Development of the microcirculation of the secondary ossification center in rat humeral head
9
Development of the Microcirculation of the Secondary Ossification Center in Rat Humeral Head SERGIO MORINI, 1 MARIA A. CONTINENZA, 2 GIUSEPPE RICCIARDI, 2 EUGENIO GAUDIO, 1,3 * AND LUIGI PANNARALE 3 1 Department of Biomedical Sciences, University Campus Bio-Medico of Rome, Rome, Italy 2 Department of Surgical Sciences, State University, L’Aquila, Italy 3 Department of Human Anatomy, State University “La Sapienza,” Rome, Italy ABSTRACT This work investigated the origin and development of microcirculation in the rat humeral head and the expression of vascular endothelial growth factor (VEGF) as a factor supporting the vascular growth and the development of the secondary ossification centers. Sixty rats aging 1, 3– 4, 6 – 8, 11, and 21 days, 5 weeks, and 4 and 8 months were used. Samples of humeral head were collected for histolgy and immunohistochemistry for VEGF. Some animals were perfused with Mercox resin in order to obtain vascular corrosion casts (vcc) observed by scanning electron microscopy (SEM). No cartilage canals were present at birth. At 6 days postnatal, blood vessels coming from the perichondrium and the region near the capsule attachment invaded the carti- lage; at 11 days postnatal, signs of calcification were present and within the third week some bone trabeculae were formed. Just before the vascular invasion of the epiphysis, a positive reaction for VEGF was localized in chondrocytes of the epiphyseal cartilage close to the capsule insertion. During the development and expansion of the secondary ossification center, VEGF expression was higher in chondrocytes but decreased when epiphysis was diffusely ossified. VEGF was expressed also by mesenchymal cells present in and around the fibrous tissue where the second- ary ossification center will develop. SEM vcc confirmed that vessels penetrating into the epiph- ysis arose merely from the periosteal and the capsular networks, and vascular connections with the diaphyseal circulation were not evident. These observations demonstrated that VEGF pro- duction by chondrocytes begun some days after birth, supported the rapid vascular growth from the surrounding soft tissues, and was chronologically related to the development of the secondary ossification center in rat proximal humerus. Finally, the possible role of VEGF as mediator of angiogenesis and, at least indirectly, as a trigger factor also in the ossification and the bone remodeling of the secondary ossification centers has been discussed. Anat Rec Part A 278A: 419 – 427, 2004. © 2004 Wiley-Liss, Inc. Key words: bone; cartilage; ephiphysis; vasculature; vascular endothelial growth factor The vascularization of the epiphysis of long bones pre- sents particular interest because of its interactions with the ossificating tissue and with the development of the secondary ossification centers. In most of the cases, ossification has been considered to take place immediately adjacent to preexisting cartilage canals inside hyaline cartilage (Wilsman and Van Sickle, 1970; Kugler et al., 1979). These were described as thin loose connective spaces that extend into the cartilage and contain a small muscular arteriole, a venule, and a termi- nal capillary glomerulus (Stockwell, 1971; Wilsman and Van Sickle, 1972) not directly connected with the diaphy- seal circulation (Levene, 1964; Skawina et al., 1994). Car- Grant sponsor: Italian Ministero dell’Istruzione, Universita `e Ricerca (MIUR). *Correspondence to: Dr. Eugenio Gaudio, Department of Hu- man Anatomy, University of Rome “La Sapienza,” Via Alfonso Borelli, 50, 00161 Rome, Italy. Fax: 39-06-49-91-80-62. E-mail: [email protected] Received 11 August 2003; Accepted 8 September 2003 DOI 10.1002/ar.a.20016 THE ANATOMICAL RECORD PART A 278A:419 – 427 (2004) © 2004 WILEY-LISS, INC.
-
Upload
sergio-morini -
Category
Documents
-
view
212 -
download
0
Transcript of Development of the microcirculation of the secondary ossification center in rat humeral head

Development of the Microcirculationof the Secondary Ossification Center
in Rat Humeral HeadSERGIO MORINI,1 MARIA A. CONTINENZA,2 GIUSEPPE RICCIARDI,2
EUGENIO GAUDIO,1,3* AND LUIGI PANNARALE3
1Department of Biomedical Sciences, University Campus Bio-Medico of Rome,Rome, Italy
2Department of Surgical Sciences, State University, L’Aquila, Italy3Department of Human Anatomy, State University “La Sapienza,” Rome, Italy
ABSTRACTThis work investigated the origin and development of microcirculation in the rat humeral
head and the expression of vascular endothelial growth factor (VEGF) as a factor supporting thevascular growth and the development of the secondary ossification centers. Sixty rats aging 1,3–4, 6–8, 11, and 21 days, 5 weeks, and 4 and 8 months were used. Samples of humeral headwere collected for histolgy and immunohistochemistry for VEGF. Some animals were perfusedwith Mercox resin in order to obtain vascular corrosion casts (vcc) observed by scanning electronmicroscopy (SEM). No cartilage canals were present at birth. At 6 days postnatal, blood vesselscoming from the perichondrium and the region near the capsule attachment invaded the carti-lage; at 11 days postnatal, signs of calcification were present and within the third week some bonetrabeculae were formed. Just before the vascular invasion of the epiphysis, a positive reaction forVEGF was localized in chondrocytes of the epiphyseal cartilage close to the capsule insertion.During the development and expansion of the secondary ossification center, VEGF expressionwas higher in chondrocytes but decreased when epiphysis was diffusely ossified. VEGF wasexpressed also by mesenchymal cells present in and around the fibrous tissue where the second-ary ossification center will develop. SEM vcc confirmed that vessels penetrating into the epiph-ysis arose merely from the periosteal and the capsular networks, and vascular connections withthe diaphyseal circulation were not evident. These observations demonstrated that VEGF pro-duction by chondrocytes begun some days after birth, supported the rapid vascular growth fromthe surrounding soft tissues, and was chronologically related to the development of the secondaryossification center in rat proximal humerus. Finally, the possible role of VEGF as mediator ofangiogenesis and, at least indirectly, as a trigger factor also in the ossification and the boneremodeling of the secondary ossification centers has been discussed. Anat Rec Part A 278A:419–427, 2004. © 2004 Wiley-Liss, Inc.
Key words: bone; cartilage; ephiphysis; vasculature; vascular endothelialgrowth factor
The vascularization of the epiphysis of long bones pre-sents particular interest because of its interactions withthe ossificating tissue and with the development of thesecondary ossification centers.
In most of the cases, ossification has been considered totake place immediately adjacent to preexisting cartilagecanals inside hyaline cartilage (Wilsman and Van Sickle,1970; Kugler et al., 1979). These were described as thinloose connective spaces that extend into the cartilage andcontain a small muscular arteriole, a venule, and a termi-nal capillary glomerulus (Stockwell, 1971; Wilsman andVan Sickle, 1972) not directly connected with the diaphy-seal circulation (Levene, 1964; Skawina et al., 1994). Car-
Grant sponsor: Italian Ministero dell’Istruzione, Universita eRicerca (MIUR).
*Correspondence to: Dr. Eugenio Gaudio, Department of Hu-man Anatomy, University of Rome “La Sapienza,” Via AlfonsoBorelli, 50, 00161 Rome, Italy. Fax: 39-06-49-91-80-62. E-mail:[email protected]
Received 11 August 2003; Accepted 8 September 2003DOI 10.1002/ar.a.20016
THE ANATOMICAL RECORD PART A 278A:419–427 (2004)
© 2004 WILEY-LISS, INC.

tilage canals have been commonly described in larger an-imals (Haines, 1933; Hurrel, 1934; Levene, 1964; Wilsmanand Van Sikle, 1972; Visco et al., 1990; Ganey et al., 1992)and only exceptionally in smaller animals (Kugler et al.,1979).
In small animals, an epiphyseal ossification that takesorigin from a peripheral vascular proliferation derived bythe periosteal and perichondrial vascular network in theabsence of cartilage canals was described in murine distalfemur (Floyd et al., 1987) and in the rat proximal tibia(Lee et al., 2001). This finding agreed with our previousobservation on vascular corrosion cast of adult rat wherethe microvasculature of the humeral head appeared notdirectly connected to the shaft microcirculation (data notshown). All these data supported the hypothesis of a vas-cular proliferation starting from the surrounding soft tis-sues in rat humerus. Thus, the key point in mice and ratswas the secondary center formation immediately followingthe emergence of the cartilage canals.
The development of the vasculature inside the tissuesdepends from the production of different growth factors(Beck and D’Amore, 1997), among which the most effec-tive is the vascular endothelial growth factor (VEGF) thatpromotes angiogenesis (Ferrara and Henzel, 1989; Leunget al., 1989). Recently, the immunolocalization of VEGFwas demonstrated in proliferative and hypertrophic chon-drocytes of the growth plates (Horner et al., 1999; Ichiga-tani et al., 2001). For this reason, VEGF was supposed tosupport vascular growth in the metaphysis during boneformation. However, the presence of VEGF inside theepiphysis and the correlations with the development of thevasculature that support the secondary ossification cen-ters were not clearly demonstrated.
Our aims were to investigate the development of thevasculature inside the growing rat humeral head cartilagein order to understand the origin of the microcirculationand the connections with the surrounding vascular net-works, and to recognize the possible presence of VEGF inthe areas of vascular proliferation correlated to the sec-ondary ossification center formation in order to study thepossible role of VEGF as a factor supporting the vasculardevelopment inside the rat humeral head.
MATERIALS AND METHODSSixty male Wistar rats aging postnatal 1, 3–4, 6–8, 11,
and 21 days, 5 weeks (100 g body weight), and 4 and 8months (350–650 g body weight) were used in this study,with the approval of the Ethical Committee of the Depart-ment of Human Anatomy of the University of Rome “LaSapienza.”
Light Microscopy (LM)Four to six rats of each group of age were sacrificed
using an intravenous overdose of sodium thiopental. Afterperfusion of the vascular tree by injection of 4% bufferedformaldehyde commercial solution (Bio-Optica, Milan, It-aly) through the left ventricle, the anterior limbs weredissected. Shoulder muscles were removed in order toexpose the articular capsule that was cut open at itsscapular insertion. Both humeri were then fixed by im-mersion in 4% buffered formaldehyde solution for 24 hr atroom temperature and subsequently decalcified throughthe use of Histodec-F commercial solution (Cytochemica,Avellino, Italy). The proximal epiphysis was then sepa-
rated from the shaft and cut into two symmetrical piecesalong a longitudinal section plane including the axis of theanatomical neck of the humerus. Specimens were embed-ded in paraffin and 5 �m sections were cut following theprevious section plane. Finally, specimens were stainedwith hematoxylin and eosin and Masson’s trichromic.
ImmunohistochemistryFor the immunohistochemical study, sections of 3–5 �m
were immunoperoxidase stained for VEGF. The indirectimmunohistochemical technique was used. In order toquench the endogenous peroxidase activity, often high inchondrocytes, samples were incubated with 3% H2O2 for10 min; CAS Block (Zymed Laboratories, San Francisco,CA) serum was applied in order to reduce nonspecificbackground staining. Sections were overnight incubatedwith primary mouse monoclonal anti-VEGF sc-7269 anti-body (Santa Cruz Biothecnologies, Santa Cruz, CA) di-luted 1:200 at 4°C with Antibody Diluent (Dako, Carpin-teria, CA). This antibody corresponds to amino acids1–140 of VEGF and does not react with VEGF-B andVEGF-C. As secondary antibody, we used biotinylatedswine antibody labeled with the streptavidin-peroxidasecomplex LSAB� (Dako) at room temperature. The subse-quent development was performed with diaminobenzidineto yield a brown-color reaction. Sections were counter-stained with Mayer’s hematoxylin. Controls were includedin each batch of slides. Negative controls were performedon bone samples applying only the diluent without theprimary antibody. Positive controls were performed oncolonic carcinoma that is known to produce positive VEGFreaction (Takahashi et al., 1998).
Immunohistochemistry for the detection of endothelialcells was performed following the same protocol using asprimary antibody polyclonal anti-von Willebrand factorsc-8068 antibody (Santa Cruz Biothecnologies).
Scanning Electron Microscopy VascularCorrosion Casts (SEM vcc)
In order to study the connections of the epiphyseal vas-cular network, rats aged 5 weeks as well as 4 and 8months were employed for obtaining vascular casts. Sixanimals from each group of age were infused with resinaccording with our previous studies (Pannarale et al.,1997; Morini et al., 1999). The animals were preanesthe-tized with diethyl ether and immediately after with anintraperitoneal injection of 15–20 I.U. sodium thiopental.In preparation for introduction of the resin material, ad-equate blood drainage was facilitated by i.v. injection ofheparin and acetylcholine.
A median thoracic and abdominal incision was per-formed. The abdominal aorta was retrogradely cannulated(Inpharven-Int cannula, Ø � 1.7 mm) and the ascendingaorta was closed by tying it. The right atrium was cutopen. The vascular bed was initially cleaned out by pre-warmed 0.9% normal saline and then injected with 50 mldiluted Mercox CL-2R resin (4:1) for about 7 min. Perfu-sion was monitored by means of an electronic manometerin order to maintain a pressure of 200–250 mm Hg in thecannula. Pressure value was obtained by trying increasinginjection pressures till consistent and satisfactory filling ofthe vessels was achieved according to our previous expe-rience (Pannarale et al., 1997; Morini et al., 1999).
After perfusion, the animals remained for 24 hr at roomtemperature for a better polymerization of the resin, then
420 MORINI ET AL.

the anterior limbs were dissected and the humeri wereisolated. Remnants of muscle and connective tissue wereremoved by leaving bones in 20% NaOH solution for 24 hr;decalcification was obtained by processing the bones with5% HCl solution. In order to obtain a better cleaning of thecast surface from precipitates, the samples were alterna-tively exposed to repeated baths in NaOH and HCl solu-tions. At the end, the samples were placed in 5% trichlo-roacetic acid for complete removal of tissue remnants. Theobtained casts were frozen in distilled water and freeze-dried. Finally, specimens were glued onto stubs, coveredwith platinum, and examined by a Hitachi S4000 fieldemission SEM operating at an accelerating voltage of 7kV.
RESULTSLM Observations
The stages of the developing humeral head are chrono-logically drawn in Figure 1.
One day.The epiphysis is made up mostly of restingcartilage (Fig. 2). Isogenous groups and multiple-cell
groups are present inside the cartilage. In the most super-ficial layer, chondrocytes flatten and dispose themselvesparallel to the articular surface. The physeal plate is notyet developed but chondrocytes tend to constitute initialcell columns. Elements of proliferation and hypertrophyzone chondrocytes are present. Neither cartilage canalsnor initial secondary ossification centers are present.VEGF reaction fails to stain both the cartilage and theperichondrium in the epiphysis proper.
Three to four days.The epiphysis appears structur-ally similar to the previous time point and vascular pres-ence inside the cartilage is not evident (stage 0, Fig. 1a).The physeal plate is formed and a light positive reactionfor VEGF stains the growing chondrocytes of the physealplate (Fig. 3). Chondrocytes in the region corresponding tothe capsule attachment show the first sign of immunore-action to VEGF stain (Fig. 4).
Six to eight days.A fibrovascular proliferation startsfrom the region corresponding to the capsule attachmentand the perichondrium very close to the capsule (Fig. 5),
Fig. 1. Schematic drawing showing the architecture of the rat hu-meral head from 4 days to 5 weeks of age as obtained by tissuesections. The small white ellipses correspond to groups of hypertrophicchondrocytes indicative of endochondral ossification in the epiphysis.The direction of marrow space (light gray) enlargement and cartilage(white) resorption are indicated by arrows. The dark sprinkle-spottedzones represent areas where chondrocytes show VEGF-positive stain.
a: Intact humeral head, stage 0; 4 days. b: Invasion of the cartilage byexcavations, stage 1; 6 days. c: Fusion of canals and formation of themarrow space, stage 2; 7–8 days. d: Expansion of the marrow spaceand appearance of the secondary ossification center, stage 3; 11 days.e: Expansion of the ossification center and complete appearance of thesecondary growth plate, stage 4; 21 days. f: Adult aspect of the humeralhead; 5 weeks.
421VASCULATURE IN RAT HUMERAL HEAD

Figures 2–9.
422 MORINI ET AL.

forming small canals that grow in a radial direction (stage1, Fig. 1b). These canals are lined by a continuous endo-thelial cell lining that can be labeled with endothelial cellmarker (von Willebrand factor). In short time the deepends of the canals tend to joint. The vascular proliferationis accompanied by resorption of the cartilage matrix; aprogressive invasion of mesenchymal cells showingVEGF-positive reaction replace the chondrocytes (Fig. 6),forming the marrow space (stage 2, Fig 1c). VEGF reactionfurther stains chondrocytes around the marrow spaces(Fig. 7). The immunostaining appears highly specific asdemonstrated by negative controls where neither chondro-cytes nor mesenchymal cells in the marrow spaces areshowing positive reaction (Fig. 8). Inside the marrowspace, some structures of variable size appear as emptyspaces lined by endothelial cells (Fig. 9). These structurescorrespond to blood vessels with very thin and not com-pletely structured walls, which are proliferating and or-ganizing to form the sinusoidal rich marrow vascular net-work.
Eleven days.The marrow space enlarges (stage 3, Fig.1d) with progressive resorption of the cartilage and ap-pearance of groups of hypertrophic chondrocytes (Fig. 10).VEGF reaction stains most of the chondrocytes that sur-round the marrow space, including hypertrophic chondro-cytes, and many mesenchymal cells inside the marrowspace (Fig. 11). The reaction appears highly specific asdemonstrated by controls (Fig. 12).
Twenty-one days.The ossification center extends fromone-third to half of the epiphysis and presents areas ofcalcification and thin trabeculae of bone tissue (stage 4,Fig. 1e). In the side facing the metaphyseal plate, theresorption proceeds, encircling portions of cartilage thatare replaced by calcified tissue (Fig. 13). Along the marginfacing the articular cartilage, hypertrophic chondrocytesassume a continuous growth plate-like aspect, forming theso-called secondary growth plate (Fig. 14). However, col-umns of proliferating chondrocytes typical of the physealplate are never forming in this area. Besides most bonemarrow cells, and chondrocytes around the secondary os-sification center, osteocytes also appear positive for VEGFreaction (Fig. 15).
Five weeks.The epiphysis is diffusely ossified exceptfor a very superficial layer below the articular cartilageand the capsular border (Fig. 1f). The bone of the epiphy-sis is clearly distinguished from the diaphysis by the pres-ence of a continuous layer of physeal cartilage. No fibro-vascular or bone tissue extends through the physeal plate.Highly vascularized fibrous tissue clearly extends fromthe capsular circumference into the epiphysis (Fig. 16). Inthe remaining cartilage, chondrocytes located under thearticular surface does not immunostain with VEGF (Fig.17).
SEM vcc Observations
Larger afferent and efferent vessels are placed along theanatomical neck of the epiphysis, all around the humeralneck (Figs. 18 and 19). These vessels are arteries andveins that penetrate into the humeral head. In theyounger animals (5 weeks), the diameter of the arteries isabout 40–60 �m, while at 4 months and later it rangesfrom 50 to 100 �m. Superficially, periosteal and capsularnetworks take rise, both composed by a polygonal mesh ofprecapillary, mostly capillary and postcapillary vessels,lying on one plane.
Via side branching, the arterial tree gives rise tosmaller branches, which take rise after dichotomous divi-sion. From the larger vessels in the neck region, smallerarteries deepen into the humeral head. In younger ani-mals, most of the smaller arteries run on a more superfi-cial plane, crossing a space without capillaries, whichcorresponds to the cartilage, before giving rise to the cap-illaries of the marrow space (Figs. 20 and 21). The capil-lary network of the epiphysis is constituted by a continu-ous three-dimensional net of sinusoids that grows denserwith age.
Postcapillary venules take rise from a bunch of sinu-soids joining together. They generally run at a shortdistance into a larger venule that ends into a moresuperficial capsular or periosteal circle, where they runparallel to the arteries. No connection with the diaphy-seal circulation are seen at any age, except for tinyperiosteal vessels.
Fig. 2. a: Proximal humeral epiphysis of rat at 1 day of age; longitu-dinal section through a frontal plane. The epiphysis is made up ofcartilage. Neither cartilage canals nor secondary ossification centers arepresent. A real front of ossification is not yet organized but chondrocytestend to constitute initial cell columns (arrowheads). H, humeral head; T,greater tuberosity; C, capsule. LM, original magnification 25�. b: Highermagnification of the area framed in a. Positive VEGF reaction is presentin the bone tissue and calcified cartilage (below) and in chondrocytes(arrowheads) close to the zone of endochondral ossification.
Fig. 3. Marginal physeal plate at 4 days of age. VEGF-positive reac-tion stains hypertrophic and proliferating chondrocytes of the growingcartilage. p, perichondrium. LM. Original magnification 125�.
Fig. 4. Capsule insertion at 4 days of age. VEGF-positive reaction isevident in chondrocytes (arrowheads) of the region corresponding to thecapsule attachment. C, capsule. LM. Original magnification 120�.
Fig. 5. Marginal cartilage close to the capsule insertion at 6 dayspostnatal. Some small vessels lined by endothelial cells (arrowheads)coming from the perichondrium (p) proliferate and grow into the carti-lage. VEGF reaction stains chondrocytes surrounding the vascular pro-liferation. LM. Original magnification 160�.
Fig. 6. A zone of cartilage resorption in the rat humeral head at 7days of age. Some areas of variable size, lined by flattened endothelium(arrowheads), can be interpreted as proliferating vessels into the carti-lage. They are surrounded by mesenchymal cells showing positive VEGFreaction. LM. Original magnification 100�.
Fig. 7. Zones of cartilage resorption in the rat humeral head at 7 daysof age. VEGF reaction stains groups of chondrocytes (arrows) aroundand cells inside the area of fibrovascular proliferation. The arrowheadsindicate vessels of variable size lined by endothelial cells. LM. Originalmagnification 80�.
Fig. 8. Zones of cartilage resorption inside the rat humeral head at 7days of age. Neither chondrocytes nor mesenchymal cells immunostainin this negative control for VEGF reaction. Note the presence of a smallvessel lined by endothelium (arrowheads). LM. Original magnification80�.
Fig. 9. Zones of cartilage resorption at 7 days of age containingsome empty spaces of variable size very similar to that of Figures 6–8.These spaces are clearly lined by endothelial cells (arrowheads) indicat-ing that vessels are growing inside the cartilage. LM, hematoxylin andeosin. Original magnification 100�.
423VASCULATURE IN RAT HUMERAL HEAD
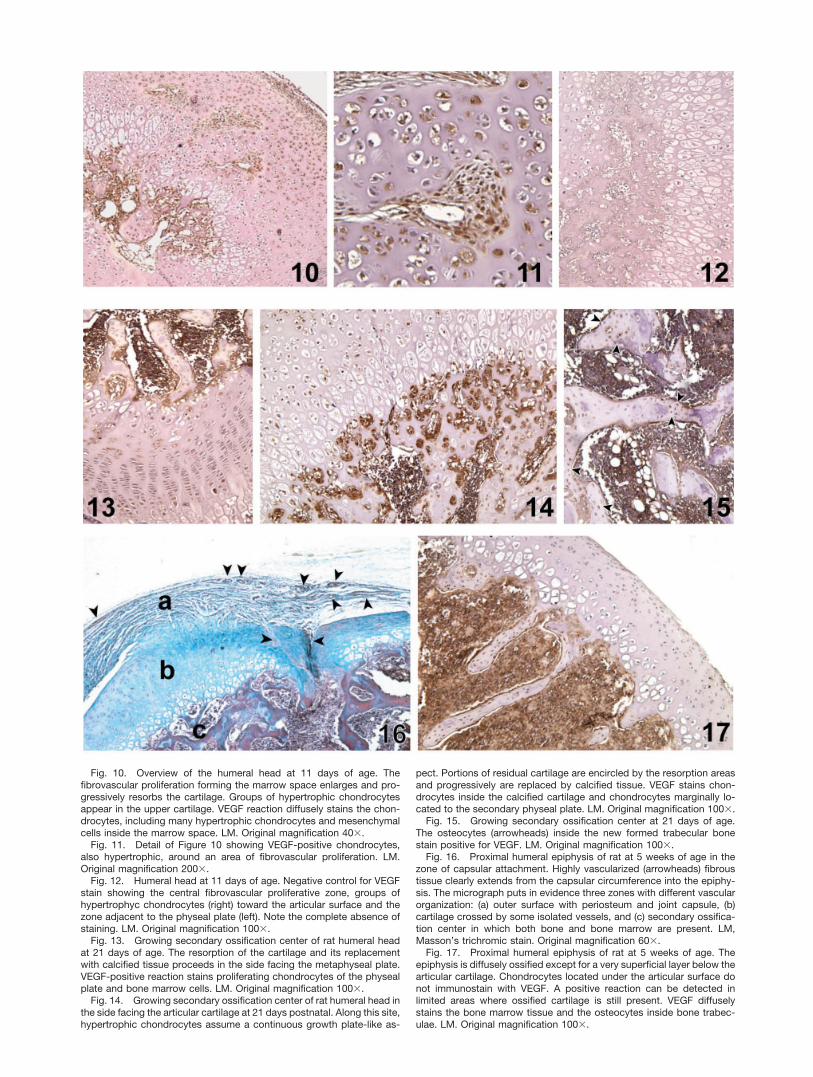
Fig. 10. Overview of the humeral head at 11 days of age. Thefibrovascular proliferation forming the marrow space enlarges and pro-gressively resorbs the cartilage. Groups of hypertrophic chondrocytesappear in the upper cartilage. VEGF reaction diffusely stains the chon-drocytes, including many hypertrophic chondrocytes and mesenchymalcells inside the marrow space. LM. Original magnification 40�.
Fig. 11. Detail of Figure 10 showing VEGF-positive chondrocytes,also hypertrophic, around an area of fibrovascular proliferation. LM.Original magnification 200�.
Fig. 12. Humeral head at 11 days of age. Negative control for VEGFstain showing the central fibrovascular proliferative zone, groups ofhypertrophyc chondrocytes (right) toward the articular surface and thezone adjacent to the physeal plate (left). Note the complete absence ofstaining. LM. Original magnification 100�.
Fig. 13. Growing secondary ossification center of rat humeral headat 21 days of age. The resorption of the cartilage and its replacementwith calcified tissue proceeds in the side facing the metaphyseal plate.VEGF-positive reaction stains proliferating chondrocytes of the physealplate and bone marrow cells. LM. Original magnification 100�.
Fig. 14. Growing secondary ossification center of rat humeral head inthe side facing the articular cartilage at 21 days postnatal. Along this site,hypertrophic chondrocytes assume a continuous growth plate-like as-
pect. Portions of residual cartilage are encircled by the resorption areasand progressively are replaced by calcified tissue. VEGF stains chon-drocytes inside the calcified cartilage and chondrocytes marginally lo-cated to the secondary physeal plate. LM. Original magnification 100�.
Fig. 15. Growing secondary ossification center at 21 days of age.The osteocytes (arrowheads) inside the new formed trabecular bonestain positive for VEGF. LM. Original magnification 100�.
Fig. 16. Proximal humeral epiphysis of rat at 5 weeks of age in thezone of capsular attachment. Highly vascularized (arrowheads) fibroustissue clearly extends from the capsular circumference into the epiphy-sis. The micrograph puts in evidence three zones with different vascularorganization: (a) outer surface with periosteum and joint capsule, (b)cartilage crossed by some isolated vessels, and (c) secondary ossifica-tion center in which both bone and bone marrow are present. LM,Masson’s trichromic stain. Original magnification 60�.
Fig. 17. Proximal humeral epiphysis of rat at 5 weeks of age. Theepiphysis is diffusely ossified except for a very superficial layer below thearticular cartilage. Chondrocytes located under the articular surface donot immunostain with VEGF. A positive reaction can be detected inlimited areas where ossified cartilage is still present. VEGF diffuselystains the bone marrow tissue and the osteocytes inside bone trabec-ulae. LM. Original magnification 100�.

DISCUSSIONA first aim of this work was to investigate the develop-
ment of the vasculature of the secondary ossification cen-ter in rat humeral head. In order to demonstrate thesource of vessels that supplied the epiphysis, we observedvascular casts by scanning electron microscopy. This tech-nique provided a useful three-dimensional visualization ofthe fine microvascular arrangements and connections.Furthermore, a number of different criteria, such as thedimension, the pattern of vessel division, and the partic-ular imprints left by endothelial cell nuclei on the castsurface, allowed one to distinguish arteries from veins andcapillaries (Lametschwandtner et al., 1991; Pannarale etal., 1997). However, considering the difficulty of visualiz-
ing the vascular network of the humeral head during itsdevelopment, because the injection of resin into not well-structured vessels easily led to the formation of artifacts,we have used young and adult animals in which the epiph-yseal vasculature was supposed to be more complete.
In spite of the different ages of the rats for vcc withrespect to those for the histological observations, thesource of vessels and their connections could be demon-strated. In fact, our observations have clearly shown thatthe vasculature of the humeral head arose exclusivelyfrom vessels connected with the periosteal and the capsu-lar microvascular networks, without any connections withthe diaphyseal circulation. In young and adult rats, therewas only one vascular peduncle made up of arteries and
Fig. 18. Larger vessels of the humeral neck (corresponding to zone ain Fig. 16) at 5 weeks as viewed from the outside surface of the cast afterthe removal of the capsular network. Small capsular vessels are stillpresent (c). The origin of the arterioles (arrowheads) looping into thehead region (H) is demonstrated. SEM vcc. Scale bar � 200 �m.
Fig. 19. Humeral neck at 4 months as viewed from the outsidesurface of the cast (zone a in Fig. 16). Coming from the periostealnetwork, an artery (A) accompanied by a vein (V) is directed toward thehead. A smaller artery (a) that supplies some capillaries (c) of the cap-sular network and a small venule (v) are also visible. SEM vcc. Scalebar � 100 �m.
Fig. 20. Afferent vessels as viewed from the joint surface at 4months. Some small arteries (arrowheads) arise from the circumferentialvessels (zone a in Fig. 16), run through a superficial space (asterisk) onceconstituted by cartilage (zone b in Fig. 16), and give rise to the sinusoidalnetwork (s) of the bone marrow tissue inside the epiphysis (zone c in Fig.16). SEM vcc. Scale bar � 200 �m.
Fig. 21. Detail of some small vessels (arrowheads) located in zone bof Figure 16 that directly connect the outer circulation (asterisk) with thesinusoids (s) of the humeral head (zone c in Fig. 16) at 5 weeks. SEM vcc.Scale bar � 100 �m.
425VASCULATURE IN RAT HUMERAL HEAD

veins that run in the inner part of the epiphysis and aftersubsequent divisions supplied the sinusoids of the medul-lary spaces. This finding seemed to confirm observationsby Floyd et al. (1987) of a vascular proliferation comingfrom the perichondrium without preexisting cartilage ca-nals.
The source of vessels that we found in the rat humeralhead was different with respect to the rat femoral head(Morini et al., 1999) in which the afferent and efferentvessels were connected with the diaphyseal vasculature.Therefore, by comparing the origin of the developing mi-crovasculature and its outside connections, two differentways existed at the same time in the rat proximal humeraland femoral epiphysis. This difference has made the hu-meral head a more viable general model for studying thedevelopment of the secondary ossification centers.
Our major aim was to relate cellular VEGF distributionto the excavations of the cartilaginous epiphysis followingchronological events. These events involved a series ofstages initially summarized by Lee et al. (2001) in the ratproximal tibia. We have observed similar stages in thestructural development of the proximal humeral epiphy-sis.
Vascular proliferation was associated with the in-creased mass of tissue during cartilage growth (Ganey etal., 1992). In the past, we have also underlined the influ-ence of bone thickness in the development of differentmicrovascular patterns in flat bones (Pannarale et al.,1997). These observations have suggested that vascularinvasion of the cartilage could proceed from very differentdistricts in response to the metabolic needs of the carti-lage, mediated by growth factors.
The direct correlation observed between increased massof cartilage during growth and its invasion by vessels hassuggested the production of growth factors by chondro-cytes. Vascular proliferation and growth could be sup-ported by a number of factors (Beck and D’Amore, 1997).Among these factors, VEGF played a unique role in theregulation of angiogenesis (Ferrara and Henzel, 1989;Leung et al., 1989) by means of a potent mitotic, chemio-tactic (Leung et al., 1989), and antiapoptotic effect onendothelial cells (Gerber et al., 1998; Nor et al., 1999).VEGF expression was increased by hypoxia (Shweiki etal., 1992), while cytokines and growth factors could mod-ulate angiogenesis by altering VEGF expression (Neufeldet al., 1999; Minet et al., 2000).
Recently, chondrocytes were shown to produce factorsthat can regulate angiogenesis (Descalzi Cancedda et al.,1995; Alini et al., 1996; Horner et al., 1996, 1998; Carle-varo et al., 1997). VEGF produced by hypertrophic chon-drocytes has been localized in neonatal human growthplates (Horner et al., 1999; Garcia-Ramirez et al., 2000).In the present work, a clear reaction for VEGF waspresent in the chondrocytes of the growth plate. Thisfinding can explain angiogenesis and vascular prolifera-tion observed under the physeal plate (Arsenault, 1987;Hunter and Arsenault, 1990; Aharinejad et al., 1995;Morini et al., 1999) where new bone was developing dur-ing growth.
However, as demonstrated in the present work bymeans of vascular corrosion casts, the development of thesecondary ossification centers was supplied by vesselsthat were not directly connected with the diaphyseal cir-culation. Hence, the presence of factors supporting thevascular development from the surrounding tissues
should be expected. However, the immunolocalization ofVEGF in zones of the epiphysis other than the physealplate was not clearly demonstrated until now.
In the present work, a marked positive reaction forVEGF within the cartilage of the humeral head was dem-onstrated in peripheral chondrocytes located very close tothe zones of capsule attachment to the epiphysis (stage 0,Fig. 1a). These zones corresponded to the areas of initialvascular proliferation. Moreover, our observations havedemonstrated that the immunolocalization of VEGF in-side chondrocytes of the epyphiseal cartilage became evi-dent just before the vascular invasion of the epiphysis(Fig. 1a and b). Later on, chondrocytes that surroundedthe marrow space were positive for VEGF during the rapidexpansion of the secondary ossification center (Fig. 1c–e),but they lost immunopositivity when expansion decreasedor stopped (Fig. 1f). However, the vascular proliferationand penetration processes were accompanied by positiveVEGF reaction in mesenchymal cells, bone and bone mar-row cells, and supported the rapid ossification of the hu-meral head.
The discovery of VEGF in hypertrophic chondrocytes ofthe growth plate (Horner et al., 1999) close to numerousblood vessels of the subchondral region has suggested arole in the regulation of vascular invasion by means of aparacrine action with invading endothelial cells as a tar-get (Carlevaro et al., 2000). Moreover, when VEGF wasdemonstrated inside the cartilage (Horner et al., 1999;Ichigatani et al., 2001), such as in the present study,vascular proliferation was constantly and rapidly followedby the ossification of the cartilage. This seems to confirmin vitro studies by Roach et al. (1998). These authorsobserved that, when at least two growing vessels are inapposition, chondrocytes become hypertrophic and a newossification center initiates. This has suggested a possibletrigger role of the cumulative release of diffusible factorsin the initiation of bone formation.
VEGF receptor and signal transduction have been local-ized in chondrocytes, supporting the hypothesis of a VEGFautocrine activity also in morphogenesis and differentia-tion of mesoderm-derived cells (Carlevaro et al., 2000).Our observations have shown an immunolocalization ofVEGF in the mesenchymal cells inside the areas of carti-lage absorption and vascular proliferation, besides itspresence into the surrounding chondrocytes. These obser-vations strengthened a possible role of VEGF in osteogen-esis, as was recently hypothesized in the literature. Infact, VEGF was recognized to take part in bone marrow(Hattori et al., 2001; Tordjman et al., 2001) and bone celldifferentiation (Yeh and Lee, 1999) and to support angio-genesis at sites of endochondral and intramembranousossification (Gerber et al., 1999; Horner et al., 2001). Fi-nally, VEGF was supposed to play a role not only as acoordinator of chondrocytes vital cycle, chondroclast func-tion, and extracellular matrix remodeling, but also as oneof the trigger factors for osteogenesis and bone formation(Gerber et al., 1999; Horner et al., 2001).
In conclusion, this study has clearly demonstrated thatthe cartilage of the growing rat’s humeral head was avas-cular at birth and did not need vessels for tissue nutrition.When the humeral head grew, VEGF was expressed bothby actively growing chondrocytes and chondrocytes in se-lected region of the epiphysis. Subsequently, vessels com-ing from the circulation of the capsule and the surround-ing soft tissues proliferated and invaded the epiphyseal
426 MORINI ET AL.

cartilage. Chondrocyte expression of VEGF was chrono-logically related to the development and expansion of thesecondary ossification center and decreased when epiphy-sis was diffusely ossified. VEGF expression was recog-nized also in mesenchymal cells in and around the areas ofcartilage resorption, supporting a role in angiogenesisand, at least indirectly, in the ossification and the boneremodeling of the secondary ossification centers.
LITERATURE CITEDAharinejad S, Marks SC, Bock P Jr, Mackay CA, Larson EK, Taha-
mtani A, Mason-Savas A, Firbas W. 1995. Microvascular pattern inthe metaphysis during bone growth. Anat Rec 242:111–122.
Alini M, Marriott A, Chen T, Abe S, Poole AR. 1996. A novel angio-genic molecule produced at the time of chondrocytes hypertrophyduring endochondral bone-formation. Dev Biol 176:124–132.
Arsenault AL. 1987. Microvascular organization at the epiphysealmetaphyseal junction of growing rats. J Bone Miner Res 2:143–149.
Beck L, D’Amore PA. 1997. Vascular development: cellular and mo-lecular regulation. FASEB J 11:365–373.
Carlevaro MF, Albini A, Ribatti D, Gentili C, Benelli R, Cermelli S,Cancedda R, Cancedda FD. 1997. Transferrin promotes endothelialcell migration and invasion: implication in cartilage neovascular-ization. J Cell Biol 136:1375–1384.
Carlevaro MF, Cermelli S, Cancedda R, Descalzi Cancedda F. 2000.Vascular endothelial growth factor (VEGF) in cartilage neovascu-larization and chondrocytes differentiation; auto-paracrine roleduring endochondral bone formation. J Cell Sci 113:59–69.
Descalzi Cancedda F, Melchiori A, Benelli R, Gentili R, Masiello L,Campanile G, Cancedda R, Albini A. 1995. Production of angiogen-esis inhibitors and stimulators is modulated by cultured growth-plate chondrocytes during in-vitro differentiation: dependence onextracellular matrix assembly. Eur J Cell Biol 66:60–68.
Ferrara N, Henzel WJ. 1989. Pituitary follicular cells secrete a novelheparin-binding growth factor specific for vascular endothelial cells.Biochem Biophys Res Commun 161:851–858.
Floyd WE, Zaleske DJ, Schiller AL, Trahan C, Mankin HJ. 1987.Vascular events associated with the appearance of the secondarycenter of ossification in the murine distal femoral epiphysis. J BoneJoint Surg 69:185–190.
Ganey TM, Love SM, Ogden JA. 1992. Development of vascularizationin the chondroepiphysis of the rabbit. J Orthopaediatr Res 10:496–510.
Garcia-Ramirez M, Toran N, Andaluz P, Carrascosa A, Audi L. 2000.Vascular endothelial growth factor is expressed in human fetalgrowth cartilage. J Bone Mine Res 15:534–540.
Gerber HP, Dixit V, Ferrara N. 1998. Vascular endothelial growthfactor induces expression of the antiapoptotic proteins Bcl-2 and A1in vascular endothelial cells. J Biol Chem 273:13313–13316.
Gerber HP, Vu TH, Ryan AM, Kowalsky J, Werb Z, Ferrara N. 1999.VEGF couples hypertrophic cartilage remodeling, ossification andangiogenesis during endochondral bone formation. Nat Med 6:623–628.
Haines RW. 1933. Cartilage canals. J Anat 68:45–64.Hattori K, Dias S, Heissig B, Hackett NR, Lyden D, Tateno M, Hicklin
DJ, Zhu Z, Witte L, Crystal RG, Moore MA, Rafii S. 2001. Vascularendothelial growth factor and angiopoietin-1 stimulate postnatalhematopoiesis by recruitment of vasculogenic and hematopoieticstem cells. J Exp Med 193:1005–1014.
Horner A, Bord S, Kemp P, Grainger D, Compston JE. 1996. Distri-bution of platelet-derived growth factor (PDGF) A chain mRNA,protein, and PDGF-alpha receptor is rapidly forming human bone.Bone 19:353–362.
Horner A, Kemp P, Summers C, Bord S, Bishop NJ, Kelsall AW. 1998.The expression and distribution of transforming growth factor beta(TGF�) isoforms and their signaling receptors in growing humanbone. Bone 23:95–102.
Horner A, Bishop NJ, Bord S, Beeton A, Kelsall AW, Coleman N,Compston JE. 1999. Immunolocalization of vascular endothelialgrowth factor (VEGF) in human neonatal growth plate cartilage. JAnat 19:519–524.
Horner A, Bord S, Kelsall AW, Coleman N, Compston JE. 2001. Tie2ligands angiopoietin-1 and angiopoietin-2 are coexpressed withVascular Endothelial cell Growth Factor in growing human bone.Bone 28:65–71.
Hunter WL, Arsenault AL. 1990. Vascular invasion of the epiphysealgrowth plate: analysis of metaphyseal capillary ultrastructure andgrowth dynamics. Anat Rec 227:223–231.
Hurrel DJ. 1934. The vascularization of cartilage. J Anat 69:47–63.Ichigatani M, Saga T, Yamani K, Yoshizuka M. 2001. Appearance of
vascular endothelial growth factor (VEGF) in femoral head in thegrowing rat. Histol Histopathol 16:463–468.
Kugler JH, Tomlinson A, Wagstaff A, Ward SM. 1979. The role ofcartilage canals in the formation of secondary centers of ossifica-tion. J Anat 129:493–506.
Lametschwandtner A, Lametschwandtner U, Weiger T. 1991. Scan-ning electron microscopy of vascular corrosion casts: technique andapplications: updated review. Scanning Microsc 4:889–941.
Lee ER, Lamplugh L, Davoli MA, Beauchemin A, Chan K, Mort JS,Leblond CP. 2001. Enzymes active in the areas undergoing carti-lage resorption during the development of the secondary ossifica-tion center in the tibia of rats aged 0–21 days: 1, two groups ofproteinases cleave the core protein of aggrecan. Dev Dyn 222:52–70.
Leung DW, Cachianes G, Kuang WJ, Geoddel DW, Ferrara N. 1989.Vascular endothelial growth factor is a secreted angiogenic mito-gen. Science 246:1306–1309.
Levene C. 1964. The patterns of cartilage canals. J Anat 98:515–538.Minet E, Michel G, Remacle J, Michiels C. 2000. Role of HIF-1 as a
transcription factor involved in embryonic development, cancer pro-gression and apoptosis. Int J Mol Med 5:253–259.
Morini S, Pannarale L, Franchitto A, Donati S, Gaudio E. 1999.Microvascular features and ossification process in the femoral headof growing rats. J Anat 195:225–233.
Neufeld N, Cohen T, Ghengrinovitch S, Poltorak Z. 1999. Vascularendothelial growth factor (VEGF) and its receptors. FASEB J 13:9–22.
Nor JE, Christensen J, Mooney DJ, Polverini PJ. 1999. Vascularendothelial growth factor (VEGF)-mediated angiogenesis is associ-ated with enhanced endothelial cell survival and induction of Bcl-2expression. Am J Pathol 154:375–384.
Pannarale L, Morini S, D’ubaldo E, Gaudio E, Marinozzi G. 1997.S.E.M. corrosion-casts study of the microcirculation of the flat bonesin the rat. Anat Rec 247:462–471.
Roach HI, Baker JE, Clarke NM. 1998. Initiation of the bone epiph-ysis in long bones: chronology of interactions between the vascularsystem and the chondrocytes. J Bone Mine Res 13:950–961.
Shweiki D, Itin A, Soffer D, Keshet E. 1992. Vascular endothelialgrowth factor induced by hypoxia may mediate hypoxia-initiatedangiogenesis. Nature 359:843–845.
Skawina A, Litwin JA, Gorezyca J, Miodonski AJ. 1994. Blood vesselsin epiphyseal cartilage of human fetal bone: a scanning electronmicroscopic study of corrosion casts. Anat Embryol 189:457–462.
Stockwell RA. 1971. The ultrastructure of cartilage canals and thesurrounding cartilage in the sheep fetus. J Anat 109:397–410.
Takahashi Y, Bucana CD, Cleary KR, Ellis LM. 1998. p53, vesselcount, and vascular endothelial growth factor expression in humancolon cancer. Int J Cancer 79:34–38.
Tordjman R, Delaire S, Plouet J, Ting S, Gaulard P, Fichelson S,Romeo PH, Lemarchandel V. 2001. Erytrhoblasts are a source ofangiogenic factors. Blood 97:1968–1974.
Visco DM, Hill MA, Van Sickle DC, Kincaid SA. 1990. The develop-ment of centers of ossification of bones forming elbow joints inyoung swine. J Anat 171:25–39.
Wilsman NJ, Van Sickle DC. 1970. The relationship of cartilagecanals to the initial osteogenesis of secondary centers of ossification.Anat Rec 168:381–392.
Wilsman NJ, Van Sickle DC. 1972. Cartilage canals, their morphologyand distribution. Anat Rec 173:79–94.
Yeh LC, Lee JC. 1999. Osteogenic protein-1 increases gene expressionof vascular endothelial growth factor in primary cultures of fetal ratcalvaria cells. Mol Cell Endocrinol 153:113–124.
427VASCULATURE IN RAT HUMERAL HEAD