
The initiation of yeast DNA replication Questions? Contact michael.weinreich@vai

Developmental Regulation of DNA Replication Initiationin Drosophila
by
Fang Xie
B.S. in Biology (2001)Beijing University, Beijing, China
Submitted to the Department of Biologyin Partial Fulfillment of the Requirement for the Degree of
Doctor of Philosophy in Biology
at the
Massachusetts Institute of Technology
August, 2007
© 2007 Fang Xie. All rights reserved.
The author hereby grants to MIT permission to reproduce and to distribute publiclypaper and electronic copies of this thesis document in whole or in part
in any medium now known or hereafter created.
Signature of Author……………………………………………………………………Department of Biology
August 17, 2007
Certified by…………………………………………………...……………………….Terry L. Orr-WeaverProfessor of Biology
Thesis Supervisor
Accepted by………………………………………………..………………………….Stephen P. Bell
Chair, Committee of Graduate StudentsDepartment of Biology

2
Developmental Regulation of DNA Replication Initiationin Drosophila
byFang Xie
Submitted to the Department of Biologyon August 17, 2007 in Partial Fulfillment of the Requirement for
the Degree of Doctor of Philosophy in Biology
ABSTRACT
Developmental gene amplification in the ovarian follicle cells of Drosophilaprovides a powerful system for the study of metazoan DNA replication. Amplificationproduces 100kb gradients of amplified DNA through repeated rounds of origin firing andbidirectional movement of replication forks from these origins. The Drosophila FollicleCell Amplicon at the cytological location 62D, DAFC-62D, is uniquely regulated, withtwo separate rounds of amplification in developmental stages 10 and 13 of egg chamberdevelopment. We investigated mechanisms that control the unusual timing of DAFC-62Dorigin activation. We first defined origin sequences in DAFC-62D by analyzing theamount of nascent replicative DNA across this amplicon. Surprisingly, the origincoincides with the coding region of a gene named yellow-g2. ORC2 localizes to theorigin, as well as two other sites that do not confer origin activity. Both ORC2 andMCM2-7 display differential association with these sequences, corresponding to the tworounds of amplification. All three elements, dispersed in a 7kb central amplified region,are required for either round of DAFC-62D amplification, because deleting any onecompletely abolished amplification in transposon experiments. Preceded by transcriptionyellow-g2 in stage 12, the late round of origin firing was ablated by the RNAPII inhibitorα-amanitin. This effect was absent from other amplicons and insulated transposons, andwas stage-13 specific for amplification at either the endogenous DAFC-62D orheterologous transposons that did not have functional insulators. Therefore amplificationat DAFC-62D in late follicle cell differentiation depends on transcription in cis.Molecularly, blocking RNAPII transcription compromises MCM2-7 recruitment.Additional transposon and histone modification analyses confirmed the involvement ofRNAPII in amplification control, which may be facilitated by favorable chromatinstructure. This work provides insights in developmental regulation of origin firing,revealing one mechanism for initiation of metazoan DNA replication: recruitment ofMCM2-7 by RNA polymerase II transcription.
Thesis Suporvisor: Terry L. Orr-WeaverTitle: Professor of Biology

3
Dedicated to
Lin Li
李 林
Xingui Liang
梁新桂
and
Shangfa Xie
谢尚发

4
Acknowledgements
This thesis work would have been impossible without the full support of myadvisor Terry Orr-Weaver. She accepted me (and my ideas) with an open mind, herguidance throughout the years has made graduate school less obscure, and she alwaysinspires me to achieve more. I thank her for being the best advisor I could ever haveasked for.
I am thankful to all past and current Orr-Weaver lab members with whom I sharednumerous memorable moments, scientifically and non-scientifically. Julie Claycombmade the initial observations and lent tremendous help in establishing the DAFC-62Dproject. Discussions with Eugenia Park and Jane Kim, the replication subgroup people,have been inspiring. Everyone else, especially Tama Resnick, Jillian Pesin, DavidDoroquez, Yingdee Unhavaithaya, Lena Kashevsky and Raissa Formina, have made theTOW zone such a fun workplace.
My current thesis committee members, Steve Bell, Jianzhu Chen, and TroyLittleton, have been fantastic mentors and incredibly supportive. I am grateful forProfessor Nick Dyson’s tremendous help as the outside member of my defensecommittee. I also thank Ilaria Rebay and Paul Garrity for their advice in past committeemeetings. The regular Drosophila replication meetings with David MacAlpine, Cary Laiand Steve Bell have been a constant driving force of my research.
I would like to express my gratitude to my husband, Lin Li, and my parents,Xingui Liang and Shangfa Xie, for their unconditional love. I never said “thank you”enough, and could never thank you enough. You are my rock. You will stay my rock. Inthe pursuit of my dreams.

5
TABLE OF CONTENTSChapter OneIntroduction: Activation Control of Replication and Amplification Origins 6
Gene amplification as a model for DNA replication 7Origins of DNA replication 11Origins of developmental gene amplification 15Amplification control elements 23The involvement of transcription factors in gene amplification 27Chromatin context and amplification activity 34A general link between replication and transcription 36Summary of thesis 40
Chapter TwoIdentification of a Drosophila Replication Origin Developmentally Controlled by Transcription 50
Summary 51Introduction 52Results 55
Identification of the replication origin and ORC binding sites in DAFC-62D 55Differential pre-RC binding in DAFC-62D 59ORC-binding sequences are required for amplification 62The two rounds of origin firing are interspersed by transcription 66α-amanitin specifically inhibits DAFC-62D stage 13 amplification 70Inhibition of transcription affects MCM2-7 localization 75
Discussion 78Experimental procedures 86
Chapter ThreeConclusions and Future Directions 101
Differential localization of pre-RC 103Transcriptional regulation of replication initiation 106Distinct mechanisms of replication regulation 109Insulators and their insensitivity to α-amanitin 112Transcription factories 114
Appendix One: Analyses of the ACE3-ori62 Transposon 120
Appendix Two: Histone Acetylation and Amplification Activity 136
Appendix Three: Table of Acronyms 156

6
Chapter One
Introduction:
Activation Control of Replication and Amplification Origins

7
Gene amplification as a model for DNA replication
Developmental gene amplification is a process that increases the number of DNA
molecules as template for transcription at specific developmental points. It has been
reported in a variety of organisms as an alternate strategy to produce large quantities of
transcripts over relatively short periods of time. Among the first observed examples is
the amplification (about a thousand fold) of the genes that code for ribosomal RNA
(rRNA) during the development of Xenopus oocytes, in order to stockpile the egg with
the translational machinery necessary for rapid embryonic development (Brown and
Dawid, 1968; Gall, 1968). Electron microscopy studies suggest that the Amphibian
rDNA is amplified via a rolling-circle mechanism (Hourcade et al., 1973). Another
example of extrachromosomal gene amplification is the rDNA in the transcriptionally
active macronucleus of the protist Tetrahymena (Gall, 1974; Yao et al., 1979). The 21kb
minichromosome in the form of a palindrome comprises two copies of the rDNA and is
amplified up to 10,000 copies. This differs from the Amphibian oocyte rDNA however,
because these palindromic minichromosomes are not produced by a rolling-circle
mechanism, but rather by bidirectional movement of the replication forks initiated from a
defined origin (Figure 1A) (Kapler et al., 1996; Prescott, 1994).
Dipteran flies, including Rhynchosciara americana (Glover et al., 1982),
Bradysia hygida (Laicine et al., 1984), and Sciara coprophila (Wu et al., 1993) all utilize
gene amplification at multiple loci throughout the genome in the larval salivary glands,
presumably for the production of large quantities of the structural proteins for the

8
construction of the cocoon. Note that unlike Tetrahymena rDNA amplification, in these
organisms the gene clusters are replicated above the copy number of surrounding
sequences without forming extrachromosomal molecules. The same strategy is employed
by another Dipteran fly, Drosophila melanogaster, to amplify at least four groups of
genes in the ovarian follicle cells (Claycomb et al., 2004; Spradling, 1981; Spradling et
al., 1980). Two of these gene clusters contain genes that encode the major structural
proteins of the chorion (eggshell) (Spradling et al., 1980). It is possible that in Dipteran
flies the intrachromosomal structures generated by the amplification process may be
tolerated, as both the larval salivary gland and ovarian follicle cells are terminally
differentiated tissues and are lost during further development. Because these cell types
are nondividing, genomic aberrations accumulated during developmental gene
amplification would not be passed on to daughter cells.
Developmental gene amplification in both Tetrahymena and Dipteran flies has
been consistently shown to utilize the normal replication machinery to repeatedly initiate
DNA replication from dominant origins, resulting in an “onionskin” structure of nested
replication bubbles/forks (Figure 1B) (Claycomb and Orr-Weaver, 2005; Tower, 2004).
This provides an advantageous model for studying metazoan DNA replication for several
reasons. First, amplified regions (amplicons) are relatively well defined especially given
the recent employment of Comparative Genomic Hybridization (CGH) arrays (Claycomb
et al., 2004). Second, the repeated firing generates a gradient of DNA copy number with
the central origin(s) being the most abundant (Claycomb et al., 2002). This allows
focused analysis of cis-regulatory elements, including both origins and control sequences,

9
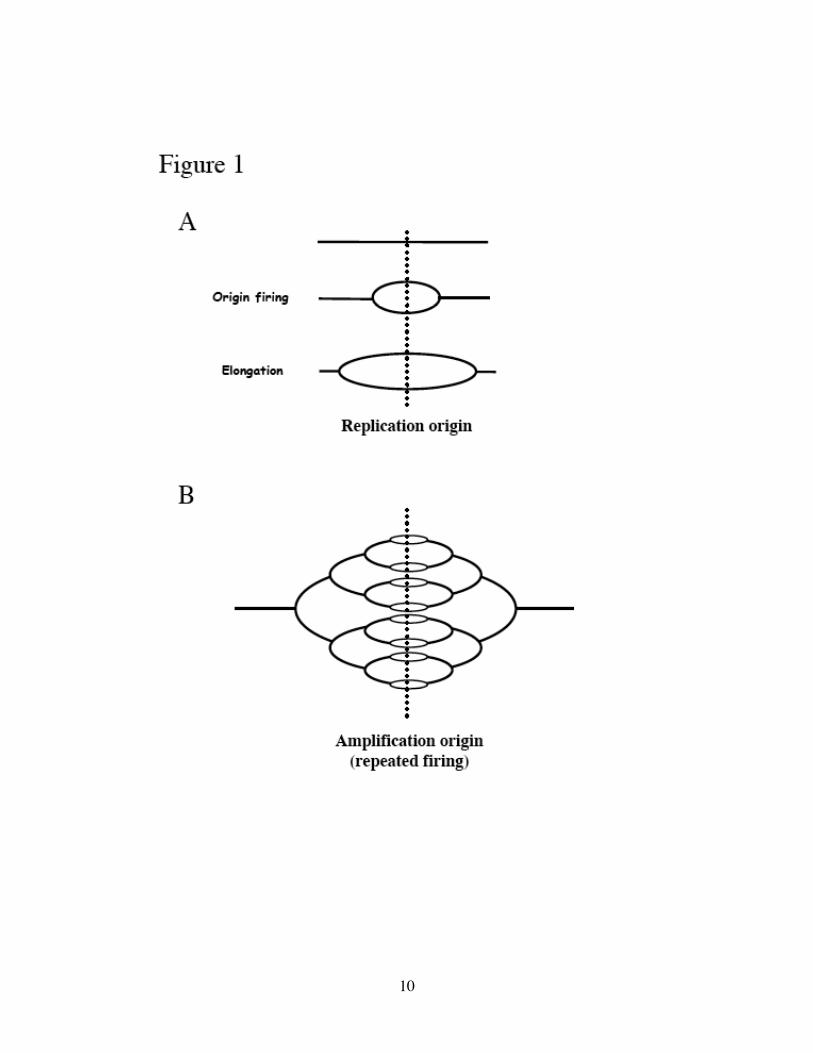
Figure 1. Models of bidirectional replication versus amplification.
(A) Replication origin fires once and only once per cell cycle, followed by bidirectional
movement (elongation) of replication forks. (B) During developmental amplification, the
origin is activated multiple times consecutively. The resulting multiple replication forks
form an “onionskin” structure, with the highest DNA copy number at the amplification
origin.
10

11
and provides insights into the mechanisms by which the usual once-per-cell-cycle control
of DNA replication can be thwarted. Third, developmental gene amplification occurs at
strategic developmental times, providing the opportunity to study how DNA replication
responds to developmental cues. Finally, a range of molecular and genetic tools are
available in these model organisms.
Origins of DNA replication
The best-studied eukaryotic origins are those in the yeast Saccharomyces
cerevisiae. Specific, well-defined origins of DNA replication have been revealed
primarily through genetic analyses (Newlon and Theis, 1993). Consisting of an 11bp A-
T-rich autonomously replicating sequence (ARS) consensus sequence (ACS) and other
elements (B1 and B2) (Figure 2A), the yeast replication origins first recruit a variety of
factors known as the pre-replication complex (pre-RC) (Figure 2A) to initiate DNA
replication. As a component of the pre-RC, the six-subunit ORC specifically recognizes
the ACS and the B1 element. Following loading of ORC, the other pre-RC factors and
additional replication factors are recruited and origins are subsequently activated (Bell
and Dutta, 2002). Although the protein factors appear to be highly conserved from yeasts
to higher eukaryotes, the DNA sequences that define origin activity in different
organisms are not (Cvetic and Walter, 2005). Furthermore, in vitro studies in higher
eukaryotes suggest that the metazoan ORC does not rely on sequence specificity to bind
DNA (Remus et al., 2004; Vashee et al., 2003).

12
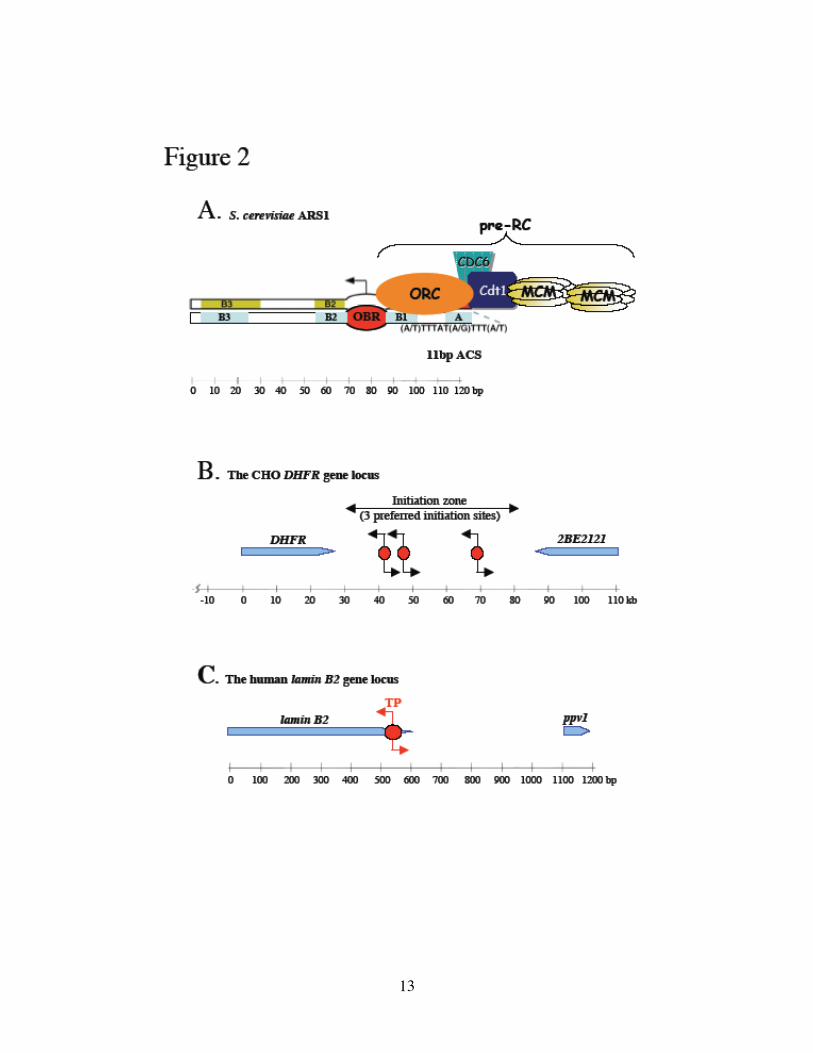
Figure 2. Classes of eukaryotic replication origins and composition of the pre-RC.
(A) S. cerevisiae ARS1 is a well-defined replication origin. The 11bp ACS (and the B1
element) is specifically recognized by ORC, which together with Cdc6, Cdt1 and MCM2-
7 constitute the pre-RC. The Origin of Bidirectional Replication (OBR) has been mapped
immediately adjacent to the ORC-binding site. (B) The CHO DHFR locus contains a
broad initiation zone, spanning the entire 55kb intergenic region between the DHFR and
2BE212 genes. Three sites show higher initiation activity. (C) The human lamin B2
replicon is markedly less complicated. The exact Transition Point (TP) from
discontinuous to continuous DNA synthesis has been determined by RIP mapping at the
nucleotide resolution. See text for references. Blue pointed bars represent the coding
frame of genes.
13

14
Physical mapping techniques have been developed to locate origins of DNA replication.
Two-Dimensional (2D) gels separate replicating from nonreplicating molecules and
allows the analysis of replication intermediates. This method is of relatively low
resolution, revealing origins of replication as small as 2kb (DePamphilis, 1999). More
sensitive approaches such as nascent strand analysis that employ PCR to determine the
abundance of nascent strands improved the resolution of initiation sites to a few hundred
basepairs (Giacca et al., 1994; Kobayashi et al., 1998a). Recently developed Replication
Initiation Point (RIP) mapping has achieved nucleotide resolution by accurately defining
the Transition Point (TP) from discontinuous to continuous DNA synthesis (Bielinsky
and Gerbi, 1998; Gerbi and Bielinsky, 1997), and positioned the origin of bidirectional
replication (OBR) immediately adjacent to the ORC-binding site in yeast (Figure 2A)
(Bielinsky and Gerbi, 1998; Bielinsky and Gerbi, 1999).
In contrast to our knowledge of yeast origins, what constitutes a replication origin
in higher eukaryotes is poorly understood (Bielinsky et al., 2001; DePamphilis et al.,
2006; Gilbert, 2004). A handful of metazoan model replicons have been studied in detail
in tissue culture cells (Cvetic and Walter, 2005; Gerbi, 2005). The fact that these systems
lack convenient genetic assays has restricted metazoan origin studies to physical
biochemical mapping methods. It has been shown by 2D gels that replication of the
Chinese hamster DHFR (Burhans et al., 1990; Vaughn et al., 1990) and human rDNA
loci (Little et al., 1993; Yoon et al., 1995) initiates in broad regions. These data suggest
the existence of large initiation zones (Gilbert, 2001), although in the DHFR locus two to
three specific sites are preferred over other initiation sites spread throughout a 55kb

15
region (Figure 2B) (Dijkwel et al., 2002; Kobayashi et al., 1998a). On the other hand,
studies of human lamin B2 (Figure 2C) and β-globin origins have identified much more
defined sites of replication initiation, consistent with the classic replicon/replicator model
(Gilbert, 2004; Jacob and Brenner, 1963). Thus, there seem to be two classes of
mammalian origins depending on the locus.
Origins of developmental gene amplification
As previously discussed, developmental gene amplification provides a powerful
system for the study of metazoan DNA replication in vivo. In the development of the
Tetrahymena macronucleus, the 10.3kb rDNA locus is specifically excised from the
genome, converted to a ~21kb head-to-head palindrome, and telomeres are added to
generate stable linear minichromosomes (Figure 3A). Then over the course of twelve
hours, the rDNA minichromosomes are preferentially amplified up to 10,000-fold
(Kapler et al., 1996; Prescott, 1994). Amplification initiates from two 430bp sites in
Tetrahymena thermophila (Figure 3A) (MacAlpine et al., 1997), or a single 900bp region
in T. pyriformis (Yue et al., 1998), all located within the 5’ Nontranscribed Spacer region
(5’NTS) that is in the center of the palindrome. It appears that some of the amplified
molecules separate from each other, as FISH studies demonstrate the presence of several
hundred rDNA loci in nucleoli throughout the macronucleus (Ward et al., 1997). Given
the small size (21 kb) of these extrachromosomal molecules, it is conceivable that at least
some minichromosomes are fully replicated and some portion of the onionskin structures
resolve. In contrast, amplification in Dipteran flies only represents a small portion of the

16
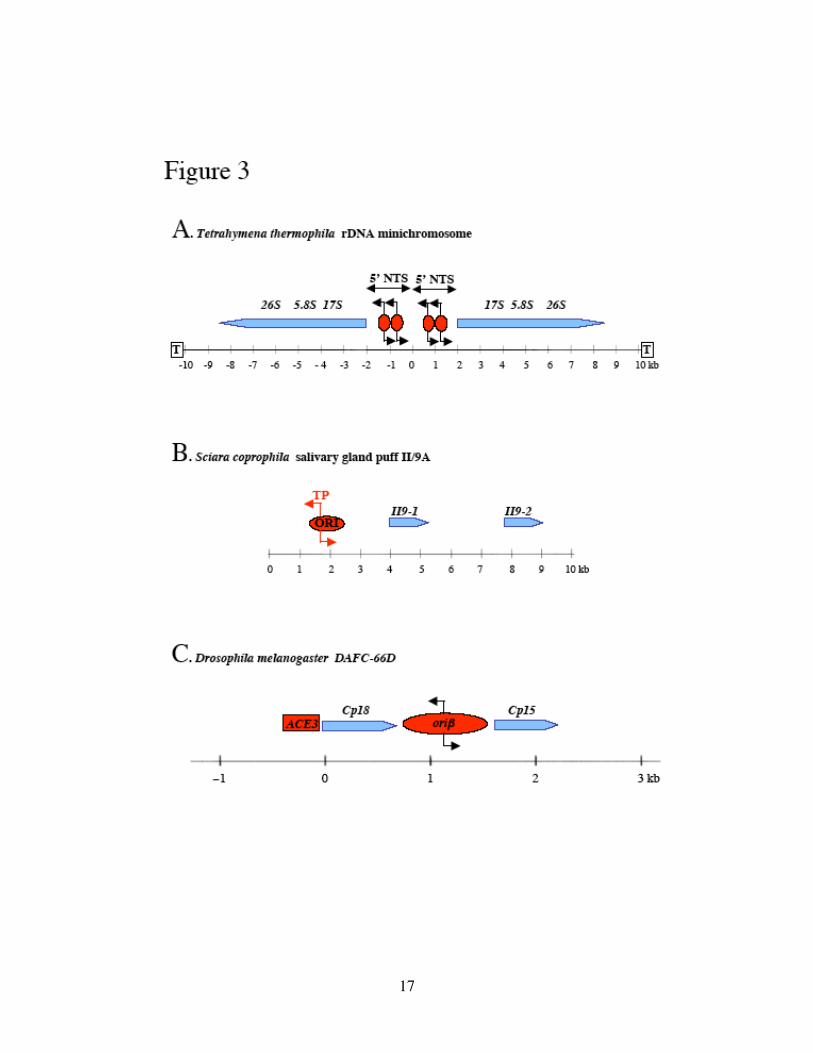
Figure 3. Origins of developmental gene amplification.
(A) Tetrahymena thermophila rDNA minichromosome. T represents the telomere at
either end. Red ovals are nucleosome-free regions within the 5’ NTS (Nontranscribed
Spacer) of the rRNA genes that show initiation activity. (B) Sciara coprophila salivary
gland DNA puff II/9A. A 1kb origin (ORI) has been mapped upstream of II/9A genes by
2D and 3D gel analyses. ORI contains an ORC-binding site, immediately adjacent to the
Transition Point (TP) from discontinuous to continuous DNA synthesis. (C) Drosophila
melanogaster DAFC-66D. The majority of origin activity resides in the intergenic oriβ.
ACE3 is an amplification control element necessary for amplification, and it is
functionally separable from the Cp18 promoter. See text for references. Blue pointed bars
represent the coding frame of genes.
17

18
giant polytene chromosomes in which the amplicons (each about 100kb in size) reside.
Furthermore, suggested by FISH (Calvi et al., 1998) the onionskin structures remain held
together without subsequent rearrangements.
In the Sciara salivary puff II/9A, 2D and 3D gel analyses indicate that initiation
occurs over a 5.5kb region, within which a preferred 1kb portion (ORI) accounts for the
majority of the origin firing (Figure 3B) (Liang and Gerbi, 1994; Liang et al., 1993). The
precise nucleotide (Transition Point, TP) within the 1kb region at which DNA synthesis
initiates has been determined by RIP mapping, and both recombinant ORC2 protein from
Drosophila and endogenous Sciara ORC2 have been shown to bind to an 80bp segment
adjacent to this initiation site (Bielinsky et al., 2001). In the related Sciarid fly,
Rhynchosciara, 2D gel analyses demonstrate that replication initiates in the salivary puff
C3 from at most 3 sites in a zone of about 6kb, and that this zone resides approximately
2kb upstream of the amplified gene C3-22 (Bielinsky et al., 2001).
In Drosophila, gene amplification of four genomic loci in the somatic follicle cells
of the ovary occurs at specific stages of egg chamber development (stages 10 to 13,
Figure 4A). During stages 9 and 10, the follicle cells surrounding the developing oocyte
cease genomic DNA replication and begin to amplify four clusters of genes, which can be
visualized as four foci by immunofluorescence following BrdU (bromodeoxyuridine, a
thymidine analog) incorporation (Calvi et al., 1998). These Drosophila Amplicons in
Follicle Cells (DAFCs) are named according to their cytological locations. Two of the
amplicons are the X chromosome (at 7F, DAFC-7F) and the 3rd chromosome (at 66D,
DAFC-66D) chorion (eggshell) genes. Chorion gene amplification is needed to meet the

19
demand for the rapid synthesis of chorion proteins (Orr-Weaver, 1991). The other two
loci of amplification, DAFC-30B and DAFC-62D, were recently identified by CGH array
studies (Claycomb et al., 2004). These amplicons contain genes encoding a variety of
proteins, including transporters, proteases, chitin-binding proteins and two putative
enzymes Yellow-g and Yellow-g2, thought to be necessary for crosslinking proteins of
the vitelline membrane or eggshell (Claycomb et al., 2004).
The third chromosome chorion amplicon DAFC-66D is the best studied (Figure
3C). 2D gel analysis has identified three potential replication origins within the peak
amplified region, with one of these, oriβ, the 884bp sequence downstream of the Cp18
chorion protein gene being the preferred site of initiation that contains 70-80% of the
origin activity (Delidakis and Kafatos, 1989; Heck and Spradling, 1990). Oriβ has ten
out of eleven base pair matches to the Saccharomyces cerevisiae ARS consensus
sequence that serves as an essential part of the origin of replication in yeast (Levine and
Spradling, 1985). However, the significance of this sequence similarity is not clear, and
notably, S. cerevisiae ARS1 origin sequences can not substitute for oriβ, thereby
confirming the sequence specificity of oriβ (Zhang and Tower, 2004). Deletion
mapping of oriβ identified a 140 bp 5' element and a 226 bp A/T-rich 3' element called
the β region that are necessary and sufficient to induce amplification of transposons
(Zhang and Tower, 2004). The high A/T content of the β region might be important,
because ORC has been shown to preferentially bind to A/T rich sequences in many
species (for review see Bell, 2002). The replication initiation protein ORC2 binds

20
directly to oriβ during gene amplification (Austin et al., 1999). However, despite its tight
association with oriβ in vivo, ORC does not preferentially bind DAFC amplification
origin sequences in in vitro assays (Remus et al., 2004). This is similar to observations in
other metazoan systems (Vashee et al., 2003), although contrasting Saccharomyces
cerevisiae in which specific sequences (the ARS Consensus Sequence or ACS) within the
origins are recognized (Newlon and Theis, 1993).
The developmental timing of amplification initiation appears to be highly
regulated and specific to particular amplicons. Real-time PCR suggested that DAFC-62D
behaves distinctly from the other three amplicons in the timing of origin firing (Figure
4B) (Claycomb et al., 2004). In DAFC-66D, -7F and -30B, origin firing occurs only in
stages 10 and 11. Afterwards there is no more increase in copy number at the central
initiation sites. However, for DAFC-62D, an additional round of origin firing is observed
in stage 13 within a defined 4kb region. Therefore, DAFC-62D provides a distinct model
for studying not only the mechanisms of origin selection and activation, but also its
developmental regulation. By identifying cis-regulatory elements in DAFCs that direct
amplification as origin(s), and those that regulate amplification as enhancers by sensing
differential developmental signals in different stages, we have gained important insights
in understanding metazoan DNA replication in vivo.

21
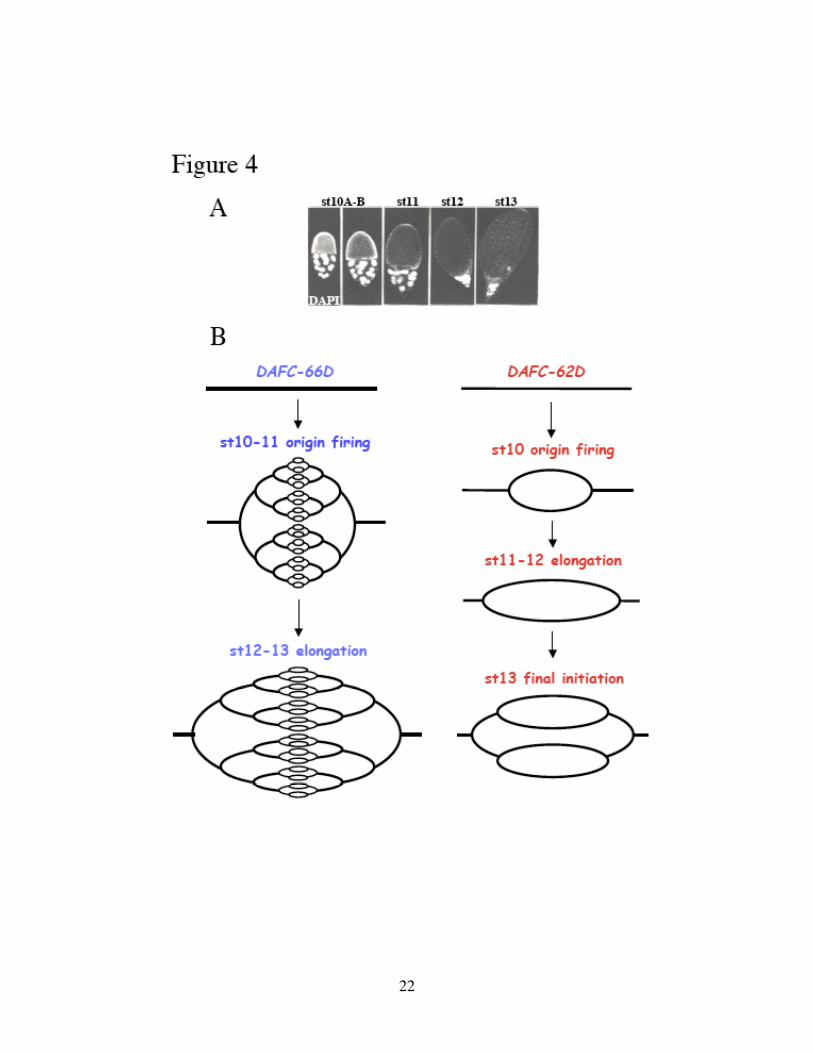
Figure 4. Developmental timing of DAFC amplification.
(A) DAPI staining of egg chambers in stages 10 to 13. Follicle cells surround the
developing oocytes. Adapted from A. C. Spradling, 1993. (B) Schematic drawing of the
developmental timing of DAFC-66D and DAFC-62D amplification. About 30 fold of
amplification in the center of DAFC-66D indicates 5 rounds of origin firing, all taking
place in stages 10-11. By contrast, amplification of DAFC-62D is activated in two
distinct stages, 10 and 13, separated by an elongation-only phase.
22

23
Amplification control elements
The relative ease of genetically manipulating Drosophila has greatly facilitated
the mapping of cis-regulatory elements in DAFCs. In the P-element mediated
transformation systems, transposons that contain proper cis elements are able to amplify
at ectopic genomic loci, although levels of amplification are dramatically affected by
chromosomal position effects (de Cicco and Spradling, 1984; Orr-Weaver and Spradling,
1986). The introduction of insulators (Suppressor of Hairy-Wing Binding Sites
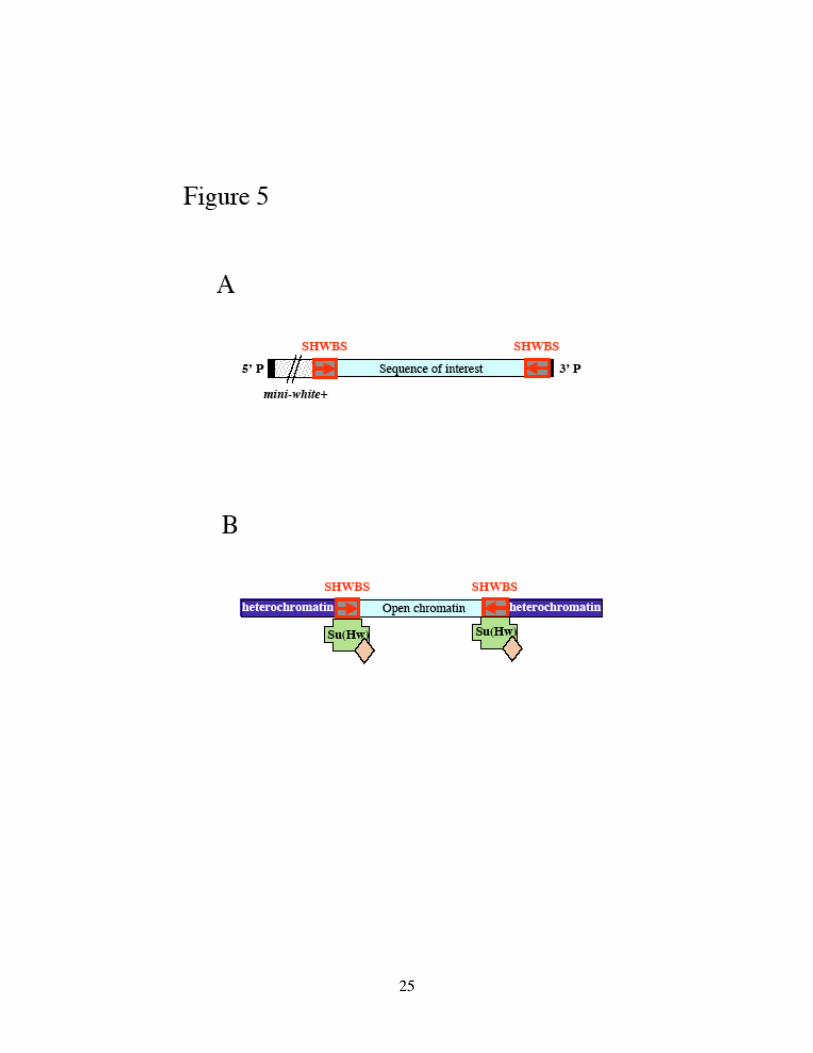
(SHWBS), Figure 5A) helps to remove position effects (Lu and Tower, 1997). The
SHWBS insulators recruit proteins including Su(Hw) (Suppressor of Hairy Wing) that
has been suggested to function as barriers between heterochromatin and open chromatin
(Figure 5B) (Capelson and Corces, 2004; Gerasimova and Corces, 2001).
A number of studies have delineated in DAFC-66D the 320bp Amplification
Control Element on the third chromosome, ACE3, required for high levels of
amplification (Figure 3C) (de Cicco and Spradling, 1984; Delidakis and Kafatos, 1989;
Orr-Weaver et al., 1989; Orr-Weaver and Spradling, 1986). ACE3 is evolutionarily well
conserved, located approximately 1.5kb upstream of oriβ and at the 5’ end of the s18
chorion gene. Demonstrated by 2D gel analyses (Delidakis and Kafatos, 1989; Heck and
Spradling, 1990), ACE3 itself does not function as an origin of DNA replication, as it is
not sufficient to support amplification in transposons protected by SHWBS insulators (Lu
et al., 2001). Similarly, DAFC-7F contains an ACE element (ACE1) that is important for
the amplification of this gene cluster (Spradling et al., 1987).

24
Figure 5. The SHWBS insulators remove position effects in P-element mediated
transposon systems.
(A) Structure of transposon within the P-element sequences (black boxes). DNA
sequence of interest (light-blue box) is flanked by SHWBS insulators. Arrows indicate
orientation of SHWBS. mini-white (stippled box) is a reporter gene for the selection of
transformation lines. (B) SHWBS binds Su(Hw) (Supressor of Hairy Wing) and
additional proteins to form a barrier against surrounding heterochromatin (dark-blue
boxes), so that the open chromatin structure (light-blue box) of the transposon is not
affected by chromosomal position effects. See text for references.
25

26
It has been proposed that ACE serves as a developmental control element by
stimulating replication initiation at nearby origins (Carminati et al., 1992; Delidakis and
Kafatos, 1989; Heck and Spradling, 1990). A 142bp highly evolutionarily conserved
“core” region of ACE3 has been determined responsible for the majority of ACE3’s
replication stimulatory activity by deletion studies (Zhang and Tower, 2004).
Furthermore, ACE3 is necessary in cooperation with oriβ to achieve high levels of gene
amplification, as a SHWBS insulator placed between ACE3 and oriβ in transposons
nearly eliminates amplification. Removal of this insulator element by FLP/FRT-
mediated recombination then restores amplification (Lu et al., 2001). Additionally,
elimination of either ACE3 or oriβ from transposon constructs dramatically reduces
amplification levels, indicating that together, ACE3 and oriβ are necessary and sufficient
to drive developmental amplification (Lu et al., 2001).
Recent molecular studies provide some clues of how ACE3 might function as an
amplification enhancer. ORC binds directly not only to oriβ but also to ACE3 and ACE1
in a site-specific manner, by either in vivo Chromatin Immunoprecipitation (ChIP) or in
vitro binding assays (Figure 6A) (Austin et al., 1999; Royzman et al., 1999). It has hence
been suggested that ACE3 serves as a nucleating site for ORC to spread along the
chromatin, thus influencing the ability of the region to replicate (Austin et al., 1999; Lu et
al., 2001). By immunofluorescence, transposons of ACE3 multimers are capable of
recruiting ORC2 in vivo (Austin et al., 1999), and support amplification presumably

27
initiated from proximal origins (Carminati et al., 1992). The amplification of a minimal
transposon buffered by SHWBS containing only ACE3, Cp18, and oriβ is dependent on
the orc2 gene product by mutant analysis, though without detection of ORC2 in
immunofluorescence (Lu et al., 2001). Therefore it appears that a certain threshold
amount of ORC2 must be recruited. It may not always be detectable by staining methods,
but in more sensitive assays such as ChIP ORC2 clearly associates with amplification
origins and enhancers (Austin et al., 1999). Cumulatively, these data indicate that ACE3
and oriβ are functionally separable, but act cooperatively to drive gene amplification. The
current working model is that ACE3 may nucleate ORC that then spreads along the
chromatin to initiate replication at oriβ.
The involvement of transcription factors in gene amplification
Genetic, cytological, and biochemical approaches have also contributed to the
understanding of the trans factors involved in developmental gene amplification. It has
been clearly demonstrated that the proteins involved in DNA replication during a normal
cell cycle are also involved in replication during gene amplification (Claycomb and Orr-
Weaver, 2005; Tower, 2004). Hypomorphic mutations in Drosophila genes encoding
essential components of the replication machinery lead to female sterility, disrupted
eggshells, and severely decreased DAFC amplification, as measured by incorporation of
BrdU or Quantitative Southern blotting (Henderson et al., 2000; Landis et al., 1997b;
Landis and Tower, 1999; Tower, 2004; Underwood et al., 1990; Whittaker et al., 2000).

28
The archetypal DNA replication machinery includes first the formation of the pre-RC at
the origins (Bell and Dutta, 2002). The ORC and DUP/Cdt1 proteins are sequentially
recruited, which in turn load the putative replication fork helicase complex, MCM2-7
(Aparicio et al., 1997; Bell and Dutta, 2002; Labib et al., 2001).
In addition to conserved replication proteins, the components known to play a role
in Drosophila chorion gene amplification include transcription factors. While chromatin
immunoprecipitation (ChIP) experiments have shown that ORC binds directly to ACE3
and to oriβ (Austin et al., 1999; Bosco et al., 2001), additional ChIP and binding studies
have localized transcription factors E2F1/DP/Rb to ACE3 during amplification stages in a
complex containing ORC (Figure 6A) (Bosco et al., 2001). In the normal cell cycle, the
E2F1/Rb complex acts as a transcriptional repressor until at the G1 to S phase transition
phosphorylation of Rb converts E2F1 into a transcriptional activator for the expression of
multiple genes required for S phase entry (Dyson, 1998; Zhu et al., 2005). E2F1 is
required for chorion gene amplification because E2f1 mutants in which the DNA-binding
domain is disrupted display decreased amplification and no ORC localization; a
hypomorphic Rb mutation or a mutation in E2f1 that removes the Rb binding site results
in overamplification and inappropriate genomic replication (Royzman et al., 1999).
These data support a model in which E2F1/Rb binds at and/or around ACE3 and
represses replication until amplification stages, during which E2F1 positively regulates
amplification initiation, with hyperphosphorylated Rb (pRb).
There are two E2f genes in Drosophila, E2f1 and E2f2 (Frolov et al., 2001). E2F1
is a potent activator of transcription, whereas E2F2 has been shown to repress

29
transcription (Dyson, 1998). In null E2f2 mutants BrdU incorporation occurs throughout
the nucleus during DAFC amplification stages, failing to confine DNA synthesis to
DAFC sites (Cayirlioglu et al., 2001). In parallel, the distribution of pre-RC components
changes from being restricted to DAFC foci into being nuclear in these mutants,
suggesting a repressive role of E2F2 in genomic DNA replication (Cayirlioglu et al.,
2001). Although in E2f2 mutants there is a mild increase in pre-RC transcript level
(Cayirlioglu et al., 2001; Cayirlioglu et al., 2003), it does not exclude the possibility that
E2F2 functions directly at genomic origins to repress replication (see below).
Another transcription factor that associates with ACE3 is the Myb oncoprotein. A
complex containing Myb, Mip120 (Myb Interacting Protein 120, formerly p120),
Mip130, Mip40, and Caf1 p55 interacts with ORC (Figure 6A) (Beall et al., 2002;
Korenjak et al., 2004). Both the Myb and Mip120 subunits exhibit specific binding.
Within ACE3, binding sites for Myb and Mip120 have been identified, and deletion of
these sites from transposons nearly abolishes amplification compared to the non-deleted
control (Beall et al., 2002). These results indicate that the Myb and at least one of the
Mip120 binding sites are necessary for amplification. Myb is essential for viability, as
Myb mutants are lethal. Myb mutant follicle cell clones are defective in BrdU
incorporation at DAFCs, showing that Myb is necessary for gene amplification (Beall et
al., 2002). The fact that by immunofluorescence ORC2 and DUP/Cdt1 are properly
localized to DAFCs in Myb mutant clones indicates that Myb is required for initiation at a
later step (Beall et al., 2002). Mip130 mutant females are sterile and have BrdU
incorporation throughout the nucleus during amplification stages (Beall et al., 2004).

30
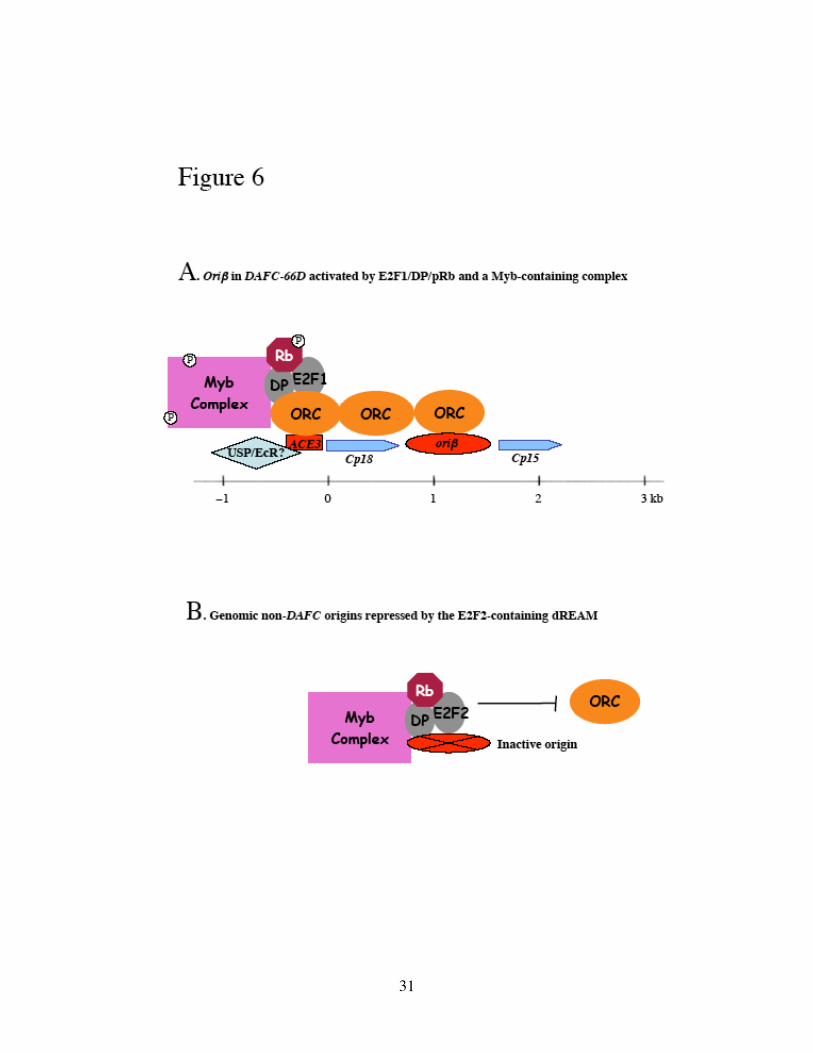
Figure 6. Transcription factors involved in DAFC-66D amplification and origin
specification.
(A) E2F1/DP/Rb and a complex containing Myb specifically associate with ACE3. The
Rb and Myb proteins may be activated through phosphorylation. ORC is site-specifically
recruited and spreads along the chromatin to initiate replication from oriβ. The
Ultraspiracle/ Ecdysone Receptor (USP/EcR) transcription factor also may regulate
amplification via an interaction with ACE3. (B) During amplification stages, genomic
replication is inhibited by the E2F2-containing dREAM complexes, which excludes ORC
from inactive non-DAFC origins.
31

32
From these data, Mip130 appears to interact with the other Mips in a complex to repress
genome-wide replication. At specific chromosomal loci Myb becomes activated in some
way, perhaps by phosphorylation (Beall et al., 2004), to initiate replication or
amplification. Such a switch from repressive to active state might be important for Myb
to specifically allow the initiation of amplification at the appropriate developmental time
at amplification origins.
Strikingly, the Myb and Mip130 mutant phenotypes are very similar to those of
E2f1 and E2f2, respectively. In addition to genetic evidence, molecular and biochemical
studies strongly suggest the E2F and Myb proteins co-regulate replication. E2F1 and the
Myb-containing complex, both localized to ACE3, may act coordinately to activate
DAFC-66D amplification (Figure 6A). Although there is no report of a physical
interaction between E2F1 and Myb, a complex containing E2F2, Myb and Mips has been
purified from Drosophila embryo extracts (Korenjak et al., 2004; Lewis et al., 2004).
These dREAM complexes (Drosophila Rb, E2F and Myb-interacting proteins) bind to
repressed chromatin (Korenjak et al., 2004). Based on the mutant phenotypes of Myb and
E2f, it has been proposed that dREAM inhibits genome-wide replication (Korenjak et al.,
2004; Lewis et al., 2004), possibly by excluding pre-RC from genomic origins (Figure
6B); at DAFC-66D this repressive effect is reversed by E2F1 and activated Myb to
achieve site-specific amplification (Figure 6A).
E2F and Myb appear to directly regulate amplification without affecting
transcription of DAFC-66D genes. The transcription factor Ultraspiracle (USP) on the
other hand, has been shown to bind to the promoter of the Cp18 chorion gene of DAFC-

33
66D (Shea et al., 1990). USP is a zinc finger protein differentially enriched in the follicle
cells, and it is a developmentally important member of the family of nuclear steroid
hormone receptors (Oro et al., 1992; Shea et al., 1990). It heterodimerizes with another
member of the nuclear receptor superfamily, ecdysteroid receptor protein (EcR), to
function as a receptor for the steroid hormone ecdysone (Christianson et al., 1992; Yao et
al., 1992). Ecdysone governs egg chamber development, and maternal EcR is required
for normal oogenesis (Buszczak et al., 1999; Carney and Bender, 2000). EcR displays
increased activity in follicle cells during amplification stages (Hackney et al., 2007).
Dominant negative mutants of EcR (DNEcR) can dimerize with USP and bind DNA, but
they do not activate target gene expression (Cherbas et al., 2003; Hackney et al., 2007).
Interestingly, introduction of DNEcR into follicle cells not only reduces chorion gene
expression, but also results in significantly decreased amplification, and the eggs laid by
mutant mother display thin eggshells and shortened dorsal appendages (Hackney et al.,
2007). Taken together, these results indicate that the USP/EcR heterodimer mediates
ecdysone regulation of chorion gene amplification and transcription (Figure 6A). These
two events may be separable from each other because ACE3, discrete from sequences
controlling transcription (Orr-Weaver et al., 1989), harbors a good match to Ecdysone
Response Element (Hackney et al., 2007).
In Sciara coprophila the amplification of salivary gland DNA puff II/9A maybe
similarly influenced by ecdysone, the master regulator of insect development (Crouse,
1968; Foulk et al., 2006). Ecdysone induces transcription of the II/9A genes (Bienz-
Tadmor et al., 1991; DiBartolomeis and Gerbi, 1989; Wu et al., 1993). In vitro

34
incubation of pre-amplification stage salivary glands with ecdysone induces premature
amplification, and injection of ecdysone into pre-amplification stage larvae results in
amplification in vivo (Foulk et al., 2006). A putative EcRE is found directly adjacent to
the ORC-binding site in the II/9A origin (Bielinsky et al., 2001) and is efficiently bound
by the Sciara EcR in in vitro binding assays (Foulk et al., 2006). The Sciara and
Drosophila results indicate that the ecdysone and EcR transcription factor control of
developmental gene amplification may be conserved in insects.
Recently proteins TIF1-4 (Type I interacting Factor) that are necessary for rDNA
amplification in Tetrahymena have been purified as complexes with differential DNA
binding activities within the initiation zone (Mohammad et al., 2000). Notably TIF1
possesses limited homology to a transcription factor in plants, p24 (Mohammad et al.,
2000). Together with data from Drosophila and Sciara, it is clear that developmental
gene amplification is under complex control that involves a number of transcription
factors, acting to modulate the use of origins for amplification. These factors may play
repressive or active roles, depending on the developmental stage, the genomic locus and
the chromatin context.
Chromatin context and amplification activity
It is not surprising that the replication and amplification machinery requires a
favorable chromatin context to access DNA (Groth et al., 2007). Eukaryotic DNA is
packaged into an organized, higher-order chromatin structure by histone proteins (Loden
and van Steensel, 2005). Post-translational modifications of histones such as acetylation

35
of histone N-terminal lysine residues induces chromosomal changes, resulting in the loss
of chromosomal repression to allow successful transcription of the underlying genes, as
well as replication of DNA molecules (Fukuda et al., 2006). Recent studies in yeast have
provided evidence that posttranslational chromatin modification can control the
efficiency and/or timing of chromosomal origin activity (Aparicio et al., 2004; Vogelauer
et al., 2002).
Using the DAFC systems, independent groups have demonstrated that histones
H3 and H4 at and around ACE3 are hyperacetylated during gene amplification (Aggarwal
and Calvi, 2004; Hartl et al., 2007), and the lysine residues that are acetylated are
associated with replication and not transcription (Hartl et al., 2007). Furthermore, the
acetylation of H3 and H4 is not the result of histone deposition after replication, as the
hyperacetylation is confined to the origins of DAFC-66D and not associated with the
replication forks (Hartl et al., 2007). Hyperacetylation of histone H4 leads to
redistribution of ORC2 from amplification foci to a genome-wide staining pattern;
tethering histone acetyl transferase (HAT) increases amplification levels of a transposon
with ACE3 and oriβ (Aggarwal and Calvi, 2004). Conversely, tethering of a histone
deacetylase (HDAC) or a chromatin repressor to ACE3 reduces amplification (Aggarwal
and Calvi, 2004). These observations suggest chromatin structure may have a definite
role in amplification origin activity and that origin firing may be facilitated by a
modification of the chromatin state.
Such modifications may be conducted through recruitment of histone-modifying
enzymes and/or chromatin-remodeling proteins by transcription factors (Kohzaki and

36
Murakami, 2005). In X. laevis eggs, injected plasmid DNA undergoes site-specific
initiation of replication in the presence of a transcription factor that is known to recruit a
chromatin-remodeling complex (Danis et al., 2004). This effect does not require active
transcription, but rather correlates with the acetylation level of histone H3 at the initiation
sites (Danis et al., 2004). The E2F/Rb and Myb complexes are good candidates that may
function at DAFC amplification origins to recruit HATs or HDACs to modulate the
accessibility of the chromatin at the origin (Beall et al., 2002; Bosco et al., 2001). For
example, Rb has been shown in a number of organisms to repress transcription by
remodeling chromatin structure through interaction with proteins involved in nucleosome
remodeling, histone acetylation/deacetylation and methylation (Giacinti and Giordano,
2006). Finally, chromatin state and nucleosomal positioning may also play a role in gene
amplification in Sciara and Tetrahymena (Clever and Ellgaard, 1970; Giacinti and
Giordano, 2006; MacAlpine et al., 1997; Mok et al., 2001).
A general link between replication and transcription
Transcription factors appear to function by several means at the amplification
origins to modulate their activity. Both E2F1 and Myb have been shown to interact with
ORC (Beall et al., 2002; Bosco et al., 2001). Possibly with some degree of redundancy,
they recruit ORC through direct interaction. Transcription factors may also directly
recruit proteins to modify chromatin structure to facilitate the assembly of the replication
machinery. It is probably not a pure coincidence, however, that some common chromatin

37
features are shared by active transcription and replication, considering the ultimate goal
of gene amplification is to augment transcript level.
There is mounting evidence for a general link between transcription and
replication. Replication origins are frequently found upstream of transcription units
(Mechali, 2001), and there are several examples in which they coincide with promoter
sequences (Kohzaki and Murakami, 2005). In the human β-globin and c-myc replicons,
transcription regulatory elements have been shown to be essential for replication
initiation (Aladjem et al., 1995; Liu et al., 2003). In addition to studies of specific
replication sites, genome-wide mapping of replication origins in eukaryotes has been
facilitated by recent advances in DNA microarray technology, and has begun to establish
the spatial and temporal program of replication initiation (MacAlpine and Bell, 2005).
Microarray analyses of genomic replication in Drosophila and human cells show a
correlation between regions undergoing active transcription and early replication (Jeon et
al., 2005; MacAlpine et al., 2004; Schubeler et al., 2002; Woodfine et al., 2004). A more
extensive study of Drosophila chromosome 2L in Kc cells uncovered a frequent
colocalization of ORC and RNA polymerase II (RNAPII) binding sites, implying a
connection between transcription and ORC localization (MacAlpine et al., 2004).
A number of studies report positive effects of transcription factors on DNA
replication (Kohzaki and Murakami, 2005). The recruitment of transcription factors
alters origin activity on episomal plasmids in both S. cerevisiae and X. laevis eggs
(Cheng et al., 1992; Danis et al., 2004). Similarly, expression of a CREB-GAL4 fusion
protein restores replication origin activity of the mutant c-myc locus where a GAL4p

38
binding cassette replaces all regulatory sequences of the c-myc gene (Ghosh et al., 2004).
These results suggest that transcription factor binding can enhance replication origin
activity. In Chinese hamster ovary (CHO) cells the dihydrofolate reductase (DHFR) gene
locus contains a 55-kb zone of potential initiation sites of replication upstream of the
gene (Burhans et al., 1990; Vaughn et al., 1990). In mutants with parts or all of the
DHFR promoter deleted such that transcription is undetectable, initiation in the intergenic
space is markedly suppressed (but not eliminated); restoration of transcription with either
the wild-type Chinese hamster promoter or a Drosophila-based construct restores origin
activity to the wild-type pattern (Saha et al., 2004).
However, 2D gel analysis of the promoterless DHFR mutants reveals that
initiation occurrs at a low level not only in the intergenic region, but also in the body of
the DHFR gene, which had never been observed in the wild-type locus (Saha et al.,
2004). Thus transcription seems to suppress replication initiation in the body of the gene,
and help define the boundaries of the downstream origin (Saha et al., 2004). In a mutant
human c-myc locus with the c-myc promoter replaced by inducible GFP-encoding genes,
replication initiation is repressed upon induction of transcription (Ghosh et al., 2004).
When basal or induced transcription complexes is slowed by the presence of α-amanitin,
origin activity depends on the orientation of the transcription unit (Ghosh et al., 2004).
These data suggest that high levels of transcription or the persistence of transcription
complexes can repress replication initiation. Taken together, the seemingly dual role of
transcription on origin firing may be important to ensure high activity of intergenic

39
origins, while suppressing initiation within the gene body to avoid disruption of pre-RCs
by the passage of the transcriptional machinery.
Another theme of transcriptional control of origin firing is the involvement of
RNAPII. Transcription factors mediate the enhancer --> activator--> mediator -->
RNAPII --> promoter pathway to initiate mRNA transcription via RNAPII in virtually all
eukaryotes (Kornberg, 2005). In Chinese hamster ovary cells it has been reported that
inhibition of RNAPII transcription by α-amanitin results in deregulation of replication
initiation at the DHFR locus (Kornberg, 2005). In Sciara salivary puff II/9A, although
transcription of the II-9-1 gene does not begin until amplification is complete, the
promoter of II-9-1 is occupied by RNAPII during amplification stages, and it is this
presence that is thought to limit the right-hand boundary of the initiation zone during
amplification (Sasaki et al., 2006). Furthermore, a direct physical interaction has been
reported between RNAPII and MCM2-7 in yeast (Gauthier et al., 2002; Holland et al.,
2002), raising the possibility that the transcriptional machinery serves to load the MCM
complex to origins in some developmental contexts. Recently the hyperphosphorylated
form of RNAPII implicated in transcriptional elongation has been shown to co-
immunoprecipitate with DNA polymerase ε (Rytkonen et al., 2006).
In addition to direct interactions with replication proteins, RNAPII has been
shown to be required for histone displacement within the transcriptionally activated
gene’s coding region preceding RNAPII (Brown and Kingston, 1997; Lee et al., 2004;
Schwabish and Struhl, 2004; Zhao et al., 2005). In the human hsp70 gene, transcription
activation leads to nucleosomal disassembly in the first 400 bp coding sequence in front

40
of RNAPII (Brown and Kingston, 1997). More recently, it has been demonstrated that
histone density throughout the entire Saccharomyces cerevisiae GAL10 coding region is
inversely correlated with RNAPII association and transcriptional activity, suggesting
efficient eviction of core histones from the DNA by transcription (Schwabish and Struhl,
2004). Additionally, MCM2-7 associated DNA is more susceptible to nuclease digestion,
indicating that these chromatin domains may be less tightly compacted, although the
causal and consequence relation is not clear (Forsburg, 2004; Holthoff et al., 1998;
Richter et al., 1998). Finally, the elongating form of RNAPII is in association with
chromatin remodeling and histone-modifying factors (Sims et al., 2004). All together,
these physical interactions between promoters, the transcriptional machinery, factors
regulating chromatin structures, replication proteins and finally replication and
amplification origins suggest a complex picture of transcription and replication regulation
in the chromatin context.
Summary of thesis
This thesis work investigated mechanisms that control the unique timing of
DAFC-62D origin activation using cytological, molecular and genetic methods. We first
defined the origin sequences in DAFC-62D by analyzing the amount of nascent
replicative DNA across this amplicon. Surprisingly the origin coincided with the coding
region of a 62D gene. ORC2 localized to the origin, as well as two other sites that did not
confer origin activity. Both ORC2 and MCM2-7 displayed differential association with
these sequences, corresponding to the two rounds of amplification in two separate

41
developmental stages (10 and 13). All three elements, dispersed in a 7kb central
amplified region, were required for either round of DAFC-62D amplification, because
deleting any one completely abolished amplification in transposon experiments. Preceded
by transcription of the 62D gene in stage 12, the late round of origin firing was ablated by
the RNAPII inhibitor α-amanitin. This effect was absent from other amplicons and
insulated transposons, and specific to the stage 13 amplification at DAFC-62D and
transposons that did not have functional insulators. Finally, blocking RNAPII
transcription compromised MCM2-7 recruitment as suggested by ChIP analysis.
Our studies of the regulation of DAFC-62D yield several unexpected findings. We
find that the positioning of ORC and MCM2-7 can be affected by differentiation stage.
Transcription via RNAPII in cis controls localization of replication factors and origin
activation. The comparative analyses of DAFC-62D and -66D demonstrate that there are
distinct mechanisms for differential regulation of amplification origins during Drosophila
follicle cell development. Transposon experiments suggest their distinct behavior than the
endogenous amplicon may be accounted for by the insulators’ unique properties.
Together our findings provide critical insights into how metazoan DNA replication is
controlled in response to developmental cues.

42
REFERENCES
Aggarwal, B. D., and Calvi, B. R. (2004). Chromatin regulates origin activity inDrosophila follicle cells. Nature 430, 372-376.Aladjem, M. I., Groudine, M., Brody, L. L., Dieken, E. S., Fournier, R. E., Wahl, G. M.,and Epner, E. M. (1995). Participation of the human beta-globin locus control region ininitiation of DNA replication. Science 270, 815-819.Aparicio, J. G., Viggiani, C. J., Gibson, D. G., and Aparicio, O. M. (2004). The Rpd3-Sin3 histone deacetylase regulates replication timing and enables intra-S origin control inSaccharomyces cerevisiae. Mol Cell Biol 24, 4769-4780.Aparicio, O. M., Weinstein, D. M., and Bell, S. P. (1997). Components and dynamics ofDNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45pduring S phase. Cell 91, 59-69.Austin, R. J., Orr-Weaver, T. L., and Bell, S. P. (1999). Drosophila ORC specificallybinds to ACE3, an origin of DNA replication control element. Genes Dev 13, 2639-2649.Beall, E. L., Bell, M., Georlette, D., and Botchan, M. R. (2004). Dm-myb mutant lethalityin Drosophila is dependent upon mip130: positive and negative regulation of DNAreplication. Genes Dev 18, 1667-1680.Beall, E. L., Manak, J. R., Zhou, S., Bell, M., Lipsick, J. S., and Botchan, M. R. (2002).Role for a Drosophila Myb-containing protein complex in site-specific DNA replication.Nature 420, 833-837.Bell, S. P. (2002). The origin recognition complex: from simple origins to complexfunctions. Genes Dev 16, 659-672.Bell, S. P., and Dutta, A. (2002). DNA replication in eukaryotic cells. Annu RevBiochem 71, 333-374.Bielinsky, A. K., Blitzblau, H., Beall, E. L., Ezrokhi, M., Smith, H. S., Botchan, M. R.,and Gerbi, S. A. (2001). Origin recognition complex binding to a metazoan replicationorigin. Curr Biol 11, 1427-1431.Bielinsky, A. K., and Gerbi, S. A. (1998). Discrete start sites for DNA synthesis in theyeast ARS1 origin. Science 279, 95-98.Bielinsky, A. K., and Gerbi, S. A. (1999). Chromosomal ARS1 has a single leadingstrand start site. Mol Cell 3, 477-486.Bienz-Tadmor, B., Smith, H. S., and Gerbi, S. A. (1991). The promoter of DNA puffgene II/9-1 of Sciara coprophila is inducible by ecdysone in late prepupal salivary glandsof Drosophila melanogaster. Cell Regul 2, 875-888.Bosco, G., Du, W., and Orr-Weaver, T. L. (2001). DNA replication control throughinteraction of E2F-RB and the origin recognition complex. Nat Cell Biol 3, 289-295.Brown, D. D., and Dawid, I. B. (1968). Specific gene amplification in oocytes. Science160, 272-280.Brown, S. A., and Kingston, R. E. (1997). Disruption of downstream chromatin directedby a transcriptional activator. Genes Dev 11, 3116-3121.

43
Burhans, W. C., Vassilev, L. T., Caddle, M. S., Heintz, N. H., and DePamphilis, M. L.(1990). Identification of an origin of bidirectional DNA replication in mammalianchromosomes. Cell 62, 955-965.Buszczak, M., Freeman, M. R., Carlson, J. R., Bender, M., Cooley, L., and Segraves, W.A. (1999). Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 126, 4581-4589.Calvi, B. R., Lilly, M. A., and Spradling, A. C. (1998). Cell cycle control of chorion geneamplification. Genes Dev 12, 734-744.Capelson M., and Corces, V. G. (2004). Boundary elements and nuclear organization.Biol Cell 96, 617-629.Carminati, J. L., Johnston, C. G., and Orr-Weaver, T. L. (1992). The Drosophila ACE3chorion element autonomously induces amplification. Mol Cell Biol 12, 2444-2453.Carney, G. E., and Bender, M. (2000). The Drosophila ecdysone receptor (EcR) gene isrequired maternally for normal oogenesis. Genetics 154, 1203-1211.Cayirlioglu, P., Bonnette, P. C., Dickson, M. R., and Duronio, R. J. (2001). DrosophilaE2f2 promotes the conversion from genomic DNA replication to gene amplification inovarian follicle cells. Development 128, 5085-5098.Cayirlioglu, P., Ward, W. O., Silver Key, S. C., and Duronio, R. J. (2003).Transcriptional repressor functions of Drosophila E2F1 and E2F2 cooperate to inhibitgenomic DNA synthesis in ovarian follicle cells. Mol Cell Biol 23, 2123-2134.Cheng, L. Z., Workman, J. L., Kingston, R. E., and Kelly, T. J. (1992). Regulation ofDNA replication in vitro by the transcriptional activation domain of GAL4-VP16. ProcNatl Acad Sci U S A 89, 589-593.Cherbas, L., Hu, X., Zhimulev, I., Belyaeva, E., and Cherbas, P. (2003). EcR isoforms inDrosophila: testing tissue-specific requirements by targeted blockade and rescue.Development 130, 271-284.Christianson, A. M., King, D. L., Hatzivassiliou, E., Casas, J. E., Hallenbeck, P. L.,Nikodem, V. M., Mitsialis, S. A., and Kafatos, F. C. (1992). DNA binding andheteromerization of the Drosophila transcription factor chorion factor 1/ultraspiracle.Proc Natl Acad Sci U S A 89, 11503-11507.Claycomb, J. M., Benasutti, M., Bosco, G., Fenger, D. D., and Orr-Weaver, T. L. (2004).Gene amplification as a developmental strategy: isolation of two developmentalamplicons in Drosophila. Dev Cell 6, 145-155.Claycomb, J. M., MacAlpine, D. M., Evans, J. G., Bell, S. P., and Orr-Weaver, T. L.(2002). Visualization of replication initiation and elongation in Drosophila. J Cell Biol159, 225-236.Claycomb, J. M., and Orr-Weaver, T. L. (2005). Developmental gene amplification:insights into DNA replication and gene expression. Trends Genet 21, 149-162.Clever, U., and Ellgaard, E. G. (1970). Puffing and histone acetylation in polytenechromosomes. Science 169, 373-374.Crouse, H. V. (1968). The role of ecdysone in DNA-puff formation and DNA synthesisin the polytene chromosomes of Sciara coprophila. Proc Natl Acad Sci U S A 61, 971-978.

44
Cvetic, C., and Walter, J. C. (2005). Eukaryotic origins of DNA replication: could youplease be more specific? Semin Cell Dev Biol 16, 343-353.Danis, E., Brodolin, K., Menut, S., Maiorano, D., Girard-Reydet, C., and Mechali, M.(2004). Specification of a DNA replication origin by a transcription complex. Nat CellBiol 6, 721-730.de Cicco, D. V., and Spradling, A. C. (1984). Localization of a cis-acting elementresponsible for the developmentally regulated amplification of Drosophila chorion genes.Cell 38, 45-54.Delidakis, C., and Kafatos, F. C. (1989). Amplification enhancers and replication originsin the autosomal chorion gene cluster of Drosophila. Embo J 8, 891-901.DePamphilis, M. L. (1999). Replication origins in metazoan chromosomes: fact orfiction? Bioessays 21, 5-16.DePamphilis, M. L., Blow, J. J., Ghosh, S., Saha, T., Noguchi, K., and Vassilev, A.(2006). Regulating the licensing of DNA replication origins in metazoa. Curr Opin CellBiol 18, 231-239.DiBartolomeis, S. M., and Gerbi, S. A. (1989). Molecular characterization of DNA puffII/9A genes in Sciara coprophila. J Mol Biol 210, 531-540.Dijkwel, P. A., Wang, S., and Hamlin, J. L. (2002). Initiation sites are distributed atfrequent intervals in the Chinese hamster dihydrofolate reductase origin of replication butare used with very different efficiencies. Mol Cell Biol 22, 3053-3065.Dyson, N. (1998). The regulation of E2F by pRB-family proteins. Genes Dev 12, 2245-2262.Forsburg, S. L. (2004). Eukaryotic MCM proteins: beyond replication initiation.Microbiol Mol Biol Rev 68, 109-131.Foulk, M. S., Liang, C., Wu, N., Blitzblau, H. G., Smith, H., Alam, D., Batra, M., andGerbi, S. A. (2006). Ecdysone induces transcription and amplification in Sciaracoprophila DNA puff II/9A. Dev Biol 299, 151-163.Frolov, M. V., Huen, D. S., Stevaux, O., Dimova, D., Balczarek-Strang, K., Elsdon, M.,and Dyson, N. J. (2001). Functional antagonism between E2F family members. GenesDev 15, 2146-2160.Fukuda, H., Sano, N., Muto, S., and Horikoshi, M. (2006). Simple histone acetylationplays a complex role in the regulation of gene expression. Brief Funct GenomicProteomic 5, 190-208.Gall, J. G. (1968). Differential synthesis of the genes for ribosomal RNA duringamphibian oogenesis. Proc Natl Acad Sci USA 60, 553-560.Gall, J. G. (1974). Free ribosomal RNA genes in the macronucleus of Tetrahymena. ProcNatl Acad Sci U S A 71, 3078-3081.Gauthier, L., Dziak, R., Kramer, D. J., Leishman, D., Song, X., Ho, J., Radovic, M.,Bentley, D., and Yankulov, K. (2002). The role of the carboxyterminal domain of RNApolymerase II in regulating origins of DNA replication in Saccharomyces cerevisiae.Genetics 162, 1117-1129.Gerasimova, T. I., and Corces, V. G. (2001). Chromatin insulators and boundaries:effects on transcription and nuclear organization. Annu Rev Genet 35, 193-208.

45
Gerbi, S. A. (2005). Mapping origins of DNA replication in eukaryotes. Methods MolBiol 296, 167-180.Gerbi, S. A., and Bielinsky, A. K. (1997). Replication initiation point mapping. Methods13, 271-280.Ghosh, M., Liu, G., Randall, G., Bevington, J., and Leffak, M. (2004). Transcriptionfactor binding and induced transcription alter chromosomal c-myc replicator activity.Mol Cell Biol 24, 10193-10207.Giacca, M., Zentilin, L., Norio, P., Diviacco, S., Dimitrova, D., Contreas, G., Biamonti,G., Perini, G., Weighardt, F., Riva, S., and et al. (1994). Fine mapping of a replicationorigin of human DNA. Proc Natl Acad Sci U S A 91, 7119-7123.Giacinti, C., and Giordano, A. (2006). RB and cell cycle progression. Oncogene 25,5220-5227.Gilbert, D. M. (2001). Making sense of eukaryotic DNA replication origins. Science 294,96-100.Gilbert, D. M. (2004). In search of the holy replicator. Nat Rev Mol Cell Biol 5, 848-855.Glover, D. M., Zaha, A., Stocker, A. J., Santelli, R. V., Pueyo, M. T., De Toledo, S. M.,and Lara, F. J. S. (1982). Gene amplification in Rhynchosciara salivary glandchromosomes. Proc Natl Acad Sci USA 79, 2947-2951.Groth, A., Rocha, W., Verreault, A., and Almouzni, G. (2007). Chromatin challengesduring DNA replication and repair. Cell 128, 721-733.Hackney, J. F., Pucci, C., Naes, E., and Dobens, L. (2007). Ras signaling modulatesactivity of the ecdysone receptor EcR during cell migration in the Drosophila ovary. DevDyn 236, 1213-1226.Hartl, T., Boswell, C., Orr-Weaver, T. L., and Bosco, G. (2007). Developmentallyregulated histone modifications in Drosophila follicle cells: initiation of geneamplification is associated with histone H3 and H4 hyperacetylation and H1phosphorylation. Chromosoma.Heck, M. M., and Spradling, A. C. (1990). Multiple replication origins are used duringDrosophila chorion gene amplification. J Cell Biol 110, 903-914.Henderson, D. S., Wiegand, U. K., Norman, D. G., and Glover, D. M. (2000). Mutualcorrection of faulty PCNA subunits in temperature-sensitive lethal mus209 mutants ofDrosophila melanogaster. Genetics 154, 1721-1733.Holland, L., Gauthier, L., Bell-Rogers, P., and Yankulov, K. (2002). Distinct parts ofminichromosome maintenance protein 2 associate with histone H3/H4 and RNApolymerase II holoenzyme. Eur J Biochem 269, 5192-5202.Holthoff, H. P., Baack, M., Richter, A., Ritzi, M., and Knippers, R. (1998). Humanprotein MCM6 on HeLa cell chromatin. J Biol Chem 273, 7320-7325.Hourcade, D., Dressler, D., and Wolfson, J. (1973). The amplification of ribosomal RNAgenes involves a rolling circle intermediate. Proc Natl Acad Sci U S A 70, 2926-2930.Jacob, F., and Brenner, S. (1963). [On the regulation of DNA synthesis in bacteria: thehypothesis of the replicon.]. C R Hebd Seances Acad Sci 256, 298-300.

46
Jeon, Y., Bekiranov, S., Karnani, N., Kapranov, P., Ghosh, S., MacAlpine, D., Lee, C.,Hwang, D. S., Gingeras, T. R., and Dutta, A. (2005). Temporal profile of replication ofhuman chromosomes. Proc Natl Acad Sci U S A 102, 6419-6424.Kapler, G. M., Dobbs, D. L., and Blackburn, E. (1996). DNA replication in Tetrahymena.In DNA replication in eukaryotic cells, M. L. DePamphilis, ed. (Cold Spring Harbor,N.Y., Cold Spring Harbor Laboratory Press), pp. 915-932.Kobayashi, T., Rein, T., and DePamphilis, M. L. (1998). Identification of primaryinitiation sites for DNA replication in the hamster dihydrofolate reductase gene initiationzone. Mol Cell Biol 18, 3266-3277.Kohzaki, H., and Murakami, Y. (2005). Transcription factors and DNA replication originselection. Bioessays 27, 1107-1116.Korenjak, M., Taylor-Harding, B., Binne, U. K., Satterlee, J. S., Stevaux, O., Aasland, R.,White-Cooper, H., Dyson, N., and Brehm, A. (2004). Native E2F/RBF complexescontain Myb-interacting proteins and repress transcription of developmentally controlledE2F target genes. Cell 119, 181-193.Kornberg, R. D. (2005). Mediator and the mechanism of transcriptional activation.Trends Biochem Sci 30, 235-239.Labib, K., Kearsey, S. E., and Diffley, J. F. (2001). MCM2-7 proteins are essentialcomponents of prereplicative complexes that accumulate cooperatively in the nucleusduring G1-phase and are required to establish, but not maintain, the S-phase checkpoint.Mol Biol Cell 12, 3658-3667.Laicine, E. M., Alves, M. A., de Almeida, J. C., Rizzo, E., Albernaz, W. C., and Sauaia,H. (1984). Development of DNA puffs and patterns of polypeptide synthesis in thesalivary glands of Bradysia hygida. Chromosoma 89, 280-284.Landis, G., Kelley, R., Spradling, A. C., and Tower, J. (1997). The k43 gene, required forchorion gene amplification and diploid cell chromosome replication, encodes theDrosophila homolog of yeast origin recognition complex subunit 2. Proc Natl Acad SciUSA 94, 3888-3892.Landis, G., and Tower, J. (1999). The drosophila chiffon gene is required for choriongene amplification, and is related to the yeast dbf4 regulator of DNA replication and cellcycle. Development 126, 4281-4293.Lee, C. K., Shibata, Y., Rao, B., Strahl, B. D., and Lieb, J. D. (2004). Evidence fornucleosome depletion at active regulatory regions genome-wide. Nat Genet 36, 900-905.Levine, J., and Spradling, A. (1985). DNA sequence of a 3.8 kilobase pair regioncontrolling Drosophila chorion gene amplification. Chromosoma 92, 136-142.Lewis, P. W., Beall, E. L., Fleischer, T. C., Georlette, D., Link, A. J., and Botchan, M. R.(2004). Identification of a Drosophila Myb-E2F2/RBF transcriptional repressor complex.Genes Dev 18, 2929-2940.Liang, C., and Gerbi, S. A. (1994). Analysis of an origin of DNA amplification in Sciaracoprophila by a novel three-dimensional gel method. Mol Cell Biol 14, 1520-1529.Liang, C., Spitzer, J. D., Smith, H. S., and Gerbi, S. A. (1993). Replication initiates at aconfined region during DNA amplification in Sciara DNA puff II/9A. Genes and Dev 7,1072-1084.

47
Little, R. D., Platt, T. H., and Schildkraut, C. L. (1993). Initiation and termination ofDNA replication in human rRNA genes. Mol Cell Biol 13, 6600-6613.Liu, G., Malott, M., and Leffak, M. (2003). Multiple functional elements comprise aMammalian chromosomal replicator. Mol Cell Biol 23, 1832-1842.Loden, M., and van Steensel, B. (2005). Whole-genome views of chromatin structure.Chromosome Res 13, 289-298.Lu, L., and Tower, J. (1997). A transcriptional insulator element, the su(Hw) binding site,protects a chromosomal DNA replication origin from position effects. Mol Cell Biol 17,2202-2206.Lu, L., Zhang, H., and Tower, J. (2001). Functionally distinct, sequence-specificreplicator and origin elements are required for Drosophila chorion gene amplification.Genes Dev 15, 134-146.MacAlpine, D. M., Rodriguez, H. K., and Bell, S. P. (2004). Coordination of replicationand transcription along a Drosophila chromosome. Genes Dev 18, 3094-3105.MacAlpine, D. M., Zhang, Z., and Kapler, G. M. (1997). Type I elements mediatereplication fork pausing at conserved upstream sites in the Tetrahymena thermophilaribosomal DNA minichromosome. Mol Cell Biol 17, 4517-4525.Mechali, M. (2001). DNA replication origins: from sequence specificity to epigenetics.Nat Rev Genet 2, 640-645.Mohammad, M., Saha, S., and Kapler, G. M. (2000). Three different proteins recognize amultifunctional determinant that controls replication initiation, fork arrest andtranscription in Tetrahymena. Nucleic Acids Res 28, 843-851.Mok, E. H., Smith, H. S., DiBartolomeis, S. M., Kerrebrock, A. W., Rothschild, L. J.,Lange, T. S., and Gerbi, S. A. (2001). Maintenance of the DNA puff expanded state isindependent of active replication and transcription. Chromosoma 110, 186-196.Newlon, C. S., and Theis, J. F. (1993). The structure and function of yeast ARS elements.Curr Opin Genet Dev 3, 752-758.Oro, A. E., McKeown, M., and Evans, R. M. (1992). The Drosophila retinoid X receptorhomolog ultraspiracle functions in both female reproduction and eye morphogenesis.Development 115, 449-462.Orr-Weaver, T. L. (1991). Drosophila chorion genes: cracking the eggshell's secrets.Bioessays 13, 97-105.Orr-Weaver, T. L., Johnston, C. G., and Spradling, A. C. (1989). The role of ACE3 inDrosophila chorion gene amplification. Embo J 8, 4153-4162.Orr-Weaver, T. L., and Spradling, A. C. (1986). Drosophila chorion gene amplificationrequires an upstream region regulating s18 transcription. Mol Cell Biol 6, 4624-4633.Prescott, D. M. (1994). The DNA of ciliated protozoa. Microbiol Rev 58, 233-267.Remus, D., Beall, E. L., and Botchan, M. R. (2004). DNA topology, not DNA sequence,is a critical determinant for Drosophila ORC-DNA binding. Embo J 23, 897-907.Richter, A., Baack, M., Holthoff, H. P., Ritzi, M., and Knippers, R. (1998). Mobilizationof chromatin-bound Mcm proteins by micrococcal nuclease. Biol Chem 379, 1181-1187.

48
Royzman, I., Austin, R. J., Bosco, G., Bell, S. P., and Orr-Weaver, T. L. (1999). ORClocalization in Drosophila follicle cells and the effects of mutations in dE2F and dDP.Genes Dev 13, 827-840.Rytkonen, A. K., Hillukkala, T., Vaara, M., Sokka, M., Jokela, M., Sormunen, R.,Nasheuer, H. P., Nethanel, T., Kaufmann, G., Pospiech, H., and Syvaoja, J. E. (2006).DNA polymerase epsilon associates with the elongating form of RNA polymerase II andnascent transcripts. Febs J 273, 5535-5549.Saha, S., Shan, Y., Mesner, L. D., and Hamlin, J. L. (2004). The promoter of the Chinesehamster ovary dihydrofolate reductase gene regulates the activity of the local origin andhelps define its boundaries. Genes Dev 18, 397-410.Sasaki, T., Ramanathan, S., Okuno, Y., Kumagai, C., Shaikh, S. S., and Gilbert, D. M.(2006). The Chinese hamster dihydrofolate reductase replication origin decision pointfollows activation of transcription and suppresses initiation of replication withintranscription units. Mol Cell Biol 26, 1051-1062.Schubeler, D., Scalzo, D., Kooperberg, C., van Steensel, B., Delrow, J., and Groudine, M.(2002). Genome-wide DNA replication profile for Drosophila melanogaster: a linkbetween transcription and replication timing. Nat Genet 32, 438-442.Schwabish, M. A., and Struhl, K. (2004). Evidence for eviction and rapid deposition ofhistones upon transcriptional elongation by RNA polymerase II. Mol Cell Biol 24,10111-10117.Shea, M. J., King, D. L., Conboy, M. J., Mariani, B. D., and Kafatos, F. C. (1990).Proteins that bind to Drosophila chorion cis-regulatory elements: a new C2H2 zinc fingerprotein and a C2C2 steroid receptor-like component. Genes Dev 4, 1128-1140.Sims, R. J., 3rd, Belotserkovskaya, R., and Reinberg, D. (2004). Elongation by RNApolymerase II: the short and long of it. Genes Dev 18, 2437-2468.Spradling, A. C. (1981). The organization and amplification of two clusters of Drosophilachorion genes. Cell 27, 193-201.Spradling, A. C., de Cicco, D. V., Wakimoto, B. T., Levine, J. F., Kalfayan, L. J., andCooley, L. (1987). Amplification of the X-linked Drosophila chorion gene clusterrequires a region upstream from the s38 chorion gene. Embo J 6, 1045-1053.Spradling, A. C., Digan, M. E., Mahowald, A. P., Scott, M., and Craig, E. A. (1980). Twoclusters of genes for major chorion proteins of Drosophila melanogaster. Cell 19, 905-914.Tower, J. (2004). Developmental gene amplification and origin regulation. Annu RevGenet 38, 273-304.Underwood, E. M., Briot, A. S., Doll, K. Z., Ludwiczak, R. L., Otteson, D. C., Tower, J.,Vessey, K. B., and Yu, K. (1990). Genetics of 51D-52A, a region containing severalmaternal-effect genes and two maternal-specific transcripts in Drosophila. Genetics 126,639-650.Vashee, S., Cvetic, C., Lu, W., Simancek, P., Kelly, T. J., and Walter, J. C. (2003).Sequence-independent DNA binding and replication initiation by the human originrecognition complex. Genes Dev 17, 1894-1908.

49
Vaughn, J. P., Dijkwel, P. A., and Hamlin, J. L. (1990). Replication initiates in a broadzone in the amplified CHO dihydrofolate reductase domain. Cell 61, 1075-1087.Vogelauer, M., Rubbi, L., Lucas, I., Brewer, B. J., and Grunstein, M. (2002). Histoneacetylation regulates the time of replication origin firing. Mol Cell 10, 1223-1233.Ward, J. G., Blomberg, P., Hoffman, N., and Yao, M. C. (1997). The intranuclearorganization of normal, hemizygous and excision-deficient rRNA genes duringdevelopmental amplification in Tetrahymena thermophila. Chromosoma 106, 233-242.Whittaker, A. J., Royzman, I., and Orr-Weaver, T. L. (2000). Drosophila double parked:a conserved, essential replication protein that colocalizes with the origin recognitioncomplex and links DNA replication with mitosis and the down-regulation of S phasetranscripts. Genes Dev 14, 1765-1776.Woodfine, K., Fiegler, H., Beare, D. M., Collins, J. E., McCann, O. T., Young, B. D.,Debernardi, S., Mott, R., Dunham, I., and Carter, N. P. (2004). Replication timing of thehuman genome. Hum Mol Genet 13, 191-202.Wu, N., Liang, C., DiBartolomeis, S. M., Smith, H. S., and Gerbi, S. A. (1993).Developmental progression of DNA puffs in Sciara coprophila: amplification andtranscription. Dev Biol 160, 73-84.Yao, M. C., Blackburn, E., and Gall, J. G. (1979). Amplification of the rRNA genes inTetrahymena. Cold Spring Harb Symp Quant Biol 43 Pt 2, 1293-1296.Yao, T. P., Segraves, W. A., Oro, A. E., McKeown, M., and Evans, R. M. (1992).Drosophila ultraspiracle modulates ecdysone receptor function via heterodimerformation. Cell 71, 63-72.Yoon, Y., Sanchez, J. A., Brun, C., and Huberman, J. A. (1995). Mapping of replicationinitiation sites in human ribosomal DNA by nascent-strand abundance analysis. Mol CellBiol 15, 2482-2489.Yue, M., Reischmann, K. P., and Kapler, G. M. (1998). Conserved cis- and trans-actingdeterminants for replication initiation and regulation of replication fork movement intetrahymenid species. Nucleic Acids Res 26, 4635-4644.Zhang, H., and Tower, J. (2004). Sequence requirements for function of the Drosophilachorion gene locus ACE3 replicator and ori-beta origin elements. Development 131,2089-2099.Zhao, J., Herrera-Diaz, J., and Gross, D. S. (2005). Domain-wide displacement ofhistones by activated heat shock factor occurs independently of Swi/Snf and is notcorrelated with RNA polymerase II density. Mol Cell Biol 25, 8985-8999.Zhu, W., Giangrande, P. H., and Nevins, J. R. (2005). Temporal control of cell cycle geneexpression mediated by E2F transcription factors. Cell Cycle 4, 633-636.

50
Chapter Two
Identification of a Drosophila Replication OriginDevelopmentally Controlled by Transcription
Fang Xie and Terry L. Orr-Weaver*
Whitehead Institute and Department of BiologyMassachusetts Institute of Technology
Cambridge, MA 02142
*Contact: [email protected]

51
Summary
We exploited developmentally induced gene amplification in Drosophila ovarian
follicle cells to identify a new metazoan origin of DNA synthesis and its cis regulatory
elements, the Drosophila Amplicon in Follicle Cells, DAFC-62D. At DAFC-62D thereplication proteins ORC2 and MCM2-7 are localized onto DNA at developmental stage-
specific binding sites. Replication initiation at DAFC-62D late in follicle cell
differentiation is preceded by transcription, and we show by α-amanitin inhibition it
requires RNA polymerase II transcription in cis to localize MCM2-7. Transposons withthe DAFC-62D replication elements bounded by chromatin insulators are resistant to α-
amanitin repression provided the Su(Hw) protein is functional. These results reveal one
mechanism for initiation of metazoan DNA replication: recruitment of MCM2-7 by RNApolymerase II transcription.

52
INTRODUCTION
Proper regulation of the initiation of DNA replication is crucial for cell division in
eukaryotes. The first step of initiation is the selection of origins by the pre-replicative
complex (pre-RC) (Bell and Dutta, 2002; Mendez and Stillman, 2003). Components of
the pre-RC are sequentially recruited to origin DNA (Bell and Dutta, 2002). The six-
subunit origin recognition complex (ORC) first binds and subsequently loads Cdc6, Cdt1
and the replicative helicase MCM2-7. Following selective binding of pre-RC in G1,
origins are activated by additional kinases and factors as cells enter S phase (Bell and
Dutta, 2002). Although the protein factors appear to be highly conserved, the DNA
sequences that define origin activity in different organisms are not (Cvetic and Walter,
2005). In the budding yeast Saccharomyces cerevisiae the well-defined autonomously
replicating sequences (ARS) are specifically recognized by ORC (Bell and Stillman,
1992; Lee and Bell, 1997). By contrast, in vitro studies in higher eukaryotes suggest that
the metazoan ORC does not rely on sequence specificity to bind DNA (Remus et al.,
2004; Vashee et al., 2003).
With recent advances in DNA microarray technology, genome-wide mapping of
replication origins in S. cerevisiae and higher eukaryotes has begun to establish the
spatial and temporal program of replication initiation (MacAlpine and Bell, 2005).
However, the mechanisms of origin selection, especially in response to developmental
cues in metazoans remain poorly understood. The reasons are at least two fold. First, only
a handful of model metazoan replicons have been studied in detail (Cvetic and Walter,
2005; Gerbi, 2005). Results from mammalian cell culture systems have suggested the

53
existence of two classes of mammalian origins: large initiation zones and localized
replicators (Gilbert, 2001; Gilbert, 2004). Second, partially due to the lack of
multicellular models, few observations of cell-type specific or developmental regulation
of replication origins have been reported (Gilbert, 2005; Norio, 2006).
Developmental gene amplification in the ovarian follicle cells of Drosophila
provides a powerful system for the study of metazoan DNA replication, and permits
analysis of developmental regulation of origin firing (Calvi and Spradling, 1999;
Claycomb and Orr-Weaver, 2005; Tower, 2004). At stage 9 of egg chamber development
the somatic follicle cells surrounding the developing oocyte cease genomic DNA
replication and begin to specifically amplify four clusters of genes across the genome
(Claycomb et al., 2004). The biological purpose is to provide high levels of DNA
templates for transcription to rapidly construct the eggshell chorion (Orr-Weaver, 1991).
Amplification occurs by repeated rounds of origin firing and bidirectional movement of
replication forks from these origins to produce 100kb gradients of amplified DNA
(Claycomb and Orr-Weaver, 2005). This process depends on the same replication
initiation and elongation proteins that are necessary for genomic replication (Calvi and
Spradling, 1999; Claycomb and Orr-Weaver, 2005; Tower, 2004).
A wide range of experimental tools is available to study amplification. The
amplified regions in the follicle cells, DAFCs, can be visualized as foci of BrdU
incorporation (Calvi et al., 1998). Immunofluorescence of ORC, DUP/Cdt1, MCM2-7,
Cdc45 and PCNA (Asano and Wharton, 1999; Claycomb et al., 2002; Loebel et al., 2000;
Royzman et al., 1999), and chromatin immunoprecipitation (ChIP) of ORC (Austin et al.,

54
1999) have shown specific association of replication proteins with DAFCs. Amplification
gradients and the developmental timing of copy number increases have been accurately
constructed by real-time PCR (Claycomb et al., 2004; Claycomb et al., 2002). P-element
mediated transformation experiments, recently facilitated by the use of insulators to
buffer transposons from chromosomal position effects (Lu and Tower, 1997), have
allowed fine dissection of cis regulatory elements for amplification, and established two
types of control elements. In the well-characterized 3rd chromosome chorion amplicon,
DAFC-66D, repeated firing occurs preferentially from oriβ, the origin element (Delidakis
and Kafatos, 1989; Heck and Spradling, 1990). A ~320bp amplification control element
(ACE) on the 3rd chromosome (ACE3) also is necessary for amplification, by stimulating
replication from proximal origins (Carminati et al., 1992; Lu et al., 2001). Moreover,
ACE3 provides the developmental specificity for amplification, acting to load ORC,
which appears to localize broadly across the amplicon, rather than strictly to the origin
(Austin et al., 1999; Zhang and Tower, 2004).
A newly identified amplicon, DAFC-62D, differs in its developmental timing
from the other DAFCs, providing the opportunity to decipher how origin firing is
influenced by differentiation events (Claycomb et al., 2004). In the other amplicons
origin firing occurs only in stages 10B and 11, followed by elongation of previously
formed replication forks, without any more initiation events during subsequent stages of
follicle cell development (Claycomb et al., 2004; Claycomb et al., 2002). DAFC-66D
undergoes about 5 rounds of origin activation to give an amplification level of 30-40 fold
at the origin (Claycomb et al., 2002). At DAFC-62D amplification initiates only once in

55
stage 10B, but in stage 13 there is an additional increase in copy number at a very precise
region (Claycomb et al., 2004). We therefore investigated mechanisms that control the
unique timing of DAFC-62D origin activation. Using cytological, molecular and genetic
methods, here we define origin sequences in DAFC-62D and additional cis regulatory
elements that are required for the developmental control of origin firing. Unexpectedly,
we find that amplification at DAFC-62D in late follicle cell differentiation depends on
transcription in cis.
RESULTS
Identification of the replication origin and ORC binding sites in DAFC-62D
To determine the site at which DNA synthesis initiates during amplification at
DAFC-62D, nascent strand analysis was performed as described (Giacca and Zentilin,
1994; Kobayashi et al., 1998b). Genomic DNA was isolated from stage 10B egg
chambers and subjected to benzoylated naphthoylated DEAE-cellulose column
chromatography, to enrich for replicative intermediate DNA molecules that are single-
stranded. Extensive λ-exonuclease treatment further purified nascent strands, because the
presence of RNA primers protects these molecules from digestion. Nascent DNA was
then size fractionated, and the levels of specific sequences in each fraction were
quantified by real-time PCR. We observed a 1kb region highly enriched in the 0.5-1kb
(Figure 1A) and 1-1.6kb (data not shown) fraction of nascent DNA, thus containing
origin activity. We have designated this region as ori62. As a control for the λ-
exonuclease digestion and uniform efficiency of PCR, DNA of size 5kb and above that is

56
not expected to contain nascent strands displayed uniformly low levels across DAFC-62D
(Figure 1A). As a positive control, we found that the known origin oriβ of DAFC-66D
was enriched in the 0.5-1kb fraction about 14-fold over a locus 5kb away (data not
shown). We also attempted to map the origin used for amplification in stage 13, but the
high levels of single-stranded DNA from apoptotic nurse cells precluded complete λ-
exonuclease digestion, creating a high background signal in the PCR reactions.
In S. cerevisiae, ORC is in close contact with the origin (Lee and Bell, 1997).
ORC also binds to key replication elements in the Sciara salivary gland amplicon DNA
puff II/9A (Bielinsky et al., 2001; Lunyak et al., 2002). A hypomorphic, female-sterile
mutation in the Drosophila orc2 gene causes a thin-eggshell phenotype due to reduced
levels of amplification of the chorion gene clusters (Landis et al., 1997b). Previous
immunofluorescence experiments have shown that ORC localizes to amplified regions
through stage 10A to 11, but it is not detectable after replication initiation has ceased at
DAFC-66D (Claycomb et al., 2002; Royzman et al., 1999). Further in vivo and in vitro
analyses demonstrated association of ORC in stage 10 with sequences required for
chorion gene amplification (Austin et al., 1999).
We used chromatin immunoprecipitation (ChIP) with antibodies against the
ORC2 subunit to test whether ORC is present at ori62 (Austin et al., 1999). As a positive
control, the presence of ORC at ACE3 was examined and found enriched over the actin
control in ChIP DNA from stage 10A but not stages 12-13 (Figure 1B). Real-time PCR
quantification also showed a ten-fold enrichment of ORC-bound ACE3 over another
nonamplified control locus on chromosome arm 3R (62C5) described in

57
Figure 1. Determination of the amplification origin and its association with ORC
(A) Nascent strand analysis across DAFC-62D. Size-fractionated nascent DNA collected
from stage 10B egg chambers was quantified over known serial standards using real-time
PCR. Abundance of nascent DNA (Y axis, normalized to arbitrary standards) in the 0.5-
1kb and 5kb above fraction (control) is shown. Numbers on the X-axis are relative
distance away from the central region in kilobases. + and − indicate orientation. The 1kb
fragment that confers origin activity is named ori62. Error bars are standard deviations
(SD) of triplicate PCR reactions. (B) Real-time PCR analysis of ChIP showing
enrichment level (±SD) of ORC2 at ACE3 and a locus 5.0 kb away over a control locus at
62C5. ACE3 is specifically pulled down from stage 10A follicular DNA but not later
stages. (C) Real-time PCR analysis of anti-ORC2 ChIP at ori62 over the same 62C5
control in different stages.
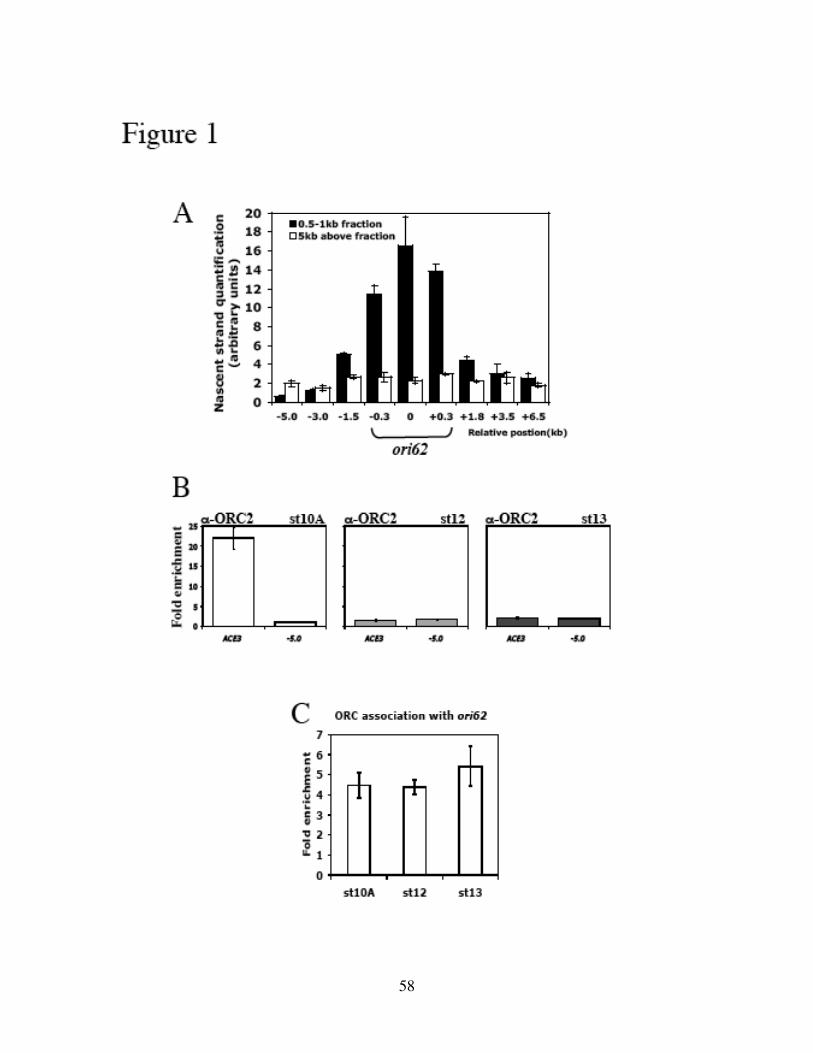
58

59
(Claycomb et al., 2002) (data not shown). The reduction in levels of ORC at ACE3 in
stages 12 and 13 correlates with the failure to detect foci of ORC localization by
immunofluorescence after stage 11 (Claycomb et al., 2002). In DAFC-62D, we detected
significant localization of ORC to ori62 by ChIP and real-time PCR quantification
(Figure 1C). In contrast to ACE3, ORC binding remained present in stages 12 and 13 at
ori62, paralleling the fact that an additional round of amplification takes place at DAFC-
62D in stage 13 (Claycomb et al., 2004).
Differential pre-RC binding in DAFC-62D
In DAFC-66D ORC binds to both ACE3 and oriβ, and the requirements for an
adequate amount of chorion amplicon DNA sequences to detect ORC binding by
immunofluorescence suggested that ORC additionally binds to multiple sites in the
amplicon (Zhang and Tower, 2004). Thus we wanted to test if ORC was present at sites
in DAFC-62D in addition to ori62. Moreover, given the two developmental time points
for amplification initiation at DAFC-62D, we also investigated whether the pattern of
ORC binding changed during follicle cell differentiation. ChIP on stage 10A, 12 and 13
egg chamber DNA suggested the binding of ORC to ori62, and also to a site about 3kb
away (–3.0) (Figure 2A, B). Another site, 3.5kb away on the opposite side of ori62, is
bound by ORC as well, but only in stage 10A (Figure 2A, B). Therefore ORC
differentially localizes to three sites at DAFC-62D, remaining associated with two of
them (ori62 and –3.0) from stage 10A on (Figure 2B).

60
Figure 2. Differential binding of pre-RC at DAFC-62D
(A). Quantitative (real-time) PCR analysis of anti-ORC2 ChIP (±SD) across DAFC-62D.
ORC2 association pattern differentially changes from stage 10A, stage 12 to stage 13 of
follicle cell development. Numbers on the X axis are relative distance away from the
center of ori62 (in kb). (B) Diagram of the 10 kb central amplified fragment in DAFC-
62D showing stage-specific ORC binding sites and the position of the single annotated
gene yellow-g2. (C) Differential MCM ChIP (±SD) in stages 10A through 13 at DAFC-
62D (upper panels) and DAFC-66D (lower panels).
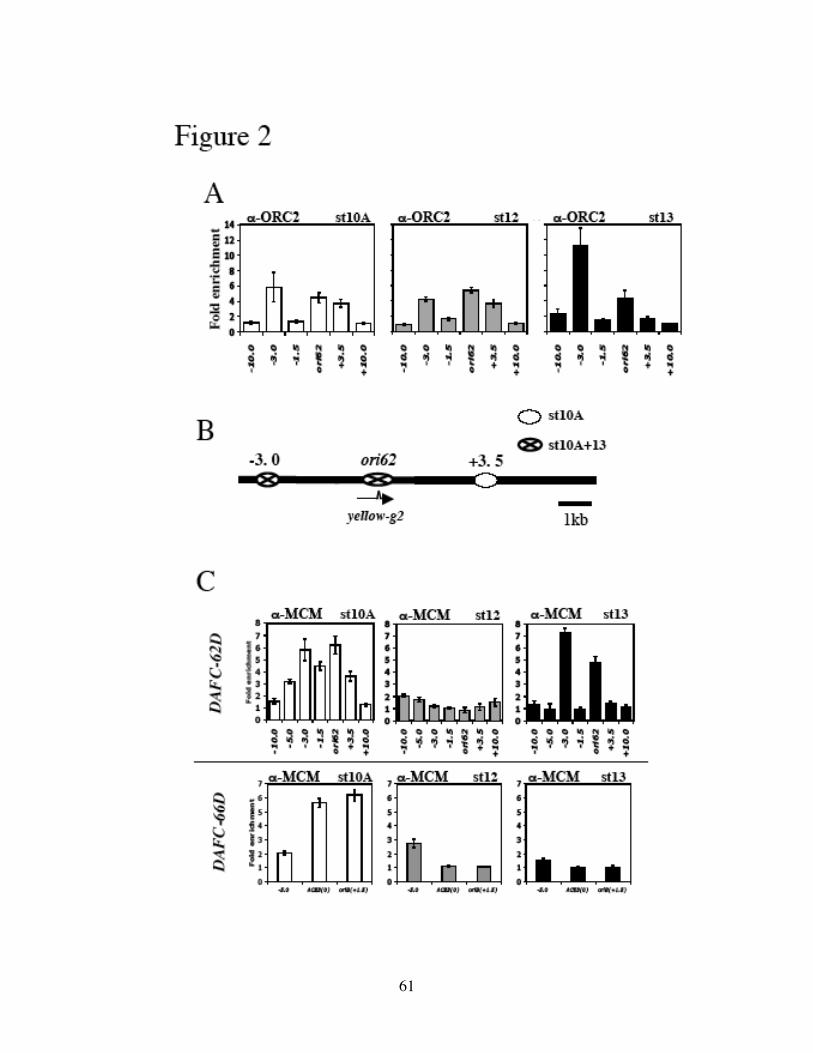
61

62
We also observed by ChIP that the MCM complex was broadly localized around
ori62 in stage 10 (Figure 2C, upper left panel), reflecting its dual role in replication
initiation and elongation. In stage 12 MCM2-7 disassociated from the origin (Figure 2C,
upper middle panel) although ORC remained bound (Figure 2A). Strikingly, the MCM
complex was reloaded to ori62 and –3.0 in stage 13 (Figure 2C, upper right panel). In
contrast, at DAFC-66D MCM2-7 associated with ACE3 and oriβ in stage 10 but not
afterwards (Figure 2C, lower panels), paralleling the binding pattern of ORC (Figure 1B).
We concluded that at DAFC-62D there is developmentally regulated pre-RC binding that
utilizes different cis-acting elements to direct origin firing in different stages (Figure 2B).
ORC-binding sequences are required for amplification
We used P-element mediated transformation to test the function of the cis
elements that associate with the pre-RC in vivo. Upon integration into ectopic sites,
transposons will amplify provided proper sequences are present, as demonstrated by
experiments on DAFC-66D and -7F (de Cicco and Spradling, 1984; Spradling et al.,
1987). Chromosomal position effects that affect levels of amplification can be buffered
away by flanking transposons with insulators (Suppressor of Hairy-wing binding sites
(SHWBS) (Lu and Tower, 1997). Using this system we found that in two out of two
transformant lines carrying the 1kb ori62 fragment the transposons did not amplify
(Figure 3A), indicating the requirement for additional sequences such as enhancer-like
elements. In contrast, a tranposon containing ori62 in cis with ACE3, the known control
element in DAFC-66D, underwent amplification at levels comparable to the endogenous

63
DAFC-62D (Figure 3A). Notably, the developmental timing of ori62 origin firing that
was activated by ACE3 recapitulated that of the DAFC-62D amplicon rather than DAFC-
66D (Figures 3A, 5B and 5C). This observation indicates that ori62 may carry intrinsic
activities that determine the extent and timing of replication initiation that cannot be
overridden by ACE3.
Given the insufficiency of ori62 to induce amplification, we tested the
amplification properties of a 10kb fragment spanning the maximally amplified region of
DAFC-62D in P-element transformant lines. By FISH/BrdU double labeling, two out of
two lines examined showed an extra 62D signal that colocalized with BrdU incorporation
(Figure 3B). In addition, real-time PCR quantification demonstrated that the
amplification level of the transposon was comparable to the endogenous amplicon, and
that proper developmental timing was preserved (Figures 3B and 5C). A transposon
containing both ACE3 and the 10kb 62D fragment, however, did not show any difference
in amplification level or developmental timing from the 10kb fragment alone (Figure
3B). Thus once again ACE3 was unable to override the amplification properties intrinsic
to DAFC-62D origin.
We tested whether the ORC binding sites were required for amplification and
found that multiple elements are essential. When either ori62 (origin) or –3.0 (control
element) was deleted from the 10kb transposon, the remaining sequences did not support
detectable amplification, as demonstrated by real-time PCR analyses on three
independent lines for each transposon (Figure 3C). Deletion of the +3.5 element also

64
Figure 3. Genetic analysis of cis control elements for DAFC-62D amplification
(A) Amplification levels of transposons containing ori62 alone or together with ACE3.
Error bars are standard errors (SE) of analyses of two or three independent transformant
lines. Structures of the transposon constructs within the 5’ and 3’ P element sequences
are depicted on the right. (B) FISH (green) and BrdU (red) double immunofluorescence
of stage 10B follicle cells that are transformed with the 10 kb central amplified DAFC-
62D fragment (construct shown on the right). The 10kb fragment was labeled for FISH
probes. The two FISH signals correspond to the endogenous amplicon and the
heterologous transposon. Scale bar = 1 µm. Amplification level (±SE) of the 10kb
transposon, alone or accompanied by ACE3, is shown in the lower panel. (C)
Amplification level (±SE) of transposons with –3.0, ori62 or +3.5 deleted from the 10kb
fragment. Deletion size and position are depicted on the right. Numbers beneath
constructs (5’ P, 3’ P, SHWBS and mini-white not shown) indicate relative distance (kb)
to the center of ori62.
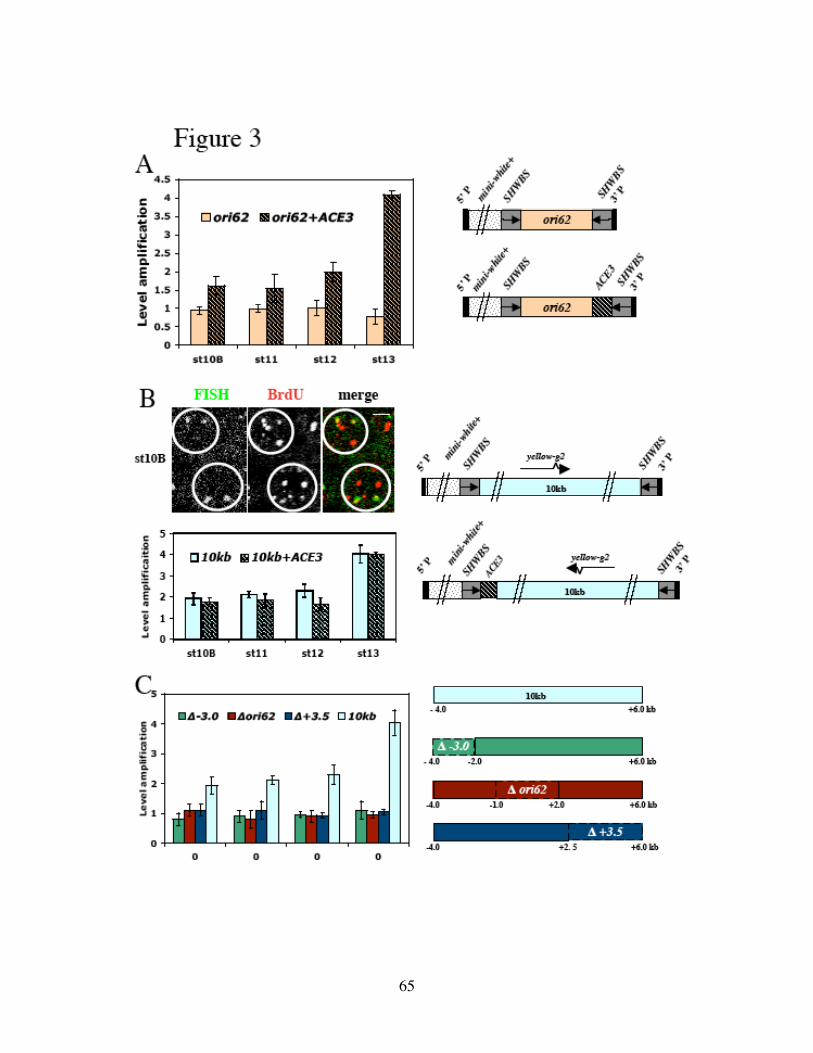
65

66
blocked amplification in all developmental stages (Figure 3C). The requirement of +3.5
for stage 13 amplification was unexpected, because +3.5 is only bound by pre-RC in
stage 10 (Figure 2). We propose that stage 10 may be the only time window during which
ORC loading is permitted and that recruiting ORC to +3.5 is a prerequisite for later origin
firing. It is possible that the –3.0, +3.5, and ori62 elements must all be present to
synergistically load ORC.
The two rounds of origin firing at DAFC-62D are interspersed by transcription
ori62 is localized within the transcription unit of the yellow-g2 (yg2) gene (Figure
2B). This localization is striking, contrasting with the fact that both ACE3 and oriβ are
intergenic and upstream of chorion genes (Delidakis and Kafatos, 1989; Heck and
Spradling, 1990; Orr-Weaver and Spradling, 1986). During genomic replication in S
phase, active origins lie close to promoter regions in fission yeast, Drosophila and
Xenopus (Gomez and Antequera, 1999; Hyrien et al., 1995; Sasaki et al., 1999). In
budding yeast (MacAlpine and Bell, 2005; Nieduszynski et al., 2005; Raghuraman et al.,
2001) and the Chinese hamster ovary dihydrofolate reductase (DHFR) gene locus,
however, initiation of replication is excluded from transcription units (Saha et al., 2004;
Sasaki et al., 2006). Moreover, it has been shown recently that transcription of the yeast
MSH4 gene in meiosis inactivates an origin contained within its open reading frame
(Mori and Shirahige, 2007). Thus we wanted to determine precisely the timing of yg2
transcription relative to the two periods of amplification origin firing.

67
By RNA in situ experiments, high levels of yg2 mRNA in the follicle cell
cytoplasm were found primarily in stage 12 (Claycomb et al., 2004) (Figure 4A). To
determine when transcription itself occurs, we used more sensitive RNA FISH to look for
nascent yg2 transcripts at a specific focus in the nucleus, as would be expected when
transcription takes place at the gene (Jolly et al., 1997; Jolly et al., 1998). We detected
such a focus of nuclear hybridization in a narrow time window of early stage 12 (Figure
4A). Slightly later, cytoplasmic yg2 message began to accumulate and nuclear staining
became undetectable (Figure 4A).
We used antibodies against RNA polymerase II (RNAPII) to further visualize
transcription during follicle cell differentiation, and to examine the localization of
RNAPII during amplification. In a Sciara coprophila amplicon the right boundary of the
initiation zone is determined by the binding of RNAPII (Lunyak et al., 2002), making it
possible that occupancy by RNAPII affects DAFC-62D amplification. RNAPII localized
to subnuclear foci in Drosophila follicle cells, but RNAPII/BrdU double labeling
indicated that RNAPII foci did not overlap significantly with amplicons in stage 10B
(Figure 4B). In stage 12, however, one of the RNAPII foci colocalized with DAFC-62D,
as shown by FISH/RNAPII double immunofluorescence to detect the DAFC-62D DNA
(Figure 4C). The colocalization was observed in stage 12, but not stage 11, (Figure 4C),
coinciding with robust transcription of yg2. Thus yg2 transcription occurs between the
two rounds of amplification origin firing. In particular, it precedes amplification in stage
13, raising the possibility for positive roles of transcription in replication.

68
Figure 4. Temporal and spatial correlation of transcription and amplification
(A) yg2 RNA FISH detects strong nascent transcripts in early stage 12 (left most panel),
weak in late stage 12 (middle panel) and none in stage 13 (right most panel) follicle cells.
Nuclei are circled. Scale bar = 1 µm. (B) Stage 10B BrdU (green) does not co-label with
RNAPII (red). Nuclei are circled. Arrowheads point to minor BrdU foci corresponding to
DAFC-62D and -30B (Claycomb et al., 2004). Scale bar = 1 µm. (C) DNA FISH to
DAFC-62D (green) colocalizes with RNAPII (red) in stage 12 (lower panels) but not
stage 11 (upper panels). Nuclei are circled. Scale bar = 1 µm. (D) BrdU (green) and
RNAPII (red) immunofluorescence in egg chambers cultured with (lower panels) or
without (upper panels) α-amanitin, in stage 10B, 12 and 13. Scale bar = 10 µm.
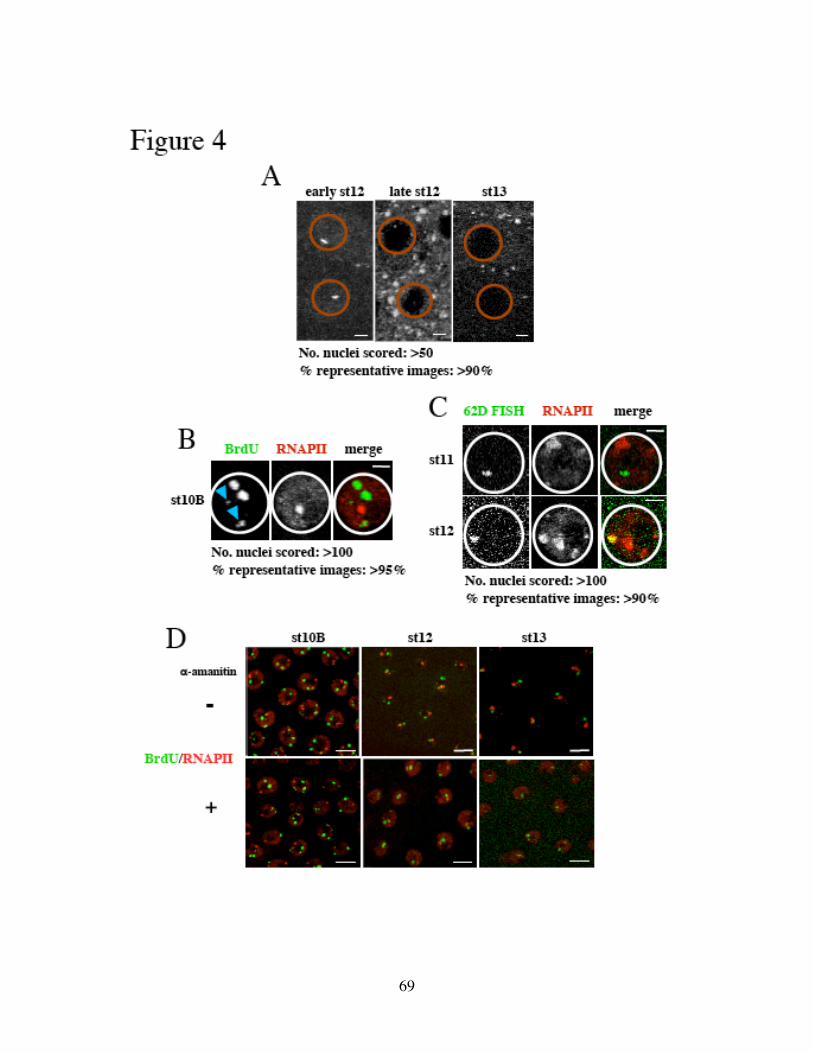
69

70
To investigate potential functional links between transcription and amplification
at DAFC-62D, we used α-amanitin, an RNAPII inhibitor (Lindell et al., 1970), to block
RNAPII elongation. Dissected ovaries were incubated in α-amanitin and allowed to
develop in vitro for 5 hours, the time window that spans stage 10B through 13 under
physiological conditions (Bosco et al., 2001). The toxin did not affect the developmental
programs in general, because the relative abundance of each developmental stage was not
significantly changed, and there was apparent progression in development compared with
dissected egg chambers that did not undergo in vitro culturing (Supplemental Figure 1).
Such treatment strongly diminished mRNA signals of the chorion gene Cp38 detected by
in situ hybridization experiments (data not shown), and completely eliminated the stage
12 FISH spot of nascent yg2 transcripts (Figure 5E). The immunostaining pattern of
BrdU and RNAPII was not affected by the toxin in stage 10B, but in subsequent stages
RNAPII lost it concentration into subnuclear foci and showed more uniform nuclear
staining (Figure 4D). These foci of RNAPII and their elimination by α-amanitin
treatment suggest that during these stages of follicle cell differentiation transcription is
localized to specific nuclear regions.
α-amanitin specifically inhibits DAFC-62D stage 13 amplification
Although after α-amanitin treatment the punctuate pattern of BrdU incorporation
at the largest chorion amplicons remained in stages 12 and 13 (when the BrdU signal for

71
DAFC-62D is often too small to visualize) (Figure 4D), there could have been subtle
changes in amplification that escaped detection by cytology. We used real-time PCR to
quantitatively measure the effect of α-amanitin, if any. The treatment did not change the
accumulative amplification levels of DAFC-66D in stage 13 (Figure 5A), indicating that
neither replication initiation nor fork progression events were affected at this amplicon.
In striking contrast, the stage 13 round of initiation at DAFC-62D was specifically
inhibited by α-amanitin (Figure 5B), whereas initiation in stage 10B was unchanged
(Figure 5B). These results suggested that transcription was required for origin activation
in stage 13.
Unexpectedly, we observed that three independent transposon insertions carrying
the 10kb fragment from DAFC-62D underwent a normal round of amplification in stage
13 in the presence of the toxin (Figure 5C). This indicated that the failure of amplification
at DAFC-62D was not due to a general block to all amplification initiation in stage 13
imposed by α-amanitin, but rather revealed a cis-specific role of transcription for
replication at the endogenous DAFC-62D site. Because all transposons were buffered
from position effects by SHWBS, we investigated whether the presence of insulators
made amplification of these transposons independent of transcription and therefore
resistant to α-amanitin.
As the name indicates, SHWBS recruits the Su(Hw) (Suppressor of Hairy-wing)
(Spana and Corces, 1990; Spana et al., 1988) and additional proteins to form insulator

72
Figure 5. Effect of α-amanitin on DAFC-62D amplification and yg2 transcription
(A) DAFC-66D stage 13 amplification level (±SD) with (stippled bars) or without (solid
bars) α-amanitin treatment. Comparable profiles suggest no obvious defects in replication
initiation or elongation were induced by α-amanitin (P≥ 0.95 in student’s T test). (B)
DAFC-62D stage 10 (upper panel, white bars) and stage 13 (lower panel, black bars)
amplification level (±SD). Stage 13 amplification is specifically inhibited by α-amanitin.
(C) Amplification of the 10kb transposon is not affected by α-amanitin in wild-type
backgrounds. Three independent transformant lines were analyzed and the amplification
levels (±SE) in stages 10B and 13 at the heterologous loci are shown. (D) The 10 kb
transposon is sensitive to α-amanitin in the su(Hw) mutant background. Two independent
lines from (C) were analyzed and amplification level (±SD) for one line is shown. (E)
Transcription from the endogenous yg2 locus but not the buffered transposon was
inhibited by α-amanitin. Panels show from left to right, respectively, RNA FISH signals
against yg2 in stage 12 follicular nuclei: One (no transposon, no α-amanitin), none (no
transposon, α-amanitin treated), two (transposon, no α-amanitin) and one (transposon, α-
amanitin treated).
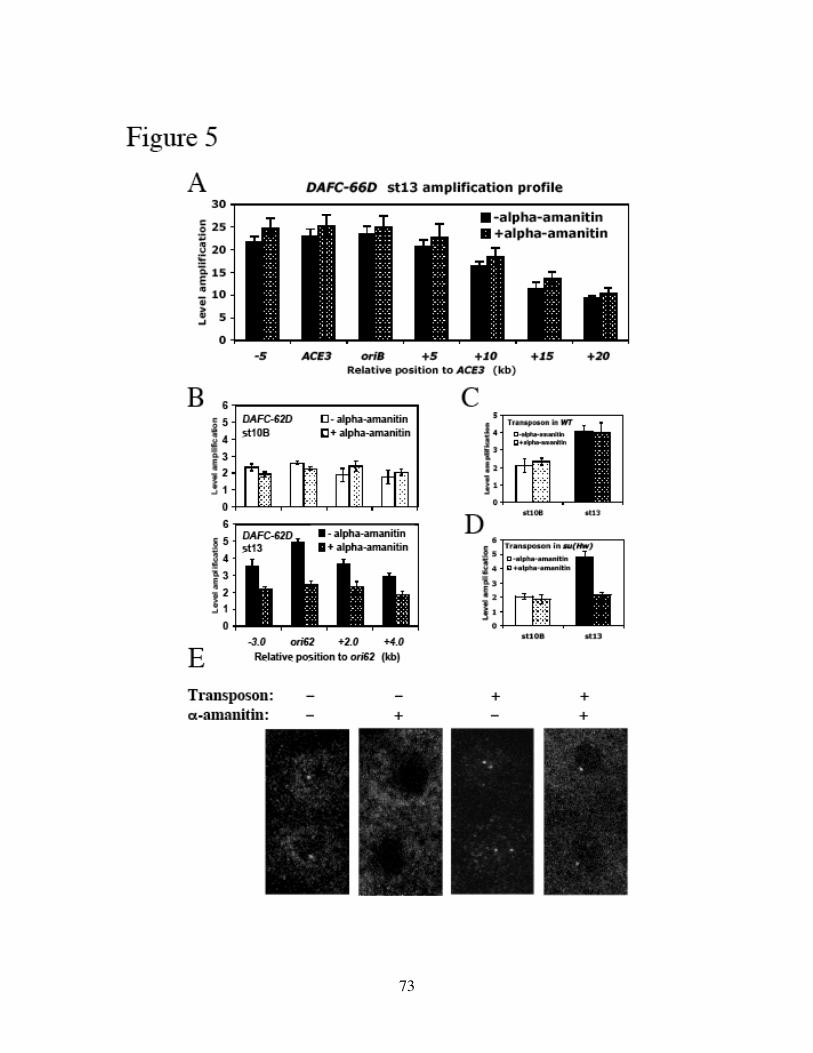
73

74
bodies that are not influenced by either positive or negative position effects (Gerasimova
and Corces, 2001). The su(Hw)v/su(Hw)f allele combination reverses the mutant
phenotype caused by insertion of insulator elements such as yellow2 (Harrison et al.,
1993). It also reduces amplification level of transposons buffered by SHWBS, because in
this su(Hw) mutant background they are subject to position effects (Lu and Tower, 1997).
Transposons containing the 10kb DAFC-62D fragment were crossed into the
su(Hw)v/su(Hw)f background, and two independent transformation lines displayed proper
transposon amplification as determined by real-time PCR analyses (Figure 5D), most
likely because their insertion sites were permissive for amplification. One line failed to
amplify in this background (data not shown). Strikingly, in the absence of Su(Hw)
insulator function, both transposons became sensitive to α-amanitin, and the stage 13
round of amplification was specifically inhibited (Figure 5D).
We also analyzed transposon transcription by RNA FISH of yg2. The ectopic
copy of yg2 carried by the transposon was actively transcribed with proper developmental
timing, as shown by the appearance of an additional locus of yg2 nascent transcripts in
stage 12 (Figure 5E), implying the presence of transcriptional machinery in the
transposon including RNAPII. After α-amanitin treatment, only one spot of yg2
transcripts was detectable, presumably from the transposon because endogenous
transcription of yg2 was completely abolished by α-amanitin in non-transformants
(Figure 5E). Taken together, these experiments suggest that neither transcription nor
amplification of transposons is responsive to α-amanitin when buffered by insulators.

75
The ability of insulated transposons to undergo amplification in the presence of the toxin
excluded the possibility that α-amanitin imposed an indirect effect in trans. Our data
therefore reveal a positive role for RNAPII and possibly other transcription factors in
origin firing in cis, specifically at DAFC-62D in the stage 13 round of amplification.
Inhibition of transcription affects MCM2-7 localization
We showed that α-amanitin treatment affected RNAPII distribution in stage 13 by ChIP
analysis across DAFC-62D. In control follicle cells RNAPII localized to upstream of yg2,
and following stage 10 also appeared at ori62, which is localized within the coding
region of yg2 (Figure 6A). The toxin prevented this redistribution into ori62 from stage
10 to 13, consistent with it blocking translocation/elongation of RNAPII across yg2
(Figure 6A). To investigate mechanisms by which RNAPII transcription could affect
replication, we also analyzed the association of pre-RC components with DAFC-62D in
the presence of α-amanitin. The binding of ORC2 in stage 10A through 13 was
unchanged by the treatment (Supplemental Figure 2). The loading of MCM2-7, however,
was completely abrogated by α-amanitin specifically in stage 13 (Figure 6B). This result
indicates that in stage 13 localization of the MCM complex at DAFC-62D, mediated
downstream of ORC binding, requires transcription in cis. By contrast, at DAFC-66D
pre-RC has disassociated at this development time (Figures 1A and 2C), and no
additional initiation events occur at this amplicon in stage 13 (Claycomb et al., 2002).

76
Figure 6. Association of RNAPII and MCM2-7 with DAFC-62D is affected by α-
amanitin
(A) The effect of α-amanitin on stage 10A-B (approximately half 10A and half 10B
combined; upper panel) and stage13 (lower panel) RNAPII binding (±SD) pattern by
ChIP. (B) MCM loading (±SD) in stage 13 (lower panel) is specifically inhibited by α-
amanitin.
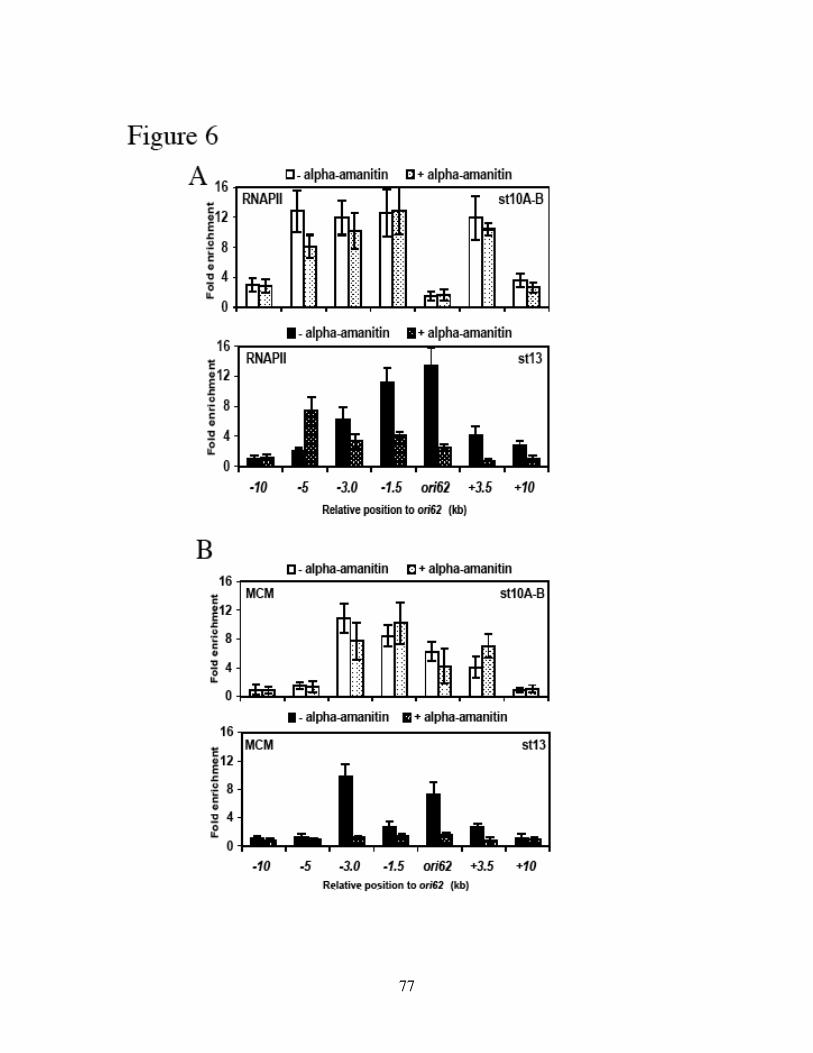
77

78
DISCUSSION
Our analysis of the regulation of DAFC-62D yielded two unexpected findings that
provide critical insights into how metazoan DNA replication is controlled in response to
developmental cues. We found that the positioning of ORC and MCM2-7 can be affected
by differentiation stage, and that MCM2-7 localization requires transcription in cis that
physiologically precedes origin firing. DAFC-62D differs from other DAFCs by
undergoing a round of amplification late in follicle cell differentiation (Figure 7). The late
round of origin activation at DAFC-62D in stage 13 follicle cells contrasts with the other
initiation events in stage 10B in that it takes place at least four hours after the cessation of
previous genomic replication. This developmental delay may have created a quiescent (or
even inhibitory) state of replication activation in stages 11 and 12 that has to be
overcome by unique mechanisms.
We showed for the first time that the pre-RC associates with DNA in a
developmentally regulated manner. Such differential control may be due to specification
of cis elements and/or trans factors such as transcription proteins that could affect ORC
binding (Royzman et al., 1999). Sequence comparison across 12 Drosophila species
showed high levels of conservation at these ORC-binding sites, especially the element at
–3.0 in which a block of 63 nucleotides shows 62% identity (data not shown). Deleting
any of the three ORC-binding sites completely ablated amplification, suggesting a
requirement for synergistic loading of ORC for a threshold level needed for origin

79
Figure 7. Coordination of replication initiation and transcription.
Developmental timing of origin firing events and transcription at DAFC-62D (bottom)
and DAFC-66D (top), as well as differential binding of ORC and the MCM complex at
the origins. Pre-RC (possibly at higher amounts to support more rounds of firing) only
associates with DAFC-66D in early stages. At DAFC-62D, ORC remains localized
through stage 13, whereas the MCM complex disassociates after the first round of origin
firing, and is reloaded in stage 13 for the late round of initiation. This later firing requires
transcription by RNAPII, because it is inhibited by α-amanitin. Candidate mechanisms
include direct interaction and recruitment of MCM2-7 by RNAPII; or indirect
recruitment that needs proximal nucleosomal disassembly at the origin (within the yg2
coding region) mediated by RNAPII transcription.
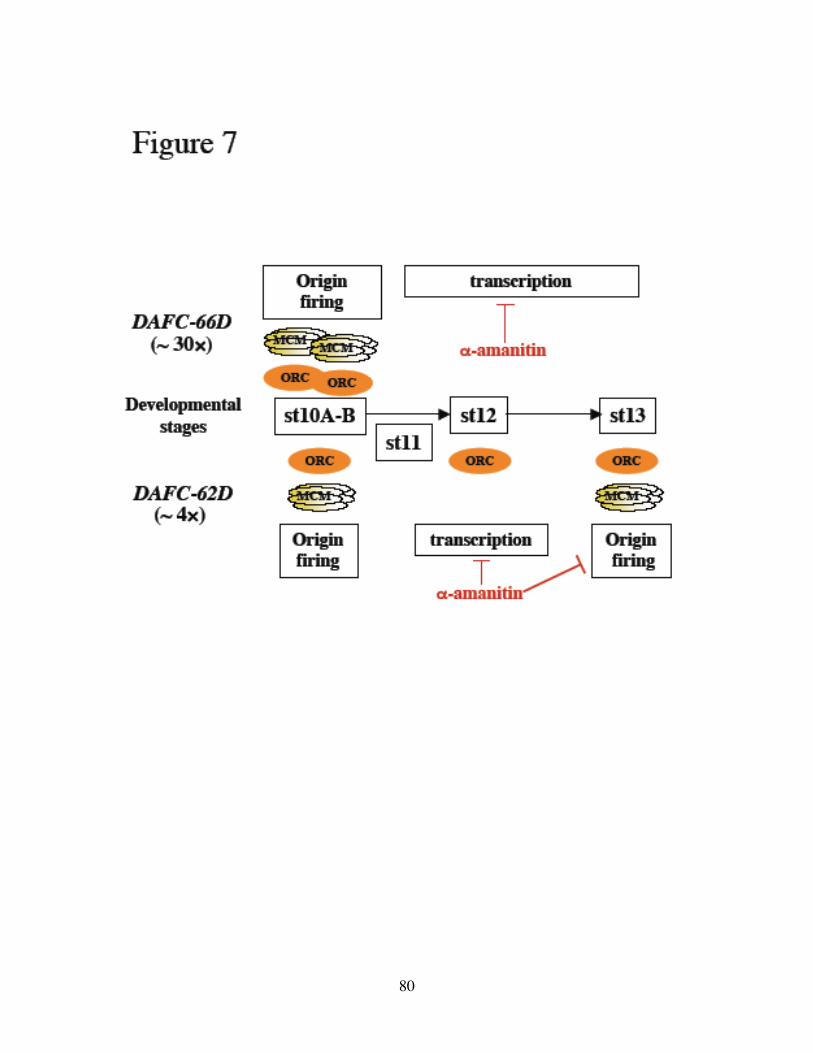
80

81
activation. It also is possible that there is a specific time window in stage 10A for ORC
binding, marked by either a developmentally unique chromatin structure, or the presence
of certain transcription factors and/or specification proteins. Once such window is
missed, ORC loading may no longer be possible, providing an explanation for the
absence of stage 13 amplification when the stage 10A-specific control element at +3.5 is
deleted.
The large control region necessary for DAFC-62D contrasts with the two small
elements of DAFC-66D, ACE3 and oriβ, separated by only 1.5kb and sufficient for
proper regulation of amplification. It is, however, analogous to one class of mammalian
origins known as large zones of initiation (Gilbert, 2004). The best-characterized
example of an initiation zone is the Chinese hamster ovary DHFR locus where a 40kb
intergenic region is composed of many potential initiation sites used with varying degrees
of efficiency (Cvetic and Walter, 2005). At DAFC-62D, the origin and other control
elements are dispersed in a 7kb fragment. Although nascent strand analysis only defined
the origin in stage 10B, the fact that the ACE3-ori62 transposon displayed proper level
and timing of amplification suggests that ori62 contains sufficient origin activity for not
only stage 10 but also stage 13 amplification. The other cis elements may help recruit the
adequate amount of ORC to license an active origin. Given that ACE3 did not cause
higher levels of amplification from either ori62 alone or the whole 10kb fragment, we
suggest that the activity determining the extent and timing of replication initiation may lie
intrinsically in the origin itself and cannot be overcome by amplification enhancers.
Mechanistically different amounts of pre-RC may be mounted onto different origins,

82
parameters for which may include A/T content (Bell, 2002), DNA topology (Remus et
al., 2004) and chromatin structure (Aggarwal and Calvi, 2004; Hartl et al., 2007).
We observed striking inhibitory effects of α-amanitin on DAFC-62D stage 13
origin firing in cis. Such inhibition is specific to the developmental stage, as well as the
genomic and/or chromatin context, since amplification of buffered transposons was not
affected. The transposon results were important, because they showed that α-amanitin did
not cause general defects in replication, such as a decrease in the amount of replication
proteins. Rather it directly affected amplification initiation by repressing transcription via
RNAPII. It is not likely that transposons are not accessible to α-amanitin, a small cyclic
octapeptide. Their resistance to α-amanitin may be due to the presence of insulators that
have established an open chromatin structure within the “insulator bodies” (Gerasimova
and Corces, 2001). Thus the inhibition or slowing down of RNAPII by α-amanitin (Rudd
and Luse, 1996) may be compensated by the favorable chromatin environment to allow
transcription of yg2 (Figure 5E) and the following round of amplification (Figure 5C), in
the presence of the toxin. When the SHWBS insulators were functionally removed, these
heterologous transposons displayed the same sensitivity to α-amanitin (Figure 5D) as the
endogenous amplicon.
In Chinese hamster ovary cells it has been reported that inhibition of transcription
by α-amanitin resulted in deregulation of replication initiation at the DHFR locus (Sasaki
et al., 2006). Our results provide a candidate molecular mechanism by which
transcription could impact replication. Because in stage 13 at DAFC-62D α-amanitin

83
appears to interrupt MCM2-7 loading without affecting the binding of ORC, a special
mechanism that involves active transcription via RNAPII may be required to reload
MCM2-7 and reactivate ori62 (Figure 7). A direct physical interaction has been reported
between RNAPII and MCM2-7 in yeast (Gauthier et al., 2002; Holland et al., 2002),
raising the possibility that such a complex serves to load the MCM complex to origins in
some developmental contexts.
Mounting evidence points to a general link between transcription and replication.
There are several examples in which replication origins coincide with intergenic regions
containing promoter sequences (Kohzaki and Murakami, 2005). In the human β-globin
and c-myc replicons, transcription regulatory elements have been shown to be essential
for replication initiation (Aladjem et al., 1995; Ghosh et al., 2004). At DAFC-66D, the
transcription factors Myb and E2F/RB associate with ORC via direct protein-protein
interaction (Beall et al., 2002; Bosco et al., 2001). In the Sciara salivary gland DNA puff
II/9A, amplification is controlled by ecdysone, potentially through direct interaction with
a putative ecdysone response element adjacent to its ORC-binding site (Foulk et al.,
2006). Similarly, a heterodimeric transcription activator containing EcR (ecdysone
receptor) mediates not only the transcription but also amplification of at least some
chorion genes in Drosophila (Hackney et al., 2007). Moreover, the recruitment of
transcription factors alters origin activity on episomal plasmids in both S. cerevisiae and
X. laevis eggs (Danis et al., 2004; Kohzaki and Murakami, 2005).
Transcription factors may modulate DNA replication through their ability to
recruit histone-modifying enzymes and/or chromatin-remodeling proteins (Kohzaki and

84
Murakami, 2005). In X. laevis eggs, injected plasmid DNA undergoes site-specific
initiation of replication in the presence of a transcription factor that is known to recruit
the chromatin-remodeling complex (Danis et al., 2004). This does not require active
transcription, but rather correlates with the acetylation level of histone H3 at the initiation
sites (Danis et al., 2004). Levels of hyperacetylated histone H4 coincide with chorion
amplicons in Drosophila and are associated with origin activation (Aggarwal and Calvi,
2004; Hartl et al., 2007). We observed no significant difference in the acetylation pattern
of histones H3 or H4 between DAFC-62D and -66D, other than higher enrichment levels
of Acetyl-K8-H4 at DAFC-66D in stage 10B (Xie and Orr-Weaver, unpublished results),
raising the intriguing possibility that acetylation levels account for the higher
concentration of ORC (Figure 1B) and higher number of rounds of initiation at oriβ.
Microarray analysis of genomic replication in Drosophila and human cells shows
a correlation between regions undergoing active transcription and early replication (Jeon
et al., 2005; MacAlpine et al., 2004; Schubeler et al., 2002; Woodfine et al., 2004). A
more extensive study of Drosophila chromosome 2L in Kc cells uncovered an association
between sites of BrdU incorporation, ORC localization and RNAPII binding (MacAlpine
et al., 2004). The involvement of RNAPII transcription in DAFC-62D amplification
regulation is a concrete example for organized domains of transcription and replication
(Chakalova et al., 2005). RNAPII has been shown to be required for histone displacement
ahead of the position of RNAPII within the transcriptionally activated gene’s coding
region in both yeast and mammalian systems (Brown and Kingston, 1997; Lee et al.,
2004; Schwabish and Struhl, 2004; Zhao et al., 2005). Activation of the human hsp70

85
gene leads to nucleosomal disassembly in the first 400 bp coding sequence in front of
RNAPII, and such chromatin disruption is resistant to α-amanitin (Brown and Kingston,
1997). Disruption of distal downstream chromatin, however, is sensitive to α-amanitin,
suggesting RNAPII movement to the vicinity is necessary to remodel chromatin (Brown
and Kingston, 1997). Such a role of RNAPII in displacing proximal histones may play
into the successful recruitment of MCM2-7 at the amplification origins (within the yg2
gene coding region) in DAFC-62D. Supporting this hypothesis, MCM2-7 associated
DNA is more susceptible to nuclease digestion, indicating that these chromatin domains
may be less tightly compacted, although the causal and consequence relation is not clear
(Forsburg, 2004; Holthoff et al., 1998; Richter et al., 1998).
The analysis of DAFC-62D and -66D demonstrates that there are distinct
mechanisms that differentially regulate amplification origins during Drosophila follicle
cell development. Our findings reveal pathways to control localization of replication
factors, license origins and activate DNA replication, which provide a conceptual
framework for defining how origin selection and activation are regulated by transcription
in metazoan development.

86
Experimental procedures
Plasmid Construction and Transformant Lines
To construct the transposons with the 1kb ori62 and 10kb central amplified
region, these DNA intervals were PCR amplified from BACR22J16 using PfuTurbo
DNA polymerase (Stratagene), blunt-ligated into pCR-Blunt vector (Stratagene), and
subcloned into pBluescript-PCRA (Lu et al., 2001) via NheI restriction sites previously
engineered into the primers. These plasmids are called PCRAori62 and PCRA10kb. The
fragment containing one SHWBS and either ori62 or 10kb was liberated and subcloned
into the Not1 and XhoI sites of Big Parent (Lu et al., 2001), to generate FXori62 and
FX10kb. To generate the ACE3 insertions, PCRAori62 or PCRA10kb were digested by
NheI to excise ori62 or 10kb. These fragments were then subcloned into Small(ori deln)
(Lu et al., 2001) to generate a construct that contains both ori62 or 10kb and ACE3
(FXACEori62 and FXACE10kb).
To generate the three deletions within the 10kb transposon, fragments from the
central amplified region including −2.0 to +6.0 (Δ-3.0), −4.0 to −1.0, +2.0 to +6.0 and
−4.0 to +2.5 (Δ+3.5) (numbers are relative distance to ori62 in kb; + and − indicate
orientation) were PCR amplified from BACR22J16 using PfuTurbo DNA polymerase
(Stratagene), and blunt-ligated into pCR-Blunt vector (Stratagene) to construct
pCRBΔ−3.0, pCRB−3.0, pCRB+3.5 and pCRBΔ+3.5, respectively. The +2.0 to +6.0
fragment was isolated by NotI digestion and subcloned into pCRB+3.5 to generate
pCRBΔori62. Δ−3.0, Δori62 and Δ+3.5 were excised and substituted for the NheI
fragment in the ori62 transposon to generate FXΔ−3.0, FXΔori62 and FXΔ+3.5.

87
All transposon constructs were individually injected into yw embryos to establish
at least three independent homozygous transformant lines per each construct. At least two
lines per each construct were analyzed for amplification level by real-time PCR (see
below). Primers targeted transposon-specific sequences to distinguish between the
endogenous DAFC-62D and the heterologous transposons. Primer sequences are
available upon request.
Transposons on either the X or 2nd chromosome were introduced into y2 sc1 w67 ct6
f1; bx34e su(Hw)v/TM6, su(Hw)f, Ubx (Harrison et al., 1993) flies by crossing. Two
independent transformation lines carrying the 10kb DAFC-62D fragment retained proper
amplification as determined by real-time PCR (see below) and were tested for sensitivity
to α-amanitin (see below).
Antibodies, Immunofluorescence and Confocal Microscopy
The anti-ORC2 antibodies were previously described and were obtained from
Stephen Bell (Royzman et al., 1999). The anti-MCM2-7 monoclonal antibody was a gift
from Stephen Bell (Claycomb et al., 2002). The anti-RNAPII antibody (Upstate)
recognizes both the phospho and non-phospho carboxyl-terminal domain of RNA
polymerase II. It was used at a 1: 250 dilution in double immunostaining with BrdU as
described (Royzman et al., 1999), with the following modifications: secondary detection
of RNAPII was with Rhodamine-RedX conjugated donkey anti-mouse at 1:100; rabbit
anti-BrdU antiserum (Accurate Chemical) was used at 1:50; and secondary detection of
BrdU was with FITC conjugated donkey anti-rabbit at 1:100.

88
All images were collected on a Zeiss Axivert 100M Meta confocal microscope
with LSM51 Software. A 63× Plan Aprochromat objective was used to capture images in
Fig. 4D and a 100× Plan Aprochromat objective was used for all others.
Chromatin Immunoprecipitation
ChIP was performed on 300 staged egg chambers per experiment as described
(Austin et al., 1999). Chromatin DNA was sheared using a Branson 250 sonicator into
100 to 500bp fragments, with most fragments at 200 to 350bp (data not shown). To
immunoprecipitate protein-bound chromatin, 1:250 diluted anti-ORC2, 1: 250 anti-
RNAPII or 1:100 diluted anti-MCM2-7 were incubated with chromatin at 4°C overnight.
For the initial screen of ORC-binding sites, primer pairs were designed to span the 10kb
central amplified region in approximately 300bp intervals (sequences available upon
request) and each was used in semi-quantitative conventional PCR together with an
internal control actin (Royzman et al., 1999). Subsequent accurate quantification of
enrichment was obtained by real-time PCR (see below).
Quantitative (Real-Time) PCR
Absolute quantitative (real-time) PCR was performed as described (Claycomb et
al., 2004; Claycomb et al., 2002). Standard curves were constructed from four tenfold
serial dilutions of stage1-8 egg chamber DNA (for amplification level), BACR22J16
DNA (for nascent strand analysis, see below), or input chromatin prior to
immunoprecipitation (for ChIP). The endogenous control was a nonamplified locus at
62C5 (Claycomb et al., 2002).

89
Relative quantitative (real-time) PCR was used to detect the difference between a
test sample and a calibrator sample wherever indicated in the text according to
manufacturer’s recommendations (Applied Biosystems 7300 Fast Real-Time PCR
System). The calibrator sample was either stage1-8 egg chamber DNA for amplification
profiling, or input chromatin for ChIP assays. The same endogenous control at 62C5 was
used (Claycomb et al., 2002).
Nascent Strand Analysis
50-100 staged egg chambers were dissected in nonsupplemented Grace’s medium
(GIBCO-BRL) and immediately frozen in −80°C until accumulatively 1000 were
collected. Nascent DNA isolation and size fractionation were performed as described
(Cotterill, 1999; Lunyak et al., 2002). The only modification was that the gel fractionized
DNA was recovered using the Qiaquick Gel Extraction Kit (Qiagen) and eluted in 30 µl
of TE buffer. Each fraction was individually analyzed for the abundance of specific
sequences by absolute quantitative real-time PCR, referenced to serial dilutions of
BACR22J16 DNA as standards, with the least concentrated standard sample designated
as 1.
Fluorescent in Situ Hybridization
DNA FISH and BrdU double labeling was performed as described (Claycomb et
al., 2004). The probe was prepared from the 10kb central amplified region previously
PCR cloned from BACR22J16, and 300 ng was used in a 40 µl hybridization reaction.

90
To detect RNA signals by FISH (Tam et al., 2002), ovaries were dissected in
nonsupplemented Grace’s medium (GIBCO-BRL) with 10mM vanadyl ribonucleoside
complex (VRC, Invitrogen) to prevent RNA degradation. Formaldehyde fixation,
formamide equilibration and pre-hybridization steps for RNA FISH were essentially the
same as in DNA FISH except that DEPC-treated ddH2O and deionized formamide
(Sigma) were used whenever applicable. The probe was prepared from yg2 cDNA using
the Invitrogen BioNick Labeling Kit. 100 ng of digoxingenin (DIG) labeled probe was
denatured at 80°C in formamide for 10 minutes together with 10 µg sonicated salmon
sperm DNA, and hybridized to pre-hybridized egg chambers at 37 overnight in 40 µl
buffer containing 50% formamide, 10% dextran sulfate (Sigma), 0.2% BSA (Sigma),
20mM VRC and 2× SSCT. Secondary detection was with goat anti-DIG FITC at 1:200
(Enzo). Samples were mounted in Vectashield (Vector Labs).
α-Amanitin Treatment
Whole ovaries were dissected from female Oregon R flies and incubated in vitro
in 333 µg/ml α-amanitin for 5 hours at room temperature as described (Bosco et al.,
2001). Egg chambers were dissected immediately after incubation to determine their
developmental stages, same stage egg chambers were pooled together for DNA
extraction, and subsequently subjected to real-time PCR analysis for amplification level
in each stage. For immunofluorescence and ChIP experiments, ovaries were washed and
formaldehyde fixed right after α-amanitin treatment. Egg chambers were then staged
based on their morphology and taken through ChIP protocols.

91
Acknowledgments
We thank David MacAlpine and Stephen Bell for supplying the ORC2 and
MCM2-7 antibodies and inspiring discussions, John Tower for providing pCaSpeR-4
constructs, Jacob Mueller for advice on RNA FISH, Bashi Raveendranathan and Anja-
Katrin Bielinsky for the nascent strand analysis protocol, as well as Jianzhu Chen, Troy
Littleton and Julie Claycomb for suggestions. The confocal microscopy was conducted
using the W.M. Keck Foundation Biological Imaging Facility at the Whitehead Institute.
Stephen Bell, Peter Reddien, Andreas Hochwagen, Cintia Hongay, Yingdee
Unhavaithaya and Jane Kim provided helpful comments on the manuscript. This work
was supported by NIH grant GM57541 to TO-W.

92
Supplemental Figure 1. Follicle cell development is not affected by α-amanitin.
After 5h incubation in 333 µg/ml α-amanitin or medium alone, whole ovaries were
dissected and the percentage of egg chambers in each developmental stage (from stage 9
to 13, about 500 egg chambers in total) was determined. Results of three independent
experiments are shown. Error bars represent standard errors. Student’s T test shows no
significant difference (P≥0.97). White bars represent similarly dissected ovaries that were
not cultured in vitro. Higher percentage of stage 10 and lower stage 14 egg chambers in
these in vivo samples than cultured ones indicate progression of development.
93

94
Supplemental Figure 2. ORC2 localization is not affected by α-amanitin.
Real-time PCR analyses of anti-ORC2 ChIP across DAFC-62D in stage 10A-B and stage
13 show insignificant changes in ORC association pattern or level with α-amanitin
treatment. Error bars are standard deviations of triplicate PCR reactions.
95

96
References
Aggarwal, B. D., and Calvi, B. R. (2004). Chromatin regulates origin activity inDrosophila follicle cells. Nature 430, 372-376.Aladjem, M. I., Groudine, M., Brody, L. L., Dieken, E. S., Fournier, R. E., Wahl, G. M.,and Epner, E. M. (1995). Participation of the human beta-globin locus control region ininitiation of DNA replication. Science 270, 815-819.Asano, M., and Wharton, R. P. (1999). E2F mediates developmental and cell cycleregulation of ORC1 in Drosophila. Embo J 18, 2435-2448.Austin, R. J., Orr-Weaver, T. L., and Bell, S. P. (1999). Drosophila ORC specificallybinds to ACE3, an origin of DNA replication control element. Genes Dev 13, 2639-2649.Beall, E. L., Manak, J. R., Zhou, S., Bell, M., Lipsick, J. S., and Botchan, M. R. (2002).Role for a Drosophila Myb-containing protein complex in site-specific DNA replication.Nature 420, 833-837.Bell, S. P. (2002). The origin recognition complex: from simple origins to complexfunctions. Genes Dev 16, 659-672.Bell, S. P., and Dutta, A. (2002). DNA replication in eukaryotic cells. Annu RevBiochem 71, 333-374.Bell, S. P., and Stillman, B. (1992). ATP-dependent recognition of eukaryotic origins ofDNA replication by a multiprotein complex. Nature 357, 128-134.Bielinsky, A. K., Blitzblau, H., Beall, E. L., Ezrokhi, M., Smith, H. S., Botchan, M. R.,and Gerbi, S. A. (2001). Origin recognition complex binding to a metazoan replicationorigin. Curr Biol 11, 1427-1431.Bosco, G., Du, W., and Orr-Weaver, T. L. (2001). DNA replication control throughinteraction of E2F-RB and the origin recognition complex. Nat Cell Biol 3, 289-295.Brown, S. A., and Kingston, R. E. (1997). Disruption of downstream chromatin directedby a transcriptional activator. Genes Dev 11, 3116-3121.Calvi, B. R., Lilly, M. A., and Spradling, A. C. (1998). Cell cycle control of chorion geneamplification. Genes Dev 12, 734-744.Calvi, B. R., and Spradling, A. C. (1999). Chorion gene amplification in Drosophila: Amodel for metazoan origins of DNA replication and S-phase control. Methods 18, 407-417.Carminati, J. L., Johnston, C. G., and Orr-Weaver, T. L. (1992). The Drosophila ACE3chorion element autonomously induces amplification. Mol Cell Biol 12, 2444-2453.Chakalova, L., Debrand, E., Mitchell, J. A., Osborne, C. S., and Fraser, P. (2005).Replication and transcription: shaping the landscape of the genome. Nat Rev Genet 6,669-677.Claycomb, J. M., Benasutti, M., Bosco, G., Fenger, D. D., and Orr-Weaver, T. L. (2004).Gene amplification as a developmental strategy: isolation of two developmentalamplicons in Drosophila. Dev Cell 6, 145-155.

97
Claycomb, J. M., MacAlpine, D. M., Evans, J. G., Bell, S. P., and Orr-Weaver, T. L.(2002). Visualization of replication initiation and elongation in Drosophila. J Cell Biol159, 225-236.Claycomb, J. M., and Orr-Weaver, T. L. (2005). Developmental gene amplification:insights into DNA replication and gene expression. Trends Genet 21, 149-162.Cotterill, S. (1999). Eukaryotic DNA replication : a practical approach (Oxford ; NewYork, Oxford University Press).Cvetic, C., and Walter, J. C. (2005). Eukaryotic origins of DNA replication: could youplease be more specific? Semin Cell Dev Biol 16, 343-353.Danis, E., Brodolin, K., Menut, S., Maiorano, D., Girard-Reydet, C., and Mechali, M.(2004). Specification of a DNA replication origin by a transcription complex. Nat CellBiol 6, 721-730.de Cicco, D. V., and Spradling, A. C. (1984). Localization of a cis-acting elementresponsible for the developmentally regulated amplification of Drosophila chorion genes.Cell 38, 45-54.Delidakis, C., and Kafatos, F. C. (1989). Amplification enhancers and replication originsin the autosomal chorion gene cluster of Drosophila. Embo J 8, 891-901.Forsburg, S. L. (2004). Eukaryotic MCM proteins: beyond replication initiation.Microbiol Mol Biol Rev 68, 109-131.Foulk, M. S., Liang, C., Wu, N., Blitzblau, H. G., Smith, H., Alam, D., Batra, M., andGerbi, S. A. (2006). Ecdysone induces transcription and amplification in Sciaracoprophila DNA puff II/9A. Dev Biol 299, 151-163.Gauthier, L., Dziak, R., Kramer, D. J., Leishman, D., Song, X., Ho, J., Radovic, M.,Bentley, D., and Yankulov, K. (2002). The role of the carboxyterminal domain of RNApolymerase II in regulating origins of DNA replication in Saccharomyces cerevisiae.Genetics 162, 1117-1129.Gerasimova, T. I., and Corces, V. G. (2001). Chromatin insulators and boundaries:effects on transcription and nuclear organization. Annu Rev Genet 35, 193-208.Gerbi, S. A. (2005). Mapping origins of DNA replication in eukaryotes. Methods MolBiol 296, 167-180.Ghosh, M., Liu, G., Randall, G., Bevington, J., and Leffak, M. (2004). Transcriptionfactor binding and induced transcription alter chromosomal c-myc replicator activity.Mol Cell Biol 24, 10193-10207.Giacca, M., and Zentilin, L. (1994). Fine mapping of a replication origin of human DNA.Proc Natl Acad Sci U S A 91, 7119-7123.Gilbert, D. M. (2001). Making sense of eukaryotic DNA replication origins. Science 294,96-100.Gilbert, D. M. (2004). In search of the holy replicator. Nat Rev Mol Cell Biol 5, 848-855.Gilbert, D. M. (2005). Origins go plastic. Mol Cell 20, 657-658.Gomez, M., and Antequera, F. (1999). Organization of DNA replication origins in thefission yeast genome. Embo J 18, 5683-5690.

98
Hackney, J. F., Pucci, C., Naes, E., and Dobens, L. (2007). Ras signaling modulatesactivity of the ecdysone receptor EcR during cell migration in the Drosophila ovary. DevDyn 236, 1213-1226.Harrison, D. A., Gdula, D. A., Coyne, R. S., and Corces, V. G. (1993). A leucine zipperdomain of the suppressor of Hairy-wing protein mediates its repressive effect onenhancer function. Genes Dev 7, 1966-1978.Hartl, T., Boswell, C., Orr-Weaver, T. L., and Bosco, G. (2007). Developmentallyregulated histone modifications in Drosophila follicle cells: initiation of geneamplification is associated with histone H3 and H4 hyperacetylation and H1phosphorylation. Chromosoma.Heck, M. M., and Spradling, A. C. (1990). Multiple replication origins are used duringDrosophila chorion gene amplification. J Cell Biol 110, 903-914.Holland, L., Gauthier, L., Bell-Rogers, P., and Yankulov, K. (2002). Distinct parts ofminichromosome maintenance protein 2 associate with histone H3/H4 and RNApolymerase II holoenzyme. Eur J Biochem 269, 5192-5202.Holthoff, H. P., Baack, M., Richter, A., Ritzi, M., and Knippers, R. (1998). Humanprotein MCM6 on HeLa cell chromatin. J Biol Chem 273, 7320-7325.Hyrien, O., Maric, C., and Mechali, M. (1995). Transition in specification of embryonicmetazoan DNA replication origins. Science 270, 994-997.Jeon, Y., Bekiranov, S., Karnani, N., Kapranov, P., Ghosh, S., MacAlpine, D., Lee, C.,Hwang, D. S., Gingeras, T. R., and Dutta, A. (2005). Temporal profile of replication ofhuman chromosomes. Proc Natl Acad Sci U S A 102, 6419-6424.Jolly, C., Mongelard, F., Robert-Nicoud, M., and Vourc'h, C. (1997). Optimization ofnuclear transcript detection by FISH and combination with fluorescenceimmunocytochemical detection of transcription factors. J Histochem Cytochem 45, 1585-1592.Jolly, C., Robert-Nicoud, M., and Vourc'h, C. (1998). Contribution of growing RNAmolecules to the nuclear transcripts foci observed by FISH. Exp Cell Res 238, 299-304.Kobayashi, T., Rein, T., and DePamphilis, M. L. (1998). Identification of primaryinitiation sites for DNA replication in the hamster dihydrofolate reductase gene initiationzone. Mol Cell Biol 18, 3266-3277.Kohzaki, H., and Murakami, Y. (2005). Transcription factors and DNA replication originselection. Bioessays 27, 1107-1116.Landis, G., Kelley, R., Spradling, A. C., and Tower, J. (1997). The k43 gene, required forchorion gene amplification and diploid cell chromosome replication, encodes theDrosophila homolog of yeast origin recognition complex subunit 2. Proc Natl Acad Sci US A 94, 3888-3892.Lee, C. K., Shibata, Y., Rao, B., Strahl, B. D., and Lieb, J. D. (2004). Evidence fornucleosome depletion at active regulatory regions genome-wide. Nat Genet 36, 900-905.Lee, D. G., and Bell, S. P. (1997). Architecture of the yeast origin recognition complexbound to origins of DNA replication. Mol Cell Biol 17, 7159-7168.

99
Lindell, T. J., Weinberg, F., Morris, P. W., Roeder, R. G., and Rutter, W. J. (1970).Specific inhibition of nuclear RNA polymerase II by alpha-amanitin. Science 170, 447-449.Loebel, D., Huikeshoven, H., and Cotterill, S. (2000). Localisation of the DmCdc45DNA replication factor in the mitotic cycle and during chorion gene amplification.Nucleic Acids Res 28, 3897-3903.Lu, L., and Tower, J. (1997). A transcriptional insulator element, the su(Hw) binding site,protects a chromosomal DNA replication origin from position effects. Mol Cell Biol 17,2202-2206.Lu, L., Zhang, H., and Tower, J. (2001). Functionally distinct, sequence-specificreplicator and origin elements are required for Drosophila chorion gene amplification.Genes Dev 15, 134-146.Lunyak, V. V., Ezrokhi, M., Smith, H. S., and Gerbi, S. A. (2002). Developmentalchanges in the Sciara II/9A initiation zone for DNA replication. Mol Cell Biol 22, 8426-8437.MacAlpine, D. M., and Bell, S. P. (2005). A genomic view of eukaryotic DNAreplication. Chromosome Res 13, 309-326.MacAlpine, D. M., Rodriguez, H. K., and Bell, S. P. (2004). Coordination of replicationand transcription along a Drosophila chromosome. Genes Dev 18, 3094-3105.Mendez, J., and Stillman, B. (2003). Perpetuating the double helix: molecular machinesat eukaryotic DNA replication origins. Bioessays 25, 1158-1167.Mori, S., and Shirahige, K. (2007). Perturbation of the activity of replication origin bymeiosis-specific transcription. J Biol Chem 282, 4447-4452.Nieduszynski, C. A., Blow, J. J., and Donaldson, A. D. (2005). The requirement of yeastreplication origins for pre-replication complex proteins is modulated by transcription.Nucleic Acids Res 33, 2410-2420.Norio, P. (2006). DNA replication: the unbearable lightness of origins. EMBO Rep 7,779-781.Orr-Weaver, T. L. (1991). Drosophila chorion genes: cracking the eggshell's secrets.Bioessays 13, 97-105.Orr-Weaver, T. L., and Spradling, A. C. (1986). Drosophila chorion gene amplificationrequires an upstream region regulating s18 transcription. Mol Cell Biol 6, 4624-4633.Raghuraman, M. K., Winzeler, E. A., Collingwood, D., Hunt, S., Wodicka, L., Conway,A., Lockhart, D. J., Davis, R. W., Brewer, B. J., and Fangman, W. L. (2001). Replicationdynamics of the yeast genome. Science 294, 115-121.Remus, D., Beall, E. L., and Botchan, M. R. (2004). DNA topology, not DNA sequence,is a critical determinant for Drosophila ORC-DNA binding. Embo J 23, 897-907.Richter, A., Baack, M., Holthoff, H. P., Ritzi, M., and Knippers, R. (1998). Mobilizationof chromatin-bound Mcm proteins by micrococcal nuclease. Biol Chem 379, 1181-1187.Royzman, I., Austin, R. J., Bosco, G., Bell, S. P., and Orr-Weaver, T. L. (1999). ORClocalization in Drosophila follicle cells and the effects of mutations in dE2F and dDP.Genes Dev 13, 827-840.

100
Rudd, M. D., and Luse, D. S. (1996). Amanitin greatly reduces the rate of transcriptionby RNA polymerase II ternary complexes but fails to inhibit some transcript cleavagemodes. J Biol Chem 271, 21549-21558.Saha, S., Shan, Y., Mesner, L. D., and Hamlin, J. L. (2004). The promoter of the Chinesehamster ovary dihydrofolate reductase gene regulates the activity of the local origin andhelps define its boundaries. Genes Dev 18, 397-410.Sasaki, T., Ramanathan, S., Okuno, Y., Kumagai, C., Shaikh, S. S., and Gilbert, D. M.(2006). The Chinese hamster dihydrofolate reductase replication origin decision pointfollows activation of transcription and suppresses initiation of replication withintranscription units. Mol Cell Biol 26, 1051-1062.Sasaki, T., Sawado, T., Yamaguchi, M., and Shinomiya, T. (1999). Specification ofregions of DNA replication initiation during embryogenesis in the 65-kilobaseDNApolalpha-dE2F locus of Drosophila melanogaster. Mol Cell Biol 19, 547-555.Schubeler, D., Scalzo, D., Kooperberg, C., van Steensel, B., Delrow, J., and Groudine, M.(2002). Genome-wide DNA replication profile for Drosophila melanogaster: a linkbetween transcription and replication timing. Nat Genet 32, 438-442.Schwabish, M. A., and Struhl, K. (2004). Evidence for eviction and rapid deposition ofhistones upon transcriptional elongation by RNA polymerase II. Mol Cell Biol 24,10111-10117.Spana, C., and Corces, V. G. (1990). DNA bending is a determinant of bindingspecificity for a Drosophila zinc finger protein. Genes Dev 4, 1505-1515.Spana, C., Harrison, D. A., and Corces, V. G. (1988). The Drosophila melanogastersuppressor of Hairy-wing protein binds to specific sequences of the gypsyretrotransposon. Genes Dev 2, 1414-1423.Spradling, A. C., de Cicco, D. V., Wakimoto, B. T., Levine, J. F., Kalfayan, L. J., andCooley, L. (1987). Amplification of the X-linked Drosophila chorion gene clusterrequires a region upstream from the s38 chorion gene. Embo J 6, 1045-1053.Tower, J. (2004). Developmental gene amplification and origin regulation. Annu RevGenet 38, 273-304.Vashee, S., Cvetic, C., Lu, W., Simancek, P., Kelly, T. J., and Walter, J. C. (2003).Sequence-independent DNA binding and replication initiation by the human originrecognition complex. Genes Dev 17, 1894-1908.Woodfine, K., Fiegler, H., Beare, D. M., Collins, J. E., McCann, O. T., Young, B. D.,Debernardi, S., Mott, R., Dunham, I., and Carter, N. P. (2004). Replication timing of thehuman genome. Hum Mol Genet 13, 191-202.Zhang, H., and Tower, J. (2004). Sequence requirements for function of the Drosophilachorion gene locus ACE3 replicator and ori-beta origin elements. Development 131,2089-2099.Zhao, J., Herrera-Diaz, J., and Gross, D. S. (2005). Domain-wide displacement ofhistones by activated heat shock factor occurs independently of Swi/Snf and is notcorrelated with RNA polymerase II density. Mol Cell Biol 25, 8985-8999.

101
Chapter Three
Conclusions and Future Directions

102
This thesis work investigated mechanisms that control the unique timing of
DAFC-62D origin activation using cytological, molecular and genetic methods. We first
defined the origin sequences in DAFC-62D, ori62, by analyzing the amount of nascent
replicative DNA across this amplicon. ORC2 localized to ori62, as well as two other
sites, -3.0 and +3.5, that did not confer origin activity. Both ORC2 and MCM2-7
displayed differential association with these sequences, corresponding to the two rounds
of amplification in two separate developmental stages (10 and 13). All three elements
were required for either round of DAFC-62D amplification, because deleting any one
completely abolished amplification in transposon experiments. Preceded by transcription
of yg2 (ori62 resides within the coding region of this gene) in stage 12, the late round of
origin firing was ablated by the RNAPII inhibitor α-amanitin. This effect was absent
from other amplicons and insulated transposons, and specific to the stage 13 round of
amplification at DAFC-62D and transposons that did not have functional insulators.
Finally, blocking RNAPII transcription compromised MCM2-7 recruitment.
Our analyses of the regulation of DAFC-62D yielded several unexpected findings
and provided critical insights into how metazoan DNA replication is controlled in
response to developmental cues. We find that the positioning of ORC and MCM2-7 can
be affected by differentiation stage. Transcription via RNAPII in cis controls localization
of replication factors and origin activation. The comparative analyses of DAFC-62D and
-66D demonstrate that there are distinct mechanisms for differential regulation of
amplification origins during Drosophila follicle cell development. Transposon
experiments suggest their distinctive amplification behavior compared to the endogenous

103
amplicon may be accounted for by the insulators’ special properties. All these and future
directions will be discussed in detail below, and in the end we will entertain the idea of
transcription “factories” based on RNAPII immunostaining patterns.
Differential localization of pre-RC
We show for the first time that components of the pre-RC associate with DNA in a
developmentally regulated manner. ORC2 remains bound to ori62 and –3.0 at all
developmental stages, even in stage 12 during active transcription through ori62. It is
possible that ORC as a complex only localizes to part of the polytene chromosome while
the other strands undergo transcription. Alternatively, the association of the six-subunit
ORC complex with chromatin is dynamic; or at least some subunit(s) such as ORC1 are
dynamically regulated. In human cells ORC1 level oscillates, accumulating in G1 and
degraded in S phase, when other ORC subunits (ORCs 2-5) remain at almost constant
levels (Tatsumi et al., 2003). ORCs 2-5 form a complex that is present throughout cell
cycle, and in G1 paralleling the elevated level of ORC1, the formation of an ORC1-5
complex temporally recruits ORCs 2-5 into nuclease-insoluable structures (Ohta et al.,
2003).
ORC1 abundance in several Drosophila tissues has been shown to generally
correlate with DNA replication activity (Asano and Wharton, 1999). In follicle cells its
localization, like ORC2, switches from nuclear during genomic replication, to foci at
amplicons in amplification stages (Asano and Wharton, 1999). Overexpression of ORC1
increases DNA synthesis throughout the nucleus, while inhibiting chorion gene
amplification (Asano and Wharton, 1999). It has been proposed that amplification may

104
be inhibited by the progression of replication forks into the amplification loci from
surrounding genomic origins activated by abundant ORC1; or activation of origins
throughout the genome may simply starve the amplification loci for scarce replication
factors (Asano and Wharton, 1999).
To understand molecularly whether ORC1 level and/or oscillation participates in
differential control of DAFC-62D amplification, ChIP experiments against ORC1 will be
the immediate next step. For example, ORC1 may be down-regulated or cleared away
from DAFC-62D in stages 11-12 when amplification initiation is quiescent. If ORC1
binding parallels that of ORC2, we will then be able to conclude that ORC association
with DAFC-62D is regulated as a whole complex rather than at the level of individual
subunits. It will also be interesting to test whether overexpressing other ORC subunits
such as ORC2 has similar inhibitory effect on amplification, as a hypomorphic orc2 allele
clearly reduces amplification (Landis et al., 1997a).
The +3.5 element is a stage 10-specific ORC2 binding site, but surprisingly it is
also required for stage 13 amplification, because deletion of +3.5 ablates amplification.
We propose that there is a specific time window in stage 10 for ORC binding that
requires the presence of all three sites to synergistically load ORC. Once such window is
missed, ORC loading may no longer be possible, resulting in complete abolishment of
amplification. The ORC-accessible period could be labeled by a developmentally unique
chromatin structure, and/or the presence of specification proteins. It would have been
informative to perform ChIP analysis and determine whether ORC is localized in these
deletions, but these transposons could not be distinguished from the endogenous
amplicon by real-time PCR. Chromatin DNA is typically sheared by sonication into

105
100bp to 1kb pieces in this assay. Thus without transposon-specific tags, we cannot
apply ChIP to analyze transposons that are larger than a couple of kilobases. In future
transformation experiments, it will be worth introducing small tag sequences into the
transposon to facilitate further analysis, as long as the tags are carefully inserted in a way
that is the least likely to interfere with functioning of cis elements carried by the
transposon.
MCM2-7 is required for both replication initiation and elongation. Consistent
with its role as a helicase, the absence of MCM2-7 in the vicinity of ori62 in stage 12
indicates that this complex has moved away from the origin with the elongating
replication forks. In stage 13, MCM2-7 is recruited back to license ori62. A similar
association pattern of MCM2-7 was found with the ACE3-ori62 transposon, which is
small enough (1.5kb) to survive sonication and provide molecules that carry both
transposon-specific and ACE3-ori62 sequences (see Appendix One). The reloading of
MCM2-7 in stage 13 may be regulated by a unique mechanism for the late round of
origin activation at DAFC-62D that is separated from earlier replication. Because the
other initiation events in stage 10B immediately follow previous genomic replication, it is
possible that little more than concentrating pre-RC onto DAFC origins is needed during
this developmental time. By contrast, the stage 13 initiation takes place at least four
hours after the initial round of amplification, and is preceded by another complex DNA-
mediated reaction, transcription, through ori62 (see below). The finely regulated
reloading of MCM2-7, therefore, may be the essential step to reactivate ori62.
Despite vigorous efforts, we have not been able to ChIP other components of the
pre-RC, probably because the antibodies are not optimal for IP. Nevertheless, we find

106
that ORC(2) remains associated with ori62, regardless of the activity of the origin; in
stage 10 and 13, two separate developmental stages, MCM2-7 is differentially recruited
to license an active origin and initiate amplification. This is reminiscent to the
observation that potential origins in yeast are marked by ORC binding throughout the cell
cycle and at the end of G1 ORC assembles the rest of the pre-RC (Mendez and Stillman,
2003).
Transcriptional regulation of replication initiation
Our findings suggest a mechanism by which origin selection and activation are
regulated by transcription in cis in Drosophila development. The striking inhibitory
effects of α-amanitin on DAFC-62D stage 13 origin firing are not due to a general defect
in replication, because it does not affect amplification of other amplicons or insulated
transposons. Therefore these data reflect a direct regulation of replication initiation by
RNAPII transcription in cis. This is of particular significance to ori62 firing, because it
exactly coincides with the coding region of yg2 that has to be transcribed in stage 12.
Although intuitively the passing through of transcriptional machinery might be imagined
to strip replication factors off the DNA and thus repress replication, our data suggest the
exact opposite.
However, a few questions remain unanswered. Firstly, is yg2 transcription THE
transcription required in cis? It is intriguing that in the 10kb central amplified region, the
1.1kb yg2 appears to be the only protein-encoding gene transcribed by RNAPII. The
only other annotated gene encodes a small Cysteine tRNA, which is usually processed by
RNA polymerase III and not affected by α-amanitin (Lindell et al., 1970; Wolffe, 1991).

107
Are there any other transcripts that may have missed annotation? We employed several
techniques to search for such unknown RNA products. Using several probes (spanning
2-3kb each) prepared from the 10kb fragment, we performed RNA FISH. While the
positive control yg2 consistently showed staining in follicle cells, other probes failed to
detect significant signals above background. In another attempt, total RNA was extracted
from whole ovaries, subjected to reverse transcription using random primers (as opposed
to polydT primers for mRNA), and screened by real-time PCR to search for positive PCR
amplification. Again no transcripts were found other than yg2, although the PCR screen
has not been saturated and small RNA products may have escaped detection. A third
method was to probe for any signals in total ovarian RNA by Northern blotting.
Preliminary results were negative, and further efforts to enrich and look for small RNAs
expressed in DAFC-62D were not a high priority, given the absence of any microRNAs
in the vicinity (the closest is about 600kb away), after a search in the small RNA
sequence database of Drosophila (Ruby and Bartel, personal communication).
We propose that RNAPII either directly recruits MCM2-7 through protein-protein
interaction, or indirectly affects the assembly of replication machinery by influencing
chromatin structure. Although not necessarily mutually exclusive, how can we
distinguish these possibilities? A direct physical interaction between RNAPII and
MCM2-7 has been reported in yeast (Gauthier et al., 2002; Holland et al., 2002). Thus a
straightforward experiment would be to test whether RNAPII and MCMs co-IP in follicle
cells, preferably in stage 13 specifically. Hand dissection of sufficient amount of stage
13 egg chambers for this experiment would be virtually impossible, but optimal
“fattening” of female ovaries may help to maximize late staged egg chambers and reduce

108
dissection work. Another added complexity is that this method requires purification of
follicle cells because the excessive proteins in the nurse cells and maturing oocytes are
likely to dilute away antibodies and interfere with follicular signals. Although follicle
cell nuclei can be enriched through FACS sorting, it may not be highly practical given the
massive amounts of samples needed.
To examine directly whether the role of RNAPII movement in displacing
proximal histones plays into the successful recruitment of MCM2-7 at ori62 (within the
yg2 gene coding region), we suggest nuclease (DNase I and MNase) sensitivity and
restriction enzyme accessibility assays. Does the chromatin structure around ori62
change in different developmental stages by displaying different sensibility/accessibility
to these enzymes? Does it become more open in stages 11-12 with increased level of
accessibility as a result of active transcription? Does α-amanitin decrease the openness of
the chromatin? How does chromatin around ACE3 and oriβ change with regard to
amplification and transcription activity, as well as developmental time? Answers to these
questions will provide definite insights into chromatin regulation of amplification
initiation.
The relevance of RNAPII and transcription (by itself) to replication initiation is
not after all surprising. Microarray-based genome-wide studies in yeast and higher
eukaryotes have revealed a recurring theme of gene-dense transcriptionally active regions
of the genome replicating before gene-sparse regions (MacAlpine and Bell, 2005). For
example, the Drosophila chromosome 2L microarray study in Kc cells uncovers
transcription/replication timing domains organized over 180kb, as suggested by an
association between sites of early/late BrdU incorporation, ORC localization and RNAPII

109
density (MacAlpine et al., 2004). These data suggest strong connections between
transcription and replication timing, although the influence of active transcription on the
process of replication is unclear. Our results reveal a positive role of RNAPII
transcription on DAFC-62D amplification, and provide a novel mechanism of
transcriptional regulation of replication initiation.
Distinct mechanisms of replication regulation
Unlike the well-characterized Saccharomyces cerevisiae origins that are defined
by an 11bp A-T-rich autonomously replicating sequence (ARS) consensus sequence and
other small elements (B1 and B2), metazoan origins and their regulation remain poorly
understood. First of all, there are different types of origins that are replicon and organism
specific: large zones of initiation and relatively defined origins (Gilbert, 2004). For
either class, no consensus sequence has been identified. Studies using DAFC as models
for replication analogously suggest the existence of both initiation zones and localized
replicators. For DAFC-66D, two small elements, ACE3 and oriβ, separated by only
1.5kb, are sufficient for proper regulation of amplification. By contrast, –3.0, ori62 and
+3.5 are dispersed in a 7kb region, suggesting a large control region necessary for
DAFC-62D origin activity.
In addition to different types of cis elements that contribute to origin identity and
activity, trans factors, especially transcription proteins, have been reported to help select
and license origins, which again, appears to vary case by case. In the Sciara salivary
gland DNA puff II/9A, amplification is controlled by ecdysone, potentially through direct
interaction with a putative ecdysone response element (EcRE) adjacent to its ORC-

110
binding site (Foulk et al., 2006). Similarly, a heterodimeric transcription activator
containing ecdysone receptor (EcR) mediates not only the transcription but also
amplification of at least some chorion genes in Drosophila (Hackney et al., 2007). At
DAFC-66D, the transcription factors Myb and E2F/RB associate with ORC via direct
protein-protein interaction (Beall et al., 2002; Bosco et al., 2001). Finally, histone-
modifying enzymes and/or chromatin-remodeling proteins may be recruited to modulate
DNA replication (Kohzaki and Murakami, 2005), as directly shown by tethering
experiments in X. laevis eggs (Danis et al., 2004) and DAFC models (Aggarwal and
Calvi, 2004).
Taken together, replication initiation is regulated at multiple levels. These
include sequence identity (especially A/T content), DNA topology, transcription factors,
and chromatin structure. Our findings that the process of transcription itself or the
movement of RNAPII prepares an origin (located within a gene’s coding region) for
firing provide yet another mechanism that is probably fine tuning origin activity in
response to developmental signals. To further our understanding, there are several
potential directions for future studies. First, the master hormone, ecdysone, may well be
the developmental cue that regulates DAFC-62D amplification in addition to the chorion
amplicons, although computational search for the highly degenerate EcRE
(PuG(G/T)T(C/G)A(N)TG(C/A)(C/A)(C/T)Py) (Antoniewski et al., 1993) did not yield
any positive hits in DAFC-62D (Xie and Orr-Weaver, unpublished results). Nonetheless,
in vitro culturing of ovaries in ecdysone titer (Buszczak et al., 1999), as well as
introduction of mutant forms of EcR (Hackney et al., 2007), combined with real-time

111
PCR analysis will allow direct examination of ecdysone’s effect on DAFC-62D
amplification level.
Second, it is important to test the involvement of the transcription factors Myb
and E2F/RB. We have formed collaboration with the Botchan lab to experimentally
search for Myb binding sites in DAFC-62D, and a genomic ChIP-chip has been
performed in Kc tissue culture cells. There were two strong and one weaker site in the
62D region that are at least 15 kb away from yg2. Binding in Kc cells may not predict
binding in follicle cells, because as previously observed, ACE3 did not appear to
associate with Myb in Kc cells, but did ChIP well in egg chambers. A ChIP-chip analysis
with staged egg chambers is underway (Lewis and Botchan, personal communication).
E2F1 mutations, on the other hand, did not significantly affect DAFC-62D amplification,
despite the presence of several predicted E2F1 binding sites (Xie and Orr-Weaver,
unpublished results).
Third, construction of a transposon carrying both ACE3 and ori62, with yg2
controlled by an exogenous promoter such as a heat-shock promoter will provide a useful
analytical tool. Without heat shock activation, such a transposon is expected to amplify
in a similar way as the endogenous DAFC-62D, as did the ACE3-ori62 transposon. If
transcriptional activity aggressively modulates replication initiation, will forced
transcription change the level and timing of transposon amplification? If so, ChIP
analyses of pre-RC components, RNAPII and histone modifications may reveal the
molecular mechanism, because this transposon will be small enough for such
manipulation.

112
Finally, after all the discussion about the uniquely activated stage 13 round of
DAFC-62D amplification, a fundamental question remains: is this strategy of cis-
transcriptional control used solely for DAFC-62D amplification initiation? Why is
amplification in stage 10 not affected by α-amanitin at DAFC-66D, -62D or 62D
transposons? A caveat to α-amanitin in vitro culturing is that stage 10 by itself is about
10 hours long, and the 5 hr α-amanitin treatment may not have been sufficient to induce
visible phenotypes. Further experiments with incubation time and α-amanitin
concentration may be needed.
Insulators and their insensitivity to α-amanitin
We used Drosophila SHWBS to protect transposons from chromosomal position
effects. Surprisingly, amplification and transcription of these insulated transposons are
not responsive to α-amanitin. This insensitivity can be reversed by introduction of the
transposon into a su(Hw) mutant background. Historically, these elements were
discovered for their enhancer-blocking activities when placed in between a transcriptional
promoter and enhancer (Geyer et al., 1986). Later Su(Hw) was reported to partially
protect transgenes from heterochromatin-mediated silencing in Drosophila (Roseman et
al., 1993). This system was then adopted in amplification analysis to reduce
chromosomal position effects (Lu and Tower, 1997). The molecular mechanism of
Drosophila insulator activity is not well understood; however, Su(Hw) has demonstrated
ability to target the chromatin fiber to insulator bodies (Gaszner and Felsenfeld, 2006;
Gerasimova et al., 2000). This protein together with two others (the POZ-domain
proteins CP190 and Mod(mdg4), modifier of mdg4), interacts with the ubiquitin ligase

113
Topoisomerase-I-interacting protein (Topors), which is bound to the nuclear lamina
(Capelson and Corces, 2005). As a consequence, these insulator elements come together
to form clustered insulator bodies. Although they are localized at the nuclear periphery
(Gerasimova et al., 2000), such localization is not essential at least to its enhancer-
blocking activity, which remains intact under heat shock conditions that have previously
been shown to disrupt the association of insulator, Su(Hw) and Mod(Mdg4) with the
nuclear periphery (Xu et al., 2004).
Studies in other systems provide clues how these elements may function to protect
against heterochromatin-mediated silencing. It has been proposed that insulators function
as chain terminators by modifying the nucleosomal substrate of the spreading
heterochromatin (Gaszner and Felsenfeld, 2006). The most extreme modification of the
template is nucleosome removal; various nucleosome-excluding sequence elements have
been shown to disrupt the spread of chromatin-mediated silencing (Bi et al., 2004). Other
forms of modification are achieved through the targeted recruitment of histone
acetyltransferases and ATP-dependent nucleosome-remodelling complexes (Oki et al.,
2004). Both nucleosome exclusion and the recruitment of histone- or nucleosome-
modifying complexes have important roles at endogenous yeast barrier elements (Donze
and Kamakaka, 2001; Oki and Kamakaka, 2005) and the complex vertebrate insulator
cHS4 in the chicken β-globin locus (Litt et al., 2001a; Litt et al., 2001b).
Therefore it is tempting to speculate that within the Drosophila insulator bodies,
there may be a relatively isolated and open chromatin structure. Supporting this idea, the
insulator itself contains several DNase I hypersensitive sites whose occurrence is
dependent on the binding of the Su(Hw) protein (Chen and Corces, 2001). The presence

114
of the insulator in the 5' region of the yellow gene increases the accessibility of the DNA
to nucleases in the promoter-proximal region (Chen and Corces, 2001). We thus propose
that the inhibition or slowing down of RNAPII by α-amanitin (Rudd and Luse, 1996)
may be compensated by the favorable chromatin environment, and therefore may allow
transcription of yg2 as well as the following round of amplification, in the presence of the
toxin. Some preliminary ChIP data analyzing histone acetylation levels of the ACE3-
ori62 transposon (see Appendix One) suggest that significantly different from the
endogenous amplicon (see Appendix Two), there is very little hyperacetylation on K8H4
in the transposon, whereas high levels of AcK8H4 are enriched in DAFC-62D. More
histone modifications need to be examined in order to understand the chromatin structure
of insulated transposons, as well as analyses of their nuclease sensitivity and restriction
enzyme accessibility.
Transcription factories
We observed that RNAPII localized to discrete subnuclear foci in Drosophila
follicle cells. Furthermore, it appears to switch from a nuclear staining to this foci pattern
at a time when these cells switch from genomic replication to localized amplification.
The RNAPII foci, however, do not significantly colocalize with BrdU incorporation spots
other than transiently with DAFC-62D in stage 12, and therefore are not likely sites
where other DAFC genes are being transcribed. Although we currently have no clue
what genes other than yg2 associate with these RNAPII loci, genes abundantly
transcribed in follicle cells at these times have been identified by microarray studies (R.
Duronio, personal communication) and provide good candidates. Eventually accurate

115
information may be collected from ChIP-chip analysis of RNAPII-associated genes.
Does the localization of RNAPII to subnuclear foci have any biological
significance? Studies of the human and mouse β-globin loci showed that promoters,
gene-proximal enhancers and far-upstream activators (which can be separated by many
kilobases) tend to co-localize within the nucleus in so-called chromatin hubs. The genes
controlled by these elements are transcribed when the hubs make contact with RNAPII
molecules, which are distributed as multimolecular aggregates (Jackson et al., 1998;
Osborne et al., 2004) within the nucleus and form “factories” for transcription. In
Drosophila it is not known whether hubs or transcriptional factories exist. Our findings
are the first evidence that such structures may be formed in at least Drosophila follicle
cells, perhaps in response to developmental regulation in order to efficiently transcribe
active genes. Intriguingly, α-amanitin treatment disrupts the foci pattern of RNAPII,
arguing that these “factories” may be dynamic structures as opposed to fixed RNAPII
aggregates.
Another remaining question is whether the RNAPII foci are composed of active or
inactive polymerases. There are two major forms of RNAPII, the active elongating form
marked by multiple phosphorylations on its C-terminal repeat domain (CTD), RNAPII0,
and the nonphosphorylated inactive form RNAPIIA that associates with inactive genes
and pauses at promoter-proximal sites (Phatnani and Greenleaf, 2006). The current
working antibodies recognizes both forms of RNAPII. Several other antibodies specific
for either form have been tested but worked poorly, providing only a nuclear staining that
looked like background noise. Optimization of fixing and staining conditions will be
necessary. Meanwhile, given the increasing number of foci seen in later stages, it will be

116
interesting to quantify them and perhaps correlate foci number with developmental
stages, and begin to search for patterns of localization within the nuclei.

117
REFERENCES
Aggarwal, B. D., and Calvi, B. R. (2004). Chromatin regulates origin activity inDrosophila follicle cells. Nature 430, 372-376.Antoniewski, C., Laval, M., and Lepesant, J. A. (1993). Structural features critical to theactivity of an ecdysone receptor binding site. Insect Biochem Mol Biol 23, 105-114.Asano, M., and Wharton, R. P. (1999). E2F mediates developmental and cell cycleregulation of ORC1 in Drosophila. Embo J 18, 2435-2448.Beall, E. L., Manak, J. R., Zhou, S., Bell, M., Lipsick, J. S., and Botchan, M. R. (2002).Role for a Drosophila Myb-containing protein complex in site-specific DNA replication.Nature 420, 833-837.Bi, X., Yu, Q., Sandmeier, J. J., and Zou, Y. (2004). Formation of boundaries oftranscriptionally silent chromatin by nucleosome-excluding structures. Mol Cell Biol 24,2118-2131.Bosco, G., Du, W., and Orr-Weaver, T. L. (2001). DNA replication control throughinteraction of E2F-RB and the origin recognition complex. Nat Cell Biol 3, 289-295.Buszczak, M., Freeman, M. R., Carlson, J. R., Bender, M., Cooley, L., and Segraves, W.A. (1999). Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 126, 4581-4589.Capelson, M., and Corces, V. G. (2005). The ubiquitin ligase dTopors directs the nuclearorganization of a chromatin insulator. Mol Cell 20, 105-116.Chen, S., and Corces, V. G. (2001). The gypsy insulator of Drosophila affects chromatinstructure in a directional manner. Genetics 159, 1649-1658.Danis, E., Brodolin, K., Menut, S., Maiorano, D., Girard-Reydet, C., and Mechali, M.(2004). Specification of a DNA replication origin by a transcription complex. Nat CellBiol 6, 721-730.Donze, D., and Kamakaka, R. T. (2001). RNA polymerase III and RNA polymerase IIpromoter complexes are heterochromatin barriers in Saccharomyces cerevisiae. Embo J20, 520-531.Foulk, M. S., Liang, C., Wu, N., Blitzblau, H. G., Smith, H., Alam, D., Batra, M., andGerbi, S. A. (2006). Ecdysone induces transcription and amplification in Sciaracoprophila DNA puff II/9A. Dev Biol 299, 151-163.Gaszner, M., and Felsenfeld, G. (2006). Insulators: exploiting transcriptional andepigenetic mechanisms. Nat Rev Genet 7, 703-713.Gauthier, L., Dziak, R., Kramer, D. J., Leishman, D., Song, X., Ho, J., Radovic, M.,Bentley, D., and Yankulov, K. (2002). The role of the carboxyterminal domain of RNApolymerase II in regulating origins of DNA replication in Saccharomyces cerevisiae.Genetics 162, 1117-1129.Gerasimova, T. I., Byrd, K., and Corces, V. G. (2000). A chromatin insulator determinesthe nuclear localization of DNA. Mol Cell 6, 1025-1035.Geyer, P. K., Spana, C., and Corces, V. G. (1986). On the molecular mechanism ofgypsy-induced mutations at the yellow locus of Drosophila melanogaster. Embo J 5,2657-2662.Gilbert, D. M. (2004). In search of the holy replicator. Nat Rev Mol Cell Biol 5, 848-855.

118
Hackney, J. F., Pucci, C., Naes, E., and Dobens, L. (2007). Ras signaling modulatesactivity of the ecdysone receptor EcR during cell migration in the Drosophila ovary. DevDyn 236, 1213-1226.Holland, L., Gauthier, L., Bell-Rogers, P., and Yankulov, K. (2002). Distinct parts ofminichromosome maintenance protein 2 associate with histone H3/H4 and RNApolymerase II holoenzyme. Eur J Biochem 269, 5192-5202.Jackson, D. A., Iborra, F. J., Manders, E. M., and Cook, P. R. (1998). Numbers andorganization of RNA polymerases, nascent transcripts, and transcription units in HeLanuclei. Mol Biol Cell 9, 1523-1536.Kohzaki, H., and Murakami, Y. (2005). Transcription factors and DNA replication originselection. Bioessays 27, 1107-1116.Landis, G., Kelley, R., Spradling, A. C., and Tower, J. (1997). The k43 gene, required forchorion gene amplification and diploid cell chromosome replication, encodes theDrosophila homolog of yeast origin recognition complex subunit 2. Proc Natl Acad Sci US A 94, 3888-3892.Lindell, T. J., Weinberg, F., Morris, P. W., Roeder, R. G., and Rutter, W. J. (1970).Specific inhibition of nuclear RNA polymerase II by alpha-amanitin. Science 170, 447-449.Litt, M. D., Simpson, M., Gaszner, M., Allis, C. D., and Felsenfeld, G. (2001a).Correlation between histone lysine methylation and developmental changes at thechicken beta-globin locus. Science 293, 2453-2455.Litt, M. D., Simpson, M., Recillas-Targa, F., Prioleau, M. N., and Felsenfeld, G. (2001b).Transitions in histone acetylation reveal boundaries of three separately regulatedneighboring loci. Embo J 20, 2224-2235.Lu, L., and Tower, J. (1997). A transcriptional insulator element, the su(Hw) binding site,protects a chromosomal DNA replication origin from position effects. Mol Cell Biol 17,2202-2206.MacAlpine, D. M., and Bell, S. P. (2005). A genomic view of eukaryotic DNAreplication. Chromosome Res 13, 309-326.MacAlpine, D. M., Rodriguez, H. K., and Bell, S. P. (2004). Coordination of replicationand transcription along a Drosophila chromosome. Genes Dev 18, 3094-3105.Mendez, J., and Stillman, B. (2003). Perpetuating the double helix: molecular machinesat eukaryotic DNA replication origins. Bioessays 25, 1158-1167.Ohta, S., Tatsumi, Y., Fujita, M., Tsurimoto, T., and Obuse, C. (2003). The ORC1 cyclein human cells: II. Dynamic changes in the human ORC complex during the cell cycle. JBiol Chem 278, 41535-41540.Oki, M., and Kamakaka, R. T. (2005). Barrier function at HMR. Mol Cell 19, 707-716.Oki, M., Valenzuela, L., Chiba, T., Ito, T., and Kamakaka, R. T. (2004). Barrier proteinsremodel and modify chromatin to restrict silenced domains. Mol Cell Biol 24, 1956-1967.Osborne, C. S., Chakalova, L., Brown, K. E., Carter, D., Horton, A., Debrand, E.,Goyenechea, B., Mitchell, J. A., Lopes, S., Reik, W., and Fraser, P. (2004). Active genesdynamically colocalize to shared sites of ongoing transcription. Nat Genet 36, 1065-1071.Phatnani, H. P., and Greenleaf, A. L. (2006). Phosphorylation and functions of the RNApolymerase II CTD. Genes Dev 20, 2922-2936.

119
Roseman, R. R., Pirrotta, V., and Geyer, P. K. (1993). The su(Hw) protein insulatesexpression of the Drosophila melanogaster white gene from chromosomal position-effects. Embo J 12, 435-442.Rudd, M. D., and Luse, D. S. (1996). Amanitin greatly reduces the rate of transcriptionby RNA polymerase II ternary complexes but fails to inhibit some transcript cleavagemodes. J Biol Chem 271, 21549-21558.Tatsumi, Y., Ohta, S., Kimura, H., Tsurimoto, T., and Obuse, C. (2003). The ORC1 cyclein human cells: I. cell cycle-regulated oscillation of human ORC1. J Biol Chem 278,41528-41534.Wolffe, A. P. (1991). RNA polymerase III transcription. Curr Opin Cell Biol 3, 461-466.Xu, Q., Li, M., Adams, J., and Cai, H. N. (2004). Nuclear location of a chromatininsulator in Drosophila melanogaster. J Cell Sci 117, 1025-1032.

120
Appendix One
Analyses of the ACE3-ori62 Transposon

121
The smallest transposon that shows amplification contains the 300bp ACE3 and
the 1kb ori62 origin. The level and timing of its amplification recapitulate the
endogenous amplicon (Figure 1A). ori62 by itself does not amplify (Figure 1A). ACE3 as
multimers has been shown to be able to stimulate amplification presumably from nearby
genomic origins, when inserted into ectopic sites (Carminati et al., 1992). The ACE3
multimer recruits ORC at such high levels that it is possible to detect a focus by
immunofluorescence (Austin et al., 1999). Both in vivo and in vitro studies show that
ORC specifically associates with ACE3 (Austin et al., 1999). Therefore the function of
ACE3 at this “minimal” transposon is probably to help to recruit an adequate amount of
ORC to license ori62. When ACE3 in the transposon with ori62 was replaced by the
500bp +3.5 element, which was bound by ORC in ChIP experiments, no amplification
occurred (Figure 1A). This may be because the +3.5 element did not bind and recruit
ORC with the same efficiency as ACE3 to achieve the threshold level needed for
amplificaiton. The +3.5-ori62 transposon may need a third ORC-binding sequence such
as the -3.0 element of DAFC-62D, also pulled down by ORC in ChIP, to fulfill the ORC
threshold requirement. It will be interesting to test whether a true minimal transposon
would be the one that contains all three ORC binding elements: -3.0, ori62 and +3.5.
The fact that the ACE3-ori62 transposon displayed the same timing of
amplification suggested that ori62 conferred sufficient origin activity for not only stage
10 but also stage 13 amplification, although by nascent strand analysis we were only able
to define the origin in stage 10B. Somewhat surprisingly, ACE3 did not cause higher
levels of amplification from ori62 (or from the whole 10kb central amplified fragment

122
Figure 1. Intrinsic origin activity not influenced by ACE3. (A) ori62 amplified in the
presence of ACE3 (but not +3.5), at the level comparable to the endogenous locus. (B) A
transposon containing multiple copies of ACE3 stimulated amplification, the timing and
extent of which appeared to be insertion-site specific.
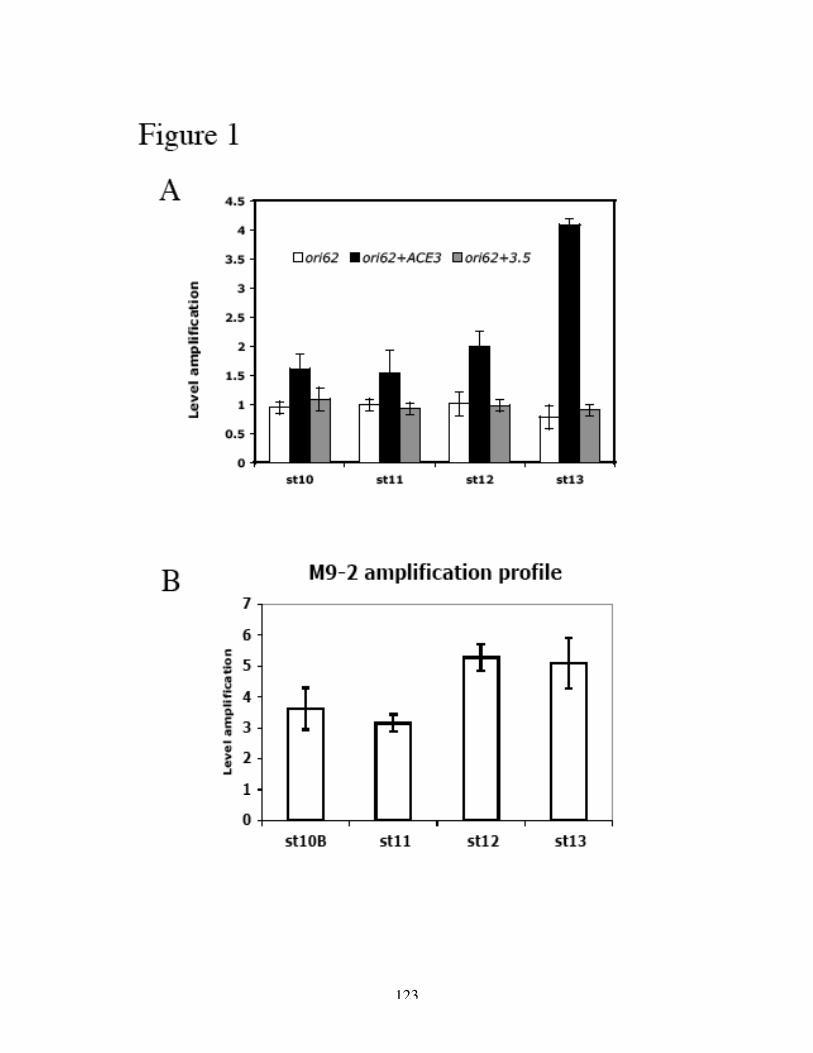
123

124
from DAFC-62D), suggesting there may be an intrinsic origin activity that determines the
extent and timing of replication initiation that is not altered by amplification enhancers.
Consistent with this hypothesis, the transposon carrying multiple copies of ACE3
(Carminati et al., 1992) causes amplification from nearby origin(s) at its insertion site in a
pattern that is once again different from DAFC-66D (Figure 1B). To begin to understand
the nature of such origin activity, we turned to ChIP to examine first the presence of
some trans factors in the transposon.
The size of the ACE3-ori62 transposon is 1.5 kb, small enough to allow ChIP
analysis of protein localization within it, because transposon-specific sequences (real-
time PCR targets, usually 50-70 bp products) are close enough to the origin so that both
are likely present in the same sheared chromatin molecule. MCM appeared to associate
with ACE3-ori62 and displayed a similar timing as at DAFC-62D, showing localization
in stages 10 and 13, but not stages 11-12 (Figure 2). The low levels of enrichment, as
well as the large error bars, were likely due to low amounts of big molecules that
contained both the PCR target and ori62 sequences (e.g. 1kb above). We have not yet
analyzed ORC, although it is expected given the observed amplification.
Next we tested whether RNAPII was recruited to the transposon, although all
upstream sequences of the yg2 gene were absent in the transposon and no active
transcription could be detected by RNA FISH of yg2 (Figure 3A, the subnuclear signal
corresponds to the endogenous yg2 transcription site). Intriguingly, RNAPII was
localized significantly to ACE3-ori62 (Figure 3B), mimicking its binding pattern to the
endogenous ACE3 locus at DAFC-66D in an independent ChIP experiment (Figure 3C).
We speculate that because ACE3 associates with the transcription factors Myb

125
Figure 2. Association of MCM2-7 with the transposon determined by ChIP and
real-time PCR. Only large enough chromatin that carried both PCR targets (transposon
specific) and ori62 (bound by MCM) could be detected. This was very likely a small pool
of molecules, as DNA was on average sheared into 100bp to 1kb pieces.
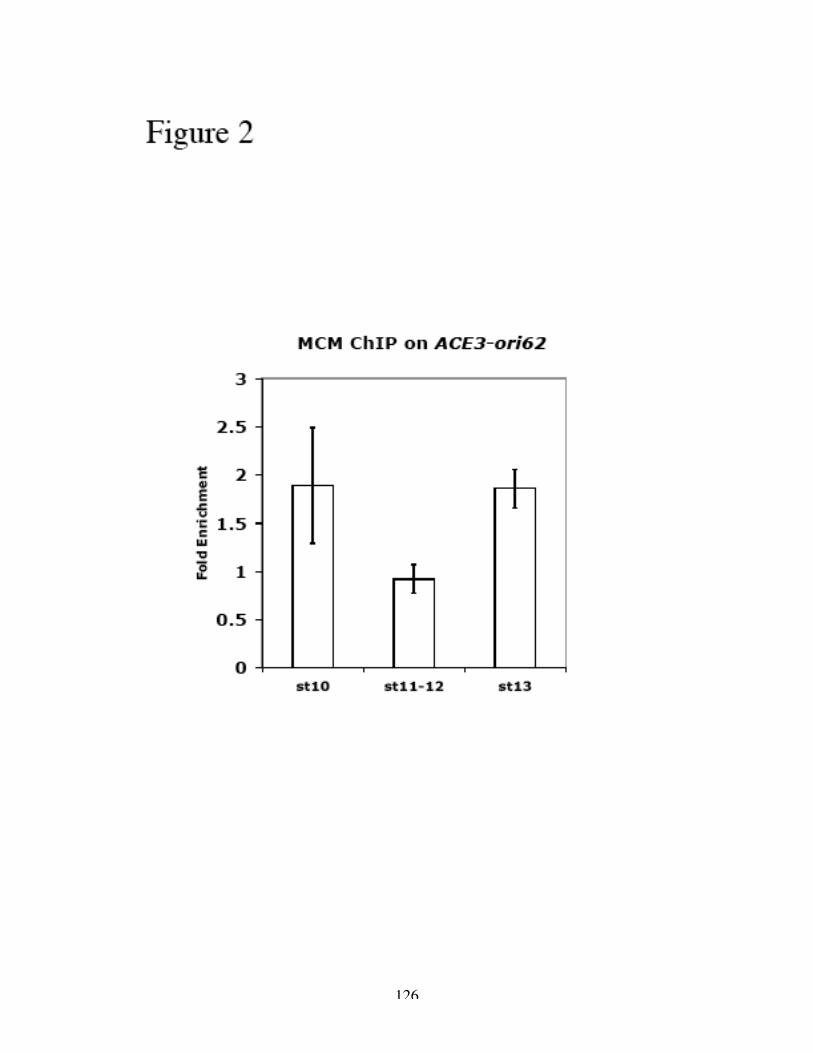
126

127
Figure 3. RNAPII associates with the transposon despite absence of active
transcription. (A) yg2 RNA FISH in stage 12 follicle cells containing the transposon.
The single nuclear signal within each cell represented nascent transcripts at the
endogenous locus. No signal was detected from the transposon. (B) RNAPII ChIP
suggested significant association with the transposon. (C) RNAPII level at the
endogenous ACE3 was similar to the transposon.
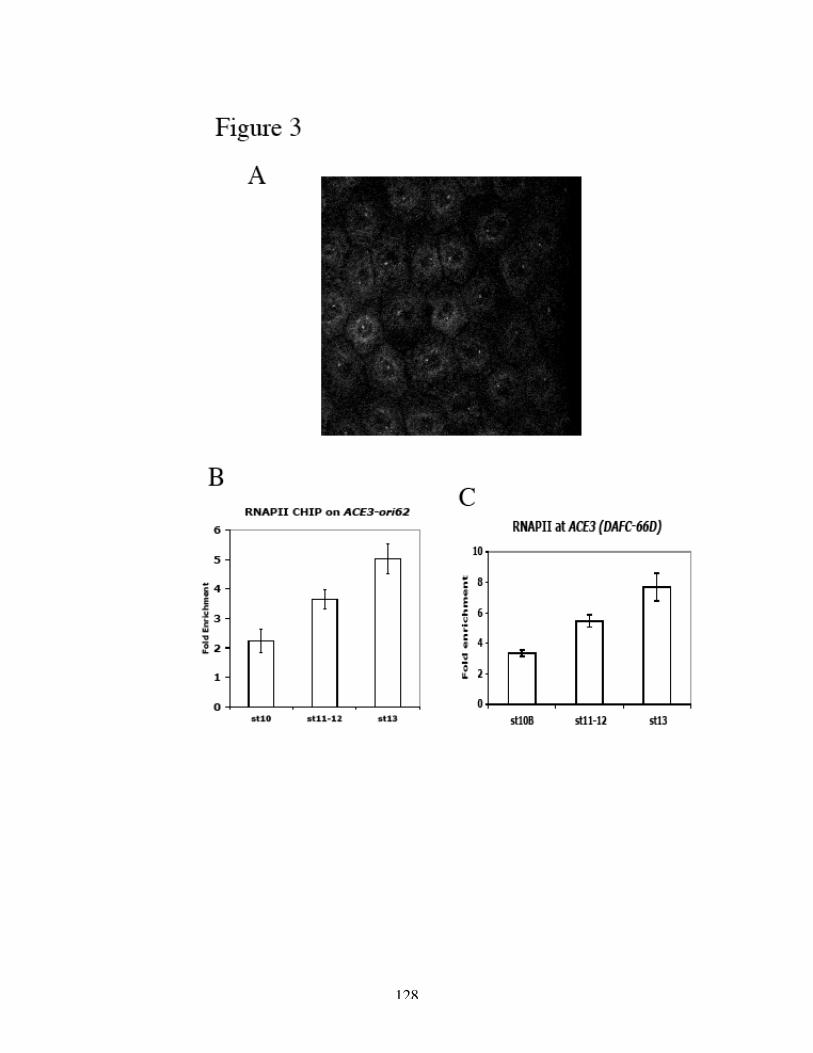
128

129
(Beall et al., 2002) and E2F1 (Bosco et al., 2001), RNAPII may ultimately be recruited
through protein-protein interactions without the requirement of promoter sequences.
Alternatively, the insulator elements have been proposed to act in a way analogous to a
promoter (Cai et al., 2001), and therefore may independently recruit RNAPII. Although
not shown for the Su(Hw) insulators, RNAPII has been clearly demonstrated to interact
with the CTCF protein that mediates the insulator activity that lies within the chicken β-
globin locus (Chernukhin et al., 2007).
Like other insulated transposons, ACE3-ori62 was not sensitive to α-amanitin
(Figure 4A). As a control, the endogenous DAFC-62D was examined in the same DNA
sample, and at least for the locus 1.5kb away from ori62, the stage 13 amplification is
specifically inhibited (Figure 4B). The association of MCM2-7 and RNAPII with the
insulated transposon remained unchanged by the toxin (Figure 4C). When crossed into
the su(Hw) mutant background, however, the only line tested so far failed to amplify,
presumably repressed by position effects (data not shown). More transformation lines
need to be examined to investigate further the insensitivity of insulated transposons to α-
amanitin, combined with ChIP analyses of protein localization.
Finally, we have begun to study the chromatin structure of the transposons using
the ChIP technique against modified histones (see Appendix Two). The first modification
tested was AcK8H4 (Figure 5). In contrast to the endogenous amplicons, very little
enrichment of hyperacetylated K8H4 was found at the ACE3-ori62 transposon. It is
possible that these insulated structures contain other chromatin characteristics, and more
modifications, including acetylation and methylation of both histones H4 and H3, need to

130
Figure 4. α-amanitin did not affect the transposon. (A) The transposon amplification
level was unchanged by α-amanitin. (B) The endogenous DAFC-62D stage 13
amplification was inhibited by α-amanitin. The DNA prep was the same as in (A). A
locus 1.5kb away from ori62 (not present in the transposon) was tested in real-time PCR.
(C) Neither MCM2-7 nor RNAPII changed association with the transposon after α-
amanitin treatment.
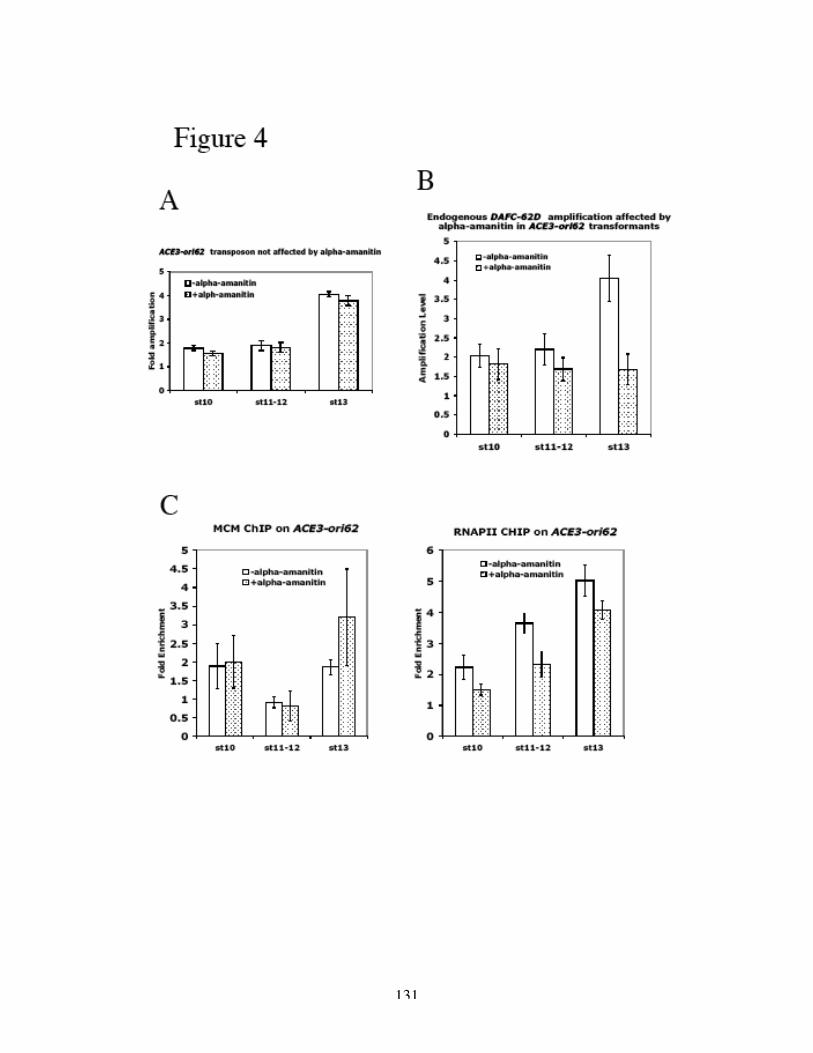
131

132
Figure 5. Histone H4 Lysine 8 was not hyperacetylated on the ACE-ori62
transposon. Very little enrichment of AcK8H4 was observed over an independent
control locus, either with or without α-amanitin treatment.
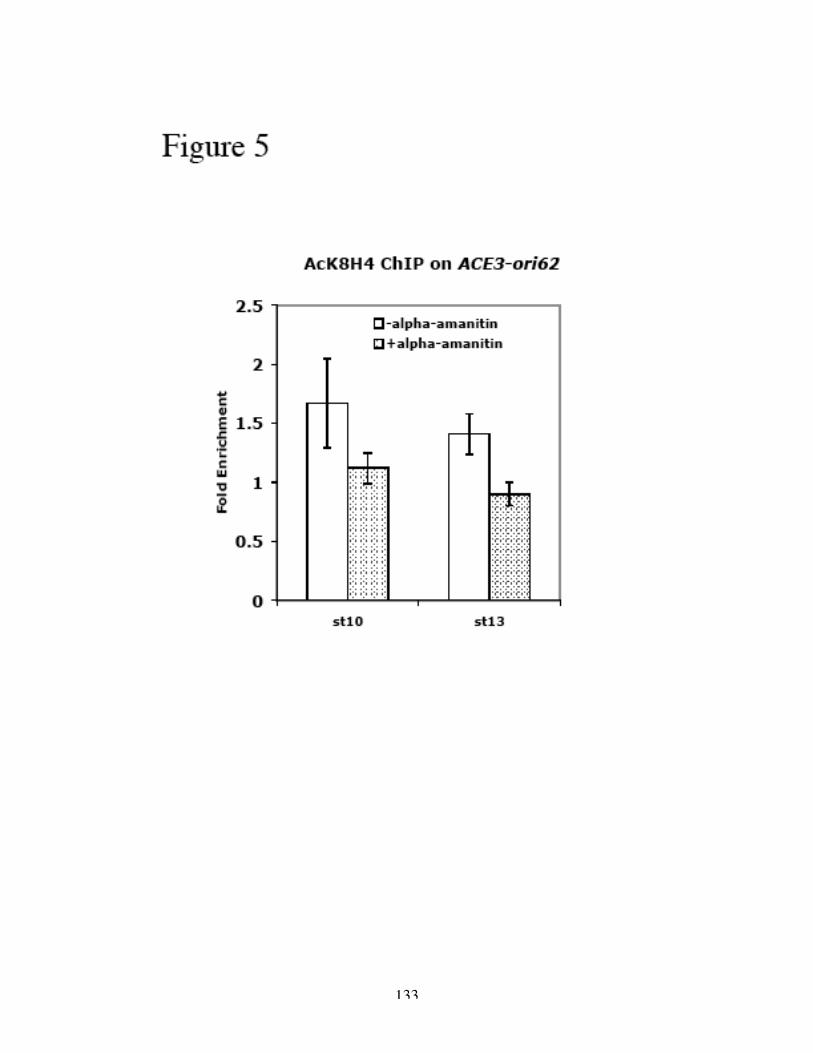
133

134
be examined. It will be of particular interest to study the unamplified ACE3-ori62 in the
mutant su(Hw) background, as well as to test how α-amanitin affects chromatin status.
Our analyses of the ACE3-ori62 transposon provided further evidence that
RNAPII regulated amplification initiation. We have previously proposed two hypotheses:
Proximal RNAPII directly recruits MCM2-7 to the origin; or RNAPII movement helps to
remodel origin chromatin to allow loading of MCM2-7. The fact that in the absence of
active transcription (at least no detectable yg2 transcription) RNAPII still localized to the
transposon, together with the accurate recapitulation of the endogenous amplification
pattern by the transposon, argues against the latter scenario. It is still possible, however,
that the transformation line tested in the RNAPII ChIP experiments had ACE3-ori62
inserted into an actively transcribing region, and investigation of more lines is required to
understand the exact mechanism.

135
REFERENCES
Austin, R. J., Orr-Weaver, T. L., and Bell, S. P. (1999). Drosophila ORC specificallybinds to ACE3, an origin of DNA replication control element. Genes Dev 13, 2639-2649.Beall, E. L., Manak, J. R., Zhou, S., Bell, M., Lipsick, J. S., and Botchan, M. R. (2002).Role for a Drosophila Myb-containing protein complex in site-specific DNA replication.Nature 420, 833-837.Bosco, G., Du, W., and Orr-Weaver, T. L. (2001). DNA replication control throughinteraction of E2F-RB and the origin recognition complex. Nat Cell Biol 3, 289-295.Cai, H. N., Zhang, Z., Adams, J. R., and Shen, P. (2001). Genomic context modulatesinsulator activity through promoter competition. Development 128, 4339-4347.Carminati, J. L., Johnston, C. G., and Orr-Weaver, T. L. (1992). The Drosophila ACE3chorion element autonomously induces amplification. Mol Cell Biol 12, 2444-2453.Chernukhin, I., Shamsuddin, S., Kang, S. Y., Bergstrom, R., Kwon, Y. W., Yu, W.,Whitehead, J., Mukhopadhyay, R., Docquier, F., Farrar, D., et al. (2007). CTCF interactswith and recruits the largest subunit of RNA polymerase II to CTCF target sites genome-wide. Mol Cell Biol 27, 1631-1648.

136
Appendix Two
Histone Acetylation and Amplification Activity

137
Eukaryotic DNA is packaged by histone proteins into chromatin, an organized,
higher-order structure. The N-terminal tails of histones are subject to post-translational
modifications such as acetylation, methylation, phosphorylation and ubiquitination
(Kouzarides, 2007). These modifications, together with DNA methylation, control the
folding of the nucleosomal array into higher-order structures that are essential for the
execution of DNA-mediated processes including transcription, DNA replication, DNA
repair and DNA recombination (Fuchs et al., 2006). The relationship between histone
acetylation and gene expression has been studied for decades. It is well established that
in the transcriptionally active portions of the genome, DNA is more accessible to
nucleases, and nucleosomes carry a combinatorial pattern of many post-translational
modifications, which include high levels of acetylation and methylation of H3K4 and
H3K79 (Groth et al., 2007).
More recently, a great deal of evidence has accumulated showing that not only
transcription but other DNA-mediated reactions also are regulated by histone
modifications (Fukuda et al., 2006). It is relatively well understood how during DNA
repair histone modifications act as signals and landing platforms for various repair
proteins (Altaf et al., 2007). Recent studies also suggest a potential role of chromatin
structure in replication control. For example, the positioning of nucleosomes is important
for replication initiation in yeast ARS (Brown et al., 1991; Lipford and Bell, 2001;
Simpson, 1990). Replication timing is regulated by histone deacetylation and acetylation
(Aparicio et al., 2004; Vogelauer et al., 2002).

138
Some instances of histone modifications regulating the selection and licensing of
replication origins have emerged from viral DNA studies, although sometimes conflicting
ones. The minimal replicator sequence of the Epstein-Barr virus (EBV) origin of plasmid
replication (OriP) is flanked by nucleosomes that in late G1 are subject to chromatin
remodeling and histone H3 deacetylation, coinciding with MCM3 loading and preceding
the onset of DNA replication (Zhou et al., 2005). On the other hand, the latent replication
origin of the viral genome of Kaposi's sarcoma-associated herpesvirus is bound by
ORC2, and is enriched in hyperacetylated histones H3 and H4. MCM3 also binds to the
origin in late-G1/S-arrested cells, which coincides with the loss of histone H3 K4
methylation (Stedman et al., 2004).
A role for histone acetylation in DNA replication has been suspected, because an
acetyltransferase, HBO1 (histone acetyltransferase binding to ORC1), is isolated as a
binding partner for ORC1 in human cell extracts (Iizuka and Stillman, 1999). A yeast
two-hybrid screen for MCM2-interacting proteins also identifies HBO1 (Burke et al.,
2001). In a separate study HBO1 is shown to augment the assembly of the pre-RC and
the recruitment of MCMs to chromatin; when Xenopus Hbo1 is immunodepleted,
chromatin binding of Mcm2-7 is lost and DNA replication is abolished in Xenopus egg
extracts (Iizuka et al., 2006). Finally, HBO1 complexes with some members of the ING
family of tumor suppressors, which are required for normal progression through S phase
and the majority of histone H4 acetylation in vivo (Doyon et al., 2006). Some of these
complexes interact with the MCM helicase and are essential for replication, because
HBO1 RNAi reduces DNA synthesis (Doyon et al., 2006). Taken together, these

139
findings suggest that HBO1, via its ability to acetylate histone H4, is required for S phase
initiation and replication initiation.
In the gene amplification model DAFC systems, it recently has been shown that
hyperacetylated histone H4 coincides with chorion amplicon origins (Aggarwal and
Calvi, 2004; Hartl et al., 2007). Tethering histone deacetylase reduced amplification of a
transposon carrying ACE3 and oriβ, whereas tethering histone acetyl transferase
(including the HBO1 homolog Chameau) increased amplification levels (Aggarwal and
Calvi, 2004). These observations suggest that histone acetylation status has a definite
role in regulating amplicon origin activity. However, the molecular mechanism remains
unclear, particularly how chromatin modification correlates with ORC binding,
subsequent pre-RC assembly and/or involvement of transcription factors.
We therefore have begun to survey systematically the acetylation level of histone
H4 across DAFC-66D and DAFC-62D, two differentially regulated amplicons, to explore
whether it could account for the reduced number of rounds of amplification at DAFC-
62D compared to -66D, the late initiation in stage 13 at DAFC-62D, and the effect of
transcription on stage 13 amplification. We performed ChIP experiments with antibodies
against pan-Acetyl-H4 (pan-AcH4), Acetyl-H4-K5 (AcK5H4) or Acetyl-H4-K8
(AcK8H4) on staged egg chambers. The immunostaining of all three show subnuclear
foci of staining at DAFCs (Hartl et al., 2007). For DAFC-66D, the level of pan-AcH4 at
ACE3 increased during follicle cell differentiation from stage 10 to 13 (Figure 1A).
AcK8H4 was enriched specifically at ACE3 and oriβ in stages 10 through 13, although
the enrichment level decreased with developmental progression (Figure 1B). AcK5H4,

140
Figure 1. Acetylation pattern of histone H4 in DAFC-66D during amplification.
(A) Pan-AcH4 in stages 10 and 13 at ACE3. More comprehensive acetylation profiles
across DAFC-66D were constructed from ChIP data against AcK8H4 in (B) and AcK5H4
in (C).
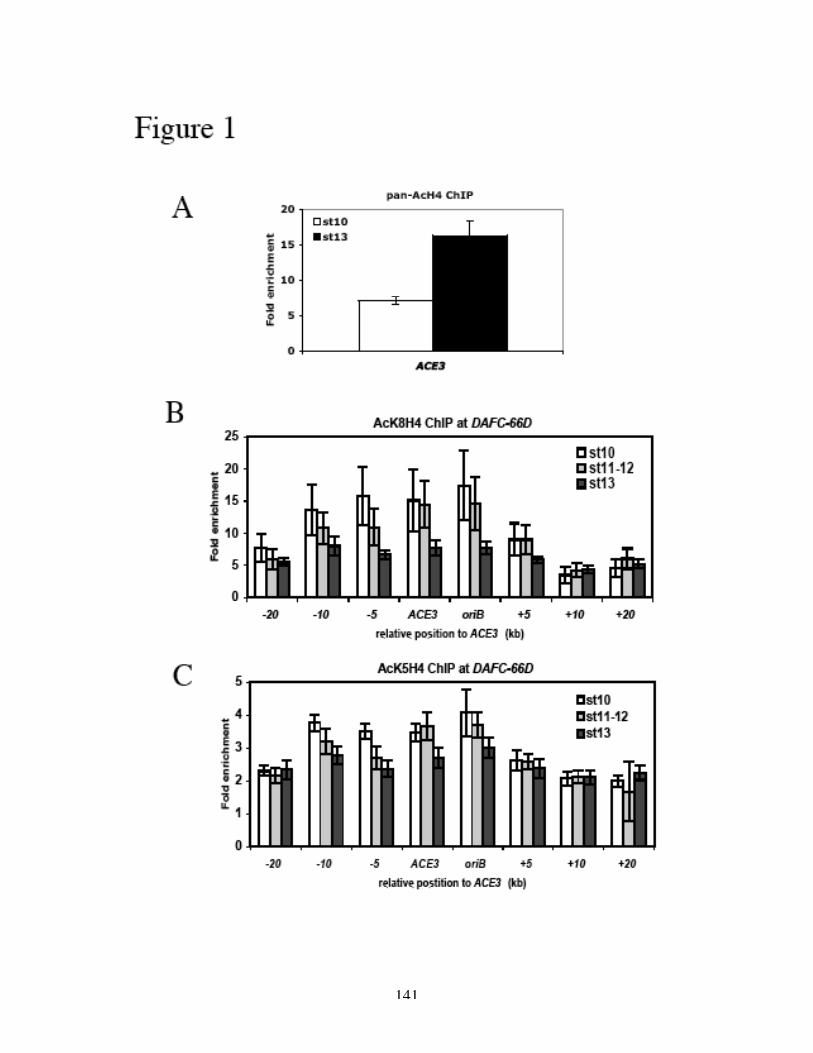
141

142
Figure 2. Association pattern of ORC2 (top panel), MCM2-7 (middle panel) and
RNAPII (bottom panel) with DAFC-66D were shown for comparison against H4
acetylation. All ChIP experiments were independently performed, and some sampling
methods may slightly differ. For example, stage 12 egg chambers were used in both
ORC2 and MCM2-7 ChIP, whereas for RNAPII it was stages 11 and 12 combined.
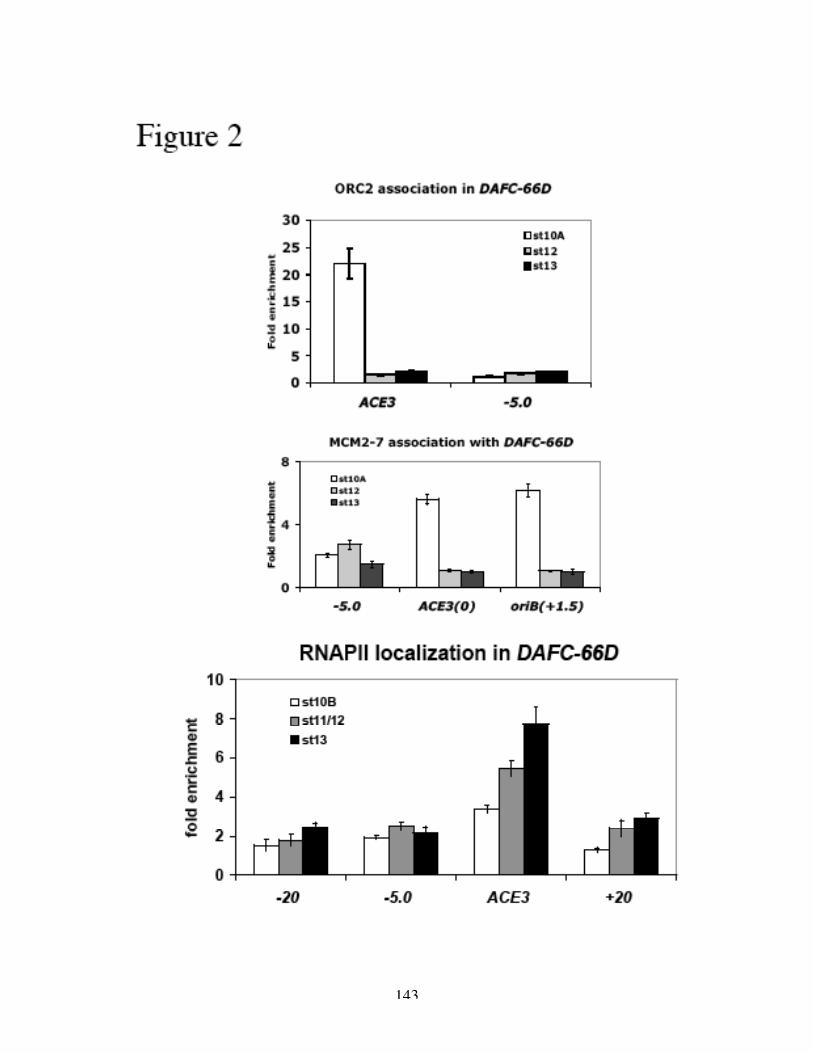
143

144
on the other hand, was merely detectable (Figure 1C, note the difference in scale of the Y
axis). AcK5H4 is associated with de novo histone deposition during replication
(Kouzarides, 2007), and thus the consistently observed two to three fold of enrichment
may reflect doubling of the chromatin at any given time.
In search for correlations between AcK8H4 and protein localization, the
association profiles for ORC2, MCM2-7 and RNAPII (all determined by ChIP) are
shown for comparison in Figure 2. Their changes with respect to developmental time are
summarized in Figure 6A. While both ORC2 and MCM2-7 diminished after stage 10,
RNAPII was detected at a higher level in later stages. Therefore it is tempting to
speculate that in stage 10 high AcK8H4 levels correlate with pre-RC assembly at DAFC-
66D. It is noteworthy, however, that AcK8H4 has been tightly linked with transcription
(Kouzarides, 2007), and the high levels in stages 11-12 may also be a marker for active
transcription.
The acetylation level of H4 was similarly analyzed for DAFC-62D. Around two
fold of enrichment of AcK5H4 (Figure 3A, top panel) and high amounts of AcK8H4
(Figure 3B, upper panel) were found. In comparison with DAFC-66D, the enrichment
level of AcK8H4 in stage 10 was two to four-fold higher for DAFC-66D over 62D
(Figures 1B and 3B). This significant difference raises an intriguing possibility that
higher acetylation levels may correspond to the much higher origin activity of oriβ that
gives rise to more rounds of replication initiation at DAFC-66D. In stages 11-12,
AcK8H4 level significantly elevated, coinciding with active transcription of yg2 (Figure
3B). The sudden drop of acetylation in stage 13 (Figure 3B) was unexpected, given
another round of amplification during this stage. However, it overlaps with loss of yg2

145
Figure 3. Acetylation pattern of histone H4 in DAFC-62D and the effect of α-
amanitin.
(A) AcK5H4 was barely detectable with or without α-amanitin treatment. (B) High levels
of AcK8H4 were found in sequences upstream of ori62 (in the yg2 gene). α-amanitin
augmented AcK8H4 levels.
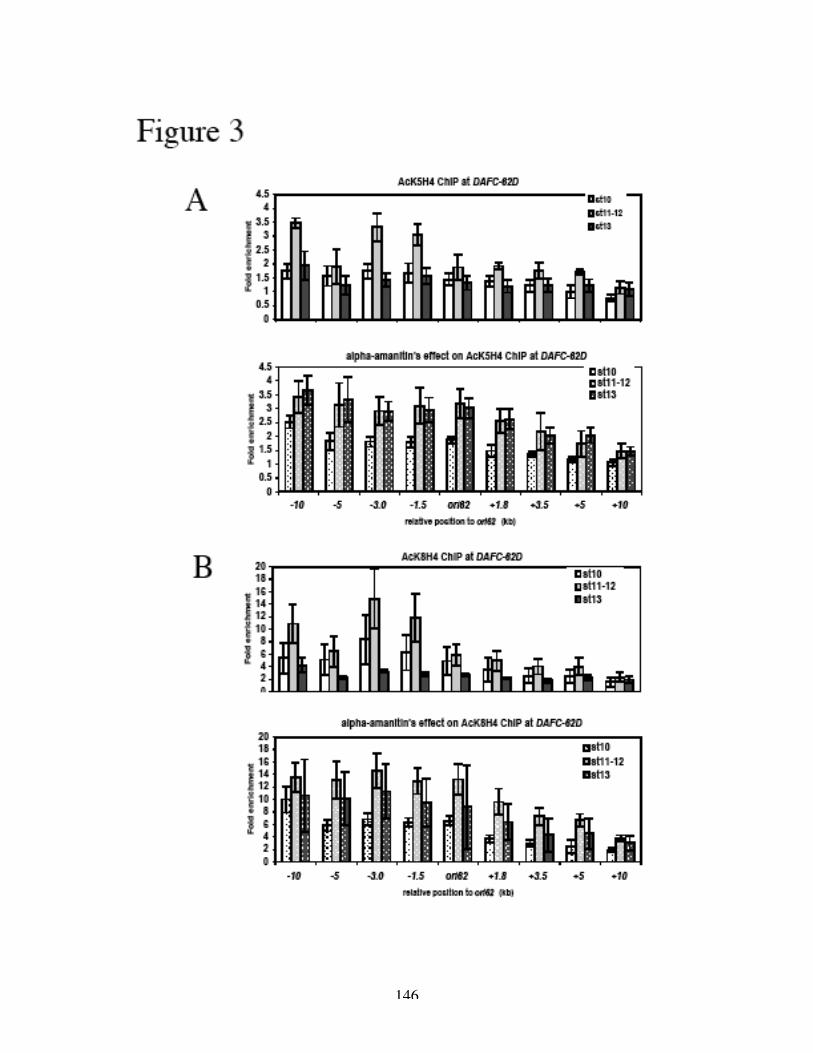
146

147
Figure 4. α-amanitin’s effect on pan-AcH4 levels in DAFC-62D.
Significantly elevated levels at several representative sites across DAFC-62D were
induced by the toxin in stage 13 (lower panel).
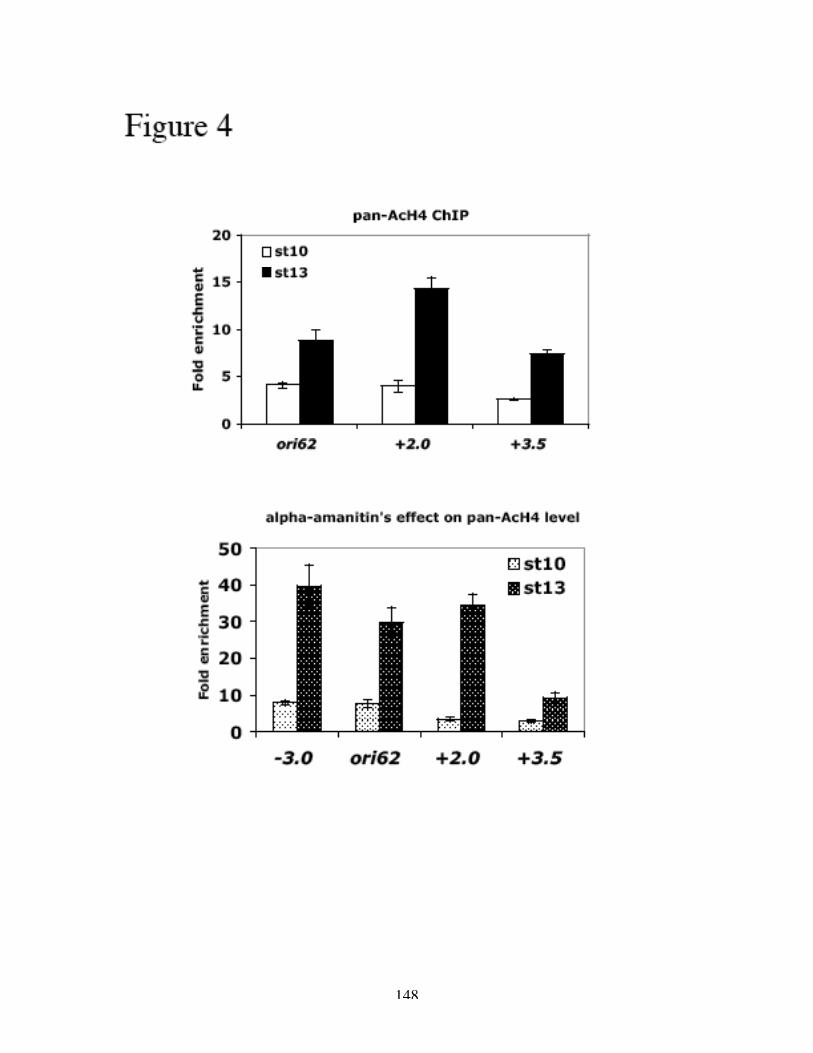
148

149
transcription, arguing that from stage 11 on, in order to coordinate transcription activation
and repression, AcK8H4 may be recognized as a transcription marker as opposed to one
specifically for replication.
We also examined whether α-amanitin affected H4 acetylation. Consistent with
the proposal that AcK8H4 does not correlate with replication activation in later stages, its
levels were actually elevated in stage 13 after α-amanitin treatment, while the stage 13
round of amplification was specifically inhibited (Figures 3B, lower panel). When pan-
AcH4 was independently examined by ChIP, an apparently augmented level was
similarly detected in the presence of α-amanitin (Figure 4), confirming the previous
observation. We therefore speculate that the suspended RNAPII machine (distant to
ori62 and unable to recruit MCM2-7) by α-amanitin, may also suspend histone
modification enzymes, leaving a previously established environment suitable for
transcription but repressive for replication. It is equally possible that another histone
modification (or a specification factor) is required to uniquely regulate this late round of
amplification of DAFC-62D. The two mechanisms do not have to be mutually exclusive.
Again in Figure 5 we show localization of ORC2, MCM2-7 and RNAPII in
different developmental stages in DAFC-62D. The association patterns of these proteins
as well as that of AcK8H4 with ori62 are depicted in Figure 6B, and Figure 6C shows a
schematic of the effect of α-amanitin. The unknown histone modification or
specification factor is labeled X in Figures 6B and 6C. Given the fact that there is no
detectable AcK8H4 in transposons (Appendix One, Figure 5) that displays regulated
amplification (Appendix One, Figure 1A), such an X marker different from AcK8H4 is
likely to exist.

150
Figure 5. Association pattern of ORC2 (top panel), MCM2-7 (middle panel) and
RNAPII (bottom panel) with DAFC-62D were shown for comparison against H4
acetylation.
All ChIP experiments were independently performed, and some sampling methods may
slightly differ. For example, stage 12 egg chambers were used in both ORC2 and MCM2-
7 ChIP, whereas for RNAPII it was stages 11 and 12 combined.
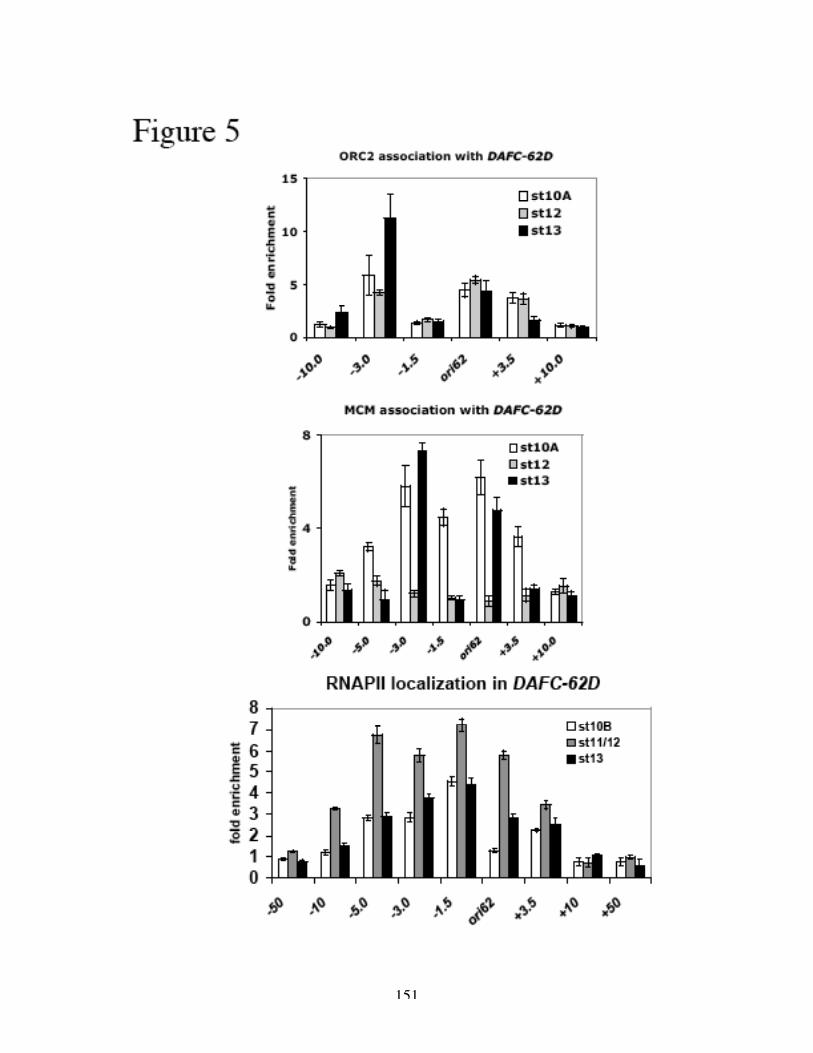
151

152
Figure 6. Changes of protein association with origins with regard to development
time.
(A) The pre-RC disassembles from oriβ of DAFC-66D during transcription (Txn) stages,
after initial amplification (Amp). (B) At ori62, Txn coincides with high AcK8H4 and loss
of MCM2-7. In the following round of Amp, Txn is probably inhibited by a drop in
AcK8H4. Facilitated by specification factor or histone modification X, RNAPII helps to
recruit MCM to ori62. (C) In the presence of α-amanitin, both Txn and the second round
of Amp are inhibited. RNAPII and MCM2-7 are no longer bound at ori62, which is
marked by high AcK8H4 and low X.

153

154
Materials and Methods
Anti-pan-acetyl-Histone H4, anti-acetyl-Histone H4 (Lys5) and anti-acetyl-
Histone H4 (Lys8) rabbit antisera (ChIP grade) were purchased from Upstate and used at
1:250 dilution for ChIP experiements.
REFERENCES
Aggarwal, B. D., and Calvi, B. R. (2004). Chromatin regulates origin activity inDrosophila follicle cells. Nature 430, 372-376.Altaf, M., Saksouk, N., and Cote, J. (2007). Histone modifications in response to DNAdamage. Mutat Res 618, 81-90.Aparicio, J. G., Viggiani, C. J., Gibson, D. G., and Aparicio, O. M. (2004). TheRpd3-Sin3 histone deacetylase regulates replication timing and enables intra-S origincontrol in Saccharomyces cerevisiae. Mol Cell Biol 24, 4769-4780.Brown, J. A., Holmes, S. G., and Smith, M. M. (1991). The chromatin structure ofSaccharomyces cerevisiae autonomously replicating sequences changes during the celldivision cycle. Mol Cell Biol 11, 5301-5311.Burke, T. W., Cook, J. G., Asano, M., and Nevins, J. R. (2001). Replication factorsMCM2 and ORC1 interact with the histone acetyltransferase HBO1. J Biol Chem 276,15397-15408.Doyon, Y., Cayrou, C., Ullah, M., Landry, A. J., Cote, V., Selleck, W., Lane, W. S.,Tan, S., Yang, X. J., and Cote, J. (2006). ING tumor suppressor proteins are criticalregulators of chromatin acetylation required for genome expression and perpetuation.Mol Cell 21, 51-64.Fuchs, J., Demidov, D., Houben, A., and Schubert, I. (2006). Chromosomal histonemodification patterns--from conservation to diversity. Trends Plant Sci 11, 199-208.Fukuda, H., Sano, N., Muto, S., and Horikoshi, M. (2006). Simple histone acetylationplays a complex role in the regulation of gene expression. Brief Funct GenomicProteomic 5, 190-208.Groth, A., Rocha, W., Verreault, A., and Almouzni, G. (2007). Chromatin challengesduring DNA replication and repair. Cell 128, 721-733.Hartl, T., Boswell, C., Orr-Weaver, T. L., and Bosco, G. (2007). Developmentallyregulated histone modifications in Drosophila follicle cells: initiation of geneamplification is associated with histone H3 and H4 hyperacetylation and H1phosphorylation. Chromosoma.Iizuka, M., Matsui, T., Takisawa, H., and Smith, M. M. (2006). Regulation ofreplication licensing by acetyltransferase Hbo1. Mol Cell Biol 26, 1098-1108.Iizuka, M., and Stillman, B. (1999). Histone acetyltransferase HBO1 interacts with theORC1 subunit of the human initiator protein. J Biol Chem 274, 23027-23034.Kouzarides, T. (2007). Chromatin modifications and their function. Cell 128, 693-705.Lipford, J. R., and Bell, S. P. (2001). Nucleosomes positioned by ORC facilitate theinitiation of DNA replication. Mol Cell 7, 21-30.

155
Simpson, R. T. (1990). Nucleosome positioning can affect the function of a cis-actingDNA element in vivo. Nature 343, 387-389.Stedman, W., Deng, Z., Lu, F., and Lieberman, P. M. (2004). ORC, MCM, and histonehyperacetylation at the Kaposi's sarcoma-associated herpesvirus latent replication origin.J Virol 78, 12566-12575.Vogelauer, M., Rubbi, L., Lucas, I., Brewer, B. J., and Grunstein, M. (2002). Histoneacetylation regulates the time of replication origin firing. Mol Cell 10, 1223-1233.Zhou, J., Chau, C. M., Deng, Z., Shiekhattar, R., Spindler, M. P., Schepers, A., andLieberman, P. M. (2005). Cell cycle regulation of chromatin at an origin of DNAreplication. Embo J 24, 1406-1417.

156
Appendix Three
Table of Acronyms
ACE3: amplification control element on the 3rd chromosomeAcH4: acetylated histone H4AcK5H4: acetylated histone H4 on Lysine 5AcK8H4: acetylated histone H4 on Lysine 8ACS: ARS consensus sequenceARS: autonomously replicating sequenceBrdU: 5’-bromo-2’-deoxyuridineChIP: chromatin immunoprecipitationCHO: Chinese hamster ovaryDAFC: Drosophila amplicon in follicle cellsDHFR: dihydrofolate reductasedREAM: Drosophila multisubunit complexes containing Rb, E2F2, Myb and MipsEcR: ecdysone receptorEcRE: ecdysone response elementFISH: fluorescent in situ hybridizationHAT: histone acetyltransferaseHDAC: histone deacetylaseMCM2-7: minichromosome maintenance proteins 2-7Mip: Myb-interecting proteinMyb: myeloblastosis oncoproteinORC: origin recognition complexori62: origin of DAFC-62Doriβ: origin of DAFC-66DPre-RC: pre-replication complexRb: retinoblastoma proteinRNAPII: RNA polymerase IIUSP: Ultraspiracleyg2: yellow-g2
~The End~










![DNA Replication Initiation Is Blocked by a Distant ... · replication and segregation of key chromosomal loci including the origin and terminus [4]. These steps also define two periods](https://static.fdocuments.net/doc/165x107/60020c885f9a0a1b3c145fd4/dna-replication-initiation-is-blocked-by-a-distant-replication-and-segregation.jpg)







