
Department of Chemistry College of Science National Taiwan ...
78
國立臺灣師範大學理學院化學研究所 碩士論文 Department of Chemistry College of Science National Taiwan Normal University Master Thesis 以單分子螢光共振能量轉移光譜研究人類端粒序列形成的鳥嘌 呤四股結構之構形變化與動力學數據分析在不同實驗因素下的影響 Single-molecule Fluorescence Resonance Energy Transfer Spectroscopy Study on the Conformational Interconversion Kinetics of Human Telomere G-quadruplexes Influenced by Local Chemical Environment 許顥頤 Hao-Yi Hsu 指導教授 : 李以仁 博士 Advisor : I-Ren Lee, Ph.D 中華民國 105 年 7 月 July, 2016
Transcript of Department of Chemistry College of Science National Taiwan ...

國立臺灣師範大學理學院化學研究所
碩士論文
Department of Chemistry
College of Science
National Taiwan Normal University
Master Thesis
以單分子螢光共振能量轉移光譜研究人類端粒序列形成的鳥嘌
呤四股結構之構形變化與動力學數據分析在不同實驗因素下的影響
Single-molecule Fluorescence Resonance Energy Transfer Spectroscopy Study
on the Conformational Interconversion Kinetics of Human Telomere G-quadruplexes
Influenced by Local Chemical Environment
許顥頤
Hao-Yi Hsu
指導教授 : 李以仁 博士
Advisor : I-Ren Lee, Ph.D
中華民國 105 年 7 月
July, 2016

i
誌謝
非常榮幸能進入李以仁老師的實驗室,記得剛進入實驗室的前半年,實驗室
一切都還在架設中,每天除了閱讀文獻還是閱讀文獻,從文獻上所理解的單分子
概念打破了以前我對化學實驗的想法。但是,在實際進行實驗之前,單分子研究
僅止於抽象的概念。從第一次自己完成實驗到之後學會自己設計實驗的成就感都
是勉勵我不斷往前的力量。當然,在實驗的過程中難免會遇到不少麻煩,但是每
周的實驗室會議中,老師總會給予我們建議,讓我可以去思考該如何解決眼前的
問題,其中讓我學到最多的是從解決問題的過程中如何找到更有效率的方式去處
理問題,我覺得這項能力是就讀研究所這兩年最大的收穫。
除了在實驗室進行的研究,非常感謝台大李弘文老師以及溫進德老師提供實
驗室之間學術交流的機會,每次的會議都讓我對實驗有許多新的想法也看到其他
同學在做的實驗和解決問題的想法。還有系上的洪偉修老師,指點我對事情的看
法可以有不同的角度,不要被自己的想法給限制,也讓我重新思考人生的方向。
感謝台大生科系的張凱鈞學長給予實驗上的建議,讓我的實驗得以順利進行。
以及感謝指導教授李以仁老師在實驗上給我寶貴的經驗以及充足的資源,在
實驗出現分歧的時候仍然耐心的傾聽我的實驗想法,並適時的給予協助。還有實
驗室的同學們,在實驗室的每一天都非常開心。最後要感謝我的父母,默默的給
予支持,讓我可以無後顧之憂的做自己想做的事,也謝謝身邊所有幫助過我的朋
友,謝謝。

ii
目錄
摘要............................................................................................................................... iv
Abstract .......................................................................................................................... v
圖目錄.......................................................................................................................... vii
表目錄........................................................................................................................... xi
第一章 緒論.......................................................................................................... 12
1-1 端粒.................................................................................................. 12
1-1.1 端粒的發現 ............................................................................ 12
1-1.2 人類端粒序列 ........................................................................ 13
1-1.3 端粒與抗癌藥物的發展 ........................................................ 15
1-2 鳥嘌呤四聚體(G-Quadruplex) ........................................................ 16
1-2.1 鳥嘌呤四聚體概述 ................................................................ 16
1-2.2 鳥嘌呤四聚體的結構與研究 ................................................ 17
1-2.3 端粒上的鳥嘌呤四聚體 ........................................................ 18
1-3 研究動機.......................................................................................... 20
第二章 實驗架構與方法...................................................................................... 21
2-1 單分子技術與發展.......................................................................... 21
2-2 螢光共振能量轉移(Förster resonance energy transfer).................. 22
2-3 全內反射式螢光顯微術(Total internal reflection fluorescence
microscopy,TIRF) ............................................................................................. 24
2-3.1 實驗樣品槽的組裝與固定樣品於修飾後的表面 ................ 24
2-3.2 全內反射原理與螢光顯微鏡 ................................................ 24
2-4 數據校正與分析.............................................................................. 25
2-5 酵素型除氧系統與顯影緩衝溶液.................................................. 26

iii
2-5.1 螢光放光與去活化機制 ........................................................ 26
2-6 酵素型除氧系統反應機制與作用.................................................. 27
2-7 石英玻片表面修飾.......................................................................... 28
2-8 螢光染料分子的標記方法.............................................................. 28
2-9 寡核苷酸黏合反應.......................................................................... 29
2-10 圓二色光譜(Circular dichroism spectroscopy) ........................... 29
2-10.1 圓二色性(Circular dichroism) ............................................. 29
2-10.2 鳥嘌呤四股結構的區別 ...................................................... 29
第三章 實驗結果與討論...................................................................................... 31
3-1 螢光共振能量轉移效率直方圖(FRET efficiency Histogram ) ..... 31
3-2 圓二色光譜...................................................................................... 31
3-3 酵素型除氧系統的改變.................................................................. 32
3-4 螢光共振能量轉移效率-時間軌跡圖 ............................................ 33
3-5 提升空間解析度的實驗設計.......................................................... 34
第四章 總結.......................................................................................................... 37
參考文獻...................................................................................................................... 67

iv
摘要
人類端粒序列為一段富含鳥嘌呤的序列,可形成 DNA 二級結構,鳥嘌呤四
股結構。先前的研究指出金屬陽離子對鳥嘌呤四股結構具有穩定其結構的效果,
在生理條件下,鉀、鈉與銨等陽離子分別能穩定鳥嘌呤四股結構,形成平行、混
和型以及反平行構形,在整體實驗和單分子實驗均已經得到證實。先前文獻證實
鎂離子和鉀離子共存可穩定此結構,因此我們想要知道單純鎂離子是否能幫助人
類端粒序列形成特定的鳥嘌呤四聚體或穩定其結構,對於已被證實存在人體的鳥
嘌呤四聚體結構應用在相關疾病的治療有所幫助。
然而,在實驗的過程中實驗控制組與文獻出現形成構形的比例不一致的情況,
為了找出實驗條件中的差異分別針對(1)酵素型除氧系統的穩定性 (2)寡核苷酸
上標記螢光染料分子的核苷酸修飾方式 (3)增加實驗的空間解析度進行改善。
在單分子螢光共振能量轉移實驗中所使用的除氧系統為葡萄糖氧化酶與過
氧化氫酶系統,可移除實驗中氧分子的存在與減緩光漂白的現象能有效的增加螢
光穩定性與提升時間解析度。但是,葡萄糖氧化酶與過氧化氫酶的反應生成物葡
萄糖酸,會造成實驗環境變複雜而影響實驗結果,藉由改變酵素型除氧系統希望
能找到適合的實驗條件。此外,單分子螢光實驗所使用的寡核苷酸,依實驗需求
標記有螢光染料分子在特定位置的核苷酸上可能會造成實驗結果穩定在特定鳥
嘌呤四股結構,但是對於不同構形之間轉換的動力學實驗結果並無顯著影響。另
外,單分子螢光實驗中常用的 DNA 架構中由互補的雙股 DNA 作為基底可能會
影響人類端粒序列在摺疊成鳥嘌呤四股結構時傾向形成特定的構形而影響實驗
結果,對此我們在寡核苷酸序列中額外加入不會參與摺疊的胸腺嘧啶,以增加人
類端粒序列可以進行摺疊的空間,同時也改善了螢光共振能量轉移的空間解析度。
綜合以上可能會影響實驗結果的因素分別進行討論,希望能對利用單分子螢光共
振能量轉移的研究提供最接近真實的生理條件且穩定的實驗環境。
關鍵字:端粒,鳥嘌呤四股結構,單分子螢光光譜,全內反射顯微鏡

v
Abstract
G-quagruplexes, secondary structures formed by guanine-rich DNA sequences in
the telomere regions of genome, are highly relevant to cancer due to the inhibition of
telomerase, an enzyme that has high activity in tumor cells and causes the proliferation
of tumor cell. Recent studies revealed that monovalent metal cations such as potassium,
sodium, and ammonium ions stabilize the G-quadruplex structure of in parallel,
antiparallel, and their hybrid configurations. In physiological conditions, the abundance
of magnesium ion is second to the concentration of potassium ion and it participates in
at least 300 biochemical reactions in cell. Previous studies have shown that the addition
of magnesium ions to the sodium or potassium solutions does further stabilize certain
G-quadruplex. Here, we examined the G-quadruplex formation and stabilization in the
presence of magnesium ions only.
Single-molecule fluorescence resonance energy transfer spectroscopy(smFRET)
was employed to study the transformation between G-quadruplex conformational states
and their unfolded counterpart. Surprisingly, our experimental data shows
inconsistency with the results in the literatures. In order to address this problem, we
varied different experimental conditions such as labeling chemistry and ingredient of
oxygen scavenging system solution. We found that the oxygen scavenging systems,
frequently used in smFRET experiment for stabilizing the fluorescent dye molecules,
and the chemistry of fluorescent dye molecule linking to the oligonucleotides sequence,
collaboratively alter the equilibrium between conformational states, hence, form a
stable dye-facilitated folded state in certain conditions, especially in the solely presence
of magnesium cations. However, the kinetics analysis by excluding the molecules in
this specific state showed that the interconversion of G-quadruplex conformational
states remain unchanged throughout the conditions we have tested.

vi
Finally, we add extra poly-thymine, which does not participate in the formation of
G-quadruplexes, to the sequence of interest to increase the distance between the
labeling dye pairs and ultimately improve the spatial resolution. We found that two
distinct states originated from the previously assigned antiparallel configuration and a
fast direct interconversion between these two states is also observed.
Keyword :Telomere,G-quadruplex,Single-molecule FRET,Total internal reflection
fluorescence microscopy

vii
圖目錄
Figure 1 線性染色體末端的複製缺陷 30。 ............................................... 40
Figure 2 哺乳類端粒結構以及與端粒結合的蛋白質複合物 2。 ........... 41
Figure 3 端粒序列中所形成的 T-loop 結構 2。 ....................................... 41
Figure 4 端粒結合蛋白質複合物。 .......................................................... 42
Figure 5 端粒酶延長端粒的機制。 .......................................................... 42
Figure 6 同源重組機制。 .......................................................................... 42
Figure 7 端粒酶在不同的細胞有著不同的活性導致端粒延長或縮短的
現象。.................................................................................................. 43
Figure 8 85~90%癌細胞中過表達的端粒酶活性的治療策略 5。 .......... 43
Figure 9 鳥嘌呤四股結構的組成以及不同金屬陽離子在結構中進行穩
定形成不同的構形。.......................................................................... 44
Figure 10 鳥嘌呤四股結構的分類。 ........................................................ 44
Figure 11 不同鳥嘌呤四股結構構形中有著不同種類的 loop以及不同數
量的鳥嘌呤異構物。.......................................................................... 45
Figure 12 螢光染料分子對的激發與放射光譜。J(λ)為供體放射與受體激
發光譜重疊部分 21。 .......................................................................... 45
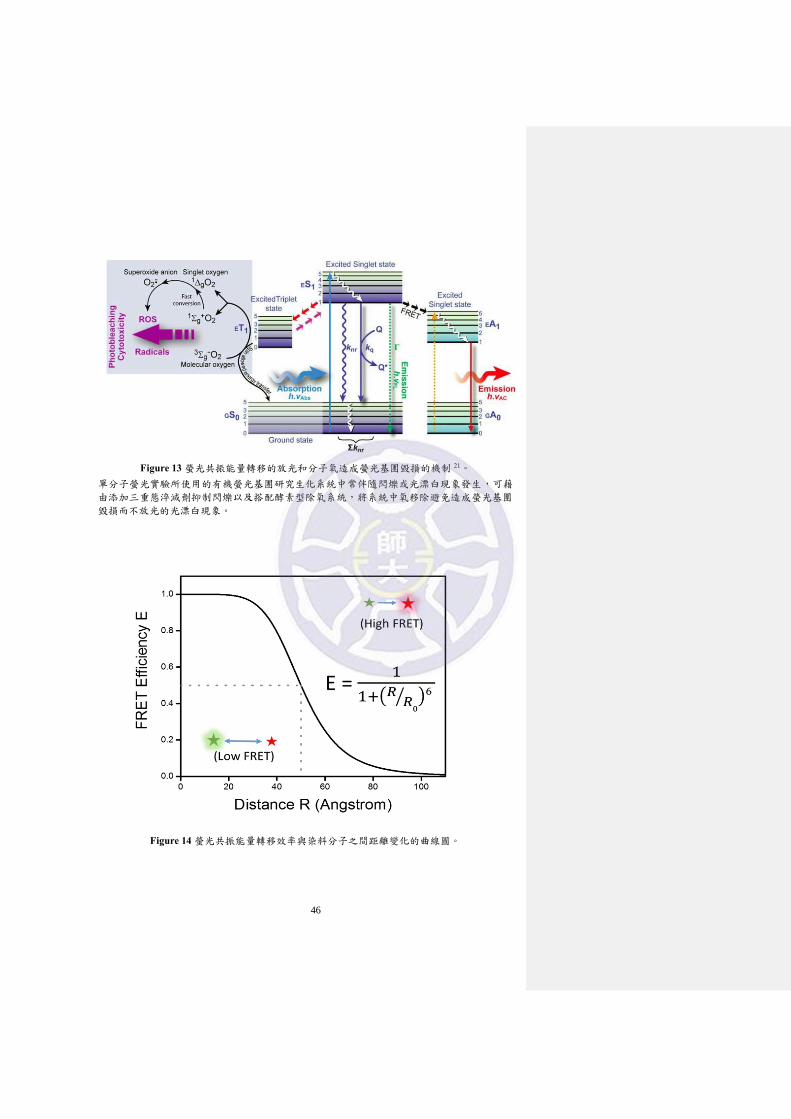
Figure 13 螢光共振能量轉移的放光和分子氧造成螢光基團毀損的機制
21。 ....................................................................................................... 46
Figure 14 螢光共振能量轉移效率與染料分子之間距離變化的曲線圖。
.............................................................................................................. 46
Figure 15 實驗數據分析與處理。 ............................................................ 47
Figure 16 不同的核苷酸修飾方式並與螢光染料分子供體 Cyanine3(Cy3)
的鍵結 31。 .......................................................................................... 48
Figure 17 石英玻片與蓋玻片組裝樣品槽之步驟(a)黏上切割好的雙面膠

viii
帶(b)放上蓋玻片(c)以環氧樹脂將縫隙填。 .................................... 49
Figure 18 石英玻片經 mPEG-SVA 和 biotin-PEG 修飾與固定 DNA 樣品
於玻片表面示意圖。.......................................................................... 49
Figure 19 PTIR(Prism-type Total Internal Reflection )稜鏡式全內反射顯微
術示意圖。.......................................................................................... 50
Figure 20 實驗光學零件架設示意圖。 ................................................... 50
Figure 21 不同的酵素型除氧系統其 pH 隨時間的變化 20。 .................. 51
Figure 22 酵素型除氧系統的反應機制(a)GOC(b)PCD(c)POC 系統 20。
.............................................................................................................. 51
Figure 23 螢光共振能量轉移效率直方圖。 ............................................ 52
Figure 24 螢光共振能量轉移效率直方圖。 ............................................ 52
Figure 25 文獻中人類端粒序列的螢光共振能量轉移直方圖與其對應之
構形以及圓二色光譜 29。 .................................................................. 53
Figure 26 圓二色光譜圖。 ........................................................................ 53
Figure 27 螢光共振能量轉移效率直方圖。 ............................................ 54
Figure 28 螢光共振能量轉移效率直方圖。 ............................................ 54
Figure 29 T20 分子在 PCD 除氧系統下隨鎂離子濃度增加產生訊號漂移
的現象。.............................................................................................. 55
Figure 30 HT21 分子在 PCD 除氧系統鉀離子濃度 50 mM 條件下錄製
120 秒的螢光共振能量轉移效率-時間軌跡圖中螢光共振能量轉移
效率穩定在特定狀態軌跡圖。.......................................................... 56
Figure 31 HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能量
轉移效率未有明顯變化的分子。...................................................... 56
Figure 32 A_HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能
量轉移效率未有明顯變化的分子。.................................................. 57

ix
Figure 33 N_ HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能
量轉移效率未有明顯變化的分子。.................................................. 58
Figure 34 將錄製的時間軌跡圖分類為(a)Dynamic 和(b)Static 在繪製成
FRET 直方圖 ....................................................................................... 59
Figure 35 螢光共振能量轉移效率直方圖,T4_HT21 序列在 PCD 除氧系
統中分別為不同濃度的鉀離子與鎂離子條件下所得到的實驗數據。
.............................................................................................................. 60
Figure 36 比較 HT21 和 T4_HT21 不同訊號下的面積比例。 ............. 60
Figure 37 T4_ HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能
量轉移效率未有明顯變化的分子。.................................................. 61
Figure 38 T4_HT21 分子的時間軌跡圖中快速轉變的訊號以及經互相關
函數分析的結果。.............................................................................. 61
FigureA 1 螢光共振能量轉移效率-時間軌跡圖。HT21 序列在原兒茶酸
與兒茶酸-3,4-雙加氧酶除氧系統中鉀離子濃度為 2mM、5mM、
10mM、20mM 和 50mM 條件下螢光共振能量轉移效率有變動的軌
跡圖。.................................................................................................. 71
FigureA 2 螢光共振能量轉移效率-時間軌跡圖。A_HT21 序列在原兒茶
酸與兒茶酸-3,4-雙加氧酶除氧系統中鉀離子濃度為 2mM、5mM、
10mM、20mM 和 50mM 條件下螢光共振能量轉移效率有變動的軌
跡圖。.................................................................................................. 72
FigureA 3 螢光共振能量轉移效率-時間軌跡圖。N_HT21 序列在原兒茶
酸與兒茶酸-3,4-雙加氧酶除氧系統中鉀離子濃度為 2mM、5mM、
10mM、20mM 和 50mM 條件下螢光共振能量轉移效率有變動的軌
跡圖。.................................................................................................. 73

x
FigureA 4 螢光共振能量轉移效率-時間軌跡圖。T4_HT21 序列在原兒茶
酸與兒茶酸-3,4-雙加氧酶除氧系統中鉀離子濃度為 2mM、5mM、
10mM、20mM 和 50mM 條件下螢光共振能量轉移效率有變動的軌
跡圖。.................................................................................................. 74
FigureA 5 HT21、A_HT21 和 N_HT21 在不同鉀離子濃度下其動力學反
應 速 率 在 未 摺 疊 狀 態 (U) 與 F1(EFRET~0.6) 和 未 摺 疊 狀 態
(U)F2(EFRET~0.8)之間的曲線圖。 ...................................................... 75
FigureA 6 HT21、A_HT21 和 N_HT21 在不同鉀離子濃度下其動力學反
應速率在 F1(EFRET~0.6)與 F2(EFRET~0.8)之間的曲線圖。 ............... 75
FigureA 7 螢光共振能量轉移效率直方圖, A_HT21、N_ HT21 和
T4_HT21 序列在葡萄糖氧化酶-過氧化氫酶除氧系統中分別為不同
濃度的鉀離子與鎂離子條件下所得到的實驗數據。...................... 76
FigureA 8 螢光共振能量轉移效率直方圖,HT21、A_HT21、N_ HT21
和T4_HT21序列在葡萄糖氧化酶-過氧化氫酶除氧系統中分別為不
同濃度的鈉離子條件下所得到的實驗數據。.................................. 77
FigureA 9 螢光共振能量轉移效率直方圖,HT21 序列在吡喃糖氧化酶-
過氧化氫酶除氧系統中分別為不同濃度的鉀離子與鎂離子條件下
所得到的實驗數據。.......................................................................... 77

xi
表目錄
Table 1 實驗所使用的藥品、儲存濃度以及最終濃度。 ........................ 62
Table 2 寡核苷酸進行黏合反應所使用的流程。 ................................... 63
Table 3 圓二色光譜之鳥嘌呤四股結構的特徵訊號位置。 ................... 63
Table 4 實驗所使用的單股寡核苷酸序列。 ............................................ 63
Table 5 HT21 不同鉀離子濃度條件長時間軌跡圖中狀態之間轉變的次
數與動力學反應速率。...................................................................... 64
Table 6 A_HT21 不同鉀離子濃度條件長時間軌跡圖中狀態之間轉變的
次數與動力學反應速率。.................................................................. 64
Table 7 N_HT21 不同鉀離子濃度條件長時間軌跡圖中狀態之間轉變的
次數與動力學反應速率。.................................................................. 65
Table 8 HT21 分子在不同鉀離子濃度下,實驗所觀察到的分子構形有改
變與構形無改變穩定存在特定狀態的比例分布。.......................... 65
Table 9 A_ HT21 分子在不同鉀離子濃度下,實驗所觀察到的分子構形
有改變與構形無改變穩定存在特定狀態的比例分布。.................. 66
Table 10 N_ HT21 分子在不同鉀離子濃度下,實驗所觀察到的分子構形
有改變與構形無改變穩定存在特定狀態的比例分布。.................. 66

12
第一章 緒論
1-1 端粒
1-1.1 端粒的發現
1930 年代,Hermann Muller 利用 X-ray 打斷果蠅染色體末端特殊結構,發現
染色體末端結構非常不穩定。因此, Hermann Muller 和 Barbara McClintock 更進
一步研究這段結構發現此特殊的結構可以保護 DNA,但是如何保護 DNA 的機
制與其基因序列卻還是無從得知。端粒一詞是由 Hermann Muller 所命名,在希
臘字裡,telo-為末端的意思而 mero 為單位。直到 1953 年,James D. Watson 和
Francis H. C. Crick 提出 DNA 為雙股雙螺旋結構與 Arthur Kornberg 發現了 DNA
複製的機制,染色體末端的特殊結構才又受到重視 1。
真核生物的染色體是由許多條線性的 DNA 纏繞而成,在細胞分裂的過程中,
染色體和 DNA 也會進行複製,複製的過程 DNA 複製酶必須遵守一定的方向(5’
至 3’)移動,雙股 DNA 分別形成連續複製的領先股及不連續複製的延滯股(又稱
岡崎片段 Okazaki fragments),延滯股的複製需要 RNA 引子引導 DNA 複製酶進
行複製,使不連續的複製最後使用連接酶將 DNA 片段接合形成連續的 DNA,但
是在延滯股末端最後一個 RNA 引子被移除後,複製 DNA 的酵素無法進行複製
而在 5’端留下一段空缺,進行一次細胞分裂會減少 50~200 個核酸序列,經過不
斷的細胞複製,染色體末端也會隨複製次數縮短,此一現象稱之為「線性 DNA
複製缺陷」(Figure 1)。但是,從一個受精卵變成一個完整的個體需要經過無數次
的細胞分裂,在細胞分裂的過程中有特別的機制與線性 DNA 複製的缺陷抗衡 1,2,3。
西元 1975 年,DNA 定序法的發明,科學家可以知道構成基因密碼的序列,
但受限於當時的 DNA 重組技術還不成熟,無法從動物細胞中分離純化端粒並取
得其基因序列。但是,Joseph Gall 等人從原生生物具有大量線性 DNA 的四膜蟲
細胞內純化並取得作為研究的 DNA 序列。美國科學家 Elizabeth H. Blackburn 等

13
人用 DNA 定序法對四膜蟲染色體末端 DNA 進行研究,發現其 DNA 序列由
TTGGGG 這六個鹼基重複 20~70 次所組成,但是沒有證據證明這段重複序列就
是端粒以及保護 DNA 完整性的功能。在相近的時空背景下,Jack W. Szostak 等
人在完成博士學位的期間利用酵母菌研究 DNA 重組的機制,畢業後他到了癌症
治療中心建立自己的研究室,延續之前的研究,他發現在酵母菌中注入線性的
DNA,線性的 DNA 會快速地降解或重組,與之前 Elizabeth H. Blackburn 等人研
究結果不同,他假設 TTGGGG 或許能保護在染色體末端線性的 DNA,並與
Elizabeth H. Blackburn 合作將親源關係很遠的四膜蟲端粒 DNA 序列注入酵母菌
中,觀察四膜蟲端粒是否能保護酵母菌的 DNA 免於降解或重組,甚至幫助酵母
菌 DNA 進行複製,這項結果成功地證明了先前的假設是正確的。他們將酵母菌
染色體進行純化後,發現酵母菌染色體末端 DNA 重複序列(TG)1-3接在先前注入
的四膜蟲端粒重複序列之後,這項實驗結果顯示酵母菌可以用本身端粒延長的機
制延長在外加的四膜蟲端粒序列上,對於真核細胞中懸而未解的端粒機制也有新
的開端。在 1980 年代,少數真核細胞端粒序列被 Elizabeth H. Blackburn 等人測
定出,並且在端粒序列發現有大量的鳥嘌呤存在。Jack W. Szostak 等人也觀察到
細胞內具有某種機制能將端粒重複序列加在染色體末端,但切確的機制仍然不清
楚。直到端粒酶在生物體內被 Carol W. Greider 等人發現,並且進一步研究得到
端粒酶是以 RNA 為模板合成 DNA 的反轉錄酶,對細胞生物學之後的研究有很
大的貢獻 1。
1-1.2 人類端粒序列
隨著生物技術的進步,科學家發現真核生物中,從原生生物、高等植物到哺
乳動物的端粒結構由一段雙股 DNA 以及一段在 3’端的單股 DNA 所組成,這是
一段富含著鳥嘌呤的 DNA,以真核生物中哺乳動物為例,其端粒序列為
(TTAGGG)n,不同的哺乳動物端粒的長度也有所不同,例如人類的端粒長度大約
10~15 千鹼基對,在單股 DNA 的部分由 150~200 核苷酸所組成 3(Figure 2)。

14
由於染色體重排、降解與融合會形成不穩定的結構,影響 DNA 複製的準確
性以及細胞的存活,端粒則是在染色體末端保護染色體避免發生上述的現象;此
外,在進行細胞有絲分裂的過程,端粒的長度會隨著分裂的次數而縮短,當端粒
長度短到無法維持染色體穩定性時,將會啟動修復機制或自我凋亡機制,因此,
端粒可作為細胞的生理鐘用以判斷細胞的壽命。
端粒重複因子 1(Telomeric repeat-binding factor, TRF1)和端粒重複因子
2(TRF2)與端粒的雙股直接結合負責調控端粒長度與功能,POT1(Protection Of
Telomeres 1)與 3’端單股端粒重複序列以及 T-loop 或 D-loop 結合,TRF1、TRF2
和 POT1 還會與其他三種蛋白形成複合物這六種蛋白複合物(Shelterin) ,在端粒
上會保護染色體末端避免被誤認為損壞的 DNA,而進行修復反應以及調控端粒
酶以維持端粒長度 2,3(Figure 2)。這些蛋白的功能缺失也可能會造成與年齡相關遺
傳疾病。除了這些端粒結合蛋白,端粒 3’端單股重複序列會與端粒雙股形成一種
特殊的環狀結構,T-loop (Figure 3)。T-loop 為染色體末端與生俱有抑制端粒酶與
端粒 3’端羥基結合的結構,當染色體末端長度未達臨界值時,可形成不同大小的
T-loop,將 3’端羥基藏匿在其結構中使端粒酶無法與端粒結合,T-loop 的大小與
功能似乎沒有太大的關係,但是,端粒長度不足以形成此環狀結構時,3’端的羥
基將裸露在外,端粒酶即可延長此染色體端粒序列造成在不同器官中,細胞可進
行分裂的次數有所不同;也可能讓端粒失去保護,意指端粒重複序列變短或失去
與端粒結合之蛋白而導致染色體末端發生融合的現象和失去細胞的可行性。端粒
酶則是避免端粒的長度短到臨界值以維持細胞可行性 3。
近期研究指出,端粒的長度與癌症的病發以及細胞老化等相關疾病有密切的
關係。在大部分正常體細胞和成人幹細胞進行細胞分裂染色體和 DNA 複製的過
程中,端粒酶在這些細胞中受到嚴密的調控,因此端粒的長度會隨細胞分裂的次
數增加而減短,當端粒的長度短到達一個臨界值或無法保護染色體末端的時候,

15
細胞將走向老化進行細胞凋亡的程序。但是在生殖細胞或癌細胞中端粒酶過表達
現象,使得在端粒酶的組成中有一段可作為反轉錄成端粒 DNA 序列的 RNA 模
板,特別是 90%的癌細胞可利用此 RNA 模板不斷地複製延長其端粒基因序列導
致癌細胞不會凋亡甚至增生擴散(Figure 5)。如何抑制端粒酶的活性已被廣泛的研
究,針對端粒酶的抑制藥物已被視為具有潛力的抗癌藥物,許多科學家也致力於
研究端粒與端粒酶相關的機制,希望對人類疾病的治療有所貢獻。
1-1.3 端粒與抗癌藥物的發展
與正常細胞相比,在癌細胞中,端粒酶展現不同的活性,可視為治療癌症的
標靶 4。在癌症初期的細胞中,85~90%的癌細胞受到端粒酶的調控而不影響周圍
正常體細胞,這是一項證據可以將端粒酶當作初期抗癌的目標,並發展抑制端粒
酶活性的治療藥物進一步抑制癌細胞的增生。另外 10~15%的癌症則是無法藉由
抑制端粒酶的活性來治療,其機制為利用具有相似序列的染色體上互補的 DNA
序列,進行端粒 DNA 的複製以延長端粒基因序列,此過程不牽涉端粒酶,稱端
粒延長替代機制(Alternative Lengthening of Telomeres,ALT,Figure 6 and Figure 8)2,5。
了解癌細胞端粒延長的機制,可以更深入的發展癌症的治療方法。現今主要
治療癌症的方式分成兩大類: (1)抑制端粒酶的活性,使端粒無法被延長 (2)抑制
端粒結合蛋白的活性,使癌細胞端粒失去保護,染色體發生融合的現象進而啟動
細胞老化或凋亡機制。前者可以在端粒酶基因進行轉錄、轉錄後修飾等過程利用
改變端粒酶前驅物影響端粒酶失去其功能而達到抑制癌症的效果。還可以利用特
定酵素將端粒酶中的 RNA 片段化,使得癌細胞增生的速度變緩慢。具有活性的
端粒酶需要 Hsp90 複合物(Chaperone complex,伴侶蛋白複合物)與端粒酶結合以
穩定端粒酶的結構處在活化位,因此可抑制此蛋白複合物的生成,同時能抑制端
粒酶的活性。除了上述這方法還有從基因組抑制端粒酶基因的表達以及免疫治療
等方法。利用端粒序列本身富含著鳥糞嘌呤的特性,可以在端粒的 3’端單股形成
鳥嘌呤四聚體的的二級結構,此結構在癌細胞染色體端粒可以抑制端粒結合蛋白

16
(TRF2 和 POT1)的作用,使得端粒結構被破壞,鳥嘌呤四聚體的形成也可以直接
抑制端粒酶作用在端粒上,因此研究在何種生理條件會形成此二級結構以及如何
穩定其結構對於發展對抗癌症的藥物有很大的幫助 5。
1-2 鳥嘌呤四聚體(G-Quadruplex)
1-2.1 鳥嘌呤四聚體概述
鳥嘌呤四聚體,由一段特殊且富含鳥嘌呤鹼基的 DNA 或 RNA 重複序列摺
疊所形成。西元 1910 年,Bang 首先發表了高濃度鳥苷酸衍生物(鳥嘌呤)可自組
裝形成膠狀結構的現象,這項研究結果可推測富含鳥苷酸衍生物(鳥嘌呤)的 DNA
序列可能可以形成更高階的 DNA 構形 6。1953 年,右手雙股雙螺旋 B-form DNA
被發表,相關研究不斷的進行中提供了生物學更多有用的資訊,除了典型的 DNA,
非典型 DNA 結構的研究也受到科學家的重視。許多體外實驗的證據都指出這類
非典型的 DNA 序列在生理條件下可以形成特殊的結構,科學家推測在細胞內它
們可能扮演著重要的功能。
1962 年,Gallert 與他的同事利用 X 射線繞射方法證明鳥苷酸衍生物(鳥嘌呤)
可形成一種四聚體的方形平面結構(G-quartet),利用與相鄰鳥嘌呤形成氫鍵穩定
其構形。這些平面結構推疊則形成鳥嘌呤四聚體(G-quadruplex)7。隨後,Sen 和
Gilbert 等學者發現某些單價陽離子(K+、Na+、NH4+ 等…)在平面和平面之間或平
面中心中和鳥嘌呤第六號位上的氧所產生的靜電斥力穩定其構形 8。在體外實驗
的生理條件下,鳥嘌呤四股結構擁有相當不錯的熱力學穩定性,被認為在細胞內
可能涉及了細胞複製和基因調控等生化過程 6。
近幾年強大的生物模擬工具提供了體外的生物資訊分析結果,依據在人體基
因組具有潛力形成鳥嘌呤四聚體的序列模型([G≧3NXG≧3NXG≧3]4,N為任意鹼基),
利用電腦模擬系統分析的結果得到高達 40,000 條的 DNA 序列存在,這些序列並
非隨機分布在基因組中,而是在特定的基因組大量出現 9。(1)在致癌基因啟動子

17
區域附近有許多能形成鳥嘌呤四聚體的 DNA 序列 10,在此形成的鳥嘌呤四聚體
可透過在上、下游調控基因轉錄起始位,(2)染色體末端端粒基因序列也是具有潛
力形成鳥嘌呤四聚體的 DNA 序列,特別是在端粒結構 3’端有一端單股的 DNA
存在,相較於啟動子附近雙股 DNA 的結構能更容易且更有機會形成鳥嘌呤四聚
體。除此之外,在不同生物中也能在轉錄因子結合位附近、有絲分裂與減數分裂
中雙股斷裂位置、核醣體 RNA 以及富含鳥嘌呤的 DNA 重複序列中找到鳥嘌呤
四聚體的存在 6,11。
關於鳥嘌呤四聚體的研究大多都是在體外實驗條件下進行,雖然用電腦模擬
分析的結果顯示可能可以形成此結構的序列非常多,現今的生物技術只有
Shankar Balsubramanian 團隊利用特殊的抗體在細胞內染色體中能與端粒所形成
的鳥嘌呤四聚體結合並放出螢光 12。2001 年,Schaffizel et.al 在體內實驗發現一
種由核醣體生成的抗體(Sty49)能與原生生物 Stylonychia lemnae 纖毛蟲端粒所形
成的鳥嘌呤四聚體結合並顯現出來 13。Paeschke et.al 使用相同的抗體(Sty49)在纖
毛蟲 DNA 進行複製的時候加入細胞內,發現端粒結合蛋白(TEBPα 和 TEBPβ)會
共同操縱鳥嘌呤四聚體的形成與構形。除此之外,免疫螢光與基因敲落(gene
knockdown)分析顯示這兩種端粒結合蛋白和端粒酶全酵素以及解旋酶(RecQ)會
在細胞週期中合成期的後期限制鳥嘌呤四聚體的形成 6。鳥嘌呤四聚體已被證實
在人體細胞中染色體末端端粒被發現 12,許多研究結果也指出此結構與許多細胞
的生化過程息息相關,除了能調控基因表現外,還會影響細胞分裂的次數以及其
他生化過程,這些體外實驗的結果都顯示了鳥嘌呤四聚體對於治療基因相關的疾
病以及抑制癌細胞的增生都有很大的潛力。
1-2.2 鳥嘌呤四聚體的結構與研究
鳥嘌呤四聚體有著構形上的多樣性,其組成由四個鳥嘌呤利用 Hoogsteen 氫
鍵形成環狀方型平面,G-quartet。再以兩個以上的 G-quartets 平面推疊形成 DNA
二級結構,鳥嘌呤四聚體(G-quardruplex),此 DNA 二級結構為右手螺旋結構,G-

18
quartet 之間的層距約 3.13~3.4 埃(Angstrom),結構扭轉的角度約 30 度、直徑 25
埃(Angstrom),以及磷酸股價所產生的四個溝槽 14(Figure 9)。
G-quadruplex 的分類方式如 Figure 10和 Figure 11可依據(1)寡核苷酸的數量(單
體(Monomer)、雙體(Dimer)、四聚體(Tetramer)),(2)寡核苷酸股的方向(平行、反
平行、混和型),(3)鹼基與五碳糖鍵結的角度χ(Syn、Anti),(4)Loop 的尺寸與串
聯鳥嘌呤的方式(Diagonal 對角、Lateral 階梯、Prepeller 螺旋槳),其中以 Loop 的
長度對最終的結構以及穩定性影響最大,當 Loop 長度太短時會限制 DNA 序列
只能藉由螺旋槳型 Loop 形成平行的結構:但是,當 Loop 長度夠長則傾向形成穩
定的反平行結構。
除此之外,環境中的陽離子種類也會影響鳥嘌呤四聚體的構形,先前的核磁
共振整體實驗研究結果得到,當環境中有鈉離子存在時,端粒 DNA 重複序列
(dAGGG(TTAGGG)3)會受到鈉離子幫助穩定在反平行的鳥嘌呤四聚體結構,在鉀
離子條件下則是得到混和型的鳥嘌呤四聚體 8。由圓二色光譜也得到相同的結果,
平行的結構在 264nm 有最大值的吸收峰,240nm 則有最小值的吸收峰;反平行
和混和型結構則是在 295nm 有最大吸收峰,265nm 有最小吸收峰(Table 3)。除此
之外,還可以用紫外光吸收光譜、電噴灑離子化質譜、單分子技術等實驗方法來
研究鳥嘌呤四聚體的結構、特性與動力學相關生化性質 14。
1-2.3 端粒上的鳥嘌呤四聚體
在人類染色體末端負責保護染色體,避免發生降解和融合現象以維持其結構
完整的端粒,也被視為 DNA 雙股斷裂的辨識位,其重複序列中富含鳥嘌呤核苷
酸,是具有潛力形成鳥嘌呤四股結構的序列之一 15。在細胞分裂過程中,DNA 複
製酶無法完整複製端粒末端 DNA 序列,在複雜的生化系統中有另一種酵素-端粒
酶,可用自身結構中的 RNA 模板複製端粒末端序列,彌補 DNA 複製酶的功能
缺失。在正常體細胞中端粒酶的活性不高,只有在幹細胞、生殖細胞以及癌細胞

19
具有顯著的活性導致癌症的病發。若能在癌細胞中端粒序列上形成鳥嘌呤四股結
構,可能可以抑制癌細胞的不正常增生,進一步使癌細胞自行老化進入細胞凋亡
的程序達到治療癌症疾病的效果 16(Figure 7)。此外,DNA 複製的過程中端粒的雙
股 DNA 先被解旋酶打開形成單股結構,此 DNA 序列富含鳥嘌呤,在此有機會
形成鳥嘌呤四股結構影響 DNA 複製的過程,以及之後的生化過程。先前的文獻
指出在 DNA 複製的過程中出現鳥嘌呤四聚體除了影響其複製的速度,甚至會造
成基因組的不穩定,對於正常體細胞進行 DNA 複製的過程雙股端粒序列,若形
成鳥嘌呤四聚體需要將此結構打開才能繼續進行複製的程序,打開此結構所需要
的解旋酶由之前體外實驗研究得知,除了大腸桿菌純化出 RecBCD 解旋酶無法
將其結構打開,剩下的 21 種解旋酶都能打開鳥嘌呤四聚體結構,這說明了此結
構對解旋酶並無特別的專一性。但是這些實驗都只是定性的實驗,鳥嘌呤四聚體
是否能在人體內被解旋酶打開,仍然需要由更多的實驗證明,還有更重要的一點,
實驗中能解開鳥嘌呤四聚體的解旋酶都與基因組不穩定所造成的遺傳疾病有關
係(BLM、WRN、RecQ、FANCj and PIFI 解旋酶)17。以 Fanconi anemia(FANCj)病
患為例,由於基因組中生成 FANCj 解旋酶的基因缺陷,導致此解旋酶失去能夠
打開鳥嘌呤四聚體以及其他相關 DNA 的功能,骨髓失去功能造血細胞無法分化
形成紅血球,而造成了先天性的貧血症狀,以及白血球與血小板的數目減少有凝
血功能的障礙,甚至增加罹患癌症的機率 18。除了端粒具有有潛力形成鳥嘌呤四
聚體的序列,在基因組啟動子附近也有相當多能形成其結構基因序列,在此處相
關的基因序列也與特定的遺傳疾病有關,在 DNA 轉錄轉譯後所形成蛋白質產物
失去了原本的功能導致疾病的發生,在此若能形成鳥嘌呤四聚體則可能減少不正
常的蛋白質產物的生成達到抑制疾病惡化的效果 10。了解鳥嘌呤四聚體在人體形
成的機制,與如何調控基因表現對於相關遺傳疾病的治療,與降低罹患癌症的風
險有很大的幫助。近期的研究除了發展能穩定鳥嘌呤四聚體結構的小分子作為抗
癌藥物,希望能得到更多在人體發現此結構的證據。

20
1-3 研究動機
先前的研究指出金屬陽離子對鳥嘌呤四聚體具有穩定其結構的效果,在生理
條件下,鉀離子和納離子分別能穩定鳥嘌呤四股結構存在平行、混和型以及反平
行構形也經由整體實驗和單分子實驗得到證實。在人體細胞中,鎂離子的含量僅
次於鉀離子而且參與 300 多種生化反應,相較於鉀離子(133pm),鎂離子(65pm)
有著較小的離子半徑能更容易的進入結構中以及帶有二價的正電可能可以更加
穩定鳥嘌呤四聚體結構;先前的文獻也指出,同時存在鉀離子和鎂離子的條件下
可以更加穩定鳥嘌呤四股結構 19。因此我們想要知道單純鎂離子是否能幫助人類
端粒序列形成特定的鳥嘌呤四聚體或穩定其結構,對於已被證實存在人體的鳥嘌
呤四聚體結構應用在相關疾病的治療有所幫助。
在實驗的過程中出現了實驗控制組與文獻在形成構形的比例不一致的情況,
為了找出實驗條件中的差異分別針對(1)酵素型除氧系統的穩定性(2)寡核苷酸上
標記螢光染料分子的核苷酸修飾方式(3)增加實驗的空間解析度 進行改善。首先,
單分子螢光實驗中所研究的分子皆標記有螢光染料,實驗所收集的訊號解析度受
到螢光染料分子的穩定性影響,因此實驗過程中會添加除去氧分子的除氧試劑,
避免在實驗過程中螢光訊號不穩定,一般常用的除氧試劑為葡萄糖氧化酶-過氧
化氫酶(簡稱 GOC),此系統因反應生成物為葡萄糖酸容易造成 pH 劇烈地下降,
參考先前發表的文獻還有另外兩種除氧系統分別為原兒茶酸-兒茶酸-3,4-雙加氧
酶系統(簡稱 PCD)及吡喃糖氧化酶-過氧化氫酶系統(簡稱 POC)具有較穩定的 pH,
能提供對 pH 靈敏的實驗系統更長的觀測時間 20。寡核苷酸上標記螢光染料分子
的核苷酸修飾的方式不同,可能會影響實驗所觀察的生化分子在形成特定構形時
得到正向或者負向的幫助。綜合以上在實驗中可能造成分子行為偏離真實情況的
因素,希望能提供穩定的實驗系統改善時間解析度以及找到最接近真實分子行為
的實驗條件對使用單分子螢光方法的相關研究有所幫助。

21
第二章 實驗架構與方法
2-1 單分子技術與發展
生物體中複雜的生化系統有非常多的蛋白質、核酸、去氧核酸等生化分子參
與酵素反應或訊號途徑。由於細胞內環境複雜,生化分子在細胞內可能存在著多
種構形且與特定大分子交互作用。為了深入了解基本的細胞生化程序,現今已有
高解析度的核磁共振光譜、X 射線-結晶學等技術可廣泛的分析生化分子的結構
以及巨觀的動力學實驗結果;而因為生物分子的個體變異性與生化反應的非同步
性,必須有更新穎的技術突破傳統整體實驗(Ensemble experiment)靜態分子的視
野及巨觀實驗的平均結果。隨著進步的光學影像及生物力學技術,提供了研究單
一分子構形及構形變化的實驗方法。
單分子技術尚未出現之前,大部分的實驗方式以整體實驗為主,整體實驗為
一次觀測整個系統中發生的各種分子行為,實驗得到的數據若無法區分系統中的
異質性,將導致實驗結果為多種狀態的平均結果,此外,整體實驗中所使用的樣
品濃度遠大於單分子實驗,在高濃度的分子環境中可能形成雙體分子或其他聚合
物,測得的實驗結果也並非真實分子的行為。加上在細胞中的生化反應皆發生地
非常快速,在整體實驗中無法定義系統中不同分子的起始點,只能取得反應達平
衡時的動力學數據;在單分子實驗中所使用的樣品濃度極低(約 10-12莫耳),因此
可以將實驗系統中的分子是為獨立個體進行觀測,得到多個單一分子行為的實驗
數據再進行分析處理,因此數據的可信度提升,此外,單分子實驗中可以定義分
子的反應起點,得到的動力學數據相較整體實驗更高。
單分子技術常與螢光或機械力結合以改善實驗中時間與空間的解析度。從增
加空間解析度的觀點切入,複雜生化系統由多個不連續且隨機(stochastic)發生的
化學反應組成,巨觀整體實驗結果只能得到多個化學反應的平均能量,但在單分
子實驗中可直接觀察到該分子正在進行的反應以及精確的空間位向。此外,在超

22
高解析螢光顯微鏡中利用單分子技術調控螢光染料分子隨機放光的特性可突破
光學繞射極限,累積 10000 個光子被偵測的情況下其精確度可達 1-2 奈米。另外
針對時間解析度的提升,在單分子螢光共振能量轉移實驗,受限於光子的通量,
當激發光的強度過強容易產生具反應性的電子三重態造成光漂白及閃爍的現象
(Figure 13),因此必須降低激發光強度並延長曝光時間,以維持可以接受的訊雜比
而限制了時間解析度,所以在此實驗過程添加的顯影緩衝溶液,包含三重態淬滅
劑,水溶性維生素 E(Trolox)以及除氧系統試劑,以增加時間解析度同時並可錄製
長時間的動力學變化軌跡圖,時間與空間解析度的改善對於研究複雜生化系統提
供了新的視野,偵測單一分子的動態行為再結合數據分析軟體可得到更真實的分
子動力學行為。
單分子技術除了與機械力或螢光結合,甚至可以結合電或電腦理論計算程式,
提供各種層面的實驗資訊,成為研究複雜生化系統的強大工具。過去的十五年單
分子技術日漸成熟也被應用在生物化學以及生物醫學等領域,對於探討生物體內
生化過程與相關疾病關聯性有極大的貢獻。
2-2 螢光共振能量轉移(Förster resonance energy transfer)
螢光共振能量轉移的現象是透過供體螢光染料分子(Donor)受到特定波長的
光激發後,因供體的放射波長與受體(Acceptor)的激發波長有部分重疊,且供體
與受體之間的距離小於 10 奈米,以及螢光染料分子對的方向非垂直方向,即可
觀察到受體受到經由不放光的螢光共振能量轉移得到能量而放出受體放射波長
的光 21。
與先前的實驗方法相比,在整體實驗中很難定義以及偵測反應發生的起點,
單分子螢光共振能量轉移實驗提供單一分子隨時間改變螢光共振能量轉移效率
的軌跡圖,雖然其訊雜比不高但可藉由累積多組實驗數據進行同步分析得到相同
起始位置的實驗結果。在單分子螢光共能量轉移實驗中所使用的蛋白質、核酸等
生化分子皆標記有螢光料分子在特定的位置,藉由偵測螢光放光的強度代入螢光

23
共振能量轉移效率對應距離的公式得到螢光染料分子的距離變化可作為欲研究
的生化分子之動力學行為或可利用螢光料分子標記的位置不同以推測生化分子
的構形。
𝐸𝐹𝑅𝐸𝑇 =𝐼𝐴
𝐼𝐴 + 𝐼𝐷=
1
1 + (𝑅 R0⁄ )
6
式中,𝐸𝐹𝑅𝐸𝑇:實驗觀測到的螢光共振能量轉移效率,𝐼𝐴、𝐼𝐷:受體、供體螢
光染料分子放出的螢光強度,R:受體、供體螢光染料分子之間的距離,𝑅0:螢
光共振能量轉移效率為 50%時對應的螢光染料分子對的距離。螢光共振能量轉移
效率與染料分子之間距離成六次方反比主要是受到偶極-偶極交互作用(Dipole-
Dipole Coupling) 所影響。
此外,R0可以用來定義實驗的有效作用距離範圍約 3-8奈米,其對應 5%-95%
的螢光共振能量轉移效率,能產生訊號變化的染料分子對距離變化在 0.5R0 -
1.5R0,如 Figure 14。但在實務中受限於雜訊的干擾可作用的範圍變小,以常用的
螢光染料分子對 Cyanine3(Cy3)與 Cyanine5(Cy5)為例,如 Figure 12,其 R0約 5.6
奈米,可觀察的作用範圍在 2.8-8.4奈米,可以提供奈米尺度的空間解析度但也
限制了研究蛋白質複合物或大分子蛋白質之間的交互作用22,23
。R0的計算受到供
體放射光譜與受體激發光譜的重疊面積(J(λ)) ,如 Figure 12,和螢光染料分子
的位向(κ2)所影響。其中螢光染料分子的位向κ
2,其範圍介在 0 至 4 之間 ,
在實驗中被假設為標記在分子上的螢光基團可自由旋轉並且旋轉的速率遠大於
螢光放光的速率,因此可將螢光基團視為點光源向四面八方放光,取其位向參數
用統計平均值κ2=2
3,若螢光基團無法自由旋轉,或旋轉速率不夠快,則必須考慮
位向因素 24。螢光有著低背景雜訊的優點,常與單分子技術結合應用在研究生物
系統中生化分子構形與構形變化動力學,有著如此高解析度的設備需要搭配有效
率的數據收集方法以及高靈敏性的偵測器。

24
2-3 全內反射式螢光顯微術(Total internal reflection fluorescence
microscopy,TIRF)
2-3.1 實驗樣品槽的組裝與固定樣品於修飾後的表面
在探討全內反射式螢光顯微術之前,必須先了解單分子實驗中以表面束縛
(surface tethering)的方式,將單分子均勻的分佈並固定於玻片的表面,簡述如下:
將欲研究之生化分子固定在經由聚乙二醇與低濃度標記有生物素-聚乙二醇修飾
過的石英玻片表面上。首先將修飾過後的石英玻片與蓋玻片用雙面膠帶及環氧樹
脂組裝成體積容量約 20uL 的微流通道,組裝流程如 Figure 17,在此空間注入抗
生物素蛋白在控制的時間內(約 2 分鐘)與表面的生物素結合再用緩衝溶液沖除,
接著將修飾有生物素的 DNA 或蛋白質注入,在相同的時間條件下,讓生化分子
上的生物素與表面的抗生物素結合後,再用緩衝溶液沖除多餘的生化分子即完成
試樣固定在石英玻片表面上的準備。(Figure 19(b)、詳細的配方請參照 Table 1)
2-3.2 全內反射原理與螢光顯微鏡
全內反射現象為光由高密度介質進到低密度介質,其入射角θ大於臨界角
θc 時,光不再折射而會形成全內反射的現象,在全內反射發生的介面會有些許
的光波穿透介面進入低密度介質,稱為漸逝場(Evanescence field),漸逝場的強度
會隨著穿透的深度成指數遞減,波長在可見光區其平均穿透深度約為 100 奈米,
由於待測分子均被固定於玻片表面上,因此可有效的消除背景造成的雜訊提升訊
雜比。
全內反射式螢光顯微鏡分為兩種:稜鏡式(Prism type)與物鏡式(Objective
type),稜鏡式的激發雷射由稜鏡折射在石英玻片上進行全反射,產生的螢光再由

25
顯微鏡物鏡所收集,考慮到樣品槽的厚度,須採用長工作距離水浸潤物鏡(Long
working distance water immersion objective)才能得到清晰的影像(Figure 19(a));物
鏡式全內反射式螢光顯微鏡的激發雷射由物鏡邊緣入射,經由大的數值光圈
(1.4NA,Numerical aperture)油浸潤物鏡內鏡片組的折射在蓋玻片上形成全反射。
這兩種全內反射式螢光顯微鏡的螢光訊號皆由物鏡接收,並經由顯微鏡內光學元
件,將影像投射至顯微鏡側面的首要影像平面,可同時偵測螢光共振能量轉移之
供體染料分子與受體染料分子的螢光訊號,在首要影像平面加裝雙視光學元件
(狹縫、透鏡、雙色鏡以及反射鏡),狹縫可裁切影像呈長條形,再用雙色鏡分離
供體染料分子與受體染料分子的螢光訊號,並將訊號透過兩個消色差透鏡傳送至
相機感測器,兩個不同波段的影像可並列且同時投影在相機偵測器表面被觀測
(Figure 20)。
相機偵測器的選擇受到偵測的螢光訊號微弱且具有相當的背景值限制,近年
來快速發展有著較高的讀取速度及價格相對便宜的科學等級互補金屬氧化物半
導體儲存器(sCOMS)相機,但因影像對比不佳無法適用於此類實驗。電子放大式
電荷耦合器(Electron Multiplied Charged Coupled Device ,EMCCD)具有可針對特
定信號強度區間放大的優點改善前者的問題,本實驗架設採用 Andor 公司背照
光(back-illumined)式 iXon Ultra 897 EMCCD,解析度為 512x512 畫素,最高全幅
譜速(frame rate)可達每秒 52 張,並具有高量子效率(Quantum efficiency,在可見
光區可達 85%以上)與高度線性(linearity, >99%)等特性,感光元件裝置有熱電
(thermoelectric)晶片可降溫至-100°C(實際實驗僅降溫至-70°C)有效抑制雜訊。
2-4 數據校正與分析
螢光訊號由電子放大式電荷耦合器接收經訊號放大後傳送至電腦,分別使用
IDL(Exelis Visual Information Solutions)、MATLAB(The Mathworks Inc.)、HaMMy
及 TDP25進行不同的數據分析處理。首先利用 IDL 程序(script)22負責將實驗拍攝
的影片進行疊圖分析並校正光學元件的誤差,排除實驗中兩點距離過近造成的異

26
常光點,將實驗得到的單分子定位,並將影像簡化分別儲存其位置與螢光強度隨
時間變化等資訊。隨後使用 MATLAB 程序,篩選螢光強度排除背景雜訊與多分
子光點之影響後,計算螢光共振能量轉移效率,並修正實驗過程中部份的受體染
料分子螢光訊號源自於供體螢光染料分子螢光通過濾光片所洩漏(Donor leakage)。
在動力學分析上使用 Ha group 所開發的 HaMMy 程式,使用 Hidden Markov 演
算法擬合長時間軌跡圖中不同狀態的變化並得到其動力學數據,再用 TDP 分析
實驗中構形改變的次數及正逆反應速率常數 25。將上述數據分析方法得到的資訊
用 Origin 繪製成直方圖、長時間軌跡圖和構形之間變化的分佈圖。
2-5 酵素型除氧系統與顯影緩衝溶液
2-5.1 螢光放光與去活化機制
單分子螢光共振能量轉移實驗中所使用的有機螢光基團,在研究生化系統的
過程中,常伴隨著閃爍或光漂白的現象,導致實驗所觀察到的訊號不穩定或限制
了觀測的時間。螢光的放光過程中,電子受激發波長的光躍遷至激發單重態再回
到基態,不斷地重複著此循環並伴隨螢光出現,但是,部分處在激發單重態的分
子可經由非放光的系統間跨越(intersystem crossing) 而轉換至激發三重態,而激
發三重態可能經過非放光的系統間跨越,或是伴隨著磷光(phosphorescence)放光
而重新返回基態,此過程的時間尺度為數毫秒到數分鐘,相較於螢光放光所需的
時間大上 104-1011倍,因此單位時間偵測到的光強度相對於螢光非常之弱而可被
忽略,當分子進入三重態時可視為進入暗態(dark state) ,重回基態後被激發至單
重激發態而可重新發出螢光,此一週期循環造成了螢光分子的閃爍,進而干擾實
驗的進行 26(Figure 13)。
基態(三重態)的分子氧是有效的三重態淬滅劑,在淬滅的過程會生成具有反
應性的單重態氧物種導致光漂白(photobleaching)現象發生,光漂白現象為光化學
過程中螢光基團進入激發與放射的循環被永久中斷,起因為螢光基團與環境中生

27
化分子反應或螢光基團毀損導致無法放光,光毀損與光化學過程主要發生在螢光
基團的電子處在生命期長且具有反應性的激發三重態,激發三重態主要與分子氧
反應,基態分子氧可翻轉激發三重態中電子自旋方向使激發三重態較容易回到基
態並伴隨具產生具反應性的單重態氧分子物種,單重態分子氧不僅造成光漂白現
象對細胞具有毒性,因此需要將實驗系統中的氧分子移除 21,27。
2-6 酵素型除氧系統反應機制與作用
酵素型的除氧系統利用反應過程中需要氧的參與,可以移除實驗中氧分子的
存在有效減緩光漂白的現象發生。但是,電子仍有可能從激發單重態經系統間跨
越到激發三重態,或者受到光破壞與光化學過程使電子停留在激發三重態無法放
出螢光,導致實驗過程中觀察到閃爍的現象而影響實驗結果,需要額外添加三重
態淬滅劑,水溶性維生素 E(Trolox)可藉由抑制三重態的出現達到減少閃爍現象
的發生,在單分子螢光共振能量轉移實驗中所使用的顯影試劑通常都會添加水溶
性維生素 E(Trolox) ,與酵素型除氧系統搭配使用可增加螢光穩定性,有效的提
升時間解析度延長實驗錄製軌跡圖的時間。
常用的酵素型除氧系統有添加葡萄糖氧化酶及過氧化氫酶(GOC)與添加原
兒茶酸與兒茶酸 -3,4- 雙加氧酶 (protocatechuic acid 與 protocatechuate-3,4-
dioxygenase,PCD) ,另外還有吡喃糖氧化酶-過氧化氫酶系統(pyranose oxidase
與 catalase,POC)20。在葡萄糖氧化酶與過氧化氫酶的反應需加入少量的葡萄糖,
反應的生成物葡萄糖酸會造成酸鹼值快速下降,影響到對 pH值敏感的實驗系統,
顯影試劑中的緩衝溶液可延緩酸鹼值快速變化。PCD 系統的除氧效率較前者快
速約 3 倍,反應過程中雖然也有酸生成,但是酸鹼值的變化也較前者緩慢,可提
升實驗系統中螢光染料分子的穩定性,我們所使用的三種酵素型除氧系統顯影試
劑的配方請參見 Table 1,其反應機制 Figure 22。一般向廠商訂購的原兒茶酸與兒
茶酸-3,4-雙加氧酶都有純度不夠高的問題,其不純物中包含有核酸酶(nucleoase),
對於研究核酸分子的實驗會造成核酸被分解的問題,本實驗室所使用的原兒茶酸

28
與兒茶酸-3,4-雙加氧酶皆經過純化處理 20,28。
2-7 石英玻片表面修飾
將石英玻片及蓋玻片依序使用丙酮、3M 氫氧化鉀以及甲醇浸泡並放入超音
波震盪儀清洗,中間過程使用超純水沖洗。用丁烷火焰以高溫再將表面有機物質
移除(蓋玻片以電漿清洗機清洗五分鐘) ,待其冷卻後浸泡在含有甲醇、醋酸、以
及 N-(2-Aminoethyl)-3-aminopropyltrimethoxysilane 的混和溶液中進行鈍化反應,
10 分鐘後以超音波震盪一分鐘,再繼續反應十分鐘,反應結束後依序使用甲醇
與超純水沖洗,再用氮氣將其吹乾,並移至底部含水以保持濕度的容器中。配製
以 100mM 碳酸氫鈉溶解 mPEG-SVA(分子量 5000,Laysan Bio,Inc)和 Biotin-
PEG(分子量 5000,Laysan Bio,Inc)後放入高速離心機,取上清液滴在玻片表面再
用蓋玻片小心地蓋上(避免氣泡產生),靜置於暗處 3 至 4 小時。最後用超純水充
分沖洗並以氮氣吹乾、抽真空後密封放入-80℃冰箱保存。
2-8 螢光染料分子的標記方法
本篇實驗所使用的寡核苷酸序列皆為人類端粒重複序列 dG3(T2AG3)3,依據標記
螢光染料位置的核苷酸修飾方式不同可分為三類,第一種為 NH2C3,在核苷酸的
磷酸根修飾碳鏈長度為三的碳的連接體(Linker)在接上螢光染料供體分子,第二
類 NH2C6 與第一類的差別在於碳鏈長度延伸為六個碳,第三類則是對核苷酸上
的胸腺嘧啶鹼基進行修改,在胸腺嘧啶上延伸同為六個碳長度的連接體再接上螢
光染料分子,如 Figure 16 所示,另外還有在寡核苷酸前後加入額外的胸腺嘧啶,
目的是為了讓鳥嘌呤四股結構在形成的時候不受到雙股的立體障礙限制,以及增
加空間解析度。Cy3 標記在寡核苷酸的步驟如下,將向廠商訂購的 2O.D 單股寡
核苷酸序列用 100mM 的硼氰化鈉 35µL 稀釋再加入 0.2 毫克 Cy3 粉末中混和均
勻,並架設在旋轉儀器上 24 小時(25℃),使 Cy3 充分結合在寡核苷酸上。加入
99.5%的乙醇及 3 M 的氯化鈉放入-80℃冰箱 30 分鐘,再放入離心機以 11200 轉

29
/分鐘、溫度控制在 4℃離心 30 分鐘後吸取上清液,再重複前一步驟一次。最後
用氮氣吹乾,產率以八成估算,再將標記有螢光染料的寡核苷酸稀釋到濃度 100
µM。
2-9 寡核苷酸黏合反應
標記有 Cy5 的互補股(取 4 µL) ,與標記有 Cy3 的寡核苷酸(取 1 µL) ,以
及不外加鹽類的 Tris緩衝溶液(pH7.8)(取 15 µL)稀釋至 20 µL,以乾浴器加熱
至 89 度五分鐘後,與加熱鋁塊一併移至暗處靜置四小時回到室溫,或是以聚合
酶連鎖反應(Polymerase chain reaction,PCR)機器依 Table 2 程序進行黏合反應,
形成類似端粒重複序列由一部分雙股與欲研究之單股序列所組成的單分子實驗
DNA 構造,最後將黏合好的 5 µM 雙股 DNA 再稀釋到 250 nM 及 10 nM 放入-20
℃冰箱保存。
2-10 圓二色光譜(Circular dichroism spectroscopy)
2-10.1 圓二色性(Circular dichroism)
利用具有掌性對稱中心的分子對於左旋與右旋的吸收度不同,在圓二色光譜
儀器中,利用 1/4 玻片讓線性偏振光形成左旋與右旋圓偏振光,因待測分子的掌
性結構影響左旋與右旋圓偏振光的吸收度不同,使訊號在重合後產生橢圓偏振光,
即圓二色性。
2-10.2 鳥嘌呤四股結構的區別
核苷酸為構形多樣性的分子,在生物體內可能以單股、雙股、三股結構甚至
能用富含多個鳥苷酸的序列折疊成四股結構,而圓二色光譜對核苷酸構形的變化
非常靈敏,常用來研究 DNA 和 RNA 的結構。
鳥嘌呤四聚體由平行與反平行結構組成,在圓二色光譜中藉由偵測 DNA 或
RNA 序列鳥苷酸糖苷鍵鍵角(χ)的不同會形成不同的結構,比較 G-quartet 平面中
鳥苷酸異構物(Syn/Anti)數目的不同會造成訊號峰值的不同。圓二色光譜中分子

30
內的平行結構在波長 260 奈米位置有最大值,在波長 240 奈米則有最小值;反平
行結構在波長 295 奈米有最大值,在波長為 265 奈米則有最小值,所有鳥嘌呤四
股結構在波長 210 奈米皆為正值,此訊號為可與 A form 雙股 DNA 作區分。圓二
色光譜也可以提供鳥嘌呤四股結構的動力學實驗數據,可以觀察在不同離子條件
下構形的變化,此優點為核磁共振光譜及 X 射線結晶學無法達到的,但還是能
結合不同的實驗方法互相驗證其實驗結果的準確性。

31
第三章 實驗結果與討論
3-1 螢光共振能量轉移效率直方圖(FRET efficiency Histogram )
由單分子螢光共振能量轉移的實驗結果繪製的的直方圖中,觀察到欲研究的
10 pM 人類端粒重複序列 (Table 4 中有完整序列) ,在使用葡萄糖氧化酶除氧系
統初始 pH 為 7.5~7.8,分別存在有不同濃度的鉀離子和鎂離子的條件下,如 Figure
23,隨著離子的濃度增加,螢光共振能量轉移的訊號由 Un-folded state(UF,
EFRET~0.3)逐漸轉變成另外兩個較高螢光共振能量轉移效率的訊號 (Folded
states:F1,EFRET~0.6 與 F2,EFRET~0.8),以鉀離子條件作為控制組用來與先前研究
的文獻做對比,此條件下得到的訊號位置與文獻相似,在濃度為 100 mM 時
UF(EFRET~0.3)完全轉變成 F1 (EFRET~0.6)和 F2 (EFRET~0.8)其對應結構為平行與反
平形構形;在鎂離子濃度為 3 mM 時,幾乎已完全轉變成較高螢光共振能量轉移
的訊號,隨著鎂離子濃度增加,F1 (EFRET~0.6)訊號也逐漸變成 F2 (EFRET~0.8)訊號
位直到濃度增加到 50 mM 只剩 F2 (EFRET~0.8)訊號。從實驗結果初步得到鎂離子
幫助 iHT21 序列摺疊相較鉀離子更有效率,用已知的鉀離子實驗結果推測在中
等濃度的鎂離子條件形成兩種鳥嘌呤四聚體結構分別為平行與反平行或混和型
與反平行構形,而在高濃度鎂離子條件則是傾向形成反平行的構形。但是,鎂離
子條件下所形成的結構仍然必須要用其他實驗方法才能得到證明。
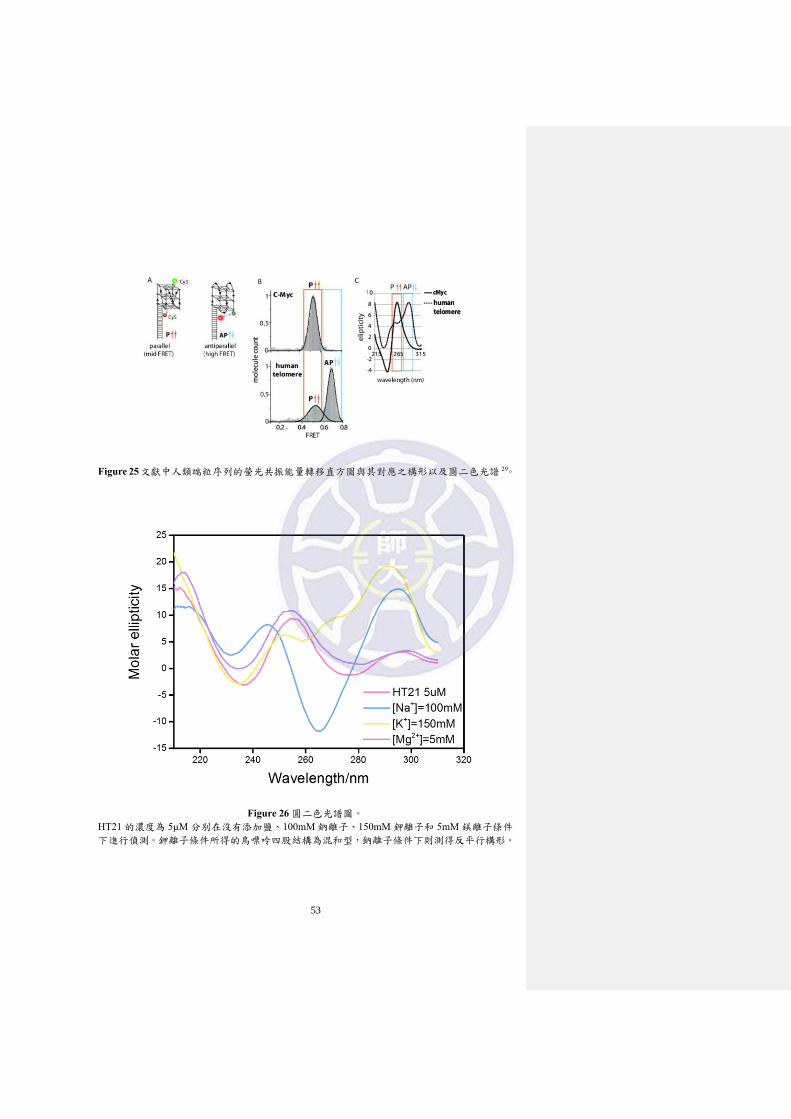
3-2 圓二色光譜
圓二色光譜偵測的 DNA 序列同為人類端粒序列 HT21(5’-G3(T2AG3)3-3’),溫
度 20℃,HT21 濃度為 5 µM 分別在鉀離子濃度 100 mM、鈉離子濃度 150 mM 以
及鎂離子濃度 5 mM 條件下進行,圓二色光譜的背景校正使用與相對應濃度條件
下含有相同的緩衝溶液(pH=7.6)以及金屬離子條件的溶液。實驗結果如 Figure 26
顯示,鉀離子與鈉離子條件結與文獻有相同的訊號位置分別為混和型與反平行的

32
鳥嘌呤四股結構,然而,鎂離子條件卻沒有鳥嘌呤四聚體構形訊號存在也無明顯
的訊號變化,顯現與單分子螢光共振能量轉移所觀察到的實驗結果不一致。
在螢光共振能量轉移觀察到的兩種訊號有可能是(1)相較於圓二色光譜在偵
測期間 pH 無明顯改變,GOC 除氧系統 pH 值急遽下降造成 DNA 序列受 pH 下
降影響實驗環境變得複雜而摺疊成其他二級結構,我們使用另外兩種除氧系統
(PCD 與 POC)改善上述問題。(2)在單分子實驗中所使用的寡核苷酸皆標記有螢
光染料分子,螢光染料分子可能會幫助寡核苷酸摺疊或穩定在特定結構。解決此
問題的方法為改變標記螢光染料位置的核苷酸修飾方式,分別有改變螢光染料分
子連接在寡核苷酸上的位置以及增加連接的長度。
3-3 酵素型除氧系統的改變
常用的酵素型除氧系統在反應過程中有羧酸的生成,造成實驗中離子強度增
加以及 pH 大幅下降影響實驗結果。為了減少羧酸生成造成的影響,將實驗中所
使用的除氧系統置換成 PCD 及 POC,先前的文獻提出不同除氧系統在反應過程
中 pH 隨時間的改變,其中,在起始 pH 為 7.8 時,GOC 系統在 10 分鐘內下降至
7.6,然而 PCD 與 POC 卻能維持 2 小時才降到 pH7.6,因此,先前鎂離子條件的
單分子實驗結果有兩根訊號出現,可能是受到顯影緩衝溶液造成實驗系統環境變
得複雜所影響。
在 pH 穩定的除氧系統可以使實驗系統環境維持穩定,得到的實驗結果(Figure 24)
觀察到隨著鎂離子濃度增加螢光共振能量轉移效率產生漂移的現象,而非先前實
驗所觀察到的數個狀態之間的消長。由對照組實驗,實驗中所使用的目標序列為
長度 20 個胸腺嘧啶的 DNA 序列(T20),此序列不會摺疊成 DNA 二級結構,因
此可作為模擬 DNA 在不同濃度的鎂離子條件下未摺疊的行為,實驗結果(Figure
34)與 HT21 相似,隨鎂離子濃度增加往高螢光共振能量轉移效率的訊號漂移。因
此我們猜測造成訊號飄移的原因,可能為 DNA 上面的電荷受到鎂離子中和而使
得斥力下降結構更趨緊密,導致螢光共振能量轉移效率隨濃度增加而逐漸提升;

33
在鉀離子條件的實驗結果與 GOC 系統相同,有兩種螢光共振能量轉移效率的訊
號出現。綜合以上結果得到在 GOC 系統中鎂離子條件下得到 EFRET 較高的訊號
主要是二價離子造成的 DNA 斥力下降所造成的,然而 F1(EFRET~0.6)的訊號產生
的原因可能與螢光染料分子幫助穩定 DNA 二級結構有關,從高濃度鉀離子條件
的實驗結果與文獻中 29(Figure 25)的差異發現原本 F2 (EFRET~0.8)應該有較強的訊
號,但在本實驗三種除氧系統(Figure 23、Figure 24 以及 FigureA 9)下卻出現
F1(EFRET~0.6)有較高的訊號強度,合理的猜測螢光染料分子在此幫助了
F1(EFRET~0.6)的構形形成或穩定其構型的存在。為了證實這項假設,針對寡核苷
酸上標記螢光染料分子的核苷酸修飾方法進行改變:分別有增加連接螢光染料分
子的碳鏈長度(NH2C3 和 NH2C6) ,以及將接在核苷酸核苷酸上的螢光染料分子
置換成接在核苷酸的磷酸骨架上,加上(AmC6dT)碳鏈長度為六個碳連接核苷酸
(Figure 16)與螢光染料分子,以增加染料分子與核酸序列間的距離期能降低其交互
作用。
A_HT21 和 N_HT21 在 PCD 除氧系統、鉀離子條件下進行實驗,得到的直
方圖(Figure 32 和 Figure 33)中 F1(EFRET~0.6)和 F2 訊號強度的比例與文獻相符證實
標記螢光染料分子的核苷酸修飾方法不同可能會影響螢光染料分子進而穩定
F1(EFRET~0.6)構形或幫助其構形的形成。為了更深入了解其動力學行為,實驗過
程中錄製了時間與螢光共振能量轉移效率變化的軌跡圖。
3-4 螢光共振能量轉移效率-時間軌跡圖
將實驗得到的螢光共振能量轉移效率-時間軌跡圖進行分類 : (1)在錄製時間內螢
光共振能量轉移效率沒有改變(以 Figure 30 為例) (2)在錄製時間內螢光共振能量
轉移效率有改變(FigureA 1 ~ FigureA 4 );首先,針對不同濃度的鉀離子其沒有改
變的軌跡圖進行分析,得到 HT21 大部分都穩定以 F1(EFRET~0.6)狀態存在(Figure
31)然而 A_HT21 和 N_HT21 則是以 F2 (EFRET~0.8)狀態穩定存在(Figure 32 和
Figure 33),這項實驗結果證明 HT21 中核苷酸修飾的方式不同會影響螢光染料分
子穩定特定的構形。此外,在錄製時間內螢光共振能量轉移效率有改變的軌跡

34
圖進行不同狀態之間的動力學分析,利用隱馬可夫演算法擬合不同狀態之間的
變化,再將 HT21 在不同鉀離子條件下的時間軌跡圖繪製成螢光共振能量轉移
直方圖(Figure 34),在錄製時間內螢光共振能量轉移效率有改變的軌跡圖所繪製
的直方圖與大部分文獻所發表的實驗結果相似,這項實驗結果更加驗證螢光染
料分子會幫助特定訊號的分子穩定形成,但是,這種訊號並非真實分子的行為
並在我們的實驗中形成誤導的訊號。另外我們找出了 UF(EFRET~0.3)和
F1(EFRET~0.6)、UF(EFRET~0.3)和 F2 (EFRET~0.8)以及 F1(EFRET~0.6)和 F2 (EFRET~0.8)
之間的正逆反應速率(FigureA 5 和 FigureA 6)與狀態之間轉換的次數(Table 5、Table
6 和 Table 7),實驗結果得到螢光染料分子標記位置核苷酸修飾方式不同的分子
對於其動力學行為趨向形成特定構形並無顯著的影響,先前在 HT21 的螢光共
振能量轉移效率直方圖得到 F1(EFRET~0.6)構形為主要訊號,造成此差異的原因
主要是受到螢光染料分子穩定其構形並且長時間存在;綜合以上分析結果得到
具有動力學行為的分子與穩定存在的分子彼此為獨立的行為,其實驗結果顯示
不同核苷酸的修飾方法對於不同構形之間的轉換並無顯著的影響可以視為獨立
的事件。
3-5 提升空間解析度的實驗設計
在單分子螢光實驗中常用的 DNA 構造由互補的雙股 DNA 作為基底,其中
一條 DNA 固定在石英玻片表面上,另一條 DNA 則包含欲研究的 DNA 序列。在
此 DNA 構造中的雙股 DNA 基座,可能會影響人類端粒序列在摺疊成鳥嘌呤四
股結構時傾向形成特定的構形而影響實驗結果。為了排除雙股 DNA 基座對 HT21
在進行折疊時造成的立體障礙,我們在目標 DNA(HT21)的前後各別多加上了 4
個和 3 個胸腺嘧啶(Thymine,T),如 Figure 16(c)和 Table 4,T4_HT21,可增加 HT21
折疊時所需的空間,也將螢光共振能量轉移效率往較高空間解析度的範圍調整。
由於螢光共振能量轉移的效率與距離非線性關係,當轉移效率落在邊界值附近時,
對空間的解析度較差,加上分子本身的距離與螢光共振能量轉移效率有相當的分
佈寬度,因此在邊界值可能無法區分距離相近的狀態。為了仔細探究 HT21 序列
中 F2(EFRET~0.8)狀態,我們加上了不會參與摺疊的數個胸腺嘧啶序列,將螢光染

35
料分子之間的距離拉長使得轉移效率下降至 0.5 附近,希望能增進空間解析度。
重複相同條件在 PCD 除氧系統的單分子螢光實驗,從螢光共振能量轉移效
率直方圖(Figure 35)可以明顯的觀察到螢光共振能量轉移效率下降至 0.1、0.3、0.5
和 0.6(UF,EFRET~0.1 為 unfolded state,F1,EFRET~0.3、F21,EFRET~0.5、F22,EFRET~0.6
為 Folded states)。大約在 10 mM 鉀離子濃度時,已無 Un-folded state 存在全都轉
變成 Folded state 而且 F1 (EFRET~0.3)的訊號強度比 F21(EFRET~0.5)與 F22(EFRET~0.6)
的訊號高,與 HT21 實驗結果有著相同的現象。值得注意的是原本在 HT21 序列
中螢光共振能量轉移效率為 F2(EFRET~0.8)的訊號在 T4_HT21 中分裂成
F21(EFRET~0.5)和 F22(EFRET~0.6)的訊號,造成此訊號分裂的原因極有可能是因為空
間的解析度提升,可以由鉀離子濃度為 50 mM 的 HT21 和 T4_HT21 螢光共振能
量轉移效率直方圖中 F1和 F2 訊號下的面積進行擬合,再計算出 HT21 的 F1 和 F2
與 T4_HT21 的 F1 和 F21 加上 F22 所佔的比例,結果得到非常相近的比例(Figure
36)。再用螢光共振能量轉移效率公式反推 T4_HT21,扣除額外加入的胸線嘧啶
所增加的長度後所對應的 FRET 值,T4_HT21 中 F21(EFRET~0.5)與 F22(EFRET~0.6)
的分別對應的距離為 5.6 奈米與 5.2 奈米以及 HT21 中 F2(EFRET~0.8)對應的距離
為 0.44 奈米,估計在 T4_HT21 序列前後額外添加的四個和三個胸腺嘧啶增長了
約 1 奈米的平均距離,依此反推 F21(EFRET~0.5)和 F22(EFRET~0.6)的 FRET 值在HT21
之差異約為 0.05,在半高寬約 0.1 的情況下即可能重疊而無法分辨。讓原本 HT21
中無法分離的訊號在此可以清楚地分辨。針對 T4_HT21 螢光共振能量轉移效率
-時間的軌跡圖進行深入的分析,將在錄製時間內螢光共振能量轉移效率沒有改
變的軌跡圖繪製成螢光共振能量轉移效率直方圖(Figure 37)得到 F1 (EFRET~0.3)的
訊號強度大於 F22(EFRET~0.6)與 HT21 得到的實驗結果相同,然而,HT21 和
T4_HT21其標記螢光染料分子之核苷酸的修飾方式不同卻出現相同的實驗結果,
造成EFRET~0.3(T4_HT21)所對應構形穩定度提升的原因可能是為了消除立體障礙
額外加入的胸腺嘧啶反而增加鳥嘌呤四聚體在形成的過程中或涉及更複雜的環

36
境因素互相牽制所造成。此外 T4_HT21 在先前所探討的鎂離子條件也出現隨著
離子強度增加螢光共振能量轉移效率產生漂移的相同現象。
從螢光共振能量轉移效率-時間的軌跡圖實驗結果顯示 F21(EFRET~0.5)和
F22(EFRET~0.6)之間有快速轉換現象(Figure 38(a)),將這種快速轉換的軌跡圖在鉀離
子濃度為 50 mM 和 200 mM 的條件下進行互相關分析(Cross-correlation ),實驗
結果如(Figure 38(b))利用單一指數遞減進行曲線擬合,得到實驗觀察到的快速變化
訊號具有關聯性,此實驗結果也在 B 細胞淋巴瘤(Bcl2)DNA 序列在生理條件下
進行單分子螢光共振能量轉移實驗中觀察到,此一快速轉換並未經過 UF 狀態,
我們推測在反平行的折疊結構下具有兩種或以上的構型,這些構型間可以快速地
互相轉換,是一動態結構而非一個穩定的靜態結構,此一發現對於致力於穩定鳥
嘌呤四股結構的研究,提供了一項重要的參考。

37
第四章 總結
單分子實驗方法提供空間與時間高解析度的工具可觀察到單一分子的精確
的構形與構形變化的動力學行為,不同於整體實驗(ensemble experiment)容易受
到分子構形異質化(heterogeneous)產生不同訊號,造成實驗結果被平均化以及高
濃度的分子彼此產生交互作用而出現實驗的誤差,在實驗過程中可直接觀測到單
一分子的行為。單分子技術藉由結合螢光共振能量轉移的原理、搭配全內反射顯
微技術、靈敏的偵測系統,以及快速的數據分析軟體經常被使用在研究複雜的生
化系統。
富含鳥嘌呤的人類端粒序列已被眾多研究證明可形成鳥嘌呤四股結構並且
與細胞老化的相關疾病以及癌症有密切的關係。在生理條件下,鳥嘌呤四股結構
可以受到金屬陽離子(K+、Na+和 NH4+等…)的幫助穩定在特定構形,先前的研究
也指出生理條件下含量最多鉀離子對於穩定鳥嘌呤四股結構有非常好的效果而
且在實驗條件中同時存在鉀離子與少量的鎂離子可以更穩定其構形。然而,使用
單分子螢光共振能量轉移的實驗結果發現單獨存在的鎂離子對於幫助鳥嘌呤四
股結構形成的效果微乎其微,訊號的變化主要源自於實驗溶液中離子強度的增加
改變溶液環境造成訊號漂移的現象。但是,在實驗的過程中發現改變顯影緩衝溶
液中的除氧系統、寡核苷酸中標記螢光料分子的核苷酸修飾方式以及為了減少立
體障礙在人類端粒序列前後額外增加胸腺嘧啶對於單分子螢光實驗產生不同層
面的影響。
不同的除氧系統其差異為去除氧分子的反應其產物改變了實驗的溶液環境
和影響螢光染料分子的穩定性。在 GOC 除氧系統中,消耗一莫耳的氧會生成 2
莫耳的葡萄糖酸,導致溶液環境的離子強度明顯增加與 pH 急遽下降,使人類端
粒序列在此系統下鉀離子的實驗數據出現不符合參考文獻的結果。將酵素型除氧
系統改用 pH 穩定的 PCD 以及 POC 系統,其實驗結果與葡萄糖氧化酵素系統相
同。但是,在鎂離子的條件下原本的 GOC 系統出現了兩個螢光共振能量轉移強

38
度訊號卻因為改用 PCD 和 POC 系統產生了訊號漂移的現象,由圓二色光譜與此
實驗結果證實單純的鎂離子條件無法幫助鳥嘌呤四股結構的形成。
為了解決鉀離子條件下 F1(EFRET~0.6)和 F2(EFRET~0.8)形成的比例與文獻不符
合的結果,針對寡核苷酸上標記螢光染料分子的核苷酸修飾方法進行修改,得到
的實驗結果無論在核苷酸的磷酸根還是鹼基上修飾有六個碳的碳鍊其螢光共振
能量轉移效率的直方圖中顯示偏向形成 F2(EFRET~0.8)的構形;然而,修飾有三個
碳的碳鍊在核苷酸的磷酸根則是傾向形成 F1(EFRET~0.6)的構形,證實了標記螢光
染料分子的核苷酸修飾方法會幫助鳥嘌呤四股結構形成特定構形,或是穩定在特
定的構形。由錄製的長時間螢光共振能量轉移效率軌跡圖針對穩定不變的螢光共
振能量轉移效率(EFRET)進行分析並繪製成直方圖也得到同樣結果。此外,對於長
時間螢光共振能量轉移效率軌跡圖 EFRET有變動的軌跡圖繪製成直方圖得到的結
果與文獻相符,另外也進行正、逆反應速率常數的計算,不同的標記螢光染料分
子核苷酸修飾方法從 UF(EFRET~0.3)轉變成 F1(EFRET~0.6)和 F2(EFRET~0.8)個別的
正逆反應速率常數數據對於直方圖中 F1(EFRET~0.6)和 F2(EFRET~0.8)比例分布並無
明顯的影響,實驗結果中不同狀態的比例與文獻不同的原因主要是受到螢光染料
分子穩定特定構形形成非真實分子行為的訊號所造成。
為了排除雙股 DNA 基座對人類端粒序列在進行折疊時造成的立體障礙以及
增加空間解析度,在人類端粒序列的前後額外加入了胸腺嘧啶進行相同條件的重
複實驗。實驗結果顯示原本 HT21 的 F2(EFRET~0.8) ,極可能為兩個狀態的疊合
T4_HT21 的 F21(EFRET~0.5)和 F22(EFRET~0.6),利用將螢光共振能量轉移效率的直
方圖中 F1 (EFRET~0.3)和 F21(EFRET~0.5)加上 F22(EFRET~0.6)曲線下的面積比例與
HT21 的 F1(EFRET~0.6)和 F2(EFRET~0.8) 曲線下的面積比例進行分析得到了近似值。
再由時間-螢光共振能量轉移效率軌跡圖也觀察到 F21(EFRET~0.5)和 F22(EFRET~0.6)
訊號之間快速的變化,證實了原本 HT21 的 F2(EFRET~0.8)是由兩種不穩定的 DNA
二級結構組成。然而,F21(EFRET~0.5)和 F22(EFRET~0.6)訊號所對應的 DNA 構形必

39
須由其他的實驗方法進行確認。此外,T4_HT21 在錄製時間內穩定不變的螢光共
振能量轉移效率的時間-螢光共振能量轉移效率軌跡圖中發現T4_HT21穩定在F1
(EFRET~0.3)狀態,此現象與 HT21 相似。但是,T4_HT21 與 HT21 中標記螢光染
料分子的核苷酸的修飾方法並不相同,實驗趨勢相同的原因可能為額外添加的胸
腺嘧啶與人類端粒序列交互作用,或胸腺嘧啶之間形成新的立體障礙所造成。
單分子技術應用在研究複雜的生化系統行之有年,結合螢光共振能量轉移原
理的單分子實驗必須考量到螢光料分子的穩定性以增加時間解析度、在研究生化
系統則需要嚴格地控制實驗,維持在相關的生理條件以及欲研究的生化分子經由
化學修飾後或實驗環境所造成的影響是否改變其本質等…。在本篇論文中,由單
純的金屬陽離子條件下,研究人類端粒序列受金屬陽離子的幫助形成鳥嘌呤四股
結構的實驗中,分別針對不同的酵素型除氧系統形成不同的溶液環境、不同的核
苷酸修飾方法用以標記螢光染料分子,以及添加額外的鹼基對實驗結果的影響進
行探討。實驗結果可能同時受到多種因素的改變而有所不同,不能單純的只考慮
單一因素,必須視欲研究的生化分子找到其適合的實驗環境與實驗條件進行調整,
達到最接近真實的生理環境提供精確的實驗結果。人類端粒序列也已被證實可作
為在癌症疾病的標靶,開發穩定人類端粒序列所形成的鳥嘌呤四股結構的小分子
也正蓬勃發展。但是,最根本的問題:人類端粒序列摺疊成鳥嘌呤四股結構的機
制仍然需要使用更穩定且更接近真實狀況的實驗條件進行深入的探討。單分子技
術除了與螢光原理結合,還可以與機械力、電或理論計算提供不同的視野對同一
分子的研究能有更精確的構形剖析及動力學數據以應用在複雜環境的生化系統。

40
圖
Figure 1 線性染色體末端的複製缺陷 30。
DNA 在進行複製的過程中,在延滯股需要 RNA 引子引導 DNA 聚合酶至 DNA 模板上進行
不連續複製,在複製的最後將 DNA 模板 5’端上的 RNA 引子移除,導致複製 DNA 的長度比
DNA 模板短。

41
Figure 2 哺乳類端粒結構以及與端粒結合的蛋白質複合物 2。
端粒重複序列由 10-15 千鹼基對的雙股與 150-200 核苷酸的單股所組成的。負責保護染色體末
端避免發生降解、融合等形成不穩定的染色體影響 DNA 複製準確性以及細胞的存亡。
Figure 3 端粒序列中所形成的 T-loop 結構 2。
多種蛋白質複合物與端粒結合可以穩定 T-loop 結構將 3’端上羥基藏匿在 T-loop 結構中避免端
粒酶結合而進行延長端粒序列。

42
Figure 4 端粒結合蛋白質複合物。
負責保護端粒末端避免被降解或融合以維持端粒完整性以及調控端粒的長度。其中端粒調控
因子 1 與端粒調控因子 2 與端粒的雙股部分結合,端粒保護因子則與端粒單股部分結合。
Figure 5 端粒酶延長端粒的機制。
端粒酶包含一段可合成端粒序列的 RNA 反轉錄酶,藉由此反轉錄酶可在癌細胞中增加端粒長
度導致癌細胞不會凋亡進而使癌症疾病病發。
Figure 6 同源重組機制。
利用具有相同序列之不同染色體上的互補序列進行 DNA 複製的行為常發生在修復損壞的
DNA。在癌細胞中可以延長端粒長度,在延長的過程不牽涉端粒酶,為少部分癌症病發的原
因。

43
Figure 7 端粒酶在不同的細胞有著不同的活性導致端粒延長或縮短的現象。
在體細胞中,端粒酶的活性受到嚴謹的調控,導致在正常體細胞中端粒長度隨細胞次數增加而
減少。在腫瘤細胞中,由於端粒酶過表達端粒可無限制增生,導致腫瘤細胞不會進入凋亡程序
而永生。若在腫瘤細胞的端粒序列利用端粒本身富含鳥嘌呤的特性可形成 DNA 二級結構,G-
quadruplex,則有機會可以抑制端粒酶與腫瘤細胞的端粒結合達到抑制癌症的效果。
Figure 8 85~90%癌細胞中過表達的端粒酶活性的治療策略 5。
第一類為抑制端粒結合蛋白複合物的活性,可以使癌細胞中端粒結構失去保護,隨細胞分裂次
數增加而自行啟動細胞凋亡程序。第二類為抑制端粒酶活性,除了直接一端粒酶活性外,還可
以針對端粒酶的前驅物進行抑制使生成的端粒酶失去延長端粒的功能達到抑制癌症的效果。

44
Figure 9 鳥嘌呤四股結構的組成以及不同金屬陽離子在結構中進行穩定形成不同的構形。
(a)四個鳥嘌呤藉由 Hoogsteen 氫鍵形成環形平面(G-quartet),再由兩個以上的 G-quartet 推疊
形成鳥嘌呤四股結構(b)人類端粒基因序列受到鈉離子或鉀離子穩定形成的不同構形的鳥嘌呤
四聚體,在鈉離子條件下形成反平行的構形,鉀離子條件下則形成混和形構形。
Figure 10 鳥嘌呤四股結構的分類。
(1)依據形成結構所需寡核苷酸股的數目可分成單體、雙體以及四聚體(2)結構中寡核苷酸股的
方向可分為平行、反平行以及混和型構形。

45
Figure 11 不同鳥嘌呤四股結構構形中有著不同種類的 loop 以及不同數量的鳥嘌呤異構物。
以混和型 2 號鳥嘌呤結構為例,(a)串聯 G-quartet 的 Loop 種類有對角型、螺旋槳型和階梯型
(b) 不同構形的鳥嘌呤四股結構中有的不同數量的鳥嘌呤異構物(Syn 和 Anti)。
Figure 12 螢光染料分子對的激發與放射光譜。J(λ)為供體放射與受體激發光譜重疊部分 21。
單分子螢光共振能量轉移的螢光染料分子對中 Cy3 的放射波長與 Cy5 的激發波長在光譜必須
有部分重疊,重疊面積為 J(λ)。
46
Figure 13 螢光共振能量轉移的放光和分子氧造成螢光基團毀損的機制 21。
單分子螢光實驗所使用的有機螢光基團研究生化系統中常伴隨閃爍或光漂白現象發生,可藉
由添加三重態淬滅劑抑制閃爍以及搭配酵素型除氧系統,將系統中氧移除避免造成螢光基團
毀損而不放光的光漂白現象。
Figure 14 螢光共振能量轉移效率與染料分子之間距離變化的曲線圖。

47
Figure 15 實驗數據分析與處理。
(a)由 EMCCD 以每幀 30 毫秒的速度拍攝的影像傳送至電腦,分別有 30 幀和 4000 幀。將影像經
由 IDL 程序進行儀器校正與疊圖分析,得到個別分子的位置與螢光強度訊號。(c)再用 MATLAB
程式進行螢光訊號強度比較並修正實驗誤差。(b)多個分子的 30 幀影像繪製的螢光共振能量轉移
效率直方圖,利用螢光共振能量轉移效率不同可計算螢光染料分子對的距離,用以判斷分子是否
有構形上的變化。(c)單一分子錄製兩分鐘的影像,可觀察到分子的構型變化動力學行為,並可進
一步分析得到動力學資訊。

48
Figure 16 不同的核苷酸修飾方式並與螢光染料分子供體 Cyanine3(Cy3)的鍵結 31。

49
Figure 17 石英玻片與蓋玻片組裝樣品槽之步驟(a)黏上切割好的雙面膠帶(b)放上蓋玻片(c)以
環氧樹脂將縫隙填。
Figure 18 石英玻片經 mPEG-SVA 和 biotin-PEG 修飾與固定 DNA 樣品於玻片表面示意圖。

50
Figure 19 PTIR(Prism-type Total Internal Reflection )稜鏡式全內反射顯微術示意圖。
Figure 20 實驗光學零件架設示意圖。
(a)532nm 綠光激發雷射光源(LASOS DPSSL serise)和 640nm 紅光激發雷射光源(Omicron LuxX
638-40),PBS:讓特定一波長光穿透與另一波長光反射。 (b)PTIR(Prism-type Total Internal
Reflection )稜鏡式全內反射(C)Home-dual view 雙視光學零件組。

51
Figure 21 不同的酵素型除氧系統其 pH 隨時間的變化 20。
Figure 22 酵素型除氧系統的反應機制(a)GOC(b)PCD(c)POC 系統 20。

52
Figure 23 螢光共振能量轉移效率直方圖。
HT21 序列在 GOC 除氧系統中分別為不同濃度的鉀離子與鎂離子條件下所得到的實驗數據。
Figure 24 螢光共振能量轉移效率直方圖。
HT21 序列在 PCD 除氧系統中分別為不同濃度的鉀離子與鎂離子條件下所得到的實驗數據。
53
Figure 25文獻中人類端粒序列的螢光共振能量轉移直方圖與其對應之構形以及圓二色光譜 29。
Figure 26 圓二色光譜圖。
HT21 的濃度為 5µM 分別在沒有添加鹽、100mM 鈉離子、150mM 鉀離子和 5mM 鎂離子條件
下進行偵測。鉀離子條件所得的鳥嘌呤四股結構為混和型,鈉離子條件下則測得反平行構形。

54
Figure 27 螢光共振能量轉移效率直方圖。
A_HT21 序列在 PCD 除氧系統中分別為不同濃度的鉀離子與鎂離子條件下所得到的實驗數
據。
Figure 28 螢光共振能量轉移效率直方圖。
N_HT21序列在PCD除氧系統中分別為不同濃度的鉀離子與鎂離子條件下所得到的實驗數據。

55
Figure 29 T20 分子在 PCD 除氧系統下隨鎂離子濃度增加產生訊號漂移的現象。

56
Figure 30 HT21 分子在 PCD 除氧系統鉀離子濃度 50 mM 條件下錄製 120 秒的螢光共振能量
轉移效率-時間軌跡圖中螢光共振能量轉移效率穩定在特定狀態軌跡圖。
Figure 31 HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能量轉移效率未有明顯變化
的分子。

57
Figure 32 A_HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能量轉移效率未有明顯變
化的分子。

58
Figure 33 N_ HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能量轉移效率未有明顯變
化的分子。

59
Figure 34 將錄製的時間軌跡圖分類為(a)Dynamic 和(b)Static 在繪製成 FRET 直方圖

60
Figure 35 螢光共振能量轉移效率直方圖,T4_HT21 序列在 PCD 除氧系統中分別為不同濃度
的鉀離子與鎂離子條件下所得到的實驗數據。
Figure 36 比較 HT21 和 T4_HT21 不同訊號下的面積比例。
計算在 PCD 系統鉀離子濃度 50 mM 條件下 HT21 的 F1 和 F2 與 T4_HT21 的 F1、F21 和 F22 經
高斯曲線擬合後,再計算 HT21 的 F1 和 F2 與 T4_HT21 的 F1、F21 加上 F22 曲面下面積所佔的
比例。

61
Figure 37 T4_ HT21 對不同鉀離子濃度條件下在觀察時間內螢光共振能量轉移效率未有明顯
變化的分子。
Figure 38 T4_HT21 分子的時間軌跡圖中快速轉變的訊號以及經互相關函數分析的結果。
已註解 [IL1]: 請把 50mM 的換成深一點的顏色

62
表格
Table 1 實驗所使用的藥品、儲存濃度以及最終濃度。

63
Table 2 寡核苷酸進行黏合反應所使用的流程。
Table 3 圓二色光譜之鳥嘌呤四股結構的特徵訊號位置。
Table 4 實驗所使用的單股寡核苷酸序列。

64
Table 5 HT21 不同鉀離子濃度條件長時間軌跡圖中狀態之間轉變的次數與動力學反應速率。
Table 6 A_HT21 不同鉀離子濃度條件長時間軌跡圖中狀態之間轉變的次數與動力學反應速
率。

65
Table 7 N_HT21 不同鉀離子濃度條件長時間軌跡圖中狀態之間轉變的次數與動力學反應速
率。
Table 8 HT21 分子在不同鉀離子濃度下,實驗所觀察到的分子構形有改變與構形無改變穩定
存在特定狀態的比例分布。

66
Table 9 A_ HT21 分子在不同鉀離子濃度下,實驗所觀察到的分子構形有改變與構形無改變穩
定存在特定狀態的比例分布。
Table 10 N_ HT21 分子在不同鉀離子濃度下,實驗所觀察到的分子構形有改變與構形無改變
穩定存在特定狀態的比例分布。

67
參考文獻
1. Blackburn, E. H. Telomeres and telomerase: the means to the end (Nobel
lecture). Angew Chem Int Ed Engl 49, 7405–7421 (2010).
2. Blasco, M. Telomeres and human disease: ageing, cancer and beyond.
Nat. Rev. Genet. 6, 611–22 (2005).
3. Palm, W. & de Lange, T. How shelterin protects mammalian telomeres.
Annu. Rev. Genet. 42, 301–34 (2008).
4. Shay, J. W. & Bacchetti, S. A survey of telomerase activity in human
cancer. Eur. J. Cancer 33, 787–791 (1997).
5. Sekaran, V., Soares, J. & Jarstfer, M. B. Telomere maintenance as a
target for drug discovery. J Med Chem 57, 521–538 (2014).
6. Bochman, M. L., Paeschke, K. & Zakian, V. A. DNA secondary
structures: stability and function of G-quadruplex structures. Nat. Rev. Genet. 13,
770–80 (2012).
7. Gellert, M., Lipsett, M. N. & Davies, D. R. Helix Formation By
Guanylic Acid. Proc. Natl. Acad. Sci. 48, 2013–2018 (1962).
8. Williamson, J. R., Raghuraman, M. K. & Cech, T. R. Monovalent
cation-induced structure of telomeric DNA: The G-quartet model. Cell 59, 871–
880 (1989).
9. Capra, J. A., Paeschke, K., Singh, M. & Zakian, V. A. G-quadruplex
DNA sequences are evolutionarily conserved and associated with distinct
genomic features in Saccharomyces cerevisiae. PLoS Comput. Biol. 6, 9 (2010).
10. Huppert, J. L. & Balasubramanian, S. G-quadruplexes in promoters
throughout the human genome. Nucleic Acids Res. 35, 406–413 (2007).
11. Bidzinska, J., Cimino-Reale, G., Zaffaroni, N. & Folini, M. G-

68
quadruplex structures in the human genome as novel therapeutic targets.
Molecules 18, 12368–12395 (2013).
12. Biffi, G., Tannahill, D., McCafferty, J. & Balasubramanian, S.
Quantitative visualization of DNA G-quadruplex structures in human cells. Nat.
Chem. 5, 182–6 (2013).
13. Schaffitzel, C. et al. In vitro generated antibodies specific for telomeric
guanine-quadruplex DNA react with Stylonychia lemnae macronuclei. Proc.
Natl. Acad. Sci. U. S. A. 98, 8572–7 (2001).
14. Małgowska, M., Gudanis, D., Teubert, A., Dominiak, G. & Gdaniec, Z.
How to study G-quadruplex structures. Biotechnologia 93, 381–390 (2013).
15. Sundquist, W. I. & Klug, A. Telomeric DNA dimerizes by formation of
guanine tetrads between hairpin loops. Nature 342, 825–829 (1989).
16. Zahler, A. M., Williamson, J. R., Cech, T. R. & Prescott, D. M.
Inhibition of telomerase by G-quartet DNA structures. Nature 350, 718–720
(1991).
17. Mohaghegh, P., Karow, J. K., Brosh, R. M., Bohr, V. A. & Hickson, I.
D. The Bloom’s and Werner's syndrome proteins are DNA structure-specific
helicases. Nucleic Acids Res. 29, 2843–9 (2001).
18. London, T. B. C. et al. FANCJ is a structure-specific DNA helicase
associated with the maintenance of genomic G/C tracts. J. Biol. Chem. 283,
36132–36139 (2008).
19. Yan, Y. Y. et al. Selective recognition of oncogene promoter G-
quadruplexes by Mg 2+. Biochem. Biophys. Res. Commun. 402, 614–618 (2010).
20. Swoboda, M. et al. Enzymatic oxygen scavenging for photostability
without ph drop in single-molecule experiments. ACS Nano 6, 6364–6369 (2012).

69
21. Ishikawa-Ankerhold, H. C., Ankerhold, R. & Drummen, G. P. C.
Advanced fluorescence microscopy techniques-FRAP, FLIP, FLAP, FRET and
FLIM. Molecules 17, 4047–4132 (2012).
22. Rahul, R., Hohng, S. & Ha, T. A Practical Guide to Single Molecule
FRET. Nat. Methods 5, 507–516 (2008).
23. Ha, T. Single-molecule fluorescence resonance energy transfer.
Methods 25, 78–86 (2001).
24. Sisamakis, E., Valeri, A., Kalinin, S., Rothwell, P. J. & Seidel, C. A. M.
Accurate Single-Molecule FRET Studies Using Multiparameter Fluorescence
Detection. Methods in Enzymology 475, (Elsevier Inc., 2010).
25. Mckinney, S. A., Joo, C. & Ha, T. Analysis of Single-Molecule FRET
Trajectories Using Hidden Markov Modeling. 91, 1941–1951 (2006).
26. Grewer, C. & Brauer, H.-D. Mechanism of the Triplet-State Quenching
by Molecular Oxygen in Solution. J. Phys. Chem 98, 4230–4235 (1994).
27. Grewer, C. & Brauer, H.-D. Mechanism of the Triplet-State Quenching
by Molecular Oxygen in Solution. J. Phys. Chem 98, 4230–4235 (1994).
28. Aitken, C. E., Marshall, R. A. & Puglisi, J. D. An oxygen scavenging
system for improvement of dye stability in single-molecule fluorescence
experiments. Biophys. J. 94, 1826–1835 (2008).
29. Tippana, R., Xiao, W. & Myong, S. G-quadruplex conformation and
dynamics are determined by loop length and sequence. Nucleic Acids Res. 42,
8106–8114 (2014).
30. Greider, C., Blackburn, E., Muller, H. & Mcclintock, B. 3 1980 2009
1982. 50–55 (2010).
31. Lee, W., Von Hippel, P. H. & Marcus, A. H. Internally labeled Cy3/Cy5

70
DNA constructs show greatly enhanced photo-stability in single-molecule FRET
experiments. Nucleic Acids Res. 42, 5967–5977 (2014).

71
附錄
FigureA 1 螢光共振能量轉移效率-時間軌跡圖。HT21 序列在原兒茶酸與兒茶酸-3,4-雙加氧酶
除氧系統中鉀離子濃度為 2mM、5mM、10mM、20mM 和 50mM 條件下螢光共振能量轉移效
率有變動的軌跡圖。

72
FigureA 2 螢光共振能量轉移效率-時間軌跡圖。A_HT21 序列在原兒茶酸與兒茶酸-3,4-雙加氧
酶除氧系統中鉀離子濃度為 2mM、5mM、10mM、20mM 和 50mM 條件下螢光共振能量轉移
效率有變動的軌跡圖。

73
FigureA 3 螢光共振能量轉移效率-時間軌跡圖。N_HT21 序列在原兒茶酸與兒茶酸-3,4-雙加氧
酶除氧系統中鉀離子濃度為 2mM、5mM、10mM、20mM 和 50mM 條件下螢光共振能量轉移
效率有變動的軌跡圖。

74
FigureA 4 螢光共振能量轉移效率-時間軌跡圖。T4_HT21 序列在原兒茶酸與兒茶酸-3,4-雙加
氧酶除氧系統中鉀離子濃度為 2mM、5mM、10mM、20mM 和 50mM 條件下螢光共振能量轉
移效率有變動的軌跡圖。

75
FigureA 5 HT21、A_HT21 和 N_HT21 在不同鉀離子濃度下其動力學反應速率在未摺疊狀態
(U)與 F1(EFRET~0.6)和未摺疊狀態(U)F2(EFRET~0.8)之間的曲線圖。
FigureA 6 HT21、A_HT21 和 N_HT21 在不同鉀離子濃度下其動力學反應速率在
F1(EFRET~0.6)與 F2(EFRET~0.8)之間的曲線圖。

76
FigureA 7 螢光共振能量轉移效率直方圖, A_HT21、N_ HT21 和 T4_HT21 序列在葡萄糖氧
化酶-過氧化氫酶除氧系統中分別為不同濃度的鉀離子與鎂離子條件下所得到的實驗數據。

77
FigureA 8 螢光共振能量轉移效率直方圖,HT21、A_HT21、N_ HT21 和 T4_HT21 序列在葡
萄糖氧化酶-過氧化氫酶除氧系統中分別為不同濃度的鈉離子條件下所得到的實驗數據。
FigureA 9 螢光共振能量轉移效率直方圖,HT21 序列在吡喃糖氧化酶-過氧化氫酶除氧系統中
分別為不同濃度的鉀離子與鎂離子條件下所得到的實驗數據。


















