
Development of an Improved Animal Model of Shigellosis in the Adult Rabbit by Colonic Infection
10
INFECTION AND IMMUNITY, Nov. 1995, p. 4350–4357 Vol. 63, No. 11 0019-9567/95/$04.0010 Copyright q 1995, American Society for Microbiology Development of an Improved Animal Model of Shigellosis in the Adult Rabbit by Colonic Infection with Shigella flexneri 2a G. H. RABBANI, 1 * M. JOHN ALBERT, 2 HAMIDUR RAHMAN, 3 MOYENUL ISLAM, 4 D. MAHALANABIS, 1 I. KABIR, 1 K. ALAM, 2 AND M. ANSARUZZAMAN 2 Clinical Research Centre, Clinical Sciences Division, 1 and Enteric Bacteriology Laboratory, 2 Animal Resources Branch, 3 and Histopathology Unit of the Laboratory Sciences Division, 4 International Centre for Diarrhoeal Disease Research, Bangladesh, Dhaka 1000, Bangladesh Received 22 May 1995/Returned for modification 21 June 1995/Accepted 10 July 1995 Rabbits are not usually susceptible to intestinal Shigella infection without extensive pretreatment, including starvation and administration of antimicrobial, antimotility, and toxic agents (carbon tetrachloride). Most animals infected this way die rapidly and do not always develop colonic lesions and signs of dysentery. We describe here a successful experimental infection in the adult rabbit which does not require preparatory treatment and which reproduced characteristic features of human shigellosis. Unstarved, untreated adult rabbits were infected by direct inoculation of virulent Shigella flexneri 2a (10 ml of 10 7 bacteria per ml) into the proximal colon after ligation of the distal cecum (cecal bypass). Within 24 h of infection, most inoculated animals consistently developed clinical dysentery, characterized by liquid stool mixed with mucus and blood, leukocytosis, anorexia, and weight loss. Histologically, there were edema, exudation, superficial ulceration, and polymorphonuclear infiltrations in the lamina propria; crypt abscess formation; focal hemorrhages; and the presence of immunohistochemically stained S. flexneri in the colonic mucosa. Successful bacterial colonization was indicated by the isolation of the challenge strain of S. flexneri 2a from the colonic contents. None of the control rabbits challenged with nonvirulent S. flexneri or without cecal bypass developed dysentery or colitis. We conclude that successful Shigella infection can be induced by direct colonic inoculation with virulent S. flexneri 2a in adult rabbits without starvation and pretreatment. The colitis is dependent on the virulence of the bacteria and on the cecal bypass. This model should be useful in studies of the immunopathogenesis of Shigella infection. The mechanisms by which Shigella organisms produce intes- tinal damage and bloody mucoid dysentery in humans are poorly understood. One reason is probably the lack of a suit- able animal model in which the disease can be reproduced and studied. To date, only a few species of laboratory animals have been reported to be experimentally infected. Furthermore, most of these experiments were designed to examine a specific characteristic of the host or the pathogen rather than the pathogenic mechanisms of the disease. The guinea pig corneal epithelium (35) has been used to examine the invasive charac- teristics of Shigella spp., and the small intestine of rabbits (2, 4) has been used to study local antibody production and immune protection. Humans, chimpanzees, and monkeys are the natural hosts for Shigella spp., whereas mice, guinea pigs, and rabbits are generally resistant to both natural and experimental infections. However, these animals can be made susceptible to Shigella infection by various forms of pretreatment manipulations, in- cluding (i) starvation; (ii) administration of antimicrobial, antimotility, and toxic agents (carbon tetrachloride); (iii) neu- tralization of gastric acid before oral challenge; and (iv) intra- peritoneal administration of opium (12, 13, 15). However, Shi- gella infections induced in this way in starved animals do not always result in symptomatic disease, there may be high mor- tality among the infected animals, and the pathologic lesions in the intestine are mostly confined to the small bowel rather than the colon, which is the primary site of Shigella infection in humans (36). We believe that shigellosis induced in this way in starved and physiologically manipulated animals represents an artificial infection and is of little practical value unless it re- flects the disease process as it naturally occurs in the normal susceptible host. In this report, we describe a simplified and improved animal model involving an adult rabbit which does not require star- vation or preparatory treatment, allows easy infection with virulent Shigella flexneri 2a, and reproduces the characteristic disease. MATERIALS AND METHODS Animals. Inbred New Zealand White rabbits weighing 1.5 to 2.8 kg of both genders were used in all experiments. Nonfasted animals were randomly selected from different breeding stocks maintained at the animal resource facilities of the International Centre for Diarrhoeal Disease Research, Bangladesh (ICDDR,B). No preparatory treatments were given. The animals were individually caged in an air-cooled room maintained at 258C. The normal feed consisted of a pelleted animal diet containing carbohydrates (52%), protein (16%), fat (3%), fiber (8.5%), and minerals (6.5%) which provided 4 kcal (;17 kJ)/g. The protocol for this study was approved by the Animal Ethics Experimenta- tion Committee of the ICDDR,B. Surgical procedures. (i) Cecal bypass. The animals were anesthetized by intravenous pentothal sodium (30 mg/kg of body weight) given into the central ear vein, and the abdomen was opened by a midline incision under proper aseptic conditions. After exposure of the cecum, a ligature of braided black silk thread (no. 1) was placed approximately 3 to 5 cm proximal to the ceco-colic junction with care so as to avoid injuries to the mesenteric capillaries (Fig. 1). The ligature was so placed that it completely obstructed the cecal lumen above the ileocecal junction while maintaining the ileo-ceco-colic communication (cecal bypass). The purpose of this ligature was to prevent the cecal contents from entering into the proximal colon. In a separate group of control rabbits (n 5 9), the cecal bypass operation was not done, but all other subsequent procedures were the same. (ii) Colonic intubation. The colon, about 15 to 20 cm in length, was then * Corresponding author. Mailing address: ICDDR,B, GPO Box 128, Dhaka 1000, Bangladesh. Phone: 880-2-600171. Fax: 880-2-883116. Electronic mail address: rabbani%[email protected]. 4350 Downloaded from https://journals.asm.org/journal/iai on 06 December 2021 by 179.227.202.56.
Transcript of Development of an Improved Animal Model of Shigellosis in the Adult Rabbit by Colonic Infection

INFECTION AND IMMUNITY, Nov. 1995, p. 4350–4357 Vol. 63, No. 110019-9567/95/$04.0010Copyright q 1995, American Society for Microbiology
Development of an Improved Animal Model of Shigellosis in theAdult Rabbit by Colonic Infection with Shigella flexneri 2a
G. H. RABBANI,1* M. JOHN ALBERT,2 HAMIDUR RAHMAN,3 MOYENUL ISLAM,4
D. MAHALANABIS,1 I. KABIR,1 K. ALAM,2 AND M. ANSARUZZAMAN2
Clinical Research Centre, Clinical Sciences Division,1 and Enteric Bacteriology Laboratory,2 Animal ResourcesBranch,3 and Histopathology Unit of the Laboratory Sciences Division,4 International Centre
for Diarrhoeal Disease Research, Bangladesh, Dhaka 1000, Bangladesh
Received 22 May 1995/Returned for modification 21 June 1995/Accepted 10 July 1995
Rabbits are not usually susceptible to intestinal Shigella infection without extensive pretreatment, includingstarvation and administration of antimicrobial, antimotility, and toxic agents (carbon tetrachloride). Mostanimals infected this way die rapidly and do not always develop colonic lesions and signs of dysentery. Wedescribe here a successful experimental infection in the adult rabbit which does not require preparatorytreatment and which reproduced characteristic features of human shigellosis. Unstarved, untreated adultrabbits were infected by direct inoculation of virulent Shigella flexneri 2a (10 ml of 107 bacteria per ml) into theproximal colon after ligation of the distal cecum (cecal bypass). Within 24 h of infection, most inoculatedanimals consistently developed clinical dysentery, characterized by liquid stool mixed with mucus and blood,leukocytosis, anorexia, and weight loss. Histologically, there were edema, exudation, superficial ulceration, andpolymorphonuclear infiltrations in the lamina propria; crypt abscess formation; focal hemorrhages; and thepresence of immunohistochemically stained S. flexneri in the colonic mucosa. Successful bacterial colonizationwas indicated by the isolation of the challenge strain of S. flexneri 2a from the colonic contents. None of thecontrol rabbits challenged with nonvirulent S. flexneri or without cecal bypass developed dysentery or colitis.We conclude that successful Shigella infection can be induced by direct colonic inoculation with virulent S.flexneri 2a in adult rabbits without starvation and pretreatment. The colitis is dependent on the virulence of thebacteria and on the cecal bypass. This model should be useful in studies of the immunopathogenesis of Shigellainfection.
The mechanisms by which Shigella organisms produce intes-tinal damage and bloody mucoid dysentery in humans arepoorly understood. One reason is probably the lack of a suit-able animal model in which the disease can be reproduced andstudied. To date, only a few species of laboratory animals havebeen reported to be experimentally infected. Furthermore,most of these experiments were designed to examine a specificcharacteristic of the host or the pathogen rather than thepathogenic mechanisms of the disease. The guinea pig cornealepithelium (35) has been used to examine the invasive charac-teristics of Shigella spp., and the small intestine of rabbits (2, 4)has been used to study local antibody production and immuneprotection.Humans, chimpanzees, and monkeys are the natural hosts
for Shigella spp., whereas mice, guinea pigs, and rabbits aregenerally resistant to both natural and experimental infections.However, these animals can be made susceptible to Shigellainfection by various forms of pretreatment manipulations, in-cluding (i) starvation; (ii) administration of antimicrobial,antimotility, and toxic agents (carbon tetrachloride); (iii) neu-tralization of gastric acid before oral challenge; and (iv) intra-peritoneal administration of opium (12, 13, 15). However, Shi-gella infections induced in this way in starved animals do notalways result in symptomatic disease, there may be high mor-tality among the infected animals, and the pathologic lesions inthe intestine are mostly confined to the small bowel rather thanthe colon, which is the primary site of Shigella infection in
humans (36). We believe that shigellosis induced in this way instarved and physiologically manipulated animals represents anartificial infection and is of little practical value unless it re-flects the disease process as it naturally occurs in the normalsusceptible host.In this report, we describe a simplified and improved animal
model involving an adult rabbit which does not require star-vation or preparatory treatment, allows easy infection withvirulent Shigella flexneri 2a, and reproduces the characteristicdisease.
MATERIALS AND METHODS
Animals. Inbred New Zealand White rabbits weighing 1.5 to 2.8 kg of bothgenders were used in all experiments. Nonfasted animals were randomly selectedfrom different breeding stocks maintained at the animal resource facilities of theInternational Centre for Diarrhoeal Disease Research, Bangladesh (ICDDR,B).No preparatory treatments were given. The animals were individually caged in anair-cooled room maintained at 258C. The normal feed consisted of a pelletedanimal diet containing carbohydrates (52%), protein (16%), fat (3%), fiber(8.5%), and minerals (6.5%) which provided 4 kcal (;17 kJ)/g.The protocol for this study was approved by the Animal Ethics Experimenta-
tion Committee of the ICDDR,B.Surgical procedures. (i) Cecal bypass. The animals were anesthetized by
intravenous pentothal sodium (30 mg/kg of body weight) given into the centralear vein, and the abdomen was opened by a midline incision under proper asepticconditions. After exposure of the cecum, a ligature of braided black silk thread(no. 1) was placed approximately 3 to 5 cm proximal to the ceco-colic junctionwith care so as to avoid injuries to the mesenteric capillaries (Fig. 1). The ligaturewas so placed that it completely obstructed the cecal lumen above the ileocecaljunction while maintaining the ileo-ceco-colic communication (cecal bypass).The purpose of this ligature was to prevent the cecal contents from entering intothe proximal colon.In a separate group of control rabbits (n 5 9), the cecal bypass operation was
not done, but all other subsequent procedures were the same.(ii) Colonic intubation. The colon, about 15 to 20 cm in length, was then
* Corresponding author. Mailing address: ICDDR,B, GPO Box 128,Dhaka 1000, Bangladesh. Phone: 880-2-600171. Fax: 880-2-883116.Electronic mail address: rabbani%[email protected].
4350
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.

flushed with 5 to 10 ml of 0.9% NaCl solution warmed at 378C with a 10-mlsyringe fitted with a 20-gauge needle. The purpose of this flushing was to removethe fecal contents from the proximal part of the colon where the cannula was tobe inserted. No other ligature was placed. At about 10 cm distal to the ceco-colicjunction, a small-caliber plastic tube (suction catheter no. 8CH; MallinckrodtLaboratories, Athlone, Ireland) was placed into the proximal colon through asmall incision and tightly secured with a silk ligature. The other end of the tubeexited through the abdominal stoma and was kept fixed on the abdominal wallunder cover of adhesive tape which prevented the animal from biting the tubeafter recovering from anesthesia. This tube was used for introducing the bacterialinoculum and test solutions into the colon and for sampling colonic contents. Theintubated colon was then repositioned into the abdomen, and the incision wasclosed in two layers with an interrupted suture.(iii) Bacterial inoculation. The intubated rabbits (n 5 16) were inoculated
with 10 ml of broth culture containing 107 virulent S. flexneri 2a bacteria per ml(see preparation below) through the transabdominal tube into the proximalcolon. The animals were subsequently observed for the development of diarrheaduring the next 96 h.A separate group of control rabbits (n 5 12) was similarly inoculated with an
avirulent strain of S. flexneri of the same inoculum size (see below). This groupserved as the bacteriologic control.Bacterial culture preparation. The virulent strain of S. flexneri 2a (strain
Y-976/92) was obtained from the National Bacteriologic Laboratory, Stockholm,Sweden (8). The strain was originally isolated from a dysenteric patient inSweden, stocked in Trypticase soy broth (GIBCO Laboratories, Grand Island,N.Y.) with 15% glycerol, and maintained at 2708C. The avirulent strain of S.flexneri was obtained from the stock culture at the ICDDR,B laboratory. Thisorganism is a nonvirulent S. flexneri serotype 5 strain, M90T-A2 (Walter ReedArmy Institute of Research) that lacks the 210-kb virulence-associated plasmid(29). The organisms were subcultured from the stock onto a Trypticase soy agar(GIBCO Laboratories) plate, and after an overnight incubation at 378C, onecolony was transferred to a 100-ml conical flask containing 20 ml of Trypticasesoy broth with 0.6% yeast extract (TSBY). This broth culture was then placed onan orbital shaker at 378C for 16 to 18 h. The next day, an appropriately dilutedculture in TSBY (to contain 107 CFU/ml as determined by bacterial colonycount) was used for animal inoculation. The final count was confirmed by plating0.1 ml of a 1026-diluted culture onto a MacConkey agar plate (Difco Labora-tories, Detroit, Mich.) and counting the number of colonies after overnightincubation at 378C.The virulence of the organisms was tested by their ability to invade the corneal
epithelium of a guinea pig, producing acute keratoconjunctivitis (Sereny test)(35). The virulent strain of S. flexneri 2a used in our experiments consistently gavea positive reaction in response to the Sereny test, whereas the avirulent strain ofS. flexneri was negative.Monitoring of the animal. Measured quantities of food and water were pro-
vided in the cages, and the leftover amounts were measured to determine theamount consumed. Body weight and rectal temperatures were recorded everymorning and evening for 96 h. Excreta were collected and examined for thepresence of blood and mucus. Rectal swabs and colonic samples were obtainedtwice daily (9 a.m. and 4:30 p.m.) and plated on MacConkey agar and Salmo-nella-Shigella agar (Difco Laboratories) for isolation of Shigella organisms. Co-lonic samples diluted in physiologic saline were also plated on these media toobtain quantitative counts of bacteria per milliliter. Colonic contents were ex-
amined microscopically for the presence of inflammatory cells. One drop of thematerial was diluted with 0.2 ml of physiologic saline on a glass slide, coveredwith a glass slip, and examined under a high-power objective microscope. In eachfield, the numbers of leukocytes, erythrocytes, and macrophages were deter-mined and expressed as the mean count in five random fields. Blood samples (3ml) were collected from the ear vein before and 48 h after inoculation fordetermination of total and differential leukocyte counts. At different time periods(24, 48, 72, and 96 h) after the bacterial inoculation, the animals were killed byintravenous pentothal sodium overdose, and the visceral tissues were taken forhistological examination.Autopsy and histologic examination. After laparotomy, the serosal surfaces of
the ileum, colon, and rectum were inspected for signs of inflammation andhemorrhage. If there was any fluid accumulation in the colon, the volume wasmeasured and samples were taken for bacteriologic and biochemical examina-tion. The segment of the intestine from the ileum to the rectum was resected,opened, and inspected for signs of mucosal edema, exudation, hemorrhage,ulceration, necrosis, and perforation. Appropriate segments of the small andlarge intestines were carefully excised, samples of the tissues were fixed in 10%neutral buffered formalin and embedded in paraffin, and then sections were cutat a 5-mm thickness and stained with hematoxylin and eosin. The slides werelabeled and read by a pathologist without prior knowledge of the experimentalconditions.Peroxidase-antiperoxidase staining for shigellae. The presence of invading S.
flexneri 2a cells in the intestinal tissue was demonstrated by light microscopy afterthe bacteria had been stained with a soluble enzyme-antienzyme immune com-plex by using a monoclonal antibody against S. flexneri 2a, MASF II-2 (9). Theimmunohistochemical method described by Mason et al. (27) was used. In thismethod, a paraffin section on a glass slide was placed in a phosphate-bufferedsaline (PBS [pH 7.2]) bath at room temperature for 5 min. After excess liquidhad been removed from the specimen, 4 to 6 drops of normal rabbit serum(dilution 1:5 to 1:20) was added and the mixture was incubated for 30 min.Rabbit anti-mouse immunoglobulin (code no. Z 259, lot no. 071) (Dakopatts,Glostrop, Denmark) diluted to 1:25 was added, and the mixture was incubatedfor 30 min at room temperature. Mouse peroxidase-antiperoxidose (monoclonalmouse PAP code no. P 850, lot 122) (Dakopatts) diluted to 1:100 was added, andthe mixture was incubated for 30 min at room temperature. Chromogenic sub-strate with diaminobenzidine (DAB peroxidase substrate tablet set, product no.D-4168; Sigma Chemicals, St. Louis, Mo.) was added, and the mixture wasincubated for 10 min. After being rinsed with PBS, the tissue was counterstainedwith standard hematoxylin and eosin, and the staining of bacteria was demon-strated by light microscopy.
RESULTS
Dysenteric illness in rabbits. The clinical and bacteriologicresponses of rabbits after colonic inoculation with shigellae areshown in Table 1. Most rabbits inoculated with virulent S.flexneri recovered from anesthesia within 3 to 4 h, appeared
FIG. 1. Diagrammatic representation of the surgical preparation of rabbitcolon for challenge with S. flexneri. The cecum is obstructed by a ligature placedapproximately 3 to 5 cm proximal to the ceco-colic junction, while the ileo-coliccommunication is maintained (cecal bypass). About 10 cm distal to the ceco-colicjunction, a small tube is inserted into the proximal colon for introduction of thechallenge organisms. After bacterial inoculation, the animal is observed for 96 h.
TABLE 1. Clinical and bacteriological responses of rabbitsinoculated with either virulent S. flexneri 2a oravirulent S. flexneri 5 injected into the colon
Clinical characteristic(24 h postinoculation)
Value for group
Virulent S. flexneri2aa-infected rabbits
(n 5 16)
Avirulent S. flexneri5b-infected rabbits
(n 5 12)
Dysentery (%) 90 11c
Blood in stool (%) 40 0Mucus in stool (%) 95 12Stool consistency Soft to liquid Hard pelletRectal temp (8C [mean 6 SD]) 40.0 6 2.6 38.8 6 1.9Total leukocyte (mean 6 SD)count (103)/mm3
14.2 6 3.6 8.3 6 3.1
Isolation of S. flexneri (%) 100 83Anorexia (% [96 h postinocula-tion])
85 15
Body wt loss (% [96 h postinocu-lation)
12.3 6.8
aWild-type strain positive by the Sereny test.b Two hundred ten-kilobase-plasmidless strain (M90T-A2) negative by the
Sereny test.c Only 2 of the 12 rabbits challenged with avirulent S. flexneri developed a
semisoft stool after 48 h of inoculation and recovered spontaneously.
VOL. 63, 1995 ADULT RABBIT MODEL FOR SHIGELLOSIS 4351
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.

sick, and remained severely anorectic for a 24-h postinocula-tion period. Thereafter, eating and drinking resumed graduallybut remained suboptimal throughout the illness. This resultedin a mean loss of 12.3% of the body weight during the periodof illness (Table 1). The rectal temperature was persistentlyelevated, and most animals had moderate leukocytosis. Within24 h of inoculation with virulent S. flexneri 2a, most animalshad developed clinical signs and symptoms of acute rabbitshigellosis characterized by frequent passage of a thick, liquidstool usually mixed with large amounts of mucus and occasion-ally blood. The dysenteric symptoms persisted with increasingseverity in most animals for up to 72 h. Thereafter, the severitytended to decline, but the animal did not recover completelywithin 96 h. After the rabbits had developed diarrhea, theirperianal regions remained constantly wet and soiled with feces.S. flexneri 2a was isolated from the colonic contents of all of theinfected rabbits at 24 and 48 h; thereafter, it was isolated from70 and 50% of the inoculated rabbits at 72 and 96 h postin-fection, respectively.Rabbits inoculated with avirulent S. flexneri showed a tran-
sient rise in body temperature and mild anorexia but no leu-kocytosis (Table 1). The loss of body weight was significantly (P, 0.05) less than in rabbits inoculated with virulent S. flexneriduring the same period of illness (Table 1). However, unlikethe rabbits challenged with virulent S. flexneri 2a, most of therabbits challenged with avirulent S. flexneri did not developdysenteric symptoms during the observation period; only 2 ofthe 12 rabbits had developed a semisoft stool without blood ormucus after 48 h, and they recovered spontaneously.At 48 h, microscopic examinations of colonic contents re-
vealed that the inoculation of the colon with virulent S. flexneri2a consistently resulted in accumulation of an alkaline exudatein the lumen (Table 2). The exudate consisted of mucopuru-
lent to mucohemorrhagic materials covering the colonic mu-cosa. Hemorrhagic inflammation of the colonic mucosa wasindicated by the presence of moderate amounts of erythrocytesand large numbers of fecal leukocytes and macrophages in thecolonic contents.However, animals inoculated with avirulent strains of S.
flexneri showed only mild, nonspecific responses, as indicatedby an absence of blood and the presence of trace amounts ofmucus. In these animals, the colonic materials contained onlyinsignificant numbers of fecal leukocytes and erythrocytes butno macrophages (Table 2).The electrolyte compositions of the colonic fluid and serum
are shown in Table 3. Virulent S. flexneri induced secretion ofcolonic fluid (5 to 10 ml) rich in electrolytes and was isotonic,with serum containing significantly higher concentrations ofbicarbonate and potassium ions and lower concentrations ofchloride ions. There were no significant differences in serumelectrolyte concentrations between the groups of rabbits in-fected with virulent S. flexneri and those infected with avirulentS. flexneri, indicating the absence of major electrolyte abnor-malities. However, in the colons of the rabbits inoculated withavirulent S. flexneri, the fecal material was scanty and too thickfor electrolyte measurements.Appearance of the colonic mucosa in rabbits challenged
with virulent S. flexneri. Necropsies done as early as 8 h afterchallenge revealed mild changes, including swelling of the co-lonic mucosa, especially at the site of intubation. At this stage,no exudate was present in the lumen, and the appearance ofthe mucosa resembled that of a collapsed, noninoculated co-lonic segment. This observation was consistent with the find-ings of mucosal edema and distended lacteals as the predom-inant histologic features (data not shown).At 24 h, the examinations revealed that the gross abnormal-
ities were confined to the colon, the colonic wall was thickened,and the mucosa appeared swollen, injected, and granular withlocalized or patchy areas of hemorrhage. However, definitemucosal necrosis was not seen. In the small intestine, no mu-cosal abnormalities were seen, although the ileocecal valve andthe site of cecal ligation appeared red and edematous.Histologically, the Shigella-infected colonic mucosa was af-
fected by an acute colitis and was markedly different from thatof an uninfected, normal colon (Fig. 2A). The colonic lesionwas characterized by severe acute hemorrhagic inflammationand necrosis in the mucosa and submucosal layers; there waspronounced edema, thrombosis in occasional blood vessels,and active vasculitis in the submucosa (Fig. 2B and C). Thesurface epithelium was eroded in many places, leading to theformation of superficial as well as deep ulcers. The laminapropria appeared congested, edematous, hyperemic, and infil-trated with numerous polymorphonuclear leukocytes. In some
TABLE 2. Light microscopic findings from the colonic contents ofrabbits inoculated with either virulent or avirulent S. flexneria
Characteristic of coloniccontents
Values for group
Virulent S. flexneri2a-infected rabbits
Avirulent S. flexneri5-infected rabbits
Blood Trace NoneMucus Heavy TracepH Alkaline AlkalineNo. of erythrocytes/hpfb 15 6 6 3 6 2No. of fecal leukocytes/hpf 356 10 5 6 3No. of macrophages/hpf 56 2 0
a The results are based on those for samples obtained after 48 h of bacterialinoculation.b hpf, high power field of microscope. Values are means 6 standard devia-
tions.
TABLE 3. Electrolyte composition and osmolality of colonic fluid and serum of rabbits inoculated with either virulent or avirulent S. flexneria
SampleElectrolyte composition (mmol/liter) Osmolality
(mOsmol)Sodium Potassium Chloride Bicarbonate
Virulent S. flexneri 2a-infected rabbitsSerum 137.6 6 6.7 4.1 6 1.0 102.3 6 11.7 24.2 6 4.8 309.2 6 25.6Colonic fluid 112.4 6 9.7 11.9 6 0.7 67.6 6 15.9 54.1 6 17.7 290.0 6 12.3
Avirulent S. flexneri 5-infected rabbitsSerum 141.8 6 8.5 5.8 6 1.9 107 6 12.4 21.8 6 5.2 295.8 6 18.2Colonic fluid Scanty fluid STH STH STH STH
a Data are based on specimens obtained after 48 h of bacterial inoculation and are means 6 standard deviations. STH, fecal material scanty and too thick forelectrolyte determination.
4352 RABBANI ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.

areas, there were aggregates of fibrinopurulent exudate, tissuedebris, and macrophages, which often appeared as a pseudo-membrane adherent to the damaged mucosal surface. In themuscle layer, there were focal hemorrhages and inflammation,and the underlying serosa appeared to be inflamed (serositis).At 48 h after infection, the colonic lumen was filled with a
mucopurulent exudate but was not distended. The colonic mu-cosa was edematous and showed scattered areas of superficialmicroulcers and hemorrhage (Fig. 2D). The most salient his-tologic features were those of an acute hemorrhagic necrotiz-ing colitis characterized by intense mucosal hyperemia. Ulcerswere pronounced and extensive, and there were hemorrhages
in the mucosal and submucosal layers. The lamina propria andsome crypts were infiltrated by inflammatory cells and tissuedebris, often resulting in the formation of crypt abscesses (Fig.3A). A patchy collection of leukocytes was observed in thesubmucosa. There was focal accumulation of inflammatorycells in the muscularis and the serosal layers.At 72 h, the appearance of the colonic mucosa to the naked
eye was markedly different from that observed earlier. Histo-logical examination revealed advanced features of acute in-flammation. Extensive superficial and deep ulcers were ob-served, and there were mucosal and submucosal hemorrhagesand focal necrosis and thrombosis in capillaries. The overall
FIG. 2. (A) Histological features of normal, uninfected rabbit colonic mucosa. Note the intact luminal epithelial surface and absence of cellular infiltrate in thelamina propria (hemotoxylin and eosin stain; magnification, 333). (B) Rabbit colonic mucosa after 24 h of infection with virulent S. flexneri 2a, showing severetransmural inflammation, formation of a microulcer (arrow), large ulcers, a pseudomembrane, and inflammatory infiltrations (magnification, 333). (C) Similarlyinfected colonic tissue showing focus of vasculitis (arrow) with aggregation of polymorphonuclear neutrophils in the underlying muscle coat (magnification, 3132). (D)Infection of rabbit colon after 48 h of challenge with virulent S. flexneri 2a. The arrow indicates a superficial microulcer (magnification, 366).
VOL. 63, 1995 ADULT RABBIT MODEL FOR SHIGELLOSIS 4353
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.
Acrobat Notes
Color plate(s) available. Click on figure caption to view.

histologic picture was that of a severe acute inflammationcharacterized by extensive and deep ulcerations, cellular infil-trations in the lamina propria, edema and congestion of thesubmucosa, crypt abscess formation, pseudomembrane forma-tion, and adhesion of clumps of bacteria on the luminal andcrypt epithelial surfaces. Abnormalities of crypt architecture,including focal crypt dilatation and microabscess formation,were more commonly seen at 72 h than in earlier specimens.Depletion of goblet cells was also more marked at 72 h (datanot shown).At 96 h, the animals became weak, lethargic, and emaciated.
The severity of dysenteric symptoms tended to decline, but theanimals were not completely recovered. Histological examina-tion of the colonic mucosa indicated an overall reduction in theinflammatory response in the lamina propria and hyperplasiaof the crypt epithelium.Immunohistochemistry. Peroxidase-antiperoxidase staining
of the infected colonic tissue with a monoclonal antibody to S.flexneri 2a revealed many positively stained bacilli in the lumenand mucosa at various stages of colonization (Fig. 3B and C).Bacilli were seen penetrating surface epithelial cells in thelamina propria, and some were lying free in the colonic lumen.Rabbits challenged with avirulent S. flexneri. Rabbits inoc-
ulated with an avirulent strain of S. flexneri did not showany inflammatory changes in the colonic mucosa. During nec-ropsy, the colonic mucosa appeared normal except for thepresence of mild edema in a few cases after 24 h of inocula-tion. No edema was observed in the colonic mucosa of rabbitsautopsied after 72 and 96 h. There was no evidence of con-gestion or hemorrhage. Microscopically, the mucosa was in-distinguishable from that of the noninoculated colonic seg-ment (Fig. 2A) in most rabbits. However, 1 of the 12 rabbitsinoculated with avirulent S. flexneri developed mild mucosalinflammation after 48 h but did not develop dysenteric illness.The two rabbits with avirulent S. flexneri infection that devel-
oped mild diarrhea did not have colonic mucosal abnormali-ties.Rabbits challenged without cecal bypass. Experiments with
rabbits challenged without cecal bypass were done to assess theeffects of cecal bypass on the development of colonic infectiondue to virulent S. flexneri. Colonic inoculation with virulent S.flexneri in rabbits without cecal bypass did not result in suc-cessful bacterial colonization. Most of the animals did notdevelop diarrhea and/or histologic evidence of colitis (data notshown). Although postsurgical symptoms (pyrexia and an-orexia) were present in five of the nine rabbits, only two de-veloped semisoft stools after 24 h of inoculation, and theyrecovered spontaneously. Microscopic examinations of colonicaspirate revealed insignificant numbers of inflammatory cells.Histologically, the colonic mucosa appeared normal in most ofthe rabbits; only two of the nine rabbits had mild congestion ofthe proximal colonic mucosa, particularly at the site of intuba-tion, after 48 h of inoculation. These two rabbits did not de-velop diarrhea.Bacteriological response. Quantitative data on the recovery
of virulent and avirulent S. flexneri from the infected colons ofrabbits with and without cecal bypass are presented in Table 4.The mean numbers of viable bacteria recovered (CFU permilliliter) from the colon were significantly (P, 0.05) higher inrabbits infected with virulent S. flexneri with cecal bypass thanin rabbits infected with avirulent S. flexneri without cecal by-pass. The mean numbers of virulent shigellae were also signif-icantly (P , 0.02) higher than those of the avirulent organismsin the colons of rabbits with cecal bypass, both 24 and 72 h afterinoculation. Moreover, when rabbits were challenged with vir-ulent shigellae, the numbers of the organisms in the colon at 72h were significantly lower (P , 0.03) in rabbits without thececal bypass than in those with it. At 24 h, although the num-bers were lower, the difference did not reach statistical signif-icance (P , 0.08).
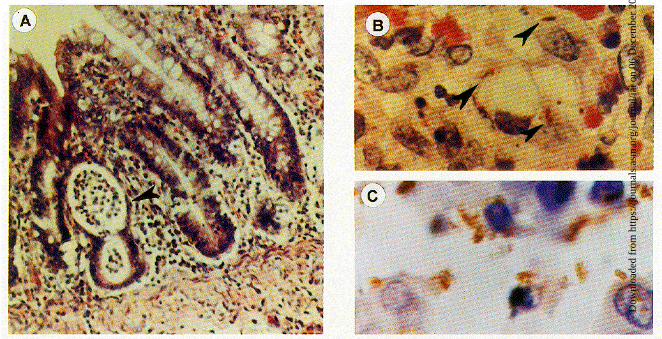
FIG. 3. (A) Rabbit colon after 48 h of infection with virulent S. flexneri 2a. There is crypt abscess formation, and there are cellular infiltrations in the lamina propria(hematoxylin and eosin stain; magnification, 366). (B) Sections of rabbit colonic mucosa showing immunoperoxidase-stained intracellular S. flexneri 2a in the laminapropria (magnification, 3330). (C) Clumps of S. flexneri within the disrupted cells (magnification, 3330).
4354 RABBANI ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.
Acrobat Notes
Color plate(s) available. Click on figure caption to view.

DISCUSSION
The findings of our study indicate that direct colonic inocu-lation of virulent S. flexneri in adult rabbits with cecal bypassresults in successful clinical infection without the need forstarvation and pretreatment of the animals. Within 24 h ofinoculation, the animals developed characteristic signs of Shi-gella dysentery similar to those observed in humans (5, 11). Thesymptoms included passage of frequent loose stool with bloodand mucus, anorexia, weight loss, rise of body temperature,and histological changes of colonic mucosa that were charac-teristics of acute colitis. The microscopic findings of inflamma-tory cells, occasional erythrocytes, and macrophages in theinflammatory exudate of the rabbit colon are also consistentwith the stool findings for children with acute shigellosis (33).Most Shigella-infected animals developed leukocytosis (in-creased peripheral leukocyte count), which is consistent withthe observations in animals given purified Shigella toxin (7) andin humans with shigellosis (6). In our study, successful bacterialcolonization of the colonic mucosa by Shigella organisms wasalso indicated by the isolation of the challenge organisms fromcolonic contents.The symptoms and tissue changes we observed in rabbits are
similar to those found in rhesus monkeys after natural as wellas experimental Shigella infections (14). Rhesus monkeys arethe only animals in which a symptomatic disease can be pro-duced by oral infection with S. flexneri and without recourse tostarvation and pretreatment (14, 32, 38). However, monkeysare costly and difficult to maintain and need to be screened fornaturally acquired Shigella infections (26). LaBrec and Formal(25) found that 18% of the monkeys have natural Shigellainfections which interfere with the development of dysentericsymptoms because of immunity (24, 32). In contrast, rabbits donot acquire natural Shigella infections, and our results indicatethat the susceptibility of the rabbits infected by colonic inocu-lation is much greater than that obtained with intracecal andoral inoculation in monkeys (24, 32). Moreover, infected rab-bits in our study developed predominantly dysenteric illness(blood and mucus), whereas other investigators have observeda combination of watery diarrhea and dysentery in both human(11) and monkey (23, 24) shigelloses.In our study, the success of colonic infection in rabbits de-
pended on several factors. This might be related to the route ofinoculation of the bacteria, which were administered directlyinto the colon in our study and into the stomach or jejunum bythe previous investigators (2, 13, 14, 38). It is understandablethat direct inoculation of the organisms into the intended site
of infection (i.e., the colon) is more likely to result in a suc-cessful colonization than when the inoculum is introduced intothe upper small intestine, requiring the organisms to surviveand travel down the whole length of the small bowel against ahost of enteric defense mechanisms. Moreover, motility in thecolon is lower than that in the small intestine and might pro-vide the opportunity for the bacteria to establish the infectionwithout the need for antimotility drugs or surgical stasis. In thisrespect, our procedures are comparable to a technique de-scribed by Spira et al. (37) as the RITARD (removable intes-tinal tie adult rabbit diarrhea) model for the studies of Vibriocholerae and enterotoxigenic Escherichia coli infections. In theRITARD model, the cecum is ligated permanently, the ileumis obstructed temporarily, and the inoculum is injected into thejejunum, bypassing the gastric juice. It is presumed that theileal tie helps to localize the bacteria within the intestinallumen and to overcome the mucosal defense mechanisms dur-ing the colonizing process.In our study, the development of colonic infection is also
facilitated by the ligation of the cecum, which prevents the flowof cecal contents into the colon. The critical role of the cecalbypass in the development of colonic infection is further indi-cated by the observation that only the rabbits with the cecalbypass developed acute dysenteric illness and colitis, whereasrabbits without the cecal bypass remained largely asymptom-atic. Nevertheless, a few rabbits (two of nine) did develop somesymptoms, including a semisoft stool, mild mucosal edema,and postsurgery pyrexia. These symptoms were mostly nonspe-cific and resolved spontaneously.The mechanism by which the cecal bypass enhances the
development of bacterial colitis in this model cannot be com-pletely ascertained. However, we postulate that fecal diversiondisrupts the normal microecology of the cecocolic flora, thusdepriving the colon of the protective and nutritive effects me-diated by the products of bacterial metabolism of the cecaldigesta in the normal animal. This hypothesis is consistent withobservations that short-chain fatty acids (SCFAs) produced bythe bacterial fermentation of ingested fibers in the relativelylarge cecum of the herbivores, such as rabbits, provide themajor metabolic sources of energy and specific nutrients forthe colonocytes (30, 31). Our observations showing that therate of isolation of the inoculated bacteria from the non-by-passed colon was significantly lower are consistent with theknown inhibitory effects of intestinal contents on pathogenicorganisms, including shigellae (17–19). In the human colon,bacterial production of SCFAs (acetate, propionate, andbutyrate) from the unabsorbed carbohydrates has been shownto be naturally protective and therapeutically effective inthe management of inflammatory bowel diseases (16, 39, 40).SCFAs have also been shown to possess a direct pH-dependentantibacterial effect against pathogenic shigellae (17–19) and adirect trophic cytoprotective effect in inflammatory bowel dis-eases (4, 40). A recent study with rats indicates that the inhi-bition of bacterial production of SCFAs by treatment withmetronidazole is associated with reduced intestinal adaptationand nutritional recovery after massive small bowel resection(1).Besides cecal ligation, the only other surgical procedure we
employed was the intubation of the proximal colon for intro-duction of the bacterial inoculum. However, this optional in-tubation can be omitted, and the inoculum can be injecteddirectly into the colon with a syringe. The latter method ofinoculation would be preferable in short-term studies in whichrepeated sampling from or introduction into the colon is notnecessary. The tube was used in our study for sampling coloniccontents and for infusing various test solutions into the colon
TABLE 4. Postinoculation quantitative recovery of virulent andavirulent S. flexneri from the colonic contents of rabbits
with and without cecal bypassa
Rabbit group
Recovery (log CFU/ml) of:
VirulentS. flexneri 2a
AvirulentS. flexneri 5
With cecal bypass (n 5 12)Inoculum dose 7.0 7.024 h postinoculation 6.626 3.21 3.41 6 2.8772 h postinoculation 4.786 1.82 2.33 6 1.60
Without cecal bypass (n 5 14)Inoculum dose 7.0 7.024 h postinoculation 4.346 3.71 2.30 6 1.4672 h postinoculation 2.556 1.54 2.17 6 1.50
a Except for inoculum dose, values are means 6 standard deviations.
VOL. 63, 1995 ADULT RABBIT MODEL FOR SHIGELLOSIS 4355
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.

over a 4-day period for the purpose of evaluating the effects ofSCFAs on Shigella-induced colitis (unpublished data).The morphological changes observed in the colonic mucosa
of the Shigella-infected rabbits are characteristic of an acuteinflammatory response with progressively increasing severityduring the first 72 h of observation. The inflammation wasusually limited to the superficial lamina propria, with decliningseverity from the epithelial surface toward the muscularis mu-cosa. Similar histologic changes due to Shigella infection havebeen described in the ligated small intestinal loops of rabbits(4) and in the colons of monkeys (21, 22). In human shigellosis,the histologic abnormalities of the colon and rectum, as de-scribed by different observers (3, 10, 20, 28), are also consistentwith our observations in experimentally infected rabbits.In our study, the colitis induced by infection with virulent S.
flexneri appears to be specific, since avirulent S. flexneri did notelicit colonic inflammation and failed to induce bacterial col-onization and symptomatic disease. Although 1 of the 12 rab-bits challenged with avirulent shigellae did develop mild colitis,this animal did not develop disease. This finding is consistentwith the observations that avirulent shigellae (mutants with thevirulence gene deleted) are incapable of colonizing the intes-tinal mucosa and fail to stimulate a specific antibody responsein human shigellosis (29, 34).To be an ideal model for the pathogenic studies of shigel-
losis, the animal should be physiologically normal and immu-nologically competent, should have a normally functioning in-testine without extensive alteration of the enteric environment,and should require a minimum of preparatory manipulations.The animal should develop dysenteric symptoms and colonicmucosal abnormalities similar to those of the human disease.The adult rabbit model that we have developed and describedhere meets these conditions to a large extent. Moreover, ourprocedures resulted in an infection in the rabbit which permit-ted repeated sampling from the site of infection over an ex-tended period of time. This allows one investigator to study thekinetics of Shigella infection, including bacteriological, mor-phological, and chemotherapeutic interventions. We believethis model or perhaps a modification of it will be useful for thestudies of toxin production and local immune response in shi-gellosis. However, this model needs to be further characterizedby specifically standardizing the size of the inoculum to pro-duce the desired intensity and duration of infection, the extentof cecal occlusion, and the grades of clinical and pathologicalseverity of the infection. Also, the susceptibility of the model toinfections due to other shigellae, including S. dysenteriae type 1and possibly non-Shigella organisms, including enteroinvasiveE. coli, needs to be studied.
ACKNOWLEDGMENTS
This study was funded by the United States Agency for InternationalDevelopment (USAID) under grant DPE-5986-A-1009-00 to the In-ternational Centre for Diarrhoeal Disease Research, Bangladesh(ICDDR,B). The ICDDR,B is supported by the aid agencies of theGovernments of Australia, Bangladesh, Belgium, Canada, China, Den-mark, Germany, Japan, The Netherlands, Norway, the Republic ofKorea, Saudi Arabia, Switzerland, the United Kingdom, and theUnited States; international organizations, including the Arab GulfFund, the Asian Development Bank, the International Atomic EnergyCentre, the United Nations Children’s Fund (UNICEF), the UnitedNations Development Programme (UNDP), the United Nations Pop-ulation Fund (UNFPA), and the World Health Organization (WHO);private foundations, including the Ford Foundation, the PopulationCouncil, the Rockefeller Foundation, and the Sasakawa Foundation;and private organizations, including American Express Bank, BayerAG, CARE, Helen Keller International, The Johns Hopkins Univer-sity, the Swiss Red Cross, and the University of California at Davis.
We thank K. A. Al-Mahmood, M. Hossain, M. Nasir, and K. M.Safiullah for helping with the animal studies. We also thank BradleySack and Z. U. Ahmed for their useful comments and Josephine Sackfor editorial help.
REFERENCES1. Aghdassi, E., H. Plapler, R. Kurian, N. Raina, D. Royall, K. N. Jeejeebhoy,Z. Cohen, and J. P. Allard. 1994. Colonic fermentation and nutritionalrecovery in rats with massive small bowel resection. Gastroenterology 107:637–642.
2. Ahmed, Z. U., M. R. Sarker, and D. A. Sack. 1990. Protection of adult rabbitand monkeys from lethal shigellosis by oral immunization with a thymine-requiring and temperature-sensitive mutant of Shigella flexneri Y. Vaccine8:153–158.
3. Anand, B. S., V. Malhotra, S. K. Bhattacharya, P. Datta, D. Datta, D. Sen,M. K. Bhattacharya, P. P. Mukherjee, and S. C. Pal. 1986. Rectal histologyin acute bacillary dysentery. Gastroenterology 90:654–660.
4. Arm, H. G., T. M. Floyd, J. E. Faber, and J. R. Hayes. 1965. Use of ligatedsegments of rabbit small intestine in experimental shigellosis. J. Bacteriol. 89:803–809.
5. Bennish, M. L., A. K. Azad, and D. Yousefzadeh. 1991. Intestinal obstructionduring shigellosis: incidence, clinical features, risk factors and outcome.Gastroenterology 101:626–634.
6. Butler, T., M. R. Islam, and P. K. Bardhan. 1984. The leukemoid reaction inshigellosis. Am. J. Dis. Child. 138:162–165.
7. Butler, T., H. Rahman, K. A. Al-Mahmood, M. Islam, P. Bardhan, I. Kabir,and M. M. Rahman. 1985. An animal model of haemolytic-uraemic syn-drome in shigellosis: lipopolysaccharides of Shigella dysenteriae 1 and Shigellaflexneri produce leucocyte-mediated renal cortical necrosis in rabbits. Br. J.Exp. Pathol. 66:7–15.
8. Carlin, N. I. A., and A. A. Lindberg. 1987. Monoclonal antibodies specific forShigella flexneri lipopolysaccharides: clones binding to type IV, V, and VIantigens, group 3,4 antigen, and an epitope common to all Shigella flexneriand Shigella dysenteriae type 1 strains. Infect. Immun. 55:1412–1420.
9. Carlin, N. I. A., M. Rahman, D. A. Sack, M. Ansari Zaman, B. Kay, and A. A.Lindberg. 1989. Use of monoclonal antibodies to type Shigella flexneri inBangladesh. J. Clin. Microbiol. 27:1163–1166.
10. Dickinson, R. J., H. M. Gilmour, and D. B. L. McCleland. 1979. Rectalbiopsy in patients presenting to an infectious disease unit with diarrhoealdisease. Gut 20:141–148.
11. DuPont, H. L., R. B. Hornick, A. T. Dawkins, M. J. Synder, and S. B. Formal.1969. The response of man to virulent Shigella flexneri 2a. J. Infect. Dis. 119:296–299.
12. Dutta, N. K., and M. K. Habbu. 1955. Experimental cholera in infant rabbits:a method for chemotherapeutic investigation. J. Pharmacol. 10:153–159.
13. Formal, S. B., G. J. Dammin, H. Schneider, and E. H. LaBrec. 1959. Exper-imental Shigella infections. II. Characteristics of a fatal enteric infection inguinea pigs following the subcutaneous inoculation of carbon tetrachloride.J. Bacteriol. 78:800–804.
14. Formal, S. B., E. H. LaBrec, and H. Schneider. 1965. Pathogenesis of bacil-lary dysentery in laboratory animals. Fed. Proc. 24:29–34.
15. Freter, R. 1956. Experimental enteric Shigella and Vibrio infections in miceand guinea pigs. J. Exp. Med. 104:411–418.
16. Harig, J. M., K. H. Soergel, R. A. Komorowski, and C. M. Wood. 1989.Treatment of diversion colitis with short-chain fatty acid irrigation. N. Engl.J. Med. 320:23–28.
17. Hentges, D. J. 1967. Influence of pH on the inhibitory activity of formic andacetic acids for Shigella. J. Bacteriol. 93:2029–2030.
18. Hentges, D. J. 1967. Inhibition of Shigella flexneri by the normal intestinalflora. I. Mechanisms of inhibition by Klebsiella. J. Bacteriol. 93:1369–1373.
19. Hentges, D. J., and B. R. Maier. 1970. Inhibition of Shigella flexneri by thenormal intestinal flora. III. Interactions with Bacteroides fragilis strains invitro. Infect. Immun. 2:364–370.
20. Islam, M. M., A. K. Azad, P. K. Bardhan, R. Raqib, and D. Islam. 1994.Pathology of shigellosis and its complications. Histopathology 24:65–71.
21. Karnell, A., F. P. Reinholt, S. Katakura, and A. A. Lindberg. 1991. Shigellaflexneri infection: a histopathologic study of colonic biopsies in monkeysinfected with virulent and attenuated bacterial strains. Acta Pathol. Micro-biol. Immunol. Scand. 99:787–796.
22. Katakura, S., F. P. Reinholt, A. Karnell, P. T. Huan, D. D. Trach, and A. A.Lindberg. 1990. The pathology of Shigella flexneri infection in rhesus mon-keys: an endoscopic and histopathological study of colonic lesions. ActaPathol. Microbiol. Immunol. Scand. 98:313–319.
23. Keusch, G. T. 1979. Shigella infections. Clin. Gastroenterol. 8:645–662.24. Kinsey, M. D., S. B. Formal, G. J. Dammin, and R. A. Giannella. 1976. Fluid
and electrolyte transport in rhesus monkeys challenged intracecally withShigella flexneri 2a. Infect. Immun. 14:368–371.
25. LaBrec, E. H., and S. B. Formal. 1961. Experimental Shigella infections. IV.Fluorescent antibody studies of an infection in guinea pigs. J. Immunol.87:562–572.
26. LaBrec, E. H., H. Schneider, T. J. Magnani, and S. B. Formal. 1964. Epi-thelial cell penetration as an essential step in the pathogenesis of bacillary
4356 RABBANI ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.

dysentery. J. Bacteriol. 88:1503–1518.27. Mason, D. Y., J. L. Cordell, Z. Abdulaziz, M. Naiem, and G. Bordenave.
1982. Preparation of peroxidase-antiperoxidase (PAP) complexes for immu-nohistological labeling of monoclonal antibodies. J. Histochem. Cytochem.30:1114–1122.
28. Mathan, M. M., and V. I. Mathan. 1986. Ultrastructural pathology of therectal mucosa in Shigella dysentery. Am. J. Pathol. 123:25–38.
29. Oberhelman, R. A., D. S. Kopecko, E. Salazar-Lindo, E. Gotuzzo, J. M.Buysse, M. M. Venkatesan, A. Yi, C. Fernandez-Prada, M. Guzman, R.Leon-Barua, and R. B. Sack. 1991. Prospective study of systemic and mu-cosal immune responses in dysenteric patients to specific Shigella invasionplasmid antigens and lipopolysaccharides. Infect. Immun. 50:2341–2350.
30. Rabassa, A. A., and A. I. Rogers. 1992. The role of short-chain fatty acidmetabolism in colonic disorders. Am. J. Gastroenterol. 87:419–423.
31. Roediger, W. E. W. 1980. Role of anaerobic bacteria in the metabolic welfareof the colonic mucosa in man. Gut 21:793–798.
32. Rout, W. R., S. B. Formal, R. A. Giannela, and G. J. Dammin. 1975. Patho-physiology of Shigella diarrhea in the rhesus monkey: intestinal transport,morphological, and bacteriological studies. Gastroenterology 68:270–278.
33. Salam, M. A., and M. L. Bennish. 1988. Therapy for shigellosis. I. Random-ized, double-blind trial of nalidixic acid in childhood shigellosis. J. Pediatr.113:901–907.
34. Sansonetti, P. J., A. Fontaine, H. d’Hauteville, and L. Bernardini. 1991.ompB (osmoregulation) and icsA (cell to cell spread) mutants of Shigellaflexneri: vaccine candidates and probes to study the pathogenesis of shigel-losis. Vaccine 9:416–422.
35. Sereny, B. 1959. Acquired natural immunity following recovery from kera-toconjunctivitis shigellosa. J. Hyg. Epidemiol. Microbiol. Immunol. III:292.
36. Speelman, P., I. Kabir, and M. Islam. 1984. Distribution and spread ofcolonic lesions in shigellosis: a colonoscopic study. J. Infect. Dis. 150:899–903.
37. Spira, W. M., R. B. Sack, and J. F. Froehlich. 1981. Simple adult rabbitmodel for Vibrio cholerae and enterotoxigenic Escherichia coli diarrhea.Infect. Immun. 32:739–747.
38. Takeuchi, A., S. B. Formal, and H. Sprinz. 1968. Experimental acute colitisin the rhesus monkey following peroral infection with Shigella flexneri. Am.J. Pathol. 52:503–526.
39. Treem, W. R., N. Ahsan, M. Shoup, and J. S. Hyams. 1994. Fecal short-chainfatty acids in children with inflammatory bowel disease. J. Pediatr. Gastro-enterol. Nutr. 18:159–164.
40. Vernia, P., R. Caprilli, G. Latella, F. Barbetti, F. M. Magliocca, and M.Cittadini. 1988. Fecal lactate and ulcerative colitis. Gastroenterology 95:1564–1568.
Editor: P. J. Sansonetti
VOL. 63, 1995 ADULT RABBIT MODEL FOR SHIGELLOSIS 4357
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.

Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 06
Dec
embe
r 20
21 b
y 17
9.22
7.20
2.56
.


















