
Decreased Degradative Enzymes in Mesangial Cells Cultured in
7
Decreased Degradative Enzymes in Mesangial Cells Cultured in High Glucose Media David J. Leehey, Ruo Hua Song, Nahid Alavi, and Ashok K. Singh Abnormalities in extracellular matrix degradation may play a pathogenetic role in diabetic nephropathy. Cul- tured renal mesangial cells are known to synthesize increased amounts of matrix proteins when incubated in high glucose media (e.g., 30 mmol/L). However, the effect of glucose loading on degradative enzymes is unknown. Primary cultures of rat mesangial cells were grown until confluent in the presence of fetal calf serum (FCS) and insulin (0.67 U/ml). Cells were then cultured for 7 days in plastic wells in either 10 or 30 mmol/1 glucose media containing neither FCS nor insulin. Collagenase activity in media were determined by zymography and quantita- tive spectrofluorometry. Cathepsin B and D activities in cell extracts were measured by spectrofluorometry (us- ing the fluorescent substrate Z-Arg-Arg-7-amido-4-meth- ylcoumarin) and 125 I-labeled hemoglobin digestion, re- spectively. Gelatin-degrading activity of live mesangial cells was also determined. mRNA levels for collagenase IV, cathepsin B, and cathepsin D were determined by Northern analysis. A major band of collagenase activity with a molecular size of 72 kDa was observed in all mesangial cell media. Exposure of cells to high glucose media resulted in significant reductions in collagenase and cathepsin B activities as well as impairment in gela- tin-degrading activity. Collagenase IV and cathepsin B and D mRNA levels were also decreased by glucose load- ing. To exclude the possibility that glucose loading was injurious to cells, 3 H-leucine uptake (as a measure of protein synthesis) and membrane alkaline phosphatase activity (as a biochemical marker of viability) were not affected by the high glucose condition. We conclude that proteinase activity is decreased in mesangial cells incu- bated in high glucose media. This may be due to changes in levels of gene expression (mRNA) of these enzymes. Diabetes 44:929-935, 1995 D iabetic nephropathy is characterized by accumu- lation of matrix material in the mesangial area of glomeruli. Normal extracellular matrix consists of several protein components, including type IV collagen, laminin, fibronectin, entactin, and proteoglycan, of which collagen IV is the major structural component (1). Accumulation of extracellular matrix in diabetes results from an alteration in the balance between matrix synthesis and degradation. Increased accumulation of extracellular matrix components and increased matrix protein gene ex- pression (mRNA) have been observed in mesangial cells incubated in high glucose media (2-6). These data support the hypothesis that the high-glucose condition of diabetes stimulates the synthesis of matrix. However, the effect of high glucose on matrix degradative pathways in these cells is unknown. Degradation of matrix is accomplished by extracellular matrix metalloproteinases (MMPs) such as collagenases and intracellular lysosomal enzymes or cathepsins, which further digest partially degraded molecules after uptake by the cells (1,7). Studies in the rat made diabetic by injection of streptozotocin have indicated that both collagenase and cathepsin activities are decreased in this model (8). Since mesangial cells are known to synthesize and secrete proteo- lytic enzymes (9-11), we believed that glucose-loaded me- sangial cells would be an excellent model to study the effects of the diabetic condition on these mechanisms of extracel- lular matrix degradation. We therefore designed experiments to answer the following questions: i ) Are collagenase and/or cathepsin B or D activities altered in glomerular mesangial cells incubated in high glucose media? 2) Is altered protein- ase activity due to changes in levels of gene expression (mRNA) of these enzymes? From the Hines Veterans Affairs Hospital, Hines; Loyola University Stritch School of Medicine, Maywood; and Section of Renal Diseases, Department of Medicine, University of Illinois at Chicago, Chicago, Illinois (N.A.). Address correspondence and reprint requests to Dr. David J. Leehey, 111-L, Veterans Affairs Hospital, Hines, IL 60141. Received for publication 6 October 1994 and accepted in revised form 20 April 1995. AMC, Z-Arg-Arg-7-amido-4-methylcoumarin; APMA, aminophenyl mercuric ace- tate; FCS, fetal calf serum; FITC, fluorescein isothiocyanate; MMP, matrix metal- loproteinase; PAI, plasminogen activator inhibitor; PBS, phosphate-buffered saline; SSC, sodium chloride-sodium citrate; SDS, sodium dodecyl sulfate; TCA, trichlo- roacetic acid; TGF, tumor growth factor; TIMP, tissue inhibitor of metalloprotein- RESEARCH DESIGN AND METHODS Culture of mesangial cells. All experiments were conducted on primary mesangial cells obtained from normal rat glomeruli as outlined below. Glomeruli were isolated from 100-150 g male Sprague-Dawley rats using a graded-sieving technique with stainless steel and nylon meshes under sterile conditions (12). Isolated glomeruli were then plated for culture in RPMI 1640 tissue culture medium containing 20% fetal calf serum (FCS), insulin (0.67 U/ml), and antibiotics. The identity of mesangial cells was confirmed by light and electron microscopy by detection of typical microfilaments. Mesangial cells were grown until confluent in the presence of FCS and insulin (0.67 U/ml) in 24-well multi-well culture dishes (Corning Glass, Corning, NY). Once cells were confluent, cells were cultured in media containing either 10 or 30 mmol/1 glucose without added insulin, hi experiments in which proteinase activities were determined, cells were incubated for 7 days in the absence of FCS. This was done because the large amount of protein in FCS would greatly decrease the ability to DIABETES, VOL. 44, AUGUST 1995 929 Downloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022
Transcript of Decreased Degradative Enzymes in Mesangial Cells Cultured in

Decreased Degradative Enzymes in Mesangial CellsCultured in High Glucose MediaDavid J. Leehey, Ruo Hua Song, Nahid Alavi, and Ashok K. Singh
Abnormalities in extracellular matrix degradation mayplay a pathogenetic role in diabetic nephropathy. Cul-tured renal mesangial cells are known to synthesizeincreased amounts of matrix proteins when incubated inhigh glucose media (e.g., 30 mmol/L). However, the effectof glucose loading on degradative enzymes is unknown.Primary cultures of rat mesangial cells were grown untilconfluent in the presence of fetal calf serum (FCS) andinsulin (0.67 U/ml). Cells were then cultured for 7 days inplastic wells in either 10 or 30 mmol/1 glucose mediacontaining neither FCS nor insulin. Collagenase activityin media were determined by zymography and quantita-tive spectrofluorometry. Cathepsin B and D activities incell extracts were measured by spectrofluorometry (us-ing the fluorescent substrate Z-Arg-Arg-7-amido-4-meth-ylcoumarin) and 125I-labeled hemoglobin digestion, re-spectively. Gelatin-degrading activity of live mesangialcells was also determined. mRNA levels for collagenaseIV, cathepsin B, and cathepsin D were determined byNorthern analysis. A major band of collagenase activitywith a molecular size of 72 kDa was observed in allmesangial cell media. Exposure of cells to high glucosemedia resulted in significant reductions in collagenaseand cathepsin B activities as well as impairment in gela-tin-degrading activity. Collagenase IV and cathepsin Band D mRNA levels were also decreased by glucose load-ing. To exclude the possibility that glucose loading wasinjurious to cells, 3H-leucine uptake (as a measure ofprotein synthesis) and membrane alkaline phosphataseactivity (as a biochemical marker of viability) were notaffected by the high glucose condition. We conclude thatproteinase activity is decreased in mesangial cells incu-bated in high glucose media. This may be due to changesin levels of gene expression (mRNA) of these enzymes.Diabetes 44:929-935, 1995
Diabetic nephropathy is characterized by accumu-lation of matrix material in the mesangial area ofglomeruli. Normal extracellular matrix consistsof several protein components, including type IV
collagen, laminin, fibronectin, entactin, and proteoglycan, ofwhich collagen IV is the major structural component (1).Accumulation of extracellular matrix in diabetes resultsfrom an alteration in the balance between matrix synthesisand degradation. Increased accumulation of extracellularmatrix components and increased matrix protein gene ex-pression (mRNA) have been observed in mesangial cellsincubated in high glucose media (2-6). These data supportthe hypothesis that the high-glucose condition of diabetesstimulates the synthesis of matrix. However, the effect ofhigh glucose on matrix degradative pathways in these cells isunknown.
Degradation of matrix is accomplished by extracellularmatrix metalloproteinases (MMPs) such as collagenases andintracellular lysosomal enzymes or cathepsins, which furtherdigest partially degraded molecules after uptake by the cells(1,7). Studies in the rat made diabetic by injection ofstreptozotocin have indicated that both collagenase andcathepsin activities are decreased in this model (8). Sincemesangial cells are known to synthesize and secrete proteo-lytic enzymes (9-11), we believed that glucose-loaded me-sangial cells would be an excellent model to study the effectsof the diabetic condition on these mechanisms of extracel-lular matrix degradation. We therefore designed experimentsto answer the following questions: i) Are collagenase and/orcathepsin B or D activities altered in glomerular mesangialcells incubated in high glucose media? 2) Is altered protein-ase activity due to changes in levels of gene expression(mRNA) of these enzymes?
From the Hines Veterans Affairs Hospital, Hines; Loyola University Stritch Schoolof Medicine, Maywood; and Section of Renal Diseases, Department of Medicine,University of Illinois at Chicago, Chicago, Illinois (N.A.).
Address correspondence and reprint requests to Dr. David J. Leehey, 111-L,Veterans Affairs Hospital, Hines, IL 60141.
Received for publication 6 October 1994 and accepted in revised form 20 April1995.
AMC, Z-Arg-Arg-7-amido-4-methylcoumarin; APMA, aminophenyl mercuric ace-tate; FCS, fetal calf serum; FITC, fluorescein isothiocyanate; MMP, matrix metal-loproteinase; PAI, plasminogen activator inhibitor; PBS, phosphate-buffered saline;SSC, sodium chloride-sodium citrate; SDS, sodium dodecyl sulfate; TCA, trichlo-roacetic acid; TGF, tumor growth factor; TIMP, tissue inhibitor of metalloprotein-
RESEARCH DESIGN AND METHODSCulture of mesangial cells. All experiments were conducted onprimary mesangial cells obtained from normal rat glomeruli as outlinedbelow. Glomeruli were isolated from 100-150 g male Sprague-Dawleyrats using a graded-sieving technique with stainless steel and nylonmeshes under sterile conditions (12). Isolated glomeruli were thenplated for culture in RPMI 1640 tissue culture medium containing 20%fetal calf serum (FCS), insulin (0.67 U/ml), and antibiotics. The identityof mesangial cells was confirmed by light and electron microscopy bydetection of typical microfilaments.
Mesangial cells were grown until confluent in the presence of FCSand insulin (0.67 U/ml) in 24-well multi-well culture dishes (CorningGlass, Corning, NY). Once cells were confluent, cells were cultured inmedia containing either 10 or 30 mmol/1 glucose without added insulin,hi experiments in which proteinase activities were determined, cellswere incubated for 7 days in the absence of FCS. This was done becausethe large amount of protein in FCS would greatly decrease the ability to
DIABETES, VOL. 44, AUGUST 1995 929
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022

GLUCOSE DECREASES MESANGIAL DEGRADATIVE ENZYMES
detect changes in proteinase activity using zymography. In experimentsin which gelatin-degrading activity was measured, live mesangial cellswere incubated for 14 days in media containing FCS (see measurementof gelatin-degrading activity). FCS was used in these experiments fortwo reasons: 1) zymographic analysis was not planned, and thus therewas no concern that FCS would impair the ability to detect proteinaseactivity and 2) prolonged incubation of cells without FCS might result insome loss of cell viability. Media was not changed during incubations.However, cells were repeatedly examined (every 1-2 days) duringincubation for evidence of detachment. Glucose concentrations ofmedia were determined using a glucometer (Glucometer 3 analyzer,Miles, Elkhart, IN) both at the beginning and at the end of the incubationperiods. To control for possible osmotic effects of the high glucosemedium, experiments in which cells were incubated in 10 mmol/1glucose plus 20 mmol/1 mannitol were also performed.Qualitative assessment of collagenase IV (gelatinase) activity.Zymography was used to demonstrate the presence of collagenase IV(gelatinase) activity in mesangial cell culture media (13). Twice-crystal-lized gelatin (1 mg/ml) was incorporated into an 8% sodium dodecylsulfate (SDS)-polyacrylamide gel electrophoresis. Samples of mediawere concentrated 25-fold by lyophilization and reconstitution. Twentymicrograms of protein was mixed with sample buffer (lacking 3-mer-captoethanol and modified to contain a final concentration of 2.5% SDS)and electrophoresed under nonreducing conditions. After electrophore-sis, SDS was eluted from the gel in 2.5% Triton X-100 for 60 min at 25°C.The gel was then incubated in substrate buffer (50 mmol/1 Tris-HCl, pH8.0, containing 5 mmol/1 CaCl2 and 1 |xmol/l ZnCl2) at 37°C for 16 h. Afterstaining with Coomassie Blue R-250, gelatin-degrading enzymes wereidentified by their ability to clear the substrate at their respectivemolecular weights.
Fluorometric assay for free collagenase activity. Cell media usuallycontain a mixture of metalloproteinases and their inhibitors, i.e., tissueinhibitors of metalloproteinases (TIMPs). Free collagenase activityrepresents that activity which is in excess of TIMP-inhibited activity andthus reflects the amount of free enzyme contained in the media. Freecollagenase activity was determined by a tube assay modified from themethod of Watanabe et al. (14). Porcine gelatin (Sigma, St. Louis, MO)was purified by crystallization with ethanol. The purified product (molwt —20 kDa) at a concentration of 10 mg/ml was mixed with a 20-molarexcess of fluorescein isothiocyanate (FITC) (Sigma) in 50 mmol/1phosphate buffer, pH 9.5. The mixture was incubated at room tempera-ture overnight and then extensively dialyzed against physiological salinefor 2-3 days at room temperature to remove free fluorescein. The finalmolar ratio of gelatin-to-fluorescein was determined to be 1:2. Thegelatin/fluorescein conjugate was diluted 1:100 (i.e., to 100 |xg/ml) in 100mmol/1 Tris-HCl, pH 7.4, containing 50 mmol/1 CaCl2. For assay, mediafrom mesangial cells incubated in phenol red-free RPMI (phenol redwas found to interfere with the assay) were used. A total of 50 |xl ofundiluted and diluted (1:5 and 1:25) samples were preincubated with12.5 mU/ml of purified plasmin (Sigma) at 37°C for 30 min. In prelimi-nary experiments, we used aminophenyl mercuric acetate (APMA) toactivate latent collagenase (13); however, we found that plasmin wasmore effective in this regard. Subsequently, all quantitative measure-ments of collagenase reported in this manuscript were performed in thepresence of plasmin. Addition of plasmin to activate latent collagenasehas been previously used by Wong et al. (15). A total of 200 |xl of dilutedconjugate was added to all tubes (20 \xg of gelatin/tube) and the tubesreincubated at 37°C for 24 h. After incubation, 1.0 ml of Tris-HCl wasadded to all tubes and fluorescence quantitated using excitation andemission wavelengths of 491 run and 519 ran, respectively. The excita-tion and emission spectra of the unhydrolyzed gelatin/fluorescein con-jugate were substantially shifted from the spectrum of free FITC; thus,at the wavelengths used, the conjugate accounted for only 5% of the totalfluorescence observed. Activity calculations were corrected for plasmin-induced hydrolysis (—10%). Clostridial collagenase (Sigma) and Trisbuffer were used as positive and negative controls, respectively. Theamount of gelatin degraded was determined by comparing samplereadings against a standard curve prepared by maximally digesting 1-20|xg/ml of FITC-gelatin with clostridial collagenase. The assay is able todetect 5 |xg/tube of digested gelatin with an intra-assay variation of
Measurement of gelatin-degrading activity by cultured mesangialcells. Primary mesangial cells were cultured in 24-well dishes in RPMI1640 tissue culture media containing 20% FCS, insulin (0.67 \jJml~) andantibiotics. Once cells were confluent, cells were incubated in phenolred-free media containing 12.5 |xg/ml FITC-labeled gelatin, FCS, plasmin
(12.5 mU/ml), and either 10 mmol/1 glucose, 30 mmol/1 glucose, or 10mmol/1 glucose plus 20 mmol/1 mannitol. Cells were incubated for 1, 2,3, 4, 7, 11, and 14 days. After incubation, 200 |xl of culture media werecollected and diluted with 1.0 ml of 100 mmol/1 Tris HC1, pH 7.4. Freefluorescence was quantitated at excitation and emission wavelengths of491 nm and 519 ran, respectively. The amount of gelatin degraded wasdetermined by comparing these readings against a standard curveprepared by maximally digesting 1-12 ixg/ml of FITC-gelatin withclostridial collagenase. Blanks included FITC-gelatin and plasmin (12.5mU/ml) incubated at 37°C in normal media in the absence of cells for asimilar number of days.Cathepsin assays
Cathepsin B. Cultured mesangial cells were washed three times withcold physiological saline and then scraped from the wells into a smallvolume (1-2 ml/well) of cold saline. These cell suspensions were madein Triton X-100 (final concentration 0.1%) and sonicated in the cold for1 min. Cathepsin B activity in these cell preparations was measuredusing a spectofluorometric method previously described by Achkar et al.(16). Protein (10 |xg) of each sample was diluted in 0.1 ml of phosphate-buffered saline (PBS) and added to an equal volume of an activationbuffer (30 mmol/1 dithiothreitol, 15 mmol/1 EDTA, pH 5.2, and pepsin 50|xg/ml) for 30 min at 25°C. After incubation, samples were mixed withthe auto-quenched fluorescent substrate Z-Arg-Arg-7-amido-4-methyl-coumarin (AMC) (Bachem Bioscience, Philadelphia, PA; 94 fxmol/1/0.2mol/1 citrate-phosphate buffer, pH 6.2). The AMC products were quanti-tated using excitation and emission wavelengths of 370 and 460 nm,respectively. Rates of increase in fluorescence intensity were comparedwith an AMC standard to obtain product concentrations.
Activation buffer by itself did not degrade the substrate.Cathepsin D. Cathepsin D was measured in cell extracts prepared inidentical manner to that described above for cathepsin B by quantitatingthe digestion of 125I-hemoglobin (17). Denatured hemoglobin (Sigma)was radiolabeled with Na 125I using the chloramine T method (18). Fivemicrograms of mesangial cell protein was mixed with 250 ng of labeledhemoglobin in citrate buffer (20 mmol/1), pH 4.8, containing 1 mmol/1CaCl2 and 0.1% Triton X-100 in a total volume of 100 |xl. The mixture wasincubated at 37°C for 18 h, after which the reaction was stopped byaddition of 0.2 ml of 10% BSA and 2 ml of 20% trichloroacetic acid (TCA).The resulting precipitate was collected by centrifugation and 7-ac-counted. The TCA soluble fraction represented the digested fraction.Alkaline phosphatase assay. Mesangial cells were extracted by soni-cation in saline containing 0.1% Triton X-100 in cold for 1 min. Onehundred microliters of each sample was added to 200 (JLI of 15 mmol/1Tris buffer pH 8.8 containing 1 mmol/1 CaCl2, 0.1% Triton X-100, and 25mmol/1 p-nitrophenyl phosphate (Sigma). The mixture was incubated at37°C for 18 h, after which the reaction was stopped by addition of 1 mlof 1.0 N NaOH. The mixture was read at 405 nm in a spectrophotometer(Beckman DU 64, Beckman, Fullerton, CA). Absorbance units wereconverted to nanomoles of p-nitrophenol using 8,800 as the molarextinction coefficient of p-nitrophenol determined experimentally bydiluting purified p-nitrophenol (Sigma) with the stopping solution.Measurement of mRNA for collagenase IV, cathepsin B, andcathepsin DcDNA probes. A cDNA probe for collagenase IV (72 kDa) was obtainedfrom Dr. G.L. Goldberg, Washington University School of Medicine, St.Louis, MO. A 6-kb insert containing the 3.1-kb 72 kDa collagenase IVcDNA was ligated at the Smal site of the vector p Bluescript KS (±)(Stratagene) and propagated in E. coli. The 72-kDa collagenase IV cDNAprobe was recovered as a 2-kb fragment after digestion of the isolatedplasmid with XbayEcoRVBaml restriction enzymes. cDNA probes forcathepsins B and D were obtained from Dr. A. Frankfater, LoyolaUniversity School of Medicine, Maywood, IL. The cathepsin B probecontains the 1.2-kb EcoRl fragment corresponding to mouse prepro-cathepsin B cloned into the pGEM2 vector. The cDNA was originallyobtained from the \-gtlO library of mouse melanoma (19). This probehas been shown to cross-react with rat mRNA (20). The cathepsin Dprobe consisted of human cathepsin D cDNA (2.3 kb). This probe hasbeen shown to cross-react with rat cathepsin D (20). Probes werelabeled with a32P-CTP using the random hexamer priming method(Promega, Madison, WI).Total RNA isolation and Northern blotting. All chemicals, includingsolvents, were of the highest analytical grade tested for negative RNAseactivity and were purchased from Sigma. Total RNA was isolated fromcultured cells by homogenization in 4 mol/1 guanidium thiocyanatecontaining p-mercaptoethanol and extraction with phenol/chloroform.The RNA was precipitated at -70°C with isopropanol and redissolved in
930 DIABETES, VOL. 44, AUGUST 1995
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022

D.J. LEEHEY AND ASSOCIATES
distilled water containing 0.1% diethylpyrocarbonate. After denaturingwith formamide and formaldehyde, 15 |xg RNA (as measured at A260)from each sample was electrophoresed in a 1.1% agarose gel at 120 V for4 h. Equivalency of sample loading was verified by ethidium bromidestaining of the 28S and 18S rRNA bands. After electrophoresis, RNA wastransferred to a nylon filter by capillary blotting for 16-20 h andcross-linked by ultraviolet light. The filter was then prehybridized in 50%formamide, 1 mol/1 NaCl, 10 mmol/1 NaH2PO4, 5 X Denhardt's solution,1% SDS, and 250 |xg/ml salmon sperm DNA at 42°C for 4 h followed byhybridization in the same buffer with 2 X 106 cpm/ml of the 32P-labeledprobe. Incubation was continued overnight at 42°C. The niters contain-ing the type IV collagenase cDNA probe were washed with 2 X sodiumchloride (0.15 mol/l)/sodium citrate (0.015 mol/1) (SSC) and 0.1% SDSthree times for 5 min at room temperature followed by two washes in 1X SSC and 0.1% SDS for 20 min at 50°C. The niters containing thecathepsin probes were washed with 0.2 x SSC and 0.5% SDS 3 x for 20min each at 65°C. Filters were subsequently exposed to X-ray film at-70°C for 24 h (cathepsin) or 72 h (collagenase).Densitometric analysis. Autoradiographic bands for each probe afterNorthern blotting were quantitated using scanning densitometry (Bio-Rad 620 scanner, Richmond, CA). To correct for differences in RNAloading, the density of the 28S ribosomal RNA band on the photographicnegative of the ethidium-bromide stained gel was also determined, andthe relative densities of the probe and the ribosomal RNA band werecompared.3H-leucine uptake. Total protein synthesis by cultured mesangial cellswas estimated by measuring the incorporation rate of 3H-labeled leucineinto cells. After 7 days of incubation in either 10 or 30 mmol/1 glucosemedium, cells were pulsed with 1 (j-Ci/ml of 3H-leucine (ICN, CostaMesa, CA) and incubated for an additional 18 h. Cells were then washedwith cold PBS and extracted in 0.1% SDS and the extracts precipitatedwith 10% TCA. After redissolving in 1 mol/1 NaOH, the precipitates werecounted in a P-scintillation counter. Incorporation of leucine into cellprotein was estimated by the counts precipitable with TCA.Data analysis. Data from cells incubated under the three-test condi-tions were compared using analysis of variance (ANOVA) followed bythe Tukey-Kramer multiple comparisons test. A P value of <0.05 wasconsidered to be significant. Data are expressed as mean ± SE.
RESULTSCell culture. By light microscopy, mesangial cell morphol-ogy was not different when cells were cultured in 10 vs. 30mmol/1 glucose media for up to 7 days. Cells were >95%viable after 7 days when tested using trypan blue exclusion.Moreover, cells remained attached to plastic throughout theincubation period when either 10 or 30 mmol/1 glucose mediawas used. Treatment of mesangial cells with 30 mmol/1glucose did not significantly change the de novo proteinsynthetic rate as measured by 3H-leucine counts incorpo-rated into cell protein (10 mmol/1 glucose: 1.23 ± 0.47 X 105
cpm/18 h/mg protein; 30 mmol/1 glucose: 1.01 ± 0.15 x 105
cpm/18 h/mg protein; n = 16 each, P > 0.1). Moreover,alkaline phosphatase in cell extracts was not different be-tween the groups (10 mmol/1 glucose: 35.3 ± 2.8nmobrT^mg"1 protein; 30 mmol/1 glucose: 44.2 ± 2.7; 10mmol/1 glucose + 20 mmol/1 mannitol: 36.6 ± 4.1, (n = 4 ineach group, P > 0.1). Media glucose concentrations deter-mined at the beginning and end of incubation periods werenot different (data not shown). Cell viability, cell attachment,leucine uptake, and alkaline phosphatase activities weremaintained for 7 days whether or not FCS was included inthe media (data not shown).Collagenase activity in mesangial cell supernatants andextracts. By zymography the major band of gelatinolyticactivity observed had a molecular size of 72 kDa, and thusrepresents the 72 kDa collagenase IV (gelatinase). Lesserbands of activity were observed at ~90 kDa and 150 kDapositions. Exposure of cells to high-glucose media resultedin a marked reduction in gelatinase activity when compared
132
94 _
6 6 _
lOmMG
30mMG
Mcontrol
FIG 1. Effect of 30 mmol/1 glucose media on collagenases secreted bymesangial cells. The picture shows typical zymograms of media obtainedfrom mesangial cells incubated for 7 days in either 10 mmol/1 glucose(G), 30 mmol/1 glucose, or 10 mmol/1 glucose plus 20 mmol/1 mannitol (Mcontrol). A decrease in the activity of the 72 kDa type IV collagenase(gelatinase) was observed with glucose loading but not with exposure tomannitol. Faint bands of activity at 92 kDa and 150 kDa were also notedin cells incubated in 10 mmol/1 glucose.
to that observed in control cells (Fig. 1). There was also asignificant (~40%) reduction in free collagenase activity(after plasmin activation) in cells treated with 30 mmol/1glucose (Fig. 2). Free collagenase activity could not bedetected unless plasmin was added to the incubation. Incu-bation of cells in media containing 10 mmol/1 glucose plus 20mmol/1 mannitol (to control for possible effects of theincreased osmolality of the high-glucose medium) did notdecrease enzyme activity. Enzyme activity from cells incu-bated in the various media were as follows: 10 mmol/1glucose: 171 ± 7.6 |xg gelatin digested/18 h/mg protein; 30mmol/1 glucose: 104 ± 4.3; 10 mmol/1 glucose + 20 mmol/1
150T
•£ 1 0 0 -oo
50-
I—I 10 mM Glucose^ 30 mM GlucoseE 3 Mannitol control
I
II
Collagenase Cathepsin B Cathepsin D
FIG. 2. Quantitation of the 72 kDa type IV collagenase (gelatinase) andcathepsins from mesangial cells treated with high-glucose media for 7days. Free collagenase activity and cathepsin activities from mesangialcells incubated in 10 mmol/1 glucose, 30 mmol/1 glucose, or 10 mmol/1glucose plus 20 mmol/1 mannitol (mannitol control) are depicted (n = 6each). The assay buffer for collagenase activity contained plasmin toactivate latent collagenase. Cells treated with 30 mmol/1 glucose secretedapproximately 40% less gelatinase into the media. Cells treated with 30mmol/1 glucose had approximately 20% less cathepsin B activity. Noeffects on enzyme activity were seen when cells were incubated in anisoosmotic mannitol-containing media. Data are presented as mean ± SE.The asterisk denotes statistical significance at P < 0.05 compared to 10mmol/1 glucose.
DIABETES, VOL. 44, AUGUST 1995 931
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022
GLUCOSE DECREASES MESANGIAL DEGRADATIVE ENZYMES
10 mM GMannitol
30 mM G
72kdColllV Cathepsin B Cathepsin 0
1 2 3 4DAYS
FIG. 3. Amount of gelatin digested by mesangial cells incubated ineither 10 mmol/1 glucose (G) (O), 30 mmol/1 glucose ( • ) , or 10 nunol/1glucose plus 20 mmol/1 mannitol (mannitol) (A) (n = 4 for each timepoint). All incubation media contained plasmin as an activator ofcollagenase. The amount of gelatin degraded was significantly decreasedby glucose loading but not by mannitol; this effect was statisticallysignificant (P < 0.05) after 4 days of incubation and was maintained forup to 14 days of incubation in the high glucose media.
mannitol: 164 ± 6.4; n = 6 per group. When enzyme activitieswere normalized for cell number rather than amount ofprotein, incubation in high glucose media was still observedto decrease activity (data not shown).
To assure that decreased collagenase activity in the mediawas not caused by impaired secretion of enzymes by the cellsinto the media, cell extracts were also examined by zymog-raphy. There was a complete absence of collagenase activityin the extracts even when lanes were loaded with up to 60 |xgof protein (data not shown).Degradation of gelatin by live mesangial cells. Livemesangial cells incubated in 30 mmol/1 glucose degradedFITC-gelatin at an ~20% slower rate than did cells incubatedin either 10 mmol/1 glucose or 10 mmol/1 glucose plus 20mmol/1 mannitol. This difference was apparent after 4 days ofmcubation and was maintained for up to 14 days of incuba-tion in the high-glucose media (Fig. 3). A linear relationshipbetween the amount of gelatin digested and the number ofdays that cells were incubated in the various media wasobserved (10 mmol/1 glucose, r = 0.97; 30 mmol/1 glucose, r= 0.97; 10 mmol/1 glucose plus 20 mmol/1 mannitol, r = 0.99).As indicated in METHODS, plasmin was added to the incuba-tion media in these experiments to activate latent collage-nase.Cathepsin activity. Cathepsin B activity was significantlyreduced by ~20% by glucose loading (Fig. 2). Cathepsin Dactivity was also somewhat decreased by glucose loading,but this difference did not reach statistical significance.Incubation with 20 mmol/1 mannitol plus 10 mmol/1 glucosegave similar results to incubation in 10 mmol/1 glucose alone.Enzyme activities from cells incubated in the various mediawere as follows: Cathepsin B: 10 mmol/1 glucose: 1,566 ± 132pmol/min/mg protein; 30 mmol/1 glucose: 1,273 ± 144; 10mmol/1 glucose + 20 mmol/1 mannitol: 1,791 ± 103; n = 6 pergroup; Cathepsin D: 10 mmol/1 glucose: 1.28 ± 0.21 jxg Hb/24h/mg protein; 30 mmol/1 glucose: 1.1 ± 0.06; 10 mmol/1glucose + 20 mmol/1 mannitol: 0.97 ± 0.03; n = 6 per group.Quantitation of mRNA. To determine whether decreaseddegradative enzyme activities were due to decreases inmRNA levels, the mRNAs for 72 kDa collagenase and cathe-psins B and D were probed with specific cDNAs using
3.1kb
NG HG
2.1 • !kb
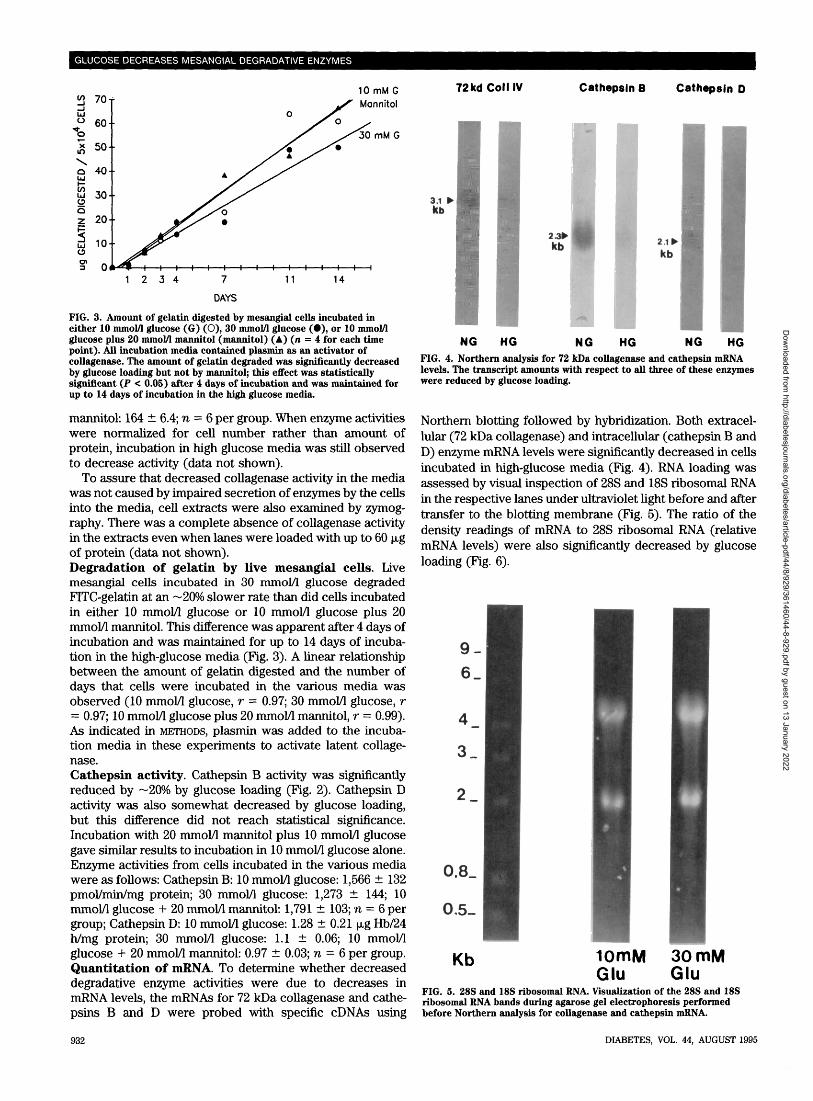
NG HGFIG. 4. Northern analysis for 72 kDa collagenase and cathepsin mRNAlevels. The transcript amounts with respect to all three of these enzymeswere reduced by glucose loading.
Northern blotting followed by hybridization. Both extracel-lular (72 kDa collagenase) and intracellular (cathepsin B andD) enzyme mRNA levels were significantly decreased in cellsincubated in high-glucose media (Fig. 4). RNA loading wasassessed by visual inspection of 28S and 18S ribosomal RNAin the respective lanes under ultraviolet light before and aftertransfer to the blotting membrane (Fig. 5). The ratio of thedensity readings of mRNA to 28S ribosomal RNA (relativemRNA levels) were also significantly decreased by glucoseloading (Fig. 6).
9-6_
3_
2_
0.8_
0.5-
Kb 10mMGlu
30 mMGlu
FIG. 5. 28S and 18S ribosomal RNA. Visualization of the 28S and 18Sribosomal RNA bands during agarose gel electrophoresis performedbefore Northern analysis for collagenase and cathepsin mRNA.
932 DIABETES, VOL. 44, AUGUST 1995
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022

D.J. LEEHEY AND ASSOCIATES
1.000T
0.800-
0.600-
® 0.400-
0.200-
0.000
I—I 10 mM GlucoseE 3 30 mM Glucose
Collogenose IV Cothepsin B Cothepsin DFIG. 6. Densitometric analysis of autoradiograms of Northern blots forcollagenase IV and cathepsins. To normalize for RNA loading conditions,mRNA values were compared to 28S ribosomal RNA values. RelativemRNA levels were calculated from the ratio of mRNA to 28S ribosomalRNA density readings.
DISCUSSIONThe etiology of extracellular matrix expansion in diabeticnephropathy remains to be fully elucidated. In animal modelsof the disease, matrix accumulation is associated with in-creased synthesis of matrix proteins (21-24). Since mesang-ial matrix expansion is the most characteristic lesion of bothexperimental and human diabetes, studies using glomerularmesangial cells in culture have been recently used in anattempt to further discern the mechanism of matrix accumu-lation. From these studies, it is clear that short-term glucoseloading increases matrix protein synthesis (2-6), althoughprolonged exposure of mesangial matrix to elevated glucosemay reduce collagen synthesis (25).
Mesangial matrix expansion in diabetes probably resultsfrom both increased synthesis and impaired degradation,since it is evident that degradative mechanisms are inade-quate to prevent matrix accumulation. Both extracellularMMPs, such as collagenases, and intracellular lysosomalenzymes or cathepsins are involved in the degradation ofextracellular matrix (1,7). MMP precursors may be activatedvia plasmin-dependent cleavage reactions (7,15). Moreover,the plasminogen/plasmin system may contribute directly tomatrix degradation independent of its role in MMP activation(7). There are various collagenases demonstrable in glomer-ular cells that are probably important in the degradation ofmatrix (9-11,13,14,26-28). Of these, the 72-kDa collagenaseIV (gelatinase) is the principal type secreted by renal me-sangial cells in culture (9-11). TIMPs serve to regulate theactivity of MMPs by forming noncovalent complexes andthus inactivating them (7). Our results demonstrate thatincubation of mesangial cells in high-glucose media results insignificant decreases in both collagenase and cathepsin Bactivity. These decrements in enzyme activity were associ-ated with decreases in enzyme mRNA levels, suggesting thatreduced gene transcription may be the etiology. The rele-vance of the reduced degradative enzyme activities wassubstantiated by the observation that live mesangial cellshad an impaired ability to degrade gelatin in the presence ofhigh glucose. These results suggest that, in addition toincreased synthesis, decreased degradation of matrix maycontribute to mesangial matrix accumulation in diabetes.The effect of glucose loading on the matrix degrading en-zymes was specific, as another marker enzyme (alkalinephosphatase) was unchanged under the same conditions.
The decrease in enzyme activity also could not be explainedby the effect of high glucose on de-novo protein syntheticrate, as the latter was not significantly altered by the exper-imental treatment.
We used two methods of assessing collagenase activity,namely zymography and quantitative assay of free collage-nase activity. When zymography is used, the high concentra-tion of SDS in the gel results in dissociation of thenoncovalent bonds between collagenases and TIMPs. SinceTIMPs do not co-electrophorese with collagenase, zymogra-phy measures total collagenase, i.e., enzyme that was previ-ously inactivated by inhibitors (TIMPs) as well as unboundor free enzyme. Thus, results using this technique are indic-ative of the amount of enzyme that is secreted by the cell intothe culture media. The marked impairment in gelatinaseactivity using zymography coupled with the marked decre-ment in 72-kDa gelatinase mRNA levels suggests that alteredsynthesis of this enzyme is at least in part responsible for thedecreased enzyme activity observed.
On the other hand, our quantitative fluorometric assaymeasures free collagenase activity, or net enzyme activity inthe presence of inhibitor substances. Thus, findings usingthis assay may better reflect actual collagenase activity in theglomerular mesangium in vivo. As noted in METHODS andRESULTS, we were unable to detect enzyme activity using thisassay unless plasmin was added to the incubation. Ourresults are similar to those of Wong et al. (15), who alsofound that the plasminogen/plasmin system was necessary inorder for mesangial cells to degrade labeled matrix substrate(Matrigel). In these studies, media from mesangial cellsgrown on labeled Matrigel in the absence of plasminogencontained two bands of gelatinase activity (mol wt 70-72kDa), but little matrix degradation occurred unless plasmin-ogen was added to the incubation. These data both suggestthat secreted collagenase is present in an inactive form as aprocollagenase. The latency of procollagenases appears tobe maintained by a linkage between an unpaired cysteineresidue in the propeptide to the zinc ion present at the activesite (7). Disruption of this bond and subsequent enzymeactivation can be achieved in several ways in vitro, includingcatalytic removal of amino acids from the aminoterminus ofthe molecule, interaction or modification of the cysteineresidue by organomercurials such as APMA, or conforma-tional change that exposes the active site by detergents suchas SDS (7). The normal mode of activation of collagenases(including the 72-kDa collagenase IV) in vivo is unknown butis believed to involve the plasminogen/plasmin system (29).
Since we did not directly assay for TIMP activity, wecannot rule out the possibility that an increase in TIMPactivity contributed to the glucose-induced inhibition of freecollagenase activity. In this regard, Kitamura et al. (30) foundthat gene expression of the metalloproteinase transin(stromelysin), which degrades casein and other matrix mol-ecules, is suppressed in mesangial cells exposed to highglucose while mRNA levels for TIMP were simultaneouslyincreased. However, increased TIMP levels cannot be thesole explanation for the decrease in free collagenase activitybecause gelatinase activity using zymography (i.e., in theabsence of TIMP) was still impaired.
It has been suggested that alterations in the plasminogen/plasmin system, e.g., an increase in plasminogen activatorinhibitors (PAIs) could impair collagenase activity in me-sangial cells exposed to high glucose (31). However, this
DIABETES, VOL. 44, AUGUST 1995 933
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022

GLUCOSE DECREASES MESANGIAL DEGRADATIVE ENZYMES
issue is controversial, as Tada et al. (32) recently reportedthat glucose-loaded human mesangial cells produced lessPAI-1 associated with increases in the synthesis of plasmin-ogen activators. In any event, our data suggests that glucose-induced impairment of free collagenase activity can occurindependently of any changes in the activity of the plasmin-ogen/plasmin system, because, in the performance of ourassay, we added plasmin to activate MMP precursors toMMPs. Addition of plasmin to the assay system would beexpected to mask any effects of glucose on the plasminogen/plasmin system.
Our data demonstrate that not only degradative enzymesynthesis and activity but also actual gelatin degradation bymesangial cells is impaired by incubation of the cells inhigh-glucose media. Similar results with respect to matrixdegradation were recently reported by McLennan and col-leagues (33). These investigators prepared radioactively la-beled mesangial matrix from mesangial cell cultures andassessed degradation by monitoring the release of radiolabelinto the culture medium. Incubation in 30 mmol/1 glucoseeither during incorporation of radioactivity into matrix orduring monitoring of degradation reduced the rate of matrixdegradation. These data are in support of those describedabove from our laboratory. Our collagenase results are inseeming contrast to those recently reported by Wang et al(34). These investigators found that glucose-loading in-creases 72-kDa MMP activity and mRNA. However, theirexperiments were done in murine mesangial cells that hadbeen incubated in glucose for only 24 h. It is possible that anearly temporary increase in collagenase activity may occurbut that activity declines to subnormal values with moreprolonged incubation. These investigators did not measurecollagenase activity at later time points; thus, it is possiblethat differences in experimental protocol explain our differ-ing results.
Lysosomal cathepsin activity has been found to be de-creased in the kidneys of diabetic animals (35-37), but hasnot previously been studied in mesangial cell culture. Similarto our findings, Monaham and Bose (35) found that cathepsinB but not cathepsin D activity was depressed in diabetic rats.Our studies are the first to document alterations in cathepsinactivity in mesangial cells exposed to high-glucose condi-tions. The presence of insulin does not enhance glucoseuptake by mesangial cells (38). Our studies were conductedin the absence of insulin to both simulate the insulin-deficient diabetic state and to obviate possible difficulties indata interpretation due to the growth-stimulating effects ofinsulin on cultured cells. In our shorter-term experiments,FCS was also omitted from the media to facilitate detectionof collagenase by zymography, which cannot be performedwhen samples contain large amounts of protein. Since theabsence of FCS can adversely affect the health of growingcells, studies were conducted using nondividing confluentcells. In such cells, removal of FCS from the media did notadversely affect the cell layer during the one-week period ofincubation as evidenced by preservation of normal morphol-ogy and ability to attach to plastic dishes, normal trypan blueexclusion, and ability to take up amino acids.
The mechanism of the effect of glucose loading on colla-genase and cathepsin activities was not addressed in thisstudy. However, it is possible that autocrine activation oftransforming growth factor p (TGF-(3) is mediating theseeffects, at least with respect to collagenase. This cytokine
has been demonstrated in a number of systems to increaseextracellular matrix accumulation. This effect is mediated byits unique ability among cytokines to stimulate matrix pro-duction while inhibiting matrix degradation and increasingcell adhesion to matrix (39). Glucose can increase TGF-(3secretion (40) and gene expression (41) by mesangial cells.In preliminary work from our laboratory, TGF-(3 inhibition bya neutralizing antibody reversed the inhibitory effect ofglucose loading on collagenase activity in mesangial cells(42). Of note, increased TGF-pJ activity and gene transcrip-tion has also been demonstrated in the streptozotocin rat(43), suggesting that these in vitro findings are relevant to thein vivo diabetic state.
ACKNOWLEDGMENTSThese studies were supported in part by a grant from theNational Institutes of Health (DK-35804). Linda Wanna, MS,aided in the preparation and maintenance of cell cultures.
This work was presented at the 1993 Annual Meetings ofthe American Society of Nephrology and has been publishedin abstract form (J Am Soc Nephrol 3:803A, 1993).
REFERENCES1. Davies M, Martin J, Thomas GJ, Lovett DH: Proteinases and glomerular
matrix turnover. Kidney Int 41:671-678, 19922. Ayo SH, Radnik RA, Garoni J, Glass WF, Kreisberg JI: High glucose
causes an increase in extracellular matrix proteins in cultured mesangialcells. Am J Path 136:1339-1348, 1990
3. Ayo SH, Radnik RA, Garoni JA, Glass WF, Rampt ER, Appling DR,Kreisberg JI: Increased extracellular matrix synthesis and mRNA inmesangial cells grown in high-glucose medium. Am J Physiol 260:F185-F191, 1991
4. Danne T, Spiro MJ, Spiro RG: Effect of high glucose on type IV collagenproduction by cultured epithelial, endothelial, and mesangial cells.Diabetes 42:170-177, 1993
5. Nahman NS Jr, Leonhart KL, Cosio FG, Hebert CL: Effects of high glucoseon cellular proliferation and flbronectin production by cultured humanmesangial cells. Kidney Int 41:396-402, 1992
6. Haneda M, Kikkawa R, Horide N, Togawa M, Koya D, Kajiwara N, MaedaS, Shigeta Y: Glucose-induced overproduction of type IV collagen incultured glomerular mesangial cells. J Diabetic Complications 5:199-200, 1991
7. Birkedal-Hansen H: Role of matrix metalloproteinases in human peri-odontal diseases. J Periodontol 64:474-484, 1993
8. Reckelhoff JF, Tygart VL, Mitias M, Walcott JL: STZ-induced diabetesresults in decreased activity of glomerular cathepsin and metalloproteasein rats. Diabetes 42:1425-1432, 1993
9. Martin J, Davies M, Thomas G, Lovett DH: Human mesangial cells secretea GBM-degrading neutral proteinase and a specific inhibitor. Kidney Int36:790-801, 1989
10. Lovett DH, Johnson RJ, Martin H, Martin J, Davies M, Couser WG:Structural characterization of the mesangial cell type IV collagenase andenhanced expression in a model of immune complex-mediated glomer-ulonephritis. Am J Pathol 141:85-98, 1992
11. Baricos WH, Shah SV: Proteinases in renal disease. Kidney Int 40:163-173, 1991
12. Kasinath BS, Singh AK, Kanwar YS, Lewis EJ: Puromycin aminonucleo-side specifically reduces heparan sulfate proteoglycan core proteincontent of cultured glomerular epithelial cells. Am J Physiol 24:F590-F596, 1988
13. Davies M, Thomas GJ, Martin J, Lovett DH: The purification andcharacterization of a glomerular-basement-membrane-degrading neutralproteinase from rat mesangial cells. Biochem J 251:419-425, 1988
14. Watanabe K, Kinoshita S, Nakagawa H: Gelatinase secretion by glomer-ular epithelial cells. Nephron 56:405-409, 1990
15. Wong AP, Cortez SL, Baricos WH: Role of plasmin and gelatinase inextracellular matrix degradation by cultured rat mesangial cells. Am JPhysiol 263:F1112-F1118, 1992
16. Achkar C, Gong Q, Frankfater A, Bajkowski AS: Differences in targetingand secretion of cathepsins B and L by BALB/3T3 fibroblasts andMoloney murine sarcoma virus-transformed BALB/3T3 fibroblasts. JBiolChem 265:13650-13654, 1990
17. Singh AK: Proteolytic machinery of the glomerular epithelial cell againstIgG. Biophys Res Comm 186:639-644, 1992
18. McConahey PJ, Dixon FJ: A method to trace iodination of proteins for invivo studies. Int Arch Allergy Appl Immunol 29:185-189, 1966
19. Qian F, Chan SJ, Gong Q, Bajkowski AS, Steiner DF, Frankfater A: The
934 DIABETES, VOL. 44, AUGUST 1995
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022

D.J. LEEHEY AND ASSOCIATES
expression of cathepsin B and other lysosomal proteinases in normaltissues and in tumors. Biomed Biochim Ada 50:531-540, 1991
20. Qian F, Frankfater A, Miller RV, Chan SJ, Steiner DF: Molecular cloningof rat precursor cathepsin H and the expression of five lysosomalcathepsins in normal tissues and in a rat carcinosarcoma. Int J Biochem22:1457-1464, 1990
21. Cohen MP, Khalifa A: Renal glomerular collagen synthesis in streptozo-tocin diabetes: reversal of increased basement membrane synthesis withinsulin therapy. Biochim, Biophys Ada 500:395-404, 1977
22. Abrass CK, Peterson CV, Raugi GJ: Phenotypic expression of collagentypes in mesangial matrix of diabetic and nondiabetic rats. Diabetes37:1695-1702, 1988
23. Fukui M, Nakamura T, Ebihara I, Shirato I, Tomino Y, Koide H: ECM geneexpression and its modulation by insulin in diabetic rats. Diabetes41:1520-1527, 1992
24. Ihm C, Lee GSL, Nast CC, Artishevsky A, Guillermo R, Levin PS, GlassockRJ, Adler SG: Early increased renal procollagen cd(rV) mRNA levels instreptozotocin induced diabetes. Kidney Int 41:768-777, 1992
25. Silbiger S, Crowley S, Shan Z, Brownlee M, Satriano J, Schlondorff D:Nonenzymatic glycation of mesangial matrix and prolonged exposure ofmesangial matrix to elevated glucose reduces collagen synthesis andproteoglycan charge. Kidney Int 43:853-864, 1993
26. Le Q, Shah S, Nguyen H, Cortez S, Baricos W: A novel metalloproteinasepresent in freshly isolated rat glomeruli. Am J Physiol 260:F555-F561,1991
27. Martin J, Knowlden J, Davies M, Williams JD: Characterization of neutralproteinase synthesis by cultured human glomerular cells (Abstract). JAm Soc Nephrol 2:579A, 1991
28. Johnson R, Yamabe H, Chen YP, Campbell C, Gordon K, Baker P, LovettD, Couser W: Glomerular epithelial cells secrete a glomerular basementmembrane-degrading metalloproteinase. J Am Soc Nephrol 2:1388-1397,1992
29. Tryggvason K, Hoyhtya M, Salo T: Proteolytic degradation of extracellu-lar matrix in tumor invasion. Biochim Biophys Ada 907:191-217, 1987
30. Kitamura M, Kitamura A, Mitarai T, Maruyama N, Nagasawa R,Kawamura T, Yoshida H, Takahashi T, Sakai O: Gene expression ofmetalloproteinase and its inhibitor in mesangial cells exposed to highglucose. Biochem Biophys Res Comm 185:1048-1054, 1992
31. Brees DK, Narita I, Ketteler M, Noble NA, Border WA: Effect of high
glucose on the expression of plasminogen activator inhibitor (PAI-1) inrat mesangial cells (Abstract). J Am Soc Nephrol 4:647A, 1993
32. Tada H, Tsukamoto M, Ishii H, Igosai S: A high concentration of glucosealters the production of tPA, uPA and PAI-1 antigens from humanmesangial cells. Diabetes Res Clin Pract 24:33-39, 1994
33. McLennan SV, Fisher EJ, Yue DK, Turtle JR: High glucose concentrationcauses a decrease in mesangium degradation: a factor in the pathogen-esis of diabetic nephropathy. Diabetes 43:1041-1045, 1994
34. Wang ZJ, Bocchicchio J, Fadly M, Sorbi D, Arbeit LA: Glucose increases72 kDa metalloproteinase activity (Abstract). J Am Soc Nephrol 5:974A,1994
35. Mohanam S, Bose SM: Influence of streptozotocin- and alloxan-induceddiabetes in the rat on collagenase and certain lysosomal enzymes inrelation to the degradation of connective tissue proteins. Diabetologia25:66-70, 1983
36. Nerurkar MA, Satav JG, Katyare SS: Insulin-dependent changes inlysosomal cathepsin D activity in rat liver, kidney, brain and heart.Diabetologia 31:119-122, 1988
37. Belfiore F, Rabuazzo AM, Iannello S, Campione R, Vasta D: Anabolicresponse of some tissues to diabetes. Biochem Med Metab Biol 35:149-155, 1986
38. Kreisberg JI, Ayo SH: The glomerular mesangium in diabetes mellitus.Kidney Int 43:109-113, 1993
39. Border WA, Noble NA: Cytokines in kidney disease: the role of trans-forming growth factor-p. Am J Kidney Dis 22:105-113, 1993
40. Jian Y, Li L: The production of total and active transforming growthfactor-3 in cultured mesangial cells and isolated glomeruli under highglucose conditions (Abstract). J Am Soc Nephrol 4:809A, 1993
41. Wolf G, Sharma K, Chen Y, Ericksen M, Ziyadeh FN: High glucose-induced proliferation of mesangial cells is reversed by autocrine TGF-(3.Kidney Int 42:647-656, 1992
42. Song RH, Singh AK, Alavi N, Leehey DJ: Decreased collagenase activity ofmesangial cells incubated in high glucose media is reversed by neutral-izing antibody to transforming growth factor-beta (Abstract). JASN4:972A, 1994
43. Yamamoto T, Nakamura T, Noble NA, Ruoslahti E, Border WA: Expres-sion of transforming growth factor (3 is elevated in human and experi-mental diabetic nephropathy. ProcNatAcad Sci USA 90:1814-1818,1993
DIABETES, VOL. 44, AUGUST 1995 935
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/44/8/929/361460/44-8-929.pdf by guest on 13 January 2022


















