
Development of rolipram-sensitive, cyclic AMP phosphodiesterase (PDE4) in rat primary neuronal...
7
Developmental Brain Research 130 (2001) 115–121 www.elsevier.com / locate / bres Research report Development of rolipram-sensitive, cyclic AMP phosphodiesterase (PDE4) in rat primary neuronal cultures a a b c Ying Ye , Keasha Jackson , Miles D. Houslay , L. Judson Chandler , d, * James M. O’Donnell a Department of Pharmacology and Therapeutics, Louisiana State University Health Sciences Center, Shreveport, LA, USA b Division of Biochemistry and Molecular Biology, University of Glasgow, Glasgow, Scotland, UK c Department of Physiology and Neuroscience, Medical University of South Carolina, Charleston, SC, USA d Department of Pharmacology, University of Tennessee Health Science Center, 874 Union Avenue, Memphis, TN 38163, USA Accepted 26 June 2001 Abstract The development of PDE4 was examined in primary neuronal cultures of rat cerebral cortex. Three days after culturing, neurons exhibited relatively low PDE4 activity (i.e., rolipram-sensitive PDE activity). It gradually increased over time, approximately doubling by day 12. The increase in activity was accompanied by an increase in the expression of the PDE4A variants, PDE4A5 and PDE4A1, as well as of the synaptic marker protein synapsin I. There was a strong correlation between the expression of the PDE4A variants with that of synapsin I, which suggests that as neurons develop and signal transduction increases there is a regulated increase in PDE4 expression and activity. Consistent with this interpretation, it was found that treatment with the sodium channel blocker tetrodotoxin, which inhibits depolarization-induced neurotransmitter release, reduced the expression of the PDE4A variants. These data demonstrate the developmen- tal regulation of PDE4 in neurons and offer a manner by which the association of PDE4 variants with particular signal transduction pathways may be studied in vitro. 2001 Elsevier Science B.V. All rights reserved. Theme: Neurotransmitters, modulators, transporters and receptors Topic: Second messengers and phosphorylation Keywords: Phosphodiesterase; Neuron; Cyclic AMP; Tetrodotoxin; Synapsin 1. Introduction ly, it has become clear that inhibitors of PDE4 have therapeutic potential. In preclinical and clinical studies, Phosphodiesterases (PDEs) comprise an 11-member such drugs have been shown to have antidepressant effects family of enzymes that hydrolyze the second messengers [18,24,25,33], memory-enhancing effects [1,14,22,40,41], cyclic AMP and cyclic GMP [3,19]. Type 4 PDE (PDE4) utility in the treatment of asthma and related disorders is a cyclic AMP-specific PDE that is encoded by four [20,27,32], and effects on immune function [4]. genes, resulting in the expression of the PDE4 subtypes, PDE4 has been shown to be involved in the signal PDE4A, B, C, and D [21]. This enzyme is characterized by transduction pathway for b-adrenergic receptors in the a high affinity for cyclic AMP, insensitivity to modulation brain. Cyclic AMP formed by stimulation of these re- by cyclic GMP, and potent inhibition by rolipram. Recent- ceptors in cerebral cortical slices is hydrolyzed predomi- nantly by this enzyme [2,34,35]. Lesion- or drug-induced changes in noradrenergic activity in the brain alter both the Abbreviations: PBS, phosphate-buffered saline; PDE, phosphodies- activity and expression of PDE4, particularly the PDE4A terase; PDE4, rolipram-sensitive, low-K , cyclic AMP phosphodiesterase; m subtype [36]; this subtype is highly expressed in brain PDHS, plasma-derived horse serum; TTX, tetrodotoxin regions associated with affect and memory [9]. Further, *Corresponding author. Tel.: 11-901-448-3621; fax: 11-901-448- antidepressant drugs that enhance central noradrenergic or 3849. E-mail address: [email protected] (J.M. O’Donnell). serotonergic neurotransmission increase the expression of 0165-3806 / 01 / $ – see front matter 2001 Elsevier Science B.V. All rights reserved. PII: S0165-3806(01)00219-X
Transcript of Development of rolipram-sensitive, cyclic AMP phosphodiesterase (PDE4) in rat primary neuronal...

Developmental Brain Research 130 (2001) 115–121www.elsevier.com/ locate /bres
Research report
Development of rolipram-sensitive, cyclic AMP phosphodiesterase(PDE4) in rat primary neuronal culturesa a b cYing Ye , Keasha Jackson , Miles D. Houslay , L. Judson Chandler ,
d ,*James M. O’DonnellaDepartment of Pharmacology and Therapeutics, Louisiana State University Health Sciences Center, Shreveport, LA, USA
bDivision of Biochemistry and Molecular Biology, University of Glasgow, Glasgow, Scotland, UKcDepartment of Physiology and Neuroscience, Medical University of South Carolina, Charleston, SC, USA
dDepartment of Pharmacology, University of Tennessee Health Science Center, 874 Union Avenue, Memphis, TN 38163, USA
Accepted 26 June 2001
Abstract
The development of PDE4 was examined in primary neuronal cultures of rat cerebral cortex. Three days after culturing, neuronsexhibited relatively low PDE4 activity (i.e., rolipram-sensitive PDE activity). It gradually increased over time, approximately doubling byday 12. The increase in activity was accompanied by an increase in the expression of the PDE4A variants, PDE4A5 and PDE4A1, as wellas of the synaptic marker protein synapsin I. There was a strong correlation between the expression of the PDE4A variants with that ofsynapsin I, which suggests that as neurons develop and signal transduction increases there is a regulated increase in PDE4 expression andactivity. Consistent with this interpretation, it was found that treatment with the sodium channel blocker tetrodotoxin, which inhibitsdepolarization-induced neurotransmitter release, reduced the expression of the PDE4A variants. These data demonstrate the developmen-tal regulation of PDE4 in neurons and offer a manner by which the association of PDE4 variants with particular signal transductionpathways may be studied in vitro. 2001 Elsevier Science B.V. All rights reserved.
Theme: Neurotransmitters, modulators, transporters and receptors
Topic: Second messengers and phosphorylation
Keywords: Phosphodiesterase; Neuron; Cyclic AMP; Tetrodotoxin; Synapsin
1. Introduction ly, it has become clear that inhibitors of PDE4 havetherapeutic potential. In preclinical and clinical studies,
Phosphodiesterases (PDEs) comprise an 11-member such drugs have been shown to have antidepressant effectsfamily of enzymes that hydrolyze the second messengers [18,24,25,33], memory-enhancing effects [1,14,22,40,41],cyclic AMP and cyclic GMP [3,19]. Type 4 PDE (PDE4) utility in the treatment of asthma and related disordersis a cyclic AMP-specific PDE that is encoded by four [20,27,32], and effects on immune function [4].genes, resulting in the expression of the PDE4 subtypes, PDE4 has been shown to be involved in the signalPDE4A, B, C, and D [21]. This enzyme is characterized by transduction pathway for b-adrenergic receptors in thea high affinity for cyclic AMP, insensitivity to modulation brain. Cyclic AMP formed by stimulation of these re-by cyclic GMP, and potent inhibition by rolipram. Recent- ceptors in cerebral cortical slices is hydrolyzed predomi-
nantly by this enzyme [2,34,35]. Lesion- or drug-inducedchanges in noradrenergic activity in the brain alter both the
Abbreviations: PBS, phosphate-buffered saline; PDE, phosphodies- activity and expression of PDE4, particularly the PDE4Aterase; PDE4, rolipram-sensitive, low-K , cyclic AMP phosphodiesterase;m subtype [36]; this subtype is highly expressed in brainPDHS, plasma-derived horse serum; TTX, tetrodotoxin
regions associated with affect and memory [9]. Further,*Corresponding author. Tel.: 11-901-448-3621; fax: 11-901-448-antidepressant drugs that enhance central noradrenergic or3849.
E-mail address: [email protected] (J.M. O’Donnell). serotonergic neurotransmission increase the expression of
0165-3806/01/$ – see front matter 2001 Elsevier Science B.V. All rights reserved.PI I : S0165-3806( 01 )00219-X

116 Y. Ye et al. / Developmental Brain Research 130 (2001) 115 –121
PDE4A and PDE4B variants [30,36,37]. Overall, these resulting pellet was washed with DMEM containing 10%data suggest that PDE4 is a dynamic component of signal PDHS and resuspended in 40 ml of the same solution. Thistransduction pathways. Changes in presynaptic activity or suspension was passed through gauze to remove any debris
6receptor responsivity result in the regulation of PDE4 to and the volume was adjusted to 3310 cells per 2 ml. Twomaintain homeostasis. ml of suspended cells were added to poly-L-lysine-coated
In addition to undergoing pharmacologically induced culture dishes and incubated at 378C in a humidifiedchanges, PDE4 expression and activity also appear to be incubator with a 5% CO –95% air atmosphere. Three days2
regulated developmentally [38,39]. Initially (1, 5 days of later, the cells were treated with 10 mM b-cytosineage), PDE4 activity in most regions of rat brain is low; as arabinoside (in DMEM containing 10% PDHS) for 2 days.animals mature, the activity increases, reaching adult levels Following this, the neurons were maintained in normalat about 21 days of age. Kinetic analysis reveals that the media for the remainder of the culturing period.change in activity is due to an increase in the V valuemax
for cyclic AMP hydrolysis, suggesting the presence of 2.2. TTX treatmentincreasing amounts of enzyme with development. Con-sistent with such an interpretation, it was found that the On day 3 after plating, neuronal cultures were treatedexpression of PDE4A variants in a number of brain regions with 10 mM TTX or its vehicle. This experiment wasincreases with age [38,39]. Such a finding suggests that as carried out in triplicate dishes using five independentneurons develop and signal transduction increases, in- neuronal preparations. On day 5, the neurons were har-creased PDE4 activity is required. vested and subjected to SDS–PAGE and immunoblot
In order to study the development of PDE4 in neurons, analysis of the PDE4A variants.its activity and expression were examined in primaryneuronal cultures of rat cerebral cortex. Examination of
2.3. PDE4 enzyme assayexpression focused on the PDE4A variants, PDE4A1 andPDE4A5 and an uncharacterized PDE4A variant, termed
PDE4 enzyme activity was measured as previouslyPDE4AX [38,39]. Previous work has shown that the PDE4described [38,39]. Dishes of neurons were washed withsubtype is regulated both developmentally and pharmaco-PBS (pH 7.4), and harvested and homogenized in 100 mllogically [30,36–39]. Consistent with the results of experi-of 40 mM Tris, pH 8.0, 2 mM PMSF. The homogenatements using rat brain tissue [38,39], it was found thatwas centrifuged at 1003g for 15 min and the supernatantPDE4 activity increased as the cultured neurons developed.was used for the PDE4 activity assay.Coincident with the increase in activity, the expression of
The assay was carried out in the presence or absence ofthe PDE4A variants also increased. The developmental100 mM rolipram (to define PDE4 activity), using sampleschange in PDE4 appeared to depend on normal synaptic 3containing 25 mg protein, 100 000 cpm [ H]cyclic AMP,function, since inhibition of neurotransmitter release with7.5 mM unlabeled cyclic AMP, 0.75 mg/ml bovine serumthe sodium channel blocker tetrodotoxin (TTX) preventedalbumin, 10 mM MgCl , 1.25 mM 2-mercaptoethanol, and2the time-dependent increase in PDE4A expression.40 mM Tris–HCl (pH 8.0) at 348C for 10 min. Thereaction was stopped by adding 400 ml of stop solution (10mM EDTA, 40 mM Tris–HCl, pH 7.4). Samples were2. Materials and methodsboiled for 2 min and then placed on ice. Two hundred mgof snake venom (Crotalus atrox) were added to each2.1. Primary neuronal culturessample. The samples were then incubated at 348C for 15min and the reaction stopped by adding 2 ml of ice-coldPrimary cerebral cortical neurons were prepared fromethanol. The incubation mixture was passed through an1-day-old rat pups as described previously [6,7]. The brainanion-exchange column (AG 1-X8, 100–200 mesh, for-was removed from each sodium pentobarbital-anesthetizedmate form). The eluant was mixed with 10 ml scintillationpup and placed in an isotonic solution containing 100 U ofcocktail, and radioactivity was measured using a scintilla-penicillin G, 100 mg of streptomycin and 0.25 mg oftion counter.amphotericin B per ml (pH 7.4). The cerebral cortex was
dissected from the rest of the brain and surface bloodvessels and pia mater were removed. Pia mater-free cortex 2.4. SDS–PAGE and immunoblot analyseswas chopped into 2-mm chunks. The cells were dissociatedby shaking the chunks in 25 ml of isotonic solution (pH SDS–PAGE and immunoblot analyses were carried out7.4) containing 0.25% trypsin at 378C for 5 min. To further as previously described [36]. In brief, dishes of neuronsdissociate the cells, 160 mg DNAase was added to the from the same origin were washed with PBS (pH 7.4), andsuspension, which was then shaken for another 5 min at harvested and homogenized in 100 ml RIPA buffer (150378C. The suspension was diluted with DMEM containing mM NaCl, 0.1% SDS, 0.5% deoxycholate, 1% NP-40, and10% plasma-derived horse serum (PDHS) to a volume of 50 mM Tris–HCl, pH 8). Samples containing 60 mg40–50 ml and then centrifuged at 10003g for 10 min. The protein were separated using SDS–PAGE and transferred
Y. Ye et al. / Developmental Brain Research 130 (2001) 115 –121 117
onto nitrocellulose membrane. The membranes wereblocked with 5% bovine serum albumin for 2 h, washed,and then blotted with anti-PDE4A-selective antibody 271(1:500 dilution in 0.1% bovine serum albumin and 0.5%normal goat serum), anti-synapsin I antibody (1:200), oranti-actin antibody (1:1000) for 1 h. Membranes werewashed and incubated for 1 h with goat anti-rabbitsecondary antisera. Either the ECL (Amersham Life Sci-ence, Little Chalfont, Bucks, UK) or alkaline phosphatase(Gibco-BRL, Gaithersburg, MD) method was used forstaining.
2.5. Protein assay
Protein content of samples was measured using thebicinchoninic acid protein assay [29], using bovine serumalbumin as a standard.
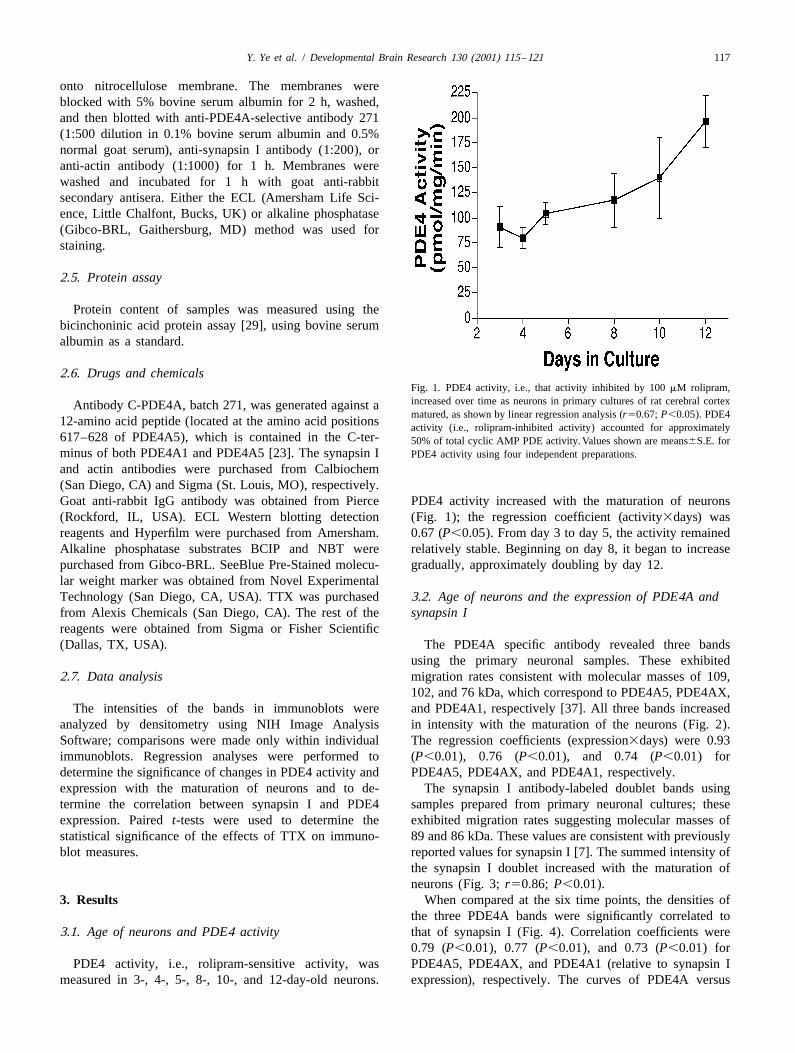
2.6. Drugs and chemicalsFig. 1. PDE4 activity, i.e., that activity inhibited by 100 mM rolipram,increased over time as neurons in primary cultures of rat cerebral cortexAntibody C-PDE4A, batch 271, was generated against amatured, as shown by linear regression analysis (r50.67; P,0.05). PDE4
12-amino acid peptide (located at the amino acid positions activity (i.e., rolipram-inhibited activity) accounted for approximately617–628 of PDE4A5), which is contained in the C-ter- 50% of total cyclic AMP PDE activity. Values shown are means6S.E. forminus of both PDE4A1 and PDE4A5 [23]. The synapsin I PDE4 activity using four independent preparations.
and actin antibodies were purchased from Calbiochem(San Diego, CA) and Sigma (St. Louis, MO), respectively.Goat anti-rabbit IgG antibody was obtained from Pierce PDE4 activity increased with the maturation of neurons(Rockford, IL, USA). ECL Western blotting detection (Fig. 1); the regression coefficient (activity3days) wasreagents and Hyperfilm were purchased from Amersham. 0.67 (P,0.05). From day 3 to day 5, the activity remainedAlkaline phosphatase substrates BCIP and NBT were relatively stable. Beginning on day 8, it began to increasepurchased from Gibco-BRL. SeeBlue Pre-Stained molecu- gradually, approximately doubling by day 12.lar weight marker was obtained from Novel ExperimentalTechnology (San Diego, CA, USA). TTX was purchased 3.2. Age of neurons and the expression of PDE4A andfrom Alexis Chemicals (San Diego, CA). The rest of the synapsin Ireagents were obtained from Sigma or Fisher Scientific(Dallas, TX, USA). The PDE4A specific antibody revealed three bands
using the primary neuronal samples. These exhibited2.7. Data analysis migration rates consistent with molecular masses of 109,
102, and 76 kDa, which correspond to PDE4A5, PDE4AX,The intensities of the bands in immunoblots were and PDE4A1, respectively [37]. All three bands increased
analyzed by densitometry using NIH Image Analysis in intensity with the maturation of the neurons (Fig. 2).Software; comparisons were made only within individual The regression coefficients (expression3days) were 0.93immunoblots. Regression analyses were performed to (P,0.01), 0.76 (P,0.01), and 0.74 (P,0.01) fordetermine the significance of changes in PDE4 activity and PDE4A5, PDE4AX, and PDE4A1, respectively.expression with the maturation of neurons and to de- The synapsin I antibody-labeled doublet bands usingtermine the correlation between synapsin I and PDE4 samples prepared from primary neuronal cultures; theseexpression. Paired t-tests were used to determine the exhibited migration rates suggesting molecular masses ofstatistical significance of the effects of TTX on immuno- 89 and 86 kDa. These values are consistent with previouslyblot measures. reported values for synapsin I [7]. The summed intensity of
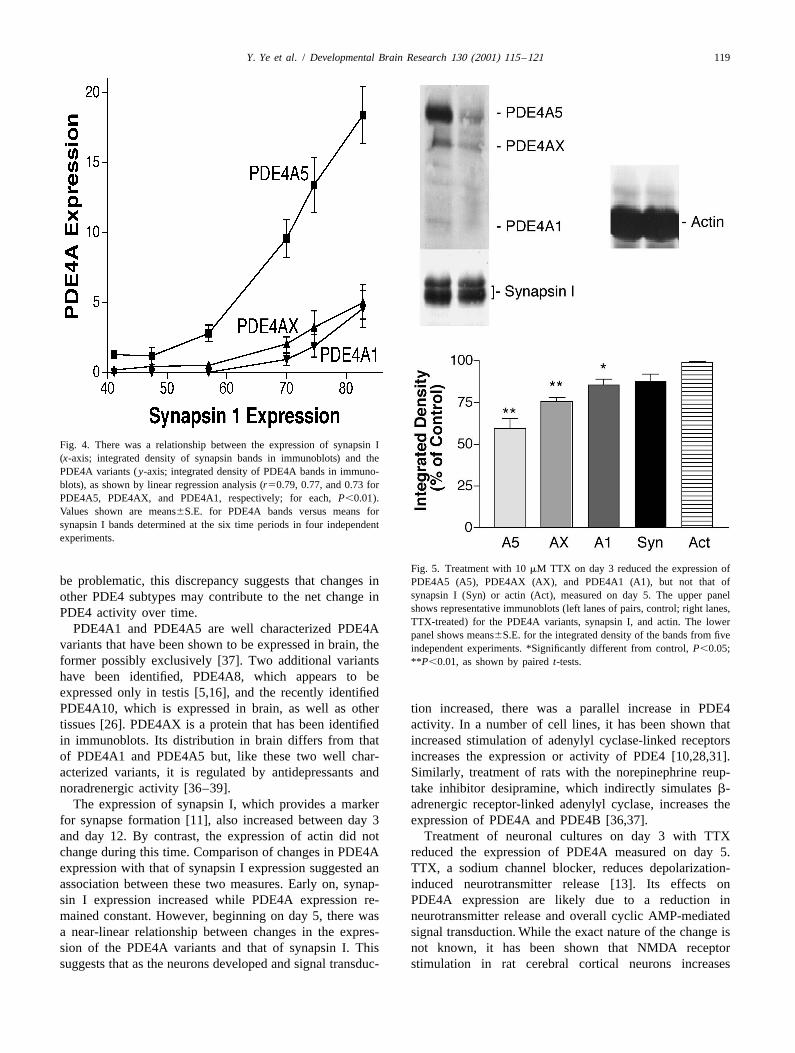
the synapsin I doublet increased with the maturation ofneurons (Fig. 3; r50.86; P,0.01).
3. Results When compared at the six time points, the densities ofthe three PDE4A bands were significantly correlated to
3.1. Age of neurons and PDE4 activity that of synapsin I (Fig. 4). Correlation coefficients were0.79 (P,0.01), 0.77 (P,0.01), and 0.73 (P,0.01) for
PDE4 activity, i.e., rolipram-sensitive activity, was PDE4A5, PDE4AX, and PDE4A1 (relative to synapsin Imeasured in 3-, 4-, 5-, 8-, 10-, and 12-day-old neurons. expression), respectively. The curves of PDE4A versus
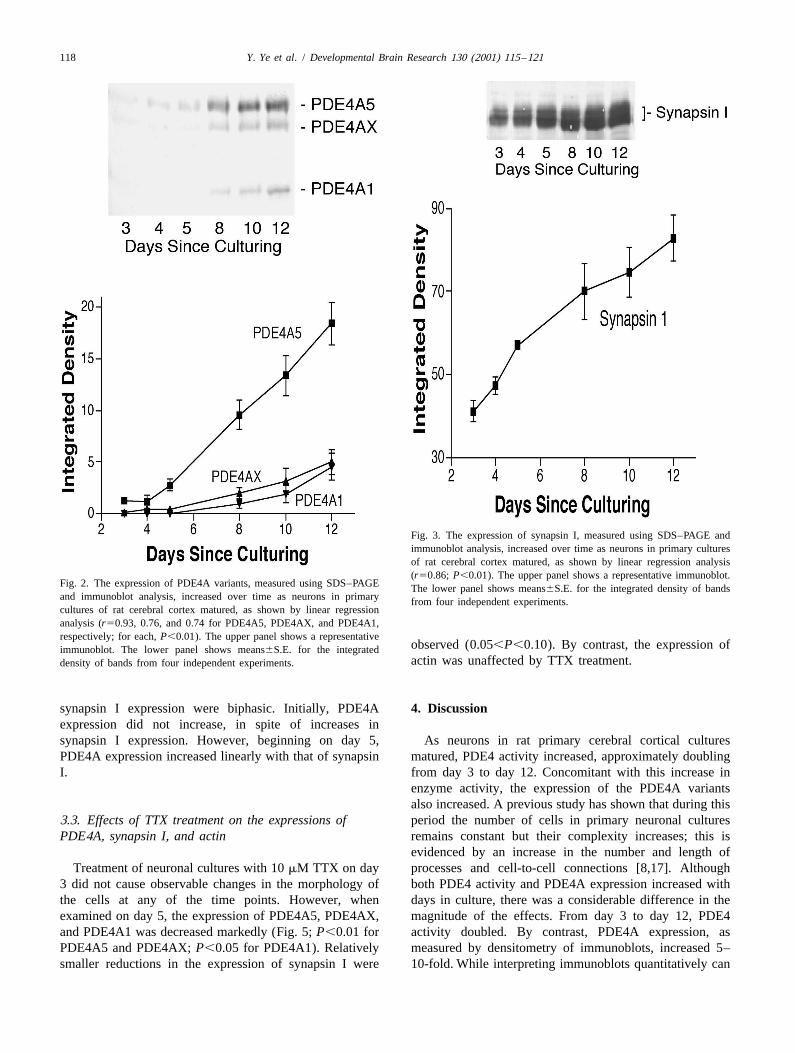
118 Y. Ye et al. / Developmental Brain Research 130 (2001) 115 –121
Fig. 3. The expression of synapsin I, measured using SDS–PAGE andimmunoblot analysis, increased over time as neurons in primary culturesof rat cerebral cortex matured, as shown by linear regression analysis(r50.86; P,0.01). The upper panel shows a representative immunoblot.
Fig. 2. The expression of PDE4A variants, measured using SDS–PAGE The lower panel shows means6S.E. for the integrated density of bandsand immunoblot analysis, increased over time as neurons in primary from four independent experiments.cultures of rat cerebral cortex matured, as shown by linear regressionanalysis (r50.93, 0.76, and 0.74 for PDE4A5, PDE4AX, and PDE4A1,respectively; for each, P,0.01). The upper panel shows a representative
observed (0.05,P,0.10). By contrast, the expression ofimmunoblot. The lower panel shows means6S.E. for the integratedactin was unaffected by TTX treatment.density of bands from four independent experiments.
synapsin I expression were biphasic. Initially, PDE4A 4. Discussionexpression did not increase, in spite of increases insynapsin I expression. However, beginning on day 5, As neurons in rat primary cerebral cortical culturesPDE4A expression increased linearly with that of synapsin matured, PDE4 activity increased, approximately doublingI. from day 3 to day 12. Concomitant with this increase in
enzyme activity, the expression of the PDE4A variantsalso increased. A previous study has shown that during this
3.3. Effects of TTX treatment on the expressions of period the number of cells in primary neuronal culturesPDE4A, synapsin I, and actin remains constant but their complexity increases; this is
evidenced by an increase in the number and length ofTreatment of neuronal cultures with 10 mM TTX on day processes and cell-to-cell connections [8,17]. Although
3 did not cause observable changes in the morphology of both PDE4 activity and PDE4A expression increased withthe cells at any of the time points. However, when days in culture, there was a considerable difference in theexamined on day 5, the expression of PDE4A5, PDE4AX, magnitude of the effects. From day 3 to day 12, PDE4and PDE4A1 was decreased markedly (Fig. 5; P,0.01 for activity doubled. By contrast, PDE4A expression, asPDE4A5 and PDE4AX; P,0.05 for PDE4A1). Relatively measured by densitometry of immunoblots, increased 5–smaller reductions in the expression of synapsin I were 10-fold. While interpreting immunoblots quantitatively can
Y. Ye et al. / Developmental Brain Research 130 (2001) 115 –121 119
Fig. 4. There was a relationship between the expression of synapsin I(x-axis; integrated density of synapsin bands in immunoblots) and thePDE4A variants ( y-axis; integrated density of PDE4A bands in immuno-blots), as shown by linear regression analysis (r50.79, 0.77, and 0.73 forPDE4A5, PDE4AX, and PDE4A1, respectively; for each, P,0.01).Values shown are means6S.E. for PDE4A bands versus means forsynapsin I bands determined at the six time periods in four independentexperiments.
Fig. 5. Treatment with 10 mM TTX on day 3 reduced the expression ofbe problematic, this discrepancy suggests that changes in PDE4A5 (A5), PDE4AX (AX), and PDE4A1 (A1), but not that of
synapsin I (Syn) or actin (Act), measured on day 5. The upper panelother PDE4 subtypes may contribute to the net change inshows representative immunoblots (left lanes of pairs, control; right lanes,PDE4 activity over time.TTX-treated) for the PDE4A variants, synapsin I, and actin. The lower
PDE4A1 and PDE4A5 are well characterized PDE4A panel shows means6S.E. for the integrated density of the bands from fivevariants that have been shown to be expressed in brain, the independent experiments. *Significantly different from control, P,0.05;former possibly exclusively [37]. Two additional variants **P,0.01, as shown by paired t-tests.
have been identified, PDE4A8, which appears to beexpressed only in testis [5,16], and the recently identifiedPDE4A10, which is expressed in brain, as well as other tion increased, there was a parallel increase in PDE4tissues [26]. PDE4AX is a protein that has been identified activity. In a number of cell lines, it has been shown thatin immunoblots. Its distribution in brain differs from that increased stimulation of adenylyl cyclase-linked receptorsof PDE4A1 and PDE4A5 but, like these two well char- increases the expression or activity of PDE4 [10,28,31].acterized variants, it is regulated by antidepressants and Similarly, treatment of rats with the norepinephrine reup-noradrenergic activity [36–39]. take inhibitor desipramine, which indirectly simulates b-
The expression of synapsin I, which provides a marker adrenergic receptor-linked adenylyl cyclase, increases thefor synapse formation [11], also increased between day 3 expression of PDE4A and PDE4B [36,37].and day 12. By contrast, the expression of actin did not Treatment of neuronal cultures on day 3 with TTXchange during this time. Comparison of changes in PDE4A reduced the expression of PDE4A measured on day 5.expression with that of synapsin I expression suggested an TTX, a sodium channel blocker, reduces depolarization-association between these two measures. Early on, synap- induced neurotransmitter release [13]. Its effects onsin I expression increased while PDE4A expression re- PDE4A expression are likely due to a reduction inmained constant. However, beginning on day 5, there was neurotransmitter release and overall cyclic AMP-mediateda near-linear relationship between changes in the expres- signal transduction. While the exact nature of the change ission of the PDE4A variants and that of synapsin I. This not known, it has been shown that NMDA receptorsuggests that as the neurons developed and signal transduc- stimulation in rat cerebral cortical neurons increases

120 Y. Ye et al. / Developmental Brain Research 130 (2001) 115 –121
adenylyl cyclase activity [41]. Given the high level of Scientist Award (MH01231) from the National Institute ofNMDA receptor stimulation observed in these cultures and Mental Health and the National Institute on Alcohol Abuseits sensitivity to inhibition by TTX [8], it is possible that and Alcoholism.glutamatergic mechanisms are involved to some degree.However, the modest tendency toward a reduction insynapsin I expression produced by the TTX treatment
Referencessuggests that a change in synaptic function may contributeto the TTX-induced reduction in PDE4A expression. The
[1] M. Barad, R. Bourtchouladze, D.G. Winder, H. Golan, E. Kandel,lack of observable changes in actin expression suggests Rolipram, a type IV-specific phosphodiesterase inhibitor, facilitatesthat a generalized TTX-induced reduction in protein the establishment of long-lasting long-term potentiation and im-expression is not responsible for the observed effects. proves memory, Proc. Natl. Acad. Sci. USA 95 (1998) 15020–
15025.The present in vitro data are consistent with data[2] R. Barber, T.J. Goka, R.W. Butcher, Cyclic AMP turnover in intactobtained previously in vivo. Examination of regions of rat
tissue: role of cyclic nucleotide phosphodiesterase, Adv. Secondbrain, including cerebral cortex, cerebellum, neostriatum,Messengers Phosphoprotein Res. 25 (1992) 1–11.
hippocampus, olfactory bulb, and brainstem, revealed a [3] J.A. Beavo, M. Conti, R.J. Heaslip, Multiple cyclic nucleotidemarked increase in PDE4 activity between postnatal day 1 phosphodiesterases, Mol. Pharmacol. 46 (1994) 399–405.
[4] B. Bielekova, A. Lincoln, H. McFarland, R. Martin, Therapeuticand day 21 [38,39]; this is a period of considerablepotential of phosphodiesterase-4 and -3 inhibitors in Th1-mediatedneuronal development and synaptogenesis that depends onautoimmune diseases, J. Immunol. 164 (2000) 1117–1124.cyclic AMP function [12]. In cerebral cortex and cere-
[5] G.B. Bolger, I. McPhee, M.D. Houslay, Alternative splicing ofbellum, this is accompanied by an increase in the expres- cAMP-specific phosphodiesterases mRNA transcripts. Characteriza-sion of PDE4, particularly PDE4A5; by contrast, PDEA5 tion of a novel tissue-specific isoform, RNPDE4A8, J. Biol. Chem.
271 (1996) 1065–1071.expression in brainstem diminishes with age [38].[6] L.J. Chandler, N.J. Guzman, C. Sumners, F.T. Crews, MagnesiumPreviously, it has been shown that 6-hydroxydopamine,
and zinc potentiate ethanol inhibition of N-methyl-D-aspartate-stimu-when administered to neonatal rats on postnatal days 1, 2,lated nitric oxide synthase in cortical neurons, J. Pharmacol. Exp.
and 3, produces noradrenergic lesions that lead to the Ther. 271 (1994) 67–75.reduction of PDE4 activity in cerebral cortex, cerebellum, [7] L.J. Chandler, D. Norwood, G. Sutton, chronic ethanol upregulates
NMDA and AMPA, but not kainate receptor subunit proteins in ratand brainstem on postnatal day 5 [39]. In cerebral cortex,primary cortical cultures, Alcohol Clin. Exp. Res. 23 (1999) 363–where the loss of noradrenergic innervation is permanent,370.PDE4 activity remains low into adulthood, as does the
[8] L.J. Chandler, G. Sutton, N.R. Dorairaj, D. Norwood, NMDAexpression of PDE4A. By contrast, in the brainstem, where receptor-mediated directional control of extracellular signal-reg-a compensatory hyperinnervation develops, PDE4 activity ulated kinase (ERK) activity in cortical neuronal cultures, J. Biol.recovers and eventually exceeds that of controls. Again, Chem. 276 (2001) 2627–2636.
[9] J.A. Cherry, R.L. Davis, Cyclic AMP phosphodiesterases arethis suggests that PDE4 is regulated in a manner tolocalized in regions of the mouse brain associated with rein-maintain overall homeostasis of receptor-mediated signalforcement, movement, and affect, J. Comp. Neurol. 407 (1999)
transduction. 287–301.The effect of TTX is much more global than effects [10] M. Conti, M.V. Toscano, L. Petrelli, R. Geremia, M. Stefanini,
produced by noradrenergic lesions or adrenergic antago- Regulation of follicle-stimulating hormone and dibutyryl adenosine39,59-monophosphate of a phosphodiesterase isoenzyme of thenists. While the former treatment reduces signal transduc-Sertoli cell, Endocrinology 110 (1982) 1189–1196.tion mediated by a number of neurotransmitters, the latter
[11] P. De Camilli, S.M. Harris Jr., W.B. Huttner, P. Greengard, Synapsinproduces much more circumscribed effects. This may I (Protein I), a nerve terminal-specific phosphoprotein. II. Itsaccount for the somewhat greater down-regulation of specific association with synaptic vesicles demonstrated by immuno-PDE4A observed in the present study than in previous cytochemistry in agarose-embedded synaptosomes, J. Cell Biol. 96
(1983) 1355–1373.studies that specifically manipulated adrenergic mecha-[12] M.A. Della Fazia, G. Servillo, P. Sassone-Corsi, Cyclic AMPnisms [15,36,37]. The ability of PDE4 to regulate in vitro
signalling and cellular proliferation: regulation of CREB andin response to changes in signal transduction indicates that CREM, FEBS Lett. 410 (1997) 22–24.the primary neuronal culture is a useful model for studying [13] J.J. Dreifuss, A review of neurosecretory granules: their contentsthe association of PDE4 subtypes with receptor-mediated and mechanisms of release, Ann. NY Acad. Sci. 248 (1975) 184–
201.signal transduction mechanisms. It should prove possible[14] T. Egawa, K. Mishima, Y. Matsumoto, K. Iwasaki, K. Iwasaki, M.to use a pharmacological approach to determine which
Fujiwara, Rolipram and its optical isomers, phosphodiesterase 4PDE4 variants are involved in transmission mediated via inhibitors, attenuated the scopolamine-induced impairments of learn-individual receptor types. ing and memory in rats, Jpn. J. Pharmacol. 75 (1997) 275–281.
[15] S.M. Farooqui, K. Zhang, M. Makhay, K. Jackson, S.Q. Farooqui,J.A. Cherry, J.M. O’Donnell, Noradrenergic lesions differentiallyalter the expression of two subtypes of low K cAMP-sensitivemAcknowledgementsphosphodiesterase type 4 (PDE4A and PDE4B) in rat brain, BrainRes. 867 (2000) 52–61.
This research was supported by research grants [16] S.M. Farooqui, F. Al-Bagdadi, M.D. Houslay, G.B. Bolger, R. Stout,(AA10983, MH40694, MH51175) and an Independent R.D. Specian, J.A. Cherry, M. Conti, J.M. O’Donnell, Surgically

Y. Ye et al. / Developmental Brain Research 130 (2001) 115 –121 121
induced cryptorchidism-related degenerative changes in sper- [29] P.K. Smith, R.I. Krohn, G.T. Hermanson, A.K. Mallia, F.H. Gartner,matogonia are associated with loss of cyclic adenosine mono- Measurement of protein using bicinchoninic acid, Anal. Biochem.phosphate-dependent phosphodiesterases type 4 in abdominal testes 150 (1985) 76–85.of rats, Biol. Reprod. 64 (2001) 1583–1589. [30] M. Takahashi, R. Terwilliger, C. Lane, P. Mezes, M. Conti, R.
[17] A. Ferreira, H.Q. Han, P. Greengard, K.S. Kosik, Suppression of Duman, Chronic antidepressant administration increases the expres-synapsin II inhibits the formation and maintenance of synapses in sion of cAMP-specific phosphodiesterase 4A and 4B isoforms, J.hippocampal culture, Proc. Natl. Acad. Sci. USA 92 (1995) 9225– Neurosci. 19 (1999) 610–618.9229. [31] T.J. Torphy, H.L. Zhou, L.B. Cieslinski, Stimulation of beta
[18] W.W. Fleischhacker, H. Hinterhuber, H. Bauer, B. Pflug, P. Berner, adrenoceptors in a human monocyte cell line (U937) up-regulatesC. Simhandl, R. Wolf, W. Gerlach, H. Jaklitsch, M. Sastre-y- cyclic AMP-specific phosphodiesterase activity, J. Pharmacol. Exp.Hernandez et al., A multicenter double-blind study of three different Ther. 263 (1992) 1195–1205.doses of the new cAMP-phosphodiesterase inhibitor rolipram in [32] D.C. Underwood, S. Bochnowicz, R.R. Osborn, C.J. Kotzer, M.A.patients with major depressive disorder, Neuropsychobiology 26 Luttmann, D.W. Hay, P.D. Gorycki, S.B. Christensen, T.J. Torphy,(1992) 59–64. Antiasthmatic activity of the second-generation phosphodiesterase 4
[19] S.H. Francis, I.V. Turko, J.D. Corbin, Cyclic nucleotide phospho- (PDE4) inhibitor SB 297499 (Ariflo) in the guinea pig, J. Phar-diesterases: relating structure and function, Prog. Nucleic Acid Res. macol. Exp. Ther. 287 (1998) 988–995.Mol. Biol. 65 (2000) 1–52. [33] H. Wachtel, H.H. Schneider, Rolipram, a novel antidepressant drug,
[20] R.J. Heaslip, D.Y. Evans, Emetic, central nervous system, and reverses the hypothermia and hypokinesia of monoamine-depletedpulmonary activities of rolipram in the dog, Eur. J. Pharmacol. 286 mice by an action beyond postsynaptic monoamine receptors,(1995) 281–290. Neuropharmacology 25 (1986) 1119–1126.
[21] M.D. Houslay, M. Sullivan, G.B. Bolger, The multienzyme PDE4 [34] M.E. Whalin, R.L. Garrett Jr., W.J. Thompson, S.J. Strada, Correla-cyclic adenosine monophosphate-specific phosphodiesterase family: tion of cell-free brain cyclic nucleotide phosphodiesterase activitiesintracellular targeting, regulation, and selective inhibition by com- to cyclic AMP decay in intact brain slices, Second Messengerspounds exerting anti-inflammatory and antidepressant actions, Adv. Phosphoproteins 12 (1988) 311–325.Pharmacol. 44 (1998) 225–342. [35] Y. Ye, J.M. O’Donnell, Diminished noradrenergic stimulation re-
[22] T. Imanishi, A. Sawa, Y. Ichimaru, M. Miyashiro, S. Kato, T. duces the activity of rolipram-sensitive, high-affinity cyclic AMPYamamoto, S. Ueki, Ameliorating effects of rolipram on experimen- phosphodiesterase in rat cerebral cortex, J. Neurochem. 66 (1996)tally induced impairments of learning and memory in rodents, Eur. 1894–1902.J. Pharmacol. 321 (1997) 273–278. [36] Y. Ye, M. Conti, M.D. Houslay, S.M. Farooqui, M. Chen, J.M.
[23] I. McPhee, L. Pooley, M. Lobban, G. Bolger, M.D. Houslay, O’Donnell, Noradrenergic activity differentially regulates the ex-Identification, characterization and regional distribution in brain of pression of rolipram-sensitive, high-affinity cyclic AMP phospho-RPDE-6 (RNPDE4A5), a novel splice variant of the PDE4A cyclic diesterase (PDE4) in rat brain, J. Neurochem. 69 (1997) 2397–AMP phosphodiesterase family, Biochem. J. 310 (1995) 965–974. 2404.
[24] J.M. O’Donnell, Antidepressant-like effects of rolipram and other [37] Y. Ye, K. Jackson, J.M. O’Donnell, Effects of repeated antidepres-inhibitors of cyclic adenosine monophosphate phosphodiesterase on sant treatment on type 4A phosphodiesterase (PDE4A) in rat brain,behavior maintained by differential reinforcement of low response J. Neurochem. 74 (2000) 1257–1262.rate, J. Pharmacol. Exp. Ther. 264 (1993) 1168–1178. [38] K. Zhang, S.M. Farooqui, J.M. O’Donnell, Ontogeny of rolipram-
[25] J.M. O’Donnell, S. Frith, Behavioral effects of family-selective sensitive, low-K , cyclic AMP-specific phosphodiesterase in ratm
inhibitors of cyclic nucleotide phosphodiesterases, Pharmacol. Bio- brain, Dev. Brain Res. 112 (1999) 11–19.chem. Behav. 63 (1999) 185–192. [39] K. Zhang, S.M. Farooqui, K. Jackson, J.M. O’Donnell, Effects of
[26] G. Rena, F. Begg, A. Ross, C. MacKenzie, I. McPhee, L. Campbell, noradrenergic lesions on the development of rolipram-sensitive, lowE. Huston, M. Sullivan, M.D. Houslay, Molecular cloning, genomic K , cyclic AMP specific phosphodiesterase in rat brain, Dev. Brainm
positioning, promoter identification, and characterization of the Res. 116 (1999) 181–189.novel cyclic AMP-specific phosphodiesterase PDE4A10, Mol. Phar- [40] H.T. Zhang, J.M. O’Donnell, Effects of rolipram on scopolamine-macol. 59 (2001) 996–1101. induced impairment of working and reference memory in the radial-
[27] C. Schudt, H. Tenor, A. Hatzelmann, PDE isoenzymes as targets for arm maze tests in rats, Psychopharmacology 150 (2000) 311–316.anti-asthma drugs, Eur Respir. J. 8 (1995) 1179–1183. [41] H.T. Zhang, A.M. Crissman, N.R. Dorairaj, L.J. Chandler, J.M.
[28] C. Sette, S. Iona, M. Conti, The short-term activation of a rolipram- O’Donnell, Inhibition of cyclic AMP phosphodiesterase (PDE4)sensitive, cAMP specific phosphodiesterase by TSH in thyroid reverses working and memory deficits associated with NMDAFRTL-5 cells is mediated by a cAMP-dependent phosphorylation, J. receptor antagonism, Neuropsychopharmacology 23 (2000) 198–Biol. Chem. 269 (1994) 9245–9252. 204.


















