taxon, probably allied to
basal ornithischians such as Lesothosaurus. Coombs et al. (1990)
Fig. 1. (a) Dentition of Echinodon becklesii paralectotype specimen BMNH 48215b in lingual view. Scale bar represents 10 mm. (b) Skeletal
reconstruction of the Lower Jurassic heterodontosaurid Heterodontosaurus tucki. Echinodon would have been similar overall to this taxon, although
different in its details. Diagram by kind courtesy of J. Headden. Scale bar represents 10 cm.
D. NAISH & D. M. MARTILL614

also regarded it as a distinct taxon, albeit a thyreophoran.
However, Galton (1975) reaffirmed the identification of both
controversial specimens as S. harrisonii.
Some unpublished S. harrisonii individuals sport curving
squamosal horns and thorn-like spikes running in rows along the
limbs (Naish & Martill 2007, fig. 1). Possibly this is evidence of
sexual dimorphism. The concept of S. harrisonii as a basal
thyreophoran was challenged by Carpenter (2001), who argued
that character evidence better places S. harrisonii as the sister-
taxon to Ankylosauria (the S. harrisonii + Ankylosauria clade
being termed Ankylosauromorpha), but this has not been
supported in a recent global analysis of ornithischian phylogeny
(Butler et al. 2008). Reports of S. harrisonii from the Rhaetian
Westbury Formation of Shepton Mallet, Somerset (Storrs 1994),
Lower Jurassic Kayenta Formation of Arizona, USA (Padian
1989) and Lufeng Formation of Yunnan, China (Lucas 1996) are
probably erroneous (Norman et al. 2007). A detailed, modern
description of S. harrisonii is still lacking.
Following S. harrisonii, thyreophorans are unknown from
Britain for perhaps 20 Ma or more. A few fossils might help fill
this gap, however. Clark (2001) described a thyreophoran from the
Bajocian of the Isle of Skye, known only from a proximal radius
and ulna. The rugose, inflated olecranon process suggests an
ankylosaurian identity more than a stegosaurian one. Hoffstetter
(1957) suggested that two armour plates from the Bathonian
Chipping Norton Formation of Stow-on-the-Wold, Gloucester-
shire, might be ankylosaurian, but Galton (1985) referred these to
the stegosaur Lexovisaurus.
Ankylosauria
Indisputable ankylosaurs do not appear in the British fossil
record until the Callovian. An incomplete lower jaw from the
Peterbrough Member of the Oxford Clay Formation of Fletton,
Cambridgeshire, was named Sarcolestes leedsi Lydekker, 1893.
In its rostrally continuous tooth row and proportionally small
predentary S. leedsi resembles nodosaurids (Galton 1983a) and it
was regarded as the sister-taxon to Nodosaurinae by Sereno
(1998). However, Vickaryous et al. (2004) proposed that S. leedsi
should be regarded as Ankylosauria incertae sedis. This poses a
problem for Sereno’s (1998) phylogenetic definition of Nodo-
saurinae, as S. leedsi was employed as a specifier. Three isolated
osteoderms from the Oxford Clay Formation were suggested by
Galton (1994) to belong to S. leedsi.
A younger ankylosaur, Cryptosaurus eumerus Seeley, 1869, is
based on a right femur from the Ampthill Clay of Great
Gransden, Cambridgeshire. A dorsal vertebra from the Oxford
Clay of Weymouth referred to the taxon was reidentified as
probably belonging to a theropod (Galton 1983b). Although
Lydekker (1889) argued that Cryptosaurus was preoccupied
(prompting the replacement name Cryptodraco Lydekker), he
was incorrect (B. Creisler, pers. comm.). Galton (1980a, 1983b)
regarded the femur as most like that of Hoplitosaurus, a North
American ankylosaur regarded as a polacanthid by those workers
who recognize this group (Kirkland 1998a; Carpenter 2001) and,
on the basis of the prominent trochanters, proposed that Crypto-
saurus may have been facultatively bipedal (Galton 1983b). The
latter idea has not been accepted by other workers. Cryptosaurus
lacks autapomorphies and should be regarded as Ankylosauria
indet.
Priodontognathus phillipsii (Seeley, 1869), known only from a
maxilla and originally described as Iguanodon phillipsii, was
reinterpreted by Galton (1980b) as a nodosaurid. The specimen
exhibits unusual teeth combining denticulate cingula and bowl-
like lingual depressions (Barrett 2001), and its recent consign-
ment to the status of nomen dubium (Vickaryous et al. 2004) is
probably unjust. It has yet to be determined whether Priodontog-
nathus is from the Upper Jurassic of Yorkshire or the Lower
Cretaceous of Sussex.
Far better represented are the Wealden ankylosaurs Hylaeo-
saurus armatus Mantell, 1833 and Polacanthus Owen in Anon.,
1865. Though once considered synonymous, they are clearly
distinct. Hylaeosaurus is from the Grinstead Clay Formation of
East Sussex (Mantell 1833) with a referred specimen from West
Sussex (Mantell 1841). The holotype consists of an articulated
anterior portion of a skeleton, still with cervical and scapular
armour in position, and possibly incorporating cranial material
(Mantell 1833; Pereda-Suberbiola 1993). Mechanical and acid
preparation has been used on this specimen, and more informa-
tion will be forthcoming.
Polacanthus is represented by two species. P. foxii Owen in
Anon., 1865 was described for a partial skeleton from the
Wessex Formation whereas P. rudgwickensis Blows, 1996 is from
the Barremian Upper Weald Clay Formation of Rudgwick, West
Sussex (Hulke 1881; Blows 1987, 1996; Pereda-Suberbiola 1993,
1994). Cranial material referred to P. foxii includes a partial
neurocranium, a possible angular, and a tooth (Blows 1987;
Norman & Faiers 1996; Naish & Martill 2001a). The characters
that differentiate P. foxii from P. rudgwickensis are not entirely
satisfactory, and unpublished specimens from the Isle of Wight
and East Sussex are intermediate between the two. In addition to
its presence in the Wessex Formation, P. foxii has been reported
from the Vectis Formation and from a Lower Greensand Forma-
tion exposure between Humble Point and Pinhay Bay, Devon
(Blows 1996). The latter record, initially reported as being from
Charmouth, Dorset, lacks characters that allow its identification
beyond Ankylosauria. Specimens from Spain and the USA have
been referred to Polacanthus, and, in addition to its occurrence
in the Weald, Hylaeosaurus has been reported from Germany,
France and the Isle of Wight, although all of these additional
records are probably erroneous (Naish & Martill 2001a). The
affinities of Hylaeosaurus and Polacanthus remain uncertain:
although often regarded as basal nodosaurids, both have been
regarded as part of a Polacanthidae clade (Kirkland 1998a;
Carpenter 2001).
Numerous ankylosaur remains from the Albian–Cenomanian
Lower Chalk, Folkestone Beds, Gault Clay and Cambridge
Greensand, described as Anoplosaurus major Seeley, 1879 and
five species of the supposed nodosaurid Acanthopholis Huxley,
1867, were reviewed by Pereda Suberbiola & Barrett (1999) and
shown to be based on non-diagnostic ankylosaurian material,
some of which had been combined with ornithopod and sauropod
material. Only one ankylosaur from these units, Anoplosaurus
curtonotus Seeley, 1879 from the Upper Gault Clay or Cam-
bridge Greensand near Reach (Cambridgeshire), is diagnosable
(Pereda Suberbiola & Barrett 1999). In its low number of sacral
vertebrae, rostral extent of its dentary tooth row and other
features, Anoplosaurus appears to be a basal nodosaurid. The
absence of dermal armour in the holotype probably reflects
juvenile status.
Stegosauria
Britain has a reasonably good stegosaur record that includes
some of the geologically oldest and youngest specimens in the
world, as well as the first stegosaur to be recognized historically.
The latter, a partial lower jaw named Regnosaurus northamptoni
Mantell, 1848 from the Berriasian–Valanginian Hastings Beds
BRITISH DINOSAURS 615

Group (the exact horizon is unknown, but the specimen was most
probably from the Grinstead Clay Formation) of Cuckfield, East
Sussex, was originally referred to Iguanodon and not recognized
as stegosaurian until recently (Olshevsky 1993; Barrett &
Upchurch 1995). Unlike stegosaurid stegosaurs (Barrett &
Upchurch 1995; Maidment et al. 2006), Regnosaurus lacks a
lateral lamina obscuring the dentary tooth row from view, and
shares two detailed features of the dentary with the most basal
stegosaur, Huayangosaurus from the Middle Jurassic of China.
Although this suggests that archaic stegosaurs survived into the
Early Cretaceous of Europe (Fig. 2), we should be cautious in
basing so much on lower jaw characters alone, given the paucity
of stegosaur cranial material. Most recently, Regnosaurus has
been regarded as a nomen dubium unclassifiable beyond Stego-
sauria (Galton & Upchurch 2004). Isolated elements from the
Hastings Beds Group, Weald Clay Group and Wessex Formation,
including an incomplete vertebra, armour plates and a pubis,
have been suggested to be stegosaurian (Galton 1981; Blows
2001; Naish & Martill 2001a) although not without controversy
or later reidentification (Galton 1985). Craterosaurus pottonensis
Seeley, 1874, from the Aptian Potton Sands of Potton, Bed-
fordshire, might be reworked from the Hastings Beds Group and
thus synonymous with Regnosaurus. A depression between the
prezygapophyses appears unique to Craterosaurus and it has
been regarded as a valid stegosaurid (Galton 1981; Galton &
Upchurch 2004).
Britain’s oldest undoubted stegosaurs are among the oldest
stegosaur body fossils in the world: they comprise teeth and
postcranial elements from the Bathonian Sharp’s Hill Formation
of Oxfordshire and Chipping Norton Formation of Gloucester-
shire (Galton & Powell 1983; Metcalf & Walker 1994; Benton &
Spencer 1995). Possible stegosaur spikes have been reported
from the Aalenian–Bajocian Inferior Oolite Formation of Brad-
ford Abbas, Dorset (Benton & Spencer 1995).
Galton (2005) argued that two partial columnar bone segments
from the Rhaetian Westbury Formation of Aust Cliff, Avon,
represent femora of Late Triassic stegosaurs. Interpreted as
femora, these bones would have been c. 1 m long when complete
and would have belonged to animals comparable in size with
Stegosaurus (total length 7–9 m). The columnar bone shape and
presence of a thin cortex and of extensive cancellous bone and/or
trabeculae were used to support a stegosaurian identity (Galton
2005). The idea that giant stegosaurs were present as early as the
Late Triassic is surprising and based on scant evidence. Butler et
al. (2006) and Norman et al. (2007) argued that these specimens
cannot be identified beyond Reptilia indet. Irmis et al. (2006)
argued that the specimens lacked features allowing identification,
that the details on internal structure employed by Galton are of
no use in taxonomic identification, and that the specimens should
be identified as Tetrapoda indet. We agree with these reapprai-
sals.
The best known British stegosaurs are the basal stegosaurid
Dacentrurus armatus (Owen, 1875) from the Kimmeridge Clay
Formation of Swindon and the stegosaurine stegosaurid Lexovi-
saurus durobrivensis (Hulke, 1887) from the Callovian Oxford
Clay Formation of Fletton. Both species were originally de-
scribed as belonging to Omosaurus Owen, 1875, a name
preoccupied by a North American phytosaur. To rectify this,
Dacentrurus Lucas, 1902 was coined for ‘O.’ armatus, and
Lexovisaurus Hoffstetter, 1957 erected for ‘O.’ durobrivensis.
Much material from Dorset, Wiltshire, Oxfordshire, Northamp-
tonshire and Cambridgeshire has been referred to these taxa
(Galton & Powell 1983; Galton 1985) and several other names
are regarded as probable junior synonyms. Among these is
‘Omosaurus’ phillipsi Seeley, 1893, based on a femur from the
Oxfordian Corallian Oolite Formation of Slingsby, North York-
shire (Galton 1983c), and deliberately given the same specific
name as Priodontognathus phillipsii (although note that Seeley
Fig. 2. Skeletal reconstructions of selected
British stegosaur taxa portrayed in their
phylogenetic context. (a) Regnosaurus
northamptoni, a possible basal stegosaur
(based only on a partial lower jaw).
(b) Dacentrurus armatus, a basal
stegosaurid. (c) Lexovisaurus durobrivensis,
a stegosaurine stegosaurid. Not to scale.
Diagrams courtesy of T. L. Ford.
D. NAISH & D. M. MARTILL616

(1893) spelt the specific name differently) because Seeley
thought it would prove synonymous with it! D. armatus was a
large stegosaur exceeding 7 m in total length: the femur of the
holotype is over 1200 mm long (Galton & Boine 1980) and
therefore similar in size to that of Stegosaurus.
Ornithopoda
The name Ornithopoda is today restricted to the clade that
incorporates the hadrosaurid Edmontosaurus and all taxa closer
to it than to the marginocephalian Triceratops (Norman et al.
2004). Basal ornithopods were small, bipedal cursors but larger
size and more robust proportions were evolved by members of
the clade Iguanodontia.
Fragmentary remains indicate the presence of basal ornitho-
pods within the British fossil record. A premaxillary tooth from
Stonesfield was identified as that of a basal ornithopod (Galton
1975), and a dentary tooth from the Kimmeridge Clay of
Weymouth, accessioned to the UCMP (as UCMP 46911), was
identified as cf. Bugenasaura (Galton 1999). Even allowing for
phylogenies indicating that basal ornithopods such as Bugen-
asaura originated in the Middle Jurassic (Weishampel et al.
2003), this is surprising given that Bugenasaura is a Maastrich-
tian North American taxon. The provenance of this specimen is
dubious, and it is likely that it did not originate in Dorset (Martill
et al. 2006).
The best known British ornithopod (after Iguanodon) is
Hypsilophodon foxii Huxley, 1869, one of the first small
ornithischians to be recognized. First identified as a juvenile
Iguanodon, Huxley (1869, 1870) showed it to be a distinct taxon.
Numerous specimens are known from the Hypsilophodon Bed
near the top of the Wessex Formation, as well as from the Vectis
Formation (Galton 1969, 1974, 1989). Reports of Hypsilophodon
sp. from East Sussex (Weishampel 1990) are erroneous. Mis-
interpreted by some 19th and 20th century workers as scansorial,
H. foxii was actually a terrestrial cursor (Galton 1971a, b, 1974).
Ornithischians similar to H. foxii persisted throughout most of
the Mesozoic and, until recently, were all united in a ‘family’
Hypsilophodontidae (e.g. Sereno 1986; Weishampel & Heinrich
1992). Phylogenetic studies indicate that this grouping is artifi-
cial (Weishampel et al. 2003; Norman et al. 2004; Butler et al.
2008).
Basal members of the ornithopod clade Iguanodontia were
superficially similar to H. foxii and were long classified with
them in ‘Hypsilophodontidae’. Among these are the dryosaurids:
gracile cursors with narrow feet lacking a hallux. Valdosaurus
canaliculatus (Galton 1975) from the Wessex Formation is
similar to Dryosaurus and was initially named as a species of
this genus (Galton 1975). V. canaliculatus was named for femora
but numerous other elements have been referred to it (Galton &
Taquet 1982; Barrett 1996; Blows 1998; Naish & Martill 2001b).
Although the Wessex Formation hindlimb elements can be
reliably referred to this species, the same cannot be said for the
others. Some of the referred material is from the Hastings Beds
Group of Heathfield and Cuckfield in East Sussex and its referral
to V. canaliculatus is questionable. Barrett (1996) suggested that
‘Camptosaurus’ valdensis Lydekker, 1889, based on an incom-
plete left femur presumed to be from the Wessex Formation,
might be a senior synonym of V. canaliculatus.
An alleged dryosaurid tibia has been reported from the
Callovian Oxford Clay of Fletton, Cambridgeshire (Galton
1977), but it cannot be reliably allocated to Dryosauridae and
should be regarded as Ornithopoda indet. A left femur, also from
the Oxford Clay of Fletton, was described as Camptosaurus
leedsi Lydekker, 1889 and, although suggested at times to be part
of Dryosauridae (Gilmore 1909; Galton 1974), was given its own
genus, Callovosaurus Galton, 1980, in Camptosauridae. Signifi-
cant in being the earliest reported iguanodontian, it has most
recently been argued to be a valid dryosaurid (Ruiz-Omenaca et
al. 2007).
Better represented is the partial camptosaurid from the
Kimmeridge Clay of Cumnor, Oxfordshire, named Iguanodon
prestwichii Hulke, 1880 but later awarded the generic name
Cumnoria Seeley, 1888. Although it has been argued that this
taxon is referable to Camptosaurus (Lydekker 1889; Galton
1980c; Galton & Powell 1980), derived characters shared by the
Kimmeridge Clay taxon and Camptosaurus have yet to be
identified and the characters used to allocate it to Camptosaurus
are unconvincing, consisting almost entirely of plesiomorphies.
Furthermore, some of the similarities that have been used to
support referral of Cumnoria to Camptosaurus (e.g. small
antorbital fenestra) are now known not to occur in the latter
taxon: the cranial material typically illustrated for Camptosaurus
actually belongs to another iguanodontian, Theiophytalia (Brill
& Carpenter 2007).
Iguanodon
Iguanodon Mantell, 1825 was first described from teeth from the
Hastings Beds Group of Cuckfield (Mantell 1825) but is today
regarded as ranging from Berriasian to Albian, and to contain six
British species (Norman 2004). Although the species-level
taxonomy of Iguanodon is regarded as non-controversial, it
requires revision. There has been considerable confusion as to
which unit yielded Mantell’s original material: Topley (1875)
stated that the ‘Tilgate Grit’ that yielded Mantell’s original
specimens was part of the Wadhurst Clay Formation, Norman
(1987) regarded the Upper Tunbridge Wells Sand Formation as
the source of Mantell’s specimens, whereas Blows (1998)
claimed that the fossils were collected from the Grinstead Clay
Formation. Although Mantell acquired specimens from various
horizons of the Hastings Beds Group, Mantell’s ‘Tilgate Grit’
was part of the Grinstead Clay Formation (Radley 2004).
Mantell’s type material of Iguanodon was later named Iguano-
don anglicus Holl, 1829 (originally I. anglicum). Unfortunately,
it is non-diagnostic so, to save the generic name, Charig &
Chapman (1998) proposed that the Belgian species Iguanodon
bernissartensis be made the new type species, and the ICZN
ruled in favour of this proposal. Although I. bernissartensis is
the earliest named valid species referred to the genus, and the
one best associated with it, it is substantially different from other
species referred to Iguanodon, and from the Hastings Beds
Group material.
The oldest species referred to Iguanodon, ‘I.’ hoggii Owen,
1874 from the Berriasian Lulworth Formation of Swanage,
Dorset, was argued by Norman & Barrett (2002) to belong to
Camptosaurus and was renamed C. hoggii. Other elements from
Dorset have been referred to C. hoggii, as have bones from
Buckinghamshire and from the Berriasian Speeton Clay Forma-
tion of Yorkshire (Norman & Barrett 2002).
In addition to I. anglicus, three species from the Hastings Beds
Group have been referred to Iguanodon. Older than I. bernissar-
tensis and more archaic, it is doubtful that they are congeneric
with this species (Lydekker 1888). Vertebrae and pelvic and
hindlimb bones, representing a large species from the Wadhurst
Clay Formation of Hastings, were named ‘I.’ dawsoni Lydekker,
1888. Several additional specimens were referred by Blows
(1998) to ‘I.’ dawsoni, with the best of them being BMNH
BRITISH DINOSAURS 617
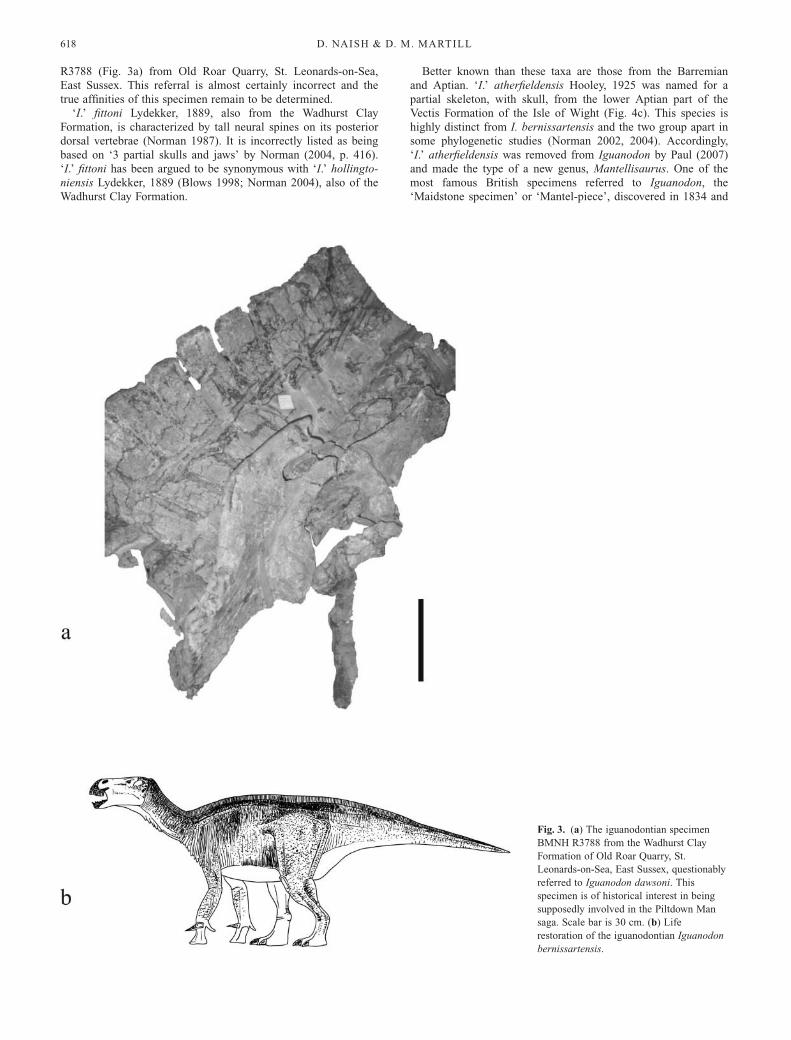
R3788 (Fig. 3a) from Old Roar Quarry, St. Leonards-on-Sea,
East Sussex. This referral is almost certainly incorrect and the
true affinities of this specimen remain to be determined.
‘I.’ fittoni Lydekker, 1889, also from the Wadhurst Clay
Formation, is characterized by tall neural spines on its posterior
dorsal vertebrae (Norman 1987). It is incorrectly listed as being
based on ‘3 partial skulls and jaws’ by Norman (2004, p. 416).
‘I.’ fittoni has been argued to be synonymous with ‘I.’ hollingto-
niensis Lydekker, 1889 (Blows 1998; Norman 2004), also of the
Wadhurst Clay Formation.
Better known than these taxa are those from the Barremian
and Aptian. ‘I.’ atherfieldensis Hooley, 1925 was named for a
partial skeleton, with skull, from the lower Aptian part of the
Vectis Formation of the Isle of Wight (Fig. 4c). This species is
highly distinct from I. bernissartensis and the two group apart in
some phylogenetic studies (Norman 2002, 2004). Accordingly,
‘I.’ atherfieldensis was removed from Iguanodon by Paul (2007)
and made the type of a new genus, Mantellisaurus. One of the
most famous British specimens referred to Iguanodon, the
‘Maidstone specimen’ or ‘Mantel-piece’, discovered in 1834 and
Fig. 3. (a) The iguanodontian specimen
BMNH R3788 from the Wadhurst Clay
Formation of Old Roar Quarry, St.
Leonards-on-Sea, East Sussex, questionably
referred to Iguanodon dawsoni. This
specimen is of historical interest in being
supposedly involved in the Piltdown Man
saga. Scale bar is 30 cm. (b) Life
restoration of the iguanodontian Iguanodon
bernissartensis.
D. NAISH & D. M. MARTILL618
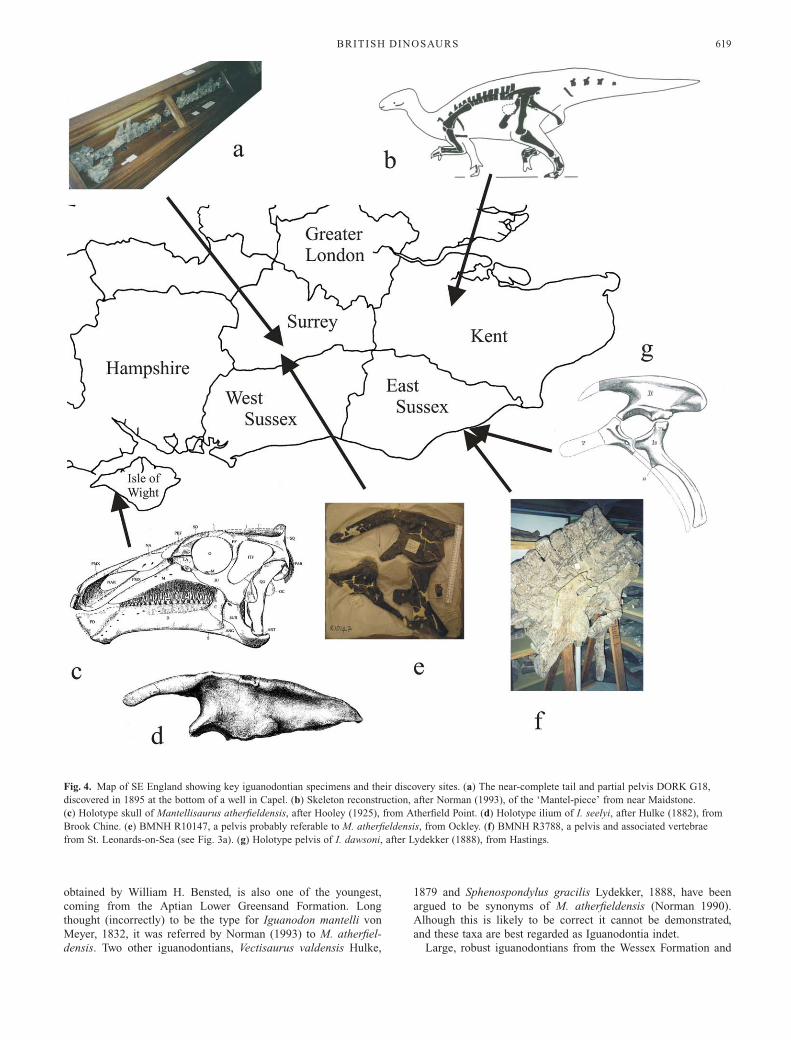
obtained by William H. Bensted, is also one of the youngest,
coming from the Aptian Lower Greensand Formation. Long
thought (incorrectly) to be the type for Iguanodon mantelli von
Meyer, 1832, it was referred by Norman (1993) to M. atherfiel-
densis. Two other iguanodontians, Vectisaurus valdensis Hulke,
1879 and Sphenospondylus gracilis Lydekker, 1888, have been
argued to be synonyms of M. atherfieldensis (Norman 1990).
Alhough this is likely to be correct it cannot be demonstrated,
and these taxa are best regarded as Iguanodontia indet.
Large, robust iguanodontians from the Wessex Formation and
Fig. 4. Map of SE England showing key iguanodontian specimens and their discovery sites. (a) The near-complete tail and partial pelvis DORK G18,
discovered in 1895 at the bottom of a well in Capel. (b) Skeleton reconstruction, after Norman (1993), of the ‘Mantel-piece’ from near Maidstone.
(c) Holotype skull of Mantellisaurus atherfieldensis, after Hooley (1925), from Atherfield Point. (d) Holotype ilium of I. seelyi, after Hulke (1882), from
Brook Chine. (e) BMNH R10147, a pelvis probably referable to M. atherfieldensis, from Ockley. (f) BMNH R3788, a pelvis and associated vertebrae
from St. Leonards-on-Sea (see Fig. 3a). (g) Holotype pelvis of I. dawsoni, after Lydekker (1888), from Hastings.
BRITISH DINOSAURS 619

Barremian Upper Weald Clay Formation have been referred to
I. bernissartensis (Naish & Martill 2001b; Norman 2004) and are
similar to Belgian specimens. However, I. seelyi Hulke, 1882,
named after Charles Seely (and not H. G. Seeley), was coined
for notably robust elements from a large Isle of Wight iguano-
dontian (Fig. 4d). Although it has been assumed that I. seelyi is a
junior synonym of I. bernissartensis, the two differ in the form
of the ilium (Hulke 1882) and this synonymy should be regarded
as provisional. Largely overlooked has been the conditional
proposal by Delair (1966) that Weald Clay material named
Streptospondylus major Owen, 1842 should be recognized as a
new iguanodontian taxon, I. major. Delair (1966) speculated that
I. major differed in enough subtle details from I. bernissartensis
to be considered distinct.
It seems that Iguanodon is the ornithopod version of Cetio-
saurus and Megalosaurus: a taxonomic dumping ground for any
iguanodontian remains that are not clearly referable to any other
named taxon. The status and phylogenetic relationships of many
of the referred species and specimens require further study.
Possible hadrosaurids
‘Iguanodon’ hillii Newton, 1892, the youngest British species
referred to this genus, has been suggested to be a hadrosaurid
(Horner et al. 2004) but is better regarded as Iguanodontia indet.
Based on only a partial maxillary tooth, it was thought by Head
(1998) to be indistinguishable from the basal iguanodontian
condition, but this is probably incorrect as the tooth is highly
derived. A second proposed British hadrosaurid is ‘Trachodon’
cantabrigiensis (not cantabrigensis, as spelt by Newton (1892))
Lydekker, 1888, a nomen dubium based on a single tooth from
the Cambridge Greensand. Although ‘T.’ cantabrigiensis is
hadrosaurid-like and has been regarded as a member of Hadro-
sauridae (Benton & Spencer 1995; Kirkland 1998b; Horner et al.
2004), Head (1998) argued that the absence of further British
hadrosaurid material, the variation observed within the tooth
morphology of iguanodontian taxa, and the geographical isola-
tion of the British archipelago from the Aptian on rendered this
identification tenuous. Further remains are needed for the
presence of hadrosaurids in Britain to be verified, and for now
‘T.’ cantabrigiensis remains a tantalizing enigma.
Marginocephalia
Marginocephalians (the pachycephalosaurs and ceratopsians) are
restricted to North America and Asia, with only a few dubious
records from elsewhere. Yaverlandia bitholus Galton, 1971, from
the Wessex Formation of the Isle of Wight and based on an
incomplete skull roof, has been regarded as a pachycephalosaur
(Galton 1971c). In lacking characters present in all definite
pachycephalosaurs it has been interpreted by some as one of the
most basal members of the group (Wall & Galton 1979; Sues
1980; Galton & Taquet 1982; Williamson & Carr 2002). How-
ever, the features that have led to its identification as a
pachycephalosaur (domed frontals, interfrontal fusion) are pre-
sent only in derived, dome-skulled members of the clade, and
consequently some workers have regarded it as a derived form
(Sues & Galton 1987; Maryanska 1990; Sereno 2000; Naish &
Martill 2001c). Furthermore, Yaverlandia differs in numerous
detailed features from definite pachycephalosaurs, leading several
workers to doubts its inclusion within the group (Giffin 1989;
Sullivan 2000, 2003, 2005). Restudy by Naish (in preparation),
incorporating computerized tomography scanning, has shown
that Yaverlandia appears, surprisingly, to be a maniraptoran
theropod. This reidentification expunges marginocephalians from
the British fossil record.
Discussion
This contribution concludes our review of the British dinosaur
assemblage (see Naish & Martill 2007). Although it is clear that
Britain is home to a rich and diverse dinosaur assemblage, we
also note again the huge contribution to the subject made by the
Geological Society of London. The well-known and well-repre-
sented British ornithischians Lexovisaurus durobrivensis, Hypsi-
lophodon foxii, Cumnoria prestwichii and Mantellisaurus
atherfieldensis were all described in the Quarterly Journal of the
Geological Society of London, and, among the more obscure
forms, Sarcolestes leedsi, Anoplosaurus curtonotus, Cratero-
saurus pottonensis and ‘Iguanodon’ dawsoni (and others) were
too.
The absence from the British Upper Triassic and Lower
Jurassic of basal ornithischians resembling Lesothosaurus,
Eocursor and members of Heterodontosauridae suggests that
Britain was not important in the early history of the clade.
However, the presence of the relict heterodontosaurid Echinodon
in Berriasian strata of Dorset shows that this mostly Early
Jurassic group survived into the Early Cretaceous. If teeth from
the Bathonian of Kirtlington are correctly identified as those of
heterodontosaurids (Norman & Barrett 2002), it would appear
that British members of the clade were not Cretaceous novelties
that invaded the region from elsewhere, but were in fact present
here throughout the Jurassic.
Britain’s oldest reported ornithischian is the thyreophoran
Scelidosaurus, one of the first dinosaurs to be described from
good, associated remains. Excellent articulated specimens mean
that this taxon has the potential to be one of the most thoroughly
described and best understood of all thyreophorans, but a
comprehensive modern analysis is still lacking. The British
record of ankylosaurs and stegosaurs is reasonably good, with a
handful of taxa, including Polacanthus, Hylaeosaurus and
Dacentrurus, represented by articulated remains.
Britain’s record of iguanodontians is particularly rich. Callovo-
saurus leedsi is the oldest global record of this clade. If correctly
identified by Ruiz-Omenaca et al. (2007) as a dryosaurid, the
Callovian age of Callovosaurus means that the ghost lineages of
many ornithopod taxa (including Hypsilophodon, Rhabdodonti-
dae and Tenontosaurus) are even longer than previously proposed
(Weishampel et al. 2003). The systematics and phylogenetic
relationships of species and specimens referred to Iguanodon
would appear to be a fertile area of further research, with
surprisingly few specimens having been adequately described. It
remains to be determined with certainty that robust iguanodon-
tians from the Wealden Group and/or Weald Clay Group are
referable to Iguanodon bernissartensis, the type species of the
genus, and several species referred to Iguanodon are almost
certainly not congeneric with I. bernissartensis.
Finally, marginocephalians (until recently thought present in
Britain because of Yaverlandia bitholus from the Wessex Forma-
tion) are absent from the British record.
For data, discussion and opinion, we thank P. Barrett, R. Butler, P. Galton,
J. Kirkland, M. Simpson, G. Paul and R. Sullivan. We thank M. Benton
and E. Buffetaut for their reviews of the manuscript. We thank
J. Headden for the heterodontosaurid reconstruction, T. Ford for the
stegosaur reconstructions, and J. Owen (Natural History Museum,
London; NHM) for use of images of NHM specimens. For access to
specimens we thank S. Chapman and A. Milner (NHM), S. Hutt and
D. NAISH & D. M. MARTILL620

M. Munt (Isle of Wight County Museum Service) and J. Cooper (Booth
Museum of Natural History, Brighton). We thank R. Strachan and A.
Hills for editorial support.
References
Barrett, P.M. 1996. The first known femur of Hylaeosaurus armatus and re-
identification of ornithopod material in The Natural History Museum,
London. Bulletin of the Natural History Museum (Geology), 52, 115–118.
Barrett, P.M. 2001. Tooth wear and possible jaw action of Scelidosaurus
harrisonii Owen and a review of feeding mechanisms in other thyreophoran
dinosaurs. In: Carpenter, K. (ed.) The Armored Dinosaurs. Indiana
University Press, Bloomington, 25–52.
Barrett, P.M. & Upchurch, P. 1995. Regnosaurus northamptoni, a stegosaurian
dinosaur from the Lower Cretaceous of southern England. Geological
Magazine, 132, 213–222.
Barrett, P.M., Clarke, J.B., Brinkman, D.B., Chapman, S.D. & Ensom, P.C.
2002. Morphology, histology and identification of the ‘granicones’ from the
Purbeck Limestone Formation (Lower Cretaceous: Berriasian) of Dorset,
southern England. Cretaceous Research, 23, 279–295.
Benton, M.J. & Spencer, P.S. 1995. Fossil Reptiles of Great Britain. Chapman
and Hall, London.
Blows, W.T. 1987. The armoured dinosaur Polacanthus foxi from the Lower
Cretaceous of the Isle of Wight. Palaeontology, 30, 557–580.
Blows, W.T. 1996. A new species of Polacanthus (Ornithischia; Ankylosauria)
from the Lower Cretaceous of Sussex, England. Geological Magazine, 133,
671–682.
Blows, W.T. 1998. A review of Lower and Middle Cretaceous dinosaurs of
England. Bulletin of the New Mexico Museum of Natural History, 14, 29–38.
Blows, W.T. 2001. Possible stegosaur dermal armor from the Lower Cretaceous of
southern England. In: Carpenter, K. (ed.) The Armored Dinosaurs. Indiana
University Press, Bloomington, 130–140.
Brill, K. & Carpenter, K. 2007. A description of a new ornithopod from the
Lytle Member of the Purgatoire Formation (Lower Cretaceous) and a
reassessment of the skull of Camptosaurus. In: Carpenter, K. (ed.) Horns
and Beaks: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press,
Bloomington, 49–67.
Butler, R.J., Porro, L.B. & Heckert, A.B. 2006. A supposed heterodontosaurid
tooth from the Rhaetian of Switzerland and a reassessment of the European
Late Triassic record of Ornithischia (Dinosauria). Neues Jahrbuch fur
Geologie und Palaontologie, Monatshefte, 2006, 613–633.
Butler, R.J., Smith, R.M.H. & Norman, D.B. 2007. A primitive ornithischian
dinosaur from the Late Triassic of South Africa, and the early evolution and
diversification of Ornithischia. Proceedings of the Royal Society of London,
Series B, 274, 2041–2046.
Butler, R.J., Upchurch, P. & Norman, D.B. 2008. The phylogeny of the
ornithischian dinosaurs. Journal of Systematic Palaeontology, 6, 1–40.
Carpenter, K. 2001. Phylogenetic analysis of the Ankylosauria. In: Carpenter,
K. (ed.) The Armored Dinosaurs. Indiana University Press, Bloomington,
455–483.
Charig, A.J. 1972. The evolution of the archosaur pelvis and hind-limb: an
explanation in functional terms. In: Joysey, K.A. & Kemp, T.S. (eds) Studies
in Vertebrate Evolution. Oliver & Boyd, Edinburgh, 121–155.
Charig, A.J. & Chapman, S.D. 1998. Iguanodon Mantell, 1825 (Reptilia,
Ornithischia): proposed designation of Iguanodon bernissartensis Boulenger
in Beneden, 1881 as the type species, and proposed designation of a
lectotype. Bulletin of Zoological Nomenclature, 55, 99–104.
Charig, A.J. & Newman, B.H. 1992. Scelidosaurus harrisonii Owen, 1861
(Reptilia, Ornithischia): proposed replacement in inappropriate lectotype.
Bulletin of Zoological Nomenclature, 49, 280–283.
Clark, N.D.L. 2001. A thyreophoran dinosaur from the early Bajocian (Middle
Jurassic) of the Isle of Skye, Scotland. Scottish Journal of Geology, 37,
19–26.
Coombs, W.P., Weishampel, D.B. & Witmer, L.M. 1990. Basal Thyreophora. In:
Weishampel, D.B., Dodson, P. & Osmolska, H. (eds) The Dinosauria.
University of California Press, Berkeley, 427–434.
Delair, J.B. 1966. New records of dinosaurs and other fossil reptiles from Dorset.
Proceedings of the Dorset Natural History and Archaeology Society, 87,
57–66.
Galton, P.M. 1969. The pelvic musculature of the dinosaur Hypsilophodon
(Reptilia: Ornithischia). Postilla, 131, 1–64.
Galton, P.M. 1971a. Hypsilophodon, the cursorial non-arboreal dinosaur. Nature,
231, 159–161.
Galton, P.M. 1971b. The mode of life of Hypsilophodon, the supposedly arboreal
ornithopod dinosaur. Lethaia, 4, 453–465.
Galton, P.M. 1971c. A primitive dome-headed dinosaur (Ornithischia: Pachyce-
phalosauridae) from the Lower Cretaceous of England and the function of the
dome of pachycephalosaurids. Journal of Paleontology, 45, 40–47.
Galton, P.M. 1974. The ornithischian dinosaur Hypsilophodon from the Wealden
of the Isle of Wight. Bulletin of the British Museum (Natural History), 25,
1–152.
Galton, P.M. 1975. English hypsilophodontid dinosaurs (Reptilia: Ornithischia).
Palaeontology, 18, 741–752.
Galton, P.M. 1977. The ornithopod dinosaur Dryosaurus and a Laurasia–
Gondwanaland connection in the Upper Jurassic. Nature, 268, 230–232.
Galton, P.M. 1980a. Armored dinosaurs (Ornithischia: Ankylosauria) from the
Middle and Upper Jurassic of England. Geobios, 13, 825–837.
Galton, P.M. 1980b. Priodontognathus phillipsii (Seeley), an ankylosaurian
dinosaur from the Upper Jurassic (or possibly Lower Cretaceous) of England.
Neues Jahrbuch fur Geologie und Palaontologie, Monatshefte, 1980, 477–
489.
Galton, P.M. 1980c. European Jurassic ornithopod dinosaurs of the families
Hypsilophodontidae and Camptosauridae. Neues Jahrbuch fur Geologie und
Palaontologie, Abhandlungen, 160, 73–95.
Galton, P.M. 1981. Craterosaurus pottonensis Seeley, a stegosaurian dinosaur
from the Lower Cretaceous of England, and a review of Cretaceous
stegosaurs. Neues Jahrbuch fur Geologie und Palaontologie, Abhandlungen,
161, 28–46.
Galton, P.M. 1983a. Sarcolestes leedsi Lydekker, an ankylosaurian dinosaur from
the Middle Jurassic of England. Neues Jahrbuch fur Geologie und Palaonto-
logie, Monatshefte, 1983, 141–155.
Galton, P.M. 1983b. Armored dinosaurs (Ornithischia: Ankylosauria) from the
Middle and Upper Jurassic of Europe. Palaeontographica Abteilung A, 182,
1–25.
Galton, P.M. 1983c. A juvenile stegosaurian dinosaur, Omosaurus phillipsi Seeley
from the Oxfordian (Upper Jurassic) of England. Geobios, 16, 95–101.
Galton, P.M. 1985. British plated dinosaurs (Ornithischia, Stegosauridae). Journal
of Vertebrate Paleontology, 5, 211–254.
Galton, P.M. 1989. Crania and endocranial casts from ornithopod dinosaurs of the
families Dryosauridae and Hypsilophodontidae (Reptilia: Ornithischia).
Geologica et Palaeontologica, 23, 217–239.
Galton, P.M. 1994. Dermal scutes of Sarcolestes, an ankylosaurian dinosaur from
the Middle Jurassic of England. Neues Jahrbuch fur Geologie und Palaonto-
logie, Monatshefte, 1994, 726–732.
Galton, P.M. 1999. Cranial anatomy of the hypsilophodontid dinosaur Bugen-
asaura infernalis (Ornithischia: Ornithopoda) from the Upper Cretaceous of
North America. Revue Paleobiologie, 18, 517–534.
Galton, P.M. 2005. Bones of large dinosaurs (Prosauropoda and Stegosauria) from
the Rhaetic Bone Bed (Upper Triassic) of Aust Cliff, southwest England.
Revue de Paleobiologie, 24, 51–74.
Galton, P.M. & Boine, G. 1980. A stegosaurian dinosaur femur from the
Kimmeridgian Beds (Upper Jurassic) of the Cap de la Heve, Normandy.
Bulletin Trimestrial de la Societe Geologique de Normandie et des Amis du
Museum du Havre, 67, 31–35.
Galton, P.M. & Powell, H.P. 1980. The ornithischian dinosaur Camptosaurus
prestwichii from the Upper Jurassic of England. Palaeontology, 23, 412–443.
Galton, P.M. & Powell, H.P. 1983. Stegosaurian dinosaurs from the Bathonian
(Middle Jurassic) of England, the earliest record of the family Stegosauridae.
Geobios, 16, 219–229.
Galton, P.M. & Taquet, P. 1982. Valdosaurus, a hypsilophodontid dinosaur from
the Lower Cretaceous of Europe and Africa. Geobios, 15, 147–159.
Galton, P.M. & Upchurch, P. 2004. Stegosauria. In: Weishampel, D.B.,
Dodson, P. & Osmolska, H. (eds) The Dinosauria, 2nd. University of
California Press, Berkeley, 343–362.
Giffin, E.B. 1989. Pachycephalosaur paleoneurology (Archosauria: Ornithischia).
Journal of Vertebrate Paleontology, 9, 67–77.
Gilmore, C.W. 1909. Osteology of the Jurassic reptile Camptosaurus, with a
revision of the species of the genus, and descriptions of two new species.
Proceedings of the United States National Museum, 36, 197–332.
Head, J.J. 1998. A new species of basal hadrosaurid (Dinosauria Ornithischia) from
the Cenomanian of Texas. Journal of Vertebrate Paleontology, 18, 718–738.
Hoffstetter, R. 1957. Quelques observations sur les stegosaurines. Bulletin de
Museum National d’Histoire Naturelle, 29, 537–547.
Hooley, R.W. 1925. On the skeleton of Iguanodon atherfieldensis sp. nov., from
the Wealden Shales of Atherfield (Isle of Wight). Quarterly Journal of the
Geological Society of London, 81, 1–61.
Horner, J.R., Weishampel, D.B. & Forster, C.A. 2004. Hadrosauridae. In:
Weishampel, D.B., Dodson, P. & Osmolska, H. (eds) The Dinosauria, 2nd.
University of California Press, Berkeley, 438–463.
Hulke, J.W. 1881. Polacanthus foxii, a large undescribed dinosaur from the
Wealden Formation in the Isle of Wight. Philosophical Transactions of the
Royal Society of London, 172, 653–657.
Hulke, J.W. 1882. Description of some Iguanodon-remains indicating a new
species, I. seelyi. Quarterly Journal of the Geological Society of London, 38,
135–144.
BRITISH DINOSAURS 621

Huxley, T.H. 1869. On Hypsilophodon, a new genus of Dinosauria. Abstracts
Proceedings of the Geological Society, London, 204, 3–4.
Huxley, T.H. 1870. On Hypsilophodon foxii, a new dinosaurian from the Wealden
of the Isle of Wight. Quarterly Journal of the Geological Society of London,
26, 3–12.
Irmis, R.B., Parker, W.G., Nesbitt, S.J. & Liu, J. 2006. Early ornithischian
dinosaurs: the Triassic record. Historical Biology, 19, 3–22.
Kirkland, J.I. 1998a. A polacanthine ankylosaur (Ornithischia: Dinosauria) from
the Early Cretaceous (Barremian) of eastern Utah. New Mexico Museum of
Natural History and Science Bulletin, 14, 271–281.
Kirkland, J.I. 1998b. A new hadrosaurid from the upper Cedar Mountain
Formation (Albian–Cenomanian: Cretaceous) of eastern Utah—the oldest
known hadrosaurid (lambeosaurine?). New Mexico Museum of Natural
History and Science Bulletin, 14, 283–295.
Lucas, S.G. 1996. The thyreophoran dinosaur Scelidosaurus from the Lower
Jurassic Lufeng Formation, Yunnan, China. Museum of Northern Arizona
Bulletin, 60, 81–85.
Lydekker, R. 1888. Note on a new Wealden iguanodont and other dinosaurs.
Quarterly Journal of the Geological Society of London, 44, 46–60.
Lydekker, R. 1889. On the remains and affinities of five genera of Mesozoic
reptiles. Quarterly Journal of the Geological Society of London, 45,
41–59.
Maidment, S.C.R., Wei, G. & Norman, D.B. 2006. Re-description of the
postcranial skeleton of the Middle Jurassic stegosaur Huayangosaurus taibaii.
Journal of Vertebrate Paleontology, 26, 944–956.
Mantell, G.A. 1825. Notice on the Iguanodon, a newly discovered fossil reptile,
from the sandstone of Tilgate Forest. Philosophical Transactions of the Royal
Society of London, 115, 179–186.
Mantell, G.A. 1833. The Geology of the South East of England. Longman,
London.
Mantell, G.A. 1841. Memoir on a portion of the lower jaw of the Iguanodon, and
on the remains of the Hylaeosaurus and other saurians, discovered in the
strata of Tilgate Forest, in Sussex. Philosophical Transactions of the Royal
Society of London, 131, 131–151.
Martill, D.M., Naish, D. & Earland, S. 2006. Dinosaurs in marine strata:
evidence from the British Jurassic, including a review of the allochthonous
vertebrate assemblage from the marine Kimmeridge Clay Formation (Upper
Jurassic) of Great Britain. In: Colectivo Arqueologico-Paleontologico
Salense, (ed.) Actas de las III Jornadas sobre Dinosaurios y su Entorno.
Salas de los Infantes, Burgos, 47–83.
Maryanska, T. 1990. Pachycephalosauria. In: Weishampel, D.B., Dodson, P. &
Osmolska, H. (eds) The Dinosauria. University of California Press, Berkeley,
565–577.
Metcalf, S.J. & Walker, R.J. 1994. A new Bathonian microvertebrate locality in
the English Midlands. In: Fraser, N.C. & Sues, H.-D. (eds) In the Shadow
of the Dinosaurs—Early Mesozoic Tetrapods. Cambridge University Press,
Cambridge, 322–331.
Naish, D. & Martill, D.M. 2001a. Armoured dinosaurs: thyreophorans. In:
Martill, D.M. & Naish, D. (eds) Dinosaurs of the Isle of Wight.
Palaeontological Association, London, 147–184.
Naish, D. & Martill, D.M. 2001b. Ornithopod dinosaurs. In: Martill, D.M. &
Naish, D. (eds) Dinosaurs of the Isle of Wight. Palaeontological Association,
London, 60–132.
Naish, D. & Martill, D.M. 2001c. Boneheads and horned dinosaurs. In:
Martill, D.M. & Naish, D. (eds) Dinosaurs of the Isle of Wight.
Palaeontological Association, London, 133–146.
Naish, D. & Martill, D.M. 2007. Dinosaurs of Great Britain and the role of the
Geological Society of London in their discovery: basal Dinosauria and
Saurischia. Journal of the Geological Society, London, 164, 493–510.
Newman, B.H. 1968. The Jurassic dinosaur Scelidosaurus harrisoni, Owen.
Palaeontology, 11, 40–43.
Newton, E.T. 1892. Note on an iguanodont tooth from the Lower Chalk
(‘Totternhoe Stone’), near Hitchin. Geological Magazine, 9(Decade 3),
49–50.
Norman, D.B. 1987. Wealden dinosaur biostratigraphy. In: Currie, P.M. &
Koster, E.H. (eds) Fourth Symposium on Mesozoic Terrestrial Ecosystems.
Boxtree Books, Drumheller, Alberta, 165–170.
Norman, D.B. 1990. A review of Vectisaurus valdensis, with comments on the
family Iguanodontidae. In: Carpenter, K. & Currie, P.J. (eds) Dinosaur
Systematics: Approaches and Perspectives. Cambridge University Press,
Cambridge, 147–161.
Norman, D.B. 1993. Gideon Mantell’s ‘Mantel-piece’: the earliest well-preserved
ornithischian dinosaur. Modern Geology, 18, 225–245.
Norman, D.B. 2000. Professor Richard Owen and the important but neglected
dinosaur Scelidosaurus harrisonii. Historical Biology, 14, 235–253.
Norman, D.B. 2001. Scelidosaurus, the earliest complete dinosaur. In: Carpenter,
K. (ed.) The Armored Dinosaurs. Indiana University Press, Bloomington,
3–24.
Norman, D.B. 2002. On Asian ornithopods (Dinosauria: Ornithischia). 4.
Probactrosaurus Rozhdestvensky, 1966. Zoological Journal of the Linnean
Society, 136, 113–144.
Norman, D.B. 2004. Basal Iguanodontia. In: Weishampel, D.B., Dodson, P. &
Osmolska, H. (eds) The Dinosauria, 2nd. University of California Press,
Berkeley, 413–437.
Norman, D.B. & Barrett, P.M. 2002. Ornithischian dinosaurs from the Lower
Cretaceous (Berriasian) of England. Special Papers in Palaeontology, 68,
161–189.
Norman, D.B. & Faiers, T. 1996. On the first partial skull of an ankylosaurian
dinosaur from the Lower Cretaceous of the Isle of Wight, southern England.
Geological Magazine, 133, 299–310.
Norman, D.B., Sues, H.-D., Witmer, L.M. & Coria, R.A. 2004. Basal
Ornithopoda. In: Weishampel, D.B., Dodson, P. & Osmolska, H. (eds) The
Dinosauria, 2nd. University of California Press, Berkeley, 393–412.
Norman, D.B., Butler, R.J. & Maidment, S.C.R. 2007. Reconsidering the status
and affinities of the ornithischian dinosaur Tatisaurus oehleri Simmons, 1965.
Zoological Journal of the Linnean Society, 150, 865–874.
Olshevsky, G. 1993. The origin and evolution of the stegosaurs. Dino-Frontline, 4,
64–103 [in Japanese].
Owen, R. 1861. A monograph of a fossil dinosaur (Scelidosaurus harrisonii,
Owen) of the Lower Lias. Palaeontographical Society Monographs, 13, 1–14.
Padian, K. 1989. Presence of the dinosaur Scelidosaurus indicates Jurassic age for
the Kayenta Formation (Glen Canyon Group, northern Arizona). Geology, 17,
438–441.
Paul, G.S. 2007. Turning the old into the new: a separate genus for the gracile
iguanodont from the Wealden of England. In: Carpenter, K. (ed.) Horns
and Beaks: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press,
Bloomington, 69–77.
Pereda-Suberbiola, J. 1993. Hylaeosaurus, Polacanthus, and the systematics and
stratigraphy of Wealden armoured dinosaurs. Geological Magazine, 130, 767–
781.
Pereda-Suberbiola, J. 1994. Polacanthus (Ornithischia, Ankylosauria), a transat-
lantic armoured dinosaur from the Early Cretaceous of Europe and North
America. Palaeontographica Abteilung A, 232, 133–159.
Pereda Suberbiola, X. & Barrett, P.M. 1999. A systematic review of
ankylosaurian dinosaur remains from the Albian–Cenomanian of England.
Special Papers in Palaeontology, 60, 177–208.
Radley, J. 2004. Demystifying the Wealden of the Weald (Lower Cretaceous,
south-east England). Open University Geological Society Journal, 25, 6–16.
Rixon, A.E. 1968. The development of the remains of a small Scelidosaurus from
a Lias nodule. Museums Journal, 67, 315–321.
Ruiz-Omenaca, J.I., Pereda-Suberbiola, X. & Galton, P.M. 2007. Callovo-
saurus leedsi, the earliest dryosaurid dinosaur (Ornithischia: Euornithopoda)
from the Middle Jurassic of England. In: Carpenter, K. (ed.) Horns and
Beaks: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press,
Bloomington, 3–16.
Sanchez-Hernandez, B., Benton, M.J. & Naish, D. 2007. Dinosaurs and other
fossil vertebrates from the Late Jurassic and Early Cretaceous of the Galve
area, NE Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 249,
180–215.
Seeley, H.G. 1888. The classification of the Dinosauria. Report of the British
Association for the Advancement of Science 1887. 698–699.
Seeley, H.G. 1893. Omosaurus phillipsi. Yorkshire Philosophical Society, Annual
Report, 1892, 52–57.
Sereno, P.C. 1986. Phylogeny of the bird-hipped dinosaurs (Order Ornithischia).
National Geographic Research, 2, 234–256.
Sereno, P.C. 1998. A rationale for phylogenetic definitions, with application to the
higher-level taxonomy of Dinosauria. Neues Jahrbuch fur Geologie und
Palaontologie, Abhandlungen, 20, 41–83.
Sereno, P.C. 2000. The fossil record, systematics and evolution of pachycephalo-
saurs and ceratopsians from Asia. In: Benton, M.J., Shishkin, M.A.,
Unwin, D.M. & Kurochkin, E.N. (eds) The Age of Dinosaurs in Russia and
Mongolia. Cambridge University Press, Cambridge, 480–516.
Storrs, G.W. 1994. Fossil vertebrate faunas of the British Rhaetian (latest
Triassic). Zoological Journal of the Linnean Society, 112, 217–259.
Sues, H.-D. 1980. A pachycephalosaurid dinosaur from the Upper Cretaceous of
Madagascar and its paleobiogeographical implications. Journal of Paleontol-
ogy, 54, 954–962.
Sues, H.-D. & Galton, P.M. 1987. Anatomy and classification of the North
American Pachycephalosauria (Dinosauria: Ornithischia). Palaeontographica
Abteilung A, 198, 1–40.
Sullivan, R.M. 2000. Prenocephale edmontonensis (Brown and Schlaikjer) new
comb. and P. brevis (Lambe) new comb. (Dinosauria: Ornithischia: Pachyce-
phalosauria) from the Upper Cretaceous of North America. New Mexico
Museum of Natural History and Science Bulletin, 17, 177–190.
Sullivan, R.M. 2003. Revision of the dinosaur Stegoceras Lambe (Ornithischia,
Pachycephalosauridae). Journal of Vertebrate Paleontology, 23, 181–207.
D. NAISH & D. M. MARTILL622

Sullivan, R.M. 2005. Pachycephalosaurs from Dinosaur Provincial Park, Alberta:
taxonomy, biostratigraphy, and paleobiogeographic implications. In: Braman,
D.R., Therrien, F., Koppelhus, E.B. & Taylor, W. (eds) Dinosaur Park
Symposium: Short Papers, Abstracts, and Program. Royal Tyrrell Museum,
Drumheller, Alberta, 121–126.
Thulborn, R.A. 1974. A new heterodontosaurid dinosaur (Reptilia: Ornithischia)
from the Upper Triassic Red Beds of Lesotho. Zoological Journal of the
Linnean Society, 55, 151–175.
Thulborn, R.A. 1977. Relationships of the Lower Jurassic dinosaur Scelidosaurus
harrisonii. Journal of Paleontology, 51, 725–739.
Topley, W. 1875. The Geology of the Weald (parts of the counties of Kent, Surrey,
Sussex and Hants.). HMSO, London.
Vickaryous, M.K., Maryanska, T. & Weishampel, D.B. 2004. Ankylosauria. In:
Weishampel, D.B., Dodson, P. & Osmolska, H. (eds) The Dinosauria, 2nd.
University of California Press, Berkeley, 363–392.
Wall, W.P. & Galton, P.M. 1979. Notes on pachycephalosaurid dinosaur
(Reptilia: Ornithischia) from North America, with comments on their status
as ornithopods. Canadian Journal of Earth Sciences, 16, 1176–1186.
Weishampel, D.B. 1990. Dinosaurian distribution. In: Weishampel, D.B.,
Dodson, P. & Osmolska, H. (eds) The Dinosauria. University of California
Press, Berkeley, 63–139.
Weishampel, D.B. & Heinrich, R.E. 1992. Systematics of Hypsilophodontidae
and basal Iguanodontia (Dinosauria: Ornithopoda). Historical Biology, 6,
159–184.
Weishampel, D.B., Jianu, C.-M., Csiki, Z. & Norman, D.B. 2003. Osteology and
phylogeny of Zalmoxes (n. g.), an unusual euornithopod dinosaur from the
latest Cretaceous of Romania. Journal of Systematic Palaeontology, 1, 65–
123.
Williamson, T.E. & Carr, T.D. 2002. A new genus of derived pachycephalosaur-
ian from western North America. Journal of Vertebrate Paleontology, 22,
779–801.
Xu, X., Forster, C.A., Clark, J.M. & Mo, J. 2006. A basal ceratopsian with
transitional features from the Late Jurassic of northwestern China. Proceed-
ings of the Royal Society of London, Series B, 273, 2135–2140.
Received 29 October 2007; revised typescript accepted 11 November 2007.
Scientific editing by Rob Strachan
BRITISH DINOSAURS 623


















